escape, resist, or tolerate? evolution of defence
TRANSCRIPT

Escape, Resist, or Tolerate? Evolution of Defence Strategies in Response
to Glyphosate Herbicide in an Agricultural Weed (Amaranthus palmeri)
by
Zachary Teitel
A Thesis
presented to
The University of Guelph
In partial fulfilment of requirements
for the degree of
PhD
in
Integrative Biology
Guelph, Ontario, Canada
© Zachary Teitel, September, 2021

ii
ABSTRACT
ESCAPE, RESIST, OR TOLERATE? EVOLUTION OF DEFENCE STRATEGIES IN
RESPONSE TO GLYPHOSATE HERBICIDE IN A NOXIOUS AGRICULTURAL
WEED
Zachary Teitel Advisor:
University of Guelph, 2021 Associate Professor C.M. Caruso
Weeds can respond to herbicide stress with three defence strategies: escape,
resistance, and tolerance. Escape occurs when a weed changes the timing of its life cycle
to avoid a herbicide; resistance occurs when a weed reduces the extent of damage it
receives from a herbicide; and tolerance occurs when a weed withstands damage from a
herbicide, but compensates for the loss of fitness resulting from that damage. Whether a
weed population evolves to escape, resist, or tolerate a herbicide depends on tradeoffs
among defence strategies, limits to the evolution of defence strategies, and selection on
defence strategies in competitive agricultural environments. I examined how each of
these factors shape the evolution of defence strategies in the noxious agricultural weed,
Amaranthus palmeri S. Wats., in response to glyphosate herbicide. First, I examined
whether escape and tolerance are correlated both within and among populations, which
would indicate whether defence strategies are trading off. Second, I tested whether
genotype by environment interactions (GEI) and estimated fitness costs have the potential
to limit the evolution of escape and tolerance. Third, I measured selection acting on
glyphosate escape and resistance within two environmental contexts: a high competition
corn (Zea mays L.) crop environment and a low competition no crop environment. I
found that both within and among populations there were no tradeoffs between escape

iii
and tolerance, suggesting that populations can potentially evolve high magnitudes of both
escape and tolerance. Further indicating this, I did not find fitness costs for escape or
tolerance in the absence of glyphosate, though one potential limit I discovered was from a
GEI, showing that variable environments could limit the evolution of escape and
tolerance. Finally, though I found no selection acting on glyphosate escape, there was
selection for increased resistance in the low competition environment, and selection
against high magnitudes of resistance in the high competition environment, indicating
that competition with crops could slow the evolution of glyphosate resistance. Overall, I
found that glyphosate escape and tolerance are less likely to evolve than resistance,
though the evolution of resistance could be managed by manipulating the competitive
crop growing environment.

iv
ACKNOWLEDGEMENTS
I first want to thank my incredible PhD advisor, Chris Caruso, for her unwavering
support, expertise, and enthusiasm for my scholarly pursuits over the last six years. I am
constantly reminded through our day-to-day interactions just how lucky I am to have
found a mentor so devoted to her students’ success. For the countless times that she has
so effortlessly showed me the way, I will always be grateful.
My advisory committee consisting of Chris Caruso, Hafiz Maherali, Brian
Husband, Clarence Swanton, Regina Baucom, and David Jordan often seems like the
world’s largest committee, and the breadth of knowledge and experience that they have
consistently brought to my thesis reinforces that notion. Thank you all for your unique
contributions that have shaped my progress and understanding of this complex and
fascinating field.
I’ve received an astonishing amount of practical and conceptual help since I
began my research from a wide variety of people that I’m so grateful to have worked
with. I wish to thank David Jordan, Andrew Hare, Stanley Culpepper, Wes Everman,
Patrick Tranel, Jeremy Kichler, Ronnie Barentine, Kurt Maertens, Eric Alinger, Kevin
Johnson, Brian Schoon, Nikki Keitner, Lanae Ringler, and Larry Uthell for help sampling
populations across the USA. Thank you to Andrew Hare, Denis Mahoney, Stephen Deal,
and David Jordan for providing constant expert support in the field when I needed it the
most in a new environment. Thank you to all the volunteers and research assistants
including Ann Lee, Sarah McDonald, Aaron Hudson, Emily Williams, Jennifer Wood,
Jasmin Dawson, Lucy Burns, Chloe Katsademas, and Kayleigh Hutt-Taylor, who
tirelessly helped me in the lab and greenhouse, and for treating my work like their own.

v
Thank you to greenhouse managers Michael Mucci and Tannis Slimmon, who were
exceptionally committed to my experiments and to so many others lucky enough to have
worked with them. You both went so far above and beyond what anyone might expect of
you, and you had fun doing it.
I thank my BSc advisor Spencer Barrett and MSc advisor Lesley Campbell for
encouraging me to continue in evolutionary research, and for providing the foundations
and tools I needed to succeed in my doctorate. Thank you to past and present members of
the Caruso and Maherali Labs for providing constant helpful insights on paper drafts,
presentations, and results. I also wish to thank Sarah Yakimowski for continually
reigniting my curiosity and fascination with this work through our collaborations.
My research was funded by a NSERC Alexander Graham Bell Canada Graduate
Scholarship – Doctoral, a NSERC Michael Smith Foreign Study Supplement Award, an
Arthur D. Latornell Graduate Travel Grant, and a University of Guelph – Department of
Integrative Biology PhD Award. Thank you to all these generous funding groups.
I thank my amazing family and friends for giving me the love, kindness, and
laughter that I undeniably needed after moving away from home. Finally, thank you to
my extraordinary life partner, Ilana Arnold, for always being there for me no matter what.
You’ve read through drafts, entered data, provided tech support, shown fascination for
and always encouraged my academic pursuits, but that does not begin to scratch the
surface of how you’ve helped me these last six years. I love you very much.

vi
TABLE OF CONTENTS
Abstract ................................................................................................................................ii
Acknowledgements .............................................................................................................. iv
Table of Contents ................................................................................................................ vi
List of Tables ..................................................................................................................... viii
List of Figures ..................................................................................................................... ix
Chapter 1: Introduction ...................................................................................................... 1
1.1 BACKGROUND ................................................................................................................... 1
1.2 STUDY SYSTEM ................................................................................................................. 3
1.3 OVERVIEW .......................................................................................................................... 4
Chapter 2: Are there trade-offs between glyphosate escape and tolerance in the
agricultural weed Amaranthus palmeri? ............................................................................ 6
2.1 ABSTRACT ........................................................................................................................... 6
2.2 INTRODUCTION ................................................................................................................. 6
2.3 METHODS .......................................................................................................................... 10 2.3.1 Study System ............................................................................................................................... 10 2.3.2 Seed Collection ............................................................................................................................ 11 2.3.3 Within-Population Correlation Experiment ................................................................................ 12 2.3.4 Among-Population Correlation Experiment................................................................................ 13 2.3.5 Measuring Escape and Tolerance ................................................................................................ 14 2.3.6 Statistical Analysis ...................................................................................................................... 16
2.4 RESULTS ............................................................................................................................ 16
2.5 DISCUSSION ...................................................................................................................... 17
2.6 FIGURES ............................................................................................................................. 22
Chapter 3: What Could Limit the Evolution of Escape and Tolerance to Glyphosate in an
Agricultural Weed (Amaranthus palmeri)? ...................................................................... 24
3.1 ABSTRACT ......................................................................................................................... 24
3.2 INTRODUCTION ............................................................................................................... 25
3.3 METHODS .......................................................................................................................... 29 3.3.1 Study System ............................................................................................................................... 29 3.3.2 Seed Collection ............................................................................................................................ 30 3.3.3 Fitness Costs Experiment ............................................................................................................ 30 3.3.4 GEI Experiment ........................................................................................................................... 33 3.3.5 Statistical Analysis ...................................................................................................................... 36
3.4 RESULTS ............................................................................................................................ 37
3.5 DISCUSSION ...................................................................................................................... 38
3.6 FIGURES ............................................................................................................................. 44

vii
Chapter 4: Crop competition causes nonlinear selection for glyphosate resistance in an
agricultural weed (Amaranthus palmeri) ......................................................................... 46
4.1 ABSTRACT ......................................................................................................................... 46
4.2 INTRODUCTION ............................................................................................................... 47
4.3 METHODS .......................................................................................................................... 50 4.3.1 Study System ............................................................................................................................... 51 4.3.2 Seed Collection ............................................................................................................................ 51 4.3.3 Experimental Design ................................................................................................................... 52 4.3.4 Statistical Analysis ...................................................................................................................... 55
4.4 RESULTS ............................................................................................................................ 57
4.5 DISCUSSION ...................................................................................................................... 60
4.6 TABLES .............................................................................................................................. 66
4.7 FIGURES ............................................................................................................................. 71
Chapter 5: Conclusion ...................................................................................................... 74
5.1 OVERVIEW ........................................................................................................................ 74
5.2 FUTURE DIRECTIONS AND SIGNIFICANCE ............................................................... 75
Chapter 6: Literature cited ............................................................................................... 78

viii
LIST OF TABLES
Table 4.6.1: ....................................................................................................................... 66
Table 4.6.2: ....................................................................................................................... 67
Table 4.6.3: ....................................................................................................................... 68
Table 4.6.4: ....................................................................................................................... 69
Table 4.6.5: ....................................................................................................................... 70

ix
LIST OF FIGURES
Figure 2.6.1: ...................................................................................................................... 22
Figure 2.6.2: ...................................................................................................................... 23
Figure 3.6.1: ...................................................................................................................... 44
Figure 3.6.2: ...................................................................................................................... 45
Figure 4.7.1: ...................................................................................................................... 71
Figure 4.7.2: ...................................................................................................................... 72
Figure 4.7.3: ...................................................................................................................... 73

1
Chapter 1: Introduction
1.1 BACKGROUND
Plants are faced with an array of stresses that impair their development, survival, and
reproduction (Larcher 2003), including biotic stresses such as herbivory, microbial
pathogens, and plant competitors (Pons et al. 1998); as well as abiotic stresses such as
drought, salinity, extreme temperature, and herbicide exposure (Jenks and Hasegawa
2008). Plants can respond to these biotic and abiotic stresses through three defence
strategies: escape, resistance, and tolerance (Xiao et al. 2007, Agrawal 2000). Resistance
occurs when a plant reduces the extent of damage it receives from a biotic or abiotic
stress (Mauricio et al. 1997). Escape occurs when a plant changes the timing of its life
cycle to avoid a biotic or abiotic stress (Hilgenfeld et al. 2004). Finally, tolerance occurs
when a plant bears damage from a biotic or abiotic stress, but compensates for the
resultant loss of fitness (Mauricio et al. 1997). Whether a plant population evolves to
escape, resist, or tolerate a biotic or abiotic stress will depend on a multitude of factors
including tradeoffs among defence strategies, limits to the evolution of defence strategies,
and selection on defence strategies in competitive agricultural environments.
For my PhD dissertation, I studied how multiple factors can affect the evolution of
escape, resistance, and tolerance in response to pesticide application. Since the
introduction of genetically engineered crops in 1996, until 2011, pesticide use in the USA
has increased by 7% (Benbrook 2012). In particular, herbicides with the active ingredient
glyphosate have globally increased 15-fold since 1996, making this herbicide one of the
strongest stresses that plants have been subject to in modern agriculture (Baylis 2000,

2
Benbrook 2016). Part of the widespread appeal of glyphosate (i.e. RoundUp®) to farmers
is that it acts as a generalist herbicide once plants have emerged, and does not remain in
the soil system after use. As well, glyphosate inhibits the chloroplast enzyme EPSPS
(Steinrücken and Amrhein 1980), a necessary component of plant growth and metabolism
(Herrmann and Weaver 1999). Because glyphosate attacks such a physiologically
essential process, it has become a widely used herbicide, with USA herbicide usage in
2001 being more than 60% glyphosate (Duke and Powles 2008).
In response to the widespread application of glyphosate herbicide, many weed
populations have evolved high magnitudes of resistance (Heap 2018), though far less is
known about escape and tolerance. Resistance to glyphosate has been found in 41 weed
species (Heap 2018). Among species, the mechanism of glyphosate resistance can vary
considerably; herbicide-resistant plants can resist through target-site mutations to the
EPSPS gene, through reduced glyphosate translocation within the plant, and through gene
amplification of the EPSPS enzyme (Powles and Preston 2006, Gaines et al. 2010,
Powles 2010). Relative to the evolution of herbicide resistance, much less is known about
the evolution of escape and tolerance as herbicide stress responses. Glyphosate escape
occurs when weeds avoid the exclusively post-emergent effects of early-season
glyphosate application by shifting the emergence time of their seedbanks to later in the
season (Hilgenfeld 2004). Seedbanks in typical agricultural fields contain a large
diversity of weed species with different phenologies (Forcella et al. 1992), so to avoid
detrimental crop-weed competition at the crop’s most vulnerable life-history stage,
herbicides will often be applied early in the growing season. This early application of
glyphosate should select for the evolution of later-emerging seeds (Jordan and Jannink

3
1997). Tolerance occurs when weeds sustain the damaging effects of glyphosate by
reallocating resources to vital reproductive systems in order minimize a loss in fitness
(Mauricio et al. 1997). In doing so, a tolerant plant sprayed with glyphosate would retain
the same or similar seed production as an unsprayed plant, while a non-tolerant plant
sprayed with glyphosate would have a diminished seed set. The evolution of tolerance
would be ineffective for plants that express no resistance since they do not survive to
reproductive maturity, or redundant on plants that express complete resistance since they
go completely undamaged. Though 41 weed species have been documented to display
glyphosate resistance (Heap 2018), few researchers have attempted to document tolerance
(e.g. Ipomoea purpurea; Baucom and Mauricio 2004) or escape (Scursoni et al. 2007) as
defense mechanisms. It is therefore unknown how common escape and tolerance stress
responses are relative to resistance in glyphosate-stressed environments.
1.2 STUDY SYSTEM
To study the evolution of glyphosate escape, resistance, and tolerance, I used the
agricultural weed, Amaranthus palmeri. I collected seed for use in my research from 22
A. palmeri populations across Illinois, North Carlina, and Georgia, USA in 2016. The C4
summer-annual A. palmeri is prevalent and ranked one of the most troublesome weeds of
cotton, corn, and soybean in the USA (Sauer 1957, Webster et al. 2001, Webster and
Nichols 2012, Ward et al. 2013). It’s rapid growth rate, competitive ability, high
fecundity, and wide genetic diversity has led to significant economic losses in crop
production, making it a heavily targeted species for both farmers and researchers. As
well, resistance to glyphosate has quickly spread far in A. palmeri, making it an ideal

4
system to study rapid evolution in response to a post-emergent herbicide. Confirmed
glyphosate-resistant A. palmeri was first detected in 2005 in Georgia (Culpepper et al.
2006) and has since been found in 27 states and Brazil (Heap 2018). No fitness costs
were found in assessing glyphosate-resistance in A. palmeri (Giacomini et al. 2014),
which could help explain why resistance is prevalent. Somewhat uniquely among weeds,
A. palmeri is both dioecious (Grant 1959) and wind pollinated. These traits could
exacerbate the spread of resistance genes by enforcing and facilitating obligate
outcrossing, respectively (Sosnoskie et al. 2007).
1.3 OVERVIEW
To determine what factors influences the evolution of glyphosate escape,
resistance, and tolerance in A. palmeri, I examined tradeoffs among defence strategies,
limits to the evolution of defence strategies, and selection on defence strategies in
competitive agricultural environments. First, I determined whether glyphosate escape and
tolerance are trading off by measuring correlations between them (Chapter 2).
Correlations were measured at both the population and family level in common garden
field experiments in 2017 and 2018. Tradeoffs resulting from negative genetic
correlations between escape and tolerance would indicate that both defence strategies are
unlikely to evolve together. I then determined if there were limits to evolving glyphosate
defence strategies by measuring fitness costs and genotype by environment interactions
(GEI) for escape and tolerance (Chapter 3). Fitness costs of escape and tolerance in the
absence of glyphosate stress were measured in the field in 2017, and GEI were looked for

5
by comparing field and greenhouse measures of escape and tolerance in 2017. Finally, I
determined whether competitively stressful environments can influence selection for
defence strategies (Chapter 4). Selection on glyphosate resistance and tolerance was
measured in the field in 2018 for A. palmeri grown in in both a competitive corn-crop
environment, and a non-competitive no-crop environment.

6
Chapter 2: Are there trade-offs between glyphosate escape and
tolerance in the agricultural weed Amaranthus palmeri?
2.1 ABSTRACT
Plants can evolve multiple defence strategies in response to environmental stress,
including escape (i.e., avoiding a stress by changing the timing of life history events), and
tolerance (i.e., compensating for damage from the stress through the reallocation of
resources). These two defence strategies can evolve independently of each other or be
trading off such that the expression of one may depend on the expression of the other. To
test for tradeoffs between escape and tolerance, I measured the expression of these
defence strategies at the population and family level in response to glyphosate herbicide
in the agricultural weed, Amaranthus palmeri. This was done using 22 populations as
well as a separate 30-family synthetic population in a common garden agricultural field in
North Carolina, USA. At the population level, escape and tolerance were not correlated,
at least indicating that populations with high expression of one defence strategy did not
have high expression of the other strategy. At the family level, escape and tolerance were
also not correlated, indicating that plants could evolve both high magnitudes of escape
and tolerance. The potential evolution of high magnitudes of both of these herbicide
defence strategies could make it more challenging for weed suppression efforts.
2.2 INTRODUCTION
Plants in natural and agricultural environments can evolve multiple defence

7
strategies in response to the biotic and abiotic stresses they face (Larcher 2003, Powles
and Yu 2010, Agrawal 2011, Lipiec et al. 2013). The ability of plants to evolve more than
one defence strategy can enhance and diversify their overall defensive function. Multiple
modes of defence can in turn create an evolutionary trajectory that selects for stronger
and more varied types of herbivore or human-mediated herbicide stresses (Leimu et al.
2012). In plant systems where multiple alternative defence strategies exist, there are two
possible outcomes for their evolution. One possibility is that plants express a single
defence strategy. This could occur if alternative defence strategies are costly to express or
because multiple strategies are redundant (Herms and Mattson 1992, Rosenthal and
Kotanen 1994, Fineblum and Rausher 1995, Baucom and Mauricio 2008a). A second
possibility is that plants express multiple defence strategies. This could occur if multiple
defence strategies are not costly to express or because multiple strategies are not
redundant (Steward and Keeler 1988, van der Meijden et al. 1988, Koricheva et al. 2004,
Leimu and Koricheva 2006, Agrawal 2011, Romeo et al. 2013). Distinguishing between
these two possibilities can provide insight into how defence strategies will impact each
other’s evolutionary trajectories.
Whether a single defence strategy or multiple defence strategies evolve will
depend on the correlation between defence strategies. Multiple defence strategies could
persist in a population if they do not trade off with each other. Tradeoffs can be identified
by testing for a negative genetic correlation between two defence strategies (Leimu and
Koricheva 2006). For example, negative genetic correlations between induced and
constitutive herbivore defences in common milkweed were interpreted as evidence of
tradeoffs between types of defence (Bingham and Agrawal 2010). If there are negative

8
genetic correlations between defence strategies within a population, then selection for one
strategy should indirectly select against the other strategy, making it unlikely that both
strategies will evolve simultaneously at the population level (Fineblum and Rausher
1995). However, if there is not a correlation between defence strategies within a
population, then evolution in response to selection for one strategy will be independent of
selection for the other strategy (Bingham and Agrawal 2010). Thus, if both defence
strategies incur positive fitness benefits, there will be selection for both strategies. With
selection for both, populations can simultaneously evolve multiple defence strategies,
potentially strengthening plant defence. Although much evidence suggests that negative
genetic correlations should not always be expected and that tradeoffs are uncommon
(Mauricio et al. 1997, Stinchcombe and Rausher 2002), they are still sometimes detected
(Fineblum and Rausher 1995, Baucom and Mauricio 2008a) and should still be tested for.
This is especially true in systems with new stresses where tradeoffs have rarely been
tested, and in new environments that can influence tradeoffs (Sgro and Hoffmann 2004).
To determine whether there are tradeoffs between defence strategies, I studied
defences against herbicide application in an agricultural weed. Two alternative defence
strategies that agricultural weeds can evolve in response to herbicide application are
escape and tolerance. Escape occurs when the timing of a plant’s life cycle allows it to
avoid herbicide stress (Hilgenfeld et al. 2004). For example, late seedling emergence in
agricultural weeds allowed for escape from the post-emergent glyphosate herbicide
applications by avoiding contact with it (Scursoni et al. 2007). Alternatively, tolerance
occurs when a plant can sustain herbicide damage while maintaining its overall fitness
(Mauricio et al. 1997, Baucom and Mauricio 2004). For example, tolerance to glyphosate

9
herbicide in an agricultural weed was determined by the ability of plants to produce as
many seeds following herbicide damage as unsprayed plants (Baucom and Mauricio
2004). Although defence tradeoffs have been commonly studied in response to other
stresses like herbivory and drought, examples of studies measuring tradeoffs in response
to herbicide are rare (Baucom and Mauricio 2008a). Further, research on agricultural
weed herbicide defence typically focusses on resistance only (Duke and Powles 2009).
The evolution of herbicide escape and tolerance in agricultural weeds is very rarely
examined and no assessments have been made on tradeoffs between these two herbicide
defence strategies. However, if these two defence strategies are not genetically correlated,
and there is selection for both, then there could be populations of weeds that evolve to
both escape and tolerate herbicide application.
I estimated tradeoffs between escape and tolerance in response to glyphosate
herbicide in the agricultural weed Amaranthus palmeri. Glyphosate resistant A. palmeri
was first detected in 2005 in Georgia (Culpepper et al. 2006) and has since been found in
28 states and Brazil (Heap 2018). As such, A. palmeri is prevalent across the USA and
ranked at or near the top of many crops’ “most troublesome weed” lists, including cotton
(Gossypium hirsutum L.), soybean (Glycine max (L.) Merr.), peanut (Arachis hypogaea
L.), and corn (Sauer 1957, Webster et al. 2001, Webster and Nichols 2012). Though there
have been no tests of whether A. palmeri has evolved escape and tolerance, not finding
tradeoffs would indicate multiple glyphosate defence strategies could evolve together,
creating an especially difficult scenario to manage. Thus, examining evolutionary
tradeoffs for glyphosate escape and tolerance in A. palmeri may be vital in efforts to
control proliferation of this relatively new and noxious pest.

10
To determine whether there are trade-offs between herbicide defence strategies, I
estimated correlations between glyphosate escape and tolerance in A. palmeri. First, to
determine if selection for one defence strategy affects selection for another defence
strategy, I estimated the family-mean correlation (a proxy for the genetic correlation; Via
1984) between glyphosate escape and tolerance of A. palmeri in a common garden
agricultural field. This will tell me if there are tradeoffs between escape and tolerance
which would prevent multiple strategies from evolving simultaneously within
populations. Second, to determine whether these tradeoffs are affecting the composition
of defence strategies among populations, I estimated correlations between glyphosate
escape and tolerance across populations of A. palmeri in a common garden agricultural
field. This will tell me, for example, if high expressions of multiple strategies have
evolved. I investigated the current state and future evolutionary potential of alternative
glyphosate defence strategies in A. palmeri by asking the following two questions:
1. Is there a tradeoff between glyphosate escape and tolerance, as evidenced by a
negative genetic correlation across families of A. palmeri?
2. Are glyphosate escape and tolerance correlated across populations of A. palmeri?
2.3 METHODS
2.3.1 Study System
Amaranthus palmeri is a wind-pollinated C4 summer-annual agricultural weed
that originated in the Sonoran Desert (Sauer 1957) and has spread recently via animal
vectors, farming equipment, and seed contamination throughout midwestern and
southeastern USA (Hensleigh and Pokorny 2017). Amaranthus palmeri is dioecious and

11
an obligate outcrosser (Ward et al. 2013). In hot and sunny environments, A. palmeri can
grow up to 5 cm/day (Horak and Loughin 2000) due to its high photosynthetic rate (81
mol/m2/s, Ehleringer 1983), enabling females to produce as many as 600,000 seeds
(Keeley et al. 1987). Amaranthus palmeri mostly emerge between March and June,
typically flower 5-9 weeks after emergence, and produce viable seed as early as 2-3
weeks after flowering. Although plants can emerge as late as October, lethal November
frosts can come before they are able to set seed (Keeley et al. 1987).
2.3.2 Seed Collection
To estimate correlations between glyphosate escape and tolerance, from
September to October in 2016, I collected seeds of A. palmeri from 22 populations spread
across their Eastern USA range in Georgia, North Carolina, and Illinois. Populations were
selected by contacting agricultural extension agents and researchers. A population was
defined as a discrete farm field because herbicide is applied consistently within a farm
field, and fields tend to differ in herbicide regimes (Kuester et al. 2015). Within each
population, seeds were collected from 10-34 mature female plants. In populations with
more than 34 females, seeds were sampled by systematically selecting a female for
collection every five paces in a straight line from a haphazard point of entry into the field.
In populations with 34 or fewer females, seeds were sampled from all females. For each
female, entire inflorescences were removed with garden shears and placed in paper bags
to be threshed of seeds. For the ‘within-population correlation experiment’ (1.3.3), seed
from 1-2 randomly selected individuals per population were used to create a synthetic
population of 30 different families. For the ‘among-population correlation experiment’

12
(2.3.4), seed was bulked from all families (10-34 families) within a population to form 22
bulked populations.
2.3.3 Within-Population Correlation Experiment
To measure genetic correlations between escape and tolerance in A. palmeri, I
conducted a field experiment during the summer of 2018 at the Fountain Farm
agricultural field research station in Edgecombe County, outside of Rocky Mount, North
Carolina, USA (35.98 N , -77.77 W). Crops such as corn, cotton, peanut, and soybean
are regularly grown at this facility. The station is within the current range of A. palmeri
and there was a sizable A. palmeri population present in the soil seedbank prior to this
study (Z. Teitel, personal observation 2017). From 1985 – 2015, the station’s average
summer (May – September) temperature was 24.4 °C and its average precipitation was
26.1 mm (Time and Date AS).
To estimate glyphosate escape and tolerance in the field, A. palmeri seeds were
planted on the shoulders of crop rows in three fields, with half the rows cultivated with
corn and the other half left unplanted. Three randomized spatiotemporal complete blocks
were separated by approximately three weeks between planting and in three adjacent
farm fields. Each block contained four treatment combinations in a two-by-two factorial
design: glyphosate sprayed and glyphosate unsprayed; corn crop planted and no crop
planted. Nested within the three blocks, seeds from the 30 families in the experimental
population were planted into a randomized split-plot design. Glyphosate sprayed /
unsprayed and corn crop planted / unplanted were between plot factors, and family was a
within plot factor. Spray treatment was a between plot factor to avoid glyphosate spray
drift contamination between spray treatments (as in Baucom and Mauricio 2008a). As

13
well, crop treatment was a between plot factor to accommodate mechanized crop planting
limitations. Each of the 30 families within the synthetic population was replicated four
times and planted using a random order within each treatment combination plot.
To manipulate weed-crop competition, A. palmeri plants were spaced every 2.44
m within a row and staggered between rows such that weeds were 1.52 m away from their
nearest weed neighbours in the next row over. Crop plants were planted 0.91 m apart
within and between crop rows. To distinguish between weeds planted and those emerging
from the seedbank, experimental seeds were planted in ~10 cm diameter holes that were
filled with potting mix soil. At least five seeds were planted from the same family at each
position to ensure adequate germination. Plants were marked with plastic tags in the soil
for identification. Crop fields were cleared of non-experimental A. palmeri and other
weed species through hoeing.
To run my experiment’s herbicide treatment, both crops and A. palmeri weeds in
the glyphosate spray treatment were sprayed once with RoundUp® Weathermax. Plants
were sprayed at the basal rosette stage when they were less than 15 cm in height, using a
backpack sprayer calibrated to the field-rate of glyphosate herbicide spray at 0.6858 kg
a.e. ha-1. Having a 1.52 m distance between plots ensured there was no glyphosate drift.
2.3.4 Among-Population Correlation Experiment
To measure the across-population correlation between escape and tolerance in A.
palmeri, I conducted a field experiment during the summer of 2017 at the Fountain Farm
agricultural field research station (see site description in 2.3.3, above). Seed from all
families within a population (10-34 families) was bulked. These bulked seeds from each
population were planted on the shoulders of crop rows in three fields, with all rows

14
cultivated with soybean. Seeds were planted in six randomized complete blocks over
three days in three adjacent farm fields. Each block contained seeds from all 22
populations exposed to two treatments: glyphosate sprayed and glyphosate unsprayed. I
grew plants both in the presence and absence of glyphosate to estimate escape and
tolerance.
Nested within the six blocks, seeds from these 22 populations were planted into a
randomized split-plot design. Glyphosate spray treatment was a between plot factor, and
population was a within plot factor. The glyphosate spray treatment was applied at the
whole-plot level to avoid glyphosate spray drift contamination between spray treatments
(as in Baucom and Mauricio 2008b). Each of the 22 populations was replicated eight
times within a block and planted using a random order within a plot. A random half of the
adjacent plants in a block were assigned to the glyphosate-sprayed treatment and the
other half were assigned to the glyphosate-unsprayed treatment.
To set up my field experiment, A. palmeri plants were spaced every 2.74 m within
a row and staggered between rows such that plants were 1.65 m away from their nearest
A. palmeri neighbours in the next row over. Crop plants were spaced, seeds were planted,
and herbicide was applied as described in 1.3.3, above. Having 1.65 m distance between
plots ensured there was no glyphosate drift.
2.3.5 Measuring Escape and Tolerance
In both experiments described above, I estimated escape by measuring days from
planting to first emergence. Emergence was recorded upon daily inspections when a
shoot was visible. Positions without any seedlings that emerged before the glyphosate
spray date were excluded from the analysis. After the first seed in a position emerged, it

15
was recorded and marked; any further emerging seeds from the same position were
removed upon detection. The fewer days from planting to emergence, the greater the
likelihood of escape by growing large enough before glyphosate is applied to
substantially decrease its efficacy. Although the more days from planting to emergence
can also be seen as escape by emerging after glyphosate is applied (Scursoni et al. 2007),
the experimental setup did not allow us to capture escape in this direction.
To quantify A. palmeri tolerance, once glyphosate sprayed and unsprayed female
plants were reproductively mature and new flowers ceased to open (October 6-27 2017
for the experiment described in 1.3.4, August 24 – October 1 2018 for the experiment
described in 1.3.3), two components of fitness were measured for both male and female
plants: inflorescence number and average inflorescence length. First, the total number of
reproductive inflorescences on every plant were counted. Second, all the inflorescences
on a haphazardly selected branch of every plant were measured from the point on the
inflorescence where flowering begins to its tip to estimate inflorescence length. All
inflorescence lengths on a branch were averaged to get mean inflorescence length. These
two fitness components were then multiplied together to get the total inflorescence length
on a plant. Family means were then divided by the overall mean of all plants from both
sprayed and unsprayed treatments (as in Baucom and Mauricio 2004). To measure
tolerance, the fitness components of untreated plants relative to the mean were subtracted
from that of glyphosate-treated plants within the same family. The greater the difference
is above zero, the greater the expression of tolerance is in that family (Baucom and
Mauricio 2004).

16
2.3.6 Statistical Analysis
To determine if there are genetic correlations between glyphosate escape and
tolerance, I calculated Pearson correlation coefficients (r). To do this, I used family-mean
values which approximate genetic correlation (Via 1984) for measures of glyphosate
escape and glyphosate tolerance in both the corn (N=28), and the no crop (N=30)
environments. If r is significant and negative, then escape and tolerance are trading off
with one another. If r is significant and positive, or not significant, then escape and
tolerance are not trading off with one another.
To determine if populations can express high magnitudes of both glyphosate
escape and tolerance, I calculated the Pearson correlation coefficient (r). To do this I used
mean population values (N=22) for measures of glyphosate escape and glyphosate
tolerance. If r is significant and negative, then populations that express high magnitudes
of escape express low magnitudes of tolerance, or vice versa. If r is significant and
positive, then populations that express high or low magnitudes of escape express high or
low magnitudes of tolerance, or vice versa. Finally, if r is not significantly different from
zero, then populations can express any magnitude of escape and tolerance.
2.4 RESULTS
To determine if there is a tradeoff between glyphosate escape and tolerance within
populations, I estimated the genetic correlation between escape and tolerance. There was
no correlation between escape and tolerance for plants grown in the corn environment
(Figure 2.6.1; r=-0.096, p=0.628, N=28). As well, there was no correlation between
escape and tolerance for plants grown in the no crop environment (Figure 2.6.1; r=0.087,

17
p=0.647, N=30). These results indicate that there is not a tradeoff between escape and
tolerance.
To determine if glyphosate escape and tolerance are correlated across populations, I
estimated the population level correlation between escape and tolerance. There was no
correlation between escape and tolerance across populations (Figure 2.6.2; r = 0.376,
p=0.085, N=22). These results indicate that escape is not correlated with tolerance across
populations.
2.5 DISCUSSION
The lack of tradeoffs I found between escape and tolerance can facilitate the
evolution of multiple glyphosate defence strategies in A. palmeri. First, glyphosate escape
and tolerance in A. palmeri were not genetically correlated within a population. Not
finding any genetic correlations, or more specifically negative genetic correlations,
implies that escape and tolerance are not trading off with each other (Agrawal et al.
2010). This has considerable implications for their evolutionary trajectories, as tradeoffs
between alternative defence strategies can lead to maximal expression of one strategy, but
not both, thus providing a limit to adaptation of defence (Futuyma and Moreno 1988,
Fineblum and Rausher 1995, Leimu and Koricheva 2006). Second, I did not find any
correlation between glyphosate escape and tolerance expression among populations. This
would be the expected result if a lack of genetic correlations between escape and
tolerance allows for trajectories where high expressions of both, one, or no defence
strategies can exist in a given population. In fact, I did find a marginally significant

18
positive correlation among populations, suggesting at least the possibility that escape and
tolerance can evolve in the same population. Together, this implies that the lack of
correlations among populations could be the result of lack of genetic correlations within
populations, potentially allowing multiple glyphosate defence strategies to evolve
independently and simultaneously.
This is the first study I am aware of to measure genetic correlations between
escape and tolerance to herbicide stress. However, comparing this study to studies that
measure correlations between other defence strategies or in response to other stresses can
be beneficial for understanding the novelty of these results. Among populations, positive
correlations between drought escape and tolerance have been identified, though there are
few examples outside of drought stress (Welles and Funk 2021). Studies that have tested
for genetic tradeoffs between tolerance and resistance to herbivory have both found and
not found correlations between them (Strauss and Agrawal 1999). Both Fineblum and
Rausher (1995) and Stowe (1998) found that high herbivore resistance was genetically
correlated with low insect tolerance, and that this tradeoff constrains evolution of
resistance. There should be a negative correlation between escape and tolerance because
evolving both strategies could be redundant (Fineblum and Rausher 1995). If defence
strategies incur fitness costs (Herms and Mattson 1992, Simms and Triplett 1994),
expressing multiple strategies at once would raise fitness costs more than is necessary to
achieve optimal fitness. In contrast, both Mauricio et al. (1997) and Weinig et al. (2003)
found no negative genetic correlation between herbivory resistance and tolerance,
allowing selection for both defence strategies together. This study also found no
correlation between the expression of escape and tolerance, supporting the possibility that

19
multiple defence strategies can evolve simultaneously. Absence of a negative genetic
correlation can be due to plant vigour being great enough to support costs of both
strategies at once, or because defence strategies could have other vital functions related to
plant growth and reproduction (Strauss and Agrawal 1999, Rosenthal and Kotanen 1994).
Testing for correlations between herbicide defence strategies can help determine whether
the presence and pervasiveness of one strategy will affect that of other strategies. In this
case, having no tradeoffs between defence strategies suggests that efforts to control the
evolutionary trajectory of escape through selection for tolerance, and vice versa, would
be ineffectual.
My results indicate that if there is selection for both escape and tolerance, then
high expression of glyphosate escape and tolerance can evolve together. High expression
of both escape and tolerance would create a more challenging scenario for weed
suppression efforts aimed at mitigating the effects of herbicide defence strategies.
Further, genetic correlations were not present in the high competition corn environment,
or the low competition no crop environment, suggesting that environmental conditions
are not influencing the correlation between traits, despite evidence that the environment
can cause genetic correlations to shift in magnitude and direction (Sgro and Hoffmann
2004). Although these herbicide defence strategies can also arise in susceptible weed
populations through gene flow via migration of seed or pollen from another population,
selection for defence acting on the standing genetic variation within a population occurs
more frequently (Jasieniuk et al. 1996, Delye et al. 2003), likely due to the prolific
reproduction and extensive seed banks of weed populations in a typical agricultural field
(Lundemo et al. 2009). Thus, strong selection on standing genetic variation for

20
glyphosate escape and tolerance in most environments is likely to result in strong
expressions of both defence strategies together.
My study has two primary limitations in its experimental design. First, I only
measured glyphosate escape via earlier, not later, emergence. Positive correlations
between escape via later emergence and tolerance seem unlikely, as plants that escape via
later emergence would not be exposed to any herbicide application at all, thus creating no
adaptive reason to evolve tolerance. Despite this, negative correlations between escape
and tolerance could be missed by not measuring escape via later emergence. Second, I
measured the two defence strategies escape and tolerance, but did not measure resistance.
Although there is value in estimating two vital yet often overlooked herbicide defence
strategies, the correlation they have with resistance is also likely to inform their
evolutionary trajectories. Resistance could be negatively correlated with escape and
tolerance due to redundancy. As well, resistance could be positively correlated with
escape since plants that escape via early emergence will still benefit from resisting
glyphosate application, even if they have grown beyond the size that herbicides have
most efficacy. Finally, resistance could be positively correlated with tolerance since
plants are only able to tolerate a stress for which there is not complete or zero resistance
(Baucom and Mauricio 2008a). Future studies should aim to include alternative herbicide
defence strategies like escape and tolerance when making predictions about the
evolutionary trajectory of resistance.
I found no tradeoffs between glyphosate escape and tolerance in A. palmeri,
which could potentially lead to populations with multiple defence strategies, and high
magnitudes of both escape and tolerance together. This result is supported by my other

21
finding that expression of herbicide escape and tolerance is not currently correlated
across populations. Whether or not tradeoffs exist between strategies can have a
significant impact on the evolutionary trajectories of defence by determining whether
populations with high expressions of multiple defence strategies can persist or not. This is
of particular importance to herbicide defence in agricultural weeds. As Amaranthus
weeds with evolved resistance to multiple herbicides are more challenging to suppress
than one herbicide (Nakka 2016, Jones et al. 2019), high expression of multiple defence
strategies can be far more challenging to control and suppress than one strategy.

22
2.6 FIGURES
Figure 2.6.1:
Scatterplots of correlation in corn (r=-0.096, p=0.628, N=28) and no crop (r=0.087, p=0.647, N=30)
environments between tolerance (sprayed relative mean total inflorescence length per plant – unsprayed
relative mean total inflorescence length per plant) and escape (days to emergence) for 30 family means of
A. palmeri.

23
Figure 2.6.2:
Scatterplot of correlation (r = 0.376, p=0.085, N=22) between tolerance (sprayed relative mean total
inflorescence length per plant – unsprayed relative mean total inflorescence length per plant) and escape
(days to emergence) for 22 population means of A. palmeri.

24
Chapter 3: What Could Limit the Evolution of Escape and
Tolerance to Glyphosate in an Agricultural Weed (Amaranthus
palmeri)?
3.1 ABSTRACT
Plants can evolve multiple defence strategies in response to environmental stress,
including escape (i.e., avoiding a stress by changing the timing of life history events) and
tolerance (i.e., compensating for damage from the stress through the reallocation of
resources). The evolution of escape and tolerance in plants can be limited if plants with
higher expression of defence strategies have lower fitness in the absence of stress (i.e.,
fitness costs). There can also be limits if the expression of defence strategies is not
consistent across environments (i.e., genotype by environment interactions [GEI]), which
is particularly likely to happen as plants expand their range into new environments. To
test whether there could be limits on the evolution of defense strategies, I measured
fitness costs and GEI for escape and tolerance in response to glyphosate herbicide in
Amaranthus palmeri. To estimate fitness costs of defence strategies, I mesured the
expression of escape and tolerance, as well as the fitness of A. palmeri genotypes in the
absence of glyphosate herbicide. There was no relationship between defence strategy and
fitness in the absence of glyphosate, indicating that neither escape nor tolerance was
costly. I tested for GEI by comparing escape and tolerance of A. palmeri genotypes
grown in the greenhouse vs. the field. I found no relationship between the expression of
escape and tolerance of genotypes grown in the greenhouse and field, indicating a GEI.
These results indicate that fitness costs should not limit the evolution of herbicide escape
and tolerance, but GEI could. Therefore, the evolution of glyphosate escape and tolerance

25
may be limited as invasive weeds such as A. palmeri expand into new environments.
3.2 INTRODUCTION
In response to the biotic and abiotic stresses in natural and agricultural environments,
plants can evolve defence strategies such as escape, resistance, and tolerance (Larcher
2003, Powles and Yu 2010, Agrawal 2011, Lipiec et al. 2013). Since strong selection for
one defence strategy could cause a depletion of defence variation, the presence of genetic
variation of defence suggests that the extent defence strategies can evolve in response to
plant stresses must be limited (Marquis 1992, van der Meijden 1996). Limits on the
evolution of defence can occur at the selection stage due to fitness costs of defence
strategies in the absence of stress (Simms and Triplett 1994, Vila-Aiub et al. 2009). But
even if there is strong selection for a defence strategy, other factors can limit the extent to
which defence strategies are able to evolve in response to selection. Such factors can
include a lack of genetic variation, pleiotropic effects, and genotype by environment
interactions (GEI; Mitchell-Olds 1996, Johnson et al. 2009, Colautti et al. 2010,
Hoffmann et al. 2014). Indeed, these limits likely maintain the high variation that is
commonly observed in plant defence traits (Simms and Rausher 1987, Simms 1992,
Bergelson and Purrington 1996). Therefore, to understand how plant defence traits are
maintained in natural populations, it is necessary to examine which factors can potentially
limit their evolution.
Understanding which factors limit the evolution of weed defence strategies in
response to the widespread application of herbicides is of particular importance to

26
agricultural practices (Jasieniuk et al. 1996, Baucom and Mauricio 2008b, Baucom 2019).
Herbicide defence strategies in weeds can involve various mechanisms that will have
different resource requirements. Although resistance is the most studied herbicide
defence strategy (Baucom 2019), escape and tolerance could also have a significant effect
on herbicide efficacy. Similar to the many well documented examples of escape from
other stresses like drought (Philippi 1993, Pake and Venable 1996, Sherrard and Maherali
2006), herbicide escape is when a plant shifts the timing of its life history to avoid contact
with herbicide (Hilgenfeld et al. 2004). Escape from herbicide application in agricultural
weeds has been attributed to a wide range of seedling emergence times (Scursoni et al.
2007). Escape via earlier emergence allows weeds to grow too large to be an effective
spray target for later applications of herbicide, while later application of herbicide can
often capture late emerging plants (Arnold et al. 1997). Tolerance is when a plant endures
the damage caused by the herbicide by maintaining the plant’s overall fitness (Mauricio
et al. 1997). For example, evidence for herbicide tolerance has come from the ability of
agricultural weeds to maintain fitness through seed production following glyphosate
herbicide damage (Baucom and Mauricio 2004). Despite their potential efficacy,
herbicide escape and tolerance have rarely been measured (Baucom and Mauricio 2004,
Scursoni et al. 2007). However, knowing which factors do or do not limit the evolution of
herbicide escape and tolerance in agricultural weeds can provide insight into why there
may be increasing or decreasing instances of these two defence strategies.
One potential limit to the evolution of herbicide escape and tolerance is fitness costs.
Fitness costs for defence strategies are expected when allocating resources to those traits
reduce other fitness components such as growth and reproduction (Herms and Mattson

27
1992, Simms and Triplett 1994). These costs can limit the evolution of herbicide defence
strategies at the selection stage if the same genotypes that express escape and tolerance to
increase fitness in a herbicide-exposed environment have reduced fitness in the absence
of herbicide exposure. Fitness costs for herbicide escape have not yet been studied.
Although fitness costs of tolerance to herbivory are often studied (Mauricio et al. 1997,
Agrawal et al. 1999), there are very few studies regarding fitness costs for herbicide
tolerance (Baucom and Mauricio 2004). Fitness costs for defence strategies are more
likely to be found in the following two scenarios. First, costs may be more likely to be
detected when plants are grown in stressful environments, regardless of herbicide.
Competitively stressful environments could reveal fitness costs because resources used
for defence would have a greater impact on reproductive output (Van Etten et al. 2015).
Second, costs may be more likely to be detected when diverse genotypes are sampled.
Sampling diverse genotypes could reveal fitness costs because looking across populations
with a wide range of genotypes could show increased genetic variation for defence in at
lease some genotypes (Kuester et al. 2015, Van Etten et al. 2015).
Another potential limit to the evolution of herbicide escape and tolerance is genotype
by environment interactions (GEI, Fox et al. 1997). If GEI is coupled with gene flow or
temporal environmental variation, the evolution of herbicide escape and tolerance in
response to selection can be limited by GEI if a genotype with high defence expression in
one environment has low defence expression in a different environment. This is because
the genotypes with high expressions of escape or tolerance that are selected for in one
environment may not be selected for in a different environment. Therefore, GEI would
make it challenging for a defence strategy to evolve in new populations with different

28
environments, despite selection for that defence strategy. Genotype by environment
interactions are particularly likely to be found for herbicide escape and tolerance defence
strategies because they are likely polygenic traits that can plastically respond to changes
in their environment. Herbicide escape is determined by emergence timing, which is
dependent on many environmental factors including latitude and soil moisture (Forcella
et al. 1997). For example, drought tolerance has had 17 underlying quantitative trait loci
with GEI (El-Soda et al. 2015). Genotype by environment interactions for herbicide
escape and tolerance have not yet been measured, but could potentially limit the
evolution of these defence strategies when weeds invade new environments.
To test for potential limits to the evolution of glyphosate escape and tolerance, I
used the agricultural weed Amaranthus palmeri. This weed is prevalent across the USA
and ranked at or near the top of many crops’ “most troublesome weed” lists, including
cotton, soybean, corn, and peanut (Sauer 1957, Webster et al. 2001, Webster and Nichols
2012). Further, future global temperature increases are projected to expand the range of
A. palmeri into northern regions (Kistner and Hatfield 2018). Given the recent success of
A. palmeri, it is important to understand its potential limits to the evolution of herbicide
defence strategies.
To investigate potential limits to the evolution of herbicide defence strategies, I
estimated fitness costs and genotype by environment interactions for glyphosate escape
and tolerance in A. palmeri. To estimate fitness costs, I grew plants in an agricultural
field from 22 populations in both the presence and absence of glyphosate herbicide
application. I then measured glyphosate escape and tolerance as well as plant fitness in
the absence of glyphosate. To test for GEI, I grew genotypes from the same 22

29
populations in both the greenhouse and an agricultural field and estimated escape and
tolerance. I used this data to answer the following questions:
1. Are there fitness costs associated with glyphosate escape and tolerance in A.
palmeri in the field?
2. Does the expression of glyphosate escape and tolerance differ between
environments (i.e., GEI) in A. palmeri?
3.3 METHODS
3.3.1 Study System
Amaranthus palmeri is a wind-pollinated C4 summer-annual agricultural weed
that originated in the Sonoran Desert (Sauer 1957) and has spread recently via animal
vectors, farming equipment, and seed contamination throughout midwestern and
southeastern USA (Hensleigh and Pokorny 2017). Amaranthus palmeri is dioecious and
an obligate outcrosser (Ward et al. 2013). In hot and sunny environments, A. palmeri can
grow up to 5 cm/day (Horak and Loughin 2000) due to its high photosynthetic rate (81
mol/m2/s, Ehleringer 1983), enabling females to produce as many as 600,000 seeds
(Keeley et al. 1987). Amaranthus palmeri mostly emerge between March and June,
typically flower 5-9 weeks after emergence, and produce viable seed as early as 2-3
weeks after flowering. Although plants can emerge as late as October, lethal November
frosts can come before they are able to set seed (Keeley et al. 1987). No fitness costs for
glyphosate resistance have been found in A. palmeri (Giacomini et al. 2014), though
fitness costs for escape and tolerance have not yet been estimated.

30
3.3.2 Seed Collection
To test for potential limits to the evolution of glyphosate escape and tolerance,
from September to October in 2016, I collected seeds of A. palmeri from 22 populations
spread across their Eastern USA range in Georgia, North Carolina, and Illinois.
Populations were selected by contacting agricultural extension agents and researchers. A
population was defined as a discrete farm field because herbicide is applied consistently
within a farm field, and fields tend to differ in herbicide regimes (Kuester et al. 2015).
Within each population, seeds were collected from 10-34 mature female plants. In
populations with more than 34 females, seeds were sampled by systematically selecting a
female for collection every five paces in a straight line from a haphazard point of entry
into the field. In populations with 34 or fewer females, seeds were sampled from all
females. For each female, entire inflorescences were removed with garden shears and
placed in paper bags to be threshed of seeds.
3.3.3 Fitness Costs Experiment
To estimate fitness costs, I conducted a field experiment during the summer of
2017 at the Fountain Farm agricultural field research station in Edgecombe County,
outside of Rocky Mount, North Carolina, USA (35.98 N , -77.77 W). Crops such as
corn, cotton, peanut, and soybean are regularly grown at this facility. The site is within
the current range of A. palmeri and there was a sizable A. palmeri population present in
the soil seedbank prior to this study (Z. Teitel, personal observation 2017). From 1985 –
2015, this site’s average summer (May – September) temperature was 24.4 °C and its
average precipitation was 26.1 mm (Time and Date AS). I conducted this experiment in
an agricultural field within the weed’s range to adequately recreate the cropping

31
environment that influences fitness costs of escape and tolerance, as well as the
magnitude of their expression. Agricultural fields are stressful growing environments
with interspecific competition from both weed and crops, which could expose the costs of
escape and tolerance.
To measure fitness costs of glyphosate escape and tolerance, seed was planted on
the shoulders of crop rows in three farm fields cultivated with soybean. Seed from all
families within a population (10-34 families) were bulked and were planted in six
randomized complete blocks over approximately three days in three adjacent farm fields.
Each block contained seed from all 22 populations exposed to two treatments: glyphosate
sprayed and glyphosate unsprayed. I grew plants both in the presence of glyphosate to
estimate escape and tolerance, and in the absence of glyphosate to measure fitness costs.
Nested within the six blocks, seed from these 22 populations were planted into a
randomized split-plot design. Glyphosate spray was a between plot factor, and population
was a within plot factor. The glyphosate spray treatment had to be a between plot factor
to avoid glyphosate spray drift contamination between spray treatments (as in Baucom
and Mauricio 2008). Each of the 22 populations was replicated eight times within a block
and planted using a random order within a plot. A random half of the adjacent plants in a
block were assigned to the glyphosate-sprayed treatment and the other half were assigned
to the glyphosate-unsprayed treatment.
To set up my field experiment, crop plants were spaced 0.91 m apart within and
between crop rows. Amaranthus palmeri plants were planted every 2.74 m within a row
and staggered between rows such that plants were 1.65 m away from their nearest A.
palmeri neighbours in the next row over. To distinguish between A. palmeri planted and

32
those emerging from the fields’ seedbanks, experimental seeds were planted in holes
approximately 10 cm in diameter that were excavated and filled with potting mix soil. At
least five seeds were planted from the same population at each position to ensure
adequate germination. Plants were marked with plastic tags in the soil. Both crops and A.
palmeri plants in the glyphosate-sprayed treatment were sprayed once at the basal rosette
stage when they were less than 15 cm in height. Plants were sprayed with RoundUp®
Weathermax using a backpack sprayer calibrated to the field-rate of glyphosate herbicide
spray at 0.6858 kg a.e. ha-1. Crop fields were kept relatively clear of weeds through
hoeing.
To estimate escape, I measured days from planting to first emergence. The fewer
days from planting to emergence, the greater the likelihood of escape by growing large
enough before glyphosate is applied to substantially decrease its efficacy. Although more
days from planting to emergence can also be seen as escape by emerging after glyphosate
is applied, the experimental design did not allow us to capture escape in this direction.
Emergence was recorded upon daily inspections when a shoot was visible. Positions
without any seedlings that emerged before the glyphosate spray date were excluded from
the analysis. After the first seed in a position emerged, it was recorded and marked; any
further emerging seeds from the same position were thinned out upon detection.
To quantify A. palmeri tolerance, once glyphosate-sprayed and -unsprayed plants
were reproductively mature and new flowers ceased to open, two components of fitness
were measured. I recorded fitness components that can be consistently measured for both
male and female plants: inflorescence number and average inflorescence length. First, the
total number of reproductive inflorescences on every plant were counted. Second, all the

33
inflorescences on a haphazardly selected branch of every plant were measured from the
point on the inflorescence where flowering begins to its tip to estimate inflorescence
length. All inflorescence lengths on a branch were averaged to get average inflorescence
length per plant. These two fitness components were then multiplied together to get the
total inflorescence length on a plant. Population means were then divided by the overall
mean of all plants (as in Baucom and Mauricio 2004). To measure tolerance, the fitness
components of untreated plants relative to the mean were subtracted from that of
glyphosate-treated plants within the same population. The greater the difference is above
zero, the greater the expression of tolerance is in that population (Baucom and Mauricio
2004).
3.3.4 GEI Experiment
To estimate GEI, plants were grown from seed in the University of Guelph
Phytotron using the same 22 populations used in the fitness costs experiment (see 3.3.3,
above). Seeds were planted into 1.67 L pots filled with Sunshine® All-Purpose Potting
Mix (Sun Gro Horticulture Canada Ltd., Vancouver, British Columbia, Canada). To
ensure adequate germination, five seeds were planted per pot. Seeds began to emerge 1-2
days after planting. Ten days after planting, seedlings were randomly thinned down to
one per pot. Plants were hand watered twice daily until the majority of germination
occurred, then once daily until the plants reached maturation, then as needed. Each pot
was fertilized every 14 days beginning 14 days after planting with 50 ml of 17-5-17
fertilizer (Master Plant-Prod Inc., Brampton, Ontario, Canada). Plants were sexed upon
flowering. Greenhouses were maintained between 24.5-26.5 C during the daytime and
19.5 - 20.5 C during the nighttime, with 16 hours per day of light that was supplemented

34
with H.P.S. 600-watt lights at 30 mol m-2 s-2.
To estimate escape and tolerance to glyphosate, eight pots of plants from each of
10-34 families were grown from each of the 22 collected populations used in the fitness
costs experiment (see 3.3.3, above). Half of the eight pots within a family were randomly
assigned to a glyphosate-sprayed treatment and half were randomly assigned to a
glyphosate-unsprayed treatment. Plants were divided into two blocks separated by 60
days. A randomly selected half of the families from every population were assigned to the
first block, and the other half of the families were assigned to the second block. This
design does not allow for estimation of variation among families within populations, but
instead allows for estimation of variation in escape and tolerance among populations.
Pots were positioned randomly across the greenhouse benches in three separate rooms per
block.
From each family, four randomly-selected plants were glyphosate treated within a
spray chamber in a portable fume hood (CaptairTM Flex, ErlabTM). Using a hand-held
pressurized sprayer attached to 6 mm-diameter tubing with three nozzles, plants were
sprayed at the basal rosette stage (mean (1 SD) height = 15.53 (7.12) cm in block 1 and
25.22 (9.61) cm in block 2). Plants were sprayed with a pressure gauge calibrated to 30
psi for 30 s to the point before runoff, such that spray droplets remained on leaves. These
glyphosate spraying methods were based on suggested field guidelines but modified for
the enclosed greenhouse environment. Plants were sprayed with RoundUp® Weathermax
(glyphosate, 540 grams acid equivalent per litre, present as potassium salt) at a spray rate
of 0.2241 kg a.e. ha-1. This spray rate was selected because the next highest rate resulted
in ~91% mortality in a preliminary study (Teitel and Caruso, unpublished data). The day

35
after spraying, all plants from the glyphosate treatment were returned to the greenhouse
(as in Feng et al. 2003).
To estimate escape, the number of days from planting to emergence was
measured. Emergence was recorded when a shoot was visible. Because seedlings within
pots are not independent of each other, the average days to emergence was calculated for
each pot (N#_seedlings_emerged= 1-10, mean = 2.62, SE = 0.02). The fewer days from planting
to emergence, the greater the likelihood of escape by growing large enough before
glyphosate is applied to substantially decrease its efficacy. Although the more days from
planting to emergence can also be seen as escape by emerging after glyphosate is applied,
the experimental setup did not allow us to capture escape in this direction. Values of
family level escape were averaged within populations to get values of population level
escape. These population-level values of escape from the greenhouse environment were
compared with population-level values of escape from the field environment (see fitness
costs experiment above) to measure GEI.
To estimate tolerance, inflorescence lengths of glyphosate-treated and -untreated
female plants in the first temporal block were measured. Inflorescence length was used to
estimate fitness because it is a significant predictor of seed production of females (Teitel
and Caruso, unpublished data). Once female plants were reproductively mature and new
flowers ceased to open, the total reproductive inflorescence length was measured. The
total inflorescence length was averaged for all glyphosate-treated and untreated plants
within families (Nplants/family/treatment = 1-4, mean = 1.77, SE = 0.04). Family means were
then divided by the overall mean of all plants in both treatments (as in Baucom and
Mauricio 2004). To measure tolerance, the relative mean reproductive inflorescence

36
length of glyphosate-sprayed plants was subtracted from the mean of glyphosate
unsprayed plants within the same family. The greater the difference is above zero, the
greater the expression of tolerance is in that family (Baucom and Mauricio 2004). Values
of family-level tolerance were averaged within populations to get values of population-
level tolerance. These population-level values of tolerance from the greenhouse
environment were compared with population-level values of tolerance from the field
environment (see 3.3.3, above) to measure GEI.
3.3.5 Statistical Analysis
To estimate fitness costs of escape and tolerance, I used regression. Each of four
regression models included either fitness components of inflorescence length or
inflorescence number in the absence of glyphosate, as the dependent variable, and either
escape or tolerance as the independent variable. Prior to analysis, field values of escape
and tolerance were averaged within populations and relativized to the grand population
means (as in Baucom and Mauricio 2008b, Van Etten et al. 2015). If there are fitness
costs for expressing escape and tolerance, then the regression slope will be negative, as
populations with high escape and tolerance will have low fitness in the absence of
glyphosate. Unlike escape, regressing tolerance on fitness in the absence of glyphosate
creates an artifactual covariance because the same fitness estimates are used in both the
dependent and independent variables. I calculated this artifactual covariance (as described
in Tiffin and Rausher 1999, Baucom and Mauricio 2008b) and subtracted it from the
covariance between the independent variable, tolerance, and the dependent variable,
fitness in the absence of glyphosate, to get an unbiased estimate of covariance. This

37
corrected covariance was then used to calculate the corresponding correlation coefficient
(instead of regression) to establish significance.
To estimate GEI for glyphosate escape and tolerance, I calculated Pearson
correlation coefficients (r). Although I was unable to analyze GEI using a more
traditional ANOVA table due to a lack of power for tolerance, alternative methods have
been used (Fernandez 1991), and correlation can reveal the same interaction. Coefficients
were measured using mean population values (N=22) for measures of both escape and
tolerance between the greenhouse and field environments. If r is significant, then
genotypes with high expression of defence strategies in one environment are the same
genotypes with high expression of defence strategies in the other environment (i.e., no
GEI). If r is not significant, then genotypes with high expression of defence strategies in
one environment are not the same genotypes with high expression of defence strategies in
the other environment (i.e., GEI).
3.4 RESULTS
To test for fitness costs for escape and tolerance in the field, I estimated the
relationship between escape and tolerance and two fitness components in the absence of
glyphosate. Plants from populations with greater escape did not produce shorter (Figure
3.6.1A; =-0.036, t21=-1.026, p=0.317), or fewer (Figure 3.6.1B; =-0.076, t21=-1.161,
p=0.259) inflorescences in the absence of glyphosate. In addition, tolerance was not
correlated with inflorescence length after correcting for artifactual covariance (Figure
3.6.1C; uncorrected =-0.193, t21=-2.121, p=0.047; corrected covariance = -0.0001, r = -

38
0.002N.S.). Finally, tolerance was not correlated with number of inflorescences after
correcting for artifactual covariance (Figure 3.6.1D; uncorrected =-0.4, t21=-2.405,
p=0.026; corrected covariance = -0.016, r = 0.163N.S.). These results indicate no fitness
costs for either escape or tolerance.
To determine if there was GEI for escape and tolerance, I compared escape and
tolerance of the same populations grown in the field and greenhouse. There was no
correlation between escape in the field and in the greenhouse (Figure 3.6.2A; r=0.064,
p=0.777). As well, there was no correlation between tolerance in the field and in the
greenhouse (Figure 3.6.2B; r=0.016, p=0.942). These results indicate that there was GEI
for escape and tolerance.
3.5 DISCUSSION
I looked for evidence of fitness costs and GEI, two mechanisms that could
potentially limit the evolution of glyphosate escape and tolerance in A. palmeri. I found
no evidence of fitness costs for glyphosate escape and tolerance in the absence of
glyphosate stress, suggesting that selection will not be limited by fitness costs. In
contrast, I found evidence of GEI for both escape and tolerance: genotypes which had
high expressions of glyphosate escape and tolerance in the greenhouse did not have the
same high expressions of glyphosate escape and tolerance in the field (Figure 3.6.2). This
could limit the evolution of escape and tolerance because the same genotypes would not
be selected for in different environments. Without fitness costs, there could potentially be
no limits for the evolution of glyphosate escape and tolerance at the selection stage.

39
However, even with strong selection for glyphosate escape and tolerance, their evolution
could still be limited if there is no response to selection (Stanton et al. 2000), potentially
due to GEI with geneflow. These results display the importance in evaluating multiple
mechanisms at different stages of evolution that may influence the extent that herbicide
defence strategies can evolve, since the lack of evidence via one mechanism at the
selection stage does not necessarily mean that there are no potential limits to defence at
other stages of evolution.
Very few studies have tested for fitness costs from herbicide defence strategies
other than resistance, and none have tested for escape. Although there is some evidence
of fitness costs for glyphosate tolerance (Baucom and Mauricio 2004, Baucom and
Mauricio 2008a), my results are consistent with most studies that have tested for, but not
found, fitness costs for herbicide resistance (Vila-Aiub et al. 2009, Baucom 2019).
Evidence for costs of resistance in weeds was found in only 26% of 88 studies (Bergelson
and Purrington 1996), and costs are not consistent across different mechanisms of
herbicide resistance (Vila-Aiub et al. 2009). Testing for fitness costs in multiple strategies
is important for understanding whether single or multiple herbicide defence strategies can
evolve. The general theoretical prediction that stress defence strategies should be
mutually exclusive of each other (Abrahamson and Weis 1997, Mauricio et al. 1997)
assumes that there are fitness costs for expressing defence, and that the benefits of having
redundant multiple defence strategies do not outweigh their costs (Strauss and Agrawal
1999, Baucom and Mauricio 2008a). However, if fitness costs do not limit the evolution
of escape and tolerance, then these two defence strategies could potentially become
alternative evolutionary trajectories to resistance. Further, without fitness costs, both

40
defence strategies could evolve simultaneously, not mutually exclusively of each other or
of resistance as well (Giacomini et al. 2014), creating a formidable suite of herbicide
defence which could be more difficult to overcome. Even without any discernable fitness
costs however, other factors can limit the evolution of herbicide defence strategies.
The evidence I found for GEI in the expression of glyphosate escape and
tolerance is expected due to the polygenic nature of these two stress defences. Unlike
herbicide resistance, which is often the result of a single major effect mutation (Baucom
2019), escape and tolerance are likely to be plastic traits that are environment dependent.
Escape, as determined by variable timing in life history transitions, is a complex trait
controlled by many genes that are influenced by numerous environmental factors
(Koornneef et al. 2002), such as precipitation seasonality (Aide 1992), and habitat type
(Allcock 2004). Like escape, tolerance is a complex trait that can use different
mechanisms related to plant growth and reproduction, which can in turn depend on
environmental factors (Herms and Mattson 1992, Rosenthal and Kotanen 1994, Cushman
and Bohnert 2000). Environmental factors such as increasing stress levels and decreasing
resource availability can have a wide range of outcomes on the expression of herbivory
tolerance (Tiffin 2002, Wise and Abrahamson 2005, Rodriguez et al. 2021). Although
variation for glyphosate tolerance among maternal lines, populations, and regions has
been found in agricultural weeds (Baucom and Mauricio 2008b), this study is the first to
test for GEI with herbicide escape and tolerance. Identification of GEI for plant escape
and tolerance to other stresses is scarce, and many of the studies that examine agricultural
environments focus on crop, not weed, escape and tolerance (Van Oosterom et al. 1996,
Chapman et al. 1997, Thungo et al. 2019). The GEI for glyphosate escape and tolerance I

41
found shows evidence of potential limits for these defence strategies, though which types
of environments create this interaction remains unclear.
Although I examined GEI by comparing the field and greenhouse environment,
GEI using realistic agricultural environments only are also expected. This is because
environments that could limit the evolution of defence strategies in range expanding
agricultural weeds are likely similar to environments that would limit evolution of
defence strategies in invasive plant species, where GEI has been documented (Franks et
al. 2008, Zenni et al. 2014). Much work has been done evaluating limits to evolution of
invasive species beyond their native ranges in natural plant communities (Xu et al. 2010,
Colautti et al. 2010), and the same concepts can apply to noxious agricultural weeds that
invade new farming environments. Particularly, expansion beyond a plant’s native habitat
could expose that plant to a harsher new biotic and abiotic environment that it is not
locally adapted to, where beneficial competitive or defensive traits are no longer
expressed (Lankau 2009). Agricultural environments have similar structures for growing
crops, but differ significantly in climate, soil properties, crops grown, herbicide regimen,
and herbivores. For example, if a southern weed genotype evolved to have high
magnitudes of tolerance, that same genotype could potentially have low magnitudes of
tolerance if it migrated to a northern environment.
The study I performed that estimated potential limits to the evolution of escape
and tolerance has two significant limitations itself. First, I only measured glyphosate
escape via earlier, not later, emergence. If escape was measured via later emergence,
fitness costs for greater escape may have been revealed through late emerging plants not
having enough time to set seed before the end of the season. Although plants can evolve

42
to escape glyphosate spray through earlier emergence by growing large enough to render
the efficacy of glyphosate low by the time it is sprayed, later emergence can also be a
viable strategy to escape glyphosate spray. Second, in estimating GEI, one of the two
environments I used to compare expression of defence strategies was a greenhouse.
Although the crop field chosen to run my experiment represents a realistic growing
environment for A. palmeri, the greenhouse is an artificial growing environment that
natural populations of A. palmeri would not typically occur in. Since the two
environments I compared are likely more different from each other than two different
agricultural environments, the GEI I found is likely overestimated. For future studies, a
more useful comparison may be GEI among agricultural environments at different
latitudes with varying climatic conditions, as A. palmeri continues to migrate north.
Although plants can evolve multiple defence strategies to combat the many biotic
and abiotic stresses they face (Larcher 2003, Agrawal 2011), evolutionary limits
occurring at different stages can prevent them from achieving optimal defence (Johnson
et al. 2009, Baucom and Mauricio 2008a). I found no evidence of fitness costs which
could have potentially limited the evolution of glyphosate escape and tolerance at the
selection stage of evolution. However, I found evidence of GEI indicating that glyphosate
escape and tolerance could potentially be limited in their response to selection from
evolving in new environments. If GEI does in fact limit escape and tolerance, then weeds
that migrate to new environments or that remain in a changing environment may not be
able to consistently evolve high magnitudes of herbicide defence strategies. In contrast,
an environment that remains the same through crop selection or herbicide program, for
example, may remove any potential limits from GEI to evolving consistently high

43
magnitudes of escape and tolerance. Therefore, decisions about changing the agricultural
environment could slow the ongoing evolution of harmful herbicide defence strategies.

44
3.6 FIGURES
Figure 3.6.1:
Scatterplots showing the relationship between escape (days to emergence) and relative inflorescence length
(a, =-0.036, t21=-1.026, p=0.317) and relative inflorescence number (b, =-0.076, t21=-1.161, p=0.259);
and between tolerance (glyphosate-sprayed relative fitness – glyphosate-unsprayed relative fitness) and
relative inflorescence length (c, uncorrected =-0.193, t21=-2.121, p=0.047; corrected covariance = -0.0001,
r = -0.002N.S.) and relative inflorescence number (d, uncorrected =-0.4, t21=-2.405, p=0.026; corrected
covariance = -0.016, r = 0.163N.S.).

45
Figure 3.6.2:
Scatterplots showing the relationship between field and greenhouse population mean values of (a) escape
(days to emergence), N = 22, r=0.064N.S.; and (b) tolerance (glyphosate-sprayed relative fitness –
glyphosate-unsprayed relative fitness), N=22, r=0.164N.S.

46
Chapter 4: Crop competition causes nonlinear selection for
glyphosate resistance in an agricultural weed (Amaranthus
palmeri)
4.1 ABSTRACT
In response to biotic and abiotic stresses, plants can evolve different defence
strategies including escape, which will shift the timing of life history events to avoid a
stress; and resistance, which will reduce the extent of damage caused by a stress. Whether
a weed population primarily evolves escape or resistance may depend on the
environmental context. To test how the environmental context could affect the evolution
of defence strategies, I studied how competition with crop species can alter selection for
glyphosate herbicide escape and tolerance in the agricultural weed Amaranthus palmeri.
Glyphosate escape, resistance, and plant fitness was estimated for A. palmeri grown in
North Carolina farm fields in the presence and absence of competition with corn to
measure the effect of competition on the strength and direction of selection. Selection on
escape did not differ between the crop and no-crop environment, as there was no direct
selection for earlier seedling emergence in either environment. Unlike escape, the form of
natural selection acting on resistance differed between the crop and no-crop environment.
In the no-crop environment, I found strong linear selection for increased resistance, but
no significant nonlinear selection. In the crop environment, I found strong linear selection
for increased resistance, but also found strong negative nonlinear selection for resistance.
As such, in the competitive crop environment only, fitness increased with increasing
resistance up to a point, but then decreased at the highest magnitudes of resistance.
Overall, these results indicate that escape is unlikely to evolve as a herbicide defence

47
strategy regardless of the competitive environment, but the extent to which herbicide
resistance can evolve in agricultural weeds may be limited by competition from crops.
4.2 INTRODUCTION
Plants are faced with multiple stresses that impair their development, survival, and
reproduction (Larcher 2003). In response to stress, plants can evolve defence strategies,
such as escape and resistance (Xiao et al. 2007, Agrawal 2000b). Escape occurs when a
plant changes the timing of its life cycle to avoid a stress (Hilgenfeld et al. 2004). For
example, Arabidopsis helleri can flower earlier to escape floral herbivory from beetles
(Kawagoe and Kudoh 2010). Resistance occurs when a plant reduces the extent of
damage it receives from a stress (Mauricio et al. 1997). For example, Arabidopsis
thaliana produces jasmonic acid and ethylene in response to caterpillar herbivores, which
then reduces herbivore performance (De Vos et al. 2006). These plant defence strategies
can have their evolution shaped by interactions with other competitive and facilitative
community members (Lankau and Strauss 2008, Lau and terHorst 2019). Plant
interactions that enhance the effects of defence strategies on fitness will favour selection
for those defence strategies while interactions that diminish the effects of defence
strategies on fitness will favour selection against those defence strategies (De Meaux and
Mitchell-Olds 2003). For example, induced defence to herbivory reduced biomass of
Lepidium virginicum when it was grown at high densities, but not when it was grown at
low densities (Agrawal 2000a). Understanding how interactions between species within a

48
community influences selection for plant stress defence strategies will enable predictions
about which environmental contexts that stress defences will evolve in.
In addition to natural biotic and abiotic stresses, plants can evolve escape and
resistance in response to human-mediated stresses. One common type of human-mediated
stress is herbicide exposure (Jenks and Hasegawa 2008). In response to herbicide
exposure, plants could evolve to escape by altering the timing of their life history: plants
could either germinate later to avoid being sprayed with herbicide or germinate earlier to
reach a large enough size that the herbicide is ineffectual (Jordan and Jannink 1997,
Hilgenfeld et al. 2004). Alternatively, plants could evolve to resist by reducing the extent
of damage from a herbicide (Mauricio et al. 1997). Of these two defense strategies,
herbicide escape has been studied far less than resistance. Amongst agricultural weeds,
there are approximately 11 new cases of herbicide resistance reported every year (Heap
2014). Relative to the identified incidences of herbicide resistance, far fewer studies have
identified escape as a herbicide defence strategy (e.g. Scursoni et al. 2007). This pattern
could be because there is overall stronger selection for resistance than for escape, or
because escape is studied much less often compared to resistance. Thus, much of the
research addressing defence strategies in response to herbicide application has focused on
how to curb the rapid evolution of resistance. By focusing on the evolution of both escape
and resistance, a more complete assessment of weed defence strategies in response to
herbicide application can be made.
The evolution of herbicide escape and resistance, like other defence strategies,
should depend on the agricultural crop growing environment, which presents biotic
community interactions from other species, including other weeds and crops (Baucom

49
2019). These biotic factors combined with herbicide application will jointly influence
selection for herbicide defence traits and should therefore be considered together when
predicting the evolution of herbicide defence strategies. Biotic crop competition
influences how selection shapes herbicide defence strategies in weeds by creating a
highly competitive agricultural growing environment (Swanton and Weise 1991, Jha et
al. 2017). Agricultural environments are highly competitive because farming practices
provide attractive, resource-rich, but space-limited habitats for plants to grow in. Highly
competitive environments should select against herbicide escape because late-emerging
plants should be at a competitive disadvantage for resources relative to their early-
emerging neighbours (Miller et al. 1994, Dyer et al. 2000). Plants that emerge late should
have a better chance of avoiding herbicide spray, but will have diminished growth rates
and resources due to intensified size-dependent competition at their most vulnerable
stage. Unlike escape, highly competitive environments should not select for or against
herbicide resistance because resistance does not require resources typically limited by
competition. For example, the mechanism for glyphosate resistance in Amaranthus
palmeri uses gene amplification of the EPSPS enzyme, a process unlikely to be limited
by resources being competed for (Gaines et al. 2010). Though competition is known to
influence how plants respond to other stresses (Strauss and Agrawal 1999), little is
known about how competition affects evolution in response to herbicide (Délye et al.
2013, Baucom 2019). Further, if competition affects selection of defence strategies in
response to herbicide, then farmers could use that to their advantage in efforts to control
weed populations through the evolution of resistance and escape (Jha et al. 2017).

50
To test whether competition alters selection on herbicide escape and resistance, I
studied the agricultural weed Amaranthus palmeri and the herbicide glyphosate (e.g.
Monsanto RoundUp®). I chose to use glyphosate because there has been a rapid increase
in glyphosate herbicide use since 1996, and increasing herbicide use in general (Bridges
1994, Benbrook 2016). Many weed species, including A. palmeri, have evolved in
response to glyphosate, causing the efficacy of glyphosate to diminish (Powles 2008,
Benbrook 2016, Heap 2019). The strong and human-mediated selection pressure exerted
from the often-exclusive use of glyphosate herbicide as a weed control measure after the
adoption of glyphosate resistant crops has led to the evolution of many glyphosate-
resistant weed populations (Powles 2008), with resistance to glyphosate found in 46 weed
species to date, including A. palmeri (Heap 2019).
To estimate selection acting on defence strategies among differing competitive
environments, I measured glyphosate escape, resistance, and fitness components of A.
palmeri in both corn and no crop agricultural environments. The corn crop environment
was a high-competition agricultural treatment while the no crop environment was a low-
competition treatment. I used these data to answer the following two questions:
1. Is there selection for herbicide escape and resistance in A. palmeri?
2. Does the competitive field environment affect the magnitude and direction of
selection for herbicide escape and resistance in A. palmeri?
4.3 METHODS

51
4.3.1 Study System
Amaranthus palmeri is a C4 summer-annual agricultural weed that originated in
the Sonoran Desert (Sauer 1957) and has spread recently via animal vectors, farming
equipment, and seed contamination throughout midwestern and southeastern USA
(Hensleigh and Pokorny 2017). In hot and sunny environments, A. palmeri can grow up
to 5 cm/day (Horak and Loughin 2000) due to its high photosynthetic rate (81 mol/m2/s,
Ehleringer 1983), enabling females to produce as many as 600,000 seeds (Keeley et al.
1987). Amaranthus palmeri mostly emerge between March and June, typically flower 5-9
weeks after emergence, and produce viable seed as early as 2-3 weeks after flowering.
Although plants can emerge as late as October, lethal November frosts can come before
they are able to set seed (Keeley et al. 1987). Amaranthus palmeri is dioecious and wind
pollinated, likely making it an effective obligate outcrosser (Grant 1959, Sosnoskie et al.
2007, Ward et al. 2013). In A. palmeri, the principal mode of action of glyphosate
resistance is through gene amplification of the copy number of EPSPS (Powles and
Preston 2006, Gaines et al. 2010, Powles 2010), the chloroplast enzyme necessary for
plant growth and metabolism (Steinrücken and Amrhein 1980, Herrmann and Weaver
1999).
4.3.2 Seed Collection
To measure selection for glyphosate escape and resistance, from September to
October in 2016, I collected seeds of A. palmeri from 22 populations spread across their
Eastern USA range in the three states of Georgia, North Carolina, and Illinois. The
populations from Georgia and North Carolina represent the southern range of A. palmeri,
where glyphosate resistance originally evolved (Heap 2018); the populations from Illinois

52
represent the northern range of A. palmeri, where glyphosate resistance may have
originated from recent southern migrations of seed (P. Tranel, personal communication
2016). Populations were selected by contacting agricultural extension agents and
researchers. A population was defined as a discrete farm field because herbicide is
applied consistently within a farm field, and fields tend to differ in herbicide regimes
(Kuester et al. 2015). Within each population, seeds were collected from 10-34 mature
female plants. In populations with greater than 30 females, plants were sampled
systematically by selecting a plant for collection every five paces in a straight line from a
haphazard point of entry into the field. In populations with or less than 30 females, seeds
were sampled from all individuals. For each individual, entire inflorescences were
removed with garden shears and placed in paper bags to be threshed of seeds. Seeds from
1-2 randomly selected individuals per population were used to create a synthetic
population of 30 different families for use in my experiment.
4.3.3 Experimental Design
The experiment was conducted during the summer of 2018 at the Fountain Farm
agricultural field research station outside of Rocky Mount, North Carolina, USA (35.98
N, -77.77 W). Fountain Farm is regularly used to grow crops such as corn, cotton,
peanut, and soybean. It is within the current range of A. palmeri and there is a sizable A.
palmeri population present in the soil seedbank (Z. Teitel, personal observation 2017).
Fountain Farm’s average (1985 – 2015) summer (May – September) temperature is 24.4
°C and its average precipitation is 26.1 mm (Time and Date AS).
To measure the effect of competition with crops on selection for glyphosate
escape and resistance, A. palmeri seeds were planted on the shoulders of crop rows in

53
three fields, with half the rows cultivated with corn and the other half left unplanted.
Three randomized spatiotemporal complete blocks were separated by approximately three
weeks between planting and in three adjacent farm fields. Each block contained four
treatment combinations in a two-by-two factorial design: glyphosate-sprayed, and
glyphosate-unsprayed; and corn crop planted, and no crop planted. Nested within the
three blocks, seeds from the 30 families in the experimental population were planted into
a randomized split-plot design. Glyphosate spray and corn crop were between plot
factors, and family was a within plot factor. Spray treatment was a between plot factor to
avoid glyphosate spray drift contamination between spray treatments (as in Baucom and
Mauricio 2008a). As well, crop treatment was a between plot factor to accommodate
mechanized crop planting limitations. Each of the 30 families within the synthetic
population was replicated four times and planted using a random order within each
treatment combination plot.
To manipulate weed-crop competition, Amaranthus palmeri were planted every
2.44 m within a row and staggered between rows such that weeds were 1.52 m away from
their nearest weed neighbours in the next row over. Crop plants were planted 0.91 m
apart within and between crop rows. To distinguish between weeds planted and those
emerging from the seedbank, experimental seeds were planted in ~10 cm diameter holes
that were filled with potting mix. At least five seeds were planted from the same family at
each position to ensure adequate germination. Plants were marked with plastic tags in the
soil for identification. Crop fields were cleared of non-experimental A. palmeri and other
weed species through hoeing.

54
To run my experiment’s herbicide treatment, both crops and A. palmeri weeds in
the glyphosate spray treatment were sprayed once with RoundUp® Weathermax. Plants
were sprayed at the basal rosette stage when they were less than 15 cm in height, using a
backpack sprayer calibrated to the field-rate of glyphosate herbicide spray at 0.6858 kg
a.e. ha-1. Having a 1.52 m distance between plots ensured there was no glyphosate drift.
To estimate escape, I measured days from planting to first emergence. Emergence
was recorded upon daily inspections, when a shoot was visible. Positions without any
seedlings that emerged before the glyphosate spray date were excluded from the analysis.
After the first seed in a position emerged, it was recorded and marked; any further
emerging seeds from the same position were removed upon detection. The fewer days
from planting to emergence, the greater the likelihood of escape by growing large enough
before glyphosate is applied to substantially decrease its efficacy. Although the more
days from planting to emergence can also be seen as escape by emerging after glyphosate
is applied, my experimental setup did not allow me to capture escape in this direction.
To estimate resistance, leaf damage was measured on glyphosate-treated plants.
Glyphosate damage is visually evident through the yellowing and eventual shriveling of
sprayed leaves. Three weeks after being sprayed, plants were recorded as either dead or
alive. For plants that lived, both the total number of leaves and the number of sprayed
leaves visually judged to be necrotic were recorded (Baucom and Mauricio 2008a). From
this, the percent of damaged (i.e., necrotic) leaves was calculated, measured as the
proportion of necrotic leaves per plant following spraying. Plants that died or plants with
only necrotic leaves after being sprayed were assigned a resistance value of zero. Since I
defined resistance as one minus the proportion of necrotic leaves, the lower the

55
proportion of necrotic leaves, the higher the expression of resistance. This measure of
phenotypic resistance is correlated with resistance as measured by EPSPS copy number
(Yakimowski et al. 2021).
Since I could not know the gender of the plants that died, I was unable to measure
selection using fitness components that could only be measured on female plants (i.e.,
seed set). Therefore, to quantify A. palmeri fitness, I used fitness components that can be
consistently measured for both male and female plants: inflorescence number and
inflorescence length. These two components of fitness were measured once glyphosate-
sprayed and -unsprayed plants were reproductively mature and new flowers ceased to
open. First, the total number of reproductive inflorescences on every plant were counted.
Second, a subsample of inflorescence lengths on a single branch of every plant was
haphazardly selected, cut with shears, and measured from the point on the inflorescence
where flowering begins to the inflorescence’s tip.
4.3.4 Statistical Analysis
To estimate linear phenotypic selection on escape and resistance in the presence
and absence of crop competition, I used general linear models (Lande and Arnold 1983).
Linear selection gradients (), which estimate direct linear selection on a trait, were
estimated using a model with one of two fitness components (number of inflorescences
per plant or mean inflorescence length) as the dependent variable; both escape and
resistance as the continuous independent variables; and farm field (N = 3) as a categorical
independent blocking variable. Fitness component measures for individual plants were
relativized by dividing by mean fitness (Lande and Arnold 1983). Values of escape and
resistance were standardized to have a mean of zero and standard deviation of one (Sokal

56
and Rohlf 1995). Fitness was relativized and traits were standardized separately within
each trait combination. Because fitness was non-normally distributed, I used bias-
corrected bootstrapping to determine if selection gradients were significantly different
from zero.
To estimate nonlinear phenotypic selection on resistance and escape in the
presence and absence of crop competition, I used general linear models (Lande and
Arnold 1983). Quadratic selection differentials (), which estimate direct nonlinear
selection on a trait, were estimated using a model with one of two fitness components
(number of inflorescences per plant or mean inflorescence length) as the dependent
variable; escape and escape2, or resistance and resistance2, as the continuous independent
variables; and farm field (N = 3) as a categorical independent blocking variable. Fitness
was relativized, defence strategies were standardized, and data was bootstrapped as
described above. Shape of nonlinear selection was visualized using cubic splines.
To determine whether there were statistical differences in selection gradients and
differentials between competition treatments I used planned contrasts. To construct these
contrasts, I calculated the difference in selection between corn crop and no crop
treatments, as well as the pooled standard error (estimated as √𝑆𝐸𝑐𝑜𝑟𝑛 𝑐𝑟𝑜𝑝2 + 𝑆𝐸𝑛𝑜 𝑐𝑟𝑜𝑝
2 ).
Contrasts were constructed for both escape and resistance as well as both fitness
components. Due to non-normality of the data, all pooled standard errors were calculated
from the bootstrapped standard errors. Finally, pooled standard errors were used to test
whether differences in selection between competition treatments significantly differed
from zero.
In addition to selection analyses, to confirm my assumptions that the corn crop

57
treatment was more competitive than the no crop treatment, and that the sprayed
treatment was more stressful than the unsprayed treatment, I used analysis of variance
(ANOVA). The ANOVA model included competition treatment, spray treatment, their
interaction, and block as independent variables; and number inflorescences and mean
inflorescence length (cm) as dependent variables in separate models. Corn crop
competition and glyphosate spray would be confirmed as stressful if the crop and spray
terms in the model are significant.
To test the assumption that competition treatments did not affect the expression of
defence strategies, I used ANOVA. The ANOVA model included competition treatment
and block as independent variables; and escape and resistance as dependent variables in
separate models. If the crop term in the model is significant, then corn crop competition is
affecting the expression of defence strategies.
4.4 RESULTS
To determine if there was selection for herbicide escape via earlier emergence, I
measured linear selection gradients and nonlinear selection differentials on emergence
time. Selection for escape was considered only as selection for earlier emergence, not
later emergence. There was linear direct selection for later emergence via number of
inflorescences per plant in the no crop environment, but no linear direct selection for
earlier or later emergence in the corn crop environment (Table 4.6.1). There was no
nonlinear selection for earlier or later emergence in any environment (Table 4.6.2). I
therefore found no evidence of selection for escape.

58
To determine if there was selection for herbicide resistance, I measured linear
selection gradients and nonlinear selection differentials for resistance in both the no crop
and corn crop environments. There was linear selection for increased resistance via
number of inflorescences per plant as well as mean length of inflorescences in both the no
crop environment and the corn crop environment (Table 4.6.1). In the no crop
environment, there was nonlinear selection for resistance via inflorescence length only, in
which fitness increased with resistance up to a point, then decreased (Table 4.6.2, Figure
4.7.1). In the corn crop environment, there was nonlinear selection for resistance via
number of inflorescences per plant as well as mean length (cm) of inflorescences, in
which fitness increased with resistance up to a point, then decreased (Table 4.6.2, Figure
4.7.1). I therefore found both linear and nonlinear selection for glyphosate resistance.
To determine if the competitive environment influences selection on herbicide
escape, I compared linear selection gradients and nonlinear selection differentials on
emergence between the no crop and corn crop environments. Linear selection for later
emergence was stronger in the no crop environment than in the corn crop environment for
number of inflorescences only (Table 4.6.3). There was no significant difference in
nonlinear selection differentials for emergence between competitive environments (Table
4.6.3). I therefore found that linear selection for later emergence only was dependent on
the competitive environment.
To determine if the competitive environment influences selection on herbicide
resistance, I compared linear selection gradients and nonlinear selection differentials for
resistance between the no crop and corn crop environments. There was no difference in
linear selection gradients for resistance via either fitness component; there was selection

59
for increased resistance in both the no crop and corn crop environments (Table 4.6.3). In
contrast, the nonlinear selection differential for resistance via mean length of
inflorescences was significantly different between competitive environments (Table
4.6.3). Both crop environments showed significant negative quadratic selection
differentials, in which fitness increased with increasing resistance up to a point, followed
by a decrease in fitness with further increasing resistance (Figure 4.7.1). However, the
nonlinear selection differential for mean length of inflorescences in the more competitive
corn crop environment (=-0.763) was significantly greater than in the no crop
environment (=-0.200). Although the nonlinear selection differential for resistance via
mean number of inflorescences also showed significant nonlinear selection in the corn
crop environment only (=-0.527), this was not significantly different from the no crop
environment (Table 4.6.3, =-0.177). These results indicate that there is stronger negative
nonlinear selection in the more competitive corn crop environment.
To confirm that the corn crop environment was more competitive than the no crop
environment, and that the glyphosate spray treatment was more stressful than the
glyphosate unsprayed treatment, I compared fitness components between crop and spray
treatments. I found that both crop and spray treatments had a significant effect on each
fitness component. Plants grown in the no crop treatment had significantly greater fitness
than plants grown in the corn crop treatment, with number of inflorescences 16.745 times
greater and mean length of inflorescences 1.608 times greater (Table 4.6.4, Figures 4.7.2
and 4.7.3). Plants grown in the no spray treatment had significantly greater fitness than
plants grown in the glyphosate spray treatment, with 1.970 times more inflorescences and
1.788 times longer inflorescences (Table 4.6.4, Figures 4.7.2 and 4.7.3). Crop treatment

60
and spray treatment interacted for number of inflorescences only, such that the
combination of corn crop and glyphosate spray yielded fitness lower than the additive
effects of both treatments (Table 4.6.4, Figure 4.7.2). These results confirm that the corn
crop environment was more competitive than the no crop environment, and the
glyphosate-sprayed environment was more stressful than the glyphosate-unsprayed
environment.
To test the assumption that competition treatments did not affect the expression of
defence strategies, I compared escape and resistance between crop treatments. I found
that emergence was significantly later in the no crop environment than in the corn crop
environment, but resistance was not affected by crop treatment (Table 4.6.5). Therefore,
the assumption that crop treatment has no effect on defence strategies is correct for
resistance but not for escape.
4.5 DISCUSSION
I found that environmental context affected selection for glyphosate resistance,
but not for escape. There was strong selection for greater glyphosate resistance in A.
palmeri in the field, and the magnitude and shape of that selection was dependent on the
competitive crop growing environment. I found direct linear selection for greater
resistance in both the crop and no crop environment, though in the crop environment
only, increased resistance did not always result in increased plant fitness. Specifically,
when A. palmeri was grown amongst competitive corn crops, there was a significant
negative nonlinear component of selection. Increasing resistance resulted in increasing

61
fitness up to a point, and then resulted in decreasing fitness. In contrast, no evidence was
found for linear or nonlinear selection on escape in either competitive environment.
These results suggest that escape, but not resistance, is unlikely to evolve as a defence
strategy, though highly competitive environments may limit the magnitude of resistance
that can evolve.
Like other studies that estimated selection for resistance (Simms 1990, Núñez‐
Farfán and Dirzo1994), I found evidence of negative nonlinear selection that can limit the
extent of resistance, though this is the first study to find nonlinear selection for herbicide
resistance. Only one other study I am aware of looked for nonlinear selection for
resistance to herbicide in an agricultural weed but did not find any, though their
experiment did not have a competition treatment (Baucom and Mauricio 2008). In
contrast to herbicide studies, herbivory studies have shown nonlinear selection for
resistance from herbivores (Mauricio et al. 1997, Pilson 2000). Studies using herbivory
that have found evidence of nonlinear selection for defence traits argue that intermediate
trait values may be maintained in populations through a mechanism involving a fitness
cost or benefit (Núñez-Farfán et al. 2007, Fornoni et al. 2004). This implies that there
may be a fitness cost for evolving very high magnitudes of glyphosate resistance in A.
palmeri. In this case there should be selection against increased glyphosate resistance as
the benefits of having greater expression of resistance do not outweigh the costs. Fitness
costs for glyphosate resistance have not yet been found in A. palmeri (Giacomini et al.
2014, Vila-Aiub 2014), and only 26% of 88 studies had evidence for fitness costs in
plants (Bergelson and Purrington 1996). Indeed, there may be no fitness costs for
resistance in A. palmeri, since the mechanism of glyphosate resistance simply upregulates

62
the expression of the glyphosate inhibited EPSPS gene which is relatively resource
inexpensive (Gaines et al. 2010). However, increased expression of phenotypic
glyphosate resistance in A. palmeri only increases with EPSPS copy number until a
certain threshold, and then levels out, suggesting that there may be little benefit in having
higher EPSPS copy numbers (Yakimowski et al. 2021). Further, costs of weed herbicide
resistance have been detected when looking for fitness costs in other fitness components
and when testing for costs in competitively stressful environments (Van Etten et al.
2015). Fitness costs and nonlinear selection for herbicide resistance may be detected
more often when measured in the presence of crop competition. Alternatively, instead of
crop competition inducing fitness costs, it may be suppressing the benefits of increased
resistance.
This study provides evidence that there is some potential limit to the evolution of
high magnitudes of glyphosate resistance in A. palmeri, through strong crop competition.
Fitness levels increased with increasing resistance until a point, and then decreased with
further increases in resistance, in the competitive corn crop environment. The fitness
benefits of glyphosate resistance in A. palmeri are likely being lowered by a competitive-
induced decrease in limiting resources such as nutrients, light, and soil moisture, as
expected in weed–crop competitive interactions (Zimdahl 1980, Murphy et al. 1996,
Rajcan and Swanton 2001). Human mediated agricultural operations can manipulate how
competitive the agricultural environment is by selecting which crops to plant (Stoltenberg
and Wiederholt 1995), what density to plant them in (Teasdale 1990), and when to plant
them (Gunsolus 1990). For example, growing corn compared with growing soybean
presents greater competition for interspersed weeds due to the ability of corn to out-grow

63
and out-shade neighbouring species at critical periods (Moolani et al. 1964, Rajcan and
Swanton 2001), and could therefore weaken selection for weed resistance. Crop rotation
and crop canopy structure significantly affected growth and morphological life history
traits among populations of A. palmeri, indicating that crop competition can influence
evolution for aggressive weed traits (Bravo et al. 2017). Additionally, if selection for
herbicide resistance in weeds is weakened by crop competition, this would reduce the
need to spray fields with increasing quantities of herbicide, thus further weakening
selection for high magnitudes of herbicide resistance (Stoltenberg and Wiederholt 1995,
Boerboom 1999). If the genetic variation for resistance is similar between corn and crop
environments, then selection against high resistance in competitive environments can
limit the evolution of high magnitudes of resistance. Thus, the ability to manipulate the
crop growing environment may be just as important a tool as manipulating herbicide
spray for limiting the evolution of herbicide defence in weeds.
Despite escape being a potential defence strategy from herbicide damage, I did
not find any evidence that selection is acting on glyphosate escape via early emergence in
either competitive environment. Timing of emergence throughout the growing season has
been shown to have a key effect on fitness in response to other stresses like herbivory
(Hill and Silvertown 1997, Seiwa 1998) and drought (Dickman et al. 2019), and there can
be selection on emergence time from a multitude of other factors in the absence of
herbicide spray (e.g. Miller et al. 1994). Although this is the first study to measure
selection for escape from glyphosate spray, I had predicted for there to be strong selection
for glyphosate escape. This is because escape has had a significant impact on weed
control efforts, with glyphosate and other herbicide efficacies shown to be dependent on

64
the proximity of glyphosate spray date with emergence date (Jordan et al. 1997), and
prolonged emergence allowing weeds to avoid planned glyphosate applications (Owen
1997). One reason I did not find selection for escape could be that glyphosate escape in
A. palmeri may provide indirect benefits to crops they compete with. Crop yields have
been significantly impacted by the relative timing of weed emergence (Kropff et al.
1992), as delayed weed emergence gave a competitive advantage to rice crops (Gibson et
al. 2002), and earlier emerging weeds reduced soybean yields more than later emerging
weeds (Hock et al. 2006). My results suggest that defence strategies other than escape,
such as resistance, may be the dominant target for selection.
I have identified two main limitations to this study. First, the experiment was set
up to measure selection for glyphosate escape via earlier, not later, emergence. Although
plants can evolve to escape glyphosate spray through earlier emergence by growing large
enough to render the efficacy of glyphosate low by the time it is sprayed, later emergence
can also be a viable strategy of escape by missing the application of post-emergent
glyphosate spray. Despite this, selection for earlier emergence, especially in the
competitive corn crop environment, may be more likely because earlier emerging weeds
have a competitive advantage compared to later emerging weeds due to fewer
competitors at early growth stages, and weaker competitors once they do emerge (Kalisz
1986, Miller et al. 1994, Dyer et al. 2000). Second, although this experiment had two
cropping treatments at both competitive extremes, whether selection on escape and
resistance operates similarly in intermediate competitive environments remains unknown.
Further research should determine how less competitive crops like peanut, soybean, and
cotton influence selection on defence strategies (Jha et al. 2017). Compared with corn, a

65
less competitive crop may have resulted in weaker selection against the highest
magnitudes of resistance, indicating a possible scale of crop competition driving selection
against glyphosate resistance.
In summary, I found that the crop-growing environment can affect the evolution
of herbicide defence strategies, suggesting that there may be competitive limits to
evolving high magnitudes of glyphosate resistance. My results show that understanding
how the community context can shape evolution in response to human-mediated stresses
can provide new approaches for agricultural management. Noxious weeds compete with
crops, decrease crop yields and harvest quality, and increase the resources needed for
cultivation (Bridges 1992, Oerke 2006, Welch et al. 2016, Soltani et al. 2017). By
determining what plant community interactions can select against weed defence
strategies, agricultural managers can suppress their rapid evolution. With the evolution of
glyphosate resistant weeds becoming increasingly unmanageable (Powles 2008,
Benbrook 2016), the ability to be able to manipulate human-mediated selection pressures
such as crop selection, in addition to herbicides, may become vital to controlling resistant
weed populations.

66
4.6 TABLES Table 4.6.1:
Linear selection gradients () +/- standard error (SE) for escape and resistance under glyphosate-sprayed
treatment via the fitness components relative number inflorescences and relative mean inflorescence length
(cm). + p<0.1, * p<0.05, ** p<0.01, *** p<0.001. N=392.
Escape Resistance
No Crop Corn Crop No Crop Corn Crop
SE SE SE SE
Number of Inflorescences 0.200* 0.087 -0.072 0.095 0.471** 0.095 0.471** 0.155
Inflorescence Length 0.019 0.063 -0.062 0.074 0.341** 0.066 0.424** 0.088
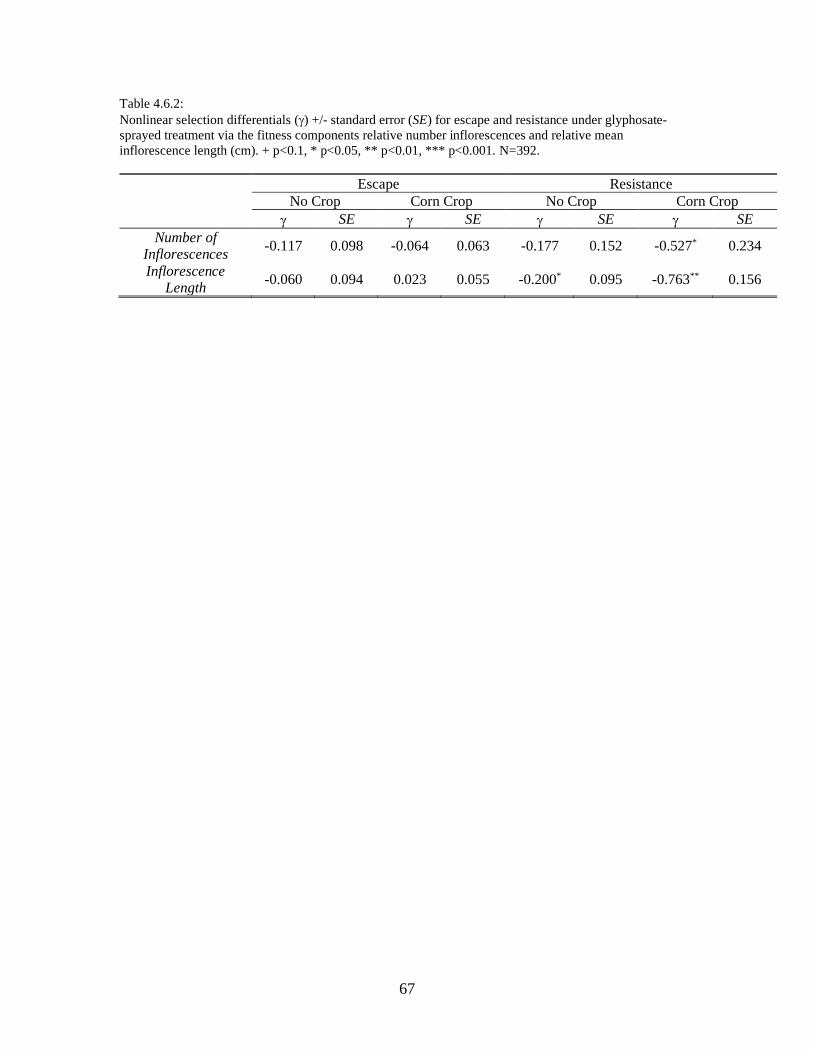
67
Table 4.6.2:
Nonlinear selection differentials () +/- standard error (SE) for escape and resistance under glyphosate-
sprayed treatment via the fitness components relative number inflorescences and relative mean
inflorescence length (cm). + p<0.1, * p<0.05, ** p<0.01, *** p<0.001. N=392.
Escape Resistance
No Crop Corn Crop No Crop Corn Crop
SE SE SE SE
Number of Inflorescences
-0.117 0.098 -0.064 0.063 -0.177 0.152 -0.527* 0.234
Inflorescence Length
-0.060 0.094 0.023 0.055 -0.200* 0.095 -0.763** 0.156

68
Table 4.6.3:
t-statistics (t) +/- standard error (SE) for the difference between linear selection gradients and nonlinear
selection differentials for escape and resistance under glyphosate-sprayed treatment in no crop vs. corn crop
treatments via the fitness components relative number inflorescences and relative mean inflorescence
length (cm).
Escape Resistance
Linear Nonlinear Linear Nonlinear
t SE t SE t SE t SE
Number of
Inflorescences -2.111* 0.129 0.455 0.116 0.000 0.182 -1.254 0.279
Inflorescence Length -0.833 0.097 0.762 0.109 0.754 0.110 -3.082* 0.183

69
Table 4.6.4:
F-statistics with degrees of freedom for ANOVA testing the effects of block, spray treatment, and crop
treatment on fitness components relative number inflorescences and relative mean inflorescence length
(cm). + p<0.1, * p<0.05, ** p<0.01, *** p<0.001.
Block Spray Crop Spray x Crop
Number of
Inflorescences 69.2912
*** 31.8691*** 185.2271
*** 19.8341***
Inflorescence Length 126.7152*** 69.5901
*** 51.1351*** 1.0621

70
Table 4.6.5:
F-statistics with degrees of freedom for ANOVA testing the effects of block and crop treatment on defence
strategies escape and resistance. + p<0.1, * p<0.05, ** p<0.01, *** p<0.001.
Block Crop
Escape 239.1842*** 35.2981
***
Resistance 2.5202+ 3.7841
+

71
4.7 FIGURES
Figure 4.7.1:
Cubic splines of both linear and nonlinear selection for glyphosate resistance in no crop (top row) and corn
crop (bottom row) environments via relative number inflorescences and relative mean inflorescence length
(cm). All resistance values are standardized to a mean of zero and standard deviation of one. All dependent
response variables are relativized to the population mean within their crop environment treatment.
Confidence intervals at 0.95 are displayed around the mean.

72
Figure 4.7.2:
Estimated mean total inflorescences (+/- 2 SE) for glyphosate-sprayed and unsprayed plants in no crop and
corn crop environments.

73
Figure 4.7.3:
Estimated mean inflorescence length (+/- 2 SE) for glyphosate-sprayed and unsprayed plants in no crop and
corn crop environments.

74
Chapter 5: Conclusion
5.1 OVERVIEW
I first determined whether multiple defence strategies can evolve simultaneously
together by testing for tradeoffs from negative genetic correlations between glyphosate
escape and tolerance for A. palmeri grown in an agricultural field (Chapter 2). Within
populations, I found no negative genetic correlations between glyphosate escape and
tolerance, indicating no evidence that escape and tolerance are trading off with each other
(Chapter 2). Among populations, I also found no correlations between glyphosate escape
and tolerance, indicating that populations can have high magnitudes of both defence
strategies (Chapter 2). Thus, selection for escape, for example, is unlikely to affect
selection for tolerance, and both herbicide defence strategies can evolve high magnitudes
simultaneously in the same weed population. With strong and independent selection for
each defence strategy, high magnitudes of multiple herbicide defence strategies can be
more challenging for agricultural weed control measures to suppress.
I next determined whether two types of evolutionary limits could potentially
suppress the extent that glyphosate escape and tolerance could evolve in A. palmeri:
fitness costs and GEI (Chapter 3). I looked for fitness costs in A. pameri by measuring
glyphosate escape and tolerance as well as fitness in the absence of herbicide stress, and
found that higher expression of defence strategies does not result in lower plant fitness
(Chapter 3). This indicates no discernable fitness cost for evolving high magnitudes of
escape and tolerance, though costs could have been revealed under stronger

75
environmental pressure or testing from a wider genotypic range. I also looked for GEI as
a potential evolutionary limit and found no relationship between the expression of escape
and tolerance of genotypes grown in the greenhouse and field, indicating a GEI (Chapter
3). Therefore, the evolution of glyphosate escape and tolerance is unlikely to be limited
by fitness costs but may be limited by invasive weeds such as A. palmeri expanding into
new environments.
Finally, I tested whether the environmental context of an agricultural field can
influence selection acting on glyphosate escape and resistance in A. palmeri (Chapter 4).
Using a synthetic population, I measured the shape and direction of selection acting on
glyphosate escape and resistance in both a competitive corn crop field environment as
well as a non-competitive no crop field environment (Chapter 4). Though there was no
selection for escape in either competitive environment, selection for resistance differed
between competitive environments (Chapter 4). Though both environments revealed
linear selection for increased resistance, only the competitive corn crop environment
revealed negative nonlinear selection for resistance, in which fitness increases with
increasing resistance up to a point and then declines with further increases in resistance
(Chapter 4). Therefore, while altering the competitive crop-growing environment is
unlikely to affect selection for glyphosate escape, competitive crops can potentially select
against high magnitudes of glyphosate resistance in A. palmeri.
5.2 FUTURE DIRECTIONS AND SIGNIFICANCE
Future research should take into account how other environmental factors in

76
agricultural fields can select for defence strategies including selection from cropping
systems, herbivores, and other herbicides. Though I studied the evolution of defence
strategies in response to glyphosate herbicide alone, a typical weed population will likely
be subject to a suite of other herbicides with varying modes of action. Population
resistance to other herbicides could potentially be correlated with glyphosate resistance
(Walsh et al. 2004, Varanasi et al. 2015), and be influencing the selection I found for and
against glyphosate resistance. Preliminary research shows a positive correlation between
population-level glyphosate and thifensulfuron resistance (Teitel et al., unpublished data).
In contrast, no population-level correlations between glyphosate resistance and atrazine,
fomesafen, glufosinate, or mesotrione resistance was found (Teitel et al., unpublished
data). If correlated, selection for resistance to other herbicide modes of action is likely to
create unexpected variation for fitness within the populations that I measured glyphosate
resistance.
My research asked which herbicide defence strategies are most likely to evolve in
weeds where multiple strategies exist, and what could potentially limit this evolution.
Though many have argued that multiple defence strategies are unlikely to evolve because
they are costly and redundant when expressed (Herms and Mattson 1992, Rosenthal and
Kotanen 1994), my research found that escape and tolerance do not tradeoff with each
other and could therefore evolve together. This may be possible because escape and
tolerance are not costly or because they provide separate functions for plant defence
(Agrawal 2011, Romeo et al. 2013). Populations that evolve to both escape and tolerate
glyphosate stress could be more challenging to control than populations with just one of
those strategies. Further, I found no discernable fitness costs for glyphosate escape and

77
tolerance despite the traditional expectation that they are present (Simms and Triplett
1994), which could potentially create evolutionary trajectories for populations with high
magnitudes of both of these defence strategies. On the other hand, I found a GEI for
escape and tolerance, indicating that gene flow and environments that change over time
could limit the evolution of these two defence strategies. Further, since I found that
highly competitive environments select against high magnitudes of glyphosate resistance,
then altering the growing environment to lower competition could decrease the spread of
resistance. Previous research and modeling have shown a close relationship between
herbicide dose, weed density, and crop yield (e.g. Kim et al. 2002), though haven’t linked
those factors to the evolution of alternative stress responses. Through this applied
evolution study, agricultural land managers will be better informed on how to manipulate
their crop-growing environments to select against the economically harmful effects of
weed resistance, thereby increasing crop yields.

78
Chapter 6: Literature cited
Abrahamson, W. G., & Weis, A. E. (2020). Evolutionary Ecology across Three Trophic
Levels: Goldenrods, Gallmakers, and Natural Enemies (MPB-29) (Vol. 96). Princeton
University Press.
Agrawal, A. A. (2000a). Benefits and costs of induced plant defense for Lepidium
virginicum (Brassicaceae). Ecology, 81(7), 1804-1813.
Agrawal, A. A. (2000b). Overcompensation of plants in response to herbivory and the by-
product benefits of mutualism. Trends in plant science, 5(7), 309-313.
Agrawal, A. A. (2011). Current trends in the evolutionary ecology of plant
defence. Functional Ecology, 25(2), 420-432.
Agrawal, A. A., Conner, J. K., & Rasmann, S. (2010). Tradeoffs and negative
correlations in evolutionary ecology. Evolution since Darwin: the first, 150, 243-268.
Agrawal, A. A., Strauss, S. Y., & Stout, M. J. (1999). Costs of induced responses and
tolerance to herbivory in male and female fitness components of wild
radish. Evolution, 53(4), 1093-1104.
Aide, T. M. (1992). Dry season leaf production: an escape from herbivory. Biotropica,
532-537.
Allcock, K. G., & Hik, D. S. (2004). Survival, growth, and escape from herbivory are
determined by habitat and herbivore species for three Australian woodland
plants. Oecologia, 138(2), 231-241.
Arnold, J. C. D., Shaw, R., & Scharer, S. M. (1997). Influence of application timing on
efficacy of glyphosate in Roundup Ready soybean. In Proc. South. Weed Sci. Soc (Vol.
50, pp. 176-177).
Baucom, R. S. (2019). Evolutionary and ecological insights from herbicide‐resistant
weeds: what have we learned about plant adaptation, and what is left to uncover?. New
Phytologist, 223(1), 68-82.
Baucom, R. S., & Mauricio, R. (2004). Fitness costs and benefits of novel herbicide
tolerance in a noxious weed. Proceedings of the National Academy of Sciences of the
United States of America, 101(36), 13386-13390.
Baucom, R. S., & Mauricio, R. (2008a). Constraints on the evolution of tolerance to
herbicide in the common morning glory: resistance and tolerance are mutually exclusive.
Evolution, 62(11), 2842-2854.
Baucom, R. S., & Mauricio, R. (2008b). The evolution of novel herbicide tolerance in a

79
noxious weed: the geographic mosaic of selection. Evolutionary Ecology, 22(1), 85-101.
Baylis, A. D. (2000). Why glyphosate is a global herbicide: strengths, weaknesses and
prospects. Pest Management Science: formerly Pesticide Science, 56(4), 299-308.
Benbrook, C. M. (2016). Trends in glyphosate herbicide use in the United States and
globally. Environmental Sciences Europe, 28(1), 3.
Bergelson, J., & Purrington, C. B. (1996). Surveying patterns in the cost of resistance in
plants. American Naturalist, 148(3), 536-558.
Bingham, R. A., & Agrawal, A. A. (2010). Specificity and trade‐offs in the induced plant
defence of common milkweed Asclepias syriaca to two lepidopteran herbivores. Journal
of Ecology, 98(5), 1014-1022.
Boerboom, C. M. (1999). Nonchemical options for delaying weed resistance to herbicides
in Midwest cropping systems. Weed Technology, 13(3), 636-642.
Bravo, W., Leon, R. G., Ferrell, J. A., Mulvaney, M. J., & Wood, C. W. (2017).
Differentiation of life-history traits among Palmer amaranth populations (Amaranthus
palmeri) and its relation to cropping systems and glyphosate sensitivity. Weed
Science, 65(3), 339-349.
Bridges, D. C. (1992). Crop losses due to weeds in the United states, 1992. Weed Science
Society of America.
Bridges, D. C. (1994). Impact of weeds on human endeavors. Weed Technology, 8(2),
392-395.
Chapman, S. C., Crossa, J., & Edmeades, G. O. (1997). Genotype by environment effects
and selection for drought tolerance in tropical maize. I. Two mode pattern analysis of
yield. Euphytica, 95(1), 01-09.
Colautti, R. I., Eckert, C. G., & Barrett, S. C. (2010). Evolutionary constraints on
adaptive evolution during range expansion in an invasive plant. Proceedings of the Royal
Society B: Biological Sciences, 277(1689), 1799-1806.
Culpepper, A. S., Grey, T. L., Vencill, W. K., Kichler, J. M., Webster, T. M., Brown, S.
M., ... & Hanna, W. W. (2006). Glyphosate-resistant Palmer amaranth (Amaranthus
palmeri) confirmed in Georgia. Weed Science, 54(4), 620-626.
Cushman, J. C., & Bohnert, H. J. (2000). Genomic approaches to plant stress
tolerance. Current opinion in plant biology, 3(2), 117-124.
Délye, C., Jasieniuk, M., & Le Corre, V. (2013). Deciphering the evolution of herbicide
resistance in weeds. Trends in Genetics, 29(11), 649-658.

80
Délye, C., Michel, S., Bérard, A., Chauvel, B., Brunel, D., Guillemin, J. P., ... & Le
Corre, V. (2010). Geographical variation in resistance to acetyl‐coenzyme A carboxylase‐
inhibiting herbicides across the range of the arable weed Alopecurus myosuroides (black‐
grass). New Phytologist, 186(4), 1005-1017.
De Meaux, J., & Mitchell-Olds, T. (2003). Evolution of plant resistance at the molecular
level: ecological context of species interactions. Heredity, 91(4), 345-352.
De Vos, M., Van Zaanen, W., Koornneef, A., Korzelius, J. P., Dicke, M., Van Loon, L.
C., & Pieterse, C. M. (2006). Herbivore-induced resistance against microbial pathogens
in Arabidopsis. Plant Physiology, 142(1), 352-363.
Dickman, E. E., Pennington, L. K., Franks, S. J., & Sexton, J. P. (2019). Evidence for
adaptive responses to historic drought across a native plant species range. Evolutionary
Applications, 12(8), 1569-1582.
Duke, S. O., & Powles, S. B. (2009). Glyphosate-resistant crops and weeds: now and in
the future.
Dyer, A. R., Fenech, A., & Rice, K. J. (2000). Accelerated seedling emergence in
interspecific competitive neighbourhoods. Ecology letters, 3(6), 523-529.
Ehleringer, J. (1983). Ecophysiology of Amaranthus palmeri, a Sonoran Desert summer
annual. Oecologia, 57(1-2), 107-112.
El‐Soda, M., Kruijer, W., Malosetti, M., Koornneef, M., & Aarts, M. G. (2015).
Quantitative trait loci and candidate genes underlying genotype by environment
interaction in the response of Arabidopsis thaliana to drought. Plant, Cell &
Environment, 38(3), 585-599.
Feng, P. C., Chiu, T., Sammons, R. D., & Ryerse, J. S. (2003). Droplet size affects
glyphosate retention, absorption, and translocation in corn. Weed Science, 51(3), 443-
448.
Fernandez, G. C. (1991). Analysis of genotype×environment interaction by stability
estimates. HortScience, 26(8), 947-950.
Fineblum, W. L., & Rausher, M. D. (1995). Tradeoff between resistance and tolerance to
herbivore damage in a morning glory. Nature, 377(6549), 517.
Forcella, F., Wilson, R. G., Dekker, J., Kremer, R. J., Cardina, J., Anderson, R. L., ... &
Buhler, D. D. (1997). Weed seed bank emergence across the Corn Belt. Weed Science,
67-76.

81
Fornoni, J., Nunez‐Farfán, J., Valverde, P. L., & Rausher, M. D. (2004). Evolution of
mixed strategies of plant defense allocation against natural enemies. Evolution, 58(8),
1685-1695.
Fox, P. N., Crossa, J., & Romagosa, I. (1997). Multi-environment testing and genotype×
environment interaction. In Statistical methods for plant variety evaluation (pp. 117-138).
Springer, Dordrecht.
Franklin, O. D., & Morrissey, M. B. (2017). Inference of selection gradients using
performance measures as fitness proxies. Methods in Ecology and Evolution, 8(6), 663-
677.
Franks, S. J., Pratt, P. D., Dray, F. A., & Simms, E. L. (2008). Selection on herbivory
resistance and growth rate in an invasive plant. The American Naturalist, 171(5), 678-
691.
Futuyma, D. J., & Moreno, G. (1988). The evolution of ecological specialization. Annual
review of Ecology and Systematics, 19(1), 207-233.
Gaines, T. A., Zhang, W., Wang, D., Bukun, B., Chisholm, S. T., Shaner, D. L., ... &
Grey, T. L. (2010). Gene amplification confers glyphosate resistance in Amaranthus
palmeri. Proceedings of the National Academy of Sciences, 107(3), 1029-1034.
Giacomini, D., Westra, P., & Ward, S. M. (2014). Impact of genetic background in
fitness cost studies: an example from glyphosate-resistant Palmer amaranth. Weed
Science, 62(1), 29-37.
Gibson, K. D., Fischer, A. J., Foin, T. C., & Hill, J. E. (2002). Implications of delayed
Echinochloa spp. germination and duration of competition for integrated weed
management in water‐seeded rice. Weed Research, 42(5), 351-358.
Grant, W. F. (1959). Cytogenetic studies in Amaranthus: I. Cytological aspects of sex
determination in dioecious species. Canadian Journal of Botany, 37(3), 413-417.
Gunsolus, J. L. (1990). Mechanical and cultural weed control in corn and
soybeans. American Journal of Alternative Agriculture, 5(3), 114-119.
Heap, I. (2014). Global perspective of herbicide‐resistant weeds. Pest management
science, 70(9), 1306-1315.
Heap, I. The International Survey of Herbicide Resistant Weeds. http://weedscience.org
[1 February 2018].
Hensleigh, P., & Pokorny, M. (2017). Palmer amaranth (Amaranthus palmeri S. Watson).
Agronomy Technical Note MT-92. USDA-NRCS.

82
Herms, D. A., & Mattson, W. J. (1992). The dilemma of plants: to grow or defend.
Quarterly Review of Biology, 67(3), 283-335.
Herrmann, K. M., & Weaver, L. M. (1999). The shikimate pathway. Annual Review of
Plant Biology, 50(1), 473-503.
Hilgenfeld, K. L., Martin, A. R., Mortensen, D. A., & Mason, S. C. (2004). Weed
management in glyphosate resistant soybean: weed emergence patterns in relation to
glyphosate treatment timing. Weed Technology, 18(2), 277-283.
Hill, B. C., & Silvertown, J. (1997). Higher-order interaction between molluscs and sheep
affecting seedling numbers in grassland. Acta Oecologica, 18(5), 587-596.
Hock, S. M., Knezevic, S. Z., Martin, A. R., & Lindquist, J. L. (2006). Soybean row
spacing and weed emergence time influence weed competitiveness and competitive
indices. Weed Science, 54(1), 38-46.
Hoffmann, A. A., Donoghue, M. J., Levin, S. A., Mackay, T. F. C., Rieseberg, L., Travis,
J., Wray, G. A. (2014). Evolutionary limits and constraints. The Princeton guide to
evolution, 247-252.
Horak, M. J., & Loughin, T. M. (2000). Growth analysis of four Amaranthus
species. Weed Science, 48(3), 347-355.
Jasieniuk, M., Brûlé-Babel, A. L., & Morrison, I. N. (1996). The evolution and genetics
of herbicide resistance in weeds. Weed science, 176-193.
Jenks, M. A., & Hasegawa, P. M. (Eds.). (2005). Plant abiotic stress. Blackwell Pub.
Jha, P., Kumar, V., Godara, R. K., & Chauhan, B. S. (2017). Weed management using
crop competition in the United States: A review. Crop Protection, 95, 31-37.
Johnson, M. T., Smith, S. D., & Rausher, M. D. (2009). Plant sex and the evolution of
plant defenses against herbivores. Proceedings of the National Academy of
Sciences, 106(43), 18079-18084.
Jones, E. A. L., Owen, M. D. K., & Leon, R. G. (2019). Influence of multiple herbicide
resistance on growth in Amaranthus tuberculatus. Weed Research, 59(3), 235-244.
Jordan, D. L., York, A. C., Griffin, J. L., Clay, P. A., Vidrine, P. R., & Reynolds, D. B.
(1997). Influence of application variables on efficacy of glyphosate. Weed
Technology, 11(2), 354-362.
Jordan, N. R., & Jannink, J. L. (1997). Assessing the practical importance of weed
evolution: a research agenda. Weed Research, 37(4), 237-246.

83
Kalisz, S. (1986). Variable selection on the timing of germination in Collinsia verna
(Scrophulariaceae). Evolution, 40(3), 479-491.
Kawagoe, T., & Kudoh, H. (2010). Escape from floral herbivory by early flowering in
Arabidopsis halleri subsp. gemmifera. Oecologia, 164(3), 713-720.
Keeley, P. E., Carter, C. H., & Thullen, R. J. (1987). Influence of planting date on growth
of Palmer amaranth (Amaranthus palmeri). Weed Science, 199-204.
Kim, D. S., Brain, P., Marshall, E. J. P., & Caseley, J. C. (2002). Modelling herbicide
dose and weed density effects on crop: weed competition. Weed research, 42(1), 1-13.
Kistner, E. J., & Hatfield, J. L. (2018). Potential Geographic Distribution of Palmer
Amaranth under Current and Future Climates. Agricultural & Environmental
Letters, 3(1).
Koornneef, M., Bentsink, L., & Hilhorst, H. (2002). Seed dormancy and
germination. Current opinion in plant biology, 5(1), 33-36.
Koricheva, J., Nykänen, H., & Gianoli, E. (2004). Meta-analysis of trade-offs among
plant antiherbivore defenses: are plants jacks-of-all-trades, masters of all?. The American
Naturalist, 163(4), E64-E75.
Kropff, M. J., Weaver, S. E., & Smits, M. A. (1992). Use of ecophysiological models for
crop-weed interference: relations amongst weed density, relative time of weed
emergence, relative leaf area, and yield loss. Weed Science, 40(2), 296-301.
Kuester, A., Chang, S. M., & Baucom, R. S. (2015). The geographic mosaic of herbicide
resistance evolution in the common morning glory, Ipomoea purpurea: Evidence for
resistance hotspots and low genetic differentiation across the landscape. Evolutionary
Applications, 8(8), 821-833.
Lambers, H., Chapin III, F. S., & Pons, T. L. (2008). Plant physiological ecology.
Springer Science & Business Media.
Lande, R., & Arnold, S. J. (1983). The measurement of selection on correlated
characters. Evolution, 37(6), 1210-1226.
Lankau, R. A., Nuzzo, V., Spyreas, G., & Davis, A. S. (2009). Evolutionary limits
ameliorate the negative impact of an invasive plant. Proceedings of the National
Academy of Sciences, 106(36), 15362-15367.
Larcher, W. (2003). Physiological plant ecology: ecophysiology and stress physiology of
functional groups. Springer Science & Business Media.
Lau, J. A., & terHorst, C. P. (2019). Evolutionary responses to global change in species‐

84
rich communities. Annals of the New York Academy of Sciences.
Leimu, R., & Koricheva, J. (2006). A meta‐analysis of tradeoffs between plant tolerance
and resistance to herbivores: combining the evidence from ecological and agricultural
studies. Oikos, 112(1), 1-9.
Leimu, R., Muola, A., Laukkanen, L., Kalske, A., Prill, N., & Mutikainen, P. (2012).
Plant‐herbivore coevolution in a changing world. Entomologia Experimentalis et
Applicata, 144(1), 3-13.
Lemen, C. (1980). Allocation of reproductive effort to the male and female strategies in
wind-pollinated plants. Oecologia, 45(2), 156-159.
Lipiec, J., Doussan, C., Nosalewicz, A., & Kondracka, K. (2013). Effect of drought and
heat stresses on plant growth and yield: a review. International Agrophysics, 27(4), 463-
477.
Lundemo, S., Falahati‐Anbaran, M., & Stenøien, H. K. (2009). Seed banks cause elevated
generation times and effective population sizes of Arabidopsis thaliana in northern
Europe. Molecular Ecology, 18(13), 2798-2811.
Marquis, R. J. (1992). Selective impact of herbivores. Plant resistance to herbivores and
pathogens: ecology, evolution, and genetics, 301. The University of Chicago Press, USA.
Mauricio, R., Rausher, M. D., & Burdick, D. S. (1997). Variation in the defense strategies
of plants: are resistance and tolerance mutually exclusive?. Ecology, 78(5), 1301-1311.
Miller, T. E., Winn, A. A., & Schemske, D. W. (1994). The effects of density and spatial
distribution on selection for emergence time in Prunella vulgaris (Lamiaceae). American
Journal of Botany, 81(1), 1-6.
Mitchell‐Olds, T. (1996). Pleiotropy causes long‐term genetic constraints on life‐history
evolution in Brassica rapa. Evolution, 50(5), 1849-1858.
Moolani, M. K., Knake, E. L., & Slife, F. W. (1964). Competition of smooth pigweed
with corn and soybeans. Weeds, 12(2), 126-128.
Murphy, S. D., Yakubu, Y., Weise, S. F., & Swanton, C. J. (1996). Effect of planting
patterns and inter-row cultivation on competition between corn (Zea mays) and late
emerging weeds. Weed science, 44(4), 865-870.
Nakka, S. (2016). Physiological, biochemical and molecular characterization of multiple
herbicide resistance in Palmer amaranth (Amaranthus palmeri). Kansas State University.

85
Núñez‐Farfán, J., & Dirzo, R. (1994). Evolutionary ecology of Datura stramonium L. in
central Mexico: natural selection for resistance to herbivorous insects. Evolution, 48(2),
423-436.
Núñez-Farfán, J., Fornoni, J., & Valverde, P. L. (2007). The evolution of resistance and
tolerance to herbivores. Annu. Rev. Ecol. Evol. Syst., 38, 541-566.
Oerke, E. C. (2006). Crop losses to pests. The Journal of Agricultural Science, 144(1),
31-43.
Owen, M. D. K. (1997). North American developments in herbicide tolerant crops.
In BRIGHTON CROP PROTECTION CONFERENCE WEEDS (Vol. 3, pp. 955-966).
BRIT CROP PROTECTION COUNCIL.
Pake, C. E., & Venable, D. L. (1996). Seed banks in desert annuals: implications for
persistence and coexistence in variable environments. Ecology, 77(5), 1427-1435.
Philippi, T. (1993). Bet-hedging germination of desert annuals: beyond the first year. The
American Naturalist, 142(3), 474-487.
Pilson, D. (2000). The evolution of plant response to herbivory: simultaneously
considering resistance and tolerance in Brassica rapa. Evolutionary Ecology, 14(4-6),
457.
Pimentel, D., Lach, L., Zuniga, R., & Morrison, D. (2000). Environmental and economic
costs of nonindigenous species in the United States. BioScience, 50(1), 53-66.
Powles, S. B. (2008). Evolved glyphosate‐resistant weeds around the world: lessons to be
learnt. Pest Management Science: formerly Pesticide Science, 64(4), 360-365.
Powles, S. B. (2010). Gene amplification delivers glyphosate-resistant weed evolution.
Proceedings of the National Academy of Sciences, 107(3), 955-956.
Powles, S. B., & Preston, C. (2006). Evolved glyphosate resistance in plants: biochemical
and genetic basis of resistance 1. Weed Technology, 20(2), 282-289.
Powles, S. B., & Yu, Q. (2010). Evolution in action: plants resistant to herbicides. Annual
review of plant biology, 61, 317-347.
Rajcan, I., & Swanton, C. J. (2001). Understanding maize–weed competition: resource
competition, light quality and the whole plant. Field Crops Research, 71(2), 139-150.
Rodríguez, J., Lorenzo, P., & González, L. (2021). Phenotypic plasticity of invasive
Carpobrotus edulis modulates tolerance against herbivores. Biological Invasions, 1-17.
Romeo, J. T., Saunders, J. A., & Barbosa, P. (Eds.). (2013). Phytochemical diversity and

86
redundancy in ecological interactions (Vol. 30). Springer Science & Business Media.
Rosenthal, J. P., & Kotanen, P. M. (1994). Terrestrial plant tolerance to herbivory. Trends
in Ecology & Evolution, 9(4), 145-148.
Sauer, J. (1957). Recent migration and evolution of the dioecious amaranths. Evolution,
11(1), 11-31.
Scursoni, J. A., Forcella, F., & Gunsolus, J. (2007). Weed escapes and delayed weed
emergence in glyphosate-resistant soybean. Crop Protection, 26(3), 212-218.
Seiwa, K. (1998). Advantages of early germination for growth and survival of seedlings
of Acer mono under different overstorey phenologies in deciduous broad‐leaved
forests. Journal of Ecology, 86(2), 219-228.
Sgrò, C. M., & Hoffmann, A. A. (2004). Genetic correlations, tradeoffs and
environmental variation. Heredity, 93(3), 241-248.
Sikkema, P. H. (2017). Perspectives on potential soybean yield losses from weeds in
North America. Weed technology, 31(1), 148-154.
Simms, E. L. (1990). Examining selection on the multivariate phenotype: plant resistance
to herbivores. Evolution, 44(5), 1177-1188.
Simms, E. L. (1992). Costs of plant resistance to herbivory. Plant resistance to
herbivores and pathogens: ecology, evolution, and genetics., 392-425.
Simms, E. L., & Rausher, M. D. (1987). Costs and benefits of plant resistance to
herbivory. The American Naturalist, 130(4), 570-581.
Simms, E. L., & Triplett, J. (1994). Costs and benefits of plant responses to disease:
resistance and tolerance. Evolution, 48(6), 1973-1985.
Sokal, R. R., & Rohlf, F. J. (1995). Biometry Freeman New York. Biometry, 3rd ed.
Freeman, New York.
Soltani, N., Dille, J. A., Burke, I. C., Everman, W. J., VanGessel, M. J., Davis, V. M., &
Sosnoskie, L. M., Webster, T. M., Kichler, J. M., MacRae, A. W., & Culpepper, A. S.
(2007, March). An estimation of pollen flight time and dispersal distance for glyphosate-
resistant Palmer amaranth (Amaranthus palmeri). In Proc South. Weed Sci. Soc (Vol. 60,
p. 229).
Stanton, M. L., Roy, B. A., & Thiede, D. A. (2000). Evolution in stressful environments.
I. Phenotypic variability, phenotypic selection, and response to selection in five distinct
environmental stresses. Evolution, 54(1), 93-111.

87
Steckel, L. E., Main, C. L., Ellis, A. T., & Mueller, T. C. (2008). Palmer amaranth
(Amaranthus palmeri) in Tennessee has low level glyphosate resistance. Weed
Technology, 22(1), 119-123.
Steinrücken, H. C., & Amrhein, N. (1980). The herbicide glyphosate is a potent inhibitor
of 5-enolpyruvylshikimic acid-3-phosphate synthase. Biochemical and Biophysical
Research Communications, 94(4), 1207-1212.
Steward, J. L., & Keeler, K. H. (1988). Are there trade-offs among antiherbivore defenses
in Ipomoea (Convolvulaceae)?. Oikos, 79-86.
Strauss, S. Y., & Agrawal, A. A. (1999). The ecology and evolution of plant tolerance to
herbivory. Trends in Ecology & Evolution, 14(5), 179-185.
Stinchcombe, J. R., & Rausher, M. D. (2002). The evolution of tolerance to deer
herbivory: modifications caused by the abundance of insect herbivores. Proceedings of
the Royal Society of London. Series B: Biological Sciences, 269(1497), 1241-1246.
Stoltenberg, D. E., & Wiederholt, R. J. (1995). Giant foxtail (Setaria faberi) resistance to
aryloxyphenoxypropionate and cyclohexanedione herbicides. Weed Science, 43(4), 527-
535.
Swanton, C. J., & Weise, S. F. (1991). Integrated weed management: the rationale and
approach. Weed Technology, 5(3), 657-663.
Teasdale, J. R. (1995). Influence of narrow row/high population corn (Zea mays) on weed
control and light transmittance. Weed technology, 9(1), 113-118.
Thungo, Z., Shimelis, H., Odindo, A. O., & Mashilo, J. (2019). Genotype-by-
environment interaction of elite heat and drought tolerant bread wheat (Triticum aestivum
L.) genotypes under non-stressed and drought-stressed conditions. Acta Agriculturae
Scandinavica, Section B—Soil & Plant Science, 69(8), 725-733.
Tiffin, P. (2002). Competition and time of damage affect the pattern of selection acting
on plant defense against herbivores. Ecology, 83(7), 1981-1990.
Tiffin, P., & Rausher, M. D. (1999). Genetic constraints and selection acting on tolerance
to herbivory in the common morning glory Ipomoea purpurea. The American
Naturalist, 154(6), 700-716.
Time and Date AS. Climate & Weather Averages in Rocky Mount, North Carolina, USA.
https://www.timeanddate.com/weather/@4488762/climate [13 January 2020].
van der Meijden, E. (1996). Plant defence, an evolutionary dilemma: contrasting effects
of (specialist and generalist) herbivores and natural enemies. In Proceedings of the 9th

88
International Symposium on Insect-Plant Relationships (pp. 307-310). Springer,
Dordrecht.
van der Meijden, E., Wijn, M., & Verkaar, H. J. (1988). Defence and regrowth,
alternative plant strategies in the struggle against herbivores. Oikos, 355-363.
Van Etten, M., Kuester, A., Chang, S. M., & Baucom, R. (2015). Reduced seed viability
and reductions in plant size provide evidence for costs of glyphosate resistance in an
agricultural weed. bioRxiv, 030833.
Van Oosterom, E. J., Mahalakshmi, V., Bidinger, F. R., & Rao, K. P. (1996). Effect of
water availability and temperature on the genotype-by-environment interaction of pearl
millet in semi-arid tropical environments. Euphytica, 89(2), 175-183.
Varanasi, V. K., Godar, A. S., Currie, R. S., Dille, A. J., Thompson, C. R., Stahlman, P.
W., & Jugulam, M. (2015). Field‐evolved resistance to four modes of action of herbicides
in a single kochia (Kochia scoparia L. Schrad.) population. Pest management
science, 71(9), 1207-1212.
Via, S. (1984). The quantitative genetics of polyphagy in an insect herbivore. II. Genetic
correlations in larval performance within and among host plants. Evolution, 896-905.
Vila‐Aiub, M. M., Neve, P., & Powles, S. B. (2009). Fitness costs associated with
evolved herbicide resistance alleles in plants. New Phytologist, 184(4), 751-767.
Walsh, M. J., Powles, S. B., Beard, B. R., Parkin, B. T., & Porter, S. A. (2004). Multiple-
herbicide resistance across four modes of action in wild radish (Raphanus
raphanistrum). Weed science, 52(1), 8-13.
Ward, S. M., Webster, T. M., & Steckel, L. E. (2013). Palmer amaranth (Amaranthus
palmeri): a review. Weed Technology, 27(1), 12-27.
Webster, T. M., Everest, A. M. P. J., Brecke, F. J. F. B., Martin, K. J. G. J., Kelly, L. E.
W. S., Griffin, J., ... & Murray, O. C. M. D. (2001). Weed survey-Southern states, 2001:
Broadleaf crops subsection (Cotton, peanut, soybean, tobacco, and forestry). In Proc.
South. Weed Sci. Soc (Vol. 54, pp. 244-259).
Webster, T. M., & Nichols, R. L. (2012). Changes in the prevalence of weed species in
the major agronomic crops of the Southern United States: 1994/1995 to 2008/2009. Weed
Science, 60(2), 145-157.
Weinig, C., Stinchcombe, J. R., & Schmitt, J. (2003). Evolutionary genetics of resistance
and tolerance to natural herbivory in Arabidopsis thaliana. Evolution, 57(6), 1270-1280.
Welch, R. Y., Behnke, G. D., Davis, A. S., Masiunas, J., & Villamil, M. B. (2016). Using
cover crops in headlands of organic grain farms: Effects on soil properties, weeds and

89
crop yields. Agriculture, Ecosystems & Environment, 216, 322-332.
Welles, S. R., & Funk, J. L. (2021). Patterns of intraspecific trait variation along an
aridity gradient suggest both drought escape and drought tolerance strategies in an
invasive herb. Annals of Botany, 127(4), 461-471.
Wise, M. J., & Abrahamson, W. G. (2005). Beyond the compensatory continuum:
environmental resource levels and plant tolerance of herbivory. Oikos, 109(3), 417-428.
Xiao, Z., Harris, M. K., & Zhang, Z. (2007). Acorn defenses to herbivory from insects:
implications for the joint evolution of resistance, tolerance and escape. Forest Ecology
and Management, 238(1-3), 302-308.
Yakimowski, S. B., Teitel, Z., & Caruso, C. M. (2021). Defense by duplication: The
relation between phenotypic glyphosate resistance and EPSPS gene copy number
variation in Amaranthus palmeri. bioRxiv.
Zenni, R. D., Bailey, J. K., & Simberloff, D. (2014). Rapid evolution and range
expansion of an invasive plant are driven by provenance–environment
interactions. Ecology letters, 17(6), 727-735.
Zimdahl, R. L. (1980). Weed-crop competition: A review. Weed-crop competition: a
review.