effect of standard bacterial vaccine on influenza virus infection and interferon production in...
TRANSCRIPT

Acta path. microbiol. scand. Section B. 81, 782-786, 1973
EFFECT OF STANDARD BACTERIAL VACCINE
ON INFLUENZA VIRUS INFECTION AND INTERFERON PRODUCTION IN
GERMFREE MICE MIKLOS D E ~ and TORE IDTVEDT
Kaptein W. Wilhelmsen og Frues Bakteriologiske Institutt, University of Oslo, Rikshospitalet, Oslo, Norway
A standard bacterial vaccine (SBV) induced interferon (IF) production and reduced the mortality due to influenza A, virus infection in germfree mice. These effects were quanti- tatively cumparable in conventional and germfree mice, The length of survival was some- what shorter in conventional mice. It is concluded, that SBV exerts a direct effect on the viral infection probably partly mediated by IF induction. The role of resident bacterial flora is discussed.
I n previous communications we reported that a standard bacterial vaccine (SBV) reduced the mortality following i d u m z a Az virus in- fection in mice (4). Furthermore, develop- ment of grass pulmonary lesions, growbh of virus in the lungs and antibody titres against the virus were also reduced by intraperitone- ally (i.p.) injected SBV, when the vaccine was given a few hours prior to viral infection. Similar protection was obtained against para- influenza 1 Sendai virus, Semliki forest virus (10, 17) and Vesicular stomatitis virus (4) . We have suggested ( 4 ) as did Singer & Har- degree (1 7 ) that the protection may at least in part be mediated by production of inter- feron or interferon-like substance.
It is well documented both in clinical and experimental studies, that viral respiratory infections are aften complicated by secondary bacterial invasion, which may influence the outcome of disease (6, 14). Reduction or
Received 17.vii.13 Accepted 1 l.vii.13
782
eliminat.icm of secondary invaders reduced the mortality and increased the length of survi- v d in Sendai virus infected mice (3, 6 ) . A possible effect of SBV on this secondary bacterial invasion should be considered. We decided 'to investigate this problem by extend- ing the previous experiments to germfree mice.
M A T E R I A L A N D M E T H O D S
Mice: Young albino, mice, both male and female, of CD 1 and NMRI strains were used. CD 1 mice were obtained germfree (GF) from the Charles River Mouse Farms, Wilmington, Mass., USA. The mice were kept under germfree conditions as described elsewhere (15). Negative cultures during and at the termination of experiments proved that the animals remained GF. The mice were 6-8 weeks of age at the time of the initiation of experiments.
NMRI specific pathogen free mice were origin- ally obtained from the National Institute of Public Health, Oslo.

TABLE 1. Mortality of Conventional and Germfree Mice Following Intranasal Inoculation of 0.1 Ml Influenza A Virus
Germfree Conventional Virus titre Dead Mean survival
Total days Dead Mean survival Total days
lo-' 4/4* 1.25 4/4 5.75 10-5 3/4 1 1.50 3/4 9.75
3/4 12.75 4/4 10.00 10-c 10-7 1 /4 0/4 10-8 0/4 0/4 LDX, 10 -Li .3 10-G.3
* Final mortality after 14 days observation.
Virus: Influenza A,/Singapore 1/57 strain was given to us by Dr. K. Herrherg, Frankfurt a.M., W.Germany. In ou r laboratory the virus was passed once in the allantoic cavity of 10 days old embryonated eggs, The allantoic fluids were harvest- ed 3 days after inoculation. Allantoic fluids with high haemagglutination titres were pooled and stored at -70" C. The infectious titre was tested in vitro in Vero cells and in vivo in mice, by in- tranasal inoculation of 0.1 mi of the virus.
Standard bacterial vaccine: The vaccine was purchased from the National Institute of Public Health, Oslo. The vaccine is composed of formalin- killed bacteria suspended in NaCl solution, and contains species regularly isolated from the re- spiratory tract, total 1.8 x 108 bacteria per ml
Interferon assay: Blood was obtained from the axillary vein at times indicated in the text. Serum was separated after clotting. All samples were ad- justed to pH 2 by 1N HCI, and kept at 4" C for two days. Before testing, the pH was readjusted to neutral by 1N NaHCO,,. Interferon activity in the sera was assayed by means of the infectivity inhibition test, employing the micro assay as described in detail elsewhere ( 2 ) . In brief, twofold dilutions of the Sera were added to freshly seeded L-F, cells in plastic microtrays. After incubation overnight the test cups and control cups without serum, were challenged with approximately 10 mean tissue culture infectious doses (TCID,,) of Vesicular stomatitis virus (VSV). A back titration of the challenge virus was run in parallel in each test. After sealing the trays they were incubated at 37" C. Microscopical reading was done after 3 days, when the end point titration of VSV was complete. The I F titre was estimated as the highest dilution which inhibited viral cytapatho- genic effect in 50 per cent of the cups, calculated by the method of Reed and Muench, The titres were calibrated to 10 TCID,, of challenge virus by means of the standard slope of regression line
(4 ) .
for mouse I F tested in the L-F,/VSV system, as described in detail elsewhere ( 2 ) .
R E S U L T S
Infectivity of Influenza Virus in GF and Conventional Mice
Mice were inoculated intranasally ( in . ) with 0.1 ml of tenfold dilutions of influenza A, virus without anaesthesia. Each dilution was given to 4 mice. The inoculated mice were observed for 14 days and mortality was recorded twice daily. The mean lethal dose of virus was practically the same in GF and in conventional mice (Table 1). The survi- val times were generally somewhat shorter in conventional mice 'than in their GF counter- parts given the same virus dose, but rhe dif- ferences were not significant (Wilcoxons signed rank test: 0.1 > p > 0.05).
Effect of SBV on Influenza Virus Caused Mortality
Twenty GF mice were injected i.p. with 0.1 ml undiluted SBV. Five hours later they were inoculated with 10 LD,, influenza A2 virus. Another group of 20 GF mice, and 30 conventional mice received the same virus inoculation but no SBV. All mice were ob- served for 14 days and mortality was recorded twice daily (Fig. 1). Total mortality after 14 days in influenza inoculated GF mice was somewhat reduced by SBV treatment from 41.5 to 25 per cent ( x 2 : 2.780,0.l>p>0.05).
783

DAYS AFTER VIRUS INOCULATION
Fig. 1. Cumulative martality of conventional and germfree mice following i.n. inoculation of 10 LD,, influenza A virus, and the influence oC i.p. in- jected standard bacterial vaccine.
The final mortality of influenza inoculated conventional mice was almost the same as that of their GF counterparts, but they seem to die after a shorter lag period than GF mice.
Interferon Induction by SBV in GF and Conuentional Mice
A group of GF mice was injected i.p. 0.1 ml undiluted SBV. At 2, 4, 6, 8, 24 and 48 hours later 2 mice were removed from the isolator, bleed from the axillary vein, and the IF content of their pooled serum was deter- mined. A parallel experiment was done in conventional mice. Interferon activity was detected both in GF and in conventional mice (Fig, 2 ) . Peak titres were comparable in the two groups, perhaps it was slighztly higher in GF mice. Peak titres were demonstrated somewhat later in GF than in conventional mice.
D T S C U S S I O N
The present data give us some information which supplements the data presented in pre- vious communications. The SBV does have
a preventive effect on influenza virus infec- tion also in GF mice. The extent of this effect is comparable to that seen in conventional animals (1 ). This finding indicates that the preventive effect of the vaccine is mainly di- rected against the viral infeotion and it is not dependent on the factor represented by the possible secondary bacterial invasion. An interaction between the infecting viral agent and the resident flora of the animals seem3 to be of minor importance in this connec- tion.
The mechanism of the preventive effect is still not clear. IF production was comparable in GF and conventional mice. In fact, it is to be expected if IF is important factor in the antiviral effect of SBV, as the prevention is also of the same magnitude. Interferon effect on viral infection in GF mice has been demonstrated earlier (12, 13). The IF levels in the present model seems to be too low to represent the entire explanation of the pre- ventive effect of vaccine. Exogenous IF with comparable titres does not influence experi- mental viral infections. Doses of viral and nonviral IF induces which gave significant protection against encephalolmyocarditis or
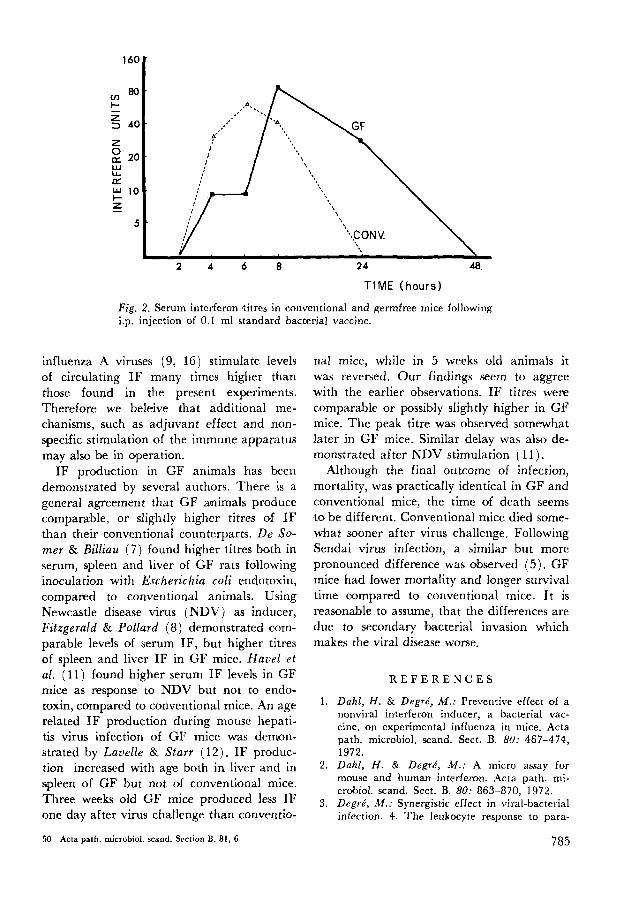
T I M E (hours)
Fig. 2. Serum interferon titres in conventional and gennfree mice following i.p. injection of 0.1 ml standard bacterial vaccine.
influenza A viruses (9, 16) stimulate levels of circulating I F many times higher than those found in the present experiments. Therefore we beleive that additional me- chanisms, such as adjuvant effect and non- specific stimulation of the immune apparatus may also be in operation.
IF production in GF animals has been demonstrated by several authors. There is a general agreement that GF animals produce comparable, or slightly higher titres of IF than their conventional counterparts. De So- mer & Billiau ( 7 ) found higher titres both in serum, spleen and liver of GF rats following inoculation with Eschericliia coli endotoxin, compared to conventional animals. Using Newcastle disease virus (NDV) as inducer, Fitzgerald & Pollard (8) demonstrated com- parable levels of serum IF, but higher titres of spleen and liver I F in GF mice. Have1 et al. ( 11 ) found higher serum IF levels in GF mice as response to NDV but not to endo- toxin, compared to conventional mice. An age related I F production during mouse hepati- tis virus infection of GF mice was demon- strated by Lavelle & Starr (12) . I F produc- tion increased with age both in liver and in spleen of GF but not of conventional mice. Three weeks old GF mice produced less IF one day after virus challenge than conventio-
50 Acta path. miciobiol. scand. Srction B. 81, 6
nal mice, while in 5 weeks old animals it was reversed. Our findings seem to aggree with the earlier observations. I F titres were comparable or possibly slightly higher in GF mice. The peak titre was observed somewhat later in GF mice. Similar delay was also de- monstrated after NDV stimulation (11).
Although the final outcome of infection, mortality, was practically identical in GF and conventional mice, the time of death seems to be different. Conventional mice died some- what sooner after virus challenge. Following Sendai virus infection, a similar but more pronounoed difference was observed (5). GF mice had lower mortality and longer survival time compared to conventional mice. I t is reasonable to assume, that the differences are due to secondary bacterial invasion which makes the viral disease worse.
R E F E R E N C E S
Dahl, H . & DegrC, M . : Preventive effect of a nonviral interferon inducer, a bacterial vac- cine, on experimental influenza in mice. Acta path. microbiol. scand. Sect. B. 80: 467-474, 1972. Dahl, H . & Degri , M.: A micro assay for mouse and human interferon. Acta path. mi- crobiol. scand. Sect. B. 80: 863-870, 1972. Degri , M.: Synergistic effect in viral-bacterial infection. 4. The leukocyte response to para-
785

influenza virus infection in the respiratory tract of mice, and the influence of antibiotics on the development of infection. Acta path.’ micmbiol. scand. Sect. B. 79: 88-94, 197 1.
4. Degrk, M . & Dahl, H.: Production d an interferon-like agent following inoculation with bacterial vaccine. Proc. SOC. Exptl. Biol. Med.
5. Degrk, M. & Midtvedt, T . : Respiratory infec- tion with parainfluenza 1, Sendai virus in gncr tobiotic and conventional mice. Acta path. microbiol. scand. Sect. B. 79: 123-124, 1971.
6. Degrk, M . & Solberg, L. A, : Synergistic effect in viral-bacterial infection. 3. Histopathologi- cal changes in the trachea of mice following viral and bacterial infection. Acta path. mi- crobiol. s a n d . Sect. B. 79: 129-136, 1971.
7 . De Somer, P . & Billiau, A , : Interferon pro- duction by spleen of rats after intravenous in- jection of sindbis virus or heat-killed Escheri- chia coli. Arch. Ges. Virusforsch. 19: 143- 154, 1966.
8. Fittgerald, G . R . & Pollard, M.: Interferon production by germfree mice. Proc. SOC. Exptl. Biol. Med. 126: 245-249, 1967.
9. Gresser, I . , Fontaine-Brouty-Boyk, D., Bourali, C . & Thomas, M.T.: A comparison of the efficacy of endogenous, exogenous, and com- bined endogenous-exogenous interferon in the treatment of mice infected with encephalo- myocarditis virus. Proc. SOC. Exptl. Biol. Med.
137: 233-236, 1971.
130: 236-242, 1969.
10. Hardegree, M . C., Singer, S . H . & Gerone, P. J . : Effect of allergenic extracts of house dust and bacterial vaccine on respiratory in- fections of mice. J. Allergy Clin. Immunol. 51: 1-10, 1973.
11. Havel, E . H., Holtermann, 0. A . & Starr, T . J . : The influence of indigenous bacterial flora upon the interferon response of mice. J. gen. Virol. 9: 93-95, 1970.
12. Lavelle, G. C. & Starr, T . ] . : Interferon re- sponse and age-related resistance of germfree mice to mouse hepatitis virus. J. Reticuloendo- thel. SOC. 5: 422-435, 1968.
13. Lavelle, G. C. & Starr, T . J . : Effects of corti- sone on interferon response of germfree mice to mouse hepatitis virus. J. Reticuloendothel.
14. Loosli, C. G.: Synergism between respiratory viruses and bacteria. Yale J. biol. Med. 40:
15. Midtvedt, T . & Trippestad, A.: Opsonizing and bactericidal effects of sera from gnoto- biotic and conventionalized rats on SSP- labelled E. coli. Acta path. microbiol. scand. 78: 1-5, 1970.
16. Portnoy, J . & Merigan, T . C.: The effect of interferon and interferon inducers on avian influenza. J. Infect. Dis. 124: 545-552, 1971.
17. Singer, S . H . & Hardegree, M . C.: Induction of interferon by bacterial vaccines and aller- genic extracts. J. Allergy 47: 332-340, 1971.
SOC. 5: 436-444, 1968.
522-540, 1967/68.
786