chapter 2. the beginnings of genomic biology classical ... · chapter 2. the beginnings of genomic...
TRANSCRIPT

Chapter 2. The beginnings of Genomic Biology –
Classical Genetics Contents
2. The beginnings of Genomic Biology – classical
genetics 2.1. Mendel & Darwin – traits are conditioned by genes
2.2. Genes are carried on chromosomes
2.3. The chromosomal theory of inheritance
2.4. Additional Complexity of Mendelian Inheritance
2.4.1. Multiple alleles
2.4.2. Incomplete dominance and co-dominance
2.4.3. Sex linked inheritance
2.4.4. Epistasis
2.4.5. Epigenetics
2.5. Genes on the Same Chromosome are Linked
2.5.1. Meiosis: chromosomes assort independently
2.5.2. Mapping genes on chromosomes
2.6. Quantitative Genetics: Traits that are Continuously Variable
2.7. Population Genetics: Traits in groups of individuals
It should be clear that the beginings of genomic biology are grounded in classical or Mendelian Genetics. Once the relationship between traits and genes was understood, the relationship between cells and genetics was investigated, leading to the discovery of chromosomes, and a quest for the substance that carried the genetic information began, culminating in the discovery of DNA. These studies constitute the contribution of classical genetics to the founding of the genomic era.
CHAPTER 2. THE BEGINNINGS OF GENOMIC
BIOLOGY –CLASSICAL GENETICS
CONCEPTS OF GENOMIC BIOLOGY Page 1
(RETURN)
Charles Darwin
Gregor Mendel
2.1. MENDEL & DARWIN –
TRAITS ARE CONDITIONDBY GENES.
The idea of genomic biology begins with a consideration of what makes up genomes. Specifically what are genes. The timeline of genetics and genomics begins with the early work of Charles Darwin and Gregor Mendel who didn’t really talk about genes per se, but who did describe the behavior of the characteristics of biological
organisms, which they referred to as traits.
In 1859 Charles Darwin published his book On the Origin of Species. In this work Darwin described a mass of descriptive support for the concept that “traits” are stably transmitted through subsequent generations, and that organisms that have superior traits survive their natural environment to pass those traits on to the next generation. However, Darwin did not describe any mechanism for such transmission of traits to the next generation.
Experimental evidence for a mechanism explaining how traits pass to subsequent generations came in 1866 when an Austrian monk, Gregor Mendel,
published his studies covering 10 years worth of work on the mechanism of inheritance of 7 characteristics in garden peas in a paper called “Experiments in Plant Hybridization”.

CONCEPTS OF GENOMIC BIOLOGY Page 2
Mendel's Experiments Video
In 1865 Mendel delivered two long lectures that were published in 1866 as "Experiments in Plant Hybridization." This established what eventually became formalized as the Mendelian Laws of inheritance:
The law of dominance. For each trait, one factor (gene) is dominant and appears as the phenotype in the first filial generation (F1). In the F2 generation the dominant trait occurs more often, in a definite 3:1 ratio. The alternative form is recessive. In Mendel's peas, tallness was dominant, shortness recessive. Therefore, three times as many plants were tall as were short. This constant ratio represents the random combination of alleles during reproduction. Any combination of alleles that includes the dominant allele will express that form of the trait.
The law of independent segregation. Inherited characteristics (such as stem length in Mendel's pea plants) exist in alternative forms (tallness, shortness)—today known as alleles. For each characteristic, an individual possesses two paired alleles—one inherited from each parent. Correspondingly, these pairs segregate (i.e. separate or assort) in germ cells and recombine during reproduction so that each parent transmits one allele to each offspring.
The law of independent assortment. Specific traits operate independently of one another. A pea plant might have a stem that is tall or short, but in either case may produce white or gray seed coats.
However, the significance of Mendel’s work and his insight into the mechanism of inheritance went unrecognized until 1900 when three European scientists, Hugo de Vries, Carl Correns, and Erich von Tschermak reached similar conclusions in their own research though they claimed to be unaware of Mendel’s earlier theory of the 'discrete units' on which genetic material resides.
The biological entity (factor) responsible for defining traits was later termed a gene by Wilhelm Johansen in 1910, but the biological basis for inheritance remained unknown until DNA was
identified as the genetic material in the 1940s. Thus,

CONCEPTS OF GENOMIC BIOLOGY Page 3
(RETURN)
Chromosomes carry genes.
The notion that Mendel’s particulate hereditary factors reside on visible structures called chromo-somes was originally independently proposed by Theodor Boveri, a German scientist, and Walter Sutton, an
American graduate student, in 1902 at about the same time that Mendel’s Laws of inheritance were being rediscovered.
(RETURN)
it was early in the 20th century that the name “gene” was given to the hereditary unity described by Mendel decades earlier, and the study of genetics and genomics began in earnest.
2.2. GENES ARE CARRIED ON CHROMOSOMES.
At about the same time that genes were coming into focus as having a role in inheritance, a series of observations at the cellular level established:
The existence of structures called chromosomes.
The developing theory stated:
More than one gene is located on each
chromosome.
Thus, chromosomes are like a string of beads with each gene represented as a bead. Along the length of the chromosome (string of beads) there are genes for many traits on each chromosome, and each gene occupies a specific position on each chromosome called a locus.
The chromosomes are passed from one generation to the next and carry genes to the next generation as they are passed.
These points were incorportated into what we now know as the Chromosomal Theory of Inheritance.

CONCEPTS OF GENOMIC BIOLOGY Page 4
(RETURN)
2.3. THE CHROMOSOMAL THEORY OF INHERITANCE.
In the early years of the 20th century Thomas Hunt Morgan, who was skeptical about the theories of the day concerning Mendel’s observations and the role of chromosomes in inheritance, began conducting a series of experiments using the fruit fly, Drosophilla melanogaster, that ultimately convinced him of the details of inheritance leading to what is called today the chromosomal theory of inheritance. The general tenets of this theory are given below:
Multiple genes conditioning the cellular and organismal traits an organism possesses are passed from one cellular or organismal generation to the next on chromosomes.
Genes for specific traits reside at specific positions on chromosomes called loci (singular locus).
Most cells of an organism have homologous pairs of chromosomes for each chromosome found in the cell.
The complete set of chromosomes an organism possesses is called it’s karyotype.
Figure 2.1. The complete set of 23 pairs of human chromosomes is shown in the karyotype above. Note that there are 22 pairs of autosomal Chromosomes, and the X and Y sex chromosome “pair”. Thus, we say that there are 22 pairs of homologous autosomal
chromosomes plus a pair of sex chromosomes (X or Y) in humans, and humans have 46 (diploid number) chromosomes in total.
The complete set of human chromosomes is shown in Figure 2.1. Humans have 22 pairs of autosomal chromosomes, and the X and Y sex chromosomes that are present in males (XY) of females (XX). Thus, we say that there are 22 pairs of homologous autosomal chromosomes plus a pair of sex chromosomes (X or Y) in humans. Humans have 46 chromosomes in total, and the diploid number of chromosomes is 26.

CONCEPTS OF GENOMIC BIOLOGY Page 5
(RETURN)
Gametes, eukaryotic cells that pass chromosomes to the next organismal generation, contain only a haploid number of chromosomes (23 in the case of homans). Thus, gametes have only 1 chromosome from each pair found in a non-gametic cell. Chromosome numbers are constant for a species, but vary from one species to another.
One of the chromosomes in each homologous pair comes from the maternal parent while the other chromosome in the pair comes from the paternal parent.
Although traits are conditioned by genes at specific loci on the chromosomes, the gene at a given locus coming from each parent may not be the same. They can be either the dominant (according to Mendel’s law of dominance) factor, ort he recessive factor. We now call the nature of the factor (gene) at each locus, an allele.
When both the maternal and paternal homologous chromosome contain the same allele, the organism is said to be homozygous, but if the alleles contained at the locus on the homologous chromosomes are different the organism is said to be heterozygous.
When an organism is homozygous, if the allele it bears is the dominant allele, the organism demonstrates a homozygous dominant genotype. While a homozygous organism bearing 2 identical recessive alleles is considered homozygous recessive genotype.
The genotype that an organism possesses in combination with environmental factors is responsible for production of the trait that we see. This is also a definition of the phenotype of an individual, i.e. the appearance of the individual resulting from the interaction of genotype and environmental factors. Thus, an organism can demonstrate a dominant phenotype or a recessive phenotype.
What Mendel observed was the phenotype of his pea plants. From observations of phenotype he proposed a model for genotypic behavior of his “factors” that we no know as genes. We also know that these genes reside on chromosomes, and the manner in which the chromosomes are passed to the next generation provides the basis for Mendel’s law of segregation that directly relates the behavior of the chromosomes bearing the genes to the phenotypic behavior that Mendel observed. However, there are a number of instances where, although Mendel’s law of segregation applies additional background is required to appreciate how such Mendel’s work applies.

CONCEPTS OF GENOMIC BIOLOGY Page 6
(RETURN)
Figure 2.2. Phenotypic description of the alleles of the C-locus for coat
color in rabbits. Note that this patterning is also found in many other animals although the names of the phenotypes may differ.
Once the simple laws of Mendel that governed inheritance had been established and related to the behavior of chromosomes, there were many examples of situations that were not fully accounted for with the simple laws. In the early 20th century there was great controversy, not just about the chromosomal theory and its relationship to inheritance of traits, but about other known examples that appeared not to be explained by Mendel and the chromosomal theory.
Resolution of these issues took decades and required careful, thorough and well-designed experiments too provide us with an understanding of many of these situation. In fact a few of these controversies were not fully resolved until the genomic era and some are still being investigated today.
2.4.1. Multiple alleles (retrun)
Note that it is possible that for some traits more than 2 alleles exist. In this case there is a hierarchy of dominance among the multiple alleles. In any given individual the more dominant allele of the 2 alleles it posses is dominant, while the more recessive one will be the recessive allele.
Examples of this phenomenon could be the ABO blood type system and the rabbit coat color example discussed shown in Figure 2.2. There are 4 unique alleles that have been found at the C-locus, which is one of 5 separate genetic loci that generate coat color patterns in rabbits. The hierarchy of dominance that has been observed at the C-locus suggests that the
wild type “large C” allele is the “most” dominant of the alleles in the dominance hierarchy, and the “most recessive” of the alleles is the “small c” locus. A rabbit whose genotype is cc has an albino phenotype while a rabbit with a CC genotype will be fully colored (e.g. agouti, or black that is really dark grey as described in the Figure 2.2). The second most dominant allele is the chinchilla allele, cch- allele, and the ch-allele is intermediate in dominance between the cch- allele and the c- allele.
2.4. ADDI TIONAL COMPLEXITY OF MENDELIAN
INHERITANCE. (RETURN)

CONCEPTS OF GENOMIC BIOLOGY Page 7
(RETURN)
2.4.2. Incomplete dominance and co-dominance (retrun)
It is also possible to have 2 alleles demonstrate an intermediate phenotype in the heterozygous condition. This phenomenon is referred to as incomplete dominance (similar to co-dominance), and can be observed in Figure 2.2. where the phenotype of a cchch or cchc heterozygous rabbit is distinct and intermediate between the homozygous (more) dominant cchcch and the homozygous (more) recessive chch or cc phenotypes.
Another example is given in Figure 2.3., where pure breeding (homozygous) red and white flowered plants are crossed to give rise to intermediate heterozygous pink plants. In some plants the
intermediate heterozygotes appear as separate distinct patches of color. This is typical of the description of co-dominant traits where the distinct alleles in a heterozygote are both visible. Thus, co-dominance and incomplete dominance may be a distinction without a difference.
Rr rr
rr rr Rr Rr
RR rr
Rr Rr Rr Rr
A. Parents
F1
Rr Rr
Rr Rr rr rr
Rr rr
rr rr
Rr Rr
B. Parents
F1
RR Rr Rr rr
Rr Rr
Rr Rr rr rr
Rr rr
rr rr
Rr rr
C. Parents
F1
Rr Rr rr rr
Figure 2.3. Example of incomplete dominance in flower color of four o’clocks. A) Red flowering x White flowering yields all pink flowers; B) pink flowering x pink flowering yields 1 red : 2 pink : 1 while flowers;
and C) pink flowering x white flowering yields half pink and half white flowers.

CONCEPTS OF GENOMIC BIOLOGY Page 8
(RETURN)
2.4.3. Sex linked interitance (retrun)
Another example that differs from typical Mendelian inheritance is sex-linked inheritance. In organisms that have X and Y choromosomes, such as Drosophila and humans, the female typically has a pair of X chromosomes (XX) while the male has an X and a Y chromosome (XY). So when a red-eyed female fruit fly is crossed with a white-eyed male, the result is all red-eyed progeny. This might seem like a normal autosomal inheritance pattern where red eyes are a dominant trait. However, in the reciprocal cross (a white-eyed female crossed to a red- eye male. All females will have red eyes, and all males will have white eyes.
This demonstrates that the eye-color trait in Drosophilla is a sex-linked trait, and it is conditioned by a gene located on the X chromosome. Males contribute an X-chromsomes only to their daughters, as their sons must get the Y-chromosome. Females contribute their X-chromsomes to both males and females.
This phenomenon is pictorially demonstrated
using Punnet’s squares in Figure 2.4. below.
Figure 2.4. Demonstration of sex linked inheritance. The outcome as demonstrated in the Punnet’s squares above is different based on whether the male bears the dominant or recessive trait.

CONCEPTS OF GENOMIC BIOLOGY Page 9
(RETURN)
2.4.4. Epistasis (retrun)
Sometimes the phenotype of an organism does not reflect the actual genotype. This can be the case when one or more genes are epistatic to others. Epistatic genes modify or eliminate the phenotype of others so that the phenotype is not apparent. An example of an epistatic gene might be a gene for baldness. This gene would be epistatic to genes for hair color, e.g. red hair or blond hair genes.
Another example of an epistatic gene is the c-allele in rabbits given above. This allele produces a phenotypically albino, white rabbit with pink eyes in
the homozygous recessive state. However, there are at least 5 additional genetic loci that condition various coat colors and patterns. Many of these other loci have multiple alleles (as does the C-locus, see above), but the rabbit will be albino if it is genotypically cc (homozygous recessive) at the C-locus. Demonstrating that the C-locus is epistatic to the other coat color loci.
2.3.5. Epigenetics (retrun)
More recently discovered phenomena involving heritable changes in gene expression that are not related to actual changes in DNA sequence, but rather are related to chromosome structure and function have also emerged. These phenomena are referred to as epigenetic inheritance, and have emerging importance in virtually all areas of biology and medicine. We will discuss them in greater molecular detail later, but they clearly had their beginning in classical genetics.

CONCEPTS OF GENOMIC BIOLOGY Page 10
(RETURN)
In his studies with garden peas Mendel observed that each of the 7 traits that he studied behaved independently of each other. The mechanism that this observation generated involved genes (hereditary factors) assorting independently of each other. Thus, when 2 factors (genes) were involved in a cross, each of them behaved independently.
However, the chromosomal theory of inheritance contradicts this observation by suggesting that genes are linked together on chromosomes, and further suggests that it is the chromosomes that are passed to the next generation. If this is the case, how can genes on the same chromosome assort indepen-dently?
Answering this question plagued the early develop-ment of genetics until the chromosomal theory of inheritance emerged and the idea of gene linkage for genes on the same chromosome were clearly shown by Thomas Hunt Morgan and his colleagues about a century ago.
Once established that it is chromosomes that assort independently, it was clear that Mendel had fortuitously
chosen 7 genes on 7 different chromosomes to work with, and as a consequence Mendel’s law of independent assortment did not necessarily apply to all genes since it was the chromosomes that assorted not the genes per se.
The question has been raised as to whether Mendel chose only data to work with that supported this theory and disregarded other data or traits that did not fit his theory to present. Whether this is true or not we
will never really know, but it surely doesn’t detract from the important contribution Mendel’s work has made to the science of genetics and genomic biology by establishing an important set of rules that govern the inheritance of traits.
2.5.1. Meiosis: chromosomes assort independently (retrun)
The theory that allows us to explain the mapping of genes begins with an understanding of the behavior of chromosomes during meiosis. During the assorting of diploid chromosomes sets like those found in somatic cells into haploid chromosomes sets like those found in gametes, it is possible to exchange parts of chromosomes between different homologous sister chromatids.
2.5. GENES ON THE SAME CHROMOMSOME ARE
LINKED.
CONCEPTS OF GENOMIC BIOLOGY Page 11
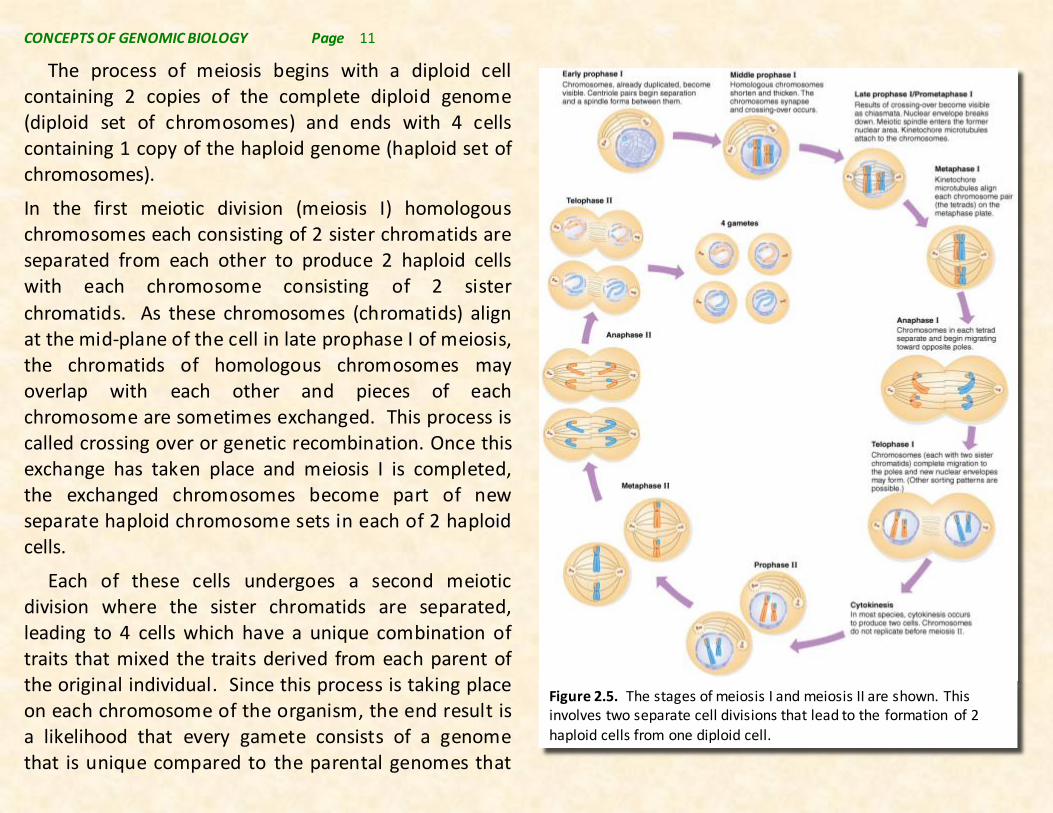
The process of meiosis begins with a diploid cell containing 2 copies of the complete diploid genome (diploid set of chromosomes) and ends with 4 cells containing 1 copy of the haploid genome (haploid set of chromosomes).
In the first meiotic division (meiosis I) homologous chromosomes each consisting of 2 sister chromatids are separated from each other to produce 2 haploid cells with each chromosome consisting of 2 sister
chromatids. As these chromosomes (chromatids) align at the mid-plane of the cell in late prophase I of meiosis, the chromatids of homologous chromosomes may overlap with each other and pieces of each chromosome are sometimes exchanged. This process is called crossing over or genetic recombination. Once this exchange has taken place and meiosis I is completed, the exchanged chromosomes become part of new separate haploid chromosome sets in each of 2 haploid cells.
Each of these cells undergoes a second meiotic division where the sister chromatids are separated, leading to 4 cells which have a unique combination of traits that mixed the traits derived from each parent of the original individual. Since this process is taking place on each chromosome of the organism, the end result is a likelihood that every gamete consists of a genome that is unique compared to the parental genomes that
Figure 2.5. The stages of meiosis I and meiosis II are shown. This involves two separate cell divisions that lead to the formation of 2 haploid cells from one diploid cell.

CONCEPTS OF GENOMIC BIOLOGY Page 12
produced the individual. This mixing of genes at loci along the length of chromosomes contributes much to the genetic diversity required to make the process of evolution work.
2.5.2. Mapping genes on chromosomes (retrun)
Using Drosophila, Thomas Hunt Morgan and his students accumulated a large collection of mutants (allele pairs) for specific traits. As the collection of mutants grew, it became clear that particular sets of
traits assorted together rather than independently as Mendel had found with his peas. Morgan concluded that genes for specific traits are linked together into 4 groups in Drosophila. This happened to equal the number of chromosomes observed in Drosophila cells in the microscope. By studying the process of meiosis as described above, it was further established that pieces of homologous chromosomes are exchanged when chromosome numbers are reduced from 2 homologous chromosomes per cell, to just a single chromosomal homolog in the gametes that are fused to produce the next generation.
From this initial idea of linkage of genes into groups on chromosomes, Alfred Sturtevant, Morgan's student, was the first scientist to make genetic or linkage maps of fruit fly chromosomes. To do this Sturtevant reasoned that since pieces of homologous
chromosomes can be exchanged during meiosis, the frequency of this exchange provides a measure of the relative distance between linked genes on the same chromosome. Distantly located genes recombine more frequently while nearby genes rarely recombine and are closely linked. By measuring the frequency of crossing-over between linked genes on the same chromosomes the distance between genes can be estimated, and genetic maps can be calculated and constructed.
From Morgan and Sturtevant’s work, the percentage crossing-over became a chromosomal distance measurement, and the definition of a unit of crossing over, became know as the Centimorgan (=1% crossing over between linked genes on the same chromosome).
Figure 2.6. Alfred Sturte- vant’s first genetic map of the Drosophila chrom-
osomes.
CONCEPTS OF GENOMIC BIOLOGY Page 13
(RETURN)
Mendel, perhaps fortuitously, chose to work with a series of traits where he could find a pair of discrete phenotypes. However, not all phenotypes are so clean producing discrete classes. Above we have already looked at examples of incomplete dominance, multiple alleles, and epistasis, but for other traits phenotypes are continuously variable between 2 extremes rather than producing discrete phenotypic classes. Examples of such traits are often related to height, weight, or amounts of things. There are several books written on the topic of quantitative inheritance, and one can link-out to online more brief treatments of the topic can be found. A number of additional references on quantitative genetics can be found at the link-out, but be aware that these may not be adequately edited, and they are certainly incomplete, although they do provide an overview of the area suitable for understanding the relationship of quantitative genetics to genomic biology. Also note that there are many more complex issues involved in understanding quantitative inheritance that require statistical background beyond that expected here.
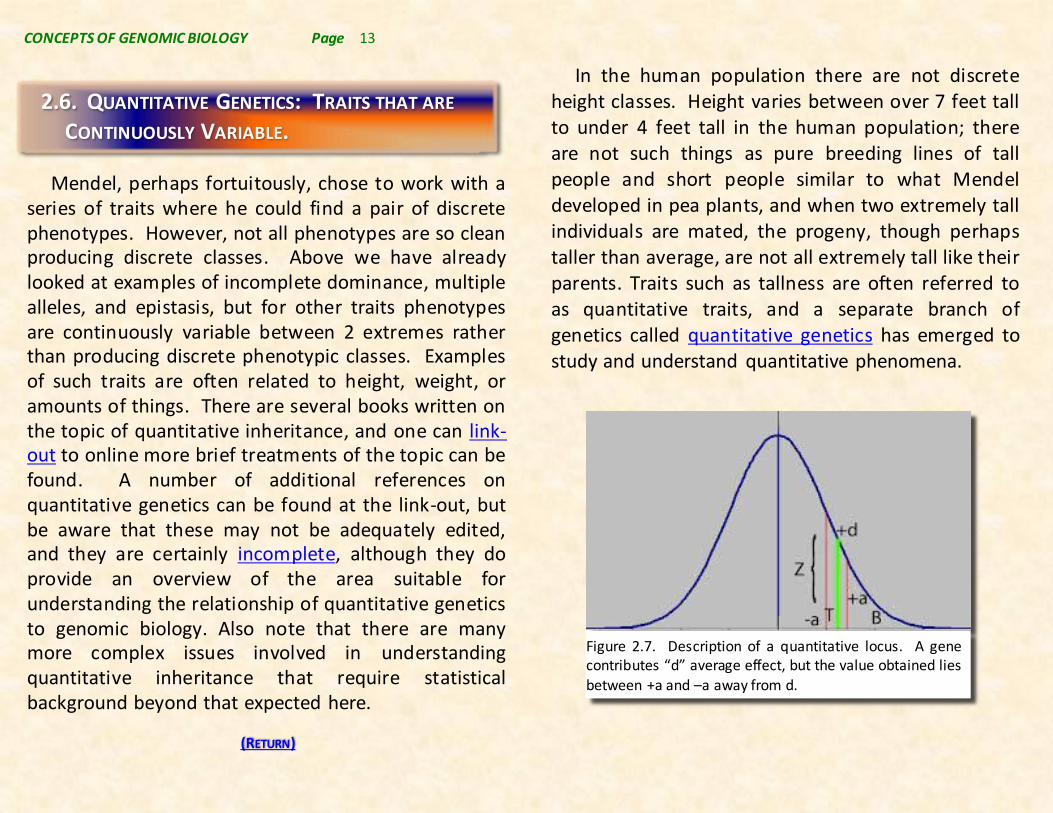
In the human population there are not discrete height classes. Height varies between over 7 feet tall to under 4 feet tall in the human population; there are not such things as pure breeding lines of tall people and short people similar to what Mendel developed in pea plants, and when two extremely tall individuals are mated, the progeny, though perhaps taller than average, are not all extremely tall like their parents. Traits such as tallness are often referred to as quantitative traits, and a separate branch of genetics called quantitative genetics has emerged to study and understand quantitative phenomena.
Figure 2.7. Description of a quantitative locus. A gene contributes “d” average effect, but the value obtained lies between +a and –a away from d.
2.6. QUANTITATIVE GENETICS: TRAITS THAT ARE
CONTINUOUSLY VARIABLE.
CONCEPTS OF GENOMIC BIOLOGY Page 14
In classical genetics, statistical approaches to quantitative inheritance have emerged that provide statistical tools for detailed analyses of quantitative inheritance. These statistical approaches focus on phenotypically defining 2 alleles at a putative “quantitative locus”. The midpoint between homozygotes of the 2 alleles is defined as +d, and the each opposing homozygotes would phenotypically deviate from the midpoint by +a or –a (see Figure 2.7.).
In a heterozygote a phenotype closer to the homozygous dominant (+a) than the midpoint (+d), indicates a dominant character to that allele, and a heterozygous phenotype closer to the homozygous recessive (–a) results from a less dominant character to the dominant allele. A measure of the heterozygote distance from the midpoint then becomes a statistical definition of incomplete dominance for such a quantitative gene. Note that in Mendel’s tall versus short pea plants the phenotype of the heterozygote is almost precisely +d, indicating 100% dominance of the tall allele over the short allele. In actual fact it is even possible to have a super dominant allele that gives a heterozygous phenotype more distant from the midpoint than +a, a phenomenon that is sometimes referred to as hybrid vigor.
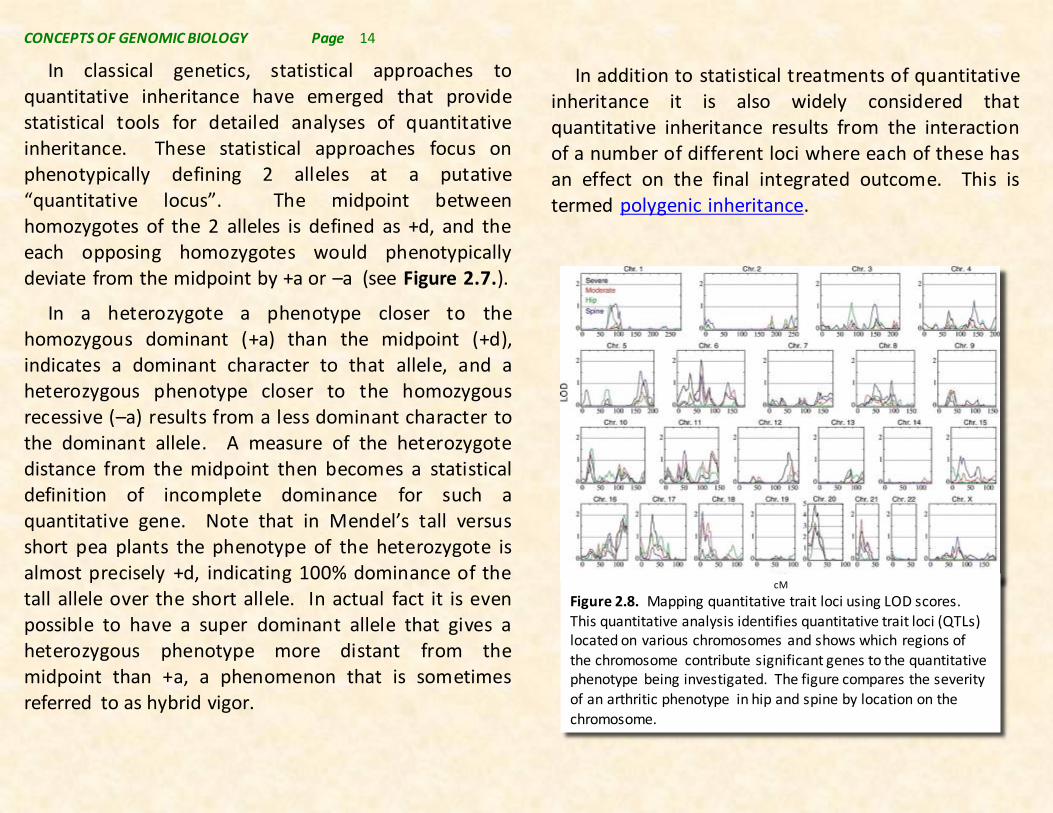
In addition to statistical treatments of quantitative inheritance it is also widely considered that quantitative inheritance results from the interaction of a number of different loci where each of these has an effect on the final integrated outcome. This is termed polygenic inheritance.
cM
Figure 2.8. Mapping quantitative trait loci using LOD scores. This quantitative analysis identifies quantitative trait loci (QTLs) located on various chromosomes and shows which regions of the chromosome contribute significant genes to the quantitative phenotype being investigated. The figure compares the severity of an arthritic phenotype in hip and spine by location on the
chromosome.

CONCEPTS OF GENOMIC BIOLOGY Page 15
(RETURN)
Gamete/Frequency
R/p r/q
R/p RR/p2 Rr/pq
r/q Rr/pq rr/q2
TABLE2.1.Femalegametes
Mal
ega
me
tes
Polygenes are nonallelic genes at a set of loci distributed in the genome that contribute to the overall quantitative phenotype observed in the organism. Figure 2.8 above shows how phenotypic data and their proximity to known marker genes allows the mapping of chromosome regions influencing quantitative phenotypes referred to as quantitative trait loci (QTLs). The distance measure used in this map is the so-called LOD score that relates phenotype to position on the chromosome. The LOD score method of locating regions of chromosomes that influence quantitative inheritance relies on having numerous closely related genetic markers on chromosomes.
Although the method has been available for some time, the advent of genomic techniques for identifying and mapping DNA sequence markers on chromosomes has markedly improved the accuracy and facility of identifying QTLs in genomes. Additionally, the availability of complete genome sequences makes it possible to not only identify regions of the chromosome related to phenotypes, but to actually identify the specific causally-related genes. The QTL approach has found wide application ranging from the mapping of human disease QTLs (example in Figure 2.8), to applications in plant and animal breeding, and to application in evolutionary and population genetics among others.
2.7. POPULATION GENETICS. (RETURN)
Statistical genetic theories have also become a major consideration in the discipline of population genetics. In the context of a population, the frequency of individuals having a given genotype is related to the frequency of each allele in the breeding population. If you assume that mating in a population is random and very large to assure that it is homogeneous, then the frequency of genotypes in the subsequent generation will be directly related to the frequency of alleles in the gamete pool that produces that generation. Thus, where there are only
2 alleles for a given locus found in the population, and p = frequency of dominant allele gametes while q = frequency of the recessive allele gametes, p + q = 1. As is shown in table 2.1., the

CONCEPTS OF GENOMIC BIOLOGY Page 16
(RETURN)
frequency of homozygous dominant individuals in the population should be p2 and the frequency of homozygous recessive individuals will be q2. Heterozygotes should then be found at a frequency of 2pq, and in total p2 + 2pq + q2 = 1. This is the binomial expansion of (p + q)2.
If this looks familiar, recall the Punnett’s squares that we did showing gene segregation in the F-2 generation. In that case since heterozygotes produce gametes, half of which carry the dominant allele and half of which carry the recessive allele, i.e. p = q = 0.5. Substituting these gamete allele frequencies into the
binomial equation above, we get the 1:2:1 segregation ratios we expect.
However, in a population, where there are both homozygotes and heterozygotes all producing gametes, p will not usually equal q, and a different equilibrium of gametes and genotypes will be established and maintained through time. This description is called a Hardy-Weinberg equilibrium.
In order for a population to sustain a Hardy-Weinberg equilibrium additional factors (assumptions) must be in place or the equilibrium will not be maintained. In addition to a large population and random mating within the population as discussed above, it is also necessary that there be no mutation
occurring in the population and that there be no natural selection taking place for the alleles or linked genes in question. Additionally, there should be no gene flow (migration into or from the population) taking place, and the population should not have gone through a dramatic change in size recently that may have related to genetic drift in the population. It should also be noted that the equations given above relate only to diploid species. Some species found in nature are natural polyploids (having more than 2 sets of chromosomes), and the equations for describing the behavior of polyploids are different
from the bionomial expansions described above. Also other changes in the equations are required for situations where there are more than 2 alleles found in a population.
As was with quantitative genetics, the introduction of tools from genomic studies into population genetics have greatly facilitated the investigation of genes in populations, and this is particularly relevant in the investigation of the human population. Population genetic studies using molecular markers for important health-related genes are now common place in Public Health studies.