disulfide mutants of barnase i: changes in stability and structure assessed by biophysical methods...
TRANSCRIPT

J. Mol. Biol. (1995) 253, 493–504
Disulfide Mutants of Barnase I: Changes in Stabilityand Structure Assessed by Biophysical Methods andX-ray Crystallography
Jane Clarke, Kim Henrick and Alan R. Fersht*
In this series of papers, we examine the effects of introducing disulfideCentre for ProteinEngineering, MRC Centre bonds on the properties, structure and thermodynamics of a small globular
protein, barnase. Three mutants have been made, in each of which a singleHills Road, CambridgeCB2 2QH, UK crosslink confers different properties. Two of the disulfide bonds, between
residues 43 and 80 (43–80) and between residues 85 and 102 (85–102),stabilise the protein, relative to both wild-type and the corresponding(reduced) dithiol forms: 85–102 is more stable than predicted from theentropic destabilisation of the unfolded state; 43–80 is less stable thanpredicted. The third disulfide bond, between residues 70 and 92 (70–92)destabilises the protein relative to both wild-type and the correspondingdithiol form, implying significant disruption of the folded protein onformation of the disulfide bond. Crystal structures of the three mutantproteins have been solved. All three proteins have essentially the same foldas wild-type, but with left-handed disulfide bonds, which have dihedralgeometries that have not been observed in naturally occurring disulfides. Inthe very stable mutant 85–102, there is no significant difference between themutant and wild-type structures: these data do not explain the large stabilityof this protein. The disulfide bond at 43–80 induces small structuralrearrangements close to the site of the disulfide bond, associated with somelocal disorder: the crosslink appears to decrease the stability of the nativeform of the protein. The destabilising disulfide bond at 70–92 inducesconsiderable structural change, with displacement of a loop and consequentdisruption of a stabilising salt-bridge. Our studies do not support the viewthat the conformation of the disulfide bond is crucial in determining thestability of the mutant proteins.
7 1995 Academic Press Limited
Keywords: mutation; protein stability; disulfide bond; barnase; crystalstructures*Corresponding author
Introduction
Naturally occurring disulfide bonds impart con-siderable stability to folded proteins. It is widelyaccepted that the stability conferred by the crosslink
can be largely related to the entropic destabilisationof the unfolded state (Poland & Scheraga, 1965; Paceet al., 1988; Chan & Dill, 1989). Disulfide bonds thathave been engineered into proteins, however, havenot always increased their stability (Wetzel, 1987),formation of a disulfide bond may have unfavour-able effects on the folded state (reviewed by Betz,1993). A complete thermodynamic and structuralstudy of a set of disulfide mutants has not previouslybeen reported. In this, and the accompanyingpapers, we analyse the global and local thermodyn-amic and structural effects of three different engin-eered disulfide bonds.
Three disulfide bonds have been separately intro-duced into barnase, the small extracellular ribonu-clease from Bacillus amyloquefaciens (Figure 1).Wild-type barnase has no Cys residues. Disulfide
Present address: K. Henrick, National Institute forMedical Research, The Ridgeway, London NW7 1AA,UK.
Abbreviations used: 43–80, mutant of barnase with adisulfide bond linking residues 43 and 80; 70–92mutant of barnase with a disulfide bond linkingresidues 70 and 92; 85–102, mutant of barnase with adisulfide bond linking residues 85 and 102; DDGU-F, thechange in free energy of unfolding upon mutation;[D]50%, the denaturant concentration at which 50% ofthe protein is denatured; m, the slope of the transitionupon denaturation; rmsd, root mean square deviation.
0022–2836/95/430493–12 $12.00/0 7 1995 Academic Press Limited

Stability and Structure of Disulfide Mutants of Barnase494
Figure 1. General view of wild-type barnase showingthe sites of introduction of disulfide bonds betweenresidues 43 and 80 (43–80), 70 and 92 (70–92) and 85and 102 (85–102). Picture prepared using the programMOLSCRIPT (Kraulis, 1991).
report the crystal structures of the disulfide pro-teins, and compare them with the wild-type struc-ture. We relate those structures to measurements ofthe relative stabilities of the proteins. In the sub-sequent papers, we report an analysis of the effectof two of the disulfide bonds on the folded protein,as assessed by hydrogen/deuterium exchange(Clarke et al., 1995, accompanying paper), and a fullthermodynamic analysis, by differential scanningcalorimetry (DSC) of the mutant proteins (C.Johnson, J.C. & A.R.F., unpublished).
Results
Stability of disulfide proteins
Nine different mutant proteins were producedand purified. The three disulfide proteins wereshown, following purification, by Ellman’s (1959)assay and by gel chromatography, to have no freethiols, and they do not form dimers. All single Cysproteins, except for S92C, dimerise to a considerableextent. A concentration of 10 mM DTTred was foundto be sufficient to maintain both single and doublemutant proteins in the reduced form.
Values of m, [D]50%, and the difference in freeenergies between wild-type and mutant proteinsat the [D]50% for wild-type, DDGD50%
U-F , are given inTable 1, for denaturations carried out in urea, and inTable 2 for denaturations using GdmCl. The value of[urea]50% for 85–102 is 07.5 M. This value is so highthat a final baseline for fully denatured proteincannot be accurately measured, and accurate mvalues cannot be determined, so GdmCl was used asa denaturant for 85–102. The values of m of all nineproteins fall within the range found for repetitivemeasurements on different preparations of wild-type barnase.
bonds between residues 43 and 80 (43–80:Ala43 : Cys, Ser80 : Cys), 70 and 92 (70–92:Thr70 : Cys, Ser92 : Cys), and between residues85 and 102 (85–102: Ser85 : Cys, His102 : Cys)were initially engineered in positions whichwould probe the pathway of folding (Clarke &Fersht, 1993). The three crosslinks have significantlydifferent effects on the stability of the mutantproteins. We use these proteins to examine theeffects that a disulfide bond can have upon a foldedprotein.
Two of the disulfide bonds in our study stabilisethe protein (Clarke & Fersht, 1993), one to an extentwhich is larger than predicted by theoretical ap-proaches, while the third destabilises the protein,relative to both the wild-type protein and to itscorresponding reduced form. In this paper, we
Table 1. Urea denaturation: changes in the free energies of unfolding upon mutation,determined by equilibrium denaturation
DDGD50%U-F
a DDGD50%U-F
a
(relative to (relative to[Urea]50% m wild-type) dithiol)
Protein (M) (kcal mol−1 M−1) (kcal mol−1) (kcal mol−1)
Wild-typeb 4.58 (20.01) 1.92 (20.03)70–92 SS 3.03 (20.04) 1.89 (20.16) 2.9 (20.2) 0.6 (20.4)70–92 (SH)2 3.40 (20.04) 1.94 (20.22) 2.3 (20.3)T70C 4.04 (20.04) 1.91 (20.17) 1.0 (20.1)S92C 3.89 (20.04) 1.92 (20.24) 1.3 (20.2)43–80 SS 5.77 (20.02) 1.73 (20.13) −2.1 (20.2) −3.2 (20.2)43–80 (SH)2 4.00 (20.03) 1.92 (20.14) 1.1 (20.1)A43C 4.04 (20.05) 2.00 (20.23) 1.1 (20.1)S80C 4.52 (20.04) 2.01 (20.20) 0.1 (20.01)85–102 SS 07.585–102 (SH)2 4.48 (20.05) 1.69 (20.18) −0.2 (20.02)S85C 4.26 (20.04) 1.82 (20.16) 0.6 (20.1)H102C 4.57 (20.05) 1.98 (20.24) 0.02 (20.02)
The experiments were performed in 50 mM MES (pH 6.3), at 25°C.The standard errors of the data are given in parentheses.a DDGD50%
U-F values were calculated using equation (11), at the concentration of denaturant at which50% of the wild-type protein is unfolded, 4.58 M urea.
b Wild-type values taken from six different measurements of different preparations of wild-typeprotein from this laboratory (Clarke & Fersht, 1993).

Stability and Structure of Disulfide Mutants of Barnase 495
Table 2. Guanidinium chloride (GdmCl) denaturation: changes in the free energies ofunfolding upon mutation, determined by equilibrium denaturation
DDGD50%U-F
a DDGD50%U-F
a
(relative to (relative to[GdmCl]50% m wild-type) dithiol)
Protein (M) (kcal mol−1 M−1) (kcal mol−1) (kcal mol−1)
Wild-type 2.00 (20.01) 4.40 (20.25)85–102 SS 3.00 (20.01) 4.30 (20.20) −4.3 (20.2) −4.8 (20.2)85–102 (SH)2 1.89 (20.01) 4.47 (20.21) 0.5 (20.1)
The experiments were performed in 50 mM MES (pH 6.3), at 25°C.The standard errors of the data are given in parentheses.a DDGD50%
U-F values were calculated using equation (11), at the concentration of denaturant at which50% of the wild-type protein is unfolded, 2.00 M GdmCl.
In all cases, the dithiol proteins are less stable thanwild-type barnase, with 70–92(SH)2 being the mostdestabilised and 85–102(SH)2 the least. (In urea85–102(SH)2 is slightly more stable than wild-type at[D]50%, this may reflect errors in the m value, whichis low.) The energy of destabilisation of the dithiolproteins is, within error, the sum of the energy ofdestabilisation of the two single Cys mutants, for allthree mutants.
Disulfide bonds at 43–80 and 85–102 are stabilis-ing, both relative to wild-type and relative tocorresponding reduced form of the protein; this isconsistent with them being sterically acceptablemutations. In contrast, the disulfide bond at 70–92destabilises the protein. The disulfide protein is lessstable than the dithiol form, by 0.6 kcal mol−1. Theimplication is that there may be considerable‘‘strain’’ on the folded protein induced by theformation of the disulfide bond, possibly includingsome structural rearrangement.
Determination of K Feq
The equilibrium constant between oxidised andreduced forms of a protein in the folded state can bedetermined relative to a reference thiol/disulfidereagent (Creighton & Goldenberg, 1984; Matsumuraet al., 1989; Mitchinson & Wells, 1989). A comparisonof the values of KF
eq for 43–80 and 85–102 allows usto compare the relative effects of disulfide bondformation upon the folded forms of the two mutantproteins.
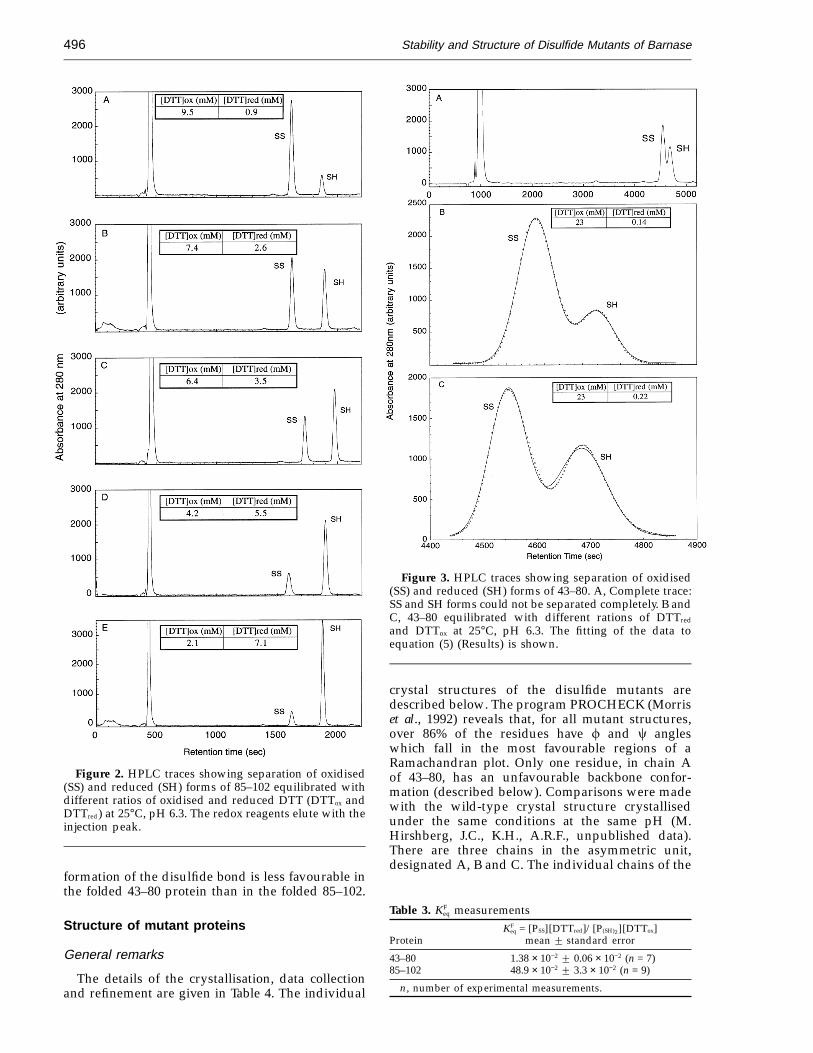
The disulfide and dithiol forms of 85–102 werecompletely separated using a linear gradient ofincreasing acetonitrile (Figure 2) and the areas ofthe peaks were calculated by the software of theHPLC apparatus. The oxidised and reduced peaksof 43–80, however, could not be completely separ-ated (Figure 3), and so the integrated peak areaswere calculated using the data analysis programKaleidaGraph, on a Macintosh computer by fittingthem to a Gaussian distribution about the maximumheight of the peak:
Y = Y0 + A
wXp2
·e−2(x−x0)2
w2 (1)
where Y is the measured absorbance, Y0 is the offset,A is the total area under the curve, x0 is the centreof the peak and w is the width of the curve at halfheight. Then
Y = Y0 + Ce−B(x−x0)2 (2)
where:
C = A
wXp2
& B = 2w2.
Thus:
w =X2B (3)
and the area under the curve, A, is
A = CXpB (4)
Two overlapping peaks were fitted to the followingequation:
Y = Y0 + Ce−B(x−x0)2 + Ee−D(x−x1)2 (5)
When measuring KFeq, the equilibrium constant
between oxidised and reduced proteins in water,DTT was used as the thiol/disulfide reagent.
PSS + DTTred~ PSSDTTSH ~ PSH
SH + DTTox (6)
Since the intramolecular reaction is very rapid,mixed disulfides do not accumulate and in anequilibrium reaction there are only two proteinspecies, disulfide (PSS) and dithiol (PSH
SH). Sampleseries of HPLC traces are shown in Figures 2 and 3.The results of at least seven experiments, usingdifferent concentrations of DTTred and DTTox wereaveraged to give a value for KF
eq as defined inequation (7):
KFeq = [PSS][DTTred]
[P(SH)2][DTTox](7)
Values for KFeq are given in Table 3.
The mean value of KFeq for 43–80, 1.4 × 10−2
(20.1 × 10−2), is significantly lower than that of85–102, 49 × 10−2 (23 × 10−2), indicating that the
Stability and Structure of Disulfide Mutants of Barnase496
Figure 2. HPLC traces showing separation of oxidised(SS) and reduced (SH) forms of 85–102 equilibrated withdifferent ratios of oxidised and reduced DTT (DTTox andDTTred) at 25°C, pH 6.3. The redox reagents elute with theinjection peak.
Figure 3. HPLC traces showing separation of oxidised(SS) and reduced (SH) forms of 43–80. A, Complete trace:SS and SH forms could not be separated completely. B andC, 43–80 equilibrated with different rations of DTTred
and DTTox at 25°C, pH 6.3. The fitting of the data toequation (5) (Results) is shown.
crystal structures of the disulfide mutants aredescribed below. The program PROCHECK (Morriset al., 1992) reveals that, for all mutant structures,over 86% of the residues have f and c angleswhich fall in the most favourable regions of aRamachandran plot. Only one residue, in chain Aof 43–80, has an unfavourable backbone confor-mation (described below). Comparisons were madewith the wild-type crystal structure crystallisedunder the same conditions at the same pH (M.Hirshberg, J.C., K.H., A.R.F., unpublished data).There are three chains in the asymmetric unit,designated A, B and C. The individual chains of the
formation of the disulfide bond is less favourable inthe folded 43–80 protein than in the folded 85–102.
Structure of mutant proteins
General remarks
The details of the crystallisation, data collectionand refinement are given in Table 4. The individual
Table 3. KFeq measurements
KFeq = [PSS][DTTred]/[P(SH)2][DTTox]
Protein mean 2 standard error
43–80 1.38 × 10−2 2 0.06 × 10−2 (n = 7)85–102 48.9 × 10−2 2 3.3 × 10−2 (n = 9)
n, number of experimental measurements.

Stability and Structure of Disulfide Mutants of Barnase 497
Table 4. Details of crystallisation and refinementProtein
43–80 70–92 85–102
A. Crystal parametersSpace group P32 P32 P32
Cell dimensions (a = b) 59.60 59.14 59.45(c) 81.79 81.79 81.52
Molecules in asymmetric unit 3 3 3Mother liquor 35–40% PEG (8K), 50 mM sodium
cacodylate, 1 mM EDTA, 50 mM sodiumacetate, 5 mM sodium phosphate (pH 7.5)
Crystal size (mm) Rod shaped100 × 600
B. Data collectionTemperature (°C) 22 −10 22Resolution (A) 13.5–2.1 10.3–2.0 15.5–2.1Reflections (observed) 57655 50660 39029Unique reflections 16295 20982 18973Completeness of data (%) 85.9 97.2 93.8Rmerge (%) 13.1 12.2 9.7
C. Refined modelCrystallographic R factor (%) 16.9 17.3 17.0Dbond (A) 0.012 0.014 0.010Dangle (°) 1.92 2.14 1.74Number of solvent molecules 230 228 216Mean B values (all atoms) (A) 16.9 13.5 17.7
mutant structures were superimposed on the corre-sponding wild-type chains, using residues outsidethe site of mutation (43–80, residues 4–40, 47–77,82–110; 70–92, residues 4–64, 72–90, 94–110; 85–102residues 4–84, 86–101, 103–110) (Figure 4). Chain Cis the best defined chain in the wild-type structureand unless otherwise stated this chain is used in thecomparisons below. Figure 5 shows the root meansquare deviations (rmsd) between the C chain ofeach mutant and wild-type. Outside the region ofthe mutation there are no significant rms differencesgreater than those between the three chains withinthe asymmetric unit, for any of the three disulfidemutants. The 85–102 mutant is the closest instructure to wild-type (over all three chains 85–102has backbone rmsd of 0.15 A, residues 4 to 110;43–80 backbone rmsd = 0.20 A outside region ofmutation; 70–92 backbone rmsd = 0.19 A outsideregion of mutation). Changes at the site of themutation are examined in more detail below. Thedisulfide bonds in all three mutant proteins haveleft-handed conformations with x3 angles close to−90° (Table 5).
43–80
At the site of the disulfide bond there is adisplacement of the peptide backbone in the regionof the disulfide bond affecting residues 43–45 and79–80 (Figure 6A). The principal change is an alteredbackbone conformation of Thr79 (Table 6). In the Achain there are two clearly defined conformations ofThr79 (Figure 6B), both were assigned an occupancyof 0.5 and included in the refinement. One of theseconformations (AII) has an associated backbone con-formation of residue 80 which falls outside the
allowed regions in a Ramachandran plot. There isessentially only one conformation of the disulfidebond (Table 5, Figure 6B).
70–92
Formation of the disulfide bond between resi-dues 70 and 92 causes a large displacement of theloop between residues 65 and 70 involving displace-ments of backbone atoms up to 7.6 A (Figures 4B,7A). There are smaller movements of residues in theturn between residue 90 and 93 (Figure 7A). Thestrands of b-sheet between residues 70 and 92 areunaffected. In all three chains of the asymmetricunit, the electron density map describes two differ-ent conformations of the disulfide bond, termed typeI and type II (Figure 7A, Table 5) which are eachgiven an occupancy of 0.5. As a consequence of therearrangement the salt-bridge between Arg69 andAsp93 is disrupted in the mutant (Figure 7B). Theside-chain of Asp93 is unaffected by the mutationbut the side-chain of Arg69 is solvent exposed andincompletely defined in the mutant structure. Thehydrophobic portion of the side-chain of Lys66 inthe 70–92 mutant occupies the cavity that remainswhen Arg69 is displaced, but the charge on Asp93remains unpaired.
85–102
The disulfide bridge between residues 85 and 102results in few changes in structure. As shown inFigure 8, there is no significant displacement of thebackbone or side-chains at the site of mutation. Thesame conformation of disulfide bond is observed inall three chains.

Stability and Structure of Disulfide Mutants of Barnase498
A
B
C
Figure 4. Superposition of the C-alpha trace ofthe mutant proteins (continuous line) with wild-type.A, 43–80; B, 70–92; C, 85–102. The site of the disulfide bondis shown.
Figure 5. Root mean square deviations (rmsd) betweenbackbone atoms of mutant crystal structures. The rmsdbetween chain C of the mutant and chain C of wild-typeis shown as a continuous line. For comparison the rmsdbetween the individual chains of the mutant asymmetricunit are shown in broken lines. A, 43–80: There issome evidence of local rearrangement. B, 70–92: There iswholesale displacement of a loop preceding residue 70.C, 85–102: There is no evidence of any significantrearrangement.
distinguish the effect of mutation from the effect ofthe introduction of the disulfide bond. All the dithiolproteins are destabilised by the sum of the destabil-isation energies of the corresponding single mutants,to within experimental error. Thus, the Cys side-chains do not interact energetically before cross-linking, and any observed changes in energy are adirect result of the disulfide bond formation.
43–80 is stabilised by 3.2 kcal mol−1 and 85–102by 4.8 kcal mol−1, compared with their respectivedithiol forms. These values, when compared withother engineered disulfides (Villafranca et al., 1987;Matsumura et al., 1989; Kanaya et al., 1991), indicatethat the disulfide bonds are formed with little strain.An analysis of the effect of disulfide bonds has twocomponents, the effect on the unfolded state (as-sumed to be largely entropic), and the effect on thefolded, native state. Theoretical approaches havesuggested that the stabilising entropic effect (DS)will be related to the size of the loop enclosed by thecrosslink (n). A commonly used approximation isthat derived by Pace et al. (1988):
DS = −2.1 − (3/2)R ln n (8)
where R is the gas constant.According to equation (8), the expected entropic
stabilisation of loops of 38 and 18 residues is −3.8and −3.2 kcal mol−1 at 25°C, for 43–80 and 85–102,respectively. If these were correct estimations ofthe effect on the unfolded state then we could inferthat the native state of 43–80 is destabilised by0.6 kcal mol−1 and that the native state of 85–102is stabilised by 1.6 kcal mol−1, a difference of
Discussion
Stability of the mutant proteins
Direct comparison of the disulfide proteins withthe corresponding dithiol form makes it possible to

Stability and Structure of Disulfide Mutants of Barnase 499
2.2 kcal mol−1. The values of KFeq measure the equi-
libria between the disulfide and dithiol forms of thefolded proteins, relative to the equilibrium betweenoxidised and reduced DTT, independently of anyeffects in the unfolded protein (equation (9)). KF
eq for85–102 is some 35 times larger than that for 43–80(Table 3); that is, the folded, native disulfide form of85–102 is considerably more stable relative to itscorresponding dithiol than is 43–80. This couldresult from stabilisation of the folded protein by thecrosslink at 85–102, or by strain induced in themolecule on formation of the bridge at 43–80, or, asis most likely, a combination of the above. Therelative effects of the formation of the crosslink,DDGSSbond, can be determined from equation (9):
DDGSSbond = −RT ln0KF85-102eq
KF43-80eq 1 (9)
DDGSSbond = −2.1 kcal mol−1, this value is very closeto the sum of the ‘‘predicted’’ effects on the nativestate.
Formation of the disulfide bond at 70–92 is de-stabilising, indicating that the final form of theprotein is considerably strained, as a result ofstructural changes (Katz & Kossiakoff, 1990). The
equation of Pace et al. (1988) (equation (8)), predictsthat the expected stabilisation conferred by en-closing a loop of 23 residues would be 3.4 kcal mol−1
at 25°C. The observed value of DDGD50%U-F is
+0.6 kcal mol−1 (Table 1). It appears that the for-mation of a crosslink between residues 70 and 92causes the folded protein to be destabilised byapproximately 4 kcal mol−1. However, following pro-longed dialysis (at least four days), the purifiedprotein was found entirely in the disulfide form. Inview of the structural changes that could be inferredfrom such a large destabilisation, further studieswere confined to structural analysis.
Relationship between structural changesand stability
In a structural analysis of the effect of disulfidebond formation on a folded protein one wouldideally use reduced protein for comparison. Crystalsof the protein could not be obtained in the presenceof 20 mM DTTred, so we use wild-type as the model.
43–80
We inferred from the thermodynamic studies thatthe folded state of 43–80 is destabilised to some small
Table 5. Details of disulfide bond geometry in the mutant crystal structuresDistance (A) Disulfide bond torsion angles (°)
Protein Ca–Ca Cb–Cb S–S x1 x2 x3 x'2 x'1
43–80Chain AI 5.51 3.93 2.04 −69 −179 −93 −47 −56Chain AII 5.70 4.03 2.08 −60 +176 −99 −51 −81(WT A) (5.16) (3.67)Chain B 5.56 3.88 2.04 −76 −177 −89 −52 −70(WT B) (5.12) (3.74)Chain C 5.45 3.69 2.03 −85 −174 −88 −47 −74(WT C) (5.08) (3.51)70–92Chain AI 4.82 3.98 2.06 −99 −40 −98 +123 −63Chain AII 4.95 4.13 2.05 −180 +86 −105 −58 +53(WT A) (5.22) (4.47)Chain BI 5.11 4.28 2.06 −104 −39 −117 +120 −47Chain BII 5.11 4.22 2.05 −180 +93 −106 −61 +54(WT B) (5.66) (5.00)Chain CI 4.82 3.97 2.00 −94 −41 −93 +121 −65Chain CII 4.89 4.23 2.03 −172 +79 −90 −62 +53(WT C) (5.27) (4.50)85–102Chain A 4.55 3.93 2.04 −91 −52 −90 102 −87(WT A) (4.80) (4.47)Chain B 4.38 3.94 2.04 −90 −46 −92 94 −83(WT B) (4.45) (4.38)Chain C 4.45 3.95 2.04 −87 −47 −93 97 −87(WT C) (4.43) (3.97)
The geometry of a disulfide bond is represented by five torsion angles, as described below:
The details are included for all three chains in the crystal asymmetric unit. Correspondingdistances in the wild-type (WT) crystal structures are given in parentheses.

Stability and Structure of Disulfide Mutants of Barnase500
Figure 6. The site of the disulfide bond of 43–80. A, Superposition of chain C of 48–80 (yellow) on chain C of wild-type(red). There are small displacements of the backbone atoms in the mutant. B, Superposition of chain A of 48–80 on chainA of wild-type (red). There are two different conformations of mutant observed, type I (yellow) and type II (green), thatdiffer in the backbone conformation of residues 79 and 80.
extent upon formation of the disulfide bond. Acomparison of the crystal structure with wild-typereveals that there is structural reorganisation atthe site of the disulfide bond. The backbone atomsof residues 43 and 44 have rms deviations fromwild-type of 0.48 and 0.63 A, respectively (backbonermsd = 0.20 A), and there is a rearrangement of thebackbone atoms of residue 79 and 80, associatedwith disorder in chain A. Away from the site of themutation there is little difference between 43–80 andwild-type. We infer that these small structuralreorganisations account for the lower than predictedstability of this protein.
85–102
The structure of 85–102 is remarkably similar towild-type, there is little structural reorganisation,even at the site of the disulfide bond. All the localhydrogen bonding interactions are maintained, andno new interactions are observed. The disulfide bondis found in a single conformation (Table 5) that is thesame as one of the two conformations adopted by thedisruptive disulfide bond at 70–92. There is noapparent structural basis for the larger than pre-dicted stabilisation of this protein.
70–92
Formation of the disulfide bond between resi-dues 70 and 92 results in considerable structuralrearrangement. There is some movement of thebackbone of residues 91 to 93, but the most notableeffect is that the loop between residues 65 and70 adopts a new conformation. This results inthe loss of the salt-bridge between Arg 69 andAsp 93. Mutagenic studies have shown that deletionof this salt-bridge leads to considerable loss instability, with an interaction energy determinedto be 3.3 kcal mol−1 (A. Tissot, S. Vuilleumier, M.Oliveberg, A.R.F., unpublished data). Some cautionis required in interpretation since the 60–70 loop is
Table 6. Backbone torsion angles of residue Thr79, inwild-type barnase and disulfide mutant 43–80
Backbone torsion angles residue Thr79
v (78–79) f c v (79–80)
43–80Chain AI −178 −119 −71 178Chain AII −177 −150 55 176Chain B −178 −94 −83 176Chain C 180 −104 −68 177
Wild-typeChain A 174 −125 −44 180Chain B 173 −128 −42 −179Chain C 177 −129 −61 178

Stability and Structure of Disulfide Mutants of Barnase 501
Figure 7. The site of the disulfide bond of 70–92.A, Superposition of chain C of 70–92 on chain C of wild-type (red). There are two different conformations ofmutant observed, type I (yellow) and type II (green)(Table 5). These differ in the conformation of the disulfidebond. B, Superposition of chain C of 70–92, type I only(yellow), on chain C of wild-type (red), some side-chain N atoms are shown in blue. The salt-bridge betweenArg 69 and Asp 93 in wild-type is disrupted in the mutant.In the mutant crystal structure the side-chain is solvatedand incompletely defined. In the mutant the side-chain ofLys66 occupies the cavity left by Arg69, but no hydrogenbonds are formed with Asp93.
Figure 8. The site of the disulfide bond of 85–102.Superposition of chain C of 85–102 (yellow) on chain C ofwild-type (red).
experiments on the pH dependence of the stabilityof this protein (J.C., M. Oliveberg & A.R.F., unpub-lished data) indicate that the salt-bridge is indeeddisrupted, and that this observation is not a crystalpacking artifact. Two conformations of disulfidebond are seen in all three chains of 70–92. The impli-cation of crystallographic evidence for two differentconformers is that there are two energetic minimawhich can be occupied by the disulfide bond. Oneof these conformations is similar to that found in85–102 (Table 5), implying that this is not of itself ahighly destabilising conformation. The loss of stab-ility upon formation of the disulfide bond in thismutant is consistent with the considerable structuralrearrangement and loss of this salt-bridge which isobserved.
The structures of a number of proteins withengineered disulfide bonds have been solved byX-ray crystallography (Katz & Kossiakoff, 1986, 1990;Villafranca et al., 1987; Pjura et al., 1990; Jacobsonet al., 1992). The studies reveal that disul-phide bonds can be formed with strained dihedralangles. In a survey of the protein data base Katz &Kossiakoff (1986) observed that naturally occurringleft-handed disulfides, with x3 angles around −90°fell into five categories, defined by the dihedralangles x1, x2, x'2 and x'1 (Table 5). They suggested thatthe high strain energies associated with disulfidebond formation play a role in decreasing the stabilityof these mutant proteins. Our studies do not supportthe view that the conformation of the disulfide bondis important in determining the stability of themutant proteins. We observe non-native disulfidebond geometry in all three mutants. It is possible thatthese are destabilising conformations, but there is noobserved correlation between the final stability ofthe protein and the calculated dihedral strain energyof the disulfide bonds themselves (calculated by themethod described by Katz & Kossiakoff, 1986).
involved in some crystal packing interactions, andhas been shown to have a different conformationin the solution structure (M. Hirshberg, M. Bycroft& A.R.F., unpublished data). However, preliminary

Stability and Structure of Disulfide Mutants of Barnase502
This study demonstrates clearly that the effects ofthe introduction of a disulfide bond on a protein arefar from predictable. The biophysical evidence ofstructural rearrangement (from equilibrium denat-uration and from comparison of KF
eq values) has beenconfirmed by structural studies. Biophysical dataalone is thus sufficient to infer large or small scalestructural rearrangements, in particular when morethan one mutant protein is being compared to wild-type. The structural data are important to determinethe nature of structural rearrangements. The confor-mation of the disulfide bond itself has no clear effectupon the stability of the mutant proteins, and thusmust be taken to have a minor effect. The dataprovide no explanation for the larger than predictedstability conferred by the disulfide bond at 85–102.There are no significant structural rearrangements inthis protein, and no new interactions which mightaccount for the increase in stability; indeed thedisulfide bond, having a non-native, apparently‘‘strained’’ conformation, would appear to be de-stabilising. The inference is that either the theoreticalapproach underestimates the effect of the disulfidebond on the unfolded protein, or that the effect ofthis disulfide bond on the folded protein is to affectlocal stability in a manner which cannot be observedby X-ray crystallography.
Materials and Methods
Materials
The urea and guanidinium chloride (GdmCl) used indenaturation experiments were ultra pure enzyme gradefrom BRL. 2-(N-morpholino)ethanesulphonic acid (MES)buffer and molecular biology grade dithiothreitol (DTTred)were from Sigma. Oxidised DTT (DTTox) was obtainedfrom Calbiochem and purified as described by Creighton(1984). Other chemicals and reagents were obtained fromSigma, BDH or Fisons.
Selection of suitable sites for SS bonds
The criteria for selection of the site of the disulfidebonds, on the basis of the known folding pathway ofbarnase, has been described earlier (Clarke & Fersht,1993). In brief, the X-ray crystal co-ordinates of barnase(Mauguen et al., 1982; A. Cameron & K. Henrick,unpublished) were examined to select suitable sites. Thesites were selected according to the following criteria.(1) Within regions of interest, pairs of residues were iden-tified that satisfied the inter-atomic differences, Ca–Ca, 4.4to 6.8 A, and Cb–Cb, 3.45 to 4.5 A (Richardson, 1981;Thornton, 1981; Hazes & Dijkstra, 1988). (2) Where poss-ible, the side-chain to be replaced would not be involvedin any stabilising interactions (hydrogen bonds, hydro-phobic interactions). (3) If possible, adverse steric effectswere minimised by replacing a residue with a side-chainof a similar size to Cys.
Mutagenesis, expression and purification ofmutant proteins
The mutagenesis, expression and production of thedisulfide mutant proteins have been described (Clarke &Fersht, 1993). Proteins with single Cys mutations were
produced in the same way. The double-mutant proteinswere purified in the absence of any reducing agent, thesingle Cys mutants were purified in the presence of10 mM DTTred. The free-thiol content of the proteins wasassessed by standard methods, Ellman’s (1959) assay,using DTNB (dithio-nitrobenzoic acid), and non-reducingacid-urea gel electrophoresis. All three disulfide mutantswere shown to be completely oxidised following purifi-cation and dialysis (results not shown). Incubation for 30minutes at 37°C with 10 mM DTTred was shown to besufficient to maintain the thiols of all nine mutant proteinsin a reduced state. After purification, the proteins wereextensively dialysed against de-ionised water, flash frozenand stored in aliquots in liquid nitrogen.
Equilibrium denaturation
The equilibrium denaturation of 43–80 and 85–102, bothoxidised and reduced, has been previously described(Clarke & Fersht, 1993). The change in free energy ofunfolding (DDGU-F) of 70–92 and of the single Cys mutantproteins was performed in the same way. Unfolding wasmonitored by fluorescence spectroscopy (Hartley, 1975),with excitation at 290 nm and emission at >315 nm.The intrinsic fluorescence of barnase decreases by about80% on unfolding (Kellis et al., 1989). For each datapoint collected, 100 ml of a solution of 09 mM barnase in450 mM MES (pH 6.3) was added to a microfuge tubecontaining 800 ml of the appropriate denaturant solution(urea or guanidinium chloride) and incubated at 25°C forat least two hours. The stability of the oxidised proteinswas measured in the presence of 1 mM DTTox. To obtainreduced proteins, DTTred was added to 180 mM to the sol-ution of protein, which was then incubated for 30 minutesat 37°C before being added to the denaturant solution(final [DTTred] = 20 mM). Fluorescence was measured inthermostatted cuvette holders at 25°C, monitored by athermocouple in the cuvette held above the light beam.The data were fitted to equation (10) by non-linear leastsquares analysis using the general curve fit option of theKaleidaGraph (Abelbeck Software) program, which givesthe calculated standard errors for individual experimentalmeasurements of m and [D]50% (Clarke & Fersht, 1993).
F =(aF + bF[D]) + (aU + bU[D])exp4m([D] − [D]50%)/RT5
1 + exp4m([D] − [D]50%)/RT5(10)
where F is the fluorescence at the given [denaturant], aF
and aU are the intercepts and bF and bU are the slopes ofthe baselines at low (F) and high (U) denaturant concen-trations, [D] is the denaturant concentration, [D]50% is thedenaturant concentration at which 50% of the protein isdenatured, m is the slope of the transition and T is thetemperature in K. The difference free energies betweenany two proteins at the [D]50% for wild-type, DDGD50%
U-F , weredetermined using equation (11);
DDGD50%U-F = −m'([D ']50% − [D]50%) (11)
where [D]50% is for wild-type, and m' and [D ']50% are thevalues for the mutant. It has been shown that DDGD50%
U-F isthe most reliable estimate of free energy differences, sinceit is not based on a long extrapolation to 0 M denaturant:for a large number of mutants of barnase, DDGD50%
U-F plottedagainst DDG determined by calorimetry, gives a slope of 1(Matouschek et al., 1994), and DDG values for thesedisulfide mutants determined by calorimetry are thesame as those determined here (C. Johnson, J.C. &A.R.F., unpublished data). There is increasing evidence(Matouschek et al., 1994; Johnson & Fersht, 1995) that m is

Stability and Structure of Disulfide Mutants of Barnase 503
not constant with [urea] and, therefore, extrapolations arenot linear as previously assumed.
Measurement of K Feq between disulfide and
dithiol proteins
The equilibrium constants between disulfide anddithiol forms of the protein, KF
eq, were measured underconditions favouring folding. The stock solutions of DTTox
and DTTred, were prepared gravimetrically, and theirconcentrations were determined spectrophotometrically(Cleland, 1964; Huyghues-Despointes & Nelson, 1992).Water, 1 M MES (pH 6.3) and DTT stock solutions weredegassed by extensive bubbling with nitrogen. Reactionmixtures (100 ml), in 0.5 ml microfuge tubes, each contain-ing 40 mM protein, 50 mM MES (pH 6.3), 0 to 20 mMDTTred and 0 to 25 mM DTTox were individually overlaidwith argon and incubated at 25°C overnight, to allowequilibration. A 50 ml aliquot of the equilibrium mixturewas withdrawn, mixed with 5 ml 0.5 M HCl to bring thepH to 02, to quench the exchange reaction (Weissman& Kim, 1991) and the sample was immediately loadedonto a previously equilibrated Dynamax C8 300 A(10 mm × 250 mm) reverse phase HPLC column. Disulfideand dithiol proteins were separated by HPLC with awater:acetonitrile gradient, in the presence of 0.1% (43–80,flow rate 0.4 ml/min) or 0.05% (85–102, flow rate 0.5 ml/min) tri-fluoroacetic acid. Elution from the column wasdetected by measuring absorbance at 280 nm. The proteinpeaks were identified by adding pure oxidised or purereduced protein to test samples. DTT eluted with theinjection peak under these conditions. Identical volumesof equal concentrations of oxidised and reduced proteinsgave peaks with the same integrated volumes, withinexperimental error, and so were assumed to have the sameextinction coefficient. The relative integrated volumes ofthe peaks were taken to represent the relative percentageof oxidised and reduced protein in the reaction mixture.The data were transferred to a Macintosh computer andprocessed using the KaleidaGraph program (Abelbecksoftware). The concentration of DTTox was taken to be thatadded. The final concentration of DTTred was determinedusing DTNB (Ellman, 1959; Creighton, 1989). The proteinwas assumed to have a negligible effect on the relativeconcentrations of thiol/disulfide reagents (040 mM pro-tein versus 10 to 40 mM DTT).
Crystallisation and data collection
Barnase has previously been crystallised in the pres-ence of zinc sulphate (Mauguen et al., 1982; Buckle et al.,1993). An incomplete factorial search (Jancarik & Kim,1991) was performed to screen for initial crystallisationconditions in the absence of zinc sulphate, to avoidthe chelation of the S atom by the Zn ions. A largerange of precipitant, salt and buffer was tested. Crystalswere grown using the hanging drop method. In the finalconditions, equal volumes of protein, between 5 and20 mg/ml, in 10 mM Na2HPO4, 25 mM Tris (pH 8), 1 mMEDTA, 1 mM DTTox, were mixed with the mother liquorwhich was 35% PEG8000, 50 mM sodium acetate, 50 mMsodium cacodylate (pH 7.5). The crystals were mounted incapillary tubes and X-ray data were recorded on a MARResearch image plate, using an in-house X-ray source(43–80 and 85–102) or the Synchrotron Radiation Source atDaresbury (70–92).
Structure calculation and refinement
The orientation and position of molecules in the unitcell were calculated by standard molecular replacementmethods using the wild-type structure (Mauguen et al.,1982) as a search model, using programs in the CCP4 suite(CCP4, 1979). The structure was refined by repeatedrounds of model building (using the program FRODO(Jones, 1985), on an Evans and Sutherland workstation)and refinement using PROLSQ (CCP4, 1979; Hendrickson& Konnert, 1980). The final R factor for each structure isgiven in Table 4. The stereochemistry of the final modelswas assessed using the program PROCHECK (Morriset al., 1992).
AcknowledgementsWe are grateful to Ashley Buckle for his help in
preparing this manuscript. The co-ordinates of all themutant structures have been released by the BrookhavenProtein Data Bank, and have been assigned ID codes asfollows: 43–80, 1BNE; 70–92, 1BNF; 85–102, 1BNG.
ReferencesBetz, S. F. (1993). Disulfide bonds and the stability of
proteins. Protein Sci. 2, 1551–1558.Buckle, A. M., Henrick, K. & Fersht, A. R. (1993). Crystal
structural analysis of mutations in the hydrophobiccores of barnase. J. Mol. Biol. 234, 847–860.
CCP4 (1979). The SERC (UK) Collaborative ComputingProject No. 4, Daresbury Laboratory, Warrington, U.K.
Chan, H. S. & Dill, K. A. (1989). Intrachain loops inpolymers: effects of excluded volume. J. Chem. Phys.90, 492–509.
Clarke, J. & Fersht, A. R. (1993). Engineering disulfidebonds as probes of the folding pathway of barnase:increasing the stability of proteins against the rate ofdenaturation. Biochemistry, 32, 4322–4329.
Clarke, J., Hounslow, A. M. & Fersht, A. R. (1995).Disulphide mutants of barnase II: changes in struc-ture and local stability identified by hydrogen ex-change. J. Mol. Biol. 253, 505–513.
Cleland, W. W. (1964). Dithiothreitol, a new protectivereagent for SH groups. Biochemistry, 3, 480–486.
Creighton, T. E. (1984). Disulfide bond formation inproteins. Methods Enzymol. 107, 305–329.
Creighton, T. E. (1989). Disulphide bonds betweencysteine residues. In Protein Function. A PracticalApproach (Creighton, T. E., ed.), pp. 155–168, IRLPress, Oxford.
Creighton, T. E. & Goldenberg, D. P. (1984). Kinetic role ofa meta-stable native-like 2-disulphide species in thefolding transition of bovine pancreatic trypsin inhibi-tor. J. Mol. Biol. 179, 497–526.
Ellman, G. L. (1959). Tissue sulfhydryl groups. Arch.Biochem. Biophys. 82, 70–77.
Hartley, R. W. (1975). A two state conformational transitionof the extracellular ribonuclease of Bacillus amylolique-faciens (barnase) induced by sodium dodecyl sulfate.Biochemistry, 14, 2367–2370.
Hazes, B. & Dijkstra, B. W. (1988). Model building ofdisulfide bonds in proteins with known three-dimensional structure. Protein Eng. 2, 119–125.
Hendrickson, W. A. & Konnert, J. H. (1980). In Computingin Crystallography (Diamond, R., Ramesseshan, S. &

Stability and Structure of Disulfide Mutants of Barnase504
Venkatesan, K., eds), pp. 1301–1323, Indian Academyof Sciences, Bangalore.
Huyghues-Despointes, B. M. P. & Nelson, J. W. (1992).Stabilities of disulfide bond intermediates in thefolding of apamin. Biochemistry, 31, 1476–1483.
Jacobson, R. H., Matsumura, M., Faber, H. R. & Matthews,B. W. (1992). Structure of a stabilising disulphidebridge mutant that closes the active-site cleft of T4lysozyme. Protein Sci. 1, 46–57.
Jancarik, J. & Kim, S. (1991). Sparse matrix sampling:a screening method for crystallisation of proteins.J. Appl. Crystallog. 24, 409–411.
Johnson, C. M. & Fersht, A. R. (1995). Protein stabilityas a function of denaturant concentration: The ther-mal stability of barnase in the presence of urea.Biochemistry, 34, 6795–6804.
Jones, T. A. (1985). Interactive computer graphics: FRODO.Methods Enzymol. 115, 157–171.
Kanaya, S., Katsuda, C., Kimura, S., Nakai, T., Kitakuni,E., Nakamura, H., Katayanagi, K., Morikawa, K. &Ikehara, M. (1991). Stabilisation of Escherichia coliribonuclease H by introduction of an artificial disul-phide bond. J. Biol. Chem. 266, 6038–6044.
Katz, B. A. & Kossiakoff, A. (1986). The crystallographi-cally determined structures of atypical straineddisulfides engineered into subtilsin. J. Biol. Chem. 261,15480–15485.
Katz, B. A. & Kossiakoff, A. (1990). Crystal structures ofsubtilisin BPN' variants containing disulfide bondsand cavities: concerted structural re-arrangementsinduced by mutagenesis. Proteins: Struct. Funct. Genet.7, 343–357.
Kellis, J. J., Nyberg, K. & Fersht, A. R. (1989). Energeticsof complementary side-chain packing in a proteinhydrophobic core. Biochemistry, 28, 4914–4922.
Kraulis, P. (1991). MolScript, a program to produce bothdetailed and schematic plots of protein structures.J. Appl. Crystallog. 24, 946–950.
Matouschek, A., Matthews, J. M., Johnson, C. M. & Fersht,A. R. (1994). Extrapolation to water of kinetic andequilibrium data for the unfolding of barnase in ureasolutions. Protein Eng. 7, 1089–1095.
Matsumura, M., Becktel, W. J., Levitt, M. & Matthews,B. W. (1989). Stabilisation of T4 lysozyme byengineered bonds. Proc. Natl Acad. Sci. USA, 86,6562–6566.
Mauguen, Y., Hartley, R. W., Dodson, E. J., Dodson, G. G.,Bricogne, G., Chothia, C. & Jack, A. (1982). Molecularstructure of a new family of ribonucleases. Nature,297, 162–164.
Mitchinson, C. & Wells, J. A. (1989). Protein engineeringof disulfide bonds in subtilisin BPN'. Biochemistry, 28,4807–4815.
Morris, A. L., MacArthur, M. W., Hutchinson, E. G. &Thornton, J. M. (1992). Stereochemical quality ofprotein structure coordinates. Proteins: Struct. Funct.Genet. 12, 345–364.
Pace, C. N., Grimsley, G. R., Thomson, J. A. & Barnett, B. J.(1988). Conformational stability and activity of ribo-nuclease T1 with zero, one and two intact disulfidebonds. J. Biol. Chem. 263, 11820–11825.
Pjura, P. E., Matsumura, M., Wozniak, J. A. & Matthews,B. W. (1990). Structure of a thermostable disulfide-bridge mutant of phage T4 shows that an engineeredcross-link in a flexible region does not increasethe rigidity of the folded protein. Biochemistry, 29,2592–2598.
Poland, D. C. & Scheraga, H. A. (1965). Statisticalmechanics of noncovalent bonds in polyamino acids.VII. Covalent loops in proteins. Biopolymers, 3,373–399.
Richardson, J. S. (1981). The anatomy and taxonomy ofprotein structure. Advan. Protein Chem. 34, 167–339.
Thornton, J. M. (1981). Disulphide bridges in globularproteins. J. Mol. Biol. 151, 261–287.
Villafranca, J. E., Howell, E. E., Oatley, S. J., Xuong,N.-H. & Kraut, J. (1987). An engineered disulfidebond in dihydrofolate reductase. Biochemistry, 26,2182–2189.
Weissman, J. S. & Kim, P. S. (1991). Reexamination ofthe folding of BPTI: predominance of native inter-mediates. Science, 253, 1386–1393.
Wetzel, R. (1987). Harnessing disulphide bonds usingprotein engineering. Trends Biochem. Sci. 12, 478–482.
Edited by R. Huber
(Received 24 May 1995; accepted 17 August 1995)