cardiology copyright © 2019 chronic hypoxia–induced …liu et al., sci. transl. med. 11, eaat8406...
TRANSCRIPT

Liu et al., Sci. Transl. Med. 11, eaat8406 (2019) 24 April 2019
S C I E N C E T R A N S L A T I O N A L M E D I C I N E | R E S E A R C H A R T I C L E
1 of 14
C A R D I O L O G Y
Chronic hypoxia–induced Cirbp hypermethylation attenuates hypothermic cardioprotection via down-regulation of ubiquinone biosynthesisYiwei Liu1*, Junyue Xing1,2*, Yongnan Li1,3, Qipeng Luo1, Zhanhao Su1, Xiaoling Zhang1,2, Hao Zhang1,2,4,5†
Therapeutic hypothermia is commonly used during cardiopulmonary bypass (CPB) to protect the heart against myocardial injury in cardiac surgery. Patients who suffer from chronic hypoxia (CH), such as those with certain heart or lung conditions, are at high risk of severe myocardial injury after cardiac surgery, but the underlying mecha-nisms are unknown. This study tested whether CH attenuates hypothermic cardioprotection during CPB. Using a rat model of CPB, we found that hypothermic cardioprotection was impaired in CH rats but was preserved in nor-moxic rats. Cardiac proteomes showed that cold-inducible RNA binding protein (CIRBP) was significantly (P = 0.03) decreased in CH rats during CPB. Methylation analysis of neonatal rat cardiomyocytes under CH and myocardium specimens from patients with CH showed that CH induced hypermethylation of the Cirbp promoter region, resulting in its depression and failure to respond to cold stress. Cirbp-knockout rats showed attenuated hypothermic cardio-protection, whereas Cirbp-transgenic rats showed an enhanced response. Proteomics analysis revealed that the cardiac ubiquinone biosynthesis pathway was down-regulated during CPB in Cirbp-knockout rats, resulting in a significantly (P = 0.01) decreased concentration of ubiquinone (CoQ10). Consequently, cardiac oxidative stress was aggravated and adenosine 5′-triphosphate production was impaired, leading to increased myocardial injury during CPB. CoQ10-supplemented cardioplegic solution improved cardioprotection in rats exposed to CH, but its effect was limited in normoxic rats. Our study suggests that an individualized cardioprotection strategy should be used to fully compensate for the consequences of epigenetic modification of Cirbp in patients with CH who require ther-apeutic hypothermia.
INTRODUCTIONChronic hypoxia (CH) affects patients with cyanotic congenital heart disease (CHD), chronic obstructive pulmonary disease (COPD), or obstructive sleep apnea (OSA), as well as individuals who live at high altitudes (hereafter collectively referred to as patients with CH). Accu-mulating evidence shows that these patients are at high risk of compli-cations after cardiac surgery, presenting as severe cardiopulmonary bypass (CPB)–related myocardial injury and worsened cardiac function. Compared with noncyanotic CHD, patients with cyanotic CHD (CCHD) had a two- to threefold increased risk for postoperative low cardiac out-put syndrome (1). Increased incidence of major adverse cardiovascular events after open-heart surgery was also associated with COPD or OSA (2, 3). Recently, patients from a Tibetan high-altitude area were found to have a higher risk of postoperative cardiac dysfunction when compared with low- altitude patients (4). Previous studies showed that chronically hypoxic myocardium was vulnerable to free radical in-jury and suffered impaired adenosine 5′-triphosphate (ATP) production during CPB (1, 5), but the underlying mechanism is still unknown.
We posited that the root to this mechanism might lie in a protein that responds to both hypoxia and another form of physiological stress: hypothermia. Hypothermia is common strategy used to pro-tect the heart against myocardial injury during CPB in open-heart surgery. It is well known that low temperature confers cardioprotec-tion by reducing metabolic activity and oxygen demands; however, hypothermia is associated with some disadvantages at the cellular level (6). To counter its negative effects, cold shock proteins are activated by hypothermia. In recent studies, cold-inducible RNA binding protein (CIRBP) has been reported to be a critical cold shock protein induced under cold stress (7, 8). CIRBP is a member of RNA binding proteins located in the nucleus. Under cold stress, CIRBP translocates from the nucleus to the cytoplasm to regulate its target mRNAs and protects the cell from apoptosis (8). A previous study showed that CIRBP exerts neuroprotective effects during therapeutic hypothermia (9). However, the role of CIRBP in hypothermic cardioprotection has not been studied.
In addition to cold stress, hypoxia also regulates the expression of CIRBP. Upon acute hypoxia, CIRBP expression is up-regulated (10), whereas CH depresses the expression of CIRBP by an unknown mechanism (11). Several studies have reported that hypoxia can lead to hypermethylation of gene promoter regions by reducing ten eleven translocation enzyme (TET) activity, which represses transcription of these genes (12, 13). TET is able to oxidize 5-methylcytosine (5mC) to 5-hydroxymethylcytosine (5hmC), which is further oxidized and replaced with an unmodified C to achieve demethylation. This process depends on oxygen supply, and it could be inhibited by hypoxia (12). Therefore, we hypothesized that CH might induce epigenetic modifications in Cirbp, which would lead to the attenuation of the
1State Key Laboratory of Cardiovascular Disease, Fuwai Hospital, National Center for Cardiovascular Diseases, Chinese Academy of Medical Sciences and Peking Union Medical College, Beijing 100037, China. 2Heart Center of Henan Provincial People Hospital and Key Laboratory of Cardiac Regenerative Medicine, National Health Commission, Zhengzhou 451464, China. 3Department of Cardiac Surgery, Lanzhou University Second Hospital, Lanzhou University, Lanzhou 730030, China. 4Center for Pediatric Cardiac Surgery, Fuwai Hospital, Chinese Academy of Medical Sci-ences and Peking Union Medical College, Beijing 100037, China. 5Heart Center and Shanghai Institute of Pediatric Congenital Heart Diseases, Shanghai Children’s Med-ical Center, Shanghai Jiaotong University School of Medicine, Shanghai 200127, China.*These authors contributed equally to this work.†Corresponding author. Email: [email protected]
Copyright © 2019 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works
by guest on April 26, 2020
http://stm.sciencem
ag.org/D
ownloaded from

Liu et al., Sci. Transl. Med. 11, eaat8406 (2019) 24 April 2019
S C I E N C E T R A N S L A T I O N A L M E D I C I N E | R E S E A R C H A R T I C L E
2 of 14
cardioprotective effect of hypothermia during the CPB procedure in patients with CH who received open-heart surgery.
Here, we established a rat model that mimics clinical CPB, in which ice slush and cold cardioplegic solutions were applied to pro-tect the myocardium. We found that cardiac CIRBP was induced by hypothermia during CPB; however, the extent of this induction was decreased in rats that had been subjected to CH due to hypermethyla-tion in the Cirbp promoter region. This epigenetic modification led to impaired biosynthesis of cardiac CoQ10 during CPB, resulting in worse postoperative cardiac function in rats and patients with CH.
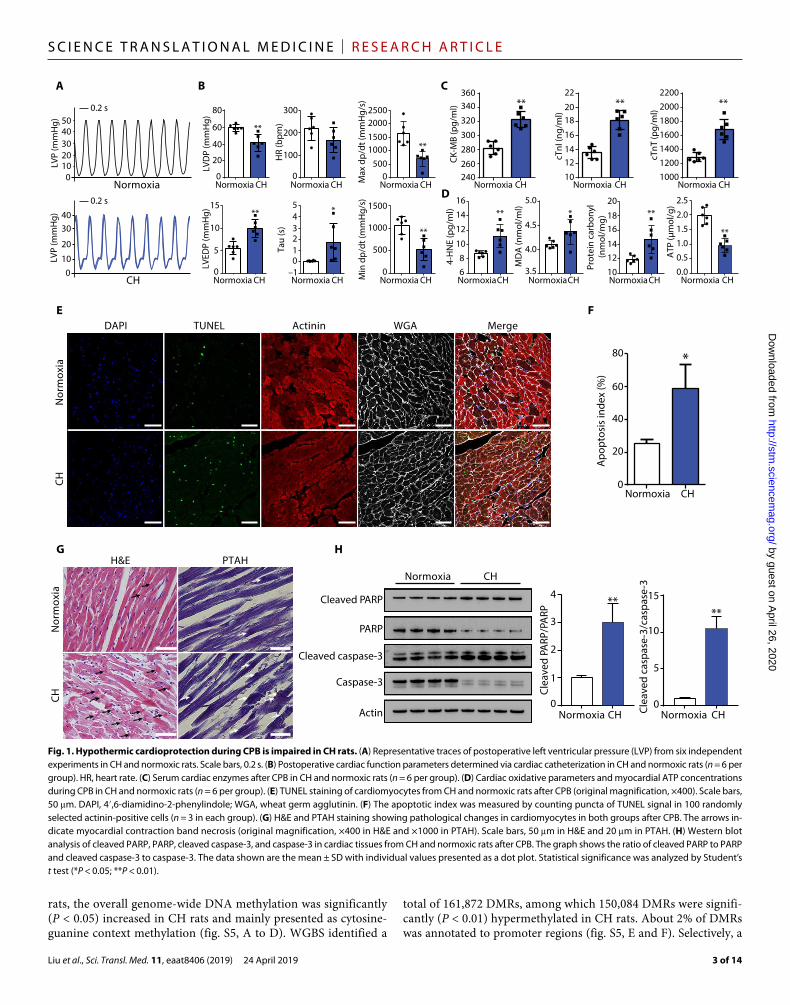
RESULTSCardioprotective effects of hypothermia during CPB is impaired in CHWe established a rat model of CH (hereafter referred to as CH rats) as previously described (14). Arterial blood gas analysis confirmed that our CH rat model was successfully established (table S1). At baseline, echocardiography showed no differences in cardiac function between CH rats and normoxic controls (fig. S1). We then subjected both groups to CPB and applied hypothermia to protect the heart during this procedure (fig. S2 and movie S1). During cardiac arrest, the hearts were cooled with local ice-slush application and perfused with cold cardioplegic solutions. Postoperative cardiac functions were measured by cardiac catheterization. Relative to normoxic rats, CH rats had reduced left ventricular developed pressure (LVDP) (P < 0.01), maximum (max) dp/dt (a measure of contractile func-tion) (P < 0.01), and minimum (min) dp/dt values (a measure of the ability of the heart to relax) (P < 0.01), and increased left ventric-ular end-diastolic pressure (LVEDP) (P < 0.01) and time constant of left ventricular isovolumic relaxation (Tau) values (P < 0.05), indicating that CH rats suffered from more severe impairment of cardiac function (Fig. 1, A and B). Compared with normoxic rats, postoperative serum cardiac enzymes in CH rats showed signifi-cantly higher concentrations of creatine kinase–myocardial band (CK-MB) (P < 0.01), cardiac troponin I (cTnI) (P < 0.01), and car-diac troponin T (cTnT) (P < 0.01), indicating profound myocardial injury in CH rats (Fig. 1C). Moreover, cardiac 4-hydroxy-2-nonenal (4-HNE) (P < 0.01), malondialdehyde (MDA) (P < 0.05), and car-bonyl proteins (P < 0.01) were significantly increased and ATP concentration was significantly decreased (P < 0.01) in CH rats (Fig. 1D), indicating that the chronically hypoxic myocardium suf-fered aggravative oxidative stress and impaired ATP production during CPB.
TUNEL (terminal deoxynucleotidyl transferase–mediated deoxyu-ridine triphosphate nick end labeling) staining showed that CH rats had more apoptotic cardiomyocytes than normoxic rats (58.7% versus 25.3%; P < 0.05) (Fig. 1, E and F). We next performed histo-pathological analyses using hematoxylin and eosin (H&E) and phosphotungstic acid hematoxylin (PTAH) staining (Fig. 1G). Rel-ative to normoxic rats, CH rats showed loss of cross-striations, more frequent sarcolemmal rupture, and reduced clarity of nuclei in myocardial fibers. Furthermore, prominent myocardial contraction band necrosis was observed in CH rats but not in normoxic rats. In addition, compared with normoxic rats, Western blot analysis showed that the ratio of cleaved to full-length poly(ADP-ribose) polymerase (PARP) and cleaved to full-length caspase-3 were signifi-cantly increased in CH rats (P < 0.01), indicating that the apoptosis signaling pathways were significantly activated in chronically hypoxic
myocardium during CPB (Fig. 1H). Together, our findings demon-strated that CH attenuated cardioprotective effects of hypothermia during CPB.
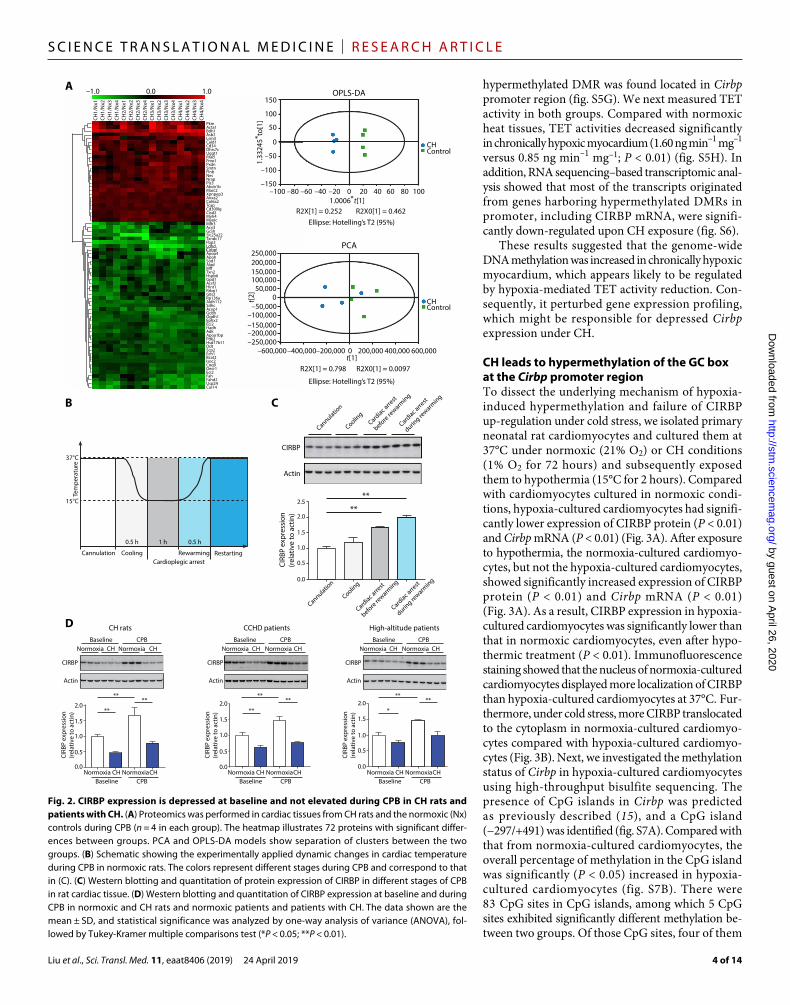
CIRBP depression underlies poor hypothermic cardioprotection during CPB in CHTo explain why CH impaired hypothermic cardioprotection during CPB, we analyzed proteomes in hearts that were harvested while CH and normoxic rats were undergoing CPB. A total of 2524 proteins were identified, where 72 proteins displayed significantly different expression between the two groups (fold change <0.83 or fold change >1.2; P < 0.05; table S2). Principal components analysis (PCA) and orthogonal projections to latent structures discriminant analy-sis (OPLS-DA) exhibited an obvious separation of clusters between the two groups (Fig. 2A). Among 45 significantly decreased proteins, CIRBP ranked 12th and was significantly decreased in the CH group (fold change = 0.77; P = 0.03). CIRBP is a cold shock protein induced by hypothermia and has been reported to play a role in hypothermic neuroprotection (8, 9).
We recorded cardiac temperature and measured CIRBP expres-sion at various stages of CPB in normoxic rats. In the cooling stage, cardiac temperature fell from 37° to 15°C in 0.5 hours and was maintained at about 15°C for 1 hour during cardiac arrest (Fig. 2B). During this process, the expression of CIRBP increased significantly (P < 0.01), indicating that CIRBP might behave as an acute-phase protein in cardiomyocytes under cold stress (Fig. 2C).
We also analyzed cardiac CIRBP expression at baseline (before CPB) and its response to cold stress during CPB in CH rats and patients with CH (all patient characteristics are summarized in table S3). At baseline, Western blot analysis showed that cardiac CIRBP expression was significantly lower in CH rats than in normoxic controls (P < 0.01). Similar findings were observed in patients with CCHD (P < 0.01) and in high altitude–dwelling patients (P < 0.05) (Fig. 2D). Moreover, compared with baseline, cardiac CIRBP expres-sion increased significantly during CPB in normoxic rats (P < 0.01) and patients (P < 0.01) undergoing open-heart surgery. By contrast, CIRBP expression was not significantly increased during CPB in CH rats, patients with CCHD, and high altitude–dwelling patients, indicating a failed response to cold stress. Consequently, cardiac CIRBP expression during CPB in CH rats and patients with CH was significantly lower than that in normoxic controls (P < 0.01).
To clarify whether there was a sex-specific effect, we performed additional animal experiments using female CH rats. We found that sex did not alter the trends of the CH-induced decreased expression of CIRBP and subsequent impairment in postoperative cardiac func-tion (fig. S3, A and B). These results suggested that the failure of CIRBP expression might be responsible for the impaired hypothermic cardio-protection during CPB observed in CH rats and patients with CH.
CH causes genome-wide DNA hypermethylation and alters transcriptional profiling in myocardiumPrevious studies have reported that hypoxia can lead to DNA hyper-methylation by reducing TET activity, which can regulate gene expression at the transcriptional level (12, 13), so we used whole- genome bisulfite sequencing (WGBS) to analyze the genome-wide DNA methylation profile of cardiac tissues harvested from normoxic and CH rats. The Circos plot shows the global methylation patterns and locations of differentially methylated regions (DMRs) in different chromosomes between two groups (fig. S4). Compared with normoxic
by guest on April 26, 2020
http://stm.sciencem
ag.org/D
ownloaded from
Liu et al., Sci. Transl. Med. 11, eaat8406 (2019) 24 April 2019
S C I E N C E T R A N S L A T I O N A L M E D I C I N E | R E S E A R C H A R T I C L E
3 of 14
rats, the overall genome-wide DNA methylation was significantly (P < 0.05) increased in CH rats and mainly presented as cytosine- guanine context methylation (fig. S5, A to D). WGBS identified a
total of 161,872 DMRs, among which 150,084 DMRs were signifi-cantly (P < 0.01) hypermethylated in CH rats. About 2% of DMRs was annotated to promoter regions (fig. S5, E and F). Selectively, a
0
20
40
60
80
0
100
200
300
0500
1000150020002500
–1012345
0
500
1000
1500
**
** ** **
*** * ** **
****
LVP
(mm
Hg)
LVP
(mm
Hg)
*
Normoxia CH
CH
240260
280
300
320
340360
16
14
12
10
86
Normoxia CH Normoxia CH Normoxia Normoxia CHNormoxia CHNormoxia
CHNormoxia
CH
Normoxia CH Normoxia CH Normoxia CH
0
5
10
15LV
EDP
(mm
Hg)
LVD
P (m
mH
g)
HR
(bpm
)Ta
u (s
)
Min
dp/
dt (m
mH
g/s)
Max
dp/
dt (m
mH
g/s)
CK-M
B (p
g/m
l)
cTnI
(ng/
ml)
cTnT
(pg/
ml)
4-H
NE
(pg/
ml)
MD
A (n
mol
/ml)
Prot
ein
carb
onyl
(n
mol
/mg)
01020304050
010
20
3040
Nor
mox
ia
Normoxia
Normoxia
Nor
mox
ia
H&E PTAH
Normoxia
CH
CH
CH Normoxia CH
CH
CH
Normoxia CH1000
10
1214
1618
20
120014001600180020002200
B
E
D
F
G
Normoxia CH10
3.5
4.0
4.5
5.0
1214
16
1820
22
DAPI TUNEL Actinin WGA Merge
0
0 0
5
10
15
1
2
3
4
20
40
60
80
Apo
ptos
is in
dex
(%)
Cleaved PARP
PARP
Cleaved caspase-3
Caspase-3
Actin
Clea
ved
PARP
/PA
RP
Clea
ved
casp
ase-
3/ca
spas
e-3
0.2 s
0.2 s
**
** **
0.0
0.5
1.0
1.5
2.0
2.5
CHNormoxia
ATP
(μm
ol/g
)
CA
H
Fig. 1. Hypothermic cardioprotection during CPB is impaired in CH rats. (A) Representative traces of postoperative left ventricular pressure (LVP) from six independent experiments in CH and normoxic rats. Scale bars, 0.2 s. (B) Postoperative cardiac function parameters determined via cardiac catheterization in CH and normoxic rats (n = 6 per group). HR, heart rate. (C) Serum cardiac enzymes after CPB in CH and normoxic rats (n = 6 per group). (D) Cardiac oxidative parameters and myocardial ATP concentrations during CPB in CH and normoxic rats (n = 6 per group). (E) TUNEL staining of cardiomyocytes from CH and normoxic rats after CPB (original magnification, ×400). Scale bars, 50 m. DAPI, 4′,6- diamidino-2-phenylindole; WGA, wheat germ agglutinin. (F) The apoptotic index was measured by counting puncta of TUNEL signal in 100 randomly selected actinin- positive cells (n = 3 in each group). (G) H&E and PTAH staining showing pathological changes in cardiomyocytes in both groups after CPB. The arrows in-dicate myocardial contraction band necrosis (original magnification, ×400 in H&E and ×1000 in PTAH). Scale bars, 50 m in H&E and 20 m in PTAH. (H) Western blot analysis of cleaved PARP, PARP, cleaved caspase-3, and caspase-3 in cardiac tissues from CH and normoxic rats after CPB. The graph shows the ratio of cleaved PARP to PARP and cleaved caspase-3 to caspase-3. The data shown are the mean ± SD with individual values presented as a dot plot. Statistical significance was analyzed by Student’s t test (*P < 0.05; **P < 0.01).
by guest on April 26, 2020
http://stm.sciencem
ag.org/D
ownloaded from
Liu et al., Sci. Transl. Med. 11, eaat8406 (2019) 24 April 2019
S C I E N C E T R A N S L A T I O N A L M E D I C I N E | R E S E A R C H A R T I C L E
4 of 14
hypermethylated DMR was found located in Cirbp promoter region (fig. S5G). We next measured TET activity in both groups. Compared with normoxic heat tissues, TET activities decreased significantly in chronically hypoxic myocardium (1.60 ng min–1 mg–1 versus 0.85 ng min–1 mg–1; P < 0.01) (fig. S5H). In addition, RNA sequencing–based transcriptomic anal-ysis showed that most of the transcripts originated from genes harboring hypermethylated DMRs in promoter, including CIRBP mRNA, were signifi-cantly down-regulated upon CH exposure (fig. S6).
These results suggested that the genome-wide DNA methylation was increased in chronically hypoxic myocardium, which appears likely to be regulated by hypoxia-mediated TET activity reduction. Con-sequently, it perturbed gene expression profiling, which might be responsible for depressed Cirbp expression under CH.
CH leads to hypermethylation of the GC box at the Cirbp promoter regionTo dissect the underlying mechanism of hypoxia- induced hypermethylation and failure of CIRBP up-regulation under cold stress, we isolated primary neonatal rat cardiomyocytes and cultured them at 37°C under normoxic (21% O2) or CH conditions (1% O2 for 72 hours) and subsequently exposed them to hypothermia (15°C for 2 hours). Compared with cardiomyocytes cultured in normoxic condi-tions, hypoxia-cultured cardiomyocytes had signifi-cantly lower expression of CIRBP protein (P < 0.01) and Cirbp mRNA (P < 0.01) (Fig. 3A). After exposure to hypothermia, the normoxia-cultured cardiomyo-cytes, but not the hypoxia-cultured cardiomyocytes, showed significantly increased expression of CIRBP protein (P < 0.01) and Cirbp mRNA (P < 0.01) (Fig. 3A). As a result, CIRBP expression in hypoxia- cultured cardiomyocytes was significantly lower than that in normoxic cardiomyocytes, even after hypo-thermic treatment (P < 0.01). Immunofluorescence staining showed that the nucleus of normoxia-cultured cardiomyocytes displayed more localization of CIRBP than hypoxia-cultured cardiomyocytes at 37°C. Fur-thermore, under cold stress, more CIRBP translocated to the cytoplasm in normoxia-cultured cardiomyo-cytes compared with hypoxia-cultured cardiomyo-cytes (Fig. 3B). Next, we investigated the methylation status of Cirbp in hypoxia-cultured cardiomyocytes using high-throughput bisulfite sequencing. The presence of CpG islands in Cirbp was predicted as previously described (15), and a CpG island (−297/+491) was identified (fig. S7A). Compared with that from normoxia-cultured cardiomyocytes, the overall percentage of methylation in the CpG island was significantly (P < 0.05) increased in hypoxia- cultured cardiomyocytes (fig. S7B). There were 83 CpG sites in CpG islands, among which 5 CpG sites exhibited significantly different methylation be-tween two groups. Of those CpG sites, four of them
–1.0 0.0 1.0
PkmActa1Bdh1Asb2Lsm3Cald1Cd34Dhrs7cUggt1RradFmo1PxdnSmtnFlnbNesNrapPls3Abcb1bMarc2Xpnpep3Anxa2Col6a2TcapCd300lgCisd2Myh4MyolcMlh3Acy3GcshSlc25a22Txndc17Fbp2LdhdCirbpApoa4ApohSod1AladM�Txn2Hspb6Gpd1Acsf2Hint1Rrbp1G0s2Rp136aAldh112SdhcAcyp1GcdhOgdh1Ephx2Eci2HadhAdkApoa1bpPdk2Hsd17b11DdtScp2Ech1Bcat2Isoc2Coq9Decr1Eci2FahFahd2Usp24Car14
150
100
50
0
–50
–100
–150–100 –80 –60 –40 –20 0 20 40 60 80 100
1.33
245
to[1
]*
*1.0006 t[1]R2X[1] = 0.252 R2X0[1] = 0.462
Ellipse: Hotelling’s T2 (95%)
CHControl
–250,000–200,000
CHControl
–150,000–100,000
–50,0000
50,000100,000150,000
–600,000–400,000–200,000
200,000250,000
200,000 400,000 600,0000
R2X[1] = 0.798 R2X0[1] = 0.0097
Ellipse: Hotelling’s T2 (95%)
t[1]
t[2]
A
CoolingCardioplegic arrest
Rewarming Restarting
37°C
15°C
0.5 h 1 h 0.5 h
Tem
pera
ture
Cannulation
CH1/
Nx1
CH1/
Nx2
CH1/
Nx3
CH1/
Nx4
CH2/
Nx1
CH2/
Nx2
CH2/
Nx3
CH2/
Nx4
CH3/
Nx1
CH3/
Nx2
CH3/
Nx3
CH3/
Nx4
CH4/
Nx1
CH4/
Nx2
CH4/
Nx3
CH4/
Nx4
BCard
iac arre
st
before rewarm
ing
Cardiac a
rrest
during re
warming
Cooling
Cannulation
Cardiac a
rrest
before rewarm
ing
Cardiac a
rrest
during re
warming
Cooling
Cannulation
CIRB
P ex
pres
sion
(rel
ativ
e to
act
in) **
**
CIRBP
Actin
0.0
0.5
1.0
1.5
2.0
2.5
C
Normoxia
CH rats CCHD patients High-altitude patients
**
0.0
0.5
1.0
1.5
2.0
CH Normoxia CH
CIRBP CIRBP
ActinActin
CIRB
P ex
pres
sion
(rel
ativ
e to
act
in)
**
0.0
0.5
1.0
1.5
2.0
CIRB
P ex
pres
sion
(rel
ativ
e to
act
in)
0.5
0.0
1.0
1.5
2.0
CIRB
P ex
pres
sion
(rel
ativ
e to
act
in)
CIRBP
Actin
**
**
** *
** ****
Baseline CPB
Normoxia CH NormoxiaCHBaseline CPB
Normoxia CH NormoxiaCHBaseline CPB
Normoxia CH NormoxiaCHBaseline CPB
Normoxia CH Normoxia CHBaseline CPB
Normoxia CH Normoxia CHBaseline CPB
D
OPLS-DA
PCA
Fig. 2. CIRBP expression is depressed at baseline and not elevated during CPB in CH rats and patients with CH. (A) Proteomics was performed in cardiac tissues from CH rats and the normoxic (Nx) controls during CPB (n = 4 in each group). The heatmap illustrates 72 proteins with significant differ-ences between groups. PCA and OPLS-DA models show separation of clusters between the two groups. (B) Schematic showing the experimentally applied dynamic changes in cardiac temperature during CPB in normoxic rats. The colors represent different stages during CPB and correspond to that in (C). (C) Western blotting and quantitation of protein expression of CIRBP in different stages of CPB in rat cardiac tissue. (D) Western blotting and quantitation of CIRBP expression at baseline and during CPB in normoxic and CH rats and normoxic patients and patients with CH. The data shown are the mean ± SD, and statistical significance was analyzed by one-way analysis of variance (ANOVA), fol-lowed by Tukey-Kramer multiple comparisons test (*P < 0.05; **P < 0.01).
by guest on April 26, 2020
http://stm.sciencem
ag.org/D
ownloaded from

Liu et al., Sci. Transl. Med. 11, eaat8406 (2019) 24 April 2019
S C I E N C E T R A N S L A T I O N A L M E D I C I N E | R E S E A R C H A R T I C L E
5 of 14
A
Normoxia
Nor
mox
ia
CH
CH
Nor
mox
iaCH
Normoxia CH
Normoxia CHC
B
D
E F
Normoxia CH Normoxia CHPerc
enta
ge o
f met
hyla
tion
in
GC
box
Perc
enta
ge o
f met
hyla
tion
i
n G
C bo
x
Perc
enta
ge o
f met
hyla
tion
in G
C bo
x
Perc
enta
ge o
f met
hyla
tion
in G
C bo
x
0.0
0.1
0.2
0.3
0.4
0.5
0.0
0.2
0.4
0.6
0.8
***
CH rats CCHD patients High-altitude patients
CIRBP
Actin
DAPI CIRBP Actinin Merge DAPI CIRBP Actinin Merge
0.0
0.5
1.0
1.5
2.0
2.5
0.0
0.2
0.4
0.6
0.8
1.0
TET
act
ivity
(ng
min
–1 m
g–1)
** ******
** *
** **
mRN
A e
xpre
ssio
n
NormoxiaCH
NormoxiaCH
NormoxiaCH
NormoxiaCH
Prot
ein
expr
essi
on
% In
put
% In
put
% In
put
0
10
20
30
40
50
0
5
10
15
Nuc
lear
CIR
BP
(rel
ativ
e to
act
in)
(rel
ativ
e to
GA
PDH
)
Nuc
lear
SP1
Cyto
plas
mic
CIR
BP
0
20
40
60
80
100
Normoxia CH
**
0.0
0.2
0.4
0.6
0.8
1.0
Normoxia CH
Normoxia CH
**
0.0
0.2
0.4
0.6
0.8
1.0
G H
JI
LK
Nuclear extract
Nuclear extractSp1 Ab
IgG
150× cold competitorGC box probe
GC box probeMethylated GC box probe
1 2 3 4 5 6 7 8 9 1011 1 2 3 4 5 6 7 8 9101112
+–––
–+–––
+
++– – – + + + – – – + + +
+ + – – – + + + – – –+ + + + + + + + + + +
+–––
++–––
+
–––
+
–––
+
–––
++ + + +
+––
+++
–––
+–+–
++––+
+Normoxia
Normoxia
CH
CH
Supershift
Free probes Free probes
Shift banding Shift banding
Normoxia CH Normoxia CH
SP1
* ****
**
****
**
*
*
SP1
expr
essi
on(r
elat
ive
to h
isto
ne H
3)
SP1
exp
ress
ion
(rel
ativ
e to
act
in)
Nor
mox
iaCH
Nor
mox
iaCH
DAPI SP1 Actinin Merge DAPI SP1 Actinin Merge
IgG SP10
10
20
30
40
50
Histone H3
0
50
100
150
IgG 5mC 5hmC
**** **
**
SP1
Actin
NC
NC
SiSp1
SiSp10.0
0.5
1.0
1.5
IgG 5mC 5hmC0
20
40
60
NCSiSp1
NS
NS
4/40 12/40
0
1
2
0
1
237°C 15°C
37°C 15°C 15°C37°C 37°C 15°C
37°C/Normoxia 15°C/Normoxia15°C/CH37°C/CH
37°C/Normoxia
15°C/Normoxia37°C/CH
15°C/CH
37°C 15°C
37°C 15°C 15°C37°C37°C 15°C
37°C/Normoxia
15°C/Normoxia37°C/CH
15°C/CH
Fig. 3. CH-induced hypermethylation of the GC box inhibits SP1 binding to the Cirbp promoter region at baseline and after hypothermia. (A) Western blotting and real-time quantitative PCR (qPCR) of CIRBP expression in neonatal rat cardiac myocytes cultured under normoxic or chronic hypoxic conditions with or without hypo-thermic exposure. GAPDH, glyceraldehyde-3-phosphate dehydrogenase. (B) Immunofluorescence staining of CIRBP in cells in (A) (original magnification, ×800). Scale bars, 25 m. (C) DNA extracted from normoxia- and hypoxia-cultured cardiomyocytes was subjected to bisulfite sequencing PCR combined with TA cloning. Each row of circles indicates the sequence of an individual clone. The open circle represents an unmethylated CpG site, and the shaded circle represents a methylated CpG site. The arrow indicates a CpG site in the GC box. (D) DNA was extracted from the cardiac tissues of CH rats (n = 6 per group), patients with CCHD (n = 8 per group), and high- altitude patients (n = 8 per group) and subjected to bisulfite sequencing PCR combined with TA cloning to assess the methylation status of the CpG site in the GC box. (E) TET activity quantification performed in cells described in (A). (F) The enrichment of 5hmC and 5mC in the GC box was detected by MeDIP-qPCR and hMeDIP-qPCR in cells described in (A). MeDIP, methylated DNA immunoprecipitation; hMeDIP, hydroxymethylated DNA immunoprecipitation. (G) EMSA analysis of nuclear extracts using a 5′-biotin–labeled GC box probe. Ab, antibody; IgG, immunoglobulin G. (H) EMSA analysis of nuclear extracts using unmethylated or methylated probes. (I) Western blotting and quantitation of nuclear abundance of SP1 in cells described in (A). (J) Immunofluorescence staining of SP1 in cells described in (A). (K) SP1 binding was determined by ChIP assays. (L) SP1 protein was knocked down in cardiomyocytes, and the enrichment of 5hmC and 5mC in the GC box was detected by MeDIP-qPCR and hMeDIP-qPCR.The data shown are the mean ± SD from three independent experiments. Differences between two groups were examined by a Student’s t test and one-way ANOVA, followed by Tukey-Kramer multiple comparisons test were used to evaluate the differences among four groups (*P < 0.05; **P < 0.01). NS, not significant.
by guest on April 26, 2020
http://stm.sciencem
ag.org/D
ownloaded from

Liu et al., Sci. Transl. Med. 11, eaat8406 (2019) 24 April 2019
S C I E N C E T R A N S L A T I O N A L M E D I C I N E | R E S E A R C H A R T I C L E
6 of 14
were located in the gene-body region, and one CpG site (no. 22) was located in a GC box element (5′-GGGCGGG-3′) in the promoter re-gion of Cirbp and showed significantly increased methylation (fig. S7, A and C). Considering that hypermethylation in the promoter region is well known to silence genes and GC box is an important transcrip-tional regulatory element (16, 17), we further verified the hypermeth-ylation status of GC box in hypoxia- cultured cardiomyocytes by using bisulfite sequencing polymerase chain reaction (PCR) combined with TA cloning (~30% versus ~10% in normoxic group; P < 0.05; Fig. 3C). Similar findings were obtained in CH rats (~48% versus ~16% in normoxic group; P < 0.01), patients with CCHD (~77% versus ~32% in normoxic group; P < 0.01), and high altitude–dwelling patients (~64% versus ~32% in normoxic group; P < 0.01) (Fig. 3D).
We next measured TET activity in normoxia- and hypoxia-cultured rat cardiomyocytes. Compared with the normoxia cultures, TET activity was significantly decreased in hypoxia-cultured cardio-myocytes (0.72 ng min–1 mg–1 versus 0.23 ng min–1 mg–1; P < 0.05) (Fig. 3E). When we compared the contents of 5mC and 5hmC in the GC box between two groups (Fig. 3F), the results showed that 5mC was significantly increased (~2.17 fold; P < 0.01) and 5hmc was signifi-cantly decreased (~0.17 fold; P < 0.01) in hypoxia-cultured cardio-myocyte, which indicates lower TET activity. Moreover, the 5mC and 5hmC contents in both groups were not affected by hypothermia (Fig. 3F).
Methylation inhibits SP1 binding to the GC box in the promoter region of CirbpTo identify the proteins that bind to the GC box and regulate Cirbp expression, we analyzed the promoter region (−1 to −2000) with the cis-element cluster finder software (https://zlab.bu.edu/~mfrith/cister.shtml), which predicted that transcription factor (TF) SP1 binds the GC box. Therefore, we performed electrophoretic mobility shift assay (EMSA) to assess the binding of cellular SP1 to the GC box se-quence (Fig. 3G). The presence of the high–molecular weight shift bands shows that the nuclear protein extracts from both normoxia- and hypoxia-cultured cardiomyocytes contained proteins that bound to the 5′-biotin–labeled GC box probe. Binding could be competed out by the addition of 150× cold competitor, indicating that the interaction was specific. The addition of an SP1-specific antibody produced a super-shift of the band, demonstrating that SP1 could bind the GC box in vitro. To determine whether the methylation of the GC box could inhibit SP1 binding, EMSA was performed using a methylated GC box probe. Binding was significantly decreased in normoxia- and hypoxia- cultured cardiomyocytes, which indicated that SP1 binding was inhibited by methylation (Fig. 3H). Western blot analysis revealed that hypo-thermia caused a significant increase in nuclear abundance of SP1 in the normoxia-cultured (P < 0.05) and hypoxia-cultured cardio-myocytes (P < 0.05) (Fig. 3I). There was no significant difference in SP1 abundance between normoxia- and hypoxia-cultured cardio-myocytes at baseline or after hypothermia, indicating that CH had no influence on SP1 expression. Immunofluorescence also showed that more SP1 localized in the nucleus of normoxia- and hypoxia- cultured cardiomyocytes after they were exposed to 15°C (Fig. 3J).
To further clarify the relationship between the SP1 binding and hypermethylation in the GC box, we performed chromatin immu-noprecipitation (ChIP). Hypoxia caused a significant decrease in SP1 binding to the GC box at 37°C (Fig. 3K). Moreover, in the pres-ence of hypothermia, SP1 binding to the GC box was significantly increased in the normoxia-cultured cardiomyocytes, whereas these
changes were not observed in the hypoxia-cultured cardiomyocytes after hypothermia. We tested the contents of 5mC and 5hmC in GC box while SP1 was silenced by RNA interference, which revealed that SP1 protein expression in cardiomyocyte was knocked down by small interfering RNA to ~0.2 fold, and both the 5mC and 5hmC remained unchanged (P = 0.46 and 0.82) (Fig. 3L). These results showed that CH caused failure of SP1 binding to the GC box in cardiomyocytes and that decreased SP1 binding to GC box would not lead to hypermethylation in GC box.
To determine whether the role of GC box methylation in the down- regulation of CIRBP expression was causative, we first treated hypoxia- cultured cardiomyocytes with increasing concentrations of the DNA methylation inhibitor 5-aza-2′-deoxycytodine (5-Aza). 5-Aza treatment inhibited the CH-induced decrease in Cirbp mRNA in a concentration- dependent manner (fig. S8A). The inhibition of hypoxia- induced methylation of the GC box by 5-Aza (30 mol/liter) was demonstrated using bisulfite sequencing PCR combined with TA cloning (7.5% versus 30.0%; P < 0.05; fig. S8B). After application of 5-Aza (30 mol/liter) in hypoxia-cultured cardiomyocytes, SP1 binding to the GC box was significantly increased before and after cooling (fig. S8C). As a result, CIRBP protein expression and its response to hypothermia were re-stored (fig. S8D). Collectively, these results showed that hypoxia- induced methylation of the GC box inhibited SP1 binding to the Cirbp promoter regions, leading to decreased CIRBP expression at baseline and after hypothermia.
Hypothermic cardioprotection is attenuated in Cirbp−/− rats but enhanced in Cirbp-transgenic ratsTo explore the role of CIRBP in hypothermic cardioprotection during CPB, we generated Cirbp-knockout (Cirbp−/−) and Cirbp-transgenic (Cirbp-Tg) rats (fig. S9). At baseline assessment, Cirbp−/− and Cirbp- Tg rats have no developmental defects and no spontaneous abnormal-ities in terms of animal physiology. Echocardiographic and his-tological analysis revealed no abnormalities in cardiac function or pathological changes in Cirbp−/− and Cirbp-Tg rats (fig. S10). Western blot analysis showed that CIRBP was not expressed in cardiac tissues from Cirbp−/− rats but was overexpressed in Cirbp-Tg rats (Fig. 4A). Compared with wild-type rats, Cirbp−/− rats had decreased LVDP (P < 0.05), max dp/dt (P < 0.05), and min dp/dt (P < 0.05) values and increased LVEDP (P < 0.05) and Tau values (P < 0.01), indicat-ing that Cirbp−/− rats suffered from more severe impairment of sys-tolic and diastolic cardiac function after CPB (Fig. 4, B and C). However, Cirbp-Tg rats had elevated LVDP (P < 0.01), max dp/dt (P < 0.05), and min dp/dt values (P < 0.01) and reduced LVEDP values (P < 0.05), indicating that the overexpression of CIRBP attenuated impairment of systolic and diastolic cardiac function after CPB (Fig. 4, B and C). Postoperative serum concentrations of myocardial enzyme were significantly higher in Cirbp−/− rats than wild-type rats, but they were decreased in Cirbp-Tg rats (Fig. 4D), indicating differential cardiac injury profiles between these rats.
TUNEL staining (Fig. 4, E and F) showed more apoptotic cardiomy-ocytes in Cirbp−/− rats than wild-type rats (60.3% versus 25.7%; P < 0.01). By contrast, the number of TUNEL-stained cardiomyocytes in Cirbp-Tg rats was less than wild-type rats (12.3% versus 25.7%; P < 0.05). H&E and PTAH staining showed myocardial contraction band necrosis in myocardial fibers accompanied by cardiac sarcolemmal rupture and loss of cross-striations in Cirbp−/− rats (Fig. 4G). However, these pathological changes were decreased in wild-type rats and were minimally observed in Cirbp-Tg rats. In addition, the ratio of cleaved to full-length PARP
by guest on April 26, 2020
http://stm.sciencem
ag.org/D
ownloaded from

Liu et al., Sci. Transl. Med. 11, eaat8406 (2019) 24 April 2019
S C I E N C E T R A N S L A T I O N A L M E D I C I N E | R E S E A R C H A R T I C L E
7 of 14
C D**
* * *
WT KO TG WT KO TG WT KO TG
***
WT KO TG0
50
100
150*
WT KO TG0
200
400
600
800 **
WT KO TG0
2000
4000
6000 **
WT KO TG–1
0
12
34
5 ***
WT KO TG0
1000
2000
3000
4000
200
250
300
350
0
500
1000
1500
2000**
WT KO TG0
5
10
15
0
5
10
15
20
25
A
CIRBP
Actin
KOWT TG
KOWT TG0.0
0.5
1.0
1.5
2.0 ***
CIRB
P pr
otei
n ex
pres
sion
(rel
ativ
e to
act
in)
B
LVP
(mm
Hg)
LVP
(mm
Hg)
LVP
(mm
Hg)
01020304050
WT0
10203040
020406080
KO TG
LVD
P (m
mH
g)
HR
(bpm
)
Max
dp/
dt (m
mH
g/s)
LVED
P (m
mH
g)
Tau
(s)
Min
dp/
dt (m
mH
g/s)
DAPI
WT
KO
TG
TUNEL Actinin MergeE F
HG
***
WT KO TG
H&E
WT KO TG
PTAH
WT KO TG
Cleaved PARP
PARP
Cleaved caspase-3
Caspase-3
ActinWT KO TG
0
1
2
3
4
Clea
ved
PARP
/PA
RP
Clea
ved
casp
ase-
3/ca
spas
e-3
** *****
WT KO TG0
1
2
3
4
5
WGA
0
20
40
60
80
Apo
ptos
is in
dex
(%)
0.2 s 0.2 s 0.2 s
CK-M
B (p
g/m
l)
cTnI
(ng/
ml)
cTnT
(pg/
ml)
Fig. 4. Hypothermic cardioprotection was attenuated in Cirbp−/− rats but enhanced in Cirbp-Tg rats. (A) Cardiac CIRBP expression in the wild-type (WT), Cirbp−/−, and Cirbp-Tg (TG) rats assessed by Western blotting. KO, knockout. (B) Representative traces of postoperative LVP from six independent experiments in the wild-type, Cirbp−/−, and Cirbp-Tg rats. Scale bars, 0.2 s. (C) Cardiac function parameters evaluated by cardiac catheterization (n = 6 per group). (D) Serum cardiac enzymes after CPB in three groups (n = 6 per group). (E) TUNEL staining of cardiomyocytes from Cirbp−/−, Cirbp-Tg, and wild-type rats after CPB (original magnification, ×400). Scale bars, 50 m. (F) The apoptotic index was measured by counting puncta of TUNEL signal in 100 randomly selected actinin-positive cells (n = 3 in each group). (G) H&E and PTAH staining showing pathological changes in cardiomyocytes after CPB in three groups. The arrows indicate myocardial contraction band necrosis (original magnification, ×400 in H&E and ×1000 in PTAH). Scale bars, 50 m in H&E and 20 m in PTAH. (H) Western blot analysis of cleaved PARP, PARP, cleaved caspase-3, and caspase-3 in cardiac tissues from three groups after CPB. The graph shows the ratio of cleaved PARP to PARP and cleaved caspase-3 to caspase-3. The data shown are the mean ± SD, and statistical significance was analyzed by one-way ANOVA, followed by Tukey-Kramer multiple comparisons test (*P < 0.05; **P < 0.01).
by guest on April 26, 2020
http://stm.sciencem
ag.org/D
ownloaded from

Liu et al., Sci. Transl. Med. 11, eaat8406 (2019) 24 April 2019
S C I E N C E T R A N S L A T I O N A L M E D I C I N E | R E S E A R C H A R T I C L E
8 of 14
and cleaved to full-length caspase-3 were significantly increased in Cirbp−/− rats (both P < 0.01) and significantly decreased in Cirbp-Tg rats (P < 0.05 and P < 0.01, respectively), when compared with that in the wild-type rats (Fig. 4H), indicating that apoptosis signaling pathways were significantly activated in Cirbp-null cardiomyocytes during CPB.
To investigate the compound effect of hypoxic status and Cirbp expression in cardioprotection, we housed wild-type, Cirbp−/−, and Cirbp-Tg rats in 10% O2 for 4 weeks and then performed CPB experi-ments. The results showed that Cirbp knockout in CH rats further attenuates hypothermic cardioprotection during CPB, as demonstrated by more severe cardiac injuries, worse cardiac functions, and higher rates of cardiac apoptosis after CPB, whereas overexpression of Cirbp in CH rats reverses their impaired cardioprotective effects (fig. S11).
Ubiquinone biosynthesis is down-regulated in Cirbp−/− rats and impairs ATP production during cardiac arrestTo explain why hypothermic cardioprotection was impaired in Cirbp−/− rats during CPB, we collected cardiac tissues from Cirbp−/− and wild-type rats during CPB before restarting the heart and ana-lyzed their proteomes. The analysis showed that 18 proteins were significantly decreased (fold change < 0.83; P < 0.05), and 26 pro-teins were significantly increased in Cirbp−/− rats (fold change > 1.20; P < 0.05) relative to wild-type rats (table S4 and Fig. 5A). Pathway analysis revealed that the ubiquinone [coenzyme Q10 (CoQ10)] bio-synthetic process was significantly perturbed in Cirbp−/− rats, in-volving decreased expression of coenzyme Q3 methyltransferase (COQ3), ubiquinone biosynthesis monooxygenase COQ6 (COQ6), ubiquinone biosynthesis protein COQ7 (COQ7), and ubiquinone biosynthesis protein COQ9 (COQ9) (Fig. 5, B and C). Western blot analysis showed that cardiac COQ6 and COQ9 expression were sig-nificantly decreased during CPB in Cirbp−/− rats and were signifi-cantly increased in Cirbp-Tg rats relative to wild-type rats (Fig. 5D), whereas COQ3 and COQ7 expression remained unchanged. Further-more, Coq6 and Coq9 mRNA expression were comparable in three groups (Fig. 5E), indicating that CIRBP posttranscriptionally modu-lated the expression of Coq6 and Coq9. Last, concentrations of CoQ10 were measured using ultra performance liquid chromatography– multiple reaction monitoring–tandem mass spectrometry (UPLC-MRM- MS/MS) analysis. Cardiac CoQ10 contents during CPB were significantly decreased in Cirbp−/− rats (P < 0.05) but were signifi-cantly increased in Cirbp-Tg rats relative to wild-type rats (P < 0.01) (Fig. 5F). Because CoQ10 can scavenge reactive oxygen species and promote ATP synthesis, we assessed cardiac oxidative stress changes and ATP concentrations (Fig. 5G). Compared with wild-type rats, cardiac tissue from Cirbp−/− rats had decreased ATP concentrations (P < 0.01) and increased cardiac contents of 4-HNE (P < 0.01), MDA (P < 0.01), and carbonyl protein (P < 0.01), indicating impaired ATP production and severe cardiac oxidative stress during CPB in this group. By contrast, relative to wild-type rats, Cirbp-Tg rats had in-creased ATP concentrations (P < 0.01) and decreased contents of 4-HNE (P < 0.05), MDA (P < 0.05), and carbonyl protein (P < 0.05), indicating that the overexpression of CIRBP attenuated cardiac oxidative stress and promoted ATP production during CPB.
To directly demonstrate that COQ6 and COQ9 are major down-stream effectors of CIRBP in CoQ10 biosynthesis under hypother-mia during CPB, we used AAV9–short hairpin RNA to inhibit cardiac expression of COQ6 or COQ9 in the wild-type rats. After CPB, cardiac- specific knockdown of COQ6 or COQ9 led to de-creased cardiac function (fig. S12, C and D), decreased ATP con-
centration, increased oxidative stress, impaired CoQ10 biosynthesis, and increased postoperative myocardial injuries (fig. S12, E to J). Furthermore, we used AAV9 to overexpress cardiac expression of COQ6 or COQ9 in Cirbp−/− rats (fig. S13, A and B) and found that cardiac functions, CoQ10 biosynthesis, and cardiac injuries were partially rescued compared to Cirbp−/− rats injected with vehicle (fig. S13, C to J).
Last, we added CoQ10 to cardioplegic solution (6 mg/100 ml) and administered to Cirbp−/− rats during CPB. We found that the CoQ10 supplementation in Cirbp−/− rats alleviated cardiac oxidative stress and improved the cardiac ATP production during CPB, accompanied by less cardiac injury, better cardiac function, and lower rate of cardiac apoptosis after CPB (fig. S14). These results showed that the dis-turbed CIRBP-COQ6/9-CoQ10 axis is responsible for severe myo-cardial injury and postoperative cardiac dysfunction in Cirbp−/− rats.
CIRBP binds to the 3′ untranslated region of Coq6 and Coq9 mRNAs and facilitates their translationsTo test whether CIRBP is capable of binding to Coq6 and Coq9 mRNA, we used biotinylated fragments of Coq6 and Coq9 mRNA (Fig. 6A) and whole-cell lysates of neonatal rat cardiomyocytes for pull-down analysis. We found that three Coq6 fragments (full length, 3′ and 3′-b) and four Coq9 fragments (full length, 3′, 3′-c, and 3′-c2) interacted with CIRBP, whereas the other fragments did not (Fig. 6B). This was specific to CIRBP, because the negative control protein, tubulin, did not interact with any of the fragments used in the assay (Fig. 6B). These results indicated that the CIRBP binding element might be located in the regions corresponding to Coq6 3′-b and Coq9 3′-c2.
To further confirm the interaction between CIRBP and Coq6 and Coq9 mRNA, we performed an RNA immunoprecipitation assay, which showed that Coq6 and Coq9 mRNA binds to CIRBP in vivo (Fig. 6C). To test whether this interaction with CIRBP influences COQ6 and COQ9 expression, we constructed a series of pGL3- derived reporters bearing fragments (Fig. 6A) and tested their activity in cardiomyocytes from wild-type and Cirbp−/− rats. Cirbp knockout selectively decreased the activity of reporters bearing Coq6-3′, Coq6-3′b, Coq9-3′, Coq9-3′c, and Coq9-3′c2 (P < 0.01), whereas the ac-tivity of other reporters was not affected (Fig. 6D). These results suggest that binding to CIRBP enhances the expression of Coq6 and Coq9.
To further explore mechanisms underlying the posttranscrip-tional modulation of COQ6 and COQ9 by CIRBP, we assessed their mRNA contents in whole-cell lysates and polysomal fractions from wild-type and Cirbp-null cardiomyocytes. Knockout of Cirbp signifi-cantly decreased the presence of COQ6 and COQ9 mRNAs in poly-somal fractions (by ~54% for COQ6, P < 0.01; by ~62% for COQ9, P < 0.01), but did not affect mRNA contents in whole-cell lysates (Fig. 6E). These findings indicate that the assembly of COQ6 and COQ9 mRNAs in the polysome was increased by CIRBP binding. Our data showed that CIRBP binds to Coq6 and Coq9 mRNAs and posttranscriptionally modulates the expression of COQ6 and COQ9 by facilitating their translations.
Hypothermia-induced cardiac CoQ10 biosynthesis is impaired in CH rats and patients with CHAs hypothermia stimulated CIRBP expression and CIRBP enhanced cardiac CoQ10 biosynthesis, we hypothesized that hypothermia pro-moted cardiac CoQ10 biosynthesis during CPB. To test this hypothesis, we measured cardiac expression of COQ6, COQ9, and CoQ10 at baseline and during CPB in normoxic rats. Western blot analysis
by guest on April 26, 2020
http://stm.sciencem
ag.org/D
ownloaded from

Liu et al., Sci. Transl. Med. 11, eaat8406 (2019) 24 April 2019
S C I E N C E T R A N S L A T I O N A L M E D I C I N E | R E S E A R C H A R T I C L E
9 of 14
showed that COQ6 and COQ9 were significantly increased during CPB rel-ative to baseline (P < 0.01), and UPLC- MRM-MS/MS analysis showed that the cardiac concentration of CoQ10 was sig-nificantly elevated during CPB (P < 0.05) (fig. S15A). These results demonstrated that hypothermia enhanced cardiac CoQ10 biosynthesis during CPB. Because CH- induced hypermethylation of the Cirbp promoter region silenced the response of CIRBP to cold stress, we next evaluated the influence of CH on cardiac CoQ10 biosynthesis during CPB.We found that the protein expressions of COQ6 and COQ9 and the concentration of CoQ10 were significantly decreased during CPB in CH rats and patients with CH (P < 0.01 for COQ6 and COQ9 in CH rats, COQ9 in patients with CH, and CoQ10 in pa-tients with CCHD; P < 0.05 for CoQ10 in CH rats, COQ6 in patients with CH, and CoQ10 in high-altitude patients) (fig. S15B), which indicates that hypothermia- induced cardiac CoQ10 biosynthesis was impaired by CH. We also housed Cirbp-Tg rats in a hypoxic chamber for 1 month and subsequently subjected them to CPB.Compared with CH-treated wild-type rats, the CH- treated Cirbp-Tg rats showed sig-nificantly higher expression of CIRBP (P < 0.01), COQ6 (P < 0.05), and COQ9 (P < 0.01), increased concentration of CoQ10 (P < 0.05), as well as reduced car-diac oxidative stress (P < 0.01 for 4-HNE and protein carbonyl) and improved ATP production (P < 0.01), indicating that impaired hypothermic cardioprotection in CH rats could be rescued by CIRBP overexpression (fig. S15, C and D). Col-lectively, these results showed that hypo-thermia conferred cardioprotection by improving the CoQ10 biosynthesis during CPB, but this process was impaired in CH rats and patients with CH.
CoQ10-supplemented cardioplegic solution improves cardioprotection in CH ratsBecause we observed compromised car-diac CoQ10 biosynthesis during CPB in CH rats, we added CoQ10 to cardioplegic solution (6 mg/100 ml). In CH rats, the ad-dition of CoQ10 to cardioplegic solution significantly improved cardiac function, as indicated by increased LVDP, max dp/dt, and min dp/dt values (P < 0.01), and significantly decreased LVEDP (P < 0.01) and Tau values (P < 0.05) (Fig. 7A).
KO KO KO WT WT WTSlc8a1Tpm4Coq7Timm44Dynlrb1Itfg3Fhl2Rpl36aCoq9Ndufaf3Acaa2Tnni3kHspb7CxadrRps26Coq3Coq6Tfrcigg-2aigh-1aAcox3Ccbl2Fabp5Fam162aPrelpMcpt1Ces1dCtsdCapn1Serpi3nAcox1Cd38Entpd2Ca2Mrpl38A1i3Ckmt2PpifHpxTmlheHmgclAhsgAlbGstm1
−1.5−1−0.500.511.5
Coq3Prelp
Ndufaf3Tnni3k
Coq9
Hpx
Capn1
Fabp5 Ces1d
Ctsd
TfrcAlb
Cd38
Ccbl2
Coq7
IgH-1a
Tpm4
Coq6
Gstm1 IgG-2a
Rpl36a
Timm44
Tmlhe
Ppif Acox3
Acox1
Acaa2
Hmgcl
Ckmt2
Hspb7Slc8a1
Ahsg
Ca2
Rps26
–1.80
0.00
1.30
A B
C D KOWT TG
CIRBP
Actin
COQ3
COQ6
COQ7
COQ9
CIRBP COQ3 COQ6 COQ7 COQ9
WTKOTG
WTKOTG
Prot
ein
expr
essi
on (
rela
tive
to a
ctin
)
**** **
***
*
0.0
0.5
1.0
1.5
2.0
2.5
F
E
*
**
WT KO TG0
10
20
30
40
Coen
zym
e Q
10 (n
g/m
g)
mRN
A e
xpre
ssio
n(r
elat
ive
to G
APD
H)
0.0
0.5
1.0
1.5
Coq6 Coq9
G
0
5
10
15
20**
** **
**** *
**
WT KO TG
WT KO TG
WT KO TG
WT KO TG
3.5
4.0
4.5
5.0
0
5
10
15
20
25
0.0
1.0
2.0
3.0
4.0
ATP
(μm
ol/g
)
4-H
NE
(pg/
ml)
MD
A (n
mol
/ml)
Prot
ein
carb
onyl
(nm
ol/m
g)
Ubiquinone biosynthetic processResponse to nutrient
Fatty acid beta-oxidationFatty acid beta-oxidation using acyl-CoA oxidase
Response to retinoic acidCellular response to hypoxia
Positive regulation of bone resorptionFatty acid beta-oxidation using acyl-CoA dehydrogenase
Positive regulation of release of cytochrome c from mitochondriaRegulation of heart rate
Acute-phase responseLipid homeostasis
Positive regulation of B cell proliferationNegative regulation of apoptotic process
B cell receptor signaling pathwayResponse to drug
Cellular iron ion homeostasisResponse to hypoxia
Mitochondrion organizationLipid metabolic process
Cellular response to drugResponse to hormone
Osteoblast di erentiationMetabolic process
Proteolysis
Response to oxidative stressResponse to organic substance
Positive regulation of cytosolic calcium ion concentration
Oxidation-reduction process
Translation
Negative regulation of transcription, DNA templatedNegative regulation of transcription from RNA polymerase II promoter
0 1.0 2.0 3.0 4.0Counts
Fig. 5. The ubiquinone biosynthesis pathway was down-regulated in Cirbp−/− rats during CPB but was enhanced in Cirbp-Tg rats. (A) Proteomics was performed in cardiac tissues from Cirbp−/− (KO) rats and the wild type during CPB (n = 3 in each group). The heatmap illustrates 44 proteins with significant differences between two groups. (B) The proteins with significant differences were searched in the STRING database to visualize potential protein networks. Highlighted in yellow are proteins involved in ubiquinone biosynthesis. (C) Pathway enrichment analysis by gene ontology biological process in DAVID. (D) Western blotting was used to assess the expression of COQ3, COQ6, COQ7, and COQ9 in the cardiac tissues from wild-type, Cirbp−/− and Cirbp-Tg rats during CPB. (E) Real-time qPCR was used to assess the mRNA expressions of Coq6 and Coq9 in the cardiac tissues from wild-type, Cirbp−/−, and Cirbp-Tg rats during CPB (n = 6 per group). (F) CoQ10 concentrations in the cardiac tissues from different groups during CPB were measured using UPLC-MRM-MS/MS analysis (n = 6 per group). (G) Cardiac oxidative parameters and myocardial ATP concentrations during CPB were assessed in cardiac tissues from the wild-type, Cirbp−/−, and Cirbp-Tg rats (n = 6 per group). The data shown are the mean ± SD, and statistical significance was analyzed by one-way ANOVA, followed by Tukey-Kramer multiple comparisons test (*P < 0.05; **P < 0.01).
by guest on April 26, 2020
http://stm.sciencem
ag.org/D
ownloaded from

Liu et al., Sci. Transl. Med. 11, eaat8406 (2019) 24 April 2019
S C I E N C E T R A N S L A T I O N A L M E D I C I N E | R E S E A R C H A R T I C L E
10 of 14
These results showed that the significant improvements in post-operative systolic and diastolic cardiac functions were achieved by CoQ10 supplementation. However, except for LVDP values, the im-provements in other cardiac function parameters were not significant in the normoxic rats (Fig. 7A).
In CH rats, the addition of CoQ10 to cardioplegic solution signifi-cantly decreased postoperative serum myocardial enzymes CK-MB, cTnI, and cTnT (P < 0.01) (Fig. 7B) and also mitigated cardiac oxidative damage and promoted ATP production during CPB (P < 0.01 for MDA and ATP; P < 0.05 for 4-HNE and protein carbonyl) (Fig. 7C). Furthermore, the apoptosis of cardiomyocytes was significantly de-creased in CH rats, as shown by lower apoptosis index (18.5% versus 52.8%; P < 0.01) and reduced activation of apoptotic pathways. HE
and PTAH staining showed that the pathological changes of cardiac sarcolemmal rupture and myocardial contraction band necrosis were decreased after CoQ10 supplementation, indicating that the addition of CoQ10 relieved myocardial injury during CPB (Fig. 7, D to G). To-gether, our data showed that applying a cardioplegic solution sup-plemented with CoQ10 improved its cardioprotective effect in CH rats, but its effect was limited in normoxic rats.
DISCUSSIONNumerous studies have reported that patients with CH have worse myocardial reperfusion injury than normoxic patients after similar pe-riods of ischemic cardioplegic arrest (1, 18), but the precise mechanism
CIRBP
Tubulin
Coq9
Coq9
CIRBP
Tubulin
Coq9
CIRBP
Tubulin
B
A
CIRBP
Tubulin
Coq6
D
E
Coq6
CIRBP
Tubulin
Coq6 mRNA Coq9 mRNA
1 1 10 949 172926 1457 1762
5’UTR Coding sequence (CDS) 3’UTR 5’UTR Coding sequence (CDS) 3’UTR
5’UTR + CDS (1–1457) 5’UTR + CDS (1–949)
3’UTR (1457–1762) 3’UTR (949–1729)a (1421–1570) a (901–1200)
b (1551–1660) b (1181–1490)c (1651–1737) c (1471–1701)
c1 (1471–1600)c2 (1581–1701)
Full length (1–1762) Full length (1–1729)
0.0
0.5
1.0
1.5WT
Cirbp–/–
WT
Cirbp–/–
** **
0.0
0.5
1.0
1.5
**
** **
C
Coq6
Input InputPolysome Polysome
Coq90
5
10
15 IgGCIRBP
WT
Cirbp–/–
WT
Cirbp–/–
*
****
**
Luci
fera
se a
ctiv
ity (f
old
of c
ontr
ol)
Luci
fera
se a
ctiv
ity (f
old
of c
ontr
ol)
RN
P-IP
(fold
of c
ontr
ol)
0.0
0.5
1.0
1.5
Coq6
mRN
A le
vel
(fol
d of
con
trol
)
Coq9
mRN
A le
vel
(fol
d of
con
trol
)
0.0
0.5
1.0
1.5
5’ + CDS
5’ + CDS
FL
FL
Input Input
Input
3’ 3’
3’
3’-a 3’-b 3’-c
Input Input3’ 3’-a 3’-b 3’-c 3’-c 3’-c1 3’-c2
PGL3PGL3
Coq6
-5’ + CDS
Coq9
-5’ + CDS
Coq6
-3’
Coq9
-3’
Coq6
-3’a
Coq6
-3’b
Coq6
-3’c
Coq9
-3’a
Coq9
-3’b
Coq9
-3’c
Coq9
-3’c1
Coq9
-3’c2
Fig. 6. CIRBP binds to Coq6 and Coq9 mRNAs and enhances their translations. (A) Schematic representation depicting the Coq6 and Coq9 mRNA fragments used for biotin pull-down assays. 5′UTR, 5′ untranslated region; 3′ UTR, 3′ untranslated region. (B) Biotin pull-down assays using the biotinylated fragments shown in (A) to detect bound cellular CIRBP by Western blotting. Whole-cell lysate (input) and binding to tubulin were included as controls. (C) Neonatal rat cardiomyocytes were used for an ultraviolet cross-link ribonucleoprotein immunoprecipitation (RNP-IP) assay by using anti-CIRBP and IgG antibody. (D) Cardiomyocytes from neonatal wild-type and Cirbp−/− rats were transfected with a pGL3-derived reporter, bearing the fragments shown in (A); 48 hours later, firefly luciferase activity relative to Renilla luciferase activity was analyzed. (E) Whole-cell lysates and polysomal fractions were prepared from neonatal wild-type and Cirbp−/− rats. RNA isolated from the whole-cell lysates (input) and polysomal fractions were subjected to real-time qPCR analysis to assess the presence of Coq6 and Coq9 mRNAs. Data represent the mean ± SD from three inde-pendent experiments; significance was analyzed by Student’s t test (*P < 0.05; **P < 0.01).
by guest on April 26, 2020
http://stm.sciencem
ag.org/D
ownloaded from

Liu et al., Sci. Transl. Med. 11, eaat8406 (2019) 24 April 2019
S C I E N C E T R A N S L A T I O N A L M E D I C I N E | R E S E A R C H A R T I C L E
11 of 14
A B
C
D
F G
E
LVP
(mm
Hg)
LVP
(mm
Hg)
010
20
30
40
CH
LVP
(mm
Hg)
LVP
(mm
Hg)
0 01020
20
40
60
20
40
60
304050
Normoxia
CH + CoQ10
Normoxia + CoQ10
0
Norm
oxia
Nx + C
oQ10
CH +
CoQ
10
Nx + C
oQ10
CH +
CoQ
10
Nx + C
oQ10
CH +
CoQ
10
Nx + C
oQ10
CH +
CoQ
10
Nx + C
oQ10
CH +
CoQ
10
Nx + C
oQ10
CH +
CoQ
10
Norm
oxia
Norm
oxia
Norm
oxia
Norm
oxia
Norm
oxiaCH
CH
CH
CH
CH CH
CH CH
CH
CH
CH CH
CH CH
CH CH*
**
**
* *NS **
**
**
**
–1
01
2
3
4
5
0
50
100
150
0
100
200
300
400
0
1000
2000
3000
0
5
10
15
0
500
1000
1500
2000
260
280
300
320
340
360
0
500
1000
1500
2000
2500
12
14
16
18
20
22
** ** **
*
**
*
**
**
3.0
3.5
4.0
4.5
5.0
0
5
10
15
8
10
12
14
16
18
20
DAPI TUNEL Actinin Merge
CHCH
+ C
oQ10
CH
0
20
40
60
80
Apo
ptos
is in
dex
(%)
0.0
0.5
1.0
1.5
2.0
ATP
(μm
ol/g
)
LVD
P (m
mH
g)
HR
(bpm
)
Max
dp/
dt (m
mH
g/s)
LVED
P (m
mH
g)
Tau
(s)
Min
dp/
dt (m
mH
g/s)
0.2 s 0.2 s
0.2 s0.2 s
WGA
H&E PTAH
PARP
Cleaved PARP
Cleaved caspase-3
Caspase-3
Actin 0.0
0.5
1.0
1.5
Clea
ved
PARP
/PA
RP
0.0
0.5
1.0
1.5
Clea
ved
casp
ase-
3/ca
spas
e-3CH
NS
NS NS NS
CK-M
B (p
g/m
l)
cTnI
(ng/
ml)
cTnT
(pg/
ml)
4-H
NE
(pg/
ml)
MD
A (n
mol
/ml)
Prot
ein
carb
onyl
(nm
ol/m
g)
CH +
CoQ
10
CH + CoQ10
CH + CoQ10 CH + CoQ10
CH + CoQ10
CH + CoQ10 CH + CoQ10
CH + CoQ10CH + CoQ10
CH + CoQ10 CH + CoQ10 CH + CoQ10
Fig. 7. CoQ10-supplemented cardioplegic solution improves cardioprotection in CH rats more than in normoxic rats. (A) Representative traces of postoperative LVP and cardiac function parameters in CH and normoxic (Nx) rats after CoQ10 supplementation (n = 6 per group). Scale bars, 0.2 s. (B) Postoperative serum cardiac enzymes in CH rats after CoQ10 supplementation (n = 6 per group). (C) Cardiac oxidative parameters and myocardial ATP concentrations during CPB were assessed in cardiac tissues from CH rats after CoQ10 supplementation (n = 6 per group). (D) TUNEL staining of cardiomyocytes after CPB in CH rats with or without the addition of CoQ10 (original magnification, ×400). Scale bars, 50 m. (E) The apoptotic index was measured by counting puncta of TUNEL signal in 100 randomly selected actinin-positive cells (n = 4 in each group). (F) H&E and PTAH staining showing pathological changes in cardiomyocytes after CPB in CH rats with or without the addition of CoQ10 (original magnification, ×400 in H&E and ×1000 in PTAH). Scale bars, 50 m in H&E and 20 m in PTAH. (G) Western blot analysis of cleaved PARP, PARP, cleaved caspase-3, and caspase-3 in cardiac tissues from CH rats with or without the addition of CoQ10 after CPB. The data shown are the mean ± SD, and statistical significance was analyzed by Student’s t test (*P < 0.05; **P < 0.01).
by guest on April 26, 2020
http://stm.sciencem
ag.org/D
ownloaded from

Liu et al., Sci. Transl. Med. 11, eaat8406 (2019) 24 April 2019
S C I E N C E T R A N S L A T I O N A L M E D I C I N E | R E S E A R C H A R T I C L E
12 of 14
was unclear. Here, we identified a mechanistic pathway that links CH with compromised hypothermic cardioprotection during CPB (fig. S16). First, we found that CH induced hypermethylation of the GC box at the Cirbp promoter region and inhibited SP1 from binding to it, resulting in depressed CIRBP expression at cold temperatures. Sec-ond, hypothermia during CPB significantly increased cardiac CIRBP expression, which bound to the Coq6 and Coq9 mRNAs and post-transcriptionally enhanced their expression. COQ6 and COQ9 play important roles in CoQ10 biosynthesis, and the cardiac concentra-tion of CoQ10 was significantly increased during CPB. In contrast, CIRBP failed to respond to cold stress in chronically hypoxic myo-cardium, leading to decreased concentration of CoQ10 and impaired cardioprotective effects of hypothermia during CPB.
DNA methylation is a well-known epigenetic modification that regulates gene expression by altering the accessibility of DNA to TFs (16). Emerging evidence suggests that a number of genes are hyper-methylated under hypoxic conditions by reducing TET activity, given that the oxidation of 5mC depends on oxygen supply (12, 19). In our study, we found that CH inhibits TET activity and causes genome-wide hypermethylation in chronically hypoxic myocardium. Consequently, the promoter region of Cirbp is hypermethylated, and its expression is depressed. A previous study reported that Cirbp was up-regulated in cancer cells cultured under hypoxic condi-tion for 24 hours (10), but we found that Cirbp expression was depressed in cardiomyocytes cultured under hypoxic condition for 72 hours. It is possible that the proliferative state of tumor cells used in these studies may affect their maintenance of DNA methylation pattern (20). Furthermore, to mimic the chronic hypoxic condition, in vitro culture should exceed 48 hours under hypoxic condition (21); therefore, 24 hours might not be sufficient to establish a chronically hypoxic cell model or to observe significant epigenetic changes in Cirbp. However, after prolonged exposure to hypoxia up to 72 hours, the GC box at the Cirbp promoter region was significantly hyper-methylated. Previous studies have reported that the GC box element is a binding site for SP1, and its high methylation status interferes with SP1 binding (17). In our study, we found that a methylated GC box inhibited SP1 from binding to the Cirbp promoter region, which influenced its transcriptional activity under hypoxic conditions. Al-though nuclear SP1 protein abundance was significantly increased during hypothermia, this increase was relatively ineffective in chron-ically hypoxic cardiomyocytes, because increased methylation of the GC box caused an even more significant decrease in SP1 binding to the Cirbp promoter region. This is consistent with a previous study showing that DNA methylation inhibited TF binding even when the expression of TF is significantly increased (13). Consequently, CH depressed the expression of CIRBP at baseline and inhibited its re-sponse to cold stress.
CIRBP belongs to the family of cold shock proteins that are activated in response to cold stress (7). In our study, the protein expression of CIRBP was significantly elevated within a short time after cardiac cooling during CPB, indicating that CIRBP might act as an acute-phase protein in cardiomyocytes under cold stress. We developed Cirbp−/− and Cirbp-Tg rats and demonstrated the pivotal role of CIRBP in hypothermic cardioprotection during CPB. Cardiac pro-teomics showed that COQ6 and COQ9 were significantly decreased in Cirbp−/− rats in comparison with the wild-type. CIRBP is an RNA binding protein and has been reported to modulate gene expression posttranscriptionally (22). Our study showed that CIRBP can enhance COQ6 and COQ9 expression at the posttranscriptional level by
binding to their mRNAs and facilitating translation. COQ6 is an evolutionarily conserved monooxygenase that is responsible for C5- hydroxylation of the quinone ring in CoQ10, and COQ9 controls the deamination step in CoQ10 biosynthesis. Although previous studies have demonstrated that mutations in human COQ6 and COQ9 genes can lead to primary CoQ10 deficiency (23), the relationship between COQ6/9 and cardiac homeostasis has not been reported. Here, on the basis of evidence from in vivo gene manipulation experiment, we found that cardiac-specific COQ6/9-knockdown rats suffered se-vere myocardial injury during CPB and overexpression of COQ6/9 in Cirbp−/− rats could attenuate the myocardial injury, which sug-gests that COQ6 and COQ9 are important downstream regulators in CIRBP- mediated hypothermic cardioprotection.
Despite the progress in understanding the CoQ10 biosynthesis pathway, there are still very few data regarding its regulation. The phosphorylation and dephosphorylation status of synthetic enzymes could regulate the CoQ10 biosynthesis in bacteria and yeast (24). How-ever, it has not been verified in vertebrates and the information on which proteins could regulate CoQ10 biosynthetic enzymes is limited. In this study, we reported that CIRBP is a regulator of the CoQ10 biosynthesis pathway. It could enhance cardiac CoQ10 biosynthesis under cold stress through up-regulation of COQ6 and COQ9, two critical enzymes for CoQ10 maturation. Furthermore, it might be a potential therapeutic target to increase the contents of endogenous CoQ10 in patients with primary CoQ10 deficiency.
As an essential component of the electron transport chain and a potent antioxidant, CoQ10 can protect cells against oxidative stress and promote ATP production (23). Cells with high energy demands, such as cardiomyocytes, are extremely sensitive to CoQ10 deficiency (25). A previous study showed that the depletion of cardiac CoQ10 could lead to severe oxidative damage in myocardium, leading to myocardial infarction and heart failure (26). In our study, we found that CoQ10 biosynthesis was significantly decreased during CPB in CH rats, leading to reduced ATP content and increased oxidative stress in cardiomyocytes, and this finding was further verified in patients with CH. This strongly suggested that impaired CoQ10 bio-synthesis under hypothermia is responsible for compromised cardio-protection in chronically hypoxic myocardium. Some studies have reported that CoQ10 supplementation could exert cardioprotective effects during CPB in patients undergoing open-heart surgery (27). However, other studies failed to demonstrate its cardioprotective effects (28), so it has not been widely used in clinical practice. These contradictory results may be attributed to differing hypoxic states of the patients, because we found that the addition of CoQ10 to cardio-plegic solution significantly improved postoperative cardiac function in CH rats rather than normoxic rats. It is possible that cold-induced CoQ10 biosynthesis is impaired in CH rats but is relatively normal in normoxic rats, making the effect of CoQ10 supplementation much more prominent in CH rats than in normoxic rats.
There are some limitations in this study. First, our studies were performed on neonatal rat cardiomyocytes, which are relatively resis-tant to hypoxia. However, we did observe that the expression of CIRBP in neonatal rat cardiomyocytes was decreased under CH, which sug-gests that the phenomenon of tolerance to hypoxia in neonatal cardiomyocytes does not alter our main finding. Furthermore, cardiac tissues from CH rats and patients with CH demonstrated similar trends of CIRBP expression, which adds another layer of evidence to our finding. Second, in addition to the effects on cardiomyocytes, CH can lead to pulmonary artery hypertension (PAH) (29), which may
by guest on April 26, 2020
http://stm.sciencem
ag.org/D
ownloaded from

Liu et al., Sci. Transl. Med. 11, eaat8406 (2019) 24 April 2019
S C I E N C E T R A N S L A T I O N A L M E D I C I N E | R E S E A R C H A R T I C L E
13 of 14
be a potential confounding factor in our study. It is commonly known that the influence of PAH on heart mainly presents with increased workload for the right ventricle. Therefore, to reduce its impact on our conclusion, the cardiac tissues used for analysis were harvested from the left ventricle of CH rats and from the right atrial appendage tis-sues in patients with CH. Moreover, CH-cultured neonatal rat car-diac myocytes are free from PAH, because these cells are exempt from the direct impact of in vivo pressure load. Together, this leads us to conclude that depressed CIRBP expression was a result of CH rather than PAH.
Our findings have strong clinical implications. They provide insight into the potential provision of individualized cardioprotec-tion strategies for patients with CH undergoing open-heart surgery. Patients with CCHD, COPD, or OSA are subject to the chronic effects of hypoxia due to the underlying pathophysiological changes involved in the respective diseases. These patients are under the chronic physiological effects of hypoxia and constitute a large pop-ulation in clinical practice. In addition, on the basis of an estimate from a United Nations Environment Programs report, 12% of the human population resides in mountainous regions with sustained exposure to hypoxic environment (19). CH-induced hypermethyla-tion of Cirbp and its failure to respond to cold stress were also demonstrated in myocardial specimen harvested from high-altitude patients in our study. Overall, for these patients with CH, applying a cardioplegic solution supplemented with CoQ10 during CPB to compensate the epigenetic modification of Cirbp may translate into better perioperative cardioprotection and less myocardial injury. Moreover, therapeutic hypothermia is widely applied in various clinical settings, including organ preservation during acute cardiac arrest and brain protection in neurosurgery. For patients with CH treated in these fields, the protective effects of hypothermia may also be compromised. Our findings may extend to these fields and could potentially improve hypothermic protection by supplementa-tion with CoQ10 in these specific populations.
MATERIALS AND METHODSStudy designThe objective of this study was to explore the underlying mechanism of impaired hypothermic cardioprotection during CPB among patients with CH and investigate the translational values. Using rodent mod-els of CH and animal models of CPB, as well as proteomic analysis, we sought to identify the most relevant protein in this process. Fur-thermore, we used genetically modified rats, in vivo genetic manipu-lation by AAV9, and in vitro cell experiments to validate the central role of the identified protein and its downstream effectors. For the CH rat model, rats were randomized to the experimental or control groups. In the rat model of CPB, animals were allocated to different groups based on their hypoxic states and genotypes. The sample size in animal studies was determined on the basis of previous study reports and our past experience using this animal model. All in vitro experiments were conducted three times. Blinding approaches were used in histopathological analyses. Primer details can be found in tables S5 to S7; primary data are reported in data file S1. All animal protocols were approved by the Fuwai Hospital Animal Care and Use Committee. All human studies conform to the principles outlined in the Declaration of Helsinki and was approved by Fuwai Hospital Research Ethics Committee. Informed consent was obtained from all patients or their families.
StatisticsStatistical analysis was performed using SPSS 16.0 software (SPSS Inc.). Normality of distributions was tested by Shapiro-Wilk test. Differences between two groups were examined by a Student’s t test, and one-way ANOVA followed by Tukey-Kramer multiple compari-sons test, was used to evaluate the differences among three or more groups. All P values were two sided, and P < 0.05 was considered statistically significant. All data are expressed as the mean ± SD.
SUPPLEMENTARY MATERIALSstm.sciencemag.org/cgi/content/full/11/489/eaat8406/DC1Materials and MethodsFig. S1. Cardiac function of CH and normoxic rats at baseline.Fig. S2. Schematic diagram of rat CPB model.Fig. S3. CIRBP expression and postoperative cardiac function in male and female CH rats.Fig. S4. Circos plot showing global methylation patterns and locations of DMRs in different chromosomes.Fig. S5. Genome-wide DNA methylation analysis of cardiac tissues from CH rats.Fig. S6. Transcriptomic analysis of cardiac tissues from CH rats.Fig. S7. The methylation degree of CpG island in Cirbp was evaluated by high-throughput bisulfite sequencing.Fig. S8. 5-Aza restored CIRBP expression and its response to hypothermia.Fig. S9. Generation of Cirbp−/− and Cirbp-Tg rats.Fig. S10. Baseline assessment of organ histology and cardiac function of wild-type, Cirbp−/−, and Cirbp-Tg rats.Fig. S11. Cirbp knockout in CH rats further attenuates hypothermic cardioprotection during CPB and overexpression of CIRBP reverses this process.Fig. S12. Cardiac-specific knockdown of COQ6 or COQ9 in wild-type rats attenuates hypothermic cardioprotection during CPB.Fig. S13. Cardiac-specific overexpression of COQ6 or COQ9 in Cirbp−/− rats partially reverses impaired hypothermic cardioprotection during CPB.Fig. S14. CoQ10 addition in cardioplegic solution during CPB significantly improves cardioprotection in Cirbp−/− rats.Fig. S15. CH impaired hypothermia-induced cardiac CoQ10 biosynthesis during CPB.Fig. S16. Schematic summarizing mechanism.Table S1. Arterial blood gas analysis in the animals at the end of a 4-week exposure to a normoxic or hypoxic environment.Table S2. Significantly different proteins (fold change < 0.83 or fold change > 1.2; P < 0.05) in cardiac tissues from CH rats versus normoxic rats.Table S3. Baseline characteristics of patients.Table S4. Significantly differently expressed proteins (fold change < 0.83 or fold change > 1.2; P < 0.05) in cardiac tissues from Cirbp−/− rats versus wild-type rats.Table S5. Primer sequences used in the high-throughput bisulfite sequencing.Table S6. Primer sequences used in bisulfite sequencing PCR combined with TA cloning.Table S7. Primer pairs used in the biotin pull-down assay.Data file S1. Primary data.Movie S1. Movie of rat CPB model.
REFERENCES AND NOTES 1. H. K. Najm, W. J. Wallen, M. P. Belanger, W. G. Williams, J. G. Coles, G. S. Van Arsdell,
M. D. Black, C. Boutin, C. Wittnich, Does the degree of cyanosis affect myocardial adenosine triphosphate levels and function in children undergoing surgical procedures for congenital heart disease? J. Thorac. Cardiovasc. Surg. 119, 515–524 (2000).
2. M. Nagappa, G. Ho, J. Patra, J. Wong, M. Singh, R. Kaw, D. Cheng, F. Chung, Postoperative outcomes in obstructive sleep apnea patients undergoing cardiac surgery: A systematic review and meta-analysis of comparative studies. Anesth. Analg. 125, 2030–2037 (2017).
3. A. S. Adabag, H. S. Wassif, K. Rice, S. Mithani, D. Johnson, J. Bonawitz-Conlin, H. B. Ward, E. O. McFalls, M. A. Kuskowski, R. F. Kelly, Preoperative pulmonary function and mortality after cardiac surgery. Am. Heart J. 159, 691–697 (2010).
4. Y. Pan, J. Niu, G. Qin, L. Wang, F. Zhang, E. Wang, Perioperative safety of Tibetan children with congenital heart disease undergoing cardiac surgery and anesthesia in low-altitude area. Zhong Nan Da Xue Xue Bao Yi Xue Ban 42, 1288–1292 (2017).
5. R. K. Li, D. A. Mickle, R. D. Weisel, L. C. Tumiati, G. Jackowski, T. W. Wu, W. G. Williams, Effect of oxygen tension on the anti-oxidant enzyme activities of tetralogy of Fallot ventricular myocytes. J. Mol. Cell. Cardiol. 21, 567–575 (1989).
6. A. J. Poncelet, M. van Steenberghe, S. Moniotte, T. Detaille, C. Beauloye, L. Bertrand, M. C. Nassogne, J. E. Rubay, Cardiac and neurological assessment of normothermia/warm blood cardioplegia vs hypothermia/cold crystalloid cardioplegia in pediatric cardiac
by guest on April 26, 2020
http://stm.sciencem
ag.org/D
ownloaded from

Liu et al., Sci. Transl. Med. 11, eaat8406 (2019) 24 April 2019
S C I E N C E T R A N S L A T I O N A L M E D I C I N E | R E S E A R C H A R T I C L E
14 of 14
surgery: Insight from a prospective randomized trial. Eur. J. Cardiothorac. Surg. 40, 1384–1390 (2011).
7. T. Kaneko, K. Kibayashi, Mild hypothermia facilitates the expression of cold-inducible RNA-binding protein and heat shock protein 70.1 in mouse brain. Brain Res. 1466, 128–136 (2012).
8. H.-T. Zhang, J.-H. Xue, Z.-W. Zhang, H.-B. Kong, A.-J. Liu, S.-C. Li, D.-G. Xu, Cold-inducible RNA-binding protein inhibits neuron apoptosis through the suppression of mitochondrial apoptosis. Brain Res. 1622, 474–483 (2015).
9. L. Wu, H.-l. Sun, Y. Gao, K.-l. Hui, M.-m. Xu, H. Zhong, M.-l. Duan, Therapeutic hypothermia enhances cold-inducible RNA-binding protein expression and inhibits mitochondrial apoptosis in a rat model of cardiac arrest. Mol. Neurobiol. 54, 2697–2705 (2017).
10. S. Wellmann, C. Bührer, E. Moderegger, A. Zelmer, R. Kirschner, P. Koehne, J. Fujita, K. Seeger, Oxygen-regulated expression of the RNA-binding proteins RBM3 and CIRP by a HIF-1-independent mechanism. J. Cell Sci. 117, 1785–1794 (2004).
11. X. Chen, X. Liu, B. Li, Q. Zhang, J. Wang, W. Zhang, W. Luo, J. Chen, Cold inducible RNA binding protein is involved in chronic hypoxia induced neuron apoptosis by down-regulating HIF-1 expression and regulated by microRNA-23a. Int. J. Biol. Sci. 13, 518–531 (2017).
12. B. Thienpont, J. Steinbacher, H. Zhao, F. D’Anna, A. Kuchnio, A. Ploumakis, B. Ghesquière, L. Van Dyck, B. Boeckx, L. Schoonjans, E. Hermans, F. Amant, V. N. Kristensen, K. P. Koh, M. Mazzone, M. L. Coleman, T. Carell, P. Carmeliet, D. Lambrechts, Tumour hypoxia causes DNA hypermethylation by reducing TET activity. Nature 537, 63–68 (2016).
13. A. J. Patterson, M. Chen, Q. Xue, D. Xiao, L. Zhang, Chronic prenatal hypoxia induces epigenetic programming of PKC{epsilon} gene repression in rat hearts. Circ. Res. 107, 365–373 (2010).
14. A. F. Corno, G. Milano, M. Samaja, P. Tozzi, L. K. von Segesser, Chronic hypoxia: A model for cyanotic congenital heart defects. J. Thorac. Cardiovasc. Surg. 124, 105–112 (2002).
15. M. Gardiner-Garden, M. Frommer, CpG islands in vertebrate genomes. J. Mol. Biol. 196, 261–282 (1987).
16. A. P. Feinberg, Phenotypic plasticity and the epigenetics of human disease. Nature 447, 433–440 (2007).
17. R. Araki, S. Nishida, Y. Hiraki, K. Matsumoto, T. Yabe, DNA methylation of the GC box in the promoter region mediates isolation rearing-induced suppression of srd5a1 transcription in the prefrontal cortex. Neurosci. Lett. 606, 135–139 (2015).
18. N. A. Silverman, J. Kohler, S. Levitsky, D. G. Pavel, R. B. Fang, H. Feinberg, Chronic hypoxemia depresses global ventricular function and predisposes to the depletion of high-energy phosphates during cardioplegic arrest: Implications for surgical repair of cyanotic congenital heart defects. Ann. Thorac. Surg. 37, 304–308 (1984).
19. J. Nanduri, G. L. Semenza, N. R. Prabhakar, Epigenetic changes by DNA methylation in chronic and intermittent hypoxia. Am. J. Physiol. Lung Cell. Mol. Physiol. 313, L1096–L1100 (2017).
20. C. Z. Huang, T. Yu, Q. K. Chen, DNA methylation dynamics during differentiation, proliferation, and tumorigenesis in the intestinal tract. Stem Cells Dev. 24, 2733 (2015).
21. H. S. Silverman, S.-k. Wei, M. C. P. Haigney, C. J. Ocampo, M. D. Stern, Myocyte adaptation to chronic hypoxia and development of tolerance to subsequent acute severe hypoxia. Circ. Res. 80, 699–707 (1997).
22. J. Li, D. Xie, J. Huang, F. Lv, D. Shi, Y. Liu, L. Lin, L. Geng, Y. Wu, D. Liang, Y.-H. Chen, Cold-inducible RNA-binding protein regulates cardiac repolarization by targeting transient outward potassium channels. Circ. Res. 116, 1655–1659 (2015).
23. M. J. Acosta, L. Vazquez Fonseca, M. A. Desbats, C. Cerqua, R. Zordan, E. Trevisson, L. Salviati, Coenzyme Q biosynthesis in health and disease. Biochim. Biophys. Acta 1857, 1079–1085 (2016).
24. I. González-Mariscal, E. García-Testón, S. Padilla, A. Martín-Montalvo, T. Pomares-Viciana, L. Vazquez-Fonseca, P. Gandolfo-Domínguez, C. Santos-Ocaña, Regulation of coenzyme Q biosynthesis in yeast: A new complex in the block. IUBMB Life 66, 63–70 (2014).
25. A. Kumar, H. Kaur, P. Devi, V. Mohan, Role of coenzyme Q10 (CoQ10) in cardiac disease, hypertension and Meniere-like syndrome. Pharmacol. Ther. 124, 259–268 (2009).
26. A. Khorrami, A. Garjani, S. Ghanbarzadeh, S. Andalib, Reduction of coenzyme q10 content: A possible effect of isoproterenol on heart failure and myocardial infarction in rat. Drug Res. 64, 177–181 (2014).
27. J. Tanaka, R. Tominaga, M. Yoshitoshi, K. Matsui, M. Komori, A. Sese, H. Yasui, K. Tokunaga, Coenzyme Q10: The prophylactic effect on low cardiac output following cardiac valve replacement. Ann. Thorac. Surg. 33, 145–151 (1982).
28. J. Moludi, S. Keshavarz, A. S. Tabaee, S. Safiri, R. Pakzad, Q10 supplementation effects on cardiac enzyme CK-MB and troponin in patients undergoing coronary artery bypass graft: A randomized, double-blinded, placebo-controlled clinical trial. J. Cardiovasc. Thorac. Res. 8, 1–7 (2016).
29. V. Sauzeau, M. Rolli-Derkinderen, S. Lehoux, G. Loirand, P. Pacaud, Sildenafil prevents change in RhoA expression induced by chronic hypoxia in rat pulmonary artery. Circ. Res. 93, 630–637 (2003).
Acknowledgments: We appreciate the help in specimen collections from G. Zhang and X. Hu from the Department of Cardiac Surgery, Fuwai Yunnan Cardiovascular Hospital and the help in bioinformatic analysis from W. Xiong from the Tsinghua University. Funding: The study was funded by the National Science Fund for Distinguished Young Scholars (81525002), Chinese Academy of Medical Sciences Innovation Fund for Medical Sciences (CIFMS 2017-IZM-1-00), Graduate Innovation Fund Project of Peking Union Medical College (2015-1002-02-21), and Program for Professors in Peking Union Medical College. Author contributions: H.Z. and Y. Liu conceived and designed the study. Y. Liu and Y. Li performed the animal study and analyzed the data. Q.L. and Z.S. collected the human samples. J.X. and X.Z. performed the cell study. Y. Liu drafted the manuscript. H.Z. was the principal investigator, obtained funding, and assisted with drafting of the manuscript. All authors reviewed, contributed to, and approved the final manuscript. Competing interests: The authors declare that they have no competing interests. Data and materials availability: All data associated with this study are present in the paper or the Supplementary Materials. All data generated in this study are available from the authors upon reasonable request. The sequence data have been submitted to GEO with accession no. GSE123948.
Submitted 10 April 2018Resubmitted 25 October 2018Accepted 25 March 2019Published 24 April 201910.1126/scitranslmed.aat8406
Citation: Y. Liu, J. Xing, Y. Li, Q. Luo, Z. Su, X. Zhang, H. Zhang, Chronic hypoxia–induced Cirbp hypermethylation attenuates hypothermic cardioprotection via down-regulation of ubiquinone biosynthesis. Sci. Transl. Med. 11, eaat8406 (2019).
by guest on April 26, 2020
http://stm.sciencem
ag.org/D
ownloaded from

cardioprotection via down-regulation of ubiquinone biosynthesis hypermethylation attenuates hypothermicCirbpinduced −Chronic hypoxia
Yiwei Liu, Junyue Xing, Yongnan Li, Qipeng Luo, Zhanhao Su, Xiaoling Zhang and Hao Zhang
DOI: 10.1126/scitranslmed.aat8406, eaat8406.11Sci Transl Med
chronic hypoxic conditions.withexposed to chronic hypoxia. This study helps explain why hypothermia may not be cardioprotective in patients
abrogating cardioprotective effects. Supplementing ubiquinone during CPB could restore cardioprotection in ratsregion, reducing protein expression in response to cold stress and antioxidant ubiquinone synthesis and
promoterCirbpCPB under normoxic and hypoxic conditions. Chronic hypoxia caused hypermethylation of the role of cold-inducible RNA binding protein (CIRBP), a stress-induced protein known to be neuroprotective, during
. studied theet alincur injury after cardiopulmonary bypass (CPB) performed with therapeutic hypothermia. Liu Hypothermia reduces a tissue's metabolic demand and can be cardioprotective; however, some patients
Cardioprotection loses its cool
ARTICLE TOOLS http://stm.sciencemag.org/content/11/489/eaat8406
MATERIALSSUPPLEMENTARY http://stm.sciencemag.org/content/suppl/2019/04/22/11.489.eaat8406.DC1
CONTENTRELATED
http://stm.sciencemag.org/content/scitransmed/11/494/eaat4407.fullhttp://stm.sciencemag.org/content/scitransmed/9/408/eaan0117.fullhttp://stm.sciencemag.org/content/scitransmed/10/441/eaan4886.fullhttp://stm.sciencemag.org/content/scitransmed/10/438/eaao0303.full
REFERENCES
http://stm.sciencemag.org/content/11/489/eaat8406#BIBLThis article cites 29 articles, 5 of which you can access for free
PERMISSIONS http://www.sciencemag.org/help/reprints-and-permissions
Terms of ServiceUse of this article is subject to the
registered trademark of AAAS. is aScience Translational MedicineScience, 1200 New York Avenue NW, Washington, DC 20005. The title
(ISSN 1946-6242) is published by the American Association for the Advancement ofScience Translational Medicine
of Science. No claim to original U.S. Government WorksCopyright © 2019 The Authors, some rights reserved; exclusive licensee American Association for the Advancement
by guest on April 26, 2020
http://stm.sciencem
ag.org/D
ownloaded from