down-regulation of scatter factor in mrc 5 fibroblasts by...
TRANSCRIPT

Journal of Cell Science 101, 323-332 (1992)Printed in Great Britain © The Company of Biologists Limited 1992
323
Down-regulation of scatter factor in MRC 5 fibroblasts by epithelial-derived
cells
A model for scatter factor modulation
TAHEREH KAMALATI, BIRUNTHI THIRUNAVUKARASU, ANDREW WALLACE,
NIGEL HOLDER, ROBERT BROOKSAnatomy and Human Biology Croup, Biomedical Sciences Division, King's College London, Strand, London WC2R 2LS, UK
TOSHIKAZU NAKAMURADepartment of Biology, Faculty of Science, Kyusyu University, Fukuoka 812, Japan
MICHAEL STOKER, ERMANNO GHERARDIImperial Cancer Research Fund and Department of Pathology, University of Cambridge, Cambridge CB2 1QP, UK
and LAKJAYA BULUWELADepartment of Biochemistry, Charing Cross and Westminster Medical School, Fulham Palace Road, London W6 8RF, UK
Summary
Scatter factor/hepatocyte growth factor (SF/HGF) is amultifunctional cytokine produced by embryonic fibro-blasts and other mesenchymal cells that affects thegrowth and/or the movement of certain epithelia. Herewe report that expression of scatter factor activity byMRC 5 cells, a strain of normal human embryonic lungfibroblasts, is greatly reduced as a result of co-culture ofthese cells with SVK14, an SV40-transformed humankeratinocyte cell line. Using a cDNA probe to the /Jchainof human HGF, we have found that the fall in SF activity
in MRC 5/SVK14 co-cultures is accompanied by the lossof SF/HGF transcripts. As the inhibition of SF activitycoincides with the disappearance of SF/HGF transcript,we conclude that inhibition of the SF activity expressedby MRC 5 cells by co-culture with SVK14 involvestranscriptlonal regulation.Key words: scatter factor (SF), human hepatocyte growthfactor (h-HGF), c-met, MRC 5, SVK14, motUity factor,growth factor, co-culture, epithelial cells, mesenchymalcells, cell-cell interactions.
Introduction
Epithelial-mesenchymal interactions are of great im-portance during development as they control inductiveevents and can involve the passage of instructiveinformation that controls the future differentiation ofone or both cell types. They may also be involved in thecontrol of specific alterations in cell behaviour, particu-larly in events such as de-epithelialisation and increasedcell motility. Such phenomena occur in major develop-mental processes such as gastrulation, somitic disper-sion of dermatome, myotome and sclerotome, andneural crest migration. Further, morphological andfunctional transitions from epithelia to mesenchyme arebelieved to occur during fusion of palatal shelves(Ferguson, 1988). Little is known about the control ofsuch dramatic alterations in cell behaviour in theembryo. There is, therefore, a great deal of interest in
the identification and characterisation of proteins thataffect de-epithelialisation and cell motility. Motilityfactors, characterised as cytokines with a significantstimulating effect on cell movement but no effect on cellgrowth, are one of a series of specific factors that canregulate cell movement (Warn and Dowrick, 1989).Until recently, scatter factor, SF, was considered alegitimate member of the "motility factor" group ofproteins. SF, a paracrine effector of mesenchymal-epithelial interactions, was originally identified byStoker and Perryman (1985) as an activity released intothe culture media by MRC 5 fibroblasts, which actsspecifically on epithelial cells by breaking their inter-cellular connections and increasing their mobility, thusresulting in "scattering" of cohesive epithelial coloniesinto isolated cells (Stoker et al. 1987). The assignmentof SF to the motility factor group of proteins was basedon the observation that SF did not appear to elicit a

324 T. Kamalati and others
mitogenic effect in Swiss 3T3 fibroblasts nor MDCKepithelial cells (Gherardi et al. 1989; Stoker andGherardi, 1989).
Purification and biochemical characterisation ofmouse (Gherardi et al. 1989), and human (Weidner etal. 1990) SF has shown it to be a 92 kDa glycoprotein,which is converted by limited proteolysis into twocovalently bonded subunits (mouse: 57 kDa and 30kDa; human: 62 kDa and 34 kDa). Sequence analysis(Gherardi and Stoker, 1990; Weidner et al. 1990),together with biological and immunological assays(Furlong et al. unpublished), has established that SF isidentical to a molecule independently characterised ashepatocyte growth factor, HGF (Gherardi and Stoker,1991; Weidner et al. 1991).
HGF has been cloned and sequenced, and shown tohave structural similarity to plasminogen and relatedserine proteases (Miyazawa et al. 1989; Nakamura et al.1989). Liver damage induced by chemical treatment orpartial hepatoectomy, has been used to show that HGFis synthesised and secreted by the non-parenchymalliver cells and stimulates the growth of parenchymalhepatocytes in a paracrine fashion (Kinoshita et al.1989; Noji et al. 1990). HGF has also been purified fromhuman lung fibroblast culture medium and shown to actas a mitogen for melanocytes, endothelial cells and avariety of epithelial cells including human and mousekeratinocytes (Matsumoto et al. 1991; Rubin et al.1991). HGF expression has been found in a variety oforgans (Kinoshita et al. 1989; Tashiro et al. 1990;Zarnegar et al. 1990; Seki et al. 1990; Rubin et al. 1991).These findings together with the evidence of HGFexpression in fibroblasts derived from adult skin (Rubinet al. 1991) suggest that HGF may act as a paracrinemediator of proliferation of a broad spectrum of celltargets.
In earlier studies of epithelial-mesenchymal interac-tions we have observed that co-culture of an SV40-transformed keratinocyte cell line, SVK14, with mesen-chymally derived co-culture partners, including MRC 5,results in the re-expression of markers characteristics ofdifferentiating, stratifying epithelia (Kamalati et al.1989). As MRC 5 is a "rich source" of SF in culture, wehave begun to investigate whether SVK14 cells can inturn modulate the expression of SF when co-culturedwith MRC 5. Here we report that co-culture of SVK14,with MRC 5 cells results in inhibition of SF expressionin MRC 5 cells and show that this inhibition involvestranscriptional regulation of SF/HGF.
These results are interesting in that they define aunique culture model for studying regulation ofSF/HGF expression. This may have a significantbearing on understanding of cell-cell interactions aswell as cell movement and positioning.
Materials and methods
CellsSV40-transformed human epidermal keratinocytes, SVK14(Taylor-Papadimitriou et al. 1982) were a gift from Dr J.
Taylor-Papadimitriou (ICRF, Lincolns Inn Fields, London,UK). Sources of MRC 5, human embryonic lung fibroblasts(Jacobs et al. 1970) and MDCK, Madin canine kidneyepithelial cells (Madin and Darby, 1958), Swiss 3T3/4A, clone5, mouse fibroblast (Brooks et al. 1984); BHK21, clone 13,baby hamster kidney fibroblasts (Macpherson and Stoker,1962); Cos, SV40-transformed African green monkey kidneycells (Gluzman, 1981); HeLa, human cervical carcinoma cells(Scherer et al. 1953); and A431, human squamous carcinomacells (Giard et al. 1973) were as described previously(Kamalati et al. 1989). SCC146, human squamous carcinomacells (Rupniak et al. 1985) were a gift from Dr T. Rupniak(Glaxo, Greenford, UK). Primary human keratinocytes werederived from newborn fore-skin.
Cell cultureSVK14
Stock cultures of SVK14 were grown in RPMI 1640supplemented with 10% foetal calf serum (FCS), 50 ng/mlcholera toxin, 5 /<g/ml hydrocortisone, and sodium hydrogencarbonate to give a final concentration of 3.7 g per liter.
MDCKStock cultures of MDCK-were grown in Dulbecco's modifiedEagle's medium (DMEM) containing 5% FCS.
Other cellsStock cultures of all others cells were grown in DMEMcontaining 10% FCS. All cells were grown at 37°C in ahumidified 12% CO2 atmosphere.
Co-culturesCo-cultures were initiated by seeding a well-mixed single cellsuspension containing 1.5 x 105 SVK14 cells together with 1.5x 105 MRC 5 or the cell type to be co-cultured, in 2 mlDMEM containing 10% FCS in a 3.5 cm dish. The controlswere composed of 1.5 x 105 MRC 5 cells seeded in 2 mlDMEM plus 10% FCS in a 3.5 cm dish. The co-cultures andtheir controls were left to condition their media for a period of7 days unless otherwise stated. The conditioned media werecentrifuged at 350 g for 5 min to remove cell debris and storedat 4°C until use.
Scatter factor stability in the presence of SVK14Samples (1 ml) of MRC 5 conditioned medium (7 days) wereexposed to a dense monolayer of SVK14 cells (1.3 x 105
cells/cm2) freshly seeded into 24-well dishes for a period oftime ranging from 0 to 72 h. At each time point a 1 ml samplewas removed and assayed for scatter factor activity.
Titration of MRC 5 and co-culture conditionedmediumMRC 5 conditioned medium (6 days) was mixed with SVK14conditioned medium (6 days) in varying ratios (0-100%) andthen assayed for SF activity. Likewise MRC 5 conditionedmedium (7 days) was mixed with MRC 5/SVK14 co-cultureconditioned medium (7 days) and assayed for SF activity. Ascontrols the respective MRC 5 conditioned media were mixedwith DMEM plus 10% FCS in the same ratios as used above.
MRC 5 conditioning of SVK14 conditioned mediumSamples (2 ml) of SVK14 conditioned medium (7 days) andDMEM plus 10% FCS were exposed to dense monolayers ofMRC 5 cells (1 x 104 cells/cm2) freshly seeded in 3.5 cm dishesfor a period of 7 days and then assayed for SF activity.

Down-regulation of scatter factor 325
Segregated co-culturesSegregated co-cultures were initiated by seeding 1.5 x 105 ofeach cell type independently on different areas of the culturedish in order to allow coexistence of the two cell types withvarying degrees of cell contact. The controls were composedof MRC 5 cells only, seeded in the same pattern as that oftheir co-culture partner. All cultures were seeded into 3.5 cmdishes containing 2 ml DMEM plus 10% FCS, and assayed forSF activity after 7 days of culture.
MRC 5/SVK14 co-cultures with varying SVK14 celldensitiesKeeping the MRC 5 cell numbers constant while varying thatof SVK14, co-cultures were initiated by seeding 1.5 X 105
MRC 5 cells with 1.5 x 104 to 1.5 x 105 SVK14 cells in 3.5 cmculture dishes with 2 ml DMEM + 10% FCS. The control wascomposed of 1.5 x 105 MRC 5 cells only. The cultures wereassayed for SF after 7 days.
Fixing SVK14 for co-culturesA total of 1.5 x 105 SVK14 cells were seeded into a 3.5 cmdish with 2 ml medium, left to settle for 24 hours and thenfixed with freshly made acetone:methanol (1:1, v/v) or 4%formaldehyde in phosphate buffered saline (PBS). The cellswere fixed by adding the fixative dropwise onto the cellswithout removing the culture medium, and left for 5 min. Themedium was then replaced with 2 ml fixative and the cells left(20 min for acetone:methanol and 10 min for 4% formal-dehyde) at room temperature. The fixed cells were thenwashed well in PBS followed by a final 5 min PBS wash, afterwhich 1.5 x 105 MRC 5 cells were seeded on top of the fixedSVK14. The controls composed of 1.5 x 105 MRC 5 cells ontheir own.
Mitomycin C treatmentA total of 1.5 x 105 SVK14 cells were mitomycin C treated 24hours post-seeding by adding 4 ng of mitomycin C permillilitre of medium. The cells were then incubated at 37°C for2 hours and washed with PBS. 1.5 x 105 MRC 5 cells werethen seeded on top of the mitomycin C-treated SVK14. MRC5/SVK14 co-cultures were likewise mitomycin C-treated 24hours post-seeding. MRC 5 control cultures were initiated bymitomycin C treatment of 1.5 x 105 MRC 5 cells only. Allcultures were seeded in 3.5 cm dishes with 2 ml DMEM plus10% FCS and assayed for SF activity after 7 days.
Scatter factor activity assayThis was carried out as described by Stoker and Perryman(1985). Briefly, using 96-well plates, twofold serial dilutions oftest samples were incubated with 3000 MDCK cells at 37°C for24 hours. The plates were then fixed with 4% formaldehyde inPBS, stained with 0.2% Coomassie blue (15 min) followed by1% crystal violet (60 min) and each well was assessed forscattering. The highest dilution at which scattering could beobserved was recorded as the end point of the assay. Since theassay is performed in serial dilutions of 2, the results arepresented as Iog2, i.e. SF activity at a dilution of 1/32 ispresented as (lofo l/32=)-5.
FACScan analysisThe fluorescein diacetate succinimidyl ester, 5- (and 6)-carboxyfluorescein diacetate succinimidyl ester (CFSE) (Mol-ecular Probes, USA) was used to label MRC 5 cellsspecifically. CFSE is a non-fluorescent ester that diffusesfreely across the cell membrane into the cell, where non-specific intracellular esterases remove the acetate group,
releasing the succinimidyl ester chromophore. The negativelycharged chromophore then reacts with protein amines and isretained within the cell for considerable periods of time(Bronner-Fraser, 1985). In our hands CSFE at 0.8 uM doesnot effect the viability or the metabolic activity of the MRC 5cells, nor is it passed on to neighbouring cells by leakage.Thus MRC 5 cells were incubated in phosphate bufferedsaline (PBS) containing 0.8 pM CSFE at 37°C for 30 min. Thecells were then washed twice in PBS, resuspended in DMEM+ 10% FCS and co-cultured with SVK14 as above. Thenumber of fluorescent MRC 5 cells present in co-culture wasthen determined by subjecting the CSFE-MRC 5/SVK14 co-cultures to FACScan analysis (Beckton and Dickinson) 24, 48and 72 hours post co-culture initiation (Flow cytometry).
Northern analysis of total RNARNA extraction, Northern blotting and hybridisation pro-cedures were performed as described previously (Boehm etal. 1988). Northern filters were probed with a 0.7 kb cDNAfragment covering the /5 chain region of the h-HGF (humanHGF) mRNA (Nakamura et al. 1989), washed to a stringencyof 5xSSC + 0.1% SDS at 65°C and autoradiographed for 5days using X-OMATfilm. The presence of fibroblast-derivedRNA in co-cultures was demonstrated by rehybridisation ofNorthern blots to a 1 kb cDNA fragment for the humanvimentin gene derived from clone L3A7A (Ferraris et al.1986); 25 jig total RNA from each sample was used unlessotherwise stated.
Results
Scatter factor expression in MRC 5/SVK14 co-culturesSF activity released into the culture medium by MRC 5cells appears to be greatly reduced on co-culture ofMRC 5 with SVK14 cells, where the co-cultures areinitiated as well mixed suspensions of both cell types(Fig. 1). SVK14 cells do not produce SF nor do theyrespond to the activity, in that they continue to remainin their epithelial cell colonies in the presence of thefactor (data not shown). Further, SVK14 cells do notdegrade or remove SF from the culture medium.Stability of SF in the presence of SVK14 cells is
MRC 5/SVK14co-culture
Fig. 1. Inhibition of scatter factor activity released by MRC5 cells in co-culture with SVK14 cells. (H) Scatter factoractivity in MRC 5/SVK14 co-cultures; and (•) MRC 5controls after 7 days of culture. As the assay is performedin twofold serial dilutions the scatter factor activity ispresented as Iog2 (Materials and methods). Bars representthe standard deviation, with n=l2.

326 T. Kamalati and others
demonstrated in Fig. 2A, where MRC 5 conditionedmedium with a high SF titer, was exposed to a densemonolayer of SVK14 cells for a period of up to 72 hourswithout any subsequent fall in SF activity. In order toestablish whether the reduced SF activity in the MRC5/SVK14 co-cultures was due to some factor(s) releasedinto the culture medium by SVK14, which effectivelyremoves SF from the co-culture medium, we mixedMRC 5 conditioned medium with varying proportionsof SVK14 conditioned medium. The observed fall in SFactivity (Fig. 2B) was due entirely to dilution, andidentical to the DMEM control, showing that SVK14 donot release products that remove SF from the culture
media. Similarly, to determine whether the co-cultureitself results in release of some product(s), which in turnremove SF from the culture media, the co-cultureconditioned medium was mixed with MRC 5 con-ditioned medium. The results presented in Fig. 2Cdemonstrate that the co-culture of SVK14 cells withMRC 5 per se does not result in release of an inhibitor ofSF. Further, MRC 5 cells are able to condition SVK14conditioned medium to precisely the same degree asfresh DMEM + 10% FCS (Fig. 2D), illustrating thatthe greatly reduced SF activity resulting from their co-culture is not due to SVK14 cells producing a factor thatinhibits the release of SF by the MRC 5 cells.
9
B
7 _
6 .
1
A
9
8
7-
0 20 40 60
1 Time (mln)
40
Time(h)60 80 100 80 60 40 20
% MRC 5 conditioned medium
100 80 60 40 20% MRC 5 conditioned medium
Fig. 2. Stability of scatter factor in the presence of SVK14 cells. A. Stability of scatter factor (MRC 5 conditioned medium;7 days) in the presence of a dense monolayer of SVK14 cells, over an initial period of 60 min (• ) , and the following 72 h(D). B. Titration of MRC 5 condition medium (6 days) against SVK14 conditioned medium (7 days) (•) and DMEM +10% FCS (•) . C. Titration of MRC 5 conditioned medium (7 days) against MRC 5/SVK14 co-culture conditioned medium(7 days) ( • ) , and DMEM + 10% FCS (S). D. Release of scatter factor by MRC 5 cells into SVK14 conditioned medium(7 days) (•), and fresh DMEM + 10% FCS (•) over a period of 7 days.
Down-regulation of scatter factor 327
Extensive cell-cell association is a requirement for theinhibition of scatter factor activity in MRC 5/SVK14co-culturesSegregated co-cultures of MRC 5/SVK14, where thetwo cell types are seeded independently and ondifferent areas of the same dish, do not exhibit aninhibition in scatter activity, unlike the co-cultures of awell-mixed suspension of both cell types. This isirrespective of whether the two cell types do notestablish cell-cell contact or whether they are allowed asmall degree of contact, Fig. 3. This experimentdemonstrates that inhibition of expression of scatteractivity requires more than merely having the two celltypes present in the same co-culture dish, suggesting arequirement for intimate cell-cell associations.
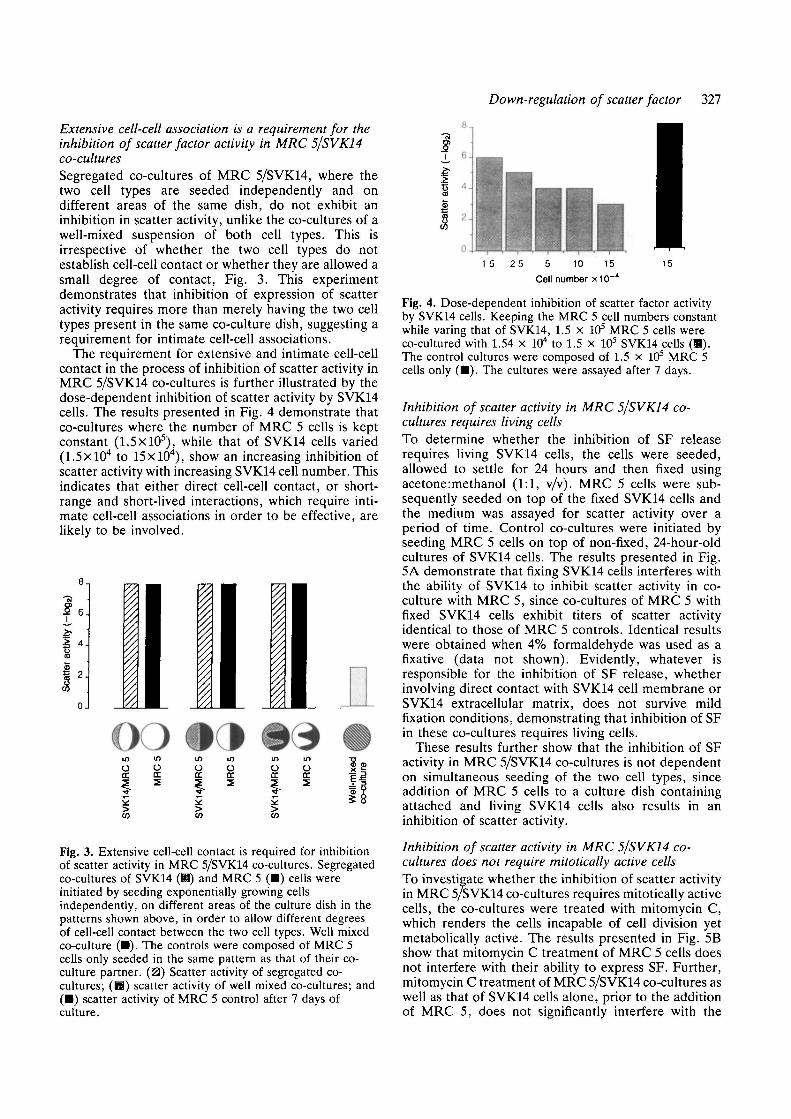
The requirement for extensive and intimate cell-cellcontact in the process of inhibition of scatter activity inMRC 5/SVK14 co-cultures is further illustrated by thedose-dependent inhibition of scatter activity by SVK14cells. The results presented in Fig. 4 demonstrate thatco-cultures where the number of MRC 5 cells is keptconstant (1.5X105), while that of SVK14 cells varied(l.SxlO4 to 15xlO4), show an increasing inhibition ofscatter activity with increasing SVK14 cell number. Thisindicates that either direct cell-cell contact, or short-range and short-lived interactions, which require inti-mate cell-cell associations in order to be effective, arelikely to be involved.
I 6 .
4 .
inODC
§CO
inOoc2
inO<rfiCO
inODC2
inOCE
f̂
VK
1
CO
inOcc2
Fig. 3. Extensive cell-cell contact is required for inhibitionof scatter activity in MRC 5/SVK14 co-cultures. Segregatedco-cultures of SVK14 (H) and MRC 5 (•) cells wereinitiated by seeding exponentially growing cellsindependently, on different areas of the culture dish in thepatterns shown above, in order to allow different degreesof cell-cell contact between the two cell types. Well mixedco-culture (•). The controls were composed of MRC 5cells only seeded in the same pattern as that of their co-culture partner. (0) Scatter activity of segregated co-cultures; (B) scatter activity of well mixed co-cultures; and(•) scatter activity of MRC 5 control after 7 days ofculture.
15 25 5 10 15Cell number x10
Fig. 4. Dose-dependent inhibition of scatter factor activityby SVK14 cells. Keeping the MRC 5 cell numbers constantwhile varing that of SVK14, 1.5 X 105 MRC 5 cells wereco-cultured with 1.54 x 104 to 1.5 x 105 SVK14 cells (B).The control cultures were composed of 1.5 x 10s MRC 5cells only (•) . The cultures were assayed after 7 days.
Inhibition of scatter activity in MRC 5/SVK14 co-cultures requires living cellsTo determine whether the inhibition of SF releaserequires living SVK14 cells, the cells were seeded,allowed to settle for 24 hours and then fixed usingacetone :methanol (1:1, v/v). MRC 5 cells were sub-sequently seeded on top of the fixed SVK14 cells andthe medium was assayed for scatter activity over aperiod of time. Control co-cultures were initiated byseeding MRC 5 cells on top of non-fixed, 24-hour-oldcultures of SVK14 cells. The results presented in Fig.5A demonstrate that fixing SVK14 cells interferes withthe ability of SVK14 to inhibit scatter activity in co-culture with MRC 5, since co-cultures of MRC 5 withfixed SVK14 cells exhibit titers of scatter activityidentical to those of MRC 5 controls. Identical resultswere obtained when 4% formaldehyde was used as afixative (data not shown). Evidently, whatever isresponsible for the inhibition of SF release, whetherinvolving direct contact with SVK14 cell membrane orSVK14 extracellular matrix, does not survive mildfixation conditions, demonstrating that inhibition of SFin these co-cultures requires living cells.
These results further show that the inhibition of SFactivity in MRC 5/SVK14 co-cultures is not dependenton simultaneous seeding of the two cell types, sinceaddition of MRC 5 cells to a culture dish containingattached and living SVK14 cells also results in aninhibition of scatter activity.
Inhibition of scatter activity in MRC 5/SVK14 co-cultures does not require mitotically active cellsTo investigate whether the inhibition of scatter activityin MRC 5/SVK14 co-cultures requires mitotically activecells, the co-cultures were treated with mitomycin C,which renders the cells incapable of cell division yetmetabolically active. The results presented in Fig. 5Bshow that mitomycin C treatment of MRC 5 cells doesnot interfere with their ability to express SF. Further,mitomycin C treatment of MRC 5/SVK14 co-cultures aswell as that of SVK14 cells alone, prior to the additionof MRC 5, does not significantly interfere with the

328 T. Kamalati and others
A
-a
1 2 3 4 7SVK14Not fixed
1 2 3 4 7 Time (days)MRC5Control
10-,
§ 8-
£> 6-|
I 4H
I 2- IFig. 5. Requirement of living but not mitotically activeSCK14 for inhibition of scatter factor activity in MRC5/SVK14 co-cultures. A. Effect of fixing SVK14 on theirability to inhibit scatter factor activity; (H) scatter factoractivity in co-cultures of MRC 5 cells with fixed(acetone imethanol) SVK14 in comparison to co-culturewith living SVK14; (•) MRC 5 control. B. Effect ofmitomycin C treatment of SVK14 on their ability to inhibitscatter factor activity; (ffl) scatter factor activity in co-cultures of MRC 5 cells with mitomycin C-treated SVK14(+) in comparison to non-mitomycin C-treated SVK14 (—)after 7 days in culture; (•) scatter factor activity inmitomycin C-treated MRC 5/SVK14 co-cultures (+) incomparison to non-mitomycin C-treated co-cultures ( -)after 7 days in culture; (•) scatter factor activity inmitomycin C-treated (+) and non-mitomycin C treated (—)MRC 5 controls after 7 days in culture.
capacity of SVK14 to cause inhibition of the scatteractivity expressed by MRC 5 cells [-l(iog2) and ~3(iog2)are equivalent to dilutions of 1/2 and 1/8, respectively,in comparison to -9(iog,), which is equivalent to adilution factor of 1/512] (Fig. 5B). These resultsdemonstrate that inhibition of scatter activity in MRC5/SVK14 co-cultures does not require mitotically activecells. Furthermore, these results demonstrate that lackof scatter activity in the culture media of MRC 5/SVK14co-cultures is not merely due to preferential prolifer-ation of the SVK14 over the MRC 5 and the subsequentloss of the MRC 5 cells.
Inhibition of expression of MRC 5 scatter factor is notspecific to SVK14 cellsIn order to determine whether the capacity to inhibitexpression of SF activity by MRC 5 is unique to SVK14cells, we co-cultured MRC 5 cells with two fibroblastic
cell lines and a variety of epithelial cells includingnormal human keratinocytes. The results presented inFig. 6 demonstrate that the two fibroblastic cell linestested do not appear to inhibit the expression of scatteractivity by MRC 5 cells (Fig. 6A). However, normalhuman keratinocytes as well as four other epithelial celllines tested appear to be able to inhibit the aboveactivity by more than 90%, when co-cultured with MRC5 cells (Fig. 6B). These results are interesting in thatthey demonstrate the regulation of expression of amesenchymal activity by epithelial cells.
SVK14 cells inhibit expression of scatter factortranscript in MRC 5We have shown that co-culture of SVK14 with MRC 5cells results in an inhibition of SF activity released byMRC 5. In order to determine at which level thisregulation is achieved total RNA was prepared fromMRC 5/SVK14 co-cultures and probed for SF/HGFmessage using a DNA probe to h-HGF. This probe waschosen in the light of sequence identity demonstrated
1 0 , A
Time (days)
Fig. 6. Scatter factor activity in co-cultures of MRC 5 cellswith a variety of mesenchymally derived and epithelial cellstrains. (A) Scatter factor activity in co-cultures of MRC 5with two fibroblast cell lines (H), in comparison to MRC 5controls (•) after 7 days in culture. (B) Scatter factoractivity in co-cultures of MRC 5 with a variety of epithelialcell lines (H) in comparison to MRC 5 control cultures(•) , after 7 days in culture. All co-cultures were initiatedas in MRC 5/SVK14 co-cultures (Materials and methods).

between the SF and h-HGF (Gherardi and Stoker,1990,1991; Weidner et al. 1990,1991). The results showthat this probe detects a 6 kb transcript in total RNAprepared from MRC 5 cells (Fig. 7). This is inagreement with the size of the HGF transcript ident-ified in human liver (Nakamura et al. 1989), placenta(Miyazawa et al. 1989) and fibroblasts (Rubin et al.1991).
In order to ascertain whether the SF transcript fromthe MRC 5 cells, equivalent in number to those presentin the co-culture during the initial 3 days, can bedetected using this probe, RNA concentrations equival-ent to the relative numbers of MRC 5 cells present inthe co-culture for each day were also probed alongsidethat of the co-culture. The number of MRC 5 cellspresent in the co-cultures were estimated using FACS-can analysis; 8.9, 7.4 and 4.8 jug RNA thereforerepresent the RNA equivalent of MRC 5 cells presentin the co-cultures on days 1, 2 and 3, respectively. Theresults show that the probe used here can indeed detecta signal from cell numbers equivalent to those of MRC5 cells present in co-culture over the first 3 days of co-culture (Fig. 7A). From the Northern blot presented inFig. 7A it is evident that SVK14 cells do not produceany scatter transcript, in agreement with their inability
Down-regulation of scatter factor 329
to release SF shown above. Further, it appears that thescatter factor transcript is absent from MRC 5/SVK14co-cultures as early as 24 hours after culture initiationand the following 48 hours (Fig. 7A). The presence ofMRC 5-derived RNA in the co-cultures is confirmed bydetection of the transcript for vimentin (a cytoskeletalprotein expressed by fibroblasts but not SVK14) (Fig.7B). The disappearance of this 6 kb message in MRC5/SVK14 co-cultures correlates with the greatly reducedSF activity observed in these co-cultures (Figs 5A, 6B).These results show that the inhibition of SF activity inMRC 5 cells involves transcriptional regulation of theSF message in these co-cultures.
Discussion
Inhibition of SF activity in MRC 5 cellsWe have found that the level of SF activity released intothe culture media by MRC 5 is greatly reduced by co-culture with SVK14. This is not due to active degra-dation of SF by SVK14 cells or to generation of solublefactors, by either SVK14 or the co-cultures, whicheffectively remove scatter activity from the culturemedia or prevent release of the factor by MRC 5 cells in
B *
2
MR
Cco
-cul
inOrr2
rs 2 2
cvj 9 CO 8.9
7.4
4.8
2inc\j
9 kb
7.5 kb
4.4 kb
sz• * •
CVJ
MR
C
no-oo 1
szCVJ
SV
K14
— 9 kb
— 7.5 kb
— 4.4 kb
— 2.4 kb
— 1.4 kb
Fig. 7. Expression of SF/HGF in MRC 5/SVK14 co-cultures. Northern blots: (A) total RNA extracted from MRC 5/SVK14co-cultures, SVK14 and MRC 5 cells (25 fig) was probed with the h-HGF (30 kDa) probe. 8.9, 7.4 and 4.8 fig RNArepresent the RNA equivalent of MRC 5 cells present in co-cultures on days 1, 2 and 3, respectively. The 4.2 kDa band isribosomal RNA. (B) The first four tracks of the same blot rehybridised to a 1 kb cDNA fragment for the human vimentingene.

330 T. Kamalati and others
co-culture. We have observed that the inhibition of SFactivity in MRC 5/SVK14 co-cultures has a requirementfor intimate cell-cell associations established betweenliving, but not mitotically active cells. This suggests thatshort-range and short-lived interactions are involved,possibly involving direct cell-cell contact.
Using FACScan analysis we have found that althoughSVK14 proliferated more rapidly than MRC 5 cells inco-culture, MRC 5 cells are not dislocated from the co-cultures, at least over the initial 3 days. The observedinhibition of SF activity in MRC 5/SVK14 co-cultures,therefore, is not simply due to rapid proliferation of theSVK14 over that of the MRC 5 cells. This conclusion isreinforced by the observation that mitomycin C-treatedco-cultures exhibit an inhibition of SF activity almostidentical to that seen in non-mitomycin C-treated co-cultures.
Since SF and HGF are identical we have used a DNAprobe to the 30 kDa sub-unit of h-HGF to detect the SFtranscript in MRC 5 cells. We have found that thisprobe detects a 6 kb message in total RNA preparedfrom MRC 5 cells. This is in agreement with themessage size of h-HGF identified in human liver(Nakamura et al. 1989), placenta (Miyazawa et al. 1989)and lung fibroblasts (Rubin et al. 1991). Further, wehave found that the 6 kb transcript detected by thisprobe is absent from MRC 5/SVK14 co-cultures overthe initial 72 hours of culture. As the inhibition of SFactivity in MRC 5/SVK14 co-cultures correlates with thedisappearance of the 6 kb transcript, these data demon-strate that the inhibition of SF activity in MRC 5/SVK.14co-cultures involves transcriptional regulation.
Regulation of HGF/SFHGF is a potent hepatotrophic factor that triggers liverregeneration after partial hepatectomy and liver injuryand was first purified and characterised from the sera ofpartially hepatectomised rats (Nakamura et al. 1984).Subsequently HGF was detected (Russel et al. 1984)and purified (Nakamura et al. 1987) from rat platelets.Purification of HGF from serum of human (Goda et al.1988) and rabbit (Zarnegar et al. 1988) has also beenreported. Derman et al. (1981) have demonstrated thatwhen mouse liver cells are disaggregated and placed inculture overnight, synthesis of hepatocyte-specificRNAs decline substantially, while synthesis of actin andtubulin RNAs remains unchanged. However, liverslices cultured under the same conditions maintain anear-normal liver-specific transcription pattern at leastfor 12-14 hours. It therefore appears that even in "fullydifferentiated" hepatocytes specific cell-cell or cell-matrix interactions are necessary to maintain theirspecialised transcription pattern. Biochemical and bio-logical characterisation of HGF have shown HGF to bea paracrine stimulator of proliferation of matureparenchymal hepatocytes, synthesised and secreted bythe non-parenchymal liver cells (Kinoshita et al. 1989;Noji et al. 1990). Further, it has been shown that HGFmRNA expression in the non-parenchymal liver cells isinduced by liver damage, and that this inductioncorrelates well with the degree of damage (Kinoshita et
al. 1989; Noji et al. 1990). It thus appears that theexpression of mHGF in the non-parenchymal cells ofliver is down-regulated by the presence of parenchymalhepatocytes and up-regulated by their absence. In thiscontext the down-regulation of expression of SF/HGFtranscript in MRC 5 observed as a result of co-culture ofthese fibroblasts with SVK14 cells, which are epithelialin origin, is analogous to the mode of control of mHGFexpression in liver. In this light, characterisation of theinteractions of SVK14 with MRC 5, and identificationof the components involved in regulation of SF/HGFtranscript may provide an insight into the regulation ofexpression of SF/HGF in vivo. Such regulatory cell-cellinteractions are important in early development wheregene control signals requiring direct contact betweencells are a prerequisite for embryonic determination ofmost specialised cells. An example of this is thedevelopment of liver, lungs and pancreas, which isdependent on mesodermal-endodermal interactions.
The capacity to inhibit SF activity of MRC 5 cellsdoes not appear to be restricted to SVK14 cells. Normalhuman keratinocytes, from which SVK14 cells wereoriginally derived, also exhibit an ability to inhibit SFactivity, when co-cultured with MRC 5 cells. Recentwork has demonstrated that (i) HGF is mitogenic forkeratinocytes (epithelial cells of skin) and melanocytes(pigment producing cells of skin) (Rubin et al. 1991;Matsumoto et al. 1991), (ii) HGF is expressed byfibroblasts isolated from skin (Rubin et al. 1991) andthat (iii) SF can induce MDCK and a sub-group ofhuman carcinoma cells to invade collagen gels (Weidneret al. 1990). In view of the identity between SF andHGF, our data together with those of others mentionedabove, indicate that SF/HGF may play a role in normaldevelopment of skin as well as metastasis of epithelialtumours. It is also interesting that the four transformedepithelial cell lines tested in co-culture with MRC 5 alsoexhibit an ability to inhibit scatter activity, while thefibroblasts tested do not. This raises the interestingpossibility that the capacity to inhibit SF activity ofMRC 5 cells is specific to epithelial cells. However,more cell types must be tested in order to verify thispossibility conclusively.
The expression of HGF/SF (Kinoshita et al. 1989;Rubin et al. 1991; Tashiro et al. 1990; Zarnegar et al.1990) together with that of its cellular receptor, the c-met proto-oncogene (Naldini et al. 1991; Bottaro et al.1991; Chan et al. 1988), has been demonstrated in avariety of tissues. However, very little is known aboutthe control of regulation of expression of this factor. Inthis context the use of MRC 5/SVK14 co-culture as amodel for studies of identification of factors that areimportant in regulating SF expression, together withfurther characterisation of the interactions of SVK14with MRC 5 cells, may provide insight into the generalfeatures that govern the control of epithelial-mesenchy-mal interactions during development and differen-tiation.
The authors thank Mrs M. Perryman for much help, Mr M.Howard for excellent technical assistance and critical dis-

Down-regulation of scatter factor 331
cussion, Dr R. Dover and Ms E. Gale for critical discussion,Mr D. Davies for FACscan analysis and Ms S. Murphy forexcellent technical advice. The human vimentin probe wasmade available by the UK Human Genome Mapping Projectthrough Mr A. King. T.K. and N.H. thank the SERC forfinancial support. L.B. is supported by the MRC.
References
Boehm, T., Buluwcla, L., Williams, D., White, L. and Rabbitts, T. H.(1988). A cluster of chromosome IIP 13 translocation found viadistinct D-D, D-D-J rearrangement of human T cell receptor <5chain gene. EMBO J. 7, 2011-2017.
Bottaro, D. P., Rubin, J. S., Faletto, D. L., Chan, A. M.-L., Kmledk,T. E., Vande Woude, G. F. and Aaronson, S. A. (1991).Identification of the hepatocyte growth factor receptor as the c-metproto-oncogene product. Science 251, 802-804.
Bronner-Fraser, M. (1985). Alterations in neural crest migration by amonoclonal antibody that affects cell adhesion. J. Cell Biol. 101,610-617.
Brooks, R. F., Richmond, F. N., Riddle, P. N. and Richmond, K. M.V. (1984). Apparent heterogeneity in the response of quiescentSwiss 3T3 cells to serum growth factors: Implications for thetransition probability model and parallels with "cellularsenescence" and "Competence". / . Cell. Physiol. 121, 341-350.
Chan, A. M.-L., King, H. W. S., Deakln, E. A., Tempest, P. R.,Hllkens, J., Kroezen, V., Edwards, D. R., Wills, A. J., Brookes, P.and Coope, C. S. (1988). Characterisation of the mouse met proto-oncogene. Oncogene 2, 593-599.
Derman, E., Krauter, K., Walling, C , Weinberger, C , Ray, M. andDarnell, J. E. Jr (1981). Transcriptional control in the productionof liver specific mRNAs. Cell 23, 731-739.
Ferguson, M. W. J. (1988). Palate development. Development 103,Supplement, 41-60.
Ferraris, S., Battlnl, R., Kaczmarek, L., Rlttllng, S., Calabretta, B.,de Rlel, J. K., Philipois, V., Wei, J.-F. and Baserga, R. (1986).Coding sequence and growth regulation of the human vimentingene. Mol. Cell. Biol. 6, 3614-3620.
Gherardi, E., Gray, J., Stoker, M., Perryman, M. and Furlong, R.(1989). Purification of scatter factor, a fibroblast derived basicprotein that modulates epithelial interaction and movement. Proc.Nat. Acad. Sci. USA. 86, 5844-5848.
Gherardi, E. and Stoker, M. (1990). Hepatocyte and scatter factor.Nature 346, 228.
Gherardi, E. and Stoker, M. (1991). Hepatocyte growth-factor:Mitogen, Motogen and Met. Cancer Cells 3, 227-232.
Glard, D. J., Aaronson, S. A., Todaro, G. J., Arrsteln, P. J. H.,Keresey, P. H., Dosik, P. and Parks, W. P. (1973). In vitrocultivation of human tumors: Establishment of cell lines derivedfrom a series of solid tumors. J. Nat. Cancer Inst. USA 51, 1417-1423.
Gluzman, Y. (1981). SV40 transformed simian cells support thereplication of early SV40 mutants. Cell 23, 175-182.
Goda, E., Tsubouchi, H., Nakayama, H. S., Hlrono, H., Sakiyama,O., Takahashi, K. H., Mlyakakl, H., Hashimoto, S. and Daikuhar,Y. (1988). Purification and partial characterisation of hepatocytegrowth factor from plasma of a patient with fulminant hepaticfailure. J. Clin. Invest. 81, 414-419.
Jacobs, J. P., Jones, C. M. and Bailie, J. P. (1970). Characteristics ofa human diploid cell line designated MRC-5. Nature 227,168-170.
Kamalati, T., Mclvor, Z., Howard, M., Green, M. R. and Brooks, R.F. (1989). Expression of markers of differentiation in a transformedhuman keratinocyte line induced by co-culture with a fibroblastline. Exp. Cell Res. 185, 453-463.
Klnoshlta, T., Tashlro, K. and Nakamura, T. (1989). Markedincrease of HGF mRNA in non-parenchymal liver cells of ratstreated with hepatotoxins. Biochem. Biophys. Res. Commun. 165,1229-1234.
Macpherson, I. A. and Stoker, M. G. P. (1962). Polyomatransformation of hamster cell clones- an investigation of geneticfactors affecting cell competence. Virology 16, 147-151.
Madln, S. H. and Darby, N. B. (1958). Established kidney cell lines of
normal adult bovine and ovine origin. Proc. Soc. Exp. Biol. Med.98, 574-576.
Matsumoto, K., Hashimoto, K., Yoshikawa, K. and Nakamura, T.(1991). Marked stimulation of growth and motility of humankeratinocytes by hepatocyte growth factor. Exp. Cell Res. 196,114-120.
Miyazawa, K., Tsubouchi, H., Naka, D., Takahashi, K., Okigaki, M.,Arakakl, N., Nakayama, H., Hlrono, S., Sakiyama, ()., Takahashi,K., Gohda, E., Daikuhara, Y. and Kltamura, N. (1989). Molecularcloning and sequence analysis of cDNA for human hepatocytegrowth factor. Biochem. Biophys. Res. Commun. 163, 967-973.
Nakamura, T., Nawa, K. and Ichlhara, A. (1984). Partial purificationand characterization of hepatocyte growth factor from serum ofhepatectomized rats. Biochem. Biophys. Res. Commun. 122, 1450-1459.
Nakamura, T., Nawa, K., Ichihara, A., Kalse, N. and Nishino, T.(1987). Purification and subunit structure of hepatocyte growthfactor from rat platelets. FEBS Lett. 224, 311-316.
Nakamura, T., Nishizawa, T., Hagiya, M., Sekl, T., Shlmoni, M.,Suglmura, A., Tashiro, K. and Shimlzu, S. (1989). Molecularcloning and expression of human hepatocyte growth factor. Nature342, 440-443.
Naldlni, L., Vigna, E., Narsimhan, R., Gaudlno, G., Zarnegar, R.,MIchalopoulos, G. K. and Comoglio, P. M. (1991). Hepatocytegrowth factor (HGF) stimulates the tyrosine kinase activity of thereceptor encoded by the proto-oncogene c-met. Oncogene 6, 501-504.
Noji, S., Tashlro, K., Koyama, E., Nohno, K., Taniguchi, S. andNakamura, T. (1990). Expression of hepatocyte growth factor genein endothelial and Kupffer cells of damaged rat liver, as revealed byin situ hybridisation. Biochem. Biophys. Res. Commun. 173,42^7.
Rubin, J. S., Chan, M.-L., Bottaro, D. P., Burgess, W. H., Taylor,W. G., Cech, A. C , Hirschfield, D. W., Wong, J., Miki, T., Finch,P. W. and Aaronson, S. A. (1991). A broad-spectrum human lungfibroblast-derived mitogen is a variant of hepatocyte growth factor.Proc. Nat. Acad. Sci. USA 88, 415-419.
Rupniak, H. T., Rowlatt, C , Lane, B. E., Steel, J. G., Trejdosiewicz,L. K., Laskiewicz, B., Povey, S. and Hill, B. T. (1985).Characterisation of four new human cell lines derived fromsquamous cell carcinoma of the head and neck. /. Nat. Cancer Inst.75, 621-635.
Russel, W. E., McGowan, J. A. and Bucher, N. L. R. (1984). Partialdcharacterization of a growth factor from rat platelets. / . Cell.Physiol. 119, 183-192.
Scherer, W. F., Syverton, J. T. and Gey, G. O. (1953). Studies on thepropagation in vitro of poliomyelitis viruses: IV. Viralmultiplication in a strain of human malignant epidermal cells (strainHeLa) derived from an epidermoid carcinoma of the cervix. /. Exp.Med. 97, 695-710.
Sekl, T., Ihara, I., Sugimura, A., Shimonishi, M., Nishizawa, T.,Asami, O., Hagiya, M., Nakamura, T. and Shimizu, S. (1990).Isolation and expression of cDNA for different forms of hepatocytegrowth factor from human leukocytes. Biochem. Biophys. Res.Commun. 172, 321-327.
Stoker, M. and Gherardi, E. (1989). Scatter factor and otherregulators of cell mobility. British Med. Bulletin 45, 481-491.
Stoker, M., Gherardi, E., Perryman, M. and Gray, J. (1987). Scatterfactor is a fibroblast-derived modulator of epithelial cell mobility.Nature 327, 239-242.
Stoker, M. and Perryman, M. (1985). An epithelial scatter factorreleased by embryo fibroblasts. J. Cell Sci. 77, 209-223.
Tashlro, K., Hagiya, M., Nishizawa, T., Sekl, T., Shlmomshi, M.,Shimizu, S. and Nakamura, T. (1990). Deduced primary structureof rat hepatocyte growth factor and expression of the mRNA in rattissue. Proc. Nat. Acad. Sci. USA 87, 3200-3204.
Taylor-Papadimitriou, J., Purkis, P., Lane, B. E., McKay, J. A. andChang, S. E. (1982). Effect of SV40 transformation on thecytoskeleton and behavioural properties of human keratinocytes.Cell Diff. 11, 169-180.
Warn, R. M. and Dowrick, P. (1989). Motility factors on the march.Nature 340, 186-187.
Weldner, K. M., Arakai, N., Vandekerchove, J., Weingari, S.,Hartmann, G., Rleder, H., Fonatsch, C , Tsubouchi, H., Hishida,

332 T. Kamalati and others
T., Daikuhara, Y. and Birchmeier, W. (1991). Evidence for theidendity of human scatter factor and human hepatocyte growthfactor. Proc. Nat. Acad. Sci. USA 88, 7001-7005.
Weldner, K. M., Behrens, J., Vandekerckhove, J. and Birchmeier,W. (1990). Scatter factor: molecular characterisation and effect onthe invasiveness of epithelial cells. / . Cell Biol. I l l , 2097-2108.
Zarnegar, R., Muga, S., Enghild, J. and Michalopoulos, G. (1988).NH2-terminal amino acid sequence for rabbit hepatopoietin A, a
heparin-binding polypeptide growth factor for hepatocytes.Biochem. Biophys. Res. Commun. 163, 1370-1376.
Zarnegar, R., Muga, S., Rahtfa, R. and Michalopoulos, G. (1990).Tissue distribution of hepatopoientin A: A heparin-bindingpolypeptide growth factor for hepatocytes. Proc. Nat. Acad. Sci.USA 87, 1252-1256.
(Accepted 22 October 1991 - Accepted 14 November 1991)