conference review the evolution of protein interaction ...eprints.gla.ac.uk/152191/1/152191.pdf ·...
TRANSCRIPT

Comparative and Functional GenomicsComp Funct Genom 2004; 5: 79–84.Published online in Wiley InterScience (www.interscience.wiley.com). DOI: 10.1002/cfg.365
Conference Review
The evolution of protein interactionnetworks in regulatory proteins
Gregory D. Amoutzias1, David L. Robertson1 and Erich Bornberg-Bauer2*1School of Biological Sciences, University of Manchester, 2.205 Stopford Building, Oxford Road, Manchester M13 9PT, UK2Bioinformatics Group, Biology Department, University of Munster, Schlossplatz 4, D-48149 Germany
*Correspondence to:Erich Bornberg-Bauer,Bioinformatics Group, BiologyDepartment, University ofMunster, Schlossplatz 4,D-48149 Germany.E-mail: [email protected]
Received: 13 November 2003Revised: 18 November 2003Accepted: 25 November 2003
AbstractInteractions between proteins are essential for intracellular communication. Theyform complex networks which have become an important source for functionalanalysis of proteins. Combining phylogenies with network analysis, we investigatethe evolutionary history of interaction networks from the bHLH, NR and bZIPtranscription-factor families. The bHLH and NR networks show a hub-like structurewith varying γ values. Mutation and gene duplication play an important rolein adding and removing interactions. We conclude that in several of the proteinfamilies that we have studied, networks have primarily arisen by the development ofheterodimerizing transcription factors, from an ancestral gene which interacts withany of the newly emerging proteins but also homodimerizes. Copyright 2004 JohnWiley & Sons, Ltd.
Keywords: protein–protein interaction networks; eukaryotic transcription factors;gene duplication; homo/heterodimerization; small world networks
Introduction
Genome, proteome and other ‘ome’ projects havegenerated a vast amount of data over the lastfew years. These data can be analysed by com-parative analysis. Proteome analysis became mostimportant for studying regulatory proteins, sincemany signalling proteins, from membrane-boundreceptors down to the DNA-binding regulatory ele-ments (transcription factors), interact directly witheach other via so-called protein–protein interac-tions (PPIs). Some of these interactions are ‘unspe-cific’, e.g. both SH3 and SH2 domains interactwith many other domains [8,27]. Since some ofthe interaction partners interact with yet other pro-teins, this gives rise to a ‘network of interactions’where proteins can be imagined as nodes thatare connected by edges, representing their physi-cal interactions. However, interactions can also bevery specific and limited to only few partner pro-teins, e.g. many transcription factors heterodimer-ize with several partners under specific physiolog-ical conditions and expression states, while they
may homodimerize under other conditions. Anexample is the leucine zipper-mediated interaction,e.g. the competing Jun–Fos/Jun–Jun interactionsin the bZIP family [7,19] or the much weakerMad–Max/Max–Max interactions in the bHLHfamily [7,12,17].
The molecular details of dimerization are verycomplicated and are not the focus of this study.Several groups have begun to investigate the‘global’ features of PPI networks, trying to inferfunctional properties by applying statistical meth-ods to the analysis of the networks [9,14,24,25].Among the most intriguing of these findings aretheir small-world characteristics. Although the net-work is very big, each protein is linked to everyother by chains of only a few edges. This is possi-ble because some proteins interact with many oth-ers, representing so-called ‘hubs’, and many haveonly very few interactions [14]. Furthermore, theseinteractions appear to be confined to cellular com-partments [24].
Copyright 2004 John Wiley & Sons, Ltd.

80 G. D. Amoutzias, D. L. Robertson and E. Bornberg-Bauer
The reliability of experimental PPI data is con-troversially discussed [13,22,23]. However, oneshould consider that binding is not an all-or-nothingprocess and at least in vivo interactions are fre-quently competitive, as shown in the bZIP net-work [18]. Furthermore, the combination of datafrom various sources can significantly improvetheir quality. In our research efforts we have col-lated database information with data mined fromthe literature to generate more reliable, ‘confirmed’,datasets [1]. Several models of network evolu-tion have been developed recently [20,21,26,29].Some models are based on gene duplication, someon domain rearrangements. Others assume that anexisting initial network is duplicated when all genescoding for the interacting proteins are duplicatedsimultaneously. This could happen, for example,via a whole-genome duplication or other large-scaleduplication events, such as tandem duplications. Inthe following a certain fraction of interactions isassumed to be lost again.
The goal of this study is to complement exist-ing perspectives on network evolution with stud-ies based on phylogenies and comparative analysisfrom genomic and proteomic data. We have chosento work on several families of eukaryotic transcrip-tion factors for which many data from differentsources are known and for which phylogenies areeither known or can be computed with a reason-able reliability. In particular, we concentrate on thequestion of how the evolution of interaction speci-ficity, such as homo- vs. heterodimerization, mayreveal the evolutionary dynamics of network evo-lution. Accordingly, results on the families of NR,bZIP and bHLH proteins are reported and discussedin the following.
Methods
Interactions for all three networks were extractedfrom a literature search in PubMed, with the focuson mammalian transcription factors (http://www4.ncbi.nlm.nih.gov/PubMed/). Specifically for thebZIPs, interactions were extracted from Newmanand Keating [18], in which protein arrays wereused to test 492 pairings of a nearly complete setof coiled-coil strands from all known human bZIPproteins. We included interactions that were sym-metrical in the consensus interaction matrix, mean-ing that the use of the protein in the array surface or
as a probe was not inhibiting the interaction. Also,we disregarded interactions with Z < 1, which isthe threshold value for the signal : noise ratio. Datafor bHLH proteins are the same as in Amoutziaset al. [1] but with close family members collapsedto one node. For the NR family we excluded alldata for which interactions were shown not tobe direct.
Results
bHLH proteins
We first focus on bHLH proteins. They represent anancient family of transcription factors being presentin all eukaryotic clades, that expanded since theemergence of multicellularity [16]. bHLH proteinsare involved in cell-cycle regulation, metabolicsensing and tissue-specific development. Theirmain constituent is a ≈60 amino acid-long domaincomprising the DNA-binding basic region andthe HLH motif which mediates interaction. How-ever, they usually also contain additional dimeriza-tion domains which contribute to the specificationof homo- and heterodimerization. While furtherdetails on their evolution are reported elsewhere[4,16], the feature which is most relevant for thisstudy is that five major groups exist. These can bedistinguished by their domain architecture, in par-ticular the presence of the additional dimerizationdomain (leucine zipper — LZ, PAS, Orange), suchthat their clustering into groups is fairly reliable.Recently, we found that PPIs between bHLH pro-teins form two hub-based networks. One of themcan be divided into two hub-based subnetworks.The two networks have a striking similarity in theirtopology but no interactions between any memberof one family to any member of the other familyare known to exist [1].
Analysing these networks of protein families(Figure 1), the most prominent feature appeared tobe that, just like the overall network, they havea hub-like topology. Interaction data have beenobtained from a number of different sources andthe low number of interactions for the majorityof the nodes has been explicitely confirmed byexperiments on these nodes. Hubs are a feature ofscale-free networks and such properties have beenobserved in social networks, the world-wide web,the western US power grid, citations of scientific
Copyright 2004 John Wiley & Sons, Ltd. Comp Funct Genom 2004; 5: 79–84.

Evolution of protein interaction networks in regulatory proteins 81
RAR
FXR
LXR
COUP
VDR
TR
PPAR
NGFIB
RXR
ERERR
B
Max MlxMad
Mnt
Mga
MondoAc-Myc
Id1
MyoDMist1
NeuroD
Ngn2
Math1
Twist
Tal1
Nscl1N-Twist
Paraxis
eHand
Usf1
Bmal1
Hey2
Dec1
Hes5
Hes6
Hes1
Amt
Npas2Hif1
Sim1
Npas1
Ahr
E2A
Ash1
A
S-Maf
Par-b
FosJun
Creb
Atf6
C/ebp
Oasis
Atf4
Atf2Cnc
L-MafPar-a
C
Figure 1. Protein interaction networks of eukaryotic transcription factors: bHLH (A), NR (B) and bZIP (C).Homodimerization is indicated by larger dots
publications, metabolic networks, protein domains,protein interaction networks and the distributionof proteins in sequence space [3,5,6,31,32]. Forthe bHLH protein-interaction network, we calcu-lated the frequency of nodes with K interactions,P(K ), and plotted this against the number of inter-actions, K .
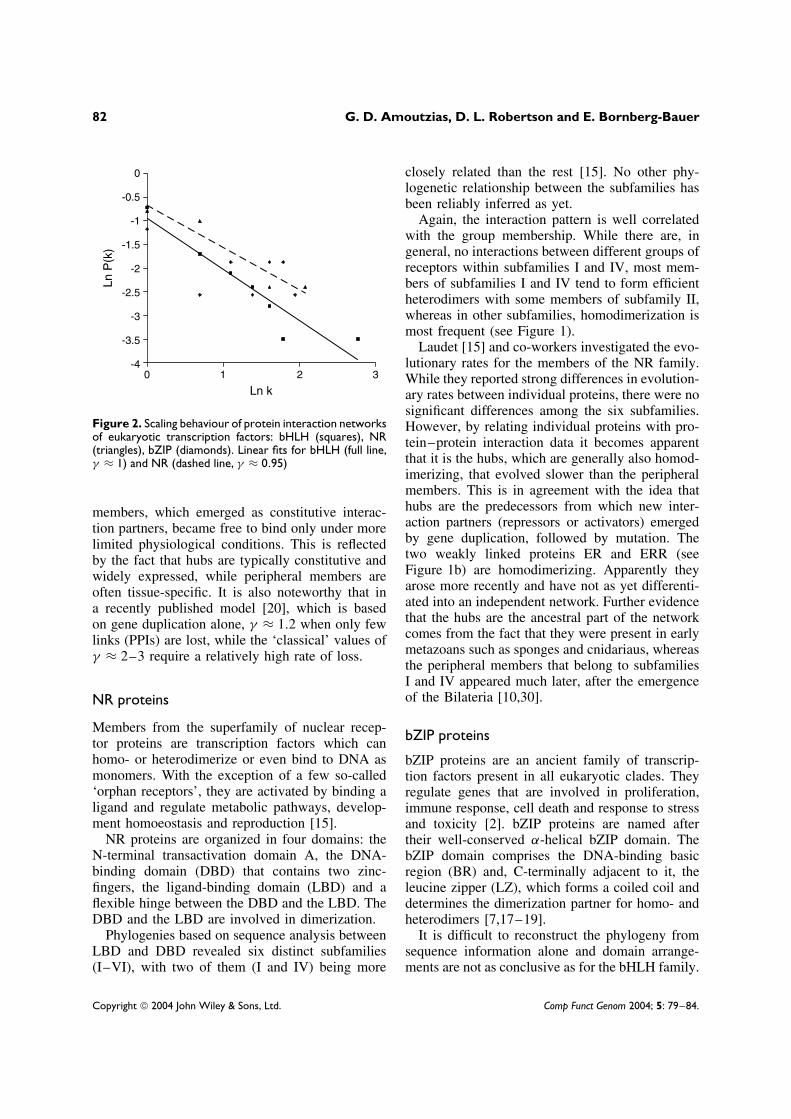
The plot (Figure 2.) of ln[P(K )] − ln(K ) showsclearly that the bHLH PPI network is scale-free,because the distribution of connectivity decays asa power-law P(K ) = K −γ , where γ ≈ 1. A scale-free network is a non-homogeneous network witha few highly connected nodes (the hubs) and many
poorly connected nodes (the peripheral members).Other analyses on scale-free networks estimatedthat γ is usually in the range 2–3 [11]. The biologi-cal significance of this lower value for γ appears tobe a direct consequence of the fact that gene dupli-cation events (single or large-scale) have generatednew peripheral proteins that then interact preferen-tially with the hub. Apparently the homodimerizingfactors are the most highly linked (at least withintheir networks) and represent hubs (Figure 2). Con-sequently, we conclude that homodimerization wasthe ancestral function. This feature was maintainedwith the emergent hub, even though the peripheral
Copyright 2004 John Wiley & Sons, Ltd. Comp Funct Genom 2004; 5: 79–84.
82 G. D. Amoutzias, D. L. Robertson and E. Bornberg-Bauer
-4
-3.5
-3
-2.5
-2
-1.5
-1
-0.5
0
Ln k
Ln P
(k)
1 2 30
Figure 2. Scaling behaviour of protein interaction networksof eukaryotic transcription factors: bHLH (squares), NR(triangles), bZIP (diamonds). Linear fits for bHLH (full line,γ ≈ 1) and NR (dashed line, γ ≈ 0.95)
members, which emerged as constitutive interac-tion partners, became free to bind only under morelimited physiological conditions. This is reflectedby the fact that hubs are typically constitutive andwidely expressed, while peripheral members areoften tissue-specific. It is also noteworthy that ina recently published model [20], which is basedon gene duplication alone, γ ≈ 1.2 when only fewlinks (PPIs) are lost, while the ‘classical’ values ofγ ≈ 2–3 require a relatively high rate of loss.
NR proteins
Members from the superfamily of nuclear recep-tor proteins are transcription factors which canhomo- or heterodimerize or even bind to DNA asmonomers. With the exception of a few so-called‘orphan receptors’, they are activated by binding aligand and regulate metabolic pathways, develop-ment homoeostasis and reproduction [15].
NR proteins are organized in four domains: theN-terminal transactivation domain A, the DNA-binding domain (DBD) that contains two zinc-fingers, the ligand-binding domain (LBD) and aflexible hinge between the DBD and the LBD. TheDBD and the LBD are involved in dimerization.
Phylogenies based on sequence analysis betweenLBD and DBD revealed six distinct subfamilies(I–VI), with two of them (I and IV) being more
closely related than the rest [15]. No other phy-logenetic relationship between the subfamilies hasbeen reliably inferred as yet.
Again, the interaction pattern is well correlatedwith the group membership. While there are, ingeneral, no interactions between different groups ofreceptors within subfamilies I and IV, most mem-bers of subfamilies I and IV tend to form efficientheterodimers with some members of subfamily II,whereas in other subfamilies, homodimerization ismost frequent (see Figure 1).
Laudet [15] and co-workers investigated the evo-lutionary rates for the members of the NR family.While they reported strong differences in evolution-ary rates between individual proteins, there were nosignificant differences among the six subfamilies.However, by relating individual proteins with pro-tein–protein interaction data it becomes apparentthat it is the hubs, which are generally also homod-imerizing, that evolved slower than the peripheralmembers. This is in agreement with the idea thathubs are the predecessors from which new inter-action partners (repressors or activators) emergedby gene duplication, followed by mutation. Thetwo weakly linked proteins ER and ERR (seeFigure 1b) are homodimerizing. Apparently theyarose more recently and have not as yet differenti-ated into an independent network. Further evidencethat the hubs are the ancestral part of the networkcomes from the fact that they were present in earlymetazoans such as sponges and cnidariaus, whereasthe peripheral members that belong to subfamiliesI and IV appeared much later, after the emergenceof the Bilateria [10,30].
bZIP proteins
bZIP proteins are an ancient family of transcrip-tion factors present in all eukaryotic clades. Theyregulate genes that are involved in proliferation,immune response, cell death and response to stressand toxicity [2]. bZIP proteins are named aftertheir well-conserved α-helical bZIP domain. ThebZIP domain comprises the DNA-binding basicregion (BR) and, C-terminally adjacent to it, theleucine zipper (LZ), which forms a coiled coil anddetermines the dimerization partner for homo- andheterodimers [7,17–19].
It is difficult to reconstruct the phylogeny fromsequence information alone and domain arrange-ments are not as conclusive as for the bHLH family.
Copyright 2004 John Wiley & Sons, Ltd. Comp Funct Genom 2004; 5: 79–84.

Evolution of protein interaction networks in regulatory proteins 83
However, a classification has been suggested, basedon the amino acid composition and on dimeriza-tion partners [28]. More comprehensive interac-tion data from bZIP proteins have been analysedmost recently, using protein chip technology [18].Applying network analysis as above reveals a moreeven distribution of connectivities; however, thereis still a fair amount of clustering (Figure 2). Thisis an indication of an evolutionary mechanism forthis family which is different from the bHLH andNR families. Also, there is no such clear differenti-ation between homo- and heterodimerization, sincemost proteins have at least a limited capacity tohomodimerize.
Discussion
In this study we have combined network analysisand phylogenies to investigate the emergence ofnew interactions in the gene networks of eukaryotictranscription factors.
In all three families we have studied (bHLH,bZIP and NR), there is an indication that homod-imerization preceded the development of het-erodimerization. In NRs, strong evidence comesfrom phylogenetic studies and the distribution ofNR families in early metazoans [10,30]. The evo-lution of the bHLH networks is also consistent withthe ancestral nature of homodimerization. Typ-ically, the ability for homodimerization appearsto be conserved, such that hubs emerge fromthe ancestral homodimerizing proteins. Subsequentgene duplication (large- or small-scale) and muta-tion results in changes in dimerization properties,thus forming a complex network. In particular, thebHLH family have apparently evolved by repeatedsingle-gene duplications which led to the initialnetwork topology [1]. Subsequent large-scale geneduplications may have increased the complexity ofthe bHLH network. While the role of gene duplica-tion was also important in the evolution of the NRand bZIP networks, the central role of single geneduplication cannot be confirmed with current data.
The basic principles, i.e. the hub-like structureof the interaction networks, comply with the globalfeatures as they were shown by other groups. How-ever, the statistical properties (γ ) for the sub-networks differ between the families and deviatemore or less from the global properties of PPInetworks as they have been analysed previously.
Obviously, these differences reflect different evo-lutionary dynamics, such as the relative frequencyof gene duplication, large-scale duplication eventsand loss of interactions. The loss of interactionsappears to be particularly important in the initialstages of network development and its influenceon the value of γ appears to be in good agree-ment with the predictions by Pastor-Satorras andco-workers [20].
Our results have obvious implications for theunderstanding of network evolution. Further the-oretical studies and models of network evolutionshould consider these variations in γ and the factthat, at least in many cases, heterodimerizationemerges from homodimerization.
Acknowledgements
GA is the recipient of a CASE studentship from theEPSRC and AstraZeneca plc. EBB acknowledges supportthrough an MRC international recruitment grant. Thanksto Chris Lockwood, Michaela Falb and Anna Divoli forhelpful discussions. GA gratefully acknowledges supportby Dimitris and Vasiliki Amoutzias.
References
1. Amoutzias GD, Robertson DL, Oliver SG, Bornberg-Bauer E.2003. Convergent evolution of gene networks by single-geneduplication in higher eukaryotes (submitted). Supplementarydataon:www.uni-muenster.de/Biologie.Botanik/ebb/bHLH/.
2. Angel P, Karin M. 1991. The role of Jun, Fos and the AP-1 complex in cell proliferation and transformation. BiochimBiophys Acta 1072: 129–157.
3. Apic G, Gough J, Teichmann SA. 2001. Domain combina-tions in archaeal, eubacterial and eukaryotic proteomes. J MolBiol 310: 311–324.
4. Atchley WR, Fitch WM. 1997. A natural classification of thebasic helix–loop–helix class of transcription factors. ProcNatl Acad Sci USA 94: 5172–5176.
5. Barabasi A. 2002. Linked: The New Science of Networks.Perseus Press: Oxford.
6. Bornberg-Bauer E. 2002. Randomness, structural uniqueness,modularity and neutral evolution in sequence space of modelproteins. Z Phys Chem 216: 139–154.
7. Bornberg-Bauer E, Rivals E, Vingron M. 1998. Computa-tional approaches to identify leucine zippers. Nucleic AcidsRes 26: 2740–2746.
8. Brannetti B, Via A, Cestra G, Cesareni G, Helmer-Citterich M. 2000. SH3-SPOT: an algorithm to predict pre-ferred ligands to different members of the SH3 gene family.J Mol Biol 298: 313–328.
9. Bu DB, Zhao Y, Cai L, et al. 2003. Topological structureanalysis of the protein–protein interaction network in buddingyeast. Nucleic Acids Res 31: 2443–2450.
Copyright 2004 John Wiley & Sons, Ltd. Comp Funct Genom 2004; 5: 79–84.

84 G. D. Amoutzias, D. L. Robertson and E. Bornberg-Bauer
10. Escriva H, Safi R, Hanni C, et al. 1997. Ligand binding wasacquired during evolution of nuclear receptors. Proc Natl AcadSci USA 94: 6803–6808.
11. Goh KI, Oh E, Jeong H, Khang B, Kim D. 2002. Classifi-cation of scale-free networks. Proc Natl Acad Sci USA 99:12 583–12 588.
12. Hurst H. 1994. bZIP proteins. Protein Profile 2: 105–106.13. Ito T, Ota K, Kubota H, et al. 2002. Roles for the two-hybrid
system in exploration of the yeast protein interactome. MolCell Proteom 1: 561–566.
14. Jeong H, Mason SP, Barabasi A, Oltvai Z. 2001. Lethalityand centrality in protein networks. Nature 411: 41–42.
15. Laudet V. 1997. Evolution of the nuclear receptor superfam-ily: early diversification from an ancestral orphan receptor. JMol Endocrinol 19: 207–226.
16. Ledent V, Vervoot M. 2001. The basic helix–loop–helixprotein family: comparative genomics and phylogeneticsanalysis. Genome Res 11: 754–770.
17. Muhle-Goll C, Nilges M, Pastore A. 1995. The leucinezippers of the HLH-LZ proteins max and c-myc preferentiallyform heterodimers. Biochemistry 34: 13 554–13 564.
18. Newman JR, Keating AE. 2003. Comprehensive identificationof human bzip interactions with coiled-coil arrays. Science300: 2097–2101.
19. O’Shea EK, Rutkowski R, Kim PS. 1992. Mechanism ofspecificity in the fos-jun oncoprotein heterodimer. Cell 68:699–708.
20. Pastor-Satorras R, Smith E, Sole RV. 2002. Evolving proteininteraction networks through gene duplication. J Theoret Biol222: 199–210.
21. Rzhetsky A, Gomez SM. 2001. Birth of scale-free molecularnetworks and the number of distinct DNA and protein domainsper genome. Bioinformatics 17: 988–996.
22. Salwinski L, Eisenberg D. 2003. Computational methods ofanalysis of protein–protein interactions. Curr Opin Struct Biol13: 377–382.
23. Schachter V. 2002. Bioinformatics of large-scale proteininteraction networks. Biotechniques S16: 16–18.
24. Schwikowski B, Uetz P, Fields S. 2000. A network ofprotein–protein interactions in yeast. Nature Biotechnol 18:1257–1261.
25. Vazquez A, Flammini A, Maritan A, Vespignani A. 2003a.Global protein function prediction from protein–proteininteraction networks. Nature Biotechnol 21: 697–700.
26. Vazquez A, Flammini A, Maritan A, Vespignani A. 2003b.Modeling of protein interaction networks. ComPlexUs 1:38–44.
27. Villanneva J, Fernandez-Ballester G Querol E, Aviles FX,Serrano L. 2003. Lig and screening by exoproteolysis andmass spectrometry in combination with computer modelling.J Mol Biol 330: 1039–1048.
28. Vinson C, Myakishev M, Acharya S, et al. 2002. Classifica-tion of human b-zip proteins based on dimerization properties.Mol Cell Biol 22: 6321–6335.
29. Wagner A. 2003. How the global structure of proteininteraction networks evolves. Proc R Soc Lond B 270:457–466.
30. Wiens M, Batel R, Korzhev M, Muller W. 2003. Retinoid xreceptor and retinoic acid response in the marine spongeSuberites domuncula . J Exp Biol 206: 3261–3271.
31. Wolf YI, Karev G, Koonin EV. 2002. Scale-free networks inbiology: new insights into the fundamentals of evolution.Bioessays 24: 105–109.
32. Wuchty S. 2001. Scale-free behaviour in protein domainnetwork. Mol Biol Evol 18: 1694–1702.
Copyright 2004 John Wiley & Sons, Ltd. Comp Funct Genom 2004; 5: 79–84.

Submit your manuscripts athttp://www.hindawi.com
Hindawi Publishing Corporationhttp://www.hindawi.com Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttp://www.hindawi.com Volume 2014
Hindawi Publishing Corporation http://www.hindawi.com
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttp://www.hindawi.com Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttp://www.hindawi.com Volume 2014
The Scientific World JournalHindawi Publishing Corporation http://www.hindawi.com Volume 2014
Hindawi Publishing Corporationhttp://www.hindawi.com Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttp://www.hindawi.com Volume 2014
Hindawi Publishing Corporationhttp://www.hindawi.com Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttp://www.hindawi.com Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttp://www.hindawi.com Volume 2014
Hindawi Publishing Corporationhttp://www.hindawi.com Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttp://www.hindawi.com Volume 2014
Hindawi Publishing Corporationhttp://www.hindawi.com Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttp://www.hindawi.com Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttp://www.hindawi.com
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttp://www.hindawi.com Volume 2014
Hindawi Publishing Corporationhttp://www.hindawi.com Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttp://www.hindawi.com Volume 2014
International Journal of
Microbiology