disaccharidase deficiencies in gerbils (meriones...
TRANSCRIPT

•
•
•
DISACCHARIDASE DEFICIENCIES IN
GERBILS (MERIONES UNGUICULATUS)
IMMUNE TO GIARDIA LAMBLIA
by
SHAWN RASHEED MOHAMMED
A Thesis submitted to the Faculty of Graduate Studies and Research in
partial fui fi Il ment of the requirements for the degree of
Institute of Parasitology
McGill University, Montreal Quebec, Canada
Master of Science
<CShawn Mohammed August 1994

Nome SüawtJ R..AS..tf~"b MDHAtnIlÎ~~ . Dissertation Abstracts Intemational i~ arranged by broad, general sublect categories Pleo.e select the one sublect which most neor\y describes the content of your dis~rtahon Enter the correspondlrlg four-digit code ln the spaces provided
/. lol~ I~ lozl U·M·I J..!il[QÙtJQL..O ~~ IJECTTERM SUIJECT CODE
Subiect Categories
'nI1 HUMAN.I.I. AND IOCIAL ICIINCII COMIIUNICATIOIIS AND THE AIT< Psychology 0525 'HILOSO'HY, IEUGION AND Anclent 0579 keh,loctu,. 0729 Reod,ng 0535 THEOlOGY Medieval 0581 Art Hillory 0377 Rellglaus 0527 Phllosophy 0422 Modern 0582 Cinema 0900 Sciences 0714 Reh~n Black 0320 Darw:. 0378 SecandOlY 0533 neral 0318 Alncan 0331 Fine Arts 0357 Social Sciences 0534 B,bllCal Stud,es 0321 As·a, Auwal,a and Oceonlo 0332 Information Science 0723 SoclolC!9)' of 0340 Clergy 0319 Car.ad,an 0334 Jou,noillm 0391 ~Ial 0529 Hisiory al 0320 European 0335 llbral)' Science 0399 Teacher Training 0530 Phllasophyal 0322 Lahn Amencan 0336 Mou CommunICations 0708 Technaljlkea 0710 Theology 0469 Middle Easlern 0333 MuSiC 0413 Tesls an surem~nts 0288 United States 0337 C~ Communication 0459 Vocallonal 0747 SOCIAL SCIENCES Hisiory 01 Science 0585
0465 Amencan Stud'es 0323 Law 0398 LANGUAGE, UnlATUIE AMD Anthrapol~ Pol,hcol Science
EDUCATION LlNG"ISTICS Archoeoogy 0324 General 0615 General 0515 Internallonallaw nnd Ad""nllirahon 0514 lon~ (ulturul 0326 RelatiOns 0616 Adul. and Canhnuln9 0516 ne,al 0679 Fhyslcal 0327 PublIC Admm"lratlon 0617 AnClent 0289 Business Admlnlstrahon Agrlculturel 0517 lln9uIshcs 0290 General 0310 Recreahon 0814 Art 0273 SocIQI Work 0452 Bllinguoi and Mulltculturol 0282 MOéIern 0291 Accountm9 0272 Soclology literature eoOnklng 0770 Buslne" 0688 General 0401 Management 0454 General 0626 CommullIty College 0275 ClasslCal 0294 Morkeltng 0338 Cnmlnol~ and Penolagy 0627 Currtculum and tns'ruel.on 0727 Comporotlve 0295 Canod,an Stud,es 0385 Demograp l. 0938 Early Ch,ldhood 0518 Medieval 0297 EcanomlCs Ethnlc and aCial Stud,es 0631 Elemenlory 0524 Mod1rn 0298 General 0501 Ind,vlduol and fam"y FlIlonce 0277 AI" ln 0316 Agrlcultural 0503 Slud,es 0628 Guidance and Coum.eltng 0519 Am"ncan 0591 Commerce Business 0505 Industnal and labor Heclth 0680 ASlon 0305 Finance 0508 Relations 0629 Higher 0745 Canad,an IEngllsh) 0352 Hlstory 0509 PublIC and Social Wellare 0630 History of 0520 Social Structure and Home EcanomlCs 0278 Canad,an french) 0355 lobar 0510 Develapment 0700 En911sh 0593 Theory 0511 Indu~tnal 0521 G<irmantc 0311 folklore 0358 Theory and Melhod~ 0344 ~uoge and llteroture 0279 lahn Amencan 0312 (;eograp'hy 0366 T ramporlatlon 0709
ernatlCs 0280 Middle Eastern 0315 Gerontology 0351 Urban and R~,onal Plannln9 0999 MusIC 0522 Romance 0313 H,s'ory Women's Stu les 0453 Ph,loWf?hy 01 0998 SlavlC and East European 0314 Gèneral 0578 PhyslCal 0523
THI ICIINCII AND INGINIIRING 1IOI.0GKAl KIEtKlS ~sy 0370 Speech f'athology 0460 Englneenn~ Agncultunt Geolqgy 0372 Toxlcology 0383 Genera 0537
General 0473 Geçpliyslcs 0373 Home EconomlCs 0386 Aerospace 0538 Ag,onomy 0285 ~drol<?9Y 0388 Agrlcultural 0~39 Anund CulhJre and Ineralogy 0411 'HYSICAL SCIENCES Autamohve 0540
Nuhillan 0475 Paleobolony 0345 Pure Sciences BiomedICal 0541 Anlmell PatholO!JY. 0476 Paleoecology 0426 Chemlstry ChemlCol 0542 Food ~icl.nce ana Poleontol99Y 0418 CiVil 0543 Tec~ln~r. 0359 Paleozoalagy 0985 General 0485 Electronlcs and Electrlcal 0544
Foresa one W,ldlt" 0478 Polynol~ 0427 Agrlcultural 0749 Heat and ThermodynamlCs 0348 Plant ultun! 047'>' Phys.col rophy 0368 Analytlcol 0486 Hyd,aullC 0545 Plon. PotholÇl9Y 0480 PhyslCal Oceonagraphy 0415 S,odiemlstry 0487 Inauslnal 0546 PIont Physldogy 0817 Inor~anlC 0488 Manne 0547 Rangt! Mon~ment 0777 HIALTH AND ENVIIONMEMTAL Nucear 0738 Motenals Science 0794 woOd Techn ogy 0746 SClltKlS OrganlC 0490 Meehar.lCal 0548
81~ Pharmaceultcal 0491 Metallurgy 0743
neral 0306 Envlronmenlal Sciences 0768 Phr,SlCai 0494 Mlning 0551 Heolth Sciences Poy.mer 0495 Anolomy 0287 General 0566 Raolatlon 0754 Nucleer 0552 8,01IotI511" 0308
Audlol::W 0300 Mathemahcs 0405 Packagln9 0549 Botony 0309 Chemot erapy 0992 Physlcs Petroleum 0765 Cell 0379 Son.to')' and Ml'~IClpal 0554 Ecalogy' 0329 Denhstry 0567 General 0605 System SCience 0790 Education 0350 AcoustlCs 0986 EntanlOlogy 0353 Hospital Managemenl 0769 Astronomy and Geotechnol(~w 0428 Genelles 0369 Human Development 0758 Astro~hyslCs 0606 Operations eseorch 0796 ltm~ 0793 Immunolagy 0982 Atmasp enc Science 0608 PlastICS T echnalogy 0795 MlCrob, 0410 Medicine and Surgery 0564 AtomlC 0748 Text"e T echnology 0994 MoIeculor 0307 NeuroKlence 0317 Menlol Heolth "'347 fleetronlCs and ElectnCl~ 0607 PSYCHOLOGY Oceo~aphy 0416 Nurstn9 0569 Elemenlllry Partlcles an General 0621 Nutnllon 0570 H;ph Energy 0798 pt,yslology 0433 Ob,tetncs and GynecoljlY 0380 Flui and Plasma 0759 Behavloral 0384 ROOlahon 0821 Occupaltonal Health an Molecular 0609 CllnlCal 0622 Vet.nnory Science 0778
Thera!? 0354 Nucleor 0610 Cevelopmenlal 0620 Zoolagy 0472 Oph"'a molagy 0381 OpllCs 0752 E':fu:nmental 0623
Blop!lyslCs Path.alagy 0571 Rad,at,an 0756 ln ustnol 0624 General 0786 Phormocology 0419 Soltd Stote 0611 Penonallty 0625 Medical 0760 Pharmoffi 0572 Stah.tlCs 0463 Physlalpglcal 0989
WTH SClltKlS PhbslCal eropy 0382 AppIied Sciences Psychob Iology 0349
Pu he Heolth 0571 Psychometncs 0632 Bloge:ochemlstry 0425 Roolology 0574 Applted Mechanlcs 0346 Social 0451 GeOchemlsrry 0996 Recreohon 0575 Computer Science 0984 &)
~----

SHORT TITLIl~:
DISACCHARIDASE DEFICIENCIES 1'N GIARDIA SIS
•

•
•
•
ii
TABLE OF CONTENTS
Title Page . . . . . . . . . , . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1 • • • • i
Table of Contents .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . il
Abstract . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . vi
Abrégé ................................... , .... , ... .. vii
Acknowledgements ... . . . . . . . . . . . . . • . . . . . . . . . . . . . . . . . . . . . ix
Thesis Office Statement ................................... x
Statement of Contribution .................................. xii
List of Figures . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . XIII
List of Tables .. , ...................................... xv
List of Abbreviations . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . X vi
GENERAL INTRODUCTION . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
REVIEW OF LITERATURE . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3
Taxonomy ....................................... 3
Life Cycle and Morphology ............................ 4
ln Vitro Cultivation of Giardia Trophozoites ... . . . . . . . . . . . . . .. 6
Animal Models for Human Giardiasis ...................... 8
Humoral Immune Responses ............................ 10
Cellular Immune Responses ............................ 13
Antigens of G. lamblia ............................... 16
Pathology ....................................... 18

• iii
Disaccharidase Deficiencies in Giardiasis .... . . . . . . . . . . . . . . . . 21
MANUSCRIPT 1. DISACCHARIDASE
DEFICIENCIES IN MONGOLIAN GERBILS (MERlON ES
UNGUICULA TUS) PROTECTED AGAINST GIARDIA LAMBLIA ....... .. 24
ABSTRACT . . . . . . . . . . . . . . . . . , . . . . . . . . . . . . . . . . . . . . 25
INTRODUCTION ... . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. 26
MATERIALS AND METHODS ....................... .. 29
Parasites .................................... 29
Animals ...... . . . . . . , . . . . . . . . . . . . . . . . . . . . . . . 29
Preparation of the G. lamblia
and E. histolytica Trophozoite Extracts . . . . . . . . . . . . . . . . . 30
• Quantification of Trophozoites in the Gerbil Small Intestine .... 30
Measurement of Intestinal Disaccharidases ............ . . . 31
Preparation of Intestinal Homogenate . . . . . . . . . . . . . . 31
Assay for Intestinal Disaccharidases . . . . . . . . . . . . ... . 31
Reagents .............................., 32
Collection of G. lamblia Excretory/Secretory Products ....... 32
Measurement of Protein Concentration ................. 33
Statistical Analysis . . . . . . . . . . . . . . . . .. ;. . . . . . . . . . . . 33
RESULTS ....................................... 34
Primary Infection with Live G. lamblia Trophozoites ........ 34
Challenge wlth Live G. lamblia Trophozoites .... . . . . . . . . . 36
• Challenge with the Soluble Extract of G. lamblia Trophozoites ...... 36

• iv
Challenge with the Soluble Extract of E. histolytic:a Trophozoites ... 39
Challenge with G. lamblia Excretory/Secretory Products .. .... 41
DISCUSSION . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . ...... 43
REFERENCES .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .... 49
CONNECTING STATEMENT ............................... 52
MANUSCRIPT 2. IDENTIFICATION OF A FRACTION OF
GIARDIA LAMBLIA TROPHOZOITE EXTRACT ASSOCIATED
WITH DISACCHARIDASE DEFICIENCIES IN MONGOLIAN GERBILS ... 53
ABSTRACT . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . ...... 54
INTRODUCTION .............................. .... 55
MATERIALS AND METHODS ......................... 58
• Parasites .................................... 58
Anirnals . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 58
Preparation of the erude Extract of G. lamblia Trophozoites . . . . 59
Column Chromatography .......................... 59
Sodium Dodecyl Sulfate-
Polyacrylamide Gel Electrophoresis (SDS-PAGE) ........... 60
Measurement of Intestinal Disaccharidases . . . . . . . . . . . . . . . 60
Preparation of Intestinal Homogenate . . . . . . . . . . . . . . 60
Assay for Intestinal Disaccharidases . . . . . . . . . . . . . . . 61
Reagents ..........................,.... 61
Measurement of Protein Concentration ................. 62
• Statistical Analysis .............................. 62

• v
RESULTS ................... , ................... 63
Fractionation of the Soluble Extract of G. lamblia Trophozoites ..... 63
Challenge with the Fractions of the G. lamblia Soluble Extract. ..... 63
Fractionation of FI. . . . . . . . . . . . . . . . . . . . . . . . . . . . . 68
Challenge with Fractions Fla and Flb ................. 68
SOS-PAGE .................................. 71
DISCUSSION . . . . . . . . . . . . . . . . . . . . . . . . . . . .......... 73
REFERENCES ............ . . . . . . . . . . . . . . ......... 1 78
GENERAL DISCUSSION .................................. 81
REFERENCES ........................................ 86
•
•

•
•
•
vi
ABSTRACT
Studies using Mongolian gerbils found that during a primary infection with
Giardia lamblia trophozoites, disaccharidase activities were decreased from day 10
post-infection (p.i.) untiJ weIl past elimination of the parasite. However, during a
challenge infection, enzyme deficiencies were short-lived. A challenge with a soluble
extract of G. lamblia trophozoites also resulted in reductions in disaccharidase activity.
The degree of these reductions in ~nzyme activity was dependent on the extract dose.
Gel filtration of the trophozoite eTUde extract resulted in fractions FI, F2, and F3.
However, only a challenge with FI led to disaccharidase deficiencies. Further
separation of FI resulted in fractions Fla and Flb. Impairments of enzyme activity
were obtained only in gerbils challenged with Flb. Protein analysis of Flb revealed
several high and low molecular weight bands. When gerbils previously exposed to G.
lamblia were challenged with an extract of Entamoeba histolytica trophozoites,
disaccharidase activities remained comparable to controls. Moreover, enzyme levels
in gerbils challenged with excretory/secretory G. lamblia products were affected i:t a
manner which was inconsistent with the live parasitic chaI.lenge. Results suggest that
the disaccharidase deficiencies in giardiasis are parasite-specifie and are induced by a
heat-stable constituent(s) of fraction Flb, possibly through an immune resiJÜnse to an
antigenic component of this parasite fraction.
1

• vii # #
ABREGE
Des études effectuées chez des gerboises mongoliennes ont montré que lors
d'une infection primaire avec des trophozoites de Giardia lamblia, l'activité des
disaccharidases diminue à partir du IDe jour après mfection jusqu'au-delà de
l'élimination du parasite. Cependant, lors d'une infection secondaire, la diminution de
l'activité enzymatique a été de courte durée. Une deuxième exposition, cette fois avec
un extrait soluble de trophozoites de G. lamblia, a résulté aussi en une réduction de
l'activité des disaccharidases. Le taux de réduction est lié à la dose de l'extrait. Les
fractions FI, F2 et F3 sont obtenues par filtration sur gel de l'extrait non-purifié de
• trophozoites. Cependant, seulement l'inoculation avec la fraction FI a induit une
déficience de l'activité des disaccharidases. Une séparation supplémentaire de la
fraction FI, a mené à deux autres fractions, soit Fla et Flb. La diminution de
l'activité enzymatique est observée seulement chez les gerboises inoculées avec la
fraction F lb. L'analyse protéique de la fraction Flb a révélé plusieurs bandes de poids
moléculaires variés. Lorsque les gerboises, précédemment infectées avec G. lamblia,
sont inoculées avec un extrait soluble de trophozoites de Entamoeba hisrolytica,
l'activité des disaccharidases est demeurée semblable à celle des gerboises témoines.
De plus, le niveau enzymatique était différent dans les gerboises inoculées avec les
produits secrémnt/excrétant de G. lamblia comparativement à celles infectées avec le
parasite vivant. Ces résultats suggèrent que la diminution de l'activité des
• disaccharidases dans les giardioses est spécifique à ce parasite. De plus, cette

•
•
•
VIII
diminution est induite par un (des) composant(s), stable(s) à la chaleur, contenu~ dans
la fraction Flb, et est possiblement causée par une réponse immune à un ou des
composants antigéniques contenus dans cette fraction .

•
•
1.
IX
ACKNOWLEDGEMENTS
A number of people were of assistance to me during my stay at the Institute.
Firstly, 1 would Iike to thank my research/thesis supervisor, Dr. Gaétan Faubert, for
his support and guidance throughout my graduate studies. Special thanks also go to my
advisors: Dr. Kris Chadee, for his advice and encouragement, and Dr. Dick MacLean.
ln addition, 1 would like to extend my appreciation to Dr. Elias Georges, Dr. James
Smith, Dr. Marilyn Scott and Dr. Charles Tanner for the help they provided.
Many thanks go to Darren Campbell, Dr. Nasreen Bughio, Dr. Wayne Butscher,
Vivian Lewis, Nancy Laporte, Norma Bautista-L6pez and Kis Djamiat', 1 for their
assistance, friendship and moral support during the many hours spent together in the
laboratory.
1 am grateful to animal-care technicians Susan Frappier, Brenda Lepitzki,
Martha Robinson and Joanne Tansey for the kind treatment of my many gerbils over
the years. My appreciation also goes out to Dr. George Lubega, Kathy Keller, Dr.
Bakela Nare, Dr. Carl Lowenberger, Dr. Dwayne Lepitzki, Elida Campos, Sil-king
Tse, Christiane Trudeau and Rosanne Séguin for their assistance in various ways. The
secretarial help of Mary LaDuke and Shirley Mongeau is sincerely acknowledged.
1 would like to express my gratitude to the Institute of Parasitology of McGill
University and Dr. Faubert for providing me with financial help.
Finally, 1 thank my parents and brother, Stephan, for their constant love, moral
support and encouragement.
----- ---- - -

•
•
•
x
THESIS OFFICE STATEMENT
The following is cited in accordance with the regulations of the Faculty of
Graduate Studies and Research, McGiIl University:
"Candidates have the option, subject to the approval of their Department, of
including, as part of their thesis, copies of the text of a paper(s) submitted for
publication, or the clearly-duplicated text of a published paper(s), provided that these
copies are bound as an integral part of the thesis. If this option is chosen, connecting
texts, providing logical bridges between the different papers, are mandatory.
"The thesis must still conform to ail other requirements of the "Guidelines
Concerning Thesis Preparation Il and should be in a Iiterary form that is more than a
Mere collection of manuscripts published or to be published. The thesis must include,
as separate chapters or sections: (1) a Table of Contents, (2) a general abstract in
English and French, (3) an introduction which clearly states the rationale and objectives
of the study, (4) a comprehensive general review of the background Iiterature to the
subject of the thesis, when this review is appropriate, and (5) a final ove rail conclusion
and/or summary. Additional material (procedural and design data, as weil as
descriptions of equipment used) must be provided whcre appropriate and in sufficient
detail (eg. in appendices) to a110w a clear and precise judgement to be made of the
importance and originality of the research rcported in the thesis.
"In the case of manuscripts co-authored by the candidate and others, the
candidate is required to make an explicit statement in the thesis of who contributed
-~

•
•
xi
to such work and to what extent; supervisors must attest to the accuracy of such
claims at the Ph.D. Oral Defense. Since the task of the examiners is made more
difticuJt in these cases, it is in the candidate's interest to make perfectly clear the
responsibilities of the different authors of co-authored papers."

•
•
•
xii
STATEMENT OF CONTRIBUTION
The experimental work reported herein (Manuscript 1 and Manuscript 2) was done
by Shawn R. Mohammed.
This thesis was written by Shawn R. Mohammed.
Dr. G. M. Faubert acted as researchlthesis supervisor.
l

•
•
•
xiii
LIST OF FIGURES
MANUSCRIPT 1:
Figure 1. Disaccharidase activities over time in gerbils
with a primary infection of live G. lamblia trophozoites .... 35
Figure 2. Disaccharidase activities over time
in gerbils challenged with live G. lamblia trophozoites . . . . . 37
Figure 3. Disaccharidase activities in gerbils
challenged orally with varying amounts of the
soluble extract (antigen) of G. lamblia trophozoites 50
days after a primary infection with 1 x lO6 live trophozoites .... 38
MANUSCRIPT 2:
Figure 1. Chromatograph of the soluble
extract of sonicated G. lamblia trophozoites ........... 64
Figure 2. Disaccharidase activities in gerbils
challenged with different fractions of
the soluble extract of G. lamblia trophozoites ........ .. 65
Figure 3. Disaccharidase activities in gerbils
challenged with varying amounts of fraction
FI of the soluble extract of G. lamblia trophozoites ....... 67
Figure 4. Chromatograph of fraction FI of
the soluble extract of sonicated G. lamblia trophozoites .. .. 69
Figure 5. Disaccharidase activities in gerbils
challenged with fractions Fla and Flb ...........•... 70

l
• Figure 6. SOS-PAGE of the crude extract of G. lamblia
xiv
tfophozoites and partially purified fractions of the extract ... 72
•
• .J

• xv
LIST OF TABLES
MANUSCRIPT 1:
Table 1. Disaccharidase activities in gerbils
previously exposed to G. lamblia and challenged
with a soluble extract of E. histolytica trophozoites . . . . . . . . 40
'fable 2. Disaccharidase activities in gerbils
previously exposed to G. lamblia on day 4 post-
challenge with in vitro-released excretorylsecretory products ...... 42
•
•

• xvi
LIST OF ABBREVIATIONS
Ald Aldolase
BD Blue Dextran 2000
BSA Bovine serum albumin
ELISA Enzyme-linked immunosorbent assay
Fig. Figure
g Gravit y
• GVHR Graft-versus-host reaction
Hel Hydrochloride
HEPES N-2-hydroxyethy11,iperazine-N'-2-ethanesulfonic acid
HR High resolution
IEL Intraepithelial lymphocyte
IFN-ex/{J Interferon-ex/ {J
Ig Immunoglobulin
kDa Kilodaltons
MW Molecular weight
•

• xvii
n Number in sample
NK Natural killer
O.D. Optical density
Oval Ovalbumin
PBS Phosphate-buffered saline
p.c. Post-challenge
p.L Post-infection
RnA Ribonuclease A
• SDS-PAGE Sodium dodecyl sulfate-polyacrylamide gel electrophoresis
S.E. Standard error
SEM Scanning electron microscopy
sIgA Secretory IgA
Tel. T cytotoxic/suppressor
Th T helper
TYI-S-33 Trypticase yeast extract iron seru!1l #33
U Unit
VSP Variant surface protein
•

•
•
•
GENERAL INTRODUCTION
Giardia lamblia is a single-celled parasite capable of infecting the
gastrointestinal tract of humans, a~ '~ei1 a~ ')ther animais. This organism is endemic
throughout the world, with the highest prevalence occurring in the tropics and sub
tropics (Wolfe, 1992). The flagellated trophozoite form of the parasite colonizcs the
host's small intestine by attaching to the epithelium of the mucosal villi via its ventral
dise. Trophozoites may then become encysted within a protective wall and the resultant
cysts subsequently pass from the host in the feces. The cysts are transmitted throllgh
contaminated food or water, or direct person-to-person contact. Althollgh not fatal, the
morbidity associated with giardiasis can be considerable, especially in children. The
symptoms of giardiasis vary from none to chronic diarrhea with malabsorption. The
reason for the spectrum of clinical manifestations is not weil understood, but may
include differences in host factors, as weil as the pathogenicity of different G. lamblia
strains (Adam, 1991). Individuals may eliminate the parasite after a variable length of
time without therapy. However, a number of drugs, such as quinacrine, metronidazole,
and furazolidone, are effective in the treatment of infected patients (Wolfe, 1992).
Disaccharidase deficiencies have been a commonly reported manifestation of
giardiasis (Jennings et al., 1976), contributing to the mal absorption of nutrients seen
in this disease. The aim of this researeh was to examine the process by which G.
lamblia causes disaccharidase deficiencies in the small intestine of Mongolian gerbils
and, ultimately, to assist in exploring mechanisms for inhibiting the enzyme deficiencies

•
1-
-
2
and preventing the Jarger probJem of malabsorption. This, in turn, may help to further
define the relationship between the parasite and the host intestinal mueosa, and lead to
the development of strategies for interrupting the )ife cycle of G. lamblia. The short
term objectives of this project were to: (a) characterize the effects on disaccharidase
activity following primary and challenge inoculations with the live parasite and erude
extract of the trophozoites; (b) attempt to purify the fraction of the parasite responsible
for the decreases in disaccharidase activity; (c) determine the specificity of the effects
on disaccharidase activity in immune animais; and (d) examine the effects of G. lamblia
excretory/secretory products on disaccharidase activity.
-

•
•
•
3
REVIEW OF LITERA TURE
Taxonomy
The tirst reported observation of G. lamblia is normally attributed to van
Leeuwenhoek, who described it in 1681. While van Leeuwenhoek undoubtedly saw
sorne sort of motile protozoan, the identification as Giardia has been a matter of debate.
It was next reported in 1859 by Lambl, and subsequently a large number of diffcrent
giardias were described (Ackers, 1980). Unfortunately there is much confusion
regarding the nomenclature of these protozoa. Both Giardia and Lamblia are used as
genus names. Giardia is the name used in the Western World and Lamblia is usually
used in Eastern European countries (Ackers, 1980). The number of species within the
genus Giardia has been a subject of controversy, with at least 40 having been
described. Many of these species are indistinguishable from each other on
morphological grounds (Faubert, 1988). Filice (1952) concluded that the only stable
morphological feature of the trophozoite is the median body, and he used the shape of
this structure to div ide the genus into three groups. They are G. agilis, which is found
in tadpoles and frogs; G. duodenalis (which includes G. intestinalis/G. lamblia), which
infects humans and other mammals; and G. muris, which is found in rodenls and birds.
In North America, G. lamblia is used to refer to the human parasite, whereas G.
intestinalis is often used in Europe. Studies, including work on cross transmission
using laboratory animais (Grant and Woo, 1978), average dimensions of trophozoites

•
•
•
4
(Solovjev, 1975), and isoenzyme profiles (Bertram et al., 1983), are being used to
further eharaeterize Giardia inter- and intra-strain differences.
G. lamblia is plaeed 10 the foHowing taxonomie classification:
Phylum: Protozoa (Honigberg, 1964)
Subphylum: Sarcomastigophora (Honigberg and Balamuth, 1963)
Superclass: Mastigophora (Diesing, 1866)
Class: Zoomastigophora (Calkins, 1909)
Order: Diplomonadida (Wenyon, 1926)
Family: Hexamitidae (Kent, 1880)
Subfamily: Octomitinae (Von Prowazek, 1904)
Lire Cycle and Morphology
There are two stages in the life cycle of G. lamblia: the trophozoite, which is
the vegetative form, and the cyst, which is the infective form. After cysts are ingested,
gastric acidity induces excystment in the stomach. The trophozoites then move into the
small intestine and divide by binary fission, resulting in mature trophozoites. These
trophozoites attach to the epithelium of the gut by 10eans of their sucking dise and
evade enzymatic degradation by unknown mechanisms. Further down the smaU
intestine, encystation oecllrs on exposure to bile salts. During this proeess, the flagella
are 10st and a hyaline wall is secreted. Cysts are eventually excreted and are infective
to the next host after a latent period of 3 to 20 days (Shandera, 1990) .
The trophozoite of G. lamblia measures approximately 12-15 I-tffi in length and

•
•
•
5
6~8 #Lm in width, and resembles a pear cut in half lengthwise. There arc two nuclei in
the anterior half of the organism. It exhibits bilateral symmetry and has four pairs of
flagella extending from basal granules at the anterior pole of the !)uelei (Fee\y ('t al.,
1984). The dorsal surface is convex and may have s1lght depressions rctaled to
underlying cytoplasmic vacuoles. Its ventral surface is concave and contains a structure
caUed the sucking disc, which is refined for adherence to surfaces. Transmission
electron micrographs show a microtubular network that seems to be associated with the
attachment disc, and a series of vesicles that line up along the dorsal surface and may
be associated with nutrition of the orgamsm (Stevens, 1982). The median body, a
structure unique to the genus Giardia, is located in the posterior half of the organism
and is described as claw~hammer in shape in G. lamblia trophozoites. This structure
is composed of microtubules and is of unknown function. The cytoplasm is known to
contain axonemes, glycogen particles, free ribosomes, rough endoplasmic reticulum and
vacuoles, but no mitochondria. In addition, a structure identified as a golgi apparatus
has been reported in in vitro encysting trophozoites (Reiner et al., 1990).
The cyst is oval in shape and is approximately 8-12 #Lm in size. The cyst wall
is approximately 0.3 #Lm thick and is composed of fibrillar elements (Sheffield and
Bjorvatn, 1977). The eyst eontains two to four nuclel, basal bodies and axoncmcs of
flagella, axostyles associated with caudal flagella, rough endoplasmic reticulum,
cytoplasmic masses, vacuoles, and fragmented pieces of the sueking dise and median
bodies (Feely et al., 1984). in addition, each cyst usually eontains two trophozoites.
The chemical nature of the cyst wall is not completely understood. Although chitin has

•
•
•
6
been suggested as a major component of the cyst wall (Ward et al., 1985), studies by
Jarron and coneagues (1989) have found that N-acetylgalactosamine is the primary
amino sugar of the cyst wall. They did not detect N-acetylglucosamine, the primary
amino sugar of chitin. The cyst wall is resistant to water and changes in temperature,
and the cyst can survive in the environ ment for over 20 days when it is free of fecaI
debris (Faubert et al., 1986).
ln Vitro Cultivation of Giardia Trophozoites
A method for the long term cultivation of G. lamblia trophozoites in vitro was
developed in 1960, when Karapetyan was able to culture the trophozoites for seven
months, in the presence of the yeast-like fungus Candida guillermondi. The complex
culture medium included serum, chick fibroblasts, tryptic meat digest, and Hank's or
Earle's balanced salt solutions. Subsequently, Karapetyan (1962) modified the method
and maintained G. lamblia for five months by replacing C. guillermondi with
Saccharomyces cerevisiae. However, he was unable to axenize the culture of Giardia.
In 1970, Meyer reported axenically culturing Giardia trophozoites isolated from
rabbits, chinchillas, and cats. The cultures were axenized by separating the
trophozoites from S. cerevisiae using a U-shaped culture tube. The yeast remained on
one side while the motile trophozoites moved to the opposite arm of the tube. Then,
in 1976, Meyer reported the establishment ofaxenic cultures of Giardia trophozoites
isolated from humans. The medium used, called HSP-l, contained human serum,
Hank's balanced salt solution, phytone peptone (a papaic digest), and cysteine.

•
•
•
7
In 1980, Visvesvara adapted the Giardia trophozoites grown in HSP-l medium
to Diamond's TPS-l medium - a medium introduced by Diamond in 1968 to axenically
culture Entamoeba histolytica trophozoites. Due to difficullies in obtaining Panmede
(for the TPS-l medium), which supported growth, Gillin and Diamond (1980)
established axenic cultures of Giardia in Diamond's TYI-S-33 (trypticase, yeast extract,
iron, serum). This medium originally consisted of a nutrient broth, bovine serum, and
a vitamin-Tween 80 mixture. Later, mammalian bile was added to the medium because
it was shown to promote the growth of the trophozoites (Farthing et al., 1983; Keister,
1983). It has a1so been shown that the reducing agent L-cysteine is required for the
growth of the trophozoites, as weIl as for attachment of these trophozoites to the culture
vessel (Gillin and Reiner, 1982) .
It is of note that only the trophozoites of G. lamblia (duodenalis type) have been
successfully cultured in vitro. Although TYI-S-33 is now the standard medium used
for the axenic cultivation of Giardia trophozoites, it remains a complex and chemically
undefined medium. There are isolates of G. lamblia which will not grow in the
currently available culture media (Meloni and Thompson, 1987) and thercfore the
development of a defined medium would be helpful for studying the in vitro
requirements of the parasite. Gillin and colleagues (1986) have shown that biliary
lipids can support the in vitro growth of G. lamblia in the absence of serum. In
addition, Bifulco and Schaefer (1992) found that the serum in TYI-S-33 can be reptaced
by Ultroser Q, a characterized mixture, without affecting the growth of the parasite .
____ J

•
•
•
8
Animal Models for Human Giardiasis
Systematic study of human giardiasis requires a satisfactory animal model.
Roberts-Thomson et al. (1976) developed a reproducible mouse model for giardiasis
using G. muris cysts. Unfortunately G. muris differs physiologicaBy and pathogenically
from G. lamblia, and it does not infect humans. Domestic animaIs such as mongrel
dogs (Hewlett et al., 1982), cats (Kirkpatrick and Greene, 1985), and kittens (WOO and
Paterson, 1986) have been tested for experimental infection by G. lamblia, but they
show only low levels of susceptibility - making it difficult to reproduce experiments.
Sehgal et al. (1976) reported that adult and weanling rats become infected at low
rates when inoculated with cysts of G. lamblia. A significantly higher infection rate
was observed once the parasite had been passaged through rats. Craft (1982) reported
an infection rate of 100% in weanling Sprague-Dawley rats following oral inoculation
with 150 G. lamblia cysts. Anand et al. (1980) studied the pathogenesis of
malabsorption during giardiasis, using adult Wistar albini> rats, and found a significant
fall in the transport of glucose and glycine in intestinal segments, but observed no
mucosal abnormalities. Contrary to these findings, Woo and Paterson (1986) were
unable to infect adult Wistar rats with G. duodenalis. These studies show that rats have
a poor susceptibility to infection and seem to lack pathology at the gut level, and
therefore are not very sui table hosts to study human giardiasis.
Vinayak et al. (1979) proposed a mouse model of G. lamblia infection using
weanling Swiss mice and reported a 100% infection rate. Hill et al. (1983) showed that
suckling CF-l mice could be infected through an oral inoculation with cultured Giardia

•
•
•
9
trophozoites. Swiss mice between 2 to 4 weeks old have been used to study the
immune response to a Giardia infection (Kanwar et al., 1985; Kanwar et al., 1986;
Vasudev et al., 1982). Unfortunately, successful infections with G. lamblia can only
occur in weanling and young mice and therefore these studies on the immune response
to Giardia must take imo account the immunologically immature status of the animais
used. The age-dependent susceptibility of mice to G. lamblia limits their usefulness as
an animal model.
In 1983, Belosevic et al. demonstrated that adult Mongolian gerbils (Meriolle.(j
unguiculatus) are highly susceptible to infection with cysts or cultured tropholoites of
G. lamblia. A reproducible pattern of infection was observed and gerbils inoculated
with cultured Giardia trophozoites produced infections which were similar to those
observed following inoculation with cysts isolated from patients. Cyst release was
intermittent tiuoughout the infection and most gerbils eliminated the trophozoites from
the small intestine in six to seven weeks. The animais were protected against challenge
infections for up to eight months, following a primary infection. Faubert et al. (1983)
demonstrated the uniform susceptibility of gerbils to G. duodenalis-type organisms from
beavers and cats, as weil as to G. muris. Wang el al. (1986) studied the pathology in
gerbils infected with G. lamblia and concl uded that the gerbil-Giardia model paralleled
sorne of the clinical manifestations and histopathological changes seen in humans. So
the Mongolian gerbil seems to be a useful model for studying human giardiasis because
the animal can be infected wh en its immune system is fully competent and the gerbils
exhibit characteristics of the disease which are similar to those seen in humans.

•
•
•
10
Humoral Immune Responses
There is much evidence to suggest that antibodies play an important role in
imrnunity to Giardia. It has been shown that infections with G. lamblia are more
corn mon arnong hypogammaglobuUnemic individuals (Hughes el al., 1971; Ament and
Rubin, 1972; Hermans et al., 1976). Thompson et al. (1977) found that the nurnber
of Ig-bearing plasma cells in the lamina propria is significantly higher in G. lamblia
infected individuals. Ridley and Ridley (1976) demonstrated the presence of anti
Giardia antibodies in the serum of infected humans. Il has been suggested that these
antibodies may play a role in the elimination of the parasite from the gut. Radulescu
and Meyer (1981) found that opsonization of trophozoites of G. lamblia with serum
obtained frorn rabbits irnmunized with antigens of the trophozoites resulted in a
significantly higher phagocyto~is of the parasite by peritoneal macrophages of rabbits.
Hill et al. (1984) found that human serum containing anti-Giardia antibodies killed up
to 98% of the trophozoites in vitro. Il was suggested that this killing was complement
mediated. In addition, the attachment of G. lamblia to intestinal epithelial ceUs can be
inhibited by trophozoite-specific antibodies (Inge el al., 1988).
Anti-Giardia IgG can he detected, using an enzyrne-Iinked immunosorbent assay
(ELISA), in more than 80% of patients with syrnptomatic infection and antibody titres
appear to rernain elevated for rnonths or even years after a primary infection (Goka et
al., 1986). Anti-Giardia IgG titres may be elevated in asymptomatic individuals in
endemic areas, indicating previous exposure to the parasite (Goka et al., 1986). These
litres are presumably maintained by repeated exposure lo Giardia antigens without

•
•
•
Il
production of symptomatic infections. The relationship between the presence of anti-
Giardia IgG and protective immunity h,as not been established (Farthing, 1990).
Studies suggest that IgG antibodÏl!s may hdp in the clearance of G. muris infections in
mice. For example, Heyworth (986), usung an immunofluorescence assay, identitied
both IgG and IgA antibodies on the surface of G. muri.\' trophowites isolated from
BALB/c mice, starting from day 10 of the infection. Other studies have demonstrated
the in vitro killing of Giardia trophozoites by specifie IgG 1 (Nash et al. 1 1988) and
IgG3 (Heyworth, 1992) monoclonal antibodies, in the presence of complement. Nash
and Aggarwal (1986) raised IgG monoclonal antibodies which reacted with a 170 kDa
surface antigen on WB, RS, and Isr strains. Two of these monoclonal antibodies were
cytotoxic for trophozoites which possessed this antigen. The mechanism of killing is
unknown, but was shown to be complement independent.
Using an immunodiffusion test, Jokipii and Jokipii (1982) found that total serum
IgM levels were slightly elevated in patients with recently acquired giardiasis. Anti
Giardia serum IgM has been detected in patients by indirect immunofluorescence and
byan ELISA (Gok.a et al., 1986). Anti-Giardia IgM titres appear to increase early in
infection and then decline within 3 weeks. Sharma and Mayrhofer (1988a) found a
transient IgM response in rats during primary and secondary infections with two isolates
of G. lamblia. Deguchi et al. (1987) demonstrated the capability of anti-G. lamblia
IgM to sensitize the parasite in vitro for complement Iysis by the classical pathway, as
weil as by a unique pathway that requires Cl and factor B, but not C2 and C4. Lysis
of trophozoites can be achieved by C5b to C8, without C9. In 1988, Butscher and

•
•
•
12
Faubert produced five IgM monoclonal antibodies against G. muris trophozoites which
were able to kill these trophozoites in vitro, in the presence of complement. When
there was no source of complement, these monoclonal antibodies were capable of
agglutinating the trophozoites and impairing fIagellar movement. One of the
monoclonal antibodies, when injected intraperitoneally into mice, was found to reduce
the intestinal Giardia burden.
Roberts-Thomson and Anders (1981) have shown the presence of an ti-Giardia
serum IgA in human giardiasis. A more recent study in Indian and United Kingdom
patients suggests that only one third of patients with active infection have detectable
anti-Giardia IgA (Goka et al., 1989). However, raised titres were not found in local
control subjects, suggesting that the presence of specifie anti-Giardia IgA is indicative
of current infection (Farthing, 1990). Although there is little information on the role
of secretory IgA (sIgA) in human giardiasis, there is evidence to suggest that
individuals with sIgA deficiency are more susceptible to giardiasis (Zinneman and
Kaplan, 1972). ln a clinical study, mothers with giardiasis were found to have anti
Giardia sIgA antibodies in their milk (Nayak et al., 1987). Their breast-feeding
children had a lower incidence of giardiasis than infants born to uninfected mothers.
Experimental infections in mice with G. muris support the view that production of sIgA
is impor tant for eradication and prevention of infections (Snider and Underdown, 1986;
Heyworth el al., 1988). 80th sIgA and IgG antibodies have been demonstrated on the
surface of G. muris trophozoites (Snider and Underdown, 1986) and termination of the
infection was c10sely related to increased concentrations of anti-Giardia sIgA in

•
•
•
13
intestinal fluid (Heyworth, 1986). A biliary IgA response has also been documented
in rats after inoculation of live G. lamblia trophozoites into the intestine (Loftness (JI
al., 1984). It was found that, in these rats, sIgA coated the surface of the trophozoites.
ineluding the flagella and dorsal and ventral surfaces. Studying rats infected with G.
duodenalis, Sharma and Mayrhofer (1988b) found IgA antibodies attached to 3 % of the
trophozoites on day 7 post-inoculation and to 70% of trophozoites on day 10. In
addition, jejunal biopsies from human patients have shown sIgA on the surface of G.
lamblia trophozoites (Briaud el al., 1981). AIso, studies in G. lamblia-infected rats
have suggested that sIgA antibodies can agglutinate the parasite and/or prevent
attachment of the trophozoites to intestinal epithelial eells (Inge et al.. 1988).
However, there are sorne findings which conflict with the view that sIgA is uniformly
associated with an ability to clear the infection. For example, specifie anti-Giardia
sIgA concentrations are normal in C3H/He mice, yet these mice develop ehronic
infections (denHollander et al., 1988).
Allergie manifestations only occasionally occur in giardiasis, in association with
inereased total serum IgE (Farthing et al., 1984). Farthing (1990) suggests that either
Giardia has little ability to elicit a type 1 hypersensitivity response or there is liule
systemic exposure of Giardia antigens. The latter explanation is consistent with the
biology of the parasite, Giardia being mainly a lumenal and non-invasive pathogen.
Cellular Immune Responses
An inflammatory response in the small intestinal mucosa often accompanies
j

•
•
•
14
human infection with Giardia (Wright and Tomkins, 1977; Hartong et al., 1979).
There is an increase in lymphocyte nurnbers both within the lamina propria and in the
epithelium which, when associated with partial or sub-total villous atrophy, can be so
severe as to resemble untreated coeliac disease. Intraepithelial lymphocyte numbers
decrease when the infection resolves. However, there has been no detailed study of
lymphocyte phenotypes in human giardiasis (Farthing, 1990).
Cellular immune responses have been studied in detail during experimental
infection with G. muris in mice. As in sorne human infections, G. muris in mice
induces increased numbers of lymphocytes in the small intestinal epithelium, an event
which closely parallels the reduction in parasite numbers in the intestinal lumen (Gillon
et al., 1982; MacDonald and Ferguson, 1978). Intraepithelial lymphocytes are mainly
T ceUs, whereas in the lamina propria there is an increase in both Band T ceUs.
During a G. muri:;- infection, lymphocyte numbers in murine Peyer's patches have been
found to be more than twice the basal levels, but return to normal following resolution
of the infection. Approximately 30% of these Peyer's patch lymphocytes are T helper
(TJ ceUs and 6% are T cytotoxic/suppressor (Te/J ceUs (Carlson et al., 1986).
Hypothymic nude T cell-defi.:ient mice experience a markedly prolonged infection with
G. muris compared to immunocompetent strains (Roberts-Thomson and Mitchell, 1978;
Stevens et al., 1978). In these hypothymic mice the number of L3T4 + /Th ceUs are
profoundly reduced, whereas the numbers of Tels ceUs and macrophages are relatively
normal (Carlson et al., 1987). It is suggested, therefore, that Th cells are critically
important for the ability of mice to cIear G. muris infections. Th ceUs may be involved

•
•
•
15
in switching B cell IgM to IgA production during infection (Clark and Holberton.
1986). The role of T ceUs in the antibody response to G. lamblia in humans remains
unclear. One limited study in humans by Gottstein and colleagues (1991) found that
Th cells from Giardia-infected individuals proliferate in vitro in response to G. lamblia
antigens. This proliferation was associated with CD4 + peripheral blood mononllc1ear
ceUs depleted of CD8+ cells, but not with periphera1 blood mononuclear cells depletcd
of CD4 + ceUs.
Congenitally mast cell-deficient mice (Wf/Wf) have prolonged experimental G.
muris infections lasting 8 weeks or more, compared with BALB/c mice which c1ear the
infection in 4 to 5 weeks (Erlich el al., 1983). These observations sllggest that mast
cells play a role in controUing infection. It is possible that degranulating mast cells
release mediators which are directly toxic to the parasite or that their release iflcreases
the access of other effector cells through changes in vascular permeability (Farthing,
1990).
Although lymphocytes and granulocytes do not exhibit spontaneous cytotoxicity
for G. lamblia, Smith et al. (1983) found that granulocytes are cytotoxic for
trophozoites in the presence of serum containing anti-G. lamblia antibodies.
Neutrophils from patients with giardiasis were shown to effe.ct antibody-dependent
cellular cytotoxicity against G. lamblia in vitro. Anti-Giardia IgG was found to be the
main antibody responsible for sensitization. These antibodies were not cytotoxic for
Giardia in the absence of granulocytes, even in the presence of complement.
Natural killer (NK) ceUs, however, are not likely to be invûlved in the expulsion

•
•
•
16
of the parasite. Studies using NK cell-deficient beige mice found that they were able
to clear G. muris infections as quickly as immunocompetent C57BL/6J mice (Heyworth
et al., 1986).
Tissue macrophages have a critical role in the mucosal immune response since
they present antigens to T lymphocytes. In addition, there is evidence that macrophages
act as effector cells for the clearance of the parasite during experimental G. muris
infection. Tissue macrophages have been observed in contact with and engulfing G.
muris trophozoites (Owen et al., 1981) and rabbit peritoneal macrophages will engulf
opsonized G. lamblia trophozoites in vitro (Radulescu and Meyer, 1981). Mouse
peritoneal macrophages have been shown to kill G. muris trophozoites in vitro, a
process which can be enhanced by the addition of immune serum or milk containing
anti-Giardia IgG and IgA. Belosevic and Faubert (1986) found that macrophages from
the G. muris-resistanl BIO.A mice were more phagocytically active and more
chemotactically responsive during a G. muris infection, as compared to those from the
susceptible A/J mice. Peripheral bl00d monocytes have also been shown to exhibit
spontaneous cytotoxicity against G. lamblia (Smith et al., 1982a).
Antigens of G. lamblia
lt has become clear that isolates of G. lamblia, although morphologically
identical, are different both genotypically and phenotypically. There are both inter- and
intra- strain antigenic differences (Smith et al., 1982b; Aggarwal and Nash, 1988) .
These variations may explain the broad spectrum of clinical disease observed in

•
•
•
17
giardiasis, as weIl as the absence of a reliable diagnostic test.
A variety of antigens have been detected, but little is known about their structure
and properties. Some of them have been shown to be glycoproteins with hydrophobie
dornains. Einfeld and Stibbs (1984) identtfied an 82 kDa surface glycoprotein in four
different G. lamblia isolates. Others have reported both 82 and 56 kDa surface
antigens common to P-l, Isr and WB strains of G. lamblia (Kumkum et al., 1988a).
Sorne patients with giardiasis developed an IgM response to the 82 kDa antigen, which
was associated with the resolution of the infection (Kumkum et al., 1988b). Char and
colleagues (1991) found that there was a 57 kDa antigen which was recognized by
serum IgG from Giardia patients. In addition, patients with non-persistent or
asymptomatic giardiasis have been shown to respond to a 56 kDa antigen with much
higher antibody titres th an individuals with persistent infections (Vinayak et al., 1989).
The immunization of Swiss mice with this 56 kDa antigen leads to resistance to G.
lamblia infections (Vinayak et al., 1992). Antibodies to a 170 kDa surface antigen in
G. lamblia are known to be cytotoxic to the parasite (Nash and Aggarwal, 1986). Each
G. lamblia isolate usually expresses one major variant surface protein (VSP) which
covers the entire trophozoite surface (Pimenta et al.! 1991). Thes.:! VSPs undergo
spontaneous variation in vitro and are likely a family of cysteine rich proteins which
can be secreted into the growth medium (Aggarwal el al., 1989). G. lamblia isolates
with different VSPs vary in their su sceptib ilit Y to intestinal proteases and this may lead
to differences in virulence (Nash et al., 1991).
Certain internaI antigens have also becn identified (32-170 kDa), sorne of which

•
•
•
18
appear to be highly immunogenic and associated with the cytoskeleton (Torian et al.,
1984). One of these proteins is the tubulin-associated protein "giardin", which has a
rnolecular weight of approximately 30 kDa. A variety of low molecular weight proteins
have also been idf'ntified, although their precise location in the parasite is uncertain
(Farthing, 1990). Sorne Giardia antigens are excreted/secreted during in vitro growth.
These antigens have been found to range in size from 94 kDa to 225 kDa (Nash et al.,
1983).
Considerable attention has been devoted to cyst antigens, which in sorne
instances appear to be different from the major trophozoite antigens, although a 65 kDa
antigen is corn mon to both (Rosoff and Stibbs, 1986a). This 65 kDa antigen is resistant
to proteolytic degradation and to prolonged storage at 40 C and -200 C in 10% formalin
and distilled water (Rosoff and Stibbs, 1986b). A group of antigens ranging from 21
to 39 kDa have been shown to appear early during in vitro encystation, and 66, 78, 92
and 103 kDa antigens are observed later (Reiner et al., 1989). Cyst antigens of 66, 78,
94, 103 and 120 kOa have been recognized by secretory and serum IgG, IgM and IgA
antibodies from patients with giardiasis (Reiner and Gillin, 1992).
Pathology
Light microscopy of the slnall intestinal rnucosa frorn Giardia infected
individuals frequently reveals histological changes. These changes inc1ude varying
degrees of infiltration of polymorphonuciear leukocytes and lymphocytes into the
epithelium, accumulation of mononuc1ear leukocytes in the lamina propria, shortened

•
•
•
19
villi (decreased ratio of villous to crypt cells), loss of the brush border, damage to
epithelial ceUs, and an increase in epithelial cell mitosis (Yardley (If ai., 1964: Takano
and Yardley, 1964; ~-Ioskms et al., 1967; Wright and Tomkins, 1977). These changes
range from minimal injury to extensive histological damage wilh total villous atrophy,
flattening of the epithelial cells, and dense mononuclear cell infiltration (Levinson and
Nastro, 1978). Studies by Saha and Ghosh (1977) and others have sllggested that G.
lamblia can invade the intestinal mllcosa. These researchers demonstrated the presence
of trophozoites within the mucosa. However, such reports of intestinal invasion by
Giardia are not universally accepted (Owen et aL, 1979).
Electron microscopy has also provided much information on histological changes
occurring in giardiasis. Scanning electron microscopy (SEM) of the intestinal mllcosa
from mice infected with G. muris reveals circlIlar indentations on the epithelial surface
where trophozoites had been attached (Owen et al., 1979). Using SEM, Erlandsen
(1974) observed that the trophozoites of G. muris in rats heavily infected with the
organism almost completely coyer the apical two thirds of the villi. Transmission
electron microscopy of the epithelium of jejunal mucosa from persons with giardiasis
shows that ultrastructural changes in epithelial ceUs accompany intlamed as weil as
noninflamed regions of the mucosa (Takano and Yardley, 1964). Thesc changes
include swelling of membrane-bound cytoplasmic sh uctures, distortion of nuclei, and
reduction of the height and number of epithelial cell microvilli. Studies of G. muris
infections in mice by Roberts-Thomson and colleagues (1976) found that mice which
were inoculated with high numb~rs of cysts had a greater impairment of weight gain
l

20
and more severe alterations in small intestinal architecture than mice given lower cyst
dosages. However, these changes were reported to be transient, with normal mucosal
structure retuming foUowing elimination of the parasite.
Changes in intraepithelial lymphocyte (IEL) numbers have also been observed
in giardiasis. Miee infected with G. muris have been reported as having significantly
higher IEL counts than those normally observed, from 3 to 10 weeks post-infection
(Gillon et al., 1982). These increased IEL counts persist even after the parasite is
eliminated from the intestine. The IELs are mainly T cells and it has been proposed
that these T cells secrete lymphokines which can contribute to villous damage (Gillon
el al., 1982). In addition, IELs have been found to exhibit direct cytotoxicity during
G. lamblia infections in micc (Kanwar et al., 1986). However, a more recent study
suggested that the IELs involved in the decline phase of a G. lamblia infection are
mainly Tb' not Tc/., cells (Vinayak et al., 1991). The specifie mediators secreted by
IELs in a G. lamblia infection have yet to he examined.
Other a1terations that have been described in human giardiasis inelude fat and
vitamin B'2 malabsorption, indicating that intestinal dysfunetion can extend into the
ileum (Hoskins et al., 1967; Wright et al., 1977). Vitamin A deficieney has been
reported to be associated with G. lamblia infections in children (Mahalanabis et al.,
1979). In sorne cases, protein-Iosing enteropathy and redueed serum carotene levels
have been observed (Sherman, 1980). AIso, individuals with reduced gastric acidity
have been found to be more likely to develop protein-energy malnutrition during a G.
lamblia infection, as nutrients are more difficult to absorb in these patients (Slonim et

•
•
•
21
al., 1976). It has been proposed that the induction of bacterial proliferation in the
smal1 intestine concurrent with a G. lamblia infection could result in the malabsorption
found in giardiasis (Tomkins et al .• 1978). However, this mechanism has not been
proven.
Based on the histological alterations observed in human and animal giardiasis.
a number of pathogenic mechanisms have been proposed as explanations for intestinal
dysfunction. These include the presentation of a mucosal barrier to the passage of
nutrients due to the presence of a high number of trophozoites, cellular in jury and
inflammatory reactions, and mucosal cell invasion by the trophozoites (Solomons,
1982) .
Disaccharidase Deticiencies in Giardiasis
The final stage of carbohydrate digestion occurs on the luminal surface of small
intestinal epithelial cells, which is known as the brush border or microvillar membrane.
In this membrane several glycosidases, which hydrolyze di- and oligo- saccharides
originating from ingested nutrients, are present. These glycosidases are referred to as
disaccharidases. The activity of disaccharidases is known to be modified by various
factors such as age, nutritional status, hormones, pancreatic secretions, and dietary
composition (Goda and Koldovsky, 1988). Diseases that damage the small intestinal
mucosa, such as giardiasis, also affect this enzymatic function (Banai et al., 1990).
Disaccharidase deficiencies have been consistently identified as one of various
abnormalities of small intestinal pathology in giardiasis (Jennings et al., 1976) .
J

•
•
•
22
Quantification of brush border disaccharidase enzyme levels during human giardiasis
has shown that lactase, sucrase, and leucylnaphthylamidase are reduccd (Hartong et al.,
1979). MacDonald and Ferguson (1978) reported that, in chronic murine giardiasis,
tissue sucrase activity is reduced - as seen in the jejunum in the "mild" human disease.
Measurement of tissue disaccharidases in murine giardiasis showed the most profound
decreases to be in lactase levels, which reached a peak 2 weeks after infection, at which
time the activities of sucrase and maltase were also below normal (Ferguson et al.,
1980). By the fourth week after infection, activities for all enzymes examined were
similar to controi levels. In c1inical terms, it has been noted that the lactase deficiency
can lead to osmotic diarrhea after ingestion of milk (Ferguson et al., 1980). Studies
by Belosevic et al. (1989) found that G. lamblia induced a transient decrease in
disaccharidase activity during the acute phase of a primary infection in gerbils. There
was a 30% to 85% decrease in enzyme activity on days JO and 20 post-infection. This
decrease was related temporally to the highest trophozoite burden in the small intestine.
These authors also found that challenge inoculation of gerbils with G. lamblia
trophozoites resulted in a sharp and immediate (24 hours after challenge) decrease in
the activity of the enzymes. This reduction in enzyme activity was found to be
independent of the size of the challenge inoculum and it occurred in the absence of
trophozoites in the intestine. In addition, challenge of gerbils previously exposed to G.
lamblia with the soluble extract of the trophozoites also resulted in disaccharidase
deficiencies, comparable with that induced by a challenge with live trophozoites. These
authors concluded that the disaccharidase deficiency associated with the primary

•
•
•
23
infection probably results from a direct effect of the parasite on the brush border of the
small intestine, while the deficiency in the secondary infection is probably due to the
local immune responses of the host. The suggestion that G. lamb/lU causes brush
border injury with consequent reduction in enzyme levels is supported by the finding
that elimination of the parasite reverses both the morphological and functional brush
border changes (Hartong et al., 1979). However, other resCc'lrchers have reported
decreases in brush border enzyme activity in the absence of microvillous damage
(Jennings et al., 1976). A more recent study by Daniels and Belosevic (1992)
examined disaccharidase activities in susceptible and resistant mice during G. muris
infections. They observed decreases in enzyme activity in susceptible C3H/HeN mice
after challenging with an antigen extract of the trophozoites. However, liule change
occurred in disaccharidase activity after a similar challenge in resistant C57Bl/6 mice.
Therefore these authors concluded that more severe disaccharidase deficiencies occur
in susceptible hosts following parasite antigen challenge.
[References for the General Introduction and Review of Literature follow the
General Discussion.]

•
•
•
MANUSCRIPT 1
DISACCHARIDASE DEFICIENCIES IN
MONGOLIAN GERBILS (MERIONES
UNGUICULATUS) PROTECTED AGAINST
GIARDIA LAMBLIA
Shawn R. Mohammed and G. M. Faubert
Institute of Parasitology
McGill University
Macdonald Campus
21 , 111 Lakeshore Road
Ste. Anne de Bellevue, Quebec
Canada H9X 3V9
(submitted for publication)
24

25
ABSTRACT
The activities of the disaccharidases lactase, maltase, sucrase and trehalase were
examined in gerbils during Giardia lamblia infections. In a primary infection with
trophozoites, the activities of al} 4 enzymes were reduced from day 10 post-infection
(p.i.) and remained at low levels well past the elimination phase of the infection.
However, during a challenge infection, the decreases in disaccharidase activity were
short-lived, with impairments only on days 2 and/or 4 post-challenge (p.c.). Sucrase
activity was not affected by a challenge infection. When 'll mg of a soluble extract
of G. lamblia trophozoites was used to challenge gerbils previously exposed to the live
parasite, the pattern and duration of enzyme deficiencies were comparable to the
challenge wÏth the live parasite. In addition, decreasing the extract dose used to
challenge the gerbils led to smaller disaccharidase deficiencies. Gerbils previously
exposed to G. lamblia were also challenged with a soluble extract of Entamoeba
histolytica trophozoites and this had no effect on the disaccharidase activities.
Therefore. the presence of the intact parasite was not necessary to induce reductions in
enzyme activitv in immune animals. As weil, the effects in the secondary infection are
parasite-specifi: and may involve the host's immune response to Giardia antigens.
Immune gerbils were further challenged with the in vitro-released excretory/secretory
products of G. lamblia. Under our experimental conditions, disaccharidase activities
were found to be affected by the~,e products in a manner which was inconsistent with
the live parasitic challenge and this merits further study.

•
•
•
26
INTRODUCTION
Giardia lamblia is an anaerobic protozoan parasite of worldwide distribution
which is found in the small intestine of humans, as weil as other animals. It is the most
frequent cause of defined waterborne outbreaks of diarrhea in developed countries
(Craun and Jakubowski, 1987; Wilson et al., 1982). The parasite exists in two forms:
the vegetative, flage.llated trophozoite and the infective cyst. Individuals infected with
Giardia may remain asymptomatic carriers or may develop sorne of the clinical
manifestations of giardiasis, such as chronic diarrhea, nausea, abdominal cramps and
distension, and malabsorption (Adam, 1991) .
The intestinal epithelium is the site of interaction between Giardia and the host,
and a mucosal immune response follows. However, the precise nature of this immune
response remain~ unknown. Several abnormalities of the small intestine have been
consistently identified in human giardiasis and these include increased intraepithelial
lymphocyte counts (Wright and Tomkins, 1977), crypt hyperplasia with shortened villi,
increased lamina propria cellularity (Yardley et al., 1964), and disaccharidase
deficiencies (Jennings el al., 1976). The pathogenic mechanisms by WhlCh Giardia
causes these abnormalities of the gut remain undefined.
Intestinal brush border enzyme deficiencies can be clearly shown during
protozoan disease and are commonly reported abnormalities of the gut (Buret et al.,
1990). Quantification of brush border disaccharidase levels during human giardiasis by
Hartong el al. (1979) showed that lactase, sucrase, and leucylnaphthylamidase were

•
•
•
27
reduced. Measurement of tissue disaccharidases in murine giardiasis showed the most
profound changes to be in lactase levels, which reacherl a peak two weeks after
infection, at which time the activities of sucrase and maltase wt'r~ also below normal
(Ferguson et al., 1980). In addition, in the gerbil animal model, decreases in enzyme
activity were observed not only in a primary G. lamblia infection, but also in challenge
infections (Belosevic et al., 1989). This reduction in enzyme activity in the immune
animaIs was found to be independent of the size of the challenge inoculum and it
occurred even in the absence of trophozoites in the intestine. 8elosevic and colleagues
also found that a challenge with a crude extract of the trophozoites resulted in lowered
disaccharidase activity, suggesting the involvement of the host's immune response in
disaccharidase deficiencies. A more recent study examined the variability in the
decreases in disaccharidase activity during G. muris infections in susceptible and
resistant mice. It was found that after challenging the immune mice with an extract of
the trophozoites, the susceptible mice showed more severe decreases in disaccharidase
activity than the resistant strain (Daniels and Belosevic, 1992). Studying the effects of
G. muris on the mouse mucosal ultrastructure, Buret el al. (1990) found that brush
border disaccharidase deficiencies are linked to a loss of microvillus surface area.
However, the precise mechanisms invoived have yet to be elucidated.
The disaccharidase deficiencies ohserved in giardiasis contribute to host
malabsorption of carbohydrates, with subsequent fluid accumulation in the intestinal
lumen and diarrhea. These deficiencies may involve the immune response to the
parasite and it has been clearly established that gerbils exhibit a long lasting immunity

•
•
•
28
to challenge infection with G. lamblia (Lewis et al., 1987; Belosevic et al., 1983). We
therefore undertook a more extensive characterization of the effects on disaccharidase
activity in gerbils previously exposed to G. lamblia. In this study, our objectives were
to: (1) examine the activity of four disaccharidases at various times following a
challenge infection with live trophozoites; (2) measure enzyme levels following a
challenge inoculation with known amounts of the soluble extract of G. lamblia
trophozoites; (3) determine the specificity of the effects that the soluble extract of G.
lamblia trophozoites has on the disaccharidases by challenging gerbils previously
exposed to G. lamblia with the soluble extract of Entamoeba histolytica trophozoites;
and (4) examine the effects of the excretorylsecretory products of G. lamblia
trophozoi tes , released in vitro, on disaccharidase activity .

• 29
MATERIALS AND METHODS
Parasites
G. lamblia trophozoites, WB strain (American Type Culture Collection no.
30957), originally isolated from a patient with chronic symptomatic giardiasis (Smith
et al., 1982) were used in this study. Trophozoites were maintained axenically in filter-
sterilized TYI-S-33 (trypticase, yeast extract, iron, and serum) medium adapted for
a.lamblia (Gillin and Diamond, 1979), with 10% adult bovine serum (Sigma Chemical
Co., St. Louis, U.S.A.), and supplemented with 100 units/mL of penicillin (Sigma) and
100 ILg/mL of dihydrostreptomycin sulphate salt (Sigma), in the absence of bovine bile.
• They were passaged twice weekly.
Pathogenic E. histolytica trophozoites, strain HM 1 :IMSS, originally isolated by
L. Diamond (National Institutes of Health, Bethesda, U.S.A.) and cultured in Dr. K.
Chadee' s laboratory (Institute of Parasitology, McGill University, Montreal, Canada)
were used for the amebic study. Trophozoites were maintained axenicaJly in autoclaved
TYI-S-33 (biosate, iron, and ~erum) medium (Diamond et al., 1978), sllpplemented
with 100 units/mL of penicillin and 100 #Lg/mL of dihydrostreptomycin slilphate salt.
Animais
Six to 10 week old male Mongolian gerbils (Meriones unguiculatus) , from
Tumblebrook Farms (West Brookfield, U.S.A.), were used throughout this study. •

•
•
•
30
They were maintained under standard laboratory conditions and provided with
laboratory chow and water ad libitum. One week after arrivaI, each gerbil was trea.ted
by oral gavage with 15 mg of metronidazole (Rhône-Poulenc, Montreal, Canada) for
three consecutive days and then allowed to rest for 10 days. This ensured that the
gerbils were free of intestinal infections.
Ail inoculations were done orally, to unanaesthetized gerbils. With live G.
lamblia inoculations" ~~h gerbil was infected with 1 x Hf trophozoites.
Preparation of the G. lamblia and E. histolytica Trophozoite Extracts
Trophozoites (from a 72-h culture) were dislodged from the walls of culture
vessels by immersion in an ice bath for 15-30 min. They were then sedimented by
centrifugation at 800 g for 10 min, at 4° C, and washed 5 times in sterile phosphate
buffered saline (PBS) (pH 7.2). Trophozoites were then lysed in sterile PBS by
sonication, using a Sonic Dismembrator (Fisher Scientific, Montreal, Canada) at
maximum output, for 10 min, in an ice bath. The sonicated material was cleared of
insoluble matter by eentrifuging at 23,300 g for 20 min, at 4° C, and the supemat.ant
eollected as the erude soluble extract.
Quantification of Trophozoites in the Gerbil Smalt Intestine
The method of Belosevic and Faubert (1983), with modifications, was ll)ed 'to
determine trophozoite numbers in the smalt intestine. Briefly, gerbils were killed and
the small intestine removed and divided into three equal sections. The segments were

•
•
•
31
slit longitudinally and each placed in 6 mL cold PBS. They were left in a shaking iee
bath for 1 h to dislodge the trophozoites from the intestinal mucosa. The intestinal
sections were then removed, the liquid centrifuged at 800 g for 10 min and the pellet
resuspended in cold PBS. The total number of trophozoites recovered from each
segment was then determined using a haemocytometer.
Measurement of Intestinal Disaccharidases
- Preparation of Intestinal Homogcnate -
Homogenates were prepared as previously described (Belosevic et al., (989),
with modifications. Briefly, the gerbils were killed and the smalt intestine removed and
divided into three sections. The segments were placed in ice cold distilled water and
were each flushed with 50 mL of distilled water. The segments wcre then slit
longitudinally and the mucosa scraped off with a glass microscope slide. The mucosa
from all three sections were combined, weighed (wet weight), and placed in four
volumes of ice cold distilled water. The mucosal scrapings were then homogenized
using a Con Torque power unit at maximum speed (Eberbach Corp., Ann ArboT,
U.S.A.). Homogenates were stored, without prior centrifugation, at - 7(J' C until used
in the assay.
- Assay for Intestinal Disaccharidases -
Disaccharidase activity of homogenated mucosa was measured using the glucose
oxidase peroxidase assay (Dahlqvist, 1968), as modified by Belosevic el al. (1989),
which is based on a colour reaction with the glucose liberated by a disaccharidase in
, j

•
•
•
32
60 min. The assay consisted of adding to e.ach weIl of a 96-well Nunc microwell plate
(Gibco BRL, Burlington, Canada) 0.01 mL of an appropriately diluted mucosal sample
(hornogenate) and 0.01 mL substrate-buffer solution of a disaccharidase to be measured,
in quadruplicate. The plate was then incubated at 37° C in humidified atmosphere for
60 min. After incubation, 0.3 mL tris-glucose oxidase reagent was added to each weIl
and the plate incubated for an additional 60 min. The plate was then read at 415 nm
using a Bio-Tek microplate reader (Mandel Scientific, Guelph, Canada). For each
assay, 8 wells of reagent blank and a glucose standard series (2, 6, and 10 p.g glucose)
in quadruplicate were also done. Disaccharidase activity is expressed as units/mg
protein in the mucosal sample (U/mg), where units represent p.moles of disaccharide
hydrolyzed/min .
- Reagents -
AIl chemicals were obtained commercially. 0-(+ )-Glucose, 11-lactose, maltose,
0-( + )-trehalose, maleic acid, glucose oxidase, o-dianisidine, peroxidase, and triton X-
100 were purchased from Sigma. Sucrose was obtained from Fisher Scientific and tris
(ultra pure) was purchased from ICN Biomedicals Canada Ltd. (St. Laurent, Canada).
Collection of G. lamblia Excretory /Secretory Products
The me.dium from a 72-h trophozoite culture was dr..canted and, in order to
remove any residual TYI-S-33 medium, the culture flask was quickly washed with
warm maintenance medium. This medium consisted of RPMI 1640 with L-glutamine
(Gibco BRL), supplemented with 11.4 mM L-cysteine Hel (Fisher Scientific), 10% fetaI

•
•
•
33
bovine serum (Flow Laboratories, Toronto, Canada), 20 mM N-2-
hydroxyethylpiperazine-N'-2-ethanesulfonic acid (HEPES) (Sigma), 100 units/mL of
penicillin (Sigma) and 100 JLg/mL of dihydrostreptomycin sulphate sait (Sigma) (Guy
et al., 1991). The culture vessel, with the attached trophozoites, was then tilled with
the maintenance medium and placed at 37° C for 6 h, in order to allow for the
maximum production of trophozoite products. Next, trophozoites were sedimented by
centrifugation at 800 g for 10 min. The supernatant was collected, a sample was
examined microscopically, and it was centrifuged again iftrophozoites were seen. This
was repeated until no trophozoites were found in the supernatant. The final supernatant
was then lyophilized and resuspended in PBS, for use as the in vitro-rele-ased products
of the parasite.
Measurement of Protein Concentration
Protein concentrations were determined ,B described by Lowry el al. (1950,
using 0.2 mL of sample and a final reaction volume of 1.3 mL. A standard curvc was
prepared using freshly dissolved bovine serum albumin (BSA).
Statisticaj Analysis
Statistical significance was determined using the Mann-Whitney V-test. The
probability level of P < 0.05 was considered significant.

•
•
•
34
RESULTS
Primary Infection with Live G. lamblia Trophozoites
In order to establish the basis of disaccharidase deficiencies in giardiasis and
provide a reference for co m pari son , the effects of a primary G. lamblia infection on the
activities of lactase, maltase, sucrase, and trehalase in gerbils were examined. For ail
4 disaccharidases, there was a 43 % to 78 % reduction in activity, starting on day 10 p. L
(Fig. 1). This time corresponds to the beginning of the acute phase of the infection,
as the highest trophozoite burden in the small intestine was found from day 9 to day 12
p. i. (data not shown). The reduction in enzyme activity was maintained well past the
elimination phase of the infection (around day 30 p.L), as it was detected even on day
50 p. i. This pattern of decreased activity was found with ail disaccharidases measured,
with the exception of maltase. Although the activity of maltase was decreased from day
10 p.L. by day 50 p.i. it had returned to normal levels. The reduction in sucrase
activity varied the most over the course of the infection (43% to 78% reduction), white
that of trehalase remained the most consistent (61 % to 76% reduction).
For ail the experiments in our study, there was a great deal of variation between
individual gerbils in a particular test group and also over time. The activity of
disaccharidases is known to be modified by a number of factors such as age, nutritional
status, hormones and pancreatic secretions (Goda and Koldovsky, 1988). In addition,
it is important to note that although the 4 enzymes examined are intestin" •
disaccharidases, they are not all affected in the same way as they are located in different
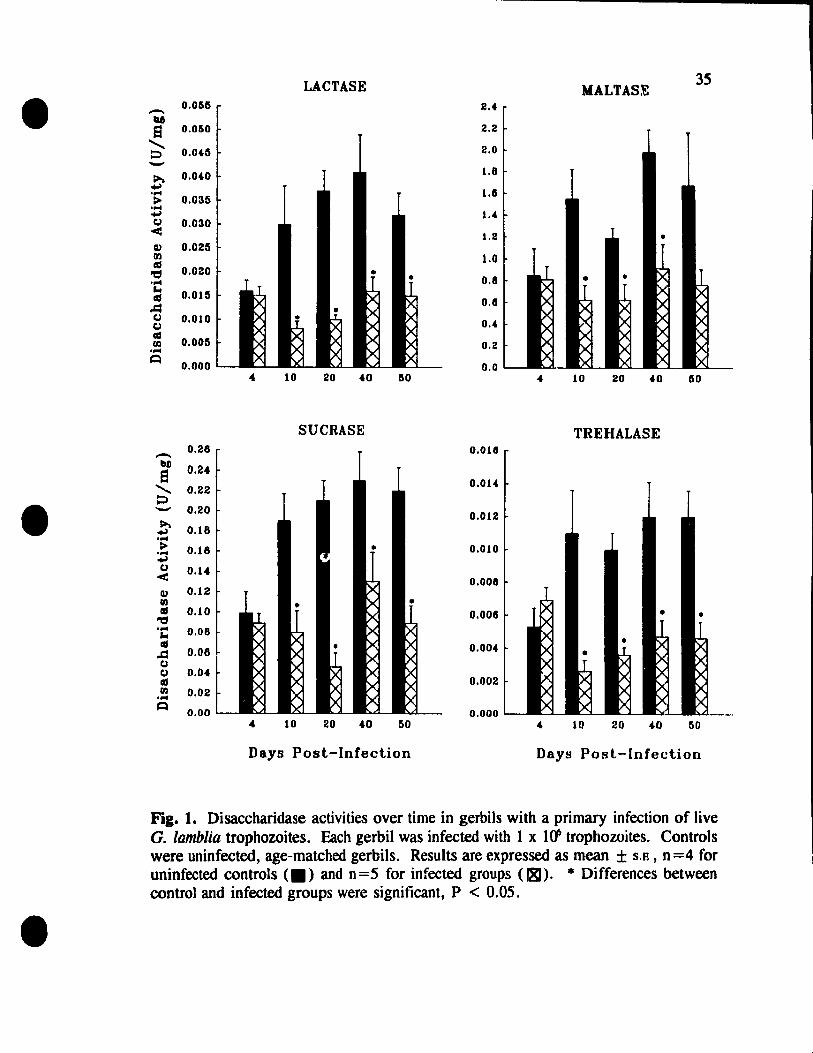
•
•
•
LACTASE MALTAS~ 35
- 0.066 2.' lUI
S 0.060 2.2
'- 0.0'6 2.0 :;:, -~ O.O~O 1.8 ~ '.-4 1.11 :> 0.035
• .-4 ~ 1.4 c:,) 0.030 < 1.2 • CI) 0.025 !Il 1.0 Ils
OC 0.020 • • '.-4 • 0.8 • ... 0.016 Ils 0.11 ..c:::
c:,) c:,)
0.010 0.4 Ils
0.005 !Il 0.2 '.-4
~ 0.000 0.0 .. 10 20 40 50 • 10 20 .0 60
SUCRASE TREHALASE 0.26 0.018 -~ 0.24 S
"- 0.22 0.014
:;:, - 0.20 0.012 ~ ~ 0.18 '.-4 :> 0.16 • 0.010 '.-4 ~ c:,) O." < 0.008 CI) 0.12 !Il Ils 0.10 0.008 • • OC .... 0.06 ... Ils
..c::: 0.06 0.004
t.l t.l O.Of Ils 0.002 !Il 0.02 .... ~ 0.00 0.000 .. 10 20 40 50 • 10 20 40 50
Days Post-Infection Days Post-Infection
Fig. 1. Disaccharidase activities over time in gerbils with a primary infection of live G. lamblia trophozoites. Each gerbil was infected with 1 x 1 rJ trophowites. Con trois were uninfected, age-matched gerbils. Results are expressed as mean ± S.E , n =4 for uninfected controis <.) and n=5 for infected groups (181). * Differences between control and infected groups were significant, P < 0.05.

•
•
•
36
areas of the mucosal brush border (Dahlqvist and Semenza, 1985).
Challenge with Live G. lamblia Trophozoites
Since gerbils are immune to challenge infections, we investigated the effects on
the activities of lactase, maltase, sucrase, and trehalase over lime in gerbiJs challenged
with G. lamblia. We were particularly interested in determining how carly after a
challenge infection enzyme activities could be affected. Therefore, disaccharidase
activities were measured as carly as 1 h (0.042 days) post-challenge (p.c.), and
compared to age-matched controls which received a primary infection but were not
challenged. Sucrase activity remained comparable to controllevels (Fig. 2). However,
the activities of lactase and trehalase were reduced on day 4 p.c. A decrease in the
activity of maltase was observed on both days 2 and 4 p.c. The reduction in lactase,
maltase, and trehalase activities in the challenged animais ranged between 33% and
59%. These decreases in enzyme activity occurred in the absence of trophozoites, as
the challenge infection does not establish in the gerbil gut (no trophozoites detected).
Challenge with the Soluble Extract of G. lamblia Trophozoites
Gerbils were challenged with 0.1 mg of the soluble extract of G. lamblia
trophozoites and the activities of the 4 enzymes under study were measured on days l,
2, 4, and 10 p.c. The activity of sucrase did not differ from controllevels on any of
these days. Howcver, the activities of lactase, maltase, and tre:.alase were reduced by
as much as 60% on day 4 p.c. (Fig. 3). In addition, the activity of maltase was also
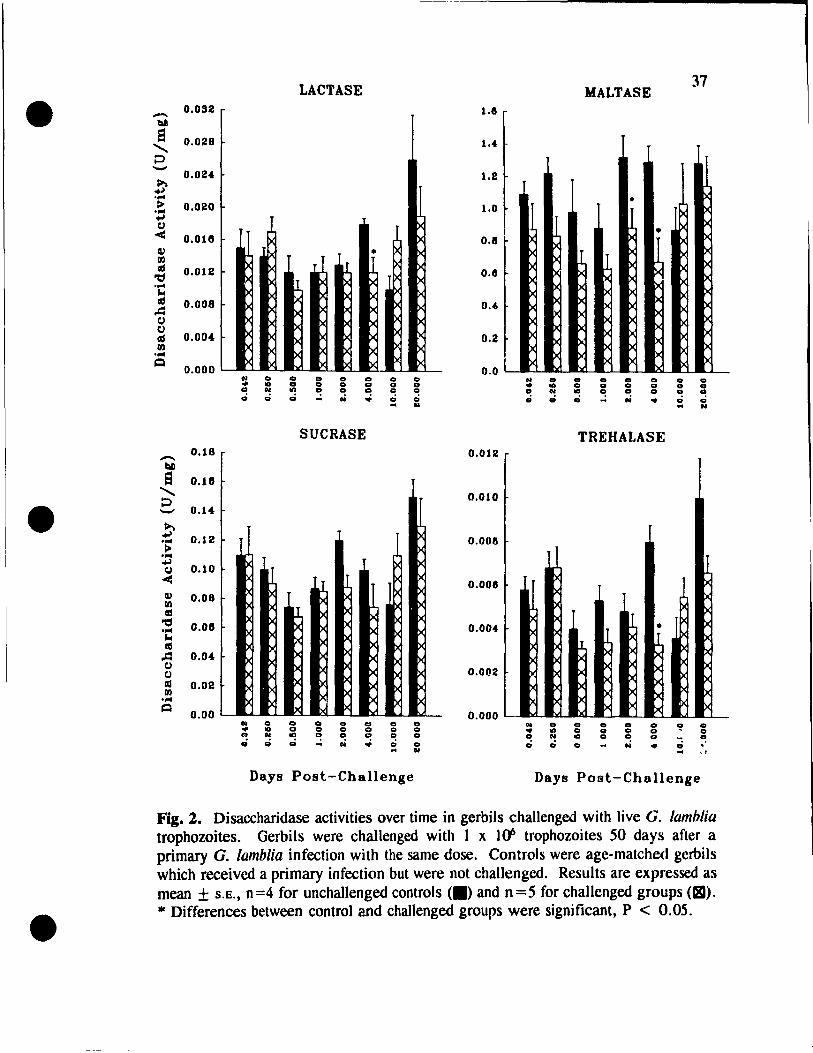
LACTASE MALTASE 37
• - 0.032 tI!
1.8
S 0.028 '-P - 0.02. ~ .. J .... > 0.020 .... 1.0
[ • 1><
1.2
+J t.)
< O.OUS C!) • III «1 0.012 't:I .... ~ «1 0.008
.cl t.)
•
L l [ IX ><:
IX )(
>< IX ><: x Xi ><: Xi x x x x
>< X x IX X X
>< IX IX >< ><
0.8
0.8
0.4
t.) 0.00. «1
III .... ~
0.000
I)c I)c IX 1>< 1>< IX I)c lx rx 1>< I)c pc
I)c lx t>< 1><
0.2
0.0 CIl .. co co .. co .. co Ct .. .. Ct .. .. ... .. CI CI .. CI CI N on "! .. CI
... on Ct .. .. .. .. .. .. N III "! .. CI CI .. .; .; .; ... oi .. .; .; .; ... N .. .; .; .. N
SUCRASE TREHALASE
- 0.18 0.012 bD e 0.18 '- 0.010 ::J • - 0.14
~ ~ 0.12 .. 0.008 > ..
~ ~ 0.10 t.)
< 0.008
~ X
1 )(
C!) 0.08 Xi
><: III X «1
L ><
'tI 0.06 IX IX IX .. 0.004 IX l
• ~ x l x «1 II
..c:I 0.0. lx 1>< IX t.) ~ lx 1>< (.) 0.002 lx 1>< i><
lx lx IX >< CIl 0.02 1>< )< III 1><
I>c lx IX 1>< .,.. IX lx i>C
~ I)c IX IX 1>< 0.00 0.000 i>C
N .. co .. .. co Ct N Ct Ct .. Ct Ct ~ .. ... la CI .. .. co CI ... on .. CI Q CI .. ft N ... ' "! .. co CI .. N ... CI .. CI "! .; .; .; ... oi .. CI .; .; CI N .. .; . N .. "
Days Post-Challenge Days Post-Challenge
Fig. 2. Disaccharidase activities over time in gerbils challenged with live G. lamblia trophozoites. Gerbils were challenged with 1 x H1 trophozoites 50 days after a primary G. lamblia infection with the same dose. Controls were age-matched gerbils which received a primary infection but were not challenged. Results are expressed as mean ± S.E., n=4 for unchallenged controls (.) and n=5 for challcnged groups (~).
• * Differences between control and challenged groups were significant, P < 0.05 .

•
•
•
0.022 -~ Si 0.020
~ 0.018 -!' 0.0111
'> o.oa .... ~
Q 0.01~ <
-ea.I
0.010
0.008
0.0011
0.00.
0.002
0.000
0.1.
e '" 0.12 ~ -~ 0.10 ~ .... ~ .... t: 0.08
-< 0.08
0.02
0.00
38 LACTASE MALTASE
1.11
1..
1.2
1.0
0.11
0.11
o .•
0.2
0.0 CONTROL 0.1 0.075 0.05 CONTROL 0.1 0.075 0.05
SUCRASE TREHALASE 0.010
0.008
0.0011
0.00.
0.002
0.000 CONTROL 0.1 0.075 0.05 CONTROL 0.1 0.0'15 0.05
Antigen Dose (mg) Antigen Dose (mg)
Fig. 3. Disaccharidase activities in gerbils challenged orally with varying amounts of the soluble extract (antigen) of G. lamblia trophozoites 50 days after a primary infection with 1 x 1(1 live trophozoites. Controi gerbils were age-matched and received a primary infection. but were not challenged. Results are for day 4 post-challenge and are expressed as mean + SE. n =4 for unchallenged controls <.) and n =5 for antigenchallenge.d groups (lX!). .je Differences between control and challenged groups were significant. P < 0.05.

•
•
•
39
reduced on day 2 p.c. (data not shown). These patterns, observed with the 0.1 mg
antigen dose, are similar to those found with the live challenge.
In order to determine whether the effect of the soluble extract on disaccharidase
activities could be obtained with lower doses, immune gerbils were challenged with
0.075 mg or 0.05 mg total of the extract. Enzyme activities were then measured on
day 4 p.c., as this was the time when the disaccharidases were most affected by the 0.1
mg extract challenge. When the antigen dose was 0.075 mg, only the lactase and
maltase activities were significantly lower th an the unchallenged controls (Fig. 3).
However, the mean reductions in these enzyme activities were not as great as in the
group of animais challenged with 0.1 mg of the soluble extract of the trophozoites.
With an antigen dose of 0.05 mg, there were no significant differences found between
challenged and control groups for any of the disaccharidases under study. However,
sucrase activity remained unaffected by the antigen, regardless of the dosage.
Challenge with the Soluble Extract of E. histolytica Trophozoites
To determine whether the effects of the G. lamblia soluble extract on the
disaccharidases are specifie to this parasite, gerbils immune to Giardia were challenged
with 0.1 mg of the soluble extraet of E. histolytica trophozoites. Disaccharidase
activities were measured on days 2 and 4 p.c. and for ail 4 enzymes the levels observed
in E. histolytica-challenged groups were comparable to control values (Table 1) .

•
•
•
40
Table 1. Disaccharidase activities in gerbils previously exposed to G. ~amblia and challenged with a soluble extract of E. histolytica trophozoites. Disaccharidase activities are the average values (n =4, for controls; n =5, for test groups) in U/mg ± SE. Gerbils were challenged orally with 0.1 mg of the soluble extract of E. histolytica trophowites 50 days following a primary infection with 1 x 106 G. lamblia trophowites. Controls were age-matched gerbils which received a primary G. lamblia infection, but were unchallenged.
Enzyme Days Post- U nchallengr.d Challenged with E.
Challenge Control histolytica*
2 0.013 ± 0.001 0.015 ± 0.002
Lactase --------------------------------------------------------------4 0.018 + 0.001 0.013 ± 0.003
2 1.32 + 0.13 0.96 ± 0.16
Maltase 4 1.29 + 0.10 1.02 ± 0.08
2 0.12 ± 0.01 0.12 ± 0.02
Sucrase 4 0.10 ± 0.01 0.098 ± 0.011
2 0.0048 ± 0.0008 0.0059 ± 0.0005
Trehalase 4 0.0080 + 0.0008 0.0059 ± 0.00 15
III Differences between control and test groups were not significant, P < 0.05

•
•
•
41
Challenge with G. lamblia Excretory ISecretory Products
Since the mainly somatie antigens of the soluble extract were able to affect
disaccharidase activities, we wanted to study whether metaboIic excretory/secretory
products released by live G. lamblia trophozoites could also affect the enzyme
activities. Therefore, previously infected gerbils were challenged .')rally with O. l mg
of the in vitro-released products of the parasite. Challenging immune gerbils with the
excretory/secretory products did not affect the activities of maltase and trehalase (Table
2). However, on day 4 p.c. the activity of lactase was decreased by 27%, while there
was a 33% reduction in sucrase activity.
In addition, uninfected gerbils were inoculated with O. 1 mg of the
excretorylsecretory products, to determine whether these products could affect
disaccharidase activities in naive animais. Enzyme activities measured on day 10 p.L
(the time when activities were first reduced in the primary infe.ction) were comparable
to controllevels (data not shown).

•
•
1.
42
Table 2. Disaccharidase activities in gerbils previously exposed to G. lamblia on day 4 post-challenge with in vitro-released excretory/secretory products. Disaccharidase activities are the average values (n=5) in U/mg ± SE. Fifty days following a primary infection with 1 x 1 ()6 G. lamblia trophozoites, gerbils were challenged with 0.1 mg of the supernatant of a fully grown G. lamblia culture maintained in modified RPMI for 6 h. Controls were age-matched gerbils, previously infected with G. lamblia, which were challenged with 0.1 mg of unconditioned modified RPMI.
- = Enzyme RPMI Control Culture Supernatant
Lactase 0.015 ± 0.001 0.011 + 0.001*
Maltase 0.77 ± 0.14 0.64 ± 0.10
Sucrase 0.11 ± 0.01 0.074 + 0.009*
8Trehalase 0.0048 ± 0.0004 0.0046 + 0.0008
* Differences between control and test groups were significant, P < 0.05

•
•
•
43
DISCUSSION
Since diarrhea is a major consequence of G. lamblia infections, it is important
to examine the malabsorption underlying much of it. Unabsorbed sugars lead to
diarrhea by causing net fluid accumulation in the intestinal lumen (Launiala, 1968). In
this respect, disaccharidase deficiencies play a key role in understanding the pathogenic
mechanisms which lead to intestinal dysfunction in giardiasis. In fact, symptotns of
sugar malabsorption in other intestinal diseases/disorders are identical to several found
in Giardia infections, including abdominal distension, cramps, and flatulence, as weil
as diarrhea (Ravich and Bayless, 1983). Disaccharidase enzymes are located on or
within the microvilli of the small intestinal brush border and are involved in the final
stage of carbohydrate digestion. In this study, we focused particularJy on differences
in disaccharidase activity following various challenge inoculations in gerbils immune
to G. lamblia.
The activities of lactase, maltase, sucrase and trehalase decreased during a
primary G. lamblia infection in gerbils, beginning on day 10 p.L and remaining low
for the duration of the study (with the exception of maltase). These results differ from
the findings of Belosevic et al. (1989), who reported a decrease in trehalase activity
beginning only on day 20 p.i. Furthermore, they reported that ail disaccharidases had
recovered by day 40 p.i., except for lactase. The longer period of decreased enzyme
activity in our study may be due to the larger infective dose (l x 1()6 trophozoites) given
to each gerbil, compared to that used by Belosevic and co-workers (2 x lOS

•
•
•
44
trophozoites). It is important to note that the presence of the parasite in the small
intestine was not found to be necessary in order to maintain the disaccharidase
deficiencies. The parasite was eliminated from the gut by day 30 p.i., but the decreases
in enzyme activity persisted beyond this time. These findings are supported by work
done on malabsorption in Giardia-infected humans. Clinical studies have shown that
there are patients in which the parasite has been eradicated and yet the malabsorption
persists (Brasitus, 1983). Il is possible that these impairments of enzyme activity are
due to a combination of direct damage of enterocytes by the attachment of G. lamblia
trophozoites, followed by an effective host mucosal immune response. However, the
precise mechanism has yet to be elucidatcd.
During a challenge infection with live G. lamblia trophozoites, we also observed
decreases in the activities of lactase, maltase and trehalase. These deficiencies were of
short duration, compared to the primary infection. Although enzyme levels were
measured as early as 1 h p.c. and up to 20 days p.c., thcy were depressed only on days
2 or 4 p.c. Belosevic and colleagues (1989), also using G. lamblia-challenged gerbils,
reported lactase, sucrase and trehalase deficiencies beginning on day 1 p.c. and
remaining low for several weeks. However, their results were compared to uninfected
gerbils as controls. We chose to use gerbils which had received a primary infection,
but no challenge, as our control group. In this case, any decreases in enzyme activity
due to the primary infection were not taken into account in our analysis of the
secondary exposure to the parasite. For the disaccharidases which were affected, the
deficiencies occurred even though G. lamblia trophozoites did not establish in the gerbil

•
•
•
45
gut upon challenge. Perhaps this led to the impairments not being persistent. Several
studies have examined the link between Iltucosal damage by the parasite and intestinal
dysfunction, to explain disaccharidase deficiencies. Buret and colleagues (1992), llsing
the gerbil model for G. lamblia, concluded that a loss of brllsh border surface area led
to impaired digestion and absorption in giardiasis. Other studies, examining the
enzyme deticiencies in Giardia infections, found that villous atrophy resulted in the
decreases in disaccharidase activity (Buret et al., 1990; Khanna (II al., 1990).
However, Jennings and co-workers (1976) found humans suffering from giardiasis who
had morphologically normal mucosa and low lactase levels, and it has been establishcd
that villous shortening is only variably present in both human and experimental
giardiasis (Adam, 1991). Although the process leading to the decline in disaccharidase
activities is unknown, our results indicate that the effects are not immediate.
The challenge of immune gerbils with 0.1 mg of the soluble extract of G.
lamblia trophozoites resulted in reductions in disaccharidase acti vit y which were
comparable in terms of the levels and patterns to those induced by the live parasite in
a challenge infection. These gerbils were challenged with considerably less Giardia
protein compared to the amounts used by Belosevic et al. (1989). In this study, we
wanted to see how closely we could correlate the amount of the extract given with the
d{lsage used in the live challenge and still obtain impairments of disaccharidase activity.
\\ e decide.d lo tirst challenge the animais with 0.1 mg of the cxtract. Under the
method used for the preparation of the soluble extract, this amount was equivalent to
4 times the number of trophozoites lIsed for the live challenge. The effr,ct on the

•
•
•
46
enzymes was lost when challenging with 0.05 mg of the extract. However, our results
also show that it is possible to link the amount of the extract dose with the effects on
the disaccharidase activities. Decreasing the antigen dose used to challenge the gerbils
led to smaller enzyme deficiencies. E. histolytica, like G. lamblia, is an intestinal
prOlozoan parasite which produces energy by fermentation. They are two of the few
eukaryotes with an anaerobic metabolism (Adam, 1991). Despite these similarities,
disaccharidase activities were not affected when gerbils previously exposed to G.
lamblia were challenged with a soluble extraet of E. histolytica trophozoites. A study
using an in vitro rabbit model of acute intestinal amebiasis found that exposing colon
segments to E. histolytica trophozoite Iysates resulted in distinct mucosal damage
(Navarro-Garcfa et al., 1993). Therefore, the inability of the E. histolytica extract to
induce reductions in enzyme aetivity in our study quite strongly suggests that the
decreases observed with a Giardia challenge are specifie to this parasite.
The fact that we did obtain reduetions ln disaecharidase activity in Giardia
extract-challenged gerbils indieates that the live parasite is not needed and suggests that
there may be an immunologieal basis for the decreases. Specifie G. lamblia surface
antigens, which are not found on E. histolytica trophozoites, may be targets of the anti
Giardia immune response. Other researchers, using a murine model, have found that
T eells, upon reaeting with an antigen, can release lymphokines whieh damage
rmcrocytes (Mowat and Ferguson, 1981). Such damage could result in a decline in
disaccharidase activities. However, although T cells are known to play a role in the
immune response to Giardia, this process has yet to be examined in giardiasis .

•
•
•
47
Macrophage interaction with Giardia antigens could also affect enzyme levels, since
mediators su ch as superoxide radicals and lysosomal proteases, released by
macrophages, have been implicated in mucosal damage in G. lamblia-infected mice
(Goyal et al., 1993).
The results obtained with the in vitro-released excretory/secrctory prodllcts of
the parasite were not consistent with the live parasite challenge and the Giardia extract
challenged findings. Sucrase activity was decreased, whercas it was not affccted by a
ch?Jlenge with either the live parasite or the soluble trophozoite extract. Similarly,
maltase and trehalase were not impaired by a challenge with the metabolic
excretory/secretory products, yet they had been when the parasite was llsed. These
results suggest that Giardia metabolic products do not likely play a major role in the
disaccharidase deficiencies observed in giardiasis. Perhaps these t~xcretory/secretory
products contain proteolytic activity which affects the enzymes in a differential manncr.
The excretory/secretory products have been reported to include antigenic, slirfacc
derived mateT/al, with a major polydisperse substance (Nash el al., 1983). Howcvcr,
the presence of a proteinase has not been detcrmined. Flirthermore, althuligh intestinal
damage related to a Giardia exotoxin is an attractive hypothesis, there is no evidcncc
of such a product (Chavez et al., 1986). Further characterization of the effects of the
excretory/secretory products on enzyme activity, as weil as the components of these
products, is necessary.
The data presented here demonstrate that the disaccharidase deficiencies in
gerbils previously exposed to G. lamblia are a parasite-specific cffect and can occur in

•
•
•
48
response to antigen challenge in a dose-dependent manner. Although there are
indications that the deriine in disaccharidase levels in the challenge infection is the
result of the host's mucosal immune response to a Giardia antigen(s), the precise
mechanism by which Giardia produces diarrhea and malabsorption have yet to be
characlerized. Specifically, therc is a need to find the mediators involved in this
process .

•
•
•
49
REFERENCES
Adam R.D. 1991. The biology of Giardia spp. Microbiol. Rev. 55: 706-732.
Belosevic M. and Faubert G.M. 1983. Giardia muris: correlation between oral dosage, cOllfse of infection and trophozoite distribution in the mOllse small intestine. Exp. Parmitol. 56: 93-100.
Belosevic M., Faubert G.M. and Mac1ean J.O. 1989. Disaccharidase activity in the small intestine of gerbils (Meriones un~uiculalus) during primary and chalienge infections with Giardia lamblia. GUI 30: 1213-1219.
Belosevic M., Faubert G.M., Mac1ean J.O., Law C. and Croll N.A. 1983. Giardia lamblia infections in Mongolian gerbils: an animal mode\. J. Infi'ct. Dis. 147: 222-226.
Brasitus T.A. 1983. Parasites and malabsorption. Clin. Gastromterol. 12: 495-510.
Buret A., Gall D.G., Nation P.N. and OIson M.E. 1990. Intestinal prolozoa and epithelial ceU kinetics, structure and function. Parasitol. Today 6: 375-380 .
Buret A., Hardin J.A., Oison M.E. and Gall D.G. 1992. Pathophysiology of small intestinal mal absorption in gerbils infected with Giardia lamblia. Gastroenterology 103: 506-513.
Chavez B., Knaippe F., Gonzalez-Mariscal L. and Martinez-Palomo A. 1986. Giardia lamblia: electrophysiology and ultrastructure of cytopathology in culture.d epithelial cells. Exp. Parasitol. 61: 379-389.
Craun a.F. and Jakubowski W. 1987. Status of watel'borne giardiasis outbreaks and monitoring methods. In: Procel~dings o/the International Symposium on Wa/(>r Related Hea/th Issues. Tate C.L. Jr. (ed.). American Watei Resources Association: Bethesda, pp. 167-174.
Dahlqvist A. 1968. Assay of intestinal disaccharidases. Anal. Biochem. 22: 99-107.
Dahlqvist A. and Semenza G. 1985. Disaccharidases of srnall-intestinal mucosa. J. Pediatr. Gastroenterol. Nulr. 4: 857-865.
Daniels C.W. and Belosevic M. 1992. Oisaccharidase activity in the small intestine of susceptible and resistant mice after primary and challenge infections with Giardia muris. Am. J. Trop. Med. Hyg. 46: 382-390 .
Diamond L.S., Harlow D.R. and Cunnick C.C. 1978. A new medium for the axenic

•
•
•
50
cultivation of Entamoeba hiltolytica and other Entamoeba. Trans. R. Soc. Trop. Med. Hyp,. 72: 431-432.
Ferguson A., Gillon J. and Ai Thamery D. 1980. Intestinal abnormalities in murine giardiasis. Trans. R. Soc. Trop. Med. Hyg. 74: 445-448.
Gillin F.D. and Oiamond L.S. 1979. Axenically cultivated Giardia lamblia: growth, attachment and the role of L-cysteine. In: Waterborne Transmission of Giardiasis. Jakubowski W. and HoffJ.C. (OOs.). U.S. Environmental Protection Agency: Cincinnati, pp. 270-272.
Goda T. and Koldovsky O. 1988. Dietary reglilation of small intestinal disaccharidases. Wld. Rev. NUlr. Diel. 57: 275-329.
Goyal R., Mahajan R.C., Ganguly N.K., Sehgal R., Gorowara S. and Singh K. 1993. Macrophage-mediated enterocyte damage in BALBlc mice infected with different strains of Giardia lamblia. Scand. J. Gastroenterol. 28: 845-848.
Guy R.A., Bertrand S. anrl Faubert G.M. 1991. Modification of RPMI 1640 for use in in vitro immur,ological studies of host-parasite interactions in giardiasis. .1. Clin. Microbici. 29: 627-629.
Hartong W.A., Gourley W.K. and Arvanitakis C. 1979. Giardiasis: clinical spectrum and fllnctional-structural abnormalities of the small intestinal mucosa. Gastroenterology 77: 61-69.
Jennings W., Rowland R., Hecker R., Gibson G.E., Fitch R.J. and Reid D.P. 1976. The significance of lower jejunal disaccharidase levels. Aust. N.Z. J. Med. 6: 556-560.
Khanna R., Joshi K., KumKum, Malik A.K. and Vinayak V.K. 1990. An ultrastructural analysis of changes in surface architecture of intestinal mucosa following Giardia lamblia infection in mice. Gastroenterol. Jpn. 25: 649-658.
Launiala K. 1968. The effect of unabsorbed suc rose and mannitol on small intestinal flow rate and mean transit time. Scand. J. Gastroenterol. 3: 665-671.
Lewis P.D. Jr., Belosevic M., Faubert G.M., Curthoys L. and Mac1ean J.O. 1987. Cortisonc-induced recrudescence of Giardia lamblia infections in gerbils. Am. J. Trop. Med. Hyg. 36: 33-40.
Lowry O.H., Rosebrough NJ., Farr A.L. and Randall R.J. 1951. Protein measurements with folin phenol reagent J. Biol. Chem. 193: 265-275.

•
•
•
51
Mowat A.MeL. and Ferguson A. 1981. Hypersensitivity reaetions in the small intestine. 6. Pathogenesis of the graft-versus-host reaetion in the small intestinal mueosa of the mouse. Transplalllation 32: 238-243.
Nash T.E., Gillin F.O. and Smith P.D. 198j. Excretory-secretory products of Giardia lamblia . ./.lmmunol. 131: 2004-2010.
Navarro-Garcia F., L6pez-Revilla R. and Tstusumi V. 1993. Dose- and time-dependent functional and structural damage to the colon mucosa by EntamOl'ba histolytica trophozoite lysates. P{lfasitol. Res. 79: 517-522.
Ravich W.J. and Bayless T.M. 1983. Carbohydrate absorption and malabsorption. Clin. Gasfroenterol. 12: 335-356.
Smith P.D., Gillin F.D., Spira W.M. and Nash T.E. 1982. Chronic giardiasis: studies on drug sensitivity, toxin production and host immune responsc. Gastroenterology 83: 797-803.
Wilson H.S.P., Stauffer S.J. and Walker T.S. 1982. Waterborne giardiasis outbrcakAlberta. Cano Dis. Wkly. Rpt. 8: 97-98.
Wright S.G. and Tomkins A.M. 1977. Quantification of the lymphocyte infiltrate in jejunal epthelium in giardiasis. Clin. Exp. Immunol. 29: 408-412.
Yardley J.H., Takano J. and Hendrix T.R. 1964. Epithelial and other mucosallesions of the jejunum in giardiasis. Jejunal biopsy studies. Bull. Johns Hopkins Ho.\'p. 115: 389-406.

•
•
•
52
CONNECTING STATEMENT
ln Manuscript l, the effects on disaccharidase activity in gerbils during G.
lamblia infections was described. Il was found that challenging previously-infected
animais with a erude soluble extract of G. lamblia trophozoites resulted in decreases
in enzyme activity similar to those obtained with a live challenge infection. Therefore,
in the study presented in Manuscript 2, an attempt was made to purify the specifie
fraction of the trophozoite extract responsible for these disaccharidase deficiencies .

•
•
•
MANUSCRIPT 2
IDENTIFICATION OF A FR.ACTION OF
GIARDIA LAMBLIA TROPHOZOITE EXTRA CT
ASSOCIATED WITH DISACCHARIDASE
DEFICIENCIES IN MONGOLIAN GERBILS
Shawn R. Mohammed and G. M. Faubert
Institute of Parasitology
McGiII University
Macdonald Campus
21 ,111 Lakeshore Road
Ste. Anne de Bellevue, Quebec
Canada H9X 3V9
(submitted for publication)
53

•
•
•
54
ABSTRACT
The effects on disaccharidase activities of challenging gerbils previously exposed
to Giardia lamblia with fractions of the crllde trophozoite extract were examined. Gel
filtration by Sephacryl S-200 HR chromatography of the soluble extract rcsulted in 3
major protein fractions (FI, F2, F3). Only a challenge with fraction FI (0.1 mg total
dose) was found to induce disaccharidase deficiencies. Boiling FI prior to challenge
did not change this effect on the enzyme activities. However, the decTeases were not
obtained when the total FI dose was reduced to 0.05 mg. Column chromatography of
fraction FI under dissociating and reducing conditions resulted in 2 further fractions:
Fla and Flb. Challenging immune gerbils with Flb led to impairments of
disaccharidase activity similar to those obtained with FI. A challenge with Fla had no
effect on disaccharidase ac:tivities. Protein analysis of the crude extrac~, as weIl as the
fractions of the extract, revealed severa! high and low moleclliar weight bands, with
proteins of the cTude extract ranging from 14 to 200 kDa. These findir.gs indicate that
a constituent(s) of fraction Flb is the portion of the parasite which induces
disaccharidase deficiencies in immune gerbils. This fraction consists of proteins
ranging in molecular weight from 32 to 200 kDa. In addition, the G. lamblia fraction
involved in the decreases in enzyme activity is heat-stable .

•
•
•
55
INTRODUCTION
Giardia lamblia is a gastrointestinal parasite of humans and other mammalian
species, with a worldwide distribution. It is a common cause of diarrhea in day-care
centres, institutionalized persons, homosexuals and travellers to developing nations
(Key stone el al., 1978; Brodsky el al., 1974; Schmerin ('1 al., 1978). This tlagellated,
binucleate parasite adheres to the brush border of the host's enterocytes by means of
a ventral sucking disk. In the gut lumen, the trophozoites differentiate into the
intermediate encysting trophozoite form and then to cysts (Campbell and Faubert,
1994). One nuclear division OCCUTS in the cyst or during its formation, so that
tetranucleate cysts are excreted with the feces. Many giardial infections are weil
tolerdted and asymptomatic in human patients. However, wh en iIIness is evident, the
spectrum of symptoms is wide and ranges from acute, self-limiting gastroenteritis to
protracted and debilitating malabsorption (Shandera, 1990).
In giardiasis, it has been established that there can be considerable
malfunctioning of the epithelium of the small intestine. For example, fat and vitamin
B12 malabsorption has been described in human giardiasis (Hoskins et al., 1967; Wright
et al., 1977). Vitamin A deficiency has been found in children infected with G.
lamblia (Mahalanabis el al., 1979). Also, temporary disaccharidase deficiencies have
becn well documented in this disease. Buret et al. (1990) reported decreases in maltase
and sucrase activities in mice, following a primary infection with G. murÎ.'l. ft was
suggested that these disaccharidase deficiencies were due to a diffuse shortening of

•
•
•
56
brush border mierovilli. Studies on G. lamblia infections in mice found decreases in
several brush border enzymes throughout the eour~e of infection (Nain et al., 1991).
The degree of decline in these enzyme aetivities correlated weIl with the number of
trophozoites in the jejunum, with the most severe reductions in enzyme activity
oceurring during the peak phase of infection. Using the gerbil as an animal model,
Belosevic and eol!eagues (1989) observed decreases in disaceharidase activity during
both primary and challenge infections with G. lamblia. The iovolvement of the host's
immune response in the deficiencies was suggested by the ability of a erude extraet of
the trophozoites to ioduee these impairments of enzyme activity in immune gerbils
(Belosevic et al., 1989). Recent work in our laboratory has confirmed these previous
findings (Manuseript 1). We also showed that decreasing the extract dose used to
challenge the gerbils led to smaller reduetions in enzyme aetivity. In addition, it was
determined that the effects of the extraet in a challenge inoculation are specifie to the
parasite, since challenging animals previously exposed to G. lamblia with a erude
extract of Entamoeba histolytica did not result in impairments of enzyme activity
(Manuscript 1). However, the component(s) of the G. lamblia crude extract involved
in the disaccharidase deficicncies, as weIl as the mechanism(s) leading to sueh
reductions in enzyme activity, are unknown. G. lamblia is consldered to be
antigenically complex, yet little is known about the structures and properties of these
antigens (Chaudhuri et al., 1988).
Cevallos and Farthing (1992) reported a strain-dependent reduction in
disaeeharidase aetivities in G. lamblia-infected rats. The antigenic differences between

•
•
•
57
these strains may have led to the variable effects on the enzyme activities. The present
study was undertaken in order to identify the specifie fraction of the G. lamblia crude
antigen extract responsible for inducing the disaccharidase deficiencies observed in
immune gerbils. ln addition, we undertook an initial characterization of this fraction .

•
•
•
58
MATERIALS AND METROnS
Parasites
G. lamblia trophozoites, WB strain (American Type Culture Collection no.
30957), originally isolated from a symptomatic patient (Smith et al., 1982) were used
throughout this study. Trophozoites were cultured axenically in filter-sterilized TYI-S-
33 (trypticase, yeast extract, iron, and serum) medium adapted for G.lamblia (Gillin
and Diamond, 1979), wW, 10% adult bovine serum (Sigma Chemical Co., St. Louis,
U.S.A.), and supplemented with 100 units/mL of penicillin (Sigma) and 100 Ilg/mL of
dihydrostreptomycin sulphate salt (Sigma), in the absence of bovine bile. Twice
weekly, the trophozoites were passaged.
Animais
Six to 10 week old male Mongolian gerbils (Meriones unguiculatus) , originating
from Tumblebrook Parms (West Brookfield, U.S.A.), were used in this study. They
were maintained under standard laboratory conditions and provided with laboratory
chow and water ad libitum. One week after arrivai, gerbils were each tre.ated with 15
mg of metronidazole (Rhône-Poulenc, Montreal, Canada), by oral gavage, for 3
consecutive days and th en allowed to rest for 10 days. This treatment was done to
ensure that the gerbils were free of intestinal infections.
AU inoculations were done orally, to unanaesthetized gerbils. In the primary

•
•
•
59
infection, each gerbil was inoculated with 1 x 1()6 live G. lamblia trophozoites.
Preparation of the Crude Extract of G. lamblia Trophozoites
Trophozoites (from a 72-h culture) were dislodged from the wans of culture
vessels by immersion in an ice bath for 15-30 min. They were sedimented by
centrifugation at 800 g for 10 min, at 4° C, and washed 5 times in sterile phosphate
buffered saline (PBS) (pH 7.2). Trophozoites, suspended in sterile PBS, were th en
lysed by discontïnuous sonication in an ice bath, using a Sonic Dismembrator (Fisher
Scientific, Montreal, Canada) at maximum output, for 10 min. The sonicated material
was cleared of insoluble matter by centrifuging at 23,300 g for 20 min, at 4()C, and the
supernatant collected as the crude soluble extract.
Column Chromatography
Samples were fractionated by gel filtration chromatography. For this purpose,
approximately 80 mL of preswollen Sephacryl S-200 HR gel (Pharmacia LKB
Biotechnology, Uppsala, Sweden), wet bead diameter 25-75 #-tm, were packed into a 1.8
x 50 cm glass column (Bio-Rad, Mississauga, Canada). The void volume of the
column was determined by applying Blue Dextran 2000 and the column calibrated with
known molecular weight (MW) marker of gel filtration proteins (Pharmacia). For each
run, at 4° C, 1-2 mL of sample (with 10% glycerol) was applied to the gel bed and a
flow rate of 5 mL/h maintained using a peristaltic pump (Pharmacia). Fractions of 2
mL each were collected and an elution profile obtained by measuring the optical density

•
•
•
60
(0.0.) at 280 nm and plotting these values against the fraction numbers. According
to the 0.0. values, e.}(·h peak and trailing eluates were pooled separately. These
pooled fractions were th en dialyzed, using Spectra/Por 6 Membrane (MW Cut-Off:
1(00) (Spectrum Medical Industries Inc., Los Angeles, U.S.A.), against distilled water
for 24 h, at 4° C. Following dialysis, fractions were concentrated by lyophilization and
resuspended in PHS. The fractions were stored at - 70° C until used.
Sodium Dodecyl Sulfate-Polyacrylamide Gel Electrophoresis
(SDS-PAGE)
Samples were electrophoresed in a Mini-PROTEAN II vertical slab cell (Bio
Rad) using the discontinuous system of Laemmli (1970). Protein samples were
separated through a 4% stacking gel and 10% separating gel. Bio-Rad molecular
weight standards were run simultaneously. The gel was stained with 0.25% Coomassie
Brilliant Blue R-250 dye (Bio-Rad).
Measurement of Intestinal Disaccharidases
- Preparation of Intestinal Homogenate -
Homogenates were prepared as previously described (Belosevic et al., 1989),
with modifications. Briefly, the gerbils were killed and the small intestine removed and
divided into three sections. The segments were placed in ice cold distilled water and
each was flushed with 50 mL of distilled water to clear it of intestinal debris. The
segments were then slit longitudinally and the mucosa scraped off with a glass

l • 61 1
microscope slide. The mucosa from all three sections were combined, weighed (wct
weight), and placed in four volumes of ice cold distilled water. The mucosal scrapings
were then homogen\zed using a Con Torque power unit at maximum speed (Eberbach
Corp., Ann Arbor, U.S.A.). Homogenates were stored, wlthout prior centrifugation.
at - 7(1 C until used in the assay.
- Assay for Intestinal Disaccharidases -
Disaccharidase activity of homogenated mllcosa was measured using the glucose
oxidase peroxidase assay of Dahlqvist (1968), as modified by BelosevÎC el al. (1989),
which is based on a colollr reaction with the glucose liberated by a particular
disaccharidase in 60 min. The assay consisted of adding to each weil of a 96-well
• Nunc microwell plate (Gibco BRL, Burlington, Canada) 0.01 mL of appropriately
diluted mucosal sample ~homogenate) and 0.01 mL sub~trate-buffer solution of a
disaccharidase to be measured, in quadruplicate. The plate was th en incubatcd at 37"
C in humidified atmosphere for 60 min. After incubation, 0.3 mL tris-glucose oxidase
reagent was added to each well and the plate incubated for an additional 60 min. The
plate was then read at 415 nm using a Bio-Tek microplate reader (Mandel Scicntitic,
Guelph, Canada). For each assay, 8 wells of reagent blank and a glucose standard
series (2, 6, and 10 Ilg glucose) in quadruplicate were also donc. Disaccharidase
activity is expressed as units/mg protein in the mucosal sample (U/mg), where units
represent f,tmoles of disaccharide hydrolyzed/min.
- Reagents -
• AlI chemicals were obtained commercially. 0-( + )-G1ucose, fi-lactose, maltose,

•
•
•
62
D-( + )-trehalose, maleic acid, glucose oxidase, o-dianisidine, peroxidase, and triton x-
100 were purchased from Sigma. Sucrose was obtained from Fisher Scientific and tris
(ultra pure) was purchased from ICN Biomedicals Canada Ltd. (St. Laurent, Canada).
Measurement of Protein Concentration
PrC'tein concentrations were determined as described by Lowry et al. (1951),
using 0.2 mL of sample and a final reaction volume of 1.3 mL. For each protein
as say , a standard curve was prepared using freshly dissolved bovine serum albumin
(BSA).
Statistical Analysis
Statistical significance was determined using the Mann-Whitney U-test.
Significancc was assigned at the probability level of P < 0.05 .

•
•
•
63
RESULTS
Fractionation of the Soluble Extract of G. lamblia Trophozoites
The crude soluble extract of G. lamblia trophozoites was subjected to column
chromatography using Sephacryl S-200 HR gel filtration in order to fractionate its
components. Three light-absorbing peaks were observed, one of which appeared in the
void volume and another at the end of total column volume (Fig. 1). Eluted materials
were cotlected to give fractions FI (> 150,000 MW), F2 (- 67,000 MW), and F3
« 14,000 MW), as indicated in Fig. 1. These were pooled from fractions 16-22, 23-
29, and 34-43, respectively.
Challenge with the Fractions of the G. lamblia Soluble Extract
To determine whether any one of the fractions of the soluble extract (described
above) could affect disaccharidase activity in gerbils previously exposed to G. lamblia,
each animal was challenged with 0.1 mg of FI, F2, or F3. The animais wcrc
challenged on day 50 post-infection, which is past the elimination of the parasite from
the gerbil small intestine (Manuscript 1). The activities of the enzymes lactase,
maltase, sucrase, and trehalase were then measured on days 2 and 4 post-challenge
(p.c.), as these were the days on which disaccharidase activity was affecte.d during
challenge inoculations with the whole crude extract (Manuscript 1). As shown in Fig.
2, the activity of sucrase was not affected by any of the 3 fractions of the extract, as

•
•
•
64
~ c::I ~
~ fil ~
rQ -< ~ = 1 1 1 1 0.8 Ft 0.6 t-------t
~ 0.4 ,...., a 0.2
~ 0.09
0 0.08 ~ N ........ 0.07 ~
o+J 0.06 .,.. rn ~ 0.05 4> ~ ..... 0.04-as (J 0.03 .,..
o+J ~ 0.02 0
0.01
0.00 5 la 15 20 25 30 35 40 45 50
Fraction Number
Fig. 1. Chromatograph of the soluble extract of sonicated G. lamblia trophozoites. Approximately 1.8 mg of the soluble extract were run through a Sephacryl S-200 HR column at a flow rate of 5 mLlh, using a buffer of 0.15 M PBS, 0.02% sodium azide (pH 7.2). Fractions were collected and pooled to give FI, F2, and F3, as indicated. The column was calibrated with Blue Dextran 2000 (BD; > 2 x 106 MW), Aldolase (Ald; 158,000 MW), Bovine Serum Albumin (BSA; 67,000 MW), and Ribonuclease A (RnA; 13,700 MW) .
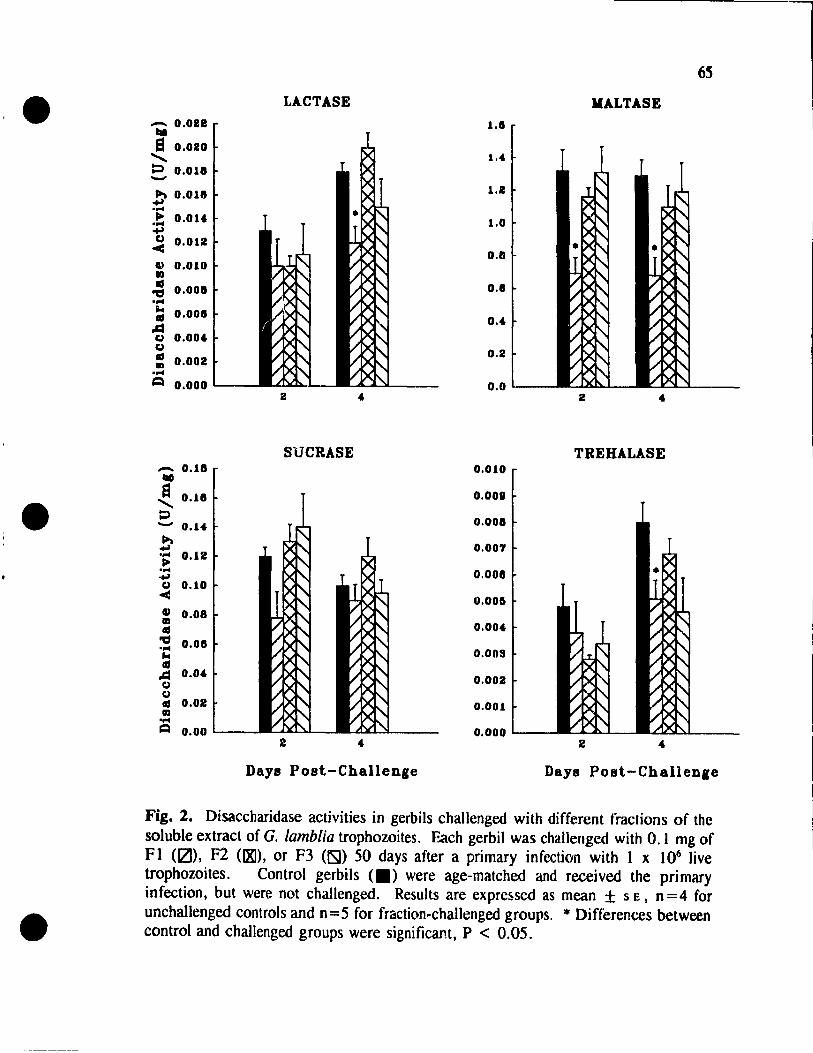
•
•
•
- 0.012 taG S 0.010
....... P 0.018 -~ 0.018 '004
.!: O.OU ~
~ 0.012
" 0.010 ID Ils
'1:1 .... ... Ils .cl g g Ils ID .... =
0.008
0.008
0.00'
0.002
o.ooo~-
_ 0.18 '-II
~ 0.18 ;:, - 0.1'
~ .... 0.12 ~ ....
+1 Co) 0.10 < : 0.08 at ~ 0.08 ... as .cl 0.0.
Co) Co) Ils 0.02 ln .... ~ 0.001---
LACTASE
2
SUCRASE
2
Days Post-Challenge
65
MALTASE
1.8
1.' 1.2
1.0
0.8
0.8
o .•
0.2
O.OL_-2
TREHALASE 0.010
0.0011
0.008
0.007
0.008
0.000
0.00'
0.003
0.002
0.001
0.0001---2
Days Post-Challen,e
Fig. 2. Disaccharidase activities in gerbils challenged with different fractions of the soluble extract of G. lamblia trophozoites. Each gerbil was cha.llenged with 0.1 mg of Ft (IZ]), F2 (IXI), or F3 (ISI) 50 days after a primary infection with 1 x 106 live trophozoites. Control gerbils <.) were age-matched and received the primary infection, but were not challenged. Results are expressed as mean ± SE, n =4 for unchallenged controls and n =5 for fraction-challenged groups. * Differences between control and challenged groups were significant, P < 0.05.

•
•
•
66
levels on both days 2 and 4 p.c. remained comparable to controls. However, the
activities of lactase, maltase, and trehalase were reduced by as much as 47% on day
4 p.c. in Fl-challenged gerbils. In addition, an FI challenge also led to a deerease in
maltase activity on day 2 p.c. On the other hand, challenging immune gerbils with
either F2 or F3 had no effeet on any of the disaccharidase activities.
In order to determine whether the effeet of fraction FIon enzyme activities is
dose-dependent, immune gerbils were challenged with 0.075 mg or 0.05 mg total of FI.
Disaccharidase activity was th en measured on day 4 p.c., the time when the most
extensive effects on the enzymes were observed with the 0.1 mg FI challenge. Sucrase
activity remained unaffeeted by lhis fraction of the soluble extract, regardles~ of the
dosage (Fig. 3). However, lactase and maltase activities were significantly reduced
with an FI ,Jose of 0.075 mg, compared to unchallenged controls. These decreases in
disaccharidase activity were similar to the reduced levels observed in the gerbils
challenged with 0.1 mg of FI. There were no reductions in the activities of any of the
4 enzymes when an FI dose of 0.05 mg was given to the animais.
Gerbils which had a prior Infection with G. lamblia were also challenged with
0.1 mg of FI which had previously been boiled at 100° C for 5 min. Disaccharidase
activity was then measured on day 4 p.c. and compared to age-matched control animais
which had reeeived a primary infection, but were not challenged. Deereases in lactase,
maltase, and trehalase activities were deteeted, while sucrase activity did not
significantly differ from control levels (data not shown). These findings were similar
to those obtained with FI which was not boiled.

•
•
•
_ 0.011 III El 0.010
....... P 0.0111 -~ O.OUI +' .... .. 0.010& .... ..., ~ 0.012
CU 0.010 la
~ 0.0011 .... :; 0.0011
~ 0.000& C)
= 0.002 .... .::1 0.000
_ 0.10& lai S ~ 0.12 '-'"
b 0.10 .... ~ .... t 0.011 ..:
CU la 0.01 .. ~ .... J.4 0.00& .. ~ C) CJ 0.02 .. la ....
.::1 0.00
LACTASE
CONTROL 0.1 0.075 0.06
SUCRASE
CONTROL 0.1 0.076 0.011
Fraction Fl DOlic (ml)
67 MALTASE
1.11
1.0&
1.2
1.0
0.8
0.1
0.0&
0.2
0.0 CONTROL 0.1 0.0711 0.011
TREHALASE 0.010
0.001
0.008
0.007
0.001
0.0011
0.000&
O.OOS
0.002
0.001
0.000 CONTROL 0.1 0.076 0.01
Fraction Fl Doae (ml)
Fig. 3. Disaccharidase activities in gerbils challengcd with varying amounts of fraction FI of the soluble extract of G. lamblia trophozoites. Gerbils were challenged 50 days following a primary infection with 1 x 106 live trophozoites. Control gerbils were agematched and received a primary infection, but were !lot challenged. Results are for day 4 post-challenge and are exprcssed as mean ± S.E , n =4 for unchallenged controls (.> and n=5 for Fl-challenged groups (~). * Differences between control and challenged groups were significant, P < 0.05.

•
•
•
68
Fractionation of FI
In order to further purify the fraction of the G. lamblia extract responsible for
inducing the disaccharidase deficiencies, fraction FI was subjected to gel filtration
chromatography using a Sephacryl S-200 HR column. A buffer consisting of the
dissociating agent guanidine Hel (Sigma) and the reducing agent DL-dithiothreitol
(Sigma) was used to achieve further separation of FI, by disrupting non-covalent
interactions and breaking disulfide bonds in this fraction. Two peaks were observed,
one of which eluted with the void volume and the other also appeared in the high
molecular weight (> 67,000 MW) region of the separation (Fig. 4). Fractions 15-19
and 20-25 were collected separately to give pooled fractions Fla and Flb, respectively.
Challenge with Fractions Fla and Flb
To determine whether disaccharidase activity could be affected by one of the
fractions of FI (obtained as described above), immune gerbils were each challenged
with 0.1 mg total of either Fla or Flb. Enzyme activities were then measured on day
4 p.c. The activities of ail 4 enzymes remain<~ comparable to controllevels in Fla
challenged animais (Fig. 5). However, challenging with fraction Flb resulted in
significant decreases in the activities of lactase, maltase, and trehalase. These
reductions ranged from 39% to 71 %. Sucrase activity was also unaffected by an Flb
challenge.
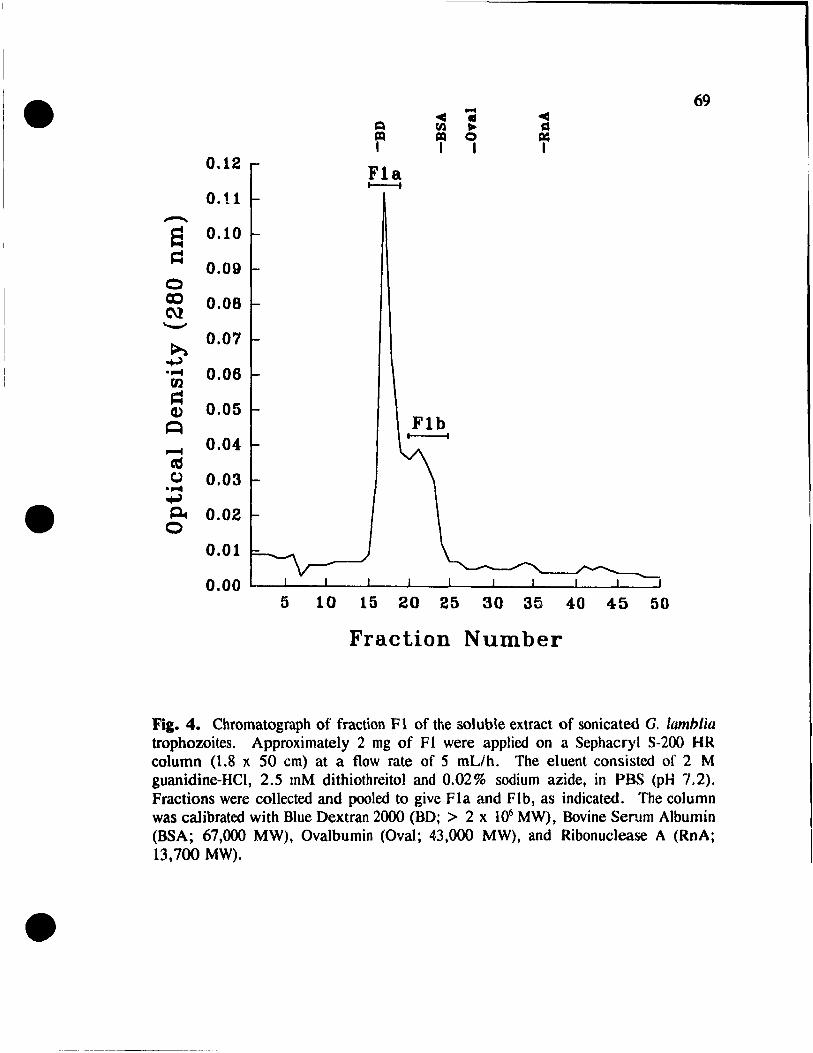
•
•
•
69 ~ - ~ Il
Q fil ~ Â IZI IZI Q = 1 1 1 1
0.12 Fla t---t
0.11 -... a 0.10
= 0.09 0 cc 0.08 ru '-'
~ 0.07
+J ...... 0.06 rn ~
0.05 Q)
~ Flb
~ 0.04-
= t.> ...... 0.03 ~ ~ 0.02
0 0.01
0.00 5 10 15 20 25 30 35 40 45 50
Fraction Number
Fig. 4. Chromatograph of fraction FI of the soluble extract of sonicated G. lamblia trophozoites. Approximately 2 mg of Ft were applied on a Sephacryl S-200 HR column (1.8 x 50 cm) at a f10w rate of S mL/ho The eluent consisted of 2 M guanidine-HCl, 2.5 mM dithiothreitol and 0.02% sodium azide, in PBS (pH 7.2). Fractions were collected and pooled to give Fla and Flb, as indicated. The column was calibrated with Blue Dextran 2000 (BD; > 2 x 106 MW), Bovine Serum Albumin (BSA; 67,000 MW), Ovalbumin (aval; 43,000 MW), and Ribonuclease A (RnA; 13,700 MW).

•
•
•
_ 0.010 ... S 0.011
"'" ~ _ 0.011
!' 0.01" .... ~
::: 0.012 g
~ 0.010 CU : 0.001 ." .... ~ .. ~ CI CI .. ., .... ~
0.001
0.001
0.000
_ 0.11 III S 0.10
'"', ~ 0.01 -!' 0.01 .... ~ 0.07 .... .., ~ 0.01
CU 0.05 ., .. ." .... ~ .. ~ CI CI .. ., .... ~
0.0"
o.os
0.01
0.01
0.00
CONTROL
CONTROL
70 LACTASE MALTASE
l."
l.2
l.0
• 0.8
0.1
0.4
0.8
0.0 'la 'lb CONTROL ria 'lb
SUCRASE TREHALASE 0.001
0.001
0.007
0.008
0.001 •
Il.003
1~.002
0.001
0.000 rla 'lb CONTROL rla 'lb
Fraction Fraction
Fig. 5. Disaccharidase activities in gerbils challenged with fractions Fla and Flb. Gerbils were each challengro with 0.1 mg of Fla or Flb 50 days following a primary infection with 1 x 1()61ive trophozoites. Control gerbils were age-matched and received a primary infection, but were not challenged. Results are for day 4 post-challenge and are expressed as mean ± SE, n=4 for unchallenged controls <.) and n=5 for FlalFlb-challenged groups (181). * Differences between control and challenged groups were significant, P < 0.05.

•
•
•
71
SDS-PAGE
In order to analyze the protein constituents of the crude extract of sonicated G.
lamblia trophozoites and of the partially purified extract fractions, samples were
compared following SDS-PAGE. Several high and low molecular weight bands were
seen (Fig. 6). The erude extraet showed approximately 20 discernable protein bands,
with molecular weights ranging from 14 to 130 kDa. However, the banding pattern of
fraction FI was in the 19 to 200 kDa range and F2 was in the smaller regiol1 of 30 to
55 kDa. Fraction F3 showed only one protein band, of 20 kDa. For both Fla and
Flb, a number of bands were detected, mai ni y in the molecular weight region of 32 to
200 kDa. However, there were 2 faint yet discrete bands seen in fraction Fla which
were absent from Flb. These had molecular masses of 25 kDa and 27 kDa (arrows).
In addition, major bands of 30 kDa and 33 kDa were secn in ail samples, with the
exception of fraction F3.

•
•
•
kDa 200-
97.4-
21.5-
1 2 3
72
4 5 6
Fig. 6. SDS-PAGE of the crude extract of G. lamblia trophozoites and partially purified fractions of the extract. The gel was stained with Coomassie brilliant blue R-250. Molccular weight standards are shown on the left. Lane 1: erude extract; 2: FI; 3: F2; 4: F3; 5: Fla; 6: Flb .

•
•
•
73
DISCUSSION
G. lamblia is the most corn mon intestinal protozoan parasite of humans and the
acute to chronic diarrhea which it can cause is often associated with intestinal
malabsorption (Farthing, 1992). Even when Giardia is not sllspected, cases of
malabsorption may be due to latent giardiasis, as the organism can be found in the
intestine withoutappearing in feces (Kamath and Murugasu, 1974). The disaccharidase
deficiencies which contribute to the malabsorption of nutricnts in Giardia infections are
therefore an important area of investigation.
In this study, we examined the contribution of the parasite to host malabsorption
of disaccharides in giardiasis. Our chromatographie profile of Giardia protcins is
supported by work done by Chaudhuri and colleagues (1988), who obtained a similar
3·peak separation of the G. lamblia soluble extract, using Sephacryl S-300 gel fi Itration.
Moreover, these researchers found that maximum antigenic activity, as determined by
counter immunoelectrophoresis, was associated with their high molecular weight
fraction, which eluted in the void volume and corresponds to our fraction FI.
Following separation of the G. lamblia soluble extract through Sephacryl S-200 HR
chromatography, we wer~ able to clearly implicate fraction FI in the disaccharidasc
deficiencies in immune gerbils. The patterns of lactase, maltase and trehalase
reductions on days 2 and 4 p.c. in the FI-challenge mimic those obtained in previous
studies with live- and extract-challenged animaIs (Manuscript 1). The fact that we were
able to decrease the amount of F l used to challenge gerbils and lose the effect on ail

•
•
•
74
the enzymes with a 0.05 mg dose, as was found in an identical study using the cru de
ex tract (Manuscript 1), served to confirm the importance of fraction FI in the
disaccharidase deticiencies. In addition, the constituent of FI involved in the
impairments of disaccharidase activity is heat-stable since the effects of Fion the
enzyme activities were not altered by boiling FI prior to the challenge.
We were able to partially purify FI into 2 fractions, Fla and Flb, again usillg
a Sephacryl S-200 HR column. The use of an eluent containing the dissociating agent
guanidine HCI and the reducing agent dithiothreitol allowed us to disrupt non-covalent
interactions and break disulfide bonds in FI, and therefore further separate giardial
proteins. The ability of a challenge with Flb, but not Fla, to induce the sa me
disaccharidase deficiencies in immune gerhils as when challenging with FIor the
complete trophozoite extract (Manuscript 1), indicates that we have isolated a fraction
of the parasite with a constituent(s) that is intimately linked to disaccharide
malabsorption.
The complexity of the soluble extract of G. lamblia was demonstrated by the
SDS-PAGE protein analysis. The present study revealed that approximately 20 protein
bands, ranging from 14 to 130 kDa, cou Id be detected. These findings are supported
by those of other researchers, who have reported between 20 to 28 protein bands with
molecular weights ranging from a low of 10 kDa up to 140 kDa (Moore et al., 1982;
Smith et al .• 1982; Chaudhuri et al .• 1988). We also observed sorne very faint bands
in the rcgion between 130 and 200 kDa. Such poorly discernable high molecular
weight bands have been reported by Smith and colleagues (1982), as weIl. Using an

•
•
•
75
extract of the Pl strain of G. lamblia trophozoites, Kumkum and co-workers (1988)
were able to clearly detect bands with molecular weights up to 205 kDa. Such
differences in the bands observed through SDS-PAGE analysis are likely the result of
inter-strain antigenic variation. It should be noted that both inter- and intra-strain
antigenic variation could contribute to the variability of symptoms seen in giardiasis.
Perhaps infections with Giardia strains which possess the relevant protel\1(s) present in
Flb lead to disaccharidase deficiencies in the hosto Conversely, an infœtion wilh il
strain which lacks the key portion of fraction Flb would not affect disaccharide
absorption. The protein profiles also revealed that the separation of the trophozoite
extract and fraction FI, using gel filtration, was not ideal. There were fractions
obtained which shared proteins of similar molecular weight. This probably rcsuItcd due
to contamination from preceding fractions during the chromatographie separation.
However, given the inability of fraction F2 proteins to affect enzyme activity, il is
likely that any proteines) in Flb responsible for the disaccharidase deficieneies is in the
55 to 200 kDa region.
The role of immune mechanisms III producing malabsorption and intestinal
symptoms is unclear. Daniels and Belosevic (1992) found that challenging micc with
an extract of G. muris led to more depressed disaccharidase activity in susceptiblc
animais as compared to resistant mice. These authOls suggested that this finding could
be due to an immune responsc to a particular parasite antigen in the susceptible micc
which differs from the response to anothcr antigen by the reslstant mice. Thereforc,
selective immune recognition of the relevant parasite antigen, present in fraction FI b,

•
•
•
76
could ultimately result in enzyme deficiencies. The specific type of immune response
to Flb which could cause a decline in disaccharidase activities rema:ns unknown.
Based on other studies on intestinal damage, the disaccharidase deficiencies in giardiasis
may be linked to a mucosal mast cell response to an :-, h antigen. Perdue and Gall
(1987) implicated mast cell mediators in the abnorma:ities of intestinal function
observed in anaphylactic rats. In these rats, there is significant disruption of the
intestinal mucosa which is linked tl' mllcosal mast ccli degranulation (Patrick et al.,
1988). In addition, Curtis et al. (1990) demonstrated decreased disaccharidase activity,
as weil as decreased brush border microvillus surface area and vilIus height, in rats
following chronic antigen challenge. Mucosal mast cell involvement was sllggested by
mast cell proliferation and degranulation. lt is certain, however, that since the Flb
challenge in the present study was done orally this would imply that in order to
stimulate any type of host mucosal immune response, fraction Ftb remained
immunogenically intact upon arrivai in the smalt intestine.
It is important to note that there are possible non-immune mechanisms which
could contribute to disaccharidase deficiencies. For example, Giardia trophozoites are
known to possess proteinase activity (Hare el al., 1989). A particular proteinase could
be present in fraction Flb which affects the enzyme levels as observed. Giardia
trophozoites also exhibit lectin activity. Plant lectins have been shown to be capable
of directly damaging rat enterocytes, producing brush border abnormalities similar to
those seen in giardiasis (Lorenzsonn and Olsen, 1982). It is therefore possible that
Giardia lectins are present in fraction Flb and contribute to host microvillus damage,

•
•
•
77
with subsequent disaccharidase impairment. In addition, it has been found that ~he
variant surface proteins (VSPs) which are present on Giardia trophozoites contain
metal-binding domains, sorne of which bind zinc (Thompson et al., 1993). Fraction
Flb may contain a VSP(s) which competes with the host for zinc in the intestine,
causing a decrease in enzyme activity. It is apparent, though, that whatever the
constituents of FI b which are involved in the disaccharidase deficiencies, they are able
to evade degradation by the enzymes of the gut, through an unknown mechanism.
The present study has implicated fraction Flb of the G. lamblta trophozoite
extract in the disaccharidase deficiencies observed in immune gerbils. It seems
probable that without the relevant component of Flb, Giardia strains cannot cause
disaccharidase deficiencies in infected hosts. However, further investigation of the
mechanism(s) of interaction between Flb and the mucosal epithelial ceUs, inc1uding the
specifie portion of fraction Flb involved in the intestinal dysfunction, is necessary .

•
•
•
78
REFERENCES
Belosevic M., Faubert G.M. and Maclean J.O. 1989. Oisaccharidase activity in the smalt intestine of gerbils (Meriones unf.:lIiculatus) during primary and challenge infections with Giardia lamblia. Gur 30: 1213-1219.
Brodsky R.E., Spencer H.C. and Schultz M.G. 1974. Giardiasis in American travelers to Soviet Union. J. Infect. Dis. 130: 319-323.
Buret A., Gall D.G. and OIson M.E. 1990. Effects of ruurine giardiasis on growth, intestinal morphology, and disaccharidase activity. J. Parasitol. 76: 403-409.
Campbell J.O. and Faubert G.M. 1994. Comparative studies on Giarclia lamblia encystation in vitro and in vivo. J. ParasilOl. 80: 36-44.
Cevallos A.M. and Farthing M.J .G. 1992. Small intestinal functional damage in experimental giardiasis is strain-dependent. Gastroenterology 102: A602.
Chaudhuri P.P., Pal S., Pal S.C. and Das P. 1988. Studies on Giardia lamblia trophozoite antigens using sephacryl S-300 column chromatography, polyacrylamide gel electrophoresis and enzyme-linked immunosorbent assay. In: Advances in Giardia Research. Wallis P.M. and Hammond B.R. (cds.). University of Calgary Press: Calgary, pp. 191-194.
Curtis G.H., Patrick M.K., Catto-Smith A.G. and Gall D.G. 1990. Intestinal anaphylaxis in the rat: effect of chronic antigcn exposure. Gastroent(lrolo~y 98: 1558-1566.
Dahlqvist A. 1968. Assay of intestinal disaccharidases. Anal. Biochem. 22: 99-107.
Daniels C. W. and Belosevic M. 1992. Oisaccharidase activity in the small intestine of susct::ptible and resistant mice after primary and challenge infections with Giardia muris. Am. J. Trop. Med. Hyg. 46: 382-390.
Farthing M.J.G. 1992. New perspectives in giardiasis. J. Med. Microbiol. 37: 1-2.
Gillin F.D. and Diam(lnd L.S. 1979. Axenically cultivated Giardia lamblia: growth, attachment and the role of L-cysteine. In: Waterborne Transmission of Giardiasis. Jakubowski W. and HoffJ.C. (eds.). U.S. Environmental Protection Agency: Cincinnati, pp. 270-272.
Hare O.F" Jarroll E.L. and Lindmark D.G. 1989. Giardia lamblia: characterization of proteinase activity in trophozoites. Exp. Parasitol. 68: 168-175.
J

•
•
•
79
Hoskins L.C., Winawer S.1., Broitman S.A., Gottlieb L.S. and Zarncheck N. 1967. Clinical giardiasis and intestinal malabsorption. Gastroenterology 53: 265-279.
Kamath K.R. and MJrugasu R. 1974. A comparative study of four methods for detecting Giardia lamhlia in children with diarrheal disease and malabsorption. Guslroenterology 66: 16-21.
Keystone J .S., Krajdell S. and Warren M.R. 1978. Person-to-person transmission of Giardia lamblia in day-care nurseries. Cano Med. Assoc. J. 119: 241-248.
Kumkum, Khanna R., Khuller M., Mehta S. and Vinayak V.K. 1988. Plasma membrane associated antigens of trophozoites ofaxenic Giardia lamblia. Trans. R. Soc. Trop. Med. Hyg. 82: 439-444.
Laemmli U.K. 1970. Cleavage of structural proteins during assembly of the head of bacteriophage T4. Nature 227: 680-685.
Lorenzsonn V. and Olsen W.A. 1982. ln vivo responses of rat intestinal epithelium to intraluminal dietary lectins. Gustroenterology 82: 838-848.
Lowry O.H., Rosebrough N.J., Farr A.L. and Randall R.J. 1951. Protein measurements with folin phenol reagent. J. Biol. Chem. 193: 265-275.
Mahalanabis D., Simpson T.W., Chakraborty M.L., Ganguli C., Bhattacharjee D.K. and Mukherjee K.L. 1979. Malabsorption of water miscible vitamin A in children with giardiasis and ascariasis. Am. J. Clin. NUlr. 32: 313-318.
Moore G.W., Sogandares-Bernal F., Dennis M.V., Root D.M., Beckwith D. and van Voorhis D. 1982. Characterization of Giardia lamblia trophozoite antigens using polyacrylamide gel electrophoresis, high-performance liquid chromatography, and enzyme-Iabeled immunosorbent assay. Vel. Parasitol. 10: 229-237.
Nain C.K., Dutt P. and Vinayak V.K. 1991. Alterations in enzymatic activities of the intestinal mucosa during the course of Giardia lamblia infection in mice. Ann. Trop. Med. Parasitol. 85: 515-522.
Patrick M.K., Dunn I.J., Buret A., Miller H.R.P., Huntley J.F., Gibson S. and Gall D.G. 1988. Mast cell protease release and mucosal ultrastructure during intestinal anaphylaxis in the rat. Gastroenterology 94: 1-9.
Perdue M.H. and Gall D.G. 1987. Mucosal mast cells and the intestinal epithelium. Adv. Exp. Med. Biol. 216A: 645-652 .

•
•
•
80
S,chmerin M.J., Jones T.C. and Klein H. 1978. Giardiasis: association with homosexuality. Ann. lm. Med. 88: 801-803.
Shandera W.X. 1990. From Leningrad to the day-care center - The ubiquitous Giardia lamblia. West. J. Med. 153: 154-159.
Smith p.n., Gil1.in F.O., Kaushal N.A. and Nash T.E. 1982. Antigenic analysls of Giardia lamblia from Afghanistan, Puerto Rico, Ecuador, and Oregon. II/fi'et. Immun. 36: 714-719.
Smith p.n., Gillin F.O., Spila W.M. and Nash T.E. 1982. Chronic giardiasis: studies on drug sensitivity, toxin production and host immune rt~sponse.
Gastroenterology 83: 797-803.
Thompson R.C.A., Reynoldson J.A. and Lymbery AJ. 1993. Giardia - from molecules to disease and beyond. Parasitol. Today 9: 313-315.
Wright S.G., Tpmkins A.M. and Ridley O.S. 1977. Giardiasis: clinical and therapeutic aspects. Gut 18: 343-350.

•
•
•
81
GENERAL DISCUSSION
Infections with G. lamblia are now considered an important cause of
gastroenteritis (Shandera, 1990) and dis:a.ccharidase deficiencies have been consistently
identified as one of various abnormalitr\~s of small intestinal pathology in giardiasis
(Jennings et al., 1976). Since unabsorbea sugars lead to diarrhea by causing net fluid
accumulation in the intestinal lumen (Launl\\la, 1968), disaccharidase deficiencies play
a key role in understanding the pathogeol'Ic mechanisms which lead to intestinal
dysfunction in giardiasis. ft has been suggest\,j that the changes in the small intestine
of Giardia-infected individuals, including the 1 mpairments of disaccharidase activity,
are due to both a direct effect of the parasite and to antigen-specific host immune
responses, such as immune complexes and delayed-type hypersensitivity (Gillon et al. ,
1982). However, other host factors, such as diet, gut motility and nutritional status
could also contribute to the intestinal pathology (Ferguson et al., 1980). At present,
there is Iittle knowledge of the relative contributions of parasite and host-related factors
to the disaccharidase rleficiencies in giardia;'ls. Therefore, this project involved a closer
examination of the process by which G. lamblia causes these reductions in enzyme
activity, using the gerbil model of the human disease.
In Manuscript l, a more extensive characterization of the effects on
disaccharidase activity in G. lamblia-infected gerbils was reported on. For all 4
disaccharidases examined, the activities were reduced during a primary G. lamblia
infection. These enzyme deficiencies were first observed during the acute phase of the

•
•
•
82
infection, but persisted weB past the elimination of the parasite from the smalt intestine.
These findings indicate that the disaccharidase deficiencies are not dependent on the
persistent presence of the parasite in the gut. In facto studies on giardiasis in humans
have found that there are patients with continued malabsorption following eradication
of the parasite (Brasitus, 1983). It is possible that the enzyme deficiencies continue due
to the initial enterocyte destruction by trophozoite attachment and to indirect intestinal
damage by an effective host mucosal immune response. Mucosal disruption would
necessarily affect the disaccharidases present there. In a challenge infection with live
G. lamblia trophozoites, the activities of the disaccharidases (with the exception of
sucrase) were also decreased. However, the reductions in enzyme activity in this case
were of short duration, occurring only on days 2 and 4 p.c., perhaps bccause G.
lamblia trophozoites do not establish in the gerbil gut upon challenge. In addition,
similar impairments of disaccharidase activity were obtained when gerbils were
challenged with 0.1 mg of the soluble extract of G. lamblia trophozoites. The effects
of the extract challenge were dose-dependent and indicate that the live parasite is not
needed to obtain the enzyme deficiencies. These findings suggest that there may lle an
immunological basis for the decreases, involving a host response to G. lamblia surface
antigens. Since challenging gerbils previously expost'.d te G. lamblia with an extract
of E. histolytica had no affect on disaccharidase activities, it seems that the decreases
observed with a Giardia challenge are specifie to this parasite. Also, it seems that G.
lamblia metabolic products do not play a major role in the disaccharidase deficiencies
observed in giardiasis as the trophozoite excretory/secretory products affected these

•
•
•
83
enzymes in a manner which was inconsistent with the live parasite challenge.
Manuscript 2 focused on studies involving the partial puriftcation and
characterization of a specific fraction of the G. lamblia crude extract responsible for
inducing the disaccharidase deficiencies in immune gerbi:s. Gel filtration using a
Sephacryl S-200 HR column allowed for the se!Jaration of the crude extract of G.
lamblia trophozoites into 3 fractions. Only a challenge with fraction FI affected
disaccharidase activities in immune gerbils, with decreased levels observed on days 2
and 4 p.c. The patterns of the reductions in enzyme activity with the FI-challenge
were similar to those obtained with live- and extract- challenged animais and indicated
the importance of fraction FI in the disaccharidase deficiencies. The component of FI
involved in the impairments of enzyme activity was determined to be heat-stable .
Sephacryl 5-200 HR gel chromatography of fraction FI, using an eluent co.mlÏnirz the
dissociating agent guanidine-HCI and the reducing agent dithiothreitol, allowed for the
further separation of G. lamblia proteins. Of the 2 fractions obtained following this
separation, only a challenge with fraction Flb resultui in disaccharidase deficiencies,
similar to those observed in F 1- or crude extract- challenged gerbils. These results
indicate that with Flb we have partially purified the fraction of the parasite responsible
for inducing disaccharidase deficiencies in immune animals. nrotein analysis of the
erude extract and the fractions of the extract reve.aled many high and low molecular
weight banùs, with any protein(s) in Flb responsible for the decreases in enzyme
activity in the 55 to 200 kDa region.
The role of immune mechanisms in producing disaccharidase deficiencies in G .

•
•
•
84
lamblia infections is not known. Garside and colle,aglles (1991) found that interferon-
a/{3 (IFN-a/{3) contributes to intestinallesions in l1lice with graft-verslls-host re,lction
(GVHR). It was suggested that IFN-a/j3 callsed mucosal damage through the induction
of natural killer (NK) ceU activity. Intestinal damage by NK cells could thcrcfore lcaù
to reductions in disaccharidase activity in giardiasis. Another possible immune
mechanism which could result in disaccharidase deficiencies in Giardia infections is the
disruption of the intestinal l1lucosa by mast cell mediators. Mast cell degranulation may
occur in response to a Giardia antigen and lead to intestinal abnormalities, as has bcen
found in rats following chronic challenge with egg albumin (Curtis l'I al., 1990). Otllcr
researcher~, using rnice with GVHR, have found that T cells, after reacting wlth an
antigen, can release lymphokines which damage enterocytes (Mowat and Fcrguson,
1981). Such damage could again result in a decline in disaccharidase activities.
Moreover, there are many other possible immune mechanisms whlch could produce
impairments of enzyme activity, however there has yet to be a link found bctwccn a
particular ho st l1lucosal immune response and the disaccharidase deti il~ncics in
giardiasis.
There are several possible- avenues for future research in this arca. For
example, more sophisticated purification techniques can be used in studies to isolate the
specifie component of G. lamblia which is responsible for the decreascs in enzyme
activity. Also, disaccharidase levels can be measured following the lIlhibition of factors
such as free radical::;, mast ceIl degranulation/n1ediators, IFN-al{3, or Iysosomal
proteases. This would aid in determining which immune mediators are involved in the

•
•
•
85
disaccharidase deficiencies. Studying the cellular infiltration in the small intestine
during challenge inoculations would also assist in specifying the type of immune
mechanism associated with the reductions in disaccharidase activity. Another area
which deserves further investigation is the effects of G. lamblia excretory/secretory
products on the small intestine.
Although many aspects of the process leading to disaccharidase deficiencies in
giardiasis have yet to be examined, it is hoped that the findings presented here can be
used in developing therapeutic strategies for malabsorption and, ultimately, for G.
lamblia infections .

•
•
•
86
REFERENCES
Ackers J.P. 1980. Giardiasis: basic parasitology. Trans. R. Soc. Trop. Met!. Hyg. 74: 427-429.
Adam R.O. 1991. The biology of Giardia spp. Microhiol. Rev. 55: 706-732.
Aggarwal A., Merritt J.W. and Nash T.E. 1989. Cysteine-rich variant surface proteins of Giardia lamblia. Mol. Biochem. Parasirol. 32: 39-48.
Aggarwal A. and Nash T.E 1988. Antigenic variation of Giardia lamhlia ;n l'ivo. Infect. Immun. 56: 1420-1423.
Ament M.E. and Rubin C.E. 1972. Relation of giardiasis to abnormal intestinal structure and function in gastrointestinal immunodeficiency syndromes. Gastroenterology 62: 216-226.
Anand 8.S., Kumar M., Chakravarti R.N., Sehgal A.K. and Chhuttani P.N. 1980. Pathogenesis of malabsorption in Giardia infection: an expcrimental study in rats. Trans. R. Soc. Trop. Met!. Hyg. 74: 565-569.
Banai J., Szanto 1., Nagy 1. and Kun M. 1990. Measurement and demonstration of lactase and sucrase activities in jejunal mucosa. Am. J. Ga.\/r()('IlI('ml. 85: 157-160.
Belosevic M. and Faubert G.M. 1986. Comparative studies of Inflamnmtory responses in susceptible and resistant mice infected with Giardia mllris. Clin. Exp. Immllnol. 65: 622-630.
Belosevic M., Faubert G.M. and Maclean J.O. 1989. Oisacchandase acti\ity in the small intestine of gerbils (Meriones unguicularus) durin6 primary and challenge infections with Giardia lamblta. GUl30: 1213-1219.
Belosevic M., Faubert G.M., Maclean J.O., Law C. and Croll N.A. 1983. Giardia lamblia infections in Mongolian gerbils: an animal model. J. Infect. Di.\'. 147: 222-226.
Bertram M.A., Meyer E.A., Lile J.O. and Morse S.A. 1983. A comparison of isoenzymes of five axenic Giardia isolates. J. Parasitol. 69: 793-801.
Bifulco J.M. and Schaefer F.W. 1992. Seriai subcultivation of Giardia lamhlia in Keister's modified TYI-S-33 medium containing Ultroser G. J. Protozool. 39:

•
•
•
87
211-213.
Brasitus T.A. 1983. Parasites and malabsorption. Clin. Gastroenterol. 12: 495-510.
Briaud M., Morichau-Beauchant M., Matuchansky C., Touchurd G. and Sabin P. 1981. Intestinal immllne responses in giardiasis. Lancet 2: 358.
Butscher W.G. and Faubert G.M. 1988. The therapeutic action of monoclonal antibodies against a surface glycoprotein of Giardia muris. Immunology 64: 175-180.
Carlson J.R., Heyworth M.F. and Owen R.L. 1986. Response of Peyer's patch lymphocyte subsets to Giardia muris infection in BALB/c mice. CeU. Immunol. 97: 44-50.
Carlson J.R., Heyworth M.F. and Owen R.L. 1987. T lymphocyte subsets in nude mice with Giardia muris. Thymu.\ 9: 189-196.
Char S., Shetty N., Narashimha M., Elliott E., Macaden R. and Farthing M.J.G. 1991. Serum antibody response in childrell with Giardia lamblia infection and identificatIOn of an immunodominant 57 kD antigen. Parasile Immul1ol. 13: 329-337.
Clark J. T. and Holberton D. V. 1986. Plasma membrane isolated from Giardia lamblia: identification of membrane proteins. Eur. J. CeU Biol. 42: 200-206.
Craft J .C. 1982. Experimental infection with Giardia lamblia in rats. J. Infect. Dis. 145: 495-498.
Curtis G.H., Patrick M.K., Catto-Smith A.G. and Gall D.G. 1990. Intestinal anaphylaxis in the rat: effect of chronic antigen exposure. Gastroenterology 98: 1558-1566.
Daniels C. W. and Belosevic M. 1992. Disaccharidase activity in the smaH intestine of susceptible and resistant mice after primary and challenge infections with Giardia ml/ris. Am. J. Trop. Med. Ilyg. 46: 382-390.
Deguchi M., Gillin F.D. and Gigli 1. 1987. Mechanism of k:lling of Giardia lamblia trophozoites by complement. J. Clin. IlIveSf. 5: 1296-1302.
denHollander N., RIley D. and Befus D. 1988. Immunology of giardiasis. ParasilOl. TodllY 4: 124-131.
Diamond L.S. 1968. Techniques ofaxenic cultivation of Entamoeba histolytica

•
•
•
88
(Schaudinn, 1903) and E. hisfOl)'IÎCalikcamocbae. J. Pan.\ilol.54: 1047-1056.
Einfeld D.A. and Stibbs H.H. 1984. Identitication and charactt'rintion of a major surface antigen of Gicm/w lamhlia. b{/l'CI. 1/1111/11". 46: 377-383.
Erlandsen S.L. 1974. Scanning electron microscopy of intestlllal gianliasis: lcsions of the microvillus border of villus epitheltal cells produccd by trophOLOitcs of Giardia. Sca"ning Electron Mlcro.\c. 1974: 775-782.
Erlich J.H., Anders R.F., Roberts-Thomson I.e .. Schradcr J. W. and Mitchell G. F. 1983. An examination of diffcrences III serum antibody spcciticitICs and hypersensitivlty reactions as contributing factors to chronic infection wlth the intestinal protozoan parasite, Giardw mllris in 1111ce. Aust. J. Erp. Ilial. Mn/. Sei. 61: 599-615.
Farthing M.J.G. 1990. Immunopathology of giardiasis. Sprillgel' Sem;". Immunopathol. 12: 269-282.
Farthing M.J.G., Chong S. and Walker-Smith J.A. phenomena in giardiasis. Lancer 2: 1428-1430.
1984. Acute allergie
Farthing M.J.G., Yaron S.R. and Keusch G.T. 1983. Mammalian bile promotcs growth of Giardia lamblia in axenic culture. Trans. R. Soc. Trop. Mnl. f1yg. 77: 467-469.
Faubert G.M. 1988. Evidence that giardiasis is a zoonosis. Para.\'iwl. Today 4: 66-68.
Faubert G.M., Belosevic M., Walker T.S., Maclean J.O. and Meerovitch E. 1983. Comparative studies on the pattern of infection with Giardia spp. in Mongolian gerbils. J. Parasitol. 69: 802-805.
Faubert a.M., Leziy S.S., Bourassa A. and Mac1can J.O. 1986. Effccts of environ mental conditions and standard chlorinatlon praetlcc~ on the infectivity of Giardia cysts. Di.\. Aquat. Org. 2: 1-5.
Feely D.E., Erlandsen S.L. and Chase D,G. 1984. Structure of the trophomite and cyst. In: Giardia and Glllrdia!)ÎS. Erlandsen S.L. and Meyer E.A. (cds.). Plenum Press: New York, pp. 3-31.
Ferguson A., Gillon J. and Al Thamery D. 1980. Inte~tinal abnormalitics in murine giardiasis. Trans. R. Soc. Trop. Med. HYR. 74: 445-448.
Pilice F.P. 1952. Studies on the cytology and life history of a Giardia from the

•
•
•
89
laboratory rat. Univ. Calif. Publ. Zool. 57: 53-146.
Garside P., Felstein M. V., Green E.A. and Mowat A.Mel. 1991. The role of interferon a/{J in the induction of intestinal pathology in mice. Immunology 74: 279-283.
Gmin F.O. and Oiamond L.S. 1980. Clonal growth of Giardia lamblia trophozoites in a semisolid agarose medium. J. Parasitol. 66: 350-352.
Gillin F.O., Gault M.J., Hofmann A.F., Gurantz D. and Saueh J.F. 1986. Biliary lipids support serum free growth of Giardia lamblia. Infect. Immun. 53: 641-645.
Gillin F.O. and Reiner O.S. 1982. Attaehment of the flagellate Giardia lamblia: the role of reducing agents, serum, temperature and the ionic composition. Mol. Cel/. Biol. 2: 369-377.
Gillon J., AI Thamery D. and Ferguson A. 1982. Features of small intestinal pathology (epithelial cell kinetics, intraepitheliallymphocytes, disaceharidases) in primary Giardia muris infection. Gut 23: 498-506 .
Goda T. and Koldovsky O. 1988. Dietary regulation of small intestinal disaccharidases. Wld. Rev. Nutr. Diet. 57: 275-329.
Goka A.K.J., Rolston D.D.K., Mathan V.1. and Farthing M.J.G. 1986. Diagnosis of giardiasis by specific IgM antibody enzyme-linked immunosorbent assay. Lancet 2: 184-186.
Goka A.K.J., Roiston D.D.K., Mathan V.1. and Farthing M.J.G. 1989. Serum IgA response in human Giardia lamblia infection. Serodiag. Immunother. 3: 273-277.
Gottstein B., Stocks N.I., Sheurer G.M. and Nash T.E. 1991. Human cellular immune responses to Giardia lamblia. Infection 19: 421-426.
Grant D.R. and Woo P.T.K. 1978. Comparative studies of Giardia spp. in small mammaJs in southern Ontario. II. Host specificity and infectivity of stored cysts. Cano J. Zool. 56: 1360-1366.
Hartong W.A., Gourley \V.K. and Arvanitakis C. 1979. Giardiasis: clinical spectrum and functional-structural abnormalities of the smalt intestinal mucosa. Gastroenterology 77: 61-69 .
Hermans P.E., Diaz-Buxo J.A. and Stobo J.O. 1976. Idiopathie late-onset

•
•
•
90
immunoglobulin deficiency: clinicat observations in 50 patients. Am. J. Med. 61: 221-237.
Hewlett E.L., Andrews J.S., Ruffier J. and Schaefer F.W. 1982. Experimental infection of mongrel dogs with Giardia lamblia cysts and cultured trophozoites. J. Infect. Dis. 145: 89-93.
Heyworth M.F. 1986. Antibody response to Giardia muris trophozoites in mOllse intestine. Infect. Immun. 52: 568-571.
Heyworth M.F. 1992. Relative susceptibility of Giardia murü trophozoites to killing by mouse antibodies of different isotypes. J. Parasitol. 78: 73-76.
Heyworth M.F., Kung J.E. and Caplin A.B. 1988. Enzyme-linked immunosorbent assay for Giardia-specifie IgA in mouse intestinal secretions. Parasite Immunol. 10: 713-717.
Heyworth M.F., Kung J.E. and Eriksson E.C. 1986. Clearance of Giardia murü infection in mice deficient in natural killer cells. Infect. Immun. 54: 903-904.
Hill D.R., Burge J.J. and Pearson R.D. 1984. Susceptibility of Giardia lamblia trophozoites to the lethal effect of human serum. J. Immunol. 132: 2046-2052.
Hill D.R., Guerrant R.L., Pearson R.D. and Hewlett E.L. 1983. Giardia lamblia infection in suckling mice. J. Infect. Dis. 147: 217-221.
Hoskins L.e., Winawer S.J., Broitman S.A., Gottlieb L.S. and Zamcheck N. 1967. Clinical giardiasis and intestinal malabsorption. Gastroenterology 53: 265-279.
Hughes W.S., Cedra J.J., Holfzapple P. and Brooks F.P. 1971. Primary hypogammaglobulinema and malabsorption. Ann. Intern. Med. 74: 903-910.
Inge P.M.G., ~son C.M. and Farthing M,J.G. 1988. Attachment of Giardia lamblia to rat intestinal epithelial cells. Gut 29: 795-801.
Jarroll B.L., Manning P., Lindmark D.G., Coggins J.R. and Erlandsen S.L. 1989. Giardia cyst wall specific carbohydrate: evidence for the presence of galactosamine. Mol. Biochem. Parasitol. 32: 121-132.
Jennings W., Rowland R., Hecker R., Gibson G.E., Fitch R.J. and Reid D.P. 1976. The significance of lowered disaccharidase levels. Aust. N.Z. J. Med. 6: 556-560.
Jokipii A.M.M. and Jokipii L. 1982. Serum IgG, IgA, IgM and IgD in giardiasis: the

•
•
•
91
most severely iII patients have tiule IgD. J. Infect. Dis. 5: 189·193.
Kanwar S.S., Ganguly N.K., Walia B.N.S. and Mahajan R.C. 1985. Changes in mouse splenic lymphocyte sub-population during primary and secondary course of Giardia lamhlia infection. Indian J. Med. Res. 82: 393-397.
Kanwar S.S., Ganguly N.K., Walia B.N.S. and Mahajan R.C. 1986. Direct and antibody dependent cclI mediated cytotoxicity against Giardia lamblia by splenic and intestinal lymphoid cells in mice. GUI 27: 73-77.
Karapetyan A.E. 1960. Methods of Lamblia cultivation. Tsitologiia 2: 379-384.
Karapetyan A.E. 1962. In vitro cultivation of Giardia duodenalis. J. Parasitol. 48: 337-340.
Keister O. 1983. Axenic culture of Giardia lamblia in TYI-S-33 medium supplemented with bile. Trans. R. Soc. Trop. Med. Hyg. 77: 487-488.
Kirkpatrick C.E. and Greene G.A. 1985. Susceptibility of domestic cats to infections with G. lamblia cysts and trophozoites from human sources. J. Clin. Microbiol. 21: 678-680 .
Kumkum R., Khanna M., Khuller M., Mehta S. and Vinayak V.K. 1988a. Plasma membrane associated antigens of trophozoites ofaxenic Giardia. Trans. R. Soc. Trop. Med. Hyg. 82: 439-444.
Kumkum R., Khanna R., Nain C.K., Mehta S. and Vinayak V.K. 1988b. Depressed humoral immune responses to surface antigens of Giardia lamblia in persistent giardiasis. Pediatr. Infect. Dis. J. 7: 492-498.
Launiala K. 1968. The effect of unabsorbed suc rose and mannitol on small intestinal tlow rate and mean transit time. Scand. J. Gastroenterol. 3: 665-671.
Levinson J.D. and Nastro L.J. 1978. Giardiasis with total villous atrophy. Gastroenœrology 74: 271-275.
Loftness T.E., Erlandsen S.L., Wilson 1.0. and Meyer E.A. 1984. Occurrence of specifie secretory immunoglobulin A in bile after inoculation of Giardia lamblia trophOloites into rat duodenum. Gastroenterology 87: 1022-1029.
MacDonald T.T. and Ferguson A. 1978. Small intestinal epithelial cell kineties and protoloal infection in mice. Gastroenterology 74: 496-500 .
Mahalanabis D., Simpson T.W., Chakraborty M.L., Ganguli C., Bhattacharjee O.K.

•
•
•
92
and Mukherjee K.L. 1979. Malabsorption of water miscible vitamin A in children with giardiasis and ascariasis. Am. J. Clin. NUlr. 32: 313-318.
Meloni B.P. and Thompson R.C.A. 1987. Comparative studies on the axenic in vitro cultivation of Giardia of human and ca.nine OI'igin: evidence for intra specifie variation. l'rans. R. Soc. Trop. Med. Hy~. 81: 637-640.
Meyer E.A. 1970. Isolation and axe nie cultivation of Giardia from the rabbit, chinchillaandcat. Exp. Parasitol. 27: 179-183.
Meyer E.A. 1976. Giardia lamblia: isolation and axenie cultivation. Exp. Parasitol. 39: 10 1-105.
Mowat A.McL. and Ferguson A. 1981. Hypersensitivity reactions in the small intestine. 6. Pathogenesis of the graft-versus-host reaction in the small intestinal mucosa of the mouse. Transplantation 32: 238-243.
Nash T.E. and Aggarwal A. 1986. Cytotoxicity of monoclonal antibodies to a subset of Giardia isolates. J. Immllnol. 136: 628-632.
Nash T.E., Aggarwal A., Adam R.D., Conrad J.T. and Merrit J.W. 1988. Antigenic variation in Giardia lamblia. J. Immunol. 141: 636-641.
Nash T.E., Gillin F.O. and Smith P.D. 1983. Excretory-secretory products of Giardia lamblia. J. Immunol. 131: 2004-2010.
Nash T.E., Merritt Jr. J.W. and Conrad J.T. 1991. Isolate and epitope variability in susceptibility of Giardia lamblia to intestinal proteases. Infect. Immun. 59: 1334-1340.
Nayak N., Ganguly N.K., Walia B.N.S., Wahi V" Kanwar S.S. and Mahajan R.C. 1987. Specifie secretory IgA in the milk of Giardia lamblia infectec:J and uninfected women. J. Infect. Dis. 155: 724-727.
Owen R.L., Allen C.L. and Stevens D.P. 1981. Phagocytosis of Giardia murü by macrophages in Peyer's patch epithelium in mice. Itifect. Immun. 33: 591-601.
Owen R.L., Nemanic P.C. and Stevens D.P. 1979. Ultrastructural observations on giardiasis in a murine model. 1. Intestinal distribution, attachment, and relationship to the immune system of Giardia muris. Gastmenteroio1!.Y 76: 757-769.
Pimenta P.F,P" da Silva P.P. and Nash T. 1991. Variant surface antigens of Giardia lamblia are associated with the presence of a thick cell coat: thin section and

•
•
•
93
label fracture immunocytochemistry survey. Infect. Immun. 59: 3989-3996.
Radulescu S. and Meyer E.A. 1981. Opsonization in vitro of Giardia lamblia trophozoites. Infect. Immun. 32: 852-856.
Reiner O.S., Douglas H. and Gillin F.O. 1989. Identification and localization of cy~tspecifie antigens of Giardia lamblia. Infect. Immun. 57: 963-968.
Reiner O.S. and Gillin F.O. 1992. Human secretory and serum antibodies ncognize environmentally induced antigens of Giardia lamblia. Infect. Immun. 60: 637-643.
Reiner O.S., McCaffery M. and Gillin F.n. 1990. Sorting of cyst wall proteins to a regulated secretory pathway during differentiation of the primitive eukaryote Giardia lamblia. Eur. J. Cell Biol. 53: 142-153.
Ridley M.J. and Ridley O.S. 1976. Serum antibodies and jejunal histology 10
giardiasis associated with malabsorption. J. Clin. Pathol. 29: 30-34.
Roberts-Thomson I.C. and Anders R.F. 1981. Serum antibodies in adults with giardiasis. Gastroenterology 80: 1262 .
Roberts-Thomson I.C. and Mitchell G.F. 1978. Giardiasis in mice: 1. Prolonged infections in certain mouse strains and in hypothymic (nude) rats. Gastroenterology 75: 42-46.
Roberts-Thomson I.C., Stevens D.P., Mahmoud A.A.F. and Warren K.S. 1976. Giardiasis in the mouse: an animal mode!. Gastroenterology. 71: 57-61.
Rosoff J.O. and Stibbs H.H. 1986a. Isolation and identification of a Giardia lambliaspecifie stool antigen (GSA 65) useful in coprodiagnosis of giardiasis. J. Clin. Microbiol. 23: 905-910.
Rosoff J.O. and Stibbs H.H. 1986b. Physical and chemical characterization of a Giardia lamblia-specifie antigen useful in the coprodiagnosis of giardiasis. J. Clin. Microbiol. 24: 1079-1083.
Saha T.K. and Ghosh T.K. 1977. Invasion of small intestinal mucosa by Giardia lamblia. Gastroenterology 73: 402-405.
Sehgal A.K., Grewal M.S., Chakravarti R.N., Broor S.L., Deka N.C. and Chuttani P.N. 1976. Experimental giardiasis in albino rats. Indian J. Med. Res. 64: 1015-1018 .

•
•
•
94
Shandera W.X. 1990. From Leningrad to the day-care center - the ubiquitous Giardia lamblia. West. J. Med. 153: 154-159.
Sharma A.W. and Mayrhofer G. 1988a. A comparative study of infections with rodent isolates of Giardia duodenalis in inbred strains of rats and \l'lice and in hypothymic nude rats. Parasite Immullo/. 10: 169-179.
Sharma A. W. and Mayrhofer G. 1988b. Biliary antibody response in rats infected with rodent Giardia duodenalis isolates. Parasill' Imnlll1lol. 10: 181-191.
Sheffield H.G. and Bjorvatn B. 1977. Ultrastructure of the cyst of Giardia lilfllblia. Am. J. Trop. Med. Hyg. 26: 23-30.
Sherman P. 1980. Apparent protein losing enteropathy associated with giardiasis. Am. J. Dis. Chi/do 134: 893-894.
Slonim J.J., Ireton H.J.C. and Smallwood R.A. 1976. Giardiasis following gastric surgery. Aust. N.Z. J. Med. 6: 479-480.
Smith P.O., Elson C.E .. Keister O.B. and Nash T.E. 1982a. Human host response to Giardia lamblia. 1. Spontaneous killing by mononuclear leukocytes in vitro. J. Immunol. 128: 1372-1376.
Smith P.D., GilIin F.O., Kaushal N.A. and Nash T.E. 1982b. Antigenic analysis of Giardia lamblia from Afghanistan, Puerto Rico, Ecuador and Oregon. !tf/h·t. Immun. 36: 714-719.
Smith P.D., Keister O.B. and Elson c.a. 1983. Human host response to Giardia lamblia. II. Antibody-dependent killing in vitro. Cell. Immuno/. 82: 308-315.
Snider D.P. and Underdown B.J. 1986. Quantitative and temporal analyses of murine antibody response in serum and gut secretions to infection with Giardia muri.\'. Infect. Immun. 52: 271-278.
Solomons N.W. 1982. Giardiasis: nutritional implicatior.s. Rev. Infc'ct. Dis. 4: 859-869.
Solovjev M.M. 1975. Biometrical studies of the trophozoites of Lamhlia from mammals in regard to the taxonomy of the genus Lamblia. Parasitologia 9: 449-456.
Stevens D.P. 1982. Giardiasis: host-pathogen biology. Rev. Infect. Dis. 4: 851-858.
Stevens O.P., Frank O.M. and Mahmoud A.A.F. 1978. Thymus dependency ofhost

•
•
•
95
resistance to Giardia muris infectiun: studies in nude mice. J. Immunol. 120: 680-682.
Takano 1. and Yardley J. H. 1964. Jejunal lesions in patients with giardiasis and malabsorption. An electron microscopie study. Bull. Johns Hopkins Hosp. 116: 413-429.
Thompson A., Roland R., Hecker R., Gibson G.E. and Reid P.P. 1977. Immunoglobulin-bearing cells in giardiasis. J. Clin. Pathol. 30: 292-294.
Tomkins A.M., Wright S.G., Drassar B.S. and James W.P.T. 1978. Bacterial colonization of jejunal mucosa in giardiasis. Trans. R. Soc. Trop. Med. Hyg. 72: 33-36.
Torian B.E., Barnes R.C., Stephens RS. and Stibbs H.H. 1984. Tubulin and highmolecular weight polypeptides as Giardia lamblia antigens. Infect. Immun. 46: 152-158.
Vasudev V., Ganguly N.K., Anznd B.S., Krishna V.R., Dilawari J.B. and Mahajan R.C. 1982. A study of Giardia infection in irradiated and thymectomized mice. J. Trop. Med. Hyg. 85: 119-122 .
Vinayak V.K., Khanna R. and Kum Kum. 1991. Kinetics of intraepithelium and lamina propria lymphocyte responses during Giardia lamblia infection in mice. Microb. Pathog. 10: 343-350.
Vinayak V.K., Kumkum and Khanna R. 1989. Serum antibodies to giardial surfacf: antigens: lower titres in persistent than in non-persistent giardiasis. J. Med. Microbiol. 30: 207-212.
Vinayak V .K., Kumkum, Khanna R. and Khuller M. 1992. Systemic oral immunization with a 56 kD molecule of Giardia lamblia affords protection in experimental mice. Vaccine 10: 21-27.
Vi!.layak V.K., Sharma G.L. and Niak S.R. 1979. Experimental Giardia lamblia infection in Swiss mice, a preliminary report. Indian J. Med. Res. 70: 195-198.
Visvesvara G.S. 1980. Axenic growth of Giardia lamblia in Diamond's TPS-l medium. Trans. R. Soc. Trop. Med. Hyg. 74: 213-215.
Wang Z.Y., Lu S.Q., Zhang Y.K., Song J.L., Wen Y. and Gui S.H. 1986. Investigations on the prevalence, immunodiagnosis and experimental model of giardiasis. Chin. Med. J. 99: 961-968 .

•
•
•
96
Ward H.n., Alroy J., Lev B.I., Kel'sch G.T. and Pereira M.E. 1985. Identitication of chitin as a structural cornponent of Giardia cysts. lnfl'cl. Immun. 49: 629-634.
Wolfe M.S. 1992. Giardiasis. Clin. Microbiol. Rev. 5: 93-100.
Woo P.T.K. and Paterson W.B. 1986. Giardia lamblia in children in daycare centres in southem Ontario, Canada, and susceptibility of animais to G. lamblia. Trans. R. Soc. Trop. Med. Hyg. 80: 56-59.
Wright S.G. and Tomkins A.M. 1977. Quantification of the lymphocyte intiltrate in jejunal epithelium in giardiasis. Clin. Exp. lmmunol. 29: 408-412.
Wright S.G., Tomkins A.M. and Ridley O.S. 1977. Giardiasis: clinical and therapeutic aspects. GUI 18: 343-350.
Yardley J.H., Takano J. and Hendrix T.R. 1964. Epithelial and other mucosallesions of the jejunum in giardiasis. Jejunal biopsy studies. Bull. Johns Hopkins f/osp. 115: 389-406.
Zinneman H.H. and Kaplan A.P. 1972. The association of giardiasis with reduced intestinal secretory immunoglobulins. Am. J. Digest. Dis. 17: 793-797.