caulerpa taxifolia in seagrass meadows: killer or opportunistic weed?
TRANSCRIPT

ORIGINAL PAPER
Caulerpa taxifolia in seagrass meadows: killeror opportunistic weed?
Tim M. Glasby
Received: 28 May 2012 / Accepted: 17 October 2012
� Springer Science+Business Media Dordrecht 2012
Abstract Seagrass habitats are being lost throughout
the world and the invasive alga C. taxifolia has often
been implicated in seagrass declines. Although C. taxi-
folia can impact a variety of species, evidence for its
effects on seagrasses is largely correlative. This study
combined observational studies and manipulative
experiments done over many years to test hypotheses
about effects of C. taxifolia on two Australian
seagrasses, namely Posidonia australis and Zostera
capricorni. Results indicated that C. taxifolia is not
having adverse impacts on the coverage of these
seagrasses in the sites studied. Rather, C. taxifolia
appears to be an opportunist, persisting longer and its
coverage being greater in previously non-vegetated
sediments than amongst seagrasses. C. taxifolia
co-existed with P. australis and did not cause reduc-
tions in the cover of the seagrass. Outcomes of
experimental manipulations of C. taxifolia amongst
Z. capriconi were less clear due to losses of Z. capri-
coni in all plots, regardless of the presence of
C. taxifolia. It was possible that C. taxifolia may have
enhanced the decline in canopy cover of Z. capricorni,
but the presence of alga did not alter the final fate of
Z. capricorni. There was also no evidence that long-
term areal coverage of P. australis or Z. capriconi has
been affected by the introduction of C. taxifolia in the
embayments studied. A review of literature on effects
of species of Caulerpa on seagrasses provided limited
experimental evidence for negative impacts of this
genus on seagrass abundance.
Keywords Invasive species � Caulerpa taxifolia �Marine algae � Seagrass � Experimental � Review
Introduction
There are numerous examples of negative correlations
between abundances of introduced and native species
(Parker et al. 1999; Ruiz et al. 1999; Bruno et al. 2005;
Simberloff 2009), but there are fewer instances for
which there is experimental evidence that an intro-
duced species has caused the decline of a native (i.e.
been the driver of change; MacDougall and Turking-
ton 2005). Removal experiments can be useful for
testing whether an introduced species is driving
ecological change (removal would result in the
recovery of the native assemblage, e.g. Bulleri et al.
2010), although results could be confused if impacts
are long-term (e.g. recovery is slow, Glasby and
Underwood 1996), or if there have been historical or
multiple interacting drivers of change (Didham et al.
2005). Moreover, due to varying degrees of hysteresis,
recovery after the removal of an invader might elicit
community responses that are quite different from
T. M. Glasby (&)
New South Wales Department of Primary Industries,
Port Stephens Fisheries Institute, Locked Bag 1,
Nelson Bay, NSW 2315, Australia
e-mail: [email protected]
123
Biol Invasions
DOI 10.1007/s10530-012-0347-1

those associated with the initial invasion (Dıaz et al.
2003). A more direct test of impacts of invasion,
therefore, is to use experimental additions of an
introduced species, noting that this can potentially
raise ethical concerns and impacts might still take a
long time to manifest.
Marine macroalgae account for some 20 % of the
known introductions of marine species globally
(Schaffelke et al. 2006). The ecological effects of
introduced marine macroalgae have been reviewed
comprehensively over the last few years (e.g. Schaff-
elke et al. 2006; Williams and Smith 2007; Thomsen
et al. 2009) and the consensus is that there is a dearth of
experimental data and hence limited understanding of
their impacts. The few high profile invasive macroal-
gae that have been studied experimentally can have
negative effects on native biota, but impacts are not
always detected (Williams and Smith 2007; Thomsen
et al. 2009). In comparison to other marine habitats,
the effects of introduced species in seagrass has
received relatively little attention (Williams 2007).
When seagrass beds are invaded, it is often by
macroalgae and in situations where the seagrass beds
have already been disturbed (Williams 2007). Species
of the genus Caulerpa are the best known macroalgal
invaders of seagrass beds and one of the better studied
species is Caulerpa taxifolia.
Much has been written about C. taxifolia since its
rise to prominence in the mid 1980s. C. taxifolia is a
highly invasive green alga, capable of spreading
rapidly over large areas and creating dense mats
(Meinesz et al. 2001; Wright and Davis 2006).
C. taxifolia can invade sheltered or exposed marine
habitats (Meinesz et al. 1993), including seagrass beds
(Ceccherelli and Cinelli 1998), non-vegetated soft
sediments (Jaubert et al. 1999) and rocky reefs
(Bellan-Santini et al. 1996; Ceccherelli et al. 2002).
Beds of C. taxifolia can affect the feeding behaviour
and distribution of benthic fishes (Levi and Francour
2004; Longepierre et al. 2005) and support assem-
blages of fish (York et al. 2006) and invertebrates
(McKinnon et al. 2009; Gribben et al. 2009a) that
differ from those in adjacent native habitats. The
recruitment of clams can be enhanced in beds of
C. taxifolia relative to non-vegetated habitats, but the
growth, survivorship and reproductive capacity of
those clams can be reduced by the alga (Gribben and
Wright 2006; Wright et al. 2007; Gribben et al. 2009b;
Byers et al. 2010).
Various authors have asserted that C. taxifolia can
cause the regression of seagrasses (e.g. Boudouresque
et al. 1995; Glardon et al. 2008; Francour et al. 2009;
Lapointe and Bedford 2010; Peirano et al. 2011), but
most evidence for this comes from correlative studies.
Observations of Posidonia oceanica beds in the
Mediterranean Sea soon after they were invaded by
C. taxifolia indicated that the seagrass showed signs of
etiolation, chlorosis and had reduced numbers of leaves
compared to an uninvaded site (Meinesz et al. 1993; de
Villele and Verlaque 1995). After C. taxifolia invaded
sites in southern California, Williams and Grosholz
2002) documented considerably reduced biomass
of the seagrass Ruppia maritima in patches where
C. taxifolia was abundant. Manipulative experiments
in Italy demonstrated that the shoot density of
the seagrass Cymodocea nodosa was decreased by
C. taxifolia over a period of 4 months (Ceccherelli and
Cinelli 1997), but a subsequent 13 month study
indicated no significant effects (Ceccherelli and Sechi
2002).
There is evidence that abundances of seagrasses at
some sites in the Mediterranean declined prior to the
introduction of C. taxifolia (Chisholm et al. 1997) and
that seagrass coverage has not in fact declined since
the introduction of the alga (Jaubert et al. 1999;
Ceccherelli and Sechi 2002). Given the limited
experimental evidence for impacts of C. taxifolia on
seagrasses (see Table 1), it is quite possible that the
alga may, at least in some cases, be responding
opportunistically to reductions of seagrass that are
caused by some other disturbance. What seems clear is
that it is inappropriate to generalise about any impacts
of C. taxifolia on seagrasses.
Caulerpa taxifolia was first discovered in the
temperate waters of New South Wales (NSW), Aus-
tralia, in 2000 and is currently present in 13 estuaries or
coastal lakes. It appears that the strain of C. taxifolia in
NSW is different from that in the Mediterranean
(Meusnier et al. 2004), although genetic data suggest
there have been several introductions of C. taxifolia
into NSW (Schaffelke et al. 2002). C. taxifolia was
declared noxious in NSW after its appearance in
isolated patches some 750 km south of native sub-
tropical populations in Queensland, and its subsequent
rapid spread (Glasby and Creese 2007). Observations
in NSW indicated that, as in the Mediterranean
(Ceccherelli and Cinelli 1999), there were negative
associations between abundances of seagrasses
T. M. Glasby
123
(P. australis and Z. capricorni) and C. taxifolia. A
possible explanation for such a pattern is that sparse
(\50 % coverage) seagrass is susceptible to impacts
from C. taxifolia, whereas dense ([50 %) seagrass is
not because it negatively affects C. taxifolia (e.g.
through competition for resources such as light).
Table 1 Putative effects of Caulerpa spp. on seagrasses based on mensurative studies
Seagrass
species
Putative
effect
Evidence Apparent response of seagrass Reference
Posidoniaoceanica
-ve C Shorter, narrower leaves at one site with C. taxifolia cf another without.
Necrosis of leave base where C. taxifolia smothering seagrass
de Villele and
Verlaque
(1995)
-ve C P. oceanica leaves produced more phenolic compounds (presumed to be a
stress response) at one site with C. taxifolia cf another without
Cuny et al.
(1995)
-ve C Longevity of P. oceanica leaves reduced where C. taxifolia was dense. At
some times of sampling, shorter leaves at one site with C. taxifolia cf
another without. Leaf productivity greater (thought to be a stress
response) in presence of C. taxifolia
Dumay et al.
(2002)
n C Areal coverage of seagrass unchanged 8 years after invasion of
C. taxifoliaJaubert et al.
(2003)
n C Number of leaves similar in areas with or without C. taxifolia Dumay et al.
(2002)
n C No indication that rhizome growth or shoot density where C. taxifoliapresent were worse than uninvaded sites
Peirano et al.
(2005)
-ve C Tendency for reduced meristematic activity in P. oceanica when mixed
with C. taxifolia or C. proliferaGarcias-Bonet
et al. (2008)
-ve C Leaves of P. oceanica smaller (and leaf turnover enhanced) when in the
presence of C. taxifoliaPergent et al.
(2008)
-ve C More plagiotropic shoots (thought to be a stress response) at one seagrass
site with C. taxifolia cf another without
Molenaar et al.
(2009)
-ve C Negative associations between P. oceanica shoot density and biomass of
three species of Caulerpa (C. prolifera, C. racemosa, C. taxifolia)
Holmer et al.
(2009)
Posidoniaaustralis
n E Cover of seagrass not affected by C. taxifolia Current study
Cymodoceanodosa
-ve E Reduced seagrass shoot density in presence of C. taxifolia (over 4 months) Ceccherelli and
Cinelli (1997)
-ve E Reduced seagrass shoot density in presence of C. racemosa (over
14 months)
Ceccherelli and
Campo (2002)
n E No effect of C. taxifolia on seagrass shoot density after 13 months Ceccherelli and
Sechi (2002)
Zosteranoltii
?ve E Increased seagrass shoot density in presence of C. racemosa (over
14 months)
Ceccherelli and
Campo (2002)
Zosteracapricorni
n(?) E Some indication that decline of seagrass canopy was exacerbated by
C. taxifolia, but Z. capricorni died in the absence of C. taxifoliaCurrent study
Ruppiamaritima
-ve C Biomass of R. maritima less in presence of C. taxifolia Williams and
Grosholz
(2002)
Halodulewrightii
-ve E Shoot density and biomass of H. wrightii were reduced in the presence of
C. prolifera (native) over 6 months
Taplin et al.
(2005)
-ve E Where C. prolifera (native) was particularly abundant, the alga colonised
gaps before H. wrightii and potentially excluded the seagrass over
15 month
Stafford and Bell
(2006)
Negative effect (-ve), positive (?ve) or neutral (n)
C correlative, E experimental
Caulerpa taxifolia in seagrass meadows
123

Hypotheses derived from this model were tested using
small-scale manipulative experiments (with limited
spatial replication given the official declaration of the
species as noxious), in conjunction with descriptions of
changes in abundance of C. taxifolia and seagrasses
over larger spatial and temporal scales.
Materials and methods
Study sites and invasion history
All experiments were done in two estuaries near
Sydney in SE Australia, namely the commercial port
of Botany Bay (34�000S, 151�110E) and Port Hacking
(34�040S, 151�080E), the latter being bounded by
residential properties to the north and national park to
the south. Two sites were used in both Port Hacking
(Gunnamatta Bay and Fishermans Bay, separated by
2.5 km) and Botany Bay (both in Quibray Bay,
separated by 800 m), each site being sheltered from
oceanic swell. The predominant native seagrasses
were the perennials Posidonia australis and Z. capri-
corni, and the annual Halophila ovalis, all of which
grew to a maximum depth of *5 m. The invasive alga
C. taxifolia was first verified in Port Hacking in 2000
and in Botany Bay in 2001 (Glasby et al. 2005),
although there is anecdotal evidence that it was present
in parts of Port Hacking in 1998 (Grey 2001). Estuary-
wide surveys indicated that C. taxifolia reached its
peak abundance in Botany Bay in 2003 and has
declined steadily ever since (unpublished data). In Port
Hacking, the area of C. taxifolia has fluctuated greatly
among years, but its abundance has been trending
downwards since 2004. In each site, C. taxifolia grew
amongst seagrass and was particularly dense immedi-
ately adjacent to seagrass beds (i.e. on soft sediment
that was previously non-vegetated). C. taxifolia sto-
lons intertwined to form a matrix *40 mm thick, with
fronds extending up a further 30–65 mm. In the field,
invasive strains of C. taxifolia are known only to
reproduce via asexual fragmentation (Wright 2005).
Temporal patterns of seagrasses and C. taxifolia
at the bay scale
To test the hypothesis that seagrass coverage has
declined since C. taxifolia was introduced, boundaries
of seagrass beds were identified from orthorectified
aerial photographs (at 1:1,500) for multiples times
before and after C. taxifolia was discovered in Quibray
Bay and Gunnamatta Bay (see Fig. 1). Field-based
ground truthing was used to validate and augment
maps for all years except 1970 and 1980 (hence species
could not be distinguished for these years). Ground
truthing was done using a real-time computer mapping
system, a GPS, an underwater video system and an
acoustic single beam depth sounder. Data on seagrass
areas for Gunnamatta Bay from 1977 to 1999 were
obtained from Meehan et al. (2005) who used the same
mapping methods. While the majority of seagrass beds
were monospecific, area estimates for P. australis
presented herein include mixtures of P. australis and
H. ovalis or Z. capricorni, whilst areas for Z. capricorni
include mixtures of Z. capricorni and H. ovalis.
C. taxifolia areal coverage was estimated in summer
and winter in 2003–2006 by towing divers behind a
boat on a manta board in a zigzag pattern around the
perimeter of each bay and recording bed boundaries.
This method overestimated the amount of C. taxifolia
as the alga rarely covered 100 % of the substratum and
was typically sparsely distributed within each bay.
Associations between C. taxifolia and seagrasses
To investigate spatial relationships between C. taxi-
folia and seagrasses, percentage covers of each were
estimated in 50 9 50 cm quadrats using a grid of 100
regularly-spaced points. Covers of primary foliage
(i.e. shoots) and canopy foliage (i.e. leaves) were
estimated. Observations suggested that the length of
C. taxifolia was greater amongst dense seagrass than
where there was little or no seagrass. This was tested
by measuring in situ the lengths of 15 haphazardly-
chosen C. taxifolia fronds (stolon to tip of frond) per
quadrat to the nearest mm using a ruler. Average
length of frond per quadrat was calculated for
comparisons with percentage cover of seagrass can-
opy. Quadrats were positioned haphazardly in areas
where C. taxifolia was present, at depths of 0.5–2 m
below mean low water. Sampling was done between
March and May 2003 in a total of 136 quadrats across
the four sites (*34 per site).
Pearson correlations were used to test for associ-
ations between C. taxifolia cover and seagrass cover.
One factor permutational ANOVA (PERMANOVA)
was used to compare C. taxifolia cover and frond
length among plots with no seagrass, sparse (1–49 %
T. M. Glasby
123

canopy cover) seagrass or dense (50–100 % canopy
cover) seagrass. Data were transformed to square roots
before calculating a Euclidian distance resemblance
matrix and using 9999 unrestricted permutations of
the raw data with Type III partial sums of squares.
Differences among treatments were investigated using
pair-wise t-tests.
Monitoring of fixed plots
To monitor vegetative expansion of C. taxifolia beds
and test for corresponding declines in seagrass,
permanent plots (50 9 50 cm) were established in
two sites in Quibray Bay in May 2003 and sampled 15
times over 8 years (until March 2011). Plots were
marked with stakes in two diagonally opposite
corners, which enabled a quadrat to be re-located in
the same place over each plot so percentage covers of
vegetation and bare space could be estimated using a
grid of 100 points. Plots were on the boundaries of
beds of C. taxifolia and beds of seagrass.
At each site, n = 3 replicate plots were posi-
tioned in each of the following habitats: primarily
P. australis, P. australis mixed with C. taxifolia, and
primarily C. taxifolia. Habitats were contiguous, with
plots in each habitat not more than 2 m apart. At one of
the sites and additional habitat, Z. capricorni mixed
with C. taxifolia, was also sampled. At the time, no
areas could be found in Quibray Bay that had
Z. capricorni but no C. taxifolia. Plots were sampled
approximately every 3–6 months in the first 3 years,
then less regularly (8–18 mo intervals) thereafter (see
Fig. 3). At each time of sampling, percentage covers
of seagrass or C. taxifolia canopies were estimated and
lengths of up to 15 haphazardly-chosen C. taxifolia
fronds (or all fronds if there were \15 in total) were
measured per quadrat.
Experimental transplantation of C. taxifolia
Two separate transplant experiments were done, one in
Quibray Bay, the other in Gunnamatta Bay, to test the
hypothesis that C. taxifolia impacts beds of sparse
seagrass (P. australis or Z. capricorni) by reducing
their canopy covers (i.e. numbers and/or lengths of
leaves), but not dense seagrass beds. C. taxifolia was
sourced from a 5 9 5 m area (*50–80 % cover of
C. taxifolia), 0.8 m deep some 15 m from the exper-
imental plots. Intact C. taxifolia (with attached
rhizomes, stolons and fronds) was removed by hand
and kept in seawater on the boat for a maximum of
30 min. Stolons were cut into 15 cm fragments (with
8–14 fronds) and four were planted in each experi-
mental plot (described below) by gently pushing
rhizoids into the sediment and using two U-shaped
plastic coated wire ‘‘stakes’’ to pin the stolon to the
sediment (as per Ceccherelli and Cinelli 1999). These
fundamentals of each experiment were the same, but as
described below there were slight differences in
treatments.
Transplant experiment 1 was set up in Quibray Bay
in water 0.5–1 m below mean low tide and ran from
December 2003 to January 2008. Eleven treat-
ments were established, with n = 3 replicate plots
(50 9 50 cm) per treatment, each separated by
10–50 m. Four treatments involved transplanting
four fragments of C. taxifolia (to create a C. taxifolia
coverage of *10 % in plots) into sparse seagrass
(P. australis and Z. capricorni; canopy cover
20–30 %) and dense seagrass (P. australis and Z. cap-
ricorni; canopy cover 80–90 %). To test for the effects
of C. taxifolia in relation to the seagrass canopy per se,
C. taxifolia was also transplanted into plots of
P. australis or Z. capricorni that had had their
canopies cut to change them from dense to sparse.
This involved removing leaves (cutting with scissors)
at the tops of some of the sheaths such that canopy
cover in the plot was reduced from 80 to 90 % down to
20–30 %. These canopy cutting treatments helped
distinguish effects of canopy cover per se from other
effects (e.g. sediment properties) that might be asso-
ciated with dense seagrass. There were also two
seagrass control treatments (one P. australis, one
Z. capricorni each 50 % canopy cover) that had no
C. taxifolia added. The final three treatments were
controls to test for artefacts associated with handling
the C. taxifolia; namely undisturbed C. taxifolia plots
(*10 % cover), C. taxifolia that had been cut into
15 cm sections in situ (without removing algae from
the sediment), and C. taxifolia fragments that had been
moved to non-vegetated plots (to test for the effects of
moving C. taxifolia to a ‘new’ place, as was being
done when it was transplanted to seagrass). Plots
without C. taxifolia were ‘weeded’ whenever they
were sampled and a 30 cm C. taxifolia exclusion
buffer was maintained around those plots. C. taxifolia
was nevertheless able to invade (e.g. via drifting
fragments) most plots at some stage of the experiment.
Caulerpa taxifolia in seagrass meadows
123

Transplant experiment 2 was done to test whether
results were consistent at a second site and included
more controls to test for natural changes in sparse
versus dense seagrass. This second experiment was set
up in Gunnamatta Bay (November 2004–September
2009) in water 0.3–0.8 m below mean low tide and did
not involve P. australis treatments. Seven treatments
were established, with n = 3 replicate plots
(50 9 50 cm) per treatment, each separated by
10–50 m. C. taxifolia was transplanted into naturally
sparse Z. capricorni (20–30 % canopy cover) and
dense Z. capricorni (80–90 % canopy cover). C. taxi-
folia was also transplanted into plots of Z. capricorni
that had had their canopies cut to change them from
dense to sparse. Z. capricorni control treatments were:
naturally sparse Z. capricorni with no C. taxifolia
added, dense Z. capricorni with no C. taxifolia added,
and cut sparse Z. capricorni with no C. taxifolia added.
The latter treatment enabled a test of the effects of
cutting the Z. capricorni canopy on the growth of
the seagrass. The final treatment consisted of undis-
turbed C. taxifolia plots (with no seagrass). Proce-
dural controls to test for effects of manipulating the
C. taxifolia were not included as results from exper-
iment 1 indicated no such experimental artefacts (see
results). All other procedures were the same as the first
experiment.
Repeated measures analyses were used to compare
percentage covers of seagrasses and C. taxifolia
among treatments over time. Analyses were done
using PERMANOVA with Euclidean distance resem-
blance matrices, as described above.
Results
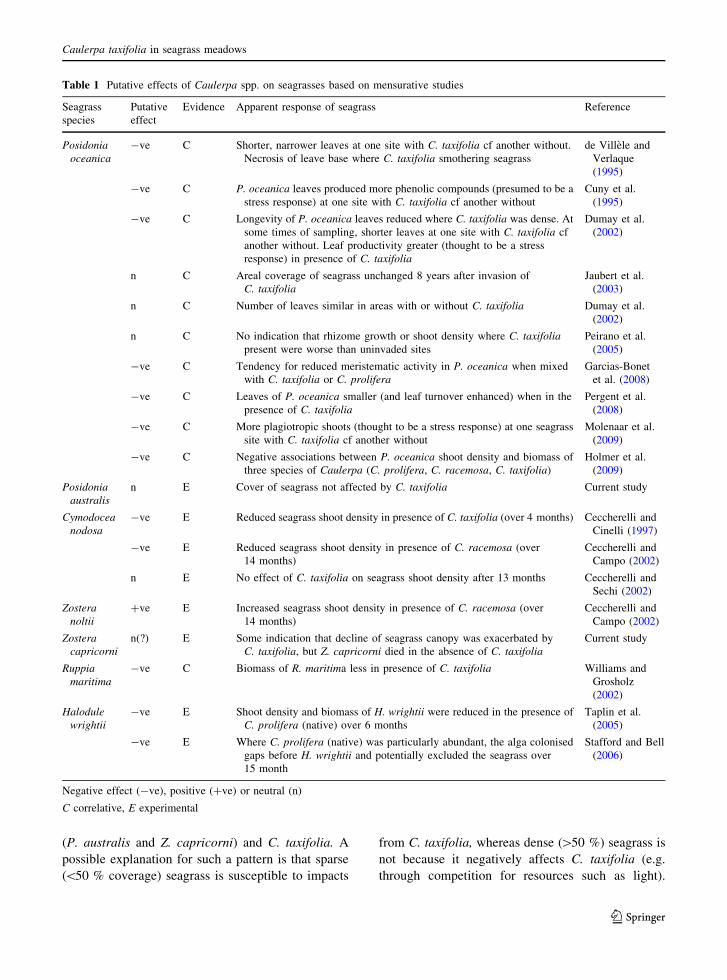
Temporal patterns of seagrasses and C. taxifolia
at the bay scale
Fluctuations in the total area of seagrass in Quibray
Bay since 1970 were driven by changes in
Z. capricorni (Fig. 1a). Such infrequent mapping of
Z. capricorni may not accurately depict changes in its
abundance, but nevertheless, the emergence and
spread of C. taxifolia occurred sometime during the
period that Z. capricorni appears to have declined.
Notably, the area of Z. capricorni in 2008 was
essentially the same as that estimated some 20 years
before the introduction of C. taxifolia. When the
coverage of C. taxifolia was maximal (25.9 ha) in
2005, it covered 10 % of Quibray Bay compared to
seagrass which covered *30 % of the bay over the
four mapping times.
In Gunnamatta Bay the total area of seagrass was
almost totally driven by P. australis (hence it was not
distinguished from total seagrass in Fig. 1b), with the
area of Z. capricorni being trivial in comparison
(Fig. 1b). The areas of P. australis and Z. capricorni
remained relatively stable during the period that
C. taxifolia appeared and became abundant. At its
peak, C. taxifolia was found in 20.5 % (25 ha) of
Gunnamatta Bay which was substantially more than
the average seagrass coverage (12 ha, 10 % of bay) or
the maximal seagrass coverage (18 ha, 15 % of bay).
This latter result was due to C. taxifolia colonis-
ing large areas of previously non-vegetated soft
sediments.
It is noteworthy that the Z. capricorni beds at both
the Gunnamatta and Quibray Bay sites used in this
study appear to have disappeared in the past. The small
patch of Z. capricorni sampled in the current study in
Gunnamatta Bay was not mapped by West et al.
(1985), while maps for Quibray Bay indicate that the
beds of Z. capricorni used in Quibray Bay were
present in 1942 and 1985 (Larkum and West 1990),
but not in the early 1970s (Larkum 1976).
Associations between C. taxifolia and seagrasses
In areas where C. taxifolia and seagrass co-occurred,
there was a negative correlation between the percent-
age cover of C. taxifolia and canopy covers of
P. australis (r = -0.42, 47 df, P \ 0.01) and
Z. capricorni (r = -0.41, 48 df, P \ 0.01). Similar
negative correlations existed between C. taxifolia and
primary (shoot) covers of P. australis (r = -0.40, 47
df, P \ 0.01) and Z. capricorni (r = -0.56, 48 df,
P \ 0.01). Percentage cover of C. taxifolia differed
significantly among habitat categories (pseudo
F4,131 = 15.98, P = 0.0001), being greatest where
there was no seagrass or a sparse canopy of seagrass
(Fig. 2a). C. taxifolia coverage was restricted to
\40 % where seagrass canopy was dense ([50 %;
Fig. 2a).
The average maximum length of C. taxifolia fronds
differed significantly among habitats (pseudo
F4,106 = 13.29, P = 0.0001), with longer fronds
occurring in plots with a dense P. australis canopy
T. M. Glasby
123
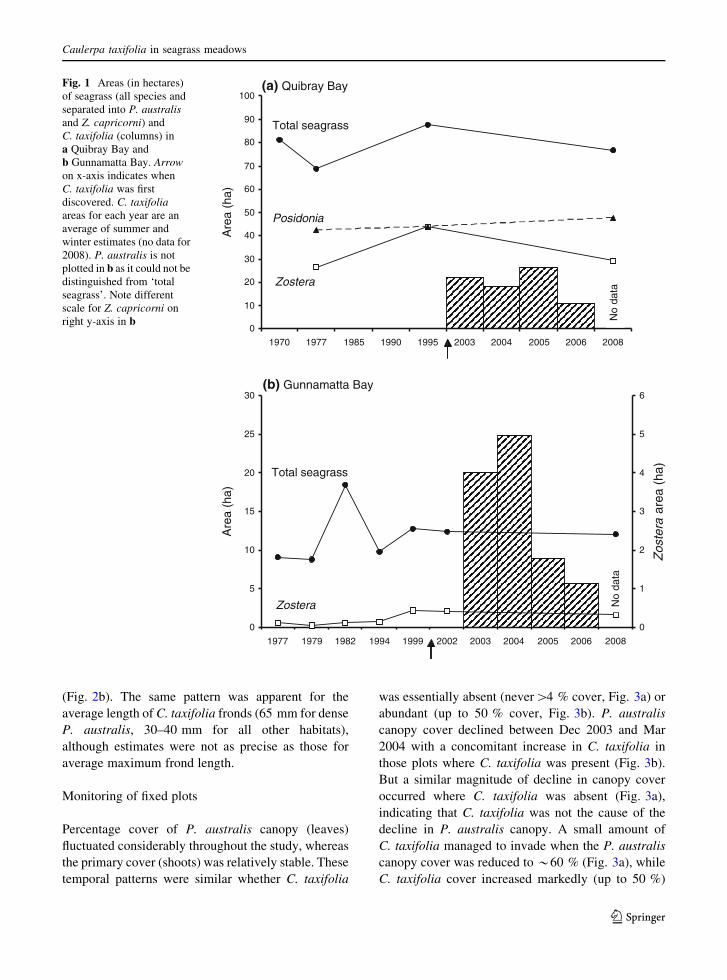
(Fig. 2b). The same pattern was apparent for the
average length of C. taxifolia fronds (65 mm for dense
P. australis, 30–40 mm for all other habitats),
although estimates were not as precise as those for
average maximum frond length.
Monitoring of fixed plots
Percentage cover of P. australis canopy (leaves)
fluctuated considerably throughout the study, whereas
the primary cover (shoots) was relatively stable. These
temporal patterns were similar whether C. taxifolia
was essentially absent (never[4 % cover, Fig. 3a) or
abundant (up to 50 % cover, Fig. 3b). P. australis
canopy cover declined between Dec 2003 and Mar
2004 with a concomitant increase in C. taxifolia in
those plots where C. taxifolia was present (Fig. 3b).
But a similar magnitude of decline in canopy cover
occurred where C. taxifolia was absent (Fig. 3a),
indicating that C. taxifolia was not the cause of the
decline in P. australis canopy. A small amount of
C. taxifolia managed to invade when the P. australis
canopy cover was reduced to *60 % (Fig. 3a), while
C. taxifolia cover increased markedly (up to 50 %)
0
10
20
30
40
50
60
70
80
90
100
1970 1977 1985 1990 1995 2003 2004 2005 2006 2008
Total seagrass
(a) Quibray Bay
No
data
Posidonia
Zostera
Are
a (h
a)
0
5
10
15
20
25
30
1977 1979 1982 1994 1999 2002 2003 2004 2005 2006 2008
0
1
2
3
4
5
6
Are
a (h
a)
(b) Gunnamatta Bay
No
data
Total seagrass
Zos
tera
are
a (h
a)
Zostera
Fig. 1 Areas (in hectares)
of seagrass (all species and
separated into P. australisand Z. capricorni) and
C. taxifolia (columns) in
a Quibray Bay and
b Gunnamatta Bay. Arrowon x-axis indicates when
C. taxifolia was first
discovered. C. taxifoliaareas for each year are an
average of summer and
winter estimates (no data for
2008). P. australis is not
plotted in b as it could not be
distinguished from ‘total
seagrass’. Note different
scale for Z. capricorni on
right y-axis in b
Caulerpa taxifolia in seagrass meadows
123
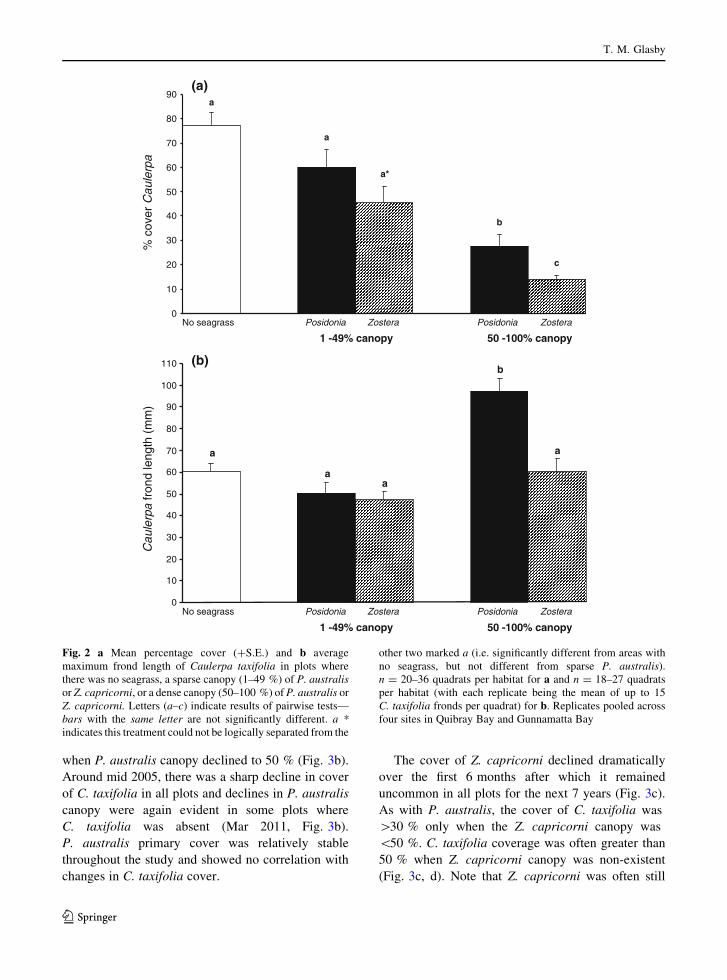
when P. australis canopy declined to 50 % (Fig. 3b).
Around mid 2005, there was a sharp decline in cover
of C. taxifolia in all plots and declines in P. australis
canopy were again evident in some plots where
C. taxifolia was absent (Mar 2011, Fig. 3b).
P. australis primary cover was relatively stable
throughout the study and showed no correlation with
changes in C. taxifolia cover.
The cover of Z. capricorni declined dramatically
over the first 6 months after which it remained
uncommon in all plots for the next 7 years (Fig. 3c).
As with P. australis, the cover of C. taxifolia was
[30 % only when the Z. capricorni canopy was
\50 %. C. taxifolia coverage was often greater than
50 % when Z. capricorni canopy was non-existent
(Fig. 3c, d). Note that Z. capricorni was often still
(b)
0
10
20
30
40
50
60
70
80
90
100
110
Cau
lerp
a fr
ond
leng
th (
mm
)
No seagrass Posidonia Zostera Posidonia Zostera
a
a
a
a
b
1 -49% canopy 50 -100% canopy
(a)
0
10
20
30
40
50
60
70
80
90
b
a
c
a
a*
% c
over
Cau
lerp
a
No seagrass Posidonia Zostera Posidonia Zostera
1 -49% canopy 50 -100% canopy
Fig. 2 a Mean percentage cover (?S.E.) and b average
maximum frond length of Caulerpa taxifolia in plots where
there was no seagrass, a sparse canopy (1–49 %) of P. australisor Z. capricorni, or a dense canopy (50–100 %) of P. australis or
Z. capricorni. Letters (a–c) indicate results of pairwise tests—
bars with the same letter are not significantly different. a *indicates this treatment could not be logically separated from the
other two marked a (i.e. significantly different from areas with
no seagrass, but not different from sparse P. australis).
n = 20–36 quadrats per habitat for a and n = 18–27 quadrats
per habitat (with each replicate being the mean of up to 15
C. taxifolia fronds per quadrat) for b. Replicates pooled across
four sites in Quibray Bay and Gunnamatta Bay
T. M. Glasby
123
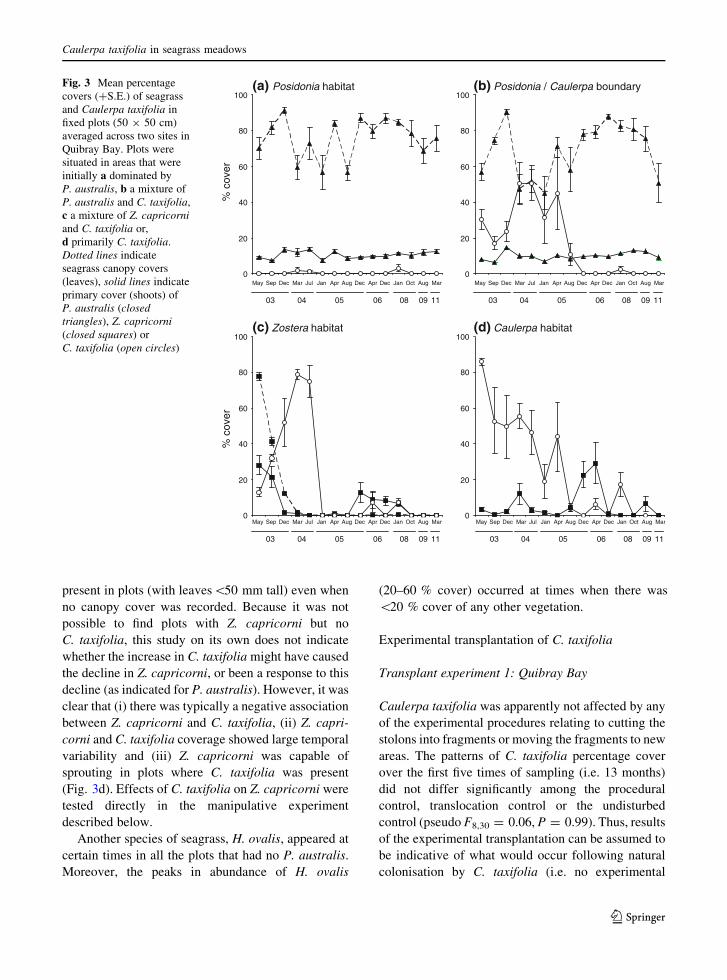
present in plots (with leaves\50 mm tall) even when
no canopy cover was recorded. Because it was not
possible to find plots with Z. capricorni but no
C. taxifolia, this study on its own does not indicate
whether the increase in C. taxifolia might have caused
the decline in Z. capricorni, or been a response to this
decline (as indicated for P. australis). However, it was
clear that (i) there was typically a negative association
between Z. capricorni and C. taxifolia, (ii) Z. capri-
corni and C. taxifolia coverage showed large temporal
variability and (iii) Z. capricorni was capable of
sprouting in plots where C. taxifolia was present
(Fig. 3d). Effects of C. taxifolia on Z. capricorni were
tested directly in the manipulative experiment
described below.
Another species of seagrass, H. ovalis, appeared at
certain times in all the plots that had no P. australis.
Moreover, the peaks in abundance of H. ovalis
(20–60 % cover) occurred at times when there was
\20 % cover of any other vegetation.
Experimental transplantation of C. taxifolia
Transplant experiment 1: Quibray Bay
Caulerpa taxifolia was apparently not affected by any
of the experimental procedures relating to cutting the
stolons into fragments or moving the fragments to new
areas. The patterns of C. taxifolia percentage cover
over the first five times of sampling (i.e. 13 months)
did not differ significantly among the procedural
control, translocation control or the undisturbed
control (pseudo F8,30 = 0.06, P = 0.99). Thus, results
of the experimental transplantation can be assumed to
be indicative of what would occur following natural
colonisation by C. taxifolia (i.e. no experimental
0
20
40
60
80
100
May
03
Sep Dec Mar Jul Jan Apr
04 05
Aug Dec Apr
06
Dec Jan Oct
08
Aug Mar
09 11
(b) Posidonia / Caulerpa boundary
0
20
40
60
80
100
May
03
Sep Dec Mar Jul Jan Apr
04 05
Aug Dec Apr
06
Dec Jan Oct
08
Aug Mar
09 11
% c
over
(a) Posidonia habitat
0
20
40
60
80
100
May
03
Sep Dec Mar Jul Jan Apr
04 05
Aug Dec Apr
06
Dec Jan Oct
08
Aug Mar
09 11
(d) Caulerpa habitat
0
20
40
60
80
100
May
03
Sep Dec Mar Jul Jan Apr
04 05
Aug Dec Apr
06
Dec Jan Oct
08
Aug Mar
09 11
% c
over
(c) Zostera habitat
Fig. 3 Mean percentage
covers (?S.E.) of seagrass
and Caulerpa taxifolia in
fixed plots (50 9 50 cm)
averaged across two sites in
Quibray Bay. Plots were
situated in areas that were
initially a dominated by
P. australis, b a mixture of
P. australis and C. taxifolia,
c a mixture of Z. capricorniand C. taxifolia or,
d primarily C. taxifolia.
Dotted lines indicate
seagrass canopy covers
(leaves), solid lines indicate
primary cover (shoots) of
P. australis (closedtriangles), Z. capricorni(closed squares) or
C. taxifolia (open circles)
Caulerpa taxifolia in seagrass meadows
123
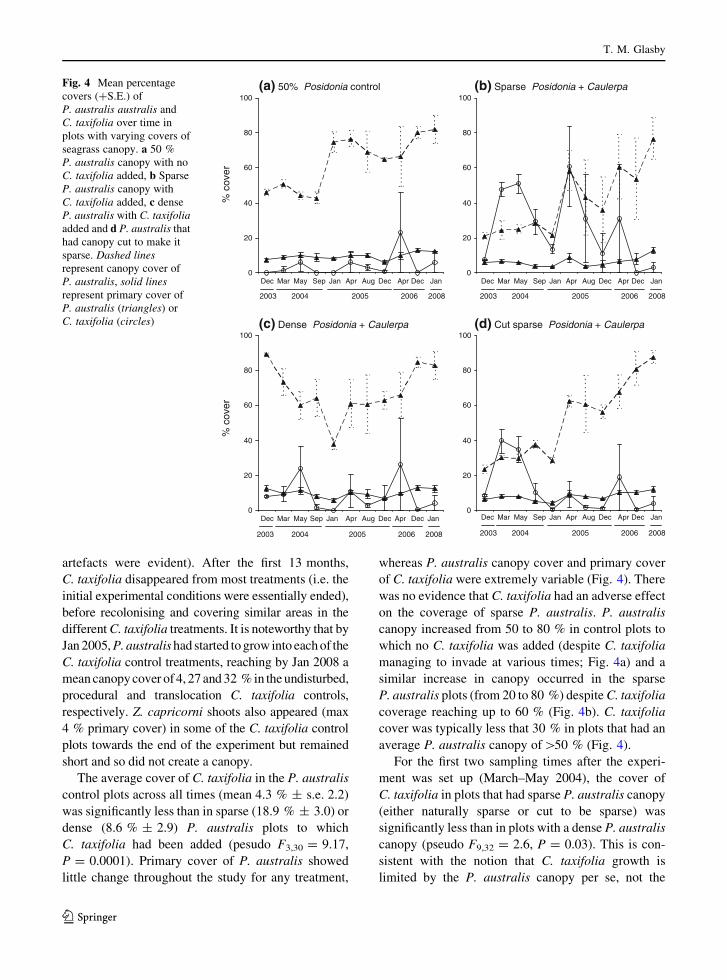
artefacts were evident). After the first 13 months,
C. taxifolia disappeared from most treatments (i.e. the
initial experimental conditions were essentially ended),
before recolonising and covering similar areas in the
different C. taxifolia treatments. It is noteworthy that by
Jan 2005, P. australis had started to grow into each of the
C. taxifolia control treatments, reaching by Jan 2008 a
mean canopy cover of 4, 27 and 32 % in the undisturbed,
procedural and translocation C. taxifolia controls,
respectively. Z. capricorni shoots also appeared (max
4 % primary cover) in some of the C. taxifolia control
plots towards the end of the experiment but remained
short and so did not create a canopy.
The average cover of C. taxifolia in the P. australis
control plots across all times (mean 4.3 % ± s.e. 2.2)
was significantly less than in sparse (18.9 % ± 3.0) or
dense (8.6 % ± 2.9) P. australis plots to which
C. taxifolia had been added (pesudo F3,30 = 9.17,
P = 0.0001). Primary cover of P. australis showed
little change throughout the study for any treatment,
whereas P. australis canopy cover and primary cover
of C. taxifolia were extremely variable (Fig. 4). There
was no evidence that C. taxifolia had an adverse effect
on the coverage of sparse P. australis. P. australis
canopy increased from 50 to 80 % in control plots to
which no C. taxifolia was added (despite C. taxifolia
managing to invade at various times; Fig. 4a) and a
similar increase in canopy occurred in the sparse
P. australis plots (from 20 to 80 %) despite C. taxifolia
coverage reaching up to 60 % (Fig. 4b). C. taxifolia
cover was typically less that 30 % in plots that had an
average P. australis canopy of[50 % (Fig. 4).
For the first two sampling times after the experi-
ment was set up (March–May 2004), the cover of
C. taxifolia in plots that had sparse P. australis canopy
(either naturally sparse or cut to be sparse) was
significantly less than in plots with a dense P. australis
canopy (pseudo F9,32 = 2.6, P = 0.03). This is con-
sistent with the notion that C. taxifolia growth is
limited by the P. australis canopy per se, not the
0
20
40
60
80
100
Dec
2003
Mar May Sep Jan Apr Aug
2004 2005
Dec Apr Dec
2006
Jan
2008
% c
over
(a) 50% Posidonia control
0
20
40
60
80
100
Dec
2003
Mar May Sep Jan Apr Aug
2004 2005
Dec Apr Dec
2006
Jan
2008
(b) Sparse Posidonia + Caulerpa
0
20
40
60
80
100
Dec
2003
Mar May Sep Jan Apr Aug
2004 2005
Dec Apr Dec
2006
Jan
2008
% c
over
(c) Dense Posidonia + Caulerpa
0
20
40
60
80
100
Dec
2003
Mar May Sep Jan Apr Aug
2004 2005
Dec Apr Dec
2006
Jan
2008
(d) Cut sparse Posidonia + Caulerpa
Fig. 4 Mean percentage
covers (?S.E.) of
P. australis australis and
C. taxifolia over time in
plots with varying covers of
seagrass canopy. a 50 %
P. australis canopy with no
C. taxifolia added, b Sparse
P. australis canopy with
C. taxifolia added, c dense
P. australis with C. taxifoliaadded and d P. australis that
had canopy cut to make it
sparse. Dashed linesrepresent canopy cover of
P. australis, solid linesrepresent primary cover of
P. australis (triangles) or
C. taxifolia (circles)
T. M. Glasby
123
primary (shoot) cover or other features of areas where
dense P. australis happens to grow. After this time,
covers of C. taxifolia declined in most treatments and
typically remained small (but variable) in plots that
had a dense canopy cover ([50 %) of P. australis
(Fig. 4).
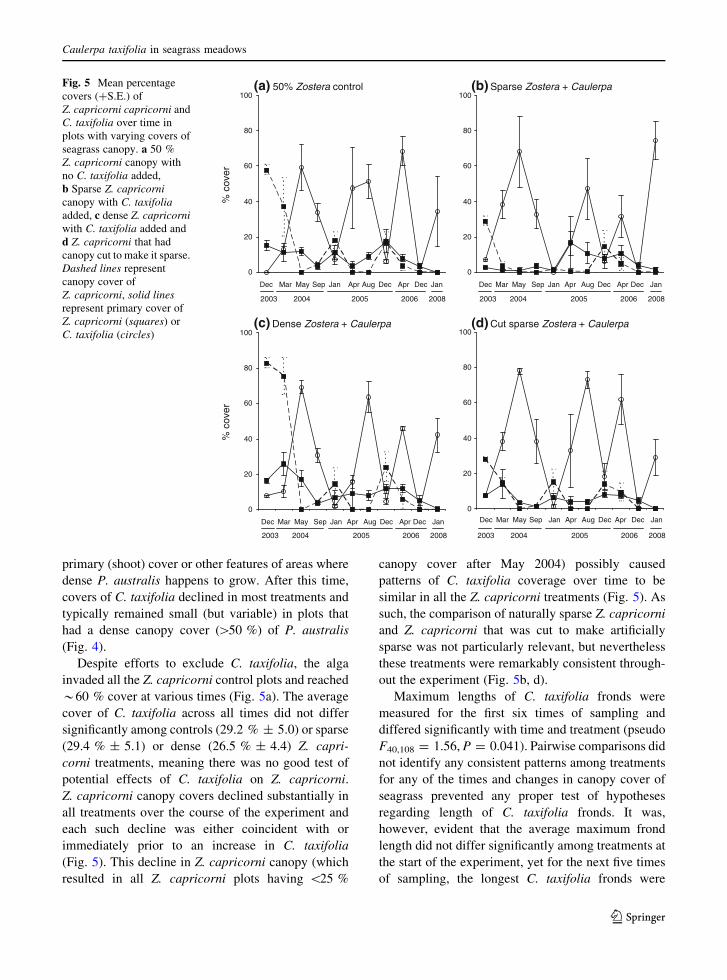
Despite efforts to exclude C. taxifolia, the alga
invaded all the Z. capricorni control plots and reached
*60 % cover at various times (Fig. 5a). The average
cover of C. taxifolia across all times did not differ
significantly among controls (29.2 % ± 5.0) or sparse
(29.4 % ± 5.1) or dense (26.5 % ± 4.4) Z. capri-
corni treatments, meaning there was no good test of
potential effects of C. taxifolia on Z. capricorni.
Z. capricorni canopy covers declined substantially in
all treatments over the course of the experiment and
each such decline was either coincident with or
immediately prior to an increase in C. taxifolia
(Fig. 5). This decline in Z. capricorni canopy (which
resulted in all Z. capricorni plots having \25 %
canopy cover after May 2004) possibly caused
patterns of C. taxifolia coverage over time to be
similar in all the Z. capricorni treatments (Fig. 5). As
such, the comparison of naturally sparse Z. capricorni
and Z. capricorni that was cut to make artificially
sparse was not particularly relevant, but nevertheless
these treatments were remarkably consistent through-
out the experiment (Fig. 5b, d).
Maximum lengths of C. taxifolia fronds were
measured for the first six times of sampling and
differed significantly with time and treatment (pseudo
F40,108 = 1.56, P = 0.041). Pairwise comparisons did
not identify any consistent patterns among treatments
for any of the times and changes in canopy cover of
seagrass prevented any proper test of hypotheses
regarding length of C. taxifolia fronds. It was,
however, evident that the average maximum frond
length did not differ significantly among treatments at
the start of the experiment, yet for the next five times
of sampling, the longest C. taxifolia fronds were
0
20
40
60
80
100
0
20
40
60
80
100
0
20
40
60
80
100
Dec
2003
Mar May Sep Jan Apr Aug
2004 2005
Dec Apr Dec
2006
Jan
2008
% c
over
(a) 50% Zostera control
Dec
2003
Mar May Sep Jan Apr Aug
2004 2005
Dec Apr Dec
2006
Jan
2008
% c
over
(c) Dense Zostera + Caulerpa
Dec
2003
Mar May Sep Jan Apr Aug
2004 2005
Dec Apr Dec
2006
Jan
2008
(b) Sparse Zostera + Caulerpa
0
20
40
60
80
100
Dec
2003
Mar May Sep Jan Apr Aug
2004 2005
Dec Apr Dec
2006
Jan
2008
(d) Cut sparse Zostera + Caulerpa
Fig. 5 Mean percentage
covers (?S.E.) of
Z. capricorni capricorni and
C. taxifolia over time in
plots with varying covers of
seagrass canopy. a 50 %
Z. capricorni canopy with
no C. taxifolia added,
b Sparse Z. capricornicanopy with C. taxifoliaadded, c dense Z. capricorniwith C. taxifolia added and
d Z. capricorni that had
canopy cut to make it sparse.
Dashed lines represent
canopy cover of
Z. capricorni, solid linesrepresent primary cover of
Z. capricorni (squares) or
C. taxifolia (circles)
Caulerpa taxifolia in seagrass meadows
123
always in the P. australis treatments (both sparse and
dense). Relatively few C. taxifolia fronds were
available to be measured in this experiment compared
to the initial descriptive study meaning precision was
greatly compromised.
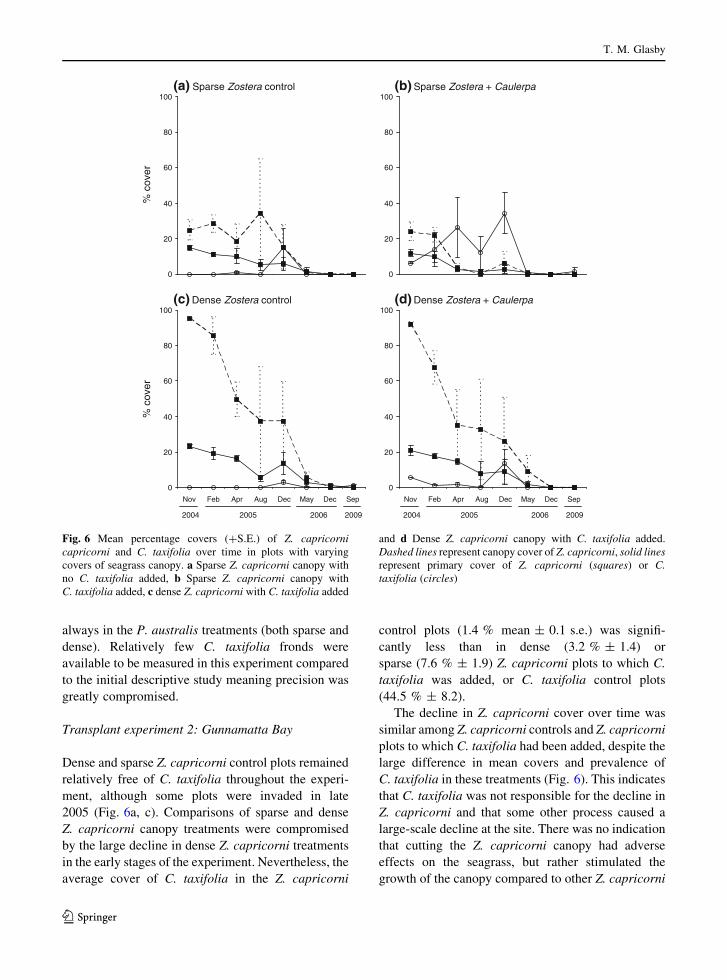
Transplant experiment 2: Gunnamatta Bay
Dense and sparse Z. capricorni control plots remained
relatively free of C. taxifolia throughout the experi-
ment, although some plots were invaded in late
2005 (Fig. 6a, c). Comparisons of sparse and dense
Z. capricorni canopy treatments were compromised
by the large decline in dense Z. capricorni treatments
in the early stages of the experiment. Nevertheless, the
average cover of C. taxifolia in the Z. capricorni
control plots (1.4 % mean ± 0.1 s.e.) was signifi-
cantly less than in dense (3.2 % ± 1.4) or
sparse (7.6 % ± 1.9) Z. capricorni plots to which C.
taxifolia was added, or C. taxifolia control plots
(44.5 % ± 8.2).
The decline in Z. capricorni cover over time was
similar among Z. capricorni controls and Z. capricorni
plots to which C. taxifolia had been added, despite the
large difference in mean covers and prevalence of
C. taxifolia in these treatments (Fig. 6). This indicates
that C. taxifolia was not responsible for the decline in
Z. capricorni and that some other process caused a
large-scale decline at the site. There was no indication
that cutting the Z. capricorni canopy had adverse
effects on the seagrass, but rather stimulated the
growth of the canopy compared to other Z. capricorni
0
20
40
60
80
100
Nov
2004
Feb Apr Aug Dec May Dec
2005 2006 2009
Sep
% c
over
(c) Dense Zostera control
0
20
40
60
80
100
Nov
2004
Feb Apr Aug Dec May Dec
2005 2006 2009
Sep
(d) Dense Zostera + Caulerpa
0
20
40
60
80
100(b) Sparse Zostera + Caulerpa
0
20
40
60
80
100(a) Sparse Zostera control
% c
over
Fig. 6 Mean percentage covers (?S.E.) of Z. capricornicapricorni and C. taxifolia over time in plots with varying
covers of seagrass canopy. a Sparse Z. capricorni canopy with
no C. taxifolia added, b Sparse Z. capricorni canopy with
C. taxifolia added, c dense Z. capricorni with C. taxifolia added
and d Dense Z. capricorni canopy with C. taxifolia added.
Dashed lines represent canopy cover of Z. capricorni, solid linesrepresent primary cover of Z. capricorni (squares) or C.taxifolia (circles)
T. M. Glasby
123
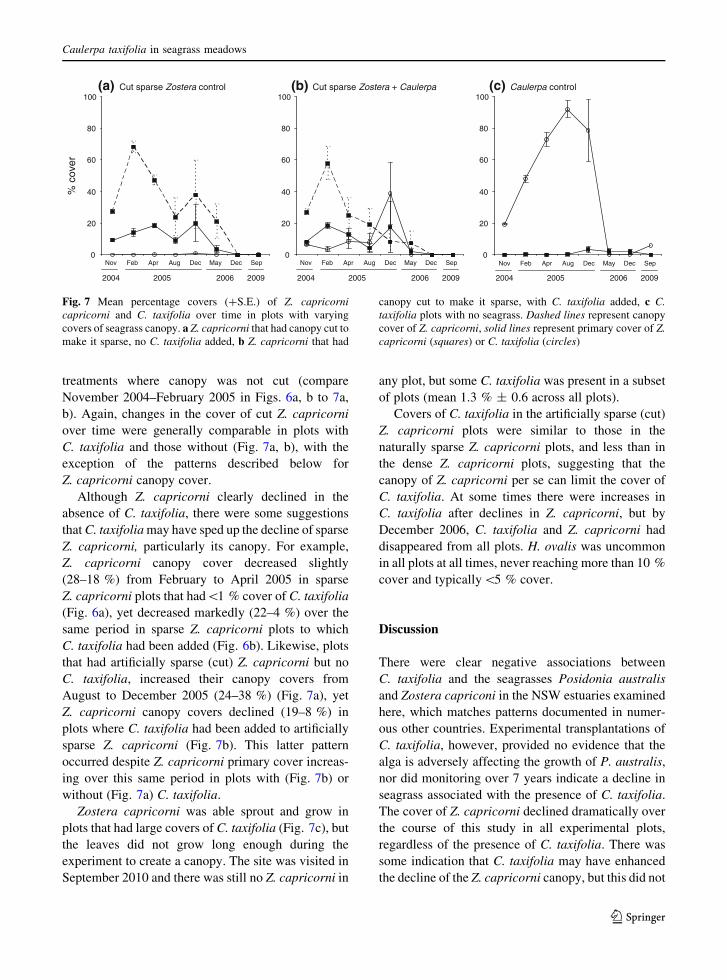
treatments where canopy was not cut (compare
November 2004–February 2005 in Figs. 6a, b to 7a,
b). Again, changes in the cover of cut Z. capricorni
over time were generally comparable in plots with
C. taxifolia and those without (Fig. 7a, b), with the
exception of the patterns described below for
Z. capricorni canopy cover.
Although Z. capricorni clearly declined in the
absence of C. taxifolia, there were some suggestions
that C. taxifolia may have sped up the decline of sparse
Z. capricorni, particularly its canopy. For example,
Z. capricorni canopy cover decreased slightly
(28–18 %) from February to April 2005 in sparse
Z. capricorni plots that had\1 % cover of C. taxifolia
(Fig. 6a), yet decreased markedly (22–4 %) over the
same period in sparse Z. capricorni plots to which
C. taxifolia had been added (Fig. 6b). Likewise, plots
that had artificially sparse (cut) Z. capricorni but no
C. taxifolia, increased their canopy covers from
August to December 2005 (24–38 %) (Fig. 7a), yet
Z. capricorni canopy covers declined (19–8 %) in
plots where C. taxifolia had been added to artificially
sparse Z. capricorni (Fig. 7b). This latter pattern
occurred despite Z. capricorni primary cover increas-
ing over this same period in plots with (Fig. 7b) or
without (Fig. 7a) C. taxifolia.
Zostera capricorni was able sprout and grow in
plots that had large covers of C. taxifolia (Fig. 7c), but
the leaves did not grow long enough during the
experiment to create a canopy. The site was visited in
September 2010 and there was still no Z. capricorni in
any plot, but some C. taxifolia was present in a subset
of plots (mean 1.3 % ± 0.6 across all plots).
Covers of C. taxifolia in the artificially sparse (cut)
Z. capricorni plots were similar to those in the
naturally sparse Z. capricorni plots, and less than in
the dense Z. capricorni plots, suggesting that the
canopy of Z. capricorni per se can limit the cover of
C. taxifolia. At some times there were increases in
C. taxifolia after declines in Z. capricorni, but by
December 2006, C. taxifolia and Z. capricorni had
disappeared from all plots. H. ovalis was uncommon
in all plots at all times, never reaching more than 10 %
cover and typically\5 % cover.
Discussion
There were clear negative associations between
C. taxifolia and the seagrasses Posidonia australis
and Zostera capriconi in the NSW estuaries examined
here, which matches patterns documented in numer-
ous other countries. Experimental transplantations of
C. taxifolia, however, provided no evidence that the
alga is adversely affecting the growth of P. australis,
nor did monitoring over 7 years indicate a decline in
seagrass associated with the presence of C. taxifolia.
The cover of Z. capricorni declined dramatically over
the course of this study in all experimental plots,
regardless of the presence of C. taxifolia. There was
some indication that C. taxifolia may have enhanced
the decline of the Z. capricorni canopy, but this did not
0
20
40
60
80
100
0
20
40
60
80
100
0
20
40
60
80
100
Nov
2004
Feb Apr Aug Dec May Dec
2005 2006 2009
SepNov
2004
Feb Apr Aug Dec May Dec
2005 2006 2009
SepNov
2004
Feb Apr Aug Dec May Dec
2005 2006 2009
Sep
(a) Cut sparse Zostera control
% c
over
(b) Cut sparse Zostera + Caulerpa (c) Caulerpa control
Fig. 7 Mean percentage covers (?S.E.) of Z. capricornicapricorni and C. taxifolia over time in plots with varying
covers of seagrass canopy. a Z. capricorni that had canopy cut to
make it sparse, no C. taxifolia added, b Z. capricorni that had
canopy cut to make it sparse, with C. taxifolia added, c C.taxifolia plots with no seagrass. Dashed lines represent canopy
cover of Z. capricorni, solid lines represent primary cover of Z.capricorni (squares) or C. taxifolia (circles)
Caulerpa taxifolia in seagrass meadows
123

alter the final fate of Z. capricorni, which disappeared
from all plots. Importantly, both Z. capricorni and
P. australis were found to sprout in plots that had large
covers of C. taxifolia for over a year (although notably
the newly-sprouted Z. capriconi did not grow large
enough to form a canopy). At the bay-wide scale
(*1 km2), the areal coverages of Z. capricorni and
P. australis were not different from what they were
many years prior to the introduction of C. taxifolia.
C. taxifolia in NSW is most likely behaving as an
opportunistic weed which rapidly colonises previously
non-vegetated areas (where it reaches large densities
and affects infauna; Wright and Gribben 2008;
Gribben et al. 2009b; McKinnon et al. 2009; Byers
et al. 2010) and periodically colonises gaps in seagrass
beds with no apparent affects on seagrass coverage.
This study appears to be the first experimental
investigation (albeit with limited replication) of
effects of C. taxifolia on the growth of a species of
Posidonia. Moreover, by using experimental additions
of C. taxifolia, this study provides a direct test of the
effects of invasion by the alga on seagrasses. A review
of the published literature (Table 1) indicates that all
the evidence for negative impacts of C. taxifolia on
Posidonia spp. is correlative. In fact, the only
published experimental evidence for negative impacts
of C. taxifolia on seagrass density seems to be a
4 month study by Ceccherelli and Cinelli (1997), and
importantly the same authors reported no such effect
over the longer-term (Table 1). This is not to say that
C. taxifolia might not be having sub-lethal effects on
seagrasses, for example by altering sediment charac-
teristics which can affect seagrass growth in some way
(Calleja et al. 2007). But as yet, evidence for such
sediment effects by C. taxifolia comes only from
correlative studies (e.g. Holmer et al. 2009; Eyre et al.
2011). Experimental studies with other species of
Caulerpa have also provided limited evidence for
negative effects on seagrasses (Table 1). C. racemosa
caused decreased shoot density of Cymodocea nodosa
over 14 months, yet increased shoot density of the
smaller seagrass Zostera noltii (Ceccherelli and
Campo 2002). In its native habitat, C. prolifera was
implicated in declines in shoot densities of the
sympatric Halodule wrightii (Taplin et al. 2005) and
outcompeted the seagrass for space over 15 months
(Stafford and Bell 2006). Thus, the published litera-
ture provides little evidence that species of Caulerpa
are capable of driving changes in seagrass abundance.
There are indications that species of Caulerpa com-
pete with seagrasses, either via interference competi-
tion, (i.e. pre-empting space), or exploitative
competition (i.e. utilising limited resources), as has
been demonstrated for C. prolifera in its native
environment. More experimental investigations are
needed to determine whether competition between
invasive Caulerpa and seagrasses could be any more
severe than competition among sympatric native
species (e.g. Williams 1987).
The thickness of the C. taxifolia beds in the present
study was around 40 mm of stolons, plus fronds. It is
possible that far thicker stolon mats, as have occurred
at some Mediterranean sites (Thibaut et al. 2004),
could have negative effects on seagrasses by creating
anoxic conditions (Holmer et al. 2009; Eyre et al.
2011). Much thicker mats of C. taxifolia (e.g.
200–300 mm high mats of stolons) have been
observed in some NSW estuaries, but never amongst
seagrasses. In some Mediterranean sites where
C. taxifolia was so abundant that it smothered
seagrass, P. oceanica showed signs of necrosis at the
base of the shoots (de Villele and Verlaque 1995).
There is certainly evidence that smothering is an
important means by which Caulerpa spp. impact
macroalgae and invertebrates (Piazzi et al. 2005;
Gribben et al. 2009b; Cebrian et al. in press).
Additionally, ambient nutrient levels might influence
the outcome of competition between Caulerpa spp.
and seagrasses, given that the former can potentially
thrive in eutrophic conditions (Lapointe and Bedford
2010; Burke and Grime 1996; Gennaro and Piazzi
2011; but see Ceccherelli and Sechi 2002), while
many seagrasses can be adversely affected (Chisholm
et al. 1997; Cardoso et al. 2004). Propagule density
could also influence the outcome between C. taxifolia
and seagrasses. It is possible that if more C. taxifolia
had been added to seagrass plots in the present study
there could have been detectable negative effects on
the seagrasses. But this seems unlikely given that
C. taxifolia increased from 10 % of the substratum
(initial experimental coverage) to 50 % in sparse
P. australis plots in Quibray Bay, yet coverage of the
seagrass was not affected.
There was evidence from this study of a threshold of
seagrass canopy cover (*50 %), which tended to
restrict C. taxifolia cover to less than 40 % of the
substratum. This was likely a shading effect of the
native canopy; scouring of the sediment by the canopy
T. M. Glasby
123

was unlikely as the denser the seagrass canopy, the
more intertwined the leaves resulting in reduced
physical contact with the substratum. Experimental
reductions in the canopy of P. australis and Z. capri-
corni resulted in increased covers of C. taxifolia
relative to plots with naturally dense seagrass canopies.
Thus, the native canopy reduces survivorship of the
invader, as demonstrated for the invasive alga Sargas-
sum muticum (Britton-Simmons 2006) and some
terrestrial plants (Corbin and D’Antonio 2004). The
native P. australis canopy also appeared to cause the
length of C. taxifolia fronds to increase, as described
for the Mediterranean (de Villele and Verlaque 1995;
Ceccherelli and Cinelli 1998); probably an effect of
reduced light levels (i.e. etiolation). Thus, it could be
argued that dense seagrass beds could provide optimal
conditions and hence a refuge for C. taxifolia (Stafford
and Bell 2006), but for the reasons discussed below, the
results of this study do not support such a conclusion.
Various authors have reported that C. taxifolia and
C. racemosa can invade only sparse seagrass (Meinesz
et al. 1993, de Villele and Verlaque 1995, Ceccherelli
and Cinelli 1999; Ceccherelli et al. 2000) or degraded
seagrass beds (Chisholm et al. 1997; Occhipinti-
Ambrogi and Savini 2003; Ruitton et al. 2005; Bulleri
et al. 2011; Kiparissis et al. 2011). Indeed plant
invasions are often correlated with the physical
structure of the recipient community, for example
forests with an intact canopy can have relatively few
invaders (Mack et al. 2000), while non-vegetated
ground can be more susceptible to invasion than well
vegetated areas (Harrison and Bigley 1982; Crawley
1986; Burke and Grime 1996; but see Thomsen et al.
2006). Dense seagrass beds seem better able to resist
colonisation by C. taxifolia, suggesting that maintaining
healthy populations of seagrasses will help minimise
colonisation of these beds by C. taxifolia and other
invaders (Williams 2007). But importantly, C. taxifolia
can rapidly colonise non-vegetated sediments, so
invasion of estuaries by this alga need not be related
at all to the health of seagrass beds. Indeed the present
study found that previously non-vegetated sediments
seem a better habitat for C. taxifolia than are seagrass
beds—the alga typically persisted longer and reached
larger densities in the former habitat (see also Holmer
et al. 2009). In this regard, patterns of C. taxifolia
abundance were similar to those of the small native
seagrass Halophila ovalis. The major difference was
that C. taxifolia bloomed more frequently than did H.
ovalis and the invader reached greater percentage
covers. The patterns of colonisation and abundance of
C. taxifolia in NSW estuaries seem similar to those of
species of Caulerpa in their native habitats (Williams
et al. 1985; Kirkman and Kuo 1990; Short et al. 2006;
Burfeind and Udy 2009), i.e. colonising non-vegetated
soft sediments, co-occurring at low densities with
seagrasses and having stochastic population dynamics.
In some cases, the previously non-vegetated areas that
C. taxifolia invades may have been non-vegetated
historically (i.e. the empty niche hypothesis, e.g. Elton
1958; MacArthur 1970), while in others they may be
periodically non-vegetated (i.e. the fluctuating resource
hypothesis, e.g. Johnstone 1986; Davis et al. 2000) due
to natural fluctuations in seagrasses (Duarte et al. 2006)
or anthropogenic disturbances (Walker et al. 2006). But
importantly, both of the aforementioned invasion
hypotheses describe situations where the invader per-
sists and dominates, whereas C. taxifolia in NSW often
disappears from sites, or in some cases, entire estuaries
(Glasby unpublished data).
Experimental work has demonstrated that other
invasive macroalgae such as Undaria pinnatifida and
Caulerpa racemosa are opportunistic invaders of algal
beds on rocky reefs (Valentine and Johnson 2003,
2004; Bulleri et al. 2010). Both of these introduced
algae colonised gaps that were created amongst native
canopy-forming algae. But U. pinnatifida subsequently
died off after the native algal canopy re-grew (Valen-
tine and Johnson 2003), whilst C. racemosa was
capable of persisting and preventing the recovery of the
native assemblage (Bulleri et al. 2010). In the present
study, C. taxifolia amongst P. australis behaved
similarly to U. pinnatifida in that it could occupy
space when the native canopy cover was low, but was
then outcompeted as the native canopy increased (see
also Burfeind and Udy 2009). The most typical
outcome involving the smaller seagrass species (Z. cap-
ricorni), however, was that both it and C. taxifolia
eventually disappeared from experimental plots. As
yet, neither the native seagrass nor the invader has
recovered and come to dominate these plots. There are
many documented cases of competitively inferior
invasive habitat-formers responding favourably to
disturbances (such as removal of vegetation) and
preventing the recovery of a native (e.g. Hobbs and
Huenneke 1992; Corbin and D’Antonio 2004; Bando
2006; Williams 2007), but it would seem that other
factors are at play in these NSW sites.
Caulerpa taxifolia in seagrass meadows
123

This study combined small-scale observational and
manipulative experiments with larger-scale habitat
mapping done over many years in two estuaries.
Overall, results indicated that C. taxifolia at these
NSW sites is an opportunistic invader of seagrass beds
that is not having adverse effects on the coverage of
P. australis. Possible effects of C. taxifolia on the
growth of Z. capricorni leaves (canopy) need to be
investigated, but large declines in Z. capricorni
occurred in the absence of C. taxifolia. C. taxifolia in
NSW is primarily colonising previously non-vegetated
soft sediments at the deeper margins of seagrass beds. In
most cases it is unclear whether these areas supported
seagrasses in the past, but abundances of native
seagrasses in various NSW estuaries have declined
since the early 1900s (Larkum and West 1990; Walker
and McComb 1992), as they have worldwide (e.g. Short
and Wyllie-Echeverria 1996; Green and Short 2003;
Orth et al. 2006; Boudouresque et al. 2009). There are
ecological differences between C. taxifolia and seag-
rasses, but future studies should investigate whether, in
certain situations, C. taxifolia could perform an eco-
logical role that is roughly intermediate between non-
vegetated sediments and small native seagrass (see
Posey 1988). There is evidence that C. taxifolia can
function similarly to (albeit significantly differently
from) some seagrasses in terms of trapping sediments
(Hendriks et al. 2010), facilitating some chemical
processes in the sediment (Holmer et al. 2009; Eyre
et al. 2011) and as a habitat for fishes (York et al. 2006)
or epibenthic invertebrates (McKinnon et al. 2009;
Tanner 2011). However, C. taxifolia can have reduced
numbers of infaunal invertebrates relative to non-
vegetated sediments, possibly due to increased anoxia
in sediments (McKinnon et al. 2009).
The potential negative effects of invasive species
are important to focus on, but invasive habitat forming
species may have some beneficial roles in situations
where there have been large-scale losses of structur-
ally similar native habitat (Crooks 2002; Rodriguez
2006). This is worth considering when, as demon-
strated for the sites studied here, there is no evidence
that the invasive habitat former is driving the loss of
native vegetation nor does it appear to be preventing
the recovery of the native There are many direct
threats to seagrass beds throughout the world (Short
et al. 2011) and there is little experimental evidence
that C. taxifolia should rank highly on this list, despite
the alga’s capacity to impact a range of other species.
Acknowledgments This research was funded by the NSW
Department of Primary Industries. Thank you to Peter Gibson,
Roger Laird, Brett Louden, Alan Genders and Brett Rankin for
help with field work and data entry. Bob Creese, Paul Gribben,
Peter Gibson, Skye Taylor and Nick Otway provided useful
discussion and comments on the manuscript. Seagrass mapping
data were compiled by Greg West using data from numerous
departmental projects, the principal investigators for which were
Ron West and Robert Williams. I gratefully acknowledge the
contribution of the many staff involved in generating seagrass
maps for past projects, in particular Trudy Walford. Thank you
to Kathy Bown for help with obtaining references.
References
Bando K (2006) The roles of competition and disturbance in a
marine invasion. Biol Invasions 8:755–763
Bellan-Santini D, Arnaud PM, Bellan G, Verlaque M (1996)
The influence of the introduced tropical alga Caulerpataxifolia on the biodiversity of the Mediterranean marine
biota. J Mar Biol Assoc UK 76:235–237
Boudouresque CF, Meinesz A, Riberta MA, Ballesteros E
(1995) Spread of the green alga Caulerpa taxifolia (Caul-
erpales, Chlorophyta) in the Mediterranean: possible con-
sequences of a major ecological event. Scientia Marina
59:21–29
Boudouresque CF, Bernard G, Pergent G, Shili A, Verlaque M
(2009) Regression of Mediterranean seagrasses caused by
natural processes and anthropogenic disturbances and
stress: a critical review. Bot Mar 52:395–418
Britton-Simmons KH (2006) Functional group diversity,
resource preemption and the genesis of invasion resistance
in a community of marine algae. Oikos 113:395–401
Bruno JF, Fridley JD, Bromberg K, Bertness MD (2005)
Insights into biotic interactions from studies of species
invasions. In: Sax DF, Stachowicz JJ, Gaines SD (eds)
Species invasions: insights into ecology, evolution, and
biogeography. Sinauer Associates, Sunderland, MA,
pp 13–40
Bulleri F, Balata D, Bertocci I, Tamburello L, Benedetti-Cecchi
L (2010) The seaweed Caulerpa racemosa on Mediterra-
nean rocky reefs: from passenger to driver of ecological
change. Ecology 91:2205–2212
Bulleri F, Alestra T, Ceccherelli G, Tamburello L, Pinna S,
Sechi N, Benedetti-Cecchi L (2011) Determinants of
Caulerpa racemosa distribution in the north-western
Mediterranean. Mar Ecol Prog Ser 431:55–67
Burfeind DD, Udy JW (2009) The effects of light and nutrients
on Caulerpa taxifolia and growth. Aquat Bot 90:105–109
Burke MJW, Grime JP (1996) An experimental study of plant
community invasibility. Ecology 77:776–790
Byers JE, Wright JT, Gribben PE (2010) Variable direct and
indirect effects of a habitat-modifying invasive species on
mortality of native fauna. Ecology 91:1787–1798
Calleja ML, Marba N, Duarte CM (2007) The relationship
between seagrass (Posidonia oceanica) decline and sulfide
porewater concentration in carbonate sediments. Estuar
Coast Shelf Sci 73:583–588
Cardoso PG, Pardal MA, Lillebo AI, Ferreira SM, Raffaelli D,
Marques JC (2004) Dynamic changes in seagrass
T. M. Glasby
123

assemblages under eutrophication and implications for
recovery. J Exp Mar Biol Ecol 302:233–248
Cebrian E, Linares C, Marschal C, Garrabou J (in press) Exploring
the effects of invasive algae on the persistence of gorgonian
populations. Biol Invasions. doi:10.1007/s10530-012-0261-6
Ceccherelli G, Campo D (2002) Different effects of Caulerparacemosa on two co-occurring seagrasses in the Mediter-
ranean. Bot Mar 45:71–76
Ceccherelli G, Cinelli F (1997) Short-term effects of nutrient
enrichment of the sediment and interactions between the
seagrass Cymodocea nodosa and the introduced green alga
Caulerpa taxifolia in a Mediterranean bay. J Exp Mar Biol
Ecol 217:165–177
Ceccherelli G, Cinelli F (1998) Habitat effect on spatio-tem-
poral variability in size and density of the introduced alga
Caulerpa taxifolia. Mar Ecol Prog Ser 163:289–294
Ceccherelli G, Cinelli F (1999) Effects of Posidonia oceanicacanopy on Caulerpa taxifolia size in a north-western
Mediterranean bay. J Exp Mar Biol Ecol 240:19–36
Ceccherelli G, Sechi N (2002) Nutrient availability in the sed-
iment and the reciprocal effects between the native sea-
grass Cymodocea nodosa and the introduced rhizophytic
alga Caulerpa taxifolia. Hydrobiologia 474:57–66
Ceccherelli G, Piazzi L, Cinelli F (2000) Response of the non-
indigenous Caulerpa racemosa (Forsskal) J. Agardh to the
native seagrass Posidonia oceanica (L.) Delile: effect of
density of shoots and orientation of edges of meadows.
J Exp Mar Biol Ecol 243:227–240
Ceccherelli G, Piazzi L, Balata D (2002) Spread of introduced
Caulerpa species in macroalgal habitats. J Exp Mar Biol
Ecol 280:1–11
Chisholm JRM, Fernex FE, Mathieu D, Jaubert JM (1997)
Wastewater discharge, seagrass decline and algal prolif-
eration on the Cote d’ Azur. Mar Pollut Bull 34:78–84
Corbin JD, D’Antonio CM (2004) Competition between native
perennial and exotic annual grasses: implications for an
historical invasion. Ecology 85:1273–1283
Crawley MJ (1986) The population biology of invaders. Philo-
sophical transactions of the royal society of London. B Biol
Sci 314:711–731
Crooks JA (2002) Characterizing ecosystem-level conse-
quences of biological invasions: the role of ecosystem
engineers. Oikos 97:153–166
Cuny P, Serve L, Jupin H, Boudouresque CF (1995) Water
soluble phenolic compounds of the marine phanerogam
Posidonia oceanica in a Mediterranean area colonised by
the introduced chlorophyte Caulerpa taxifolia. Aquat Bot
52:237–242
Davis MA, Grime JP, Thompson K (2000) Fluctuating resources
in plant communities: a general theory of invasibility.
J Ecol 88:528–534
de Villele X, Verlaque M (1995) Changes and degradation in a
Posidonia oceanica bed invaded by the introduced tropical
alga Caulerpa taxifolia in the north western Mediterranean.
Bot Mar 38:79–87
Dıaz S, Symstad AJ, Stuart CF, Wardle DA, Huenneke LF
(2003) Functional diversity revealed by removal experi-
ments. Trends Ecol Evol 18:140–146
Didham R, Tylianakis JM, Hutchison MA, Ewers RM, Gemmell
NJ (2005) Are invasive species the drivers of ecological
change? Trends Ecol Evol 20:470–474
Duarte CM, Fourqurean JW, Krause-Jensen D, Olesen B
(2006) Dynamics of seagrass stability and change. In:
Larkum AWD, Orth RJ, Duarte CM (eds) Seagrasses:
biology, ecology and conservation. Springer, Dordrecht,
pp 271–294
Dumay O, Fernandez C, Pergent G (2002) Primary production
and vegetative cycle in Posidonia oceanica when in com-
petition with the green algae Caulerpa taxifolia and
Caulerpa racemosa. J Mar Biol Assoc UK 82:379–387
Elton CS (1958) The ecology of invasions by animals and plants,
p 175
Eyre BD, Maher D, Oakes JM, Erler DV, Glasby TM (2011)
Differences in benthic metabolism, nutrient fluxes, and
denitrification in Caulerpa taxifolia communities com-
pared to uninvaded bare sediment and seagrass (Zosteracapricorni) habitats. Limnol Oceanogr 56:1737–1750
Francour P, Pellissier V, Mangialajo L, Buisson E, Stadelmann
B, Veillard N, Meinesz A, Thibaut T, De Vaugelas J (2009)
Changes in invertebrate assemblages of Posidonia ocea-nica beds following Caulerpa taxifolia invasion. Vie et
Milieu Life Environ 59:31–38
Garcias-Bonet N, Marba N, Holmer M, Duarte CM (2008)
Effects of sediment sulfides on seagrass Posidonia ocea-nica meristematic activity. Mar Ecol Prog Ser 372:1–6
Gennaro P, Piazzi L (2011) Synergism between two anthropic
impacts: Caulerpa racemosa var. cylindracea invasion and
seawater nutrient enrichment. Mar Ecol Prog Ser
427:59–70
Glardon C, Walters LJ, Quintana-Ascencio P, McCauley L,
Stam WT, Olsen JL (2008) Predicting risks of invasion of
macroalgae in the genus Caulerpa in Florida. Biol Inva-
sions 10:1147–1157
Glasby TM, Creese RG (2007) Invasive marine species man-
agement and research. In: Connell SD, Gillanders BM
(eds) Marine ecology. Oxford University Press, Mel-
bourne, pp 569–594
Glasby TM, Underwood AJ (1996) Sampling to differentiate
between pulse and press perturbations. Environ Monit
Assess 42:241–252
Glasby TM, Creese RG, Gibson PT (2005) Experimental use of
salt to control the invasive marine alga Caulerpa taxifoliain New South Wales, Australia. Biol Conserv 122:573–580
Green EP, Short FT (2003) World atlas of seagrasses. University
of California Press, Berkeley
Grey D (2001) Caulerpa taxifolia: invasive weed prompts
response actions. Fisheries NSW Mag 4:4–5
Gribben PE, Wright JT (2006) Invasive seaweed enhances
recruitment of a native bivalve: roles of refuge from pre-
dation and habitat choice. Mar Ecol Prog Ser 318:177–185
Gribben PE, Byers JE, Clements M, McKenzie LA, Steinberg
PD, Wright JT (2009a) Behavioural interactions between
ecosystem engineers control community species richness.
Ecol Lett 12:1127–1136
Gribben PE, Wright JF, O’Connor WA, Doblin MA, Eyre B,
Steinberg PD (2009b) Reduced performance of native
infauna following recruitment to a habitat-forming inva-
sive marine alga. Oecologia (Berlin) 158:733–745
Harrison PG, Bigley RE (1982) The recent introduction of the
seagrass Zostera japonica Aschers. and Graebn. to the
Pacific Coast of North America. Can J Fish Aquat Sci
39:1642–1648
Caulerpa taxifolia in seagrass meadows
123

Hendriks I, Bouma T, Morris E, Duarte C (2010) Effects of
seagrasses and algae of the Caulerpa family on hydrody-
namics and particle-trapping rates. Mar Biol 157:473–481
Hobbs RJ, Huenneke LF (1992) Disturbance, diversity, and
invasion: implications for conservation. Conserv Biol
6:324–337
Holmer M, Marba N, Lamonte M, Duarte CM (2009) Deterio-
ration of sediment quality in seagrass meadows (Posidoniaoceanica) invaded by macroalgae (Caulerpa sp.). Estuar
Coasts 32:456–466
Jaubert JM, Chisholm JRM, Ducrot D, Ripley HT, Roy L,
Passeron-Seitre G (1999) No deleterious alterations in
Posidonia beds in the Bay of Menton (France) eight
years after Caulerpa taxifolia colonization. J Phycol 35:
1113–1119
Jaubert JM, Chisholm JRM, Minghelli-Roman A, Marchioretti
M, Morrow JH, Ripley HT (2003) Re-evaluation of the
extent of Caulerpa taxifolia development in the northern
Mediterranean using airborne spectrographic sensing. Mar
Ecol Prog Ser 263:75–82
Johnstone IM (1986) Plant invasion windows: a time-based
classification of invasion potential. Biol Rev 61:369–394
Kiparissis S, Fakiris E, Papatheodorou G, Geraga M, Kornaros
M, Kapareliotis A, Ferentinos G (2011) Illegal trawling
and induced invasive algal spread as collaborative factors
in a Posidonia oceanica meadow degradation. Biol Inva-
sions 13:669–678
Kirkman H, Kuo J (1990) Pattern and process in southern
Western Australian seagrasses. Aquat Bot 37:367–382
Lapointe BE, Bedford BJ (2010) Ecology and nutrition of
invasive Caulerpa brachypus f. parvifolia blooms on coral
reefs off southeast Florida, USA. Harmful Algae 9:1–12
Larkum AWD (1976) Ecology of Botany Bay I. Growth of
Posidonia australis (Brown) Hook. f. in Botany Bay and
other bays of the Sydney basin. Aust J Mar Freshw Res
27:117–127
Larkum AWD, West RJ (1990) Long-term changes of seagrass
meadows in Botany Bay, Australia. Aquat Bot 37:55–70
Levi F, Francour P (2004) Behavioural response of Mullussurmuletus to habitat modification by the invasive macro-
alga Caulerpa taxifolia. J Fish Biol 64:55–64
Longepierre S, Robert A, Levi F, Francour P (2005) How an
invasive alga species (Caulerpa taxifolia) induces changes
in foraging strategies of the benthivorous fish Mullus sur-muletus in coastal Mediterranean ecosystems. Biodivers
Conserv 14:365–376
MacArthur RH (1970) Species packing and competetive equi-
librium for many species. Theor Popul Biol 1:1–11
MacDougall AS, Turkington R (2005) Are invasive species the
drivers or passengers of change in degraded ecosystems?
Ecology 86:42–55
Mack RN, Simberloff D, Lonsdale WM, Evans H, Clout M,
Bazzaz FA (2000) Biotic invasions: causes, epidemiology,
global consequences, and control. Ecol Appl 10:689–710
McKinnon JG, Gribben PE, Davis AR, Jolley DF, Wright JT
(2009) Differences in soft-sediment macrobenthic assem-
blages invaded by Caulerpa taxifolia compared to unin-
vaded habitats. Mar Ecol Prog Ser 380:59–71
Meehan AJ, Williams RJ, Watford FA (2005) Detecting
trends in seagrass abundance using aerial photograph
interpretation: problems arising with the evolution of
mapping methods. Estuaries 28:462–472
Meinesz A, de Vaugelas J, Hesse B, Mari X (1993) Spread of the
introduced tropical green alga Caulerpa taxifolia in the
northern Mediterranean waters. J Appl Phycol 5:141–147
Meinesz A, Belsher T, Thibaut T, Antolic B, Ben Mustapha K,
Boudouresque CF, Chiaverini D, Cinelli F, Cottalorda JM,
Djellouli A, El Adeb A, Orestano C, Grau AM, Ivesa L,
Jaklin A, Langar H, Massuti-Pascual E, Peirano A, Tunesi
L, de Vaugelas J, Zavodnik N, Zuljevic A (2001) The
introduced green alga Caulerpa taxifolia continues to
spread in the Mediterranean. Biol Invasions 3:201–210
Meusnier I, Valero M, Olsen JL, Stam WT (2004) Analysis of
rDNA ITS1 indels in Caulerpa taxifolia (Chlorophyta)
supports a derived, incipient species status for the invasive
strain. Eur J Phycol 39:83–92
Molenaar H, Meinesz A, Thibaut T (2009) Alterations of the
structure of Posidonia oceanica beds due to the introduced
alga Caulerpa taxifolia. Scientia Marina 73:329–335
Occhipinti-Ambrogi A, Savini D (2003) Biological invasions as
a component of global change in stressed marine ecosys-
tems. Mar Pollut Bull 46:542–551
Orth RJ, Carruthers TJB, Dennison WJ, Duarte CM, Fourqurean
JW, Heck KL Jr, Hughes AR, Kendrick GA, Kenworthy WJ,
Olyarink S, Short FT, Waycott M, Williams SL (2006) A
global crisis for seagrass ecosystems. Bioscience 56:987–996
Parker IM, Simberloff D, Lonsdale WM, Goodell K, Wonham
M, Kareiva PM, Williamson MH, Von Holle B, Moyle PB,
Byers JE, Goldwasser L (1999) Impact: toward a frame-
work for understanding the ecological effects of invaders.
Biol Invasions 1:3–19
Peirano A, Damasso V, Montefalcone M, Morri C, Bianchi CN
(2005) Effects of climate, invasive species and anthropo-
genic impacts on the growth of the seagrass Posidoniaoceanica (L.) Delile in Liguria (NW Mediterranean Sea).
Mar Pollut Bull 50:817–822
Peirano A, Cocito S, Banfi V, Cupido R, Damasso V, Farina G,
Lombardi C, Mauro R, Morri C, Roncarolo I, Saldana S,
Savini D, Sgorbini S, Silvestri C, Stoppelli N, Torricelli L,
Bianchi CN (2011) Phenology of the Mediterranean sea-
grass Posidonia oceanica (L.) Delile: medium and long-
term cycles and climate inferences. Aquat Bot 94:77–92
Pergent G, Boudouresque CF, Dumay O, Pergent-Martini C,
Wyllie-Echeverria S (2008) Competition between the
invasive macrophyte Caulerpa taxifolia and the seagrass
Posidonia oceanica: contrasting strategies. BMC Ecol 8:20
Piazzi L, Balata D, Ceccherelli G, Cinelli F (2005) Interactive
effect of sedimentation and Caulerpa racemosa var. cy-lindracea invasion on macroalgal assemblages in the
Mediterranean Sea. Estuar Coast Shelf Sci 64:467–474
Posey MH (1988) Community changes associated with the
spread of an introduced seagrass, Zostera japonica. Ecol-
ogy 69:974–983
Rodriguez L (2006) Can invasive species facilitate native spe-
cies? Evidence of how, when, and why these impacts occur.
Biol Invasions 8:927–939
Ruitton S, Javel F, Culioli J-M, Meinesz A, Pergent G, Verlaque
M (2005) First assessment of the Caulerpa racemosa(Caulerpales, Chlorophyta) invasion along the French
Mediterranean coast. Mar Pollut Bull 50:1061–1068
T. M. Glasby
123

Ruiz GM, Fofonoff P, Hines AH, Grosholz ED (1999) Non-
indigenous species as stressors in estuarine and marine
communities: assessing invasion impacts and interactions.
Limnol Oceanogr 44:950–972
Schaffelke B, Murphy N, Uthicke S (2002) Using genetic
techniques to investigate the sources of the invasive alga
Caulerpa taxifolia in three new locations in Australia. Mar
Pollut Bull 44:204–210
Schaffelke B, Smith JE, Hewitt CL (2006) Introduced macro-
algae—a growing concern. J Appl Phycol 18:529–541
Short FT, Wyllie-Echeverria S (1996) Natural and human-
induced disturbance of seagrasses. Environ Conserv 23:
17–27
Short FT, Fernandez E, Vernon A, Gaeckle JL (2006) Occur-
rence of Halophila baillonii meadows in Belize, Central
America. Aquat Bot 85:249–251
Short FT, Polidoro B, Livingstone SR, Carpenter KE, Bandeira
S, Bujang JS, Calumpong HP, Carruthers TJB, Coles RG,
Dennison WC, Erftemeijer PLA, Fortes MD, Freeman AS,
Jagtap TG, Kamal AHM, Kendrick GA, Judson Kenworthy
W, La Nafie YA, Nasution IM, Orth RJ, Prathep A,
Sanciangco JC, Tussenbroek BV, Vergara SG, Waycott M,
Zieman JC (2011) Extinction risk assessment of the
world’s seagrass species. Biol Conserv 144:1961–1971
Simberloff D (2009) Invasions of plant communities—more of
the same, something very different, or both? Am Midl Nat
163:220–233
Stafford NB, Bell SS (2006) Space competition between sea-
grass and Caulerpa prolifera (Forsskaal) Lamouroux fol-
lowing simulated disturbances in Lassing Park, FL. J Exp
Mar Biol Ecol 333:49–57
Tanner JE (2011) Utilisation of the invasive alga Caulerpataxifolia as habitat by faunal assemblages in the Port
River–Barker Inlet Estuary, South Australia. Estuaries
Coasts 34:831–838
Taplin KA, Irlandi EA, Raves R (2005) Interference between the
macroalga Caulerpa prolifera and the seagrass Halodulewrightii. Aquat Bot 83:175–186
Thibaut T, Meinesz A, Coquillard P (2004) Biomass seasonality
of Caulerpa taxifolia in the Mediterranean Sea. Aquat Bot
80:291–297
Thomsen MA, D’Antonio CM, Suttle KB, Sousa WP (2006)
Ecological resistance, seed density and their interactions
determine patterns of invasion in a California coastal
grassland. Ecol Lett 9:160–170
Thomsen MS, Wernberg T, Tuya F, Silliman BR (2009) Evi-
dence for impacts of nonindigenous macroalgae: a meta-
analysis of experimental field studies. J Phycol 45:812–819
Valentine JP, Johnson CR (2003) Establishment of the intro-
duced kelp Undaria pinnatifida in Tasmania depends on
disturbance to native algal assemblages. J Exp Mar Biol
Ecol 295:63–90
Valentine JP, Johnson CR (2004) Establishment of the intro-
duced kelp Undaria pinnatifida following dieback of the
native macroalga Phyllospora comosa in Tasmania, Aus-
tralia. Mar Freshw Res 55:223–230
Walker DI, McComb AJ (1992) Seagrass degradation in Aus-
tralian coastal waters. Mar Pollut Bull 25:191–195
Walker DI, Kendrick GA, McComb AJ (2006) Decline and
recovery of seagrass ecosystems—the dynamics of change.
In: Larkum AWD, Orth RJ, Duarte CM (eds) Seagrasses:
biology, ecology and conservation. Springer, Dordrecht,
pp 551–565
West RJ, Thorogood C, Walford T, Williams RJ (1985) An
estuarine inventory for New South Wales, Australia.
Fisheries Bulletin 2. Department of Agriculture, Cronulla,
p 140
Williams SL (1987) Competition between the seagrasses Thal-assia testudinum and Syringodium filiforme in a Caribbean
lagoon Mar. Ecol Prog Ser 35:91–98
Williams SL (2007) Introduced species in seagrass ecosystems:
status and concerns. J Exp Mar Biol Ecol 350:89–110
Williams SL, Grosholz ED (2002) Preliminary reports from the
Caulerpa taxifolia invasion in southern California. Mar
Ecol Prog Ser 233:307–310
Williams SL, Breda VA, Anderson TW, Nyden BB (1985)
Growth and sediment disturbances of Caulerpa spp.
(Chlorophyta) in a submarine canyon. Mar Ecol Prog Ser
21:275–281
Williams SL, Smith JE (2007) A global review of the distribu-
tion, taxonomy, and impacts of introduced seaweeds. Annu
Rev Ecol Evol Syst 38:327–359
Wright JT (2005) Differences between native and invasive
Caulerpa taxifolia: a link between asexual fragmentation
and abundances in invasive populations. Ecology 147:
559–569
Wright JT, Davis AR (2006) Demographic feedback between
clonal growth and fragmentation in an invasive seaweed.
Ecology 87:1744–1754
Wright JT, Gribben PE (2008) Predicting the impact of an
invasive seaweed on the fitness of native fauna. J Appl Ecol
45:1540–1549
Wright JT, McKenzie LA, Gribben PE (2007) A decline in the
abundance and condition of a native bivalve associated
with Caulerpa taxifolia invasion. Mar Freshw Res
58:263–272
York PH, Booth DJ, Glasby TM, Pease BC (2006) Fish
assemblages in habitats dominated by Caulerpa taxifoliaand native seagrasses in south-eastern Australia. Mar Ecol
Prog Ser 312:223–234
Caulerpa taxifolia in seagrass meadows
123