effect of continuous light on daily levels of plasma ...ruoff/salmon2.pdf · effect of continuous...
TRANSCRIPT

EFFECT OF CONTINUOUS LIGHT ON DAILY LEVELS OF PLASMAMELATONIN AND CORTISOL AND EXPRESSION OF CLOCKGENES IN PINEAL GLAND, BRAIN, AND LIVER IN ATLANTICSALMON POSTSMOLTS
Tien-sheng Huang,1 Peter Ruoff,2 and Per G. Fjelldal3
1National Institute of Nutrition and Seafood Research (NIFES), Nordnes, Bergen,Norway2Centre for Organelle Research, University of Stavanger, Stavanger, Norway3Institute of Marine Research (IMR), Matredal, Norway
Continuous light is a common practice in salmon farming, where it is used toenhance growth, induce smoltification, and regulate puberty. However, knowledgeabout how different tissues receive information about daylength is limited. The aimof the present study was to evaluate the daily expression of clock (Per1-like, Cry2, andClock), the nuclear transcription factor (peroxisome proliferator-activated receptor,PPAR; CCAAT/enhancer binding protein, C/EBP), and the endoplasmic reticulum(ER) stress (protein disulfide isomerase associated 3, PDIA3) genes in the pinealgland, brain, and liver of Atlantic salmon postsmolts reared under 12-h light:12-hdark (LD) regimes or under continuous light (LL) for 6 wks following transfer to sea-water. All measured clock mRNAs displayed daily variations in one or more organsunder LD, as well as plasma levels of melatonin. Similar variations were noted in theliver c/ebpα, pineal c/ebpδ, and pdia3 mRNAs. Under LL, the clock and nuclear tran-scription factor mRNAs did not show any daily variation in the studied organs, withthe exception of pineal pdia3. Furthermore, LL had the opposite effect on the levelsof melatonin and cortisol, as observed by the increase in pineal Clock, Per2, pparα,and c/ebpα and c/ebpδ mRNAs and decrease in liver Clock, Per2, and pparα mRNAscompared to those under LD. The present findings show that the expression of clockgenes is affected by the light across organs and that there is a relation between PPAR,C/EBP, and clock mRNAs; however, the functional role of the individual nucleartranscription factors related to this observation remains to be established in the pinealgland and liver. (Author correspondence: [email protected])
Keywords Atlantic salmon; Clock gene; Cortisol; Light; Melatonin; Nucleartranscription factor
1715
Address correspondence to Dr. Tien-sheng Huang, National Institute of Nutrition and SeafoodResearch (NIFES), P. O. Box 2029 Nordnes, N-5817 Bergen, Norway. Tel: (47) 98859588; Fax: (47)55905299; E-mail: [email protected]
Submitted April 7, 2010, Returned for revision May 7, 2010, Accepted August 11, 2010
Chronobiology International, 27(9–10): 1715–1734, (2010)Copyright © 2010 Informa Healthcare USA, Inc.ISSN 0742-0528 print/1525-6073 onlineDOI: 10.3109/07420528.2010.521272
Chr
onob
iol I
nt D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y pe
ter
ruof
f on
10/
29/1
0Fo
r pe
rson
al u
se o
nly.

INTRODUCTION
Molecular clocks control the daily variations of wakefulness, stresshormones, lipid metabolism, immune function, and cell division, amongother functions and processes (Duez & Staels, 2008). Genetic studies haveidentified components of the circadian pacemaker, including negativeregulators, period (PER) and cryptochrome (CRY), as well as positive reg-ulators, CLOCK and BMAL1, which are conserved in mammals wherethey constitute the core of the circadian clock described as transcrip-tional-translational feedback loops (Duguay & Cermakian, 2009; Gauger& Sancar, 2005). These feedback loops control the central and peripheralclocks that generate and maintain the functions and properties of varioustissues and cell types during constant environmental conditions(Balsalobre, 2002; Ralph et al., 1990). Circadian clocks can be induced insome cultured cell lines and stem cells in vitro by different stimuli(Balsalobre et al., 1998; Huang et al., 2009a; Mendez-Ferrer et al., 2008).In contrast to the mammalian biological clock located in the suprachias-matic nucleus (SCN) of the brain, to date the location of biological clockin fish is not known. Although the zebrafish is a useful model to study theeffects of environmental stimuli, including light and temperature on cir-cadian clock mechanisms (Hirayama et al., 2005; Lahiri et al., 2005),related studies are still lacking in other teleosts. Recently, Clock and Per1has been mapped to a quantitative trait locus to act as potential candi-dates for the timing of spawning in rainbow trout (Leder et al., 2006),and daily variations in clock gene expression have been shown in Atlanticsalmon parr brains (Davie et al., 2009) and pineal glands (Huang et al.,2010b). These studies suggest the existence of a clock gene system thatcouples inputs from the environment to complex biological functions andcontrols the timing of lifecycle transitions in salmonids.
In seasonal mammals, robust annual cycles of adiposity, food intake,and energy metabolism are driven by daylength, which generates adiurnal melatonin profile and acts on neuroendocrine pathways(Rousseau et al., 2003). Salmonids also exhibit behavioral and physiologi-cal rhythms, of which the generation of the biological timing system isthought to control their daily and annual functions in relation to develop-ment, locomotor activity, sedation, skin pigmentation, oxygen consump-tion, and thermoregulation (Falcon et al., 2010). Seasonal changes inphotoperiod are the most predictable environmental cues to modulatedaily and annual rhythms. Daylength controls the release of pineal mela-tonin in the blood, with low and high levels during the light and darkphases, respectively (Falcon, 1999). In Atlantic salmon, photoperiod influ-ences behavior (Oppedal et al., 2001); appetite (Nordgarden et al.,2003a); muscle lipid composition (Nordgarden et al., 2003b); develop-ment and growth of the axial muscle (Johnston et al., 2003), vertebral
T.-s. Huang et al.1716
Chr
onob
iol I
nt D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y pe
ter
ruof
f on
10/
29/1
0Fo
r pe
rson
al u
se o
nly.

column (Fjelldal et al., 2005), and gonads (Taranger et al., 1998); as wellas plasma levels of melatonin (Porter et al., 1999), cortisol (Thorpe et al.,1987), growth hormone (GH), insulin-like growth, factor 1 (IGF-I)(Nordgarden et al., 2005), and sex steroids (Taranger et al., 1998).Cortisol and GH act together during smoltification and seawater adaptionin regulating gill sodium-potassium adenosine triphosphatase (Na+,K+-ATPase) activity (McCormick, 2001). However, light-induced effects onthe temporal relationship between physiological clocks and plasma levelsof melatonin and cortisol are not clear in Atlantic salmon postsmolts.
CCAAT/enhancer binding protein (C/EBP) in the mollusc eye(Hattar et al., 2002), C/EBPδ in the hamster pineal gland (Marondeet al., 2007), and peroxisome proliferator-activated receptor (PPARα) inthe rat liver (Lemberger et al., 1996) have been identified as clock-controlled genes with diurnal expression. In addition, PPARα is alsoinvolved in the circadian regulation of lipid metabolism (Oishi et al.,2005). In Atlantic salmon, PPAR and C/EBP display important roles inregulating lipid metabolism in hepatocytes, adipocytes, and myocytes(Huang et al., 2010a; Kleveland et al., 2006; Vegusdal et al., 2004), andstress-induced pdia3 mRNA expression was found in the liver of smoltssuffering from hypoxia (Huang et al., 2009b). Less is known about theassociation of clock and nuclear transcription factor mRNAs in teleost.
The clock gene system has previously been shown in Atlantic salmonparr and smolts (Davie et al., 2009; Huang et al., 2010b), whereas theexpression of clock and clock-controlled genes and their response to lighthas not been reported in neural and peripheral tissues in postsmolts. Inorder to better understand the daily expression of clock and nuclear tran-scription factor mRNAs in Atlantic salmon in seawater, we described theplasma levels of melatonin and cortisol, in addition to the expression ofclock (Per1-like, Per2, Cry2, and Clock), nuclear transcription factor ( pparand c/ebp), and endoplasmic reticulum (ER) stress ( pdia3) mRNAs in thepineal gland, brain, and liver of postsmolts reared under 24 h of continu-ous light (LL) or a 12-h light:12-h dark (LD) cycle.
MATERIALS AND METHODS
Fish Stock
The underyearling Atlantic salmon (Salmo salar L.) smolts used in thepresent experiment were produced at the Institute of Marine Research(IMR), Matre Research Station, western Norway. The fish were from theAquagen strain, and smoltification was induced by an artificial photo-period commonly used in commercial salmon farming. The fish hadbeen reared under LD for 6 wks, followed by LL for 6 wks in freshwateruntil December 14, 2007, when they were transferred to experimental
Clock Gene Expression in Postsmolts 1717
Chr
onob
iol I
nt D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y pe
ter
ruof
f on
10/
29/1
0Fo
r pe
rson
al u
se o
nly.

seawater tanks. On December 14, the smolts had a low condition factor,lacked parr marks, and were silvery, and the mean gill Na+,K+-ATPaseactivity was 9.2 mmol adenosine diphosphate (ADP)/mg protein/h (minvalue 6.9, max value 12.5), which confirmed complete smoltification.
Experimental Design
On December 14, 2007, 300 underyearling smolts (mean weight196 g) were randomly allocated to six square, white, covered seawatertanks (1 × 1 × 0.43 m), with 50 fish/tank. For the following 6 wks (fromDecember 14, 2007 to January 25, 2008), triplicate groups in separatetanks were employed with LD or LL. For illumination, two 18-W fluor-escent daylight tubes (OSRAM L 18W/840 LUMILUX; OSRAM,Ausburg, Germany) were used to produce 960 lux underwater in thecenter of the tank. The light of the LD tanks was switched on at 08:05 hand off at 20:05 h. Both the LD and LL groups were fed by disc feedersthat continuously release pellets into the water during the lighted periodfor the LD group. The feed used was a commercial salmon feed (NutraOlympic; Skretting, Averøy, Norway), and the groups were fed by auto-matic feeders (ARVO-TEC T Drum 2000; Arvotec, Huutokoski, Finland).The fish were starved 24 h prior to sampling. The light and feeding werecontrolled automatically by a PC-operated system (Normatic, Norfjordeid,Norway). The temperature was stable at 8.9°C, and the oxygen saturationof the outlet was kept >80%.
Sampling of Atlantic Salmon Tissues for Expression Analysis
The samplings were performed on January 24–25, 2008, withsampling performed every 4 h; the first sampling started at 12:00 h onday 1 and the last started at 12:00 h on day 2 (day 1: 12:00, 16:00, 20:00,24:00 h, and day 2: 04:00, 08:00, and 12:00 h). At each sampling, 3 fish/tank were anesthetized with benzokain (40 mg L−1) (Benzoak® Vet;A.C.D., Braine-L’alleud, Belgium) and measured for fork length andbody weight. Blood samples were then taken from the caudal vessel usinga heparinized tuberculin syringe fitted with a 23-gauge needle. Plasmawas separated by centrifugation and stored at −80°C until analysis. Thenight sampling of individual fish of the LD tanks was performed under adim-red light. The fish were then decapitated with a scalpel, and thetissues were dissected from the carcass and immediately frozen in liquidnitrogen. During the sampling, all fish were anesthetized at the sametime, and then 1 fish/tank was sequentially measured and dissected toomit systematic errors caused by time-related differences in geneexpression between the tanks and light groups within the subsamplings.The experimental protocol and procedures were performed in
T.-s. Huang et al.1718
Chr
onob
iol I
nt D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y pe
ter
ruof
f on
10/
29/1
0Fo
r pe
rson
al u
se o
nly.

accordance with both international ethical standards (Portaluppi et al.,2008) and the National Animal Research Authority in Norway.
Levels of Plasma Melatonin and Cortisol
Melatonin levels were determined by radioimmunoassay (RIA) kit, inwhich the plasma sample was incubated with assay buffer, 125I-melatonin,and rabbit anti-melatonin antiserum, as described in the manufacturer’sinstructions (IBL, Hamburg, Germany). The cortisol concentration wasmeasured by RIA kit (DiaSorin, Minnesota, USA), in which 10 μL plasmasamples were applied in 1 mL tracer-buffer before incubation with125I-cortisol and rabbit anti-corticosterone antibody followed by thestandard protocol (Stefansson et al., 1989).
Isolation of RNA and qRT-PCR Analysis
RNA isolation and DNase treatment of the tissue samples were per-formed using a standard RNA extraction method (Invitrogen, Paisley,UK) and DNA-free™ DNase following the manufacturer’s protocol. Thetotal RNA, 500 ng/sample, was reverse transcribed using TaqMan RTreagents (ABI, Foster City, CA, USA) according to the manufacturer’sinstructions. A standard curve was obtained from a two-fold serial dilutionof total RNA from the control fish, with six RNA concentrations coveringbetween 31.25 ng and 1000 ng. The quantitative reverse transcriptase–-polymerase chain reaction (qRT-PCR) primer sequences (Table 1) weredesigned from expressed sequence tag (EST) sequences of Per2, Cry2,and Clock, obtained from the Salmon Genome Project (SGP) database,www.salmongenome.no, as described by Davie et al. (2009). In order toidentify salmon Per ortholog from the EST sequence, PCR primers, 5′-CCCAGTGACCTACTGGACCTG-3′ and 5′-TGGTGCTCTCTGGTAAGG-3′, were designed to amplify a partial cDNA fragment (576 bp) fromAtlantic salmon liver mRNA by PCR reaction. We refer to this gene as thePer1-like gene. The PCR primers for PPAR and C/EBP genes weredesigned from published sequences of Atlantic salmon (Huang et al.,2010a). The amplified PCR products of all three clock genes weresequenced to ensure correct mRNA sequences for quantification. Theprimers shown in Table 1 were used for SYBR Green assays (RocheDiagnostics, Germany) in a total volume of 20 μL containing 1 μL cDNA,10 μL SYBR Green master, 0.2 μL (50 μM) of each primer, and 8.6 μLH2O. Reactions were run on a Roche Lightercycler 480 (RocheDiagnostics) with 96 wells. The cycling conditions were composed of pre-incubation at 95°C for 5 min, followed by 45 cycles of amplification. Eachcycle included 10 s of denaturation at 95°C, 20 s of primer annealing at60°C, and 30 s of extension/synthesis at 72°C. The gene expression
Clock Gene Expression in Postsmolts 1719
Chr
onob
iol I
nt D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y pe
ter
ruof
f on
10/
29/1
0Fo
r pe
rson
al u
se o
nly.

TABLE 1 Primer sequences used in qRT-PCR of the reference and clock genes studied
Gene Accession number Forward primer Reverse primer
pdia3 EJ457023 5′-CCAACGCCATGATCAAGAA-3′ 5′-CAGCAGGTCCTTGTCCTTCA-3′
c/ebpα EU668995 5′-AGACCTCGGCGAGATTTGT-3′ 5′-TGTGGAATAGATCAGCCAGGAA-3′
c/ebpδ EU668997 5′-TTGGGCGGTGGAGCCTAT-3′ 5′-TTTCCTCGCCCGTGTCAT-3′
pparα DQ294237 5′-TCCTGGTGGCCTACGGATC-3′ 5′-CGTTGAATTTCATGGCGAACT-3′
Per1-like DW576689 5′-CTGTCTCCTTGGGCACTGTGT-3′ 5′-GAGTCGATGCTGCCAAAGTACTT-3′
Clock CA038738 5′-GTTAGACGGCTTCTTCCTAGCAAT-3′ 5′-CCACCAGGTCAGAAGGAAGATG-3′
Cry2 DY730105 5′-CCACAACATGGCAGGTGAATT-3′ 5′-CAGACATCCAAAGCGGAGGTA-3′
Per2 FM877775 5′-GCTCCCAGAATTCCTAGTGACAAG-3′ 5′-GAACAGCCCTCTCGTCCACATC-3′
β-Actin BG933897 5′-CCAAAGCCAACAGGGAGAAG-3′ 5′-AGGGACAACACTGCCTGGAT-3′
EF1AB AF321836 5′-TGCCCCTCCAGGATGTCTAC-3′ 5′-CACGGCCCACAGGTACTG-3′
ARP AY255630 5′-GAAAATCATCCAATTGCTGGATG-3′ 5′-CTTCCCACGCAAGGACAGA-3′
Chr
onob
iol I
nt D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y pe
ter
ruof
f on
10/
29/1
0Fo
r pe
rson
al u
se o
nly.

stability (M) was calculated with geNorm, and the three reference genes(ARP, β-actin, and EF1AB) revealed a low M value. The geNorm tool wasused to determine a normalizing factor from the three reference genes(acidic ribosomal phosphoprotein [ARP], β-actin, and elongation factor 1AB [EF1AB]) to postsmolts (LD and LL), and it was also used to calculatethe normalized expression for the target genes (Vandesompele et al.,2002).
Statistical Analysis
Significant differences in both length and weight were tested by two-way nested analyses of variance (ANOVAs), with the tank as the randomfactor nested in the light group, and those in the plasma levels of melato-nin and cortisol and in the mRNA expression of each clock, nuclear tran-scription factor, and pdia3 genes were evaluated by one-way ANOVA(Statistica; Statsoft, Tulsa, USA). If the ANOVA test showed statistical sig-nificance ( p< .05), a Newman-Keuls test was applied to determine allpairwise comparisons. Because ANOVA cannot assess rhythmicity, thecosinor method was used to test for circadian rhythmicity using normaliz-ing data for each variable in three individual analyses (the average of the 3fish/tank, n= 3 each timepoint) and in one cosinor test using the meansfor each timepoint (means of total fish from all the tanks). These analyseswere performed by the cosinor periodogram (www.circadian.org/softwar.html), with significance of the periodicity determined by the zero-ampli-tude test of the cosinor procedure. The rhythm characteristics estimatedby this method include the mesor (middle value of the fitted cosine repre-senting a rhythm-adjusted mean), amplitude (half the difference betweenthe minimum and maximum of the fitted cosine function), and acrophase(the time of the peak value expressed in h and min from local 00:00 h)(Table 2).
RESULTS
Survival, Somatic Growth, and Plasma Levels of Melatonin andCortisol
There was no mortality in either of the groups during the 6-wkexperimental period, and no significant differences in fork length andbody weight between the fish under LD (27.9± 0.3 cm, 249± 7.3 g) andLL (27.5± 0.2 cm, 227± 6.1 g) at the end of the experiment. Under LDconditions, plasma melatonin levels were typically low during the lightphase and gradually increased during the dark phase (cosinor, p< .01)(Figure 1) in contrast with no significant difference at any timepoint inmelatonin levels under LL (ANOVA, p = .55). There was no significant
Clock Gene Expression in Postsmolts 1721
Chr
onob
iol I
nt D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y pe
ter
ruof
f on
10/
29/1
0Fo
r pe
rson
al u
se o
nly.
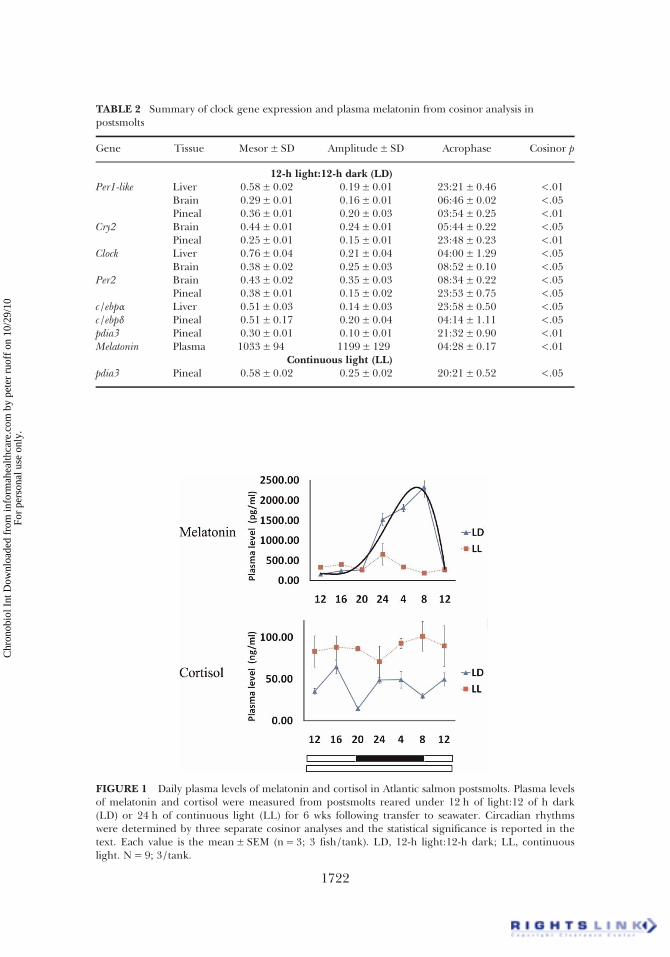
TABLE 2 Summary of clock gene expression and plasma melatonin from cosinor analysis inpostsmolts
Gene Tissue Mesor± SD Amplitude± SD Acrophase Cosinor p
12-h light:12-h dark (LD)Per1-like Liver 0.58± 0.02 0.19± 0.01 23:21± 0.46 <.01
Brain 0.29± 0.01 0.16± 0.01 06:46± 0.02 <.05Pineal 0.36± 0.01 0.20± 0.03 03:54± 0.25 <.01
Cry2 Brain 0.44± 0.01 0.24± 0.01 05:44± 0.22 <.05Pineal 0.25± 0.01 0.15± 0.01 23:48± 0.23 <.01
Clock Liver 0.76± 0.04 0.21± 0.04 04:00± 1.29 <.05Brain 0.38± 0.02 0.25± 0.03 08:52± 0.10 <.05
Per2 Brain 0.43± 0.02 0.35± 0.03 08:34± 0.22 <.05Pineal 0.38± 0.01 0.15± 0.02 23:53± 0.75 <.05
c/ebpα Liver 0.51± 0.03 0.14± 0.03 23:58± 0.50 <.05c/ebpδ Pineal 0.51± 0.17 0.20± 0.04 04:14± 1.11 <.05pdia3 Pineal 0.30± 0.01 0.10± 0.01 21:32± 0.90 <.01Melatonin Plasma 1033± 94 1199± 129 04:28± 0.17 <.01
Continuous light (LL)pdia3 Pineal 0.58± 0.02 0.25± 0.02 20:21± 0.52 <.05
FIGURE 1 Daily plasma levels of melatonin and cortisol in Atlantic salmon postsmolts. Plasma levelsof melatonin and cortisol were measured from postsmolts reared under 12 h of light:12 of h dark(LD) or 24 h of continuous light (LL) for 6 wks following transfer to seawater. Circadian rhythmswere determined by three separate cosinor analyses and the statistical significance is reported in thetext. Each value is the mean± SEM (n= 3; 3 fish/tank). LD, 12-h light:12-h dark; LL, continuouslight. N= 9; 3/tank.
1722
Chr
onob
iol I
nt D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y pe
ter
ruof
f on
10/
29/1
0Fo
r pe
rson
al u
se o
nly.

daily variation in plasma cortisol in fish reared under LL (ANOVA, p =.86) or LD (ANOVA, p < .01; cosinor, p = .97), although the level wastwo-fold higher under LL compared to LD (ANOVA, p < .001)(Figure 1).
Variations of Daily Clock Gene Expression in Salmon Postsmolts
Under LD, there was significant daily variation of Per1-like in thepineal gland, brain, and liver; of Cry2 and Per2 in the pineal gland andbrain, and of Clock in the brain and liver (Figure 2). The mesor andamplitude were similar in the pineal gland and brain for Per2, Per1-like,and Cry2, and their acrophases, respectively, occurred in the pineal gland2 h before (23:53 h), 2 h after (03:54 h), and 2 h before (23:48 h) themiddle of the dark period (02:00 h), respectively, whereas their
FIGURE 2 Daily expression of clock gene mRNAs in the pineal gland, liver, and brain of Atlanticsalmon postsmolts. Expression of clock gene mRNAs in the pineal gland, brain, and liver harvestedfrom postsmolts reared under 12 h of light:12 h of dark (LD) or 24 h of continuous light (LL) for 6wks following transfer to seawater and normalized to three endogenous control genes, ARP, β-actin,and EF1AB. Circadian rhythms were determined in three separate cosinor analyses, and the statisticalsignificance is shown in Table 2. Each value is the mean± SEM (n= 3; 3 fish/tank). LD, 12-hlight:12-h dark; LL, continuous light; n= 9, 3/tank.
Clock Gene Expression in Postsmolts 1723
Chr
onob
iol I
nt D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y pe
ter
ruof
f on
10/
29/1
0Fo
r pe
rson
al u
se o
nly.

acrophases in the brain for Per1-like and Cry2 occurred late in the darkperiod at 06:46 and 05:44 h, respectively (Table 2). In contrast, the acro-phase of brain Per2 and Clock was early in the light phase, at 08:34 and08:52 h, respectively (Table 2). The acrophases of liver Per1-like and Clockoccurred 2.5 h before (23:21 h) and 2 h (04:00 h) after the middle of thedark period, respectively (Table 2). Of the studied genes and organs, thepineal and liver Per1-like and the pineal Cry2 demonstrated the most pro-nounced oscillation ( p < .01; Table 2), and only pineal Per1-like demon-strated a strong response after the shift between the light and dark spans(marked increase between 20:00 and 24:00 h; Figure 2).
Under LL, none of the clock RNAs showed significant daily variationsin any of the analyzed tissues, but the overall levels of the Clock and Per2increased in the pineal gland and decreased in the liver of LL-reared fish(Figure 2).
Rhythmic Expression of Nuclear Transcription Factor and ERStress-Related Genes
The mRNA expression of c/ebpα and c/ebpδ displayed significant dailyvariations in the liver and pineal gland, respectively, with acrophases 2 hbefore (23:58 h) and 2.2 h (04:14 h) after the middle of the dark period(Figure 3, Table 2). The pparα expression did not show significantrhythms in either the pineal gland or liver. There were no rhythms ofthese mRNAs in fish under LL, but mRNA levels of the liver pparα wererepressed by LL and those of the pineal pparα, c/ebpα, and c/ebpδ wereup-regulated by LL when compared to those under LD. The pdia3expression showed significant daily variations in the pineal gland of fishreared under both LD and LL, but their respective acrophases weredifferent under the two light conditions (Figure 3, Table 2).
DISCUSSION
The present study is the first to demonstrate the effect of light ondaily variations in the expression of genes encoding for clock proteins,nuclear transcription factors, and ER stress protein in seawater postsmoltsof Atlantic salmon. We observed daily variations of plasma melatoninlevels under LD, which were elevated in the dark phase and suppressedin the light phase. This nocturnal rise in plasma melatonin disappearedunder LL. These results are in accordance with reports on the diurnalrhythms of plasma melatonin levels in salmonids (Iigo et al., 1997;Randall et al., 1995). Although significant daily variations of cortisollevels were not seen in postsmolts (LD), lower levels occurred during thetransition between the day and night spans, which was in contrast withthe nocturnal increase in plasma cortisol reported in smolts under
T.-s. Huang et al.1724
Chr
onob
iol I
nt D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y pe
ter
ruof
f on
10/
29/1
0Fo
r pe
rson
al u
se o
nly.

natural winter photoperiods (Ebbesson et al., 2008). Some studies havereported daily variations of cortisol with different acrophases in fish, suchas rainbow trout (Polakof et al., 2007), goldfish (Singley & Chavin, 1975),carp (Kuhn et al., 1986), eel (Li & Takei, 2003), and sea bass (Cerda-Reverter et al., 1998), which may be due to different light regimes andspecies. LL induced overall higher plasma cortisol levels compared toLD. In rainbow trout, glucocorticoid receptors are present in the pineal
FIGURE 3 Daily expression of nuclear transcription factor and pdia3 mRNAs in the pineal glandand liver of Atlantic salmon postsmolts. Expression of PPAR, C/EBP, and pdia3 mRNAs in the pinealgland and liver harvested from postsmolts reared under 12 h of light:12 h of dark (LD) or 24 h ofcontinuous light (LL) for 6 wks following transfer to seawater and normalized to three endogenouscontrol genes, ARP, β-actin, and EF1AB. Circadian rhythms were determined in three separatecosinor analyses and the statistical significance is shown in Table 2. Each value is the mean± SEM(n= 3; 3 fish/tank). LD, 12-h light:12-h dark; LL, continuous light; n= 9, 3/tank.
Clock Gene Expression in Postsmolts 1725
Chr
onob
iol I
nt D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y pe
ter
ruof
f on
10/
29/1
0Fo
r pe
rson
al u
se o
nly.

gland and melatonin production is inhibited by administration of the syn-thetic glucocorticoid dexamethasone (Benyassi et al., 2001). Similarresults were found in mammals, in which adrenocorticotropin-stimulatedcortisol was inhibited by the administration of melatonin (Torres-Farfanet al., 2003). Moreover, maternal melatonin suppression increases cortisolproduction in the newborn rat (Torres-Farfan et al., 2004). Therefore,there may be an inverse relationship between melatonin and cortisol inAtlantic salmon under LL, which needs to be studied in more detail.Cortisol is known to be involved in the endocrine response to stress(Boujard & Leatherland, 1992). In the present study, however, it is unli-kely that the cortisol levels were a result of stress during the capture pro-cedure, since they did not reveal high levels in both LD and LL. Inteleost, it is known that cortisol increases salinity tolerance through itseffect on mineral corticoids and ion transport proteins (Kiilerich et al.,2007; McCormick et al., 2008). Thus, cortisol levels appear to be influ-enced by light, although the biological function of the constant increaseof levels induced by LL is as yet unknown in postsmolts.
Here, the rhythmic expression of the pineal and liver Per1-like peakedat midnight, which is in accordance with reports in goldfish retina Per1(Velarde et al., 2009). The peak of brain Per1-like that occurred at dawnwas similar to that of Per1b (previously referred to as Per4) in the goldenrabbitfish brain (Park et al., 2007). In zebrafish, rhythmic expression ofPer1a (previously referred to Per1), Per1b, Per2, and Per3—with differenttiming of their maximal expression—was reported in an in vitro embryosystem, in which those of Per1a, Per1b, and Per3 were activated beforelight-onset (Pando et al., 2001; Vallone et al., 2004). In mice, three Pergenes—Per1, Per2, and Per3—are rhythmically expressed with differentlight responses in the SCN, but only Per1 and Per2 have been identifiedas important elements of the feedback loops (Takumi et al., 1998a,1998b; Zylka et al., 1998). We have described the expression of Atlanticsalmon Per1-like gene in parr and smolts (Huang et al., 2010b), in whichthe putative amino acid sequence derived from a cDNA fragment of Per1-like showed greatest homology with golden rabbitfish PER1b (Park et al.,2007). Since the full sequence data were not obtained, it is necessary toobtain a large sequence in order to clarify the identity of Per1 expression.
Similar to light-dependent Per2 expression in zebrafish (Delaunayet al., 2000; Ziv et al., 2005), golden rabbitfish (Sugama et al., 2008), andgoldfish (Velarde et al., 2009), brain Per2 mRNAs oscillated under LDwith a peak of expression at 08:34 h, which corresponds to the time oflight-onset. However, the peak expression of pineal Per2 occurred in thedark phase, as described in parr brain Per2 under a short-day photo-period (SD) by Davie et al. (2009). It is likely that Per2 expression in thewhole brain does not occur in parallel with that in the pineal gland,which does not contain a functional circadian clock as in other teleosts
T.-s. Huang et al.1726
Chr
onob
iol I
nt D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y pe
ter
ruof
f on
10/
29/1
0Fo
r pe
rson
al u
se o
nly.

(Falcon et al., 2010). We have unpublished data on pituitary clock geneexpression in smolts showing that light-dependent Per2 expression is anti-phasic to Per1-like; this suggests that other brain areas contribute tointernal circadian oscillators and that the regulation of Per2 is different insalmonids compared to other teleosts (Sugama et al., 2008; Velarde et al.,2009; Ziv et al., 2005). Although we cannot explain why Per2 is light-induced in the pineal gland but not in the liver and brain under LL inthe present study, this finding can provide important information aboutthe clock gene system in salmonids.
Pineal Cry2 peaked at midnight and brain Cry2 peaked at dawn,which is similar to the nocturnal expression of Cry2 in goldfish retina(Velarde et al., 2009), Atlantic salmon parr brain (Davie et al., 2009), andzebrafish larva (Lahiri et al., 2005). The collection of these resultssuggests that Atlantic salmon Per1-like and Cry2 mRNAs are expressedwith daily variations in the brain and peripheral tissues of postsmolts.The peak expression of brain Clock was in the beginning of the lightphase (08:52 h), and in liver Clock it was in the late night phase; in con-trast, Atlantic salmon parr Clock peaked at midnight under SD (Davieet al., 2009), whereas it peaked during the light phase in mouse SCN(Abe et al., 1999). These variations of the peak time may imply that thereis a different response of Clock expression in various organs, species, andunder different photoperiods. Given that Clock expression is not rhythmicin the pineal gland under LD, we suggest that Per1-like and Cry2expression is not dependent on the rhythmic expression of Clock in thepineal gland, although it is unknown whether this organ acquires specificregulation of clock gene expression in postsmolts. The mechanisms regu-lating gene expression in the rat pineal gland are complex, because genetranscription in this organ is controlled by both the central clock in theSCN and the circadian clocks present in the pineal gland (Abe et al.,2002; Fukuhara et al., 2005). In contrast to light up-regulated Clock anti-phasic to dark up-regulated Per1-like and Cry2 in the pineal gland of parrunder LD (Huang et al., 2010b), the brain transcriptional repressors andactivators peaked before and after onset of daylight in postsomlts, respect-ively. Although it is uncertain if clock gene RNA levels can be comparedbetween parr and postsmolts, the differences of expression under LDmay be due to developmental changes. The acrophases of brain Per1-like,Per2, and Cry2 occurred a few hours later than those of the pineal genesand brain Per2, and Clock peaked in the beginning of the light phase,suggesting that a set of genes that peaked during the day or later at nightmay be regulated by pathways that are indirectly activated by melatoninor perhaps by a “master clock” located within a SCN‐like structure in thebrain. Yet, in salmonids, pineal melatonin is directly regulated by lightbut not by a circadian clock (Iigo et al., 2007), and salmonids may haveevolve a decentralized system in the pineal gland, independently of the
Clock Gene Expression in Postsmolts 1727
Chr
onob
iol I
nt D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y pe
ter
ruof
f on
10/
29/1
0Fo
r pe
rson
al u
se o
nly.

eyes (Migaud et al., 2007). There is to date no evidence for the presenceof a functional circadian clock either in the pineal gland or retina,although they express clock genes in salmonids. Thus, further studiesshould clarify the role of the clock genes in the growth and stress axisand their expression in the hypothalamus.
The expression of c/ebpα and c/ebpδ exhibited daily variations in theliver and pineal gland, respectively, consistent with observations of rhyth-mic mRNA expression of c/ebp in the eye of the mollusc (Hattar et al.,2002), c/ebpβ in murine epididymal adipose tissue (Bray & Young, 2007),and c/ebpδ in the hamster pineal gland (Maronde et al., 2007). It hasbeen shown that C/EBP is involved in the regulation of arylalkylamineN-acetyltransferase (AANAT) mRNA in response to cyclic adenosinemonophosphate (cAMP) activation via the cAMP-response element-CCAAT complex in the promoter region (Baler et al., 1997) and that arole of adrenergic/cAMP signal transduction in regulating rhythmic geneexpression has been studied in rat pineal function (Bailey et al., 2009).These reports will provide a foundation for studies of the function of thesalmon c/ebpδ in the pineal gland. In a report by Bozek et al. (2009), C/EBP is a suggestive candidate for circadian transcriptional regulators in alarge-scale analysis of the CCG promoters, and circadian clock genes havebeen identified as targets of C/EBP in mouse NIH 3T3 cells (Gery et al.,2005). Thus, the rhythmic expression of salmon c/ebpα, c/ebpδ, and Per1-like supports the idea of a link between C/EBP proteins and the clockgene system. In addition, we found that expression of liver pparα mRNAshowed a dual peak, one in the evening and the other at dawn, whosevalues were statistically significant different from other timepoint valuesby ANOVA, but not by 24-h waveform cosinor analysis. This may bebecause the variable we are measuring varies over time, but not in a circa-dian manner, as not all biological rhythms are circadian. Targets of liverPPARα are involved in cellular lipid catabolism (Yamazaki et al., 2002),and the daily variation in the expression of lipogenic and cholesterogenicgenes were found to be attenuated or abolished in PPARα-null mice(Patel et al., 2001). As described in mammals (Jump, 2004), Atlanticsalmon PPAR and C/EBP have been suggested to be fatty acid–regulatedtranscriptional factors that play important roles in lipid metabolism inhepatocytes, adipocytes, and myotubes (Huang et al., 2010a; Klevelandet al., 2006; Vegusdal et al., 2004). PPARα and C/EBPα may act as poss-ible mediators for the daily rhythms of lipid metabolism in the liver,although this requires further study. Different sets of clock-controlledgenes across tissues were observed in the study of cycling transcripts inmice (Panda et al., 2002; Storch et al., 2002), suggesting liver c/ebpα,pparα, and pineal c/ebpδare putative clock-controlled genes in Atlanticsalmon. PPARs are expressed in the rat brain, of which the specificlocation, degree of expression, and physiological roles vary among the
T.-s. Huang et al.1728
Chr
onob
iol I
nt D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y pe
ter
ruof
f on
10/
29/1
0Fo
r pe
rson
al u
se o
nly.

receptors (Ramanan et al., 2010). Consequently, it is important to furtherelucidate the role of individual nuclear transcription factors when inter-acting with tissue-specific ligands to perform to their own functioncoupled with clock gene expression in the pineal gland and liver.
The expression of C/EBP, PPAR, and clock genes did not show sig-nificant daily variations in the studied tissues under LL, with the excep-tion of pineal pdia3. The mRNA expression of pineal Clock, Per2, pparα,c/ebpα, and c/ebpδ increased under LL, whereas the liver Clock, Per2, andpparα expression decreased compared to under LD. Nonetheless, theincreasing or decreasing clock mRNAs were not observed in the brain,indicating a different role of clock genes across tissues. Although pinealPer2, c/ebpα, and c/ebpδ did not express at high levels in the studiedorgans during the day under LD, the increase of expression was observedunder LL in the pineal gland, but not in the liver. It is unknown whythere were opposite effects of LL on the expression of Per2, Clock, andpparα in the pineal gland and liver, and future studies should addressthis. These results will provide insight into clock gene system in salmonidsand provide considerable clues as to what is happening under LL con-ditions. Contrary to a lack of daily variations in the plasma levels of mela-tonin and cortisol and the expression clock genes, continuous light canenhance growth and muscle fiber recruitment (Johnston et al., 2003),affect mineralization, and delay osteoid incorporation in Atlantic salmonpostsmolts (Wargelius et al., 2009). Similar results have reported that con-tinuous light exposure affects the growth rate, timing of smoltification,and reproduction (Krakenes et al., 1991; Saunders & Henderson, 1988;Stefansson et al., 1991). Light and feeding effects on the daily rhythmshave been reported in clock genes of zebrafish (Per1 and Cry) by Sanchezand Sanchez-Vazquez (2009) and humoral daily rhythms of sea bream byLopez-Olmeda et al. (2009). In our experiments, LL-reared fish wereexposed to continuous light but 12 h cycles of feeding and starvationeach day and showed no variation in daily clock gene expression, indicat-ing that light has a stronger effect on clock gene expression than feedingin salmon. However, the connection between the earlier observed effectsof LL on growth and development and a disrupted rhythm in theexpression of clock genes imposed by LL as observed here remainselusive.
The most interesting effect of light was observed on pdia3 expression.The pdia3 mRNA appears to be rhythmically expressed in the pinealgland under both LD and LL, but the acrophases differed. Multiple func-tions of PDIA3 are known, i.e., to catalyze the oxidation, reduction, andisomerization of intra- and intermolecular disulfide bond formation in ER(Khanal & Nemere, 2007). We previously demonstrated that liver-expressed pdia3 and c/ebpδ may be important biomarkers for oxidant- andtoxicant-induced stress in smolts (Huang et al., 2009b). Although
Clock Gene Expression in Postsmolts 1729
Chr
onob
iol I
nt D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y pe
ter
ruof
f on
10/
29/1
0Fo
r pe
rson
al u
se o
nly.

arrhythmic in the liver, the daily expression of pdia3 and c/ebpδ was onlypresent in the pineal gland under LD. Although it is unknown whetherthe significance of pineal pdia3 and c/ebpδ transcripts is related to physio-logical function, their expression may be coupled with clock geneexpression under LD in Atlantic salmon. Whether the observation thatpdia3 expression is not correlated with the arrhythmic expression of clockgenes under LL is related to the idea that an endogenous growth rhythmexists in Atlantic salmon under LL (Oppedal et al., 2006) remainsunknown. This connection and the importance of the rhythmic expressionof pineal pdia3 under both LD and LL need to be studied in more detail.
The present study indicates that light exerts an influence on the dailyvariation and overall expression of clock and nuclear transcription factorgenes in the pineal gland, brain, and liver of Atlantic salmon postsmolts,as well as levels of melatonin and cortisol. It is possible that light-dependent variations of expression reported here reflect interplaybetween daily and annual regulation. These changes can promote newapproaches for achieving a better understanding of how the clock genesystem is associated with the environmental, physiological, and geneticinteractions that occur in lifecycle transitions and their signaling pathwaybetween the brain and periphery. On the other hand, the integration ofcontinuous light for the timing of events, such as smoltification andsexual maturation, and its effect on the seasonal growth pattern, need tobe studied in more detail.
ACKNOWLEDGMENTS
We wish to thank Eva Mykkeltvedt, Jacob Wessels, and LeiknyFjeldstad for their excellent analytical work at NIFES and Dr. BenteE. Torstensen for reading through the manuscript. This work was sup-ported by the NIFES Internal Clock.
Declaration of Interest: The authors report no conflicts of interest.The authors alone are responsible for the content and writing of thepaper.
REFERENCES
Abe H, Honma S, Namihira M, Tanahashi Y, Ikeda M, Yu W, Honma K. (1999). Phase-dependentinduction by light of rat Clock gene expression in the suprachiasmatic nucleus. Brain Res. Mol.Brain Res. 66:104–110.
Abe M, Herzog ED, Yamazaki S, Straume M, Tei H, Sakaki Y, Menaker M, Block GD. (2002).Circadian rhythms in isolated brain regions. J Neurosci. 22:350–356.
Bailey MJ, Coon SL, Carter DA, Humphries A, Kim JS, Shi Q, Gaildrat P, Morin F, Ganguly S,Hogenesch JB, Weller JL, Rath MF, Moller M, Baler R, Sugden D, Rangel ZG, Munson PJ,Klein DC. (2009). Night/day changes in pineal expression of >600 genes: central role of adre-nergic/cAMP signaling. J. Biol. Chem. 284:7606–7622.
T.-s. Huang et al.1730
Chr
onob
iol I
nt D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y pe
ter
ruof
f on
10/
29/1
0Fo
r pe
rson
al u
se o
nly.

Baler R, Covington S, Klein DC. (1997). The rat arylalkylamine N-acetyltransferase gene promoter—cAMP activation via a cAMP-responsive element-CCAAT complex. J. Biol. Chem. 272:6979–6985.
Balsalobre A. (2002). Clock genes in mammalian peripheral tissues. Cell Tissue Res. 309:193–199.Balsalobre A, Damiola F, Schibler U. (1998). A serum shock induces circadian gene expression in
mammalian tissue culture cells. Cell 93:929–937.Benyassi A, Schwartz C, Ducouret B, Falcon J. (2001). Glucocorticoid receptors and serotonin
N-acetyltransferase activity in the fish pineal organ. NeuroReport 12:889–892.Boujard T, Leatherland JF. (1992). Circadian-rhythms and feeding time in fishes. Environ. Biol. Fish.
35:109–131.Bozek K, Relogio A, Kielbasa SM, Heine M, Dame C, Kramer A, Herzel H. (2009). Regulation of
clock-controlled genes in mammals. PLoS One 4:pe4882.Bray MS, Young ME. (2007). Circadian rhythms in the development of obesity: potential role for the
circadian clock within the adipocyte. Obes. Rev. 8:169–181.Cerda-Reverter JM, Zanuy S, Carrillo M, Madrid JA. (1998). Time-course studies on plasma glucose,
insulin, and cortisol in sea bass (Dicentrarchus labrax) held under different photoperiodic regimes.Physiol. Behav. 64:245–250.
Dardente H, Cermakian N. (2007). Molecular circadian rhythms in central and peripheral clocks inmammals. Chronobiol. Int. 24:195–213.
Davie A, Minghetti M, Migaud H. (2009). Seasonal variations in clock-gene expression in Atlanticsalmon (Salmo salar). Chronobiol. Int. 26:379–395.
Delaunay F, Thisse C, Marchand O, Laudet V, Thisse B (2000). An inherited functional circadianclock in zebrafish embryos. Science 289:297–300.
Duez H, Staels B. (2008). Rev-erb alpha gives a time cue to metabolism. FEBS Lett. 582:19–25.Duguay D, Cermakian N. (2009). The crosstalk between physiology and circadian clock proteins.
Chronobiol. Int. 26:1479–1513.Dunlap JC. (1999). Molecular bases for circadian clocks. Cell 96:271–290.Ebbesson LO, Bjornsson BT, Ekstrom P, Stefansson SO. (2008). Daily endocrine profiles in parr and
smolt Atlantic salmon. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 151:698–704.Falcon J. (1999). Cellular circadian clocks in the pineal. Prog. Neurobiol. 58:121–162.Falcon J, Migaud H, Munoz-Cueto JA, Carrillo M. (2010). Current knowledge on the melatonin
system in teleost fish. Gen. Comp. Endocrinol. 165:469–482.Fjelldal PG, Nordgarden U, Berg A, Grotmol S, Totland GK, Wargelius A, Hansen T. (2005).
Vertebrae of the trunk and tail display different growth rates in response to photoperiod inAtlantic salmon, Salmo salar L., post-smolts. Aquaculture 250:516–524.
Fukuhara C, Yamazaki S, Liang J. (2005). Pineal circadian clocks gate arylalkylamine N-acetyltrans-ferase gene expression in the mouse pineal gland. J. Neurochem. 93:156–162.
Gauger MA, Sancar A. (2005). Cryptochrome, circadian cycle, cell cycle checkpoints, and cancer.Cancer Res. 65:6828–34.
Gery S, Gombart AF, Yi WS, Koeffler C, Hofmann WK, Koeffler HP. (2005). Transcription profilingof C/EBP targets identifies Per2 as a gene implicated in myeloid leukemia. Blood106:2827–2836.
Hattar S, Lyons LC, Dryer L, Eskin A. (2002). Circadian regulation of a transcription factor, Ap C/EBP, in the eye of Aplysia californica. J. Neurochem. 83:1401–1411.
Hirayama J, Kaneko M, Cardone L, Cahill G, Sassone-Corsi P. (2005). Analysis of circadian rhythmsin zebrafish. Methods Enzymol. 393:186–204.
Huang T-S, Grodeland G, Sleire L, Wang MY, Kvalheim G, Laerum OD. (2009a). Induction of circa-dian rhythm in cultured human mesenchymal stem cells by serum shock and cAMP analogs invitro. Chronobiol. Int. 26:242–257
Huang T-S, Olsvik PA, Krovel A, Tung HS, Torstensen BE. (2009b). Stress-induced expression ofprotein disulfide isomerase associated 3 (PDIA3) in Atlantic salmon (Salmo salar L.). Comp.Biochem. Physiol. B Biochem. Mol. Biol. 154:435–442.
Huang T-S, Todorcevic M, Ruyter B, Torstensen BE. (2010a). Altered expression of CCAAT/enhan-cer binding protein (C/EBP) and FABP11 genes during ddipogenesis in vitro in Atlantic salmon(Salmo salar) Aquaculture Nutr. 16:72–80.
Huang T-S, Ruoff P, Fjelldal PG. (2010b). Diurnal expression of clock genes in pineal gland andbrain and plasma levels of melatonin and cortisol in Atlantic salmon parr and smolts. Chronobiol.Int. In press.
Clock Gene Expression in Postsmolts 1731
Chr
onob
iol I
nt D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y pe
ter
ruof
f on
10/
29/1
0Fo
r pe
rson
al u
se o
nly.

Iigo M, Hara M, Ohtani-Kaneko R, Hirata K, Tabata M, Aida K. (1997). Photic and circadian regu-lations of melatonin rhythms in fishes. Biol. Signals 6:225–232.
Iigo M, Abe T, Kambayashi S, Oikawa K, Masuda T, Mizusawa K, Kitamura S, Azuma T, Takagi Y,Aida K, Yanagisawa T. (2007). Lack of circadian regulation of in vitro melatonin release fromthe pineal organ of salmonid teleosts. Gen. Comp. Endocrinol. 154:91–97.
Johnston IA, Manthri S, Smart A, Campbell P, Nickell D, Alderson R. (2003). Plasticity of musclefibre number in seawater stages of Atlantic salmon in response to photoperiod manipulation.J. Exp. Biol. 206:3425–3435.
Jump DB. (2004). Fatty acid regulation of gene transcription. Crit. Rev. Clin. Lab. Sci. 41:41–78.Khanal RC, Nemere I. (2007). The ERp57/GRp58/1,25D(3)-MARRS receptor: multiple functional
roles in diverse cell systems. Curr. Med. Chem. 14:1087–1093.Kiilerich P, Kristiansen K, Madsen SS. (2007). Cortisol regulation of ion transporter mRNA in
Atlantic salmon gill and the effect of salinity on the signaling pathway. J. Endocrinol.194:417–427.
Kleveland EJ, Ruyter B, Vegusdal A, Sundvold H, Berge RK, Gjoen T. (2006). Effects of 3-thia fattyacids on expression of some lipid related genes in Atlantic salmon (Salmo salar L.). Comp.Biochem. Physiol. B Biochem. Mol. Biol. 145:239–248.
Krakenes R, Hansen T, Stefansson SO, Taranger GL. (1991). Continuous light increases growth-rateof Atlantic salmon (Salmo-Salar L) postsmolts in sea cages. Aquaculture 95:281–287.
Kuhn ER, Corneille S, Ollevier F. (1986). Circadian variations in plasma osmolality, electrolytes,glucose, and cortisol in carp (Cyprinus carpio). Gen. Comp. Endocrinol. 61:459–468.
Lahiri K, Vallone D, Gondi SB, Santoriello C, Dickmeis T, Foulkes NS. (2005). Temperature regu-lates transcription in the zebrafish circadian clock. PLoS Biol 3:e351.
Leder EH, Danzmann RG, Ferguson MM. (2006). The candidate gene, Clock, localizes to a strongspawning time quantitative trait locus region in rainbow trout. J. Hered. 97:74–80.
Lemberger T, Saladin R, Vazquez M, Assimacopoulos F, Staels B, Desvergne B, Wahli W, Auwerx J.(1996). Expression of the peroxisome proliferator-activated receptor alpha gene is stimulated bystress and follows a diurnal rhythm. J. Biol. Chem. 271:1764–1769.
Li Y, Takei Y. (2003). Ambient salinity-dependent effects of homologous natriuretic peptides (ANP,VNP, and CNP) on plasma cortisol level in the eel. Gen. Comp. Endocrinol. 130:317–323.
Lopez-Olmeda JF, Montoya A, Oliveira C, Sanchez-Vazquez FJ. (2009). Synchronization to light andrestricted-feeding schedules of behavioral and humoral daily rhythms in Gilthead Sea Bream(Sparus aurata). Chronobiol. Int. 26:1389–1408.
Maronde E, Pfeer M, Glass Y, Sthele JH. (2007). Transcription factor dynamics in pineal gland andliver of the Syrian hamster (Mesocricetus auratus) adapts to prevailing photoperiod. J. Pineal Res.43:16–24.
McCormick SD. (2001). Endocrine control of osmoregulation in teleost fish. Am. Zool. 41:781–794.McCormick SD, Regish A, O’Dea MF, Shrimpton JM. (2008). Are we missing a mineralocorticoid in
teleost fish? Effects of cortisol, deoxycorticosterone and aldosterone on osmoregulation, gillNa+,K+-ATPase activity and isoform mRNA levels in Atlantic salmon. Gen. Comp. Endocrinol.157:35–40.
Mendez-Ferrer S, Lucas D, Battista M, Frenette PS. (2008). Haematopoietic stem cell release is regu-lated by circadian oscillations. Nature 452:442–447.
Migaud H, Davie A, Chavez CCM, Al-Khamees S. (2007). Evidence for differential photic regulationof pineal melatonin synthesis in teleosts. J. Pineal Res. 43:327–335.
Nordgarden U, Oppedal F, Taranger GL, Hemre GI, Hansen T. (2003a). Seasonally changingmetabolism in Atlantic salmon (Salmo salar L.) I—growth and feed conversion ratio. AquacultureNutr. 9:287–293.
Nordgarden U, Torstensen BE, Froyland L, Hansen T, Hemre GI. (2003b). Seasonally changingmetabolism in Atlantic salmon (Salmo salar L.) II—beta-oxidation capacity and fatty acid compo-sition in muscle tissues and plasma lipoproteins. Aquaculture Nutr. 9:295–303.
Nordgarden U, Hansen T, Hemre GI, Sundby A, Bjornsson BT. (2005). Endocrine growth regu-lation of adult Atlantic salmon in seawater: the effects of light regime on plasma growthhormone, insulin-like growth factor-I, and insulin levels. Aquaculture 250:862–871.
Oishi K, Shirai H, Ishida N. (2005). CLOCK is involved in the circadian transactivation of peroxi-some-proliferator-activated receptor alpha (PPAR alpha) in mice. Biochem. J. 386:575–581.
T.-s. Huang et al.1732
Chr
onob
iol I
nt D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y pe
ter
ruof
f on
10/
29/1
0Fo
r pe
rson
al u
se o
nly.

Oppedal F, Juell JE, Taranger GL, Hansen T. (2001). Artificial light and season affects vertical distri-bution and swimming behaviour of post-smolt Atlantic salmon in sea cages. J. Fish Biol.58:1570–1584.
Oppedal F, Berg A, Olsen RE, Taranger GL, Hansen T. (2006). Photoperiod in seawater influenceseasonal growth and chemical composition in autumn sea-transferred Atlantic salmon (Salmosalar L.) given two vaccines. Aquaculture 254:396–410.
Panda S, Antoch MP, Miller BH, Su AI, Schook AB, Straume M, Schultz PG, Kay SA, Takahashi JS,Hogenesch JB. (2002). Coordinated transcription of key pathways in the mouse by the circadianclock. Cell 109:307–320.
Pando MP, Pinchak AB, Cermakian N, Sassone-Corsi P. (2001). A cell-based system that recapitulatesthe dynamic light-dependent regulation of the vertebrate clock. Proc. Natl. Acad. Sci. U. S. A.98:10178–10183.
Park JG, Park YJ, Sugama N, Kim SJ, Takemura A. (2007). Molecular cloning and daily variations ofthe Period gene in a reef fish Siganus guttatus. J Comp. Physiol. A Neuroethol. Sens. Neural Behav.Physiol. 193:403–411.
Patel DD, Knight BL, Wiggins D, Humphreys SM, Gibbons GF. (2001). Disturbances in the normalregulation of SREBP-sensitive genes in PPAR alpha-deficient mice. J. Lipid Res. 42:328–337.
Polakof S, Ceinos RM, Fernandez-Durán B, Mıguez JM, Soengas JL. (2007). Daily changes in par-ameters of energy metabolism in brain of rainbow trout: dependence of feeding. Comp. Biochem.Physiol. A 146:265–273.
Porter MJR, Duncana NJ, Mitchellb D, Bromage NR. (1999). The use of cage lighting to reduceplasma melatonin in Atlantic salmon (Salmo salar) and its effects on the inhibition of grilsing.Aquaculture 176:237–244.
Portaluppi F, Touitou Y, Smolensky MH. (2008). Ethical and methodological standards for laboratoryand medical biological rhythm research. Chronobiol. Int. 25:999–1016.
Ralph MR, Foster RG, Davis FC, Menaker M. (1990). Transplanted suprachiasmatic nucleus deter-mines circadian period. Science 247:975–978.
Ramanan S, Zhao W, Riddle DR, Robbins ME. (2010). Role of PPARs in Radiation-Induced BrainInjury. PPAR Res. 2010:234975.
Randall CF, Bromage NR, Thorpe JE, Miles MS, Muir JS. (1995). Melatonin rhythms in Atlanticsalmon (Salmo salar) maintained under natural and out-of-phase photoperiods. Gen. Comp.Endocrinol. 98:73–86.
Rousseau K, Atcha Z, Loudon AS. (2003). Leptin and seasonal mammals. J. Neuroendocrinol.15:409–414.
Sanchez JA, Sanchez-Vazquez FJ. (2009) Feeding entrainment of daily rhythms of locomotor activityand clock gene expression in zebrafish brain. Chronobiol. Int. 26:1120–1135.
Saunders RL, Henderson EB. (1988). Effects of constant day length on sexual-maturation andgrowth of Atlantic salmon (Salmo-Salar) parr. Can. J. Fish. Aquat. Sci. 45:60–64.
Singley JA, Chavin W. (1975). Serum cortisol in normal goldfish (Carassius auratus L.). Comp. Biochem.Physiol. 50A:77–82.
Stefansson SO, Naevdal G, Hansen T. (1989). The influence of 3 unchanging photoperiods ongrowth and parr-smolt transformation in Atlantic salmon, Salmo-Salar L. J. Fish Biol.35:237–247.
Stefansson SO, Bjornsson BT, Hansen T, Haux C, Taranger GL, Saunders RL. (1991). Growth, parr-smolt transformation, and changes in growth-hormone of Atlantic salmon (Salmo-Salar) rearedunder different photoperiods. Can. J. Fish. Aquat. Sci. 48:2100–2108.
Storch KF, Lipan O, Leykin I, Viswanathan N, Davis FC, Wong WH, Weitz CJ. (2002). Extensive anddivergent circadian gene expression in liver and heart. Nature 417:78–83.
Sugama N, Park JG, Park YJ, Takeuchi Y, Kim SJ, Takemura A. (2008). Moonlight affects nocturnalPeriod2 transcript levels in the pineal gland of the reef fish Siganus guttatus. J. Pineal Res.45:133–141.
Takumi T, Matsubara C, Shigeyoshi Y, Taguchi K, Yagita K, Maebayashi Y, Sakakida Y, Okumura K,Takashima N, Okamura H. (1998a). A new mammalian period gene predominantly expressedin the suprachiasmatic nucleus. Genes Cells 3:167–176.
Takumi T, Taguchi K, Miyake S, Sakakida Y, Takashima N, Matsubara C, Maebayashi Y, OkumuraK, Takekida S, Yamamoto S, Yagita K, Yan L, Young MW, Okamura H. (1998b). A light-independent oscillatory gene mPer3 in mouse SCN and OVLT. EMBO J. 17:4753–4759.
Clock Gene Expression in Postsmolts 1733
Chr
onob
iol I
nt D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y pe
ter
ruof
f on
10/
29/1
0Fo
r pe
rson
al u
se o
nly.

Taranger GL, Haux C, Stefansson SO, Bjornsson BT, Walther BT, Hansen T. (1998). Abruptchanges in photoperiod affect age at maturity, timing of ovulation and plasma testosterone andoestradiol-17 beta profiles in Atlantic salmon, Salmo salar. Aquaculture 162:85–98.
Thorpe JE, McConway MG, Miles MS, Muir JS. (1987). Diel and seasonal changes in resting plasmacortisol levels in juvenile Atlantic salmon, Salmo salar L. Gen. Comp. Endocrinol. 65:19–22.
Torres-Farfan C, Richter HG, Rojas-Garcia P, Vergara M, Forcelledo ML, Valladares LE, Torrealba F,Valenzuela GJ, Seron-Ferre M. (2003). mt1 Melatonin receptor in the primate adrenal gland:inhibition of adrenocorticotropin-stimulated cortisol production by melatonin. J. Clin. Endocrinol.Metab. 88:450–458.
Torres-Farfan C, Richter HG, Germain AM, Valenzuela GJ, Campino C, Rojas-Garcia P, ForcelledoML, Torrealba F, Seron-Ferre M. (2004). Maternal melatonin selectively inhibits cortisol pro-duction in the primate fetal adrenal gland. J. Physiol. 554:841–856.
Vallone D, Gondi SB, Whitmore D, Foulkes NS. (2004). E-box function in a period gene repressedby light. Proc. Natl. Acad. Sci. U. S. A. 101:4106–4111.
Vandesompele J, De Preter K, Pattyn F, Poppe B, Van Roy N, De Paepe A, Speleman F. (2002).Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of mul-tiple internal control genes. Genome Biol. 3:RESEARCH0034.
Vegusdal A, Ostbye TK, Tran TN, Gjoen T, Ruyter B. (2004). Beta-oxidation, esterification, andsecretion of radiolabeled fatty acids in cultivated Atlantic salmon skeletal muscle cells. Lipids39:649–658.
Velarde E, Haque R, Iuvone PM, Azpeleta C, Alonso-Gomez AL, Delgado MJ. (2009). Circadianclock genes of goldfish, Carassius auratus: cDNA cloning and rhythmic expression of period andcryptochrome transcripts in retina, liver, and gut. J. Biol. Rhythms 24:104–113.
Wargelius A, Fjelldal PG, Nordgarden U, Hansen T. (2009). Continuous light affects mineralizationand delays osteoid incorporation in vertebral bone of Atlantic salmon (Salmo salar L.). J. Exp.Biol. 212:656–661.
Yamazaki K, Kuromitsu J, Tanaka I. (2002). Microarray analysis of gene expression changes inmouse liver induced by peroxisome proliferator activated receptor alpha agonists. Biochem.Biophys. Res. Commun. 290:1114–1122.
Ziv L, Levkovitz S, Toyama R, Falcon J, Gothilf Y. (2005). Functional development of the zebrafishpineal gland: light-induced expression of period2 is required for onset of the circadian clock.J. Neuroendocrinol. 17:314–320.
Zylka MJ, Shearman LP, Weaver DR, Reppert SM. (1998). Three period homologs in mammals:differential light responses in the suprachiasmatic circadian clock and oscillating transcriptsoutside of brain. Neuron 20:1103–1110.
T.-s. Huang et al.1734
Chr
onob
iol I
nt D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y pe
ter
ruof
f on
10/
29/1
0Fo
r pe
rson
al u
se o
nly.