characterization of human adenovirus type 5 early region
TRANSCRIPT

CHARACTERIZATION. OF THE HOMAN ADENOVIRUS
TYPE 5 EARLY REGION IB 1.76R PROTEIN
,.
By
CATHERINE JANE MCGLADE
A Thesis
Submitted to the School of Graduate Studies
in Partial Fulfilment of the Requirements
for the Degree
Doctor of Philosophy
McMaster University
t' .
."i "

c CHARACTERIZATION OF THE HUMAN ADENOVIRUSTYPE 5 EARLY REGION 18 176R PROTEIN
,.'
.-.. ' ~

DOCTOR OF PHiLOSOPHY (1990)(Medical Science)
McMaster universityHamilton, ontario
TITLE:
AUTHOR:
Characterization of Human AdenovirusType 5 Early Region lB 176R Protein
Catherine Jane McGlade
SUPERVISOR: Dr. Philip E. Branton
.'
NUMBER OF PAGES: 238
.'
,",<.
(I
ii
c'

::..:
ABSTRACT
Cellular t~ansformation by human adenoviruses requires the
expression of two early transcription units, E~ (0 to 4.5% of
the genome) and EIB (4.5 to 11.2%). The products of both genes
are required to disrupt normal cellular growth control and
morphology. Early region 1A products alone can induce cellular
immortalization, but establishment of fully transformed cells
requires cooperation with EIB. Early region IB produces two
unrelated proteins, 496R and 176R, each of which can act
independently with E1A to induce cellular transformation.
176R is a membrane associated protein whose molecular role
in transformation is not known. Anti-peptide sera specific for. ~---
the amino- and carboxy-termini of 176R were produced. These two
sera were used for further purification and biochemical
characterization of the protein. The 176R migrated as two species
termed 18.5 and 19K on 5DS-PAGE • The "generatio~\ of the two
species was not due to proteolysis, nor any post-translational
modification investigated.
Two post-translational modifications of 176R were
identified: phosphorylation and acylation. The phosphorylation
site was mapped to serine residue 164, and a mutant virus in
which this amino acid was altered, ~2204, was produced by
oligonucleotide-directed mutagenesis. Characterization of ~2204
iii

as well as an El plasmid carrying this mutation suggested that
phosphorylation is not important for the biological activity of
176R. 176R was also found to contain covalently bound palmitate
and myristate. Both tryptic and chymotryptic peptide mapping as
well as CNBr cleavage were used to localize the acylation site.
These experiments suggested that multiple acylation sites exist,
and at least one site was localized to the amino terminal region"-by tryptic peptide mapping. The linkage of fatty acid to 176R was
o found to be via an amide, or an unusually stable ester bond to an
internal amino acid. Acylation of 176R likely facilitates its,~
membrane association, but it was not possible to test\this"
hypothesis without precise mapping of the acylation site(s) and
construction of specific mutants.
/''..
CJ
c
iv

\.
l' ~I ',',~
c
ACKNOWLEDGEMENTS
I am deeply indebted to my supervisor Dr. Philip Branton,
not only for his constant support and encouragement throughout
the course of my Ph.D. work, but for making the experience one
which I will look back on with fond memories. I am especially
grateful for his advice and friendship which helped me through
some frustrating times and made completion of this work possible.
Thanks also to Dr. Stan Bayley and Dr. David Johnson for
their advice over the years and for their careful ~~ading of this
thesis.
To Siu-Pok Yee, a special thank you for being so generous~
with his time and knowledge while getting me started in the lab.
I am also indebted to Michel Temblay and Bruce Rowley for
their friendship and for many helpful discussions. Their
suggestions and advice contributed greatly to this work •
.r am also grateful to the other·-members of the lab, past and
present, with whom I spent many enjoyable hours having a few.~;
beers and talking science.""
It was a pleasure to work in the cooperative environment of
the Cancer Research Group and I thank all its members, both
faculty and students, f~r many helpful discussions.
Finally, I would like to thank my parents, who gave me the
confidence to pursue this work, and Jim who had to put up with me
while I completed it.
v
.'..,\'

c:
This thesis is dedicated to
my parents,
and to Jim.;~".
ex, (, ..':-
S\
,.
",:
.-
Q
l'
o
vi

TABLE OF CONTENTS
Page
126
1113
172224
2529 "
35
41
531.3 Thesis proposal •••••••••••••.•••••••••••••••••
1.2 Cellular transformation by adenoviruses.1.2.1 General features •••••••••••••••••••••••1.2.2 Transforming genes •••••••••••••••••••••1.2.3 Role of E1 products
a) structure and function of E1Apolypeptides••••••••••••••••••.••.•••
b) structure and function of E1Bpolypeptides •••••••••••.••••.••••••.•
Chapter 1: Introduction.1.1 Adenovirology.
1.1.1 Classification of human adenoviruses •••1.1.2 Structure of the virion••••••••••••.••.1.1.3 Organization of the genome•••••••••••••1.1.4 Lytic infection by adenoviruses
a} adsorption and uncoating••••.••••••••b) early gene expression••••••••••••••••c) replication and the early to late
switch••••••••....••.•.•••...•.•••••.d) asseJDbly•••••••••••••••••••••••••••••e) cytopathology .
Chapter 2: Materials and Methods. ,2.1 Cells•••••• of 552 • 2 Viruses ~ '. •• 552.3 Infection. • • • • • • • • • • • • • • .. • • . • • • • • • • • • • • • •• 562.4 Synthetic peptide antisera••••••••••••••••••• 562.5 Radioactive labeling••.•••••••••••••••••••••• 572.6 Immunoprecipitation•••••••••••••••••••••.•••• 592.7 Polyacrylamide gel electrophoresis••••••••••• 602.8 western immunoblot••••••••••••••••••••••••••• 612.9 Immunofluorescence••••••••••••••••••••••••••• 622.10 Cleveland peptide mapping••••••••••••••••••• 632.11 Protein extraction from acrylamide••••••••••• 642.12 Phosphoamino acid analysis •••••••••••••••••••.,652.13 Chemical cleavage of proteins using CNBr•••••· 652.14 Immunoprecipitation of v-a digests ••••••••••• 66
2.15 DNA cloning and maniplation.2.15.1 bacterial strains and culture••••••••• 662.15.2 restriction digests, ligation reactions,
vii

=:
agarose and acrylamide gels for DNA,transformation of bacteria and DNA. .m~n~-pre~s. • • • • • • • • • • • • • • • • • • • • • • • • • • •• 67
2.15.3 large scale preparation of plasmidDNA by CsCl Banding•••••••••••••••••••• 68
2.15.4 isolation of DNA fragments fromacrylamide gels•..•......•..•.....•.... 69
2.16 site directed mutagenesis ••••••••••••••••••••. 702.17 Rescue of mutations into pXC38 •••••••••••••••• 722.18 Mutant virus rescue.
2.18.1 transfection of 293 cells •••••••••••••• 732.18.2 i~olation and screening of recombinant
v~ruses. . • • . . . . • . . • • . . . • • . . . . • . . . . . . . •. 742.19 Transformation assay.
2.19.1 preparation of BRK cells••••••••••••••• 752.19.2 transfection of BRK cells and scoring
.transformants. • • • • • • • • • • • • • • • • • • • • • • • •• 762.20 Biochemical characterization fatty acid
protein interaction.2.20.1 hydroxylamine treatment of gels •••••••• 772.20.2 methanol-KOH treatment of gels ••••••••• 77
2.21 Methanol hydrolysis and analysis of fattyacids by HPLC 78
2.22 Peptide mapping by HPLC.2.22.1 Performic acid oxidation and protease
digestion. . . . . . . . . . . . . . . . . . . . . . . . . . . . .. 792.22.2 Separation of peptide by reverse-
phase HPLC.............................. 792.22.3 ~solation ~f.T1 ~sing 19-N1
1mmunoprec~p1tat1on•••••••••••••••••••• 802.23 Ion exchange chromatography of CNBr cleavage
products. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. 81
Chapter 3: Analysis of AdS r/6R using antipeptide sera.3.1 preparation of antipeptide sera •••••••••••••••• 823.2 Immunoprecipitation of 176R polypeptides••.••••• 853.3 Cleveland peptide mapping of the 18.5 and' '::.
19 kDa species of 176R••••••••••••••••••••••••• 883.4 western blot analysis of 176R polypeptides
produced in bacteria••••••••••••••••••••••••••• 913.5 Localization of 176R by indirect
immunofluorescent antibody staining•••••••••••• 943.6 Glycosylation as a possible post-
translational modification••••••••••••••••••••• 94
Chapter 4: Phosphorylation of AdS 176R.4.1 Detection of phosphorylated forms of 176R•••••• I004.2 Identification of the phosphorylated amino
viii

acid in 176R l034.3 Estimation of the level of 176R
phosphorylation••••..•••.•••..•.••........••... l034.4 Mapping of the phosphorylation site.
4.4.1 chemical cleavage with CNBr••.•••••••••• 1064.4.2 digestion with V-8 protease and
immunoprecipitation••••••••••••••••••••• 1124.5 confirmation of ser-164 phosphorylation
site by site-directed mutagenesis.4.5.1 oligonucleotide directed
mutagenesis•...•..•..••...•.••......••.• 1184.5.2 rescue of mutation into pXC38 ••••••••••• 1234.5.3 rescue of mutation into infectious
adenovirus type 5•••••..•••••..••••.•••• 1264.6 Biological role of phosphorylation of 176R.
4.6.1 protein stability••••••••••••••••••••••• 1354.6.2 virus growth, and cyt/deg phenotype .•••• 1354.6.3 cellular transformation••••••••••••••••• 138
4.7 Rescue of mutation into a 496R- background.4.7.1 subcloning of mutations into pXC38 •••••• 1384.7.2 cellular transformation•••• ~ •••••••••••• 143
Chapter 5: Acylation of Ad5 176R.5.1 Detection of acylated forms of 176R•••••••••••• 1495.2 Identification of the fatty acid bound
to 176R••••••••.••••••••••••••••••••.••••.•••.• 1525.3 Analysis of the protein-fatty acid linkage
5.3.1 hydroxylamine treatment of 176R••••••••• 1555.3.2 chloroform:methanol extraction and
treatment with KOH•••••••••••••••••••••• 1565.4 Localization of the acylation site.
5.4.1 CNBr cleavage••••••••••••••••••••••••••• 1615.4.2 analysis of 176R tryptic p~ptides••••••• 164 ~5.4.3 ion-exchange chromatography of
176R CNBr cleavage products ••••••••••••• 178~> 5". 4 ~ 4 analysis of 176R chymotryptic
peptides 184
.' Chapter 6:' DisCussion••••••••••••••••••••••••••••••••••• 192·
REFEREtICES •••••••••••••••••••••••••••••••••••••••••••••• 213
ix

Figure No.
1
2
3
5
6
7
8
9
10
11
LIST OF FIGURES
Title
Virion structure of human adenovirus.
strllcture of the adenovirus genome.
Transcription map of the E1 region.
Functional domains of the E1a polypeptides.
Transcription map of the E1B region.
Predicted amino and carboXY,terminalsequences of AdS E1B-176R andsynthetic peptides.
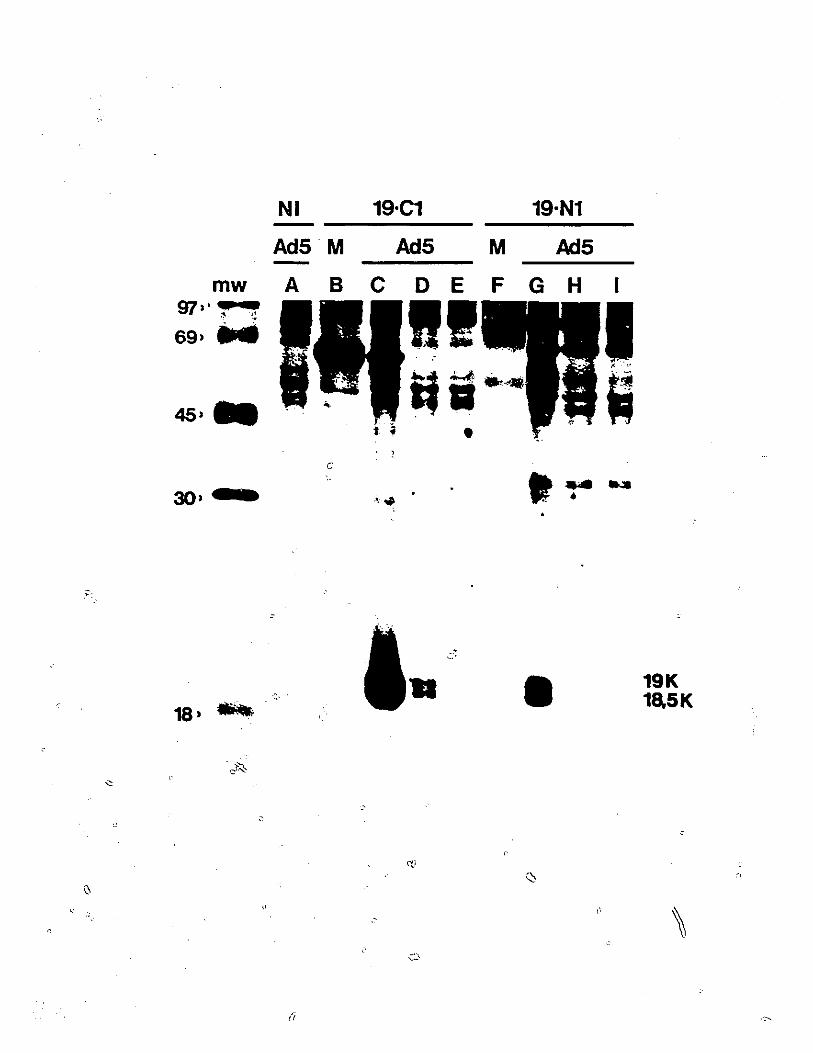
Immunoprecipitation of 176R with19-N1 and 19-Cl antipeptide sera.
Partial proteolysis peptide mappingof the 18.5 and 19 kDa.176R species.
Western blot "analysis of 176Rproteins produced in bacteria.
Indirect immunofluorescent stainingusing 19-C1 serum.
Effect of tunicamycin treatment on176R species.
Page
4
9
30
36
42
83
86
89
92
95
97
12
13
'.i
I ~ J
SDS-PAGE of 176R time course labelingwith [35s]methionine and [32p]orthophosphate.
Two-dimensional phosphoamino acidanalysis.
x
101
104

14
15
'16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
Amino Acid sequence of Ad5 E1B-176Rshowing CNBr and V-8 proteasecleavage sites.
Cleavage pattern of 176R by CNBr.
Cleavage pattern of 176R with s.aureus V-8 protease.
v-a protease cleavage pattern of3H-amino acid labeled 176R.
Generation of mutant pm2204.
Single lane sequencing of M13 phage.
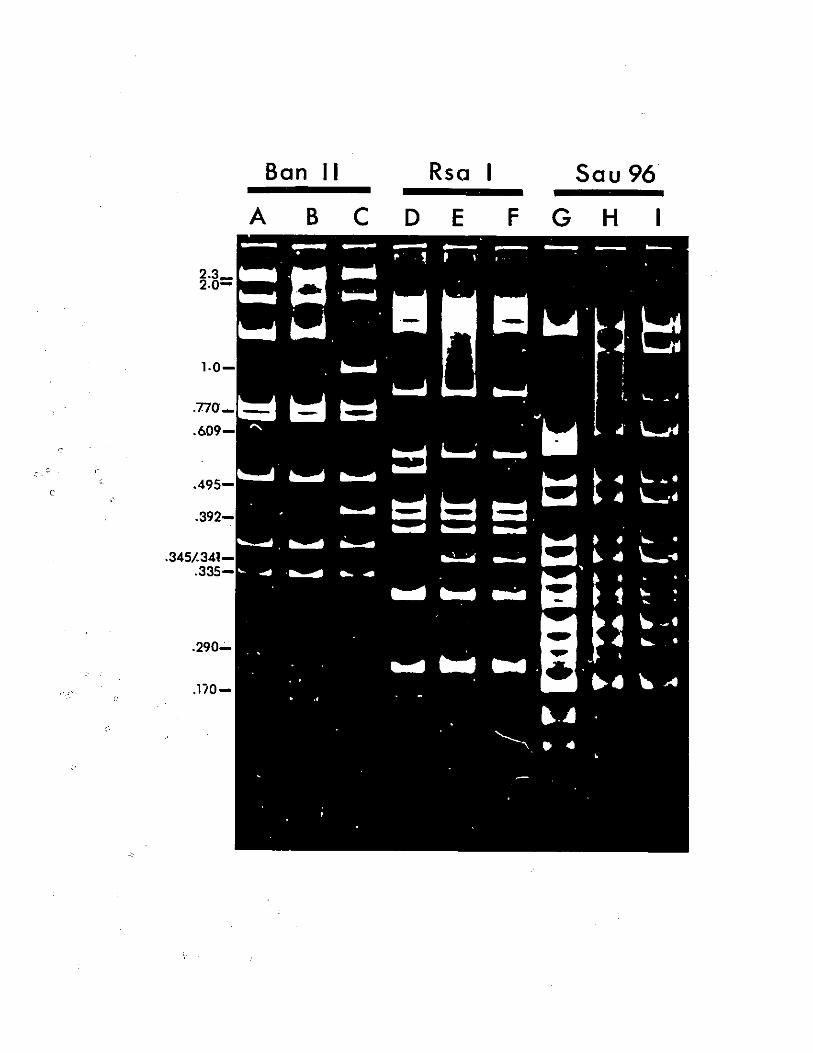
BanII digest of recombinant pXC38.
southern blot of recombinant Ad5plaques.
Restriction digests of recombinantAd5 DNA.
Phosphorylation of 176R in cellsinfected by wt Ad5 or mutant pm2204.
Pulse-chase labeling of 176R fromwt Ad5 or pm2204 infected cells.
Analysis of Hirt extracted viral DNAto detect the deg+ phenotype.
Generation of pXC2204(!j4).
Restriction analysis of pXC2204(lj4).
Labeling of 176R with (3H]fatty acids.
Analysis of the fatty acid presentin 176R.
Sensitivity of fatty acid.linkage toliydroxylamine (HA) treatment.
Sensitivity of fatty acid linkage tomethanol-KOH treatment.
CNBr cleavage of (3H]myristate labeled176R.
Trypsin cleavage sites of Ad5 176R.
xi
108
110
114
116
119
121
124
127
130
133
136
140
144
146
150
153
157
159
162
165

34
\~
35
36
37
38
39
40
"
Separation of t~tic peptides of 176Rlabelled with [3 ]methionine [35]_cysteine, [3H]arginine and [~H]lysine.
Identification of T1 by immunoprecipitation.
Tryptic peptides of the 176R mutant \)pm2072.
Separation of [3H]myristate labelled176R tryptic peptides.
Ion-exchange chromatography of 176RCNBr cleavage products.
Chymotryptic cleavage sites of Ad5176R.
separation of 176R chymotrypticpeptides.
I,'
(1
xii
170
174
176
179
182
185
188

Table No.
1
2
3
4
S
,... :':.
i·
F
LIST OF TABLES
Title
Estimation of the level ofphosphorylation of ~76R.
Plaguing efficiency on HeLa and293 cells.
Transformation of. baby rat kidneycells.
Tr}~tic peptides of AdS-E1B 176R.
Chymotryptic peptides of AdS-E1B 176R.
r
Page
~07
139
142
167
187
(,
.~.:""--::-
//..:::-;;

AdS
Ad2--.
Ad12
BSA
dATP,\\
.::cDNA
dCTP
CNBr
CPE
DMSO
DNA
EDTA
EtBr
HA
.-~ -:;";' ::~.:' ... :.......
.-'
"
HPLC
KOH
mRNA
min
m.u.
",Ci
:
LI:ST OF ABBREVIATIONS
adenovirus type 5
adenovirus type 2
adenovirus type 12
bovine st:rum albumin -
deoxyadenosine S' triphosphate
complimentary deoxyribonucleic acid
deoxycytosine S' triphosphate
cyanogen bromide
cytopathic effect
dimethylsulfoxide
deoxyribonucleic acid
ethylenediaminetetraacetic acid
ethidium bromide
hydroxyl amine
high performance liquid chromatography
potassium hydroxide
mess~nger ribonucleic acid
minute.,','
map unit
micro-CUrie
microgram
microlitre
xiv