crop adaptation to climate change (yadav/crop adaptation to climate change) || wild relative and...
TRANSCRIPT

P1: SFK/UKS P2: SFK Color: 1C
BLBS082-27 BLBS082-Yadav July 12, 2011 17:16 Trim: 246mm X 189mm
Chapter 27
Wild Relative and Transgenic Innovationfor Enhancing Crop Adaptation to Warmerand Drier ClimateGang-Ping Xue and C. Lynne McIntyre
Introduction
Climate change scenarios suggest a warmer andfrequently drier climate in the future. In agri-culture, such a scenario would require a shift offocus in plant breeding for most regions of theworld from increasing yield potential to improv-ing yield stability in drought- and heat-proneenvironments through the increased stress tol-erance. Drought and heat stresses can cause amarked reduction in crop productivity. Droughtand heat, in combination, are known to have aneven more severe impact on carbon assimilationand hence crop yield (Barnabas et al. 2008). Ex-tensive selection for yield potential in agricul-tural environments in the last century has greatlyreduced genetic variation, especially for agro-nomic traits necessary to meet the challenge ofabiotic stresses. Introduction of stress-resistancegenes from wild relatives (WRs) that currentlygrow in existing harsh environments is an essen-tial part of crop breeding for the future. How-ever, it is generally considered that breeding forenhancing yield in drought and heat-prone envi-ronments is technically difficult (Richards 1996;Rauf 2008), as many morphological, physiologi-
cal, and biochemical attributes contribute to yieldand each is a multigene trait. Many of droughtand heat-resistance traits are expressed throughinteraction between genetics and environment.There is a strong need to elucidate the molecularand genetic bases of drought and heat resistancein crop species, to identify beneficial genes andalleles, and to utilize them in breeding programsfor efficient selection of superior varieties in thefuture.
The understanding of the molecular mecha-nisms underpinning each stress-resistance traitis required to achieve a more rapid and substan-tial progress in the improvement of crop abi-otic stress resistance. It is generally thought thatsuccess in pyramiding of the best stress adapta-tion genes in future crop varieties in a relativelyshort time is likely to result from an integratedapproach of trait-based breeding and molecu-lar technologies. Recent molecular and genomicstudies on plant adaptation to drought and heatstresses and the development of transgenic croptechnologies provide a foundation for potentiallyrapid incorporation of the crucial stress adapta-tion genes into crop species in future breedingprograms. Furthermore, crop WRs that inhabit
Crop Adaptation to Climate Change, First Edition. Edited by Shyam S. Yadav, Robert J. Redden, Jerry L. Hatfield,Hermann Lotze-Campen and Anthony E. Hall.c© 2011 John Wiley & Sons, Ltd. Published 2011 by John Wiley & Sons, Ltd.
522

P1: SFK/UKS P2: SFK Color: 1C
BLBS082-27 BLBS082-Yadav July 12, 2011 17:16 Trim: 246mm X 189mm
ADAPTATION TO WARMER AND DRIER CLIMATE 523
Protective proteincapacity
Antioxidativecapacity
Osmoticadjustment
General abioticstress protection
Droughtescape
Transpirationefficiency
Stomatalconductance
Carbonassimilation
Water-useefficiency
Crop productivityunder water deficit
Stem carbonreserve
Deep soil waterextraction
Maintainingphotosynthesis
Delayed leafsenescence
Earlyphenology
Osmotic protectioncapacity
Fig. 27.1. Physiological and biochemical mechanisms relevant to crop productivity indrought-prone environments.
extreme drought and heat environments couldprovide some superior alleles for crop improve-ment. This chapter will outline our current under-standing in the molecular mechanisms of droughtand heat resistance in crop species and theirWRs, examine genes that are associated withstress resistance, and review recent progress intransgenic innovation for enhancing crop adap-tation to drought and heat stresses.
The molecular mechanisms ofdrought and heat resistance
For understanding molecular mechanisms un-derlying drought and heat resistance, first weneed to understand physiological and biochem-ical mechanisms. It should be aware that notall stress adaptive mechanisms are beneficial toyield. Some traits that enhance crop survival mayonly produce a small or no yield improvementunder stress (Blum 2005).
Some useful physiological and biochemicalmechanisms relevant to crop productivity un-der water-limited conditions are summarized
in Fig. 27.1. Antioxidative, osmotic protectionand protective protein capacities are importantbiochemical mechanisms that have not been dis-cussed in the previous chapters. The antioxi-dant system is required to remove toxic reac-tive oxygen species (ROS) (e.g., O2
−, H2O2,and OH−), which are overproduced in plantsduring abiotic stress, through both enzymatic(e.g., superoxide dismutase, ascorbate peroxi-dase, and glutathione peroxidase) or nonenzy-matic reactions (e.g., ascorbic acid, glutathione,and α-tocopherol as well as some sugar alco-hols: proline, mannitol, etc). The osmotic pro-tection system serves as osmoprotectant to sta-bilize proteins and cell structures in dehydratedplants and consists of various osmolytes: sug-ars (e.g., hexoses and sucrose), polyhydric al-cohols (e.g., mannitol and pinitol), and amines(e.g., glycine betaine and proline), although thetypes of osmolytes that are accumulated dur-ing drought stress vary among species (Bartelsand Sunkar 2005; Xue et al. 2008a). The pro-tective protein system consists of mainly twoimportant protein families—late embryogenesis

P1: SFK/UKS P2: SFK Color: 1C
BLBS082-27 BLBS082-Yadav July 12, 2011 17:16 Trim: 246mm X 189mm
524 CROP ADAPTATION TO CLIMATE CHANGE
Protective proteincapacity
Antioxidativecapacity
General abioticstress protection
Heatescape
Leaf coolingcapacity
Crop productivityunder heat stress
Stem carbonreserve Membrane
lipidcomposition
Proteinthermostability
Deep soil waterextraction
Cellularthermostability
Maintainingphotosynthesis
Delayed leafsenescence
Earlyphenology
Osmotic protectioncapacity
Fig. 27.2. Physiological and biochemical mechanisms relevant to crop productivity in heat-proneenvironments.
abundant (LEA) protein and heat shock protein(HSP)/chaperone. LEA proteins include a num-ber of loosely related groups of proteins andmany of them are produced in maturing seeds anddehydrated vegetative organs. Certain LEA pro-teins are often referred to as dehydrin. It has beenproposed that LEA proteins are able to protectother proteins or membranes, in a fashion simi-lar to sugars, perhaps by acting as water replace-ment molecules. HSPs/chaperones also consistof several groups and most of them are inducedduring heat stress (Wang et al. 2004). How-ever, a few members are induced during droughtstress (Bartels and Sunkar 2005). Many HSPsare responsible for protein folding, assembly,translocation, and degradation in normal cellularprocesses, functioning as molecular chaperones.Under drought and heat stress, HSPs/chaperonescan stabilize membrane, assist protein refolding,and prevent protein denaturation. However, thesebiochemical mechanisms are rarely exploited bybreeders in current breeding practices. In con-trast, the physiological mechanisms (or traits)
listed in Fig. 27.1 have often been used in breed-ing programs (Cattivelli et al. 2008; Reynoldsand Tuberosa 2008; previous chapters). Manyof these physiological mechanisms are deter-mined by morphological characters (Reynoldsand Tuberosa 2008).
The physiological and biochemical mech-anisms for crop adaptation to heat stressshare some similarity with drought adaptation(Fig. 27.2). Three most important differencesare: (1) leaf cooling, (2) thermostability of pro-teins involved in photosynthesis and metabolicreactions, and (3) membrane thermostability. Anincrease in water evaporation helps cooling theleaf temperature. However, it depends on wa-ter availability. A deep root system may pro-vide more water for leaf cooling during heatstress in drought-prone environments. Proteinthermostability is an important issue for cropproductivity under high temperatures, as manyproteins in plants are thermolabile. For ex-ample, high growth temperature leads to themultimeric dissociation and/or denaturation of

P1: SFK/UKS P2: SFK Color: 1C
BLBS082-27 BLBS082-Yadav July 12, 2011 17:16 Trim: 246mm X 189mm
ADAPTATION TO WARMER AND DRIER CLIMATE 525
Rubisco activase in plants (Salvucci and Crafts-Brandner 2004b). Rubisco activase is a regula-tory protein for keeping Rubisco at an activatedstate. Other important thermolabile enzymes aresoluble starch synthase from wheat endosperm(Keeling et al. 1993) and ADP-glucose py-rophosphorylase from maize endosperm (Greeneand Hannah 1998). Membrane thermostabilityis important for maintenance of the integrity ofphotosynthesis apparatus and in general relieson lipid composition in the membrane. A highlevel of saturated fatty acid such as palmiticacid in chloroplast membranes is known to beassociated with heat resistance in Arabidopsis(Kunst et al. 1989). The thermolabile property ofRubisco activase and chloroplast membrane is amajor factor responsible for a heat-induced de-cline in photosynthesis. During heat stress, someHSPs/chaperones play an important role in sta-bilizing thermolabile proteins and membrane.
The molecular bases ofdrought adaptation
Genotypic variation in physiological and bio-chemical traits of drought adaptation is de-termined by differences either in protein con-centrations or protein sequence. Each of theadaptive traits is underlain by multiple genes.Recent molecular studies on plant response todrought using genome-wide expression profil-ing have shown that more than a thousand geneschange their mRNA levels (Rizhsky et al. 2004),which usually lead to changes in their pro-tein levels. Most of these genes are not di-rectly related to drought resistance, but are asecondary consequence of drought stress, whichresults from extensive metabolic changes includ-ing those involved in photosynthesis and car-bohydrate metabolism (Xue et al. 2008a). Todate, drought upregulated genes have attractedthe most attention. These genes are involved ei-ther in the known physiological and biochemicalmechanisms of drought resistance or in the reg-ulation of other drought-responsive genes, in-cluding those involved in abscisic acid (ABA)
synthesis. ABA is the major phytohormone thatmediates plant drought response (stomatal clo-sure and induction of drought-responsive genes).
The molecular studies of drought stress genesduring last 20 years have led to our currentknowledge on extensive gene regulatory net-works involved in plant response or adapta-tion to drought stress, mediated by both ABA-dependent and independent pathways (Shinozakiand Yamaguchi-Shinozaki 2007), although muchof the details of the connection within and be-tween individual networks still needs to be deter-mined. A brief summary of drought stress pro-tection genes, regulators, and their associationwith drought adaptation is given below.
Drought adaptation genes
Some drought adaptation genes, particularlydrought upregulated ones, can be directly as-sociated with the known physiological or bio-chemical traits of drought resistance. The mostcharacterized groups are genes encoding: (1) en-zymes for osmolyte synthesis, (2) LEA proteinsor HSPs/chaperones, and (3) antioxidative en-zymes or enzymes for the synthesis of antioxi-dants. A comprehensive review of this topic iscovered by Bartels and Sunkar (2005). How-ever, many drought adaptation genes that un-derlie the morphological and physiological traitsof drought resistance, such as a deep root sys-tem and water-use efficiency (WUE), are stillunknown.
The knowledge about the role of indi-vidual genes in drought resistance is mainlyderived from genetic or transgenic studies. Anumber of genes that encode LEA proteins andenzymes involved in osmolyte synthesis or ROSremoval have been shown to improve droughtresistance when they are overexpressed in trans-genic plants (see Section “Genetic engineering ofcrops for adaptation to drought and heat stresses”of this chapter). The positive association of somedrought upregulated genes at the mRNA levelwith drought resistance has been observed in sev-eral crop species. For example, the levels of some

P1: SFK/UKS P2: SFK Color: 1C
BLBS082-27 BLBS082-Yadav July 12, 2011 17:16 Trim: 246mm X 189mm
526 CROP ADAPTATION TO CLIMATE CHANGE
LEA protein transcripts are positively associatedwith genotypic variation in drought resistance(Lopez et al. 2003). Comparative functional ge-nomic studies have shown at least a hundredgenes that are differentially expressed betweengenotypes with a contrasting drought-resistancetrait (Xue et al. 2006; Degenkolbe et al. 2009).Some of these differentially expressed genes areinvolved in the drought response, such as dehy-drin, DnaJ (molecular chaperone), and enzymesinvolved in proline synthesis (Xue et al. 2006;Schafleitner et al. 2007; Degenkolbe et al. 2009).However, many genes that are positively associ-ated with drought-resistance traits encode un-known proteins. This indicates that many morenovel genes related to drought resistance stillawait functional characterization.
Regulatory genes
Genotypic variation in drought resistance is oftenunderpinned by differences in expression levelsof multiple genes (often several gene networks).The requirement of simultaneous alterations inthe mRNA levels of multiple genes is obviousin views of known biochemical processes suchas ROS removal, which involves a number ofantioxidative enzymes (Møller et al. 2007). Theexpression of these genes involved in the samemetabolic or detoxification pathway is often con-trolled by a specific group of regulatory genes.Therefore, a regulatory gene can play a more im-portant role than a single stress protection genein drought resistance.
Regulatory genes that are involved in droughtresponse include: (1) signalling proteins (e.g.,protein kinases and phosphatases), (2) transcrip-tion factors (TFs) that are involved in tran-scriptional regulation through directly binding tocis-elements in promoters, (3) transcriptional co-factors that are capable of modifying or inter-acting with TFs, (4) proteins involved in post-transcriptional regulation (i.e., RNA processing,mRNA degradation, and translation), (5) proteinsinvolved in regulating the turnover rate of regula-tory proteins, and (6) enzymes involved in ABA
synthesis. This category should also includegenes for small interfering RNAs and microRNAs (Shukla et al. 2008) and genes involved inepigenetic regulation through DNA methylationor histone modification (Chinnusamy and Zhu2009). A large amount of information relatedto drought-induced gene regulation is coveredin recent reviews by Shinozaki and Yamaguchi-Shinozaki (2007) and Nakashima et al. (2009).
Evidence for the important contribution ofregulatory genes to drought resistance is de-rived mainly from genetic and transgenic studies.A large number of regulatory genes have beenshown to confer drought resistance in transgenicplants (see Section “Genetic engineering of cropsfor adaptation to drought and heat stresses”).
The molecular bases of heat adaptation
A comparative expression analysis between heat-and drought-responsive genes has revealed somecommon abiotic stress protection genes, butthe majority of the heat-responsive genes dif-fer from drought-responsive ones (Rizhsky et al.2004). This suggests that there are both com-mon and unique molecular mechanisms for heatand drought resistance. The common-responsivegenes cover most of those involved in ac-tive cellular stress protection, such as ROS re-moval, osmolyte synthesis, and some membersof HSPs/chaperones. However, plant adaptationto heat differs from drought in several aspectsas described earlier. This section will discuss thegenes encoding heat adaptation proteins under-lying biochemical and physiological attributes toheat resistance as well as known regulators con-trolling the expression of heat adaptation genes.
Heat adaptation genes
Heat resistance of plants can be altered bychanges in the expression of genes that areinvolved in cellular heat protection. This isparticularly evident for acquired thermotoler-ance through preexposure of plants to permis-sive temperature, during which a large number

P1: SFK/UKS P2: SFK Color: 1C
BLBS082-27 BLBS082-Yadav July 12, 2011 17:16 Trim: 246mm X 189mm
ADAPTATION TO WARMER AND DRIER CLIMATE 527
of heat protection genes are induced, such asHSPs/chaperones, enzymes for ROS, and os-molyte synthesis (Larkindale and Vierling 2008;Qin et al. 2008). A higher level accumulation ofthese gene transcripts during heat stress in heat-acclimated plants than nonacclimated ones is ac-companied with a better survival rate at subse-quent exposure to high temperatures (Larkindaleand Vierling 2008). The recent comparative ge-nomic studies also suggest that genotypic differ-ences in heat tolerance are generally associatedwith the higher mRNA levels of some criticalprotection genes (Zhang et al. 2005b; Qin et al.2008). Hsp70 in Festuca is one of the examples(Zhang et al. 2005b).
It is also perceivable that genotypes or specieswith genes encoding a biochemically superiorform of a critical stress protection or metabolicprotein can have some improvement of thermo-tolerance. One of the better-studied examplesis Rubisco activase. A 10◦C difference in theheat susceptibility of this enzyme was observedbetween creosote bush and Antarctic hairgrass(Salvucci and Crafts-Brandner 2004a). Otherperceivable superior forms of stress protectionproteins that are likely to exist are higher spe-cific activity or high substrate-binding affinity ofsome important enzymes such as those involvedin ROS removal. The superior thermostability ofcritical metabolic enzymes is likely to be moreimportant than their protein concentrations foreffective adaptation to heat stress. However, dis-covery in the superior forms of the genes at theprotein structure/function level is relatively slow,compared to the identification of genes based onthe expression level.
Regulatory genes
Similar to regulators involved in drought stress,a heat-responsive regulatory gene can play amore important role than a single heat protec-tion gene in contributing to heat-stress resistance.If the regulatory process of a gene has a linkto more than one of morphological, physiolog-ical, or biochemical mechanisms of heat resis-
tance, the gene could have a significant impactof heat resistance. Some members of HSPs suchas Hsp90 can modulate the activity of a num-ber of regulatory proteins in plants, leading to anumber of morpho-physiological changes suchas leaf shape and flowering time (Sangster et al.2007; Yamada et al. 2007).
Among regulators involved in heat-stress re-sponse in plants, heat shock factors (Hsf) havebeen well studied (von Koskull-Doring et al.2007). The Hsf family consists of 21 membersin Arabidopsis and >23 in rice (von Koskull-Doring et al. 2007). Although most Hsf genes areupregulated by heat, a few are regulated by ROS(Davletova et al. 2005), ABA (Kotak et al. 2007),or by both drought and heat (Schramm et al.2008). The downstream genes of Hsf proteinsinclude various HSPs/chaperones, antioxidativeenzymes, and enzymes involved in osmoprotec-tant synthesis (Busch et al. 2005; Nishizawa et al.2006; von Koskull-Doring et al. 2007). Severalstudies have shown that overexpression of someHsf genes or a TF upstream of an Hsf can im-prove heat resistance in transgenic plants (seeSection “Genetic engineering of crops for adap-tation to drought and heat stresses”). The Hsffamily is not the only TF group that is involvedin heat response in plants. A few genes fromother families have been shown to be upreg-ulated by heat, such as Arabidopsis DREB2A(AP2) and bZIP28 (Sakuma et al. 2006a; Gaoet al. 2008), and rice ZFP177 (C2H2 zinc fin-ger) and WRKY11 (Huang et al. 2008; Wu et al.2009). Some of these TFs have been shown tohave an important role in heat resistance fromtransgenic studies (see Section “Genetic engi-neering of crops for adaptation to drought andheat stresses”).
Simultaneous drought and heat stress
Many crops in the field often encounter simulta-neous drought and heat stresses at certain stagesof their growth period. Particularly, high tem-perature speeds up water evaporation from bothplant and soil and hence promotes or intensifies

P1: SFK/UKS P2: SFK Color: 1C
BLBS082-27 BLBS082-Yadav July 12, 2011 17:16 Trim: 246mm X 189mm
528 CROP ADAPTATION TO CLIMATE CHANGE
drought stress, which in turn increases leaf tem-perature due to the reduced rate of transpiration.A combination of drought and heat stresses hasa much greater detrimental effect on the growthand productivity of field crops than a single stress(Barnabas et al. 2008).
There are some unique features in the phys-iological response of plants to the combinationof drought and heat stresses. The most notableone is transpiration rate, which is higher in heat-stressed plants to cool the leaf, but becomeslower during the combined stresses (Rizhskyet al. 2002). The drought and heat stress com-bination can markedly reduce the ratio of photo-synthesis to respiration due to the reduced pho-tosynthesis combined with increased respiration(Mittler 2006). This is likely attributed to thecombination of several factors such as stomatalclosure, the reduced activation status of Rubisco,and differential changes in the relative solubilityof Rubisco substrates: CO2 and O2. High tem-perature leads to a more pronounced decreasein the solubility of CO2 than that of O2, thusfavoring Rubisco activity toward the photores-piration pathway (Rennenberg et al. 2006). Inaddition, simultaneous drought and heat stress islikely to exacerbate the reduction in the variouscomponents of photosynthesis including chloro-phyll and the Calvin cycle enzymes as observedin drought or heat stress.
Some metabolic changes in plants during ex-posure to the simultaneous drought and heatstress are also different from individual stresses.The simultaneous stress leads to an accumulationof sucrose, maltose, and glucose in Arabidopsis,but not proline (Rizhsky et al. 2004). Proteomicanalysis by Koussevitzky et al. (2008) identi-fied 45 proteins that specifically accumulated inArabidopsis, in response to the drought and heatstress combination. These specifically accumu-lated proteins include enzymes involved in ROSremoval, malate metabolism, and the Calvin cy-cle. Expression profiling analysis showed a newset of stress-responsive genes in plants exposedto the drought and heat stress combination, inaddition to stress-responsive genes that overlap
with those induced by either drought or heat(Rizhsky et al. 2004). However, in general, themajority of the biochemical and molecular mech-anisms that are involved in drought or heat adap-tation are used in plant response to the stresscombination.
Some common molecular mechanisms areused in both drought and heat adaptation. For ex-ample, some regulatory genes have been shownto be involved in both drought and heat stressessuch as DREB2A in Arabidopsis and maize(Sakuma et al. 2006a; Qin et al. 2007) andWRKY11 in rice (Wu et al. 2009). Over-expression of these TFs in transgenic plantsshowed improved resistance to both drought andheat stresses (see Section “Genetic engineer-ing of crops for adaptation to drought and heatstresses”). Improvement of crop yield under si-multaneous drought and heat stresses is chal-lenging, but it is also likely to attract the interestof more plant stress biologists in the future.
Drought and heat resistance inwild relatives of crop species
Genetic variation in many crop species is rela-tively limited due to the focus on yield poten-tial during the domestication of crop species. Alack of allelic variation will restrict the develop-ment of drought- and heat-resistant cultivars suit-able for future warmer and drier environmentsthrough traditional breeding. The WRs of cropspecies potentially provide a valuable source ofgenes and traits for crop improvement. WRs thatinhabit harsh environments are usually betteradapted to abiotic stress than cultivated species.However, crosses between crop species and WRsfrequently have a negative effect on crop yield inagricultural production environments.
A recent review by Hajjar and Hodgkin (2007)indicated that of 19 species surveyed, wild geneincorporation reaching the released cultivar stagewas found in 13. They found that more than 60wild species had contributed more than 100 ben-eficial traits to 13 crops. While the most commonuse of WRs in the past was as a source of pest and

P1: SFK/UKS P2: SFK Color: 1C
BLBS082-27 BLBS082-Yadav July 12, 2011 17:16 Trim: 246mm X 189mm
ADAPTATION TO WARMER AND DRIER CLIMATE 529
disease resistance, WRs have also contributed toabiotic stress-resistance traits.
Drought-resistance traits inwild relatives
Superior drought-resistance traits have beenidentified in WRs of many important cropspecies, such as rice (Liu et al. 2004), bar-ley (Nevo 2007), wheat (Reynolds et al. 2007),chickpea (Toker et al. 2007; Canci and Toker2009), potato (Coleman 2008), sunflower (Rauf2008), and groundnut (Nautiyal et al. 2008). Thedrought-resistance traits reported in or derivedfrom wild wheat species are high WUE (Nevo2007; Reynolds et al. 2007), early vigor (TerSteege et al. 2005), high productive tillers, highgrain weight, and better yield stability underwater-limited conditions (Baalbaki et al. 2006).In wild rice species, a number of useful traitsfor the improvement of upland rice droughtresistance have been observed, such as highWUE, greater membrane stability, increased rootbiomass, and better maintenance of leaf elonga-tion under drought stress (Liu et al. 2004). Thesuperior root traits such as deep root and highroot biomass have also been observed in WRsof chickpea (Gaur et al. 2008). One of the mostcommon drought-resistance traits is high WUE,which is also observed in WRs of other cropspecies such as wild potato (Coleman 2008) andwild sunflower (Rauf 2008). Other potentiallyuseful drought-resistance traits include a betterrecovery rate after severe drought stress in wildCicer species (Toker et al. 2007) and high harvestindex under drought in wild sunflower species(Rauf 2008). Some crop WRs such as Glycinelatifolia are able to tolerate a very low soil mois-ture and low leaf water content (McLean, James,and Xue, unpublished). The critical relative leafwater content of G. latifolia is about 23%, whilethat of the soybean cultivars is about 35%. Thecritical relative leaf water content is the thresholdat which a leaf is no longer able to be rehydrated.This cellular drought-tolerant property is rare incultivated species.
Only a small number of cultivars have beenreleased with abiotic stress resistance that hasbeen contributed by WRs (Hajjar and Hodgkin2007). WRs have contributed drought toleranceto released cultivars of tomato, chickpea, barley,rice, sorghum, sunflower, and wheat (Hajjar andHodgkin 2007).
Heat-resistance traits in wild relatives
Only a limited number of reports describe thecomparative analysis of heat resistance betweencrop species and their WRs. However, superiorheat resistance has been observed in some cropWRs. For example, Solanum gandarillasii ex-hibits remarkably higher membrane thermosta-bility than cultivated potato (Coleman 2008).Within wild species, a wide genetic variationfor heat resistance also exists among accessionsoriginated from a diverse range of environments(Nautiyal et al. 2008). Heat-resistance traits canalso be derived from inter- or intraspecific hy-brids such as synthetic wheats (Yang et al. 2002;van Ginkel and Ogbonnaya 2007). Some pri-mary synthetic wheats have higher chlorophyllcontent under high temperature growth condi-tions and more resistance to heat-induced ac-celeration of grain-filling duration than wheatcultivars (Yang et al. 2002). One interesting as-pect of heat-resistance genetics from crop WRsis that some WRs inhabiting hot environmentsmight provide superior alleles for thermostableproteins. A marked difference in the thermosta-bility of Rubisco activase has been observedbetween two C3 species inhabiting contrast-ing thermal environments (Salvucci and Crafts-Brandner 2004a). Khanna-Chopra and Sabari-nath (2004) have shown the remarkable ther-mostability of a chloroplastic Cu/Zn superoxidedismutase from Chenopodium murale that inhab-its the environment of up to 45◦C.
To date, there are few reports of released cul-tivars with heat tolerance derived from WRs.Hajjar and Hodgkin (2007) cite the release of achickpea cultivar with thermotolerance derivedfrom a related species.

P1: SFK/UKS P2: SFK Color: 1C
BLBS082-27 BLBS082-Yadav July 12, 2011 17:16 Trim: 246mm X 189mm
530 CROP ADAPTATION TO CLIMATE CHANGE
Molecular basis of drought andheat-resistance traits in wild relatives
Although a number of drought and heat-resistance traits have been characterized in cropWRs, the molecular basis for these traits isessentially unknown. The identification of crit-ical genes underpinning the trait can be ex-tremely useful for the introgression of stress-resistance genes from wild species into cropsby either marker-assisted breeding or transgenictechnology. Recent advance in the elucidationof molecular mechanisms of abiotic stressesand progress in functional genomics have pro-vided appropriate tools for the identification ofgenes underpinning these adaptive traits in WRs.Perceivably, two approaches can be usedfor identification of critical genes underlyingdrought and heat-resistance traits in WRs: analy-sis of known candidate genes and genome-widesearch (comparative functional genomics).
Candidate gene approach
This approach is based on the current knowledgeabout hundreds of stress adaptation and regula-tory genes that are involved in plant responseto drought and heat stresses. The effect of WRgenes on drought or heat resistance can be envis-aged as either a difference in protein sequence orthe concentration of a given protein that is oftenreflected by its mRNA level.
The comparative studies of gene expressionor sequences of drought and heat stress genesin WRs are relatively few to date. Suprunovalet al. (2004) have shown that some drought-upregulated genes such as dehydrins are ex-pressed higher in drought resistant genotypesof Hordeum spontaneum than susceptible ones.Chen et al. (2006) have shown that the mRNAlevel of a drought upregulated TF (DREB) ishigher in a drought-resistant Glycine soja thana sensitive soybean cultivar. We have also ob-served a few stress-protection genes (e.g., su-peroxide dismutase and dehydrin) that are ex-pressed at much higher levels in highly droughtresistant G. latifolia than two soybean cultivars
(McLean et al. unpublished). Few comparativestudies have been reported about allelic variationin the sequences of stress-protection proteins be-tween WRs and crop species. One example forthis is the observation of allelic variation in Dhn4between H. spontaneum and barley (Nevo 2007).
The candidate-gene approach has a seri-ous limitation, as the molecular basis of manydrought- or heat-resistance traits are still un-known even in crop species. A seemingly appar-ent trait can actually be a consequence of anothertrait or be linked to a change in an unknownbiochemical reaction. Therefore, genome-widesearch is generally more appropriate for the iden-tification of genes associated with the trait.
Comparative functional genomics
Comparative functional genomics is a rapidly ex-panding branch of functional genomics. With theavailability of large amounts of EST sequencesin major crop species (e.g., rice, wheat, and bar-ley) and large-scale expression profiling plat-forms (e.g., cDNA microarray and Affymetrixarrays), genome-wide searches for differentiallyexpressed genes between stress resistant and sus-ceptible genotypes has become feasible. To date,a number of comparative functional genomicstudies in this area are based on the identifi-cation of genes associated with genotypic vari-ation in drought or heat resistance (Schafleit-ner et al. 2007; Qin et al. 2008). There arealso some studies on a defined drought adap-tive trait, such as osmotic adjustment in rice(Hazen et al. 2005), transpiration efficiency inwheat (Xue et al. 2006), photosynthetic mainte-nance in potato plants under drought (Vasquez-Robinet et al. 2008), and stem water soluble car-bohydrates (WSC) in wheat (Xue et al. 2008b).
These types of studies provide a list of po-tential candidate genes that are associated witha trait for further investigation. Many of thesedifferentially expressed genes are only casuallyassociated with the trait, particularly when only afew genotypes are used for comparison. It shouldalso be noted that most of trait-associated genesidentified by expression analysis do not possess

P1: SFK/UKS P2: SFK Color: 1C
BLBS082-27 BLBS082-Yadav July 12, 2011 17:16 Trim: 246mm X 189mm
ADAPTATION TO WARMER AND DRIER CLIMATE 531
causal allelic variation in sequence, as geno-typic differences in the expression levels of manygenes are merely a reflection of allelic variationin their upstream regulator.
Comparative functional genomic studies be-tween a crop species and its WRs have not beenreported to date for several reasons. Firstly, genesequence information on WRs is scarce. Sec-ondly, the sequences of many gene transcriptsmay differ considerably between a crop speciesand its WR, which prevents a direct comparisonof expression levels between crop species and itsWR using the current genome array platformsbased on sequences from a crop species. Thirdly,genetic differences between a crop species andits WR are generally too large to pinpoint the as-sociation of differentially expressed genes witha drought or heat-resistance trait. This problemis also relevant to the comparison of expres-sion profiling between two unrelated genotypeswithin a crop species. The relevance of differ-entially expressed genes to a trait under studyincreases with the number of genotypes contrast-ing with the trait. Furthermore, precision in theidentification of the candidate genes can be im-proved by focusing on analysis of known func-tion genes involved in the process if the physi-ological and biochemical bases of the adaptivetrait are known. Such a case study is well illus-trated in the identification of genes associatedwith genotypic variation in the stem WSC by ex-amining the genes involved in WSC metabolicpathways (Xue et al. 2008b).
Introgression of drought- andheat-resistance genes fromwild relatives
A direct means of exploitation of a novel wildgene pool is to cross elite material with its WRthat is adapted to drought or heat stress envi-ronments. Introgression from compatible wildgermplasm has resulted in the release of somemore abiotic stress-resistant cultivars of severalcrop species (Hajjar and Hodgkin 2007). Moredrought- and heat-resistant lines have also beenreported in synthetic wheats derived from cross
between tetraploid wheat and Aegilops tauschii(Reynolds et al. 2007; Ortiz et al. 2008). Ex-ploitation of wild gene pools from sexually in-compatible distant WR species can be achievedby cell-fusion technique–somatic hybridization(Liu et al. 2005). Introgression of WR genes byinterspecific or intergeneric somatic hybridiza-tion has been achieved in many crop species,such as rice, wheat, Brassica, and potato (Liuet al. 2005; Liu et al. 2006; Zhao et al. 2008;Thieme et al. 2008). This technique providesa powerful means to further increase allelic di-versity in crop species. However, generation ofdrought- or heat-resistant somatic hybrids with-out compromising yield and quality traits isstill a challenging task for the future. Molecu-lar marker-assisted or genomic-guided selectiontools are likely to be necessary for acceleratingprogress in this area.
Genetic engineering of cropsfor adaptation to drought andheat stresses
The advantage of genetic engineering for im-provement of a plant trait or acquisition of anew trait is that any genetic modification canbe designed and tailored by introduction of oneor a few known gene(s). This is in contrast tothe conventional crossing or somatic hybridiza-tion, which has a severe drawback of uninten-tional introduction of a large chunk of unknownDNA with undesirable characteristics. Anotheradvantage of transgenic technology is its abilityto greatly amplify the expression (hundreds foldsif necessary) of the gene of interest in a trans-genic line compared to the parental genotype,while natural allelic variation in gene expressionlevels is generally within a few folds (Xue et al.2006, 2008b).
Despite the advantages, transgenic technol-ogy requires known candidate genes for thetraits. For drought- or heat-resistance traits, somecandidate genes underlying certain physiologicaland biochemical mechanisms have been identi-fied, such as genes encoding dehydrins, antiox-idant enzymes, or Rubisco activase. However,

P1: SFK/UKS P2: SFK Color: 1C
BLBS082-27 BLBS082-Yadav July 12, 2011 17:16 Trim: 246mm X 189mm
532 CROP ADAPTATION TO CLIMATE CHANGE
candidate genes for most drought- or heat-resistance traits are still unknown. In the pastdecades, many of drought- or heat-responsivegenes, particularly the upregulated ones, havebeen selected as potential candidates for func-tional evaluation in transgenic plants. In this sec-tion, we are going to examine the studies demon-strating improved resistance.
Enhanced drought resistance
Numerous studies have demonstrated the en-hanced drought resistance in transgenic plantsunder laboratory and greenhouse conditions. Anumber of recent reviews have covered thisarea (Shinozaki and Yamaguchi-Shinozaki 2007;Cattivelli et al. 2008; Gosal et al. 2009). Thegenes that have a positive effect on droughtresistance cover several groups as listed inTable 27.1.
Most of these genes are involved either in cel-lular stress protection or in drought-stress sig-naling and transcriptional activation for upreg-ulation of stress-protection genes. Some genesare known to be linked to specific physio-logical traits, such as AtMYB61 for reducedtranspiration by positively controlling stomatalclosure (Liang et al. 2005; Hu et al. 2006),NADP-malic enzyme for the improvement ofWUE through reduction of stomatal conduc-tance (Laporte et al. 2002), WXP1 for leaf waxdeposition which affects nonstomatal leaf wa-ter loss rate (Zhang et al. 2005a). Improve-ment of transpiration efficiency through re-duced stomatal conductance was also observedin transgenic peanut overexpressing ArabidopsisDREB1A (Bhatnagar-Mathur et al. 2007). In ad-dition to stress-upregulated genes, the improve-ment of drought resistance can also be achievedby nonstress genes such as isopentenyltrans-ferase which catalyses the rate-limiting step incytokinin synthesis (Rivero et al. 2007) and 14-3-3 protein GF14λ (Yan et al. 2004). The overex-pression of isopentenyltransferase in transgenictobacco delays leaf senescence (“stay green”like) and confers drought resistance (Rivero et al.
2007). The transgenic cotton with overexpres-sion of the Arabidopsis GF14λ also exhibits a“stay green”-like trait and the transgenic linesalso have a higher photosynthesis rate and aremore drought-resistant under water deficit condi-tions (Yan et al. 2004). The 14-3-3 protein familyis one group of regulatory proteins through inter-action with other proteins including TFs, antiox-idative enzymes, and HSPs. The improvementof WUE can be also achieved by the enhance-ment of carbon-fixation rate by overexpress-ing a C4-phosphoenolpyruvate carboxylase geneas shown in transgenic maize under moderatedrought conditions (Jeanneau et al. 2002). Re-cently, Yu et al. (2008) have shown that the rootsystem can be manipulated by overexpression ofa homeodomain-START TF. The overexpress-ing transgenic tobacco plants have a more exten-sive and deeper root system, as well as reducedleaf stomatal density and improved droughtresistance.
These transgenic studies have shed some lighton the genes underpinning some physiologicaland biochemical mechanisms of drought adapta-tion. However, most physiological adaptive traitssuch as stay green and WUE can arise from al-lelic variation in completely different genes. Notall these genes (possibly only a small portion)have been identified by genetic and transgenicstudies to date. Furthermore, the ultimate goalof crop improvement of drought resistance is toimprove yield in drought-prone environments.This requires not only the genes for combatingcellular stress, but also identifying genes under-pinning plant ability of extracting water in deepsoil (e.g., a deep root system) or better parti-tioning of assimilated carbon to grains. Genesassociated with some of these important drought-adaptive traits are still unknown. It is likely thatsome traits require manipulation of more thanone gene. Further scope for improvement of cropyield in drought-prone environment may lie inthe improvement of efficient use of energy un-der stress through metabolic engineering as pro-posed by Dobrota (2006). The outcome of thistype of genetic manipulation may be seen as a

P1: SFK/UKS P2: SFK Color: 1C
BLBS082-27 BLBS082-Yadav July 12, 2011 17:16 Trim: 246mm X 189mm
ADAPTATION TO WARMER AND DRIER CLIMATE 533
Table 27.1. Transgenes confer drought resistance.
Protein group encoded by transgene Expression mode Examples of transgene
Late embryogenesis proteins Over-expression (OE) Barley HVA1 (Bahieldin et al. 2005) andrice LEA3 (Xiao et al. 2007)
Heat shock proteins/chaperones OE HSP70-1 (Cho and Hong 2006) and BiP(Alvim et al. 126)
Enzymes for the synthesis ofosmoprotectants
OE Mannitol-1-phosphate dehydrogenase(Abebe et al. 2003) and�1-pyrroline-5-carboxylate synthetase(Kavi Kishor et al. 1995)
Antioxidative enzymes for removal ofreactive oxygen species
OE Mn-superoxide dismutase (McKersie et al.1996) and antiquitin-like protein(antiquitin catalyses acetaldehydeoxidation) (Rodrigues et al. 2006)
Enzymes for polyamine synthesis OE Arginine decarboxylase (Capell et al.,2004) and S-adenosylmethioninedecarboxylase (Waie & Rajam 2003)
TFs involved in drought response OE DREB2A (Sakuma et al. 2006b) and ABF3(Oh et al. 2005)
TFs involved in cold response OE DREB1A/CBF3 (Kasuga et al. 1999)
TFs controlling leaf cuticular waxproduction
OE WXP1 (Zhang et al. 2005a)
TFs involved in regulation of stomatalconductance
OE AtMYB61 (Liang et al. 2005), SNAC1 (Huet al. 2006) and AtMYB44 (Jung et al.2008)
Protein kinases or phosphatase involved instress signalling
OE NPK1 (Shou et al. 2004), OsCDPK7 (Saijoet al. 2000), and protein phosphatase 2a(Xu et al. 2007)
Enzymes for ABA synthesis OE AtNCED3 (Iuchi et al. 2001)
Farnesyltransferases involved in regulationof ABA sensing
Repression ERA1 (Wang et al. 2005) and FTA (Wanget al. 2009)
Regulatory proteins not involved indrought response
OE Isopentenyltransferase (Rivero et al. 2007),14-3-3 protein GF14� (Yan et al. 2004)and homeodomain-START TF(Yu et al.2008)
reduction in respiration rate, consequently im-proving carbon-use efficiency and WUE.
Another underexploited area is to use supe-rior alleles from drought-resistant wild or ex-tremophilic species that inhabit extreme droughtenvironments. However, this requires priormolecular/biochemical studies of these genesto characterize their superior function, such asbetter enzymatic properties or stronger regula-tory activity.
It should be noted that not all of thesetransgenic experiments resulted in a yield
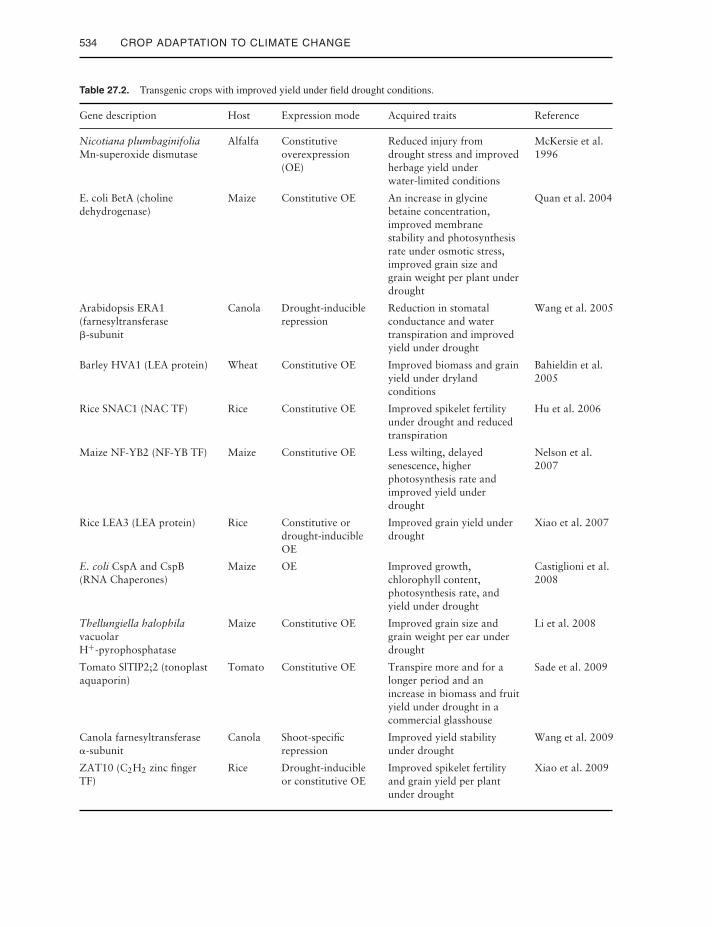
advantage under field drought conditions orhave been tested in a range of relevant fieldenvironments. In fact, many overexpressingtransgenic plants showed a slow growth rate(Kasuga et al. 1999; Iuchi et al. 2001), whichis likely to lead to lower crop productivity undernormal growth conditions. Increasingly, trans-genic experiments are being undertaken in fieldconditions to obtain a more realistic and rele-vant evaluation of the performance of the trans-gene; those undertaken to date are listed inTable 27.2.
P1: SFK/UKS P2: SFK Color: 1C
BLBS082-27 BLBS082-Yadav July 12, 2011 17:16 Trim: 246mm X 189mm
534 CROP ADAPTATION TO CLIMATE CHANGE
Table 27.2. Transgenic crops with improved yield under field drought conditions.
Gene description Host Expression mode Acquired traits Reference
Nicotiana plumbaginifoliaMn-superoxide dismutase
Alfalfa Constitutiveoverexpression(OE)
Reduced injury fromdrought stress and improvedherbage yield underwater-limited conditions
McKersie et al.1996
E. coli BetA (cholinedehydrogenase)
Maize Constitutive OE An increase in glycinebetaine concentration,improved membranestability and photosynthesisrate under osmotic stress,improved grain size andgrain weight per plant underdrought
Quan et al. 2004
Arabidopsis ERA1(farnesyltransferase�-subunit
Canola Drought-induciblerepression
Reduction in stomatalconductance and watertranspiration and improvedyield under drought
Wang et al. 2005
Barley HVA1 (LEA protein) Wheat Constitutive OE Improved biomass and grainyield under drylandconditions
Bahieldin et al.2005
Rice SNAC1 (NAC TF) Rice Constitutive OE Improved spikelet fertilityunder drought and reducedtranspiration
Hu et al. 2006
Maize NF-YB2 (NF-YB TF) Maize Constitutive OE Less wilting, delayedsenescence, higherphotosynthesis rate andimproved yield underdrought
Nelson et al.2007
Rice LEA3 (LEA protein) Rice Constitutive ordrought-inducibleOE
Improved grain yield underdrought
Xiao et al. 2007
E. coli CspA and CspB(RNA Chaperones)
Maize OE Improved growth,chlorophyll content,photosynthesis rate, andyield under drought
Castiglioni et al.2008
Thellungiella halophilavacuolarH+-pyrophosphatase
Maize Constitutive OE Improved grain size andgrain weight per ear underdrought
Li et al. 2008
Tomato SlTIP2;2 (tonoplastaquaporin)
Tomato Constitutive OE Transpire more and for alonger period and anincrease in biomass and fruityield under drought in acommercial glasshouse
Sade et al. 2009
Canola farnesyltransferase�-subunit
Canola Shoot-specificrepression
Improved yield stabilityunder drought
Wang et al. 2009
ZAT10 (C2H2 zinc fingerTF)
Rice Drought-inducibleor constitutive OE
Improved spikelet fertilityand grain yield per plantunder drought
Xiao et al. 2009

P1: SFK/UKS P2: SFK Color: 1C
BLBS082-27 BLBS082-Yadav July 12, 2011 17:16 Trim: 246mm X 189mm
ADAPTATION TO WARMER AND DRIER CLIMATE 535
Table 27.2. (Continued)
Gene description Host Expression mode Acquired traits Reference
NPK1 (protein kinase) Rice Drought-inducibleor constitutive OE
Improved spikelet fertilityand grain yield per plantunder drought
Xiao et al. 2009
CBF3 (C-repeat binding TF) Rice Drought-inducibleOE
Improved spikelet fertilityand grain yield per plantunder drought
Xiao et al. 2009
LOS5 (molybdenumcofactor sulfurase)
Rice Drought-inducibleor constitutive OE
Improved spikelet fertilityand grain yield per plantunder drought
Xiao et al. 2009
Enhanced heat resistance
Many transgenic studies documenting the im-provement of heat resistance have been coveredin a recent review by Singh and Grover (2008). Ingeneral, transgenes that have been shown to con-fer heat resistance cover the following groups(Table 27.3): (1) HSPs/chaperones, (2) antiox-idative enzymes, (3) enzymes affecting the sat-uration of membrane fatty acids, (4) enzymes
involved in photosynthesis, (5) enzymes in-volved in osmoprotectant synthesis, (6) enzymesinvolved in polyamine synthesis, (7) Hsf, and(8) other regulators which control heat-shock re-sponse or antioxidative enzymes. In addition, arecent study has shown that the overexpression ofa cold-inducible TF (AtDREB1A) in Chrysan-themum also confers heat resistance and in-duce heat upregulated genes in transgenic plants(Hong et al. 2009).
Table 27.3. Transgenes confer heat resistance.
Protein group Expression mode Examples of transgenic studies
Heat shock proteins(HSP)/chaperones
Overexpression (OE) Hsp17.7 (Malik et al. 1999; Sato and Yokoya, 2008) andchaperones (DjA2 and DjA3) (Li et al. 2007)
Antioxidative enzymes OE Ascorbate peroxidase (Tang et al. 2006) and superoxidedismutase (Tang et al. 2006)
Enzymes affecting thesaturation of membranefatty acids
Repression or OE Repression of �-3-fatty acid desaturase (Murakami et al.2000; Sohn and Back 2007) and over-expression ofglycerol-3-phosphate acyltransferase (Yan et al. 2008)
Enzymes involved inphotosynthesis
OE Thermostable Rubisco activase (Kurek et al. 2007) andSedo-heptulose-1,7-bisphosphatase (Feng et al. 2007)
Enzymes involved in thesynthesis of osmoprotectants
OE Betaine aldehyde dehydrogenase (Yang et al. 2005) andcholine oxidase (Alia et al. 1998)
Enzymes involved inpolyamine synthesis
OE S-adenosylmethionine decarboxylase (Cheng et al. 2009)
Heat shock factor (Hsf) TFfamily
OE Hsf1 (Lee et al. 1995) and HSFA2e (Yokotani et al. 227)
Other TF families regulatingheat shock response
OE WRKY11 (Wu et al. 2009), DREB2C (Lim et al. 2007),and ZFP177 (Huang et al. 2008)
Other regulatory genes OE Multiprotein bridging factor 1c (Suzuki et al. 2008) andnucleotide diphosphate kinase 2 (Tang et al. 2008)

P1: SFK/UKS P2: SFK Color: 1C
BLBS082-27 BLBS082-Yadav July 12, 2011 17:16 Trim: 246mm X 189mm
536 CROP ADAPTATION TO CLIMATE CHANGE
All these transgenes appear to have knownphysiological and biochemical bases for im-proving heat resistance. However, the molecularmechanisms of these regulatory genes includingTFs and some HSPs/chaperones in heat resis-tance are not fully understood yet. Despite manyheat-resistant transgenic plants that have beengenerated, no studies describing the field per-formance of the transgenic plants have been re-ported to date. Furthermore, some specific issuesrelated to heat resistance at the reproductive stageof temperate cereals have not been addressed bythe current transgenic studies. Improvement ofheat resistance at the vegetative stage by thesegenes may not solve the yield problem at thereproductive stage of heat stress for some cropspecies. For example, soluble starch synthase inwheat endosperm is markedly inhibited at tem-peratures above 30◦C (Keeling et al. 1993). An-other detrimental effect of heat stress at the re-productive stage on the grain yield and weight isthrough shortening of the grain-filling duration(Barnabas et al. 2008). The grain-filling dura-tion in some heat-resistant genotypes appears tobe less sensitive to high temperatures. Gene thatcan minimize the effect of heat on the grain-filling duration and improve the thermostabilityof enzymes involved in starch synthesis shouldbe useful.
Enhanced drought and heat resistance
As drought and heat stresses often occur con-currently in the field conditions for many cropproduction regions, global warming further stim-ulates the demand for breeding crop varietieswith the improvement of both drought and heatresistance. Most of transgenic studies focus ongenes involved in a single stress to date. Re-cently, a number of reports describe the resis-tance of transgenic plants to both drought andheat stresses and in some cases to simultaneousdrought and heat stress (Table 27.4).
Many of these genes in Table 27.4 are up-regulated under both drought and heat condi-tions, indicating their role in adaptation to these
stresses. A single transgene that has an effecton both drought and heat resistance in mostcases appears to have a regulatory role, includingtrehalose-6-phosphate synthase and trehalose-6-phosphate phosphatase, which are involved insugar sensing and signaling as well as regu-lating plant architecture and cell shape (Charyet al. 2008). The overexpression of these reg-ulatory genes upregulates an array of drought-and heat-inducible genes (Sakuma et al. 2006a,2006b; Qin et al. 2007). Besides the regulatorygenes, enhancing the antioxidant capacity alsoimproves both drought and heat resistance (DeRonde et al. 2004; Kocsy et al. 2005; Tognettiet al. 2006), suggesting an important role of an-tioxidative capacity in both drought and heat re-sistance. Improvement of both drought and heatresistance by a single transgene is scientificallyinteresting. However, whether these genes rep-resent better genes for the improvement of cropyield stability under drought- and heat-prone en-vironments awaits field evaluation.
Enhanced stress resistance linked toyield improvement
Although numerous transgenic studies havedemonstrated the improvement of stress resis-tance, many drought- or heat-resistant transgeniclines with constitutive overexpression of a trans-gene exhibit a slower rate of growth under non-stress conditions as mentioned in the previoussection. We have also noticed a reduction in thegrain weight of transgenic wheat plants consti-tutively overexpressing a drought upregulatedTF at a high level under well-watered condi-tions, even though the growth rate of transgenicplants at the vegetative stage is not impaired (Xueet al., unpublished data). Field trials have alsoshown that some trangenes tend to have a nega-tive effect on grain yield under normal growthconditions (Xiao et al. 2009; Bahieldin et al.2005). This phenomenon can theoretically resultfrom the following two causes: (1) genes thatare induced during stress generally have a nega-tive impact on the growth and yield and (2) the

P1: SFK/UKS P2: SFK Color: 1C
BLBS082-27 BLBS082-Yadav July 12, 2011 17:16 Trim: 246mm X 189mm
ADAPTATION TO WARMER AND DRIER CLIMATE 537
Table 27.4. Transgenic plants conferring drought and heat resistance.
Gene description Expression mode Host species Transgenic phenotypes Reference
OsWRKY11(a drought and heatupregulatedWRKY TF)
Over-expression(OE)
Rice The slower leaf wiltingand higher survivalrate undersimultaneous droughtand heat. A slower rateof water loss indetached leaves.
Wu et al. 2009
Arabidopsis ABF3(an ABA upregulatedbZIP TF)
OE Arabidopsis andtobacco
A high survival rateunder either droughtor heat stress. A slowerrate of water loss fromplants under waterdeprivation.
Kim et al. 2004
Arabidopsis ABF3(an ABA upregulatedbZIP TF)
OE Agrostis mongolica A small stomatalaperture size and highleaf water contentunder drought. A highsurvival rate undereither drought or heat
Vanjidorj et al.2006
Arabidopsis ABF9(an ABA upregulatedbZIP TF)
OE Arabidopsis An improvedphotosynthesis rateunder simultaneousdrought and heat
Zhang et al. 2008
Maize DREB2A(a drought and heatupregulated AP2 TF)
OE Arabidopsis A slower rate of waterloss from plants underdrought and a highersurvival rate underdrought or heat
Qin et al. 2007
Arabidopsis DREB2A(a drought & heatupregulated AP2 TF)
OE Arabidopsis A slower rate of waterloss from plants underdrought and a highersurvival rate underdrought or heat
Sakuma et al.2006a, 2006b
Capsicum CaPF1(an AP2 TF)
OE Potato A higher survival rateunder drought or heat
Youm et al. 2008
A chimeric protein ofyeast trehalose-6-Psynthase &trehalose-6-Pphosphatase
OE Arabidopsis A slower rate of waterloss from plants underdrought and a highersurvival rate underdrought or heat
Miranda et al. 2007
Bacillus CspB (RNAchaperone)
OE Rice Improved growth rateunder heat or drought
Castglioni et al.2008
Poly(ADP-ribose)polymerase
OE Brassica napus &Arabidopsis
A higher survival rateand higher biomassunder simultaneousdrought and heat
De Block et al. 2005

P1: SFK/UKS P2: SFK Color: 1C
BLBS082-27 BLBS082-Yadav July 12, 2011 17:16 Trim: 246mm X 189mm
538 CROP ADAPTATION TO CLIMATE CHANGE
Table 27.4. (Continued)
Gene descriptionExpressionmode Host species Transgenic phenotypes Reference
�-pyrroline-5-carboxylate reductaseinvolved in prolinesynthesis
OE Soybean Reduced injury andhigher leaf watercontent and anenhanced antioxidativecapacity undersimultaneous droughtand heat
De Ronde et al.2004; Kocsy et al.2005
Cyanobacterialferredoxin
OE Tobacco Reduced injury and anenhanced antioxidativecapacity under heat, alower rate of plantwater loss underdrought
Tognetti et al. 2006
energetic cost of transgene expression or its cat-alytic products. The first instance is typified bydrought upregulated regulatory genes such asArabidopsis DREB2A (Kasuga et al. 1999). Re-duction in growth during drought stress is a ben-eficial physiological process for plant survivalunder stress conditions, which allows plants toredistribute water, carbon, and energy for stressprotection as well as effective accumulation ofthe high osmolyte concentrations through a re-duced rate of dilution by cell expansion. For thesecond cause, the mRNA and protein synthesis ofa transgene or enhanced osmolyte accumulationby overexpression of an osmolyte-synthetic en-zyme requires extra carbon and energy. However,if the expression level of a transgene is low and itdoes not impact the expression of other genes, thecarbon and energy cost may not be significant. Ineither case, the expression of a transgene needsto be tailored to meet the requirement for plantstress adaptation if the crop yield is concerned.Any reduction of crop yield under normal growthconditions could potentially override a markedyield advantage under stress. This is because thecrop productivity during good years representsfarmers’ major income in the most crop produc-tion regions.
The expression of a transgene can be tai-lored to stress adaptation by using a drought-or
heat-inducible promoter for driving the ex-pression of a stress-responsive gene. Forexample, transgenic Arabidopsis plants carryinga drought-inducible, promoter-driven DREB2Aexhibit the improved drought resistance with nosignificant difference in growth rate under nor-mal growth conditions (Sakuma et al. 2006b).Similarly, an oxidative stress-inducible SWPA2promoter has successfully been used for driv-ing the overexpression of Arabidopsis nucleotidediphosphate kinase 2 gene in potato for improve-ment of heat and drought tolerance (Tang et al.2008). Other aspects for consideration of min-imizing the negative impact of transgene ex-pression on growth and yield include the ap-propriate expression level of a transgene andcell specificity. In most cases, the expressionof a stress-protection gene should be avoidedin the endosperm of cereal crops or the tubesof potato.
Concluding remarks
Breeding crop varieties for adaptation to warmerand drier climate relies on our understand-ing of drought and heat-resistance mechanismsand their impacts on crop productivity. Sub-stantial progress in unveiling these resistancemechanisms among and within plant species

P1: SFK/UKS P2: SFK Color: 1C
BLBS082-27 BLBS082-Yadav July 12, 2011 17:16 Trim: 246mm X 189mm
ADAPTATION TO WARMER AND DRIER CLIMATE 539
has been made through extensive physiological,biochemical, and molecular studies over the pasttwo decades. Some improvement of drought orheat resistance can now be achieved through en-hancing the capacity of individual mechanismsby genetic manipulation of a critical underlaingene. However, drought and heat-induced cellu-lar injuries in plants are multifacets and there ex-ist multiple weak physiological and biochemicalcomponents in current crop varieties in combat-ing drought and heat stresses. Hence, simulta-neous enhancement of several stress-protectionmechanisms is required in order to achieve sub-stantial improvement of crop adaptive capacity.Natural variation in drought or heat resistanceobserved among plant species generally adoptsmultiple resistance mechanisms and multiplegene networks. In particular, global warmingwill lead to more frequent simultaneous droughtand heat stress events in the future agriculturalenvironments. Therefore, enhancement of bothdrought and heat resistant capacities needs to beconsidered for breeding future adaptive crop va-rieties, which would exclude some mechanismsthat are beneficial to one stress and undesirablefor the other. For example, lowing stomatal con-ductance is a most effective mean for enhancingdrought resistance, but reduces the leaf coolingcapacity during heat stress. On other hand, somecommon stress avoidance and protection mech-anisms such as a deep root system, and an an-tioxidant capacity can be used for improvementof both drought and heat adaptation.
Improvement of crop productivity in drought-and heat-prone environments still remains achallenging task. It requires two seeminglyopposing physiological processes: enhancingstress-resistant capacity and maximizing carbonassimilation. Enhancement of stress-protectioncapacity or deep-soil water extraction capac-ity generally leads to delayed stress-inducedleaf senescence and maintaining better photo-synthesis capacity and water/carbon/energy useefficiencies under drought and heat stress condi-tions. However, some drought-resistance mech-anisms such as low stomatal conductance or
slow growth rate generally result in a reduc-tion in carbon assimilation and hence cropproductivity. Even a big root system demandsan extra amount of carbon and would reduceabove-ground biomass growth in favorable en-vironments if the carbon assimilation capac-ity is not improved. Similarly, improvement ofdrought and heat resistance using the stress es-cape mechanism (early phenology) would alsoreduce yield potential in many crop speciesif the growth rate is not enhanced. Therefore,the manipulation of stress-resistant capacity isonly one side of the story for improvementof yield in the future cropping environments.Simultaneous enhancement of stress-resistantcapacity and carbon assimilation should be con-sidered for future breeding, as this might coun-teract some negative effects of enhanced stress-resistant capacity on plant productivity underfavorable conditions. This might be achieved bygenetic manipulation of multiple genes to en-hance both stress-resistant capacity and carbonassimilation efficiency. Some genes that havedemonstrated to improve stress resistance or car-bon assimilation in transgenic studies may bepyramided into a single line to examine theireffects on yield in drought- and heat-prone en-vironments. In some crop species, enhancementof temporary carbon reserve capacity can signif-icantly improve yield in drought and heat-proneenvironments, such as stem WSC in temperatecereals, which is an important carbon source forgrain filling, particularly under terminal droughtand heat stresses.
Tailoring stress resistance also depends oncrop species and their cropping environments aswell as the timing and extent of expression ofindividual resistant capacities and the develop-mental stages of crops. Improvement of stresssurvival by reducing the number of stomata andtightly closing stomata or shedding old leaves isuseful for crop adaptation to the stress at thevegetative stage, but is not suitable for manycrops, particularly those which grow in terminaldrought and heat stress environments. Althoughconstitutive reduction in the stomatal number and

P1: SFK/UKS P2: SFK Color: 1C
BLBS082-27 BLBS082-Yadav July 12, 2011 17:16 Trim: 246mm X 189mm
540 CROP ADAPTATION TO CLIMATE CHANGE
conductance is undesirable for crop productiv-ity, a certain degree of enhancement of stress-inducible reduction of stomatal conductance thatmatches with root water uptake might be bene-ficial for improvement of yield in drought- andheat-prone environments. Several genes that un-derlie the stomatal conductance trait have beenidentified. The current transgenic technology iscapable of the manipulation of stomatal conduc-tance by controlling both the expression leveland drought-driven induction. However, the ap-propriate level and timing of this modificationneed to be verified through many years of fieldtrials in the whole system, which includes naturalcropping environments with both good and badyears and the genetic background of the genotypeinto which the new trait is introduced. Similarly,a drought-inducible, strong, and deep root sys-tem is also likely to be better than the constitutiveone in term of crop productivity.
The most challenging task is to improve yieldunder simultaneous drought and heat stress atthe reproductive stage. A substantial improve-ment requires enhancing the thermostability ofmany critical proteins involved in photosynthe-sis, grain starch synthesis, or other biologicalprocesses that might be thermolabile, besidesgeneral drought and heat stress avoidance andprotection mechanisms. Pyramiding of multiplegenes required for achieving substantial resis-tance to simultaneous stresses without affectingyield potential is still a formidable task throughgenetic engineering at the present. In some cir-cumstances, if a WR that is adapted to an ex-tremely dry and hot environment is available,introgression of stress-resistant genes from thewild species may provide a better option forbringing all these necessary genes into a newcrop variety. Modern molecular technologies in-cluding comparative functional genomics canprovide powerful tools in identification of allelicgenes associated with traits or screening the traitsin breeding populations at individual gene levels.In addition, further investigation into drought andheat-resistance mechanisms and genes underpin-ning individual resistance traits is likely to pro-
vide us better knowledge to tackle this complexdrought and heat problem in the future.
Acknowledgment
This work was supported in part by a grant fromAustralian Grains Research and DevelopmentCorporation.
References
Abebe T, Guenzi AC, Martin B et al. (2003) Tolerance ofmannitol accumulating transgenic wheat to water stressand salinity. Plant Physiology 131: 1748–1755.
Alia, Hayashi H, Sakamoto A et al. (1998) Enhancementof the tolerance of Arabidopsis to high temperatures bygenetic engineering of the synthesis of glycinebetaine.The Plant Journal 16: 155–161.
Alvim EC, Carolino SMB, Cascardo JCM et al. (2001) En-hanced accumulation of BiP in transgenic plants con-fers tolerance to water stress. Plant Physiology 126:1042–1054.
Baalbaki R, Hajj-Hassan N, Zurayk R (2006) Aegilopsspecies from semiarid areas of Lebanon: Variation inquantitative attributes under water stress. Crop Science46: 799–806.
Bahieldin A, Mahfouz HT, Eissa HF et al. (2005) Field eval-uation of transgenic wheat plants stably expressing theHVA1 gene for drought tolerance. Physiologia Plantarum123: 421–427.
Barnabas B, Jager K, Feher A (2008) The effect of droughtand heat stress on reproductive processes in cereals. PlantCell and Environment 31: 11–38.
Bartels D, Sunkar R (2005) Drought and salt tolerance inplants. Critical Reviews in Plant Sciences 24: 23–58.
Bhatnagar-Mathur P, Devi MJ, Reddy DS et al. (2007) Stressinducible expression of AtDREB1A in transgenic peanut(Arachis hypogaea) increases transpiration efficiency.Plant Cell Reports 26: 2071–2082.
Blum A (2005) Drought resistance, water-use efficiency, andyield potential—are they compatible, dissonant, or mu-tually exclusive? Australian Journal of Agricultural Re-search 56: 1159–1168.
Busch W, Wunderlich M, Schoffl F (2005) Identification ofnovel heat shock factor-dependent genes and biochemicalpathways in Arabidopsis thaliana. The Plant Journal 41:1–14.
Canci H, Toker C (2009) Evaluation of annual wild Cicerspecies for drought and heat resistance under field con-ditions. Genetic Resources and Crop Evolution 56: 1–6.
Capell T, Bassie L, Christou P (2004) Modulation of thepolyamine biosynthetic pathway in transgenic rice con-fers tolerance to drought stress. Proceedings of the Na-tional Academy of Sciences USA 101: 9909–9914.

P1: SFK/UKS P2: SFK Color: 1C
BLBS082-27 BLBS082-Yadav July 12, 2011 17:16 Trim: 246mm X 189mm
ADAPTATION TO WARMER AND DRIER CLIMATE 541
Castiglioni P, Warner D, Bensen RJ et al. (2008) BacterialRNA chaperones confer abiotic stress tolerance in plantsand improved grain yield in maize under water-limitedconditions. Plant Physiology 147: 446–455.
Cattivelli L, Rizza F, Badeck FW et al. (2008) Drought tol-erance improvement in crop plants: An integrated viewfrom breeding to genomics. Field Crops Research 105:1–14.
Chary SN, Hicks GR, Choi YG et al. (2008) Trehalose-6-phosphate synthase/phosphatase regulates cell shape andplant architecture in Arabidopsis. Plant Physiology 146:97–107.
Chen Y, Chen P, de los Reyes BG (2006) Differential re-sponses of the cultivated and wild species of soybean todehydration stress. Crop Science 46: 2041–2046.
Cheng L, Zou Y, Ding S et al. (2009) Polyamine accumula-tion in transgenic tomato enhances the tolerance to hightemperature stress. Journal of Integrative Plant Biology51: 489–499.
Chinnusamy V, Zhu JK (2009) Epigenetic regulation of stressresponses in plants. Current Opinion in Plant Biology 12:133–139.
Cho EK, Hong CB (2006) Over-expression of tobaccoNtHSP70-1 contributes to drought-stress tolerance inplants. Plant Cell Reports 25: 349–358.
Coleman WK (2008) Evaluation of wild Solanum speciesfor drought resistance: 1. Solanum gandarillasii Car-denas. Environmental and Experimental Botany 62:221–230.
Davletova S, Rizhsky L, Liang H et al. (2005) CytosolicAscorbate Peroxidase 1 is a central component of thereactive oxygen network of Arabidopsis. The Plant Cell17: 268–281.
De Block M, Verduyn C, de Brouwer D et al. (2005)Poly(ADP-ribose) polymerase in plants affects energyhomeostasis, cell death and stress tolerance. The PlantJournal 41: 95–106.
De Ronde JA, Cress A, Kruger GHJ et al. (2004) Photo-synthetic response of transgenic soybean plants contain-ing an Arabidopsis P5CR gene, during heat and droughtstress. Journal of Plant Physiology 61: 1211–1244.
Degenkolbe T, Do PT, Zuther E et al. (2009) Expressionprofiling of rice cultivars differing in their tolerance tolong-term drought stress. Plant Molecular Biology 69:133–153.
Dobrota C (2006) Energy dependant plant stress acclimation.Reviews in Environmental Science and Biotechnology 5:243–251.
Feng L, Wang K, Li Y et al. (2007). Overexpression ofSBPase enhances photosynthesis against high temper-ature stress in transgenic rice plants. Plant Cell Reports26: 1635–1646.
Gao H, Brandizzi F, Benning C et al. (2008) A membrane-tethered transcription factor defines a branch of theheat stress response in Arabidopsis thaliana. Proceed-ings of the National Academy of Sciences U.S.A. 105:16398–16403.
Gaur PM, Krishnamurthy L, Kashiwagi J (2008) Improvingdrought-avoidance root traits in chickpea (Cicer ariet-inum L.)—Current Status of Research at ICRISAT. PlantProduction Science 11: 3–11.
Gosal SS, Wani SH, Kang MS (2009) Biotechnology andDrought Tolerance. Journal of Crop Improvement 23:19–54.
Greene TW, Hannah LC (1998) Enhanced stability of maizeendosperm ADP-glucose pyrophosphorylase is gainedthrough mutants that alter subunit interactions. Proceed-ings of the National Academy of Sciences U.S.A. 95:13342–13347.
Hajjar R, Hodgkin T (2007) The use of wild relatives in cropimprovement: A survey of developments over the last 20years. Euphytica 156: 1–13.
Hazen SP, Pathan MS, Sanchez A et al. (2005) Expressionprofiling of rice segregating for drought tolerance QTLsusing a rice genome array. Functional and IntegrativeGenomics. 5: 104–116.
Hong B, Ma C, Yang Y et al. (2009) Over-expression ofAtDREB1A in Chrysanthemum enhances tolerance toheat stress. Plant Molecular Biology 70: 231–240.
Hu H, Dai M, Yao J et al. (2006) Overexpressing a NAM,ATAF, and CUC (NAC) transcription factor enhancesdrought resistance and salt tolerance in rice. Proceed-ings of the National Academy of Sciences U.S.A. 103;12987–12992.
Huang J, Wang MM, Jiang Y et al. (2008) Expression analysisof rice A20/AN1-type zinc finger genes and characteri-zation of ZFP177 that contributes to temperature stresstolerance. Gene 420: 135–144.
Iuchi S, Kobayashi M, Taji T et al. (2001) Regulationof drought tolerance by gene manipulation of 9-cis-epoxycarotenoid dioxygenase, a key enzyme in abscisicacid biosynthesis in Arabidopsis. The Plant Journal 27:325–333.
Jeanneau M, Gerentes D, Foueillassar X et al. (2002) Im-provement of drought tolerance in maize: Towards thefunctional validation of the Zm-Asr1 gene and increaseof water use efficiency by over-expressing C4-PEPC.Biochimie 84: 1127–1135.
Jung C, Seo JS, Han SW et al. (2008) Overexpression ofAtMYB44 enhances stomatal closure to confer abioticstress tolerance in transgenic Arabidopsis. Plant Physi-ology 146: 623–635.
Kasuga M, Liu Q, Miura S et al. (1999) Improving plantdrought, salt, and freezing tolerance by gene transferof a single stress inducible transcription factor. NatureBiotechnology 17: 287–291.
Kavi Kishor PB, Hong Z, Miao G-H et al. (1995) Over-expression of �‘-pyrroline 5-carboxylate synthetase in-creases proline production and confers osmotolerance intransgenic plants. Plant Physiology 108: 1387–1394.
Keeling PL, Bacon PJ, Holt DC (1993) Elevated temperaturereduces starch deposition in wheat endosperm by reduc-ing the activity of soluble starch synthase. Planta 191:342–348.

P1: SFK/UKS P2: SFK Color: 1C
BLBS082-27 BLBS082-Yadav July 12, 2011 17:16 Trim: 246mm X 189mm
542 CROP ADAPTATION TO CLIMATE CHANGE
Khanna-Chopra R, Sabarinath S (2004) Heat-stable chloro-plastic Cu/Zn superoxide dismutase in Chenopodium mu-rale, Biochemical and Biophysical Research Communi-cations 320: 1187–1192.
Kim JB, Kang JY, Kim SY (2004) Over-expression of atranscription factor regulating ABA-responsive gene ex-pression confers multiple stress tolerance. Plant Biotech-nology Journal 2: 459–466.
Kocsy G, Laurie R, Szalai G et al. (2005) Genetic manipula-tion of proline levels affects antioxidants in soybean sub-jected to simultaneous drought and heat stresses. Physi-ologia Plantarum 124: 227–235.
Kotak S, Larkindale J, Lee U et al. (2007) Complexity of theheat stress response in plants. Current Opinion in PlantBiology 10: 310–316.
Koussevitzky S, Suzuki N, Huntington S et al. (2008) Ascor-bate peroxidase 1 plays a key role in the response ofArabidopsis thaliana to stress combination. The Journalof Biological Chemistry 283: 34197–34203.
Kunst L, Taylor DC, Giblin M et al. (1989) Analysis of seedlipid elongation using Arabidopsis thaliana mutants. InAbstracts of Symposium on Genetics and Molecular Bi-ology of Arabidopsis, Indiana University, Bloomington,IN.
Kurek I, Chang TK, Bertain SM et al. (2007) Enhanced Ther-mostability of Arabidopsis Rubisco activase improvesphotosynthesis and growth rates under moderate heatstress. The Plant Cell 19: 3230–3241.
Laporte MM, Shen B, Tarczynski MC (2002) Engineeringfor drought avoidance: Expression of maize NADP-malicenzyme in tobacco results in altered stomatal function.Journal Experimental Botany 53: 699–705.
Larkindale J, Vierling E (2008) Core Genome Responses In-volved in Acclimation to High Temperature. Plant Phys-iology 146: 748–761.
Lee JH, Hubel A, Schoffl F (1995) Derepression of the activ-ity of genetically engineered heat shock factor causesconstitutive synthesis of heat shock proteins and in-creased thermotolerance in transgenic Arabidopsis. ThePlant Journal 8: 603–612.
Li GL, Chang H, Li B et al. (2007) The roles of the atDjA2and atDjA3 molecular chaperone proteins in improvingthermotolerance of Arabidopsis thaliana seedlings. PlantScience 173: 408–416.
Li B, Wei A, Song C et al. (2008) Heterologous expression ofthe TsVP gene improves the drought resistance of maize.Plant Biotechnology Journal 6: 146–159.
Liang YK, Dubos C, Dodd IC et al. (2005) AtMYB61,an R2R3-MYB transcription factor controlling stomatalaperture in Arabidopsis thaliana. Current Biology 15:1201–1206.
Lim CJ, Hwang JE, Chen H et al. (2007) Over-expression ofthe Arabidopsis DRE/CRT-binding transcription factorDREB2C enhances thermotolerance. Biochemical Bio-physical Research Communications 362: 431–436.
Liu H, Shi L, Zhao JS et al. (2006) Genetic characteris-tic of high molecular weight glutenin subunits in so-
matic hybrid wheat lines—potential application to wheatbreeding. Journal Agricultural and Food Chemistry 54:5007–5013.
Liu J, Xu X, Deng X (2005) Intergeneric somatic hybridiza-tion and its application to crop genetic improvement.Plant Cell, Tissue and Organ Culture 82: 19–44.
Liu L, Lafitte R, Guan D (2004) Wild Oryza species as po-tential sources of drought-adaptive traits. Euphytica 138:149–161.
Lopez CG, Banowetz GM, Peterson CJ et al. (2003) De-hydrin expression and drought tolerance in seven wheatcultivars. Crop Science 43: 577–582.
Malik MK, Slovin JP, Hwang CH et al. (1999) Modifiedexpression of a carrot small heat shock protein gene,hsp17.7, results in increased or decreased thermotoler-ance. The Plant Journal 20: 89–99.
McKersie BD, Bowley SR, Harjanto E et al. (1996) Wa-ter deficit tolerance and field performance of transgenicalfalfa overexpressing superoxide dismutase. Plant Phys-iology 111: 1177–1181.
Miranda JA, Avonce N, Suarez R et al. (2007) A bifunctionalTPS-TPP enzyme from yeast confers tolerance to mul-tiple and extreme abiotic-stress conditions in transgenicArabidopsis. Planta 226: 1411–1421.
Mittler R (2006) Abiotic stress, the field environmentand stress combination. Trends in Plant Science 11:15–19.
Møller IM, Jensen PE, Hansson A (2007) Oxidative modifi-cations to cellular components in plants. Annual Reviewof Plant Biology 58: 459–481.
Murakami Y, Tsuyama M, Kobayashi Y et al. (2000) Trienoicfatty acids and plant tolerance of high temperature. Sci-ence 287: 476–479.
Nakashima K, Ito Y, Yamaguchi-Shinozaki K (2009) Tran-scriptional regulatory networks in response to abioticstresses in Arabidopsis and grasses. Plant Physiology149: 88–95.
Nishizawa A, Yabuta Y, Yoshida E et al. (2006) Arabidopsisheat shock transcription factor A2 as a key regulator inresponse to several types of environmental stress. ThePlant Journal 48: 535–547.
Nautiyal PC, Rajgopal K, Zala PV et al. (2008) Evaluationof wild Arachis species for abiotic stress tolerance: I.Thermal stress and leaf water relations. Euphytica 159:43–57.
Nelson DE, Repetti PP, Adams TR et al. (2007) Plant nuclearfactor Y (NF-Y) B subunits confer drought toleranceand lead to improved corn yields on water-limited acres.Proceedings of the National Academy of Sciences U.S.A.104: 16450–16455.
Nevo E (2007) Evolution of wild wheat and barley and cropimprovement: Studies at the Institute of Evolution. IsraelJournal of Plant Science 55: 251–262.
Oh SJ, Song SI, Kim YS et al. (2005) ArabidopsisCBF3/DREB1A and ABF3 in transgenic rice increasedtolerance to abiotic stress without stunting growth. PlantPhysiology 138: 341–351.

P1: SFK/UKS P2: SFK Color: 1C
BLBS082-27 BLBS082-Yadav July 12, 2011 17:16 Trim: 246mm X 189mm
ADAPTATION TO WARMER AND DRIER CLIMATE 543
Ortiz R, Braun H-J, Crossa J et al. (2008) Wheat geneticresources enhancement by the International Maize andWheat Improvement Center (CIMMYT). Genetic Re-sources and Crop Evolution 55: 1095–1140.
Qin F, Kakimoto M, Sakuma Y et al. (2007) Regulationand functional analysis of ZmDREB2A in response todrought and heat stress in Zea mays L. The Plant Journal50: 54–69.
Qin D, Wu H, Peng H et al. (2008) Heat stress-responsivetranscriptome analysis in heat susceptible and tolerantwheat (Triticum aestivum L.) by using Wheat GenomeArray. BMC Genomics 9: 432.
Quan R, Shang M, Zhang H et al. (2004) Engineering of en-hanced glycine betaine synthesis improves drought toler-ance in maize. Plant Biotechnology Journal 2: 477–486.
Rauf S (2008): Breeding sunflower (Helianthus annuus L.)for drought tolerance. Communications in Biometry andCrop Science 3: 29–44.
Rennenberg H, Loreto F, Polle A et al. (2006) Physiologi-cal responses of forest trees to heat and drought. PlantBiology 8: 556–571.
Reynolds M, Dreccer F, Trethowan R (2007) Drought-adaptive traits derived from wheat wild relatives and lan-draces. Journal of Experimental Botany 58: 177–186.
Reynolds M, Tuberosa R (2008) Translational researchimpacting on crop productivity in drought-prone en-vironments. Current Opinion in Plant Biology 11:171–179.
Richards R.A (1996) Defining selection criteria to im-prove yield under drought. Plant Growth Regulation 20:157–166.
Rivero RM, Kojima M, Gepstein A et al. (2007) Delayedleaf senescence induces extreme drought tolerance in aflowering plant. Proceedings of the National Academy ofSciences U.S.A. 104: 19631–19636.
Rizhsky L, Hongjian L, Mittler R (2002) The combined effectof drought stress and heat shock on gene expression intobacco. Plant Physiology 130: 1143–1151.
Rizhsky L, Liang H, Shuman J et al. (2004) When defensepathways collide. The response of Arabidopsis to a com-bination of drought and heat stress. Plant Physiology 134:1683–1696.
Rodrigues SM, Andrade MO, Gomes APS et al. (2006) Ara-bidopsis and tobacco plants ectopically expressing thesoybean antiquitin-like ALDH7 gene display enhancedtolerance to drought, salinity, and oxidative stress. Jour-nal of Experimental Botany 55: 301–308.
Sade N, Vinocur BJ, Diber A et al. (2009) Improving plantstress tolerance and yield production: Is the tonoplastaquaporin SlTIP2;2 a key to isohydric to anisohydricconversion? New Phytologist 181: 651–661.
Saijo Y, Hata S, Kyozuka J et al. (2000) Over-expressionof a single Ca2+-dependent protein kinase confers bothcold and salt/drought tolerance on rice plants. The PlantJournal 23: 319–327.
Sakuma Y, Maruyama K, Quin F et al. (2006a) Dual func-tion of an Arabidopsis transcription factor DREB2A in
water-stress-responsive and heat-stress-responsive geneexpression. Proceedings of the National Academy of Sci-ences U.S.A. 103: 18822–18827.
Sakuma Y, Maruyama K, Osakabe Y et al. (2006b) Func-tional analysis of an Arabidopsis transcription factor,DREB2a, involved in drought-responsive gene expres-sion. The Plant Cell 18: 1292–1309.
Salvucci ME, Crafts-Brandner SJ (2004a) Relationship be-tween the heat tolerance of photosynthesis and the ther-mal stability of rubisco activase in plants from con-trasting thermal environments. Plant Physiology 134:1460–1470.
Salvucci ME, Crafts-Brandner SJ (2004b) Inhibition of pho-tosynthesis by heat stress: The activation state of Rubiscoas a limiting factor in photosynthesis. Physiologia Plan-tarum 120: 179–186.
Sangster TA, Bahrami A, Wilczek A et al. (2007) Pheno-typic diversity and altered environmental plasticity inArabidopsis thaliana with reduced Hsp90 levels. PLoSONE 2: e648.
Sato Y, Yokoya S (2008) Enhanced tolerance to droughtstress in transgenic rice plants overexpressing a smallheat-shock protein, sHSP17.7. Plant Cell Reports 27:329–334.
Schafleitner R, Rosales ROG, Gaudin A et al. (2007) Cap-turing candidate drought tolerance traits in two nativeAndean potato clones by transcription profiling of fieldgrown plants under water stress. Plant Physiology andBiochemistry 45: 673–690.
Schramm F, Larkindale J, Kiehlmann E et al. (2008)A cascade of transcription factor DREB2A and heatstress transcription factor HsfA3 regulates the heatstress response of Arabidopsis. The Plant Journal 53:264–274.
Shinozaki K, Yamaguchi-Shinozaki K (2007) Gene networksinvolved in drought stress response and tolerance. Jour-nal of Experimental Botany 58: 221–227.
Shou H, Bordallo P, Wang K (2004) Expression of the Nico-tiana protein kinase (NPK1) enhanced drought tolerancein transgenic maize. Journal of Experimental Botany 55:1013–1019.
Shukla LI, Chinnusamy V, Sunkar R (2008) The role ofmicroRNAs and other endogenous small RNAs in plantstress responses. Biochimica et Biophysica Acta - GeneRegulatory Mechanisms 1779: 743–748.
Singh A, Grover A (2008) Genetic engineering for heat tol-erance in plants. Physiology and Molecular Biology ofPlants 14: 155–166.
Sohn SO, Back K (2007) Transgenic rice tolerant to hightemperature with elevated contents of dienoic fatty acids.Biologia Plantarum 51: 340–342.
Suprunova T, Krugman T, Fahima T et al. (2004) Differentialexpression of dehydrin genes in wild barley, Hordeumspontaneum, associated with resistance to water deficit.Plant Cell and Environment 27: 1297–1308.
Suzuki N, Bajad S, Shuman J et al. (2008) The transcriptionalco-activator MBF1c is a key regulator of thermotolerance

P1: SFK/UKS P2: SFK Color: 1C
BLBS082-27 BLBS082-Yadav July 12, 2011 17:16 Trim: 246mm X 189mm
544 CROP ADAPTATION TO CLIMATE CHANGE
in Arabidopsis thaliana. Journal of Biological Chemistry283: 9269–9275.
Tang L, Kim MD, Yang KS et al. (2008) Enhanced toler-ance of transgenic potato plants overexpressing nucleo-side diphosphate kinase 2 against multiple environmentalstresses. Transgenic Research 17: 705–715.
Tang L, Kwon SY, Kim SH et al. (2006) Enhanced toler-ance of transgenic potato plants expressing both super-oxide dismutase and ascorbate peroxidase in chloroplastsagainst oxidative stress and high temperature. Plant CellReports 25: 1380–1386.
Ter Steege MW, den Ouden FM, Lambers H et al. (2005)Genetic and physiological architecture of early vigorin Aegilops tauschii, the D-genome donor of hexaploidwheat. a quantitative trait loci analysis. Plant Physiology139: 1078–1094.
Thieme R, Rakosy-Tican E, Gavrilenko T et al. (2008)Novel somatic hybrids (Solanum tuberosum L.+Solanumtarnii) and their fertile BC1 progenies express extremeresistance to potato virus Y and late blight. Theoreticaland Applied Genetics 116: 691–700.
Tognetti VB, Palatnik JF, Fillat MF et al. (2006) Functionalreplacement of ferredoxin by a cyanobacterial flavo-doxin in tobacco confers broad-range stress tolerance.The Plant Cell 18: 2035–2050.
Toker C, Canci H, Yildirim T (2007) Evaluation of peren-nial wild Cicer species for drought resistance. GeneticResources and Crop Evolution 54: 1781–1786.
Van Ginkel M, Ogbonnaya F (2007) Novel genetic diversityfrom synthetic wheats in breeding cultivars for chang-ing production conditions. Field Crops Research 104:86–94.
Vasquez-Robinet C, Mane SP, Ulanov AV et al. (2008) Physi-ological and molecular adaptations to drought in Andeanpotato genotypes. Journal of Experimental Botany 59:2109–2123.
Von Koskull-Doring P, Scharf KD, Nover L (2007) The di-versity of plant heat stress transcription factors. Trendsin Plant Science 12: 452–457.
Waie B, Rajam MV (2003) Effect of increased polyaminebiosynthesis on stress responses in transgenic tobaccoby introduction of human S-adenosylmethionine gene.Plant Science 164: 727–734.
Wang Y, Beaith M, Chalifoux M et al. (2009) Shoot-specificdown-regulation of protein farnesyltransferase (-subunit)for yield protection against drought in canola. MolecularPlant 2: 191–200.
Wang Y, Ying J, Kuzma M et al. (2005) Molecular tailoringof farnesylation for plant drought tolerance and yieldprotection. The Plant Journal 43: 413–424.
Wang W, Vinocur B, Shoseyov O et al. (2004) Role of plantheat-shock proteins and molecular chaperones in the abi-otic stress response. Trends in Plant Science 9: 244–252.
Wu X, Shiroto Y, Kishitani S et al. (2009) Enhanced heatand drought tolerance in transgenic rice seedlings over-expressing OsWRKY11 under the control of HSP101 pro-moter. Plant Cell Reports 28: 21–30.
Xiao B, Huang Y, Tang N et al. (2007) Over-expression ofa LEA gene in rice improves drought resistance underthe field conditions. Theoretical Applied Genetics 115:35–46.
Xiao B-Z, Chen X, Xiang C-B et al. (2009) Evaluation ofseven function-known candidate genes for their effectson improving drought resistance of transgenic rice underfield conditions. Molecular Plant 2: 73–83.
Xu C, Jing R, Mao X et al. (2007) A wheat (Triticum aes-tivum) protein phosphatase 2a catalytic subunit gene pro-vides enhanced drought tolerance in tobacco. Annals ofBotany 99: 439–450.
Xue GP, McIntyre CL, Glassop D et al. (2008a) Use ofexpression analysis to dissect alterations in carbohydratemetabolism in wheat leaves during drought stress. PlantMolecular Biology 67: 197–214.
Xue GP, McIntyre CL, Jenkins CLD et al. (2008b) Molec-ular dissection of variation in carbohydrate metabolismrelated to water soluble carbohydrate accumulation instems of wheat (Triticum aestivam L.). Plant Physiology146: 441–454.
Xue GP, McIntyre CL, Chapman S et al. (2006) Differentialgene expression of wheat progeny with contrasting levelsof transpiration efficiency. Plant Molecular Biology 61:863–881.
Yamada K, Fukao Y, Hayashi M et al. (2007) CytosolicHSP90 regulates the heat shock response that is responsi-ble for heat acclimation in Arabidopsis thaliana. Journalof Biological Chemistry 282: 37794–3804.
Yan J, He C, Wang J et al. (2004) Overexpression of theArabidopsis 14-3-3 protein GF14 lambda in cotton leadsto a “stay-green” phenotype and improves stress toler-ance under moderate drought conditions. Plant and CellPhysiology 45: 1007–1014.
Yan K, Chen N, Qu YY et al. (2008) Overexpression ofsweet pepper glycerol-3-phosphate acyltransferase geneenhanced thermotolerance of photosynthetic apparatus intransgenic tobacco. Journal of Integrative Plant Biology50: 613–621.
Yang J, Sears RG, Gill BS et al. (2002) Growth and senes-cence characteristics associated with tolerance of wheat-alien amphiploids to high temperature under controlledconditions. Euphytica 126: 185–193.
Yang X, Liang Z, Lu C (2005). Genetic engineering of thebiosynthesis of glycinebetaine enhances photosynthesisagainst high temperature stress in transgenic tobaccoplants. Plant Physiology 138: 2299–2309.
Yokotani N, Ichikawa T, Kondou Y et al. (2008) Expres-sion of rice heat stress transcription factor OsHsfA2e en-hances tolerance to environmental stresses in transgenicArabidopsis. Planta 227: 957–967.
Youm JW, Jeon JH, Choi D et al. (2008) Ectopic expressionof pepper CaPF1 in potato enhances multiple stressestolerance and delays initiation of in vitro tuberization.Planta 228: 701–708.
Yu H, Chen X, Hong YY et al. (2008) Activatedexpression of an Arabidopsis HD-START protein

P1: SFK/UKS P2: SFK Color: 1C
BLBS082-27 BLBS082-Yadav July 12, 2011 17:16 Trim: 246mm X 189mm
ADAPTATION TO WARMER AND DRIER CLIMATE 545
confers drought tolerance with improved root systemand reduced stomatal density. The Plant Cell 20: 1134–1151.
Zhang JY, Broeckling CD, Blancaflor EB et al. (2005a)Overexpression of WXP1, a putative Medicago trun-catula AP2 domain-containing transcription factor gene,increases cuticular wax accumulation and enhancesdrought tolerance in transgenic alfalfa (Medicago sativa).The Plant Journal 42: 689–707.
Zhang X, Wollenweber B, Jiang D et al. (2008) Waterdeficits and heat shock effects on photosynthesis of a
transgenic Arabidopsis thaliana constitutively express-ing ABP9, a bZIP transcription factor. Journal of Exper-imental Botany 59: 839–848.
Zhang Y, Mian MA, Chekhovskiy K et al. (2005b) Differen-tial gene expression in Festuca under heat stress condi-tions. Journal of Experimental Botany 56: 897–907.
Zhao ZG, Hu TT, Ge XH et al. (2008) Production andcharacterization of intergeneric somatic hybrids betweenBrassica napus and Orychophragmus violaceus andtheir backcrossing progenies. Plant Cell Reports 27:1611–1621.