description of a new genus and species of rodent (murinae
TRANSCRIPT

PUBLISHED BY THE AMERICAN MUSEUM OF NATURAL HISTORY
CENTRAL PARK WEST AT 79TH STREET, NEW YORK, NY 10024
Number 3517, 41 pp., 14 figures, 4 tables May 17, 2006
Description of a New Genus and Species of Rodent(Murinae, Muridae, Rodentia) from the Tower Karst
Region of Northeastern Vietnam
GUY G. MUSSER,1 DARRIN P. LUNDE,2 AND NGUYEN TRUONG SON3
ABSTRACT
Tonkinomys daovantieni, a new genus and species of murid rodent in the Dacnomys Division, isdescribed. It is represented by 14 adults collected from talus habitats in the forested tower karstlandscape of the Huu Lien Nature Reserve of northeastern Vietnam. The combination ofsemispinous, dense, grayish black fur covering upperparts; a dark gray venter; gray ears; a thick,bicolored tail considerably shorter than length of head and body; and large, extremely bulbousfootpads is unlike any other species of Indomalayan murid. Body size and build of the new rat,along with some cranial features, are similar to the Thai Leopoldamys neilli, but other cranial traitscoupled with molar occlusal patterns resemble morphology in species of the IndomalayanNiviventer, Chiromyscus, and Saxatilomys. The new species is petricolous, includes insects in itsdiet, and was found only in talus composed of large limestone blocks. Its distribution in the reserveis likely patchy. Whether this limestone rat is restricted to the extensive karst regions ofnortheastern Vietnam or also occurs in southern China and elsewhere in the northern karstlandscapes of Indochina, and Vietnam in particular, will be known only by conducting surveys inlimestone regions outside of northeastern Vietnam.
INTRODUCTION
During Paleozoic times, a succession ofwaterways covered what is now northernVietnam. Lithification, followed by tectonic
deformation and uplift, and spurred by sub-sequent erosional processes, transformedthose ancient tidal flows and marine land-scapes into a spectacular terrestrial topogra-phy of tower karst riddled by fissures and
Copyright E American Museum of Natural History 2006 ISSN 0003-0082
1 Division of Vertebrate Zoology (Mammalogy), American Museum of Natural History. Current address: 1714 HenleyLane, Charleston, SC 29412 ([email protected]).
2 Division of Vertebrate Zoology (Mammalogy), American Museum of Natural History ([email protected]).3 Department of Zoology, Institute of Ecology and Biological Resources, 18 Hoang Quoc Viet Road, Cau Giay, Hanoi,
Vietnam ([email protected]).

honeycombed with caverns. Beautiful cycadrosettes secured high on sheer limestone cliffs,and a forest cover in which many of thecomponents are adapted to karstic habitats,are the living botanical link to those ancientseas. A dark gray rat, its fur colorationindistinguishable from the limestone, its lairconcealed deep within fissured cliffs andlimestone talus, is the zoological connection.
We discovered this limestone rat duringa survey (April, 2004) of small mammals ina forested tower karst region that forms muchof Lang Son Province in northeastern Vietnam(Głazek, 1966; Tuyet, 1998) (fig. 1). Our intentwas to sample, using a variety of traps andother techniques, as many native species aspossible. Our past field experience inIndochina had taught us that setting trapson the ground in regular lines would quicklysample the generalists of the more commonterrestrial habitats but not those specializedfor life on substrates above the forest floor orin unusual petricolic environments. With this
in mind we made special efforts to set a portionof our traps on tree limbs, woody vineslooping through the understory, and on highlimestone ledges. At one place, chunks oflimestone had long ago fallen from two highforested towers, filling the space between themwith huge blocks, their surfaces now partiallycovered with tropical vegetation, and riddledbeneath with tunnels and fissures detectable atthe surface only by small scattered openings(fig. 2). We set traps on top of this block pile,and also tied traps to lengths of rope andcarefully lowered them through the openingsto the bottom, to depths of at least 8 m.Unsure if we would catch anything differentfrom the common rats already encountered innearby different habitats—Niviventer fulves-cens, N. langbianis, and Rattus andamanen-sis—we were very pleased, and somewhatsurprised, to capture a dark gray rat on topof the rubble and two from deep within thelabyrinth. Additional individuals were cap-tured during the days that followed.
Fig. 1. Tower karst landscape where we captured our series of Tonkinomys daovantieni within the HuuLien Nature Reserve. Tropical moist deciduous forest covers the steep slopes, but the valley bottom has beencleared for agriculture. Photographed by D. P. Lunde, April 2004.
2 AMERICAN MUSEUM NOVITATES NO. 3517

We knew nothing like this animal had everbeen caught in Vietnam or elsewhere inSoutheast Asia. Except for their fur colorationand short tails, the rats resembled species ofLeopoldamys, but no member of that genushas dark gray fur and a tail shorter than headand body. Those two attributes also pointedto Berylmys, but none of them have dark grayunderparts. We knew our sample of thelimestone rat represented an undescribedspecies, but only after study in the museumdid we confirm our suspicion that it could notbe placed in any named genus.
Here we describe the limestone rat as a newgenus and species, and summarize our limitedobservations about its natural history. Wefocus on describing the animal and comparingit to murines with which it is morphologicallymost similar, members of a cluster containinggenera that Musser and Carleton (2005) referto as the Dacnomys Division: extant species ofthe Indomalayan Chiromyscus, Dacnomys,Leopoldamys, and Niviventer; the Sri Lankan
Srilankamys; and the recently describedLaotian Saxatilomys (Musser et al., 2005).We also contrast Tonkinomys with members ofthe Dacnomys Division represented by fossilsrecovered from Indochinese Plio-Pleistocenesediments: extinct species of Leopoldamysand Niviventer, and the extinct generaWushanomys and Qianomys. Of these, thenew rat requires close and careful compari-sons only with Leopoldamys, Niviventer,Chiromyscus, Saxatilomys and the Plio-Pleistocene Wushanomys and Qianomys.
Descriptions of new taxa, and in particularthose occurring in gradually diminishingtropical forest refuges, should be publishedas soon as possible. Phylogenetic inquiriestake time—incorporating the new rat intophylogenetic analyses employing samples ofall native Indochinese murines will be thesubject of a future report.
Our discovery joins those of several othersmall-bodied mammals living in Indochinathat have been discovered and described, with
Fig. 2. Habitat where we trapped 13 of the 14 examples of Tonkinomys daovantieni showing large blocksof limestone overgrown with creeping vines (left) and a portion of the steep slopes of an adjacent karst tower(right). Our trail to the site is visible between these two structures. Photographed by D. P. Lunde, April 2004.
2006 MUSSER ET AL.: NEW RODENT FROM VIETNAM 3

little fanfare, during the past five years: twoshrews, Chodsigoa caovansunga and Crocidurakegoensis (Lunde et al., 2003, 2004); threebats, Hipposideros scutinares (Robinson et al.,2003), Rhinolophus stheno microglobosus(Csorba and Jenkins, 1998) and Myotisannamiticus (Kruskop and Tsytsulina, 2001);and a rabbit, Nesolagus timminsi (Averianov etal., 2000; Can et al., 2001). These can be addedto an assemblage of large mammals occurringin Vietnam, and some in nearby Laos, namedand described as new within the last decade orso, but with attendant worldwide publicity:two primates, Trachypithecus ebenus andPygathrix cinerea (summarized in Groves,2001); three muntjacs, Muntiacus truongsonen-sis, M. vuquangensis, and M. puhoatensis (seereviews in Amato et al., 2000; Groves andSchaller, 2000; Grubb, 2005); and the saola,Pseudoryx nghetinhensis (see review and refer-ences in Groves and Schaller, 2000).4 All area testimony to the significant results derivedfrom past biodiversity surveys, discoveriesthat provide compelling stimulus for zoolo-gists to continue detecting the real diversity ofsmall mammals in Vietnam.
MATERIALS AND METHODS
All 14 examples of the new rat were fixed in10% formalin in the field and subsequentlytransferred to 70% ethanol for storage. Liversamples were collected prior to fixation andplaced in lysis buffer in the field, and laterstored in liquid nitrogen at the AmericanMuseum of Natural History (AMNH). Skullswere subsequently removed from 10 of 13specimens (one of the 14 preserved is repre-sented by only an ear with a bit of fur-coveredskin attached, the only remnant left in the trapby a hungry predator) and cleaned by a der-mestid colony, American Museum. The ele-ments of each specimen are identified bya catalog number in the Department ofMammalogy, AMNH. Each liver sample wastransferred to the American Museum CryoCollection (AMCC), where it is registered
under a second number in addition to itsdepartmental catalog number. Some fluid-pre-served specimens and accompanying skulls willbe retained at AMNH; others will be returnedto the Department of Zoology at the Instituteof Ecology and Biological Resources in Hanoi.
Comparative material we consulted, pri-marily samples of Leopoldamys, Niviventer,Chiromyscus, and Saxatilomys, are in collec-tions at AMNH; the Field Museum, Chicago;and the National Museum of Natural History,Washington, DC.
Values (in millimeters, mm) for total length,length of tail (LT), length of hind foot,including claw (LHF), and length of ear fromintertragal notch to crown (LE) are those weobtained in the field and recorded in our fieldjournals. Values for length of head and body(LHB) were determined by subtracting lengthof tail from total length. Weights were obtainedin the field with a Pesola spring scale (300-gramcapacity). The distal portion of the tail is white(unpigmented) in all specimens of the limestonerat. We measured the length of this white tip onthe dorsal surface of each tail and calculated itas a percentage of total tail length. Values forcranial and dental measurements were obtainedby Lunde using digital calipers accurate to thenearest 0.01 mm; however, values were round-ed to the nearest 0.1 mm. The following cranialand dental dimensions (listed in the sequencethey appear in tables) were measured; theirlimits are illustrated in figure 3 and defined inMusser and Newcomb (1983).
ONL occipitonasal length (5 greatestlength of skull)
ZB zygomatic breadthIB interorbital breadthLR length of rostrumBR breadth of rostrumBBC breadth of braincaseHBC height of braincaseBZP breadth of zygomatic plateLD length of diastemaLIF length of incisive foraminaBIF breadth of incisive foraminaLBP length of bony palate (palatal bridge)BBP breadth across bony palate at first
molarsPPL postpalatal lengthBMF breadth of mesopterygoid fossaLB length of bullaCLM1-3 crown length of maxillary molar rowBM1 breadth of first upper molar
4 Viverra tainguensis, another supposed newly discoveredspecies living in Vietnam, was described by Sokolov et al.(1997). Its specific integrity, however, was questioned byWalston and Veron (2001), and the name is currentlyincluded in the synonymy of the large Indian civet, Viverrazibetha (Wozencraft, 2005).
4 AMERICAN MUSEUM NOVITATES NO. 3517

Fig. 3. Views of cranium and molars of an adult Bunomys chrysocomus showing limits of cranial anddental measurements. See text for abbreviations and additional information.
2006 MUSSER ET AL.: NEW RODENT FROM VIETNAM 5

Old adults, adults, and young adults (asdefined by Musser and Heaney, 1992: 5) werelumped as ‘‘adults’’ and measurements derivedfrom them were used to calculate standarddescriptive statistics (mean, standard devia-tion, and observed range). Small sample sizedictated that we combine sexes in the analyses.Anatomical terminology follows Brown (1971)and Brown and Yalden (1973) for externalfeatures of the head and limbs; Bugge (1970)for the cephalic arteries; Wahlert (1985) forthe cranial foramina; and Carleton (1980),Carleton and Musser (1984), Musser andDurden (2002), Musser and Heaney (1992),Musser et al. (1998), and Voss (1988) forcranial morphology. Names of cusps andcusplets of upper and lower molars are notedin figure 13; sources of this terminology areexplained by Musser and Newcomb (1983:332).
SYSTEMATIC DESCRIPTION OF THENEW GENUS AND SPECIES
Tonkinomys, new genus
TYPE SPECIES: Tonkinomys daovantieni, thenew species described below.
DIAGNOSIS: A genus of Muridae in theDacnomys Division (Musser and Carleton,2005) of the subfamily Murinae (as delimitedby Carleton and Musser, 1984) that is setapart from all other described murid genera bythe following combination of morphologicaltraits: (1) fur covering head and body semi-spinous and grayish black, spattered witha white blaze on forehead in most individuals,underparts dark gray with white patch on thechest, ears gray, rhinarium and lips and chinunpigmented; (2) mystacial and superciliaryvibrissae very long; (3) dorsal surfaces of frontfeet white, hind feet white with brown hairs onmetatarsal region; (4) tail much shorter thanlength of head and body, thick and round incross-section and well-haired, the proximalone-half to three-fourths of its dorsal andlateral surfaces dark brown, the distal one-fourth to one-half white (unpigmented);(5) palmar and plantar pads large, swollen,and set close together; (6) four pairs of teats;(7) robust skull with moderately long and widerostrum, prominent postorbital and temporalridges, sturdy zygomatic arches, and deep
occiput; (8) squamosal root of each zygomaticarch situated high on side of cranium where itsposterior ridge-like portion runs horizontallyjust below posteroventral margin of theparietal and projects ventrad to form dorso-lateral side of cranium; (9) lateral cranial wallintact anterior to occiput, without largesubsquamosal foramen; (10) narrow alisphe-noid struts in most specimens; (11) wide andmoderately long incisive foramina, their pos-terior borders located just before anteriormargins of molar rows, even with them, orpenetrating slightly between; (12) posteriorlydivergent maxillary molar rows; (13) wide andlong bony palate projecting beyond molarsto form a wide platform; (14) moderatelyspacious sphenopalatine vacuities; (15) wideand shallowly excavated pterygoid fossae;(16) small ectotympanic bulla relative to skullsize, incompletely covering periotic so thatdorsal and slanting posterodorsal wall ofcarotid canal formed by periotic and notectotympanic; (17) large foramen for stapedialartery, no sphenofrontal foramen or squamo-soalisphenoid groove, indicating the widespreadmurine cephalic arterial pattern; (18) coronoidprocess of dentary small, condyloid and angularprocesses joined by shallow concave posteriormargin of dentary, and alveolar incisorcapsule only slightly evident on lateral surfaceof dentary; (19) upper incisors opisthodontrelative to rostrum; (20) first upper molar withfour large roots (anterior, two lingual, andposterior), second and third molars each withthree roots (anterior, lingual, and posterior);(21) each lower molar with two roots;(22) molars brachydont, with cusp rows form-ing uncomplicated occlusal patterns resemblingthose in species of Niviventer, Chiromyscus, andSaxatilomys.
ETYMOLOGY: From Tonkin, used duringthe French colonial period (1883–1954) forthe protectorate occupying the northern thirdof Vietnam, and the Greek mys, referring tomouse, or rat in this case. Although mostoften negatively associated with Vietnam’sdark French administration, the nameTonkin has an illustrious origin. It is derivedfrom the Vietnamese Don Kinh, which means‘‘eastern capital’’, and was the name given towhat is now Hanoi by Emperor Le Loi who,in 1427, along with his band of Vietnamese
6 AMERICAN MUSEUM NOVITATES NO. 3517
rebels, expelled from Vietnam the repressiveMing occupiers. By the early 1800s, the capitalwas moved to Hue, and the Emperor MinhMang renamed the ‘‘eastern capital’’ Hanoi,meaning ‘‘this side of the river’’.
Tonkinomys daovantieni, new species
HOLOTYPE: AMNH 275618, an adult fe-male collected by the authors (DPL 1706) on 17April, 2004, in the vicinity of Lan Ðat Village(21u409520W/106u209280E), 150 m, Hu,u LienNature Reserve, Hu,u Lien Commune, Hu,uLung District, Lang So,n Province, Vietnam.The specimen was fixed in 10% formalin forseveral days before being transferred to 70%ethanol for storage. A liver sample wascollected prior to fixation and placed in lysisbuffer before its ultimate transfer to the
American Museum Cryo Collection (AMCC125060). The skull (figs. 10, 11) was laterextracted and cleaned by dermestids in a colonyat the American Museum.
REFERRED MATERIAL: In addition to theholotype, 13 additional adults of T. daovantieniwere collected from the vicinity of Lan ÐatVillage during April, 2004, on the days in-dicated (AMCC catalog numbers tied to liversamples are in brackets). April 13: AMNH275575 [125016], male; 275576 [125017], male;275577 [125018], male. April 14: 275586[125027], female; 275593 [125034], male. April16: 275602 [125044], male. April 19: 275627[125069], male; 275644 [125086], male. April 23:275688 [125130], male. April 24: 275692[125134], male; 275693 [125135], ear withattached bit of furry skin only. April 25:275711 [125153], male; 275712 [125154], female.
TABLE 1
Summary Statistics for Cranial and Dental Measurements (mm) and Weight (m) of Tonkinomys daovantienifrom Vietnam
(Mean, plus or minus one SD, observed range in parentheses, and number of specimens in each sampleare listed for each measurement. Sample contains both sexes and a range in age from young to old
adults. Individuals measured and weighed are identified in the footnote.)
Holotypea Sample (with holotype)b
Length of head and body 213 204.6 6 10.34 (184–217) 13
Length of tail 177 169 6 7.0 (156–183) 13
Length of hind foot 40 39.0 6 1.1 (37–41) 13
Length of ear 30 30.0 6 0.8 (29–31) 14
Weight 160 171 6 22.8 (140–205) 13
Occipitonasal length 51.2 50.0 6 1.6 (47.1–52.4) 7
Zygomatic breadth 21.6 21.3 6 0.2 (20.9–21.6) 7
Interorbital breadth 6.7 6.9 6 0.2 (6.6–7.3) 7
Length of rostrum 17.7 17.1 6 0.6 (15.9–17.7) 7
Breadth of rostrum 7.7 7.8 6 0.4 (7.1–8.5) 7
Breadth of braincase 17.7 17.7 6 0.2 (17.4–18.1) 7
Height of braincase 11.7 11.8 6 0.2 (11.4–12.0) 7
Breadth of zygomatic plate 4.9 4.8 6 0.1 (4.7–5.0) 7
Length of diastema 13.3 13.6 6 0.5 (12.9–14.6) 7
Length of incisive foramina 9.1 9.5 6 0.5 (8.9–10.2) 7
Breadth of incisive foramina 3.4 3.5 6 0.2 (3.4–3.9) 7
Length of bony palate 10.7 10.5 6 0.3 (10.0–10.7) 7
Breadth across bony palate at M1 9.6 9.6 6 0.1 (9.5–9.8) 7
Postpalatal length 16.2 15.7 6 0.6 (14.6–16.4) 7
Breadth of mesopterygoid fossa 3.9 3.8 6 0.2 (3.5–3.9) 7
Length of bulla 6.2 6.3 6 0.1 (6.2–6.5) 7
Crown length of M1-3 8.3 8.3 6 0.2 (8.0–8.8) 7
Breadth of first upper molar 2.1 2.1 6 0.1 (2.0–2.2) 7
a AMNH 275618.b AMNH 275575-275577, 275586, 275593, 275602, 275618, 275627, 275644, 275688, 275692, 275711, and 275712.
2006 MUSSER ET AL.: NEW RODENT FROM VIETNAM 7

GEOGRAPHIC DISTRIBUTION: Tonkinomysdaovantieni is currently known only from thetype locality, but the species probably occursin suitable forested habitats throughout thekarst landscapes of northern Vietnam (identi-fied as the ‘‘Vietbac Karst Zone’’ by Tuyet,1998: 187; or the Caobang and Bacson regionsby Głazek, 1966), and possibly in southernChina (see the distributions of karst outcropsmapped in Tuyet, 1998, and Zhang, 1989).
DIAGNOSIS: Because daovantieni is the onlyknown species of Tonkinomys, generic andspecific diagnoses are the same.
ETYMOLOGY: We are honored to name thisdistinctive gray-furred and short-tailed lime-stone endemic after the late Ðao Van Tien,who when he died on May 3, 1995, at the ageof 78, was emeritus professor of biology at theNational University of Hanoi. Our esteem forthis Vietnamese scholar is reflected in theobituary prepared by Groves and Weitzel(1995: 15216):
Professor Tien was educated in Hanoi under the Frenchcolonial Administration. He taught several generations ofVietnamese scientists and carried his Biology Facultythrough the difficult years after his country gained itsfreedom. Though Professor Tien’s principal works andmany of his papers were in mammalian zoology, his advice,his academic research, and most importantly his studentsupport was never limited to this. Certainly, all seniorbiologists in northern Vietnam today gladly call ProfessorTien their teacher. It is hard to calculate the importance ofthis gentle and thoughtful man in the history of Vietnam’sscience. He was the father of his field in Vietnam. His losswas a great blow for us, though he left for the world anincalculable legacy of studies of the rich biology of Vietnam,and a scholarly tradition for those who will follow in hisfootsteps.
The biodiversity surveys undertaken innorthern Vietnam during the past decade thathave contributed so significantly to knowledgeof mammalian diversity in that country can beviewed as extensions of earlier vertebratesurveys in northern Vietnam from 1957 to1971 initiated and managed by Professor Tien.His results generated large and important
Fig. 4. A live Tonkinomys daovantieni (AMNH 275593). Note the unpigmented distal portion of the tailand the blaze of white on the forehead. Other external traits are described in the text. Photographed by D. P.Lunde, April 2004.
8 AMERICAN MUSEUM NOVITATES NO. 3517

collections of voucher specimens (currentlyhoused at the University of Hanoi), and weresummarized by him in 1985 (Ðao, 1985). Thatbody of field data and museum specimens, andthe scientific results derived from them formedthe foundation of ‘‘Checklist of Mammals inVietnam’’ published nine years later (Ðang etal., 1994), which at the time stood as the finestand most comprehensive annotated list ofVietnam mammals, and today remains notonly an important reference, but a tribute toProfessor Tien, his students, and his collea-gues.
MORPHOLOGICAL DESCRIPTION: Large headand eyes, a stocky body, dark gray ears, shortand bicolored tail, grayish black (gunmetal)upperparts, and dark gray underparts charac-
terize live Tonkinomys daovantieni (figs. 4, 5;table 1). Detailed descriptions of its externalfeatures, skull, and dentition are providedbelow.
External traits: The dense fur coveringupperparts of head and body is grayish blackwith burnished highlights, and consists ofthree types of hairs. The soft underhairs (upto 12 mm long) are gray for their entire length.The overfur layer (up to 15 mm thick) isformed by wide, flexible, and grooved spines,each pale gray for most of its length andtipped with brown or black. Scattered guardhairs project 5210 mm beyond the overhairs,the basal half of each is gray, the distal halfblack. The combination of hair types and theirpigmented banding patterns imparts a thickand semispinous texture, and a burnished,grayish black tone to the upperparts. Thedorsal pelage is less spiny along the sides ofthe body, appears especially soft and gray overthe shoulders and hips, and gradually blendsinto the softer and shorter (up to 8 mm thick)dark gray fur covering the underparts. Thatcoat is formed by soft underhairs mixed withsoft, wide, and grooved spines; both kinds ofhairs are gray for their entire lengths.Although paler than the dorsal coat becauseit has no dark brown guard hairs, no sharpdemarcation exists between dorsal and ventralfur. A white patch (size ranges from 10 mmlong and 5 mm wide to 30 mm by 12 mm)interrupts the dark gray fur over the chest in11 of the 13 specimens; the other two lack thischest mark. On the forehead a blaze of whitehairs (expression ranges from a few hairs toa patch 15 mm long and 5 mm wide) con-trasting with surrounding dark fur is presentin nine specimens; four, including the holo-type, lack the blaze. Sides of the face andthroat are dark gray; the lips, chin, andrhinarium are unpigmented. Fur covering thelower limbs is dark gray everywhere, butshorter than that on the torso.
Eyes, relative to head area, are large,proportionally similar to species of Rattus. Awide unpigmented ring surrounds each narrowand circular, dark brown eyelid. Beyond theeyes, the fur is dark gray covering this part ofthe head, and although somewhat moresuffused with black, this darker tone doesnot define a facial mask.
Fig. 5. Underparts of Tonkinomys daovantieni(AMNH 275593). The dark grayish black dorsalcoat merges gradually into the dark gray venter,which is broken by a white pectoral patch (foundin ten other specimens of the sample of 13).Photographed by D. P. Lunde, April 2004.
2006 MUSSER ET AL.: NEW RODENT FROM VIETNAM 9

The pinnae (external ears) are moderatelylarge, but not disproportionately expansiverelative to body size. They are dark gray,somewhat oval in outline, and rubbery intexture. Short brown hairs (visible only undermagnification) are sparsely scattered overboth outer and inner surfaces, but do not tintthe gray background.
The mystacial vibrissae are either darkbrown or unpigmented, fine, and very long(up to 75 mm); when laid against the head thelongest projects up to 20 mm beyond thedistal margin of each pinna. The longest ineach pair of superciliary (or supraorbital)vibrissae (up to 60 mm long) also extend wellpast the pinna when appressed against thehead. The short submental and interramalvibrissae are unpigmented. The few short (upto 25 mm) genal vibrissae are inconspicuousand unpigmented for most of their lengths.The base of the wrist supports four moderate-ly long (up to 10 mm) unpigmented ulnarcarpal vibrissae, and from the inside of eachheel springs a longer (up to 15 mm) unpig-mented tarsal vibrissa.
The tail is appreciably shorter than thelength of head and body (LT/LHB 5 83%,average of 13 individuals) and it is thick (rangein 13 specimens is 5.7 mm to 6.3 mm, mea-sured laterally at midpoint of tail length) andround in cross section (table 1; fig. 6). Theproximal one-half to three-fourths is darkbrown on dorsal and lateral surfaces (pro-duced by the brown pigment in scales andscale hairs). The ventral surface is white, as areall surfaces of the distal one-fourth to one-half(25%–44% of total tail length among the 13specimens). In a few specimens, scatteredscales in the unpigmented regions retain verypale brown pigment, which provides a slightspeckling to the white portion, usually overthe ventral surface. Scales in the overlappingannuli are moderately large (10 annuli ofscales per centimeter, counted along basalone-third of the tail), and beneath each scaleemerge three hairs. These are bristle-like andabout as long as two scale annuli (1.0 mm)near the base of the tail, but those at the tailtip are softer and longer (526 mm); there isa gradual increase in hair length and texturefrom base to tail tip. Because the hairs aremostly longer than the scale annuli, the tail
Fig. 6. Holotype of Tonkinomys daovantieni.Dorsal (left) and ventral (right) views of the tail.Note increasing length of scale hairs from base totail tip. Basal two-thirds of dorsal and lateralsurfaces of tail is dark brown, produced by brownpigment in both scales and scale hairs (a); scales andscale hairs (c) on underside and distal one-third ofthe tail are unpigmented. See text for discussion ofpattern variation within the complete sample. Tailis round in cross-section (b).
10 AMERICAN MUSEUM NOVITATES NO. 3517
appears somewhat hairy, especially the distalhalf (fig. 6).
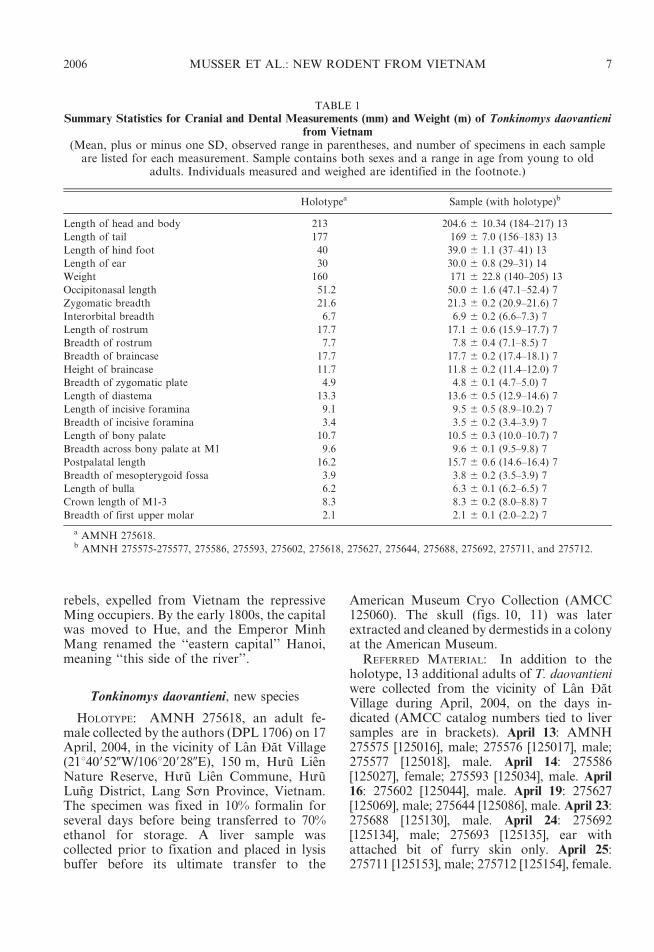
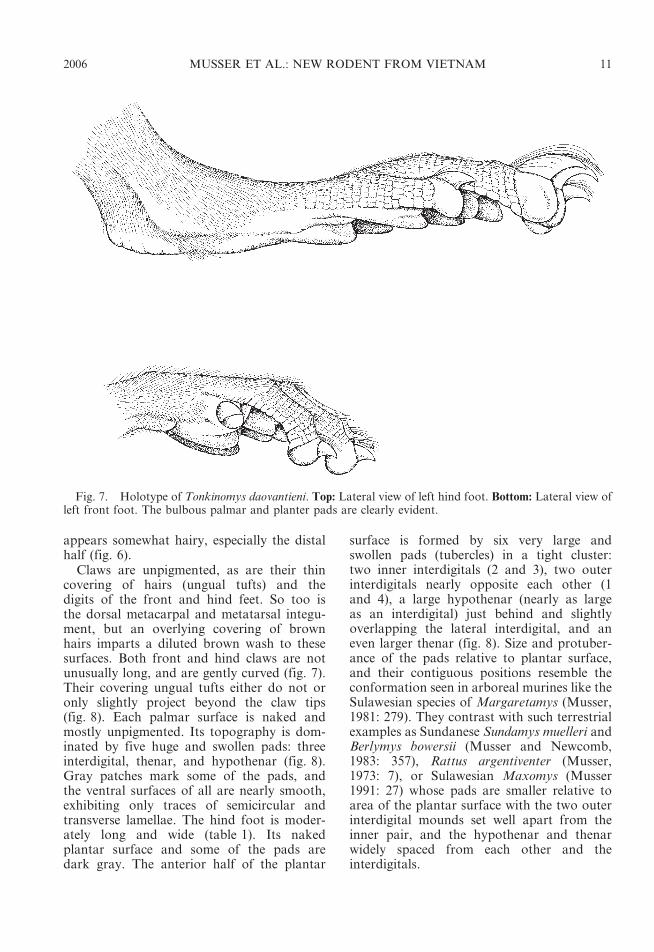
Claws are unpigmented, as are their thincovering of hairs (ungual tufts) and thedigits of the front and hind feet. So too isthe dorsal metacarpal and metatarsal integu-ment, but an overlying covering of brownhairs imparts a diluted brown wash to thesesurfaces. Both front and hind claws are notunusually long, and are gently curved (fig. 7).Their covering ungual tufts either do not oronly slightly project beyond the claw tips(fig. 8). Each palmar surface is naked andmostly unpigmented. Its topography is dom-inated by five huge and swollen pads: threeinterdigital, thenar, and hypothenar (fig. 8).Gray patches mark some of the pads, andthe ventral surfaces of all are nearly smooth,exhibiting only traces of semicircular andtransverse lamellae. The hind foot is moder-ately long and wide (table 1). Its nakedplantar surface and some of the pads aredark gray. The anterior half of the plantar
surface is formed by six very large andswollen pads (tubercles) in a tight cluster:two inner interdigitals (2 and 3), two outerinterdigitals nearly opposite each other (1and 4), a large hypothenar (nearly as largeas an interdigital) just behind and slightlyoverlapping the lateral interdigital, and aneven larger thenar (fig. 8). Size and protuber-ance of the pads relative to plantar surface,and their contiguous positions resemble theconformation seen in arboreal murines like theSulawesian species of Margaretamys (Musser,1981: 279). They contrast with such terrestrialexamples as Sundanese Sundamys muelleri andBerlymys bowersii (Musser and Newcomb,1983: 357), Rattus argentiventer (Musser,1973: 7), or Sulawesian Maxomys (Musser1991: 27) whose pads are smaller relative toarea of the plantar surface with the two outerinterdigital mounds set well apart from theinner pair, and the hypothenar and thenarwidely spaced from each other and theinterdigitals.
Fig. 7. Holotype of Tonkinomys daovantieni. Top: Lateral view of left hind foot. Bottom: Lateral view ofleft front foot. The bulbous palmar and planter pads are clearly evident.
2006 MUSSER ET AL.: NEW RODENT FROM VIETNAM 11

Fig. 8. Holotype of Tonkinomys daovantieni. Left: Palmar view of left front foot. Right: Plantar view ofleft hind foot (compare with the plantar views of Chiromyscus, Niviventer, and Saxatilomys in fig. 9).Abbreviations: hy, hypothenar pad; po, pollex; th, thenar pad; 1—4, first through fourth interdigital pads.
12 AMERICAN MUSEUM NOVITATES NO. 3517
Each female in our sample has four pairs ofmammae: one pectoral, one postaxillary, andtwo inguinal.
Skull: Tonkinomys daovantieni has an elon-gate skull, sturdy in appearance, the visualimpression reflecting a long rostrum, narrowcranium, and robust zygoma. In dorsal view(fig. 10), the rostrum is moderately long andwide, its parallel sides dimpled by a smallnasolacrimal capsule ( just anterior to theanterior spine of each zygomatic plate). Thedistal portion of the rostrum is slightlytapered, the anterior margin of the nasals istriangular, and the adjacent premaxillariesproject well beyond the posterior nasal-frontalsuture. A moderately deep zygomatic notch isevident between the rostral wall and theprojecting anterior spine of the zygomaticplate. The dorsal portion of each lacrimal
bone (in the anterolateral pocket of theorbit) is small, rectangular, and fused withthe dorsal maxillary zygomatic root (noshared suture with the frontal). Dorsolateralmargins of the interorbit, which is about aswide as the rostrum, are defined by highridges that are prominent as they sweepback along the dorsolateral postorbital mar-gins of the frontals; these ridges becomeweaker in expression and less pronouncedwhere they extend (as temporal ridges) ontothe parietal margins (accentuating dorsolater-al rims of the cranium) to about the level ofthe posterior edge of the squamosal zygomaticroot. Here the temporal ridge is barelydiscernable, and the parietal extends ventrallyto form the dorsolateral cranial wall betweenthe posterior margin of the zygomatic rootand the lamboidal ridge; a bony swelling
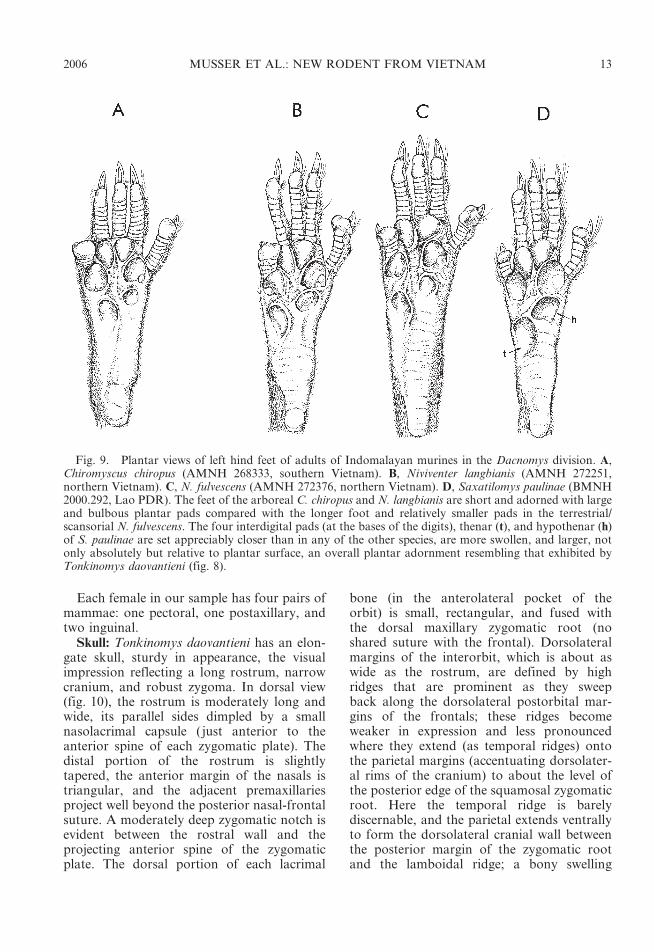
Fig. 9. Plantar views of left hind feet of adults of Indomalayan murines in the Dacnomys division. A,Chiromyscus chiropus (AMNH 268333, southern Vietnam). B, Niviventer langbianis (AMNH 272251,northern Vietnam). C, N. fulvescens (AMNH 272376, northern Vietnam). D, Saxatilomys paulinae (BMNH2000.292, Lao PDR). The feet of the arboreal C. chiropus and N. langbianis are short and adorned with largeand bulbous plantar pads compared with the longer foot and relatively smaller pads in the terrestrial/scansorial N. fulvescens. The four interdigital pads (at the bases of the digits), thenar (t), and hypothenar (h)of S. paulinae are set appreciably closer than in any of the other species, are more swollen, and larger, notonly absolutely but relative to plantar surface, an overall plantar adornment resembling that exhibited byTonkinomys daovantieni (fig. 8).
2006 MUSSER ET AL.: NEW RODENT FROM VIETNAM 13

outlines the parietal-squamosal connection allthe way to the lamboidal ridge. The interpar-ietal is long (in anterior-posterior plane) andmoderately wide (its lateral margins are withinthe parietal and supraoccipital sutures, notextending to the parietal-squamosal suture)and its posterior two-thirds roofs a deepocciput with a slightly convex or chevron-shaped posterior margin. Zygomatic archesare robust and arranged nearly parallel to one
another; neither zygoma bows out far fromthe skull.
The skull appears long, low, and withoutsignificant basicranial flexion when viewedfrom lateral perspective (fig. 11). The rostrumis rectangular in side view, and extends slightlybeyond the incisor faces to enclose the nasalaperature in a short tube in which nasalsprovide the roof and premaxillaries the sides.Much of the ventral maxillary root of each
Fig. 10. Holotype of Tonkinomys daovantieni. Dorsal (left) and ventral (right) views of cranium.Measurements are listed in table 1.
14 AMERICAN MUSEUM NOVITATES NO. 3517

zygomatic arch lies anterior to the molar row,with the posterior margin of the zygomaticplate extending behind the front margin of thetoothrow (0.521.0 mm is the range in oursample of skulls). The zygomatic plate is wide,its convex anterior margin projecting beyondthe dorsal maxillary zygomatic root to forma moderately deep zygomatic notch (best seenin dorsal view). This anterior spine of the plate
does not project far enough forward toconceal the nasolacrimal capsule, which issmall and only slightly inflated. Maxillary andsquamosal zygomatic processes are joined bya long jugal to form a thick and robustzygomatic arch. The squamosal root of eacharch is situated high on the side of the craniumwhere its posterior ridge2like portion extendshorizontally just below the posteroventral
Fig. 11. Holotype of Tonkinomys daovantieni. Top: Lateral view of cranium. Middle: Lateral view of leftdentary with incisor and molars. Bottom: Medial view of same dentary.
2006 MUSSER ET AL.: NEW RODENT FROM VIETNAM 15

margin of the parietal, which projects ventradto form the dorsolateral side of the cranium.The low ridge of the squamosal zygomaticroot disappears 10–15 mm anterior to thevertical ridge formed by the squamosal-exoc-cipital suture. The configuration is similar to,but not identical with, that documented forspecies of Leopoldamys (see Musser, 1981:fig.18) and Niviventer (Musser et al., 2005: figs. 7and 8). Dorsal to the auditory (ectotympanic)bulla and posterior to the squamosal zygo-matic root, the squamosal forming the side ofthe cranium is intact except for a minutesubsquamosal foramen at the prominent ridgeformed by the squamosal-exoccipital suture.This foramen is the only landmark identifyingwhere the squamosal and a wide hamularprocess of the squamosal have completelyfused to form the solid cranial wall. Aspacious postglenoid foramen that is confluentin some specimens with a wide postalar fissure(ventral to the postglenoid foramen) separatesthe squamosal margin from the periotic flangeand dorsoanterior surface of the ectotympanic(auditory) bulla. That capsule is attached tothe squamosal dorsally by the posteroventralportion of that bone, which represents thecranial wall fused with a wide hamularprocess. There is no anterior attachment,which is the usual pattern among mostmurines. An anterior flange of the periotic,the tegmen tympani, is present (and partlyseparates the dorsal postglenoid foramenfrom the ventral postalar fissure), but isseparated by a narrow or moderately widegap from the posterior margin of the squa-mosal in most specimens; there is no posteriorsuspensory process of the squamosal towhich the tegmen tympani could attach (insome cricetid rodents, the tegmen tympanioverlaps a posterior projection of the squa-mosal to form one of the suspensory attach-ments between ectotympanic capsule andsquamosal; see the diagrams in Voss, 1993:19). The moderately inflated mastoid is tightlysutured to squamosal and exoccipital. It iscompletely ossified, without the moderate orlarge fenestrae found in some murines(Crunomys celebensis, for example; Musser,1982: 38) and sigmodontines (see Carletonand Musser, 1989: 34); only a very smallopening penetrates the dorsal mastoid-exocci-
Fig. 12. Holotype of Tonkinomys daovantieniillustrating details in enlarged view of left basicra-nial region. Abbreviations: ab, auditory bulla; al,alisphenoid; bo, basioccipital; bp, bony palate; cc,carotid canal; fo, foramen ovale; hp, hamularprocess; iog, groove for the infraorbital artery;mlf, middle lacerate foramen; pp, pterygoid plate(its ventral surface is the pterygoid fossa); ppm,posterolateral palatal margin; pt, periotic; spv,sphenopalatine vacuity; sq, squamosal; stf,stapedial foramen.
16 AMERICAN MUSEUM NOVITATES NO. 3517

pital suture. The foramen magnum is directedcaudad (which reflects the horizontal ratherthan arched axis of the skull—no significantbasicranial flexion), and the deep occiputprojects slightly beyond posterior surfaces ofthe occipital condyles. The ethmoid foramenwithin the orbit is tiny, and the opticforamen moderately large. The dorsal palatineforamen is separate from the sphenopalatineforamen, a configuration usual amongAsian murines, with a few exceptions wherethe two foramina are coalesced (Musser, 1982:22). Eight specimens have a bony alisphenoidstrut (ranging from moderately wide toslender in the sample) on each side of theskull posterior to the orbit and dorsad to thepterygoid ridge (the lateral edge of thepterygoid plate). The strut separatesthe foramen ovale accessorius from thecombined buccinator-masticatory foramenon the side of the cranium, and obscures theanterior opening of the alisphenoid canal. Onespecimen exhibits a strong strut on the leftside, but only a bony thread on the right. Onlyone individual in our sample of skulls lacksstruts (which are represented by only a dorsalnubbin on each side). In this specimen, theopening to the alisphenoid canal is evident, theforamen ovale accessorius and buccinator-masticatory foramina coalesce (basically dis-appear), and the course of the buccinator andmasticatory nerves is marked by a shallowgroove extending from the foramen ovale ontothe side of the alisphenoid.
The short rostral tube anterior to faces ofthe incisors, wide rostrum, long incisiveforamina and bony palate, diverging molarrows, moderately wide mesopterygoid fossaand pterygoid plates, and small bullae are thetopographic highlights when the skull isstudied from ventral perspective (figs. 10,12). Just behind the incisors is a tiny inter-premaxillary foramen, and beyond it therostrum is perforated by long and spaciousincisive foramina, their posterior rims situatedslightly before (0.5 mm) anterior margins ofthe first molars, even with those molarsurfaces, or projecting slightly (not more than0.5 mm) between them. The bony palate iswide and long, extending 1.5–2.0 mm beyondthe molar rows to form a broad platform witha slightly concave and smooth posterior
margin (up to 20% of the palate extends pastthe third molars) unmarked by any bony ridgein most specimens, slightly thickened inothers. Each posterolateral margin of thisshelf extends as a flat, bony surface onto theanterior portion of each pterygoid plate whereits sharp diagonal border marks the antero-medial margin of each pterygoid fossa. Theventral surface of the bony palate is smoothexcept for a pair of oblong posterior palatineforamina (in the maxillary-palatine sutureopposite the middle or posterior half of eachsecond molar), a pair of shallow grooves, andsmall or minute nutrient foramina scatteredover the palatal shelf. The mesopterygoidfossa is wide, and dorsolateral margins of itsroof are perforated by long and moderatelywide sphenopalatine vacuities. On each side ofthe pterygoid fossa, a robust hamular processof the pterygoid marks the boundary betweenthe mesopterygoid fossa and each adjacentpterygoid plate, its ventral surface sculpturedby a shallow pterygoid fossa. The bonypterygoid plate is breached by the largeventral opening of the foramen ovale in itsposterior half; its anteromedial portion alongthe rim of the bony palatal extension is slightlyfenestrated (irregular traces of the sphenopter-ygoid vacuity). A shallow trough (in which theinfraorbital artery passes) scores the ventralmargin of the pterygoid plate between theventral opening of the foramen ovale and themiddle lacerate foramen. Small relative tooverall size of the skull, the globular and onlyslightly inflated ectotympanic (auditory) bulladoes not conceal nearly all of the periotic as itdoes in some other murine groups, species ofRattus, for example. In Tonkinomys daovan-tieni, a wide, wedge-shaped segment of theperiotic is exposed between the dorsal capsularmargin and basioccipital, and this wedge ofperiotic extends anteriorly to form the dorsaland slanting posterodorsal wall of the carotidcanal. The conformation resembles that illus-trated by Carleton and Musser (1989: 33) forthe sigmodontine, Oligoryzomys, and is unlikemany other muroid rodents in which a moreinflated ectotympanic capsule covers most ofthe periotic and the anteromedial margin ofthis capsule meets the basioccipital, so theopening to the carotid canal is bounded by theectotympanic capsule and adjacent basiocci-
2006 MUSSER ET AL.: NEW RODENT FROM VIETNAM 17

pital (the condition in Oryzomys palustrisdepicted by Carleton and Musser, 1989: 33;and species of Rattus and its allies illustratedin Musser and Heaney, 1992: 99). Taperingrostrally from the ectotympanic capsule isa moderately short and wide bony eustachiantube. A spacious middle lacerate foramenseparates the anterior slope of the ectotympa-nic capsule from posterior margin of thepterygoid plate.
The cephalic arterial pattern possessed byTonkinomys daovantieni is not unique amongmembers of Murinae (as that subfamily isdelineated by Carleton and Musser, 1984;Musser and Carleton, 2005). As in all de-scribed species of Niviventer, Chiromyscus,Saxatilomys, and Leopoldamys, T. daovantienilacks a sphenofrontal foramen and relatedsquamosoalisphenoid groove, the petrotym-panic fissure between bullar capsule andperiotic is perforated by a spacious stapedialforamen continuous laterally with a deepgroove on dorsal surface of the periotic, andthere is a shallow but wide groove on theposterior ventral surface of the pterygoid plate(fig. 12). This set of osteological landmarkssignals a cephalic arterial pattern in which thesupraorbital branch of the stapedial artery isabsent and the orbital circulation derives fromthe distal portion of the infraorbital branch.The infraorbital is a continuation of thestapedial artery, which passes over the dorsalgroove in the periotic, through the middlelacerate foramen, continues in the shallowgroove ventral to the posterior margin of thepterygoid plate to course through the ali-sphenoid canal, and emerges onto the orbitalfloor through the anterior alar fissure.Common to most murines (Carleton andMusser, 1984; Musser and Newcomb, 1983;Musser and Heaney, 1992), this pattern ofvessels and associated osteological landmarksis also characteristic of some sigmodontinerodents (described and diagramed forOligoryzomys by Carleton and Musser, 1989;pattern 2 in Voss, 1988), and is derivedcompared with the cephalic arterial patternhypothesized to be primitive for muroidrodents (Bugge, 1985; Carleton, 1980).
Mandible: The sturdy dentary supportsa delicate coronoid process, relative to extentof the deep ascending ramus, a moderately
deep sigmoid notch between coronoid andprominent condyloid processes, and a shallow-ly concave posterior margin connecting con-dyloid and angular processes (fig. 11). Incontrast with the wide and conspicuousventral masseteric ridge, the dorsal ridge iseither undetectable or indicated by a shallow,elongate depression. Besides the ventral mas-seteric ridge, the only other significant struc-ture that marks an otherwise smooth lateraldentary surface is a slight bulge below theanterior root of the coronoid process that inturn marks termination of the incisor alveoluswithin the bone. In no specimen did the incisorcapsule extend farther caudad to project intothe condyloid process (checked by shininga high-intensity light through the dentary) as itdoes in some other murines (Musser andHeaney, 1992: 84). Topography of the lingualdentary surface resembles that characteristicof most other murines, the primary landmarksbeing a prominent lingual ridge above whichis the large mandibular foramen (as inPapagomys armandvillei, for example; seeMusser et al., 1986: 5). The body of thedentary projects anteriorly beyond the molarrow as a moderately short and stocky tubecontaining the incisor. Just in front of themolar row on the dorsolateral labial surface isa small mental foramen.
Dentition: Enamel faces of upper and lowerincisors are pale orange and smooth, withoutgrooves or shallow sulci. Thickness of theenamel relative to the dentine is similar to thatin species of Rattus and many other murines(Musser and Heaney, 1992: 79). The upperincisors curve caudad after emerging fromthe rostrum (opisthodont conformation; seeThomas, 1919; Hershkovitz, 1962: 103).Gnawing edges of the uppers are at rightangles to the long axis of the skull, so theircombined tips form a straight cutting edge (asin species of Rattus; see Musser and Heaney,1992: 79), not curved or V-shaped (as in somePhilippine and Sulawesi endemics; see Musserand Heaney, 1992; Musser and Durden, 2002).The lower incisors are moderately long andcurved, their cutting tips either straight orarcuate.
Alveolar patterns for roots of upper molarsare distinctive. A large anterior root, tworobust lingual roots, and a large posterior
18 AMERICAN MUSEUM NOVITATES NO. 3517

holdfast anchor each first upper molar. Nolabial root is present between the largeanterior and posterior anchors. Three rootshold the second and third molars in place:a large anterior and posterior, and smalllingual. Two large roots anchor each lowermolar. Except for the first upper molars, thepattern of roots in all our examples ofTonkinomys daovantieni is primitive for mur-ines (Musser and Newcomb, 1983; Musser andHeaney, 1992).
Upper molar rows are moderately long(about 17% of occipitonasal length) andstrongly diverge posteriorly (width of bonypalate is considerably greater between thirdmolars than between the first; see fig. 10).Occlusal surfaces of the brachydont (low-crowned) molars are simple in topography,consisting of tightly pressed rows exposingchunky chewing surfaces in the shapes ofnarrow chevrons and diamonds (fig. 14).Occlusal surface of each first upper molarconsists of two anterior rows of cusps, each inthe form of a tight chevron, and a largeroughly diamond-shaped posterior surface.The most anterior chevron is comprised ofcusps t1, t2, and t3; cusps t2 and t3 are socoalesced that their limits are obliterated, atleast in adults. The blended cusps t4, t5, and t6form the second chevron, although the labialand lingual cusps retain some definition. Thelarge, chunky posterior third of the molarconsists primarily of cusp t8 merged witha smaller, indistinct cusp t9 to form a roughlydiamond-shaped chewing surface. On theocclusal surface of each second upper molar,the anterior chevron is represented by onlya large cusp t1 forming the anterolingualportion of the tooth. In seven out of 10specimens, a small cusp t3 sits on theanterolabial margin; in two others, the cuspis missing and the anterolabial border ismarked by only an inconspicuous cingulum;and presence or absence of cusp t3 in the tenthspecimen cannot be determined because themolars are too worn. The complete chevronand chunky posterior portion of the secondmolar, in size and shape, resembles compara-ble rows in the first molar. Each third molarhas an occlusal pattern that is basicallya compacted form of that characterizing thesecond molar: a conspicuous cusp t1; minute
cusp t3 (seen in young rats but unidentifiablein older animals with more worn chewingsurfaces); a complete arcuate lamina derivedfrom the total coalesence of cusps t4, t5, andt6; and small posterior chunk either diamond-shaped or oblong in outline (formed primarilyof cusp t8; cusp t9 is either absent orunidentifiable due to its total incorporationinto cusp t8). We could not detect a cusp t7(on the lingual cingulum between cusps t4and t8 when present; see Musser andNewcomb, 1983: 333), posterior cingulum(pressed against the posterior margin of cuspt8, if present; see Musser and Newcomb, 1983:333), accessory cusplets, or longitudinal crestsconnecting the cusps to form a garlandconfiguration (‘‘stephanodonty’’; see Schaub,1938, and Misonne, 1969) on any of the uppermolars. When present these structures providecomplexity to the occlusal planes and in somemurines significantly increase chewing sur-faces.
Occlusal planes of the lower molars are alsocomprised of uncomplicated patterns (fig. 14).An anterolingual cusp coalesced with a smalleranterolabial forms the anteroconid, the mostanterior cusp row, which is narrower than thelamina behind it. The second and third are inthe shape of narrow chevrons and representunions of protoconid-metaconid and hypoco-nid-entoconid, respectively. A roughly trian-gular posterior cingulum forms the entireposterior fourth of the molar. Except forlacking an anteroconid, occlusal cusp patternsof the second molar resemble patterns seen inthe first. The anterior half of each third molarconsists of a front chevron formed by union ofprotoconid and metaconid and behind thata large oblong structure likely representing thecomplete fusion of hypoconid and entoconid.Anterolabial cusps are absent from secondand third molars in most specimens, but weare not certain about others in the samplebecause their molars are worn to a level suchthat, if present, these cusps are now indistinctbecause they have completely coalesced withthe adjacent lamina; we did find evidence ofa small anterolabial cusp on the third molar ofthe holotype (fig. 14). We had the samefrustration determining the frequency of pos-terior labial cusplets on some specimens. Thissmall cone is prominent and forms the
2006 MUSSER ET AL.: NEW RODENT FROM VIETNAM 19

anterolabial margins of the posterior laminaeof first and second molars in the holotype(fig. 14). The cusplets are present but in-conspicuous on first and second molars inthree other specimens (the laminar outlinereflects the blending of a cusplet with theanterolabial laminar margin), and are eitherabsent or undetectable in seven rats (becausethe cusplet has completely coalesced with theadjacent cusp). A posterolabial cusplet clearlydoes not occur on any third molar in any ofthe specimens at hand.
COMPARISONS
The morphological attributes of Ton-kinomys daovantieni place it in the Dac-nomys Division of Musser and Carleton(2005), which contains the IndomalayanChiromyscus (one species), Dacnomys (one),Leopoldamys (six), and Niviventer (17); SriLankan Srilankamys (one); and the recently
described Lao PDR Saxatilomys (one species).Musser (1981), Musser and Newcomb (1983),Musser and Heaney (1992), Musser andCarleton (2005), and Musser et al. (2005) havechronicled the geographic distributions ofthese genera, along with their taxonomies,descriptions of morphologies, and postulatedphylogenetic relationships. In conformation ofbody and limb proportions, and in character-istics of skull and teeth, T. daovantieni mostclosely resembles species of Leopoldamys,Niviventer, Chiromyscus, and Saxatilomys.Our comparisons contrast Tonkinomys firstwith extant examples of Leopoldamys, thenwith Niviventer and Chiromyscus, and finallywith Saxatilomys.
A large body of literature has accumulatedover the past two decades reporting fossilmurines, mostly isolated molars and incisors,which have been recovered from Pliocene,Pleistocene, and Holocene sediments in theIndomalayan region. Some material has been
Fig. 13. Diagram of maxillary (left) and mandibular (right) molar rows from right side of Bunomyschrysocomus illustrating structural terms. Maxillary molars: cusps are numbered according to Miller’s (1912)scheme and referred to in the text with the prefix ‘‘t’’. Mandibular molars: a-lab, anterolabial cusp; a-ling,anterolingual cusp; alc, anterior labial cusplet; ed, entoconid; hd, hypoconid; md, metaconid; pc, posteriorcingulum; pd, protoconid; plc, posterior labial cusplet.
20 AMERICAN MUSEUM NOVITATES NO. 3517

described as new taxa whose diagnostic traitsidentify them as members of the DacnomysDivision, and other samples have been re-garded as examples of extant members of thatgroup. We examined pertinent publications todetermine whether any of the new taxa thatare based on fossils represent examples ofTonkinomys. The significance for nomencla-ture is obvious. Equally important is thezoogeographic context. If some samples offossils that have been identified as eitherextinct or extant species are actually examplesof Tonkinomys, or close phylogenetic relatives,they would provide a window through whichcould be glimpsed the past distribution of thisunique limestone rat, and the geologic depthof its evolutionary history.
Leopoldamys: The smallest extant memberof the six currently recognized species ofLeopoldamys (Musser and Carleton, 2005) isthe Thai L. neilli (only the extinct L. minutus issmaller, described from samples recoveredfrom late Pliocene–early Pleistocene fissureand cave sediments; Chaimanee, 1998), andthe head and body proportions, body size,
weight, and posture of the live animal closelyresemble Tonkinomys daovantieni (Marshall,1988: 485, provided a photograph of a liveLeopoldamys). Although the two species aresimilar in body size (table 2), they are strik-ingly dissimilar in other particulars of externalmorphology. Leopoldamys neilli has: (1) short,sleek brown fur over upperparts of head andbody that is sharply demarcated from whiteunderparts (long, semispinous, grayish blackfur over the dorsum and dark gray venter in T.daovantieni); (2) slightly smaller external pin-nae, both absolutely and relative to head andbody length; (3) a long, slender, tapered tailthat is much longer than length of head andbody, LT/LHB 5 124% on average, andsimilar to an inverted U in cross-section—flatventral surface and arched across sides andtop (less tapered in T. daovantieni, chunky andmuch shorter than head and body, LT/LHB 583%, and round in cross-section); (4) a veryslender, pointed tail tip that is covered withshort scale hairs no longer than 122 mm (tipof tail is stubby in T. daovantieni, covered bymuch longer hairs, 527 mm, and brush-like in
Fig. 14. Occlusal views of maxillary (left) and mandibular (right) molar rows from right side of theholotype of Tonkinomys daovantieni (CLM1-3 5 8.3 mm, CLm1-3 5 8.3 mm). See figure 13 for terminologyof cusps and cusplets.
2006 MUSSER ET AL.: NEW RODENT FROM VIETNAM 21

appearance); and (5) swollen plantar tuberclessituated in distal two-thirds of plantar surface,as seen in Tonkinomys (figs. 9, 10), but slightlysmaller relative to plantar area—the thenarpad, for example, forms about 17% of plantersurface (T. daovantieni has relatively moreexpansive plantar pads—the thenar constitu-tes about 20% of plantar surface). Short, sleekfur with the coat that covers upperparts of thehead and body strikingly contrasting with theventral covering is characteristic of all extantspecies of Leopoldamys. Also common to allextant species in that genus is a very long andtapered tail, with its cross section resemblingan inverted U, and its tip covered by veryshort bristles. Elongate hind feet are commonto all Leopoldamys, as are the number ofplantar tubercles, their size relative to plantarsurface, and distribution in the distal two-thirds of the plantar surface.
The skull of L. neilli is similar to that of T.daovantieni in overall size (table 2), and thetwo are remarkably alike when viewed froma dorsal perspective (compare the cranialphotographs of Leopoldamys in Musser,1981: 259, and the drawing on p. 264, withthe renditions in fig. 10). In ventral view,however, the two species are strikingly dissim-ilar. Compared with the Vietnam species, L.neilli (and all other extant species ofLeopoldamys) has: (1) significantly shorter
incisive foramina, their posterior margins wellanterior to faces of first upper molars (longerin T. daovantieni, the posterior edges locatedjust before anterior margins of first molars,even with them, or slightly projecting betweenthe first molars); (2) a much shorter bonypalate, its posterior margin aligned somewherebetween the middle or end of each third uppermolar, depending upon the species and vari-ation within each sample (longer in T.daovantieni, extending beyond the molar rowsto form a wide platform); (3) longer postpalatallength, which reflects the shorter bony palate(much shorter in T. daovantieni); (4) wider,chunky molars relative to size of skull, whichis correlated with an absolutely narrower bonypalate (smaller, gracile molars relative to sizeof skull, and an attendant absolutely widerbony palate in T. daovantieni); (5) molar rowsthat barely diverge posteriorly (molar rowsdiverge appreciably along anterior-posterioraxis in T. daovantieni); (6) a small, moderatelyinflated ectotympanic bulla relative to size ofthe skull (more inflated capsule in T. daovan-tieni, and absolutely larger in most examples);and (7) a very long, bony eustachian tube,especially contrasted with the small bulla(much shorter eustachian tube in T. daovan-tieni). These differences also separate T.daovantieni from specimens we examinedrepresenting the other five species of
TABLE 2
Summary Statistics for Cranial and Dental Measurements (mm), and Tail-to-Body Ratio of Tonkinomys,Leopoldamys, and Niviventer Selected to Demonstrate Contrasts in Relative Tail Length, and Size of Body,
Skull, and Maxillary Molar Rowa
(Mean, observed range in parentheses, and number of specimens are listed for each measurement;values for L. neilliare means derived from a sample of 12. LT/LHB expressed as %.)
T. daovantieni L. neillib N. andersoni c N. confucianusc
(Vietnam) (Thailand) (Sichuan, Yunnan) (Sichuan)
LHB 204.6 (184–217) 13 217 171.6 (150–198) 46 143.2 (125–170) 24
LT 168.9 (156–183) 13 270 229.7 (194–269) 44 182.7 (160–204) 23
LT/LHB 83 124 134 128
LHF 39.0 (37–41) 13 42 35.1 (31–40) 47 29.8 (28–32) 20
LE 30.0 (29–31) 13 27 26.4 (22–28) 19 21.0 (20–23) 24
ONL 50.0 (47.1–52.4) 7 51.9 41.7 (38.7–45.7) 37 36.9 (33.9–40.5) 25
CLM 8.33 (8.0–8.8) 7 8.3 7.3 (6.9–7.5) 18 5.8 (5.3–6.3) 59
a Leopoldamys neilli is the smallest in body size of any extant Leopoldamys, N. andersoni is among the largest within
Niviventer, and N. confucianus represents most species in the genus in body size, along with body and tail proportions.b Values are from Marshall (1988: 485), and are means derived from 12 adults.c Lengths of head and body, tail, hind foot, ear, and ONL are from Musser and Chiu (1979: 584); lengths of maxillary
molar rows are unpublished summaries by Musser and Lunde (ms.) derived from AMNH samples collected in China.
22 AMERICAN MUSEUM NOVITATES NO. 3517

Leopoldamys. Our study of those compara-tive samples also reveals that L. neilli and theother species of Leopoldamys have wider andsturdier alisphenoid struts relative to skullsize, and these struts are present in everyindividual of each species (relatively weakerstruts in T. daovantieni, and absent fromsome specimens).
Patterns of roots beneath certain molarsthat are characteristic of T. daovantieni aredissimilar to those exhibited by species ofLeopoldamys. Both extant samples (material inAMNH and USNM; also see Musser, 1981:260) of Leopoldamys and those represented byfossils (Chaimanee, 1998; Zheng, 1993) havefour-rooted upper molars: large posterior,single large lingual (we have never encoun-tered a specimen with two lingual roots), largeanterior, and a small labial, which is some-times divided (also four roots in T. daovantie-ni, but there are two large lingual holdfastsinstead of one, and no labial root). AllLeopoldamys have a second upper molar thatis anchored by either three or four roots (onlythree roots in all our T. daovantieni). BothLeopoldamys and Tonkinomys have three-rooted third upper molars. All species ofLeopoldamys have four-rooted first lowermolars: two large anterior and posterioranchors, and smaller labial and lingual hold-fasts (all examples of T. daovantieni have onlytwo roots, an anterior and posterior coequalin size). Second and third lower molars inLeopoldamys have two roots (large anteriorand posterior), which is also the pattern in T.daovantieni.
In addition to the marked contrasts inrobustness and width of molars betweenLeopoldamys and Tonkinomys, occlusal pat-terns of molars are different (compare illustra-tions of molar rows for Leopoldamys inMusser, 1981: 262, with those of Tonkinomysin fig. 14). Coronal surfaces of maxillarymolars are characterized by simple patternsin both genera (no cusp t7 or posteriorcingulum on any molar, no accessory cusplets,no longitudinal crests connecting the primarycusps), but the conformations of laminaeforming the occlusal surfaces are dissimilar.The anterior lamina of the first molar inLeopoldamys consists of a nearly horizontalsegment, formed by the complete fusion of
cusps t2 and t3, and a discrete cusp t1 on itsposterolingual margin. The second lamina isnot as straight but gently arcuate. These arediagnostic molar traits for Leopoldamys. Incontrast, the two comparable laminae inTonkinomys are tight chevrons—cusps t1 andt3 are opposite each other in the first lamina,as are cusps t4 and t6 in the second.Furthermore, cusps t4 and t6 in T. daovantie-ni, although coalesced with the central cusp t5,are large relative to overall dimensions of thelamina, and retain part of their identity ascusps. Comparable cusps in the second laminain Leopoldamys are relatively smaller and sobroadly merged with cusp t5 that theirboundaries are not determinable. The largechunky posterior lamina in Leopoldamys iswider than long, but roughly diamond-shapedin Tonkinomys. Contrasts in second uppermolars are similar to those seen in the first.The third upper molars in Leopoldamys havea stretched, ridge-like cusp t4 compared withthe shorter version in Tonkinomys; otherwise,the occlusal patterns in the two genera aresimilar.
Cusps form either nearly straight or gentlyarcuate lamina in the lower (mandibular)molars of Leopoldamys. The posterior cingulaare wide and elliptical in cross-section in allmembers of that genus, prominent posteriorcusplets adorn the labial margin of the firstand second molars, and two nearly straightlaminae form the occlusal surface of each thirdmolar (see the illustrations in Musser, 1981:260). In parallel with the upper molars ofTonkinomys, the two cusps forming thelaminae of its lower molars are diagonallyoriented so that after wear they form chevronsin which the arms are very close, the posteriorcingula are roughly triangular, labial cuspletsare indistinct or not present on most speci-mens (prominent only in the holotype), andthe occlusal surface of the third molar isformed by a distinct anterior chevron-shapedlamina and a posterior oblong chunk.
Niviventer and Chiromyscus: We examinedsamples of all 17 of the currently recognizedspecies of Niviventer and the single species ofChiromyscus (taxonomy and geographic dis-tributions for both genera are summarized inMusser and Carleton, 2005; specimens form-ing the comparative samples we utilized are
2006 MUSSER ET AL.: NEW RODENT FROM VIETNAM 23

listed in the Appendix by Musser et al., 2005).We preface our comparisons by explainingthat the present contents of Chiromyscus andNiviventer will likely be altered after carefulsystematic revision of these genera. Musser(1981) characterized them, provided provi-sional diagnoses and contents, but his exposi-tion was more of a taxonomic summary andmeant to be a working hypothesis, not a sys-tematic revision. That kind of study has yet tobe realized. Most species will remain inNiviventer, but two sets may eventually bedifferently allocated. Except for its nail-likeclaw on the hallux, shelf-like ridges borderingdorsolateral margins of the postorbital regionand braincase, pattern of roots beneath the firstupper molar (table 3), color pattern of thedorsal pelage, and large and very bulbousplantar pads set close together, other externaland most cranial and dental characteristics ofthe arboreal C. chiropus are similar to thosefeatures in the species of Niviventer, particular-ly N. langbianis (Musser, 1981; Musser andLunde, ms.). The latter is also arboreal(observations by Lunde and Musser derivedfrom trapping surveys in Vietnam), and shareswith C. chiropus comparable prominent post-orbital and temporal ridges, the number ofroots anchoring each first upper molar, a nail-like claw on the hallux, a pelage color patternthat does not match in details but is similar,and equal expression of plantar pads. Arevisionary inquiry may either move N. lang-bianis to Chiromyscus, or C. chiropus will besubsumed within Niviventer (that name wouldthen be a synonym of the older Chiromyscus).
Niviventer andersoni and N. excelsior formanother set now allocated to Niviventer thatwill probably be removed. Musser (1981)described a suite of cranial and dental traitsheld by these two species that contrast withthe characters common to the other species inNiviventer. Should information from otheranatomical systems, and that derived fromchromosomal and biochemical surveys, sup-port the cranial and dental distinctions, he(p 328) noted ‘‘… that the phylogenetic re-lationship between Niviventer and the N.andersoni division [would be] better expressedby excluding the latter from Niviventer andplacing N. andersoni and N. excelsior ina genus of their own.’’
We accept the current, although provision-al, contents of Niviventer for our compara-tive context. We will also refer only to‘‘Niviventer’’, understanding that the contrastsbetween species in that genus and Tonkinomysare much the same as between the latter andC. chiropus.
While the body conformation of Ton-kinomys daovantieni recalls small-bodiedLeopoldamys, no such resemblance existsbetween the Vietnamese animal and anyspecies of Niviventer. Among the largestspecies of Niviventer in body size is N.andersoni, which is still appreciably smallerthan T. daovantieni (table 2). In all species ofNiviventer, dorsal head and body fur rangesfrom buffy or brownish gray through brownto reddish brown (Musser, 1981). Niviventerbrahma and N. eha have grayish white under-parts (Musser, 1970), N. hinpoon has a buffypale gray venter (Marshall, 1988), but all otherspecies have white venters. No example ofNiviventer has Tonkinomys’s unique dorsaland ventral pelage coloration. Some speciesof Niviventer, however, do have semispinouspelage similar in texture to that possessed byT. daovantieni (N. fulvescens and N. tenasterare good examples). The conformation oftail and proportional relationship betweenlengths of tail and body in Tonkinomys(stocky, with a stubby tip, and significantlyshorter than combined head and bodylength) does not occur in any species ofNiviventer. The tail is tapered to a pointedtip and gracile in appearance in all species ofNiviventer, and much longer than length ofhead and body in all species except N. hinpoon,in which the tail is equal to or slightly longerthan head and body length (table 2; Marshall,1988; Musser, 1970, 1973, 1981; Musser andChiu, 1979; Lunde and Son, 2001). Theplantar pads of Tonkinomys are larger,closer together, and more swollen relative toplantar surface than in any terrestrial orscansorial species of Niviventer, including N.hinpoon, which is thought to be closelyassociated with limestone cliff habitats(Marshall, 1988), or even the arboreal N.langbianis and Chiromyscus chiropus (fig. 9);N. fulvescens (fig. 9) exhibits the configurationof plantar pads that is common to mostspecies of Niviventer.
24 AMERICAN MUSEUM NOVITATES NO. 3517

The skull of Tonkinomys is appreciablylarger (as indexed by occipitonasal length)and more robust than all species of Niviventer,even N. andersoni, which has the largest skullamong the species in the genus (table 2).The skull of Tonkinomys, however, is nota giant version of Niviventer; compared withNiviventer, Tonkinomys has: (1) insignificantbasicranial flexion (most species of Niviventerdisplay marked basicranial flexion); (2) max-illary molar rows that are situated fartherforward relative to posterior margin of thezygomatic plate (front of molar row eithereven with back of zygomatic plate or wellbehind it in most specimens of Niviventer);(3) the squamosal root of the zygoma origi-nating higher on side of the braincase (setlower in Niviventer, particularly N. andersoniand N. excelsior); (4) more slender alisphenoidstruts relative to size of skull, with somespecimens lacking struts (relative to skull size,alisphenoid struts are more substantial in allspecies of Niviventer, and present in all speci-mens we examined); (5) strongly posteriorlydiverging molar rows (parallel or only slightlydiverging in Niviventer); (6) a bony palateprojecting appreciably beyond posterior mar-gins of third molars to form a wide platform(posterior margin of bony palate endsabout 0.5 mm anterior to posterior surfacesof upper third molars, even with them, orno more than about 0.5 mm beyond in allspecies of Niviventer); (7) a posterior marginof the bony palate that is smooth or onlyslightly thickened (the palatal border is de-fined by a prominent and thick bony ridgein Niviventer); and (8) a thick, bony, andflat posterolateral projection of the bonypalate covering the anteromedial portion ofeach pterygoid plate, its sharp diagonalborder (ppm in fig. 12) defining the anterio-medial edge of the pterygoid fossa (no suchconformation in any species of Niviventer).Other than the larger size of each dentaryin Tonkinomys, we did not detect any signif-icant differences with those of Niviventer inoverall shape; positions of foramina; ex-panse of coronoid, condyloid, and angularprocesses relative to body of the dentary;expression of masseteric and lingual ridging;or position of the posterior terminus of theincisor capsule.
Shapes of the upper and lower incisors inTonkinomys, their enamel color and thickness,and the opisthodont conformation of theuppers resemble the incisors in all species ofNiviventer. The larger size of these structuresin Tonkinomys provides the obvious contrast.Reflecting the much greater skull dimensionsof Tonkinomys when compared with species ofNiviventer are the elongate molar rows of theformer, which are much longer than even thelargest-bodied species of Niviventer (table 2).Occlusal patterns of the upper molars fitwithin the range of variation we observedamong most species of Niviventer, except thatcusps t4 and t6 on the second lamina of thefirst molar in Tonkinomys tend to retain moreof their identity, and cusp t9 is smaller andindistinct relative to the huge central cusp t8.The anterior lamina on the first upper molarin N. andersoni and N. excelsior resemblesthe shape in Leopoldamys, and contrasts withthe comparable chevron-shaped lamina inTonkinomys, but those two species also differfrom most other species of Niviventer in thistrait. Coronal patterns of lower molars inTonkinomys are also closely similar to those inthe species of Niviventer.
While occlusal patterns on upper and lowermolars in our sample of Tonkinomys fallwithin the range of variation of the patternswe observed among the species of Niviventer,the number of roots anchoring most of thosemolars do not—this set of structures bythemselves identifies Tonkinomys as the rep-resentative of a monophyletic group separatefrom Niviventer, a position only strengthenedby the external and cranial contrasts betweenthe two genera already described. Except forthe first upper molar, which has four roots,Tonkinomys exhibits the primitive number ofroots anchoring upper (three per tooth) andlower (two per tooth) molars. All species ofNiviventer, in contrast, have multi-rootedmolars (table 3; also see Chaimanee, 1998,and Zheng, 1993). First upper and lowermolars can be used as examples. Most speciesof Niviventer have five-rooted first uppermolars: large anterior, two smaller lingual,large posterior, and small labial (some speci-mens have one or more labial rootlets inaddition to the primary labial anchor). In N.langbianis, and also in Chiromyscus chiropus,
2006 MUSSER ET AL.: NEW RODENT FROM VIETNAM 25

four roots anchor each molar: large singlelingual, large anterior and posterior roots, andsmall labial. Tonkinomys also possesses fourroots beneath each first upper molar, but thepattern is different from that in N. langbianisand Chiromyscus. No specimen in our sample
of Tonkinomys has a labial anchor, and thefour-rooted pattern is formed by large anteriorand posterior roots along with two prominentlingual roots. A small labial root, not a dividedlingual, forms the fourth root in N. langbianisand Chiromyscus. Although our survey resultsfor second and third upper molars are notsummarized in table 3, we also determinedthat second upper molars in Niviventer havefour roots (three in Tonkinomys), and thirdupper molars have three (as in Tonkinomys,which is the usual number for the third molarin most murines, even when the first andsecond molars have more than three roots).
Four roots (large anterior and posterior,smaller labial and lingual) anchor each firstlower molar in all species of Niviventer (andChiromyscus), but only two are present in theVietnamese limestone rat. Two to four is therange in number of roots anchoring eachsecond lower molar among species ofNiviventer, and each third lower molar haseither two or three roots (two roots beneathsecond and third molars of Tonkinomys).
Saxatilomys: Described from two wholeanimals along with 14 individuals representedby cranial, mandibular, and dental fragmentsrecovered from owl pellets, S. paulinae hasbeen recorded only from the KhammouanLimestone National Biodiversity Conser-vation Area in Lao PDR (Musser et al., 2005).
Body size, build, and proportions of limbs,pinnae, and tail that are characteristic of S.paulinae resemble most species of Niviventer;‘‘S. paulinae simply looks like a Niviventer andnot like any other species in any other genus’’(Musser et al., 2005: 20). That Lao speciesstrikingly contrasts with the large-bodied andshort-tailed Vietnamese animal. Tonkinomysdaovantieni is more robust in build and muchlarger in body size than S. paulinae (range inlength of head and body is 184–217 mm for13 examples of T. daovantieni versus 144–150 mm for two S. paulinae). Its thick, stubbytail is shorter than head and body length(LT/LHB 5 83%, average of 13 specimens)compared with the slim, tapered, and graciletail of S. paulinae that is appreciably longerthan body length (LT/LHB 5 112%–116% fortwo individuals). The tail of both species isbicolored, but the dark brown and whitepattern is different in each. In T. daovantieni,
TABLE 3
Number of Roots on M1 and m1 in Selected Speciesof Niviventer, and the Species of Chiromyscus and
Saxatilomys(Number opposite species indicates specimensexamined. Symbol key: 4/4, M1 with largeanterior, single lingual, and posterior roots, anda small labial root; m1 with large anterior andposterior roots, and small labial and lingual roots.5/4, M1 with large anterior and posterior roots, twolingual, and a small labial root; m1 with largeanterior and posterior roots, and small labial andlingual anchors. 5/2, M1 with large anterior andposterior roots, two lingual, and a small labial root;m1 with a large anterior root and large posteriorroot. Specimens surveyed are in AMNH, BMNH,FMNH, and USNM.)a
Species
Roots (M1/m1)
4/4 5/4 5/2
N. andersoni (China: Sichuan, Yunnan) — 46 —
N. excelsior (China: Sichuan) — 12 —
N. brahma (northern Myanmar) — 7 —
N. eha (northern Myanmar) — 17 —
N. langbianis (Vietnam) 40 — —
N. cremoriventer (Sunda Shelf ) — 53 —
N. hinpoon (Thailand) — 15 —
N. confucianus (China, northern
Myanmar)
— 53 —
N. tenaster (China: Hainan, Vietnam) — 55 —
N. conninga (Taiwan) — 53 —
N. rapit (northern Borneo) — 6 —
N. fraternus (Sumatra) — 20 —
N. lepturus (Java) — 21 —
N. cameroni (Malay Peninsula) — 42 —
C. chiropus (Vietnam) 2 — —
S. paulinae (Lao PDF) — — 11
a Labial roots are divided into two or more rootlets on
some upper and lower molars, but we did not count these
projections as independent roots. In some specimens, the
posterior root on the lower molar is furrowed down the
middle suggesting separate posterolingual and poster-
olabial anchors, but we scored this configuration as one
large root. The sample of S. paulinae consists of two whole
animals and fragments recovered from owl pellets, so the
count is derived from 3 first upper molars and 8 first lower
molars. Sample sizes for all the other species are based on
complete specimens.
26 AMERICAN MUSEUM NOVITATES NO. 3517

the tail is dark brown except for the ventralsurface, which is white, and the distal one-fourth to one-half, which is white on allsurfaces; the tail of S. paulinae is dark brownon dorsal and lateral surfaces from base to tip,and white (with slight speckling) over only itsventral surface. Both species have dense anddark gray fur covering upperparts and under-parts of the head and body. Saxatilomyspaulinae, however, lacks a white foreheadblaze and white pectoral patch, a patternexhibited by most examples of T. daovantieni,and its venter is lightly frosted in contrast tothe total gray of the Vietnamese animal.Tonkinomys and Saxatilomys do share bul-bous plantar pads that are very large relativeto plantar surface and arranged in a tightcluster (compare the plantar surface of T.daovantieni in fig. 8 with that of S. paulinae infig. 9).
Tonkinomys and Saxatilomys possess anelongate skull, wide zygomatic plate relativeto cranial size, narrow alisphenoid strutspresent in most specimens, bony palateprojecting past the third molars to forma broad shelf with either a smooth or slightlythickened (not ridged) posterior margin,strongly divergent maxillary molar rows, anda similar mandibular conformation (comparefigs. 10 and 11 with the views of Saxatilomysin Musser et al., 2005: fig. 6).
The two species differ in size and somequalitative traits. Tonkinomys has a morerobust and larger skull (occipitonasal length,for example, ranges from 47.1 to 52.4 mm inseven T. daovantieni, and 40.3 to 40.5 mm forthe two intact skulls of S. paulinae). Size of thegracile skull in the Lao genus resembles thelarger-bodied species of Niviventer (theIndochinese N. tenaster, for example; comparethe values listed in tables 1 and 4 of Musser etal., 2005, with those presented in table 1).Tonkinomys further contrasts with Saxa-tilomys by having a less tapered rostrum(when viewed from dorsal or ventral aspect),significantly less basicranial flexion (markedflexion in Saxatilomys; see fig. 6 in Musseret al., 2005), and spacious incisive foramina(not the narrow slits of Saxatilomys.
Three other qualitative features providemarked contrasts between the two genera. InTonkinomys, the squamosal root of the zygo-
matic arch is situated high on the side of thecranium where its ridge-like portion runshorizontally just below the posteroventralmargin of the parietal, which projects ventradto form the dorsolateral side of the braincase;the low ridge of the squamosal root disappears10–15 mm anterior to the vertical ridgeformed by the squamosal-exocciptal suture(fig. 11). The configuration is similar to thatcharacteristic of Leopoldamys and Niviventer.The squamosal root of each arch inSaxatilomys also originates high on the sideof the cranium, but is oriented diagonally sothat its posterior ridge-like portion extendsdiagonally to merge with the squamosal-parietal suture (which forms the temporalridge) well anterior to the vertical ridgecreated by the squamosal-exoccipital suture(see figs. 6 and 8 in Musser et al., 2005). Theparietal does not drop below the temporalridge, and the upper portion of the braincaseconsists only of the squamosal.
Tonkinomys daovantieni and S. paulinaehave a wide and long bony palate that projectsbeyond the molar rows to form a broadplatform. In Tonkinomys, the posterolateraledges of this shelf extends as a flat, bonysurface onto the anterior portion of eachpterygoid plate where its sharp diagonalborder (ppm in fig. 12) marks the anterome-dial margin of each pterygoid fossa. No suchconformation exists in S. paulinae (or in anyspecies of Niviventer).
The globular and only slightly inflatedectotympanic (auditory) bulla of Tonkinomysdoes not conceal all of the periotic. A wide,wedge-shaped segment of that bone is exposedbetween the dorsal capsular margin andbasioccipital, and this wedge of perioticextends anteriorly to form the dorsal andslanting posterodorsal wall of the carotidcanal (fig. 12). The conformation is primitiveamong murines and closely resembles themorphology depicted by Carleton andMusser (1989: 33) for the sigmodontine,Oligoryzomys, and by Musser and Heaney(1992: 99) for some of the Philippine OldEndemic murines. By contrast, the globularectotympanic capsule exhibited by Saxa-tilomys covers all but a narrow wedge of theperiotic, and the anteromedial margin of thecapsule meets the basioccipital, so that it is the
2006 MUSSER ET AL.: NEW RODENT FROM VIETNAM 27

ectotympanic (or auditory capsule) that formsthe posterior wall of the opening into thecarotid canal. This structural relationshipbetween ectotympanic and periotic is special-ized, and is particularly well illustrated inCarleton and Musser (1989: 33) for thesigmodontine Oryzomys palustris. Species ofRattus and its allies have a comparable pattern(Musser and Heaney, 1992: 99).
Primary dental differences between Ton-kinomys and Saxatilomys involve size, oneproportional feature, and pattern of molarroots. Tonkinomys has larger upper and lowerincisors than does Saxatilomys, but otherwisepigmentation and thickness of enamel, theirshapes, and opisthodont configuration of theuppers relative to the rostrum are similar.Tonkinomys also has longer molar rows (asindexed by lengths of maxillary rows: 8.0–8.8 mm in 7 specimens of Tonkinomys, 7.2–7.3 mm for three Saxatilomys); length ofmaxillary row relative to occipitonasal lengthfor the two genera, however, is, on average,not significantly different (the ratio, CLM1-3/ONL, using mean values from each genericsample, is 17% for Tonkinomys and forSaxatilomys). In contrast to molar length,the individual molars in Tonkinomys are notsignificantly wider than are the molars ofSaxatilomys (using breadth of the first uppermolar as a guide, the mean is 2.1 mm for 7examples of Tonkinomys and for 3 specimensof Saxatilomys; contrast the values listedin table 1 with those figs. in table 4 inMusser et al., 2005: 21), indicating significant-ly narrower molars relative to length of themaxillary row (the ratio, BM1/CLM1-3, usingmean values from each generic sample, is 25%for Tonkinomys and 29% for Saxatilomys).Two roots (large anterior and posterior)anchor each lower molar in all specimensexamined of Tonkinomys and Saxatilomys,but the root patterns for the first andsecond maxillary molars are dissimilar inthe two genera. In Tonkinomys, the firstmaxillary molar has four roots (anterior,posterior, and divided lingual), the secondhas three. Saxatilomys has five-rooted firstmolars (table 3) and four-rooted second mo-lars; in samples of both Tonkinomys andSaxatilomys, three roots anchor each thirdmolar.
Tonkinomys and Saxatilomys (along withmost species of Niviventer) share similarmaxillary and mandibular molar occlusalpatterns. Distinctions between the Viet-namese and Lao limestone rats are presentbut not spectacular (contrast fig. 14 with theocclusal views of Saxatilomys in Musser et al.,2005: fig. 10): compared with Saxatilomys,cusps t4 and t6 on the second lamina of thefirst maxillary molar in Tonkinomys tend toretain more of their identity (less distinct inSaxatilomys and in most species ofNiviventer), second and third mandibularmolars lack anterolabial cusps in most speci-mens (most species of Niviventer also lackthese cusps, but they are present inSaxatilomys), and the cusps on upper andlower molars form more distinct and tighterchevrons (cusp rows more arcuate, withgreater space between arms of each chevronin Saxatilomys).
Other genera: We also compared examplesof Dacnomys and Srilankamys, otherIndomalayan members of the DacnomysDivision, with our sample of Tonkinomys.Dacnomys is a giant version of Niviventer,especially N. andersoni, in its external andcranial traits, as well as root patterns of themolars (see description and illustrations inMusser, 1981). Those molars, however, arelarge, chunky, and hypsodont, with compli-cated cuspidate occlusal patterns, and strik-ingly contrast with the brachydont and gracilemolars with their simple occlusal patternspossessed by all species of Niviventer.Tonkinomys bears no close resemblance toDacnomys in external, cranial, or dental traits.
A combination of traits, some of whichrecall features in Niviventer, others that re-semble Maxomys, and some that are unique,characterizes the morphological definition ofSrilankamys (see the description and illustra-tions in Musser, 1981). Its membership in theDacnomys Division is provisional, pendingreassessment of its phylogenetic relationshipsby additional morphological characters incombination with molecular data (Musserand Carleton, 2005). Srilankamys shares thesame pattern of molar roots with Tonkinomys,but otherwise its diagnostic external andcranial traits, along with molar occlusalpatterns, are unlike those of Tonkinomys.
28 AMERICAN MUSEUM NOVITATES NO. 3517

Musser and Carleton (2005) included thePhilippine Mindoran endmic Anonymomysmindorensis in the Dacnomys Division. Itsplacement there, however, is very provisional.Some morphological traits of Anonymomysrecall those of genera in the DacnomysDivision, particularly Niviventer, but otherssuggest a closer affinity to Bornean andSulawesian Haeromys, a member of theMicromys Division of Musser and Carleton(2005). Tonkinomys is morphologically distantfrom Anonymomys and requires no compar-isons with it.
Our comparative study also included theIndomalayan Maxomys, although that genusis not contained within the Dacnomys Group(Musser and Carleton, 2005). Species ofMaxomys are prominent members of theIndochinese murine fauna (Corbet and Hill,1992), and some share with Tonkinomys thesame number of roots anchoring each molar(Musser et al., 1979). Any close similarity endswith that character complex. No species ofMaxomys has the pelage coloration possessedby Tonkinomys, or the thick, short tail, or thecrowded, fleshy palmar and plantar pads.Additionally, all Maxomys have short, wideincisive foramina that have the outline of aninverted heart, a bony palate ending wellanterior to posterior margins of the molarrows, a very long postpalatal region, and othercranial attributes, as well as molar occlusalpatterns (see descriptions and illustrations inMusser, 1991) that are not shared withTonkinomys. We found no derived charactersthat identified Tonkinomys as either a memberor close relative of Maxomys.
Fossils: Reports covering murines recoveredfrom late Pliocene to Pleistocene sediments incaves and limestone fissures in Thailandand China contain descriptions of samplesidentified as species of Leopoldamys, Nivi-venter, and related extinct genera. The bulkof the material is comprised of isolatedmolars; a few samples contain cranial andmandibular fragments. Only some of thepublications contain information pertinent toour inquiry.
In her detailed and comprehensive treatiseon the Plio-Pleistocene rodents of Thailand,Chaimanee (1998) described samples repre-senting extant species of Leopoldamys (L.
sabanus) and Niviventer (N. fulvescens) andnew species in those genera that are extinctand represented only by fossils (L. minutusand N. gracilis). Judged from her descriptions,measurements, and illustrations, all thesesamples are correctly allocated to genus; nonerepresents Tonkinomys.
Pleistocene samples from Guangxi Provincein southern China were determined by Chen etal. (2002) to contain examples of Niviventerand Leopoldamys, but no identifications tospecies were provided. Zheng et al. (1997)reported the extant Niviventer confucianus andextinct N. preconfucianus from Plio-Pleistocenefissure fillings in Shandong Province. Mea-surements and drawings of molar occlusalpatterns presented in these reports convinceus their generic attributions are correct.
Zheng (1993) has provided perhaps themost comprehensive and thorough survey offossil murines recovered from late Plioceneand Pleistocene cave and fissure fillings. Allcome from the northern margin of theYunnan-Guizhou Plateau within an areabounded by 104u to 110uE and 26u to 31uNin the provinces of Sichuan and Guizhou. Byclear descriptions and a wealth of measure-ments, Zheng meticulously documented thoseparts of the samples representing extantLeopoldamys edwardsi and extinct L. edward-sioides; extant Niviventer fulvescens, N. con-fucianus, N. andersoni, and possibly N. excel-sior; and the extinct N. preconfucianus. Hisdescriptive and quantitative data, along withclear illustrations, provide unquestionablesupport for his identifications.
Zheng (1993) also described three newspecies in two new genera that seem morpho-logically similar to Niviventer and othermembers of the Dacnomys Division. Wusha-nomys brachyodus is represented by a largecranial fragment containing intact molar rows,dentary fragments with complete or partialmolar rows, and numerous isolated upper andlower molars. The fossils come from sedimentsthought to be no older than 2.5 million yearsago, and no younger than 1.0 million yearsago. Judged by length of upper (9.30 mm, 1specimen) and lower (8.80–9.20 mm, 5 speci-mens) molar rows, this extinct species is muchlarger than Tonkinomys daovantieni (uppers,8.03–8.78 mm, 7 specimens; lowers, 7.64–
2006 MUSSER ET AL.: NEW RODENT FROM VIETNAM 29

8.52 mm, 10 specimens). Range in measure-ments of first upper (4.00–4.90 mm by 2.14–2.66 mm, 85 examples) and lower (3.40–3.95 mm by 1.90–2.20 mm, 97 specimens)molars of W. brachyodus overlaps with thoseof T. daovantieni (table 4), but average longerand wider (see Zheng, 1993: 190). Most of thefirst upper molars in W. brachyodus have threeroots, but Zheng noted that some molars havea divided lingual (see Zheng, 2004: 175), whichis the pattern in T. daovantieni. First lowermolars have two roots, the primitive patternthat is also typical of the Vietnamese limestonerat.
Molar occlusal patterns of Wushanomys areunlike those of Tonkinomys. The anteriorlamina on the first upper molar inWushanomys consists of a straight segmentformed by the complete fusion of cusps t2 andt3, and a posteriorly offset cusp t1; the secondlamina is arcuate in outline; and cusp t9 isdistinct in the posterior part of the tooth (seethe illustration in Zheng, 1993: 192). Theseocclusal outlines are strongly dissimilar tothe strong, tight chevrons formed by the twofront laminae on the first upper molar inTonkinomys (fig. 14) and its cusp t9, socoalesced with the large central cusp t8 thatit resembles a low ridge. Laminar outlines onlower molars of Wushanomys are also nearlystraight or form wider-armed chevrons, a con-trast with the tight chevrons in Tonkinomys.
The cranial and mandibular fragments ofW. brachyodus provide additional compara-tive information. Zheng did not illustrate thespecimen, but described it as having very longincisive foramina that terminate 1.7 mm pos-terior to anterior faces of first molars, a bonypalate extending about 0.7 mm beyond themolar rows to form a short platform, a verynarrow interorbit (6 mm), especially for whatseems to be a large skull judging from lengthsof molar rows, and a flat, wide zygomaticplate (no measurement listed). Zheng didinclude a drawing of a partial dentary (thebody of the ramus behind the molar row isgone, p. 204). The portion anterior to themolar row, along with the incisor, is long andslender, and the dorsal outline from anteriormargin of the molar to incisor tip is gentlyconcave. This conformation usually accom-panies a long rostrum, and the mandibular
configuration recalls that typical of the long-nosed Sulawesian murines, Bunomys prolatusand B. chrysocomus (Musser, 1991: 12–13). InTonkinomys, posterior margins of the incisiveforamina end just in front of anterior faces ofthe first molars, even with them, or extendonly slightly beyond (no more than 0.5 mm);the bony palate extends 1.5–2.0 mm beyondthe molar rows forming a much more expan-sive platform, and the interorbit is wide (6.6–7.3 mm; table 1), which is the usual pro-portion in skulls the size of Tonkinomys (andother murines, Leopoldamys neilli, for exam-ple). The bony projection containing the lowerincisor in the dentary of Tonkinomys is shorterand thicker, and the dorsal outline from molarrow to incisor tip is more deeply concave,a shape common to species of Niviventer,Leopoldamys, and other murines with lessspecialized dentaries (Musser, 1981: 272;Zheng, 1993: 204).
Zheng’s description of the cranial traits inWushanomys brachyodus tantalizingly resem-bles the conformation in other extinct mur-ines, such as the Canary Islands Malpaisomysinsularis, which was likely adapted to livingand climbing about in the creviscular systemof lava fields (Boye et al., 1992; Hutterer et al.,1988). Perhaps W. brachyodus lived andclimbed about in karstic fissures, had a longrostrum, and included invertebrates in its diet.Whatever its habitat and habits, the summa-tion of morphological traits characterizing thecomplement of fossil remains identifies, inour view, a monophyletic group (at the levelof genus) separate from that representedby Tonkinomys daovantieni; other thanWushanomys likely being a member of theDacnomys Group, there seems no closerphylogenetic link between Wushanomys andTonkinomys.
Zheng (1993) described a second species ofWushanomys, W. hypsodontus, based on a fewisolated molars that are high-crowned, haveslightly more complex occlusal surfaces, andgreater dimensions than found in W. brachyo-dus. The time span in which the fossils lie is 2.0to 1.8 million years ago.
Zheng’s other new genus and species,Qianomys wui, is based on a small series ofdentary fragments containing full or partialmolar rows, an isolated first lower molar, and
30 AMERICAN MUSEUM NOVITATES NO. 3517

two upper molars. These were recovered fromsediments thought to be late Pleistocene,400,000 to about 200,000 years ago. The twoupper molars (4.40 mm and 4.60 mm long)fall within the high end and slightly beyondthe range of variation in molar length werecorded for Tonkinomys daovantieni, but theyare much wider (2.35 mm and 2.40 mm) thanthe molars of T. daovantieni (table 4). Theocclusal pattern does not match that ofTonkinomys, and five roots are present. Theuppers are not examples of the Vietnameselimestone rat. Even Zheng noted their closesimilarity to Niviventer in crown outline andnumber of roots, and suggested they may notbe correctly associated with the lower molars,to which the generic and specific name isnomenclaturally attached. We agree. For us,Zheng’s diagram (p. 199) of a first uppermolar closely resembles N. andersoni inocclusal pattern; however, the size is slightlyoutside the range of variation we obtainedfrom our sample of that species (table 4) andthat which Zheng derived from his material ofN. andersoni (p. 181).
The three intact lower molar rows ofQianomys for which measurements are pro-vided (8.60, 9.40, and 9.43 mm) are apprecia-bly longer than those in Tonkinomys (7.64–8.52 mm, 10 specimens) as are length andwidth dimensions of the first lower molars(3.66–4.20 mm by 2.10–2.34 mm, 8 specimensof Qianomys; 3.40–3.50 mm by 1.90–2.00 mmin Tonkinomys, table 4). If molar dimensionsare any indication of skull and body size, Q.wui was a much larger rat than T. daovantieni.Qianomys and Tonkinomys share the sameprimitive root pattern, two beneath each lowermolar. Occlusal patterns formed by the rowsof laminae fall within the range of patternsexhibited by Tonkinomys as well as species ofNiviventer. Zheng, however, claimed that noneof the molars he assigned to Qianomys haveanterolabial cusps on the second and thirdmolars or posterolabial cusplets on any molar,features, in addition to number of roots, whichhe used to distinguish Qianomys fromNiviventer. Comparable cusps do occur onthe lower molars of T. daovantieni. Anillustration of a partial dentary (Zheng, 1993:204) indicates the portion anterior to themolar row to be similar in shape to that of
Tonkinomys, Niviventer, Leopoldamys, andSaxatilomys.
Qianomys wui is certainly a different speciesthan Tonkinomys daovantieni. Whether thegross similarities between the two in molarocclusal patterns signal close phylogeneticrelationship or convergence cannot be de-termined until we can compare the fossilsdirectly with our extant samples, and untilcranial material representing Qianomys isrecovered and described. Resolving the am-biguous association between the present sam-ple of upper molars and lowers allocated toQianomys is also critical. That genus wouldresemble only Saxatilomys among members ofthe Dacnomys Division if the five-rooted firstupper molars with their Niviventer-like occlu-sal patterns actually belong to Qianomys withits two-rooted lower molars.
A recent report by Zheng (2004) documentsidentities of rodents, represented mostly byisolated molars, collected from sedimentarylayers in Longgudong Cave (30u39914.90N/110u04929.10E), the ‘‘Jianshi Homind Site’’ inwestern Hubei Province of central China.Geological age was given as Early Pleisto-cene, but correlation between the 11 sedimen-tary layers sampled and the geomagneticpolarity timescale indicated a range extendingfrom the Olduvai Event of the MatuyamaChron (the Pleistocene/Pliocene boundary)back to earlier than 2.15 million years beforethe present, reflecting initial deposition duringthe Late Pliocene (see Flynn et al., 1997, andQiu, 1989, for illuminating expositions ofChinese geochronology at the end of theNeogene). Among the murines recovered werethree species in the Dacnomys Division. One isthe extinct Niviventer preconfucianus, whichZheng himself had named and described in1993, and another is represented by isolatedmolars identified only as an undeterminedspecies of Leopoldamys. The third is a newspecies of the extinct Wushanomys, W. ultimus,described from molars recovered from all butthe top two layers of the cave sediments, mostfrom layers dated between 1.95 to earlier than2.15 million years before the present (LatePliocene). Molars from W. ultimus are in-termediate in size between the earlier describedW. brachyodus and W. hypsodontus (Zheng,1993), express greater hypsodonty, and differ
2006 MUSSER ET AL.: NEW RODENT FROM VIETNAM 31

slightly in coronal patterns. Ranges in lengthand breadth values for first upper (3.90–4.92 mm by 2.20–2.65 mm, 22 examples) andlower (2.90–4.30 mm by 1.98–2.30 mm, 18specimens) molars of W. ultimus (see Zheng,2004: 173) overlap with those of Tonkinomysdaovantieni (table 4), but average slightlylonger and wider. Each first lower molar ofW. ultimus has two roots, the pattern commonto the other two species of Wushanomys, butfirst upper molars have either four (anterior,divided lingual, posterior) or five roots (asmall labial holdfast or nubbin in addition tothe four primary roots). The pattern of thefour primary roots beneath the first uppermolars and two anchoring first lower molarsalso characterizes Tonkinomys daovantieni, butmolars of that Vietnamese rat are not hypso-dont, and although the extinct and extantspecies share molars similar in size, theirocclusal patterns are dissimilar. The occlusalcusp patterns characteristic of W. ultimusresemble, except for minor details, those inW. brachyodus and differ from T. daovantieniin the configurations we previously describedcontrasting W. brachyodus and the Vietnamesespecies.
These new samples of the DacnomysDivision recovered from the Jianshi HomindSite provide additional information on LatePliocene geographic distributions of Niviventerand Leopoldamys. The geographic range ofWushanomys is also expanded as is thediversity of species in that genus. Judgedfrom Zheng’s (2004) descriptions, measure-ments, and illustrations, none of the identifiedfossil molars represent the VietnameseTonkinomys.
Our final survey of fossils assesses theidentification of murines recovered fromPleistocene sediments in Ma U’Oi cave innorthern Vietnam, south of Hanoi (Bacon etal., 2004). One mandible containing a completemolar row was identified as Niviventer fulves-cens; an upper first molar and a lower firstmolar were attributed to N. andersoni; anda dentary containing all molars, and isolatedfirst and second molars, were determined asLeopoldamys sabanus. The molars were iden-tified by comparing them with Chaimanee’s(1998) treatise on fossil Plio-Pleistocene Thairodents. Unfortunately, we are not treated to
drawings of molar occlusal cusp patterns, orto measurement values for the sample of N.fulvescens.
We cannot verify the identification of N.fulvescens. That species is common in northernVietnam and in many places occurs togetherwith N. tenaster (results of our trappingsurveys in several different regions of northernVietnam), which is somewhat larger in dimen-sions of body and skull, as well as length ofmolar row and breadths of molars (Musserand Lunde, ms.). Niviventer tenaster alsooccurs in northern Thailand (Musser andCarleton, 2005), but Chaimanee (1998) didnot include it in her report. The materialreported by Bacon et al. (2004) should bereexamined to determine whether it is really N.fulvescens or possibly N. tenaster.
Measurements listed by Bacon et al. (2004)for two first lower molars of Leopoldamys (4.0by 2.6 mm and 4.7 by 2.8 mm) fall within therange of variation for the larger species ofthat genus (Chaimanee, 1998; Zheng, 1993),and well outside the range for our sampleof Tonkinomys (table 4). The lower secondand third molars that were identified as Leo-poldamys are also large and likely representthat genus. But these fossil molars may notbelong to L. sabanus. Leopoldamys requirestaxonomic revision because more species existthan are usually recognized (four were listedby Musser and Carleton, 1993, and six byMusser and Carleton, 2005, which still under-estimates the number of species), and thesituation in northern Vietnam is especiallycomplex. At least two species occur there. Onemay be L. edwardsi, the other is usuallyidentified as L. sabanus, but is actuallya different species (Gorog et al., 2004;Musser and Carleton, 2005).
An upper first molar and lower first molarwere identified as Niviventer andersoni inBacon et al. (2004). Ma U’Oi cave is far southof the present geographic distribution of N.andersoni, which extends from eastern Xizang(Tibet) through northern Yunnan, westernSichuan, and northern Ghizhou to southernShaanxi, and over an altitudinal range from6000 to 10,000 ft (Musser and Chiu, 1979;Musser and Carleton, 1993, 2005). We soughtto determine whether these two fossil molarsbelong to Tonkinomys. Dimensions of the first
32 AMERICAN MUSEUM NOVITATES NO. 3517

upper molar (3.7 by 2.0 mm) do fall within therange of variation of our values (table 4) andthose tabulated by Zheng (1993: 181) for largesample sizes of N. andersoni. Breadth of thefossil molar fits within our observed range forfirst upper molars of Tonkinomys, but thelength is much too short—based on thesedata, the molar did not come from theVietnamese limestone rat. However, it maynot represent N. andersoni. The width, forexample, matches our range of values for N.excelsior, and the length is only 0.1 mmshorter (table 4). Niviventer excelsior is a closerelative of N. andersoni that presently occursonly in high mountains north of the YangtzeRiver (Musser and Chiu, 1979). For N.excelsior to have once occurred in northernVietnam, however, seems as implausible to usas the presence of N. andersoni there, espe-cially because all the fossils of other mammalsfound in situ in the Ma U’Oi cave define ‘‘…a relatively modern fauna …’’ (Bacon et al.,2004: 312). Niviventer tenaster currently livesin northern Vietnam in forest habitats.Lengths and breadths of first upper molarsfrom this species in our sample from thatcountry nearly embrace dimensions of thefossil molar (table 4), and that tooth justmight be from a large individual of N.tenaster. Both N. andersoni and N. excelsiorcan easily be distinguished from N. tenaster byocclusal patterns. In the former pair, theanterior lamina on the first upper molar
consists of a nearly straight segment, formedby the coalesence of cusps t2 and t3, anda posteriorly displaced lingual cusp t1 (see figs.in Musser, 1981, and Zheng, 1993). Thecomparable lamina has an arcuate outline onthe upper molar in N. tenaster, similar to thatfound in N. confucianus (Musser, 1981). Thefossil molar needs to be reexamined; it is notan example of Tonkinomys, and we questionits identity as N. andersoni.
The first lower molar determined to be N.andersoni in Bacon et al. (2004) is large (4.1 by2.5 mm), and its dimensions fall far outsidethe range of variation in our samples ofTonkinomys, Niviventer andersoni, N. excelsi-or, and N. tenaster (table 4). The tooth isactually about the same size as the two firstlower molars (4.0 by 2.6 mm, 4.7 by 2.8 mm)they identified as Leopoldamys. The valuesare either misprints, or the molar doesbelong to Leopoldamys; it, and the first uppermolar assigned to N. andersoni, warrantsrestudy.
Summary: There would have been anelegant symmetry in discovering ancient,fossilized remnants hidden in limestone fis-sures that represented the same murine speciesliving today in creviscular habitats within thetower karst landscape forming northernVietnam. Unfortunately, we cannot unambig-uously tie any published identifications offossilized fragments representing Indochinesemurines to the extant Tonkinomys daovantieni.
TABLE 4
Summary Statistics for Lengths and Breadths (mm) of First Upper and Lower Molars of Niviventer andersoni(China, Yunnan and Sichuan), N. excelsior (China, Sichuan), N. tenaster (Vietnam), Pleistocene First Molars
Identified as N. andersoni by Bacon et al. (2004), and Tonkinomys daovantieni (Vietnam)(Mean, plus or minus one SD, and observed range in parentheses are listed for each measurement. Samples
consist entirely of AMNH specimens except those reported by Bacon et al., 2004.)
N. andersoni N. excelsior N. tenaster Bacon et al. T. daovantieni
(N 5 45) (N 5 13) (N 5 16) (N 5 1) (N 5 7)
Length of M1 3.7 6 0.1 3.3 6 0.1 3.2 6 0.1 3.7 4.2 6 0.1
(3.5–4.0) (3.1–3.6) (3.0–3.6) (4.1–4.5)
Breadth of M1 2.1 6 0.1 1.9 6 0.1 1.7 6 0.1 2.0 2.1 6 0.1
(2.0–2.2) (1.8–2.0) (1.6–1.8) (2.0–2.2)
Length of m1 3.3 6 0.1 3.0 6 0.1 2.8 6 0.1 4.1 3.5 6 0.1
(3.1–3.5) (2.8–3.2) (2.7–3.1) (3.4–3.5)
Breadth of m1 1.8 6 0.1 1.7 6 0.1 1.6 6 0.1 2.5 2.0 6 0.1
(1.7–1.9) (1.6–1.9) (1.5–1.7) (1.9–2.0)
2006 MUSSER ET AL.: NEW RODENT FROM VIETNAM 33

Its past distribution and phylogenetic allianceto extinct taxa, as might have been illuminatedby fossils, remain a mystery.
Unresolved also is the phylogenetic positionof Tonkinomys with respect to Leopoldamys,Niviventer, and Saxatilomys, the three extantgenera with morphologies most like that ofTonkinomys. The differences enumeratedabove between Tonkinomys daovantieni onone hand and species of Leopoldamys andNiviventer on the other reflect a combinationof primitive and derived features. Relative tothe pelage coloration, shape of tail, and itslength relative to length of head and body inLeopoldamys and Niviventer, the contrastingcondition in Tonkinomys is derived. Alsorepresenting specializations in Tonkinomysare the position of the squamosal zygomaticroots high on the sides of the braincase,slender or no alisphenoid struts, its longincisive foramina, a very long bony palatethat projects beyond the molar rows to forma broad shelf, posteriorly diverging maxillarymolar rows, simple molar occlusal patterns,and a divided lingual root beneath the firstupper molar. Presence of three roots beneatheach second and third molar, and two rootsanchoring each lower molar represent theprimitive condition in Tonkinomys comparedwith the derived state in Leopoldamys andNiviventer. Tonkinomys daovantieni resemblesthe Indochinese Leopoldamys neilli in bodysize, and it along with the other Indomalayanspecies of Leopoldamys in general cranialconformation and a few cranial particulars(position of the squamosal zygomatic roots,for example); with Niviventer, Tonkinomysshares long incisive foramina, comparableinflation of the auditory bulla relative to theunderlying periotic bone, and similar molarocclusal patterns. Other traits exclude T.daovantieni from each of those genera asthey are currently diagnosed (Musser, 1981).No species of Leopoldamys or Niviventer (orChiromyscus), whether occurring in Indochinaor on the islands on the Sunda Shelf, hasTonkinomys’s combination of dark grayishblack upperparts and dark gray underparts;thick tail with a stubby tip that is muchshorter than length of head and body; large,bulbous, and contiguous plantar pads; bonypalate projecting well beyond the third molars
to form a shelf with a smooth posteriormargin; strongly diverging (along the ante-roposterior axis) maxillary molar rows; de-rived root pattern for the first upper molarcoupled with the primitive root patternsunderlying the other uppers and all the lowermolars.
Both Tonkinomys daovantieni and Saxa-tilomys paulinae are associated with forestedkarstic habitats. Both genera display darkgray pelage, large and swollen plantar tuber-cles, a bony palate that projects beyond themolar rows to form a broad platform,maxillary molar rows that appreciably divergeposteriorly (along the anterior-posterior axis)and are long relative to length of skull, andtwo-rooted mandibular molars. All but theroot pattern are specialized traits. This com-bination of attributes is not combined in anyspecies of either Leopoldamys or Niviventer (orChiromyscus). We do not know whether themorphological specializations shared by T.daovantieni and S. paulinae phylogeneticallylink these species closer to each other than tospecies of Leopoldamys and Niviventer, orrather reflect independent evolutionary adap-tations to forested karstic environments in theIndochinese tropics. In opposition to theshared specializations, external morphologyof Tonkinomys (except for relative length oftail) recalls a small-bodied Leopoldamys, whilethat of Saxatilomys resembles most speciesof Niviventer. Overall cranial form inTonkinomys also resembles Leopoldamys, andexcept for those cranial traits that distinguishS. paulinae from species of Niviventer (com-parisons are enumerated by Musser et al.,2005), the general cranial conformation ofSaxatilomys is not unlike most species ofNiviventer. Inferring the phylogenetic relation-ship of Tonkinomys among species now allo-cated to the Dacnomys Division in general, andespecially to species of Leopldamys, Niviventer(including Chiromyscus), and Saxatilomys willrest on results derived from future phylogeneticanalyses of information derived from bothmorphological and molecular sources.
HABITAT AND HABITS
The Dao village of Lan Dat lies withina spectacular tower karst landscape, its houses
34 AMERICAN MUSEUM NOVITATES NO. 3517

and adjoining agricultural clearings scatteredover flat depressions nestled among highlimestone hills with steep sides and sheer,expansive cliffs. The area epitomizes theclassic description of karst landforms:‘‘Karstic terrain is commonly characterizedby closed depressions, subterranean drainageand caves’’ (Gillieson, 1996: 59). The climate ismonsoonal, with most of the year’s rainconcentrated from April to October. Averageannual rainfall (measured at Lang Son over50 years) is 1395 mm, and even though coldtemperatures dominate from Novemberthrough March, humidity remains high, theannual relative humidity averaging 81% (Ðieu,1995).
Most of the flat areas, once supportingtropical moist deciduous forest, have beencleared by the Dao for rice cultivation andshifting agriculture. Forest remains at thebase of the cliffs, in ravines between the hightowers, and on the steep slopes, and it is notunlike Whitmore’s (1984: 174) description:‘‘… limestone slopes have a dense, irregularforest with trees clinging precariously, theirroots penetrating to great depths in crevices…’’ The forest consists of a low canopy (upto 15 m high) with taller (up to 25 m ormore) emergents scattered over the steepslopes. Most trees are small (1–3 ft in di-ameter near the base), except in placesallowing accumulation of sufficient soilwhere huge emergents are able to grow.Here and there a grove of emergent speciesforms a small area of high canopy. Palmsand bamboo clumps are scattered throughthe understory, strangler figs are uncommon,and woody vines drape over high limestoneledges and form tangles beneath the forestcanopy. The limestone towers are denselyforested except for the sheer cliffs. Clumps ofPandanus are common at the base of thesewalls, and sometimes grow higher where theycan obtain purchase in joints and fissures.We did not encounter cycads in the forest,and saw them only high on the vertical cliffs.Isolated clumps of bamboo remain in someof the clearings, and dense bamboo forestcovers the steep slopes and tops of somelimestone towers.
Most of the hillsides support primary forest,old and tall secondary growth covers hillsides
close to houses and fields, and scrub habitatsdominate clearings not utilized for crops(fig. 1). Forest once growing on the few flator gently sloping places above the valley andbetween limestone towers has been eitherremoved or thinned. The primary forest coveris also not undisturbed by human activity.Trails cut through the understory usually leadto remnants of a huge emergent tree felled andprocessed for lumber. Natural disturbance inthe form of treefalls and landslips is eventuallycovered by shrubs and then taller second-growth.
The day we broke camp and climbed out ofthe valley our usual trail chatter was silencedby a loud rumble, which had been generatedby a massive rockslide. This is a periodic,unpredictable event, according to the villagers,and potentially dangerous. Weathering ofthe high cliff faces through the combinedforces of water dissolving the limestone,differences in temperature, and pressure fromlarge tree roots eventually dislodge chunksof cliff wall, some small, others huge. Suchan event formed the habitat in which wetrapped most of the limestone rats. Abovethe valley floor, a wide ravine between twocontiguous limestone towers had been filledby a chunk of cliff wall the size of a smallhouse along with smaller pieces—basicallya talus of large blocks between the towers.The openings among the pieces were eventu-ally enlarged, mostly by water flowing intothe rubble pile from the top, and under-neath it along the bottom of the old ravine.The result is a honeycomb of passageways,some extending vertically to small openingson top of the rubble, others opening to anuncovered section of the ravine. Walls ofthe labyrinth are wet and the air is cool.Shrubs, vines, and moss cover the surface ofthe rubble, hill forest persists at the cliffbases of each tower, and small trees arescattered over the talus (fig. 2). The surfaceof the rubble is difficult to negotiate. Rainhas sculpted the limestone into small depres-sions surrounded by knife-sharp edges; smallto large openings are everywhere, some asdeep as 8 meters that expand into largecavities; and some openings are concealedby loose limestone fragments that shift un-derfoot.
2006 MUSSER ET AL.: NEW RODENT FROM VIETNAM 35

Thirteen of the T. daovantieni were caughtin this talus. Two were trapped on the surface,one beneath a shrub, the other one at themargin of the rubble beneath a rotting trunk.Five were taken in traps set at the bottom ofthe ravine beneath the main overhanginglimestone chunk (the configuration resembleda small cave, but was really only a low passagebeneath the straddling rock). Another wascaptured on a moss-covered ledge just outsideof this cave-like passage. Five others werecaught in traps lowered by ropes throughsmall and large holes in the talus surface to thebottom of the labyrinth, which ranged from 4to 8 m, depending on how close the verticalpassages were to sides of the ravine. In thesesituations, most live and snap-traps wereplaced directly on the passageway floors.Traps placed on limestone ledges above thisrubble near bases of the towers, or in the denseforest covering steep slopes away from thetalus, yielded only Niviventer fulvescens and N.langbianis.
Tonkinomys daovantieni was not the onlyinhabitant of this talus. We captured a Rattusandamanensis and a Niviventer fulvescens atthe mouth of the cave-like passage, an N.fulvescens on the surface, and one at thebottom of the deepest chamber.
The only limestone rat not trapped in or onthe talus was taken in mature second growthhillside forest behind camp, which is at the otherend of the valley from the rubble where all theothers were caught. Here between the massivecliffs and valley floor is a steep middle region oflimestone talus, formed of small to massivechunks that broke away from the cliff above.This is the only talus-like structure we encoun-tered here. While soil has filled in most of thespaces among the blocks, apparently enoughinterstices and underground extensions exist toprovide the proper habitat for T. daovantieni.Either that or the animal wandered from placeson the tower we did not survey. The forestcovering the talus is dense and festooned withwoody vines. Niviventer fulvescens and thearboreal N. langbianis were common.
Populations of T. daovantieni apparentlyhave a patchy distributional pattern. Despiteour sampling efforts on other hillsides, wheretraps were placed beneath rocky outcrops, onledges, at the bottom of deep fissures in cliffs,
and on the forest floor, we did not encounterany T. daovantieni. A large cave was accessi-ble, but it was being used as a corral for cattle;the interior was dry, and mostly scrub andyoung secondary forest persisted near theentrance. Traps placed on ledges inside thecave and outside near the entrance yieldednothing. We also set traps in a small, wet caveused as a roosting site by bats. No limestonerats were caught. Caves occur throughout thearea, but the openings to most are high in thecliffs and inaccessible to us.
Certainly more field study is required toassess the habitat requirements of T. daovan-tieni, but our trapping experience suggests thisforest species to be petricolous. It seems torequire large areas of rock debris or talus thatprovides sufficient passageways and chambersfor protected den sites, and a cool, dampenvironment. Long vibrissae; large, bulbouspalmar and plantar pads; and a tail that isconsiderably shorter than head and body, thatis thick and stubby-tipped, and that has a well-haired distal half and tip are adaptationsimportant in a creviscular environment. Thevibrissae monitor the cracks and walls forprey, surface information, and passage width;the fleshy pads provide purchase on rocksurfaces; and the tail is used for balance asthe rat scurries up and over the rocks andthrough the crevices and passageways, or asa fifth appendage when pressed against thelimestone (the hairs would aid in purchase bypressing against the rough surface). We seesimilar adaptations in other petricolous mur-oid rodents. The bushy-tailed woodrat,Neotoma cinerea, for example, is a NorthAmerican cricetid analog to the Vietnamesemurid, T. daovantieni. This species of Neotomadens in caves, fissures in high cliffs, and deepin talus slopes (Finley, 1958; Grayson, 1999;Musser’s trapping experience). Size and flesh-iness of its footpads relative to foot size,length of tail relative to head and bodylength, and even overall body size is closelysimilar to these qualities in the limestone rat(see measurements provided by Finley, 1958:322–325).
A cool and damp environment of crevicesand larger passageways may be important tothe diet of T. daovantieni because the diversityand numbers of arthropods is probably higher
36 AMERICAN MUSEUM NOVITATES NO. 3517

there than in dry situations. Of two limestonerats kept captive, one devoured two kinds ofcockroaches: a large-bodied species caught atthe talus, and a smaller species collected invillage houses and at camp. This rat refusedoffers of corn, manioc, snails, earthworms,grasshoppers, beetles, and small frogs. Ourother captive rejected cockroaches offeredduring daytime, but headless bodies we piledin its cage were gone by morning. We are notsure whether they were dragged away byinsects or eaten by the rat. We extractedstomach contents from two rats. In one, thestomach was distended with bait (mixture ofpeanut butter, bacon pieces, oatmeal, andraisins) in which were scattered pieces of verysmall beetles, which were likely on the bait andinadvertently ingested by the rat. The stomachof the other was partly full, containingfragments of sclerites intermingled with softunidentifiable animal tissue. Tonkinomys dao-vantieni includes insects in its diet, but othercomponents remain to be discovered.
The limestone rat is probably nocturnal, butmay also be active at times during the day. Wewere drawn to the talus because during onelate morning Lunde had glimpsed a large,dark gray rodent scurrying across the lime-stone and disappearing into the rubble.Results from the first night’s traps revealedthe rodent to be the limestone rat. We keptour traps in place during day and night, andoften checked them in the evening, but nevercaught any rats during the day.
CLOSING OBSERVATIONS
Other species of Indochinese murids live inthe tropical forests of karst regions but, exceptfor Saxatilomys paulinae, none have any ofthe diagnostic morphological attributes ofTonkinomys daovantieni and none are soecologically tied to the kind of creviscularhabitat in which we discovered the Vietnamesespecies. Two Thai murids are generally con-sidered closely associated with limestone.Leopoldamys neilli is documented by a fewspecimens captured on the upper sections offorested limestone cliffs in Saraburi Provinceand adjacent Kanchanaburi Province insouthwestern Thailand (Marshall, 1988; speci-mens in AMNH and USNM). Wiles (1981:
49), however, captured three individuals inlowland bamboo forest in a different part ofKanchanaburi Province. The trap sites aver-aged 115 m from the base of the nearestlimestone mountain, and L. neilli was notencountered in the other habitat types Wilessurveyed, indicating the species to preferlowland bamboo forest, which ‘‘… challengesthe belief that L. neilli is strictly a cliff-dwelling rat.’’
Niviventer hinpoon is apparently endemic tothe Korat Plateau in Saraburi Province ofsouthern Thailand (Marshall, 1988; specimensin AMNH and USNM). One sample, whichincludes the holotype, was collected halfwayup the face of a forested limestone cliff outsidethe entrance to a cave. Another series camefrom ‘‘… high above the valley floor, inscrubby vegetation at the base of verticallimestone cliffs … ,’’ (Marshall, 1988: 459).Other species of murids were captured in theforested stream valley below the cliffs, but notN. hinpoon. Marshall (1988) also noted thatone individual had been taken inside a lime-stone cave at a different locality. Whether N.hinpoon dens in clefts in cliffs or in talus is notrecorded. Except for its tail, which is equal toor slightly longer than length of head andbody, its buffy gray underparts, and itssmaller body size (Marshall, 1988), the mor-phological traits of N. hinpoon are similar tothose of N. fulvescens; our multivariate anal-yses of morphometric cranial and dental dataplace it closer to N. fulvescens than to anyother species (Musser and Lunde, ms.).Relative to palmar and plantar areas, thepads are no larger or set no closer togetherthan in N. fulvescens (see fig. 9) or any of theother species of Niviventer. Nothing in themorphology of N. hinpoon points to anobligate limestone cliff-dweller. We need tolearn details of its actual microhabitat andother aspects of its life history.
Nothing comparable to the VietnameseTonkinomys, or to the Lao Saxatilomys, inmorphology or habits has so far been found inthe karst landscapes of Thailand.
Whitmore (1984: 148) noted that karstlandscapes covered only a small part oftropical Asia. In Indochina they occur insouthern China, north and central Vietnam,central Lao PDR, southern Cambodia,
2006 MUSSER ET AL.: NEW RODENT FROM VIETNAM 37

Thailand, and northeastern Burma (Mid-dleton and Waltham, 1986; Zhang, 1989;Tuyet, 1998; also see the references cited inWhitmore, 1984). Except for the large lime-stone plateau in southern China, karst land-scapes in Indochina are small in area, scat-tered over the region, and constitute isolatedgeological islands (see the informative distri-bution maps in Middleton and Waltham,1986). They are difficult places in which towork, and most Indochinese karst habitatshave been inadequately sampled for muridrodents.
Saxatilomys paulinae and Tonkinomys dao-vantieni are the only petricolous Indochinesemurines that to date have been found to beendemic to tropical karstic environments. TheKhammouan Limestone in central Lao PDRdefines the known distribution of S. paulinae.That tower karst landscape is part of the QuyDat massif (or Kebang karst block; Tuyet,1998: 189), which extends eastward at thesame latitude into Bienh Tri Thien Province ofnorth-central Vietnam. In the Lao segment, S.paulinae, the gymnure, Hylomys megalotis,and the hystricomorph rodent, Laonastesaenigmamus form a small community thatmay be restricted to forested, rocky habitatswithin the extensive and massive karst land-scape; this same mammalian assemblage mayalso occur in the Vietnamese segment of theQuy Dat massif (Musser et al., 2005). The QuyDat outcrop forms a karstic island approxi-mately 475 km south of the expansive Vietbactower karst region in northern Vietnam whereTonkinomys daovantieni occurs. That lime-stone rat, isolated on a geological islandseparate from the one on which S. paulinaewas found, has been recorded only from thetype locality in northeastern Vietnam but mayhave a wider geographic distribution inforested habitats throughout the karst land-scapes of northern Vietnam and adjacentsouthern China. Unlike the KhammouanLimestone region, no other petricolous mam-malian limestone endemics have been discov-ered in the same landscape occupied by T.daovantieni.
We have asked ourselves if the Vietnameseand Lao limestone rats are the only murinecomponents of a pattern reflecting evolution-ary adaptation of ancestral murines to forest-
ed karstic environments, or if other karsticislands in Indochina support different andunique murine endemics. Surveys for smallmammals, such as ours that yieldedTonkinomys, would uncover the mammaliandiversity in these extraordinary isolated land-scapes and provide an answer to our question.
ACKNOWLEDGMENTS
Our work in Vietnam has its origins ina National Science Foundation Grant(No. 9870232) to the Center for Biodiversityand Conservation at the American Museum ofNatural History. Funds for the survey duringApril of 2004 were provided by the Division ofVertebrate Zoology (Mammalogy) at theAmerican Museum, and we are especiallygrateful to Nancy Simmons for her help insecuring that money. Dr. Nguyen Xuan Dang,Head of the Department of Zoology at theInstitute of Ecology and Biological Resourcesin Hanoi, allowed us to use the department asa home base and offered the assistance ofmembers of the scientific staff: Nguyen XuanNghia (mammalogist) and Doan Van Kien(herpetologist). Mr. Le Dinh Thanh served ascamp cook and agreeable companion. Manyresidents of Lang Son Province supported ourendeavors in the field, including Mr. TranVan Chuyen, our local forest guide, and TrieuSinh Duong, the head of the Dao villagewhere we worked. Neil Duncan prepared theskulls at the American Museum. The brillianttalent and competence of Patricia J. Wynneare reflected in her line drawings. Themorphological comparisons described in ourreport are built on results of biologicalsurveys in the field, and the innumerable days(over a span of three decades for Musser)working in collections of museums in theUnited States, Europe, and Asia. Custodiansof those collections, some now deceased,allowed us to study specimens, and neverhesitated to loan material whenever we re-quested it. This report, those already pub-lished under our names, and future contribu-tions now in manuscript, reflect in part theresults of original research, but also theunselfish contributions of those persons whoare responsible for maintaining research col-lections—we thank you.
38 AMERICAN MUSEUM NOVITATES NO. 3517

REFERENCES
Amato, G., M.G. Egan, and G.B. Schaller. 2000.Mitochrondrial DNA variation in muntjac:evidence for discovery, rediscovery, and phylo-genetic relationships. In E.S. Vrba and G.B.Schaller (editors), Antelopes, deer, and rela-tives: fossil record, behavioral ecology, system-atics, and conservation: 285-295. New Haven:Yale University Press.
Averianov, A.O., A.V. Abramov, and A.N.Tikhonov. 2000. A new species of Nesolagus(Lagomorpha, Leporidae) from Vietnam withosteological description. Contributions fromthe Zoological Institute, St. Petersburg 3: 1–22.
Bacon, A-M., F. Demeter, M. Schuster, V.T. Long,N.K. Thuy, P-O. Antoine, S. Sen, H.H. Hga,and N.M. Huong. 2004. The Pleistocene MaU’Oi cave, northern Vietnam: palaeontology,sedimentology and palaeoenvironments. Geo-bios 37: 305–314.
Boye, P., R. Hutterer, N. Lopez-Martınez, and J.Michaux. 1992. A reconstruction of the lavamouse (Malpaisomys insularis), an extinctrodent of the Canary Islands. Zeitschrift furSaugetierkunde 57: 29–38.
Brown, J.C. 1971. The description of mammals 1.The external characters of the head. MammalReview 1: 151–168.
Brown, J.C., and D.W. Yalden. 1973. The de-scription of mammals 2. Limbs and locomotionof terrestrial mammals. Mammal Review 3:107–134.
Bugge, J. 1970. The contribution of the stapedialartery to the cephalic arterial supply in muroidrodents. Acta Anatomica 76: 313–336.
Bugge, J. 1985. Systematic value of the carotidarterial pattern in rodents. In W.P. Luckett andJ.L. Hartenberger (editors), Evolutionary rela-tionships among rodents: a multidisciplinaryanalysis: 355-379. New York: Plenum Press.
Can, D.N., A.V. Abramov, A.N. Tikhonov, andA.O. Averianov. 2001. Annamite striped rabbitNesolagus timminsi in Vietnam. Acta Therio-logica 46(4): 437–440.
Carleton, M.D. 1980. Phylogenetic relationships inneotomine-peromyscine rodents (Muroidea)and a reappraisal of the dichotomy withinNew World Cricetinae. Miscellaneous Publi-cations Museum of Zoology, University ofMichigan 157: 1–146.
Carleton, M.D., and G.G. Musser. 1984. Muroidrodents. In S. Anderson and J.K. Jones, Jr.(editors), Orders and families of Recent mam-mals of the world: 289-379. New York: Wiley.
Carleton, M.D., and G.G. Musser. 1989. System-atic studies of oryzomyine rodents (Muridae,Sigmodontinae): a synopsis of Microryzomys.
Bulletin of the American Museum of NaturalHistory 191: 1–83.
Chaimanee, Y. 1998. Plio-Pleistocene rodents ofThailand. Thai Studies in Biodiversity 3: 1–303.
Chen, G.-J., W. Wang, J.-Y. Mo, Z.-T. Huang, F.Tian, and W.-W. Huang. 2002. Pleistocenevertebrate fauna from Wuyun Cave ofTiandong County, Guangxi. VertebrataPalAsiatica 40(1): 42–51.
Corbet, G.B., and J.E. Hill. 1992. Mammals of theIndomalayan region. A systematic review.Oxford: Oxford University Press. 488 pp.
Csorba, G., and P.D. Jenkins. 1998. First recordsand a new subspecies of Rhinolophus stheno(Chiroptera, Rhinolophidae) from Vietnam.Bulletin of the Natural History Museum,London (Zoology) 66(2): 207–211.
Ðang, H.H., V.T. Ðao, V.S. Cao, T.A. Pham, andM.K. Hoang. 1994. [Checklist of mammals inVietnam.] Hanoi: Publishing House ‘‘Scienceand Technics’’. 167 pp. [in Vietnamese]
Ðao, V.T. 1985. [Scientific results of some mammalsurveys in North Vietnam (1957–1971)]. Hanoi:Scientific and Technical Publishing House.330 pp. [in Vietnamese]
Ðieu, N.T. 1995. Geography of Vietnam: natural—human—economic. Hanoi: The Gio,I Pub-lishers.
Finley, R.B., Jr. 1958. The wood rats of Colorado:distribution and ecology. University of KansasPublications of the Museum of Natural History10: 213–552.
Flynn, L.J., W. Wu, and W.R. Downs, III. 1997.Dating vertebrate microfaunas in the lateNeogene record of northern China. Palae-ogeography, Palaeoclimatology, Palaeoecology133: 227–242.
Gillieson, D. 1996. Caves: Processes, developmentand management. Oxford: Blackwell.
Głazek, J. 1966. On the karst phenomena in NorthVietnam. Bulletin de l’Acemie Polonaise desSciences, Serie des Science Geologie etGeographie 14(1): 45–51.
Gorog, A.J., M.H. Sinaga, and M.D. Engstrom.2004. Vicariance or dispersal? Historical bio-geography of three Sunda shelf murine rodents(Maxomys surifer, Leopoldamys sabanus andMaxomys whiteheadi). Biological Journal of theLinnean Society 81: 91–109.
Grayson, D.K. 1999. Bushy-tailed woodrat—Neotoma cinerea. In D.E. Wilson and S. Ruff(editor.). The Smithsonian book of NorthAmerican mammals: 598-600. Washington, DC:Smithsonian Institution Press.
Groves, C.P. 2001. Primate taxonomy. Wash-ington, DC: Smithsonian Institution Press.
2006 MUSSER ET AL.: NEW RODENT FROM VIETNAM 39

Groves, C.P., and G.B. Schaller. 2000. Thephylogeny and biogeography of the newlydiscovered Annamite artiodactyls. In E.S.Vrba and G.B. Schaller (editors), Antelopes,deer, and relatives. Fossil record, behavioralecology, systematics, and conservation: 261-282. New Haven, Yale University Press.
Groves, C.P., and V. Weitzel. 1995. Obituary: DaoVan Tien. Laboratory Primate Newsletter34(3): 15–16.
Grubb, P. 2005. Order Artiodactyla. In D.E.Wilson and D.M. Reeder (editors), Mammalspecies of the world: a taxonomic and geo-graphic reference, 3rd ed.: 637-722. Baltimore:Johns Hopkins University Press.
Hershkovitz, P. 1962. Evolution of Neotropicalcricetine rodents (Muridae), with special refer-ence to the phyllotine group. Fieldiana Zoology46: 1–524.
Hutterer, R., N. Lopez-Martınez, and J. Michaux.1988. A new rodent from Quaternary depositsof the Canary Islands and its relationships withNeogene and Recent murids of Europe andAfrica. Palaeovertebrata 18: 241–262.
Kruskop, S.V., and K.A. Tsytsulina. 2001. A newbig-footed mouse-eared bat Myotis annamiticussp. nov. (Vespertilionidae, Chiroptera) fromVietnam. Mammalia 65(1): 63–72.
Lunde, D.P., and N.T. Son. 2001. An identificationguide to the rodents of Vietnam. New York:Center for Biodiversity and Conservation,American Museum of Natural History.
Lunde, D.P., G.G. Musser, and N.T. Son. 2003. Asurvey of small mammals from Mt. Tay ConLinh II, Vietnam, with the description of a newspecies of Chodsigoa (Insectivora: Soricidae).Mammal Study 28: 31–46.
Lunde, D.P., G.G. Musser, and T. Ziegler. 2004.Description of a new species of Crocidura(Soricomorpha: Soricidae, Crocidurinae) fromKe Go Nature Reserve, Vietnam. MammalStudy 29: 27–36.
Marshall, J.T., Jr. 1988. Family Muridae. InB. Lekagul, and J.A. McNeely (authors),Mammals of Thailand, 2nd ed.: 397-487.Bangkok: Association for the Conservationof Wildlife, Saha Karn Bhaet Co.
Middleton, J., and T. Waltham. 1986. The un-derground atlas. A gazetteer of the world’s caveregions. New York: St. Martin’s Press.
Miller, G.S., Jr. 1912. Catalogue of the mammals ofWestern Europe (Europe exclusive of Russia)in the collection of the British Museum.London: British Museum (Natural History).
Misonne, X. 1969. African and Indo-AustralianMuridae: evolutionary trends. Annales MuseeRoyal de l’Afrique Centrale, Tervuren,
Belgique, Serie IN-8, Sciences Zoologiques172: 1–219.
Musser, G.G. 1970. Species-limits of Rattus brah-ma, a murid rodent of northeastern India andnorthern Burma. American Museum Novitates2406: 1–27.
Musser, G.G. 1973. Species-limits of Rattus cre-moriventer and Rattus langbianis, murid ro-dents of Southeast Asia and the Greater SundaIslands. American Museum Novitates 2525:1–65.
Musser, G.G. 1981. Results of the ArchboldExpeditions. No. 105. Notes on systematics ofIndo-Malayan murid rodents, and descriptionsof new genera and species from Ceylon,Sulawesi, and the Philippines. Bulletin of theAmerican Museum of Natural History 168(3):225–334.
Musser, G.G. 1982. Results of the ArchboldExpeditions. No. 110. Crunomys and thesmall-bodied shrew rats native to thePhilippine islands and Sulawesi (Celebes).Bulletin of the American Museum of NaturalHistory 174: 1–95.
Musser, G.G. 1991. Sulawesi rodents: descriptionsof new species of Bunomys and Maxomys(Muridae, Murinae). American MuseumNovitates 3001: 1–41.
Musser, G.G., and M.D. Carleton. 1993. FamilyMuridae. In D.E. Wilson and D.M. Reeder(editors), Mammal species of the world:a taxonomic and geographic reference, 2nded.: 501-755. Washington DC: SmithsonianInstitution Press.
Musser, G.G., and M.D. Carleton. 2005.Superfamily Muroidea. In D.E. Wilson andD.M. Reeder (editors), Mammal species of theworld: a taxonomic and geographic reference,3rd ed.: 894-1531. Baltimore: Johns HopkinsUniversity Press.
Musser, G.G., and S. Chiu. 1979. Notes ontaxonomy of Rattus andersoni and R. excelsior,murids endemic to western China. Journal ofMammalogy 60(3): 581–592.
Musser, G.G., and L.A. Durden. 2002. Sulawesirodents: description of a new genus and speciesof Murinae (Muridae, Rodentia) and itsparasitic new species of sucking louse (Insecta,Anoplura). American Museum Novitates 3368:1–50.
Musser, G.G., and L.H. Heaney. 1992. Philippinerodents: definitions of Tarsomys and Limnomysplus a preliminary assessment of phylogeneticpatterns among native Philippine murines(Murinae, Muidae). Bulletin of the AmericanMuseum of Natural History 211: 1–138.
Musser, G.G., and C. Newcomb. 1983. Malaysianmurids and the giant rat of Sumatra. Bulletin of
40 AMERICAN MUSEUM NOVITATES NO. 3517

the American Museum of Natural History174(4): 327–598.
Musser, G.G., J.T. Marshall, Jr., and Boeadi. 1979.Definition and contents of the Sundaic genusMaxomys (Rodentia, Muridae). Journal ofMammalogy 60(3): 592–606.
Musser, G.G., A.L. Smith, M.F. Robinson, andD.P. Lunde. 2005. Description of a new genusand species of rodent (Murinae, Muridae,Rodentia) from the Khammouan LimestoneNational Biodiversity Conservation Area inLao PDR. American Museum Novitates 3497:1–31.
Musser, G.G., A. Van de Weerd, and E. Strasser.1986. Paulamys, a replacement name forFloresomys Musser, 1981 (Muridae), and newmaterial of that taxon from Flores, Indonesia.American Museum Novitates 2850: 1–10.
Musser, G.G., L.R. Heaney, and B.R. Taba-ranza, Jr. 1998. Philippine rodents: redefini-tions of known species of Batomys (Muridae,Murinae) and description of a new species fromDinagat Island. American Museum Novitates3237: 1–51.
Qiu, Z. 1989. The Chinese Neogene mammalianbiochronology—Its correlation with the Euro-pean Neogene mammalian zonation. In E.H.Lindsay, V. Fahlbusch and P. Mein (editors),European Neogene Mammal Chronology: 527-556. New York: Plenum Press.
Robinson, M.F., P.D. Jenkins, C.M. Francis, andA.J.C. Fulford. 2003. A new species of theHipposideros pratti group (Chiroptera, Hippo-sideridae) from Lao PDR and Vietnam. ActaChiropterologica 5(1): 31–48.
Schaub, S. 1938. Tertiare und Quartare Murinae.Abhandlungen der Schweizerischen Paleon-tologischen Gesellschaft 61: 1–39.
Sokolov, V.E., V.V. Rozhnov, and P.T. Anh. 1997.New species of viverrids of the genus Viverra(Mammalia, Carnivora) from Vietnam.Zoologicheskii Zhurnal 76: 585–589.
Thomas, O. 1919. The method of taking the incisiveindex in rodents. Annals and Magazine ofNatural History, Series 9(4): 289–290.
Tuyet, Do. 1998. Overview on karst of Vietnam. InYuan Daoxian and Liu Zaihua (editors),Global karst correlation: 179-192. Hanoi:Science Press and VSP BV.
Voss, R.S. 1988. Systematics and ecology ofichthyomyine rodents (Muroidea): patterns ofmorphological evolution in a small adaptiveradiation. Bulletin of the American Museum ofNatural History 188(2): 259–493.
Voss, R.S. 1993. A revision of the Brazilian muroidrodent genus Delomys with remarks on‘‘Thomasomyine’’ characters. American Mu-seum Novitates 3073: 1–44.
Wahlert, J.H. 1985. Cranial foramina of rodents. InW.P. Luckett and J.-l. Hartenberger (editors),Evolutionary relationships among rodents,a multidisciplinary analysis: 311-332. NewYork: Plenum Press.
Walston, J., and G. Veron. 2001. Questionablestatus of the ‘‘Taynguyen civet’’, Viverratainguensis Sokolov, Rozhnov and PhamTrong Anh, 1997. (Mammalia: Carnivora:Viverridae). Mammalian Biology 66: 181–184.
Whitmore, T.C. 1984. Tropical rain forests of theFar East, 2nd ed. Oxford: Clarendon Press.
Wiles, G.J. 1981. Abundance and habitat prefer-ences of small mammals in southwesternThailand. Natural History Bulletin of theSiam Society 29: 44–54.
Wozencraft, W.C. 2005. Order Carnivora. In D.E.Wilson and D.M. Reeder (editors), Mammalspecies of the world: a taxonomic and geo-graphic reference, 3rd ed.: 532-628. Baltimore:Johns Hopkins University Press.
Zhang, S. 1989. Paleokarst of China. In P. Bosak,D.C. Ford, J. Głazek and I. Horack (editors),Paleokarst, a systematic and regional review:297-310. Amsterdam and Praha: Elsevier andAcademia.
Zheng, S. 1993. [Quaternary rodents of Sichuan-Guizhou area, China] Beijing: Science Press.270 pp.
Zheng, S. 2004. Mammalia: Rodentia. In S. Zheng(editor), Jianshi Hominid Site. State KeyProject of the 9th Five Year Plan—Origin ofEarly Humans and Environmental Back-ground: 121-181. Series Monograph I. Beijing:Science Press. [in Chinese, with English sum-mary]
Zheng, S., Z. Zhang, and L. Liu. 1997. [Pleistocenemammals from fissure-fillings of SunjiashanHill, Shandong, China.] Vertebrata PalAsiatica35(3): 201–216.
2006 MUSSER ET AL.: NEW RODENT FROM VIETNAM 41