contenidos0b3945371a06d9a2.jimcontent.com/download/version/1308626853/module...gloria gonzález...
TRANSCRIPT

Contenido
Diagnóstico
Ensayo en fragmentos de hojas de bananos y plátanos (Musa spp.) para el estudio a nivel monocíclico de la evolución de los síntomas de la sigatoka negra causada por Mycosphaerella fijiensis Morelet 3Luis Pérez Vicente, Michel Pérez Miranda, María Isabel Jiménez y María Jama
Fitopatógenos en los cultivos de pastos y forrajes en Cuba 11Gloria González Arias, María O. López Mesa, Zenaida Amat Novo, Giselle Estrada Vilardel, Danay López Manes, Blanca Bernal Areces, Ana Granda, Giselle Rodríguez Gutiérrez, Leidys Figueredo González, Ana D. Pupo Zayas, María Ramos, Mercedes González, Martha Ruiz Guardado, Idiel Pérez Guevara, César Nápoles Albanés, Graciela García Rivero, Carmen R. Sánchez, Carmen Buchillón y Mirtha López
Causas de la aparición de la enfermedad mancha verde en hojas de tabaco (Nicotiana tabacum L.) 19Danneys Armario Aragón, Sinesio Torres García, Oguelis Rodríguez Pérez, Iban Arredondo Quevedo, Yunetsy Girado López y Ángel Mollineda Trujillo
Ecología
Efecto de los medios de cultivo y la temperatura en el crecimiento de Cladosporium fulvum Cooke (Sin. Passalora fulva) 25Alexander Bernal Cabrera, Benedicto Martínez Coca, Manuel Díaz Castellanos, Lidcay Herrera Isla y Yanelys Alonso Díaz
Control biológico
Reproducción de Heterorhabditis indica en cultivos bidimensionales elaborados con proteína animal 29Yirina Valdés Vázquez, Antonio A. Lobaina Audevert, María E. Márquez Gutiérrez, Maylen Gómez Pacheco y Mercedes Escobar Hernández
Introducción de la avispa de Costa de Marfil Cephalonomia stephanoderis Betrem (Hymenoptera: Bethylidae), parasitoide de la broca del fruto del cafeto Hypothenemus hampei Ferrari (Coleoptera: Scolytidae) en Cuba 33Eliel Peña Marrero, Mario García Hernández, Eliazar Blanco Rodríguez y Juan F. Barreras Gaytán
Manejo de plagas
Variabilidad de Mycosphaerella fijiensis Morelet. Estabilidad de la resistencia a sigatoka negra de los clones híbridos de la FHIA 37Michel Pérez Miranda, Luis Pérez Vicente, Roberto Trujillo y Dulce M. Betancourt
Efectos de una cobertura viva de Teramnus labialis (L.f.) Sprengel sobre las arvenses en campos citrícolas 49Iván R. Gutiérrez Rojas, María de los A. Felipe López, Rafael Pérez Carmenate, Dayamí Fontes y Vernon I. Daniel
Reseña
Manejo convencional y alternativo de la sigatoka negra en bananos: estado actual y perspectivas 55Luis Pérez Vicente
Comunicación corta
Nuevos aislados de Bacillus spp. antagonistas a Sclerotium rolfsi, Rhizoctonia solani y Pythium aphanidermatum 73Acenet I. Sosa López, Victoria Pazos Álvarez-Rivera y Marleny González García
Insectos del orden Lepidoptera detectados en plantas de noni (Morinda citrifolia L.) 75Nancy González García, Rubén Avilés Pacheco, Bienvenido Cruz Díaz y Yolanda Martínez Suárez
Resumen de tesis
Conservación de artrópodos benéficos en un sistema de producción agrícola urbano 77Yaril Matienzo Brito
Epidemiología de Erwinia spp. en el cultivo de la papa (Solanum tuberosum L.) y medidas para su control 79Yolanda Morales Gómez

2/fitosanidad
Contents
Phytosanitary diagnosis
An Assay in Banana and Plantain (Musa spp.) Leaf Fragments to Study the Evolution of Symptoms of Black Sigatoka Disease Caused by Mycosphaerella fijiensis Morelet at Monocyclic Level 3Luis Pérez Vicente, Michel Pérez Miranda, María Isabel Jiménez and María Jama
Phytopatogens in Grass and Forage Crops from Cuba 11Gloria González Arias, María O. López Mesa, Zenaida Amat Novo, Giselle Estrada Vilardel, Danay López Manes, Blanca Bernal Areces, Ana Granda, Giselle Rodríguez Gutiérrez, Leidys Figueredo González, Ana D. Pupo Zayas, María Ramos, Mercedes González, Martha Ruiz Guardado, Idiel Pérez Guevara, César Nápoles Albanés, Graciela García Rivero, Carmen R. Sánchez, Carmen Buchillón and Mirtha López
Causes of Green Spot Disease Appearance in Tobacco (Nicotiana tabacum L) 19Danneys Armario Aragón, Sinesio Torres García, Oguelis Rodríguez Pérez, Iban Arredondo Quevedo, Yunetsy Girado López and Ángel Mollineda Trujillo
Ecology
Effect of Culture Media and Temperature on the Growth of Cladosporium fulvum Cooke (Sin. Passalora fulva) 25Alexander Bernal Cabrera, Benedicto Martínez Coca, Manuel Díaz Castellanos, Lidcay Herrera Isla and Yanelys Alonso Diaz
Biological control
Reproduction of Heterorhabditis indica in Bidimentional Culture Media Elaborated with Animal Protein 29Yirina Valdés Vázquez, Antonio A. Lobaina Audevert, María E. Márquez Gutiérrez, Maylen Gómez Pacheco and Mercedes Escobar Hernández
Introduction of Ivory Coast Wasp Cephalonomia stephanoderis Betrem (Hymenoptera: Bethylidae), Parasitoid of Coffee Berry Borer Hypothenemus hampei Ferrari (Coleoptera: Scolytidae) in Cuba 33Eliel Peña Marrero, Mario García Hernádez, Eliazar Blanco Rodríguez and Juan F. Navarro Gaytán
Pest management
Variability of Mycosphaerella fijiensis Morelet. Durability of the Resistance to Black Sigatoka in the FHIA Hybrids Cultivars 37Michel Pérez Miranda, Luis Pérez Vicente, Roberto Trujillo and Dulce M. Betancourt
Effects of a Cover Crop Teramnus labialis (L.f.) Sprengel on Weed Populations in Citric Fields 49Iván R. Gutiérrez Rojas, María de los A. Felipe López, Rafael Pérez Carmenate, Dayamí Fontes and Vernon I. Daniel
Review
Conventional and Alternative Management of Black Sigatoka Disease in Banana. Current Status and Perspectives 55Luis Pérez Vicente
Short communication
New Bacillus spp. Isolates Antagonists to Sclerotium rolfsi, Rhizoctonia solani and Pythium aphanidermatum 73Acenet I. Sosa López, Victoria Pazos Álvarez-Rivera and Marleny González García
Lepidoptera Insects Detected on Noni Plants (Morinda citrifolia L.) 75Nancy González García, Rubén Avilés Pacheco, Bienvenido Cruz Díaz and Yolanda Martínez Suárez
Thesis abstract
Preservation of Benefits Arthropods in an Urban System of Agricultural Production 77Yaril Matienzo Brito
Epidemiology of Erwinia spp. in Potato Crop (Solanum tuberosum L.) and Control Measures 79Yolanda Morales Gómez

fitosanidad/3
FITOSANIDAD vol. 10, no. 1, marzo 2006
Dia
gnós
tico ENSAYO EN FRAGMENTOS DE HOJAS DE BANANOS
Y PLÁTANOS (MUSA SPP.) PARA EL ESTUDIO A NIVELMONOCÍCLICO DE LA EVOLUCIÓN DE LOS SÍNTOMASDE LA SIGATOKA NEGRA CAUSADA POR MYCOSPHAERELLAFIJIENSIS MORELET
Luis Pérez Vicente,1 Michel Pérez Miranda,1 María Isabel Jiménez2 y María Jama2
1 Instituto de Investigaciones de Sanidad Vegetal. Calle 110 no. 514 e/ 5a. B y 5a. F, Playa, Ciudad de La Habana, CP 116002 Centro Biotecnológico del Ecuador (CIBE) de la Escuela Politécnica del Litoral en Guayaquil, Ecuador
RESUMEN
Se describe el desarrollo de un ensayo de inoculación artificial so-bre fragmentos de hojas de clones de bananos y plátanos con dife-rentes niveles conocidos de resistencia parcial a sigatoka negramantenidos in vitro. Se colectaron fragmentos de hojas de 3,5 x 3,5 cmde la primera hoja completamente expandida (10 frag/clon), los cua-les se lavaron con agua estéril y colocaron en una placa de Petri de10 cm con agar agua + benzimidazol (20 g agar.L–1 + 40 µg.mL–1 debenzimidazol). Se inocularon con diferentes concentraciones deconidios de M. fijiensis obtenidos de cultivos in vitro del patógeno eincubaron bajo luz fluorescente. La cantidad de lesiones y su evolu-ción por estados durante un ciclo infeccioso se evaluó en días con-secutivos. Se determinó la concentración óptima de inóculo para eldesarrollo del ensayo. Existió una fuerte correspondencia entre lascurvas de aparición de lesiones en relación con el tiempo en losfragmentos de hoja in vitro a nivel monocíclico y las curvas de desarro-llo de la enfermedad en el campo a nivel policíclico en los clonesrespectivos, por lo que el método es aplicable para estimar el nivelde resistencia parcial de los clones frente a sigatoka negra y paracomparar la agresividad de los aislamientos bajo condiciones con-troladas. La duración de la vida verde de las hojas en el ensayo nopermitió determinar la intensidad de la reproducción sexual del pató-geno.
Palabras clave: resistencia parcial, sigatoka negra, Mycosphaerellafijiensis, ensayo monocíclico
ABSTRACT
An assay of artificial inoculation on banana leaf fragments in vitro wasstudied. Fragments of 3.5 x 3.5 cm of the first fully expanded leaf (10fragments/cultivar) were washed with sterilized water and placed in 10cm Petri dishes containing 20 g agar.L–1 + 40 µg.mL–1 of benzimidazol.The fragments were inoculated with different concentrations of M.fijiensis conidia obtained from in vitro cultures the pathogen and wereincubated under fluorescent light. The number of lesions and itsevolution to different stages of development were assessed daily in asingle infection cycle. The optimal conidial concentration for thedevelopment of the assay was determined. A strong correlation betweenthe curves of development of lesions on time in the fragments in vitroat monocyclic level and the curves of development of the disease ontime in different cultivars at polycyclic level in the field was observedallowing sustain that the method is useful to compare the level ofpartial resistance of Musa cultivars to Black Sigatoka disease and theaggressiveness of the populations under controlled conditions. Theduration of the fragments green life in vitro in the experiments do notallowed assess the intensiveness of the sexual reproduction of thepathogen. Further research is required to increasing the green life ofthe fragments of banana leaves to allowing the formation of the sexualreproduction of the pathogen in the spots.
Key words: partial resistance, black sigatoka, Mycosphaerellafijiensis, monocyclic assay
INTRODUCCIÓN
La aparición de la sigatoka negra en Cuba causadapor Mycosphaerella fijiensis Morelet, a finales de 1990[Vidal, 1992], tuvo un impacto marcado en los costosde producción y especialmente en la estructura clonalde la superficie del país plantada de musáceas [Pérez etal., 2002].
El uso de variedades resistentes es sin duda la estrate-gia más económica y ambientalmente sostenible de lu-cha contra esta enfermedad. Se han llevado a cabo nu-
merosos estudios sobre la resistencia de los clones debananos y plátanos a M. fijiensis [Meredith yLawrence, 1970; Firman, 1972; Fouré et al., 1984; Fouréet al., 1990; Fouré, 1994; Hernández y Pérez, 2001; Rowe,1998].
Fouré (1994) describió dos tipos de reacciones en Musafrente a sigatoka negra: la reacción de incompatibili-dad –resistencia muy alta o hipersensibilidad, observa-da en Yangambi Km 5 del subgrupo Ibota, AAA–, don-

Pérez Vicente y otros
4/fitosanidad
de no ocurre reproducción del patógeno, y la reacciónde compatibilidad con desarrollo de los síntomas y re-producción del patógeno, en la que se diferencian laresistencia parcial, que se expresa por un alargamientodel ciclo y disminución de la reproducción del patógenocon una baja tasa de incremento de la enfermedad, y lasensibilidad observada en los clones Cavendish y pláta-nos (AAB), caracterizada por un ciclo de la enferme-dad corto, reproducción intensa, alta tasa de incrementode la enfermedad y grandes afectaciones del áreafotosintética de la planta.
Hernández y Pérez (2001) determinaron la reacción ycomponentes de la resistencia a sigatoka negra de losclones FHIA 23, FHIA 2, FHIA 3, FHIA 18, SH 3436y de un grupo importante de clones naturales del ban-co de germoplasma de Cuba, entre los que se encontra-ban diferentes clones pertenecientes al subgrupoCavendish (AAA), plátanos (AAB), el Yangambi Km 5(subgrupo Ibota, AAA), el Paka (AB) y el BurroCEMSA (ABB), mantenidos sin tratamiento confungicidas durante todo el tiempo que duraron las ob-servaciones. Los clones FHIA con resistencia parcial asigatoka negra mostraron un alargamiento esta-dísticamente significativo de la duración en días entrela emergencia de las hojas y la aparición y duración dela evolución de las lesiones, en relación con el clon Granenano susceptible. La resistencia parcial de estos clonesse expresa mediante el alargamiento de la evolución otransición de los síntomas de rayas a necrosis y en lareducción del número de pseudotecios por mancha[Pérez, 1998]. En el Yangambi Km 5 las aloinfeccionesa partir de inóculo producido en parcelas vecinas de-terminaron la aparición de numerosas manchas ydefoliaciones, pero no la formación de espermogonios ypseudotecios.
En Cuba más de 11 000 ha de superficie de bananos yplátanos están cultivadas de híbridos FHIA [Pérez etal., 2002]. Por cuanto la resistencia parcial es de carác-ter cuantitativo, el ambiente tiene una fuerte influen-cia en el resultado de la evaluación de la reacción de losclones, por lo que es altamente deseable disponer de unensayo bajo condiciones controladas que permita de-terminar la reacción de los clones y los cambios de agre-sividad de las poblaciones del patógeno bajo condicio-nes ambientales estables y comparables.
El Hadrami et al. (1998, 2000) desarrollaron ensayos enfragmentos de hojas in vitro de diferentes cultivares debananos y plátanos, pero no publicaron los detalles delos procedimientos de trabajo.
El objetivo del presente trabajo fue el estudio de laevolución de los síntomas de sigatoka negra causadapor M. fijiensis sobre fragmentos de hojas in vitro bajocondiciones controladas (estudio de la epidemiología desigatoka negra a nivel monocíclico y su efecto sobre loscomponentes que caracterizan la resistencia parcial).
MATERIALES Y MÉTODOS
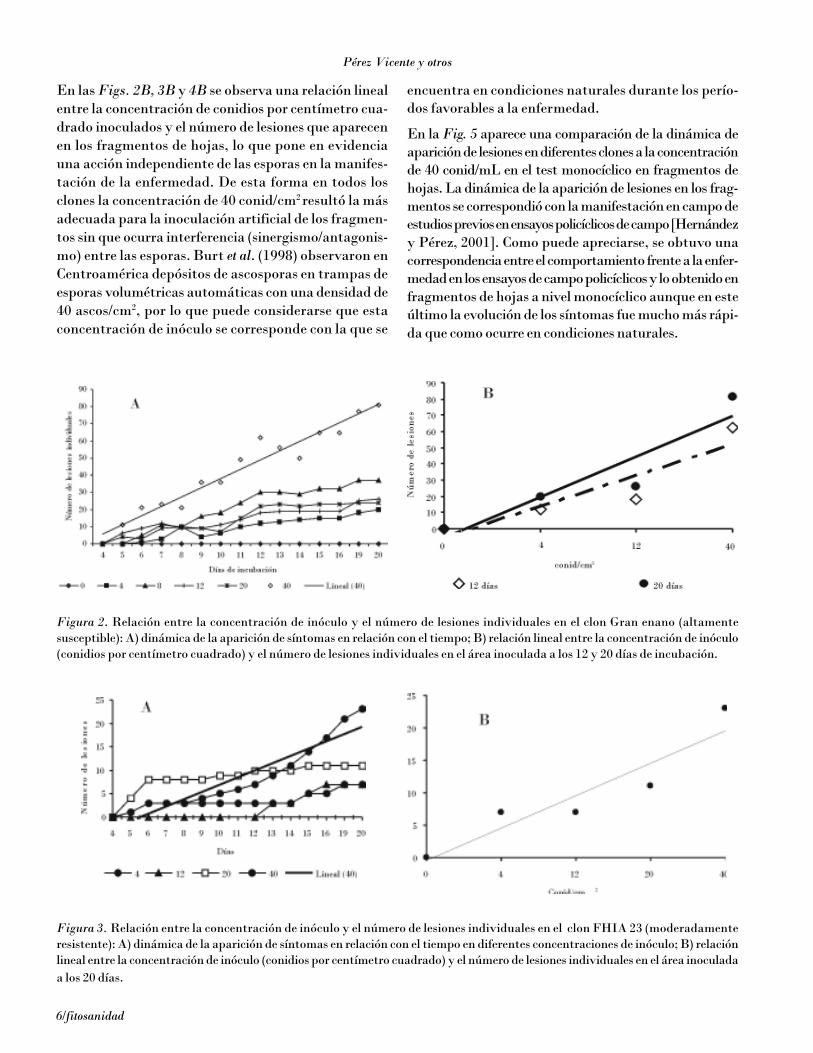
Con el objetivo futuro de la comparación de la agresivi-dad de diferentes aislamientos de M. fijiensis y de eli-minar la influencia ambiental en los ensayos cuantita-tivos de desarrollo de la enfermedad en diferentes clones,se estudió un procedimiento de inoculación en fragmen-tos de hojas, similar al desarrollado por El Hadrami etal. (1998, 2000) a nivel monocíclico sobre un ciclo deinfección, para lo cual se plantaron plantas de los clonesGran enano, FHIA 2, FHIA 18, FHIA 23 y YangambiKm 5 en macetas dentro de un aislador. Se colectaronfragmentos de la primera hoja completamente expan-dida de 3,5 x 3,5 cm y se lavaron con agua destiladaestéril, luego se colocaron en una placa de Petri conagar-agua + 40 µg.mL–1 de benzimidazol, con el haz de lahoja en contacto con el agar (Fig. 1A). Para evitar quelos extremos del fragmento de hoja se separaran delagar, se colocó sobre este una lámina transparente plás-tica estéril del mismo tamaño de la superficie interiorde la placa, a la que se le había eliminado un cuadradode 3 x 3 cm en su centro (Fig. 1B), de forma que esesector de la hoja quedó expuesto para poder realizarlas inoculaciones [J. Carlier, comunicación personal].
La superficie expuesta de los fragmentos se inoculó con50 µL de suspensiones conidiales de M. fijiensis. Paradeterminar las concentraciones de inóculo más adecua-das para el ensayo se inocularon en 10 fragmentos dehoja tomados de los clones Gran enano, FHIA 23,FHIA 18, FHIA 2 y FHIA 21, conidios –procedentesde cultivos de 14 días de edad– a las concentraciones de0, 4, 8, 12, 20 y 40 conid/cm2. Una vez selladas las pla-cas con parafilm se incubaron en un cuarto climatizado(25-27°C) bajo dos tubos de luz fluorescente (Fig. 1C).Diariamente se contó el número de lesiones que apare-cieron por estadios de evolución (Figs. 1D y 1E). Secomparó el número de lesiones en el fragmento inocula-do de cada clon a la concentración de 40 conid/cm2 dehoja. Se midió además la duración en días de la evolu-ción de los síntomas.
Los resultados de las pruebas monocíclicas en fragmen-tos se compararon con los de las evaluaciones del de-

fitosanidad/5
Ensayo en fragmentos de hojas...
sarrollo de la enfermedad en los estudios policíclicos encampo previamente realizados por el primer autor[Hernández y Pérez, 2001].
RESULTADOS Y DISCUSIÓN
Cualquier procedimiento de inoculación de patógenospara evaluar la reacción de las variedades requiere: a)una evaluación previa de la relación entre el número deesporas depositadas por centímetro cuadrado de su-perficie del hospedante y el número de lesiones que seproducen en consecuencia; b) demostrar a diferentescargas de inóculo la tendencia lineal de las curvas paraevitar los efectos antagónicos y sinérgicos entre las es-poras en relación con la manifestación de la enferme-dad; c) que la carga de esporas utilizada sea correspon-diente en gran medida con la que ocurre naturalmentepara poder estimar de forma real cuál será el compor-tamiento de los genotipos en estudio bajo condicionesnaturales [Van der Plank, 1963, 1968].
En todos los experimentos realizados la aparición desíntomas y su evolución en los fragmentos de hojas fuemás rápida que en condiciones naturales en los mismosclones en el campo. En las Figs. 2A, 3A y 4A aparece ladinámica de aparición de lesiones en relación con el tiem-po en el test monocíclico en fragmentos de hojas deGran enano (altamente susceptible), FHIA 23 (resis-tencia parcial moderada) y FHIA 2 (alta resistenciaparcial), respectivamente, para diferentes concentra-ciones de inóculo. En el clon Gran enano se observó unincremento del número de lesiones en el tiempo en to-das las concentraciones de inóculo. La concentraciónde 40 conid/cm2 de fragmento de hoja inoculada brindóla mayor cantidad de lesiones y se alcanzó a los 20 díashasta 90. En el caso del FHIA 02 también hay un in-cremento lineal en el tiempo, pero de menor pendiente,por lo que a los 20 días a la concentración de 40 conid/cm2
aparecieron 23 lesiones en contraste con más de 90 ob-servadas en el Gran enano. El clon FHIA 23 mostróvalores intermedios entre ambos.
Figura 1. Fragmentos de hoja en placas con agar + benzimidazol.
Fragmento sobre el agar Lámina plástica para evitar separación delfragmento del agar durante incubación.
Placas selladas con parafilm incubando a27°C bajo dos tubos de luz fluorescente.
Aparición de los primeros síntomas de laslesiones.
Desarrollo de síntomas en diferentes es-tados.
A B
C ED
Pérez Vicente y otros
6/fitosanidad
En las Figs. 2B, 3B y 4B se observa una relación linealentre la concentración de conidios por centímetro cua-drado inoculados y el número de lesiones que aparecenen los fragmentos de hojas, lo que pone en evidenciauna acción independiente de las esporas en la manifes-tación de la enfermedad. De esta forma en todos losclones la concentración de 40 conid/cm2 resultó la másadecuada para la inoculación artificial de los fragmen-tos sin que ocurra interferencia (sinergismo/antagonis-mo) entre las esporas. Burt et al. (1998) observaron enCentroamérica depósitos de ascosporas en trampas deesporas volumétricas automáticas con una densidad de40 ascos/cm2, por lo que puede considerarse que estaconcentración de inóculo se corresponde con la que se
encuentra en condiciones naturales durante los perío-dos favorables a la enfermedad.
En la Fig. 5 aparece una comparación de la dinámica deaparición de lesiones en diferentes clones a la concentraciónde 40 conid/mL en el test monocíclico en fragmentos dehojas. La dinámica de la aparición de lesiones en los frag-mentos se correspondió con la manifestación en campo deestudios previos en ensayos policíclicos de campo [Hernándezy Pérez, 2001]. Como puede apreciarse, se obtuvo unacorrespondencia entre el comportamiento frente a la enfer-medad en los ensayos de campo policíclicos y lo obtenido enfragmentos de hojas a nivel monocíclico aunque en esteúltimo la evolución de los síntomas fue mucho más rápi-da que como ocurre en condiciones naturales.
Figura 2. Relación entre la concentración de inóculo y el número de lesiones individuales en el clon Gran enano (altamentesusceptible): A) dinámica de la aparición de síntomas en relación con el tiempo; B) relación lineal entre la concentración de inóculo(conidios por centímetro cuadrado) y el número de lesiones individuales en el área inoculada a los 12 y 20 días de incubación.
Figura 3. Relación entre la concentración de inóculo y el número de lesiones individuales en el clon FHIA 23 (moderadamenteresistente): A) dinámica de la aparición de síntomas en relación con el tiempo en diferentes concentraciones de inóculo; B) relaciónlineal entre la concentración de inóculo (conidios por centímetro cuadrado) y el número de lesiones individuales en el área inoculadaa los 20 días.
fitosanidad/7
Ensayo en fragmentos de hojas...
Figura 4. Relación entre la concentración de inóculo y el número de lesiones individuales en el clon FHIA 2 (altamente resistente):A) dinámica de la aparición de síntomas en relación con el tiempo en diferentes concentraciones de inóculo; B) relación lineal entre laconcentración de inóculo (conidios por centímetro cuadrado) y el número de lesiones individuales en el área inoculada a los 20 días.
Figura 5. Comparación de los resultados de la reacción de los clones Gran enano, FHIA 02, FHIA 18 y FHIA 23 a las infeccionesnaturales en las pruebas policíclicas de campo [Hernández y Pérez, 2001] y a las inoculaciones con conidios en pruebas monocíclicassobre fragmentos de hojas in vitro.
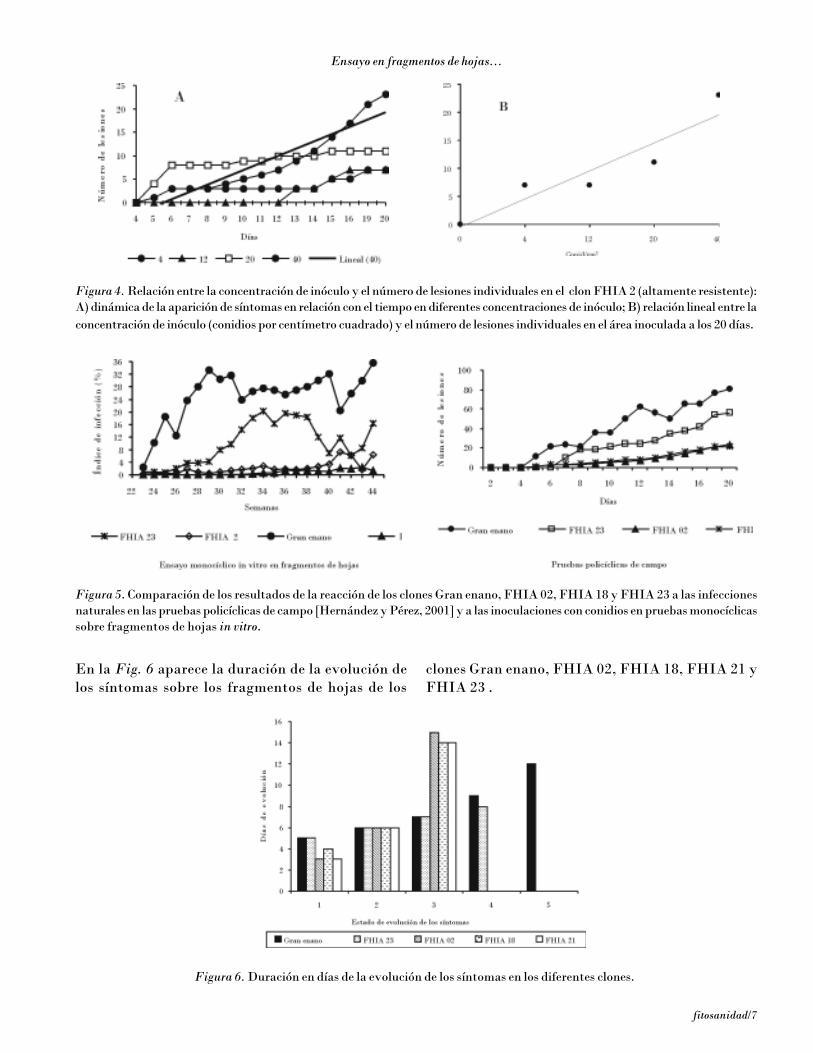
En la Fig. 6 aparece la duración de la evolución delos síntomas sobre los fragmentos de hojas de los
clones Gran enano, FHIA 02, FHIA 18, FHIA 21 yFHIA 23 .
Figura 6. Duración en días de la evolución de los síntomas en los diferentes clones.

Pérez Vicente y otros
8/fitosanidad
Se hizo evidente el efecto de la resistencia parcial sobrela evolución de los síntomas. La evolución de laincubación y de la transición a manchas fue más cortaen el Gran enano (altamente susceptible) que en losclones con resistencia parcial, en correspondencia conlos informes de Fouré et al. (1984 y 1990) y con las ob-servaciones previas en clones FHIA [Hernández yPérez, 2001]. En el FHIA 18 y el FHIA 21 la evoluciónde los síntomas hasta el estado 3 fue el doble más largaque en el Gran enano. Pudo observarse la aparición desíntomas 4 en el Gran enano y FHIA 23, y de estado 5solo en Gran enano. El FHIA 02 detuvo su evoluciónen el estado 3 de los síntomas, mientras que el FHIA 23(FHIA 1-1) lo hizo en el estado 4. Se logró mantenerdurante 25 días las hojas verdes, lo que es insuficientepara poner en evidencia toda la evolución de los sínto-mas, en especial cuando los clones tienen una resisten-cia parcial elevada, pues no es posible visualizar los es-tados más avanzados de los síntomas y la formación defructificaciones.
Los resultados muestran la potencialidad del ensayoen fragmentos de hojas para la evaluación de la reac-ción de resistencia parcial de clones de bananos; sinembargo, como aspecto negativo hay que considerar quemuestra variación de la supervivencia de los fragmen-tos de hojas, aún provenientes de una misma hoja, porlo que debe continuarse el estudio en el futuro de losfactores que inciden en esto para la optimización delensayo y poder evaluar la producción potencial de cuer-pos fructíferos.
Es por primera vez que se documentan las condicionesespecíficas para la conducción de los ensayos en frag-mentos de hojas. Los informes anteriores de El Hadramiet al. (1998, 2000) no establecen las condiciones especí-ficas para desarrollarlos.
CONCLUSIONES
• La evolución de la enfermedad es más rápida en losfragmentos de hojas in vitro que la ocurrida en lasplantas en campo.
• La curva del desarrollo de lesiones con el tiempo enun ciclo de infección en los fragmentos de hojas delos diferentes clones muestra una alta correspondenciacon la curva de desarrollo policíclico de la enferme-dad en relación con el tiempo en el campo, por lo quees posible estimar el nivel de resistencia parcial delos clones a partir de este ensayo.
• Este método puede utilizarse para comparar la agre-sividad en un solo ciclo infectivo de diferentes pobla-ciones de M. fijiensis bajo condiciones controladasde inoculación, así como para evaluar el nivel de laresistencia parcial de los clones al patógeno, basadaen la velocidad de evolución de las lesiones. Ello re-quiere posterior afinamiento para lograr alargar la vidaverde de los fragmentos con el objetivo de observar lareproducción sexual del patógeno en las manchas.
Los autores desean hacer constar y expresar su agrade-cimiento al INIBAP por financiar parte de los estudiosrealizados; al doctor J. Carlier por las sugerencias rea-lizadas en relación con la metodología del ensayo; al es-timado amigo, ya fallecido, doctor Rodolfo Maribona, porsu acogida en el Laboratorio de Fitopatología del CIBEen la ESPOL en Guayaquil, donde se replicaron partede los ensayos durante un entrenamiento ofrecido por elprimer autor a investigadores de esa institución.
REFERENCIAS
Burt, P. J. A.; J. Rutter; F. Ramírez: «Airborne Spore Loads and MesoscaleDispersal of the Fungal Pathogens Causing Sigatoka Diseases inBanana and Plantain», Aerobiology 14:209-214, 1998.
El Hadrami, A.; M. F. Zapater; F. Lapeyre; C. Abadie; J. Carlier: «Evaluationsur fragments foliaires en survie de la résistance partielle du bananieret de l’agressivité de Mycosphaerella fijiensis agent causal de lacercosporiose noire», CIRAD-FLHOR, Montpellier, Francia, FA176790,INIBAP’s data base Musalit 2001, 1998.
El Hadrami, A.; C. Abadie; J. Carlier: «Evaluation de la résistance partielledes bananiers à Mycosphaerella fijiensis (maladie des raies noires)en conditions contrôlées et au champ», Journées Jean Chevaugeon,Rencontres de Mycologie-phytopathologie, Aussois, Francia, 2000/03/05-09, 1p, INIBAP’s database Musalit 2001, 2000.
Firman, I. D.: «Susceptibility of Banana Cultivars to Fungus Diseases inFiji», Trop. Agric. Trinidad, 49:189-196, 1972.
Fouré, E.; A. Mouliom-Pefoura; X. Mourichon: «Etude de la sensibilitévarietale des bananiers et des plantains a Mycosphaerella fijiensisau Cameroun. Caractérization de la résistance au champ desbananiers et des plantains appartenent au divers groupsgénétiques», Fruits 45:339-345, 1990.
Fouré, E.: «Leaf Spot Diseases of Bananas and Plantain Caused byMycosphaerella musicola and M. fijiensis», The Improvement andTesting of Musa: a Global Partnership. Proceedings of the First Glo-bal Conference of the International Musa Testing Program, INIBAP,Held at FHIA, Honduras, April 27-30, 1994.
Fouré, E.; M. Grisoni; R. Zurfluh: «Les cercosporioses du bananier etleurs traitements. Comportement des variétés. II. Etude de la sensibilitédes bananiers et plantains á Mycosphaerella fijiensis Morelet et desquelques caractéristiques biologiques de la maladie des raies noiresau Gabon», Fruits 39:365-378, 1984.
Hernández, A.; L. Pérez: «Reaction of Banana and Plantain Cultivars toBlack Sigatoka Caused by Mycosphaerella fijiensis Morelet.Epidemiological components of the resistance», Fitosanidad 5 (3):9-16, 2001.
Pérez, L.: «Sigatoka negra (Mycosphaerella fijiensis Morelet) de

fitosanidad/9
Ensayo en fragmentos de hojas...
bananos y plátanos (Musa spp.) en Cuba. Biología, epidemiología ymanejo integrado de la enfermedad», Simposium Internacional sobresigatoka negra, Manzanillo, Colima, México, 8-10 de julio de 1998.
Pérez, L.; J. M. Álvarez; M. Pérez: «Economic Impact and Managementof Black Leaf Streak Disease in Cuba», Mycosphaerella Leaf SpotDiseases of Bananas: Present Status and Outlook. Proceedings ofthe International Workshop on Mycosphaerella Leaf Spot Diseases,San José, Costa Rica, 2002, pp. 71-83.
Rowe, P.: «Latest Developments in the FHIA Banana and PlantainBreeding Program: Bred Hybrids Are Now Being Grown Commercially»,
Memorias del Seminario Internacional sobre Producción de Plátano,Armenia Quindío, Colombia, 4-8 de mayo de 1998.
Meredith, D. S.; J. S. Lawrence: «Black Leaf Streak of Bananas(Mycosphaerella fijiensis). Susceptibility of Cultivars», Trop. Agric.Trinidad, 47:275-287, 1970.
Van der Plank, J. E.: Plant Disease Epidemiology, Academic PressNew York, 1963, pp.180-181.
––––: Disease Resistance In Plants, Academic Press New York, 1968.
Vidal, A.: «Sigatoka negra en Cuba. En nuevos focos de plagas yenfermedades», Boletín Fitosanitario, FAO 40:1-2, 1992.

10/fitosanidad

fitosanidad/11
FITOSANIDAD vol. 10, no. 1, marzo 2006
FITOPATÓGENOS EN LOS CULTIVOS DE PASTOSY FORRAJES EN CUBA
Gloria González Arias,1 María O. López Mesa,1 Zenaida Amat Novo,1 Giselle Estrada Vilardel,1 DanayLópez Manes,1 Blanca Bernal Areces,2 Ana Granda,3 Giselle Rodríguez Gutiérrez,4 Leidys FigueredoGonzález,4 Ana D. Pupo Zayas,4 María Ramos,5 Mercedes González,5 Martha Ruiz Guardado,5 IdielPérez Guevara,6 César Nápoles Albanés,6 Graciela García Rivero,6 Carmen R. Sánchez,7 CarmenBuchillón7 y Mirtha López8
1 Instituto de Investigaciones de Sanidad Vegetal. Calle 110 no. 514 e/ 5a. B y 5a. F, Playa, Ciudad de La Habana, CP 11600, [email protected] Instituto de Investigaciones Hortícolas Liliana Dimitrova. Carretera a Bejucal Km 33½, Quivicán, La Habana3 LAPROSAV. Ave. 25 no. 23011 e/ 230 y 234, La Coronela, Playa, Ciudad de La Habana4 LAPROSAV. Antonio Maceo 22 e/ J. Agüero y A. Guardia, Las Tunas, Cuba5 LAPROSAV. Carretera a Palmira Km 4, Cienfuegos, Cuba6 LAPROSAV. Ave Finlay Km 2½ e/ Planta de Nitrógeno y Circunvalación Norte, Camagüey, Cuba7 LAPROSAV. Calle Siguanea Km 2½, El Abra, Nueva Gerona, Isla de la Juventud, Cuba8 Estación de Pastos y Forrajes Niña Bonita, Carretera de Cangrejeras, Bauta, La Habana
RESUMEN
Mundialmente se señala la presencia de hongos, bacterias y virus enleguminosas y gramíneas utilizadas como pastos y forrajes, los quecausan efectos negativos en el crecimiento, en la calidad nutricionaly en su capacidad reproductiva. En el presente trabajo se analizaron,por diferentes técnicas de diagnóstico, muestras de estos cultivosprovenientes de la Estación de Pastos y Forrajes Niña Bonita y Labo-ratorios Provinciales de Sanidad Vegetal. Se registraron, por primeravez en Cuba, 23 especies de hongos, la bacteria Pseudomonas fluo-rescente del grupo 1b, el virus del mosaico de la soya (soybeanmosaic virus) y el virus del moteado amarillo del frijol (bean yellowmottle virus).
Palabras clave: fitopatógenos, pastos, forrajes
ABSTRACT
The presence of fungi, bacteria and virus attacking graminous andleguminous plants used as grass and forage, causing negatives effectson their growth, nutritional quality and reproductive capacity is a factknown all over the world. Samples o this kind of plants from NiñaBonita Grass and Forage Institute and Provincial Health PlantLaboratories were analyzed by different diagnosis techniques in thiswork. 23 fungal species, a fluorescent Pseudomonas group 1b, soybeanmosaic virus and bean yellow mottled virus Cuba are reported for thefirst time to.
Key words: phytopathogens, grass, forages
INTRODUCCIÓN
El establecimiento de los pastos y forrajes se afectapor la adaptación al clima, la resistencia al pisoteo delganado y la infección de hongos, bacterias y virus. Loshongos son los que poseen una mayor participación, nosolo por los daños que producen sobre los rendimien-tos, sino porque provocan alteraciones importantes enlos parámetros del producto cosechado, como lalignificación de los tallos, la disminución de ladigestibilidad de las paredes celulares [Abe y Okumura,1972] y del contenido de los aminoácidos libres en lashojas y raíces de las plantas afectadas [Hodges yRobinson, 1977], así como la producción de micotoxinas
durante el proceso de parasitismo [Delgado y Alonso,1994], mientras que los patógenos restantes dan lugara síntomas que afectan los rendimientos como enanis-mo y marchitez de las plantas.
En Cuba las enfermedades fúngicas reconocidas comoimportantes son las pertenecientes a los génerosPuccinia, Bipolaris y Piricularia, las cuales perjudicanel follaje de las gramíneas. En Panicum maximun Jacq.las principales patologías que afectan el follaje son cau-sadas por especies de Drechslera, y en las espiguillaspor el verdadero carbón causado por Conidiosporomycesayresii (Berk.), que produce índices de afectaciones

González y otros
12/fitosanidad
superiores al 40% [Bernal y Díaz, 1988; Delgado et al.,1990].
Respecto a las enfermedades virales, la literatura recogesolamente la presencia, en varios ecotipos de Centro-sema spp., de partículas virales semejantes a Potyvirus[Delgado y Machado, 1994], mientras que de las enfer-medades bacterianas están presentes las bacteriasXanthomonas axonopodis pv. sojensis en soya [Albornoz,1978], Erwinia sp. en Leucaena sp. [Delgado et al., 1989]Dickeya chrysanthemi [Samson et al., 2004] y Erwiniachrysanthemi en el sorgo [Caraballo, 1990].
Debido a los escasos estudios en las condiciones de Cubasobre las patologías que afectan a los pastos y forrajes y,al considerar que existen programas agrícolas para laproducción de semillas con bajos insumos de las especiesde gramíneas y leguminosas utilizadas como alimentospara el ganado; con el objetivo de incrementar la recupe-ración ganadera, se llevó a cabo la identificación defitopatógenos que las infectan, lo que constituye un co-nocimiento de gran utilidad para los interesados.
MATERIALES Y MÉTODOS
Para la determinación de hongos, bacterias y virus secolectaron muestras frescas y de semillas de especiespratenses y forrajeras del Instituto de Investigacionesde Pastos y Forrajes Niña Bonita, y las procedentes dediferentes estaciones analizadas en los laboratorios pro-vinciales de sanidad vegetal de las provincias de La Ha-bana, Cienfuegos, Camagüey, Las Tunas, Granma y elmunicipio especial de Isla de la Juventud.
Diagnóstico de hongos
Se analizaron porciones de las áreas afectadas de cadamuestra, las que se lavaron con abundante agua corrien-te durante 5 min, se desinfectaron en una solución dehipoclorito de sodio al 2% y se colocaron en placas quecontenían agar-agua. De las colonias desarrolladas enlas placas de agar-agua se transfirieron discos de 5 mma cuñas de papa dextrosa agar, incubados a 25°C conalternancia de luz-oscuridad y se observaron a partirde las 48-72 h, según se desarrollaron los hongos. Lasdeterminaciones específicas se realizaron al microsco-pio óptico y al estereoscopio según los criterios de Ellis(1971, 1976), Domsch et al. (1980), Nelson et al. (1983)y Mercado et al. (1997).
Diagnóstico de bacterias
El procedimiento se realizó mediante el método de ex-tracción por trituración del tejido por una hora en PBS
y siembra por agotamiento en medio de cultivo agarnutriente. Las placas se incubaron durante cuatro díasy posteriormente se realizó transferencia de las colo-nias a las que se les realizaron las pruebas de produc-ción de pigmentos en medio B de King [King et al.,1954], reacción de hipersensibilidad Klement (1963), me-tabolismo oxidativo fermentativo de la glucosa, tinciónde flagelos y pruebas de LOPAT [Lelliott et al., 1966],presencia de nitrato reductasa, protopectinasa, utili-zación de malonato, de tartrato de sodio y producciónde catalasa [Delgado et al., 1989].
Para el análisis de las semillas se aplicaron los métodosde cámara húmeda tradicional, del papel enrollado ydel triturado y remojado de estas durante 24 h a 4oC, yel método peso-agua del grifo [Van Bovenkamp, 1985].
Diagnóstico de virus
Se realizaron inoculaciones mecánicas a plantas sanasde cada especie de pasto y forraje, después de una se-mana de germinadas, mediante el método de frotacióndel macerado. Como inóculo se utilizaron hojas con lossíntomas de amarillamiento, reducción de la láminafoliar, mosaico y moteado. La maceración se realizó enmorteros sumergidos en hielo con tampón fosfato0,0025M y pH 7,8, que contenía mercaptoetanol a0,5% + carbón activado en una relación 3:1:1. Las plan-tas testigos solo se inocularon con tampón, y todas semantuvieron en condiciones aisladas.
En los casos en que fue necesario se realizó la transmi-sión mediante saltahojas, para lo cual se utilizaron aque-llos que procedieron de colonias establecidas en plan-tas de maíz y que fueron previamente identificadas comoPeregrinus maidis (Ashm) y Dalbulus maidis (De Longy Wolcott) [Rodríguez-León, comunicación personal].Las pruebas se realizaron en cubículos o en pequeñasjaulas donde se colocaron de 2-20 macetas con dos plan-tas sanas cada una, a la vez de contar con plantas in-fectadas de maíz que sirvieron de inóculos, sobre lasque se mantuvieron altas poblaciones de los insectos.
La microscopia óptica se realizó mediante las técnicasde tiras de epidermis [Christie y Edwardson, 1977], deDienes [June et al., 1979] y de abrasión [Ko et al., 1985]en un rango de 100-1 000X.
Las observaciones en microscopía electrónica se reali-zaron por el método de tinción negativa [Hitchorny yHills, 1965] y por la técnica de corte e inclusión. Lasmuestras se observaron en un rango de 20 000X. Latécnica serológica fue ELISA-DAS, y se utilizaron jue-

fitosanidad/13
Fitopatógenos en los cultivos...
gos de inmunosueros de la firma AGDIA, con las dilucio-nes de IgG y conjugados recomendados por el fabricante.
RESULTADOS Y DISCUSIÓN
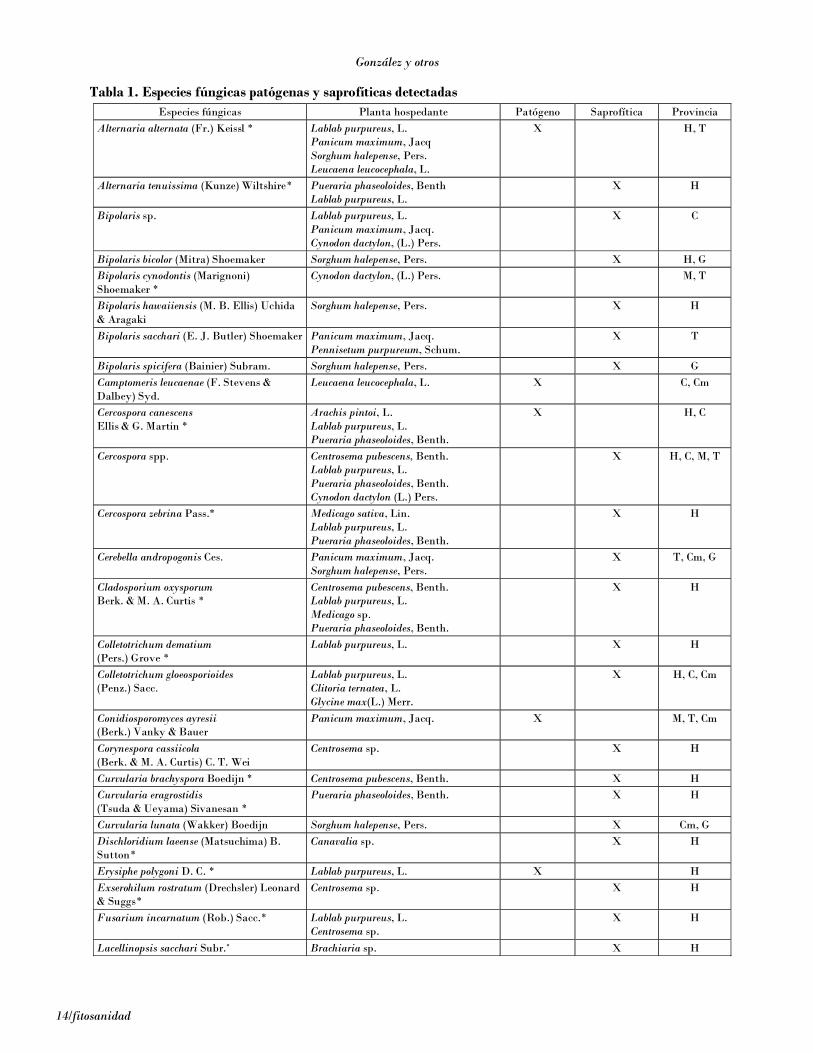
Hongos
De la totalidad de especies de hongos detectados, 23 deellas se informan por primera vez en Cuba en los cultivosde pastos y forrajes (Tabla 1), entre las se encuentranAlternaria alternata (Fr.) Keissl, cuyo aislamiento se rea-lizó a partir de pequeñas manchas pardo oscuras con unhalo fino amarillento, una especie cosmopolita que apa-rece en muchas clases de plantas y sustratos, incluyen-do suelo, alimentos y textiles [Ellis, 1971; Domsch et al.,1980]. Aunque este hongo es considerado de poca impor-tancia en los cultivos que afecta [Shew y Lucas, 1991],en las plantas en que se encontró provocó abundantessíntomas, además de estar muy bien distribuido. Porotra parte, existen registros de toxicidad en animales desangre caliente que han ingerido alimentos contamina-dos con este hongo [Domsch et al., 1980]. Bipolariscynodontis (Maringnoni) Shoemaker es cosmopolita ycausa manchas foliares en varias especies de gramíneas[Farr et al., 1995]. En el área del Caribe solo se habíaregistrado en Puerto Rico [Minter et al., 2001].
Cercospora canescens Ellis & G. Martín es muy frecuen-te sobre diferentes leguminosas en trópicos ysubtrópicos [Ellis, 1976]. En las muestras analizadasse observó en las hojas provocando manchas redondea-das pardas con el centro grisáceo, margen pardo-rojizooscuro y halo amarillo. Cercospora zebrina se ha regis-trado en plantas de Medicago, Trifolium y otras en elhemisferio norte, África y Sudamérica [Farr et al.,1995], pero no se han encontrado registros enCentroamérica. En el diagnóstico realizado se detectóen Medicago spp. al causar manchas pardo-grisáceasmuy oscuras, con el centro más claro, y en algunas unhalo amarillento no muy marcado. Colletotrichumdematium (Pers.) Grove es un saprófito común de teji-do vegetal en descomposición, pero también es causan-te de pudriciones en frutos, manchas foliares y damping-off en varias especies de plantas, especialmente enleguminosas [Domsch et al., 1980]. En este caso se de-tectó sobre las hojas con síntomas típicos deantracnosis, donde estaba presente también la faseteleomórfica Glomerella cingulata (Stonem) Spauldingand Schrenk. Fusarium semitetectum Berck & Rau esuna especie cosmopolita, pero su mayor importanciaeconómica radica como agente causal de pudriciones
poscosecha en varios cultivos tropicales, y puede cau-sar damping-off en posturas de tomate. En Cuba estehongo es frecuente sobre restos vegetales de numero-sas plantas, con acción saprofítica fundamentalmen-te [Gerlach y Nirenberg, 1982; López et al., 1993]. Al-gunas cepas de esta especie son toxicogénicas y puedenproducir beauvericina, equisetina, fusapirona yzearalenona [Thrane, 1999]. La zearalenona es un com-puesto similar al 17ß-estradiol, y su consumo puedeproducir problemas estrogénicos en el procesoreproductivo. Se ha comprobado la virulencia de estecompuesto, especialmente en cerdos, donde se ha vis-to que la presencia de esta toxina en alimentos causahiperestrogenismo, vulvovaginitis en cerdas, abortosespontáneos, agalactia e infertilidad, debido a sus mar-cadas propiedades estrogénicas y anabólicas.
Entre las otras especies detectadas y señaladas conanterioridad en Cuba se encuentra B. bicolor, que cau-só pudrición en la base del tallo y lesiones foliares enS. halepense y S. verticilliflorum, Bipolaris hawaiiensis,detectados en diferentes especies de plantas, así comoen suelo, textiles y otros sustratos, y B. sacchari, quees el agente causal de la enfermedad mancha de ojo enhojas de la caña de azúcar; pero en condiciones de ex-trema humedad también puede producir marchitamien-to en posturas de esta planta [Ellis, 1971].
Bacterias
Se detectaron siete especies de bacterias en los pastos yforrajes analizados (Tabla 2). De estas se registran porprimera vez en Cuba cepas oxidasa, levan, proto-pectinasa y arginina negativas y positivas frente a lareacción de hipersensibilidad en tabaco, correspondientea Pseudomonas fluorescentes del grupo 1b en G. max, apartir de hojas afectadas con manchas grasientas, loque coincide con Lelliot et al. (1966).
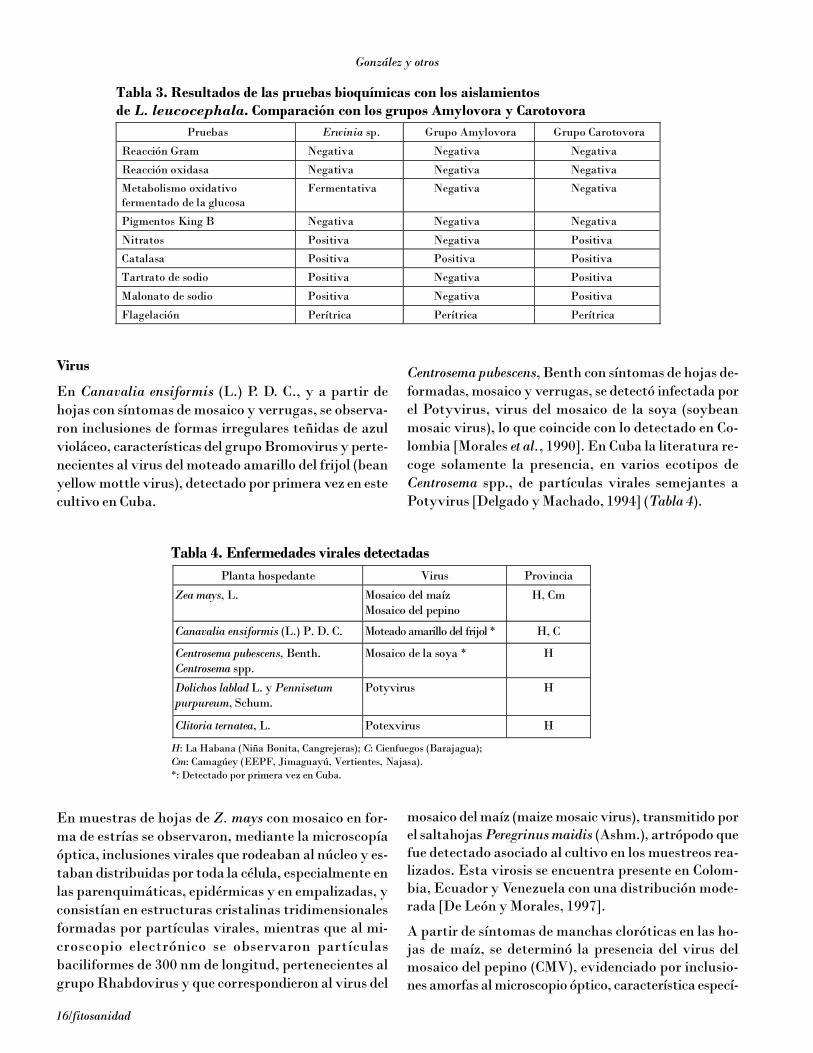
Por otra parte, se observaron cepas no fluorescentes apartir de las muestras tomadas de L. leucocephala, lasque según las pruebas llevadas a cabo correspondían algénero Erwinia. En la Tabla 3 se observan los resulta-dos de las pruebas bioquímicas para este género, y secompara con los grupos Amylovora y Carotovora, loque demuestra que L. leucocephala está infectada poruna bacteria que tiene características de ambos gru-pos. Hasta el momento, en Cuba estaba definida paraeste cultivo la presencia de una bacteria del géneroErwinia, asociada a hongos del género Fusarium [Del-gado et al., 1989].
González y otros
14/fitosanidad
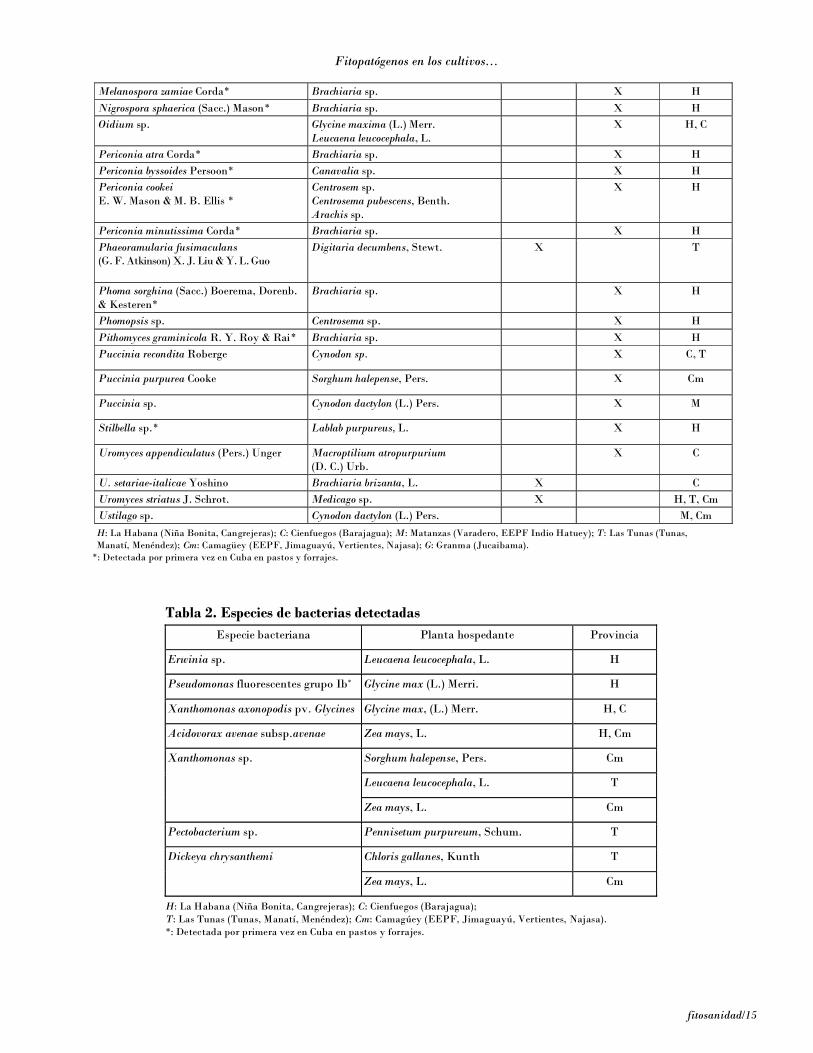
Tabla 1. Especies fúngicas patógenas y saprofíticas detectadas
Especies fúngicas Planta hospedante Patógeno Saprofítica Provincia Alternaria alternata (Fr.) Keissl * Lablab purpureus, L.
Panicum maximum, Jacq Sorghum halepense, Pers. Leucaena leucocephala, L.
X
H, T
Alternaria tenuissima (Kunze) Wiltshire* Pueraria phaseoloides, Benth Lablab purpureus, L.
X H
Bipolaris sp. Lablab purpureus, L. Panicum maximum, Jacq. Cynodon dactylon, (L.) Pers.
X C
Bipolaris bicolor (Mitra) Shoemaker Sorghum halepense, Pers. X H, G Bipolaris cynodontis (Marignoni) Shoemaker *
Cynodon dactylon, (L.) Pers. M, T
Bipolaris hawaiiensis (M. B. Ellis) Uchida & Aragaki
Sorghum halepense, Pers. X H
Bipolaris sacchari (E. J. Butler) Shoemaker Panicum maximum, Jacq. Pennisetum purpureum, Schum.
X T
Bipolaris spicifera (Bainier) Subram. Sorghum halepense, Pers. X G Camptomeris leucaenae (F. Stevens & Dalbey) Syd.
Leucaena leucocephala, L. X C, Cm
Cercospora canescens Ellis & G. Martin *
Arachis pintoi, L. Lablab purpureus, L. Pueraria phaseoloides, Benth.
X
H, C
Cercospora spp. Centrosema pubescens, Benth. Lablab purpureus, L. Pueraria phaseoloides, Benth. Cynodon dactylon (L.) Pers.
X H, C, M, T
Cercospora zebrina Pass.* Medicago sativa, Lin. Lablab purpureus, L. Pueraria phaseoloides, Benth.
X H
Cerebella andropogonis Ces. Panicum maximum, Jacq. Sorghum halepense, Pers.
X T, Cm, G
Cladosporium oxysporum Berk. & M. A. Curtis *
Centrosema pubescens, Benth. Lablab purpureus, L. Medicago sp. Pueraria phaseoloides, Benth.
X H
Colletotrichum dematium (Pers.) Grove *
Lablab purpureus, L.
X H
Colletotrichum gloeosporioides (Penz.) Sacc.
Lablab purpureus, L. Clitoria ternatea, L. Glycine max(L.) Merr.
X H, C, Cm
Conidiosporomyces ayresii (Berk.) Vanky & Bauer
Panicum maximum, Jacq. X M, T, Cm
Corynespora cassiicola (Berk. & M. A. Curtis) C. T. Wei
Centrosema sp. X H
Curvularia brachyspora Boedijn * Centrosema pubescens, Benth. X H Curvularia eragrostidis (Tsuda & Ueyama) Sivanesan *
Pueraria phaseoloides, Benth. X H
Curvularia lunata (Wakker) Boedijn Sorghum halepense, Pers. X Cm, G Dischloridium laeense (Matsuchima) B. Sutton*
Canavalia sp. X H
Erysiphe polygoni D. C. * Lablab purpureus, L. X H Exserohilum rostratum (Drechsler) Leonard & Suggs*
Centrosema sp. X H
Fusarium incarnatum (Rob.) Sacc.* Lablab purpureus, L. Centrosema sp.
X H
Lacellinopsis sacchari Subr.* Brachiaria sp. X H
fitosanidad/15
Fitopatógenos en los cultivos...
Melanospora zamiae Corda* Brachiaria sp. X H Nigrospora sphaerica (Sacc.) Mason* Brachiaria sp. X H Oidium sp. Glycine maxima (L.) Merr.
Leucaena leucocephala, L. X H, C
Periconia atra Corda* Brachiaria sp. X H Periconia byssoides Persoon* Canavalia sp. X H Periconia cookei E. W. Mason & M. B. Ellis *
Centrosem sp. Centrosema pubescens, Benth. Arachis sp.
X H
Periconia minutissima Corda* Brachiaria sp. X H Phaeoramularia fusimaculans (G. F. Atkinson) X. J. Liu & Y. L. Guo
Digitaria decumbens, Stewt. X
T
Phoma sorghina (Sacc.) Boerema, Dorenb. & Kesteren*
Brachiaria sp. X H
Phomopsis sp. Centrosema sp. X H Pithomyces graminicola R. Y. Roy & Rai* Brachiaria sp. X H Puccinia recondita Roberge Cynodon sp. X C, T
Puccinia purpurea Cooke Sorghum halepense, Pers. X Cm
Puccinia sp. Cynodon dactylon (L.) Pers. X M
Stilbella sp.* Lablab purpureus, L. X H
Uromyces appendiculatus (Pers.) Unger Macroptilium atropurpurium (D. C.) Urb.
X C
U. setariae-italicae Yoshino Brachiaria brizanta, L. X C Uromyces striatus J. Schrot. Medicago sp. X H, T, Cm Ustilago sp. Cynodon dactylon (L.) Pers. M, Cm
H: La Habana (Niña Bonita, Cangrejeras); C: Cienfuegos (Barajagua); M: Matanzas (Varadero, EEPF Indio Hatuey); T: Las Tunas (Tunas, Manatí, Menéndez); Cm: Camagüey (EEPF, Jimaguayú, Vertientes, Najasa); G: Granma (Jucaibama).
*: Detectada por primera vez en Cuba en pastos y forrajes.
Tabla 2. Especies de bacterias detectadas
Especie bacteriana Planta hospedante Provincia
Erwinia sp. Leucaena leucocephala, L. H
Pseudomonas fluorescentes grupo Ib* Glycine max (L.) Merri. H
Xanthomonas axonopodis pv. Glycines Glycine max, (L.) Merr. H, C
Acidovorax avenae subsp.avenae Zea mays, L. H, Cm
Sorghum halepense, Pers. Cm
Leucaena leucocephala, L. T
Xanthomonas sp.
Zea mays, L. Cm
Pectobacterium sp. Pennisetum purpureum, Schum. T
Chloris gallanes, Kunth T Dickeya chrysanthemi
Zea mays, L. Cm
H: La Habana (Niña Bonita, Cangrejeras); C: Cienfuegos (Barajagua); T: Las Tunas (Tunas, Manatí, Menéndez); Cm: Camagúey (EEPF, Jimaguayú, Vertientes, Najasa). *: Detectada por primera vez en Cuba en pastos y forrajes.
González y otros
16/fitosanidad
Virus
En Canavalia ensiformis (L.) P. D. C., y a partir dehojas con síntomas de mosaico y verrugas, se observa-ron inclusiones de formas irregulares teñidas de azulvioláceo, características del grupo Bromovirus y perte-necientes al virus del moteado amarillo del frijol (beanyellow mottle virus), detectado por primera vez en estecultivo en Cuba.
Tabla 3. Resultados de las pruebas bioquímicas con los aislamientos de L. leucocephala. Comparación con los grupos Amylovora y Carotovora
Pruebas Erwinia sp. Grupo Amylovora Grupo Carotovora
Reacción Gram Negativa Negativa Negativa
Reacción oxidasa Negativa Negativa Negativa
Metabolismo oxidativo fermentado de la glucosa
Fermentativa Negativa Negativa
Pigmentos King B Negativa Negativa Negativa
Nitratos Positiva Negativa Positiva
Catalasa Positiva Positiva Positiva
Tartrato de sodio Positiva Negativa Positiva
Malonato de sodio Positiva Negativa Positiva
Flagelación Perítrica Perítrica Perítrica
Centrosema pubescens, Benth con síntomas de hojas de-formadas, mosaico y verrugas, se detectó infectada porel Potyvirus, virus del mosaico de la soya (soybeanmosaic virus), lo que coincide con lo detectado en Co-lombia [Morales et al., 1990]. En Cuba la literatura re-coge solamente la presencia, en varios ecotipos deCentrosema spp., de partículas virales semejantes aPotyvirus [Delgado y Machado, 1994] (Tabla 4).
Tabla 4. Enfermedades virales detectadas
Planta hospedante Virus Provincia
Zea mays, L. Mosaico del maíz Mosaico del pepino
H, Cm
Canavalia ensiformis (L.) P. D. C. Moteado amarillo del frijol * H, C
Centrosema pubescens, Benth. Centrosema spp.
Mosaico de la soya * H
Dolichos lablad L. y Pennisetum purpureum, Schum.
Potyvirus H
Clitoria ternatea, L. Potexvirus H
H: La Habana (Niña Bonita, Cangrejeras); C: Cienfuegos (Barajagua); Cm: Camagúey (EEPF, Jimaguayú, Vertientes, Najasa). *: Detectado por primera vez en Cuba.
En muestras de hojas de Z. mays con mosaico en for-ma de estrías se observaron, mediante la microscopíaóptica, inclusiones virales que rodeaban al núcleo y es-taban distribuidas por toda la célula, especialmente enlas parenquimáticas, epidérmicas y en empalizadas, yconsistían en estructuras cristalinas tridimensionalesformadas por partículas virales, mientras que al mi-croscopio electrónico se observaron partículasbaciliformes de 300 nm de longitud, pertenecientes algrupo Rhabdovirus y que correspondieron al virus del
mosaico del maíz (maize mosaic virus), transmitido porel saltahojas Peregrinus maidis (Ashm.), artrópodo quefue detectado asociado al cultivo en los muestreos rea-lizados. Esta virosis se encuentra presente en Colom-bia, Ecuador y Venezuela con una distribución mode-rada [De León y Morales, 1997].
A partir de síntomas de manchas cloróticas en las ho-jas de maíz, se determinó la presencia del virus delmosaico del pepino (CMV), evidenciado por inclusio-nes amorfas al microscopio óptico, característica especí-

fitosanidad/17
Fitopatógenos en los cultivos...
fica de este grupo, y es el único representante que afec-ta al maíz. Su presencia en Cuba es escasa, lo que coin-cide con De León y Morales (1997) quienes lo detecta-ron en algunas zonas de Brasil en una incidencia muybaja.
En D. labblab, P. purpureum y C. ternatea se observaroninclusiones citoplasmáticas pertenecientes a los gruposPotyvirus y Potexvirus.
CONCLUSIONES
• Las especies de hongos A. alternata, A. tenuissima,B. cynodontis, C. canescens, C. zebrina, C. dematium,C. brachyspora, C. eragrostidis, D. laense, E. polygoni,E. rostratum, F. incarnatum, P. byssoides, P. cookei,Stilbella sp., L. sacchari, M. zamiae, N. sphaerica,P. atra, P. minutissima, P. sorghina y P. graminicolason nuevos registros para el cultivo de los pastos enCuba.
• Entre los hongos patógenos los géneros Cercospora yUromyces fueron los más representados (tres espe-cies), seguidos por Alternaria y Colletotrichum. Sedestacó la presencia de dos especies de F. incarnatumque, aunque tiene poca importancia como fito-pátogeno, es un oportunista con gran capacidad depudrición y que además puede resultar toxigénico.
• Se determinó que L. leucocephala está infectada poruna bacteria no fluorescente del género Erwinia, cuyaespecie tiene características de E. amylovora y E. caro-tovor, y se determinó la presencia de X. axonopodispv. glycines en plantas de G. máxima y de A. avenaesubp. avenae en semillas de maíz (Z. mays, L.).
• En G. max se observaron cepas fluorescentes, corres-pondientes a Pseudomonas fluorescentes del grupoIb, detectadas por primera vez en Cuba.
• En C. ensiformis se detectó el virus del moteado ama-rillo del frijol, y en C. pubescens el virus del mosaicode la soya, no antes detectados en Cuba. En plantasde maíz se determinaron los virus del mosaico delmaíz y del mosaico del pepino, mientras que P. purpureumy C. ternatea se encontraron infectadas por unPotyvirus y un Potexvirus respectivamente.
REFERENCIAS
Abe, A.; T. Okumura: «Influence of Aphid Infestation on the ChemicalComposition and Nutritive Value of Lucerne», Bull. Nat. Inst. Anim,Husb. 25:19, 1972.
Albornoz, Alicia: «La pústula bacteriana de la soya en Cuba»,Agrotecnia de Cuba. 10 (2): 65-69, 1978.
Bernal, Blanca; J. A, Díaz. «Incidencia y distribución de las principalesenfermedades fungosas de pastos y forrajes en dos estaciones deLa Habana», Ciencia y Técnica de la Agricultura. Protección dePlantas 11 (1): 99-111, 1988.
Caraballo, E.: «Sorghum sp., nuevo hospedero de Erwinia crysanthemien la provincia de Holguín», Lista de bacterias fitopatógenas, Labo-ratorio Provincial de Sanidad Vegetal de Holguín, abril, 1990.
Christie, R. G.; J. R. Edwardson: «Light and Electron Microscopy ofPlant Vrus Inclusion», Fla. Agr.Exp. Stn. Monogr. Ser. 9, 1977.
De León, C.; F. Morales: «Determinación y efecto de enfermedadesvirosas del maíz en América del Sur», Reunión del Programa Regio-nal de Maiz, marzo, 1997.
Delgado, A.; N. Martínez; B. Rodríguez: «Estudio de la gomosis bacterianaen legumbres de Leucaena leucocephala (Lam) de Wit.», Pastos yForrajes 12:127-133, 1989.
Delgado, A.; H. Machado; G. de la Paz: «Evaluación de la resistencia ahongos de las espículas en una colección introducida de Panicummaximun Jacq.», Pastos y Forrajes 13:59, 1990.
Delgado, A.; R. Machado: «Comportamiento de 11 ecotipos deCentrosema ante un mosaico amarillo y confirmación práctica delagente causal», Pastos y Forrajes 17:55-62, 1994.
Delgado, A.; O. Alonso: «Las enfermedades fungosas en los pastostropicales», Pastos y Forrajes 17:89-93, 1994.
Domsch, K. H.; W. Gams; T. H. Anderson: Compendium of Soil Fungi,Acad. Press, vol. 1, 1980.
Ellis, M. B.: «Dematlaceous Hyphomycetes», CMI, Kew, Surrey, 1971.
––––: «More Dematlaceous Hyphomycetes», CMI, Kew, Surrey, 1976.
Farr, D. F.; F. B. Gerald; G. P. Chamuris; A. Y. Rossman: Fungi on Plantsand Plant Products in the Unites States, The AmericanPhytopathological Society, St. Paul., Minnesota, E.U., 1995, pp. 413-415.
Gerlach, W.; H. Nirenberg: The genus Fusarium. A Pictorial Atlas,Paul Parey, Berlin und Hamburg, 1982.
Hitchourny, J. H.; G. B. Hills: «The Use of Negative Staining in the ElectronMicroscopy Examination of Plant Viruses in Crude Extraits» ,Virology27:526-540, 1965.
Hodges, C. F.; P. W, Robinson: «Sugar and Amino Acid Content of Poapratensis Infected with Ustilago seriformis and Urocystis agropyri»,Phyiologia Plantarum 41:25, 1977.
June, D.; A. Stenens; R. T. Fox: «Use of Dienesstain to Detect PlantDisease Induced by Mycoplasma Like Organism», Phytopathology69 (11):1169-1171, 1979.
King, R. O.; M. K. Ward; D. E. Rany: «Two Simple Media for theDemostration of Pyocianis and Fluorescein», J. Lab. Clin. Med.44:301-304, 1954.
Klement, Z.: «Rapid Detection of the Pathogenecity of PhytopathogenicPseudomonas», Nature 199:300, 1963.
Ko, N. J.; F. W. Zettler; J. R. Edwardson; R. G. Christie: «LightMicroscopie Techniques for Detecting Orchid Viruses», Acta Hortic.116:241-243, 1985.
Lelliot, R. A.; E. Billing; A. Hayward: «A Determination Scheme for theFluorescent Plant Pathogenic Psedomonas», J. Apl. Bact. (3):470-489, 1966.
López, M. Ofelia; Ángela Estrada; América Milla; Iliana Sandoval: «Es-pecies de Fusarium frecuentes en plantas cultivadas en Cuba», IVSimposio de Botánica, La Habana, 22-26 de junio, 1993, p. 60.
Mercado, A. V.; Holubová-Jechová; J. Mena: Hifomicetes demacláceosde Cuba. Enteroblásticos. Monografia XXIII, Museo Regionale deScience Naturali, Torino, 1997.

González y otros
18/fitosanidad
Minter, D.; J. Mena; M. Rodríguez: Fungi of the Caribbean An annotatedchecklist. PDMS Publishing, Inglaterra, 2001.
Morales, F. J.; A. I. Niessen; M. D. Castaño: «Detection of A Strain ofSoybean Mosaic Virus Affecting Tropical Forage Species ofCentrosema», Plan Dis. 74:648-654, 1990.
Nelson, P. E. ; T. A. Toussoun; W. F. Marasas: Fusarium species. AnIllustrated Manual for Identification, The Pennsylvania UniversityPress, 1983.
Samson, R.; J. B. Legendre; R. Christen ; W. Achouak ; L. Gardan :«Transfer of Pectobacterium chrysanthemi [Brenner et al., 1973;Hauben et al., 1998 ] and Brenneria paradisiaca to the genus Dickeya
Gen. Nov. As Dickeya chrysanthemi Comb. Nov. and Dickeyaparadisiaca Comb. Nov and Delineation of four Novel Species:Dickeya dadantii sp. Nov., Dickeya dianthicola sp.Nov., Dickeyadieffenbachiae sp. Nov. and Dickeya zeae sp. Nov.», InternationalJournal of Systematic and Evolutionary Microbiology 54:1-13, 2004.
Shew, H. D.; G. B. Lucas: Compendium of Tobacco Diseases, AmericanPhytopatological Society, St. Paul, Minn., 1991.
Thrane, U.: Fusarium, Encyclopedia of Microbiology, Academic Press,Londres, 1999, pp. 901-906.
Van Bovenkamp, J.: «Detection of Seed-Borne Bacteria», WorkshopISTA, Versailles, 1-7, 1985.

fitosanidad/19
FITOSANIDAD vol. 10, no. 1, marzo 2006
CAUSAS DE LA APARICIÓN DE LA ENFERMEDAD MANCHAVERDE EN HOJAS DE TABACO (NICOTIANA TABACUM L.)
Danneys Armario Aragón,1 Sinesio Torres García,2 Oguelis Rodríguez Pérez,3 Iban ArredondoQuevedo,1 Yunetsy Girado López1 y Ángel Mollineda Trujillo2
1 Instituto de Investigaciones en Viandas Tropicales (INIVIT). Apdo. 6, Santo Domingo, Villa Clara, Cuba, CP 53 000, [email protected] Facultad de Ciencias Agropecuarias (UCLV). Carretera a Camajuaní, Km 5½, Santa Clara, Villa Clara, Cuba, [email protected] División Centro Oeste de Tabaco. Villa Clara, Cuba, [email protected]
RESUMEN
La enfermedad de la mancha verde del tabaco (Nicotiana tabacum L.),de origen desconocido, es responsable de considerables pérdidas enlas hojas para el tabaco torcido y para las capas que se comercializan;por lo tanto, el objetivo de este trabajo es mostrar que la aparición dela enfermedad de la mancha verde está relacionada con los fungicidascompuestos por manganeso (Mn) y zinc (Zn), como en el caso delmancozeb y con las características morfológicas de cada variedad. Elmayor número de hojas manchadas apareció en los niveles medio einferior de las plantas tratadas y no se observaron manchas en las notratadas. Las hojas hervidas de plantas sin aplicación tomaron uncolor oscuro típico de la feofitina, mientras que las hojas tratadaspresentaron tonalidades verdes. Las manchas aparecieron en las par-tes de las hojas donde la concentración de Mn era la misma o mayorque el magnesio (Mg). La enfermedad fue evidente solo en las plantastratadas, lo que demuestra que la causa de la mancha verde del tabacoestá dada por la acumulación de Mn en las regiones del limbo; o sea,tiene lugar un exceso de Mn por encima de las concentraciones míni-mas permisibles y una sustitución del Mg por el Mn en las moléculasde la clorofila. La variedad Habana 2000 resultó ser la más suscepti-ble y la de más bajo índice de esclerofilia, mientras Criollo 98 fue lamás tolerante y de mayor índice de esclerofilia, características que serelacionan directamente con la incidencia de la mancha. El estratofoliar superior fue el menos afectado
Palabras clave: mancha verde, tabaco, manganeso, Mancozeb, clorofila
ABSTRACT
Tobacco green spot disease, of unknown origin, is responsible ofconsiderable losses in leaves for twisted tobacco and layers for mar-keting, in that way it was realized a research work to show thatappearance of green spot disease is related with fungicides composedby Mn and Zn, like mancozeb, and to morphologic characteristics oftobacco varieties too. The highest number of spotted leaves appearedin middle and low levels of treated plants, and there were no spot innon-treated plants. Non-treated boiled leaves got the typical browncolor of feofitine, while treated leave presented green tonalities. Spotsappear in leaf parts where Mn concentration was the same or biggerthan Mg concentration. The disease was evident only in treated plants,showing that the cause of tobacco green spot is due to Mn accumulationin leaf limb regions, that is an excess of Mn above minimumconcentrations takes place, as well as, a replacement by Mg in thechlorophyll molecules. ´Havana 2000‘ variety was the most suscepti-ble and showed the lowest sclerophyllous rate, while, Criollo-98 varietywas more tolerant and presented the highest sclerophyllous rate whichare characteristics closely related with the spot incidence. The supe-rior leaf stratum was the least affected one.
Key words: green spot, tobacco, manganese, Mancozeb, chlorophyll
INTRODUCCIÓN
Entre los mecanismos que explican la declinación dela fotosíntesis en las plantas están el daño peroxidativode las funciones de la membrana del tilacoide –que re-percute sobre la actividad fotoquímica de losfotosistemas I y II de los cloroplastos–, el decrecimien-to de la actividad de las enzimas fotosintéticas por losproductos de la polifenoloxidación y la disminución enla actividad de carboxilación de la ribulosa 1-5 bifosfatocarboxilasa/oxigenasa (rubisco) [Kitao et al., 1997a;1998], observados en hojas de plantas de trigo y taba-co, con la aplicación de alta concentración de Mn.
Según Jordan y Ogren (1981), la sustitución del Mg porel Mn puede ocurrir, dado el cambio de las propiedadesde la enzima rubisco, donde es posible con adicción deMn ocasionar el remplazo del Mg de tal enzima por Mn yformar la rubisco Mn, con lo cual tiene una mayor pro-porción de actividad oxigenasa que carboxilasa. Por suparte, Pequeño (1964) reportó que en condiciones artifi-ciales, al añadir acetato de cobre (Cu) y zinc (Zn) a hojasfotosintéticamente activas y mediante calentamiento, seorigina una modificación en la estructura de la clorofila,los protones de hidrógeno pueden sustituir temporal-

Armario y otros
20/fitosanidad
mente al Mg de la clorofila (formación de fofitina),solo que el sitio que le correspondía al Mg es ocupadopor el Cu o el Zn y toma un color verde azulado. Ade-más, se ha observado que la tasa fotosintética de ho-jas jóvenes de tabaco Burley disminuyó hasta 60%en comparación con el control, con altos tratamientosde Mn cuando este aumentó a 5 mg.g–1 de peso seco(PS) [Nable et al., 1988].
El objetivo de este trabajo fue demostrar que la apari-ción de la mancha verde se relaciona con los fungicidascompuestos por Mn y Zn, componentes del fungicidamancozeb 80% PH, y que las características mor-fológicas inherentes a cada variedad inciden en su sus-ceptibilidad.
MATERIALES Y MÉTODOS
La investigación se llevó a cabo en los laboratorios dela Universidad Central de Villa Clara, con plantas pro-cedentes de las CCS El Vaquerito, en Santa Clara, deenero a junio del 2002. Se realizaron dos experimentossobre suelo pardo con carbonatos con diseños de blo-ques al azar y cuatro réplicas, uno para evaluar el efec-to del fungicida a base de Mn y Zn, y otro para valorarla sensibilidad varietal.
1. Estudio del efecto de la aplicación del fungicidamancozeb 80% PH en la aparición de la mancha verde
Se evaluó un tratamiento con aplicación de fungicidamancozeb 80% PH a la dosis de 3 kg.ha–1 [MINAGRI,1999] y otro sin aplicación de fungicida (testigo abso-luto) sobre la variedad de tabaco Habana 2000. Se de-terminó el número de hojas con manchas verdes (NHM)por estrato foliar, clasificadas con respecto al grado deafectación según una escala de patrón de colores elabo-rada al efecto, con la siguiente gradología:
0: No presencia de mancha.1: Manchado ligero.2: Manchado medio.3: Manchado fuerte.
De las hojas se extrajeron las áreas manchadas con losdiferentes grados y se determinó la concentración demicroelementos (Mg, Mn, Zn y Cu), para lo cual se uti-lizó un espectrofotómetro de absorción atómica marcaSP-9 Pye Unican.
Hojas verdes de ambos tratamientos se cocieron en aguadestilada a temperaturas entre 80 y 100°C para favore-cer la ruptura de las membranas y facilitar la penetra-
ción del protón H+, proveniente de la ionización de losácidos orgánicos de la vacuola, al cloroplasto. Se deter-minó el pH del jugo vacuolar en la disolución por mediode un conductímetro-pH metro INOLAB, Level 1, defabricación alemana.
2. Estudio del comportamiento varietal con aplicacióndel fungicida mancozeb 80% PH a 3 kg.ha–1
Se utilizaron las variedades Habana 2000, Criollo 98 yCorojo 99. Las plantas por evaluar se subdividieron entres niveles foliares de acuerdo con la selección de pro-ducción: inferior (libre pie y uno medio), medio (pri-mer y segundo centro ligeros) y superior (primer y se-gundo centro fino), y en ellos se determinó el númerode hojas con manchas verdes por nivel. En total se eva-luaron 36 hojas por planta y 12 hojas por nivel.
El área foliar (AL) se determinó por la fórmula:
AL = ∑ (l x a) x f
donde:
l: Longitud de la hojaa: Ancho de la hojaf: Coeficiente de área foliar para el tabaco (f = 0,70)
El índice de esclerofilia (IE) o masa seca (MS) por uni-dad de área del limbo (AL) se determinó por variedady estratos a través del método del coeficiente de áreafoliar, y se calculó mediante la ecuación:
IE = PS/AL
donde:
PS: Peso seco del limbo (g)AL: Unidad de área del limbo (dm2)
RESULTADOS Y DISCUSIÓN
1. Estudio del efecto de la aplicación del fungicidamancozeb 80% PH en la aparición de la mancha verde
Con aplicaciones del fungicida cada siete días se pre-sentaron las mayores incidencias sobre los estratosmedios e inferior (Tabla 1). Donde no se aplicó el pro-ducto no hubo aparición de la enfermedad en ningunode los estratos. De acuerdo con Kitao et al. (1997b), lacausa de la mancha verde está asociada a la capacidadde los iones de Mn y/o Zn para sustituir el Mg de lamolécula de clorofila a nivel celular. Así forma una clo-rofila que no puede ser degradada por la acción de laclorofilasa y no ser convertida en feofitina por la acti-vidad de los protones H+ del jugo celular (al menos al
fitosanidad/21
Causas de la aparición de la enfermedad...
pH presente en el tejido celular del tabaco). Kitao et al.(1997b) plantean el efecto peroxidativo sobre las membranas
de los cloroplastos y todo el sistema membranoso, con lo cualhace a las células más permeables incluso a este mismo ión.
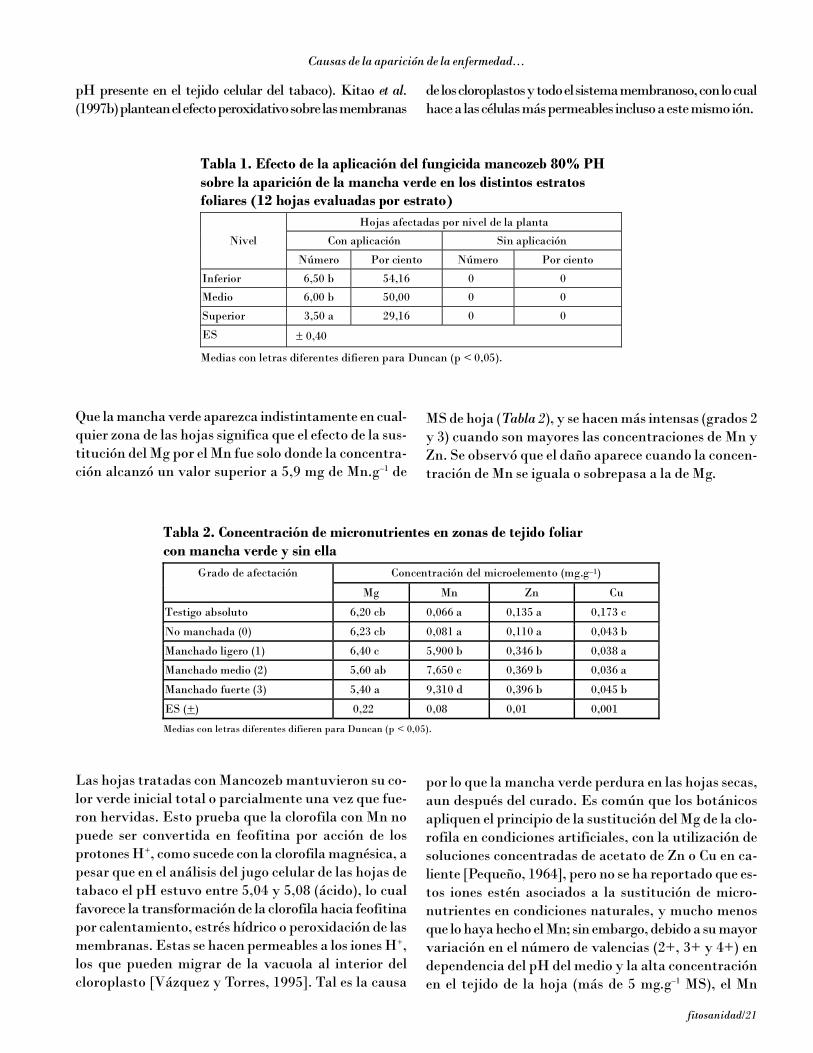
Que la mancha verde aparezca indistintamente en cual-quier zona de las hojas significa que el efecto de la sus-titución del Mg por el Mn fue solo donde la concentra-ción alcanzó un valor superior a 5,9 mg de Mn.g–1 de
MS de hoja (Tabla 2), y se hacen más intensas (grados 2y 3) cuando son mayores las concentraciones de Mn yZn. Se observó que el daño aparece cuando la concen-tración de Mn se iguala o sobrepasa a la de Mg.
Tabla 2. Concentración de micronutrientes en zonas de tejido foliar con mancha verde y sin ella
Concentración del microelemento (mg.g–1) Grado de afectación
Mg Mn Zn Cu
Testigo absoluto 6,20 cb 0,066 a 0,135 a 0,173 c
No manchada (0) 6,23 cb 0,081 a 0,110 a 0,043 b
Manchado ligero (1) 6,40 c 5,900 b 0,346 b 0,038 a
Manchado medio (2) 5,60 ab 7,650 c 0,369 b 0,036 a
Manchado fuerte (3) 5,40 a 9,310 d 0,396 b 0,045 b
ES (+) 0,22 0,08 0,01 0,001
Medias con letras diferentes difieren para Duncan (p < 0,05).
Las hojas tratadas con Mancozeb mantuvieron su co-lor verde inicial total o parcialmente una vez que fue-ron hervidas. Esto prueba que la clorofila con Mn nopuede ser convertida en feofitina por acción de losprotones H+, como sucede con la clorofila magnésica, apesar que en el análisis del jugo celular de las hojas detabaco el pH estuvo entre 5,04 y 5,08 (ácido), lo cualfavorece la transformación de la clorofila hacia feofitinapor calentamiento, estrés hídrico o peroxidación de lasmembranas. Estas se hacen permeables a los iones H+,los que pueden migrar de la vacuola al interior delcloroplasto [Vázquez y Torres, 1995]. Tal es la causa
por lo que la mancha verde perdura en las hojas secas,aun después del curado. Es común que los botánicosapliquen el principio de la sustitución del Mg de la clo-rofila en condiciones artificiales, con la utilización desoluciones concentradas de acetato de Zn o Cu en ca-liente [Pequeño, 1964], pero no se ha reportado que es-tos iones estén asociados a la sustitución de micro-nutrientes en condiciones naturales, y mucho menosque lo haya hecho el Mn; sin embargo, debido a su mayorvariación en el número de valencias (2+, 3+ y 4+) endependencia del pH del medio y la alta concentraciónen el tejido de la hoja (más de 5 mg.g–1 MS), el Mn
Tabla 1. Efecto de la aplicación del fungicida mancozeb 80% PH sobre la aparición de la mancha verde en los distintos estratos foliares (12 hojas evaluadas por estrato)
Hojas afectadas por nivel de la planta
Con aplicación Sin aplicación Nivel
Número Por ciento Número Por ciento
Inferior 6,50 b 54,16 0 0
Medio 6,00 b 50,00 0 0
Superior 3,50 a 29,16 0 0
ES ± 0,40
Medias con letras diferentes difieren para Duncan (p < 0,05).

Armario y otros
22/fitosanidad
perfectamente puede sustituir al Mg en la clorofila encondiciones naturales.
2. Estudio del comportamiento varietal con aplicacióndel fungicida mancozeb 80% PH
El comportamiento de las variedades con la aplicaciónde una dosis de 3 k.ha–1 fue diferente. La menor afecta-ción se observó en la variedad Criollo 98, seguida deCorojo 99, ambas procedentes del cruzamiento entrelas variedades Habana PR y Habana 92, cuyo origen esel cruce entre Corojo con RxT de Polonia [Espino et al.,2000]; sin embargo, la Habana 2000 fue la más afecta-da (Tabla 3). Esta variedad procede del cruzamientoentre Corojo y Habana 2.1.1, y esta última el resul-tado del cruce de Corojo con la variedad norteameri-cana Bell 61-9 [MINAGRI, 2001]. Es posible queentre las características morfológicas heredadas porla Habana 2000 se encuentren la presencia de hojasde láminas foliares más onduladas y finas, con for-mación de fosos, y conjuntamente paredes celularesy cutículas más delgadas respecto al resto, factoresque facilitan la permeabilidad a agentes externos(iones) acumulados en el limbo, junto con los pesti-cidas en altas concentraciones, durante un prolon-gado tiempo, y que a su vez esto no suceda de igualforma en otras variedades.
Las afectaciones en cada variedad se distribuyeron dediferente forma. Criollo 98 mostró mayor número dehojas manchadas en el estrato inferior y menos en elsuperior, el cual resultó estadísticamente similar a laporción media de la planta. Por su parte, en Corojo 99el daño se produjo en igual proporción en los estratosinferior y medio, y muy bajo en el superior. En Haba-na 2000 esta afectación se apreció en mayor cuantía enel estrato medio –valor que correspondió al máximo deafectación de todas las variedades– seguido del infe-rior, y también menos afectadas las hojas de la porciónsuperior, al igual que en las dos anteriores (Tabla 4). Enestos valores podría estar influyendo la arquitectura
de la planta de tabaco, lo que pudiera ser motivo deotra investigación posterior.
Índice de esclerofilia (IE) para cada variedad y estrato
La variedad Criollo 98 mostró el mayor valor deesclerofilia, y Habana 2000 el más bajo. Este últimovalor está vinculado a cutículas finas y peor estruc-turadas, células de paredes más delgadas y bajos valo-res de esclerofilia. Las condiciones de sombreo creadas,tanto por el efecto de la tela como por la superposiciónde las hojas de los estratos superiores, da lugar a unaalta humedad relativa, y por ende es menor la intensi-dad de la transpiración, factores estos que inducen laformación de cutículas y paredes celulares más delga-das [Fonseca y Medina, 2002]. Muestran además unabaja esclerosis debido a su mayor grado de suculenciapor constituir tejido joven (Tabla 5). Estas caracterís-ticas dan lugar a la presencia de hojas finas que permi-ten la permeabilidad a microelementos depositados enla superficie como Mn y Zn, los que penetran a travésde la membrana hasta el interior de los cloroplastos.
CONCLUSIONES
• La aplicación del fungicida mancozeb es la causa dela mancha verde en la hoja de tabaco al tener comoingrediente activo el Mn.
• La mayor incidencia de la aparición de la mancha ver-de es en los estratos inferiores. Los menos afectadosfueron los superiores. El testigo no tuvo afectación.
• La variedad Habana 2000 resultó ser la más sus-ceptible y la de más bajo índice de esclerofilia, mo-
Tabla 4. Número de hojas manchadas por estrato en cada variedad evaluada
Variedades Número de hojas/estrato Criollo 98 Corojo 99 Habana 2000
Estrato inferior 3,25 b 3,75 b 3,75 a
Estrato medio 2,50 a 4,25 b 10,00 b
Estrato superior 1,50 a 1,75 a 2,25 a
ES± 0,27 0,25 0,45
Tabla 5. Comportamiento del índice de esclerofilia en cada variedad
Variedades Índice de escerofilia
Criollo 98 0,74 a
Corojo 99 0,63 b
Habana 2000 0,53 c
ES± 0,01
Tabla 3 Comportamiento de las hojas encontradas con manchas por cada variedad de un total de 36 evaluadas
Variedades Número de hojas afectadas
Criollo 98 7,25 a
Corojo 99 9,75 b
Habana 2000 16,00 c
ES± 0,45

fitosanidad/23
Causas de la aparición de la enfermedad...
tivado en gran medida por sus características mor-fológicas.
• La variedad Criollo 98 fue la más tolerante y la demás alto índice de esclerofilia, por lo que este aspec-to se relaciona con incidencia de la mancha verde.
REFERENCIAS
Espino, E.; Xiomara Rey; V. García; Nilda Peñalver; L. M. Guardiola:«Habana 92 y Habana 2000. Dos variedades cubanas de tabaconegro resistentes al moho azul (Perenospora tabacina)», RevistaCubana de Agricultura 1(1):15-24, 2000.
Fonseca, I.; R. Medina: «Influencia de varios tipos de tela y densidadesde plantación sobre algunos indicadores anatómicos en hojas detabaco (Nicotiana tabacum L.) cultivado bajo tela». Memorias delCongreso del INCA (Programa de Resúmenes), Ediciones INCA, 12-15 de noviembre de 2002.
Jordan, D. B.; W. L. Ogren: «A Sensitive Assay Procedure forSimultaneous Determination of Ribulose-1,5 BisphosphateCarboxylase and Oxygenase Activities», Plant Physiol. 67:237-245, 1981.
Kitao, M.; T. T. Lei; T. Kioke: «Comparison of Photosynthetic Responsesto Manganese Toxicity of Deciduous Broad-Leaved Trees in NorthernJapan», Environmental Pollution l.97 (1-2):113-118, 1997a.
Kitao, M.; T. T. Lei; T. Kioke: «Effects of Manganese Toxicity onPhotosynthesis of White Birch (Betula platyphylla var. japonica)Seedlenys», Plant Physiol. 101:249-256, 1997b.
––––: «Application of Chlorophyll Fluorescence to Evaluate Mn Toleranceof Deciduous Broad-Leaved Tree Seedlings Native to NorthernJapan», Tree Physiology 18:135-140, 1998.
MINAGRI: «Manual técnico para la producción del tabaco negro tapa-do», Ed. Ministerio de la Agricultura, La Habana, 2001.
––––: «Programa de defensa del cultivo del tabaco campaña 1999/2000»,Centro Nacional de Sanidad Vegetal, Subdivisión de Protección dePlantas, Departamento Programas de Defensa, La Habana, 1999.
Nable, R. O.; R. L Houtz; G. M. Cheniae: «Early Inhibition ofPhotosynthesis During Development of Mn Toxicity in Tobacco», PlantPhysiol. 86:1136-1142, 1988.
Pequeño, J.: Prácticas de fisiología vegetal, Editora del Consejo Na-cional de Universidades, La Habana, 1964.
Vázquez, Edith; S. Torres. Fisiología vegetal, Ed. Pueblo y Educación,La Habana, 1995.

24/fitosanidad

fitosanidad/25
FITOSANIDAD vol. 10, no. 1, marzo 2006
Eco
logí
a EFECTO DE LOS MEDIOS DE CULTIVO Y LA TEMPERATURAEN EL CRECIMIENTO DE CLADOSPORIUM FULVUM COOKE(SIN. PASSALORA FULVA)
Alexander Bernal Cabrera,1 Benedicto Martínez Coca,2 Manuel Díaz Castellanos,1 Lidcay Herrera Isla1
y Yanelys Alonso Díaz1
1 Universidad Central de Las Villas. Carretera a Camajuaní Km 5½, Santa Clara, Villa Clara, Cuba, CP 54830, [email protected] Centro Nacional de Sanidad Agropecuaria (CENSA). Apartado 10, San José de las Lajas, La Habana
RESUMEN
El moho de la hoja causado por Cladosporium fulvum es una de lasenfermedades más comunes del tomate en los invernaderos cuba-nos. Para conocer la influencia del medio de cultivo y la temperaturase estudió el crecimiento micelial y la esporulación de tres aisla-mientos de C. fulvum (nos. 3, 6 y 10), procedentes de plantacionescomerciales de tomate bajo condiciones de invernadero, en cuatromedios de cultivo (PDA, SDA, AEM y Czapek) y cinco temperaturasen un rango entre 15 y 35ºC. Los medios de cultivos papa-dextrosaagar y sabouraud-dextrosa-agar, así como las temperaturas com-prendidas entre 20 y 25ºC, y 25 y 30ºC, fueron las condiciones ópti-mas para diferenciar los aislamientos de este hongo en cuanto a sucrecimiento micelial y esporulación.
Palabras clave: medios de cultivo, temperatura, esporulación, creci-miento micelial
ABSTRACT
Leaf mould caused by Cladosporium fulvum is one of the most commontomato diseases in Cuban greenhouses. In order to know the effect ofculture media and temperature on mycelial growth and sporulation ofthree C. fulvum isolation (numbers 3, 6 and 10), coming fromcommercial tomato plantation under greenhouse conditions, wasrealized a study in four culture media (PDA, SDA, MEA and Czapek)and five temperatures (from 15 to 35ºC). Potato-Dextrose-Agar andSabouraud-Dextrose-Agar culture media, as well as temperaturesbetween 20-25ºC and 25-30ºC, were the optimum conditions to differamong the isolations of this fungus as for their mycelial growth andsporulation.
Key words: culture media, temperature, sporulation, mycelial growth
INTRODUCCIÓN
Cladosporium fulvum (sin. Passalora fulva) [Braun etal., 2003] es uno de los principales hongos fitopatógenosque incide en el tomate en Cuba, bajo las condicionesde cultivo protegido. Este microorganismo provoca,junto a otros agentes fúngicos, pérdidas importantes[Bernal et al., 2001], y está ampliamente distribuidoen las regiones donde se siembra el cultivo [Blancard.,1992; Jones et al., 1997].
La temperatura y el medio de cultivo son aspectos fun-damentales en la fisiología y distribución de los hongos[Thomma et al., 2005]; sin embargo, existen pocas ex-periencias sobre la influencia de estos factores en elhongo mencionado, y son en la mayoría de los casosempíricas. Por ello el presente trabajo se propone estu-diar el efecto del medio de cultivo y la temperaturasobre el crecimiento de este patógeno, y así incremen-tar los conocimientos acerca de su biología con el fin depoder ejercer sobre él un control más eficaz.
MATERIALES Y MÉTODOS
Se evaluó la influencia de los medios de cultivo papadextrosa-agar (PDA), czapek dox-agar (ACZ), agarextracto de malta (AEM) y Sabouraud-dextrosa-agar(SDA) con pH 5,6 y las temperaturas 15, 20, 25, 30 y35ºC sobre el crecimiento de C. fulvum. Para ello se sem-braron discos de micelio de 7 mm de diámetro de culti-vos monospóricos de tres aislamientos de este patóge-no (Tabla 1) en cada una de cuatro placas Petri porvariante, las que contenían los medios de cultivo res-pectivos, y se incubaron a las temperaturas señaladas.El diámetro de las colonias por aislados se midió a lossiete y catorce días, y la esporulación a los catorce díasde incubación. Se vertieron 10 mL de agua destiladasobre las colonias y se barrió suavemente con la ayudade un pincel. La concentración de conidios en la sus-pensión obtenida se determinó mediante la cámara deNeubauer por observación al microscopio ópticoOlympus con 40X de aumento.
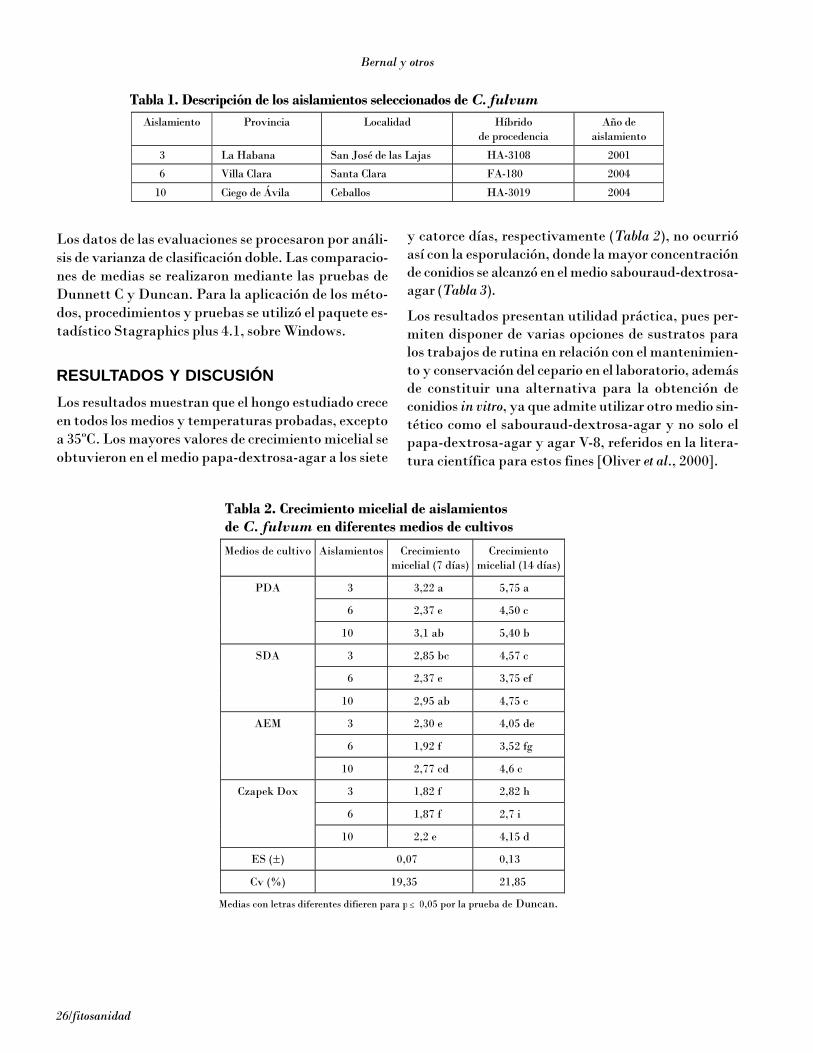
Bernal y otros
26/fitosanidad
Tabla 1. Descripción de los aislamientos seleccionados de C. fulvum
Aislamiento Provincia Localidad Híbrido de procedencia
Año de aislamiento
3 La Habana San José de las Lajas HA-3108 2001
6 Villa Clara Santa Clara FA-180 2004
10 Ciego de Ávila Ceballos HA-3019 2004
Los datos de las evaluaciones se procesaron por análi-sis de varianza de clasificación doble. Las comparacio-nes de medias se realizaron mediante las pruebas deDunnett C y Duncan. Para la aplicación de los méto-dos, procedimientos y pruebas se utilizó el paquete es-tadístico Stagraphics plus 4.1, sobre Windows.
RESULTADOS Y DISCUSIÓN
Los resultados muestran que el hongo estudiado creceen todos los medios y temperaturas probadas, exceptoa 35ºC. Los mayores valores de crecimiento micelial seobtuvieron en el medio papa-dextrosa-agar a los siete
y catorce días, respectivamente (Tabla 2), no ocurrióasí con la esporulación, donde la mayor concentraciónde conidios se alcanzó en el medio sabouraud-dextrosa-agar (Tabla 3).
Los resultados presentan utilidad práctica, pues per-miten disponer de varias opciones de sustratos paralos trabajos de rutina en relación con el mantenimien-to y conservación del cepario en el laboratorio, ademásde constituir una alternativa para la obtención deconidios in vitro, ya que admite utilizar otro medio sin-tético como el sabouraud-dextrosa-agar y no solo elpapa-dextrosa-agar y agar V-8, referidos en la litera-tura científica para estos fines [Oliver et al., 2000].
Tabla 2. Crecimiento micelial de aislamientos de C. fulvum en diferentes medios de cultivos
Medios de cultivo Aislamientos Crecimiento micelial (7 días)
Crecimiento micelial (14 días)
3 3,22 a 5,75 a
6 2,37 e 4,50 c
PDA
10 3,1 ab 5,40 b
3 2,85 bc 4,57 c
6 2,37 e 3,75 ef
SDA
10 2,95 ab 4,75 c
3 2,30 e 4,05 de
6 1,92 f 3,52 fg
AEM
10 2,77 cd 4,6 c
3 1,82 f 2,82 h
6 1,87 f 2,7 i
Czapek Dox
10 2,2 e 4,15 d
ES (±) 0,07 0,13
Cv (%) 19,35 21,85
Medias con letras diferentes difieren para p ? 0,05 por la prueba de Duncan.
≤
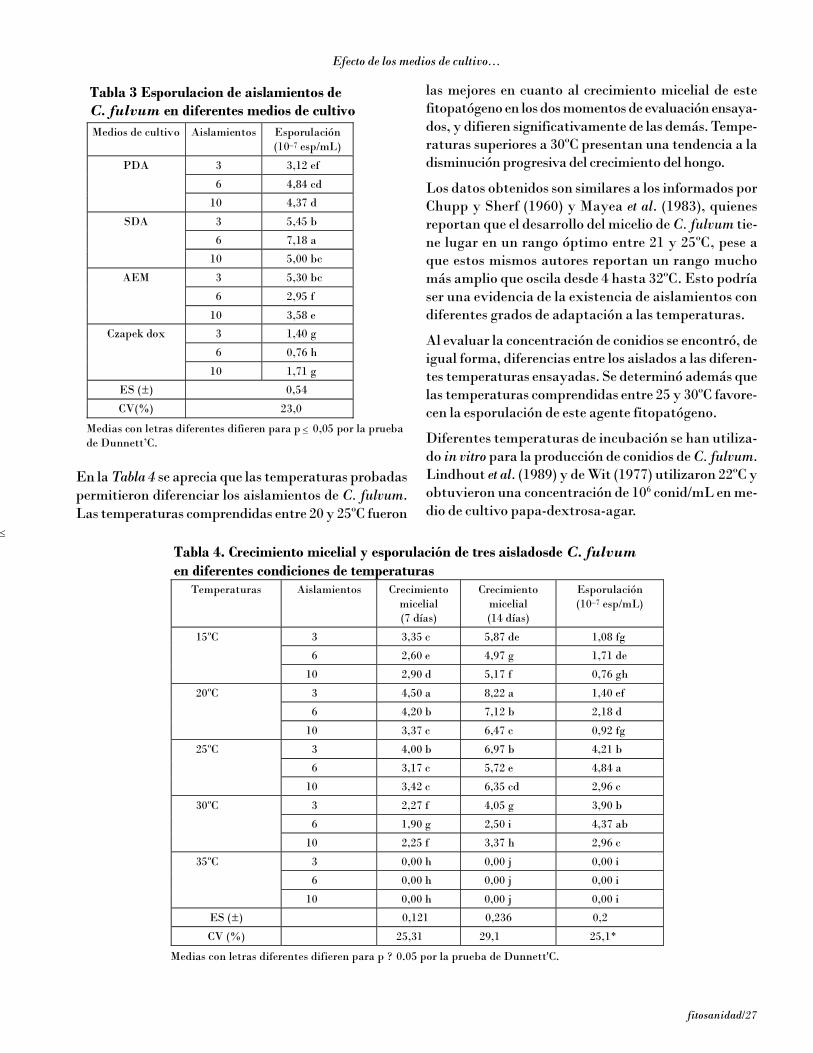
fitosanidad/27
Efecto de los medios de cultivo...
Tabla 3 Esporulacion de aislamientos de C. fulvum en diferentes medios de cultivo
Medios de cultivo Aislamientos Esporulación (10–7 esp/mL)
3 3,12 ef
6 4,84 cd
PDA
10 4,37 d
3 5,45 b
6 7,18 a
SDA
10 5,00 bc
3 5,30 bc
6 2,95 f
AEM
10 3,58 e
3 1,40 g
6 0,76 h
Czapek dox
10 1,71 g
ES (±) 0,54
CV(%) 23,0
Medias con letras diferentes difieren para p ? 0,05 por la prueba de Dunnett’C. En la Tabla 4 se aprecia que las temperaturas probadaspermitieron diferenciar los aislamientos de C. fulvum.Las temperaturas comprendidas entre 20 y 25ºC fueron
las mejores en cuanto al crecimiento micelial de estefitopatógeno en los dos momentos de evaluación ensaya-dos, y difieren significativamente de las demás. Tempe-raturas superiores a 30ºC presentan una tendencia a ladisminución progresiva del crecimiento del hongo.
Los datos obtenidos son similares a los informados porChupp y Sherf (1960) y Mayea et al. (1983), quienesreportan que el desarrollo del micelio de C. fulvum tie-ne lugar en un rango óptimo entre 21 y 25ºC, pese aque estos mismos autores reportan un rango muchomás amplio que oscila desde 4 hasta 32ºC. Esto podríaser una evidencia de la existencia de aislamientos condiferentes grados de adaptación a las temperaturas.
Al evaluar la concentración de conidios se encontró, deigual forma, diferencias entre los aislados a las diferen-tes temperaturas ensayadas. Se determinó además quelas temperaturas comprendidas entre 25 y 30ºC favore-cen la esporulación de este agente fitopatógeno.
Diferentes temperaturas de incubación se han utiliza-do in vitro para la producción de conidios de C. fulvum.Lindhout et al. (1989) y de Wit (1977) utilizaron 22ºC yobtuvieron una concentración de 106 conid/mL en me-dio de cultivo papa-dextrosa-agar.
Tabla 4. Crecimiento micelial y esporulación de tres aisladosde C. fulvum en diferentes condiciones de temperaturas
Temperaturas Aislamientos Crecimiento micelial (7 días)
Crecimiento micelial (14 días)
Esporulación (10–7 esp/mL)
3 3,35 c 5,87 de 1,08 fg
6 2,60 e 4,97 g 1,71 de
15ºC
10 2,90 d 5,17 f 0,76 gh
3 4,50 a 8,22 a 1,40 ef
6 4,20 b 7,12 b 2,18 d
20ºC
10 3,37 c 6,47 c 0,92 fg
3 4,00 b 6,97 b 4,21 b
6 3,17 c 5,72 e 4,84 a
25ºC
10 3,42 c 6,35 cd 2,96 c
3 2,27 f 4,05 g 3,90 b
6 1,90 g 2,50 i 4,37 ab
30ºC
10 2,25 f 3,37 h 2,96 c
3 0,00 h 0,00 j 0,00 i
6 0,00 h 0,00 j 0,00 i
35ºC
10 0,00 h 0,00 j 0,00 i
ES (±) 0,121 0,236 0,2
CV (%) 25,31 29,1 25,1*
Medias con letras diferentes difieren para p ? 0,05 por la prueba de Dunnett'C.
≤
≤

Bernal y otros
28/fitosanidad
Estos hallazgos resultan de importancia para trabajosgenéticos con este hongo y en la obtención de materialpara estudios de patogenicidad.
CONCLUSIONES
• Los mayores valores del crecimiento micelial yesporulación para los tres aislamientos se obtuvie-ron en los medios de cultivo papa-dextrosa-agar ysabouraud-dextrosa-agar.
• Las temperaturas entre 20 y 25ºC permitieron dife-renciar los aislamientos en cuanto a su crecimientomicelial, mientras que las comprendidas entre 25 y30ºC permitieron diferenciar los aislamientos en cuan-to a la esporulacion.
• La temperatura de 35ºC no permite diferenciar losaislamientos en cuanto al crecimiento micelial yesporulación.
REFERENCIAS
Blancard, D.: Enfermedades del tomate. Observar, identificar y lu-char, INRA, Ed. Mundi-Prensa, 1992.
Bernal, Blanca; L. Rivero; E. Fernández; Wendolyn Pérez: «Manejo deplagas en híbridos de tomate bajo condiciones de cultivo protegido»,Fitosanidad 5(1):57-61, 2001.
Braun, U.; P. W. Crous; F. Dugan; J. Z. Groenewald; G. S. de Hoog:«Phylogeny and Taxonomy of Cladosporium-like hyphomycetes,Including Davidiella Gen. Nov., the Teleomorph of Cladosporium»,Mycology Progress 2, 2003.
Chupp, C.; A. F. Sherf: Vegetable Disease and Their Control, Constable,Londres, 1960, pp. 541-545.
Jones, J. B.; J. P. Jones; R. E. Stall; T. A. Zitter: Compendium of TomatoDiseases, APS Press, E.U., 1997.
Lindhout, P.; W. Korta; M. Cislik; I. Vos; T. Gerlagh: «Further Identificationof Races of Cladosporium fulvum (Fulvia fulva) on Tomato Originatingfrom the Netherladns, France and Poland», Neth. J. Plant. Path.95:143-148, 1989.
Mayea, S.; L. Herrera; C. M. Andreu: Enfermedades de las plantas cultiva-das en Cuba, Ed. Pueblo y Educación, La Habana, 1983, pp. 234 y 235.
Oliver, R. P.; B. Henricot; G. Segers: Cladosporium fulvum, Cause ofLeaf Mould of Tomato», Fungal Pathology, Kluwer AcademicPublishers, 2000. pp. 65-91.
Thomma, B. P. H. J.; H. P. van Esse; P. W. Crous; P. J. G. M. de Wit:«Cladosporium fulvum (syn. Passalora fulva), a Highly SpecializedPlant Pathogen As Model for Functional Studies on Plant PathogenicMycosphaerellacea», Molecular Plant Pathology 6(4):379-393, 2005.
Wit, P. J. G. M. de: «Linght and Scanning-Electron Microscopic Study ofInfection of Tomato Plants by Virulent and Avirulent Races ofCladosporium fulvum», Neth. J. Plant Pathol. 83:109-122, 1977.

fitosanidad/29
FITOSANIDAD vol. 10, no. 1, marzo 2006
Con
trol
bio
lógi
co REPRODUCCIÓN DE HETERORHABDITIS INDICAEN CULTIVOS BIDIMENSIONALES ELABORADOSCON PROTEÍNA ANIMAL
Yirina Valdés Vázquez,1 Antonio A. Lobaina Audevert,1 María E. Márquez Gutiérrez1, Maylen GómezPacheco2 y Mercedes Escobar Hernández1
1 Instituto de Investigaciones de Sanidad Vegetal Calle 110 no. 514 e/ 5a. B y 5a. F, Playa, Ciudad de La Habana, CP 11600, [email protected] Instituto de Investigaciones de Fruticultura Tropical. Calle 7a. no. 3005 e/ 30 y 32, Playa, Ciudad de La Habana, [email protected]
RESUMEN
La reproducción in vivo de nematodos entomopatógenos es alta-mente costosa y laboriosa, lo que trae aparejado una gran necesi-dad de elaboración de metodologías para su reproducción in vitroa gran escala. Con este objetivo se evaluó la reproducción y desarro-llo de juveniles infectivos de Heterorhabditis indica [Poinar,Karunakar y David, 1992] sobre diferentes medios de cultivobidimensionales, que se elaboraron por la combinación de híga-do de pollo con almidón, así como hígado de cerdo con melaza,almidón y polvo de arroz. Los cuatro medios de cultivos permitie-ron la reproducción de los nematodos. Se destaca la variante conhígado de cerdo-polvo de arroz, que evidenció que la combinaciónentre la fuente de ácidos grasos y carbohidratos fue más eficazque en el resto de las variantes.
Palabras clave: Heterorhabditis indica, nematodos, reproducción
ABSTRACT
In vivo reproduction of entomopathogenic nematodes is highly costand laborious, so it necessary to develope new methodologies forhigh level in vitro reproduction. The objective of this project was toevaluate the reproduction and development of infective juveniles ofHeterorhebditis indica [Poinar, Karunakar y David, 1992] in fourbidimentional culture media. These culture media were elaboratedcombining chicken liver with starch and pork liver with molasses,starch and rice powder. Every culture medium allowed the reproductionof the nematodes primarily the variant with pork liver-rice powder dueto the combination of fatty acids and carbohydrates present in thisculture medium were more effective than in the rest culture media.
Key words: Heterorhabditis indica, nematodes, reproduction
INTRODUCCIÓN
Los nematodos entomopatógenos ocupan un lugar muyimportante en la lucha biológica como una alternativamás dentro de los métodos de control de plagas [Kayay Gaugler, 1993] debido a que el uso indiscriminado delos plaguicidas químicos causa grandes afectaciones enla diversidad biológica. A pesar de haberse estudiadosignificativamente desde 1980 y de emplearse con bue-nos resultados en cultivos como cítricos, arroz, maíz yhortalizas [Arteaga, et al., 1996], aún su producciónestá muy limitada y no satisface la demanda.
La reproducción de estos organismos sobre larvas deinsectos es muy costosa [Bedding, 1984], por lo queexisten numerosas patentes que se refieren fundamen-talmente a tecnologías para la reproducción masiva deforma artificial, como son el cultivo bidimensional, eltridimensional y la fermentación líquida. Estos méto-
dos de reproducción in vitro se deben ajustar a la dis-ponibilidad de materias primas, y que a su vez permi-tan una eficiente reproducción de los nematodos. Coneste objetivo se evaluó en este trabajo la reproduccióny desarrollo de juveniles infectivos de Heterorhabditisindica [Poinar, Karunakar y David, 1992] sobre cuatromedios de cultivo bidimensionales.
MATERIALES Y MÉTODOS
Se elaboraron cuatro medios de cultivos nutritivosbidimensionales en combinaciones de hígado de pollocon almidón e hígado de cerdo con melaza, almidón ypolvo de arroz. Por cada una de estas variantes se pre-pararon cuatro réplicas que se inocularon primeramentecon la bacteria simbionte Photorhabdus luminescens

Valdés y otros
30/fitosanidad
[Boemare, Akhurst y Mourant, 1993]. A las 48 h serealizaron dos perforaciones de 1 cm de diámetro en elcentro de cada placa, donde se inocularon 6 000 juveni-les infectivos de H. indica, previamente esterilizadoscon hipoclorito de sodio, y finalmente todo se incubó a28oC.
El diseño empleado en este experimento fue completa-mente aleatorizado. A partir de las 24 h se realizaronobservaciones diarias, y a los 13 días comenzó la reco-lección de los juveniles mediante el lavado de las placascon agua destilada esterilizada. Esta operación se repi-tió cuatro veces a intervalos de 24 h. Se contó la canti-dad de nemátodos extraídos de cada uno de los mediosde cultivo y se analizaron los datos mediante la dócimade comparación múltiple de Newman-Keuls con 5% deprobabilidad de error.
RESULTADOS Y DISCUSIÓN
Los cuatro medios evaluados permitieron la reproduc-ción de H. indica. Se observó que los juveniles inocula-dos en las placas se ubicaron en áreas con abundantecrecimiento bacteriano, es decir, alrededor de las colo-nias de P. luminescens. Esto representa la gran relaciónque existe entre los nematodos y sus simbiontes, puesellas son las responsables de transformar el sustrato yconvertirlo en una fuente asequible para su alimentación[Forst y Clarke, 2002]. Como organismos microvívoros,estos nematodos pueden además alimentarse de labiomasa bacteriana, y esto garantiza que las nuevas ge-neraciones también presenten la bacteria simbionte alo-jada en su intestino, lista para ser liberada en el interiorde otro insecto hospedero [Boemare, 2002].
Otros autores plantean también la necesidad que tieneel nematodo de su simbionte bacteriano, fundamental-mente para el género Heterorhabditis, donde no se ha lo-grado la reproducción de nematodos axénicos [Gerritseny Smits, 1993; Lunau et al., 1993], es decir, que no con-tengan la bacteria simbionte en su intestino.
A las 72 h de incubación se observaron hembras gigan-tes en todos los medios elaborados con hígado de cerdo,lo que evidenció el desarrollo de los juveniles infectivosa estado adulto. Durante la recolección de las nuevasgeneraciones de juveniles infestados (JI) la presenciade adultos fue mínima; sin embargo, en la combinaciónhígado de pollo-almidón las primeras hembras gigan-tes aparecieron a los 13 días, y el ciclo reproductivo seextendió hasta los 24. Cuando se recolectaron ademáslos nemátodos de las placas, conjuntamente con los ju-
veniles infectivos se extrajo un gran número de hem-bras en estado de gravidez.
Se conoce que cuando el nematodo llega al hemocele delinsecto, sale de su estado de juvenil infectivo parareiniciar su desarrollo, proceso que es denominado re-cuperación. La mayoría de las señales para la ocurren-cia de este evento se producen en la hemolinfa del hos-pedero; pero Grewal et al. (1997) demostraron que labacteria simbionte también produce señales que con-trolan el desarrollo del nemátodo. Muchas de estas se-ñales deben estar altamente relacionadas con la pro-ducción de metabolitos, pues cuando el simbiontebacteriano se cultiva artificialmente produce diferen-tes combinaciones de enzimas durante la etapa post-exponencial del crecimiento [Thaler et al., 1998], y en elcaso específico de la especie P. luminescens, Daborn etal. (2001) plantearon que también produce unametaloproteasa en esta misma fase del cultivo.
Todo este fenómeno antes descrito explica por qué lareproducción de H. indica sobre medios de cultivo ela-borados con hígado de pollo ocurrió con tanto tiempode diferencia con respecto a la reproducción sobre híga-do de cerdo. Como estas fuentes de proteína tienen di-ferente composición, el hígado de pollo resultó de másdifícil degradación para el simbionte bacteriano, la fasepostexponencial de crecimiento se alcanzó más tarde,se retardó la producción de metabolitos y consigo laseñal alimenticia que desencadenaría la recuperaciónde los juveniles infectivos.
En la Fig. 1 se evidencian las diferencias significativasde la reproducción de H. indica. En las variantes conhígado de cerdo el número de juveniles obtenidos fuesuperior que con hígado de pollo.
Este resultado también está relacionado con laasincronía observada en el desarrollo de los juveniles,causado por su pobre recuperación. Gaugler y Han(2002) plantearon que esto conlleva la obtención de bajosrendimientos en las producciones.
Otra causa importante es que el contenido lipídico enel hígado de cerdo es superior que en el de pollo. Eneste sentido Yoo et al. (2000) enfatizaron la importan-cia de los lípidos ricos en ácidos grasos monoinsaturadospara la elaboración de un medio de cultivo paraHeterorhabditis bacteriophora. Hatab y Gaugler (1997)plantearon además que los lípidos metabolizados porlos nematodos le proporcionan el 60% del total de suenergía, de manera que su presencia es primordial enlos medios de cultivo que se elaboren artificialmentepara su reproducción.

fitosanidad/31
Reproducción de Heterorhabditis...
Figura 1. Número de juveniles infectivos obtenidos por medio de cultivo. (Letras iguales nodifieren, p < 0,05).
Si se tiene en cuenta que los medios ensayados no fue-ron suplementados con lípidos ni sales, se puede corro-borar que los hígados de cerdo y pollo aportaron sufi-ciente cantidad de nutrientes como para permitir lareproducción de H. indica, pese a que los rendimientosno fueron tan elevados.
Finalmente estos resultados arrojaron que componen-tes del medio de cultivo como fuentes de proteínas,lípidos y carbohidratos, su origen y la forma en queellos se combinan, influyen en la reproducción de H. in-dica, lo que coincide con autores como Gaugler y Han(2002). No obstante, ellos también plantearon que elestablecimiento de todos estos parámetros depende dela especie de nemátodo por reproducir.
CONCLUSIONES
• Los cuatro medios de cultivo estudiados permitie-ron la reproducción y el desarrollo de H. indica.
• La mayor productividad se obtuvo con la combina-ción hígado de cerdo-polvo de arroz, mientras la va-riante hígado de pollo-almidón fue la menos produc-tiva.
REFERENCIAS
Arteaga, E.; M. Montes; E. Fernández; B. Chang; V. Calzadilla; O.Vázquez; G. Plumas; V. García: «Organization of the Work withEntomopathogenic Nematodes in Cuba», Third InternacionalNematology Congress, Grosier, Guadalupe Antilles, French WestIndies, 1996.
Bedding, R. A.: «Large Scale Production, Storage and Transport of theInsect-Parasitic Nematode Neoaplectana spp. and Heterorhabditisspp.», Ann. Appl. Biol. 104:117-120, 1984.
Boemare, N.: «Biology, Taxonomy and Systematics of Photorhabdusand Xenorhabdus», Entomopathogenic Nematology, New York, CABIPublishing, 2002, pp: 35-56.
Daborn, P. J.; N. Waterfield; M. A. Blight; R. French-Constant: «MeasuringVirulence Factor Expression by the Pathogenic BacteriumPhotorhabdus luminescens in Culture and During Insect Infection»,Journal of Bacteriology 183:5834-5839, 2001.
Forst, S.; D. Clarke: «Bacteria-Nematode Symbiosis», EntomopathogenicNematology, New York CABI Publishing, 2002, pp. 57-77.
Gaugler, R.; R. Han: «Production Technology», EntomopathogenicNematology, New York, CABI Publishing, 2002, pp. 289-310.
Gerritsen, L. J. M.; P. H. Smits: «Variation in Pathogenicity ofRecombinations of Heterorhabditis and Xenorhabdus luminescensstrain», Fundam. Appl. Nematol. 16:367-373, 1993.
Grewal, P. S.; M. Matsuura; V. Converse: «Mechanisms of Specificityof Association Between the Nematode Stinernema scapterisci andIts Symbiotic Bacterium», Parasitology 114:483-488, 1997.
Hatab, M. A.; R. Gaugler: «Growth-Mediated Variations in Fatty Acid ofXenorhabdus sp.», Journal of Applied Microbiology 82:351-358,1997.
Kaya, H. K.; R. Gaugler: «Entomopathogenic Nematodes», Annu. Rev.Entomol. 38:181-206, 1993.
Lunau, S.; S. Stoessel; A. J. Schmidt-Peisker; R. U. Ehlers: «Establish-ment of Monoxenic Inocula for Scaling Up for in Vitro Culture ofEntomopathogenic Nematodes Steinernema spp. andHeterorhabditis spp.», Nematologica 39:385-399, 1993.
Thaler, J. O.; B. Duvic; A. Givaudan; N. Boemare: «Isolation andEntomotoxic Properties of the Xenorhabdus nematophilus F1Lecithinase», Applied and Environmental Microbiology 64:2367-2373, 1998
Yoo, S. K.; I. Brown; R. Gaugler: «Liquid Media Development forHeterorhabditis bacteriophora: Lipid Source and Concentration»,Applied Microbiology and Biotechnology 54:759-763, 2000.

32/fitosanidad

fitosanidad/33
FITOSANIDAD vol. 10, no. 1, marzo 2006
INTRODUCCIÓN DE LA AVISPA DE COSTA DE MARFILCEPHALONOMIA STEPHANODERIS BETREM (HYMENOPTERA:BETHYLIDAE), PARASITOIDE DE LA BROCA DEL FRUTODEL CAFETO HYPOTHENEMUS HAMPEI FERRARI(COLEOPTERA: SCOLYTIDAE) EN CUBA
Eliel Peña Marrero,1 Mario García Hernández,2 Eliazar Blanco Rodríguez2 y Juan F. Barreras Gaytán3
1 Centro de Servicios Ambientales de Matanzas. CITMA. Autopista Sur, Km 14, Los Taínos, Varadero, Matanzas, Cuba2 Centro Nacional de Sanidad Vegetal. Ayuntamiento no. 231, e/ San Pedro y Lombillo, Plaza de la Revolución, Ciudad de La Habana3 El Colegio de la Frontera Sur. ECOSUR. Tapachula, Chiapas, México
RESUMEN
En mayo del 2003 se recibieron en Cuba 1 022 ejemplares adultosdel parasitoide Cephalonomia stephanoderis Betrem (Hymenoptera:Bethylidae), provenientes de los Laboratorios de Cría del Colegio dela Frontera Sur (ECOSUR) en Tapachula, Chiapas, México. Estaintroducción se realizó con el objetivo de adaptar los individuos a lascondiciones climáticas del país para el desarrollo de estudiosbioecológicos, su cría masiva en laboratorio y su posterior libera-ción y establecimiento en las áreas cafetaleras afectadas por laplaga Hypothenemus hampei Ferrari. La introducción se consideró unéxito, ya que el análisis arrojó que el índice de mortalidad total fue de6,75% (machos 31,1% y hembras 4,23% respectivamente), donde elíndice de sobrevivencia de viaje total fue de 93,2%.
Palabras clave: Cephalonomia stephanoderis, parasitoides, brocadel café, Hypothenemus hampei, control biológico
ABSTRACT
A total of 1 022 individual adults of Cephalonomia stephanoderis Betrem(Hymenoptera: Bethylidae) parasitoid were received in Cuba in Mayof 2003 from Mass Rearing Laboratories of Colegio de la FronteraSur (ECOSUR) in Tapachula, Chiapas, Mexico. This introduction wasrealized for adapting the insects to Cuban climatic conditions andthus develope bio-ecologic studies, massive rearing in laboratoriesand their subsequent release and establishment in coffee areasaffected by the pest Hypothenemus hampei Ferrari. The introductionwas considered a success, since the analysis give out that the totaldeath rate was to 6.75 % (male 31.1% and female 4.23% respectively),where the total index of journey survival was 93.2%.
Key words: Cephalonomia stephanoderis, parasitoids, coffee berryborer, Hypothenemus hampei, biological control.
INTRODUCCIÓN
En la mayoría de los países cafetaleros el control deHypothenemus hampei Ferrari se ha fundamentado enel uso de insecticidas; sin embargo, la resistencia de labroca al endosulfan, uno de los insecticidas más utili-zados para el control, así como los problemas de conta-minación ambiental y las características de la biologíade esta plaga, limitan el uso de estos productos quími-cos. Por ello, en los últimos años se han incrementadolos estudios para generar estrategias de lucha másbiorracionales, donde el control biológico es el de ma-yor nivel de investigación y aplicación en la actualidad[Brun et al., 1989; Mansingh, 1991].
Los programas de control biológico clásico para la lu-cha contra H. hampei se han sustentado en la impor-tación de parasitoides específicos, obtenidos en la re-
gión de origen de la plaga (África), en unos casos direc-tamente y en otros desde países de la región donde sehan establecido [Barrera et al., 1990a].
Murphy y Moore (1990) plantean que en el mundo sehan realizado, básicamente, introducciones de cuatroespecies: Cephalonomia stephanoderis Betrem (avispade Costa de Marfil), Prorops nasuta Waterston (avispade Uganda), Phymastichus coffea La Salle (avispa deTogo) y Heterospilus coffeicola Schmiedeknecht; sinembargo, se ha demostrado en los diferentes países enque la plaga está presente, que el control biológico conel parasitoide C. stephanoderis ha logrado disminuirconsiderablemente las poblaciones, y además se consi-dera la especie más estudiada y utilizada en estos pro-gramas, por poseer cualidades superiores a las restan-

Peña y otros
34/fitosanidad
tes, aunque existen referencias de complementación enel efecto supresor de las poblaciones de la broca, en li-beraciones combinadas de estos parasitoides específi-cos [Barrera et al., 1990b].
Las autoridades de Sanidad Vegetal del Ministerio dela Agricultura de Cuba consideran que la broca del caféconstituye el principal organismo nocivo en este culti-vo, no solo por los daños causados en diferentes áreascafetaleras del macizo montañoso oriental del país, sinoademás por su posible diseminación al resto de las áreascafetaleras de la isla, motivo por el cual se solicitó laintroducción del parasitoide C. stephanoderis, avispaespecífica de esta plaga. Este enemigo natural se consi-dera una alternativa viable y con efectividad compro-bada en diversos países de la región neotropical, por loque con su utilización se contribuirá a reducir los índi-
ces de la plaga y la utilización de insecticidas sintéti-cos. Mediante esta introducción se podrá disponer deuna nueva alternativa para la protección de áreas delagroecosistema cafetalero de montaña que puedan in-festarse con la plaga, y de obtener un sistema de mane-jo acorde con la tecnología de este cultivo e integrar eluso de esta avispa parásita al programa de manejo dela broca en Cuba.
MATERIALES Y MÉTODOS
Durante los primeros días de mayo del 2003 se introdu-jeron en Cuba los ejemplares adultos del parasitoide C.stephanoderis provenientes de los Laboratorios de Críadel Colegio de la Frontera Sur (ECOSUR) de Tapachula,Chiapas, en México. En la Tabla 1 se describen los ele-mentos esenciales de la introducción.
Tabla 1. Elementos que describen la introducción de la especie Cephalonomia stephanoderis Betrem en Cuba
Aspectos Descripción
Fechas de envío Fecha de embalaje: 8 de mayo de 2003 Fecha de transportación desde su origen hasta Ciudad de La Habana: 9 de mayo de 2003 Fecha de transportación en destino desde Ciudad de La Habana hasta los laboratorios del
Centro Nacional de Referencia Fitosanitaria para la Montaña, Buey Arriba, Granma: 10 de mayo de 2003 (seis de la tarde)
País exportador México
Lugar de procedencia Laboratorios del Colegio de la Frontera Sur. ECOSUR, Tapachula, Chiapas, México
Institución receptora Laboratorio de Cuarentena del Instituto de Investigaciones de Sanidad Vegetal (INISAV), Cuba
Destinatario Laboratorios del Centro Nacional de Referencia Fitosanitaria para la Montaña (CNRFM), Buey Arriba, Granma, Cuba
Vía de transportación Aérea desde México a Ciudad de La Habana y Terrestre desde Ciudad de La Habana a Buey Arriba, Granma
Condiciones del envío Satisfactorias
Tipo de embalaje Caja unicel de 30 x 20 x 25 cm, aditada con gel preservante de la temperatura
Vial o paquete 100 viales de vidrio de 7 x 1,5 cm provistos con tapa de malla organdí
Insecto huésped Ninguno
Especie benéfica Cephalonomia stephanoderis Betrem (Hymenoptera : Bethylidae)
Planta huésped Ninguna
Procedencia del material Colecta no aplicable Cría de laboratorio sí
Fecha de la colecta No aplicable (El material procede de un laboratorio de cría masiva)
Cantidad de individuos recibidos
1 022 ejemplares adultos alimentados con solución de miel de abeja
El material venía amparado por los documentos lega-les siguientes:
• Certificado Fitosanitario Internacional (emitido porlas autoridades de Sanidad Vegetal de México).
• Certificado de Origen y Pureza (emitido porECOSUR).
• Certificado de envío de parasitoides de la broca delcafé con vistas a satisfacer todos los requisitos parael movimiento de material biológico en territoriomexicano, los cuales son solicitados por las autori-dades de aduana, sanidad vegetal y SEMARNAP(medio ambiente) respectivamente.

fitosanidad/35
Introducción de la avispa...
• Licencia de Importación, Producción y Ensayo emi-tida por el Centro Nacional de Seguridad Biológicade Cuba.
• Licencia Ambiental del Laboratorio del Centro Na-cional de Referencia Fitosanitaria para la Montañaemitida por el Centro Nacional de Seguridad Bioló-gica de Cuba.
RESULTADOS Y DISCUSIÓN
Para el correcto desarrollo del trabajo fue necesariorealizar el Registro del Procedimiento Diario con losindividuos introducidos en el laboratorio. Este reque-ría que primeramente se examinaran por los especialis-
tas del Laboratorio Central de Cuarentena Vegetal delCentro Nacional de Sanidad Vegetal para determinarsi se trataba de la especie deseada, y mediante el em-pleo de las claves taxonómicas de Betrem (1961) secorroboró que correspondía a Cephalonomia stepha-noderis Betrem (Hymenoptera: Bethylidae), parasitoideespecífico de la broca del fruto del cafeto Hypothenemushampei Ferrari. Por otro lado, se analizó y determinóque a los individuos no les acompañaba ningún insectohuésped, y que la unidad reproductiva lo representa-ban 1 022 ejemplares adultos de la especie antes men-cionada. En la Tabla 2 se muestra la cronología de lasacciones desarrolladas durante el arribo del lote deparasitoides.
Tabla 2. Acciones desarrolladas durante el proceso de introducción y cuarentena post entrada del parasitoide Cephalonomia Stephanoderis Betrem en Cuba
Vivos Muertos Estado de Desarrollo Evolución Descendencia 09/5/03
– – Adultos Buena –
Acciones El material arribó a Cuba sobre las tres de la tarde, por la Terminal 3 del Aeropuerto Internacional José Martí de Ciudad de La Habana. Se entregó la documentación requerida por las autoridades fitosanitarias del aeropuerto y se emitió el Acta de Liberación correspondiente. Luego el material importado se llevó al Laboratorio de Cuarentena del Instituto de Investigaciones de Sanidad Vegetal para su traslado al día siguiente al Centro Nacional de Referencia Fitosanitaria para la Montaña, ubicado en Buey Arriba, Granma
Vivos Muertos Estado de Desarrollo Evolución Descendencia 10/5/03
– – Adultos Buena –
Acciones El traslado se realizó por carretera en auto climatizado, en un período aproximado de 10 h y se arribó al Centro Nacional de Referencia aproximadamente a las seis de la tarde. En el Laboratorio de Cuarentena Vegetal de ese centro se confirmó la pureza del material y su diagnóstico. Se observó, además la vitalidad de los individuos con vistas a comenzar el proceso de reproducción
Vivos Muertos Estado de desarrollo Evolución Descendencia 11/5/03
953 69 Adultos Buena No aplicable
Acciones Se determinó el índice de mortalidad total (6,75%) y por sexo (macho 31,1% y hembra 4,23% respectivamente), así como el índice de sobrevivencia de viaje total equivalente a un 93,2%. Los individuos vivos fueron liberados en frascos plásticos de 1,5 L de capacidad que contenían 400 cotiledones de café pergamino infestados con H. hampei, a razón de 40 ejemplares del parasitoide por frasco. Estos se trasladaron a un cuarto de cría masiva climatizado con temperatura entre 21-27ºC. Los individuos muertos se separaron y colocaron en cámara húmeda para su posterior observación a las 72 h. Se colectaron ejemplares de referencia para su conservación en el Laboratorio de Cuarentena Vegetal del CNSV
Vivos Muertos Estado de desarrollo Evolución Descendencia 14/5/03
– 69 Adultos Muertos No aplicable
Acciones A las 72 h se observaron bajo el estéreo los 69 individuos muertos que fueron colocados en cámara húmeda; solo se apreció en algunos individuos la presencia de hongo del género Aspergillus sp., el cual es común observar cuando el material entra en contacto con el ambiente
Se aprecia que los elementos más importante por des-tacar lo constituyen el índice de mortalidad de la espe-cie introducida, que fue de 6,75%, y el índice desobreviviencia de viaje total con 93,2%. Este últimoelemento hace considerar que la introducción fue exitosadebido al alto porcentaje de individuos vivos que so-brevivieron el viaje desde ECOSUR, México, hastaCuba, incluso, se registró un porcentaje de sobre-
viviencia de viaje mayor al alcanzado por México cuan-do se realizó la introducción de esta misma especiecon individuos procedentes de Togo en 1988, que fuede 88,5% y con 11,5% de mortalidad [Barrera et al.,1990c].
Otra de las acciones desarrolladas por los especialistascorrespondió al Registro de Desechos con los elemen-

Peña y otros
36/fitosanidad
tos que contenía la muestra recibida. En este análisisse determinó que lo integraban dos elementos con loscuales se procedió a su incineración:
1. Caja unicel de poliespuma con dimensiones de 30 x20 x 25 cm.
Causa por la que se desecha: material desechable.2. Caja de cartón con secciones divisorias. Causa por la que se desecha: material desechable.
CONCLUSIONES
• El lote de individuos de parasitoides venía ampara-do con todos los documentos legales para su salidadel país de procedencia e introducción en Cuba.
• Se comprobó que la especie introducida correspon-día a Cephalonomia stephanoderis Betrem, para-sitoide específico de la broca del fruto del cafeto Hypo-thenemus hampei Ferrari.
• Al material introducido se le realizó el proceso decuarentena postentrada en los laboratorios del Cen-tro Nacional de Referencia Fitosanitaria para la Mon-taña en Buey Arriba, Granma.
• La introducción de Cephalonomia stephanoderisBetrem en Cuba se consideró un éxito.
REFERENCIAS
Barrera, J. F.; D. Moore; Y. J. Abraham; S. T. Murphy; C. Prior:«Biological Control of the Coffee Berry Borer, Hypothenemus hampei,in Mexico and Possibilities for Further Action», Brighton Crop ProtectionConference-Pests and Diseases, 1990, International Institute ofBiological Control, Londres, 1990a, pp. 391-396.
Barrera, J. F.; F. Infante; M. Vega; O. González; E. Carrillo; O. Campos;R. Muñoz; A. Serrano; J. J. Osorto; B. Decazy; D. Moore: «Introduc-ción de Cephalonomia stephanoderis (Hymenoptera: Bethylidae) aCentroamérica para el control biológico de la broca del cafeto,Hypothenemus hampei (Coleoptera: Scolytidae)», Turrialba 40:570-574, Costa Rica, 1990b.
Barrera, J. F.; P. S. Baker; A. Schwarz; J. Valenzuela: «Introducción dedos especies de parasitoides africanos a México para el controlbiológico de la broca del cafeto Hypothenemus hampei (Ferr.)(Coleoptera: Scolytidae)», Folia Entomológica Mexicana 79:245-247, México, 1990c.
Betrem, J. G.: «Cephalonomia stephanoderis Nov. Spec. (Hymenoptera:Bethylidae)», Entomol Berichten. 21:183-184, 1961.
Brun, L. O.; C. Marcillaud; V. Gaudichon; D. Suckling: «EndosulphanResistance in Hypothenemus hampei (Coleoptera: Scolytidae) inNew Caledonia», Journal of Economic Entomology 82:1311-1316,1989.
Mansingh, A.: «Limitations of Insecticides in the Management of theCoffee Berry Borer Hypothenemus hampei Ferrari», Journal of CoffeeResearch 21 (2):67-98, 1991.
Murphy, S. T.; D. Moore: «Biological Control of the Coffee Berry Borer,Hypothenemus hampei (Ferrari) (Coleoptera: Scolytidae): PreviousProgrammes and Possibilities for the Future», Biocontrol News andInformation 11 (2):107-117, 1990.

fitosanidad/37
FITOSANIDAD vol. 10, no. 1, marzo 2006
VARIABILIDAD DE MYCOSPHAERELLA FIJIENSIS MORELET.ESTABILIDAD DE LA RESISTENCIA A SIGATOKA NEGRADE LOS CLONES HÍBRIDOS DE LA FHIA
Michel Pérez Miranda,1 Luis Pérez Vicente,1 Roberto Trujillo2 y Dulce M. Betancourt3
1 Instituto de Investigaciones de Sanidad Vegetal. Calle 110 no. 514 e/ 5a. B y 5a. F, Playa, Ciudad de La Habana, CP 11600, [email protected] LAPROSAV. Carretera Central, Extremo Oeste, Ciego de Ávila, Cuba3 ETPP Baracoa. Delegación de la Agricultura del municipio de Baracoa, Guantánamo, CubaM
anej
o d
e pl
agas
RESUMEN
La sigatoka negra (SN) causada por Mycosphaerella fijiensis Morelet(Mf) es la enfermedad más importante de los bananos y plátanos. EnCuba hay más de 12 000 ha sembradas de clones con resistenciaparcial, por lo que la presión de selección de patotipos con mayoragresividad puede considerarse elevada. Se evaluó la severidad delataque, la velocidad de evolución de la enfermedad, así como laformación de fructificaciones en las manchas de parcelas de clonessusceptibles y con resistencia parcial en dos localidades de la pro-vincia de La Habana, en la empresa La Cuba, en la provincia deCiego de Ávila, y en Baracoa, en la provincia de Guantánamo. Para-lelamente se inocularon, en condiciones controladas de laboratorio,fragmentos de hojas colocados en agar con benzimidazol en placasde Petri plásticas, con suspensiones de conidios provenientes deaislamientos obtenidos de poblaciones de Mf salvajes y de clonescon resistencia parcial, en los que se evaluó diariamente la cantidadde lesiones y la velocidad de evolución de los síntomas. Los clonesde la FHIA en las dos localidades de La Habana y en Baracoa mos-traron una resistencia elevada a la enfermedad, mientras que losniveles de ataques y la producción de pseudotecios en plantas deFHIA 18 de La Cuba, en Ciego de Ávila, fue similar a la de los clonessusceptibles, y mucho más intensa que la observada en 1994-1995,cuando los clones con resistencia parcial se introdujeron por prime-ra vez en Cuba. Las inoculaciones sobre fragmentos de hojas delclon FHIA 18 y de Gran enano con aislados salvajes demostraron lamayor agresividad de los últimos sobre el clon FHIA 18. Este consti-tuye el primer informe de cambios de la agresividad de las poblacio-nes del patógeno o de erosión de la resistencia parcial a sigatokanegra del clon FHIA 18.
Palabras clave: variabilidad patogénica, sigatoka negra, Mycos-phaerella fijiensis, test monocíclico
ABSTRACT
Black Sigatoka (BS) caused by Mycosphaerella fijiensis Morelet (Mf)is the most important disease of banana and plantains. In Cuba existmore than 12 thousand planted of FHIA hybrids with partial resistanceand hence, the pressure of selection of pathotypes with an increasedaggressiveness could be considered to be high. Experimental plotswith susceptible and partially resistant cultivars in Alquízar and Güi-ra de Melena in Havana province, La Cuba in Ciego de Avila, andBaracoa in Guantánamo were established where disease severityand the fructifications development in the spots were assessed. Alaboratory experiment under controlled conditions using leaf fragmentsof Grand Nain and FHIA 18 placed on agar + benzimidazole in plasticPetri plates and inoculated with conidial suspensions belonging tosingle ascospore isolates from wild Mf populations and partial resistantcultivars showing strongly BS attack were carried out where he amountof lesions and the speed of evolution of symptoms were daily assessed.FHIA cultivars in the two localities of Havana and in Baracoa showeda high resistant reaction to the disease meanwhile the level of attackand the pseudothecia production in Ciego de Avila was similar toCavendish cultivars, and more severe than the observed in 1994/95,when the cultivars these cultivars were introduced in Cuba for firsttime. Conidial inoculations on Grand Nain and FHIA 18 leaf fragmentswith isolates from the strongly attacked FHIA 18 plants from Ciego deAvila and from Cavendish cultivars (wild type) showed the higheraggressiveness of the former. This is considered the first report of achange of aggressiveness of Mf populations or the erosion of thepartial resistance to BS in the FHIA 18 cultivar.
Key words: black sigatoka, pathogenic variability, Mycosphaerellafijiensis, monocyclic test
INTRODUCCIÓN
La aparición de la sigatoka negra en Cuba, causada
por Mycosphaerella fijiensis Morelet a finales de 1990[Vidal, 1992], tuvo un impacto marcado en los costosde producción, pero especialmente en la estructuraclonal de la superficie del país plantada de musáceas
[Pérez et al., 2002]. Los bananos Cavendish (AAA) fue-ron sustituidos por los clones FHIA 23 (AAAA) yFHIA 18 (AAAB), que junto al FHIA 3 (AABB) ocu-pan alrededor de 12 000 ha. Asimismo, en la actualidadsolo se mantiene el 18% de las más de 43 000 ha de

Pérez y otros
38/fitosanidad
plátanos (AAB) existentes en 1990. Los plátanos sehan sustituido por el cultivo a gran escala del clon Bu-rro CEMSA (ABB), que ocupa unas 63 000 ha, y por elFHIA 3 (AABB).
El uso de variedades resistentes es, sin duda, la estra-tegia más económica y ambientalmente sostenible delucha contra sigatoka negra. Se han realizado numero-sos estudios sobre la reacción de los clones de bananosy plátanos a M. fijiensis [Meredith y Lawrence, 1970;Firman, 1972; Fouré et al., 1984; Fouré et al., 1990;Fouré, 1994; Hernández y Pérez, 2001; Rowe, 1998].
Fouré (1994) describió tres tipos de reacciones en Musafrente a sigatoka negra: a) la reacción de resistenciamuy alta o hipersensibilidad observada en YangambiKm 5 (subgrupo Ibota, AAA); b) la resistencia parcial,que se expresa por un alargamiento del ciclo y la dismi-nución de la reproducción del patógeno; c) la reaccióncompatible o de sensibilidad observada en los clonesCavendish y plátanos (AAB).
En 1996 Hernández y Pérez [citados por Pérez, 1998]determinaron la reacción y componentes de la resisten-cia a sigatoka negra de los clones FHIA 23, FHIA 2,FHIA 3, FHIA 18, SH 3436 y de un grupo importantede clones del banco de germoplasma de Cuba, entre losque se encontraban diferentes clones pertenecientes alsubgrupo Cavendish, plátanos (AAB), el Yangambi Km 5,el Paka y el Burro CEMSA (ABB), mantenidos sin tra-tamientos con fungicidas durante todo el tiempo queduraron las observaciones. Los clones FHIA mostraronun alargamiento estadísticamente significativo de laduración en días entre la emergencia de las hojas y laaparición de los primeros síntomas, en relación con elclon Gran enano susceptible. La resistencia parcial deestos clones se expresa mediante el alargamiento de laevolución o transición de los síntomas de rayas a necrosisy en la reducción del número de pseudotecios/mancha[Pérez, 1998]. En el Yangambi Km 5 las aloinfecciones apartir de inóculo producido en parcelas vecinas determi-naron la aparición de numerosas manchas y defoliaciones,pero no la formación de espermogonios y pseudotecios.
Fullerton y Olsen (1995) informaron de la presencia enNueva Guinea y las Islas del Pacífico de poblaciones deM. fijiensis virulentas sobre los clones Musa acuminataburmannica Calcutta 4, (T8), Paka y Yangambi Km 5.Melioum (1999) informó del cambio de reacción en elclon Yangambi Km 5 a las infecciones de sigatoka ne-gra en Camerún 10 años después de los informes deFouré et al. (1990).
La aparición de nuevos genotipos virulentos depatógenos capaces de vencer la resistencia de varieda-des resistentes es un fenómeno de presión de selección,determinado en gran medida por condiciones ambien-tales y de manejo del cultivo favorables a la reproduc-ción del patógeno y por la extensión ocupada por losgenotipos resistentes, o más directamente por la su-perficie ocupada por los genes de resistencia desplega-dos en la población [Van der Plank, 1963, 1968].
En Cuba más de 12 000 ha de superficie de banano yplátanos están cultivadas de híbridos FHIA [Pérez etal., 2002]. Estos clones crecen en una gran diversidadde condiciones, y en la mayoría de los casos cerca deplantaciones de bananos y plátanos susceptibles conataques severos de sigatoka negra, por lo que estánexpuestos a una carga severa de inóculo durante la épocalluviosa. La presión de selección de patotipos con ma-yor agresividad capaces de producir infecciones severassobre estos clones es por tanto elevada. Con estos ante-cedentes se desarrolló una investigación con los siguien-tes objetivos:
a) Conocer la estabilidad de la resistencia de los cloneshíbridos de la FHIA y diferentes clones de referen-cia de reacción conocida, en un estudio de tres añoscon diseños experimentales de bloques colocados endiferentes condiciones ecológicas de Cuba.
b) Comparar la virulencia in vitro de diferentes aisla-mientos de M. fijiensis bajo condiciones estan-darizadas de incubación.
MATERIALES Y MÉTODOS
1. Medición de la virulencia/patogenicidad en campo delas poblaciones de M. fijiensis en un grupo de clones di-ferenciales e híbridos en diferentes localidades del país.Epidemiología de sigatoka negra a nivel policíclico
En mayo del 2001 se seleccionaron parcelas de más de5 ha de los clones FHIA 2, FHIA 3, FHIA 18, FHIA23 y Gran enano en la finca Pulido, en Alquízar, La Ha-bana; también de los clones FHIA 2, FHIA 3, FHIA18, FHIA 21, FHIA 23 y Gran enano en la empresa LaCuba, en Ciego de Ávila, en el occidente y centro delpaís respectivamente, ambas localidades con pluvio-metría anual promedio entre 1 100 y 1 300 mm de llu-via; además parcelas de 50 plantas en Baracoa, provin-cia de Guantánamo, en las localidades de San Luis conlos clones FHIA 18 y Robusta, y en la de Mosquiterocon los clones FHIA 2, FHIA 3, FHIA 18, FHIA 19,FHIA 20, FHIA 21, FHIA 23 y Robusta (subgrupo

fitosanidad/39
Variabilidad de Mycosphaerella...
Cavendish, AAA). Esta región presenta una pluvio-metría de más de 1 700 mm de lluvia anuales.
Debido a que el huracán Michel destruyó totalmente laplantación de Alquízar, se estableció inmediatamenteotra en la Empresa de Cultivos Varios (ECV) de la lo-calidad de Güira de Melena, en la misma provincia deLa Habana, con los clones FHIA 1, FHIA 2, FHIA 18,FHIA 20, FHIA 21, Yangambi Km 5 y Gran enano, don-de se mantuvieron las observaciones hasta junio del 2003.
Evaluación del desarrollo de la enfermedad. Se evaluó laseveridad de los ataques en las diferentes parcelas pormediciones del área foliar afectada total, la hoja másjoven manchada [Stover y Dickson, 1970]; hoja más
joven abierta promedio con 10 o más lesiones en es-tado 5 y 6 de la descripción de Fouré (1982)], asícomo del número de pseudotecios por mancha en 200manchas tomadas al azar en cada parcela. Para ga-rantizar la uniformidad de las evaluaciones se im-primieron las escalas gráficas del desarrollo de laenfermedad y se procedió a uniformar los criteriosde evaluación.
Con el objetivo de detectar diferencias en el comporta-miento de los clones, en relación con cambios depatogenicidad en las poblaciones de M. fijiensis o de laestabilidad de la resistencia, la escala de Stover (1971),transformada por Gauhl (1990), fue modificada a suvez de la siguiente forma:
Valor Características
0 Sana (libre de síntomas o rayas en estado 1 a 3) 1 Hasta el 1% de área necrosada (o hasta 10 manchas en estado 4 al 6 de la
descripción de la evolución de los síntomas de Fouré et al. (1984)
2 Hasta el 5% del área foliar necrosada 3 De 6 a 16% del área foliar necrosada 4 De 16 a 33% del área foliar necrosada
5 De 33 a 50 % de área foliar necrosada
6 Más del 50% del área foliar necrosada
Los datos obtenidos se utilizaron en el cálculo del por-centaje de hojas sanas y el índice de infección II –valormedio ponderado del área foliar afectada o índice deMcKinney [McKinney, 1923, citado también porTowsend y Heuberguer según Unterstenhöefer (1963):
II = (∑ an/6N) x 100
donde:a: Grado de la escalan: Número de hojas en cada grado de la escalaN: Total de hojas evaluadas
Se determinó además la velocidad de evolución (sumabruta (SBEE) de la enfermedad en las diferentes par-celas de acuerdo con el procedimiento descrito por Fouré(1988).
Determinación de la producción de fructificaciones. Semuestrearon en los días 28 de febrero (estación seca yfría) y 14 de septiembre de 2003 (estación caliente ylluviosa), 200 manchas en estado 6 de hojas de los clonesFHIA 18 y Gran enano, previamente marcadas en elmomento de su emergencia, para realizar conteos de lacantidad de fructificaciones presentes en las localida-des de Güira de Melena en La Habana, y La Cuba, enCiego de Ávila. Las manchas se colocaron en un beaker
con lactofenol, que a su vez se puso en un baño conagua hirviendo durante 3-5 min para aclararlas; se ob-servaron al microscopio óptico y se contó el número depseudotecios, espermogonios y conidióforos de M. fi-jiensis.
2. Comparación de la agresividad de diferentes aislamien-tos de M. fijiensis. Epidemiología de la sigatoka negra anivel monocíclico en condiciones controladas
Se obtuvieron aislamientos monoascósporicos de plan-taciones afectadas de FHIA 18 de La Cuba y de bananossusceptibles Cavendish (AAA), de zonas donde nuncase habían introducidos clones con resistencia parcial enla zona del Cerro en la Ciudad de La Habana, que semantuvieron en PDA a 27ºC y colocados durante 48 ha 16ºC para inducir la formación de conidios.
Para poner en evidencia diferencias de agresividad en-tre aislados provenientes de fincas plantadas de FHIA18 (AAAB) con fuertes afectaciones de sigatoka negracon los obtenidos de zonas donde no existen clones conresistencia parcial bajo condiciones controladas (a par-tir del hecho que la resistencia parcial es expresadacuantitativamente y que el ambiente tiene una influen-cia marcada sobre el desarrollo de la enfermedad), se
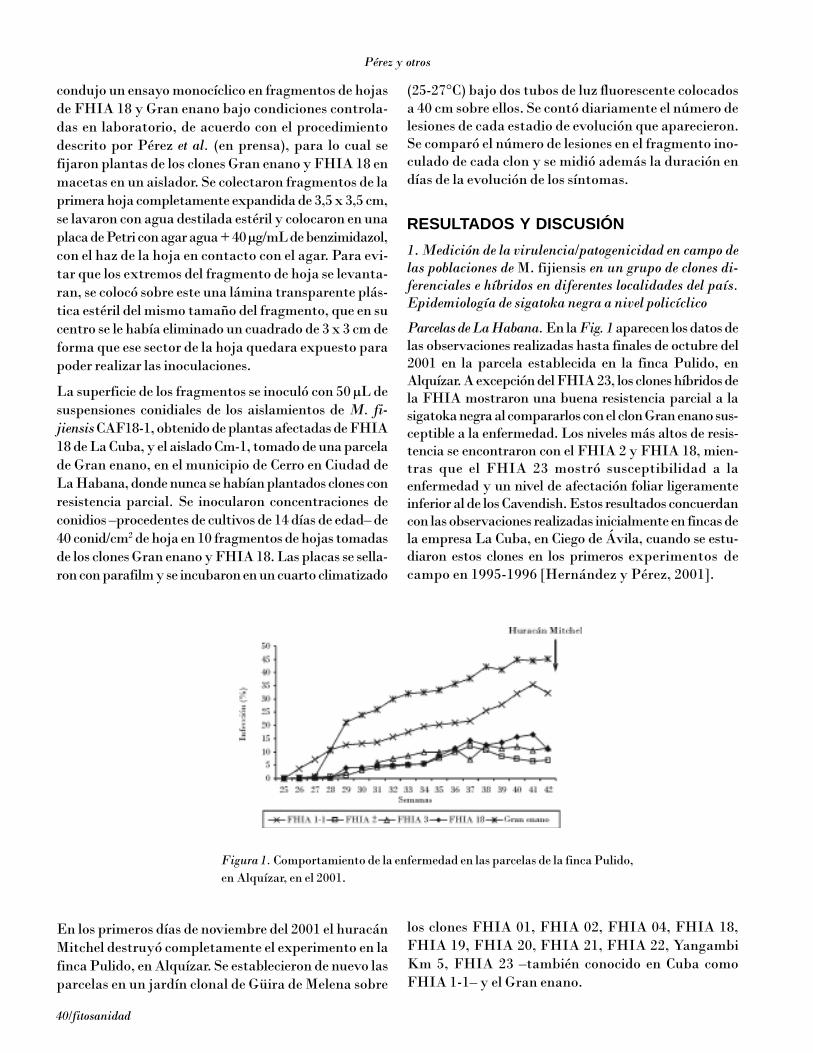
Pérez y otros
40/fitosanidad
condujo un ensayo monocíclico en fragmentos de hojasde FHIA 18 y Gran enano bajo condiciones controla-das en laboratorio, de acuerdo con el procedimientodescrito por Pérez et al. (en prensa), para lo cual sefijaron plantas de los clones Gran enano y FHIA 18 enmacetas en un aislador. Se colectaron fragmentos de laprimera hoja completamente expandida de 3,5 x 3,5 cm,se lavaron con agua destilada estéril y colocaron en unaplaca de Petri con agar agua + 40 µg/mL de benzimidazol,con el haz de la hoja en contacto con el agar. Para evi-tar que los extremos del fragmento de hoja se levanta-ran, se colocó sobre este una lámina transparente plás-tica estéril del mismo tamaño del fragmento, que en sucentro se le había eliminado un cuadrado de 3 x 3 cm deforma que ese sector de la hoja quedara expuesto parapoder realizar las inoculaciones.
La superficie de los fragmentos se inoculó con 50 µL desuspensiones conidiales de los aislamientos de M. fi-jiensis CAF18-1, obtenido de plantas afectadas de FHIA18 de La Cuba, y el aislado Cm-1, tomado de una parcelade Gran enano, en el municipio de Cerro en Ciudad deLa Habana, donde nunca se habían plantados clones conresistencia parcial. Se inocularon concentraciones deconidios –procedentes de cultivos de 14 días de edad– de40 conid/cm2 de hoja en 10 fragmentos de hojas tomadasde los clones Gran enano y FHIA 18. Las placas se sella-ron con parafilm y se incubaron en un cuarto climatizado
(25-27°C) bajo dos tubos de luz fluorescente colocadosa 40 cm sobre ellos. Se contó diariamente el número delesiones de cada estadio de evolución que aparecieron.Se comparó el número de lesiones en el fragmento ino-culado de cada clon y se midió además la duración endías de la evolución de los síntomas.
RESULTADOS Y DISCUSIÓN
1. Medición de la virulencia/patogenicidad en campo delas poblaciones de M. fijiensis en un grupo de clones di-ferenciales e híbridos en diferentes localidades del país.Epidemiología de sigatoka negra a nivel policíclico
Parcelas de La Habana. En la Fig. 1 aparecen los datos delas observaciones realizadas hasta finales de octubre del2001 en la parcela establecida en la finca Pulido, enAlquízar. A excepción del FHIA 23, los clones híbridos dela FHIA mostraron una buena resistencia parcial a lasigatoka negra al compararlos con el clon Gran enano sus-ceptible a la enfermedad. Los niveles más altos de resis-tencia se encontraron con el FHIA 2 y FHIA 18, mien-tras que el FHIA 23 mostró susceptibilidad a laenfermedad y un nivel de afectación foliar ligeramenteinferior al de los Cavendish. Estos resultados concuerdancon las observaciones realizadas inicialmente en fincas dela empresa La Cuba, en Ciego de Ávila, cuando se estu-diaron estos clones en los primeros experimentos decampo en 1995-1996 [Hernández y Pérez, 2001].
Figura 1. Comportamiento de la enfermedad en las parcelas de la finca Pulido,en Alquízar, en el 2001.
En los primeros días de noviembre del 2001 el huracánMitchel destruyó completamente el experimento en lafinca Pulido, en Alquízar. Se establecieron de nuevo lasparcelas en un jardín clonal de Güira de Melena sobre
los clones FHIA 01, FHIA 02, FHIA 04, FHIA 18,FHIA 19, FHIA 20, FHIA 21, FHIA 22, YangambiKm 5, FHIA 23 –también conocido en Cuba comoFHIA 1-1– y el Gran enano.
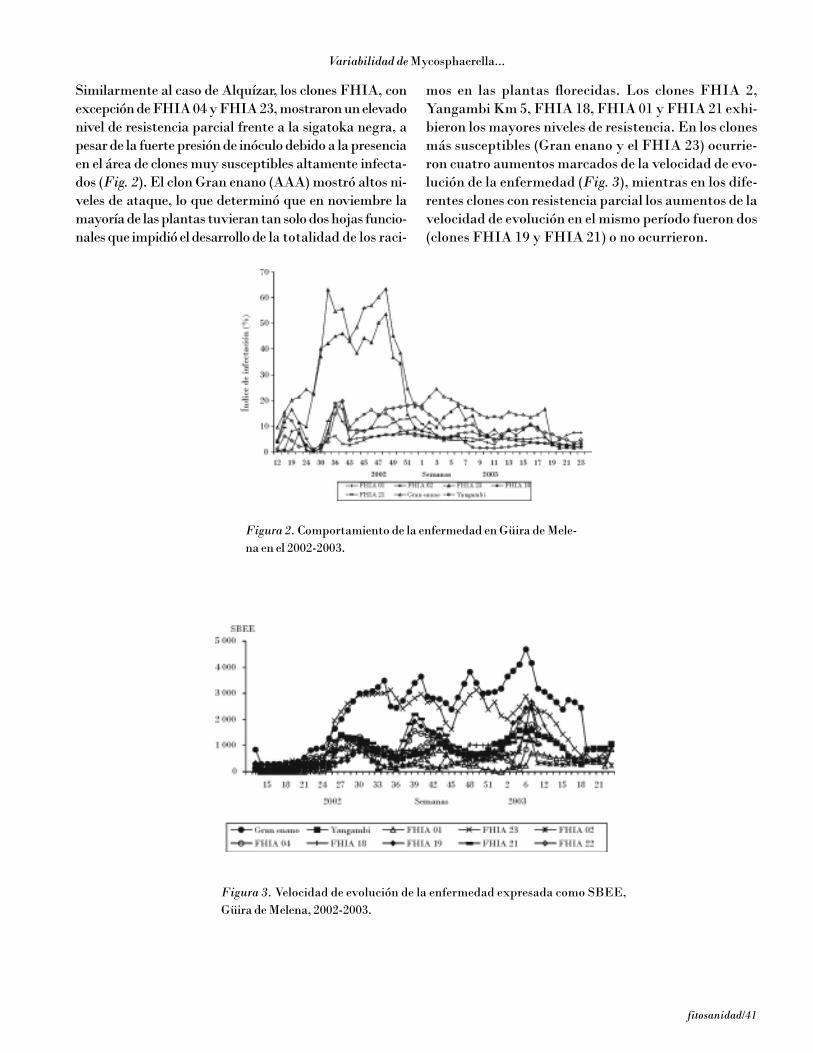
fitosanidad/41
Variabilidad de Mycosphaerella...
Similarmente al caso de Alquízar, los clones FHIA, conexcepción de FHIA 04 y FHIA 23, mostraron un elevadonivel de resistencia parcial frente a la sigatoka negra, apesar de la fuerte presión de inóculo debido a la presenciaen el área de clones muy susceptibles altamente infecta-dos (Fig. 2). El clon Gran enano (AAA) mostró altos ni-veles de ataque, lo que determinó que en noviembre lamayoría de las plantas tuvieran tan solo dos hojas funcio-nales que impidió el desarrollo de la totalidad de los raci-
mos en las plantas florecidas. Los clones FHIA 2,Yangambi Km 5, FHIA 18, FHIA 01 y FHIA 21 exhi-bieron los mayores niveles de resistencia. En los clonesmás susceptibles (Gran enano y el FHIA 23) ocurrie-ron cuatro aumentos marcados de la velocidad de evo-lución de la enfermedad (Fig. 3), mientras en los dife-rentes clones con resistencia parcial los aumentos de lavelocidad de evolución en el mismo período fueron dos(clones FHIA 19 y FHIA 21) o no ocurrieron.
Figura 2. Comportamiento de la enfermedad en Güira de Mele-na en el 2002-2003.
Figura 3. Velocidad de evolución de la enfermedad expresada como SBEE,Güira de Melena, 2002-2003.
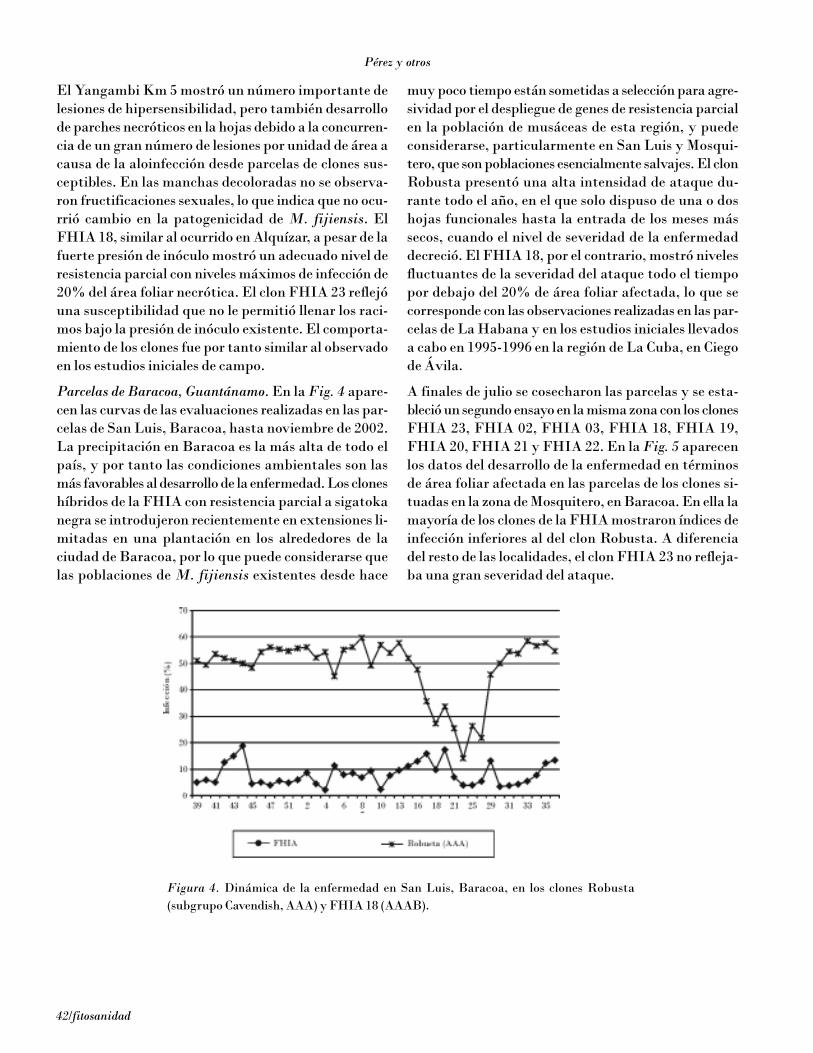
Pérez y otros
42/fitosanidad
El Yangambi Km 5 mostró un número importante delesiones de hipersensibilidad, pero también desarrollode parches necróticos en la hojas debido a la concurren-cia de un gran número de lesiones por unidad de área acausa de la aloinfección desde parcelas de clones sus-ceptibles. En las manchas decoloradas no se observa-ron fructificaciones sexuales, lo que indica que no ocu-rrió cambio en la patogenicidad de M. fijiensis. ElFHIA 18, similar al ocurrido en Alquízar, a pesar de lafuerte presión de inóculo mostró un adecuado nivel deresistencia parcial con niveles máximos de infección de20% del área foliar necrótica. El clon FHIA 23 reflejóuna susceptibilidad que no le permitió llenar los raci-mos bajo la presión de inóculo existente. El comporta-miento de los clones fue por tanto similar al observadoen los estudios iniciales de campo.
Parcelas de Baracoa, Guantánamo. En la Fig. 4 apare-cen las curvas de las evaluaciones realizadas en las par-celas de San Luis, Baracoa, hasta noviembre de 2002.La precipitación en Baracoa es la más alta de todo elpaís, y por tanto las condiciones ambientales son lasmás favorables al desarrollo de la enfermedad. Los cloneshíbridos de la FHIA con resistencia parcial a sigatokanegra se introdujeron recientemente en extensiones li-mitadas en una plantación en los alrededores de laciudad de Baracoa, por lo que puede considerarse quelas poblaciones de M. fijiensis existentes desde hace
muy poco tiempo están sometidas a selección para agre-sividad por el despliegue de genes de resistencia parcialen la población de musáceas de esta región, y puedeconsiderarse, particularmente en San Luis y Mosqui-tero, que son poblaciones esencialmente salvajes. El clonRobusta presentó una alta intensidad de ataque du-rante todo el año, en el que solo dispuso de una o doshojas funcionales hasta la entrada de los meses mássecos, cuando el nivel de severidad de la enfermedaddecreció. El FHIA 18, por el contrario, mostró nivelesfluctuantes de la severidad del ataque todo el tiempopor debajo del 20% de área foliar afectada, lo que secorresponde con las observaciones realizadas en las par-celas de La Habana y en los estudios iniciales llevadosa cabo en 1995-1996 en la región de La Cuba, en Ciegode Ávila.
A finales de julio se cosecharon las parcelas y se esta-bleció un segundo ensayo en la misma zona con los clonesFHIA 23, FHIA 02, FHIA 03, FHIA 18, FHIA 19,FHIA 20, FHIA 21 y FHIA 22. En la Fig. 5 aparecenlos datos del desarrollo de la enfermedad en términosde área foliar afectada en las parcelas de los clones si-tuadas en la zona de Mosquitero, en Baracoa. En ella lamayoría de los clones de la FHIA mostraron índices deinfección inferiores al del clon Robusta. A diferenciadel resto de las localidades, el clon FHIA 23 no refleja-ba una gran severidad del ataque.
Figura 4. Dinámica de la enfermedad en San Luis, Baracoa, en los clones Robusta(subgrupo Cavendish, AAA) y FHIA 18 (AAAB).
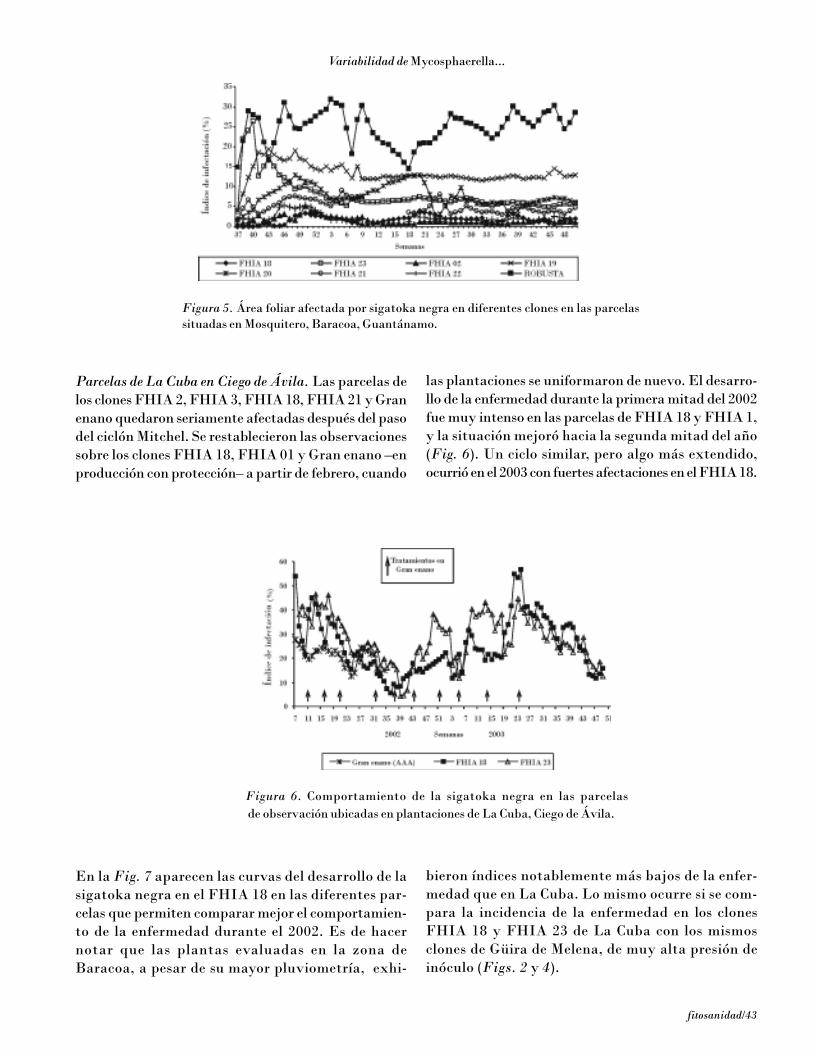
fitosanidad/43
Variabilidad de Mycosphaerella...
Figura 5. Área foliar afectada por sigatoka negra en diferentes clones en las parcelassituadas en Mosquitero, Baracoa, Guantánamo.
Parcelas de La Cuba en Ciego de Ávila. Las parcelas delos clones FHIA 2, FHIA 3, FHIA 18, FHIA 21 y Granenano quedaron seriamente afectadas después del pasodel ciclón Mitchel. Se restablecieron las observacionessobre los clones FHIA 18, FHIA 01 y Gran enano –enproducción con protección– a partir de febrero, cuando
las plantaciones se uniformaron de nuevo. El desarro-llo de la enfermedad durante la primera mitad del 2002fue muy intenso en las parcelas de FHIA 18 y FHIA 1,y la situación mejoró hacia la segunda mitad del año(Fig. 6). Un ciclo similar, pero algo más extendido,ocurrió en el 2003 con fuertes afectaciones en el FHIA 18.
Figura 6. Comportamiento de la sigatoka negra en las parcelas de observación ubicadas en plantaciones de La Cuba, Ciego de Ávila.
En la Fig. 7 aparecen las curvas del desarrollo de lasigatoka negra en el FHIA 18 en las diferentes par-celas que permiten comparar mejor el comportamien-to de la enfermedad durante el 2002. Es de hacernotar que las plantas evaluadas en la zona deBaracoa, a pesar de su mayor pluviometría, exhi-
bieron índices notablemente más bajos de la enfer-medad que en La Cuba. Lo mismo ocurre si se com-para la incidencia de la enfermedad en los clonesFHIA 18 y FHIA 23 de La Cuba con los mismosclones de Güira de Melena, de muy alta presión deinóculo (Figs. 2 y 4).
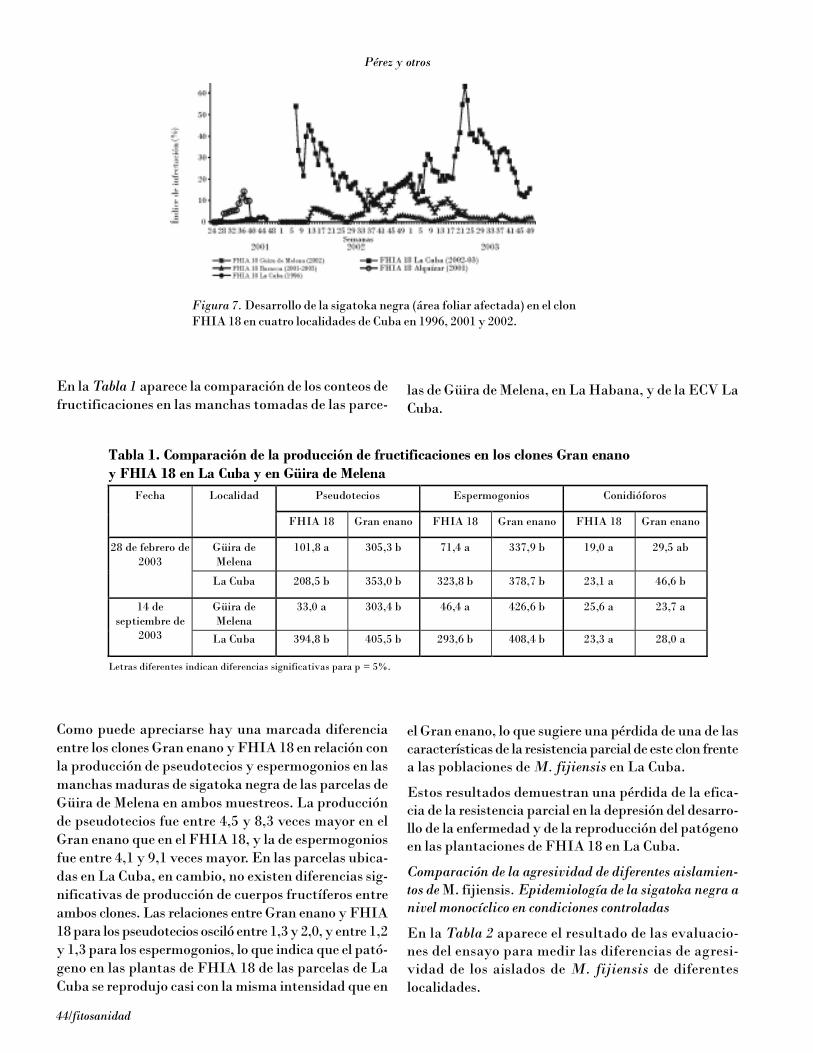
Pérez y otros
44/fitosanidad
Figura 7. Desarrollo de la sigatoka negra (área foliar afectada) en el clonFHIA 18 en cuatro localidades de Cuba en 1996, 2001 y 2002.
En la Tabla 1 aparece la comparación de los conteos defructificaciones en las manchas tomadas de las parce-
las de Güira de Melena, en La Habana, y de la ECV LaCuba.
Tabla 1. Comparación de la producción de fructificaciones en los clones Gran enano y FHIA 18 en La Cuba y en Güira de Melena
Pseudotecios Espermogonios Conidióforos Fecha Localidad
FHIA 18 Gran enano FHIA 18 Gran enano FHIA 18 Gran enano
Güira de Melena
101,8 a 305,3 b 71,4 a 337,9 b 19,0 a 29,5 ab 28 de febrero de 2003
La Cuba 208,5 b 353,0 b 323,8 b 378,7 b 23,1 a 46,6 b
Güira de Melena
33,0 a 303,4 b 46,4 a 426,6 b 25,6 a 23,7 a 14 de septiembre de
2003 La Cuba 394,8 b 405,5 b 293,6 b 408,4 b 23,3 a 28,0 a
Letras diferentes indican diferencias significativas para p = 5%.
Como puede apreciarse hay una marcada diferenciaentre los clones Gran enano y FHIA 18 en relación conla producción de pseudotecios y espermogonios en lasmanchas maduras de sigatoka negra de las parcelas deGüira de Melena en ambos muestreos. La producciónde pseudotecios fue entre 4,5 y 8,3 veces mayor en elGran enano que en el FHIA 18, y la de espermogoniosfue entre 4,1 y 9,1 veces mayor. En las parcelas ubica-das en La Cuba, en cambio, no existen diferencias sig-nificativas de producción de cuerpos fructíferos entreambos clones. Las relaciones entre Gran enano y FHIA18 para los pseudotecios osciló entre 1,3 y 2,0, y entre 1,2y 1,3 para los espermogonios, lo que indica que el pató-geno en las plantas de FHIA 18 de las parcelas de LaCuba se reprodujo casi con la misma intensidad que en
el Gran enano, lo que sugiere una pérdida de una de lascaracterísticas de la resistencia parcial de este clon frentea las poblaciones de M. fijiensis en La Cuba.
Estos resultados demuestran una pérdida de la efica-cia de la resistencia parcial en la depresión del desarro-llo de la enfermedad y de la reproducción del patógenoen las plantaciones de FHIA 18 en La Cuba.
Comparación de la agresividad de diferentes aislamien-tos de M. fijiensis. Epidemiología de la sigatoka negra anivel monocíclico en condiciones controladas
En la Tabla 2 aparece el resultado de las evaluacio-nes del ensayo para medir las diferencias de agresi-vidad de los aislados de M. fijiensis de diferenteslocalidades.

fitosanidad/45
Variabilidad de Mycosphaerella...
Tabla 2. Número de lesiones a los 14 y 20 días en los fragmentos de hojas inoculados con diferentes aislamientos del patógeno
Número de lesiones Aislamientos
A los 14 días A los 20 días
CAF 18-1/Gran enano 60 c 81c
Cm-1/Gran enano 58 c 71 c
CAF 18-1/FHIA 18 31 b 38 b
Cm-1/FHIA 18 13 a 21 a
Letras diferentes indican diferencias significativas según el test de Neumann-Keuls para p = 5%.
Ambos aislados se comportaron de manera similar enel clon Gran enano muy susceptible, pero mostraronuna agresividad diferente sobre los fragmentos de FHIA 18.El aislado CAF 18-1, obtenido de plantaciones de FHIA18 con fuertes afectaciones de sigatoka negra, presentóuna evolución más rápida sobre este clon que el obteni-do de Robusta de un área donde no se han plantadoclones FHIA. La duración del ensayo no permitió esta-blecer diferencias en relación con la producción defructificaciones debido al tiempo en que se mantienenviables los fragmentos de hojas. No obstante, estos re-sultados se corresponden con las evaluaciones de la ca-pacidad reproductiva del patógeno que aparecen en laTabla 1, en el sentido de que la reproducción de los ais-lados de La Cuba no es inhibida de la forma que ocu-rría en las evaluaciones originales de estos clones en esaECV [Pérez, 1998; Hernández y Pérez, 2001], y poneen evidencia cambios en la capacidad patogénica de lapoblación de M. fijiensis en La Cuba.
DISCUSIÓN GENERAL
Los resultados de los estudios de campo demostraronel comportamiento más susceptible frente a sigatokanegra del clon FHIA 18 en La Cuba –donde esta varie-dad se introdujo en pequeñas parcelas en 1994, cuandola empresa cultivaba esencialmente el clon Gran enanosusceptible a la enfermedad–, en relación con el com-portamiento evidentemente resistente de este clon enel resto de las localidades del país. Cuando se introdujoeste clon en el país –por primera vez en esta empresa–mostró una buena resistencia parcial frente a la enfer-medad basada en el alargamiento del ciclo de evoluciónde la enfermedad y en una reducción drástica de la ca-pacidad de producción de pseudotecios y ascósporas[Pérez, 1998; Hernández y Pérez, 2001]. Las compara-ciones de la producción de pseudotecios y espermogonios
en las manchas en hojas del FHIA 18 y en Gran enanoen fincas no tratadas de la empresa La Cuba y de Güi-ra de Melena en La Habana permitieron constatar di-ferencias marcadas entre las dos localidades en rela-ción con la inhibición de la formación de pseudotecios yespermogonios en las manchas, que es parte de la ex-presión del mecanismo de resistencia parcial de estosclones. Por último, las comparaciones de la velocidad eintensidad del desarrollo de la enfermedad bajo condi-ciones controladas en laboratorio, donde se limita lainfluencia ambiental sobre la evolución de la enferme-dad, permitieron detectar diferencias de agresividadentre aislados monoascospóricos procedentes de plan-tas severamente afectadas del clon FHIA 18 de fincasLa Cuba y de aislados procedentes de plantas Cavendishde una región donde no existen clones con resistenciaparcial, y no ha ocurrido selección para agresividad,por lo que puede considerarse una población salvaje enrelación con este carácter.
Los resultados indican por primera vez la selección depoblaciones de M. fijiensis con agresividad sobre elclon FHIA 18 con resistencia parcial en esta localidad,posiblemente por presión de selección direccional[Mather, 1953; Wolfe, 1982] debido a la extensión de lasuperficie cultivada de este genotipo y a las condicio-nes desfavorables de nutrición que han prevalecido enlos últimos años en la mayoría de las áreas de bananosdel país. Pérez et al. (2002) observaron una relaciónentre los niveles de potasio y la severidad de sigatokanegra en fincas de Ciego de Ávila. Se ha informado queel potasio tiene una influencia importante sobre el de-sarrollo de las enfermedades en diferentes cultivos porsu función como regulador móvil de la actividadenzimática y su participación en las funciones celula-res, como son la fotosíntesis, fosforización, síntesis deproteínas, engrosamiento de las paredes celulares y laacumulación de aminoácidos [Huber, 1980].

Pérez y otros
46/fitosanidad
Hasta el presente la selección de poblaciones más agre-sivas de M. fijiensis sobre clones con resistencia par-cial no se había documentado, y debe tenerse en cuentaen los programas de mejora de musáceas actuales. Elpatógeno muestra una importante capacidad de varia-ción y adaptabilidad, como lo ha demostrado la selec-ción de poblaciones resistentes a triazoles (de carácterpoligénico) en Cuba y otros países [Pérez y Batlle, 1993;Chin et al., 1996; Pérez et al., 2003] y a diferentes genesde resistencia de los clones con resistencia parcial de laFHIA desplegados en la empresa La Cuba.
Estos resultados señalan la necesidad de introducir nue-vas fuentes de resistencia a la enfermedad en los progra-mas de mejora convencional y biotecnológica y mejorarlas condiciones de nutrición de los clones que permitanpotenciar los mecanismos de defensa de los clones enexplotación comercial. Deben establecerse estrategias decultivo que disminuyan la presión de selección direccional,rompan el desarrollo poliético del patógeno y disminu-yan el estrés fisiológico para potenciar los mecanismosde defensa de las plantas a la enfermedad. El cultivoanual intensivo puede romper el desarrollo poliético dela enfermedad y disminuir la presión de selección anual.Las áreas bajo cultivo convencional, donde se ha consta-tado la presencia de poblaciones agresivas, deben some-terse a saneamiento intensivo de hojas, mejorar los nive-les de nutrición, ir al cultivo anual (siempre que seaposible), y finalmente adoptar tratamientos confungicidas siempre que las medidas anteriores no garan-ticen los niveles necesarios de control.
CONCLUSIONES
• Los clones FHIA 2, FHIA 18, FHIA 01, FHIA 21,y FHIA 20 mantienen su resistencia parcial asigatoka negra en las zonas de Baracoa y La Habanay en la mayoría de las áreas del país. En las parcelasde La Cuba el clon FHIA 18 mostró durante dosaños consecutivos afectaciones severas de la enfer-medad, muy superiores a las de Baracoa, donde elrégimen de lluvias es superior y más distribuido.
• Las comparaciones de la capacidad del patógeno deformar fructificaciones (pseudotecios, espermogoniosy conidióforos) en manchas de hojas de FHIA 18 yGran enano desarrolladas en la misma semana per-mitió establecer que en las áreas donde el FHIA 18muestra su resistencia parcial es 75-80% menor queen el clon Gran enano, mientras que en áreas dondeestá afectado no existen diferencias significativas
entre ambos clones, lo que pone en evidencia la pér-dida de este carácter.
• Se constató la existencia de diferencias de agresivi-dad entre aislados de M. fijiensis de plantas de FHIA 18de campos severamente afectadas por la enfermedady lo obtenido de plantas Cavendish de zonas libresde clones con resistencia parcial. Las diferencias encampo son sin embargo mayores que las observadasen los ensayos en fragmentos de hojas.
REFERENCIAS
Chin, K.; T. Arroyo; B. Forster; C. Steden: «Sensitivity of Mycosphaerellafijiensis to Demethylation Inhibitors in Central America: TestingMethodology and Cross Resistance», Proceedings of the XII AcorbatMeeting, Santo Domingo, 27 de oct.- 2 de nov., 1996, pp. 413-422.
Firman, I. D.: «Susceptibility of Banana Cultivars to Fungus Diseases inFiji», Trop. Agric. Trinidad, 49:189-196, 1972.
Fouré, E.: «Les Cercosporiose du bananier et leurs traitement.Comportement des variétés. Etude de la sensibilité variétale desbananiers et plantains a Mycosphaerella fijiensis Morelet au Gabon(maladie de raies noires). I. Incubation et évolution de la maladie. II.Etude de quelques paramètres», Fruits 37 (12):749-754, 1982.
––––: «Stratégies de lutte contre le Cercospora noire des bananiers etdes plantains provoquée par Mycosphaerella fijiensis Morelet.L’avertissement biologique au Cameroun. Evaluation des possibilitésd’amélioration», Fruits 43(5):269-274, 1988.
––––: «Leaf Spot Diseases of Bananas and Plantain Caused byMycosphaerella musicola and M. fijiensis», The Improvement andTesting of Musa: A Global Partnership. Proceedings of the First Glo-bal Conference of the International Musa Testing Program, INIBAPHeld at FHIA, Honduras, April 27-30, 1994
Fouré, E.; M. Grisoni; R. Zurfluh: «Les cercosporioses du bananier etleurs traitements. Comportement des variétés. II. Etude de la sensibilitédes bananiers et plantains á Mycosphaerella fijiensis Morelet et desquelques caractéristiques biologiques de la maladie des raies noiresau Gabon», Fruits 39:365-378, 1984.
Fouré, E.; A. Moulioum Pefoura; X. Mourichon: «Etude de la sensibilitévariétale des bananiers et des plantains a Mycosphaerella fijiensisMorelet au Cameroun. Caractérisation de la résistance au champdes bananiers appartenant a divers groupes génétiques», Fruits45:339-345, 1990.
Gauhl, F.: «Epidemiología y ecología de la sigatoka negra. Traducciónde la obra: Unterouchungen zur Epidemiologie und oxologie derSchwanzen Sigatoka krankheit (Mycosphaerella fijiensis Morelet)an kochbananen (Musa sp.) in Costa Rica», Tesis Doctoral presen-tada en el Instituto Sistemático-Geobotánico e Instituto de Patología yProtección de Plantas de la Universidad Georg August GottingenUPEB Ciudad de Panamá, 1990.
Hernández, A.; L. Pérez: «Reaction of Banana and Plantain Cultivars toBlack Sigatoka Caused by Mycosphaerella fijiensis Morelet. EpidemiologicalComponents of the Resistance», Fitosanidad 5 (3):9-16, 2001.
Huber, D.: «The Role of Mineral Nutrition in Defense», Plant Diseasevol. 5, Chapter 21, Academic Press, New York, 1980, pp. 381- 405.
Mather, K.: «The Genetical Structure of Populations», Symposia of theSociety for Experimental Biology, 7, Cambridge University Press,1953, pp. 66-95.
McKinney, H. H.: «Influence of Soil Temperature and Moisture on Infectionof Wheat Seedlings of Helminthosporium sativum», Journal ofAgricultural Research 26:195-218, 1923.

fitosanidad/47
Variabilidad de Mycosphaerella...
Meredith, D. S.; J. S. Lawrence: «Black Leaf Streak of Bananas(Mycosphaerella fijiensis). Susceptibility of Cultivars», Trop. Agric.Trinidad. 47:275-287, 1970.
Mouliom Pefoura, A.: «First Observation of the Breakdown of HighResistance in Yangambi Km 5 (Musa sp.) to the Black Leaf StreakDisease in Cameroon», Plant Disease 83 (1):78, 1999.
Pérez, L.: «Sigatoka negra (Mycosphaerella fijiensis Morelet) debananos y plátanos (Musa spp.) en Cuba. Biología, epidemiología ymanejo integrado de la enfermedad», Simposium Internacional sobreSigatoka Negra, Manzanillo, Colima, México, 8-10 de julio de 1998.
Pérez, L.; J. M. Álvarez; M. Pérez: «Economic Impact and Managementof Black Leaf Streak Disease in Cuba», Mycosphaerella Leaf SpotDiseases of Bananas: Present Status and Outlook. Proceedings ofthe International Workshop on Mycosphaerella Leaf Spot Diseases,San José, Costa Rica, 2002, pp. 71-83.
Pérez, L.; A. Batlle: «Monitoring Procedure to Determine PropiconazoleResistance in Mycosphaerella fijiensis Morelet», Proceedings ofthe 6th International; Congress of Plant Pathology, Montreal, 1993.
Pérez, L.; A. Batlle; A. Hernández; R. Trujillo; C. Álvarez; A. Méndez:«Evolución de la sensibilidad a fungicidas de las poblaciones deMycosphaerella fijiensis Morelet en banano en Cuba», Fitosanidad7(3):49-54. 2003.
Pérez, L.; M. Pérez; M. I. Jiménez; M. Jama: «Ensayo en fragmentos dehojas de bananos y plátanos (Musa spp.) para el estudio a nivel
monocíclico de la evolución de los síntomas de la sigatoka negracausada por Mycosphaerella fijiensis Morelet», Fitosanidad, 2006.
Rowe, P.: «Latest Developments in the FHIA Banana and PlantainBreeding Program: Bred Hybrids Are Now Being Grown Commercially»,Memorias del Seminario Internacional sobre Producción de Plátano,Armenia Quindío, Colombia, 4-8 de mayo de 1998.
Stover, R. H.; J. Dickson: «Leaf Spot of Bananas Caused byMycosphaerella musicola. Methods of Measuring Spotting Prevalenceand Severity», Trop. Agric. Trinidad 47:289-302, 1970.
Stover, R. H.: «A Proposed International Scale for Estimating Intensity ofBanana Leaf Spot (Mycosphaerella musicola Leach)», Trop. Agric.Trinidad 48:185-196, 1971.
Unterstenhöefer, G.: Las bases para los ensayos fitosanitarios decampo, Pflanzenchutz, Nachrichten, Bayer, 16, 167, 1963.
Van der Plank, J. E.: Plant Disease Epidemiology, Academic Press,New York, 1963, pp.180 y181.
––––: Disease Resistance in Plants. Academic Press, New York,1968.
Vidal, A.: «Sigatoka negra en Cuba. En nuevos focos de plagas yenfermedades», Boletín Fitosanitario. FAO 40:(1-2), 1992.
Wolfe, M. S.: «Dynamics of Pathogens Populations in Relation toFungicide Resistance», Fungicide Resistance in Crop Protection,Ed. Pudoc, Center for Agricultural Publishing and Documentation,Wageningen, 1982, pp. 139-148.

48/fitosanidad

fitosanidad/49
FITOSANIDAD vol. 10, no. 1, marzo 2006
EFECTOS DE UNA COBERTURA VIVA DE TERAMNUS LABIALIS(L.F.) SPRENGEL SOBRE LAS ARVENSES EN CAMPOS CITRÍCOLAS
Iván R. Gutiérrez Rojas,1 María de los A. Felipe López,2 Rafael Pérez Carmenate,3 Dayamí Fontes1 yVernon I. Daniel1
1 Universidad de Ciego de Ávila. Carretera a Morón, Km 9½, Ciego de Ávila, Cuba, [email protected] Laboratorio Provincial de Sanidad Vegetal. Carretera Central, Extremo Oeste, Ciego de Ávila, Cuba3 Centro de Investigaciones en Bioalimentos de Ciego de Ávila (CIBA). Carretera a Patria Km 1½, Morón, Ciego de Ávila, Cuba
RESUMEN
Los resultados en la década del noventa del pasado siglo en Ciego deÁvila, Cuba, dirigidos al estudio del uso de leguminosas como co-berturas vivas en campos citrícolas, determinó que se probara laefectividad de la introducción de la especie Teramnus labialis (L.f.)Sprengel var. semilla oscura como una vía para lograr la sostenibilidadde la producción de naranja Valencia Late (Citrus sinensis L. Osbeck),especialmente en cuanto a la conservación del suelo y el desarrolloy producción de la variedad citada, una vez comprobada su utilidaden la mayor parte de los indicadores productivos de esas plantacio-nes. A partir de estos resultados, en la Cooperativa de ProducciónAgropecuaria José Martí, en Ceballos, provincia de Ciego de Ávila,se llevó a cabo un experimento para la determinación de las bonda-des de esa leguminosa nativa de estos suelos, en cuanto a su empleopara manejar las poblaciones de arvenses. De esta manera se redu-jo el nivel de diseminación de todas las especies de arvenses en lasáreas objeto de estudio, en las que se estableció la leguminosacomo cobertura viva, donde Malvastrum coromandelianum (L.), Garkefue la especie que mostró una mayor persistencia ante lasinterferencias de la cobertura utilizada.
Palabras clave: arvenses, leguminosas, Teramnus labialis, coberturas
ABSTRACT
Results about the study of leguminous cover crops in citrus fieldsobtained during the 90´s in Ciego de Ávila, Cuba, determined toevaluate the effectiveness of introducing the cover crop plant Teramnuslabialis (L.f.) Sprengel cv dark seed as a way to obtain a sustainableproduction of Valencia Late orange (Citrus sinensis L. Osbeck),basically focused on both soil conservation and the development andimproved production of that cultivar. The introduced speciesdemonstrated its usefulness in most of productive indicators of thosecitrus plantations. From the results obtained, an experiment wascarried out in order to determine the feasibility of the use of that nativeleguminous cover crop to effectively manage weed populations. Theexperiments were conducted at the Agricultural ProductionCooperative José Martí in Ceballos in the province of Ciego de Ávila.It was achieved a decrease in dissemination of every weed species inthe areas under the cover crop treatment. Malvastrum coromandelianumwas the more persistent species.
Key words: weeds, leguminous, Teramnus labialis, cover crop
INTRODUCCIÓN
A partir de la década del cincuenta y hasta la del no-venta del pasado siglo el uso de coberturas en los cítri-cos fue un tema poco estudiado y debatido que, deacuerdo con Ingels (1994), se ha retomado por los in-vestigadores citrícolas.
No son muy abundantes las referencias en cuanto a laspropuestas sobre el uso de coberturas en frutales yespecíficamente en cítricos. En México se han llevado acabo estudios fundamentalmente dirigidos al controlde arvenses [Osorio, 2000] o agroecológicos [López,1996], así como en Estados Unidos, especialmente enCalifornia y Arizona [O’Connell, 2000; Mc Closkey yWright, 2000; Sakovich y Faber, 1996]. En este sentido
Zaragoza (1997) plantea que el manejo de arvenses puedelograrse con el establecimiento de una cobertura vege-tal, y en este caso específico puede ser a base de plantasespontáneas o introducidas, las que facilitan el paso dela maquinaria con el suelo húmedo, entre otros facto-res, aunque aclara que la competencia en años secos oen zonas semiáridas puede ser drástica. En muchospaíses altamente productores, tal vez las texturas delsuelo y las condiciones climáticas no se adapten siem-pre a los criterios técnicos de algunos especialistascitrícolas que exigen suelo desnudo. El propio Zarago-za (1994, 1997) plantea que el arrope vivo se prefierepara un manejo natural de las líneas o entrecalles, yque generalmente se buscan especies poco competiti-

Gutiérrez y otros
50/fitosanidad
vas, con los frutales sembrados a una densidad sufi-cientemente elevada para ahogar o reducir el crecimientode las arvenses.
En Costa Rica, Argel y Villareal (1998) afirman queArachis pintoi Krap. & Greg. es una leguminosa prefe-rida en plantaciones de café, cítricos, árboles maderablesy otros, y aunque se ha cuestionado en ese país porposibles infecciones de nemátodos, no hay reportes so-bre estudios concluyentes al respecto.
En Cuba se han recogido resultados sobre el uso de co-berturas vivas de leguminosas en cocoteros [Pérez etal., 1996], plátano [Gutiérrez et al., 2002a] y cítricos[Gutiérrez et al., 2002b].
Resulta necesario realizar serios esfuerzos por dominarlas técnicas de manejo de las arvenses en los cultivos, yespecialmente en los cítricos. En cuanto al período críti-co de interferencia, los daños promedio pueden alcanzara reducir los rendimientos hasta en 5 t/ha mensualmen-te cuando se «enyerban» las áreas, lo que provoca afec-taciones irreversibles en las plantaciones más jóvenes.
Casamayor (1999) informó las especies de arvenses degran importancia en un gran listado de las más abun-dantes en los suelos del país, y definió como las másproblemáticas a Ruellia tuberosa L., Struthantusdensiflorus (Benth.) Standl., Panicum maximum Jacq.y Cynodon dactylon (L.), Pers. Esta información resul-ta de extraordinaria importancia para decidir las me-didas oportunas que han de tomarse en cuanto a laposible utilización de coberturas naturales o mejoradascon leguminosas para procurar la sofocación de las es-pecies dañinas.
La principal estrategia en los cítricos para el control delas arvenses siempre ha sido la de utilizar herbicidas delarga residualidad; sin embargo, algunos investigado-res dirigen sus esfuerzos hacia la agricultura ecológica.Tal es el caso de Primavesi (1990), quien recomiendaque en el manejo de especies «invasoras», cuando no esposible disminuir los espaciamientos, se recurra a lascoberturas del suelo, con la sustitución de las hierbasindiscriminadas por la escogida. Así se obtiene un cul-tivo protector que debe ser un ecotipo adecuado paraasegurar la supresión de arvenses nativas.
Los tratamientos con herbicidas son los que imperanen la actualidad para el control de arvenses en cítricos,y en general se desestiman las técnicas agroecológicas,razones por las que se llevó a cabo el siguiente trabajocon el objetivo de comprobar los efectos del uso de una
cobertura de la leguminosa forrajera Teramnus labiales(L.f.) Sprengel var. semilla oscura en el manejo dearvenses en plantaciones de naranjos establecidas ensuelos ferralíticos rojos en la provincia de Ciego de Ávila.
MATERIALES Y MÉTODOS
El experimento se desarrolló en una plantación de na-ranja valencia (Citrus sinensis L. Osbeck) de 16 añosen producción, con un marco de plantación de 8 x 4 m,en un suelo ferralítico rojo (Eutric ferralsol según FAO),perteneciente a la Cooperativa de Producción Agro-pecuaria (CPA) José Martí, en Ceballos, Ciego de Ávila.El trabajo constituyó la fase extensiva de anterioresresultados experimentales relacionados con el uso decoberturas vivas de leguminosas y la utilización de laespecie Teramnus labialis var. semilla oscura, por ha-ber mostrado el mejor comportamiento entre las espe-cies nativas de estos suelos de la zona.
Los resultados de las evaluaciones realizadas se com-pararon con los correspondientes a las mismas callesantes de ser sembradas con la leguminosa, y se utiliza-ron como tratamiento control del experimento, en sucondición de suelo con cobertura natural inicial. Se defi-nieron tres calles para establecer la cobertura propues-ta y cinco plantas en cada calle, para un total de 15plantas evaluables. Se efectuaron 10 muestreos por ca-lle con marcos de 0,25 m2 después de determinar el áreamínima de muestreo. Se empleó el herbicida glyphosate15 días antes de la siembra con dosis de 5 L/ha paralograr el establecimiento de las leguminosas. Se utilizóuna grada ligera de 540,5 kg, con dos pases por cadacalle seleccionada de la plantación objeto de análisis.
Las especies se sembraron a voleo después de la escari-ficación de la semilla y la inoculación con las cepas es-pecíficas de Bradyrhizobium sp. a una densidad de 2 kgde semilla pura germinable por hectárea (SPG/ha). Setomaron muestras, y una vez identificadas las espe-cies, se determinaron las frecuencias relativas (Fr), asícomo las densidades relativas (Dr) de las arvenses enlas áreas objeto de estudio.
El cálculo de la frecuencia relativa [Pérez y Pedroso,1987] se realizó basado en las áreas en las que cada es-pecie se encontró o era dominante, en relación con elárea total evaluada respectivamente según la fórmula:
100(%) xAaFr =
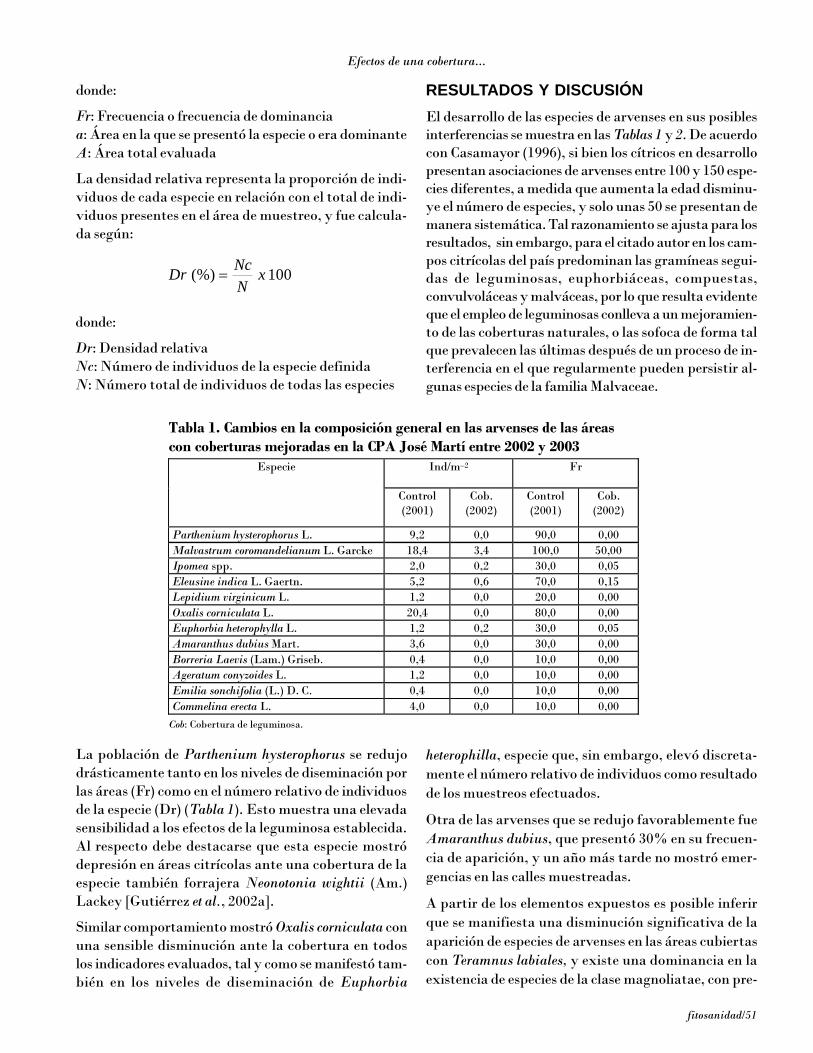
fitosanidad/51
Efectos de una cobertura...
donde:
Fr: Frecuencia o frecuencia de dominanciaa: Área en la que se presentó la especie o era dominanteA: Área total evaluada
La densidad relativa representa la proporción de indi-viduos de cada especie en relación con el total de indi-viduos presentes en el área de muestreo, y fue calcula-da según:
100(%) xNNcDr =
donde:
Dr: Densidad relativaNc: Número de individuos de la especie definidaN: Número total de individuos de todas las especies
RESULTADOS Y DISCUSIÓN
El desarrollo de las especies de arvenses en sus posiblesinterferencias se muestra en las Tablas 1 y 2. De acuerdocon Casamayor (1996), si bien los cítricos en desarrollopresentan asociaciones de arvenses entre 100 y 150 espe-cies diferentes, a medida que aumenta la edad disminu-ye el número de especies, y solo unas 50 se presentan demanera sistemática. Tal razonamiento se ajusta para losresultados, sin embargo, para el citado autor en los cam-pos citrícolas del país predominan las gramíneas segui-das de leguminosas, euphorbiáceas, compuestas,convulvoláceas y malváceas, por lo que resulta evidenteque el empleo de leguminosas conlleva a un mejoramien-to de las coberturas naturales, o las sofoca de forma talque prevalecen las últimas después de un proceso de in-terferencia en el que regularmente pueden persistir al-gunas especies de la familia Malvaceae.
Tabla 1. Cambios en la composición general en las arvenses de las áreas con coberturas mejoradas en la CPA José Martí entre 2002 y 2003
Ind/m–2 Fr Especie
Control (2001)
Cob. (2002)
Control (2001)
Cob. (2002)
Parthenium hysterophorus L. 9,2 0,0 90,0 0,00 Malvastrum coromandelianum L. Garcke 18,4 3,4 100,0 50,00 Ipomea spp. 2,0 0,2 30,0 0,05 Eleusine indica L. Gaertn. 5,2 0,6 70,0 0,15 Lepidium virginicum L. 1,2 0,0 20,0 0,00 Oxalis corniculata L. 20,4 0,0 80,0 0,00 Euphorbia heterophylla L. 1,2 0,2 30,0 0,05 Amaranthus dubius Mart. 3,6 0,0 30,0 0,00 Borreria Laevis (Lam.) Griseb. 0,4 0,0 10,0 0,00 Ageratum conyzoides L. 1,2 0,0 10,0 0,00 Emilia sonchifolia (L.) D. C. 0,4 0,0 10,0 0,00 Commelina erecta L. 4,0 0,0 10,0 0,00
Cob: Cobertura de leguminosa.
La población de Parthenium hysterophorus se redujodrásticamente tanto en los niveles de diseminación porlas áreas (Fr) como en el número relativo de individuosde la especie (Dr) (Tabla 1). Esto muestra una elevadasensibilidad a los efectos de la leguminosa establecida.Al respecto debe destacarse que esta especie mostródepresión en áreas citrícolas ante una cobertura de laespecie también forrajera Neonotonia wightii (Am.)Lackey [Gutiérrez et al., 2002a].
Similar comportamiento mostró Oxalis corniculata conuna sensible disminución ante la cobertura en todoslos indicadores evaluados, tal y como se manifestó tam-bién en los niveles de diseminación de Euphorbia
heterophilla, especie que, sin embargo, elevó discreta-mente el número relativo de individuos como resultadode los muestreos efectuados.
Otra de las arvenses que se redujo favorablemente fueAmaranthus dubius, que presentó 30% en su frecuen-cia de aparición, y un año más tarde no mostró emer-gencias en las calles muestreadas.
A partir de los elementos expuestos es posible inferirque se manifiesta una disminución significativa de laaparición de especies de arvenses en las áreas cubiertascon Teramnus labiales, y existe una dominancia en laexistencia de especies de la clase magnoliatae, con pre-

Gutiérrez y otros
52/fitosanidad
dominio de Malvastrum coromandelianum, la que per-sistió a los niveles de interferencia de la cubierta vege-tal; sin embargo, otras especies que se observaron enlas áreas son, aunque no formaron parte de las mues-tras tomadas:
Para la cobertura natural. Priva lappulacea (L.) Pers.,Solanum nigrum Jacq., Ageratum conyzoides L., Cissuscisyoides L., Emilia sonchifolia (L.) D. C., Rottboelliacochinchinensis Lour. Clayton.
Para la cobertura de leguminosas. Paspalum fimbriatumH. B. K., Cissus cisyoides L., Eleusine indica L. Gaertn,Ageratum conyzoides L., Borreria laevis (Lam) Griseb,Brachiaria fasciculata (Sw.) Blake.
Otro de los resultados de significación es el referido alas especies que muestran persistencia bajo la cobertu-ra de la leguminosa, y que en consecuencia deben sermanejadas por otros medios, de ser imprescindible porefectos negativos relacionados con el desarrollo del cul-tivo o la influencia que puedan ejercer sobre la apari-ción de otras plagas.
Malvastrum coromandelianum, Euphorbia heterophillay Eleusine indica, esta última de las poáceas, mos-traron las mayores proporciones relativas de indivi-duos en las calles, en las que se estableció la legumi-nosa citada como cobertura viva, tal y como se indicaen la Fig. 1.
Figura 1. Especies de arvenses que muestran persistencia a las interferenciasde Teramnus labialis.
Sin embargo, a pesar de no haberse obtenido un con-trol total en todas las especies de arvenses, resulta po-sible afirmar que fue significativa la disminución deindividuos por efecto de la cobertura vegetal del suelo.
CONCLUSIONES
• Todas las especies de arvenses redujeron su nivel dediseminación (Fr) en las áreas objeto de estudio enlas que se estableció la leguminosa como coberturaviva. Solo 33% de las especies mostraron incremen-tos en sus densidades relativas.
• La especie Malvastrum coromandelianum fue la quemostró una mayor persistencia ante las interferenciasde la cobertura utilizada.
• Son favorables los efectos que en general puede ejer-cer la leguminosa nativa de estos suelos Teramnuslabialis sobre la emergencia de las especies dearvenses.
REFERENCIA
Argel, P. J.; M. Villareal: «Nuevo maní forrajero perenne. Arachis pintoiKrapovickas y Gregory. Cultivar Porvenir», Ministerio de Agriculturay Ganadería (MAG) y Centro Internacional de Agricultura Tropical(CIAT), Boletín Técnico, 1998.
Casamayor, R.: «Control de malezas en el cultivo de los cítricos»,Curso Integral de Citricultura, Instituto de Investigaciones de Cítricosy Frutales (IICF), La Habana, 1996, pp. 118-137.
––––: «Comparativo de varias formulaciones de Diuron y Bromacilpara el control de malezas en el cultivo de los cítricos», Memorias delPrimer Encuentro Nacional de Ciencias de Malezas, INSV, INICA, LaHabana, 1999, p. 135.
Gutiérrez, R. I.; G. Pérez; R. Benega; Lourdes Gómez: «Coberturasvivas de leguminosas en el plátano (Musa sp.) FHIA 03», CultivosTropicales, 23(3):11-12, 2002a.
––––: «Influencia de una cobertura de Neonotonia wightii en los cam-bios florísticos de una plantación de naranja Valencia late (Citrussinensis l. Osbeck)», Cultivos Tropicales, 23(3):5-9, 2002b.
Ingels, C.: «Protecting Groundwater Quality in Citrus Production. UCSustainable Agriculture. Research and Education Program», Divisionof Agriculture and Natural Resources, Ed. SAREP, University ofCalifornia, Oakland, California, 1994.

fitosanidad/53
Efectos de una cobertura...
López, S.: «Uso de leguminosas de cobertura en plantaciones de cítri-cos», Memorias del III Simposio Internacional sobre Sistemas de Pro-ducción en Cítricos, vol. 2, Universidad Autónoma de Chapingo, Méxi-co, PIISCI, 1996.
McCloskey, W. B.; G. C. Wright: «Evaluation and Management of aSalina’ Strawberry Clover Cover Crop in Citrus: First Year PreliminaryResults», 2000,
(http://ag.arizona.edu/pubs/crops/az1138/. (con acceso en internet el10 de junio de 2001).
Osorio, R.: «Estudio comparativo de los métodos de control de malezasen cítricos prevalecientes en la cuenca de Tlacolula, Veracruz, Méxi-co». Tesis en opción al título de Ingeniero Agrónomo, UniversidadAutónoma de Chapingo, México, 2000.
O’Connell, N. «Citrus Project Explores Cover Crop Potential», (http://cetulare.ucdavis.edu/update/citrus.htm). In focus, 2000 (con acce-so el 14 de junio de 2001).
Pérez, R.; J. Carrera; A. Borroto; C. Mazorra; A. Ozuna; A. Arencibia; Z.Rodríguez; J. R. García; María del C. Santana: «Establecimiento deleguminosas como cobertura para sistemas mixtos de producciónsostenible en una finca de cocos (Cocus nucifera)», Pastos y Fo-rrajes 3(19):261, 1996.
Pérez, E.; C. Pedroso: «Malezas en cítricos en Cuba», Ciencia y Téc-nica en la Agricultura. Protección de Plantas 10(4):39-64, 1987.
Primavesi, Ana: Agricultura em regioes tropicais, Manejo Ecológico doSolo, Ed. Nobel, Brasil, 1990.
Sakovich, N.; B. Faber: «Grower Comments from Citrus Cover CropField Meeting»,
(http://www.sarep.ucdavis.edu/ccrop/ccres/1996/34.htm) (con acce-so el 20 de junio de 2001).
Zaragoza, C.: «Las coberturas del suelo», Rev. Horto Fruticultura,(5):60-62, 1994.
––––: «El laboreo de conservación en el viñedo», Fruticultura Profe-sional (91):6-13, 1997.

54/fitosanidad

fitosanidad/55
FITOSANIDAD vol. 10, no. 1, marzo 2006
MANEJO CONVENCIONAL Y ALTERNATIVO DE LA SIGATOKANEGRA EN BANANOS: ESTADO ACTUAL Y PERSPECTIVAS
Luis Pérez Vicente
Instituto de Investigaciones de Sanidad Vegetal. Calle 110 no. 514 e/ 5a. B y 5a. F, Playa, Ciudad deLa Habana, CP 11600, [email protected]
RESUMEN
La lucha contra Mycosphaerella fijiensis en bananos y plátanos se hadesarrollado dentro de sistemas de manejo que integran el sanea-miento, medidas culturales que tienden a crear condiciones másdesfavorables al desarrollo de la enfermedad y al control químico. Serevisa el estado del conocimiento de los mecanismos de acciónbioquímico de los diferentes grupos de ingredientes activos, su efec-to sobre el desarrollo del patógeno y los problemas de emergenciade poblaciones con sensibilidad reducida. Para el control deMycosphaerella fijiensis se han utilizado fungicidas de contacto comolos alquilenbisdithiocarbamatos y el clorotalonil, y sistémicosinhibidores de la mitosis como los benzimidazoles, inhibidores de labiosíntesis de ergosterol (principalmente triazoles y morfolinas),inhibidores del transporte de electrones en las mitocondrias QoI’s(estrobilurinas y oximino acetatos), inhibidores de la producción deenzimas en el patógeno (anilinopyrimidinas) e inductores de resis-tencia (ASM), los que en general se han utilizado en emulsión deaceite y agua o directamente en aceite. El patógeno muestra una altacapacidad de variación y adaptación, desarrollo de poblaciones com-petitivas con las cepas salvajes, con baja sensibilidad a ben-zimidazoles, triazoles y methoxyacrilatos en diferentes países. Lacreciente preocupación de los consumidores sobre los residuos deagroquímicos en frutos, así como la emergencia de poblaciones consensibilidad reducida y los costos crecientes de la protección contrasigatoka negra, han compulsado a la búsqueda de nuevas alternati-vas al manejo, entre las que se encuentran el uso de clones conresistencia parcial mediante mejora convencional e ingenieríagenética, el estudio del efecto secundario de agroquímicos para lanutrición, así como el uso de compostaje y lixiviados de humus, loscuales muestran eficacia en diferentes sistemas de producción parael control de la enfermedad. La lucha biológica con antagonistasbacterianos no brinda resultados de interés práctico para el manejode clones susceptibles en el trópico húmedo. Finalmente las alterna-tivas del manejo son distintas para los diferentes escenariosclimáticos epidemiológicos y sistemas de producción.
Palabras clave: Mycosphaerella fijiensis, bananos, plátanos
ABSTRACT
The management of Mycosphaerella fijiensis in banana and plantainshas been carried out integrating sanitation measures, culturalpractices conducing to create unfavorable conditions to diseasedevelopment and the chemical control. The stage of the currentknowledge is reviewed on the biochemical action mechanism ofdifferent active ingredient groups, its effects on pathogen developmentand the problem of emergence of populations with reduced sensibilityto them. For Mycosphaerella fijiensis control, contact fungicides asbisdithiocarbamates and chlorotalonil, and systemics inhibidors ofmitosis as the benzimidazole, has been used; inhibitors of ergosterolbiosíntesis (mainly triazole and morpholines); inhibitors of electrontransport in the mitocondria QoI’s (strobilurines and oximinoacetates); inhibitors of production of enzymes in the pathogen(anilinopyrimidines) and inductors of resistance (ASM), which hasbeen in general used in plain oil or as oil in water emulsions. Thepathogen shows a high capacity of variation and adaptation and hasdeveloped low sensitive competitive populations with the wild typepopulations to benzimidazole, triazole and methoxyacrilates indifferent countries. The raising concern of consumers on theagrochemical residues in fruits, as well as the emergence of resistantpopulations of the fungus and the rising costs of protection againstBlack Sigatoka has compelled the search of new alternatives ofmanagement, among them, the use of cultivars with partial resistancedeveloped by conventional breeding and genetic engineering; thestudy of secondary effect of fertilizers as well as the use of compostand lixiviates of humus which has been effective in different systemsof banana production. Biological control by use of antagonists doesnot have brought practical results for disease control in susceptiblecultivars particularly in the humid tropics. Finally, the alternative ofmanagement are different for the different climatic –scenario andproduction systems.
Key words: Mycosphaerella fijiensis, bananas, plantains
INTRODUCCIÓN
Los bananos y plátanos no pueden producirse co-mercialmente sin control de las manchas de las hojas porMycosphaerella spp. Mycosphaerella fijiensis Morelet,agente causal de la raya negra o sigatoka negra, es sinduda, uno de los patógenos que mayor impacto económi-co, social y ambiental ha tenido en la historia de la agri-
cultura, y particularmente en la producción mundial debananos y plátanos. La enfermedad puede reducir entreel 25 y el 90% los rendimientos [Fouré, 1985; Stover, 1983].Los costos de manejo de la enfermedad en plantacionescomerciales oscilan usualmente entre 15 y 25% de todoslos costos de producción [Stover, 1980, Pérez et al., 2002b].
Res
eña

Luis Pérez Vicente
56/fitosanidad
El manejo de la enfermedad se basa en la imple-mentación de medidas culturales –drenajes para redu-cir la humedad interna de la plantación, una adecuadadensidad de plantas, saneamiento de hojas enfermas,nutrición equilibrada para garantizar una rápida sali-da de hojas y potenciar las defensas naturales de laplanta, y un adecuado sistema de cosecha–, el uso declones con resistencia parcial, donde estos son acepta-dos, y el empleo de agroquímicos y compuestos de ori-gen natural para el control [Pérez, 1997; 1998; Romero,2000]. La implementación de programas de manejo efi-cientes requiere disponer además de tres tipos de in-formaciones básicas: 1) climáticas: particularmentecantidad y frecuencia de la lluvia y duración de la hu-mectación de las hojas, que permite valorar la evolu-ción epidemiológica futura [Gauhl, 1990; Pérez, 1998;Pérez et al., 2000a,b]; 2) biológicas: velocidad o ritmode salidas de hojas, hojas más jóvenes con síntomas ymanchas necróticas; severidad en la floración y a las7-9 semanas de paridas [Stover y Dickson, 1970; Robertet al., 1982]; velocidad de la evolución de la enfermedad[Fouré, 1988c]; 3) sensibilidad de las poblaciones a losprincipales fungicidas utilizados.
El manejo químico de sigatoka negra
El uso de fungicidas para la protección contra la enfer-medad recibe una atención importante, porque en áreascon una adecuada pluviometría para la producciónbananera de clones susceptibles las medidas no quími-cas no permiten alcanzar un control satisfactorio de laenfermedad. El 41,6% de los más de setecientos seten-ta artículos publicados hasta 1990 en relación conMycosphaerella musicola y M. fijiensis se referían almanejo químico de ambas enfermedades [Barrantes,1990]. Se han realizado además revisiones sobre estetópico [Meredith, 1970; Stover, 1972; 1990; Romero,2000].
El manejo químico de la sigatoka negra se ha llevado acabo con el uso de fungicidas protectores y sistémicosen suspensión acuosa, en emulsiones de aceite y agua, oen mezcla directamente con aceite mineral solo, conactivadores de mecanismos de resistencia del hos-pedante, y últimamente mediante el uso de compues-tos relacionados con la nutrición, tanto de origen quí-mico como naturales, con efecto sobre la fisiología de laplanta y el desarrollo de la enfermedad.
Fungicidas protectores. Al no penetrar al interior de lostejidos de las hojas, su actividad sobre el patógeno esprincipalmente por inhibición de la germinación y el
crecimiento de los tubos de las esporas que se deposi-tan sobre la superficie tratada antes de la penetraciónpor los estomas, así como la formación de conidios alcontaminar los conidióforos durante el tratamiento delas hojas o mediante la redistribución del ingredienteactivo que ocurre en el agua del rocío o la lluvia que sedeposita en la lámina foliar.
Fungicidas cúpricos. Entre 1930 y 1957 Mycosphaerellamusicola estuvo sometida a control mediante aspersio-nes a alto volumen de mezcla bordeleza en muchos paí-ses [Meredith, 1970]. Estas aspersiones se realizaron conla utilización de aspersores portátiles, máquinas de as-persión y por sistemas de tuberías permanentes alimen-tadas por estaciones centrales de bombeo. Se asperjaronvolúmenes de hasta 2 500 L.ha–1 a intervalos de 2-5 se-manas, de acuerdo con las condiciones locales.
Alquilenbisdithiocarbamatos. Se han utilizado amplia-mente Maneb y Mancozeb para el control de las man-chas de hojas tipo sigatoka [Guyot y Cuillé, 1954a,b;Cuillé y Guyot, 1956; Meredith, 1970; Stover, 1972;Pérez, 1978a]. En general reaccionan de formainespecífica con grupos tiol (SH) que causan bloqueode la respiración y de otros procesos metabólicos don-de median proteínas con estos grupos [Kaars Sijpesteijnet al., 1977]. En la actualidad existen diferentesformulaciones de mancozeb que permiten realizar tra-tamientos en emulsiones de aceite y agua, o directa-mente en aceite, solos o como parte de cocteles confungicidas sistémicos para reducir la presión de selec-ción de poblaciones tolerantes a los fungicidas de altoriesgo de selección de poblaciones resistentes en los pro-gramas de tratamientos contra sigatoka negra. Se hautilizado en aspersiones foliares a dosis de 1,5-2,0 kgdel ingrediente activo.ha–1. Inhiben la germinación delas esporas del patógeno que entran en contacto con lasuperficie de la hoja tratada o que se contaminan du-rante su desarrollo debido a la redistribución del ingre-diente activo por el agua libre disponible en la superfi-cie de las hojas.
Derivados de phtalimidas (clorotalonil). El clorotalonilpresenta una baja solubilidad en agua. Su mecanismode acción está relacionado con la reacción del glutatión,coenzima A, 2-mercaptoetanol y otros compuestos queforman derivados del S [Vincent y Sisler, 1968; KaarsSijpesteijn et al., 1977]. Por esto, el contenido de SH dela célula se reduce significativamente, lo que resulta eninhibición de todas las reacciones dependientes de losgrupos tiol. El glutatión es un componente importante

fitosanidad/57
Manejo convencional y alternativo...
del metabolismo celular y se afecta rápidamente por elclorotalonil, por lo que su acción es inespecífica ymultisitio. Existe la formulación Bravo 500 SC que pre-senta una fuerte resistencia al lavado por la lluvia y elrocío [Washington et al., 1998b], y en aspersiones a in-tervalos semanales inhibe la germinación de los conidiosy su formación sobre las manchas [Washington et al.,1998a,b]. La aparición de poblaciones con sensibilidadreducida a los benzimidazoles y triazoles en los últimosaños ha determinado un incremento del uso declorotalonil a dosis de 1-2 kg.ha–1, en aspersiones acuo-sas sin aceite, porque la mezcla con aceite causa dañosa las hojas. Se debe tener cuidado con los tratamientosde clorotalonil a hojas que presentan depósitos ante-riores de aceite y viceversa.
Fungicidas sistémicos. El uso de ingredientes activosque penetran las barreras estructurales naturales de laplanta y se incorporan al apoplasto, donde sontraslocados hacia los puntos de mayor transpiración,permitió una mejora importante del control de M. fi-jiensis debido a una distribución y protección de lasuperficie tratada de la hoja más completa, y en proce-so de expansión a la posibilidad de acceso a estructurasfúngicas que se desarrollan en el interior de los tejidos,y a una mayor resistencia a los factores ambientales.La absorción y la traslocación de los ingredientes acti-vos sistémicos, y por tanto sus propiedades fungicidas,dependen en gran medida de su lipofilia y solubilidaden agua. La lipofilia permite la penetración a nivel delas capas cerosas y la cutícula de la hoja; la traslocaciónsistémica está determinada por la solubilidad en agua.Mientras más alta es la lipofilia más reducida es latraslocación en el interior de la planta [Edginton yPetersen, 1977; Edginton, 1981; Pontsen y Wissfeld,1996]. Como medida de la lipofilia se utiliza el coefi-ciente de partición octanol/agua (Pow) o el log Pow, el cualmuestra una adecuada correlación con la traslocaciónde diferentes fungicidas. La eficacia y la estrategia deuso de los fungicidas sistémicos en los programas decontrol de sigatoka negra se determina no solo por laactividad tóxica intrínseca del ingrediente activo sobreel patógeno, sino también por las propiedades físico-químicas, y por tanto de traslocación de la molécula enlas hojas de la planta.
En la Tabla 1 aparecen los valores de log Pow de diferen-tes fungicidas utilizados en bananos. Entre las princi-pales familias químicas de fungicidas utilizados en elcontrol de sigatoka negra en bananos se encuentran losbenzimidazoles y tiofanatos, aparecidos a principios de
la década del setenta del pasado siglo; los triazoles eimidazoles, las morfolinas, utilizados en bananos amediados de la década del ochenta; las estrobilurinas ymethoxyacrilatos aparecidas comercialmente en 1998;la spiroketalamina (spiroxamina) y las anilinopy-rimidina (pyrimethanil, informado en 1988) con uso anivel comercial después del 2000 para el control desigatoka negra en bananos.
Derivados de benzimidazol. Se utilizaron desde iniciosde la década del setenta en el control de sigatoka negra.En este grupo se incluyen el benomyl, thiabendazol,carbendazim (metil benzimidazol carbamato o MBC) ymetil tiofanato, entre otros. Este último no es unbenzimidazol hasta que ocurre su metabolización en lacélula del hongo [Delp, 1995]. El benomyl se ha usadoampliamente a la dosis de 140-150 g.ha–1, en emulsiónde aceite agua o solo en aceite. Presenta una excelenteactividad sistémica y terapéutica, y muestra un fuerteefecto sobre la formación de fructificaciones en las man-chas. Todos, en dilución acuosa, forman por diferentesvías el compuesto MBC o carbendazim (agentefungitóxico), que bloquea la formación de microtúbulos,al unirse selectivamente con las β-tubulinas fúngicas(no así en mamíferos), e impiden el acoplamiento de losdímeros de α y β-tubulinas, y por tanto la mitosis ce-lular [Davidse, 1981]. Las ascosporas de poblacionessensibles de M. fijiensis descargadas en agar envene-nado con concentraciones letales de benzimidazoles pre-sentan inhibición total de la germinación o tubosgerminativos deformados. Los cambios de un soloaminoácido en la β-tubulina determina la pérdida desensibilidad al ingrediente activo en diferentes especiesde hongos (Tabla 2). Las mutaciones ocurridas en cam-po han sido restrictas a los codones 198 y 200, las cua-les aparentemente no tienen implicaciones en la habili-dad competitiva de las líneas [Davidse e Ishii, 1995).
Para el monitoreo de los cambios de la sensibilidad delas poblaciones usualmente se determina la germinaciónnormal o anormal de las esporas en medio de cultivo,con una concentración umbral de 5-10 µg.mL–1 debenomyl. Según recomendaciones del FungicideResistance Action Committee (FRAC) del BananaWorking Group, se deben considerar resistentes laspoblaciones que tienen >10% de germinación normal a10 µg.mL–1. Se ha desarrollado una técnica para el diag-nóstico de la resistencia al carbendazim mediante lareacción en cadena de la polimerasa (PCR) [Martin etal., 1992]. Cañas et al. (2004) amplificaron un fragmen-to de 527pb de cepas de M. fijiensis resistentes al

Luis Pérez Vicente
58/fitosanidad
benomyl, con los cebadores Resistencia 1 reverse y Diag-nóstico forward, que muestran un cambio en el codón 198de una citosina por una adenina, lo cual se corresponde conun cambio de un ácido glutámico por una alanina a niveldel de la β-tubulina, así como un fragmento de 635 pb de
cepas sensibles con los cebadores Sensible 2 forward yB2Ali. Estos dos cebadores permiten diferenciar de formaconfiable, mediante una PCR, las cepas del patógeno re-sistentes y sensibles al benomyl, por medio de la utiliza-ción de la enzima Bsh12361 (FunDII; MBI Fermentas).
Tabla 1. Valores de los coeficientes de partición octanol/agua (como log Pow) de diferentes ingredientes activossistémicos utilizados en el control de M. fijiensis enbananos y plátanos (valores de log Pow tomados de Tomlin(2003))
Fungicidas estudiados para control de Mycosphaerella spp. en banano
Log Pow Benomyl 1,4 Carbendazim 1,5 Tiofanato metilo 1,5 Azoxystrobin 2,5 Spiroketalamina 2,8 Pyrimethanil 2,8 Cyproconazol 2,9 Tetraconazol 3,1 Triadimefon 3,1 Fenbuconazol 3,2 Triadimenol 3,3 Epoxyconazol 3,4 Tebuconazol 3,7 Propiconazol 3,7 Flusilazol 3,7 Imazalil 3,8 Hexaconazol 3,9 Pyraclostrobin 3,9 Bitertanol 4,1 Tridemorph 4,2 Difenoconazol 4,3 Trifloxystrobin 4,5
Alta Baja
La pérdida de sensibilidad es un proceso gobernado porun solo par de genes, y los riesgos de selección de pobla-ciones tolerantes competitivas con las poblaciones sal-vajes son elevados [Wolfe, 1981, Georgopoulos, 1985].La selección de poblaciones de M. musicola y M. fi-jiensis con resistencia a benzimidazoles (seleccióndisruptiva) ha sido ampliamente informada en casi to-das las áreas productoras de bananos y plátanos [Stover,1977; Pérez et al., 1985; Jiménez y Rodríguez, 1985;Romero y Sutton, 1998; Marín et al., 2003]. Las pobla-ciones de M. musicola y M. fijiensis resistentes albenomyl son frecuentemente insensibles a altas dosisdel ingrediente activo y competitivas con las poblacio-nes salvajes. Puede presentarse una alta frecuencia deindividuos resistentes en la población aun después deno utilizar el ingrediente activo por muchos años [Pérez,inédito; Marín et al., 2003].
En la actualidad el FRAC recomienda para las áreasen que aún puede utilizarse no realizar más de tres tra-tamientos por año [Kuck et al., 2004].
Inhibidores de la síntesis de ergosterol (EBI). Los EBIson de dos tipos: 1) inhibidores de C14 a-demetilasa oDMI [Sisler et al., 1984; Köller y Scheinflug, 1987; Sisler,1996]. En bananos y plátanos se utilizan esencialmentetriazoles e imidazoles; 2) aminas cíclicas (inhibidoresde D8-D7 isomerasa, C14 reductasa y escualenoepoxidasa). En bananos y plátanos se utilizanmorfolinas y spiroketalaminas.
Inhibidores de demetilasa (DMI). Los DMI utilizadospara el control de Mycosphaerella spp. en Musa spp.son esencialmente triazoles. El primero y más amplia-mente utilizado de los triazoles es el propiconazol, quese introdujo en Cuba para el control de sigatoka en

fitosanidad/59
Manejo convencional y alternativo...
1982 [Pérez et al., 1983] debido a los frecuentes problemasde resistencia al benomyl y comercializado para su uso enbananos en América Central a partir de 1987 [Carlier etal., 2000]. Al inicio de su uso (a 100 g.ha–1 tanto en aceitecomo en emulsiones de aceite y agua) mostró una altaeficacia y una reducción apreciable del número de trata-mientos requeridos para obtener elevados niveles de con-trol de la enfermedad, una significativa reducción del ta-maño de las manchas en las hojas tratadas y de lafrecuencia e intensidad de la formación de fructificaciones[Pérez et. al., 1983; Pérez y Mauri, 1994]. Otros triazolesutilizados son el flusilazol, fenbuconazol, tebuconazol,hexaconazol (utilizados a 100 g.ha–1, bitertanol a 150 g.ha–1),epoxiconazol, difenoconazol y cyproconazol (80 g.ha–1), quemuestran diferencias de eficacia entre sí y del nivel de latraslocación sistémica en las hojas, demostrada por losdiferentes patrones de daños de la enfermedad [Fouré,1983; 1988a, 1988c, 1988d, 1990; Pérez et al., 1993a; Pérez,1998]. Los de mayor actividad sistémica se mueven rápi-damente hacia el borde de las hojas, así permiten dosissubletales alrededor del nervio central de la hoja y la apa-rición de un patrón típico de lesiones a ambos lados. Elfenbuconazol y el hexaconazol presentan una menortraslocación en las hojas que el propiconazol y elcypendazol, mientras el bitertanol y el difenoconazol sonesencialmente translaminares y no se observan estos pa-trones de daños alrededor de la nervadura central.
Todos inhiben la síntesis de novo de ergosterol en lacélulas fungosas al bloquear la citocromo P450 esterolC14 demetilasa [Sisler et al., 1984; Köller y Scheinflug,1987; Sisler, 1996]. Las esporas depositadas en un me-dio envenenado con propiconazol u otro IBE germinany se desarrollan a partir del ergosterol constitutivo hastasu agotamiento. En una población de M. fijiensis sepueden encontrar diferentes niveles de inhibición delcrecimiento de los tubos germinativos de ascósporas yconidios a una concentración dada del fungicida; por lotanto, la determinación de la pérdida de sensibilidad esmás complicada en los IBE que en el caso de losbenzimidazoles. En la Fig. 1A se observa la distribu-ción de frecuencias del crecimiento de los tubosgerminativos de ascósporas de M. fijiensis en PDAenvenenado con 0,01 µg.mL–1 de propiconazol. En ellase aprecian las diferencias de sensibilidad de las espo-ras en una población del patógeno. En la Fig. 1B semuestran las distribuciones de frecuencias de la inhibi-ción del crecimiento de los tubos germinativos de dosfincas, una no tratada y otra tratada con triazoles enun medio envenenado con 0,01 µg.mL–1 de tebuconazol,
donde se pueden ver las diferencias de la sensibilidadde las poblaciones de M. fijiensis de ambas [Pérez yBattle, 1993b; Pérez et al., 2000c, 2003]. En la fincatratada las mayores frecuencias de esporas se encuen-tran entre los valores más bajos de inhibición.
Se han sugerido diferentes mecanismos de resistenciapara los DMI [Buchenauer, 1995; Sisler, 1996]: pérdidade afinidad en el citocromo P45014DM, defectos en elcitocromo P45014DM, detoxificación, sobreproducción delcitocromo P450, falta de activación, la expulsión activadel ingrediente activo de la célula, protonación,circunvención al sitio de acción, tolerancia a esterolestóxicos, entre otros. La evidencia fragmentaria de mu-chos estudios ha permitido establecer, por tanto, queoperan diferentes mecanismos de resistencia, y que engeneral el proceso de pérdida de sensibilidad a los DMIes poligénico [Van Tuyl, 1977; Kalamarakis et al., 1991],por lo que ocurre gradualmente y bajo un proceso deselección direccional [Wolfe, 1981; Georgopoulos, 1985].Los cambios resultantes de la resistencia poligénica sonmenos factibles de conducir a pérdidas totales de con-trol, y el gran número de mutaciones requeridas parauna alta resistencia conducen a una alta probabilidad deque los individuos resultantes presenten una capacidadcompetitiva reducida [Buchenauer, 1995; Sisler, 1996].
La aparición de poblaciones tolerantes de M. fijiensisa benzimidazoles estableció una mayor dependencia enel uso de triazoles, y por tanto en el número de ciclospor año en la mayoría de las áreas de cultivo. Esto de-terminó cambios graduales en la composición de laspoblaciones y la aparición de fenotipos del patógenocon diferentes niveles de sensibilidad. Los primeros sín-tomas en Cuba de la pérdida de sensibilidad alpropiconazol fue la pérdida del efecto depresivo inme-diato de los tratamientos sobre la velocidad de evolu-ción de la enfermedad y de la reducción del tamaño delas lesiones junto a su inhibición de la formación defructificaciones del patógeno. En los tratamientos conaviso bioclimático, la duración del período entre dosciclos se acortó de 21-24 días hasta 14-12 días [Pérez etal., 2000c]. El monitoreo de la sensibilidad se desarro-lla por la determinación de los cambios en el tiempo delas frecuencias de ascósporas, obtenidas de muestrasde hojas de fincas tratadas y no tratadas, con diferen-tes niveles de inhibición del crecimiento de los tubosgerminativos, en PDA envenenado con concentracionesde 0; 0,001; 0,01; 0,1 y 1 µg.mL–1y de las DE50 de laspoblaciones [FRAC, 1996]. Todos los DMI presentanresistencia cruzada entre ellos [Chin et al., 1996], aun-que los factores de resistencia entre estos sean muy

Luis Pérez Vicente
60/fitosanidad
variables [Köller y Wubben, 1989]. La presencia de pér-dida de sensibilidad a los triazoles se ha evidenciado anivel mundial [Marín et al., 2003]. Una variación signi-ficativa ocurrió en Costa Rica, Panamá y el norte deGuatemala [Kuck et al., 2004], principalmente en fin-cas situadas en zonas de muy alta pluviometría, dondese requieren más ciclos [Guzmán et al., 2004]. En elresto de áreas de Centroamérica, México, Colombia,
Ecuador Camerún y Filipinas han ocurrido pérdida desensibilidad más leves. La integración del uso de lostriazoles en mezclas con tridemorph y mancozeb, y eluso de altas dosis de aceite junto a la adopción de me-didas estrictas de saneamiento y drenaje de los cam-pos, ha permitido que los triazoles se mantengan comola principal herramienta de control [Marín et al., 2003;Guzmán et al., 2004].
Tabla 2. Sustituciones de aminoácidos en β-tubulinas de mutantes de laboratorio y aislados de campo con resistencia a benzimidazoles según revisión de Davidse y Ichii (30) modificada
Codón Sustitución Organismo
His por Leu A. nidulans (a)1 6
His por Tir A. nidulans (a) y S. nodorum (b)
Tir por Asn A. nidulans (c) 50
Tir por Ser A. nidulans (c)
134 Gln por Lis A. nidulans (c)
165 Ala por Val A. nidulans (d)
167 Fen por Tir N. crassa (e)
Glu por Ala B. cinerea (f, g), N. crassa (b); P. aurantiogriseum (i); P. expansum (c); P. puberulum (i); V. inaequalis (c)
Glu por Asp A. nidulans (a)
Glu por Gln A. nidulans (a)
Glu por Gly N. crassa (b); R. secalis (i); V. inaequalis (i)
Glu por Lis A. nidulans (a); B. cinerea (g); M. fructicola (i); P. digitatum (c); P. italicum (c); R. secalis (i)
Glu por Val P. expansum (c)
198
Cit por Ade M. fijiensis2
200 Ala por Tir B. cinerea (g); P. aurantiogriseum (i); P. italicum (i)
241 Arg por His S. saccharomyces (j)
257 Met por Leu A. nidulans (c)
Datos citados por Davidse y Ichii, (1995) tomados de: 1a) Jung et al., 1992; b) Cooley y Caten, 1993; c) Koenraadt et al., 1992; d) Jung and Oakley, 1990; e) Orbach et al., 1986; f) Martin et al., 1992; g) Yarden y Katan, 1993; h) Fujimura et al., 1992; i) Wheeler et al., 1994; j) Thomas et al., 1985; 2: Cañas et al., 2004.
Figura 1. Frecuencias de inhibición del crecimiento de los tubos germinativos de ascósporas de M. fijiensis. A) De unafinca tratada –con poblaciones de M. fijiensis sensibles– en medios envenenados con diferentes concentraciones depropiconazol; B) de dos fincas, una no tratada nunca y otra tratada en un medio envenenado con 1 µg/mL–1 de tebuconazol.

fitosanidad/61
Manejo convencional y alternativo...
Aminas cíclicas. En bananos y plátanos se utilizanmorfolinas y spiroketalaminas.
Morfolinas. En control de sigatoka negra se ha utiliza-do esencialmente el tridemorph (450 g.ha–1) enemulsiones de aceite y agua y en aceite solo. La dura-ción de su eficacia para el control de la enfermedad esmás corta que la de los benzimidazoles y triazoles con-tra poblaciones sensibles a estos ingredientes activos,en tratamientos mediante aviso bioclimático; muestraexcelente eficacia en tratamientos mezclados contriazoles al 75% de las dosis de ambos ingredientes ac-tivos [Pérez, 1993a]. El tridemorph inhibe la ∆8-∆7isomerasa y la C14 reductasa [Leroux y Gredt, 1983;Kerkenaar, 1995] en la ruta metabólica de la síntesis deergosterol –presenta un riesgo reducido de selección depoblaciones resistentes–, por lo que afecta seriamentela funcionabilidad de las membranas celulares fúngicas.Su traslocación es limitada en musáceas y es esencial-mente translaminar [Cronshaw y Akers, 1990]. Con másde veinticinco años de uso en cereales no hay casos es-tablecidos de selección de poblaciones resistentes[Hollomon, 1994]. Tampoco hay casos documentadosde poblaciones resistentes a este ingrediente activo enbananeras de exportación. Es un importante compo-nente en las estrategias antirresistencia a benzi-midazoles, DMI y QoI. Las recomendaciones de FRAC[Kuck et al., 2004] establecen no realizar más de docetratamientos por año de morfolinas, ya sean solas o enmezclas. En Cuba se han estudiado además elfenpropimorph y el aldimorph, los que mostraron unaeficacia comparable al tridemorph.
Spiroketalaminas. La spiroxamina, descubierta en 1987e informada por primera vez en 1996 [Dutzmann et al.,1996], después del 2000 se comercializa para el controlde sigatoka negra en bananos en Centroamérica. Laabsorción por las hojas es rápida y su traslocaciónacropetal sistémica, y por tanto presenta una buenadistribución en las hojas [Tomlin, 2002]. Su eficacia ala dosis de 240 g.ha–1 es similar a la del tridemorph[Arias et al., 2002]. La spiroxamina inhibe de formaprincipal la C14 reductasa además de la ∆8-∆7 isomerasay la escualeno epoxidasa [Tiemann et al., 1997], por loque el riesgo de selección de poblaciones resistentes esmuy reducido. El FRAC tiene establecida una reco-mendación de uso para el control de sigatoka negra si-milar a la del tridemorph [Kuck et al., 2004].
Inhibidores del transporte de electrones en el sitio QoI.Los compuestos más utilizados en bananos en
emulsiones de aceite y agua son el azoxystrobin[Godwin et al., 1992] usado a la dosis de 100 g.ha–1 eintroducido comercialmente en 1996, y el trifloxystrobin[Margot et al., 1998], usado a dosis de 75 g.ha–1 [Pérezet al., 2000c y 2002a), los que muestran una excelenteeficacia en el control de sigatoka negra. Se introdujopara el control de sigatoka en Centroamérica en el 2002el pyraclostrobin [Ammermann et al., 2000], que hamostrado una eficacia similar a la del azoxystrobin a ladosis de 100 g.ha–1 [Farías-Larios y Orozco-Santos,2002], y el picoxystrobin a la de 250 g.ha–1 [Godwin et al.,2000]. Los derivados de estrobilurinas mantienen sumejor eficacia en tratamientos tempranos previos a lainfección. Inhiben la germinación de las esporas y elcrecimiento de las hifas, y presentan por demás unabuena eficacia sobre la formación de fructificaciones enlas manchas tratadas. Al introducir el azoxystrobin serecomendaron entre seis y ocho tratamientos por año.Poco tiempo después de su entrada al mercado se in-formaron los primeros casos de resistencia de M. fi-jiensis en bananos [Wirtz y Chin, 2000; Chin et al.;2001]. En la actualidad se han documentado casos deresistencia del patógeno a este grupo de fungicidas envarios países exportadores [Guzmán et al., 2004].
El modo de acción se basa en el bloqueo de la transfe-rencia de electrones a nivel del sitio o centro externodel citocromo bc1 de la membrana externa de lasmitocondrias, se inhibe la respiración y por tanto lasíntesis de ATP [Sauter et al., 1999]. Aunque se hanreconocido once posibles puntos de mutaciones en dosregiones del citocromo b [Geier et al., 1992, 1993], en lamayoría de los casos de pérdida de sensibilidad defitopatógenos fungosos a los QoI, esta se ha determi-nado por una mutación puntual en el gen cyt b del genmitocondrial que da lugar a un cambio de una glicina auna alanina en posición 143 (G143A) en el citocromobc1 en la mitocondria [Sierotsky et al. 2000a,b; Heaneyet al., 2000], la que se ha encontrado en poblaciones deM. fijiensis resistentes a azoxystrobin y trifloxys-trobin, y se asocia a factores de resistencia típicamentealtos (FR >100) en las poblaciones [Gisi et al., 2000;Chin et al., 2001]. Se ha planteado que esta mutaciónno afecta la eficiencia de la enzima, por lo cual no esta-blece una penalidad sobre la compatibilidad de losmutantes, como se ha comprobado en Blumeria graminiss.sp. tritici [Gisi et al., 2002], aunque esto requiere com-probación en otros patógenos.
Otro punto de mutación en el citocromo b es la susti-tución de una fenilalanina con leucina en la posición

Luis Pérez Vicente
62/fitosanidad
129 (F129L), que en contraste confiere una resistenciamás moderada con un FR entre 10-50 [Kuck y Mehl,2003]. Aunque esta mutación no se ha confirmado enM. fijiensis, la falta de correspondencia entre los re-sultados de ensayos de sensibilidad de laboratorio y dela frecuencia de la mutación G143A en las poblacionespudiera estar condicionada por la presencia de estamutación.
La enzima oxidasa alternativa (AOX) permite lacircunvención de los complejos III y IV y la aceptaciónde electrones directamente de la ubiquinona, mientrasque la dehidrogenasa alternativa permite oxidar tantoel NADH mitocondrial como citoplasmático sininvolucración del complejo I de la cadena del transpor-te de electrones en las mitocondrias [Word y Hollomon,2003]. De esta forma, existen tres posibles sistemas detransporte de electrones del NADH exógeno al oxígenoen las mitocondrias: 1) la ruta de los citocromos, lacual es sensible a los QoI (methoxyacrilatos y otros) ya los QiI (Antimycin A); 2) el sistema de la NADH-citocromo C reductasa con circunvención de la inhibi-ción de los metoxyacrilatos en el complejo bc1; 3) laoxidasa alternativa, que es funcional solo en mito-condrias aisladas de mutantes resistentes a azoxys-trobin y que es inhibida por el ácido salicilhidroxámico[Ziogas et al.,1997]. Por este motivo es importante in-cluir en el medio para el monitoreo de la sensibilidad aQoI 100 µg.mL–1 de SHAM para inhibir la oxidasa al-ternativa, la cual distorsiona los resultados de los testsde monitoreo de sensibilidad. Por cuanto la resistenciaestá controlada por genes mitocondriales, su herenciase asume que es no mendeliana [Gisi et al., 2002].
Anilinopyrimidinas (pyrimethanil). El pyrimethanil[Neumann et al., 1992] es un fungicida de más recienteintroducción en los programas de control de sigatokanegra. Este ingrediente activo inhibe la extensión de lostubos germinativos y la biosíntesis de metionina al inhi-bir la cistationa-liasa, y por tanto la síntesis de proteí-nas [Pontzen, 1997], y da lugar a la inhibición de la se-creción de cutinasas, pectinasas y celulasas, enzimasnecesarias para la degradación de la pared celular delhospedante y para la infección [Milling et al., 1993,Milling y Daniels, 1996]. Presenta actividad sistémicacon una buena absorción dentro de la hoja y retenciónen la cera, lo que previene su arrastre por la lluvia. Sudistribución es eminentemente translaminar. Se utilizaa razón de 250-300 g/ha–1 en tratamientos en aceite mi-neral, y su actividad es comparable a la del tridemorph[González et al., 2000; Orozco-Santos et al., 2004].
Un estudio realizado con 469 aislamientos de Venturiainaequalis mostró que la sensibilidad de las poblacio-nes al pyrimethanil y al myclobutanil (DMI pertene-ciente al grupo de las pyrimidinas) estuvieronsignificativamente correlacionadas, y se sugirió que unode los múltiples genes que condicionan la resistencia alos DMI pudieran también condicionar la resistencia alas anilinopyrimidinas [Köller et al., 2004]. Se recomien-da realizar el monitoreo de la sensibilidad de las pobla-ciones mediante la medición del crecimiento de los tu-bos germinativos de las ascósporas en 0; 1; 10,0; 30,0;100,0 µg.mL–1 [Duvert et al., 2002].
Inductores de reacciones de resistencia sistémica adqui-rida. Las plantas pueden inducirse localmente osistémicamente a hacerse más resistentes a las enfer-medades mediante varios estreses bióticos o abióticos(resistencia sistémica adquirida (SAR)). A través deuna red de señales, inducen un espectro de proteínasvinculadas a la resistencia (β-1,3 glucanasas yquitinasas, proteínas ricas en cisteína relacionadas a lataumatina) características de diferentes especies deplantas y sistemas de activación y por su dependenciadel ácido salicílico [AS; Gaffney et al., 1993; Delaney etal., 1994; Kessman et al., 1994a,b; Kessman et al., 1996;Friedrich et al., 1996; Oostendorp et al., 2001]. El niveldel AS endógeno se ha correlacionado estrechamentecon la inducción de proteínas PR y a una catalasa quese une al AS, la cual potencia la generación de H2O2[Chen y Klessig, 1991; Chen et al., 1993; 1994; Lawtonet al., 1996]. Se ha informado de la presencia de otraproteína de enlace con el AS con una afinidad cientocincuenta veces mayor que entre el SA y la catalasa, yde mayor nivel de producción de radicales superóxido[Du y Klessig, 1997]. Diferentes ingredientes activosmostraron inducir SAR, pero el más estudiado y cono-cido es el acybenzolar-S-methyl (ASM) [Görlach et al.,1996], el cual se trasloca sistémicamente en las plantasy que puede tomar el lugar del ácido salicílico en laruta metabólica natural del SAR, e inducir el mismoespectro de resistencia y los mismos marcadoresmoleculares. Su actividad en la inducción de SAR se harelacionado a la capacidad de unirse a la proteína paragenerar H2O2. Está demostrada la eficacia del ASM enel control de sigatoka negra y nemátodos en bananos[Madrigal y Ruess, 1998; Madrigal y Corrales, 2000] adosis de 40 g.ha–1 en mezclas con otros fungicidas entratamientos a intervalos entre 35 y 40 días utilizandodosis mínimas de aceite mineral. El ASM supera la efi-cacia de los fungicidas mejorando su nivel de control y/

fitosanidad/63
Manejo convencional y alternativo...
o extendiendo el intervalo entre tratamientos. En en-sayos realizados en Costa Rica, Corrales et al. (2002)informaron una reducción de 46% de consumo de acei-te mineral, de seis ciclos menos de fungicidas y de dosaplicaciones de nematicidas en un período de un año enlas áreas tratadas con cuatro ciclos de ASM + fungicidasque en las tratadas con fungicidas en la forma conven-cional. Las manchas en las hojas tratadas con ASMadquieren un aspecto distintivo, pierden sus márgenesbien delimitados y reducen marcadamente la velocidadde evolución, su tamaño y la formación de fructi-ficaciones [Pérez, 2000b].
Aceites y su uso en el control de Mycospherella spp. enbananos y plátanos. Los aceites como fungicidas pue-den ser clasificados de acuerdo con su origen: del petró-leo se derivan los aceites minerales constituidos pormezclas complejas o hidrocarburos; de las plantas yanimales se derivan ácidos grasos constituidos portriglicéridos de ácidos grasos; de las plantas tambiénse derivan los aceites esenciales volátiles y odoríferosconstituidos por mezclas de terpenoides, aldehídos oésteres. Los aceites minerales han sido ampliamenteempleados para el manejo de manchas de las hojas porMycosphaerella spp. en bananos, y se encuentran enestudio los aceites de ácidos grasos derivados de plan-tas, particularmente de palma aceitera (Elaeisguiinensis J.).
Los aceites tienen un triple papel en los tratamientosen bananos para el control de manchas por Mycos-phaerella spp.: 1) permiten reducir los volúmenes deaplicación al poderse producir una nube de aspersióncompuesta de pequeñas gotas sin que se volatilicen, yque garantiza una adecuada distribución o reparticióndel fungicida sobre la hoja; 2) limitan el lavado de losfungicidas por la lluvia que permite una mejor penetra-ción de los fungicidas sistémicos en las hojas; 3) inhibenlos procesos de desarrollo del patógeno y de la evolu-ción de la enfermedad, en tratamientos en las primerasfases del desarrollo de las lesiones.
Aceites minerales. Investigadores franceses introduje-ron la utilización de aceites minerales (AM) para el con-trol de sigatoka a principios de la década del cincuenta[Guyot, 1953; Guyot y Cuillé, 1954a,b, 1955 y 1956],así como la tecnología de su uso a bajo volumen en tra-tamientos aéreos [Cuillé y Guyot, 1954, 1956 y 1957].Estos estudios brindaron las bases para los métodosde control actuales en la mayoría de las plantacionesbananeras del mundo.
Los AM pueden ser definidos en función de su compo-sición química y su peso molecular, el contenido de re-siduos no sulfonados, la viscosidad y el rango de desti-lación.
Composición química y peso molecular (número de áto-mos de carbono). Los AM pueden ser de estructuraparafínica, nafténica, olefínicas y aromática [Calpouzoset al., 1961; Calpouzos y Colberg, 1964]. Los olefínicosy aromáticos contienen grupos no saturados con do-bles enlaces entre dos carbonos, mientras que losparafínicos y nafténicos tienen todos sus carbonos sa-turados y no presentan doble enlaces. Los aceitesparafínicos tienen la mayor actividad como pesticidas,y es deseable que su contenido sea alto (> 60%) en losaceites de uso agrícola con este fin. La presencia de en-laces no saturados dobles en los aromáticos los hacemás químicamente reactivos y presentan una mayorfitotoxicidad y riesgo a la salud humana [Jacques yKuhlmann, 1999].
La eficacia está relacionada con el tamaño de la molécu-la de parafina, y es óptima cuando se acerca a 23 átomosde C. El riesgo de fitotoxicidad aguda se incrementacuando el número de C es menor de 18 [Madsen yWilliam, 1967]. La fitotoxicidad crónica se incrementacon el aumento del peso molecular, especialmente porencima de 25 átomos de C.
Residuos no sulfonados (RNS). El residuo no sulfonadoes el volumen porcentual del aceite que no reaccionacon ácido sulfúrico 37 N. Los grupos no saturados o losdobles enlaces de C en las moléculas aromáticas reac-cionarán o se volverán sulfonados en presencia de ácidosulfúrico 37 N. Los aromáticos se consumen según avan-za la sulfonación. El nivel de compuestos no saturadosen un aceite tiene un significativo efecto en su poten-cial de daño agudo a las hojas. La fitotoxicidad de unaceite aumenta en la misma medida en que disminuyenlos residuos no sulfonados por debajo del 90%. Debidoa la precisión y reproducibilidad de los sistemas de aná-lisis (ASTM D 483) se ha fijado un nivel mínimo del92% para los aceites de uso en protección de plantas[Calpouzos y Colberg, 1964; Calpouzos et al., 1961;Walker, 1972].
Volatilidad. La volatilidad está considerada tambiéncomo una característica importante que ha de tenerseen cuenta en relación con la calidad de los aceites mine-rales para el control de sigatoka [Calpouzos et al., 1959;Delfel et al., 1962; Calpouzos, 1968; Walker, 1972]. Lavolatilidad del aceite determina la cantidad que queda

Luis Pérez Vicente
64/fitosanidad
en la hoja posterior al tratamiento, y por tanto tieneinfluencia sobre el control y la fitotoxicidad [Riedhart,1961]. A medida que su volatilidad es menor mejora elcontrol y son mayores los riesgos de fitotoxicidad. Lomás deseable es disponer de un aceite con un balanceóptimo de sus parámetros principales. En la Tabla 3 sereproducen las características de varios aceites mine-rales [Jacqques y Kauffman, 1999].
El efecto biológico de los AM y su papel en el control seha estudiado ampliamente [Brun, 1958; Calpouzos etal., 1959; Klein, 1961; Calpouzos, 1968; Stover yDickson, 1968; Pérez 1978a,b; Pérez et al., 1981]. LosAM se utilizan a dosis entre 5 y 15 L.ha–1. Su dosis deempleo depende de la severidad de la infección, de laepidemiología de la enfermedad y de la sensibilidad delas poblaciones de Mycosphaerella spp. a los fungicidasen uso, así como de las condiciones ambientales predo-minantes cuando se realizan los tratamientos. A dosisde 5 L/ha–1 e inferiores actúa fundamentalmente como
vehículo para permitir obtener gotas finas, bajos volú-menes de aplicación y mejorar la penetración de algu-nos ingredientes sistémicos. En el caso de los trata-mientos a dosis mayores de 10 L.ha–1 inhiben ademásla germinación, el crecimiento de los tubos germinativosy formación de apresorios, la penetración y, sobre todo,la evolución de los estados de síntomas primarios derayas a manchas. Brun (1958) informó de la inhibiciónde la germinación de conidios de M. musicola y de laevolución de los síntomas en función de su estado alrealizar el tratamiento de AM, propiedad en la que sebasó Klein (1960) para desarrollar un sistema de tra-tamientos contra sigatoka basado en conteos de rayasen hojas. En Cuba Pérez et al. (1981) encontraron entres ensayos repetidos en diferentes momentos del añouna inhibición altamente significativa de la germinaciónde los conidios y la penetración de los tubos germina-tivos por los estomas en hojas tratadas con aceite con10 y 15 L.ha–1, así como una inhibición marcada de laevolución de los síntomas de rayas 1 y 2 a manchas.
Tabla 3. Características físico-químicas de diferentes aceites minerales
Parámetro Orchex 796
Prorex 38
Banole HV
Spraytex M
RNS (%) 95,4 90 97 93,8 Parafinas (%) 67,5 65,3 75,5 60,6 Nafténicos (%) 29,8 29,6 21,1 36,4
Destilación (°C) 50% (punto medio) 227,3 216,3 216,3 225,7 90-10% (rango) 36,7 79,1 47,8 43,8 Número de carbono % < C18 (riesgo fitox. aguda) 2 5 10 3 % < C20 (menor eficacia) 9 22 22 14 % < C20-C26 (zona óptima) 77 54 70 64 % < C26 (riesgo de fitox. crónica) 14 24 8 22 % < C28 (mayor riesgo de fitox. crónica) 3 15 3 10
Pérez et al. (1993c) informaron de la eficacia de los tra-tamientos con fungicidas sistémicos en mezcla con aceitemineral de 10-12 L.ha–1 en el control de sigatoka negraal utilizar tratamientos por pronóstico bioclimático(preaviso). La eficacia de los tratamientos sobre la evo-lución de los síntomas de la enfermedad mejorósignificativamente al usar más aceite a razón de 10 L.ha–1.Guzmán y Romero (1996) encontraron una relación li-neal entre la dosis de AM utilizado y la severidad delataque de sigatoka negra en campo. Según aumentó ladosis de 5 a 10 y 15 L.ha–1, disminuyó el área foliarafectada significativamente; observaron períodos deincubación más largos y menor severidad de los ata-
ques en plantas tratadas con 10 L de aceite que las no tra-tadas, y que las tratadas con clorotalonil a 1,44 kg.ha–1.
La fitotoxicidad producida por los tratamientos conAM puede ser aguda o crónica. La aguda se manifiestaprimeramente como pequeñas pecas de color claro queinmediatamente se tornan de violáceas a oscuras, pro-ducen quemaduras necróticas y se deben principalmentea la calidad del aceite, a la acumulación de depósitosexcesivos. La crónica se manifiesta por la aparición depequeñas lesiones, primeramente acuosas, que más tardetornan la hoja de color bronceado, y se debe a la acu-mulación en tratamientos sucesivos que reduce el ren-dimiento debido a la interferencia con el intercambio

fitosanidad/65
Manejo convencional y alternativo...
de gases y la fotosíntesis [Riedhart, 1961; Israeli et al.,1993]. La aparición de este tipo de fitotoxicidad de-pende en buena medida de la calidad del aceite y delnúmero de tratamientos que se realicen y la predispo-sición de la planta por presencia de estrés hídrico y detemperatura. Con los aceites utilizados en Cuba los sín-tomas fitotóxicos característicos se hacen evidentes enplantas que han recibido alrededor de 70 L de aceiteaplicados en la estación.
Las plantas son además más susceptibles a mostrarsíntomas cuando hay estrés hídrico y de temperatura,como el que ocurre a la salida de la estación más lluvio-sa y húmeda, y el inicio de la seca y más fría. En CostaRica, Romero y Marín en 1993 [citados por Guzmán yRomero, 1996] encontraron un aumento de la fito-toxicidad cuando la dosis de aceite alcanzó los 330 L.ha–1,que se reflejó en una disminución del 5% del peso delracimo.
Aceites de palma aceitera. El aceite de palma se ob-tiene del mesocarpo del fruto de la palma de aceiteEleasis guineensis [Rutengwe, 2004; Choo, 2004]. Elárea mundial cosechada de aceite de palma en el 2004fue de 12 115 000 ha, y la producción mundial fue dealrededor de 162 000 000 t, de los cuales Malasia(43,6%) e Indonesia (37,2%) fueron los mayores pro-ductores. Centroamérica y Colombia producen cadauno 3 100 000 t [FAOSTAT data, 2005].
El aceite de palma muestra utilidad en el manejo deplagas insectiles, como emulsificantes y coadyuvantes.En tratamientos a dosis entre 9 y 12 L.ha–1 fue efectivopara el manejo de sigatoka negra tanto en mezcla confungicidas en sustitución del aceite mineral como solo.No se conocen antecedentes de fitotoxicidad hasta ladosis de 12 L.ha–1. Se informa la inhibición de lagerminación y de la esporulación en las manchas. Es-tas características lo hacen una alternativa naturaladaptable en el manejo de sigatoka negra en la produc-ción orgánica y como remplazo del aceite agrícola.
Vías alternativas al uso de fungicidas químicos. A pesarde que el control químico sigue siendo la vía más co-múnmente utilizada para el control de sigatoka negra,los costos crecientes de las nuevas tecnologías para laprotección, la cada vez más frecuente aparición de po-blaciones del patógeno con sensibilidad reducida a losingredientes activos en uso, y la casi universal preocu-pación por el contenido de residuos químicos en la fru-ta, ha establecido una creciente presión por encontrarvías alternativas al control químico. Estas son en ge-
neral solo factibles de uso en esquemas de manejo inte-grado de la enfermedad para reducir la presión del mal,ya sea junto al uso de los diferentes grupos de ingre-dientes activos en la producción convencional de bananoso solas en esquemas de producción orgánica.
Uso de clones resistentes a la enfermedad. El uso de clonesresistentes es el método más económico y ambien-talmente seguro de manejo de sigatoka negra, y prácti-camente el único método de control disponible parapequeños agricultores en áreas de media y altapluviometría [Hernández y Pérez, 2001]. Se han des-crito los tipos de interacciones existentes entreMycosphaerella fijiensis y Musa spp. [Fouré et al.,1984; Fouré et al., 1990; y Fouré, 1994]. Una de lasinteracciones más frecuentemente explotada en los pro-gramas de mejora para resistencia a sigatoka negra esla resistencia parcial utilizada ampliamente en el pro-grama de obtención de híbridos de la FHIA [Rowe yRichardson, 1975; Rowe y Rosales, 1994]. La resisten-cia parcial se manifiesta por el alargamiento de laincubación, de la transición de las lesiones de los esta-dos iniciales de rayas a manchas y de la formación decuerpos de reproducción sexual y asexual del patógenoque permite llegar con más hojas funcionales a la cose-cha [Pérez, 1998; Hernández y Pérez, 2001]. Se handesarrollado y distribuido por la FHIA y el IITA clonescon resistencia parcial aptos para los mercados locales,lo que en algunos casos ha obligado al cambio de laspreferencias gustativas. En Cuba se encuentran en pro-ducción más de 12 000 ha de clones tetraploides sinté-ticos de la FHIA (FHIA 23, FHIA, 18 y FHIA 21) conresistencia parcial, lo que ha permitido una importan-te disminución de los costos de producción [Pérez etal., 2002b].
Clones resistentes vía transgénesis. Los bananos del gru-po Cavendish, hoy comercializados a gran escala, pre-sentan alta susceptibilidad a sigatoka negra y esterili-dad femenina, por lo que son difícilmente mejorablesmediante el mejoramiento convencional. Se han puestoa punto metodologías eficientes para la transformacióngenética de bananos para resistencia a sigatoka negramediante uso de bombardeo de partículas y deplásmidos en Agrobacterium tumesfaciens [Sági et al.,1995a,b; Arinaitwe et al., 2004; Gómez-Koski et al.,2004]. En Cuba se han realizado transformaciones apartir de suspensiones de células embrionarias del clonGran enano mediante A. tumesfaciens cepa EHA 105portadora de los plásmidos pHCG59 (gen de β-glucanasa,aislado de plantas de frijol), y con los genes pHA C58 y

Luis Pérez Vicente
66/fitosanidad
pHGA91 (gen de quitinasa clase 1 y gen AP24 relacio-nado a la osmotina respectivamente, aislados de plan-tas de tabaco [Gómez-Koski et al., 2004; 2005; Bermúdezet al., 2005].
Después de obtener los permisos correspondientes dela Agencia Nacional de Bioseguridad, se llevaron al cam-po 150 líneas, junto a los clones originales no transfor-mados, y se seleccionaron un total de 15 líneastransgénicas (en total de las tres construccionesgenéticas utilizadas de proteínas PR), las que tuvieronun comportamiento superior al control en relación conel índice de infección y la severidad de la enfermedad ala floración. Se transformaron además suspensionesembrionarias del clon de plátano Navolean (AAB) y seseleccionaron un total de cuatro líneas transgénicas,tres de estas con la construcción glucanasa + AP24 yuna con la construcción quitinasa + AP 24. Los índicesde infección en los controles (clones originales no trans-formados) variaron entre el 33,7 y el 40,2%, mientrasque los de las líneas transformadas variaron entre el7,35 y el 10,4%. Estos materiales tienen un gran valorpara áreas de clones Cavendish bajo gran presión deenfermedad en regiones con altos patrones pluvio-métricos. Las perspectivas futuras del uso comercialde estos clones dependerá en gran medida de los cam-bios de la percepción pública sobre el consumo de ali-mentos procedentes de plantas transgénicas, y de lainformación futura que se genere sobre la inocuidadpara el consumo humano de los productos de los genesprocedentes de plantas de cultivo utilizados en las trans-formaciones.
Antagonistas microbianos. La lucha biológica en gene-ral se ha estudiado para el manejo de sigatoka negradebido a diferentes factores técnicos y de finan-ciamiento. Jiménez et al. (1987) evaluaron contrasigatoka negra más de doscientos aislados de bacteriasepifíticas de las hojas de plátanos, y encontraron eninvernadero un solo aislado de Pseudomonas sp. quebrindó una actividad comparable a la del clorotalonil.González et al. (1997) estudiaron la actividad de Serratiamarcesens (cepas A23 y R1), Serratia entomophyla (cepaA100) y de Bacillus cereus (cepa A30) en aplicacionesde estos microorganismos con quitina, en comparacióncon la quitina sola y el fungicida en invernadero y cam-po, y obtuvieron una eficacia insuficiente para garan-tizar el control. Tratamientos realizados con unbioproducto basado en metabolitos purificados dePseudomonas aeuroginosa, aislado Pss en estudios decampo, brindaron un control parcial e insuficiente de
la enfermedad, aun cuando fueron aplicados a interva-los cortos [Pérez, inédito]. En general, los niveles decontrol hasta el presente informados en la literaturacientífica hasta el presente no permiten considerar elcontrol biológico con antagonistas como una vía alter-nativa al control químico para el manejo de la enferme-dad en plantaciones de clones susceptibles en regionescon alta y media pluviometría (> de 200 mm/año).
Uso de compuestos vinculados a la nutrición con activi-dad sobre sigatoka negra. Entre estos se encuentran com-puestos químicos y derivados orgánicos de compostaje.
Vigor Cal-Phos. El calcio y el fósforo tienen un efectomarcado sobre el desarrollo de las enfermedades en lasplantas mediante la inhibición de enzimas pectolíticasutilizadas por los patógenos para destruir la pared ce-lular y el engrosamiento de la cutícula. Jácome et al.(2004) y Pasbergh-Gauhl et al. (2004) informaron el efec-to secundario de los tratamientos con Vigor Cal-Phosa dosis de 3 L.ha–1 en combinación con triazoles y otrosfungicidas convencionales en el control de sigatoka ne-gra, los cuales permitieron mejorar el nivel de control,alargar el intervalo entre ciclos y reducir en consecuen-cia el número total de ciclos de fungicidas por año, asícomo el volumen de aceite en la mezcla fungicida sinpérdida de eficacia, adicionalmente a un mayor tama-ño de las plantas, un mejor tamaño de los racimos y unmejor comportamiento frente a los diferentes tipos deestrés en las plantas.
El silicato de calcio y potasio induce la expresión demecanismos de defensa a la infección de Magnaporthegrisea y Tanathephorus cucumeris, Monographiellaalbensis, Cochliobolus miyabeanus en arroz [Datnoff etal., 1991; Rodrigues et al., 2002a,b; Kim et al., 2002],Sphaeroteca fuliginea y Pythium ultimum en cucurbi-táceas [Cherif, et al., 1994. Fawe et al., 1998], Cercosporacoffeicola en café [Pozza et al., 2004]. Entre los meca-nismos asociados a la inducción de resistencia se hanmencionado incrementos significativos en la producciónde quitinasas, β-1-3 glucanasa, peroxidasas, fenilalaninaamonio liasas, polifenoloxidasas y acumulación de com-puestos fenólicos, así como la formación de una capa dealta densidad de Si a los lados de la pared celular. Losresultados de los ensayos en invernadero y campo hanpermitido una inhibición marcada del desarrollo de es-tos patógenos y un aumento importante de los rendi-mientos, acompañado de una disminución marcada oabandono del uso de fungicidas. La ausencia de infor-mes en revistas científicas indica que se le ha prestadopoca o ninguna atención para su uso en el caso de las

fitosanidad/67
Manejo convencional y alternativo...
musáceas, lo que sería un aspecto importante por estu-diar en el futuro.
Derivados de compostaje y lixiviados de humus de lom-briz. Hay antecedentes del uso de bioproductos resul-tantes del compostaje y de lixiviados de la humificacióncon lombrices de estiércol y residuos de cosechas(casting) para el control eficiente de sigatoka negra enplantaciones orgánicas, tanto de clones susceptiblescomo resistentes [Cañarte, 2003; Álvarez, 2004].
Cañarte (2003) informó de la eficacia de las aplicacio-nes semanales en una finca de 300 ha de bananoCavendish en zona tropical húmeda de Guayaquil, Ecua-dor, con una mezcla en aceite mineral de 2 L.ha–1, delbioproducto resultante de la fermentación en biorreac-tores artesanales durante 21 días, de estiércol vacunoen mezcla con melaza de caña, a la que se les añadiómicroorganismos de suelo. Los tratamientos inhibierono retardaron la evolución de las manchas. Los produc-tos de las fermentaciones son ricos en macro ymicroelementos, así como en ácidos orgánicos.
En Cuba se utilizan 2 L de lixiviado de la lombrizcaliforniana Eisenia foetida obtenido de la suspensiónen agua de humus, resultante de la descomposición bajotecho de estiércol y residuos de plantas de plátano, auna concentración del 50% v/v, la cual se aplica sema-nalmente a las plantas de bananos y plátanos a alta den-sidad [Álvarez, 2004]. Las plantas tratadas muestranun excelente control de la enfermedad y retardo de laevolución de los síntomas. Se requiere de estudios paradilucidar los mecanismos de acción de los lixiviados dehumus de lombriz. Zhang et al. (1998) informaron quelos tratamientos con suspensiones acuosas de humus delombricultura indujeron SAR en cucurbitáceas yArabidiopsis. No obstante, hay evidencias de la presen-cia en los lixiviados de una flora bacteriana con una fuerteactividad antagónica [Alvarado, comunicación personal].
Balance de las alternativas para el manejode la sigatoka negra
El manejo de la sigatoka negra en el futuro seguirá en-focado en la integración de procedimientos culturales yquímicos. Los cambios en la percepción de los consu-midores de productos más sanos y en la preocupaciónpública por detener la contaminación ambiental ten-drá una influencia marcada en el futuro en las tecnolo-gías de control que se implementarán.
En este sentido hay que considerar los diferentes esce-narios de producción de musáceas. Las producciones
tradicionales de bananos Cavendish y plátanos altamen-te susceptibles a la enfermedad en zonas de altapluviometría en grandes superficies, por algunos años,mantendrán su dependencia del uso de fungicidas quí-micos, del saneamiento y de la mejora de las condicio-nes culturales para hacer más desfavorables las condi-ciones ambientales al desarrollo de la enfermedad. Enestas áreas los antagonistas hasta el momento parecentener pocas oportunidades como opción de control.
Mucho se ha avanzado en el desarrollo de grupos quí-micos más eficaces y menos riesgosos ambientalmente,y se ha ganado conocimiento en sus mecanismos de ac-ción, lo que permite establecer recomendaciones sóli-damente fundamentadas de las estrategias de utiliza-ción para disminuir la presión de selección de poblacionesresistentes a los ingredientes activos en uso. El patóge-no ha mostrado en el tiempo una gran capacidad devariación y adaptación, y desarrollado poblaciones conbaja sensibilidad a las diferentes familias de ingredien-tes activos que salen al mercado. La principal partici-pación la tendrán en el futuro los productos de contac-to apoyados por el uso de sistémicos, particularmentelos triazoles, a pesar de que hay evidencias en estasregiones de problemas crecientes de pérdida de sensibi-lidad.
A largo plazo, para estas regiones con alta pluviometríay presión de infección la resistencia genética es la únicasolución económica y ambientalmente factible. Un pocode resistencia parcial brinda grandes beneficios desdeel punto de vista epidemiológico, y puede ser integradaa los sistemas de manejo para complementar o susti-tuir los tratamientos químicos [Hernández y Pérez,2001; Pérez y Pérez, 2003]. Esta puede obtenerse en elcaso de las producciones de bananos Cavendish y plá-tanos mediante transgénesis, lo que dependerá de unaamplia concientización del efecto de los productos delos genes antifúngicos provenientes de plantas cultiva-das a la salud.
La situación de los productores pequeños de bananos yplátanos vinculados a la cadena de exportación, situa-dos dentro de los grandes macizos bananeros en el tró-pico húmedo, es muy variable y requiere de más aten-ción y asesoramiento en el manejo de fungicidas y de laenfermedad. Muchos no resisten un aumento de loscostos de las medidas de protección.
Las producciones de bananos y plátanos en trópicossemihúmedo y seco tienen más opciones mediante inte-gración de sistemas de manejo cultural (densidades de

Luis Pérez Vicente
68/fitosanidad
plantación, nutrición, drenajes, número de manos/ra-cimo), adopción de sistemas de monitoreo de la enfer-medad para optimizar los momentos de los tratamien-tos. En estas áreas hay fuertes oportunidades para lasproducciones orgánicas. Un caso exitoso que ha de se-guirse investigando cómo funcionan son los tratamien-tos con lixiviados de compost y vermicompost que hanvenido estableciéndose en alguna extensión en Ecuadory Cuba en plantaciones de clones susceptibles y conresistencia parcial.
Se requiere continuar el estudio de compuestos que in-duzcan mecanismos de defensa para integrarlos a las es-trategias de manejo de sigatoka negra. Entre estos habríaque determinar la factibilidad del uso del silicato de calcioy potasio como inductor de mecanismos de resistenciacontra patógenos de las raíces hojas y frutos en Musa.
REFERENCIAS
Álvarez, J. M.: Una nueva concepción de producción de plátano frutay vianda en Cuba, Impresiones del Ministerio de Agricultura, 2004,pp. 1-16.
Ammermann, E.; G. Lorenz; K. Schelberger; B. Mueller; R. Kirstgen; H.Sauter; «BAS 500 F–the New Broad-Spectrum Strobilurin Fungicide».Proceedings of the BCPC Conference–Pest and Diseases 2000,2:541-548, 2000.
Arias, O.; R. Ceciliano; R. González: «Use of Impulse 80 EC (Spiroxamine),for Combat of Mycosphaerella fijiensis in Bananas (Musa AAA)».Proceedings of the XV ACORBAT Meeting, Cartagena de Indias,2002, pp. 126-129.
Arinaitwe, G.; S. Remy; R. Strosse; R. Swennen; L. Sági:«Agrobacterium and Particle Bombardment-Mediated Transformationof a Wide Range of Banana Cultivars», Banana Improvement: Cellular,Molecular Biology And Induced Mutations. Proceedings of a Mee-ting Held Sept 24-28, 2001 in Leuven, Bélgica, FAO/AIEA Joint Division,INIBAP, Sience Publishers, Enfield, (NH, E.U.), 2004.
Barrantes, N.: «Bibliometric Analysis of Sigatoka Literature», SigatokaLeaf Spots Diseases of Bananas. Proceedings of an InternationalWorkshop Held at San José, Costa Rica, March 28, April 1. 1989,INIBAP, 1990, pp. 17-28.
Bermúdez, I.; R. Gómez; B. Chong; M. Reyes; J. Machado; O. Portal; B.Ocaña; Y. Alvarado; M. Leiva; M. Acosta; M. Cruz; B. Roque; R.Swennen; L. Sagi; L. Hernández: «Agrobacterium Tumefaciens-Mediated Genetic Transformation of Banana Musa spp. cv. GrandeNaine, (AAA) Using Chimerical Antifungal Genes for Black SigatokaResistance», Plant Cell Report, 2005.
Brun, J.: «Les traitements pesticides a débit réduit en culture fruitièretropicale. Etude sur l’action des fongicides huileux dans la lutte contrela Cercosporiose», Fruits 13:3-14, 1958.
Buchenauer, H.: «DMI-Fungicides-Side Effects on the Plant and Problemsof Resistance», Modern Selective Fungicides: Properties,Applications, Mechanism of Action, Gustav Fischer, Verlag, Jena,Germany,. 1995, pp. 259-290.
Calpouzos, L.; T. Theis; C. M. Rivera; C. Colberg: «Studies on the Actionof Oil in the Control of Mycosphaerella musicola on Banana Leaves»,Phytopathology 49:119-122, 1959.
Calpouzos, L.; N. E. Delfel; C. Colberg; T. Theis: «Relation of PetroleumOil Composition to Phytotoxicity and Sigatoka Disease Control onBanana Leaves», Phytopathology 51:317-321, 1961.
Calpouzos, L.; C. Colberg: «Importance of Source of Spray Oils forSigatoka Disease Control and Phytotoxicity to Banana Leaves»,Phytopathology 54:235-236, 1964.
Calpouzos, L.: «Oliz», Fungicides, vol. 2, Academic Press, New York,1968, pp. 367-393.
Cañas, G. P.; E. Rodríguez; L. F. Patiño; R. E. Arango: «Identificación decepas de Mycosphaerella fijiensis resistentes al benomyl, usandola reacción en cadena de la polymerasa PCR». Resúmenes, XVIReunión Internacional ACORBAT 2004, Oaxaca, México, sept. 26-oct-1, 2004.
Cañarte, S.: «Control de plagas en banano por la vía de la nutrición»,Memorias del Taller Internacional para el Manejo Integrado de SigatokaNegra y Otras Plagas en Musa. MUSALAC/INIBAP, Guayaquil, Ecua-dor, 2003.
Carlier, J.; E. Fouré; F. Gauhl; D. R. Jones; P. Lepoivre; X. Murichon; C.Pasbergh-Gauhl; R. A. Romero: «Black Leaf Streak», Diseases OfBanana, Abaca And Enset, CAB International, Wallingford, Inglate-rra, 2000, pp 37-79.
Chen, Z.; D. F. Klessig: «Identification of a Soluble, Salicylic Acid-BindingProtein That May Function in Signal Transduction in the Plant DiseaseResistance Response», Proc. Natl. Acad. Science, E. U. 88:8179-1886, 1991.
Chen, Z.; H. Silva; D. F. Klessig: «Active Oxygen Species in the Inductionof Plant Systemic Acquired Resistance by Salicylic Acid», Science262 (5141):1883-1886, 1993.
––––: «Involvement of Reactive Oxygen Species in the Induction ofSystemic Acquired Resistance by Salicylic Acid in Plants», Science242:1883-1886, 1994.
Chérif, M.; A. Asselin; R. R. Belanger: «Defense Responses Induced bySoluble Silicon in Cucumber Roots Infected by Pythium spp.»,Phytopathology 84:236-242, 1994.
Chin, K. M.; T. Arroyo; B. Foster; C. Steden: «Sensitivity ofMycosphaerella fijiensis to Demethylation-Inhibitors in CentralAmerica: Testing Methodology and Cross Resistance», Proceedingsof the XII ACORBAT Meeting, Santo Domingo, oct 28-nov 1, 1996.
Chin, K. M.; M. Wirz; D. Laird: «Sensitivity of Mycosphaerella fijiensisfrom Banana to Trifloxystrobin», Plant Dis. 85:1264-127, 2001.
Choo Y. M.: «Palm Oil Carotenoids», Food and Nutrition Bulletin.1993/1994 (no 2 June, http://www.unu.edu/unupress/food/8F152e/8F152E05.htm, 2004.
Corrales, O.; S. Knight; A. Madrigal: «Control of Black Sigatoka(Mycosphaerella fijiensis Morelet) and the Red Burrowing Nematode(Radopholus similis Cobb) in Banana, Using the Plant Activator Boost50 SC Within a Reduced-Input Crop Protection Program», Proceedingsof the XVth ACORBAT Meeting, Cartagena de Indias, 2002, pp. 143-147.
Cronshaw, K.; A. Akers: «Mode of Action of Tridemorph and Sensitivityof Mycosphaerella fijiensis Morelet», Sigatoka Leaf Spot Diseasesof Bananas. Proc. Int. Workshop, March 28-April 1, 1989, INIBAP,Montpellier, France. 1990.
Cuillé, J.; H. Guyot: «Les traitements fongicides des bananeraies.Utilisation des appareils de traitement en bananeraie», Fruits 9(7):269-288, 1954.
––––: «Les traitements aériens en bananeraie contre Cercosporamusae», Fruits 11:435-440, 1956.
––––: «Le matériel du traitement, son utilisation. Les réglage desappareils a débit réduit», Fruits 12 (11):461-475, 1957.
Datnoff, L. E.; R. N. Raid; G. H. Snyder; D. B. Jones: «Effect of CalciumSilicate on Blast and Brown Spot Intensities and Yields of Rice»,Plant Disease 75:729-732, 1991.

fitosanidad/69
Manejo convencional y alternativo...
Davidse, L. C.: «Benzimidazole Compounds: Selectivity and Resistance»,Fungicide Resistance in Crop Protection, Ed. PUDOC, Wageningen,1981.
Davidse, L.; H. Ichii: «Biochemical and Molecular Aspects of theMechanism of Action of Benzimidazole Fungicides, N-PhenylCarbamates and N-phenylformamidoximes and the Mechanism ofResistance to These Compounds in Fungi», Modern SelectiveFungicides: Properties, Applications, Mechanism of Action, Verlag,Jena, Alemania, 1995, pp. 305-322.
Delaney, T. P.; S. Uknes; B. Vernooij; L. Friedrich; K. Weymann; D.Negretto; T. Gaffney; M. Gut-Rella; H. Kessmann; E. Ward: «A Cen-tral Role of Salicylic Acid in Plant Disease Resistance», Science266:1247-1249, 1994.
Delfel, N. E.; L. Calpouzos; C. Colberg: «Measurement of Spray-OilVolatility and Its Relation to Sigatoka Fungus Disease Control»,Phytopathology 52:913-917, 1962.
Delp, C.: «Benzimidazole and Related Fungicides», Modern SelectiveFungicides: Properties, Applications, Mechanism of Action, Verlag,Jena, Alemania, 1995, pp. 291-303.
Du, H.; D. F. Klessig: «Identification of a Soluble High-Affinity SalicylicAcid-Binding Protein in Tobacco», Plant Physiol. 113:319-1327, 1997.
Dutzmann, S.; D. Berg; N. E. Clausen; W. Kramer; K. H. Kuck. Proc. Br.Crop Prot. Conf-Pests Dis. 1(47):395-402, 1996.
Duvert, P.; R. Milling; R. González-Quesada: «Monitoring the Sensitivityof Mycosphaerella fijiensis to Pyrimethanil», Memorias de la XVReunión de ACORBAT en Cartagena de Indias, Colombia, 27 oct.-2nov., 2002, pp. 153-157.
Edginton, L. V.; C. A. Peterson: «Systemic Fungicides: Theory, Uptakeand Translocation», Antifungal Compounds, vol. 2, Interactions inEcological Systems, Marcel Dekker, New York,. 1977, pp. 51-89.
Edginton, L. V.: «Structural Requirements of Systemic Fungicides»,Ann. Rev. of Phytopathology 19:107-124, 1981.
FAO STAT, www.fao.org/faostat, 2005.
Farías-Larios, J.; M. Orozco-Santos: «Effect of Pyraclostrobin on BlackSigatoka (Mycosphaerella fijiensis) control in Banana», Proceedingsof the XV ACORBAT Meeting, Cartagena de Indias, 2002, pp. 243-248.
Fawe, A.; M. Aboud-Zaid; J. G. Menzies; R. R. Bélanger: «Silicon-Mediated Accumulation of Flavonoid Phytoalexins in Cucumber»,Phytopathology 88:296-401, 1998.
Fouré, E.: «Les Cercosporiose du bananier et leurs traitements. Sélectiondes molécules fongicides nouvelles. Activités comparés dedifférentes molécules fongicides sur Mycosphaerella fijiensis Moreletagent de la maladie des raies noires des bananiers et plantains auGabon», Fruits 38:21-34, 1983.
Fouré, E.: «Les Cercosporiose du bananier et leurs traitements. Etudede la sensibilité variétale des bananiers et plantains á Mycosphaerellafijiensis au Gabon», Fruits 40:393-399, 1985.
––––: «Les Cercosporiose du bananier et leurs traitements. Efficacitécomparée des différentes molécules fongicides sur Mycosphaerellafijiensis Morelet., agent de la maladie des raies noires des bananierset plantains au Cameroun (I)», Fruits, 43:15-19, 1988a.
––––: «Stratégies de lutte chimique contre le Cercosporiose noire dubananier et des plantains provoqué par Mycosphaerella fijiensisMorelet, L’avertissement biologique au Cameroun. Evaluation despossibilités d’amélioration», Fruits 43:269-274, 1988c.
––––: «Les Cercosporiose du bananier et leurs traitements. Efficacitécomparée du pyrazophos et du triadimenol sur Mycosphaerellafijiensis. Sur Mycosphaerella fijiensis Morelet (agent de laCercosporiose noire des bananiers et des plantains au Cameroun lorsdes traitements du grandes surfaces», Fruits, 43:143-147, 1988d.
––––: «Stratégies de lutte chimique contre le Cercosporiose noire dubananier au Cameroun: efficacités comparées de différentesmolécules fongicides, stratégies d’alternance», Sigatoka Leaf SpotsDiseases of Bananas. Proceedings of an International WorkshopHeld at San José, Costa Rica, March 28, April 1, 1989, INIBAP, 1990,pp. 135-158.
––––: «Leaf Spot Diseases of Banana and Plantains Caused byMycosphaerella musicola and M. fijiensis», The Improvement andTesting of Musa: a Global Partnership. Proceedings of the FirstGlobal Conference of the International Testing Program held at FHIA,Honduras, April 27-30, INIBAP, Montpellier, France, 1994, pp. 37-46.
Fouré, E.; M. Grisoni; R. Zurfluh: «Les cercosporioses du bananier etleurs traitements. Comportement des variétés. II. Etude de la sensibilitédes bananaiers et plantains á Mycosphaerella fijiensis Morelet etdes quelques charactéristiques biologiques de la maladie des raiesnoires au Gabon», Fruits 39:365-378, 1984.
Fouré, E.; A. Molium Pefoura; X. Mourichon: «Etude de la sensibilitévariétale des bananiers et des plantains a Mycosphaerella fijiensisMorelet au Cameroun. Caractérisation de la résistance au champdes bananiers appartenant a divers groups génétiques», Fruits45:339-345, 1990.
FRAC: «Banana Working Group», 4th. Meeting, Summary andRecommendations, Miami Airport Marriott Hotel, Miami, 25 & 26 January,Organized by BASF, 1996.
Friedrich, L.; K. Lawton; W. Ruess; P. Masner; N. Specker; M. Gut-Rella;B. Meier; S. Dincher; T. Staub; S. Ukness; J. P. Metraux; H. Kessmann;J. Ryals: «A Benzothiadiazole Derivative Induces Systemic AcquiredResistance in Tobacco», Plant J. 10: 61-70, 1996.
Gaffney, T.; L. Friedrich; B. Vernooij; D. Negretto; G. Nye; S. Uknes; E.Ward; H. Kessmann; J. Ryals: «Requirement of Salicylic Acid for theInduction of Systemic Acquired Resistance», Science 261:754-756,1993.
Gauhl, F.: «Epidemiología y ecología de la sigatoka negra», Traducciónde la obra: «Unterouchungen zur Epidemiologie und Oxologie derSchwarzen Sigatoka Krankheit (Mycosphaerella fijiensis Morelet)an Kochbananen (Musa sp.) in Costa Rica». Tesis doctoral presen-tada en el Instituto Sistemático-Geobotánico e Instituto de Patología yProtección de Plantas de la Universidad Georg August Goettingen.UPEB, Ciudad de Panamá, 1990.
Geier B.; H. Schnagger; U. Brandt; A. M. Colson; G. Von Jagow: «PointMutation in Cytochrome-B of Yeast Ubihydroquinone: Cytochrome-COxidoreductase Causing Myxothiazol Resistance and FacilitatedDissociation of the Iron-Sulfur Subunit», European Journal ofBiochemistry 208:375-380, 1992.
Geier, B. M.; U. Haase; G. Von Jagow: «Inhibitors Binding to the Qo Siteof BC1 Complex: Comparative Studies of Yeast Mutants and NaturalInhibitor Resistant Fungi», Biochem. Soc. Trans. 22:203-209, 1993.
Georgopoulos, S. G.: «The Genetic Basis of Classification of FungicidesAccording to Resistance Risk», EPPO Bull. 15:513-517, 1985.
Gisi, U.; K. M. Chin; G. Knapova; R. Küng-Färber; U. Mohr; S. Parisi; H.Sierotski; U. Stenfield: «Recent Developments in Elucidating Modesof Resistance to Phenylamides, DMI and Strobilurin Fungicides», CropProtect. 19:863-872, 2000.
Gisi, U.; H. Sierotzki; A. Cook; A. McCaffery: «Mechanisms Influencingthe Evolution of Resistance to Qo Inhibitor Fungicides», Pest Manag.Sci. 58:859-67, 2002.
Godwin, J. R.; J. M. Anthony; J. M. Clough; C. R. A. Godfrey: «ICI 5504:A Novel, Broad Spectrum Systemic (ß-methoxyacrilate fungicide)».Proceedings of the Brighton Crop Protection Conference on Pestand Diseases, vol. 1,. 1992, pp. 435-442.
Godwin, J. R.; D. W. Bartlett; J. M. Clough; C. R. A. Godfrey; E. G.Harrison; S. J. Maund: «Picoxystrobin: A New Strobilurin Fungicidefor Use on Cereals». Proceedings of the Brit. Crop Prot. Conference-Pests and Diseases, 2:533-540, 2000.

Luis Pérez Vicente
70/fitosanidad
Gómez-Kosky, R.; B. Chong-Pérez; I. Bermúdez; Y. Alvarado; M. Leiva;J. López-Torres; J. Machado; O. Portal; O. Sweenen; L. Sági; L.Hernández: «Behavior in Field of Banana and Plantain CarryingChimeric Genes for the Resistance to Black Sigatoka (M. fijiensis)».Proceedings of the XVI International ACORBAT Meeting, Oaxaca,México, sept. 26-oct 1, 2004.
Gómez-Kosky, R.; B. Chong-Pérez; I. Bermúdez; Y. Alvarado; M. Leiva;J. López-Torres; J. Machado; O. Portal: «Obtención de líneas debanano cv. Gran Enano (AAA) con posible resistencia a la enferme-dad de la sigatoka negra (Mycosphaerella fijiensis Morelet) portransformación genética», Informe Final de Proyecto Nacional delCITMA (Código del proyecto: 00300136). Instituto de Biotecnologíade las Plantas (IBP), 2005, inédito.
González, R.; E. Bustamante; P. Shannon: «Integrated Biological Con-trol of Mycosphaerella fijiensis. Report of the International Workshopon Biological Control and Integrated Production in Banana Cultivation,Las Mercedes de Guácimo, Limón, Costa Rica, Nov.16-19, 1997,pp.132-139.
González R.; J. Bocanegra; C. Osorio; J. Valladares: «Effect of Siganex(Pyrimethanil) 60 SC in the Control of Black Sigatoka». Proceedingsof the XIV ACORBAT Meeting, San Juan, Puerto Rico Jul. 31-Aug. 4.,2000, p. 69.
Görlach, J.; S. Volrath; G. Knauf-Beiter; U. Beckhove; K. H. Kogel; M.Oostendorp; T. Staub; E. Ward; H. Kessmann; J. Ryals:«Benzothiadiazole A Novel Class of Chemical Inducers of SystemicAcquired Resistance, Activates Gene Expression and DiseaseResistance in Wheat», Plant Cell. 8:629-643, 1996.
Guyot, H.: «La lutte contre Cercospora musae dans les bananeraiesdu Guadeloupe, Essais de nébulisation (Fogging)», Fruits 8 (11):525-532, 1953.
Guyot, H.; J. Cuillé: «Les formules fongicides huileuses pour lestraitements des bananeraies», Fruits 9:289-292, 1954a.
––––: «Les formules fongicides huileuses pour les traitements desbananeraies. Essais de nébulization», Fruits 9:293-296, 1954b.
––––: «Les traitements fongicides des bananeraies. II Efficacités desdifférents modes de traitements. Rôle de l’huile», Fruits 10 (3):101-107, 1955.
––––: «Les traitements fongicides des bananeraies. III. Résultatspratiques obtenus en Guadeloupe lors applications par brouillardslégers huileux», Fruits 11 (4):141-150, 1956.
Guzmán, M.; R. A. Romero: «Eficacia de cuatro dosis de aceite agrícolaen el control de la sigatoka negra (Mycosphaerella fijiensis Morelet)en banano», Corbana 21:129-139, 1996.
Guzmán, M.; S. Knight; H. Sierotski; G. Franceshi; I. Alon: «Sensitivity toFungicides in Mycosphaerella fijiensis Morelet. A Global Overview».Memorias de la XVI Reunión Internacional de ACORBAT, Oaxaca,México, sept. 29-oct 1, 2004, pp. 251 y 252.
Heaney, S. P.; A. A. Hall; S. A. David; G. Olaya: «Resistance to Fungicidesin the QoI- STAR Cross Resistance Group: Current Perspective».Proceedings of the BCPC Conference Pest and Diseases, 2000, pp.755-762.
Hernández, A.; L. Pérez: «Reaction of Banana and Plantains Cultivarsto Black Sigatoka Disease Caused by Mycosphaerella fijiensis Moreletin Cuba. Epidemiological Components of the Resistance», Fitosanidad5 (3):9-15, 2001.
Hollomon, D. W.: «Do Morpholine Fungicides Select for FungicideResistance?», Fungicide Resistance, British Crop Protection Council,Farnham, Surrey, 1994, pp. 281-289.
Israeli, Y.; E. Shabi; W. R. Slabaugh: «Effect of Banana Spray Oil onBanana Yield in the Absence of Sigatoka (Mycosphaerella sp.)»,Scientia Horticulturae 56:116-117, 1993.
Jácome, L. H.; R. Blanco; J. L. Galindo: «Effect of Vigor-Cal-Phos (VCP)on Mitigation of Black Sigatoka Disease on Bananas». Memorias de
la XVI Reunión Internacional de ACORBAT, Oaxaca, México, sept.29-oct. 1, 2004, pp. 212-224.
Jacques, D. F.; B. Kuhlman: «Exxon/Esso Experience with Horticultural Mine-ral Oils», Sprays Oils Beyond 2000, oct. 25-29, Sydney, Australia, 1999.
Jiménez, J. M.; J. J. Galindo; C. Ramírez: «Estudios sobre combatebiológico de Mycosphaerella fijiensis var. Diiformis mediante bacte-rias epífitas». Memorias de la VII Reunión Internacional de ACORBAT,Costa Rica, 1987, pp. 105-109.
Jiménez, L. G.; R. A Rodríguez: «First Outbreak of Mycosphaerellafijiensis Tolerant of the Fungicide Benomyl in Bananas in Costa Rica»,Phytopathology 75:1177, 1985.
Kaars Sijpeisteijn, A.; H. M. Dekuijzen; J. W. Vonk: «Biologicalconversions of fungicides in plants and microorganisms», AntifungalCompounds, vol. 2, Interactions in Biological and Ecological Systems,Marcel Dekker, New York, 1977, pp. 149-208.
Kalamarakis, A. E.; M. A. De Waard; B. N. Ziogas; S. G. Georgopoulos:«Resistance to Triadimenol en Nectria haematococca var.cucurbitae», Pestic. Biochem. Physiol. 40:212-220, 1991.
Kerkenaar, A.: «Mechanism of Action of Cyclic Amines Fungicides:Morpholines and Piperidines», Modern Selective Fungicides:Properties, Applications, Mechanism of Action, Verlag, Jena, Ale-mania, 1995, pp. 185-204.
Kessmann, H.; T. Staub; J. Ligon; M. Oostendorp; J. Ryals: «Activationof Systemic Acquired Resistance in Plants», Europ. J. of Plant. Pathol.100:359-369, 1994a.
Kessmann, H.; T. Staub; C. Hoffman; T. Maetzke; J. Herzog; E. Ward; S.Uknes; J. Ryals: «Induction of Systemic Acquired Resistance in Plantsby Chemicals», Ann. Rev. of Plant. Pathol. 12:439-454, 1994b.
Kessmann, H.; M. Oostendorp; W. Ruess; T. Staub; W. Kunz; J. Ryals:«Systemic Activated Resistance: A New Technology for Plant DiseaseControl», Pestic. Outlook 7:1-13, 1996.
Kim, S. G.; K. W. Kim; E. W. Park; D. Choi: «Silicon-Induces Cell WallFortification on Rice Leaves: A Possible Mechanism of EnhancedHost Resistance in Blast», Phytopathology 92:1095-1103, 2002.
Klein, H. H.: «Control of Cercospora Leaf Spot of Bananas withApplications of Oil Sprays Based on the Disease Cycle»,Phytopathology 50:488-490, 1960.
––––: «Effects of Fungicides, Oil and Fungicide Oil Water Emulsions onDevelopment of Cercospora Leaf Spot of Banana in the Field»,Phytopathology 51:294-297, 1961.
Köller, W.; H. Scheinpflug: «Fungal Resistance to Sterol ByosinthesisInhibitors: A New Challenge», Plant Disease 71:1066-1074, 1987.
Köller, W.; J. P. Wubben: «Variable Resistance Factors of Fungicides ActingAs Sterol Demethylation Inhibitors», Pestic. Sci. 26:133-145, 1989.
Köller, W.; W. F. Wilcox; D. M. Parker: «Sensitivity of Venturia inaequalisPopulations to Anilinopyrimidine Fungicides and Their Contribution toScab Management in New York», Plant Dis. 89:357-365, 2004.
Kuck, K. H.; A. Mehl: «Trifloxystrobin: Resistance Risk and ResistanceManagement», Planzenchutz-Nachrichten Bayer 56 (74):313-325, 2003.
Kuck, K. H.; A. Kroneberg; G. Zuninga; K. Stoermer; A. Boebel; R.González; R. Guendel; A. Martínez; R. Blanco; F. Gauhl; J. Sandoval;D. MarÍn; J. F. Rodríguez; G. Leandro; H. Calle; B. Sheppard; M. V.Blanco; J. González; T. Arroyo; L. Jacome; G. YÉpez; H. Sierotzki; M.Guzmán; E. Bureau; M. J. García: «7ht. Meeting of the FRAC BananaWorking Group, Miami, Florida, 11-12 Feb.», Abstracts of the XVIACORBAT Meeting, Oaxaca, México, sept. 26-oct. 1, 2004.
Lawton, K. A.; L. Friedich; M. Hunt; K. Weymann; T. Delaney; H.Kessmann; T. Staub; J. Ryals: «Benzothiadiazole Induces DiseaseResistance in Arabidopsis by Activation of the Systemic AcquiredResistance Signal Transduction Pathway», Plant J. 10:71-82, 1996.

fitosanidad/71
Manejo convencional y alternativo...
Leroux, P.; M. Gredt: «Effets sur les inhibiteurs de la biosynthèse desstérols fongiques: I. Fungicides provoquant l’accumulation dedesmethylstérols», Agronomie 3:123-190, 1983.
Madrigal, A.; W. Ruess: «CGA 245704, A New Plant Activator to ImproveNatural Resistance of Banana Against Black Sigatoka». Proceedingsof the XIII ACORBAT Meeting, Guayaquil, Ecuador, Nov. 23-29, 1998,pp. 266-274.
Madrigal, A.; O. Corrales: «Boost 50 SC, Plant Activator Which ImprovesNatural Resistance of Banana to Black Sigatoka (Mycosphaerellafijiensis) and Nematodes (Radopholus similis)». Proceedings ofthe XIV ACORBAT Meeting, San Juan, Puerto Rico, Jul. 31- Aug. 4,2000, p. 69.
Madsen, H. F.; K. Williams: «Control of Pear Psylla with Oils and Oil-Insecticide Combinations», J. Econ. Ent. 60 (1):121-124, 1967.
Margot, P.; F. Huggenberger; J. Amrei; B. Weiss: «CGA 279202, A NewBroadspectrum Strobilurin Fungicide», Brighton Crop Protection,Conference Pest & Diseases, vol. 2, 1998.
Marín, D. H.: R. A. Romero; M. Guzmán; T. Sutton: «Black Sigatoka andIncreasing Threat to Banana Cultivation», Plant Disease 87:208-222, 2003.
Martin, L. A.; R. T. V. Fox; B. C. Valdwin; L. F. Connerton: «Use ofPolymerase Chain Reaction to the Diagnosis of MBC Resistance inBotrytis cinerea», British Crop Protection Conference Pest andDiseases, 1992, pp. 207-214.
Meredith, D. S.: «Banana Leaf Spot Disease, (Sigatoka) Caused byMycosphaerella musicola Leach», Phytopathological Paper 11,Commonwealth Mycological Institute, Kew Surrey, Inglaterra, 1970.
Milling R. J.; C. J. Richardson; A. Daniels: «Pyrimethanil Inhibits HydrolyticEnzyme Secretion by Botrytis spp. and Prevents lysis of Host Cells».Proceedings of the 6th International; Congress of Plant Pathology,Montreal, 1993.
Milling, R. J.; A. Daniells: «Effect of Pyrimethanil on the Infection Processand Secretion of Fungal Cell Wall Degrading Enzymes», ModernFungicides and Antifungal Compounds, Intercept, Andover, Inglate-rra, 1996, pp. 53-59.
Neumann G. L.; E. H. Winter; J. E. Pittis: «Pyrimethanil, A New Fungicide»,Brighton Crop Protection Conference, Pests and diseases 1:395-402, 1992.
Orozco Santos, M.; O. Pérez Zamora; J. Orozco Romero: «Control deSigatoka negra (Mycosphaerella fijiensis) en banano con aplicacionesdel fungicida pyrimethanil», Memorias de la XVI Reunión Internacional deACORBAT, Oaxaca, México, sept. 29-oct. 1, 2004, pp. 225-230.
Oostendorp, M.; W. Kunz; B. Dietrich; T. Staub: «Induced DiseaseResistance in Plants by Chemicals», European Journal of PlantPathology 107:19-28, 2001.
Pasbergh-Gauhl, C.; L. Jácome; H. Cubero: «Effect of NIR Booster andVigor Cal-Phos on Black Sigatoka and Growth of Banana».Proceedings of the XVI International ACORBAT Meeting, Oaxaca,México, sept. 29-oct 1, 2004, pp. 201-211.
Pérez, L.: «Control de Mycosphaerella musicola Leach (Sigatoka) conaceite y mezclas de fungicidas en aceite», Agrotecnia de Cuba10:83-94, 1978a.
––––: «Control de Mycosphaerella musicola Leach (sigatoka) confungicidas sistémicos mezclados en aceite mineral», Agrotecnia deCuba 10:95-104, 1978b.
––––: «Black Sigatoka Disease Control in Banana and PlantainsPlantations in Cuba. Management of the Disease Based on AnIntegrated Approach», Infomusa 7(1):27-30, 1997.
––––: «Sigatoka negra (Mycosphaerella fijiensis Morelet) de bananos yplátanos (Musa spp.) en Cuba. Biología, epidemiología y manejo integra-do de la enfermedad», Memorias del Simposio Internacional de Sigatokanegra, Manzanillo, Colima, México, 8-10 de julio, 1998, pp. 24-52.
––––: «Black Sigatoka (Mycosphaerella fijiensis Morelet) of Bananasand Plantains (Musa spp.): Advances in Disease Management».Proceedings of the XIV ACORBAT Meeting, San Juan, Puerto Rico,Jul. 31-Aug. 4, 2000b.
Pérez, L.: «Manejo integrado de plagas y enfermedades en bananosen Cuba», Memorias del Taller Internacional para el Manejo Integradode Sigatoka Negra y Otras Plagas en Musa, MUSALAC/INIBAP, Gua-yaquil, Ecuador, 2003.
Pérez, L.; L. Peñalver; M. Delgado: «Acción del aceite sobre el desa-rrollo de Mycosphaerella musicola, agente causal de la mancha dela hoja (sigatoka) en plátano», Ciencia y Técnica en la Agricultura.Serie Protección de Plantas 4 (1):49-57, 1981.
Pérez, L.; V. T. Hoa; F. Mauri: «Lucha contra Mycosphaerella musicolaLeach con fungicidas sistémicos derivados de triazol», Ciencia y Téc-nica en la Agricultura. Serie Protección de Plantas 6 (4):87-98, 1983.
Pérez, L.; M. Iglesias; F. Mauri: «Aparición de una raza resistente al benomylde Mycosphaerella musicola Leach ex Mulder, agente causal de lasigatoka en los plátanos», Agrotecnia de Cuba 17 (1):79-98, 1985.
Pérez, L.; F. Mauri; B. Barranco; G. García: «Efficacy of EBI Fungicidesin the Control of Mycospha erella fijiensis Morelet on Bananas andPlantains with Treatments Based on Stage of Evolution of the Disease(Biological Warnings) in Cuba». Proceedings of the 6th InternationalCongress of Plant Pathology, Montreal, 1993a.
Pérez, L.; A. Batlle: «Monitoring Procedure to Determine PropiconazoleResistance in Mycosphaerella fijiensis Morelet». Proceedings ofthe 6th International; Congress of Plant Pathology, Montreal, 1993b.
Pérez, L.; A. Hernández; A. Porras; F. Mauri: «Efficacy of a BiologicalWarning System for Timing Fungicide Treatments for the Control ofBlack Sigatoka Disease (Mycosphaerella fijiensis Morelet) in Bana-na Plantations in Cuba». Proceedings of the 6th International Congressof Plant Pathology., Montreal, 1993c.
Pérez, L.; F. Mauri: «Efficacy of EBI Fungicides on the Control of SigatokaDisease of Banana Caused by Mycosphaerella musicola Leach ex Mulderin Cuba», International Journal of Pest Management 40:1-5, 1994.
Pérez, L.; F. Mauri; A. Hernández; E. Abreu; A. Porras: «Epidemiologíade la Sigatoka negra (Mycosphaerella fijiensis Morelet) en Cuba. IPronóstico bioclimático de los tratamientos en bananos (Musaacuminata AAA)», Revista Mexicana de Fitopatología 18 (1):15-26, 2000a.
Pérez, L.; A. Hernández, A. Porras: «Epidemiología de la Sigatoka ne-gra (Mycosphaerella fijiensis Morelet) en Cuba. II Pronósticobioclimático de los tratamientos contra la enfermedad en plátanos(Musa spp. AAB)», Revista Mexicana de Fitopatología 18 (1):27-36, 2000b.
Pérez, L.; A. Hernández; R. Trujillo; L. Hernández; M. Pérez: «Efficacyof Azoxystrobin and Trifloxystrobin in Banana and Plantains»,Abstracts of the XIV ACORBAT Meeting, San Juan, Puerto Rico, Jul.31-Aug. 4, 2000c.
Pérez, L.; A. Hernández; L. Hernández; M. Pérez: «Effect ofTrifloxystrobin and Azoxystrobin on the Control of Black Sigatoka(Mycosphaerella fijiensis Morelet) on Banana and Plantain», CropProtection 21:17-23, 2002a.
Pérez, L.; J. M. Álvarez; M. Pérez: «Economic Impact and Managementof Black Leaf Streak Disease in Cuba», Mycosphaerella Leaf SpotDiseases of Bananas: Present Status and Outlook. Proceedings ofthe 2nd. International Workshop on Mycosphaerella Leaf SpotsDiseases, San José, Costa Rica, 2002b, pp. 71-84.
Pérez, L.; A. Batlle; A. Hernández; R. Trujillo; C. álvarez; A. Méndez:«Evolución de la sensibilidad a fungicidas de las poblaciones deMycosphaerella fijiensis Morelet en banano en Cuba», Fitosanidad7 (3):49-54, 2003.
Pérez, L.; M. Pérez: «Epidemiología de la Sigatoka negra (Mycosphaerellafijiensis Morelet) en clones híbridos de la FHIA con resistencia par-

Luis Pérez Vicente
72/fitosanidad
cial», Memorias del Taller Internacional para el Manejo Integrado deSigatoka Negra y Otras Plagas en Musa, MUSALAC/INIBAP, Guaya-quil, Ecuador, 2003.
Pontsen, R.; S. Wissfeld: «Evaluation About the Distribution and SystemicBehavior of Triazoles in and on the Banana Leaves», Abstracts of XIIACORBAT Meeting, Santo Domingo, oct. 27-nov. 2, Santo Domingo, 1996.
Pontzen, R.: «Recent Developments in the Chemical Control of PlantDiseases», Resúmenes III Seminario Científico Internacional de Sani-dad Vegetal, Ciudad de La Habana, junio 23-27, 1997, pp. 61 y 62.
Pozza, A. A. A.; E. Alves; A. Edson; «Efeito do silício no controle dacercosporiose em três variedades de cafeeiro», Fitopatol. bras.29:185-188, 2004.
Riedhart, J. M.: «Influence of Petroleum Oil in Photosynthesis of BananaLeaves», Tropical Agriculture, Trinidad 39:327-338, 1961.
Robert, C. F.; A. López; R. H. Fulton: «Quantitative Modifications inBanana Field Surveys to Improve Spray Timing and Control of BlackSigatoka Mycosphaerella fijiensis var. diiformis», Abstracts of the74th. Annual Meeting APS, Phytopathology 72:987, 1982.
Rodrigues, F. A.; F. X. R. Vale; L. E. Datnoff; A. S. Prabhu; G. H. Korndörfer:«Effect of Rice Growth Stages and Silicon on Sheath BlightDevelopment», Phytopathology 93:256-261, 2002a.
Rodrigues, F. A.; N. Benhamou; L. Datnoff; B. Jones; R. Bélanger:«Ultrastructural and Cytochemical Aspects of Silicon-Mediated RiceBlast Resistance», Phytopathology 93:533-546, 2002b.
Romero, R. A.; T. B. Sutton: «Characterization of Benomyl Resistance inMycosphaerella fijiensis, Cause of Black Sigatoka of Banana inCosta Rica», Plant Disease 82:931-934, 1998.
Romero, R. A.: «Black Leaf Streak, Control», Diseases of banana, Abaca,and Enset, CABI Publishing, Wallingford, Inglaterra, 2000, pp. 72-79.
Rowe, P. R.; D. L. Richardson: «Breeding Bananas for DiseaseResistance, Fruit Quality and Yield», Bulletin 2. Tropical AgricultureResearch Services (SIATSA), La Lima, Honduras, 1975.
Rowe, P. R.: F. E. Rosales: «Musa Breeding at FHIA», The Improvementand Testing of Musa: A Global Partnership. Proceedings of the FirstGlobal Conference of the International Testing Program held at FHIA,Honduras, Apr. 27-30, INIBAP, Montpellier, Francia, 1994, pp. 117-129.
Rutengwe, R. M.: «The Magic Power of the Natural Multiple Nutrient Oilof Africa: Red Palm Oil», African Journal of Food Agriculture, Nutritionand Development (on line) 4(1) (http://www.ajfand.net/Issue-VI-files/IssueVI-Editorial.htm), 2004.
Sági, L.; B. Panis; S. Remy; H. Schoofs; K. De Smet; R. Swennen;B.P.A. Cammue: «Genetic Transformation of Banana and Plantain(Musa spp.) Via Particle Bombardment», Bio-Technology 13: 481-485, 1995a.
Sági L.; S. Remy; B. Verelst; R. Swennen; B. Panis: «Genetic Transformationin Musa Species (Banana)», Biotechnology in Agriculture and Forestry,Plant Protoplasts and Genetic Engineering VI, vol. 34, Springer-Verlag,Berlin, Heidelberg, New York, 1995b, pp. 214-227.
Sauter, H.; W. Steglich; T. Anke: «Strobilurines, A New Class of ActiveSubstances», Angew. Chem. Int. Ed. 38:1328-1349, 1999.
Sierotzki, H.; J. Wullschleger; U. Gisi: «Point Mutation in Cytochrome bGene Conferring Resistance to Strobilurine Fungicides in Erysiphegraminis f. sp. tritici Field Isolates», Pest Biochem. Physiology 68:107-112, 2000a.
Sierotzki, H.; S. Parisi; U. Steinfeld; I. Tenser; S. Poirey; U. Gisi: «Mode ofResistance to Respiration Inhibitors at the Cytochrome bc
1 Complex
of Mycosphaerella fijiensis», Pest Management Science 56:833-841, 2000b.
Sisler, H. D.; N. Ragsdale; W. W. Waterfield: «Biochemical Aspects ofthe Fungitoxic and Growth Regulatory Action of Fenarimol and OtherPyrimidin-5-yl Methanols», Pesticide Sci., 167-176, 1984.
Sisler, H. D.: «Chemistry and Mode of Action of Sterol ByosinthesisInhibitors. Characteristics of Fungal Resistance to Sterol 14Demethylase Inhibitors», 4th Meeting of FRAC Banana Working Group,Miami, Jan. 25-26, 1996.
Stover, R. H.; J. H. Dickson: «Leaf Spots of Bananas Caused byMycosphaerella musicola: Action of Oil on the Life Cycle of thePathogen», Canadian Journal of Botany 46:1495-1505, 1968.
––––: «Leaf Spot of Bananas Caused by Mycosphaerella musicola:Methods of Measuring Spotting Prevalence and Severity», Trop. Agric.Trinidad 47:289-302, 1970.
Stover, R. H.: Banana Plantain and Abaca Diseases, CommonwealthMycological Institute, Kew, 1972.
––––: «Behavior of Benomyl Tolerant Strains of the Black SigatokaPathogen in the Field», Proc. American Phytopathological Soc. 4:180-181, 1977.
Stover R. H.: «Sigatoka Leaf Spots of Bananas and Plantains», PlantDisease Reporter 64:750-756, 1980b.
––––: «Effet du Cercospora noir sur les plantains en Amérique Centrale»,Fruits 38:326-329, 1983.
––––: «Thirty Years of Changing Control Strategies», Sigatoka LeafSpots Diseases of Bananas. Proceedings of An InternationalWorkshop Held at San José, Costa Rica, Mar. 28, Apr.1, 1989. INIBAP,1990, pp. 66-74.
Tiemann, R.; D. Berg; W. Krämer; R. Pontzen: «Biochemistry of the NewFungicide KWG 4168 (Spiroxamine)», Pflanzenchutz-NachrichtenBayer 50:29-48. 1997.
Tomlin, C. D. S.: The E-Pesticide Manual 2002-2003. A worldcompendium, twelfth edition, version 2.2., British Crop ProtectionCouncil, Wise & Loveys Information Services, 2002.
Van Tuyl, J. M.: «Genetics of Fungal Resistance to Systemic Fungicides»,Meded. Landbouwhogeschool Wagenigen 77-2, 1-136, 1977.
Vincent, P. G.; H. D. Sisler: Physiol. Plantarum, 21:1249, 1968.
Walker, L. A.: «Properties of Banana Spray Oil in Relation to SigatokaDisease Control and Phytotoxicity on Banana Leaves», Pans 18:34-42, 1972.
Washington, J.R.; J.Cruz; F. López; M. Fajardo: «Infection studies ofMycosphaerella fijiensis on banana and the control of Black Sigatokawith chlorothalonil», Plant Disease 82:1185-1190. 1998a.
Washington, J. R.; J. Cruz; M. Fajardo: «Detection of Chlorothalonil inDew Water Following Aerial Spray Application and Its Role on theControl of Black Sigatoka of Banana», Disease 82:1191-1198, 1998b.
Wirtz, M.; K. M. Chin: «Detection and Characterization of StrobilurinResistant Individuals of Black Sigatoka (Mycosphaerella fijiensisMorelet) in Costa Rica», Abstracts of the XIV ACORBAT Meeting,San Juan, Puerto Rico, Jul. 31-Aug. 4, 2000.
Wolfe, M. S.: «Dynamics of Pathogen Population in Relation to FungicideResistance», Fungicide Resistance in Crop Protection, Pudoc,Wageningen, 1981, pp. 46-52.
Wood, P. M.; D.W. Holloman: «A Critical Evaluation of the Role of AlternativeOxidase in the Performance of Strobilurin and Related FungicidesActing at the Qo Site of Complex III», Pest Management Science59:499-511, 2003.
Zhang, W.; D. Y.Han; W. A. Dick; K. R. Davis; H. A. J. Hoitink: «Compostand Water Extract-Induced Systemic Acquired Resistance inCucumber and Arabidopsis», Phytopath. 88:450-455, 1998.
Ziogas, B. N.; C. Brian; B. C. Baldwin; J. E. Young: «AlternativeRespiration: A Biochemical Mechanism of Resistance to Azoxystrobin(ICIA 5504) in Septoria tritici», Pestic. Sci. 50:28-44, 1997.

fitosanidad/73
FITOSANIDAD vol. 10, no. 1, marzo 2006
NUEVOS AISLADOS DE BACILLUS SPP. ANTAGONISTASA SCLEROTIUM ROLFSI, RHIZOCTONIA SOLANI Y PYTHIUMAPHANIDERMATUM
Acenet I. Sosa López,1 Victoria Pazos Álvarez-Rivera2 y Marleny González García1
1 Instituto de Investigaciones de Sanidad Vegetal. Calle 110 no.514 e/ 5a. B y 5a. F, Playa, Ciudad de La Habana, CP 11600, [email protected] Facultad de Biología de la Universidad de La Habana. Calle 25 no. 453 e/ J e I, Vedado, Plaza de la Revolución, Ciudad de La Habana, CP 10500
Com
unic
ació
n c
orta
El cultivo del tabaco ocupa un lugar importante en laagricultura cubana por los aportes y beneficios que re-porta anualmente al país, y al poseer amplia venta en elmercado europeo adquiere una elevada importancia, porlo que recibe una atención priorizada [Pérez et al., 2000].
Anualmente los semilleros de tabaco muestran afecta-ciones provocadas por numerosas plagas y enfermeda-des, cuyos daños severos ocasionan el colapso total delas plántulas, fundamentalmente en la fase tempranadel cultivo; por lo general los hongos fitopatógenos delsuelo ocasionan estas enfermedades. Hay estudios quemuestran que las especies pertenecientes a los génerosFusarium, Pythium, Sclerotium, Rhizoctonia yPhytophthora son las más frecuentemente encontradas,en muchas ocasiones debido a las condiciones climáticasque se registran anualmente en el país [Brada et al.,1995].
El control biológico como alternativa o como comple-mento del control químico ocupa un lugar muy impor-tante en el control de plagas y enfermedades. El grupode las Pseudomonas fluorescentes, el hongo Trichodermaharzianum y bacterias pertenecientes al género Bacillusse caracterizan por presentar un elevado potencial en elcontrol de tales patógenos. Estas últimas son produc-toras de hormonas, antibióticos y metabolitostermoestables que les confieren una alta capacidad decolonización en los ambientes agrícolas y a su vez impi-de el establecimiento de otros microorganismospatógenos [Bochow et al., 1995].
El objetivo de este trabajo fue obtener aisladosbacterianos, pertenecientes al género Bacillus, con efec-
to antagonista para el control de Sclerotium rolfsii,Rhizoctonia solani (Rs10) y Pythium aphanidermatumen condiciones in vitro, a partir de muestras de hojas,suelo, raíces y rizosferas colectadas en las provincias dePinar del Río, La Habana, Ciudad de La Habana,Sancti Spíritus, Cienfuegos, Granma, Santiago de Cubay Las Tunas.
Los patógenos empleados en este estudio para la reali-zación de las pruebas de antagonismo pertenecen a lacolección del Laboratorio de Fitopatología del INISAV.Las muestras se procesaron según procedimiento des-crito por Fernández-Larrea (1999). Para determinar lascaracterísticas morfológicas y culturales de las coloniasy de las células cultivadas en agar nutritivo se realiza-ron observaciones al microscopio estereoscópico (100X)y al microscopio óptico (1000X). Para la determina-ción del género Bacillus se utilizó el Manual de Bergey[Claus y Berkeley, 1986]. La actividad biológica de losaislados identificados como Bacillus spp. se evaluó invitro mediante un enfrentamiento dual con lospatógenos Sclerotium rolfsii, Rhizoctonia solani (Rs10)y Pythium aphanidermatum. Como testigo se empleósolo el patógeno, sembrado en el centro de las placas dePetri que contenían PDA; luego se incubó todo por elperíodo de veintiún días y se realizaron observacionesdiarias del experimento, con el fin de detectar algunamanifestación de antagonismo, y además observar elcomportamiento de ambos microorganismos duranteel período de incubación. Las evaluaciones se hicieron alos cinco, quince y veintiún días, además de repetirse elexperimento tres veces.

Sosa y otros
74/fitosanidad
Se obtuvo un total de 323 aislados nativos, con carac-terísticas semejantes a las descritas para el géneroBacillus al presentar células en forma de bastón, alar-gadas, presencia de endoesporas y esporas, por respon-der positivamente a la tinción de Gram y ser catalasapositiva. Dentro de ellos, 17 mostraron efecto antago-nista a los hongos S. rolfsii, R. solani (Rs10) y P. apha-nidermatum, ya que al tercer día de iniciado el experi-mento los testigos de los tres patógenos habían cubiertola placa de Petri completamente; sin embargo, al en-frentarse con los Bacillus fueron susceptibles a 17 deellos, al presentar diferentes manifestaciones de anta-gonismo. En una el crecimiento micelial del patógenofue diferente al testigo, con el micelio enroscado y sinllegar a cubrir la placa completa; en otra el micelio al-canzó a cubrir la placa, pero no se distinguió a simplevista, sino mediante el microscopio estereoscópico 100Xdebido a que no presentaba la misma coloración conrespecto al testigo, y además era un micelio empobreci-do pegado a la superficie del medio de cultivo. Otrocaso presentó una inhibición total del crecimientomicelial del patógeno, y por último se observó la pre-sencia de una zona clara que separaba a un microorga-nismo de otro sin crecimiento aparente por parte deambos. Todo parece indicar que tanto el hongo como labacteria segregan algún tipo de metabolito al medioque impide el crecimiento del patógeno, y aunque a sim-ple vista no se observa crecimiento bacteriano, se de-mostró experimentalmente que sí, pero que la pobla-ción de este último fue menor que en el resto de la placa,lo que prueba que este grupo es resistente al metabolitoproducido por el patógeno.
El proceso de selección de bacterias antagonistas in vitrose considera el paso más difícil en el desarrollo de uncandidato promisorio para el control biológico de en-fermedades [Talavera et al., 1998]. Estos autores plan-tean además que de 25 aislamientos a partir de larizosfera de cereales, solo uno produjo una zona de in-
hibición contra algunos de los microorganismos proba-dos, mientras que Jiménez et al. (2001) seleccionaronsolo una cepa que podía inhibir diferentes tipos depatógenos entre los 30 aislamientos obtenidos derizosfera de papa.
Este trabajo demuestra metodológicamente que a par-tir de este tipo de interacción antagónica es posibleencontrar bacterias antagonistas evaluables para elbiocontrol, a pesar de la desproporción entre el total deaislamientos obtenidos y el número de antagonistasencontrados, lo que indica que pese a que la microbiotadel suelo es abundante y diversa, el número de antago-nistas no es proporcional.
REFERENCIASBochow, H.; K. Gantcheva; A. Vanachter: «Soil Introductions of Bacillus
subtilis As Biocontrol Agent and Its Population and Activity Dynamic»,Fourth International Symposium on Soil and Substrate Infestationand Desinfestations, Leuven, Bélgica, Sept., Acta-Horticultural382:164-172, 1995.
Brada, I. E.; E. Quintana; E. Pelaya; T. Araújo: «Efecto de Bacillus spp.sobre la germinación y desarrollo de semillas de tomate (Lyco-persicum esculentum M.) infestadas con Fusarium oxysporiumSchl. var. cubensis Smith», Resúmenes Bioplag 95, INIFAT, 26-28 abr.,La Habana, 1995.
Claus, D.; R. C. W. Berkeley: «Genus Bacillus. Bergey’s», Manual ofSystematic Bacteriology, 9th ed., vol. 2, Williams & Wilkins, pp. 1105-1139, 1986.
Fernández-Larrea, O.: «Aislamiento, selección y estudio de cepas deBacillus thuringiensis para el control fitosanitario», Informe final,PNCT, Biotecnología Avícola, INISAV, Cuba. 1999.
Jiménez, Rocío; G. V. Calleros; S. T. Franco; V. O. Portugal: «Bacteriaspromotoras del crecimiento de plantas», Agro-Biotecnología. Avan-ce y Perspectivas 20:395-400, 2001.
Pérez, E.; A. Fernández; V. Andino; M. García; E. Paredes; B. L. Muiño;M. Stefanova; F. Piedra; C. Hernández; O. Rodríguez; C. M. Medina:Tecnología para la eliminación del bromuro de metilo. Semillerosde tabaco con sustrato orgánico y uso de medios biológico. Pro-yecto Eliminación total del bromuro de metilo en el cultivo del tabacoen Cuba, INISAV, MINAGRI, La Habana, 2000.
Talavera, M. E.; E. Bustamante; R. González; V. Sánchez: «Selección yevaluación en el laboratorio y campo de microorganismos glu-canolíticos antagonistas a Mycosphaerella fijiensis», Manejo Inte-grado de Plagas, 47:24-30, 1998.

fitosanidad/75
FITOSANIDAD vol. 10, no. 1, marzo 2006
INSECTOS DEL ORDEN LEPIDOPTERA DETECTADOSEN PLANTAS DE NONI (MORINDA CITRIFOLIA L.)
Nancy González García, Rubén Avilés Pacheco, Bienvenido Cruz Díaz y Yolanda Martínez Suárez
Instituto de Investigaciones Fundamentales en Agricultura Tropical Alejandro de Humboldt. Calle 1a.esq. a 2, Santiago de las Vegas, Ciudad de La Habana
El noni (Morinda citrifolia L.), conocida en Cuba tam-bién con los nombres de mora de la India, guanábanacimarrona, empella y árbol del pan, pertenece a la fa-milia Rubiaceae y está considerada como originaria deLa Polinesia [Roig, 1965, 1974].
Debido a los resultados terapéuticos positivos atribui-dos a la ingestión de los jugos de sus frutos, que contri-buyen a la disminución de afectaciones causadas pordistintas enfermedades, y a las cualidades antioxidantesque posee, así como que algunos estudios compruebanel potencial de noni para estimular el sistemainmunológico y para inhibir el crecimiento de los tu-mores, regularizar el funcionamiento de las células yregenerar las dañadas [Solomon, 2005], esta planta seha distribuido por todo el país en patios y parcelas[Rodríguez y Sánchez, 2004].
La planta de noni es atacada por un grupo de plagasy enfermedades que afectan su crecimiento y produc-ción, dentro de las cuales se destaca la bibijagua uhormiga arriera [Hidalgo, 2004]. En evaluaciones rea-lizadas a plantas sembradas en pequeñas parcelas enpatios, en la provincia de Ciudad de La Habana, seobservó un fuerte ataque a su sistema radical causa-do por nemátodos de las agallas (Meloidogyne spp.)[Cuadra et al., 2005].
Regularmente se han visto daños en las hojasatribuibles al ataque de lepidópteros, los que se dife-rencian evidentemente de los de la bibijagua, ya queesta, al comer, hace cortes en forma de media luna enlos bordes de las hojas, y los lepidópteros comen encualquier lugar de ella, sin que hayan sido identifica-dos.
En el municipio de Boyeros, en Ciudad de La Haba-na, se encontraron sobre plantas de noni de tres me-
ses de sembradas larvas de un lepidóptero de colorverdoso a marrón, con una cabeza semejante a la deun perro y cuyas características correspondían a unalarva de «primavera» perteneciente a la familiaSphingidae; en plantas de más edad se hallaron lar-vas conocidas comúnmente como ositos peludos, de colormarrón claro de la familia Arctiidae, por lo que se pro-cedió a obtener los adultos de ambas especies en ellaboratorio para su posterior identificación mediantecomparación con los ejemplares de la colección de in-sectos del INIFAT.
Los adultos de la «primavera» (Fig. 1) correspondie-ron a la especie Xylophanes pluto (F.), informada porBruner (1975) sobre plantas de jibá (Erythroxylonhavanense Jacq.). Los del osito peludo (Fig. 2) corres-pondieron a la especie Maenas jussiaeae (Poey), in-formada por Bruner (1975) sobre bagá Spilosoma sp.,ocuje (Calophyllum sp.), frutabomba (Carica papa-ya Lin.), girasol (Helianthus annuus Lin.), Ipomoeasp., clavellina (Jussiaea repens L.), Musa spp.,higuereta (Ricinus communis Lin.), aguinaldo blan-co (Rivea corymbosa (L.)), may. f., Rosa spp., por loque este trabajo constituye el primer informe delataque de estos lepidópteros en Cuba sobre las plan-tas del noni.
Las larvas de ambas especies son voraces y puedenconsumir grandes cantidades del área foliar parasu desarrollo, lo que retrasa el crecimiento de lasplantas, de ahí que se recomiende tomar las medi-das adecuadas al aparecer los primeros síntomas dehojas comidas. Pueden realizarse eliminacionesmanuales de las larvas, en caso de tratarse de pocasplantas, o en plantaciones mayores la utilización decontroles biológicos apropiados.

González y otros
76/fitosanidad
REFERENCIAS
Bruner,S. C.; L. C. Scaramuzza; A. R. Otero: Catálogo de los insectosque atacan a las plantas económicas de Cuba, Academia de Cien-cias de Cuba, 1975.
Cuadra, R.: «El noni (Morinda citrifolia L.), una planta altamente sus-ceptible al ataque de los nemátodos de las agallas», X JornadaCientífica, INIFAT, mar. 31-abr. 1, La Habana, 2005.
Hidalgo, G.: «Observaciones sobre las plagas y las enfermedades delnoni (Morinda citrifolia L.) (Rubiaceae) en Panamá», Seminario so-bre el Cultivo del Noni y la Agricultura Orgánica en Panamá, Facultadde Ciencias Agropecuarias de la Universidad de Panamá, 2004.
Rodríguez, N. A.; P. Sánchez: Especies frutales cultivadas en Cubaen la agricultura urbana, Agrinfor, La Habana, 2004.
Roig, J. T.: Diccionario de nombres vulgares de Cuba, t. 2, EditoraRevolucionaria, La Habana, 1965.
––––: Plantas medicinales aromáticas o venenosas de Cuba, EditoraCiencia y Técnica, La Habana, 1974.
Solomon, N.: El jugo Tahitian Noni, cuánto, con qué frecuencia, paraqué, Direct Source Publishing,Fibromialgia x Enfermedades Reumá-ticas 2005.
Figura 1. Xilophanes pluto, «primavera»encontrada en la planta de noni Morindacitrifolia.
Figura 2. Maenas jussiaeae «osito peludo»encontrado en la planta de noni Morindacitrifolia.

fitosanidad/77
FITOSANIDAD vol. 10, no. 1, marzo 2006
Res
umen
de
tesi
sCONSERVACIÓN DE ARTRÓPODOS BENÉFICOS EN UN SISTE-MA DE PRODUCCIÓN AGRÍCOLA URBANO
Yaril Matienzo Brito
Instituto de Investigaciones de Sanidad Vegetal. Calle 110 no. 514 e/ 5a. B y 5a. F, Playa, Ciudadde La Habana, CP 11600, [email protected]
Tesis en opción al grado académico de Máster en Agroecología y Agricultura Sostenible
Para lograr un avance eficiente en la utilización delcontrol biológico por conservación de especies benéfi-cas, es preciso realizar cambios significativos en los sis-temas agrícolas y desarrollar tecnologías de manejo deorganismos nocivos con enfoque agroecológico. Estu-diar la diversidad y el incremento de las poblaciones deartrópodos benéficos con la utilización de técnicas deconservación en un sistema de producción agrícola ur-bano, ha sido el objetivo de este trabajo de investiga-ción, realizado en el período comprendido de enero del2002 a marzo del 2004 en la UBPC Organopónico Vive-ro Alamar, perteneciente al municipio de La Habanadel Este de la provincia de Ciudad de La Habana. Serealizó una caracterización del subsistema de produc-ción de cultivos varios y se efectuaron muestreos conuna frecuencia semanal, en los que se seleccionaron plan-tas al azar, distribuidas en zig zag, para relacionar lasespecies de artrópodos benéficos y nocivos presentes enlas hortalizas Solanum melongena L., Vigna unguiculata(L.) y Cucumis sativus L., así como en las especies bo-tánicas introducidas como técnicas de conservación. Ladiversidad de artrópodos en el subsistema estudiado se
estimó con la utilización de índices estadísticos de in-formación adaptados a la ecología, se realizó un análi-sis de las relaciones de similitud en las comunidades deespecies de artrópodos en las diferentes etapas del tra-bajo y se evaluó la actividad biorreguladora de los ar-trópodos benéficos mediante los indicadores relacióndepredador-presa y porcentaje de parasitoidismo. Seevidenció el bajo nivel de conocimiento del personal vin-culado a la producción en relación con la utilización detécnicas conservacionistas, y que la inapropiada distri-bución de cultivos y la implementación de prácticasagronómicas inadecuadas limitaban la presencia y ac-tividad de los artrópodos benéficos. La capacitaciónbrindada posibilitó un mayor conocimiento de estaspoblaciones y de las prácticas que favorecen el mante-nimiento y su preservación en sistemas de producciónagrícola, así como la utilización de prácticas conser-vacionistas; posibilitó también la tendencia hacia unamayor diversidad de especies de artrópodos, en la quelos benéficos mostraron una mayor expresión cuanti-tativa, además de favorecerse su actividad biorre-guladora.

Normas EditorialesFitosanidad tiene como objetivo divulgarde forma sistemática el quehacer de losinvestigadores y especialistas del Insti-tuto de Investigaciones de Sanidad Ve-getal, así como de otros centros del paíso extranjeros, vinculados al trabajo dela sanidad vegetal.El Comité Editorial de esta publicación,que se edita trimestralmente, agradeceel envío de colaboraciones y observa-ciones que ayuden a hacer mejor nues-tra labor.Los artículos deben reflejar los resulta-dos de investigaciones básicas o de apli-cación práctica, asimismo aquellos quese encuentren en proceso de extensióno generalización.Igualmente resultan de importancia lostrabajos en que se comuniquen nuevosprocedimientos o innovaciones, asícomo los que aborden temas novedosos.• Tipo de artículosLa revista acepta manuscritos origina-les (inéditos) en cualquiera de las espe-cialidades directa e indirectamente vin-culadas a la sanidad vegetal. Estospueden ser: artículos científicos, rese-ñas, comunicaciones cortas, resúmenesde tesis, informes técnicos. Los artícu-los científicos no deben exceder de sie-te cuartillas, las reseñas de quince, lascomunicaciones de dos y los informestécnicos de cinco.• LenguajeEl lenguaje oficial de la revista es espa-ñol, aunque excepcionalmente se acep-tarán contribuciones en inglés, francésy portugués. El estilo de escritura debeser totalmente impersonal, con criteriode exactitud, brevedad y párrafos cor-tos. Debe utilizarse el sistema métricodecimal. Los nombres científicos se es-cribirán completos, incluyendo el autor,y siguiendo los códigos internacionales(ejemplo: Leucoptera coffeella GuerinMeneville). Si es necesario utilizarlos envarias partes del texto, entonces se es-cribirán completos la primera vez queaparezcan y luego se abrevian (ejemplo:L. coffeella). Se escriben en cursiva o sesubrayan.• Estructura de los artículosLos artículos científicos tendrán la es-tructura siguiente: título, autor(es),afiliación de los autores, resumen ypalabras claves (español e inglés), intro-ducción, materiales y métodos, resulta-dos y discusión, conclusiones (si lashubiera), agradecimientos (si los hubie-ra), referencias.Las reseñas adoptarán la siguiente es-tructura: título, autor(es), afiliación de
los autores, resumen y palabras claves(español e inglés), introducción, el con-tenido se estructura a criterio del au-tor, agradecimientos (si los hubiera),referencias.Las comunicaciones cortas incluirán: tí-tulo, autor(es), afiliación de los auto-res, texto de la comunicación, incluyen-do las referencias principales.La estructura de los informes técnicosserá: título, autor(es), afiliación de losautores, resumen y palabras claves (es-pañol e inglés), introducción, desarro-llo y referencias.• Elaboración del textoEl título debe ser claro y conciso, pro-curando que no sea extenso. Debe te-ner correspondencia con el contenido.No se incluirán abreviaturas.De los autores se escribirán nombres ydos apellidos. Si el autor tiene un se-gundo nombre, este se abrevia con lainicial. La afiliación de los autores seescribirá con su nombre completo, lassiglas sólo se emplearán entre parénte-sis, si lo consideran necesario. Debe in-cluirse la dirección postal, fax y correoelectrónico si los posee.El resumen no debe exceder de 250 pa-labras, y debe tener una síntesis de losmétodos y resultados, mencionándoselos nombres científicos completos yvalores cuantitativos de los resultados,es decir, debe tener el contenido sufi-ciente de forma comprimida.Las palabras claves son aquellas que per-miten identificar el contenido del artí-culo y que facilitan la identificación enlos índices de materia. Debe incluir lastaxas de los entes biológicos.La introducción debe tener una brevereferencia de los antecedentes específi-cos del trabajo, así como una revisiónbreve de las referencias más recientesque se relacionan con el tema que sepresenta. También se incluirá el objeti-vo del trabajo.Los materiales y métodos deben ser cla-ros y concretos, se redactarán según unorden lógico de los métodos empleados.De igual forma se escribirán claramentelos procedimientos analíticos y estadísti-cos utilizados. Se pueden citar los méto-dos y procedimientos, siempre que ha-yan sido publicados en revistas científicas.Los resultados se pueden expresar apo-yados en tablas y/o figuras, con una dis-cusión a partir de referencias actuales.Deben presentarse de manera lógica,interpretando las conclusiones. Las fi-guras se deben elaborar solamente enWord u otro programa compatible.
Las referencias sólo serán de publica-ciones disponibles en las bibliotecas, yse presentarán en orden alfabético. Secolocará el primer apellido del autorprincipal y luego las iniciales de los nom-bres; para los demás autores, primerola inicial y luego los apellidos, en todoslos casos separados por comas. Cuandouna obra lleve más de tres autores, sepondrá el apellido y nombre del prime-ro y a continuación et al. (en cursiva). Acontinuación, el título del artículo, elnombre de la revista, así como volumen,número, páginas y año. En el caso delos libros y folletos, después del título,número de la edición, lugar de la publi-cación (ciudad), casa editorial y pági-nas. Los títulos de los artículos y po-nencias se entrecomillarán, mientrasque los referidos a libros y publicacio-nes periódicas irán en cursiva, o en sudefecto subrayados. En el texto las refe-rencias se citan con el apellido del pri-mer autor y el año entre paréntesis; másde un autor se anota como et al.• Envío de manuscritosDeben entregarse un original mecano-grafiado, a doble espacio, en papel blan-co tamaño 28 x 21,5 cm, utilizando unasola cara, con márgenes de dos centíme-tros a los lados y tres en la parte supe-rior e inferior. Cada cuartilla debe serenumerada. La letra que ha de utilizarsedebe ser Arial, con puntaje 11. Además,debe entregar una copia en disquete,utilizando el procesador de texto Word,ya que el consejo de redacción no dispo-ne de capacidades para mecanografiar losartículos. Se acepta el envío de manus-critos por correo electrónico, como do-cumentos adjuntos al mensaje o carta depresentación, siempre que no posean fi-guras. Los manuscritos se enviarán a:Instituto de Investigaciones de SanidadVegetal. Calle 110 no. 514 e/ 5a.B y 5a.F,Playa, Ciudad de La HabanaNo se aceptan manuscritos que no es-tén acompañados de la Declaración delAutor.También pueden enviarse por correoelectrónico: [email protected]• Revisión de los manuscritosLos trabajos enviados al Comité Edito-rial serán sometidos a un proceso dearbitraje y corrección de estilo. Los au-tores colaborarán con los árbitros y co-rrectores a evacuar cualquier duda alrespecto y efectuar, si es preciso, lasmodificaciones que se le sugieran. ElComité Editorial se reserva el derechode aprobar o rechazar los trabajos pro-puestos, lo cual será notificado oportu-namente a los interesados.

fitosanidad/79
FITOSANIDAD vol. 10, no. 1, marzo 2006
EPIDEMIOLOGÍA DE ERWINIA SPP. EN EL CULTIVO DE LA PAPA(SOLANUM TUBEROSUM L.) Y MEDIDAS PARA SU CONTROL
Yolanda Morales Gómez
Facultad de Ciencias Agropecuarias. Departamento de Agronomía, Universidad Central de Las Villas,Carretera a Camajuaní, Km 5½, Santa Clara, Villa Clara, Cuba
Tesis en opción al grado académico de Máster en Sanidad Vegetal
El trabajo se desarrolló en la Estación ExperimentalÁlvaro Barba y el Laboratorio de Fitopatología perte-necientes ambos a la UCLV, y en la Empresa de Culti-vos Varios Valle del Yabú de 1988 a 1996. Se realizaronexperimentos de laboratorio donde se aisló e identificóla bacteria, se llevaron a cabo estudios de resistenciavarietal, influencia de la edad de la planta al cosecharen la manifestación de las pudriciones, fertilización ysu influencia en la manifestación de los síntomas, y es-tudios de infección latente en el peridermo de los tu-bérculos. Se realizaron además investigaciones enfrigoríficos, naves de pilón y agromercados en relación
con Erwinia spp. y su manifestación en los tubérculos,así como estudios de campo.
Se encontró que no existió variedad con respuesta in-mune. Las mayores pudriciones se presentaron a los 60días; las dosis de fertilizantes utilizadas no influyeronen la manifestación de los síntomas; en todas las mues-tras procesadas se detectó infección latente en laslenticelas del tubérculo; se reportaron cuantiosas pér-didas en frigoríficos, naves de pilón y agromercados, ylo que representan desde el punto de vista económico.Se recomendó una metodología para el control de estabacteria en el cultivo de la papa en Cuba.

Rgx
kuvc
Fit
osa
nid
ad
INIS
AX
SO
LICI
TUD
DE
SUSC
RIPC
IÓN
Rgp
qx
ct
uw
ue
tkr
ekô
p
Nw
gxc
uw
uet
kre
kôp
A
evw
cnk|
ct
fc
vqu
Aò
qu
sw
g u
qnk
ekvc
____
____
____
____
____
____
____
____
____
R
RE
CIO
DE
SU
SC
RIR
CIÓ
N
F
qto
cu f
g r
ciq
R
ci
q g
p n
c kp
uvkv
wek
ôp
fkt
ge
vco
gpvg
q r
qt
kpvg
tog
fkc
tkq
Cj
gsw
g g
p M
N<
Ipu
vkvw
vq f
g Ip
xgu
vki
ce
kqp
gu
fg
Sc
pkf
cf
Xg
igv
cn0
Cc
nng 3
32
pq
0 7
36
g1 7
c0 B
{ 7
c0 F
. Rnc
{c.
Ckw
fcf
fg
Lc
Jc
dc
pc
. C
R0
338
22
0
Cw
gp
vc N
q0
:4
4
Cj
gsw
g g
p M
LC
< E
or
tguc
CA
TE
C.
ew
gp
vc B
ICS
A N
q0
5432
33
682
22
S
g c
egr
vcp
ej
gs
wg
u g
p e
wc
nsw
kgt
oq
pg
fc
nkd
tgo
gp
vg e
qp
xgtv
kdng
.
g
zeg
rvq
eq
pvt
c c
ig
pe
kcu
dcp
ec
tkc
u e
qo
rtq
og
vkf
cu
eq
p g
n uk
uvgo
c
p
qtv
gc
og
tke
cpq
fg
rc
iq
u0
N
qo
dtg
{ A
rg
nnkf
qu1
Nc
og
cpf
Sw
tpc
og
00000
00000
000000
000000
00000
000000
00000
0000
00000
00000
00000
00000
0000
000000
00000
00000
0000
00000
00000
00000
00000
00000
00000
00000
0000
00000
000000
000000
00000
000
Ip
uvkv
wek
ôp
1Ip
uvk
vwvk
qp
000
0000
000000
00000
0000
00000
00000
00000
0000
000000
00000
00000
0000
00000
00000
00000
0000
000000
00000
000000
000000
00000
00000
00000
0000
00000
000000
000000
00000
000000
00000
0000
00000
00000
000000
0000
0
Rtq
hguk
ôp
1Rtq
hgu
ukq
p 0
00000
000000
000000
00000
00000
000000
0000
00000
00000
00000
00000
0000
000000
00000
00000
0000
00000
00000
00000
0000
000000
00000
000000
000000
00000
00000
0000
00000
00000
000000
0000
00000
00000
00000
00000
0000
Dkt
geek
ôp
1Af
ftg
uu 0
0000
000000
00000
00000
0000
00000
00000
000000
0000
00000
00000
00000
0000
00000
00000
000000
0000
00000
00000
000000
000000
00000
00000
00000
0000
00000
000000
00000
0000
00000
00000
000000
0000
00000
00000
00000
0000
00
Cô
fki
q R
quv
cn1\
kr C
qf
g 0
00000
00000
00000
000000
00000
00000
0000
00000
00000
000000
0000
00000
00000
000
Ckw
fc
f1C
kv{
0000
00000
000000
00000
00000
00000
000000
000000
00000
000000
00000
0000
00000
00000
000000
0000
00
R
cîu
1Cq
wp
vt{
00000
0000
00000
00000
00000
00000
0000
000000
00000
00000
0000
00000
00000
000000
Tg
ngh0
1Tg
ngr
jq
pg
””
””
””
””
””
””
0 T
gng
hcz
””
””
””
””
”
E
oc
kn ”
””
””
””
””
””
””
””
””
””
””
”0
N
ûo
gtq
fg
Cw
gp
vc {
Ai
gp
ekc
Bc
pe
ctkc
000
00000
00000
00000
000000
0000
0000
000000
00000
0000
00000
00000
000000
0000
0000
000000
000
N
ota
: E
nví
e p
or
corr
eo p
ost
al /
Cad
a su
scri
pció
n te
ndrá
un
acu
se d
e r
ecib
o
Dku
vtkd
we
kôp
pc
ekq
pc
n<
Eje
mp
lar
MN
: $1
0.0
0 –
US
D:
$10
.00
A
nua
l
M
N: $
40
.00
– U
SD
: $4
0.0
0
Ep
xîq
fg
glg
or
nct
uw
gnv
q c
n gz
vgtk
qt<
A
mé
rica
: $ 1
5.0
0
E
uro
pa:
$ 2
0.0
0 R
esto
del
Mu
ndo
: $
25.0
0
Sw
ue
tkr
ekô
p c
pw
cn
{ g
pxî
q c
n gz
vgtk
qt<
A
mé
rica
: $4
5.0
0
E
uro
pa:
$50
.00
R
esto
del
Mu
ndo
: $
55.0
0
(I
nclu
ye c
ost
o d
e e
nví
o)