differential regional and cellular distribution of dopamine d2-like
TRANSCRIPT

Differential Regional and CellularDistribution of Dopamine D2-Like
Receptors: An ImmunocytochemicalStudy of Subtype-Specific Antibodies in
Rat and Human Brain
ZAFAR U. KHAN,* ANTONIA GUTIERREZ, ROSA MARTIN, ANTONIO PENAFIEL,
ALICIA RIVERA, AND ADELAIDA DE LA CALLE
Department of Cell Biology, Faculty of Science, University of Malaga, Teatinos 29071,Malaga, Spain
ABSTRACTDopamine D2-like receptors (D2, D3, and D4) are major targets for action of typical and
atypical neuroleptics, commonly used in the treatment of schizophrenia. To understand theirindividual functional contribution, subtype-selective anti-peptide antibodies were raisedagainst D2, D3, and D4 receptor proteins. The antibodies were shown to be specific onimmunoblots of rat brain membranes and immunoprecipitated the solubilized native dopa-mine receptors in an antibody concentration-dependent manner. In addition, they also bindselectively to the respective recombinant D2, D3, and D4 receptor membrane proteins fromcDNA transfected cells. Immunolocalization studies show that the D2-like receptor proteinshad differential regional and cellular distribution in the cerebral cortex, hippocampus, basalganglia, cerebellum, and midbrain, thus providing anatomical substrate for area-specificregulation of the dopamine neurotransmission. In cortical neurons, D4 receptor protein wasfound in both pyramidal and nonpyramidal cells, whereas D2 and D3 seem to be mostlyassociated with nonpyramidal interneurons. In rat hippocampus, the expression pattern ofD2-like receptors (D4.D3.D2) mirrored that obtained with immunoprecipitation studies. D2and D4 receptor immunolabeling was observed in the thalamic reticular nucleus, which wasnegative for the D3 subtype. Species differences were also observed; for example, the D4subtype receptor is the most highly expressed protein in the rat cortex, whereas it issignificantly less in human cortex. Differential patterns of D2, D3, and D4 receptor expressionin rat and human brain should shed light on the therapeutic actions of neuroleptic drugs andmay lead to the development of more specifically targeted antipsychotic drugs. J. Comp.Neurol. 402:353–371, 1998. r 1998 Wiley-Liss, Inc.
Indexing terms: D2 subtype; D3 subtype; D4 subtype; immunoblot; immunoprecipitation;
immunocytochemistry
Dopamine exerts its action through four different wellknown pathways and modulates motor, cognition, emo-tional, and endocrine functions. The nigrostriatal pathwayoriginates in the substantia nigra pars compacta andprojects to the striatum. The striatal structure has beenassociated with planning, initiation, and coordination involuntary movement (Gerfen, 1992) and also receivesmotivational, sensory, and motor inputs arriving by meansof afferents from cortical and limbic areas (Graybiel et al.,1994). The loss in dopaminergic neurons of substantianigra pars compacta and consequently the lack of dopa-mine innervation to the striatum is thought to be respon-
sible for the deficits in motor function seen in Parkinson’sdisease.
The two other pathways of dopamine neurotransmissionoriginate in the ventral tegmental area and constitute the
Grant sponsor: Spanish DGICYT; Grant number: PB94–0219-C02; Grantsponsor: Junta de Andalucia; Grant number: CTS-0161; Grant sponsor:European Union BIOMED I; Grant number: BMH1-CT94–1060.
*Correspondence to: Zafar U. Khan, M.S., Ph.D., Section of Neurobiology,School of Medicine, Yale University, C303 Sterling Hall of Medicine, 333Cedar Street, New Haven, CT 06520–8001.
Received 29 January 1998; Revised 28 May 1998;Accepted 10August 1998
THE JOURNAL OF COMPARATIVE NEUROLOGY 402:353–371 (1998)
r 1998 WILEY-LISS, INC.

mesolimbic and mesocortical systems. Mesolimbic dopa-mine neurons project to the nucleus accumbens, septum,olfactory tubercle, amygdaloid complex, and pyriform cor-tex. However, the mesoaccumbens neurons are thought tobe involved in mediating positive, reinforcing effects ofdrugs of abuse (Koob, 1992). The mesocortical neuronsproject to the medial prefrontal, cingulate, and entorhinalcortices (Grace, 1993). These areas are involved in theemotional, motivational, and cognitive functions and theirdysregulation may result in psychiatric disorders. How-ever, the tuberoinfundibular pathway originates in thehypothalamus and projects to the median eminence andintermediate lobe of the pituitary gland, where dopamineis responsible for the synthesis and release of prolactinand proopiomelanocortin-derived peptide, respectively(Kelly et al., 1997).
The modulatory actions of the dopamine are transmittedto the postsynaptic side of the neurons by interaction withfive distinct dopamine receptors, each of which is encodedby different gene. These receptors belong to the superfam-ily of G-protein–coupled receptors with seven transmem-brane domains. On the basis of the amino acid sequencesand structural and pharmacologic similarities, the fivedopamine receptors have been grouped into D1-like (D1and D5) and D2-like (D2, D3, and D4) families (for reviewsee, Sibley and Monsma, 1992; Civelli et al., 1993; Gingrichand Caron, 1993; Jackson and Westlind-Danielsson, 1994;O’Dowd et al., 1994; Neve and Neve, 1997). The analysis ofdopamine neurotransmission mediated by D2-like recep-tors is the focus of current research because these recep-tors have been implicated in several brain disorders suchas schizophrenia (Davis et al., 1991; Grace, 1993), Parkin-son’s disease (Albanese, 1990; Robertson, 1992), and drugaddiction (Sen and Lee, 1988). In particular, the typicaland atypical neuroleptic drugs, which are often used in thetreatment of schizophrenia, target D2-like receptors. Neu-roleptics interact with D2-like receptors and mediatebeneficial but many adverse effects (Baldessarini andTarazi, 1996; Neve and Neve, 1997; Tarazi et al., 1997;Joyce and Meador-Woodruff, 1997). Therefore, informationabout the precise localization of dopamine D2-like receptorsubtypes is pivotal for understanding their individualcontribution and function in the brain areas where thesedrugs target.
The distribution of dopamine D2-like receptors in therat, nonhuman primate, and human brain has been ana-lyzed by different methods such as autoradiography (Boy-son et al., 1986; Bouthenet et al., 1987; Charuchinda et al.,1987; De Keyser et al., 1988; Rakic et al., 1988; Camps etal., 1989; Lidow et al., 1989; Goldman-Rakic et al., 1989;Cox and Waszczak, 1991; Murray et al., 1992; Levant etal., 1993; Herroelen et al., 1994; Nobrega and Seeman,1994; Yokoyama et al., 1994, 1995; Booze and Wallace,1995; Lahti et al., 1995; Ricci et al., 1995; Hall et al.,1996a,b; Defagot and Antonelli, 1997), in situ hybridiza-tion (Meador-Woodruff et al., 1989, 1991, 1994; Mengod etal., 1989, 1992; Weiner and Brann, 1989; Mansour et al.,1990; Weiner et al., 1991; Bouthenet et al., 1991; Huntleyet al., 1992; Landwehrmeyer et al., 1993a,b; Diaz et al.,1995; Gaspar et al., 1995), and immunocytochemistry(McVittie et al., 1991; Brock et al., 1992; Ariano et al.,1993, 1997; Boundy et al., 1993a,b; Levey et al., 1993;Ariano and Sibley, 1994; Sesack et al., 1994; Yung et al.,1995; Mrzljak et al., 1996; Defagot et al., 1997). Althoughthe studies show a distribution restricted mostly to the
basal ganglia and midbrain, many of these results areconflicting. Some of the conflicting data might have arisenfrom the cross-reactivity of the probes with other un-wanted targets or a lack of reactivity with a target protein.For example, immunostaining of D2 receptors showed noor low labeling (Levey et al., 1993 and Sesack et al., 1994)or extensive immunostaining throughout the cortical lami-nae (Ariano et al., 1993). Therefore, our first objective wasto prepare highly specific and subtype-selective antibodiesto D2, D3, and D4 receptor subtypes. Then, these antibod-ies were used to analyze and compare the distribution ofdopamine D2-like receptors in all major rat brain struc-tures and some available human brain regions. From theresults obtained, it is evident that the three receptors aredifferentially localized and show distinct patterns of rela-tive concentrations in several brain areas, suggestingpotential differences in dopaminergic neurotransmissionwithin the discrete regions of brain.
MATERIALS AND METHODS
Materials
YM-09151–2[M-methyl-3H] (81.7 Ci/ mmol) was pur-chased from DuPont-New England Nuclear (Wilmington,DE). The (1) butaclamol-HCl was from Research Biochemi-cals, Inc. (Natick, MA). Peptides were synthesized by theCore Facility at the School of Biological Sciences, Univer-sity of Missouri-Kansas City (Kansas City, MO).
Preparation of anti-peptide antisera. The rat D2peptide HGLHSNPDSPAK corresponding to residues 284–295 (Bunzow et al., 1988) common to both long and shortforms (Giros et al., 1989; Monsma et al., 1989), the rat D3peptide RQRQRKRILTRQ corresponding to residues 216–227 (Sokoloff et al., 1990), and the human D4 peptideKLHGRAPRRPSG corresponding to residues 229–240 (VanTol et al., 1991) were synthesized. The sequence for D3does not differentiate between dopamine D3 and D3nf(Schmauss et al., 1993; Liu et al., 1994). An additionalcysteine was added at the carboxy end of each peptide forcoupling to keyhole limpet hemocyanin (KLH) protein.Peptide-KLH coupling was done by using the Pierce InjectActivated Immunogen Conjugation Kit (Pierce, Rockford,IL). New Zealand White rabbits (Animal facility at CharlesRiver, France) were immunized with 100 µg of peptide-KLH conjugate emulsified in complete Freund’s adjuvant,followed by subsequent boosts of same amount of peptide-KLH conjugate suspended in incomplete Freund’s adju-vant. A solid-phase ELISA with immobilized syntheticpeptides was used to monitor antibody production. Affinity-purification of the antisera was done on the correspondingantigen peptide immobilized columns as described indetail elsewhere (Gutierrez et al., 1994; Khan et al., 1994).
Preparation of membranes. Hippocampi were col-lected from 25 adult Sprague-Dawley rat brains afterdecapitation and homogenized (10 strokes) with 10% su-crose in 50 mM Tris-HCl, pH 7.4, by using 5 ml/g wettissue. The homogenate was centrifuged at 3,000 rpm for 5minutes in a Sorvall centrifuge with an SS-34 rotor. Thesupernatant was centrifuged at 29,000 rpm for 30 minutesin a Beckman ultracentrifuge with a type 30 rotor. Thepellet was washed twice in 50 mM Tris-HCl, pH 7.4. Themembranes were suspended at 5 mg/ml concentration in50 mM Tris-HCl, pH 7.4, aliquoted, and stored at -80°Cuntil they were used. The protein concentration of mem-branes was determined by Lowry et al. (1951).
354 Z.U. KHAN ET AL.

Solubilization and immunoprecipitation of dopa-
mine receptors. The dopamine receptors were solubi-lized with 1% digitonin (Gorrisen and Laduron, 1979) in 50mM Tris-HCl, pH 7.4, containing 1 mM ethylenediamine-tetraacetic acid, 5 mM KCl, 1.5 mM CaCl2, 4 mM MgCl2,120 mM NaCl, and 0.1% ascorbate, for 30 minutes at 4°C.After centrifugation at 29,000 rpm for 1 hour, the superna-tant was used for binding assays. Aliquots of 800 µl ofsolubilized receptor membranes (0.15–0.18 pmol of [3H]YM-09151–2 binding sites) were incubated overnight at 4°Cwith 5–120 µl of antisera. The receptor-antibody com-plexes were separated after incubation for 3 hours at 4°Cwith 80 µl of 40% (vol/vol) suspension of protein-A agarosefollowed by centrifugation. The nonimmunoprecipitatedsupernatant was saved for the radioligand binding, andthe pellet was discarded. For the binding of [3H]YM-09151–2, 200 µl of supernatant was incubated with 2 nM ofradioligand for 1 hour at 25°C in total volume of 500 µl of50 mM Tris-HCl (pH 7.4) containing 1 mM ethylenedi-aminetetraacetic acid, 5mM KCl, 1.5 mM CaCl2, 4mMMgCl2, and 120 mM NaCl. The reaction was stopped byrapid filtration through 0.3% polyethylenimine-soakedWhatman GF/B filters (Bruns et al., 1983) and washedthree times with 50 mM Tris-HCl, pH 7.4. Filters weredried at room temperature and counted for radioactivity.The nonspecific binding of the dopamine receptors wasdetermined in the presence of 2 µM (1) butaclamol-HCl.
Immunoblots. Immunoblots were performed accord-ing to Khan et al. (1994). Rat hippocampus membraneproteins (20 µg/lane) were separated by 10% sodiumdodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and transferred to nitrocellulose membranes. Thetransblotted nitrocellulose membranes were incubatedwith 5 µg/ml of affinity-purified primary antibodies (anti-D4, anti-D3, or anti-D2), which were diluted in phosphatebuffer saline (PBS) containing 0.05% Tween-20 and 1%bovine serum albumin, followed by anti-rabbit immuno-globulin G (IgG) (Sigma, St. Louis, MO) diluted 1:100 andperoxidase antiperoxidase complex (Sigma) diluted 1:100.The polypeptide bands were developed with 0.05% 3–3’-diaminobenzidine tetrahydrochloride as the chromogenfor the peroxidase reaction.
For the immunoblots of recombinant D4, D3, and D2membranes of Sf9 cells (BioSignal Inc., Quebec), 40 µg oftotal protein was electrophoresed on 10% SDS-PAGE andtransblotted onto nitrocellulose membranes. Membraneswere saturated by incubating with PBS containing 10%non-fat dry milk, 5% sheep serum, 5% donkey serum, and0.1% Tween-20 for 2 hours at room temperature. Followedby incubation with 5 µg/ml antibodies to D4, D3, and D2for overnight at 4°C. The secondary antibody, anti-rabbitIgG-HRP (Amersham, Arlington Height, IL) was diluted1:2000 and incubated for 1 hour. Immunoreactive bandswere visualized by using ECL kit as instructed by manufac-turer (Amersham). Immunoblots were scanned by UMax(UC 1260), and the page was created by using the AdobePhotoshop program.
Immunocytochemistry. The protocols used with ratsfor our studies were approved by the appropriate animalcare committee at University of Malaga. Twelve adultmale Sprague-Dawley rats (Charles River, France) ofabout 250 g of body weight each were anesthetized withketamine-HCl (40 mg/kg) and acepromazine maleate (1mg/kg) and perfused transcardially with 400 ml of 4%paraformaldehyde in 0.1 M phosphate buffer (PB), pH 7.4.
Brains were then removed and postfixed in the samefixative for 3 hours at 4°C, cryoprotected with 30% sucrose,and frozen in dry ice. Parasagittal and transverse sections(25 µm) were obtained with a freezing microtome (ReitcherJung, Leica, Germany). The postmortem (7–15 hoursinterval) human brain tissues were obtained from clinicalautopsy at Carlos Haya Hospital at Malaga (Malaga,Spain). These tissues were collected from five males 35–50years old without neurologic or psychiatric disorders. Allthe studies were conducted in conformance with thepolicies and guidelines of human protection and ethics.Human brain blocks of 5 mm2 from motor cortex (area 4),visual cortex (area 17), prefrontal cortex (area 46), cerebel-lum, and striatum were immersed in 4% paraformalde-hyde in PB, pH 7.4, for 48 hours at 4°C followed by 30%sucrose at 4°C until they sank. Frozen tissues were cut (40µm) with a freezing microtome and sections were stored at4°C in PB, pH 7.4, containing 0.02% sodium azide.
Free-floating sections were processed for immunocyto-chemistry as described earlier (Gutierrez et al., 1994). Allantibodies were diluted in 0.1 M PBS with 0.3% TritonX-100. Sections were incubated 48 hours at 4°C in one ofthe primary affinity-purified antibodies, i.e., anti-D4 (15µg/ml), anti-D3 (7 µg/ml), or anti-D2 (20 µg/ml). This stepwas followed by incubation in the secondary antibodysheep anti-rabbit IgG (Sigma) diluted 1:100 and then inrabbit peroxidase antiperoxidase complex (Sigma) diluted1:100. Both incubations were for 1 hour at room tempera-ture. Between incubation the sections were washed threetimes for 15 minutes each in 0.1 M PBS. The peroxidasereaction was developed in 0.05% 3–3’-diaminobenzidinetetrahydrochloride with or without 0.03% nickel ammo-nium sulfate and 0.01% H2O2 in 0.1 M PB, pH 7.4. Thesections were washed twice in PB, mounted on gelatin-coated slides, air dried, dehydrated with ethanol, clearedin xylene, and cover-slipped with Eukit mounting medium.
Control experiments were carried out with the primaryaffinity-purified receptor antibodies previously incubatedwith their respective complementary synthetic peptideantigen (75 µg/ml of D4, 35 µg/ml of D3, or 100 µg/ml ofD2). Under these control conditions, immunostaining wasabolished in all brain areas. In addition, tissue immunola-beling was not seen when the incubation with any of theprimary antibodies was omitted or preimmune sera wereused instead of primary antibodies. Furthermore, indi-vidual variation in results among the same species wereundetectable with the methods we have used to performthe experiments.
RESULTS
Subtype-specific antibodies to D2, D3,and D4 dopamine receptors
The polyclonal antibodies were prepared against syn-thetic peptides of dopamine D2, D3, and D4 receptors.These peptide sequences were selected to have epitopescommon to both rat and human species. Computer-assisted homology analysis of amino acid sequences showedthat D2, D3, and D4 peptide sequences are subtype-specific among all identified dopamine receptor subtypesand other G-protein–coupled receptors.
In Figure 1A, the immunoblots of rat hippocampalmembrane show that antibodies to D4 and D3 receptorsbound to a single polypeptide bands of 41 kDa and 49 kDa,respectively, and antibody to the D2 subtype reacted with
ANTIBODIES TO DOPAMINE D2-LIKE RECEPTORS 355

Fig. 1. Immunoblots of membranes from rat hippocampus (A) andrecombinant membranes of Sf9 cells expressing various dopaminereceptors (B) with anti-D4, anti-D3, and anti-D2 affinity-purifiedantibodies. In A, each lane contains the transfer of 20 µg of membraneprotein. All three antibodies showed immunoreactivity with singlepolypeptide bands of expected molecular sizes. As a control for thespecificity of immunoreactive bands, the antibodies were preincubatedwith 25 µg/ml of D4, D3, and D2 synthetic peptides, respectively. In B,
each lane represent the transfer of 4 µg of recombinant membraneproteins. Antibodies to D4 (a), D3 (b), and D2 (c) immunoreacted to 40kDa, 54 kDa, and 54 kDa, respectively, and these antibodies did notshow any reactivity with other members of the dopamine receptors.The concentration of the antibodies was 5 µg/ml in all incubations. Themolecular weight of the D2 dopamine receptor subtypes was deter-mined by comparison with the relative mobility of markers of knownmolecular weight.
356 Z.U. KHAN ET AL.

bands of 93 kDa and 49 kDa. However, when the antibod-ies were preincubated with their respective D4, D3, andD2 antigen peptide, the immunoreactive protein bandscompletely disappeared, indicating the specificity of immu-noreactive bands. The molecular size of the protein bandsis in good agreement with the predicted size of Mr 40,873for the human D4 receptor (Van Tol et al., 1991), 49,495 forthe rat D3 receptor (Sokoloff et al., 1990), and 47,544 forthe rat D2 receptor (Bunzow et al., 1988). However, theadditional band of 93 kDa that immunoreacted with theD2 subtype antibodies has also been identified by othergroups (Farooqui et al., 1991; Chazot et al., 1993). Never-theless, it has been observed that ligand and lectin-affinitypurified D2 receptors from brain migrated on SDS-PAGEat 92–95 kDa (Elazar et al., 1988; Williamson et al., 1988).Furthermore, when photoaffinity labeled D2 receptorswere deglycosylated, the migration of the protein band wasreduced to 40–44 kDa (Grigoriadis et al., 1988; Jarvie etal., 1988). Thus, the 93-kDa D2 immunoreactive bandcould represent a glycosylated part of this receptor. Thereis growing evidence for the existence of D2-subtype recep-tor dimers (Zawarynski et al., 1997; Lee et al., 1997), andthe 93-kDa size is in agreement with it. The 41-kDa bandobserved in rat hippocampus crude membrane with our D4antibody coincides with the protein size found by Mrzljaket al. (1996) with an antibody to GST-D4 in primate brain.The polypeptide band at 40–42 kDa has also been reportedwith an antipeptide antisera to the rat D4 receptor (De-fagot et al., 1997). The results of immunoblots indicate thesubtype specificity of these antibodies toward native pro-teins of unique sizes, probably of dopamine D2-like familyreceptors.
The antibodies were further tested for cross-reactivityand subtype specificity on another immunoblot analysiswith membranes of recombinant baculovirus-infected Sf9insect cells expressing various dopamine receptors. Figure1B represents immunoblots of the recombinant proteins ofD4 (a), D3 (b), and D2 (c) subtypes, in which antibodiesshowed the selective reactivity. Anti-D4 showed a 40-kDaimmunoreactive band, anti-D3 reacted to a 54-kDa pro-tein, and anti-D2 to a similar-sized 54-kDa band. Theseresults further show the subtype selectivity of these anti-bodies.
The antibodies to dopamine subtypes were able toimmunoprecipitate the radioligand [3H]YM-09151–2, whichis specific for dopamine D2-like (D2, D3, and D4) receptors,binding sites of native dopamine receptors. Antibody con-centration-dependent immunoprecipitation of digitonin-solubilized dopamine receptors from rat hippocampus isshown in Figure 2.
The nonimmunoprecipitated dopamine receptors weremeasured by [3H]YM-09151–2 binding in supernatant. Tocalculate the immunoprecipitated dopamine receptors, thenonimmunoprecipitated receptor in supernatant was de-ducted from the total amount of the dopamine receptors.
Fig. 2. Immunoprecipitation of the solubilized dopamine receptorsfrom rat hippocampus with anti-D4 (A), anti-D3 (B), and anti-D2 (C)antisera. The dopamine receptors present in supernatants (opencircles) were determined by [3H]YM-09151–2 binding. The amount ofprecipitated receptors (open squares) was calculated after subtractingthe values of supernatant from total specific binding (100%) of[3H]YM-09151–2. No precipitation activity was detected in the preim-mune sera. Data are mean 6 SEM values of three experiments, eachdone in duplicate.
ANTIBODIES TO DOPAMINE D2-LIKE RECEPTORS 357

Anti-D4 showed a saturation of immunoprecipitation at 80µl of antiserum (Fig. 2C), anti-D3 at 20 µl (Fig. 2B), andanti-D2 at 40 µl (Fig. 2A). The maximal immunoprecipita-tion achieved with 80 µl of anti-D4 was 51.37 6 4.88%, 20µl of anti-D3 was 42.36 6 1.64%, and 40 µl of anti-D2 was12.38 6 3.09%. The relative immunoprecipitation of D2-like receptors in hippocampus was D4D3D2. Chazot et al.(1993) have reported 10–20% immunoprecipitation of theD2 receptors from the solubilized preparation of bovinebrain. Another finding shows that antibodies to D2 and D3receptors immunoprecipitated 88% and 4–6% of the bind-ing sites for 125I-NCQ 298 from solubilized extracts of ratcaudate, respectively (Boundy et al., 1993a,b). However,there is no study reporting immunoprecipitation of nativedopamine D2-family receptors by using highly selectiveligand as it is shown in this study. Moreover, the sum ofhippocampal dopamine D4, D3, and D2 receptors immuno-precipitation was 106.1 6 4.7%, which means that all ofthe [3H]YM-09151–2 binding sites of D2-like receptorswere immunoprecipitated with the antibodies to D4, D3,and D2 subtype, which further reflects the high specificityof these antibodies. It seems that the specific proteinsthese antibodies bind in native membrane are dopamineD2-family receptors.
Moreover, the reactivity of immunoblots, immunoprecipi-tations, immunocytochemistry with antibodies to D2, D3,
and D4 were specifically abolished when they were preab-sorbed with their respective antigen peptides. The reactiv-ity was also not seen with the omission of antibodies orwhen preimmune sera was used instead of antibodies.Taken together, all the results clearly establish the sub-type specificity and selectivity of these antibodies to dopa-mine D2-like receptors.
Localization of D2-like receptor proteins
Immunoreactive cell bodies, dendrites, and axonal pro-cesses were seen in several brain regions with the affinity-purified antibodies to D2, D3, and D4 dopamine receptors(Fig. 3). Although D2-like receptors are localized in similarbrain areas (cerebral cortex, striatum, hippocampus, hypo-thalamus, cerebellum, septum, substantia nigra pars re-ticulata, olfactory tubercle, and inferior colliculus), theyoften exhibit complementary distribution and significantsubtype-specific differences in labeling intensity and cellu-lar localization. For example, D4 immunolabeling (Fig.3A,B) was concentrated in cerebral cortex, hippocampus,caudate putamen, and inferior colliculus, whereas, D3(Fig. 3C) and D2 (Fig. 3D) proteins were more abundant inforebrain subcortical areas. D4 receptor immunoreactivitywas seen mainly as reaction product along plasma mem-brane and/or within cytoplasm of cells bodies and den-
Fig. 3. Rat brain immunocytochemistry with affinity-purified anti-bodies to D4 (A,B), D3 (C), and D2 (D) receptor proteins. D4immunolabeling is mainly concentrated in cerebral cortex, hippocam-pus, caudate putamen, and inferior colliculus, whereas D3 and D2receptors are both enriched in subcortical forebrain areas. B: Rostraltransverse section showing intense D4 immunostaining in frontal andparietal cortices, moderate in cingulate cortex, and weak in pyriformcortex; immunoreactivity in caudate putamen and corpus callosum is
also prominent. A, C, and D are parasagittal sections. Ac, nucleusaccumbens; cc, corpus callosum; Cg, cingulate cortex; Ctx, cerebralcortex; Cb, cerebellum; CPu, caudate-putamen; Fr, frontal cortex; GP,globus pallidus; H, hippocampus; Hp, hypothalamus; IC, inferiorcolliculus; M, medulla; OB, olfactory bulb; OT, olfactory tubercle; P,parietal cortex; Pir, piriform cortex; S, septum; SC, superior colliculus;Th, thalamus. Scale bar 5 2.2 mm in A,C,D, 1 mm in B.
358 Z.U. KHAN ET AL.

drites, whereas D2 and D3 receptor immunoreactivitieswere mostly concentrated in the neuropil. However, acytoplasmic immunostaining outlining neuronal cell bod-ies and initial dendrites could be seen with D2, D3, and D4antibodies (Fig. 5A-G). To analyze the distribution of theD2-like dopamine receptor proteins, we will describe thelocalization of D4 receptor protein and compare the distri-bution with those of D2 and D3 receptor proteins.
Cerebral cortex. D4 receptor protein was the mosthighly expressed D2-like protein in the rat cerebral cortex(Figs. 3, 4). This brain area and the hippocampal formationshowed the highest level of D4 receptor protein expressionin the rat brain. Moreover, we have also seen the highestamount of D4 receptor present in hippocampus by immuno-precipitation compared with D2 and D3 subtypes (Fig. 2).Neurons expressing D4 receptor protein were detected inmost areas of the cerebral cortex. Frontal, temporal,parietal, and occipital cortices showed intense D4 receptorimmunostaining (Fig. 3A,B); however, cingulate cortexshowed moderate immunolabeling (Fig. 3B). Pyriform,entorhinal, and retrosplenial cortices displayed the weak-est cortical immunolabeling. (Figs. 3B, 7D). D4 immuno-staining was observed in a considerable population ofneurons in layers II–V of cerebral cortex (Fig. 4A). Bothpyramidal and nonpyramidal neurons were labeled (Figs.4A, 5A–C). However, a bilaminar distribution of D4 posi-tive cells could be seen in all cortical areas. A prominentand intense immunoreactive cell band was located atlayers II-III and a less dense band at layers IV–V. Thelatter band was much more prevalent at the occipitalcortex compared with the other cortical areas. Immunore-active cells were weakly labeled in layer VI. Neuropilimmunostaining was seen from layers I to VI (Fig. 3A,B),but it was more prominent at layers I–III. An intense D4immunoreactivity was also seen in the corpus callosumfiber bundle (Fig. 3B). On the other hand, antibodies to D2and D3 receptor proteins labeled mostly neuropil (Fig.3C,D) and weakly outlined neural cell bodies (Fig. 4B,C) inall cortical regions. However, some neurons from layerIV–V exhibited intense D2 or D3 immunoreactivitiesassociated with somata and proximal portions of dendrites(Fig. 5D–G) as seen for D4 antibody (Fig. 5A–C).
In human brain, motor (area 4), visual (area 17) andprefrontal (area 46) cortices showed immunopositive neu-rons to D4, D3, and D2 antibodies (Fig. 6A–C). Visual andmotor cortices presented more intense D4, D3, and D2immunolabeling than prefrontal cortex. In all three areas(4, 17, and 46) D4, D3, and D2 labeled neurons werelocated in all cortical layers, but particularly in layersIV–V. Labeling was predominantly associated with neuralcell bodies and initial dendritic segments. Pyramidal andnonpyramidal D4 positive neurons were visualized (Fig.6A,D). However, D3 (Fig. 6B,E) and D2 (Fig. 6C,F) immu-nostaining was mostly associated to nonpyramidal neu-rons.
Hippocampal formation. D4, D3, and D2 receptorantibodies showed subtype-specific immunostaining pat-terns in the hippocampal formation (Fig. 7A–C). Pyrami-dal cells of CA1 field were heavily immunostained alongwith the neuropil of the stratum oriens and stratumradiatum with anti-D4 (Fig. 7A). The transition from CA1to CA2/3 fields was very clearly delimited by the low levelof expression of D4 receptor protein by CA2/3 pyramidalcells (Figs. 7A, 8A). A bundle of immunoreactive processescould be seen running along the CA2/3 pyramidal cell layer
until the hilar region (Figs. 7A, 8B). In the dentate gyrus,D4 immunoreactivity was seen in granule cell bodies andtheir dendrites were seen in the molecular layer (Figs. 7A,8B). In the subicular region, numerous neurons showed ahigh expression of D4 protein (Fig. 8D,E). The alveus andfornix also were immunostained with anti-D4 receptorantibody. In the hippocampus proper and dentate gyrus,anti-D2 and anti-D3 immunoreactivities were only local-ized to the neuropil (Fig. 7B,C). Anti-D3 immunoreactivitywas concentrated in the stratum oriens and stratumradiatum of CA1 field (Fig. 7B), and in the inner third ofthe molecular layer of the dentate gyrus (Figs. 7B, 8F).Weak D2 immunolabeling was seen in the dendritic fieldsof hippocampal pyramidal cells without clear differencesin the labeling intensity among the hippocampus propersubfields (Fig. 7C). In the dentate gyrus, the molecularlayer also showed weak D2 immunostaining. However, astrong D2 neuropil immunoreaction was detected in thehilar region and could be followed until the CA2/3 regionfollowing the same course as mossy fibers (Figs. 7C, 8C). Inthe subicular region, few cell bodies with cytoplasmicimmunostaining were observed with both anti-D2 andanti-D3 antibodies (not shown). The relative immunostain-ing of D2-like receptor was D4D3D2.
Subcortical forebrain areas. Numerous medium-sized D4 receptor positive neurons and a prominent stainedneuropil was present throughout the caudate putamen(Fig. 3A,B). Neuronal cell bodies and neuropil showed D4immunoreactivity in the shell of nucleus accumbens (Fig. 9A,B). These D4-positive cells in the nucleus accumbenswere more abundant in the most rostral portion (Fig. 9B).Intense and homogeneous neuropil immunostaining wasrevealed with anti-D2 and anti-D3 in the striatum andnucleus accumbens (Fig. 3C,D). D2 was also present in thestriatal neurons. The globus pallidus showed scattered cellbodies immunoreactive to D2, D3, and D4 receptors anti-bodies. The neuropil of the septal area appeared immu-nopositive to the three dopamine D2-like receptor subtypeantibodies (Fig. 3). Abundant D4 receptor–positive cellbodies and their processes were revealed in the amygdalarcomplex. The olfactory tubercle was also immunostainedwith the three D2-like receptor antibodies (Fig. 3 A,C,D).Weak neuropil immunolabeling was seen with anti-D4antibody in the olfactory tubercle (Fig. 3A), although fewclusters of positive cells could be observed (Fig. 9C).Scattered D4-positive cells were present in the anteriorolfactory nucleus (Fig. 9D). On contrary, intense immuno-labeling of the olfactory tubercle was found with anti-D3(Fig. 3C) and anti-D2 (Fig. 3D) antibodies. The striaterminalis and the lateral division of the bed nucleus wereimmunolabeled with D4 receptor antibody (Fig. 10B). Inthe striatal compartments of human brain caudate andputamen, a diffuse neuropil immunostaining and numer-ous small size immunostained neurons could be seen withall three D2-like receptor antibodies (Fig.11A-C).
Thalamus. The D4 and D2 immunolabeling outliningcells bodies in the reticular nucleus was observed (Figs. 9F,10B). This nucleus was negative for anti-D3 antibody.Neuropil immunostaining was seen in the lateral dorsalthalamic nucleus with the three D2-like subtype receptorantibodies (D3.D2.D4) (Figs. 3A,C,D).
Hypothalamus. This brain area showed the strongestD2-like immunoreaction product (D2.D3D4) in the dien-cephalon (Fig. 3A,C,D). D2, D3, and D4 immunostaining
ANTIBODIES TO DOPAMINE D2-LIKE RECEPTORS 359

was seen in arcuate, supraoptic, suprachiasmatic, andmammillary nuclei (Fig. 9E).
Midbrain. D4 positive cell bodies were found in thesuperficial gray layer of the superior colliculus (Fig. 10
A,C). Moderate neuropil immunostaining was seen withanti-D2 and anti-D3 antibodies in this layer. The otherlayers of the superior colliculus did not show immunostain-ing with all three dopamine receptor antibodies. The
Fig. 4. Rat parietal cerebral cortex immunostained with anti-D4(A), anti-D3 (B), and anti-D2 (C) dopamine receptor antibodies. D4immunoreactivity is much more intense than D3 and D2. Note strong
D4-immunoreaction product in cell bodies and dendrites of pyramidaland nonpyramidal cortical neurons. I–IV represent the cortical layers.Scale bar 5 67 µm (applies to A–C).
360 Z.U. KHAN ET AL.

inferior colliculus was intensely labeled with anti-D4antibody (Fig. 10A,D). Numerous D4 cells bodies and theirprocesses could be seen throughout the rostrocaudal exten-sion of the inferior colliculus dorsal cortex. D2- and lessD3-positive cells were mainly observed in the centralnucleus of the inferior colliculus (Fig. 10E). Prominentneuropil immunolabeling appeared in the substantia nigrapars reticulata with anti-D4, anti-D3, and anti-D2 antibod-ies. D4-positive processes could be followed in sagittalbrain sections from the substantia nigra to the striatum
(Fig. 10B). D2 subtype was predominant in the substantianigra-pars compacta, ventral tegmental area, and retroru-bral field.
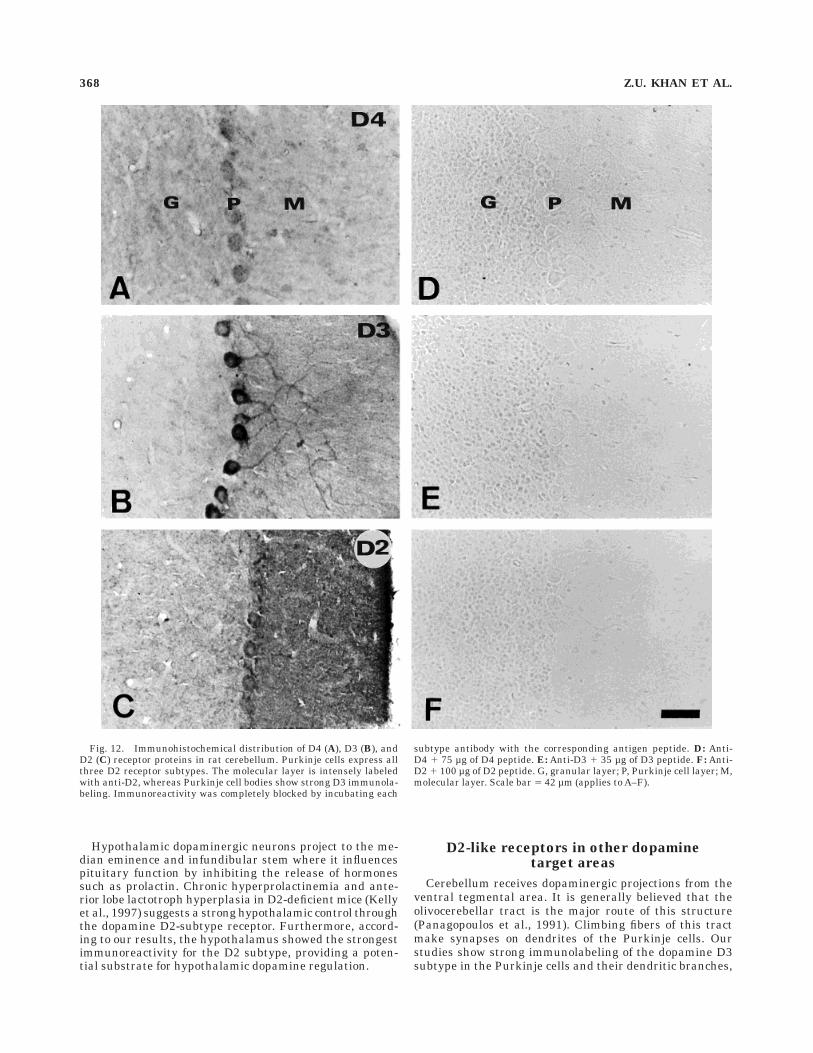
Cerebellum
D4, D3, and D2 immunoreactivities were localized toPurkinje cells (Fig. 12A-C). The strongest immunostainingof Purkinje cell bodies was seen with D3 antibody. Themolecular layer was more densely labeled with anti-D2
Fig. 5. Higher magnification photomicrographs of cortical neuronsimmunolabeled with anti-D4 (A–C), anti-D3 (D,E), and anti-D2 (F,G)affinity-purified antibodies. Punctuate cytoplasmic immunostaining
outlines cell bodies and proximal dendrites. D4 cytoplasmic immunola-beling was also seen in nonpyramidal (B) and pyramidal (C) neurons.Scale bar 5 13 µm (applies to A–G).
ANTIBODIES TO DOPAMINE D2-LIKE RECEPTORS 361

Fig. 6. Human motor cortex (Brodmann area 4) immunocytochem-istry with anti-D4 (A), anti-D3 (B), and anti-D2 (C) affinity-purifiedreceptor antibodies. D4, D3, and D2 labeled neurons are mainlylocalized to layers IV–V. Pyramidal neurons (thick arrows) are more
frequently labeled with anti-D4. Numerous nonpyramidal neurons(thin arrows) appear labeled with all three D2-like receptor antibod-ies. Higher magnification of D4 (D), D3 (E), and D2 (F) nonpyramidalcells from layer IV. Scale bar in C 5 148 µm in A–C, 15µm in D–F.
362 Z.U. KHAN ET AL.

than with any of the other two D2-like subtype antibodies.D4 receptor protein expression in the cerebellum waslower than D3 and D2 proteins. No immunoreactivity wasseen throughout the brain when the antibodies wereincubated with their respective antigen peptides beforethe experiments as shown here for the cerebellum (Fig.12D-F).
In the cerebellum of human brain, D4, D3, and D2dopamine receptor proteins were expressed by Purkinjecells (Fig.13A,C,D). D4, D3, and D2 immunoreaction prod-
ucts were concentrated within cytoplasm of Purkinje cellbodies and dendrites. No immunoreactivity was seen incerebellar molecular or granular layers. In human controlsections, immunoreaction products were not seen whenusing antibodies previously incubated with the correspond-ing peptide antigen, as shown in the cerebellum for D4antibody (Fig.13B).
DISCUSSION
The present study of dopamine receptors immunostain-ing in rat and human brain showed a distinct differences inregional distribution of D4, D3, and D2 subtypes withinD2-like family of receptors. We also found that these threereceptors are frequently expressed in different neuronalpopulations throughout the brain. Therefore, our compara-tive results of distribution of D2-like receptors provide asubstantial evidence for differential region- and cell-specific dopaminergic regulation by each receptor sub-types.
Specificity and selectivity of antibodies
The subtype-specific antibodies were produced from thepeptides of dopamine D2, D3, and D4 receptors, whichwere derived from the third and large intracellular looplocated between the putative transmembrane domains 5and 6. The specificity of antibodies was established on thebasis of several studies: (1) the sequence homology studyshow that selected peptide sequences of D2, D3, and D4are highly divergent and unique for its subtype; (2) in rathippocampal membrane immunoblots, antibodies to dopa-mine D2, D3, and D4 receptors reacted to unique polypep-tide bands of expected molecular sizes as predicted frommolecular cloning (Bunzow et al., 1988, Sokoloff et al.,1990, Van Tol et al., 1991); (3) antibodies specificallyrecognize the corresponding receptor proteins expressed inrecombinant Sf9 cells and showed no cross-reactivity withthe other member of dopamine receptor family; (4) antibod-ies also immunoprecipitated the native dopamine recep-tors from hippocampal membranes, moreover, after combin-ing these three antibodies, they were able toimmunoprecipitate all (100%) D2-like receptors activity,suggesting that these antibodies bind exclusively to D2-type receptors and also do not cross-react between eachother; and (5) immunoreactivity of immunoblots, immuno-precipitation, and brain sections was specifically elimi-nated by the preincubation of antibodies with their peptideantigen. But, when the antibodies were preincubated withnoncognate antigen peptide, they did not block the immu-noprecipitation, again reconfirming the non–cross-reactiv-ity of these antibodies between each other. From theseresults, it seems that possibility for reactivity of theseantibodies to other unrelated proteins is highly unlikely.
Cortical system of dopamineD2-like receptors
In cerebral cortex, we found that the dopamine D4receptor is expressed by both pyramidal and nonpyramidalneurons, whereas the D3 and D2 subtype were mainlypresent in the nonpyramidal interneurons. Interaction ofdopamine with its neuronal receptors play a major role inregulation of excitability of pyramidal neurons by directand indirect mechanisms (Goldman-Rakic, 1998). In pre-frontal cortex, these mechanisms control the efficacy of theworking memory functions. The direct mechanism in-
Fig. 7. Immunocytochemical localization of D4 (A), D3 (B), and D2(C) receptor proteins in rat hippocampus. CA1, CA2, and CA3,hippocampus proper fields; HL, hilus; DG, dentate gyrus. Scale bar 50.42 mm (applies to A–C).
ANTIBODIES TO DOPAMINE D2-LIKE RECEPTORS 363

Fig. 8. Anti-D4 (A, B, D, and E), anti-D2 (C) and anti-D3 (F)immunoreactivity in the rat hippocampal formation. A: Pyramidalcells from CA1 field exhibit high expression of D4 protein, whereasCA2 neurons show very low or no expression. B: Dentate gyrusgranule cells and their dendrites in the molecular layer display strongD4 immunoreactivity as well as some neurons displaced from CA2/CA3 region and the neuropil located in the pyramidal cell layer ofCA2/CA3 fields. D: Transition between the retrosplenial cortex and
the subicular region immunolabeled with anti-D4. E: Subicular neu-rons showing strong D4 immunostaining. C: Dentate gyrus immunocy-tochemistry with anti-D2. F: Diffuse neuropil immunostaining withanti-D3 in the inner third of the molecular layer in the dentate gyrus.DG, dentate gyrus; G, granule cell layer; HL, hilus; M, molecularlayer; RS, retrosplenial cortex; Sb, subiculum; SO, stratum oriens; SP,stratum pyramidale; SR, stratum radiatum. Scale bar 5 67µm inA,B,F, 133 µm in C,D, 34 µm in E.
364 Z.U. KHAN ET AL.

volves the dopamine modulation on pyramidal neurons,whereas indirect mechanism acts through nonpyramidalGABAergic neurons by feed-forward inhibition on pyrami-dal neurons. Thus, it is likely that most of the dopaminemodulation of D2-like receptors are preferentially drivenby an indirect mechanism in cortical circuits.
The distinctive regional and laminar patterns of D2-likereceptors throughout the cortical areas suggest that dopa-mine modulation is area specific. We observed a bilaminardistribution of dopamine D4 receptor in prefrontal cortexsimilar to that of the dopamine transporter (Horn, 1990).
Dopamine axons are known to make synaptic contactswith pyramidal cells and a subset of GABAergic interneu-rons of middle layer, which are parvalbumin positive(Conde et al., 1994), where we have also seen interneuronscontaining D2-like receptors. These interneurons are parv-albumin positive too (unpublished observation). Therefore,the typical antipsychotic drugs such as haloperidol, whichhas higher affinity for the D2 subtype, probably enforcesits effect indirectly through interneurons to pyramidalcells; however, an atypical drug such as clozapine, whichhas higher affinity for the D4 subtype, may act on pyrami-
Fig. 9. D4 immunoreactive neurons in the accumbens nucleus (A,B), olfactory tubercle (C,D),mamillary nucleus (E), and reticular thalamic nucleus (F) of rat brain. Ac, accumbens nucleus; AO,anterior olfactory nucleus; Ctx, cortex. Scale bar 5 400 µm in A, 80 µm in B,C,E,F, 16 µm in D.
ANTIBODIES TO DOPAMINE D2-LIKE RECEPTORS 365

dal neurons through both direct and indirect mechanisms.This may be the reason for beneficial effect of clozapine(Joyce and Meador-Woodruff, 1997) at least at the corticallevel. The antagonism effect of antipsychotic drugs on thepyramidal neurons is probably achieved by direct synapticinteraction, but this effect is reversed when the drug actionis processed through GABAergic interneurons by an indi-rect pathway.
The principal projection neurons of cerebral cortex arepyramidal cells that use glutamate as an excitatory neuro-transmitter and output projection from this structure tothe striatum are through corticostriate or corticothalamo-striate pathways. Dopamine D2-like receptors may notactively participate in this pathway because most arepresent in the GABAergic nonpyramidal neurons, exceptthe dopamine D4 receptors. Thus, our results support thehypothesis of the contribution of cerebrocortical D2 andstriatolimbic D4 receptors to the antipsychotic actions ofneuroleptic drugs (Tarazi et al., 1997).
D2-like receptors in hippocampal function
The D4 subtype is the predominant D2-like receptor inthe hippocampus, particularly in the CA1 region. Thedopaminergic fibers from the ventral tegmental area ofmidbrain target to the hippocampus, which is associatedwith memory function and is critical for everyday episodicmemory (Vargha-Khadem et al., 1997). It is well knownthat lesions of the hippocampus in human and othermammals produce severe amnesia for certain memories(Squire, 1987). Long-term potentiation (LTP) and long-term depression (LTD) function as a gate for synapticplasticity in hippocampus. Furthermore, our results showa heavy labeling of only the D4 subtype in the pyramidalcells of CA1 field. The large pyramidal cell populations ofCA1 discharge electrical activity patterns, which integrateinto specific stages of memory formation. Both LTP andLTD are postulated as cellular substrates for certain typesof learning and memory (Bliss and Collingridge, 1993). It
Fig. 10. A: D4 immunoreactivity in the rat inferior and superiorcolliculus. B: D4 immunoreactive processes can be visualized from thesubstantia nigra toward the striatum in this parasagittal section.C: Superior gray layer of superior colliculus showing D4 immunoreac-tive neurons. D4 (D) and D2 (E) immunoreactive neurons from the
inferior colliculus. CPu, caudate putamen; IC, inferior colliculus; Rt,reticular thalamic nucleus; SC, superior colliculus; St, stria termina-lis; SN, substantia nigra; Th, thalamus. Scale bar in E 5 400 µm in A,158 µm in B, 40 µm in C–E.
366 Z.U. KHAN ET AL.

has been shown that the blockade of D2-like receptorseliminated the late phase of LTP in hippocampus (Frey etal., 1990). Therefore, it is reasonable to assume that thedopamine D4 receptor subtype of the D2-like family maybe one of the required elements for LTP modulation in theCA1 area.
D2-like receptors in basal ganglia
Our results show the predominance of D2 and D4receptor protein in the caudate putamen complex, whereasthe D3 subtype was restricted to the olfactory tubercle andventral striatum. Both the D2 and D4 receptors werefound in the medium-sized spiny neurons, which are
known to be GABAergic. These medium sized-GABAergicstriatal neurons project to the substantia nigra, wherethey inhibit the tonic activity in the GABAergic neurons ofsubstantia nigra-pars reticulata (Chevalier et al., 1985;Gerfen, 1992). The functional and anatomical aspect of theD2 receptor localization in striatal GABAergic neuronshas been well described (Yung et al., 1995; Hersch et al.,1995). However, the abundance of D4 subtype was surpris-ing because Mrzljak et al. (1996) have reported a lowamount of this receptor in primate striatum. This varia-tion may be attributed to species differences. The expres-sion of the D4 receptor in GABAergic neurons suggests itsrole in modulation of the striatal output system by meansof a ‘‘direct’’ or ‘‘indirect’’ pathway, which projects to theinternal and external segment of the globus pallidus,respectively (Gerfen, 1992). However, it remains unclearwhether the D4 subtype is localized in D1-positive GABAer-gic neurons of the ‘‘direct’’ or D2-positive GABAergicneurons of the ‘‘indirect’’ pathway. In Parkinson’s disease,the dopamine transmission through an ‘‘indirect’’ path-way is increased, resulting in hypokinesia (Hersch et al.,1995).
The reinforcing properties of drugs of abuse are thoughtto be mediated through limbic dopamine projections, par-ticularly in the nucleus accumbens (Koob, 1992). Theability of drugs of abuse, including cocaine, methamphet-amine, and alcohol, to increase the dopamine level in thecerebral cortex and nucleus accumbens has been impliedas a reason for their positive reinforcing properties (Kuharet al., 1991). The inforcing and additive properties can beattenuated by selective dopamine D2 receptor antagonists(Harris and Auston-Jones, 1994), thus, indicating a role ofdopamine D2-like receptors in addictive and reward mecha-nisms of nucleus accumbens, where we have localized thethree subtypes of dopamine D2-like receptors.
Dopaminergic system of D2-like receptors
Among the three receptors, the D2 subtype was predomi-nantly found in the dopaminergic neurons of the substan-tia nigra-pars compacta. The striatonigral pathway origi-nates from the substantia nigra-pars compacta and projectsto the basal ganglia, which is primarily involved in motorfunction and where mainly the D2 receptor was found. Thelack of dopamine innervation of this pathway is believed tobe responsible for the loss in motor function seen in theParkinson’s disease, suggesting an important role for theD2 subtype of dopamine receptor.
The regulation of neurotransmitter synthesis and re-lease is crucial for normal brain function. Malfunctioningof these processes leads to a number of neuropsychiatricdiseases, and many antipsychotic drugs exert their effectsthrough this mechanism. Dopamine neurons synthesizeautoreceptor, which appears to be exclusively a D2-likedopamine receptor (Sesack et al., 1994, 1995). Our studiesshow the presence of D2 subtype in dopaminergic neuronsof the mesencephalon, which may be an autoreceptor.However, mounting evidence suggests that the D2 (Craggand Greenfield, 1997) as well as D3 (Aretha and Galloway,1996) receptor subtypes function as autoreceptors. Further-more, recent studies have shown the loss of autoreceptorfunction in D2-deficient mice (Mercuri et al., 1997), whereasthis function was preserved in D3-mutant mice (Koeltzowet al., 1998), suggesting that the D2 subtype is the onlyautoreceptor within the dopamine system.
Fig. 11. Putamen compartment of human striatum immunola-beled with anti-D4 (A), anti-D3 (B), and anti-D2 (C) antibodies. Allthree D2 receptor subtypes are enriched in the neuropil. ic, internalcapsula; Pu, putamen. Scale bar 5 417 µm in C (applies to A–C).
ANTIBODIES TO DOPAMINE D2-LIKE RECEPTORS 367
Hypothalamic dopaminergic neurons project to the me-dian eminence and infundibular stem where it influencespituitary function by inhibiting the release of hormonessuch as prolactin. Chronic hyperprolactinemia and ante-rior lobe lactotroph hyperplasia in D2-deficient mice (Kellyet al., 1997) suggests a strong hypothalamic control throughthe dopamine D2-subtype receptor. Furthermore, accord-ing to our results, the hypothalamus showed the strongestimmunoreactivity for the D2 subtype, providing a poten-tial substrate for hypothalamic dopamine regulation.
D2-like receptors in other dopaminetarget areas
Cerebellum receives dopaminergic projections from theventral tegmental area. It is generally believed that theolivocerebellar tract is the major route of this structure(Panagopoulos et al., 1991). Climbing fibers of this tractmake synapses on dendrites of the Purkinje cells. Ourstudies show strong immunolabeling of the dopamine D3subtype in the Purkinje cells and their dendritic branches,
Fig. 12. Immunohistochemical distribution of D4 (A), D3 (B), andD2 (C) receptor proteins in rat cerebellum. Purkinje cells express allthree D2 receptor subtypes. The molecular layer is intensely labeledwith anti-D2, whereas Purkinje cell bodies show strong D3 immunola-beling. Immunoreactivity was completely blocked by incubating each
subtype antibody with the corresponding antigen peptide. D: Anti-D4 1 75 µg of D4 peptide. E: Anti-D3 1 35 µg of D3 peptide. F: Anti-D2 1 100 µg of D2 peptide. G, granular layer; P, Purkinje cell layer; M,molecular layer. Scale bar 5 42 µm (applies to A–F).
368 Z.U. KHAN ET AL.

suggesting a functional role of this receptor in the cerebel-lum (Barik and deBeaurepaire, 1996). It seems that the D3subtype primarily serves the dopaminergic pathway in thecerebellum.
The differential distribution was also evident in thethalamic reticular nucleus, where the D2 and D4 subtypesof the dopamine receptor were expressed and D3 wasabsent. Reticular GABAergic neurons inhibit thalamocor-tical activity through their projections to dorsal thalamicnuclei (Steriade et al., 1993).
In conclusion, the distinct heterogeneous localization ofthe D2-like receptors at the potential dopamine interac-tion sites suggests their unique and individual role inregulation of dopaminergic neurotransmission. The pres-ence of the dopamine D4 subtype in cortical pyramidal aswell as in nonpyramidal cells suggests that the antipsy-chotic action of clozapine, a D4 antagonist, may be achievedby a balanced performance of both cell types. In contrast,haloperidol, which is a D2 agonist, presumably acts throughnonpyramidal cells and shows a less beneficial effect thanclozapine, suggesting that a direct interaction with thepyramidal cells in cortex may be the key to the beneficialeffect of clozapine in psychiatric patients.
ACKNOWLEDGMENTS
We thank Marıa Jose Lopez-Alvarez for her assistancewith the immunocytochemical experiments. During this
research, Z.U. Khan received fellowships from Junta deAndalucia, Universidad de Malaga, and European UnionBIOMED.
LITERATURE CITED
Albanese, A. (1990) Extrapyramidal system, motor ganglia and movementdisorders. Rev. Neurosci. 2:145–163.
Aretha, C.W. and M.P. Galloway (1996) Dopamine autoreceptor reserve invitro: Possible role of dopamine D-3 receptor. Eur. J. Pharmacol.305:119–122.
Ariano, M.A. and D.R. Sibley (1994) Dopamine receptor distribution in therat CNS: Elucidation using anti-peptide antisera directed against D1Aand D3 subtypes. Brain Res. 649:95–110.
Ariano, M.A., R.S. Fisher, E. Smyk-Randall, D.R. Sibley, and M.S. Levine(1993) D2 dopamine receptor distribution in the rodent CNS usinganti-peptide antisera. Brain Res. 609:71–80.
Ariano, M.A., J. Wang, K.L. Noblett, E.R. Larson, and D.R. Sibley (1997)Cellular distribution of the rat D4 dopamine receptor protein in theCNS using anti-receptor antisera. Brain Res. 752:26–34.
Baldessarini, R.J. and F.I. Tarazi (1996) Brain dopamine receptors: Aprimer on their current status, basic and clinical. Harv. Rev. Psychiatry3:301–325.
Barik, S. and R. deBeaurepaire (1996) Evidence for a functional role of thedopamine D3 receptor in the cerebellum. Brain Res. 737:347–350.
Bliss, T.V.P. and G.L. Collingridge (1993) A synaptic model of memory: Longterm potentiation in the hippocampus. Nature 361:31–39.
Booze, R.M. and D.R. Wallace (1995) Dopamine D2 and D3 receptors in therat striatum and nucleus accumbens: Use of 7-OH-DPAT and [125I]-iodosulpiride. Synapse 19:1–13.
Fig. 13. Human cerebellum immunocytochemistry with anti-D4(A), anti-D3 (C), and anti-D2 antibodies (D). Purkinje cells express allthree subtypes of D2 dopamine receptor proteins. B: Antibodiespreviously incubated with their corresponding antigen peptide yielded
no tissue immunoreaction product as shown here for anti-D4 in acerebellar section. G, granular layer; P, Purkinje cell layer; M,molecular layer. Scale bar 5 42 µm (applies to A–D).
ANTIBODIES TO DOPAMINE D2-LIKE RECEPTORS 369

Boundy, V.A., R.R. Luedtke, R.P. Artymyshyn, T.M. Filtz, and P.B. Molinoff(1993a) Development of polyclonal anti-D2 dopamine receptor antibod-ies using sequence-specific peptides. Mol. Pharmacol. 43:666–676.
Boundy, V.A., R.R. Leudtke, A.L. Gallitano, J.E. Smith, T.M. Filtz, R.G.Kallen, and P.B. Molinoff (1993b) Expression and characterization ofthe rat D3 dopamine receptor: Pharmacological properties and develop-ment of antibodies. J. Pharmacol. Exp. Ther. 264:1002–1011.
Bouthenet, M.L., M.P. Martres, N. Sales, and J.C. Schwartz (1987) Adetailed mapping of dopamine D2 receptors in rat central nervoussystem by autoradiography with [125I]iodosulpiride. Neuroscience 20:117–155.
Bouthenet, M.L., E. Souil, M.P. Martres, P. Sokoloff, B. Giros, and J.C.Schwartz (1991) Localization of dopamine D3 receptor mRNA in the ratbrain using in situ hybridization histochemistry: Comparison withdopamine D2 receptor mRNA. Brain Res. 564:203–219.
Boyson, S.J., P. McGonigle, and P.B. Molinoff (1986) Quantitative autoradio-graphic localization of the D1 and D2 subtypes of dopamine receptors inrat brain. J. Neurosci. 6:3177–3188.
Brock, J.W., S. Farooqui, K. Ross, and C. Prasad (1992) Localization ofdopamine D2 receptor protein in rat brain using polyclonal antibody.Brain Res. 578:244–250.
Bruns, R.F., K. Lawson-Wendling, and T.A. Pugsley (1983) A rapid filtrationassay for soluble receptors using polyethylenimine-treated filters. Anal.Biochem. 132:74–81.
Bunzow, J.R., H.H.M. Van Tol, D.K. Grandy, P. Albert, J. Salon, M. Christie,C.A. Machida, K.A. Neve, and O. Civelli (1988) Cloning and expressionof a rat D2 dopamine receptor cDNA. Nature 336:783–787.
Camps, M., R. Cortes, B. Gueye, A. Probst, and J.M. Palacios (1989)Dopamine receptors in human brain: Autoradiographic distribution ofD2 sites. Neuroscience 28:275–290.
Charuchinda, C., P. Supavilai, M. Karobath, and J.M. Palacios (1987)Dopamine D2 receptors in the rat brain: Autoradiographic visualizationusing a high-affinity selective agonist ligand. J. Neurosci. 7:1352–1360.
Chazot, P.L., A.J. Doherty, and P.G. Strange (1993) Antisera specific for D2dopamine receptors. Biochem. J. 289:789–794.
Chevalier, G., S. Vacher, J.M. Deniau, and M. Desban (1985) Disinhibitionas a basic process in the expression of striatal functions: I. Thestriato-nigral influence on tecto-spinal/tecto-diencephalic neurons. BrainRes. 333:215–226.
Civelli, O., J.R. Bunzow, and D.K. Grandy (1993) Molecular diversity of thedopamine receptors. Annu. Rev. Pharmacol. Toxicol. 32:281–307.
Conde, F., J.S. Lund, D.M. Jacobowitz, K.G. Baimbridge, and D.A. Lewis(1994) Local circuit neurons immunoreactive for calretinin, calbindinD-28k, or parvalbumin in monkey prefrontal cortex: Distribution andmorphology. J. Comp. Neurol. 341:95–116.
Cox, R.F. and B.L. Waszczak (1991) Autoradiographic of dopamine D2receptors using [3H]YM-09151–2. Eur. J. Pharmacol. 199:103–106.
Cragg, S.J. and S.A. Greenfield (1997) Differential autoreceptor control ofsomatodendritic and axon terminal dopamine release in substantianigra, ventral tegmental area, and striatum. J. Neurosci. 17:5738–5746.
Davis, K.L., R.S. Kahn, G. Ko, and M. Davidson (1991) Dopamine inschizophrenia: A review and reconceptualization. Am. J. Psychiatry148:1474–1486.
Defagot, M.C. and M.C. Antonelli (1997) Autoradiographic localization ofthe putative D4 dopamine receptor in rat brain. Neurochem. Res.22:401–407.
Defagot, M.C., E.L. Malchiodi, M.J. Villar, and M.C. Antonelli (1997)Distribution of D4 dopamine receptor in rat brain with sequence-specific antibodies. Mol. Brain Res. 45:1–12.
De Keyser, J., A. Claeys, J.P. De Backer, F. Roels, G. Ebinger, and G.Vauquelin (1988) Autoradiographic localization of D1 and D2 dopaminereceptors in the human brain. Neurosci. Lett. 91:142–147.
Diaz, J., D. Levesque, C.H. Lammers, N. Griffon, M.P. Martres, J.C.Schwartz, and P. Sokoloff (1995) Phenotypical characterization ofneurons expressing the dopamine D3 receptor in the rat brain. Neurosci-ence 65:731–745.
Elazar, Z., H. Kanety, C. David, and S. Fuchs (1988) Purification andcharacterization of the D2-dopamine receptor from bovine striatum.Biochem. Biophys. Res. Commun. 156:602–609.
Farooqui, S.M., J.W. Brock, A. Hamdi, and C. Prasad (1991) Antibodiesagainst synthetic peptides predicted from the nucleotide sequence of D2receptor recognize native dopamine receptor protein in rat striatum. J.Neurochem. 57:1363–1369.
Frey, U., H. Schroeder, and H. Matthies (1990) Dopaminergic antagonistsprevent long-term maintenance of posttetanic LTP in the CA1 region ofrat hippocampal slices. Brain Res. 522:69–75.
Gaspar, P., B. Bloch, and C. Le Moine (1995) D1 and D2 receptor geneexpression in the rat frontal cortex. Cellular localization in differentclasses of efferent neurons. Eur. J. Neurosci. 7:1050–1063.
Gerfen, C.R. (1992) The neostriatal mosaic: Multiple levels of compartmen-tal organization. Trends Neurosci. 15:133–139.
Gingrich, J.A. and M.G. Caron (1993) Recent advances in the molecularbiology of dopamine receptors. Annu. Rev. Neurosci. 16:299–321.
Giros, B., P. Sokoloff, M.P. Martres, J.F. Riou, L.J. Emorine, and J.C.Schwartz (1989) Alternative splicing directs the expression of two D2dopamine receptor isoforms. Nature 342:923–926.
Goldman-Rakic, P.S. (1998) The cortical dopamine system: Role in memoryand cognition. Adv. Pharmacol. 42:707–711.
Goldman-Rakic, P.S., C. Leranth, S.M. Williams, N. Mons, and M. Geffard(1989) Dopamine synaptic complex with pyramidal neurons in primatecerebral cortex. Proc. Natl. Acad. Sci. USA 86:9015–9019.
Gorrisen, H. and P. Laduron (1979) Solubilisation of high-affinity dopaminereceptors. Nature 279:72–74
Graybiel, A.M., T. Aosaki, T. Flaherty, and M. Kimura (1994) The basalganglia and adaptive motor control. Science 265:1826–1831.
Grace, A.A. (1993) Cortical regulation of subcortical dopamine systems andits possible relevance to schizophrenia. J. Neural Transm. 91:111–134.
Grigoriadis, D.E., H.B. Niznik, K.R. Jarvie, and P. Seeman (1988) Glycopro-tein nature of D2 dopamine receptors. FEBS Lett. 227:220–224.
Gutierrez, A., Z.U. Khan, and A.L. de Blas (1994) Immunocytochemicallocalization of g2short and g2long subunits of the GABAA receptor in therat brain. J. Neurosci. 14:7168–7179.
Hall, H., L. Farde, C. Halldin, Y.L. Hurd, S. Pauli, and G. Sedvall (1996a)Autoradiographic localization of extrastriatal D2-dopamine receptorsin the human brain using [125I]epidepride. Synapse 23:115–123.
Hall, H., C. Halldin, D. Dijkstra, H. Wikstrom, L.D. Wise, T.A. Pugsley, P.Sokoloff, S. Pauli, L. Farde, and G. Sedvall (1996b) Autoradiographiclocalisation of D3-dopamine receptors in the human brain using theselective D3-dopamine receptor agonist (1)-[3H]PD128907. Psychophar-macology 128:240–247.
Harris, G.C. and G. Auston-Jones (1994) Involvement of D2 dopamine-receptors in the nucleus-accumbens in the opiate withdrawal syn-drome. Nature 371:155–157.
Herroelen, L., J.P. De Backer, N. Wilczak, A. Flamez, G. Vauquelin, and J.De Keyser (1994) Autoradiographic distribution of D3-type dopaminereceptors in human brain using [3H]-hydroxy-N,N-di-n-propyl-2-aminotetralin. Brain Res. 648:222–228.
Hersch, S.M., B.J. Ciliax, C. Gutekunst, H.D. Rees, C.J. Heilman, K.K.L.Yung, J.P. Bolam, E. Ince, H. Yi, and A.L. Levey (1995) Electronmicroscopic analysis of D1 and D2 dopamine receptor proteins in thedorsal striatum and their synaptic relationships with motor corticostria-tal afferents. J. Neurosci. 15:5222–5237.
Horn, A.S. (1990) Dopamine uptake: A review of progress in the last decade.Prog. Neurobiol. 34:387–400.
Huntley, G.W., J.H. Morrison, A. Prilhozhan, and S.C. Sealfon (1992)Localization of multiple dopamine receptor subtype mRNAs in humanand monkey motor cortex and striatum. Mol. Brain Res. 15:181–188.
Jackson, D.M. and A. Westlind-Danielsson (1994) Dopamine receptors:Molecular biology, biochemistry and behavioural aspects. Pharmacol.Ther. 64:291–369.
Jarvie, K.R., H.B. Niznik, and P. Seeman (1988) Dopamine D2 receptorbinding subunit of Mr5140,000 and 94,000 in brain: Deglycosylationyields a common unit of Mr544,000. Mol. Pharmacol. 34:91–97.
Joyce, J.N. and J.H. Meador-Woodruff (1997) Linking the family of D2receptors to neuronal circuits in human brain: Insight into schizophre-nia. Neuropsychopharmacology 16:375–384.
Kelly, M.A., M. Rubinstein, S.L. Asa, G. Zhang, C. Saez, J.R. Bunzow, R.G.Allen, R. Hnasko, N. Ben-Jonathan, D.K. Grandy, and M.J. Low (1997)Pituitary lactotroph hyperplasia and chronic hyperprolactinemia indopamine D2 receptor-deficient mice. Neuron 19:103–113.
Khan, Z.U., A. Gutierrez A, and A.L. de Blas (1994) Short and long form g2
subunits of the GABAA/benzodiazepine receptors. J. Neurochem. 63:1466–1476.
Koeltzow, T.F., M. Xu, D.C. Cooper, X. Hu, S. Tonegawa, M.E. Wolf, and M.J.White (1998) Alterations in dopamine release but not dopamine autore-ceptor function in dopamine D-3 receptor mutant mice. J. Neurosci.18:2231–2238.
370 Z.U. KHAN ET AL.

Koob, G.F. (1992) Drugs of abuse: Anatomy, pharmacology and function ofreward pathways. Trends Pharmacol. Sci. 13:177–184.
Kuhar, M.J., M.C. Ritz, and J.W. Boja (1991) The dopamine hypothesis ofthe reinforcing properties of cocaine. Trends Neurosci. 14:299–302.
Lahti, R.A., R.C. Roberts, and C.A. Tamminga (1995) D2-family receptordistribution in human postmortem tissue: An autoradiographic study.Neuroreport 6:2505–2512.
Landwehrmeyer, B., G. Mengod, and J.M. Palacios (1993a) Dopamine D3receptor mRNA and binding sites in human brain. Mol. Brain Res.18:187–192.
Landwehrmeyer, B., G. Mengod, and J.M. Palacios (1993b) Differentialvisualization of dopamine D2 and D3 receptor sites in rat brain: Acomparative study using in situ hybridization histochemistry andligand binding autoradiography. Eur. J. Neurosci. 5:145–153.
Lee, S.P., G.Y.K. Ng, P. Zawarynski, P. Seeman, B.F. O’Dowd, and S.R.George (1997) Regulation of the dopamine D2 receptor dimer. Neurosci.Abst. 23:1780.
Levant, B., D.A. Grigoriadis, and E.B. De Souza (1993) [3H]Quinpirolebinding to putative D2 and D3 dopamine receptors in rat brain andpituitary gland: A quantitative autoradiographic study. J. Pharmacol.Exp. Ther. 264:991–1001.
Levey, A.I., S.M. Hersch, D.B. Rye, R.K. Sunahara, H.B. Niznik, C.A. Kitt,D.L. Price, R. Maggio, M.R. Brann, and B.J. Ciliax (1993) Localizationof D1 and D2 dopamine receptors in brain with subtype-specificantibodies. Proc. Natl. Acad. Sci. USA 90:8861–8865.
Lidow, M.S., P.S. Goldman-Rakic, P. Rakic, and R.B. Innis (1989) DopamineD2 receptors in the cerebral cortex: Distribution and pharmacologicalcharacterization with [3H]raclopride. Proc. Natl. Acad. Sci. USA 86:6412–6416.
Liu, K., C. Bergson, R. Levenson, and C. Schmauss (1994) On the origin ofmRNA encoding the truncated dopamine D3-type receptor D3nf anddetection of D3nf-like immunoreactivity in human brain. J. Biol. Chem.269:29220–29226.
Lowry, O.H., N.J. Rosebrough, A.L. Farr, and R.J. Randall (1951) Proteinmeasurement with the Folin phenol reagent. J. Biol. Chem. 193:265–275.
Mansour, A., J.H. Meador-Woodruff, J.R. Bunzow, O. Civelli, H. Akil, andS.J. Watson (1990) Localization of dopamine D2 receptor mRNA and D1and D2 receptor binding in the rat brain and pituitary: An in situhybridization-receptor autoradiographic analysis. J. Neurosci. 10:2587–2600.
McVittie, L.J., M.A. Ariano, and D.R. Sibley (1991) Characterization ofanti-peptide antibodies for the localization of D2 dopamine receptors inrat striatum. Proc. Natl. Acad. Sci. USA 88:1441–1445.
Meador-Woodruff, J.H., A. Mansour, J.R. Bunzow, H.H.M. Van Tol, S.J.Watson, and O. Civelli (1989) Distribution of D2 dopamine receptormRNA in rat brain. Proc. Natl. Acad. Sci. USA 86:7625–7628.
Meador-Woodruff, J.H., A. Mansour, D.J. Healy, R. Kuehn, Q.Y. Zhou, J.R.Bunzow, H. Akil, O. Civelli, and S.J. Watson (1991) Comparison of thedistributions of D1 and D2 dopamine receptor mRNAs in rat brain.Neuropsychopharmacology 5:231–242.
Meador-Woodruff, J.H., D.K. Grandy, H.H.M. Van Tol, S.P. Damask, K.Y.Little, O. Civelli, and S.J. Watson (1994) Dopamine receptor geneexpression in the human medial temporal lobe. Neuropsychopharmacol-ogy 10:239–248.
Mengod, G., M.I. Martinez-Mir, M.T. Vilaro, and J.M. Palacios (1989)Localization of the mRNA for the dopamine D2 receptor in the rat brainby in situ hybridization histochemistry. Proc. Natl. Acad. Sci. USA86:8560–8564.
Mengod, G., M.T. Vilaro, B. Landwehrmeyer, M.I. Martinez-Mir, H.B.Niznik, R.K. Sunahara, P. Seeman, B.F. O’Dowd, A. Probst, and J.M.Palacios (1992). Visualization of dopamine D1, D2 and D3 receptormRNAs in human and rat brain. Neurochem. Int. 20:33–43.
Mercuri, N.B., A. Saiardi, A. Bonci, R. Picetti, P. Calbresi, G. Bernardi, and E.Borrelli (1997) Loss of autoreceptor function in dopaminergic neurons fromdopamine D2 receptor deficient mice. Neuroscience 79:323–327.
Monsma, F.J., L.D. McVittie, C.R. Gerfen, L.C. Mahan, and D.R. Sibley(1989) Multiple D2 dopamine receptors produced by alternative RNAsplicing. Nature 342:926–929.
Mrzljak, L., C. Bergson, M. Pappy, R. Huff, R. Levenson, and P.S. Goldman-Rakic (1996) Localization of dopamine D4 receptors in GABAergicneurons of the primate brain. Nature 381:245248.
Murray, A.M., H.L. Ryoo, and J.N. Joyce (1992) Visualization of dopamineD3-like receptors in human brain with [125I]epidepride. Eur. J. Pharma-col. 227:443–445.
Neve, K.A. and R.L. Neve (1997) Molecular biology of dopamine receptors.In K.A. Neve, and R.L. Neve (eds): The Dopamine Receptors. Totowa,NJ: Humana Press, pp. 27–76.
Nobrega, J.N. and P. Seeman (1994) Dopamine D2 receptors mapped in ratbrain with [3H](1)PHNO. Synapse 17:167–172.
O’Dowd, B.F., P. Seeman, and S.R. George (1994) Dopamine receptors. InS.J. Peroutka (ed): Handbook of Receptors and Channels. G Protein-Coupled Receptors. New York: CRC Press, pp. 95–123.
Panagopoulos, N.T., G.C. Papadopoulos, and N.A. Matsokis (1991) Dopamin-ergic innervation and binding in the rat cerebellum. Neurosci. Lett.130:208–212.
Rakic, P., P.S. Goldman-Rakic, and D. Gallager (1988) Quantitative autora-diography of major neurotransmitter receptors in the monkey striateand extrastriate cortex. J. Neurosci. 8:3670–3690.
Ricci, A., J.A. Vega, C.L. Mammola, and F. Amenta (1995) Localization ofdopamine D3 receptor in the rat cerebellar cortex: A light microscopeautoradiographic study. Neurosci. Lett. 190:163–166.
Robertson, H.A. (1992) Synergistic interactions of D1 and D2-selectivedopamine agonists in animal models for Parkinson’s disease: Sites ofaction and implications for the pathogenesis of dyskinesias. Can. J.Neurol. Sci. 19:147–152.
Schmauss, C., V. Haroutunian, K.L. Davis, and M. Davidson (1993)Selective loss of dopamine D3-type receptor mRNA expression inparietal and motor cortices of patients with chronic schizophrenia. Proc.Natl. Acad. Sci. USA 90:8942–8946.
Sen, A.K. and T. Lee (1988) Receptors and Ligands in Neurology. Cam-bridge: Cambridge University Press.
Sesack, S.R., C. Aoki, and V.M. Pickel (1994) Ultrastructural localization ofD2 receptor-like immunoreactivity in midbrain dopamine neurons andtheir striatal targets. J. Neurosci. 14:88–106.
Sesack, S.R., S.L. Snyder, and A. Lewis (1995) Axon terminals immunola-beled for dopamine or tyrosine hydroxylase synapse on GABA-immunoreactive dendrites in rat and monkey cortex. J. Comp. Neurol.363:264–280.
Sibley, D.R. and F.J. Monsma (1992) Molecular biology of dopaminereceptors. TIPS 13:61–69.
Sokoloff, P., B. Giros, M.P. Martres, M.L. Bouthenet, and J.C. Schwartz(1990) Molecular cloning and characterization of a novel dopaminereceptor (D3) as a target for neuroleptics. Nature 347:146–151.
Squire, L.R. (1987) Memory and Brain. New York: Oxford University Press.Steriade, M., D.A. McCormick, and T.J. Sejnowski (1993) Thalamocortical
oscillations in the sleeping and arousal brain. Science 262:679–685.Tarazi, F.I., S.K. Yeghiayan, R.J. Baldessarini, N.S. Kula, and J.L. Neum-
eyer (1997) Long term effects of S(1)N-n-propylnorapomorphine com-pared with typical and atypical antipsychotics: Differential increases ofcerebrocortical D2-like and striatolimbic D4-like dopamine receptors.Neuropsychopharmacology 17:186–196.
Van Tol, H.H.M., J.R. Bunzow, H.G. Guan, R.K. Sunahara, P. Seeman, H.Niznik, and O. Civelli (1991) Cloning of the gene for a human dopamineD4 receptor with high affinity for the antipsychotic clozapine. Nature350:610–614.
Vargha-Khadem, F., D.G. Godian, K.E. Watkins, A. Konnelly, W. VanPaesschen, and M. Mishkin (1997) Differential effects of early hippocam-pal pathology in episodic and semantic memory. Science 277:376–380.
Weiner, D.M. and M.R. Brann (1989) The distribution of a dopamine D2receptor mRNA in rat brain. FEBS Lett. 253:207–213.
Weiner, D.M., A.I. Levey, R.K. Sunahara, H.B. Niznik, B.F. O’Dowd, P.Seeman, and M.R. Brann (1991) D1 and D2 dopamine receptor mRNAin rat brain. Proc. Natl. Acad. Sci. USA 88:1859–1863.
Williamson, R.M., S. Worrall, P.L. Chazot, and P.G. Strange (1988) Purifica-tion of brain D2 dopamine receptor. EMBO J. 7:4129–4133.
Yokoyama, C., H. Okamura, T. Nakajima, J. Taguchi, and Y. Ibata (1994)Autoradiographic distribution of [3H]YM-09151–2, a high-affinity andselective antagonist ligand for the dopamine D2 receptor group, in therat brain and spinal cord. J. Comp. Neurol. 344:121–136.
Yokoyama, C., H. Okamura, and Y. Ibata (1995) Dopamine D2-like recep-tors labeled by [3H]YM-09151–3 in the rat hippocampus: Characteriza-tion and autoradiographic distribution. Brain Res. 681:153–159.
Yung, K.K., J.P. Bolam, A.D. Smith, S.M. Hersch, B.J. Ciliax, and A.I. Levey(1995) Immunocytochemical localization of D1 and D2 dopamine recep-tors in the basal ganglia of the rat: Light and electron microscopy.Neuroscience 65:709–730.
Zawarynski, P., G.Y.K. Ng, T. Tallerico, S.P. Lee, B.F. O’Dowd, S.R. George,and P. Seeman (1997) Separation of dopamine D2 receptor monomersand dimers. Neurosci. Abstr. 23:1780.
ANTIBODIES TO DOPAMINE D2-LIKE RECEPTORS 371