current biology supplemental information org 1 dependent …€¦ · · 2015-04-07org-1-dependent...
TRANSCRIPT

Current Biology
Supplemental Information
Org-1-Dependent Lineage Reprogramming
Generates the Ventral Longitudinal
Musculature of the Drosophila Heart
Christoph Schaub, Johannes März, Ingolf Reim, and Manfred Frasch
Supplemental data
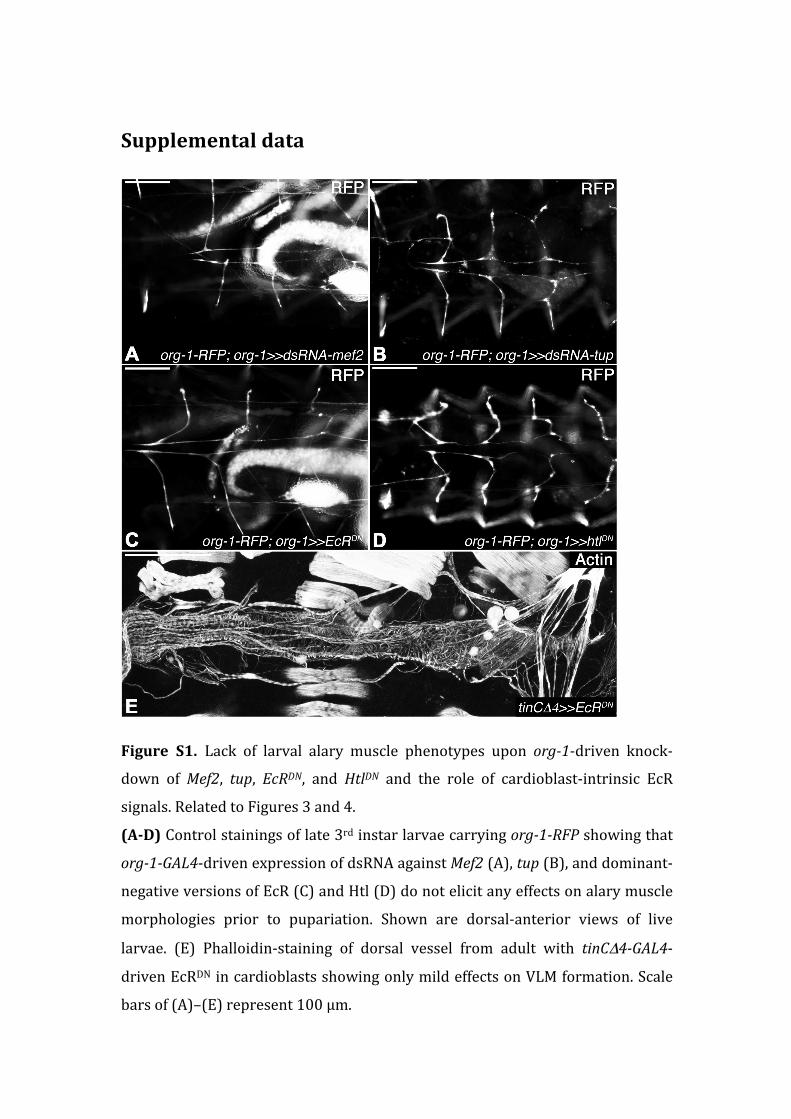
Figure S1. Lack of larval alary muscle phenotypes upon org-‐1-‐driven knock-‐
down of Mef2, tup, EcRDN, and HtlDN and the role of cardioblast-‐intrinsic EcR
signals. Related to Figures 3 and 4.
(A-‐D) Control stainings of late 3rd instar larvae carrying org-‐1-‐RFP showing that
org-‐1-‐GAL4-‐driven expression of dsRNA against Mef2 (A), tup (B), and dominant-‐
negative versions of EcR (C) and Htl (D) do not elicit any effects on alary muscle
morphologies prior to pupariation. Shown are dorsal-‐anterior views of live
larvae. (E) Phalloidin-‐staining of dorsal vessel from adult with tinCΔ4-‐GAL4-‐
driven EcRDN in cardioblasts showing only mild effects on VLM formation. Scale
bars of (A)–(E) represent 100 µm.

Supplemental Material & Methods
Analysis of pupal and adult phenotypes
Individuals were kept and GAL4/UAS-‐induced overexpression was carried out at
22°C, except for the experiments using TARGET [S1], which involved
temperature shifts to 28oC during embryonic stages. Control experiments with
the TARGET system without this temperature shift did not show any phenotype
or G-‐TRACE expression, thus ruling out that the observed effects are due to leaky
expression during later development.
Pupal stages until P4 were dissected with microsurgery scissors in toto. For
pupae from P4 and later as well as for pharate adults the pupal cases were
removed prior to dissection.
Drosophila strains
In this study the following strains were used: org-‐1-‐HN39-‐GFP [S2], hand-‐nGFP
[S3, S4], duf-‐rP298-‐lacZ [S5] and duf-‐rP298-‐GAL4 [S6], tupF4-‐nGFP [S7], tin346
[S8], tinEC40 [S9], tin-‐ABD [S10], and tinCΔ4-‐Gal4 [S11]. UAS-‐Abd-‐A was a gift from
Juan Botas (Baylor, Houston). UAS-‐lifeact-‐GFP was a gift from Frank Schnorrer
(MPI, Martinsried). UAS-‐dsRNA-‐org-‐1 (Transformant ID 104393), UAS-‐dsRNA-‐tup
(Transformant IDs 103585 and 45859) and UAS-‐dsRNA-‐Mef2 (Transformant IDs
15549 and 15550) were obtained from the VDRC (Vienna) [S12]. The two
different RNAi lines for tup and Mef2, respectively, showed essentially the same
effects. tubP-‐GAL80ts20;TM2/TM6B and UAS-‐FLP-‐Exel3,Ubi-‐p63E(FRT.STOP)
Stinger [S13] as well as UAS-‐EcR-‐B1DN [S14] and UAS-‐heartlessDN [S15] were
obtained from the Bloomington stock center (Indiana).
Construction of org-‐1-‐HN18-‐RFP, org-‐1-‐HN39-‐GAL4 and tup-‐ADME-‐GFP
reporters
For the creation of org-‐1-‐HN18-‐RFP, the genomic region chrX: 8442048...
8444555 (R6.01) was amplified using yw genomic DNA as template and cloned
into BglII/NaeI of pRed H-‐Pelican [S16].
For org-‐1-‐HN39-‐GAL4 the HN39 fragment from org-‐1-‐HN39-‐pH-‐Stinger-‐AttB [S2]
was cloned into EcoRI/BamHI of p221-‐GAL4 (a gift from C. Klämbt).

For tup-‐ADME-‐GFP the genomic region chr2L: 18899122…18900688 (R6.01)
(tup-‐ADME) [S17] was amplified using yw genomic DNA as template and cloned
into KpnI/XhoI of pH-‐Stinger-‐AttB [S18]. For the analogous creation of tup-‐
ADMEorgI-‐IIImut-‐GFP reporter the Org-‐1 binding sites within the tup-‐ADME
sequence [S17] were mutated via site directed mutagenesis as follows: tup-‐
ADME-‐orgI TAACACAT -‐> tup-‐ADME-‐orgImut TAAGCTTT, tup-‐ADME-‐orgII
GGGTGCCA -‐> tup-‐ADME-‐orgIImut GGCTCGAG and tup-‐ADME-‐orgIII TGGTGGGA -‐
> tup-‐ADME-‐orgIIImut TGTCTAGA.
All constructs were transformed into yw using standard transgenesis techniques.
Immunofluorescence
For antibody stainings the dissected animals were fixed in 3,7 % formaldehyde
for 1 hour, washed several times in PBT, and incubated in the primary antibody
for two days at 4 °C. Subsequent staining procedures were performed as
described [S19].
The following primary antibodies were used: rat anti-‐Org-‐1 (1:100, [S2]), mouse
anti-‐Isl1/Tup 40.3A4 (1:25, DSHB) guinea-‐pig anti-‐Ubx (1:400, a gift from I.
Lohmann, Heidelberg), rabbit anti-‐RFP (1:300, Millipore) and mouse anti-‐GFP
(1:100, Molecular Probes). Filamentous actin was visualized using Phalloidin-‐
Atto 647N (1:1500, Sigma-‐Aldrich). Confocal pictures were taken with a Leica
SP5 II (20x/1.3 PL APO Glycerol). Projections were done with Leica Application
Suite Advanced Fluorescence (LAS AF).
In vivo time-‐lapse imaging
Pupae were aligned on a strip of double-‐faced adhesive tape connected to a slide,
covered with a drop of halocarbone oil and a coverslip. Time-‐lapse imaging was
essentially performed as described in [S20, S21]. Movies were generated using
LAS AF.

Supplemental References S1. McGuire, S.E., Mao, Z., and Davis, R.L. (2004). Spatiotemporal gene
expression targeting with the TARGET and gene-‐switch systems in
Drosophila. Sci STKE 2004, pl6.
S2. Schaub, C., Nagaso, H., Jin, H., and Frasch, M. (2012). Org-‐1, the Drosophila
ortholog of Tbx1, is a direct activator of known identity genes during
muscle specification. Development 139, 1001-‐1012.
S3. Han, Z., and Olson, E.N. (2005). Hand is a direct target of Tinman and
GATA factors during Drosophila cardiogenesis and hematopoiesis.
Development 132, 3525-‐3536.
S4. Sellin, J., Albrecht, S., Kolsch, V., and Paululat, A. (2006). Dynamics of heart
differentiation, visualized utilizing heart enhancer elements of the
Drosophila melanogaster bHLH transcription factor Hand. Gene Expr
Patterns 6, 360-‐375.
S5. Nose, A., Isshiki, T., and Takeichi, M. (1998). Regional specification of
muscle progenitors in Drosophila: the role of the msh homeobox gene.
Development 125, 215-‐223.
S6. Menon, S.D., and Chia, W. (2001). Drosophila Rolling pebbles: a
multidomain protein required for myoblast fusion that recruits D-‐Titin in
response to the myoblast attractant Dumbfounded. Dev Cell 1, 691-‐703.
S7. Tao, Y., Wang, J.B., Tokusumi, T., Gajewski, K., and Schulz, R.A. (2007).
Requirement of the LIM homeodomain transcription factor Tailup for
normal heart and hematopoietic organ formation in Drosophila
melanogaster. Mol Cell Biol 27, 3962-‐3969.
S8. Azpiazu, N., and Frasch, M. (1993). tinman and bagpipe: two homeo box
genes that determine cell fates in the dorsal mesoderm of Drosophila.
Genes Dev 7, 1325-‐1340.
S9. Bodmer, R. (1993). The gene tinman is required for specification of the
heart and visceral muscles in Drosophila. Development 118, 719-‐729.
S10. Zaffran, S., Reim, I., Qian, L., Lo, P.C., Bodmer, R., and Frasch, M. (2006).
Cardioblast-‐intrinsic Tinman activity controls proper diversification and
differentiation of myocardial cells in Drosophila. Development 133, 4073-‐
4083.

S11. Lo, P.C., and Frasch, M. (2001). A role for the COUP-‐TF-‐related gene seven-‐
up in the diversification of cardioblast identities in the dorsal vessel of
Drosophila. Mech. Dev. 104, 49-‐60.
S12. Dietzl, G., Chen, D., Schnorrer, F., Su, K.C., Barinova, Y., Fellner, M., Gasser,
B., Kinsey, K., Oppel, S., Scheiblauer, S., et al. (2007). A genome-‐wide
transgenic RNAi library for conditional gene inactivation in Drosophila.
Nature 448, 151-‐156.
S13. Evans, C.J., Olson, J.M., Ngo, K.T., Kim, E., Lee, N.E., Kuoy, E., Patananan,
A.N., Sitz, D., Tran, P., Do, M.T., et al. (2009). G-‐TRACE: rapid Gal4-‐based
cell lineage analysis in Drosophila. Nat Methods 6, 603-‐605.
S14. Cherbas, L., Hu, X., Zhimulev, I., Belyaeva, E., and Cherbas, P. (2003). EcR
isoforms in Drosophila: testing tissue-‐specific requirements by targeted
blockade and rescue. Development 130, 271-‐284.
S15. Michelson, A.M., Gisselbrecht, S., Zhou, Y., Baek, K.H., and Buff, E.M. (1998).
Dual functions of the Heartless fibroblast growth factor receptor in
development of the Drosophila embryonic mesoderm. Dev Genet 22, 212-‐
229.
S16. Barolo, S., Castro, B., and Posakony, J.W. (2004). New Drosophila
transgenic reporters: insulated P-‐element vectors expressing fast-‐
maturing RFP. BioTechniques 36, 436-‐440, 442.
S17. Boukhatmi, H., Schaub, C., Bataillé, L., Reim, I., Frendo, J.-‐L., Frasch, M., and
Vincent, A. (2014). An Org-‐1–Tup transcriptional cascade reveals
different types of alary muscles connecting internal organs in Drosophila.
Development 141, 3761-‐3771.
S18. Jin, H., Stojnic, R., Adryan, B., Ozdemir, A., Stathopoulos, A., and Frasch, M.
(2013). Genome-‐wide screens for in vivo Tinman binding sites identify
cardiac enhancers with diverse functional architectures. PLoS Genet 9,
e1003195.
S19. Domsch, K., Ezzeddine, N., and Nguyen, H.T. (2013). Abba is an essential
TRIM/RBCC protein to maintain the integrity of sarcomeric
cytoarchitecture. J Cell Sci 126, 3314-‐3323.
S20. Hollfelder, D., Frasch, M., and Reim, I. (2014). Distinct functions of the
laminin beta LN domain and collagen IV during cardiac extracellular

matrix formation and stabilization of alary muscle attachments revealed
by EMS mutagenesis in Drosophila. BMC Dev Biol 14, 26.
S21. Reim, I., Hollfelder, D., Ismat, A., and Frasch, M. (2012). The FGF8-‐related
signals Pyramus and Thisbe promote pathfinding, substrate adhesion, and
survival of migrating longitudinal gut muscle founder cells. Dev Biol 368,
28-‐43.