correction of hypophosphatasia-associated mineralization deficiencies in vitro by...
TRANSCRIPT

Correction of Hypophosphatasia-Associated Mineralization DeficienciesIn Vitro by Phosphate/PyrophosphateModulation in Periodontal Ligament CellsThaisangela L. Rodrigues,* Brian L. Foster,†‡ Karina G. Silverio,* Luciane Martins,*Marcio Z. Casati,* Enilson A. Sallum,* Martha J. Somerman,†‡ and Francisco H. Nociti, Jr.*†
Background: Mutations in the liver/bone/kidney alkalinephosphatase (ALPL) gene in hypophosphatasia (HPP) reducethe function of tissue non-specific alkaline phosphatase (ALP),resulting in increased pyrophosphate (PPi) and a severe defi-ciency in acellular cementum. We hypothesize that exogenousphosphate (Pi) would rescue the in vitro mineralization capacityof periodontal ligament (PDL) cells harvested from HPP-diagnosed patients, by correcting the Pi/PPi ratio and modulat-ing expression of genes involved with Pi/PPi metabolism.
Methods: Ex vivo and in vitro analyses were used to identifymechanisms involved in HPP-associated PDL/tooth root defi-ciencies. Constitutive expression of PPi-associated geneswas contrasted in PDL versus pulp tissues obtained fromhealthy individuals. Primary PDL cell cultures from patientswith HPP (monozygotic twin males) were established to assayALP activity, in vitro mineralization, and gene expression.Exogenous Pi was provided to correct the Pi/PPi ratio.
Results: PDL tissues obtained from healthy individualsfeatured higher basal expression of key PPi regulators, genesALPL, progressive ankylosis protein (ANKH), and ectonucleo-tide pyrophosphatase/phosphodiesterase 1 (ENPP1), versuspaired pulp tissues. A novel ALPL mutation was identified inthe twin patients with HPP enrolled in this study. Comparedto controls, HPP-PDL cells exhibited significantly reducedALP and mineralizing capacity, which were rescued by addi-tion of 1 mM Pi. Dysregulated expression of PPi regulatorygenes ALPL, ANKH, and ENPP1 was also corrected by addingPi, although other matrix markers evaluated in our studyremained downregulated.
Conclusion: These findings underscore the importance ofcontrolling the Pi/PPi ratio toward development of a functionalperiodontal apparatus and support Pi/PPi imbalance as theetiology of HPP-associated cementum defects. J Periodontol2012;83:653-663.
KEY WORDS
Dental cementum; hypophosphatasia; inorganic phosphates;inorganic pyrophosphatase; periodontal ligament.
Biomineralization of bones andteeth requires a tight regulationof the ratio between concentra-
tions of extracellular phosphate (Pi), anion incorporated in hydroxyapatite min-eral, and pyrophosphate (PPi), a potentinhibitor of mineral precipitation.1-3 Dis-orders arising from imbalance in the Pi/PPi ratio have been reported to producedissimilar effects on developing dentalhard tissues, prompting the hypothesisthat dental tissues are differentially sen-sitive to Pi/PPi metabolism.4,5 In hypo-phosphatasia (HPP), deficiency of serumalkaline phosphatase (ALP) activity re-sults from mutations in the liver/bone/kidney alkaline phosphatase gene (ALPL;Online Mendelian Inheritance in Manidentification number 171760), encodingtissue non-specific alkaline phosphatase(TNAP). One consequence is defectiveformation of acellular cementum, result-ing in poor attachment of the periodontalligament (PDL) to the root surface and,consequently, premature tooth exfolia-tion.6 In the Akp2 null murine modelfor HPP, cementum is similarly defec-tive.7,8 However, in both humans andmice with reduced ALP, dentin has beenreported to be unaffected or less af-fected than cementum.4,8-10 Accumu-lation of the mineralization inhibitor PPi
was identified as the proximal cause fordefective mineralization in skeletal and* Department of Prosthodontics and Periodontics, Division of Periodontics, School of
Dentistry, Campinas State University, Piracicaba, Sao Paulo, Brazil.† Department of Periodontics, University of Washington, School of Dentistry, Seattle, WA.‡ Department of Oral Biology, University of Washington, School of Dentistry. doi: 10.1902/jop.2011.110310
J Periodontol • May 2012
653

dental hard tissues. In contrast, in murine models inwhich local PPi is deficient, namely loss of progressiveankylosis protein (ANK) or ectonucleotide pyrophos-phatase/phosphodiesterase 1 (ENPP1) function, acel-lular cementum is markedly increased, although dentinis unaffected.5,11
Although differences in expression of Pi/PPi regula-tors have been suggested between tissues of the peri-odontal versus pulp regions, the mechanism by whichHPP affects the periodontium has not been defined atthe cellular/molecular level. Based on evidence fromhuman patients and transgenic mouse models, wehypothesized that PPi-regulating factors would beexpressed at higher levels in periodontal versus pulptissues under normal conditions and, further, that es-tablishing an appropriate Pi/PPi balance is of criticalimportance for directing mineralization of the root sur-face, i.e., cementum formation. Here, we aim to deter-mine levels of PPi-regulating genes in PDL versus pulptissues, changes in PDL cell phenotype (i.e., gene ex-pression and mineralizing capacity) resulting fromHPP, and potential for normalization of the Pi/PPi ratioto correct mineralization defects in cells harvestedfrom HPP-diagnosed patients (monozygotic twins).
MATERIALS AND METHODS
ParticipantsA total of 16 participants (six males and 10 females;aged 17 to 22 years) were enrolled in this study, withInstitutional Review Board approval (Campinas StateUniversity, School of Dentistry, Piracicaba, Sao Paulo,Brazil, No. 065/2005). The study group consisted oftwo male monozygotic twins and 14 controls. Inclu-sion criteria included no history of smoking, diabetes,bone metabolic disorders, or other systemic disease,except for HPP in the HPP group. For the controlgroup, individuals were periodontally healthy witherupted teeth scheduled for extraction for orthodonticreasons and with serum ALP activity within the normaladult range (25 to 100 U/L). Patients with HPP weremonitored and treated as needed, and tooth extrac-tions were performed as a consequence of HPP-related pathology. The 14 control participants weresplit into two groups, with paired PDL and pulp tissuesharvested (as described below) from nine control par-ticipants, and primary PDL cell cultures established(as described below) from five control participantsand two HPP-diagnosed individuals. Patients withHPP were seen in the clinic from 1991 to the presentday, whereas cells were harvested from participantswith HPP and control participants from 2004 to 2005.
Genetic Analysis of Patients With HPPHuman deoxyribonucleic acid (DNA) from bloodsamples was purified from leukocytes.12 Twelveexons of ALPL were polymerase chain reaction
(PCR) amplified.13 Sequencing analyses were per-formed using a sequencing reaction kit using dye-labeled terminators,§ followed by detection on anautomated sequencing system.i The identified mu-tation was additionally confirmed using standard re-striction enzyme digestion and electrophoresis.
Tissue HarvestTo determine the basal levels of genes associated withPi/PPi metabolism between PDL versus pulp tissues,comparisons were made from paired tissues takenfrom the same tooth. Teeth were extracted from con-trol participants and rinsed in phosphate-buffered sa-line (PBS), and PDL tissues removed by scraping theroot surface. Next, the teeth were cracked open andpulps were removed with sterile forceps. Tissues wereimmersed in a ribonucleic acid (RNA) stabilizationsolution¶ and stored at -80�C. Total RNA was isolatedby the guanidinium thiocyanate-phenol-chloroformextraction method# in conjunction with automated ce-ramic bead homogenization.** Non-pooled PDL andpulp tissue RNA samples were analyzed for basal geneexpression using quantitative PCR.
Cell Isolation and CultureExtracted teeth were placed in biopsy media, and PDLcells were obtained by enzymatic digestion (3 mg/mLcollagenase type I and 4 mg/mL dispase††) for 1 hourat 37�C. Cells were maintained in Dulbecco’s modifiedEagle medium (DMEM) with 10% fetal bovine serum(FBS), 1% L-glutamine, and 1% penicillin/streptomycin,‡‡ and incubated at 37�C in 5% CO2.Equivalent passages 2 to 4 for control and HPP-PDL primary cells were used for all experiments.
Cell ProliferationCells were seeded at 1.5 · 104 cells/cm2 in 96-well platesinDMEMwith2%FBSfrom24hours to6days.Cellswerecounted with a hemacytometer and analyzed by a colori-metric formazan-based cell proliferation assay [3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium, inner salt; MTS].§§
ALP ActivityCells were seeded at 2.0 · 104 cells/cm2 in 60-mmplates in DMEM with 2% FBS and 50 mg/mL ascorbicacid (AA) up to 21 days. Relative ALP activity wasmeasured using a commercial kitii as reported pre-viously.14 Briefly, media were removed, cells wererinsed with PBS, and release of thymolphthalein
§ DYEnamic ET Terminator Cycle Sequencing kit, GE Healthcare,Buckinghamshire, UK.
i ABI PRISM 3100 sequencer, Applied Biosystems, Carlsbad, CA.¶ RNAlater, Ambion, Austin, TX.# TRIzol reagent, Invitrogen, Grand Island, NY.** MagNA Lyser, Roche Applied Science, Indianapolis, IN.†† Invitrogen.‡‡ Invitrogen.§§ Promega, Madison, WI.ii Labtest Diagnostica, Lagoa Santa, MG, Brazil.
Periodontal Ligament Cells in Hypophosphatasia Volume 83 • Number 5
654

from thymolphthalein monophosphate substratewas measured by absorbance readings at 590 nmafter 30 minutes of incubation at 37�C. Results wereexpressed as ALP activity normalized to total proteincontent per well.
Mineralization AssayCells were seeded at 2.0 · 104 cells/cm2 in 24-wellplates under non-mineralizing (2% FBS and AA) ormineralizing conditions up to 28 days. Mineralizingconditions included 2% FBS and AA plus a Pi source,10 mM b-glycerophosphate (bGP)¶¶ or 1 mM inor-ganic Pi (a solution of monobasic and dibasic sodiumphosphates, pH 7.4).## The dose 1 mM Pi was chosenbased on previous work with Pi and cementoblasts,15,16
as well as preliminary experiments with PDL cellsto determine a non-toxic dose of Pi that was suf-ficient to allow mineral nodule formation (data notshown). Mineral nodules were detected by vonKossa assay and alizarin–red (AR) staining*** (40mM, pH 4.2). AR stain was quantified by measuringabsorbance of bound dye (570 nm) solubilized in 10%cetylpyridinium chloride,††† as described previously.17
Gene Expression AssayCells were seeded at 2.0 · 104 cells/cm2 in 60 mmplates in non-mineralizing and mineralizing media(with added Pi), as outlined above, for up to 20 days.Total RNA was extracted as described for tissuesabove,‡‡‡ deoxyribonuclease treated,§§§ and usedforcomplementaryDNAsynthesiswitha recombinantreverse transcription kit.iii Quantitative real-time PCRreactions were performed using a cyanine dye–basedhot start PCR kit.¶¶¶ Relative quantification was per-formed using amplification efficiency correction withGAPDH as the reference gene. Assessed genes andprimers are listed in Table 1.
Statistical AnalysesAll cell and tissue experiments were performed in trip-licate and repeated at least twice. Values are given asmeans – SD. Intragroup and intergroup comparisonswere performed using the Kruskal-Wallis one-wayanalysis of variance (ANOVA) followed by the Stu-dent-Newman-Keuls method (P =0.05) for prolifera-tion and mineralization assays and gene expression.A Student t test was used for intergroup comparisonsfor ALP activity. Statistical power was ‡0.08 withP = 0.05 for any statistical test.
RESULTS
Diagnosis of HPPWhite, male identical twins were brought for dentalevaluation by their parents to the Campinas StateUniversity, School of Dentistry, Piracicaba, Sao PauloBrazil. Individuals with HPP were evaluated for possi-
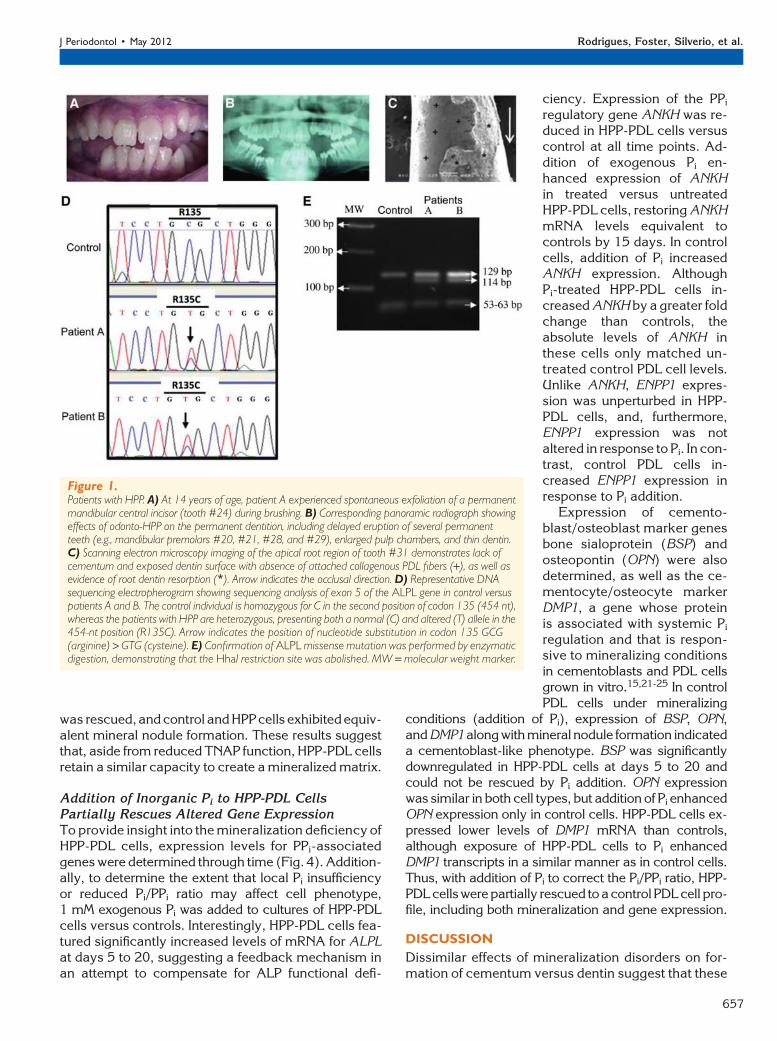
ble mineralized tissue disorders at the age of 2 yearsbecause of reported premature exfoliation of anteriorprimary teeth. Physical examination and radiography(long bones, joints, and skull) did not indicate addi-tional aberrant findings. Biochemical analysis re-vealed low serum ALP activity (patient A, 62 U/L;patient B, 63 U/L; normal range for children, 151 to471 U/L). Serum Pi and calcium serum levels were nor-mal. Based on the above, a diagnosis of HPP subtypeodonto-HPP was given.
Over subsequent years, patients suffered prema-ture exfoliation of primary and permanent teeth.Permanent dentition featured short roots, wide pulpchambers, and reduced alveolar bone height (Figs.1A through 1C). At the age of 22 years, when cellswere harvested for this study, ALP activity remainedlow, at 8 and 6 U/L for patients A and B, respectively(normal adult range is 25 to 100 U/L).
Sequencing the ALPL gene revealed a heterozy-gous transition 454C > T in exon 5 of both patients,leading to substitution of cysteine for arginine at posi-tion 135 (R135C) (Fig. 1D). The missense mutationwas confirmed by HhaI digest, demonstrating thatthe expected restriction site 59-GCGC-39 was abol-ished (Fig. 1E). Although digest of the control 306bp ALPL exon 5 produced fragments of 129, 63, 61,and 53 bp (from three restriction sites), in the patientswith HPP, loss of one of these restriction sites by ALPLmutation resulted in fragments of 129, 114, and 63 bp.
PPi Regulators Are Differentially Expressed inPDL Versus Pulp TissuesDissimilar effects of mineralization disorders on indi-vidual dental hard tissues during formation haveprompted the hypothesis that cementogenesis anddentinogenesis are regulated by disparate mecha-nisms.4,5 Constitutive gene expression was comparedin PDL versus pulp tissues harvested from healthyindividuals. PDL tissues featured significantly higher(P <0.05) messenger RNA (mRNA) levels for ALPL,progressive ankylosis protein (ANKH), and ectonucleo-tide pyrophosphatase/phosphodiesterase 1 (ENPP1),all key regulators of local PPi levels (Fig. 2). Gene ex-pression of other mineralized tissue markers (notassociated with PPi metabolism) was determined toconsider the specificity of differences between PDLand pulp in PPi-associated genes. PDL featured 10-foldhigher expression of osteocalcin (OCN) and an al-most 20-fold greater expression of COL1 versus pulp,
¶¶ Sigma-Aldrich, St. Louis, MO.## Sigma-Alrich.*** Alizarin red-S, Sigma-Aldrich.††† MP Biomedicals, Solon, OH.‡‡‡ TRIzol reagent, Invitrogen.§§§ TURBO DNA-free, Ambion.iii Transcriptor reverse transcription kit, Roche Diagnostic, Indianapolis,
IN.¶¶¶ SYBR green FastStart DNA MasterPLUS, Roche Applied Science.
J Periodontol • May 2012 Rodrigues, Foster, Silverio, et al.
655
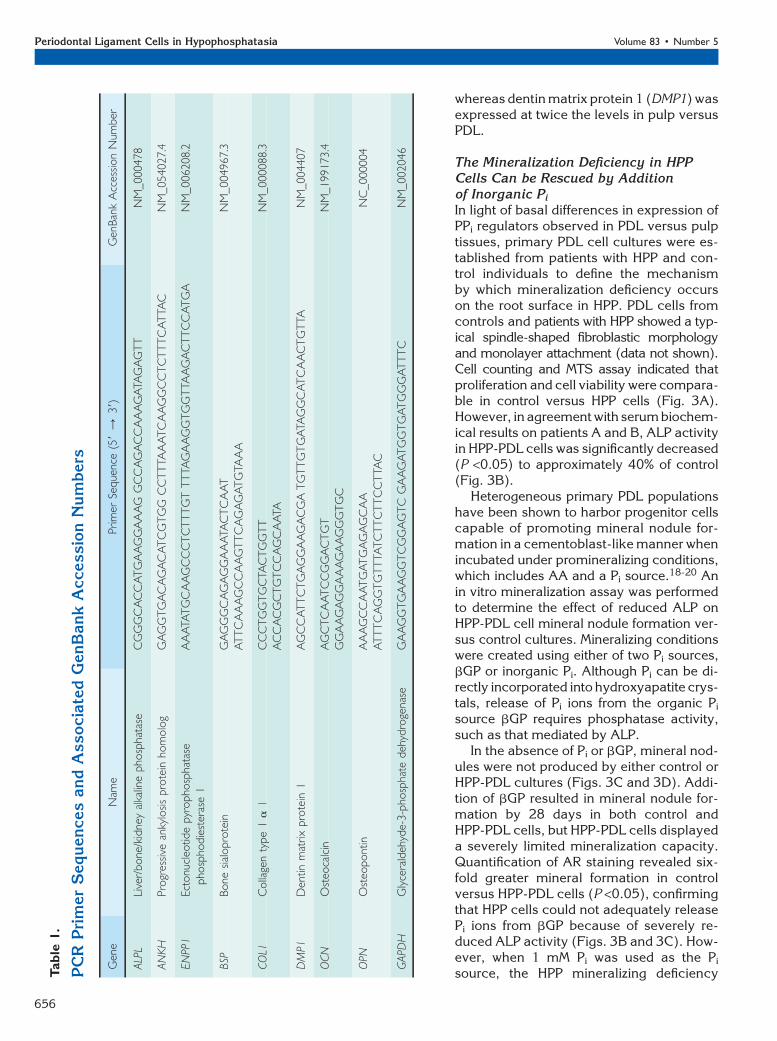
whereas dentin matrix protein 1 (DMP1) wasexpressed at twice the levels in pulp versusPDL.
The Mineralization Deficiency in HPPCells Can be Rescued by Additionof Inorganic Pi
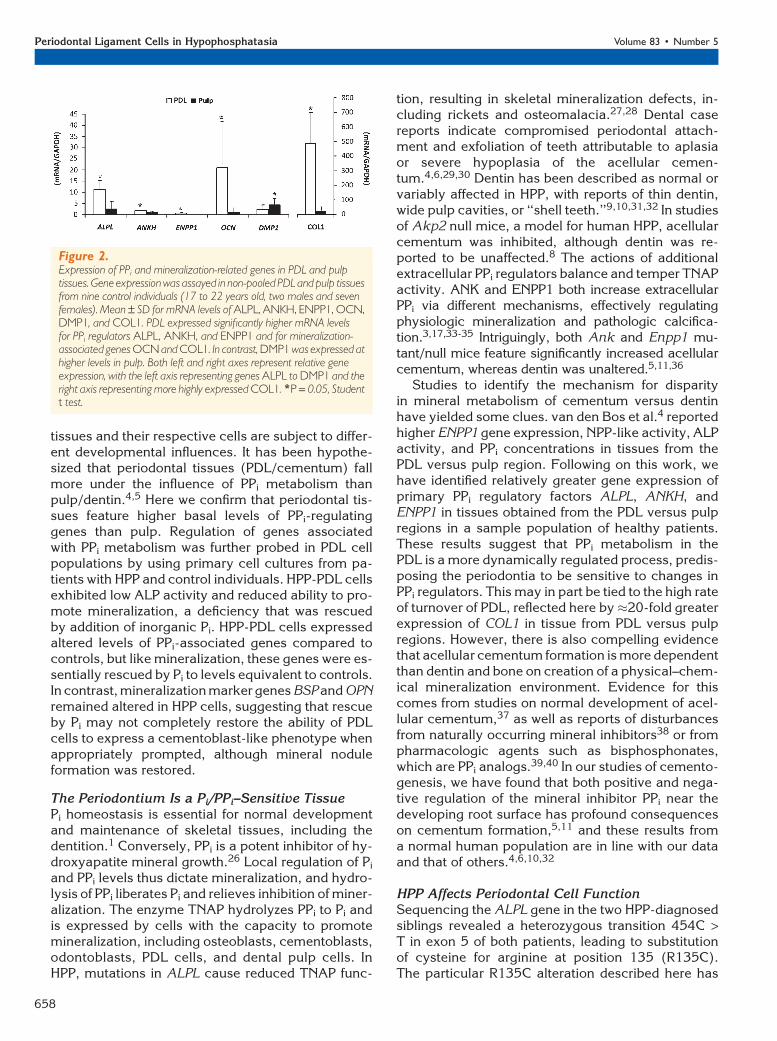
In light of basal differences in expression ofPPi regulators observed in PDL versus pulptissues, primary PDL cell cultures were es-tablished from patients with HPP and con-trol individuals to define the mechanismby which mineralization deficiency occurson the root surface in HPP. PDL cells fromcontrols and patients with HPP showed a typ-ical spindle-shaped fibroblastic morphologyand monolayer attachment (data not shown).Cell counting and MTS assay indicated thatproliferation and cell viability were compara-ble in control versus HPP cells (Fig. 3A).However, in agreement with serum biochem-ical results on patients A and B, ALP activityin HPP-PDL cells was significantly decreased(P <0.05) to approximately 40% of control(Fig. 3B).
Heterogeneous primary PDL populationshave been shown to harbor progenitor cellscapable of promoting mineral nodule for-mation in a cementoblast-like manner whenincubated under promineralizing conditions,which includes AA and a Pi source.18-20 Anin vitro mineralization assay was performedto determine the effect of reduced ALP onHPP-PDL cell mineral nodule formation ver-sus control cultures. Mineralizing conditionswere created using either of two Pi sources,bGP or inorganic Pi. Although Pi can be di-rectly incorporated into hydroxyapatite crys-tals, release of Pi ions from the organic Pi
source bGP requires phosphatase activity,such as that mediated by ALP.
In the absence of Pi or bGP, mineral nod-ules were not produced by either control orHPP-PDL cultures (Figs. 3C and 3D). Addi-tion of bGP resulted in mineral nodule for-mation by 28 days in both control andHPP-PDL cells, but HPP-PDL cells displayeda severely limited mineralization capacity.Quantification of AR staining revealed six-fold greater mineral formation in controlversus HPP-PDL cells (P <0.05), confirmingthat HPP cells could not adequately releasePi ions from bGP because of severely re-duced ALP activity (Figs. 3B and 3C). How-ever, when 1 mM Pi was used as the Pi
source, the HPP mineralizing deficiencyTab
le1.
PC
RPrim
er
Sequences
and
Associa
ted
GenB
ank
Accessio
nN
um
bers
Gen
eN
ame
Prim
erSe
que
nce
(59
/39)
Gen
Ban
kA
cces
sion
Num
ber
ALP
LLi
ver/
bone
/kid
ney
alka
line
pho
spha
tase
CG
GG
CA
CC
AT
GA
AG
GA
AA
GG
CC
AG
AC
CA
AA
GA
TAG
AG
TT
NM
_000478
AN
KH
Pro
gres
sive
anky
losis
pro
tein
hom
olo
gG
AG
GT
GA
CA
GA
CA
TC
GT
GG
CC
TT
TAA
AT
CA
AG
GC
CT
CT
TT
CA
TTA
CN
M_054027.4
EN
PP1
Ecto
nucl
eotid
epyr
opho
spha
tase
pho
spho
die
ster
ase
1A
AA
TAT
GC
AA
GC
CC
TC
TT
TG
TT
TTA
GA
AG
GT
GG
TTA
AG
AC
TT
CC
AT
GA
NM
_006208.2
BSP
Bone
sial
opro
tein
GA
GG
GC
AG
AG
GA
AA
TAC
TC
AA
TN
M_004967.3
AT
TC
AA
AG
CC
AA
GT
TC
AG
AG
AT
GTA
AA
CO
L1C
olla
gen
type
1a
1C
CC
TG
GT
GC
TAC
TG
GT
TN
M_000088.3
AC
CA
CG
CT
GT
CC
AG
CA
ATA
DM
P1D
entin
mat
rix
pro
tein
1A
GC
CA
TT
CT
GA
GG
AA
GA
CG
AT
GT
TG
TG
ATA
GG
CA
TC
AA
CT
GT
TAN
M_004407
OCN
Ost
eoca
lcin
AG
CT
CA
AT
CC
GG
AC
TG
TN
M_199173.4
GG
AA
GA
GG
AA
AG
AA
GG
GT
GC
OPN
Ost
eopont
inA
AA
GC
CA
AT
GA
TG
AG
AG
CA
AN
C_000004
AT
TT
CA
GG
TG
TT
TAT
CT
TC
TT
CC
TTA
C
GAPD
HG
lyce
rald
ehyd
e-3-p
hosp
hate
deh
ydro
gena
seG
AA
GG
TG
AA
GG
TC
GG
AG
TC
GA
AG
AT
GG
TG
AT
GG
GA
TT
TC
NM
_002046
Periodontal Ligament Cells in Hypophosphatasia Volume 83 • Number 5
656
was rescued,and control and HPPcells exhibitedequiv-alent mineral nodule formation. These results suggestthat, aside from reduced TNAP function, HPP-PDL cellsretain a similar capacity to create a mineralized matrix.
Addition of Inorganic Pi to HPP-PDL CellsPartially Rescues Altered Gene ExpressionTo provide insight into the mineralization deficiency ofHPP-PDL cells, expression levels for PPi-associatedgenes were determined through time (Fig. 4). Addition-ally, to determine the extent that local Pi insufficiencyor reduced Pi/PPi ratio may affect cell phenotype,1 mM exogenous Pi was added to cultures of HPP-PDLcells versus controls. Interestingly, HPP-PDL cells fea-tured significantly increased levels of mRNA for ALPLat days 5 to 20, suggesting a feedback mechanism inan attempt to compensate for ALP functional defi-
ciency. Expression of the PPi
regulatory gene ANKH was re-duced in HPP-PDL cells versuscontrol at all time points. Ad-dition of exogenous Pi en-hanced expression of ANKHin treated versus untreatedHPP-PDL cells, restoring ANKHmRNA levels equivalent tocontrols by 15 days. In controlcells, addition of Pi increasedANKH expression. AlthoughPi-treated HPP-PDL cells in-creased ANKH by a greater foldchange than controls, theabsolute levels of ANKH inthese cells only matched un-treated control PDL cell levels.Unlike ANKH, ENPP1 expres-sion was unperturbed in HPP-PDL cells, and, furthermore,ENPP1 expression was notaltered in response to Pi. In con-trast, control PDL cells in-creased ENPP1 expression inresponse to Pi addition.
Expression of cemento-blast/osteoblast marker genesbone sialoprotein (BSP) andosteopontin (OPN) were alsodetermined, as well as the ce-mentocyte/osteocyte markerDMP1, a gene whose proteinis associated with systemic Pi
regulation and that is respon-sive to mineralizing conditionsin cementoblasts and PDL cellsgrown in vitro.15,21-25 In controlPDL cells under mineralizing
conditions (addition of Pi), expression of BSP, OPN,andDMP1alongwithmineralnodule formation indicateda cementoblast-like phenotype. BSP was significantlydownregulated in HPP-PDL cells at days 5 to 20 andcould not be rescued by Pi addition. OPN expressionwas similar in both cell types, but addition of Pi enhancedOPN expression only in control cells. HPP-PDL cells ex-pressed lower levels of DMP1 mRNA than controls,although exposure of HPP-PDL cells to Pi enhancedDMP1 transcripts in a similar manner as in control cells.Thus, with addition of Pi to correct the Pi/PPi ratio, HPP-PDLcellswerepartially rescued toacontrolPDLcell pro-file, including both mineralization and gene expression.
DISCUSSION
Dissimilar effects of mineralization disorders on for-mation of cementum versus dentin suggest that these
Figure 1.Patients with HPP. A) At 14 years of age, patient A experienced spontaneous exfoliation of a permanentmandibular central incisor (tooth #24) during brushing. B) Corresponding panoramic radiograph showingeffects of odonto-HPP on the permanent dentition, including delayed eruption of several permanentteeth (e.g., mandibular premolars #20, #21, #28, and #29), enlarged pulp chambers, and thin dentin.C) Scanning electron microscopy imaging of the apical root region of tooth #31 demonstrates lack ofcementum and exposed dentin surface with absence of attached collagenous PDL fibers (+), as well asevidence of root dentin resorption (*). Arrow indicates the occlusal direction. D) Representative DNAsequencing electropherogram showing sequencing analysis of exon 5 of the ALPL gene in control versuspatients A and B. The control individual is homozygous for C in the second position of codon 135 (454 nt),whereas the patients with HPP are heterozygous, presenting both a normal (C) and altered (T) allele in the454-nt position (R135C). Arrow indicates the position of nucleotide substitution in codon 135 GCG(arginine) > GTG (cysteine). E) Confirmation of ALPL missense mutation was performed by enzymaticdigestion, demonstrating that the HhaI restriction site was abolished. MW = molecular weight marker.
J Periodontol • May 2012 Rodrigues, Foster, Silverio, et al.
657
tissues and their respective cells are subject to differ-ent developmental influences. It has been hypothe-sized that periodontal tissues (PDL/cementum) fallmore under the influence of PPi metabolism thanpulp/dentin.4,5 Here we confirm that periodontal tis-sues feature higher basal levels of PPi-regulatinggenes than pulp. Regulation of genes associatedwith PPi metabolism was further probed in PDL cellpopulations by using primary cell cultures from pa-tients with HPP and control individuals. HPP-PDL cellsexhibited low ALP activity and reduced ability to pro-mote mineralization, a deficiency that was rescuedby addition of inorganic Pi. HPP-PDL cells expressedaltered levels of PPi-associated genes compared tocontrols, but like mineralization, these genes were es-sentially rescued by Pi to levels equivalent to controls.In contrast, mineralization marker genes BSP and OPNremained altered in HPP cells, suggesting that rescueby Pi may not completely restore the ability of PDLcells to express a cementoblast-like phenotype whenappropriately prompted, although mineral noduleformation was restored.
The Periodontium Is a Pi/PPi–Sensitive TissuePi homeostasis is essential for normal developmentand maintenance of skeletal tissues, including thedentition.1 Conversely, PPi is a potent inhibitor of hy-droxyapatite mineral growth.26 Local regulation of Pi
and PPi levels thus dictate mineralization, and hydro-lysis of PPi liberates Pi and relieves inhibition of miner-alization. The enzyme TNAP hydrolyzes PPi to Pi andis expressed by cells with the capacity to promotemineralization, including osteoblasts, cementoblasts,odontoblasts, PDL cells, and dental pulp cells. InHPP, mutations in ALPL cause reduced TNAP func-
tion, resulting in skeletal mineralization defects, in-cluding rickets and osteomalacia.27,28 Dental casereports indicate compromised periodontal attach-ment and exfoliation of teeth attributable to aplasiaor severe hypoplasia of the acellular cemen-tum.4,6,29,30 Dentin has been described as normal orvariably affected in HPP, with reports of thin dentin,wide pulp cavities, or ‘‘shell teeth.’’9,10,31,32 In studiesof Akp2 null mice, a model for human HPP, acellularcementum was inhibited, although dentin was re-ported to be unaffected.8 The actions of additionalextracellular PPi regulators balance and temper TNAPactivity. ANK and ENPP1 both increase extracellularPPi via different mechanisms, effectively regulatingphysiologic mineralization and pathologic calcifica-tion.3,17,33-35 Intriguingly, both Ank and Enpp1 mu-tant/null mice feature significantly increased acellularcementum, whereas dentin was unaltered.5,11,36
Studies to identify the mechanism for disparityin mineral metabolism of cementum versus dentinhave yielded some clues. van den Bos et al.4 reportedhigher ENPP1 gene expression, NPP-like activity, ALPactivity, and PPi concentrations in tissues from thePDL versus pulp region. Following on this work, wehave identified relatively greater gene expression ofprimary PPi regulatory factors ALPL, ANKH, andENPP1 in tissues obtained from the PDL versus pulpregions in a sample population of healthy patients.These results suggest that PPi metabolism in thePDL is a more dynamically regulated process, predis-posing the periodontia to be sensitive to changes inPPi regulators. This may in part be tied to the high rateof turnover of PDL, reflected here by �20-fold greaterexpression of COL1 in tissue from PDL versus pulpregions. However, there is also compelling evidencethat acellular cementum formation is more dependentthan dentin and bone on creation of a physical–chem-ical mineralization environment. Evidence for thiscomes from studies on normal development of acel-lular cementum,37 as well as reports of disturbancesfrom naturally occurring mineral inhibitors38 or frompharmacologic agents such as bisphosphonates,which are PPi analogs.39,40 In our studies of cemento-genesis, we have found that both positive and nega-tive regulation of the mineral inhibitor PPi near thedeveloping root surface has profound consequenceson cementum formation,5,11 and these results froma normal human population are in line with our dataand that of others.4,6,10,32
HPP Affects Periodontal Cell FunctionSequencing the ALPL gene in the two HPP-diagnosedsiblings revealed a heterozygous transition 454C >T in exon 5 of both patients, leading to substitutionof cysteine for arginine at position 135 (R135C).The particular R135C alteration described here has
Figure 2.Expression of PPi and mineralization-related genes in PDL and pulptissues. Gene expression was assayed in non-pooled PDL and pulp tissuesfrom nine control individuals (17 to 22 years old, two males and sevenfemales). Mean – SD for mRNA levels of ALPL, ANKH, ENPP1, OCN,DMP1, and COL1. PDL expressed significantly higher mRNA levelsfor PPi regulators ALPL , ANKH, and ENPP1 and for mineralization-associated genes OCN and COL1. In contrast, DMP1 was expressed athigher levels in pulp. Both left and right axes represent relative geneexpression, with the left axis representing genes ALPL to DMP1 and theright axis representing more highly expressed COL1. *P = 0.05, Studentt test.
Periodontal Ligament Cells in Hypophosphatasia Volume 83 • Number 5
658

not been reported previously for odonto-HPP, althoughcodon 135 was linked to a case of adult-onset HPP(R135C/R167W; Societe d’Etudes et de Soins pourles Enfants Paralyses et Polymalformes - SESEP Labo-ratory,UniversityofVersailles-SaintQuentin,Versailles,France), as well as to a lethal HPP case (R135H),41 sug-gesting genotype–phenotype correlation. HPP has beenlinked with a large spectrum of mutations, with no clearpattern linking location of mutations with clinical formof HPP, onset, or severity of disease.13
Changes in cell profile associated with dysregu-lated PPi metabolism were further probed by using pri-
mary cell cultures obtainedfrom control and patients withHPP. One limitation of this ap-proach was our reliance ononly two HPP-diagnosed sib-lings for cell isolation. BecauseHPP is a rare disease, we werenot able to identify additionalindividuals for inclusion in thestudy. The patients describedhere were diagnosed with anodonto-specific subtype of HPPin which serum biochemistry isabnormal and dental defectsare the primary phenotypictraits.27,28 To our knowledge,this is the first report examiningthe mineralization capacity andgene expression profile of dentalcells isolated from patients withHPP.
No differences were noted inmorphology, proliferation, orviability between HPP and con-trol cells, in agreement withstudies of HPP dermal fibro-blasts.42-44 ALP activity andmineralization capacity forHPP-PDL cells were presentbut significantly impaired, asreported for osteoblasts fromAkp2 null mice45 and inagreement with studies ofgenotype–phenotype corre-lation indicating that humanpatients with milder forms ofHPP (e.g., odonto-HPP) exhibitresidual ALP activity.28,30
When 1 mM exogenous Pi
was added as a Pi for mineral-ization and to correct the Pi/PPi ratio, the HPP-PDL cellmineralization deficiency wascorrected to normal levels.
This result suggests that PDL cells lacking TNAPfunction are otherwise competent to promotemineralization.
To further define the behavior of periodontal cellsderived from individuals with HPP, we elected to assaygenes involved in PPi regulation (ALPL, ANKH, andENPP1) and in cementoblast/osteoblast mineraliza-tion (BSP, OPN, and DMP1). Intriguingly, expressionof ALPL and ANKH mRNAs were dysregulated inHPP-PDL cells compared to controls. Increased ALPLmay be indicative of a response to high PPi levelsas a result of reduced ALP, a corrective feedback
Figure 3.Rescue of HPP mineralization deficiency by addition of inorganic Pi. A) Control and HPP-PDL cells grown in2% FBS were counted by the MTS method. Values corresponding to relative cell numbers are shown asmean – SD (absorbance at 490 nm). Proliferation and cell viability were equivalent between HPP andcontrol cells, except at day 2. *P = 0.05, intragroup significant differences versus day 0 by Kruskal-Wallisone-way ANOVA followed by the Student-Newman-Keuls method; †P = 0.05, intergroup significantdifferences within the same period of time by the Student t test. B) Relative ALP activity was significantlylower in HPP-PDL cells versus controls. *P = 0.05, statistically different by the Student t test. C and D) ARassay for in vitro mineralization. Control and HPP-PDL cells were grown in 2% FBS, and compared tocontrols, HPP-PDL cells displayed a severely limited mineralization capacity with bGP as a Pi source. TheHPPmineralization deficiencywas rescuedwhen1mMPi was used as the Pi source. Different capital lettersindicate intergroup statistical differences (control versus HPP) for P = 0.05, using the Kruskal-Wallis testfollowed by Student-Newman-Keuls method.
J Periodontol • May 2012 Rodrigues, Foster, Silverio, et al.
659
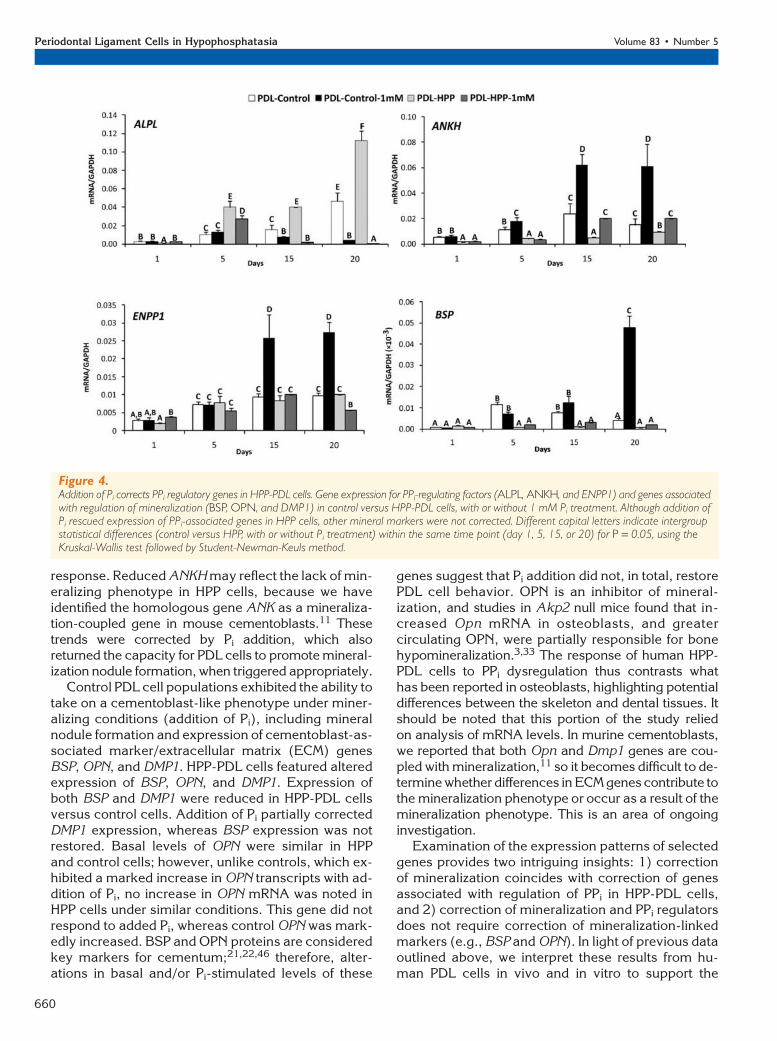
response. Reduced ANKH may reflect the lack of min-eralizing phenotype in HPP cells, because we haveidentified the homologous gene ANK as a mineraliza-tion-coupled gene in mouse cementoblasts.11 Thesetrends were corrected by Pi addition, which alsoreturned the capacity for PDL cells to promote mineral-ization nodule formation, when triggered appropriately.
Control PDL cell populations exhibited the ability totake on a cementoblast-like phenotype under miner-alizing conditions (addition of Pi), including mineralnodule formation and expression of cementoblast-as-sociated marker/extracellular matrix (ECM) genesBSP, OPN, and DMP1. HPP-PDL cells featured alteredexpression of BSP, OPN, and DMP1. Expression ofboth BSP and DMP1 were reduced in HPP-PDL cellsversus control cells. Addition of Pi partially correctedDMP1 expression, whereas BSP expression was notrestored. Basal levels of OPN were similar in HPPand control cells; however, unlike controls, which ex-hibited a marked increase in OPN transcripts with ad-dition of Pi, no increase in OPN mRNA was noted inHPP cells under similar conditions. This gene did notrespond to added Pi, whereas control OPN was mark-edly increased. BSP and OPN proteins are consideredkey markers for cementum;21,22,46 therefore, alter-ations in basal and/or Pi-stimulated levels of these
genes suggest that Pi addition did not, in total, restorePDL cell behavior. OPN is an inhibitor of mineral-ization, and studies in Akp2 null mice found that in-creased Opn mRNA in osteoblasts, and greatercirculating OPN, were partially responsible for bonehypomineralization.3,33 The response of human HPP-PDL cells to PPi dysregulation thus contrasts whathas been reported in osteoblasts, highlighting potentialdifferences between the skeleton and dental tissues. Itshould be noted that this portion of the study reliedon analysis of mRNA levels. In murine cementoblasts,we reported that both Opn and Dmp1 genes are cou-pled with mineralization,11 so it becomes difficult to de-termine whether differences in ECM genes contribute tothe mineralization phenotype or occur as a result of themineralization phenotype. This is an area of ongoinginvestigation.
Examination of the expression patterns of selectedgenes provides two intriguing insights: 1) correctionof mineralization coincides with correction of genesassociated with regulation of PPi in HPP-PDL cells,and 2) correction of mineralization and PPi regulatorsdoes not require correction of mineralization-linkedmarkers (e.g., BSP and OPN). In light of previous dataoutlined above, we interpret these results from hu-man PDL cells in vivo and in vitro to support the
Figure 4.Addition of Pi corrects PPi regulatory genes in HPP-PDL cells. Gene expression for PPi-regulating factors (ALPL, ANKH, and ENPP1) and genes associatedwith regulation of mineralization (BSP, OPN, and DMP1) in control versus HPP-PDL cells, with or without 1 mM Pi treatment. Although addition ofPi rescued expression of PPi-associated genes in HPP cells, other mineral markers were not corrected. Different capital letters indicate intergroupstatistical differences (control versus HPP, with or without Pi treatment) within the same time point (day 1, 5, 15, or 20) for P = 0.05, using theKruskal-Wallis test followed by Student-Newman-Keuls method.
Periodontal Ligament Cells in Hypophosphatasia Volume 83 • Number 5
660

hypothesis that PPi regulation may be more active inthe periodontium compared to other dental tissuessuch as pulp/dentin, and more critical to cementogen-esis, i.e., a primary requirement for this tissue is a localPi/PPi environment that can promote mineralization.
Insight into the role of the Pi/PPi ratio on dental de-velopment and function not only identifies develop-mental regulation and pathologic mechanisms butmay inform regenerative therapies, and thus we haveextended this hypothesis to cementum regenerationstudies in mice.47 The results from these regenerationstudies support our findings here. Specifically, wedemonstrated that cementum regeneration in Ankknock-out mice (which feature PPi deficiency) wasmore rapid and more extensive when compared tocomparably treated control mice. Local modulationof Pi/PPi may prove to be a promising novel approachfor promoting regeneration of cementum.
CONCLUSIONS
Ex vivo and in vitro findings in the present study un-derscore the importance of controlling the Pi/PPi ratiotoward development of a functional periodontal appa-ratus and support Pi/PPi imbalance as the etiology ofHPP-associated cementum defects. Periodontal tis-
sues featured higher constitu-tive expression of centralregulators of PPi compared topulp tissues. PDL cells isolatedfrom individuals with HPP diag-nosed with a novel ALPL muta-tion exhibited reduced ALPactivity and capacity for miner-alization and dysregulated PPi
regulatory genes, all of whichwere corrected by normalizingthe Pi/PPi ratio in vitro. Theimportant influence of PPi me-tabolism on cementum devel-opment suggests that targetedmodulation of Pi/PPi may pro-videanovelapproach forachiev-ing cementum regeneration.
ACKNOWLEDGMENTS
Studies were supported by SaoPaulo State Research Founda-tion Grants 07/08192-5 and08/00534-7 (FHN), NationalInstitutes of Health (NIH)/Na-tional Institute of Dental andCraniofacial Research GrantDE15109 (MJS), and NIH Fo-garty International ResearchCollaboration Award Grant
5R03TW007590-03 (FHN, MJS). The authorsthank Mrs. Ana Paula Giorgetti (Campinas StateUniversity, School of Dentistry, Piracicaba, SaoPaulo, Brazil) for clinical support and Dr. Marisi Aidar(Campinas State University, School of Dentistry, Pi-racicaba, Sao Paulo, Brazil) for genetic analyses. Theauthors report no conflicts of interest related to thisstudy.
REFERENCES1. Foster BL, Tompkins KA, Rutherford RB, et al. Phos-
phate: Known and potential roles during developmentand regeneration of teeth and supporting structures.Birth Defects Res C Embryo Today 2008;84:281-314.
2. Murshed M, Harmey D, Millan JL, McKee MD,Karsenty G. Unique coexpression in osteoblasts ofbroadly expressed genes accounts for the spatialrestriction of ECM mineralization to bone. Genes Dev2005;19:1093-1104.
3. Harmey D, Hessle L, Narisawa S, Johnson KA,Terkeltaub R, Millan JL. Concerted regulation of in-organic pyrophosphate and osteopontin by akp2,enpp1, and ank: An integrated model of the patho-genesis of mineralization disorders. Am J Pathol 2004;164:1199-1209.
4. van den Bos T, Handoko G, Niehof A, et al. Cementumand dentin in hypophosphatasia. J Dent Res 2005;84:1021-1025.
Figure 4. (cont.)
J Periodontol • May 2012 Rodrigues, Foster, Silverio, et al.
661

5. Nociti FH Jr., Berry JE, Foster BL, et al. Cementum:A phosphate-sensitive tissue. J Dent Res 2002;81:817-821.
6. Chapple IL. Hypophosphatasia: Dental aspects andmode of inheritance. J Clin Periodontol 1993;20:615-622.
7. McKee MD, Nakano Y, Masica DL, et al. Enzymereplacement therapy prevents dental defects in amodel of hypophosphatasia. J Dent Res 2011;90:470-476.
8. Beertsen W, VandenBos T, Everts V. Root develop-ment in mice lacking functional tissue non-specificalkaline phosphatase gene: Inhibition of acellularcementum formation. J Dent Res 1999;78:1221-1229.
9. Olsson A, Matsson L, Blomquist HK, Larsson A, SjodinB. Hypophosphatasia affecting the permanent denti-tion. J Oral Pathol Med 1996;25:343-347.
10. Liu H, Li J, Lei H, Zhu T, Gan Y, Ge L. Genetic etiologyand dental pulp cell deficiency of hypophosphatasia.J Dent Res 2010;89:1373-1377.
11. Foster BL, Nagatomo KJ, Bamashmous SO, et al. Theprogressive ankylosis protein regulates cementumapposition and extracellular matrix composition. CellsTissues Organs 2011:194:382-405.
12. Aidar M, Line SR. A simple and cost-effective protocolfor DNA isolation from buccal epithelial cells. BrazDent J 2007;18:148-152.
13. Mornet E, Taillandier A, Peyramaure S, et al. Identifi-cation of fifteen novel mutations in the tissue-nonspecific alkaline phosphatase (TNSALP) gene inEuropean patients with severe hypophosphatasia.Eur J Hum Genet 1998;6:308-314.
14. Rodrigues TL, Marchesan JT, Coletta RD, et al. Effectsof enamel matrix derivative and transforming growthfactor-beta1 on human periodontal ligament fibro-blasts. J Clin Periodontol 2007;34:514-522.
15. Foster BL, Nociti FH Jr., Swanson EC, et al. Regula-tion of cementoblast gene expression by inorganicphosphate in vitro. Calcif Tissue Int 2006;78:103-112.
16. Rutherford RB, Foster BL, Bammler T, Beyer RP, SatoS, Somerman MJ. Extracellular phosphate alterscementoblast gene expression. J Dent Res 2006;85:505-509.
17. Hessle L, Johnson KA, Anderson HC, et al. Tissue-nonspecific alkaline phosphatase and plasma cellmembrane glycoprotein-1 are central antagonisticregulators of bone mineralization. Proc Natl Acad SciUSA 2002;99:9445-9449.
18. Arceo N, Sauk JJ, Moehring J, Foster RA, SomermanMJ. Human periodontal cells initiate mineral-likenodules in vitro. J Periodontol 1991;62:499-503.
19. Nohutcu RM, McCauley LK, Koh AJ, Somerman MJ.Expression of extracellular matrix proteins in humanperiodontal ligament cells during mineralizationin vitro. J Periodontol 1997;68:320-327.
20. Kaneda T, Miyauchi M, Takekoshi T, et al. Character-istics of periodontal ligament subpopulations obtainedby sequential enzymatic digestion of rat molar peri-odontal ligament. Bone 2006;38:420-426.
21. MacNeil RL, Berry J, D’Errico J, Strayhorn C,Piotrowski B, Somerman MJ. Role of two mineral-associated adhesion molecules, osteopontin and bonesialoprotein, during cementogenesis. Connect TissueRes 1995;33:1-7.
22. McKee MD, Zalzal S, Nanci A. Extracellular matrixin tooth cementum and mantle dentin: Localization
of osteopontin and other noncollagenous proteins,plasma proteins, and glycoconjugates by electronmicroscopy. Anat Rec 1996;245:293-312.
23. D’Errico JA, MacNeil RL, Takata T, Berry J, StrayhornC, Somerman MJ. Expression of bone associatedmarkers by tooth root lining cells, in situ and in vitro.Bone 1997;20:117-126.
24. Ye L, Zhang S, Ke H, Bonewald LF, Feng JQ. Periodontalbreakdown in the Dmp1 null mouse model of hypo-phosphatemic rickets. J Dent Res 2008;87:624-629.
25. Berendsen AD, Smit TH, Schoenmaker T, et al. Inor-ganic phosphate stimulates DMP1 expression inhuman periodontal ligament fibroblasts embedded inthree-dimensional collagen gels. Cells Tissues Organs2010;192:116-124.
26. Fleisch H, Bisaz S. Mechanism of calcification: In-hibitory role of pyrophosphate. Nature 1962;195:911.
27. Whyte MP. Hypophosphatasia and the role of alkalinephosphatase in skeletal mineralization. Endocr Rev1994;15:439-461.
28. Mornet E. Hypophosphatasia. Orphanet J Rare Dis2007;2:40.
29. Bruckner RJ, Rickles NH, Porter DR. Hypophospha-tasia with premature shedding of teeth and aplasia ofcementum. Oral Surg Oral Med Oral Pathol 1962;15:1351-1369.
30. Hu JC, Plaetke R, Mornet E, et al. Characterization ofa family with dominant hypophosphatasia. Eur J OralSci 2000;108:189-194.
31. Beumer J 3rd, Trowbridge HO, Silverman S Jr.,Eisenberg E. Childhood hypophosphatasia and thepremature loss of teeth. A clinical and laboratory studyof seven cases. Oral Surg Oral Med Oral Pathol 1973;35:631-640.
32. Wei KW, Xuan K, Liu YL, et al. Clinical, pathologicaland genetic evaluations of Chinese patients withautosomal-dominant hypophosphatasia. Arch OralBiol 2010;55:1017-1023.
33. Harmey D, Johnson KA, Zelken J, et al. Elevatedskeletal osteopontin levels contribute to the hypo-phosphatasia phenotype in Akp2(-/-) mice. J BoneMiner Res 2006;21:1377-1386.
34. Gurley KA, Chen H, Guenther C, et al. Mineral formationin joints caused by complete or joint-specific loss ofANK function. J Bone Miner Res 2006;21:1238-1247.
35. Ho AM, Johnson MD, Kingsley DM. Role of the mouseank gene in control of tissue calcification and arthritis.Science 2000;289:265-270.
36. Fong H, Foster BL, Sarikaya M, Somerman MJ.Structure and mechanical properties of Ank/Ankmutant mouse dental tissues—An animal model forstudying periodontal regeneration. Arch Oral Biol2009;54:570-576.
37. Bosshardt DD, Schroeder HE. Cementogenesis re-viewed: A comparison between human premolarsand rodent molars. Anat Rec 1996;245:267-292.
38. Kaipatur NR, Murshed M, McKee MD. Matrix Glaprotein inhibition of tooth mineralization. J Dent Res2008;87:839-844.
39. Beertsen W, Niehof A, Everts V. Effects of 1-hydroxy-ethylidene-1, 1-bisphosphonate (HEBP) on the forma-tion of dentin and the periodontal attachment apparatusin the mouse. Am J Anat 1985;174:83-103.
40. Jayawardena CK, Takahashi N, Watanabae E, TakanoY. On the origin of intrinsic matrix of acellular extrinsicfiber cementum: Studies on growing cementum pearls
Periodontal Ligament Cells in Hypophosphatasia Volume 83 • Number 5
662

of normal and bisphosphonate-affected guinea pigmolars. Eur J Oral Sci 2002;110:261-269.
41. Taillandier A, Lia-Baldini AS, Mouchard M, et al. Twelvenovel mutations in the tissue-nonspecific alkalinephosphatase gene (ALPL) in patients with various formsof hypophosphatasia. Hum Mutat 2001;18:83-84.
42. Whyte MP, Vrabel LA, Schwartz TD. Alkaline phos-phatase deficiency in cultured skin fibroblasts frompatients with hypophosphatasia: Comparison of theinfantile, childhood, and adult forms. J Clin EndocrinolMetab 1983;57:831-837.
43. Whyte MP, Vrabel LA. Infantile hypophosphatasiafibroblasts proliferate normally in culture: Evidenceagainst a role for alkaline phosphatase (tissue non-specific isoenzyme) in the regulation of cell growthand differentiation. Calcif Tissue Int 1987;40:1-7.
44. Fedde KN, Whyte MP. Alkaline phosphatase (tissue-nonspecific isoenzyme) is a phosphoethanolamineand pyridoxal-59-phosphate ectophosphatase: Normaland hypophosphatasia fibroblast study. Am J HumGenet 1990;47:767-775.
45. Wennberg C, Hessle L, Lundberg P, et al. Functionalcharacterization of osteoblasts and osteoclasts fromalkaline phosphatase knockout mice. J Bone MinerRes 2000;15:1879-1888.
46. Macneil RL, Sheng N, Strayhorn C, Fisher LW,Somerman MJ. Bone sialoprotein is localized to theroot surface during cementogenesis. J Bone Miner Res1994;9:1597-1606.
47. Rodrigues TL, Nagatomo KJ, Foster BL, Nociti FH,Somerman MJ. Modulation of phosphate/pyrophos-phate metabolism to regenerate the periodontium: Anovel in vivo approach. J Periodontol 2011;82:1757-1766.
Correspondence: Dr. Francisco Nociti, Department ofProsthodontics and Periodontics, Division of Periodontics,School of Dentistry, Campinas State University, Piraci-caba, Sao Paulo, Brazil. E-mail: [email protected].
Submitted May 23, 2011; accepted for publication August20, 2011.
J Periodontol • May 2012 Rodrigues, Foster, Silverio, et al.
663