cereal seed storage proteins - journal of experimental botany
TRANSCRIPT

Cereal seed storage proteins: structures, properties and
role in grain utilization
Peter R. Shewry1 and Nigel G. Halford
IACR-Long Ashton Research Station, Department of Agricultural Sciences, University of Bristol,Long Ashton, Bristol BS41 9AF, UK
Received 18 July 2001; Accepted 8 October 2001
Abstract
Storage proteins account for about 50% of the totalprotein in mature cereal grains and have importantimpacts on their nutritional quality for humans andlivestock and on their functional properties in foodprocessing. Current knowledge of the structuresand properties of the prolamin and globulin storageproteins of cereals and their mechanisms of syn-thesis, trafficking and deposition in the developinggrain is briefly reviewed here. The role of the glutenproteins of wheat in determining the quality of thegrain for breadmaking and how their amount andcomposition can be manipulated leading to changesin dough mixing properties is also discussed.
Key words: Cereals, functional properties, gene expression,
genetic engineering, protein bodies, proteins.
Introduction
Cereals are the most important crops in the world, withtotal annual grain yields exceeding 2000 million tonnes(mt), compared with less than 250 mt for legume seeds(including pulses, soybean and groundnut) (FAO, 1999).Although a number of cereal species are grown, three(maize (604 mt in 1998), wheat (589 mt in 1998) andrice (563 mt in 1998)) together account for over 70%of the total production. Other cereals include barley,sorghum, millets (which comprise a number of small-seeded tropical species), oats and rye, in order ofdecreasing total production.
Cereal grains contain relatively little protein comparedto legume seeds, with an average of about 10–12% dry wt.Nevertheless, they provide over 200 mt of protein for thenutrition of humans and livestock, which is about three
times the amount derived from the more protein-rich(20–40%) legume seeds. In addition to their nutritionalimportance, cereal seed proteins also influence the utiliza-tion of the grain in food processing. This is particularlyimportant in wheat, which is largely consumed by humansafter processing into bread and other foods. It is notsurprising, therefore, that cereal seed proteins have been amajor topic of research for many years, with the aim ofunderstanding their structures, control of synthesis androle in grain utilization.
Cereal seed storage proteins
The scientific study of cereal grain proteins extends backfor over 250 years, with the isolation of wheat gluten firstbeing described in 1745 (Beccari, 1745). Since then moresystematic studies have been carried out, notably by TBOsborne (1859–1929) who can be regarded as the fatherof plant protein chemistry. Osborne developed a classi-fication of plant proteins based on their solubility ina series of solvents, for example, albumins in water,globulins in dilute saline. Although ‘Osborne fractiona-tion’ is still widely used, it is more usual today to classifyseed proteins into three groups: storage proteins, struc-tural and metabolic proteins, and protective proteins.Seed storage proteins fall into three different Osbornefractions and occur in three different tissues of the grain.
Storage globulins
The embryo and outer aleurone layer of the endospermcontain globulin storage proteins, and those from maizeembryos have been characterized in some detail (Kriz,1989, 1999; Kriz and Schwartz, 1986; Kriz and Wallace,1991; Wallace and Kriz, 1991). These proteins are readilysoluble in dilute salt solution and have sedimentationcoefficients of about 7. They have limited sequence
1 To whom correspondence should be addressed. Fax: q44 (0)1275 394299. E-mail: [email protected]
Journal of Experimental Botany, Vol. 53, No. 370,Inorganic Nitrogen Assimilation Special Issue, pp. 947–958, April 2002
� Society for Experimental Biology 2002
Dow
nloaded from https://academ
ic.oup.com/jxb/article-abstract/53/370/947/537249 by guest on 14 April 2019

similarity with, and may be homologous to, the 7S vicilinsof legumes and other dicotyledonous plants; they alsohave similar structures and properties (Kriz, 1999).Related proteins have been found in embryos anduoraleurone layers of wheat, barley and oats (Burgess andShewry, 1986; Yupsanis et al., 1990; Heck et al., 1993). 7Sglobulins from rice embryos have also been characterized(Horikoshi and Morita, 1975), but their relationshipsto other plant 7S globulins have not been established. The7S globulins are stored in protein bodies and appear tofunction solely as storage proteins. However, they do notappear to be absolutely required for normal seed func-tion, at least in maize, where a null mutant behavesnormally in terms of development and germination (Krizand Wallace, 1991). Furthermore, although the aleuroneand embryo are rich in proteins compared with the starchyendosperm, the globulins in these tissues have limitedimpact on the end use properties of the grain. In smallgrained cereals, such as wheat, the aleurone and embryoaccount only for about 10% of the grain dry weightand are usually removed by milling (wheat), polishing(rice), pearling (barley) or decortication (sorghum), beforehuman consumption. By contrast, the embryo of maizeaccounts for 10–11% of the grain and its high contents ofprotein and oil are important for livestock nutrition.
Storage globulins of 11–12S located in the starchyendosperm are also present in at least some cereal grains.In fact, in oats and rice these proteins form the majorendosperm storage protein fraction, accounting for about70–80% of the total protein. It is now known that theseproteins are related to the widely distributed ‘legumin’type globulins which occur in most dicotyledonousspecies (Casey, 1999). The rice proteins are not readilysoluble in dilute salt solutions and hence are classicallydefined as glutelins, but they clearly belong to the 11–12Sglobulin family. They comprize subunits of Mr approx.55 000 that are post-translationally cleaved to give acidic(Mr approx. 33 000 in oats, 28–31 000 in rice) and basic(Mr approx. 23 000 and 20–22 000, respectively) polypep-tide chains linked by a single disulphide bond (Shotwell,1999; Takaiwa et al., 1999). The oat globulin alsoresembles the legumins in forming a hexameric structurewith a sedimentation coefficient of about 12.
Proteins related to legumins, called ‘triticins’, arepresent in starchy endosperm of wheat, although theyaccount only for about 5% of the total seed protein(Singh et al., 1988). Triticins consist of large (Mr about40 000) and small (Mr about 22–23 000) polypeptidechains, but the subunits appear to form dimeric structuresrather than the typical legumin hexamers (Singh et al.,1988, 1993; Singh and Shepherd, 1985).
The high content of globulin storage proteins in oatgrain may contribute to the high nutritional value whencompared with other cereals, such as barley and wheat,an important factor in view of the widespread use of
oats for livestock feed (Lockhard and Hurt, 1986;Cuddeford, 1995).
Prolamin storage proteins: general properties
With the exceptions of oats and rice, the major endo-sperm storage proteins of all cereal grains are prolamins.This name was originally based on the observation thatthey are generally rich in proline and amide nitrogenderived from glutamine, but it is now known that thecombined proportions of these amino acids actually varyfrom about 30–70% of the total among different cerealsand protein groups. Similarly, although prolamins wereoriginally defined as soluble in alcoholuwater mixtures(e.g. 60–70% (vuv) ethanol, 50–55% (vuv) propan-1-ol orpropan-2-ol), some occur in alcohol-insoluble polymers.Nevertheless, all individual prolamin polypeptides arealcohol-soluble in the reduced state. The prolamins varygreatly, from about 10 000 to almost 100 000, in theirmolecular masses.
It is clear, therefore, that prolamin storage proteinsare much more variable in structure than the 7S and11u12S globulins, and it is possible that the major groupsof prolamins in the Triticeae (wheat, barley, rye) andthe Panicoideae (maize, sorghum, millets) have separateevolutionary origins. Nevertheless, most prolaminsshare two common structural features. The first is thepresence of distinct regions, or domains, which adoptdifferent structures to each other and may have differ-ent origins. The second is the presence of amino acidsequences consisting of repeated blocks based on one ormore short peptide motifs, or enriched in specific aminoacid residues, such as methionine. These features areresponsible for the high proportions of glutamine, pro-line and other specific amino acids (e.g. histidine, glycine,methionine, phenylalanine) in some prolamin groups.
The prolamin superfamily
Discussions of prolamin structure and properties can beconfusing for the non-expert because of the complexity ofthe fractions and their specialized nomenclature. How-ever, the availability of complete amino acid sequencesof representatives of all the major prolamin groups hasallowed the redefinition of their classification in relationto structural and evolutionary relationships (Shewry andTatham, 1990).
This new system of classification assigns all of theprolamins of the Triticeae (wheat, barley and rye) tothree broad groups: sulphur-rich (S-rich), sulphur-poor(S-poor) and high molecular weight (HMW) prolamins,with several subgroups within the S-rich group (Table 1).These groups do not correspond directly to the polymericand monomeric fractions in wheat (glutenins and gliadins,respectively) recognized by cereal chemists, as both
948 Shewry and Halford
Dow
nloaded from https://academ
ic.oup.com/jxb/article-abstract/53/370/947/537249 by guest on 14 April 2019

monomeric and polymeric forms of S-rich and S-poorprolamins occur.
Structures of typical S-rich, S-poor and HMWprolamins of wheat are summarized in Fig. 1. They allcontain extensive repeated sequences based on proline-rich and glutamine-rich motifs with the repeat motifsof the S-rich and S-poor groups being clearly related.Similarly, sequence similarity is clearly present betweenthe non-repetitive domains of the S-rich and HMWprolamins, particularly in positions of conserved cysteineresidues and amino acid residues adjacent to these. Onthe basis of such comparisons, it can be concluded thatthe S-rich, S-poor and HMW prolamins have a com-mon evolutionary origin. Wider comparisons show fur-ther evolutionary and structural relationships to severalgroups of zein proteins (see below), the prolamins of oatsand rice, 2S albumin storage proteins of dicotyledon-ous seeds, cereal seed inhibitors of a-amylase and trypsinand a range of low Mr cysteine-rich plant proteins includ-ing lipid transfer proteins and cereal grain puroindolines.These proteins are, therefore, together defined as theCereal Prolamin Superfamily of plant proteins (Kreiset al., 1985).
In wheat, the prolamins form the major components ofthe gluten protein fraction which forms a viscoelasticnetwork in doughs and is largely responsible for theability to process wheat to form bread, pasta and manyother food products.
The prolamins of maize
The prolamins of maize (called zeins) and of otherpanicoid cereals (e.g. sorghum and many millets) arecomprised of one major group of proteins (a-zeins) andseveral minor groups (b, c, d-zeins) (Coleman and Larkins,1999; Leite et al., 1999) (Fig. 2). Amino acid sequencecomparisons demonstrate that the b, c and d-zeins are allmembers of the prolamin superfamily, but only the
c-zeins contain repeated amino acid sequences (eithertwo or eight tandem repeats of Pro-Pro-Pro-Val-His-Leu). The b-zeins and d-zeins are both rich in methioninewith these residues being clustered in a region close to theC-terminus in the former.
By contrast, the a-zeins do not appear to be related toany other prolamins except the a-type prolamins of otherpanicoid cereals. They consist of two major subclassescalled the 19K and 22K zeins based on their Mr deter-mined by SDS-PAGE although they have true molecularmasses of 23–24 000 and 26 500–27 000, respectively. Bothsubclasses contain degenerate repeats of about 20 amino
Fig. 1. Schematic structures of typical HMW, S-rich and S-poorprolamins (based on sequences in Anderson et al., 1989; Bartels et al.,1986; Hsia and Anderson, 2001). Repetitive sequences are shadedand disulphide bonds between conserved cysteine residues (1–8) in thec-gliadin shown as lines. SH denotes the positions of cysteine residues inthe HMW prolamins.
Table 1. Summary of the types and characteritics of wheat grain prolamins (gluten proteins)
Components Mr (% total) Polymers or monomers Partial amino acid composition (mol%)
HMW prolaminsHMW subunits of glutenin 65–90 000
(6 –10%)Polymers 30–35% Gly, 10–16% Pro, 15–20% Gly, 0.5–1.5% Cys,
0.7–1.4% LysS-rich prolaminsc-gliadins 30– 45 000 Monomersa-gliadins (70 –80%) Monomers 30–40% Gln, 15–20% Pro, 2–3% Cys, -1.0% LysB- and C-type LMW subunits of glutenina
)Polymers
)
S-poor prolaminsv-gliadinsD-type LMW subunits of glutenina
o30–75 000(10 –20%)
MonomersPolymers
o40–50% Gln, 20–30% Pro, 8–9% Phe, 0 –0.5% Lys,
0 –-0.5% Cysb
aC-type LMW subunits are essentially polymeric forms of a- and c-gliadins and D-type LMW subunits polymeric v-gliadins. The B-type LMWsubunits constitute a discrete group of S-rich prolamins.
bCys is present in D-type LMW subunits, but not v-gliadins.
Cereal seed storage proteins 949
Dow
nloaded from https://academ
ic.oup.com/jxb/article-abstract/53/370/947/537249 by guest on 14 April 2019

acid residues, with nine such blocks present in the Z19and ten in the Z22 zeins (Fig. 2).
The a-zeins contain only one or two cysteine residuesper molecule and are present in the grain as monomersor oligomers, while the b-, c- and d-zeins are all richer incysteine and form polymers.
Synthesis and deposition of cereal seed storageproteins
Cereal seed storage proteins are produced by the secret-ory pathway and deposited in discrete protein bodies.However, the origins of the protein bodies and themechanisms that determine the pathway of storage pro-tein trafficking and deposition are still incompletelyunderstood. The 7S and 11S storage globulins, whichare present in the embryo and the aleurone layer and inthe starchy endosperm, respectively, of some cereals arebelieved to follow the same route as the homologousproteins in dicotyledonous seeds. Thus, they are synthes-ized on rough endoplasmic reticulum (ER) membranes,transported co-translationally into the lumen and thenpass via the Golgi apparatus into a specific populationof protein storage vacuoles, which differ from the lyticvacuoles that are also present in developing seeds. Theprecise details of this process have been reviewed else-where (Kermode and Bewley, 1999). The precise sortingmechanisms are incompletely understood but physicalaggregation within the Golgi appears to be important,leading to the formation of electron dense aggregateswhich form the contents of dense vesicles. Globulin stor-age proteins do not contain cleavable pro-domains whichconfer vacuolar targeting, but non-cleavable segmentswithin the mature protein sequence may be important(Kermode and Bewley, 1999).
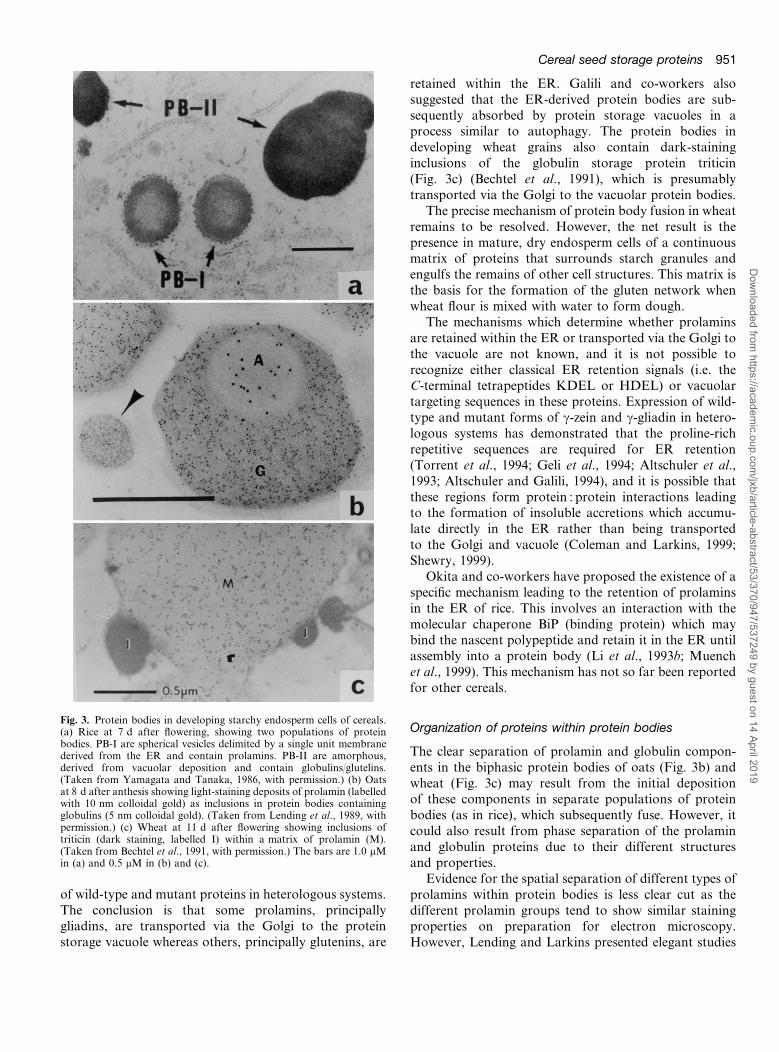
The mechanisms of prolamin transport and depositionare less well understood than those of globulins, but twopathways may occur. In maize, other panicoid cereals(sorghum, millets) and rice the prolamins appear to
accumulate directly within the lumen of the ER, leadingto the formation of discrete protein bodies surrounded bya membrane of ER origin (Coleman and Larkins, 1999;Muench et al., 1999). In rice, this leads to the presenceof two populations of protein bodies, PB-I which isof ER origin and contains prolamins and PB-II whichis vacuolar in origin and contains globulinsuglutelins(Fig. 3a) (Yamagata and Tanaka, 1986; Krishnan et al.,1986). Okita and co-workers also provided evidence thatprolamins and globulinsuglutelins are synthesized in sep-arate subdomains of the ER, with mRNAs for prolaminsbeing targeted to the rough ER associated with thedeveloping prolamin-containing protein bodies andglobulinuglutelin mRNAs to more typical cysternal ERmembranes (Li et al., 1993a; Choi et al., 2000). Morerecent work indicates that prolamin mRNA localizationresults from binding to a specific site on the tubulin andactin cytoskeleton (Muench et al., 1998).
Oats resemble rice in having a high proportion ofglobulin-type storage proteins in starchy endosperm cells,but in this case the globulins and prolamins are co-locatedin the same protein bodies, with the prolamins presentas lighter-staining inclusions (Fig. 3b) (Lending et al.,1989). The authors suggested that this results from thefusion of two populations of protein bodies, of ER origin(containing prolamins) and of vacuolar origin (containingglobulins).
Whereas the prolamins of maize, rice and probablyalso oats appear to accumulate directly within the ERwith little or no evidence for transport to the vacuole,there is now overwhelming evidence that both routes ofprotein transport and protein body formation operatein wheat, barley and probably also rye. Evidence for thiswas reviewed elsewhere (Galili, 1997), but includesimmunogold labelling of prolamins within Golgi com-plexes (see also Shewry, 1999), the observation of smallprotein bodies within or associated with the ER, theassociation of ER marker enzymes with protein bodiesprepared by subcellular fractionation and the expression
Fig. 2. (a) One-dimensional SDS-PAGE of total zeins of maize. (b) Schematic structures of the Mr 19 000 (Z19) and Mr 22 000 (Z22) a-zeins of maize.(Taken from Shewry and Tatham, 1990, with permission.)
950 Shewry and Halford
Dow
nloaded from https://academ
ic.oup.com/jxb/article-abstract/53/370/947/537249 by guest on 14 April 2019
of wild-type and mutant proteins in heterologous systems.The conclusion is that some prolamins, principallygliadins, are transported via the Golgi to the proteinstorage vacuole whereas others, principally glutenins, are
retained within the ER. Galili and co-workers alsosuggested that the ER-derived protein bodies are sub-sequently absorbed by protein storage vacuoles in aprocess similar to autophagy. The protein bodies indeveloping wheat grains also contain dark-staininginclusions of the globulin storage protein triticin(Fig. 3c) (Bechtel et al., 1991), which is presumablytransported via the Golgi to the vacuolar protein bodies.
The precise mechanism of protein body fusion in wheatremains to be resolved. However, the net result is thepresence in mature, dry endosperm cells of a continuousmatrix of proteins that surrounds starch granules andengulfs the remains of other cell structures. This matrix isthe basis for the formation of the gluten network whenwheat flour is mixed with water to form dough.
The mechanisms which determine whether prolaminsare retained within the ER or transported via the Golgi tothe vacuole are not known, and it is not possible torecognize either classical ER retention signals (i.e. theC-terminal tetrapeptides KDEL or HDEL) or vacuolartargeting sequences in these proteins. Expression of wild-type and mutant forms of c-zein and c-gliadin in hetero-logous systems has demonstrated that the proline-richrepetitive sequences are required for ER retention(Torrent et al., 1994; Geli et al., 1994; Altschuler et al.,1993; Altschuler and Galili, 1994), and it is possible thatthese regions form protein : protein interactions leadingto the formation of insoluble accretions which accumu-late directly in the ER rather than being transportedto the Golgi and vacuole (Coleman and Larkins, 1999;Shewry, 1999).
Okita and co-workers have proposed the existence of aspecific mechanism leading to the retention of prolaminsin the ER of rice. This involves an interaction with themolecular chaperone BiP (binding protein) which maybind the nascent polypeptide and retain it in the ER untilassembly into a protein body (Li et al., 1993b; Muenchet al., 1999). This mechanism has not so far been reportedfor other cereals.
Organization of proteins within protein bodies
The clear separation of prolamin and globulin compon-ents in the biphasic protein bodies of oats (Fig. 3b) andwheat (Fig. 3c) may result from the initial depositionof these components in separate populations of proteinbodies (as in rice), which subsequently fuse. However, itcould also result from phase separation of the prolaminand globulin proteins due to their different structuresand properties.
Evidence for the spatial separation of different types ofprolamins within protein bodies is less clear cut as thedifferent prolamin groups tend to show similar stainingproperties on preparation for electron microscopy.However, Lending and Larkins presented elegant studies
Fig. 3. Protein bodies in developing starchy endosperm cells of cereals.(a) Rice at 7 d after flowering, showing two populations of proteinbodies. PB-I are spherical vesicles delimited by a single unit membranederived from the ER and contain prolamins. PB-II are amorphous,derived from vacuolar deposition and contain globulinsuglutelins.(Taken from Yamagata and Tanaka, 1986, with permission.) (b) Oatsat 8 d after anthesis showing light-staining deposits of prolamin (labelledwith 10 nm colloidal gold) as inclusions in protein bodies containingglobulins (5 nm colloidal gold). (Taken from Lending et al., 1989, withpermission.) (c) Wheat at 11 d after flowering showing inclusions oftriticin (dark staining, labelled I) within a matrix of prolamin (M).(Taken from Bechtel et al., 1991, with permission.) The bars are 1.0 mMin (a) and 0.5 mM in (b) and (c).
Cereal seed storage proteins 951
Dow
nloaded from https://academ
ic.oup.com/jxb/article-abstract/53/370/947/537249 by guest on 14 April 2019

showing differences in the distribution of zeins withinprotein bodies related to their position in the developingendosperm (Lending and Larkins, 1989). Thus, the pro-tein bodies in the youngest cells in the sub-aleuroneregion contain mainly b- and c-zeins which are distrib-uted throughout the protein body. Passing into theendosperm the protein bodies increased in size, withthe appearance of centrally-located ‘locules’ containinga-zeins. Finally, these locules fused to form a continuouscentral region of a-zeins, with the b- and c-zeins beinglocated peripherally in the mature protein bodies of thecentral endosperm cells (Fig. 4).
The analysis of protein body development in maize wasfacilitated by the availability of highly specific antibodiesfor a, b and c-zeins, and similar studies have not so farbeen reported for other cereals. However, recent workcarried out with different gliadin fractions indicates thatseparation could occur, resulting in the formation ofmicrophases. Purified a-gliadin and v-gliadin fractionswere dissolved in 70% (vuv) ethanol, mixed in ratios of1 : 3, 1 : 1 and 3 : 1, applied to a mica surface and thesolvent allowed to evaporate. Analysis of the surfaceproperties of the dried films with atomic force microscopyshowed two phases in proportions approximately relatedto the ratios of the components (McMaster et al., 1999)(Fig. 5). This suggests that gliadins and glutenins couldpartition into separate microphases in protein bodies,which would not be observed by conventional electronmicroscopy.
Expression in heterologous systems also indicated thata mixture of zein classes may be required for theformation of normal protein bodies. Thus, co-expressionof c-zein was required for the accumulation of a-zein intransgenic tobacco (Coleman et al., 1996) while b- andd-zeins formed abnormal protein bodies when expressed
alone in tobacco, but apparently normal protein bodieswhen expressed together (Bagga et al., 1995, 1997). It isclear from these studies that there is still much to learnabout the mechanisms of protein body formation incereals and the roles and organization of the differentprotein groups.
Spatial distribution of prolamins withinthe starchy endosperm
Because the aleurone cells continue to divide periclinallyin the developing cereal endosperm the youngest cells are
Fig. 4. Developmental pattern of protein body formation in maize endosperm. The most immature protein body is on the left and development is fromleft to right. Greek letter designations in the lower figure indicate the location of the corresponding zein classes as determined by immunolocalization.(Taken from Coleman et al., 1999, with permission.)
Fig. 5. Topographic image determined by scanning probe microscopy ofa 1 : 1 mixture of purified a- and v-gliadins deposited from solution on amica surface. Note the presence of two clear phases. (Taken fromMcMaster et al., 1999, with permission.)
952 Shewry and Halford
Dow
nloaded from https://academ
ic.oup.com/jxb/article-abstract/53/370/947/537249 by guest on 14 April 2019

present in the sub-aleurone layer and the oldest cellsin the central part of the endosperm. In both small graincereals and panicoid species the sub-aleurone cells containfew starch granules, which tend to be smaller than thosein the central endosperm cells. Consequently, these cellscontain high proportions of proteins, although the totalprotein content per cell varies little across the wholewheat endosperm (Evers, 1970). In maize, the proteinbodies in the sub-aleurone and outer parts of the endo-sperm are enriched in c- and b-zeins and low in a-zeins,the latter being more uniformly distributed across theendosperm (Geetha et al., 1991). This distribution isconsistent with the ontogeny of maize protein bodiesdiscussed above.
Differences in protein distribution throughout theendosperm also occur in barley (and probably also inwheat), although the developmental basis for this isunclear. The sub-aleurone cells of both species are rich inproteins, but immunocytochemical and pearling studiesof barley show that they contain mainly S-rich andS-poor prolamins (principally B and C hordeins), with theHMW prolamin (D hordein) only occurring in significantamounts below the sub-aleurone (Shewry et al., 1996;Tecsi et al., 2000). This may have significance for theutilization of wheat and barley, as D hordeins are majorcomponents of the gel protein fraction that may limit themodification of barley during malting (Smith and Lister,1983), while the homologous HMW subunits of wheatglutenin are major components of the elastomeric poly-mers that underpin breadmaking and other food uses(Shewry et al., 1995).
Regulation of prolamin gene expression
Prolamin genes are subject to tissue-specific and devel-opmental regulation, being expressed exclusively in thestarchy endosperm during mid- and late-development,and nutritional regulation, responding sensitively tothe availability of nitrogen and sulphur in the grain(Duffus and Cochrane, 1992; Giese and Hopp, 1984).This control of gene expression is exerted primarily atthe transcriptional level (Bartels and Thompson, 1986;Sørensen et al., 1989).
Little, if anything, is known about the mechanisms bywhich the expression of prolamin genes responds tosulphur. However, a motif involved in the response ofS-poor and S-rich prolamin genes of wheat, barley andrye to nitrogen has been identified. This motif (theN motif or nitrogen element) (Hammond-Kosack et al.,1993; Muller and Knudsen, 1993) is present within ahighly conserved sequence called the prolamin box. Theprolamin box (sometimes called the endosperm element)was the first prolamin gene regulatory sequence to bereported and was identified through a comparison of the
promoters of several gliadin and hordein genes (Fordeet al., 1985). This revealed the presence of the conservedsequence, which is approximately 30 bp long, around300 bp upstream of the transcription start site (it was firstcalled the �300 element). The consensus sequence forthe element is: 59-TGACATGTAA AGTGAATAAGATGAGTCATG.
The N motif is at the 39 end of the box and has theconsensus sequence G(AuG)TGAGTCAT in S-rich pro-lamin genes. It is present in the reverse orientation withinthe prolamin box of S-poor prolamin genes (Shewry et al.,1999). It has similarity with the binding site of the GCN4transcription factor, which is a component of the nitrogensignalling pathway in yeast, and is sometimes called theGCN4-like motif (GLM). The prolamin box contains asecond highly conserved motif, sequence TGTAAAGT,that has been called the endosperm or E motif(Hammond-Kosack et al., 1993).
Promoter regions containing the prolamin box havebeen shown to be functional by introducing promoteruchloramphenicol acetyl transferase (CAT) reporter geneconstructs into transgenic tobacco (Colot et al., 1987;Marris et al., 1988). A regulatory role for the prolaminbox itself has been established previously (Muller andKnudsen, 1993; Hammond-Kosack et al., 1993). Mullerand Knudsen used an homologous, transient expressionsystem, involving particle bombardment of cultured barleyendosperms with C hordein promoterub-glucuronidase(GUS) constructs. These experiments confirmed that theE and N motifs are separate elements and showed thatthe N motif exerts a negative effect on gene expression atlow nitrogen levels and interacts with the E motif andother upstream elements to give high expression whennitrogen levels are adequate.
Hammond-Kosack et al. used in vivo footprinting andgel retardation assays to show that E motifs within theprolamin box and further upstream in the promoter of awheat low molecular weight (LMW) subunit gene bounda putative transcription factor, ESBF-1 (Hammond-Kosack et al., 1993). A second putative transcriptionfactor, ESBF-II, bound the N motif prior to maximumexpression of the gene. A third putative transcriptionfactor, SPA, has since been shown to recognize the Nmotif (Albani et al., 1997).
Together, the results of these experiments suggestthat the N motif is an important component of thenitrogen regulatory mechanism for S-rich and S-poorprolamin genes. Its function requires interaction withthe E motif, the two motifs together making up theprolamin box. However, the prolamin box is not presentin all prolamin genes. Zein gene promoters, for example,contain a highly conserved 15 bp element that has beensuggested to act as a tissue-specific enhancer (Quayle andFeix, 1992). It contains the sequence TGTAAAG, whichresembles the E motif, but the N motif is absent (Coleman
Cereal seed storage proteins 953
Dow
nloaded from https://academ
ic.oup.com/jxb/article-abstract/53/370/947/537249 by guest on 14 April 2019

and Larkins, 1999). The N motif is present in c-zeinpromoters, but it is separated from the E motif and itsfunction, if any, has not been investigated (Coleman andLarkins, 1999).
The prolamin box in its entirety is also not presentin HMW prolamin gene promoters (Shewry et al., 1999).Instead, HMW prolamin promoters contain a majorregulatory element (identified by Thomas and Flavell,1990) which is located in a 38 bp sequence withthe consensus: 59-GTTTTGCAAA GCTCCAATTGCTCCTTGCTT ATCCAGCT.
The location of this sequence is highly conserved inall HMW prolamin promoters, beginning at position�185 to �189 (Shewry et al., 1999). The element con-tains the sequence TGCAAAG, which is similar to theE motif sequence TGTAAAG that is also present inzein genes, but it does not contain anything resemblingthe N motif.
Sequences corresponding to parts of the N and Emotifs are present in HMW prolamin promotersupstream from the major enhancer (Lamacchia et al.,2001). However, deletion of these sequences does notappear to affect activity of the promoter, at least whendriving reporter gene expression in transgenic tobacco(Halford et al., 1989; Thomas and Flavell, 1990).
Cereal seed proteins and grain utilization
The total protein contents of cereal seeds vary from about10–15% of the grain dry weight, with about half ofthe total being storage proteins. Nevertheless, proteinshave major impacts on the end use properties of the grain.The prolamins, which form the major storage proteinfraction in all the major cereals except oats and rice,are deficient in the essential amino acids lysine and inthreonine and tryptophan (particularly in maize). Thisresults in nutritional deficiencies in these amino acidswhen the whole grain are fed to monogastric livestocksuch as pigs and poultry. It is therefore usual to combinecereals with other sources of these amino acids for animalfeed, for example, legume seeds (notably soybean), oilseedcake, fish meal or synthetic amino acids. The combinationof cereals and legume seeds is particularly favoured, asthese two types of seeds are essentially complementary intheir compositions of essential amino acids: cereals tendto be rich in sulphur-containing amino acids and low inlysine and legume seeds vice versa.
The nutritional quality of cereals is not generally animportant consideration for human diets in the developedworld, although it is still important in some developingcountries. The major consideration is the impact ofthe grain proteins on functional properties for foodprocessing, since the bulk of all cereals, except rice,are consumed in processed foods. Processing quality is
particularly important for wheat where the glutenproteins are the major determinant of end use quality.
Gluten proteins and wheat grain quality
Most of the wheat consumed by humans is processedfrom white flour, which is produced by milling to removethe germ (embryo) and bran (pericarp, testa, nucellarlayer, and aleurone layer). It therefore corresponds to thestarchy endosperm cells and contains high proportions ofstarch and gluten.
The gluten proteins form a continuous matrix in themature dry endosperm cells, as discussed above. Whenflour is mixed with water to form a dough, the proteinmatrices in the individual cells are brought together toform a continuous network. This confers visco-elasticproperties that allow the dough to be expanded byfermentation and baked into leavened bread or processedinto pasta, noodles and a range of other foods.
The molecular basis for the visco-elastic properties ofwheat gluten has fascinated cereal scientists for many yearsas a phenomenon of fundamental interest as well as inrelation to improving the end use properties of wheatflours. Of particular importance are the glutenin polymers,and it is well established that strong (i.e. highly visco-elastic) doughs contain high proportions of high molecularmass glutenin polymers (Field et al., 1983b). However, abreakthrough in understanding came about 20 yearsago when Payne and co-workers demonstrated that allelicvariation in the composition of the HMW prolamins (theHMW subunits of glutenin) was strongly correlated withdifferences in the breadmaking quality of European breadwheats (Payne, 1987). This association has been con-firmed in many laboratories worldwide and has led todetailed studies of HMW subunit structure and properties.
Cultivars of hexaploid bread wheat have six HMWsubunit genes, two each at the Glu-1 loci on the long armsof the group 1 chromosomes (1A, 1B, 1D). Each of theseloci encodes one x-type and one y-type subunit. However,differences in gene expression result in the presence ofonly three, four or five HMW subunit proteins, with 1Dx,1Dy and 1Bx subunits being present in all cultivars and1Ax anduor 1By subunits only in some cultivars. Goodbreadmaking quality is particularly associated with thepresence of a 1Ax subunit (compared with the silent ornull allele) and the chromosome 1D-encoded subunit pair1Dx5q1Dy10 (compared with the allelic subunit pairs1Dx2q1Dy12, 1Dx3q1Dy12 and 1Dx4q1Dy12).
Results from a range of studies are consistent with thehypothesis that the HMW subunits form an elastomericpolymer network which provides a ‘backbone’ for inter-actions with other glutenin subunits and with gliadins.There is little doubt that this network is stabilized byinter-chain disulphide bonds (Shewry and Tatham, 1997),but Belton has proposed that inter-chain hydrogen bonds,
954 Shewry and Halford
Dow
nloaded from https://academ
ic.oup.com/jxb/article-abstract/53/370/947/537249 by guest on 14 April 2019

formed in particular between glutamine residues presentin the repetitive domains (Fig. 1), are also important inconferring elasticity (Belton, 1999).
The biological significance of the biophysical pro-perties of gluten is unknown, as the proteins functionessentially as grain storage proteins and have no knownbiological requirement to exhibit visco-elasticity. How-ever, it is probable that the molecular interactions whichdetermine these properties are initially established in thedeveloping grain, because protein bodies isolated duringthe mid-development stage contain disulphide-linkedglutenin polymers (Field et al., 1983a) which can bedemonstrated to exhibit visco-elasticity (authors’ unpub-lished observations). The precise roles of the enzyme,protein disulphide isomerase, in catalysing disulphidebond formation between gluten proteins in the ER and ofBiP in establishing other protein : protein interactionsremain uncertain (Grimwade et al., 1996; DuPont et al.,1998; Galili, 1997; Shewry, 1999).
Manipulating HMW subunit composition andwheat grain quality
A number of factors have contributed to the choice of theHMW subunits as an early target for genetic engineeringto improve grain quality. They show clear associationswith grain quality, with both quantitative effects relatedto gene expression (e.g. 1Ax subunits) (Halford et al.,1992) and qualitative effects associated with allelic differ-ences in subunit structure and properties (e.g. subunits1Dx5q1Dy10). Furthermore, a number of genes encod-ing subunits associated with good and poor breadmak-ing quality are available, and the transgene productsare readily identified by simple SDS-PAGE of totalseed proteins.
Four laboratories have so far reported the expressionof HMW subunit transgenes in bread wheat, in each caseusing either the gene’s own promoter or that of anotherHMW subunit gene (Blechl and Anderson, 1996; Altpeteret al., 1996; Barro et al., 1997; Alvarez et al., 2000). Allreported expression of the transgenes at levels up to orexceeding those of the endogenous HMW subunit genes.Transgene expression also appeared to be restrictedto starchy endosperm cells, and this was confirmed bythe analysis of transgenic wheat lines in which theHMW subunit 1Dx5 gene promoter was used to drivethe UidA reporter gene encoding b-glucuronidase (Gus)(Lamacchia et al., 2001).
The impact of the HMW subunit transgenes on doughstrength has been determined on selected lines grown inthe glasshouse (Barro et al., 1997; Rooke et al., 1999) andfield plots (Popineau et al., 2001) using the Mixographwhich measures the energy input during dough mixing.This showed that the expression of the HMW subunit1Ax1 transgene in a poor quality background did indeed
result in the expected improvement in dough strength.However, when the HMW subunit 1Dx5 gene was highlyexpressed in either the same background or in a goodbreadmaking quality background, it had unexpectedeffects. Flours milled from these lines failed to forma normal dough on hydration and mixing, resulting indecreases in the Mixograph parameters that are usuallyassociated with dough strength. Rheological analysis ofglutens isolated from these lines indicated that the con-nectivity of the gluten network had been greatly increased,by about 1003 and 103 in the poor and good qualitybackgrounds, respectively (Popineau et al., 2001), result-ing in similar properties to those of gluten modified bytransglutaminase (Larre et al., 2000).
An increase in the degree of cross-linking betweengluten proteins in the transgenic lines may have resultedfrom the presence of a cysteine residue towards theN-terminal end of the repetitive domain of subunit 1Dx5(Fig. 1) because cysteine residues are not present inequivalent positions in other 1Dx subunits. However,there is also evidence that allelic pairs of HMW subunitssuch as 1Dx5q1Dy10 exist as dimers in the gluteninpolymers (Shewry and Tatham, 1997). The expression ofhigh levels of subunit 1Dx5 in the absence of equivalentamounts of subunit 1Dy10 could, therefore, result in adrastic restructuring of the glutenin polymers.
It is clear from the preliminary results discussed abovethat it is feasible to manipulate the structure and pro-perties of wheat gluten by genetic engineering, althoughthe current knowledge of gluten structure and function-ality is still insufficient to predict the results reliably.Bread wheat is currently being transformed with a rangeof wild-type and mutant genes encoding HMW subunitsand other gluten proteins. This should allow the deter-mination of the precise roles of specific proteins andstructural features as well as the production of lines withimproved properties suitable for incorporation into plantbreeding programmes.
Conclusions
The storage proteins of cereals are of immense import-ance in determining the quality and end use propertiesof the grain. Understanding the structures of these pro-teins, their biophysical and functional properties, and thebiological mechanisms which determine their synthesis,trafficking and deposition in the grain is important tounderpin future attempts to improve the end use qualityof grain by genetic engineering.
Acknowledgement
IACR receives grant-aided support from the Biotechnology andBiological Sciences Research Council of the United Kingdom.
Cereal seed storage proteins 955
Dow
nloaded from https://academ
ic.oup.com/jxb/article-abstract/53/370/947/537249 by guest on 14 April 2019

References
Albani D, Hammond-Kosack MCU, Smith C, Conlan S, Colot V,Holdsworth M, Bevan MW. 1997. The wheat transcriptionalactivator SPA: a seed-specific bZIP protein that recognizesthe GCN4-like motif in the bifactorial endosperm boxof prolamin genes. The Plant Cell 9, 171–184.
Altpeter E, Vasil V, Srivastava V, Vasil IK. 1996. Integra-tion and expression of the high-molecular-weight gluteninsubunit 1Ax1 gene into wheat. Nature Biotechnology14, 1151–1159.
Altschuler Y, Galili G. 1994. Role of conserved cysteines ofa wheat gliadin in its transport and assembly into proteinbodies in Xenopus oocytes. Journal of Biological Chemistry269, 6677–6682.
Altschuler Y, Harel R, Galili G. 1993. Role of the N- andC-terminal regions of a wheat c-gliadin in its transport via theendoplasmic reticulum of Xenopus oocytes. The Plant Cell5, 443–450.
Alvarez ML, Guelman S, Halford NG, Lustig S, Reggiardo MI,Ryabushkina N, Shewry P, Stein J, Vallejos RH. 2000.Silencing of HMW glutenins in transgenic wheat expressingextra HMW subunits. Theoretical and Applied Genetics100, 319–327.
Anderson OD, Greene FC, Yip RE, Halford NG, Shewry PR,Malpica-Romero J-M. 1989. Nucleotide sequences of the twohigh-molecular-weight glutenin genes from the D-genome of ahexaploid bread wheat, Triticum aestivum L. cv. Cheyenne.Nucleic Acids Research 17, 461–462.
Bagga S, Adams H, Kemp JD, Sengupta-Gopalan C. 1995.Accumulation of 15-kilodalton zein in novel protein bodiesin transgenic tobacco. Plant Physiology 107, 13–23.
Bagga S, Adams H, Rodriquez FD, Kemp JD,Sengupta-Gopalan C. 1997. Co-expression of the maized- and b-zein genes results in stable accumulation of d-zeinin ER-derived protein bodies formed by b-zein. The PlantCell 9, 1683–1696.
Barro F, Rooke L, Bekes F, Gras P, Tatham AS, Fido R,Lazzeri PA, Shewry PR, Barceo P. 1997. Transformationof wheat with high molecular weight subunit genes resultsin improved functional properties. Nature Biotechnology15, 1295–1299.
Bartels D, Thompson RD. 1986. Synthesis of messenger-RNAscoding for abundant endosperm proteins during wheat graindevelopment. Plant Science 46, 117–125.
Bartels D, Altosaar I, Harberd NP, Barker RF, Thompson RD.1986. Molecular analysis of c-gliadin gene families at thecomplex Gli-1 locus of bread wheat (T. aestivum L.).Theoretical and Applied Genetics 72, 845–853.
Beccari. 1745. De Frumento. De Bononiensi Scientiarum etArtium Instituto atque Academia Commentarii, II. Part I.,122–127.
Bechtel DB, Wilson JD, Shewry PR. 1991. Immunocytochemicallocalization of the wheat storage protein triticin in developingendosperm tissue. Cereal Chemistry 68, 573–577.
Belton PS. 1999. On the elasticity of wheat gluten. Journal ofCereal Science 29, 103–107.
Blechl AE, Anderson OD. 1996. Expression of a novel high-molecular-weight glutenin subunit gene in transgenic wheat.Nature Biotechnology 14, 875–879.
Burgess SR, Shewry PR. 1986. Identification of homologousglobulins from embryos of wheat, barley, rye and oats.Journal of Experimental Botany 37, 1863–1871.
Casey R. 1999. Distribution and some properties of seedglobulins. In: Shewry PR, Casey R, eds. Seed proteins.Dordrecht: Kluwer Academic Publishers, 159–169.
Choi S-B, Wang C, Muench DG, Ozawa K, Franceschi VR,Wu Y, Okita TW. 2000. Messenger RNA targeting of riceseed storage proteins to specific ER subdomains. Nature407, 765–767.
Coleman CE, Larkins BA. 1999. The prolamins of maize. In:Shewry PR, Casey R, eds. Seed proteins. Dordrecht: KluwerAcademic Publishers, 109–139.
Coleman CE, Herman EM, Takasaki K, Larkins BA. 1996. Themaize c-zein sequesters a-zein and stabilizes its accumulationin protein bodies of transgenic tobacco endosperm. The PlantCell 8, 2335–2345.
Colot V, Robert LS, Kavanagh TA, Bevan MW, Thompson RD.1987. Localization of sequences in wheat endosperm proteingenes which confer tissue-specific expression in tobacco. TheEMBO Journal 6, 3559–3564.
Cuddeford D. 1995. Oats for animal feed. In: Welch RW, ed.The oat crop: production and utilization. London: Chapman& Hall, 321–368.
Duffus CM, Cochrane MP. 1992. Grain structure and composi-tion. In Shewry PR, ed. Barley: genetics, biochemistry,molecular biology and biotechnology. Wallingford: CABInternational, 291–317.
DuPont FM, Hurkman WJ, Tanaka CK, Chan R. 1998. BiP,HSP70, NDK and PDI in wheat endosperm. I. Accumulationof MRNA and protein during grain development. PhysiologiaPlantarum 103, 70–79.
Evers AD. 1970. Development of the endosperm of wheat.Annals of Botany 34, 547–555.
FAO. 1999. FAO Yearbook Production 53.Field JM, Shewry PR, Burgess SR, Forde J, Parmar S, Miflin BJ.
1983a. The presence of high molecular weight aggregates inthe protein bodies of developing endosperms of wheat andother cereals. Journal of Cereal Science 1, 33–41.
Field JM, Shewry PR, Miflin BJ. 1983b. Solubilization andcharacterization of wheat gluten proteins; correlationsbetween the amount of aggregated proteins and bakingquality. Journal of the Science of Food and Agriculture34, 370–377.
Forde BG, Heyworth A, Pywell J, Kreis M. 1985. Nucleotidesequence of a B1 hordein gene and the identification ofpossible upstream regulatory elements in endosperm storageprotein genes from barley, wheat and maize. Nucleic AcidsResearch 13, 7327–7339.
Galili G. 1997. The prolamin storage proteins of wheat and itsrelatives. In: Larkins BA, Vasil IK, eds. Cellular and molecularbiology of plant seed development. The Netherlands: KluwerAcademic Publishers, 221–256.
Geetha KB, Lending CR, Lopes MA, Wallace JC, Larkins BA.1991. Opaque-2 modifiers increase c-zein synthesis andalter its spatial distribution in maize endosperm. The PlantCell 3, 1207–1219.
Geli MI, Torrent M, Ludevid D. 1994. Two structural domainsmediate two sequential events in c-zein targeting: proteinendoplasmic reticulum retention and protein body formation.The Plant Cell 6, 1911–1922.
Giese H, Hopp E. 1984. Influence of nitrogen nutrition on theamount of hordein, protein Z and b-amylase messengerRNA in developing endosperms of barley. Carlsberg ResearchCommunications 49, 365–383.
Grimwade B, Tatham AS, Freedman RB, Shewry PR, Napier JA.1996. Comparison of the expression patterns of wheatgluten proteins and proteins involved in the secretory path-way in developing caryopses of wheat. Plant MolecularBiology 30, 1067–1073.
Halford NG, Forde J, Shewry PR, Kreis M. 1989. Functionalanalysis of the upstream regions of a silent and an expressed
956 Shewry and Halford
Dow
nloaded from https://academ
ic.oup.com/jxb/article-abstract/53/370/947/537249 by guest on 14 April 2019

member of a family of wheat seed protein genes in transgenictobacco. Plant Science 62, 207–216.
Halford NG, Field JM, Blair H, Urwin P, Moore K, Robert L,Thompson R, Flavell RB, Tatham AS, Shewry PR. 1992.Analysis of HMW glutenin subunits encoded by chromosome1A of bread wheat Triticum aestivum L. indicates quantitativeeffects on grain quality. Theoretical and Applied Genetics83, 373–378.
Hammond-Kosack MCU, Holdsworth MJ, Bevan MW. 1993. Invivo footprinting of a low molecular weight glutenin gene(LMWG-1D1) in wheat endosperm. The EMBO Journal12, 545–554.
Heck GR, Chamberlain AK, Ho DT-H. 1993. Barley embryoglobulin 1 gene, Beg1: characterization of cDNA chromo-some mapping and regulation of expression. Molecular andGeneral Genetics 239, 209–218.
Horikoshi M, Morita Y. 1975. Localization of c-globulin in riceseed and changes in c-globulin content during seed develop-ment and germination. Agricultural and Biological Chemistry39, 2309–2314.
Hsia CC, Anderson OD. 2001. Isolation and characterizationof wheat v-gliadin genes. Theoretical and Applied Genetics103, 37–44.
Kermode AR, Bewley JD. 1999. Synthesis, processing anddeposition of seed proteins: The pathway of protein synthesisand deposition of the cell. In: Shewry PR, Casey R, eds. Seedproteins. Dordrecht: Kluwer Academic Publishers, 807–841.
Kreis M, Forde BG, Rahman S, Miflin BJ, Shewry PR. 1985.Molecular evolution of the seed storage proteins of barley, ryeand wheat. Journal of Molecular Biology 183, 499–502.
Krishnan HB, Franceschi VR, Okita TW. 1986. Immuno-chemical studies on the role of the Golgi complex in proteinbody formation in rice seeds Planta 169, 471–480.
Kriz AL. 1989. Characterization of embryo globulins encodedby the maize Glb genes. Biochemical Genetics 27, 239–251.
Kriz AL. 1999. 7S globulins of cereals. In: Shewry PR, Casey R,eds. Seed proteins. Dordrecht: Kluwer Academic Publishers,477–498.
Kriz AL, Schwartz D. 1986. Synthesis of globulins in maizeembryos. Plant Physiology 82, 1069–1075.
Kriz AL, Wallace NH. 1991. Characterization of the maizeGlobulin-2 gene and analysis of two null alleles. BiochemicalGenetics 29, 241–254.
Lamacchia C, Shewry PR, Di Fonzo N, Forsyth JL, Harris N,Lazzeri PA, Napier JA, Halford NG, Barcelo P. 2001.Endosperm-specific activity of a storage protein gene pro-moter in transgenic wheat seed. Journal of ExperimentalBotany 52, 243–250.
Larre C, Denery-Papini S, Popineau Y, Deshayes G, Lefebvre J.2000. Biochemical analysis and rheological properties ofgluten modified by transglutaminase. Cereal Chemistry 77,
121–127.Leite A, Neto GC, Vettore AL, Yunes JA, Arruda P. 1999.
The prolamins of sorghum, Coix and millets. In: Shewry PR,Casey R, eds. Seed proteins. Dordrecht: Kluwer AcademicPublishers, 141–157.
Lending CR, Larkins BA. 1989. Changes in the zein composi-tion of protein bodies during maize endosperm development.The Plant Cell 1, 1011–1023.
Lending CR, Chesnut RS, Shaw KL, Larkins BA. 1989.Immunolocalization of avenin and globulin storageproteins in developing endosperm of Avena sativa L. Planta178, 315–324.
Li X, Franceschi V, Okita TW. 1993a. Segregation of storageprotein mRNAs on the rough endoplasmic reticulummembranes of rice endosperm membrane. Cell 72, 869–879.
Li X, Wu Y, Zhang D-Z, Gillikin JW, Boston RS, Franceschi VR,Okita TW. 1993b. Rice prolamin protein body biogenesis: aBiP-mediated process. Science 262, 1054–1056.
Lockhart HB, Hurt HD. 1986. Nutrition of oats. In: WebsterFH, ed. Oats: chemistry and technology. St Paul, Minnesota,USA: American Association of Cereal Chemists, Inc.,297–308.
Marris C, Gallois P, Copley J, Kreis M. 1988. The 59 flankingregion of a barley B hordein gene controls tissue and develop-mental specific CAT expression in tobacco plants. PlantMolecular Biology 10, 359–366.
McMaster TJ, Miles MJ, Wannerberger L, Eliasson A-C,Shewry PR, Tatham AS. 1999. The identification of micro-phases in mixed a- and c-gliadin protein films investigated byatomic force microscopy. Journal of Agricultural and FoodChemistry 47, 5093–5099.
Muench DG, Wu Y, Coughlan SJ, Okita TW. 1998. Evidence fora cytoskeleton-associated binding site involved in prolaminemRNA localization to the protein bodies in rice endospermtissue. Plant Physiology 116, 559–569.
Muench DG, Ogawa M, Okita TW. 1999. The prolamins of rice.In: Shewry PR, Casey R, eds. Seed proteins. Dordrecht:Kluwer Academic Publishers, 93–108.
Muller M, Knudsen S. 1993. The nitrogen response of a barleyC-hordein promoter is controlled by positive and negativeregulation of the GCN4 and endosperm box. The PlantJournal 4, 343–355.
Payne PI. 1987. Genetics of wheat storage proteins and theeffect of allelic variation on bread-making quality. AnnualReview of Plant Physiology 38, 141–153.
Popineau Y, Deshayes G, Lefebvre J, Fido RJ, Tatham AS,Shewry PR. 2001. Prolamin aggregation, gluten viscoelasticityand mixing properties of transgenic wheat lines expressing1Ax and 1Dx HMW glutenin subunit transgenes. Journal ofAgricultural and Food Chemistry 49, 395–401.
Quayle T, Feix G. 1992. Functional analysis of the �300 regionof maize zein genes. Molecular and General Genetics231, 369–374.
Rooke L, Bekes F, Fido R, Barro F, Gras P, Tatham AS,Barcelo P, Lazzeri P, Shewry PR. 1999. Overexpression of agluten protein in transgenic wheat results in highly elasticdough. Journal of Cereal Science 30, 115–120.
Shewry PR. 1999. The synthesis, processing and deposition ofgluten proteins in the developing wheat grain. Cereal FoodsWorld 44, 587–589.
Shewry PR, Tatham AS. 1990. The prolamin storage proteins ofcereal seeds: structure and evolution. Biochemical Journal267, 1–12.
Shewry PR, Tatham AS. 1997. Disulphide bonds in wheat glutenproteins. Journal of Cereal Science 25, 207–227.
Shewry PR, Tatham AS, Barro F, Barcelo P, Lazzeri P. 1995.Biotechnology of breadmaking: unravelling and manip-ulating the multi-protein gluten complex. BiouTechnology13, 1185–1190.
Shewry PR, Brennan C, Tatham AS, Warburton T, Fido R,Smith D, Griggs D, Cantrell I, Harris N. 1996. Thedevelopment, structure and composition of the barley grainin relation to its end use properties. In: Cereals 96.Proceedings of the 46th Australian Cereal ChemistryConference, Sydney, September 1996, 158–162.
Shewry PR, Tatham AS, Halford NG. 1999. The prolamins ofthe Triticeae. In: Shewry PR, Casey R, eds. Seed proteins.Dordrecht: Kluwer Academic Publishers, 35–78.
Shotwell MA. 1999. Oat globulins. In: Shewry PR, Casey R, eds.Seed proteins. Dordrecht: Kluwer Academic Publishers,389–400.
Cereal seed storage proteins 957
Dow
nloaded from https://academ
ic.oup.com/jxb/article-abstract/53/370/947/537249 by guest on 14 April 2019

Singh NK, Shepherd KW. 1985. The structure andgenetic control of a new class of disulphide-linked proteinsin wheat endosperm. Theoretical and Applied Genetics71, 79–92.
Singh NK, Shepherd KW, Langridge P, Gruen LC, Skerritt JH,Wrigley CW. 1988. Identification of legumin-like proteins inwheat. Plant Molecular Biology 11, 633–639.
Singh NK, Donovan GR, Carpenter HC, Skerritt JH,Langridge P. 1993. Isolation and characterization of wheattriticin cDNA revealing a unique lysing-rich repetitivedomain. Plant Molecular Biology 22, 227–237.
Smith DB, Lister PR. 1983. Gel forming proteins in barley grainand their relationship with malting quality. Journal of CerealScience 1, 229–239.
Sørensen MB, Cameron-Mills V, Brandt A. 1989. Tran-scriptional and post-transcriptional regulation of geneexpression in developing barley endosperm. Molecular andGeneral Genetics 217, 195–201.
Takaiwa F, Ogawa M, Okita TW. 1999. Rich glutelins. In:Shewry PR, Casey R, eds. Seed proteins. Dordrecht: KluwerAcademic Publishers, 401–425.
Tecsi L, Darlington HF, Harris N, Shewry PR. 2000. Patterns ofprotein deposition and distribution in developing and maturebarley grain. In: Barley genetics, VIII. Proceedings of the8th International Barley Genetics Symposium, Adelaide,Australia, Vol. 2, 266–268.
Thomas MS, Flavell RB. 1990. Identification of an enhancerelement for the endosperm-specific expression of highmolecular weight glutenin. The Plant Cell 2, 1171–1180.
Torrent M, Geli MI, Ruiz-Avila L, Canals J, Puigdomenech P,Ludevid MD. 1994. Role of structural domains for maizec-zein retention in Xenopus oocytes. Planta 192, 512–518.
Wallace NH, Kriz AL. 1991. Nucleotide sequence of a cDNAclone corresponding to the maize Globulin-2 gene. PlantPhysiology 95, 973–975.
Yamagata H, Tanaka K. 1986. The site of synthesis andaccumulation of rice storage proteins. Plant and CellPhysiology 27, 135–145.
Yupsanis T, Burgess SR, Jackson PJ, Shewry PR. 1990.Characterization of the major protein component fromaleurone cells of barley Hordeum vulgare L. Journal ofExperimental Botany 41, 385–392.
958 Shewry and Halford
Dow
nloaded from https://academ
ic.oup.com/jxb/article-abstract/53/370/947/537249 by guest on 14 April 2019