by decorating lipid a with phosphoethanolamine accepted · lipopolysaccharide (lps) forms the outer...
TRANSCRIPT

PmrAC confers pmrHFIJKL-dependent EGTA and polymyxin resistance to msbB
Salmonella by decorating lipid A with phosphoethanolamine
Running title: EGTA and polymyxin resistance in msbB Salmonella
Key words: PmrA, PmrH, PmrF, PmrI, PmrJ, PmrK, PmrL, PagP, MsbB, Salmonella, EGTA,
polymyxin, phosphoethanolamine, lipid A
Sean R. Murray1+
, Robert K. Ernst2, David Bermudes
3/, Samuel I. Miller
2 and K.
Brooks Low4*
1 Department of Biology, Yale University, New Haven, CT
2 Department of Microbiology, University of Washington, Seattle, WA
3 Vion Pharmaceuticals, New Haven, CT
4 Department of Therapeutic Radiology, Yale University School of Medicine, New
Haven, CT
+ Present address: Department of Developmental Biology, Stanford, CA
/ Present address: Celator Pharmaceuticals, Vancouver, BC, Canada
* Corresponding author. Mailing address: Radiobiology Laboratories, Yale
University School of Medicine, 333 Cedar St., New Haven, CT 06520. Phone: (203)
785-2976. Fax: (203) 785-6309. E-mail: [email protected].
ACCEPTED
Copyright © 2007, American Society for Microbiology and/or the Listed Authors/Institutions. All Rights Reserved.J. Bacteriol. doi:10.1128/JB.01969-06 JB Accepts, published online ahead of print on 20 April 2007
on June 8, 2019 by guesthttp://jb.asm
.org/D
ownloaded from

ABSTRACT
Mutations in pmrA were recombined into msbB Salmonella ATCC 14028 to
determine if pmrA-regulated modifications of lipopolysaccharide could suppress msbB
growth defects. A mutation that functions to constitutively activate pmrA (pmrAC)
suppresses msbB growth defects on EGTA-containing media. Lipid A structural analysis
showed that msbB pmrAC Salmonella, as compared to msbB Salmonella, have increased
amounts of palmitate and phosphoethanolamine but no aminoarabinose addition,
suggesting that aminoarabinose is not incorporated into msbB lipid A. Surprisingly, loss-
of-function mutations in the aminoarabinose biosynthetic genes restored EGTA and
polymyxin sensitivity to msbB pmrAC Salmonella. These blocks in aminoarabinose 10
biosynthesis also prevented lipid A phosphoethanolamine incorporation and reduced the
levels of palmitate addition, indicating previously unknown roles for the aminoarabinose
biosynthetic enzymes. Lipid A structural analysis of the EGTA and polymyxin resistant
triple mutant msbB pmrAC pagP::Tn10, which contains phosphoethanolamine but no
palmitoylated lipid A, suggests that phosphoethanolamine addition is sufficient to confer
EGTA and polymyxin resistance to msbB Salmonella. Additionally, palmitoylated lipid
A was only observed in wild type Salmonella grown in the presence of salt in rich media.
Thus, we correlate EGTA resistance and polymyxin resistance with
phosphoethanolamine-decorated lipid A and demonstrate that the aminoarabinose
biosynthetic proteins play an essential role in lipid A phosphoethanolamine addition and 20
affect lipid A palmitate addition in msbB Salmonella.
ACCEPTED
on June 8, 2019 by guesthttp://jb.asm
.org/D
ownloaded from

3
INTRODUCTION
Lipopolysaccharide (LPS) forms the outer leaflet of the gram-negative bacterial outer
membrane. It consists of three molecular domains: lipid A, core oligosaccharide and O-antigen.
The endotoxic portion, lipid A, anchors LPS into the asymmetric outer membrane and is essential
for outer membrane barrier function and cell viability. LPS biosynthesis initiates on the
cytoplasmic side of the inner membrane before intermediates in LPS biosynthesis are transported
to their final destination in the outer membrane. 4-amino-4-deoxy-L-arabinose (hereafter simply
referred to as aminoarabinose) can be added to lipid A on the periplasmic side of the inner
membrane (32) and palmitate can be added to the lipid A portion of LPS once it arrives in the 30
outer membrane (1). One of the cytoplasmic lipid A biosynthetic enzymes, MsbB (=LpxM), adds
myristate to lipid IVA (see filled arrow pointing to the myristate moiety in Figure 1).
MsbB mutants are of particular interest in microbial pathogenesis and vaccine
development research because lipid A lacking myristate no longer has strong endotoxic activity
(16, 19, 27). Furthermore, msbB Salmonella enterica serovar Typhimurium have severe growth
defects in LB, galactose MacConkey, or EGTA-containing media that can be suppressed by
extragenic compensatory mutations that arise at high frequency (22). Thus, an understanding of
the different suppressors of msbB and their effect on cell growth and virulence is essential for
their application in the engineering of attenuated live-bacterial vaccines or bacterial vectors. Our
group has thus far identified two spontaneous suppressor mutations, somA (22) and the Suwwan 40
deletion (23). During our investigations we hypothesized that modification of lipid A could
suppress msbB growth defects. However, our analyses of lipid A from spontaneous msbB
suppressor strains YS1456 (msbB somA1) and YS1170 (msbB Suwwan) revealed no structural
changes in lipid A from unsuppressed msbB strains (23). In addition, SDS-PAGE analysis of
these strains showed no changes in LPS carbohydrate structure.
EDTA, a general chelator of divalent cations, has been proposed to disrupt the outer
membrane by increasing electrostatic repulsion between neighboring LPS molecules, leading to
the presence of phospholipid domains within the outer leaflet of the outer membrane and
impairing outer membrane barrier function (24). A more specific chelator of divalent cations is
ACCEPTED
on June 8, 2019 by guesthttp://jb.asm
.org/D
ownloaded from

4
EGTA, which preferentially binds calcium. msbB Salmonella are EGTA sensitive (i.e., dependent 50
on high calcium levels for growth) and many suppressor mutations confer EGTA resistant
phenotypes (22, 23). We hypothesized that the modification of the phosphate groups of lipid A
with either phosphoethanolamine or aminoarabinose would yield an EGTA resistant phenotype,
since the mutants would no longer be as dependent on [Ca2+
] because these modifications could
reduce the amount of electrostatic repulsion between neighboring lipid A molecules.
The addition of aminoarabinose and phosphoethanolamine to the lipid A portion of LPS
(Figure 1), mediated by the two-component system PmrA/B, contributes to virulence (11) and
resistance to polymyxin (35), which is a cyclic antimicrobial lipopeptide. The PmrA/B two-
component system is activated under either mildly acidic, low [Mg2+
/Ca2+
] (28), or high ferric
chloride growth conditions (37). The Salmonella aminoarabinose biosynthetic genes were 60
previously identified (9, 11). Since that time, Raetz and colleagues have elucidated the
biochemical functions of PmrH (also called ArnB) (3), PmrI (ArnA) (36), PmrF (ArnC, PbgP) (2)
and PmrK (ArnT) (33) in aminoarabinose biosynthesis. The pmrE (ugd) gene is physically
separated from the other genes in this pathway (pmrHFIJKLM) which form a transcriptional
regulon in Salmonella. Strains carrying loss-of-function mutations in pmrM, in contrast to strains
with loss-of-function mutations in pmrE and pmrHFIJKL, can still modify lipid A with
aminoarabinose (11). Thus far, the only lipid A phosphoethanolamine biosynthetic protein
identified in Salmonella is PmrC (18). pmrC lies directly in front of pmrAB in a transcriptional
regulon and PmrC is an inner membrane protein with a large periplasmic domain. The protein
responsible for adding phosphoethanolamine to the LPS core, CptA, has been recently identified 70
and is PmrA-regulated (29).
To determine whether the addition of aminoarabinose and/or phosphoethanolamine to
lipid A could suppress msbB growth defects, we recombined constitutive and loss-of-function
mutations in the PmrA/B two-component system into msbB Salmonella. As described below, a
constitutive mutation in pmrA (pmrA505) (10) results in an msbB suppressor phenotype. We also
confirm our hypothesis that the addition of phosphoethanolamine to lipid A can confer an EGTA
resistant phenotype to msbB Salmonella and demonstrate that the aminoarabinose biosynthetic
ACCEPTED
on June 8, 2019 by guesthttp://jb.asm
.org/D
ownloaded from

5
proteins are required for lipid A phosphoethanolamine incorporation, and affect palmitate addition,
in an msbB genetic background.
80
MATERIALS AND METHODS
Bacterial strains, phage and media. The bacterial strains used in this study are listed in Table
1. P22 mutant HT105/1int201 (obtained from the Salmonella Genetic Stock Center, Calgary,
Canada) was used for Salmonella transductions. Salmonella Typhimurium strains were grown on
LB-0 or MSB agar or in MSB broth. MSB media consists of LB (Luria-Bertani media) (21) with no
NaCl and supplemented with 2mM MgSO4 and 2mM CaCl2. LB-0 is LB media with no NaCl.
These media were used for the growth of strains under non-selective conditions. LB-0 agar was
used when performing genetic selections with antibiotics. Plates were solidified with 1.5% agar.
LB-0 agar and MSB broth were supplemented with chloramphenicol (15 µg/ml in broth; 25 µg/ml
in agar), ethylene glycol-bis(ß-Aminoethyl Ether)- N,N,N’,N’-tetraacetic acid (EGTA) (Sigma, St. 90
Louis, MO) (6mM or 6.5mM), streptomycin (200 µg/ml) or tetracycline (3, 5 or 20 µg/ml) as
needed. A 350 mM stock of EGTA at pH 8.0 (adjusted with NaOH) was dissolved and then
autoclaved. Antibiotics were added to LB-0 agar after cooling to 45 degrees Celsius.
MacConkey Agar Base (Difco) was used to prepare Galactose MacConkey agar.
Growth Analysis. Phenotypes of strains were confirmed by replica plating. Replica plating was
performed using the double velvet technique (17).
Preparation of Electroporation-Competent Cells. A standard protocol for making
electrocompetent cells (25) was modified as described previously (22). 100
Transduction and Transformation. Salmonella P22 transductions were carried out as
previously described (7), except that EGTA was not added to the medium. A Bio-rad Gene
Pulser was used for transformation, following the unique electroporation protocol described
previously (22).
ACCEPTED
on June 8, 2019 by guesthttp://jb.asm
.org/D
ownloaded from

6
Polymyxin survival. 10 mls of MSB broth was inoculated from a patch on a master plate with
phenotypes confirmed by replica plating. Cultures were grown in 2.5 cm diameter glass tubes to
an O.D.600 of 0.4 and held on ice. Once all cultures reached the appropriate O.D. and sat on ice
for at least 15 minutes, dilutions were made in ice-cold plastic eppendorf tubes containing MSB 110
broth. Cells were diluted so that approximately 300 cells would be present in a 500 µl aliquot.
Polymyxin B sulfate was added to media achieve a final concentration of 0.1 µg/ml (a 500 µl
volume of 2x polymyxin B sulfate in MSB broth was added to 500 µl aliquot of cells, giving a total
of 1 ml in each tube) in a plastic eppendorf tube for each strain and they were incubated for 1.5
hours in a 37˚C incubator without shaking. 500 µl from each tube was spread onto MSB agar.
Plates were incubated overnight at 37˚C and CFUs were determined.
Mass Spectrometry of lipid A: lipid A purification for MALDI-TOF analysis. Lipid A samples
were purified from at least two independent cultures and all independent cultures, from a given
strain, yielded nearly identical lipid A mass spectra. Strains were either grown in 500 ml cultures 120
of MSB broth to an O.D.600 of 0.10 or 100 ml cultures of MSB broth without MgSO4 (pH 8.0), were
grown to an O.D.600 of 1.0. (Cultures were stopped when grown to an O.D.600 of 0.1 in order to
decrease the chance of jackpots with derivatives. Jackpots arise when a suppressor mutation
that confers a growth advantage spontaneously occurs, early in the growth of a culture, and cells
with this secondary mutation overgrow the original clone, which lack the second mutation for
faster division. Later, we found out that this was unnecessary because the frequency of
suppressors is relatively constant in MSB broth between an O.D.600 of 0.1 and 1.0. Furthermore,
we detected no changes in lipid A structure in our strains grown under both conditions.) Cultures
in MSB broth lacking MgSO4 (pH 8.0) were inoculated with 1:500 dilutions of overnight cultures
grown in the same broth and 1mM EGTA was added after 40 minutes of incubation at 37°C. 130
MSB broth cultures were grown with 100 rpm of translational movement and MSB broth lacking
MgSO4 (pH 8.0) cultures were grown at 125 rpm of translational movement. LPS was purified by
the Mg2+
-ethanol precipitation method as previously described (5). Lipid A was purified by
ACCEPTED
on June 8, 2019 by guesthttp://jb.asm
.org/D
ownloaded from

7
hydrolysis in 1% SDS at pH 4.5 (4). Before being applied on a sample plate, the lyophilized lipid
A was dissolved in 20 µl of 5-chloro-2-mercaptobenzothiazole MALDI matrix in chloroform-
methanol (1:1). Negative-ion MALDI-TOF was performed as described previously (8).
Thin-layer chromatography of lipid A. Lipid A was labeled with P-33, purified, and analyzed by
thin-layer chromatography as previously described (33), except that cells were grown in MSB
broth to an O.D.600 of 0.60 instead of 1.0. 140
Gas Chromatographic Analysis of Lipopolysaccharide (LPS) Fatty Acids. Strains were
grown as described under “Mass spectrometry of lipid A: lipid A purification for MALDI-TOF
anaylsis” section of Materials and Methods. The rapid LPS purification technique for gas
chromatography was performed as previously described (27). Fatty acid components of LPS
were converted to methyl esters by methanolysis (27).
RESULTS AND DISCUSSION
pmrAC suppresses msbB growth defects. To determine if mutations in pmrA could suppress
msbB-growth defects, we transduced loss-of-function and constitutive alleles of pmrA into msbB 150
Salmonella ATCC 14028. As we hypothesized, a constitutive mutation (pmrA505, referred to as
pmrAC), but not a loss-of-function mutation, in pmrA partially suppresses EGTA sensitivity in an
msbB background (Figure 2). In contrast, it does not rescue growth on galactose-MacConkey
media.
Polymyxin resistance has previously been correlated with EDTA resistance in pmrA
mutants (34). Constitutive activation of pmrA has been shown to result in the modification of the
phosphate groups of lipid A with aminoarabinose and/or phosphoethanolamine (9, 13, 30, 39).
We propose that these lipid A structural modifications suppress msbB growth defects by
decreasing the electrostatic repulsion between neighboring phosphate groups, which would make
the cells less dependent on divalent cations like Mg2+
and Ca2+
. 160
ACCEPTED
on June 8, 2019 by guesthttp://jb.asm
.org/D
ownloaded from

8
Aminoarabinose biosynthetic genes are required for pmrAC to suppress msbB growth
defects. Salmonella’s aminoarabinose operon has previously been described (9, 11). To
determine if the aminoarabinose biosynthetic genes are necessary for the suppression of msbB
growth defects in the pmrAC strain, we moved our msbB::Ωtet marker into pmrA
C strains with non-
polar deletions in pmrH, pmrI, pmrJ, pmrK, and pmrL. As shown in Figure 3, these non-polar
deletions in pmrH, pmrI, pmrJ, pmrK, and pmrL have an unsuppressed msbB phenotype despite
the presence of the pmrAC mutation. In contrast, loss of pmrM, which is not believed to play a
role in the aminoarabinose biosynthetic pathway (11), does not alter the EGTA resistant
phenotype of the msbB pmrAC strain (Figure 3). The rpsL allele, which yields resistance to 170
streptomycin, present in some of these triple mutants does not alter the EGTA or galactose
MacConkey phenotypes (data not shown).
The pmrAC mutation confers polymyxin resistance to wild type and msbB Salmonella
ATCC 14028 grown in MSB broth. Our results (see below) demonstrate that the msbB mutation
confers polymyxin sensitivity to Salmonella ATCC 14028 grown in MSB broth, which is essential
to avoid enrichment for suppressor mutations. Another study, using LB broth, in comparison, also
reported that msbB conferred polymyxin sensitivity to Salmonella strain C5 (31). The pmrAC
mutation was isolated from a screen for polymyxin resistant Salmonella in LB medium (26). To
confirm that the pmrAC mutation also increases polymyxin resistance in msbB Salmonella ATCC 180
14028 grown in MSB medium, polymyxin survival for the various msbB strains tested are shown
in Figure 4. Our results show that the pmrAC mutation confers polymyxin resistance to msbB
Salmonella ATCC 14028 grown in MSB broth. In a wild type background, the pmrAC mutation
allows for ~73% of the cells to survive 1.5 hours exposure to 3.0 µg/ml polymyxin B sulfate in
comparison to ~1.3% survival in ATCC 14028 (not shown). In an msbB background, the pmrAC
mutation allows for ~53% of cells to survive a 1.5 hour exposure to 0.1 µg/ml polymyxin B sulfate
whereas ~0% of msbB Salmonella ATCC 14028 survive this treatment. Thus, although the msbB
mutation confers polymyxin sensitivity (31), the pmrAC allele significantly reduces this sensitivity.
ACCEPTED
on June 8, 2019 by guesthttp://jb.asm
.org/D
ownloaded from

9
Non-polar deletions in pmrHFIJKL restored both EGTA (Figure 3) and polymyxin (Figure
4) sensitivity to msbB pmrAC Salmonella ATCC 14028. Loss of pmrM, which has not been shown 190
to play a role in lipid A modification, does not affect either EGTA or polymyxin resistance.
Salt in LB broth stimulates the palmitoylation of lipid A in wild type Salmonella ATCC
14028. Previous reports of lipid A structure in wild type and pmrAC Salmonella were from
cultures grown in LB broth (9, 11, 18, 31, 32, 39). However, msbB Salmonella must be grown in
MSB broth to prevent rapid overgrowth by suppressor mutants (22). To determine if there were
any differences in lipid A structure based on growth conditions, we analyzed lipid A from strains
grown in LB, LB-no-salt, and MSB broth using negative-ion matrix-assisted laser desorption
ionization-time of flight (MALDI-TOF) mass spectrometry. The lipid A portion of LPS has been
well characterized by mass spectrometry, with published mass-to-charge ratios correlated with 200
distinct chemical structures (12, 38, 39). As shown in Figure 5A, ATCC 14028 has both
hexaacylated (m/z 1797) and heptaacylated lipid A (m/z 2036) when grown in LB broth.
However, when grown in LB-no-salt broth (Figure 5B) or MSB broth (not shown), lipid A lacks
heptaacylated lipid A (m/z 2036), showing that palmitoylation is upregulated by the presence of
1% NaCl.
Gas chromatography (Figure 5E - under C16 column) confirms that there is an increase
(~2-3-fold) in palmitate addition in both 14028 and 14028 pmrAC grown in LB broth compared to
LB-no-salt broth. Thus, we see that palmitoylation, in both 14028 and 14028 pmrAC, is stimulated
by the addition of 1% NaCl to LB-no-salt broth.
210
Myristoylation of lipid A by MsbB is necessary for decoration with aminoarabinose, but
not phosphoethanolamine. Similar investigations were carried out with ATCC 14028 pmrAC.
As shown in Figure 5C, the pmrAC mutation, in an msbB
+ genetic background, confers
aminoarabinose and phosphoethanolamine addition to lipid A. Peaks corresponding to
hexaacylated lipid A (m/z 1797) with phosphoethanolamine (m/z 1920) or aminoarabinose (m/z
1928) and heptaacylated lipid A (m/z 2036) with phosphoethanolamine (m/z 2159) or
ACCEPTED
on June 8, 2019 by guesthttp://jb.asm
.org/D
ownloaded from

10
aminoarabinose (m/z 2167) are apparent. Phosphoethanolamine addition shifts peaks m/z 123
units while aminoarabinose addition shifts peaks m/z 131 units. Figure 5D shows that the effect
of the pmrAC mutation on lipid A structure in rich media lacking salt (LB-no-salt broth). Under
these growth conditions, the pmrAC mutation confers phosphoethanolamine (m/z 1920) and 220
aminoarabinose addition (m/z 1928) to hexaacylated lipid A and very small amounts of
heptaacylated lipid A are present, either in LB-no-salt broth (Figure 5D) or MSB broth (not
shown).
To see if the pmrAC mutation has the same effects on both wild type and MsbB lipid A,
we isolated lipid A from msbB and msbB pmrAC Salmonella ATCC 14028. As seen in Figure 6A,
msbB lipid A contains a pentaacylated peak (m/z 1588) as its major lipid A species (expected
since its lipid A lacks myristate, which yields a m/z shift of 210) and contains a minor amount of
hexaacylated lipid A (m/z 1826 due to palmitate addition). In contrast to the effects of the pmrAC
mutation on wild type (Figure 5D), the pmrAC mutation confers only phosphoethanolamine but not
aminoarabinose addition to msbB lipid A (Figure 6B). In addition to pentaacylated lipid A (m/z 230
1588) decorated with phosphoethanolamine (m/z 1711), there is a large increase in palmitate
addition (m/z 1826) and some hexaacylated lipid A with phosphoethanolamine is observed (m/z
1949). Since the pmrAC mutation results in lipid A aminoarabinose addition in a wild type
(Figures 5C and 5D) but not in a msbB genetic background (Figure 6B), we conclude that
aminoarabinose is not be added to lipid A lacking the myristate residue added by MsbB in MSB
medium. A related finding was reported (31) from experiments using msbB Salmonella C5,
showing that aminoarabinose is not added to MsbB lipid A in LB broth. Perhaps the myristoyl
group added by MsbB is required for the localization or proper positioning of one of the
aminoarabinose biosynthetic enzymes, thereby conferring substrate specificity.
240
Lipid A from EGTA resistant (msbB pmrAC) strains grown in MSB broth have increased
levels of palmitate and phosphoethanolamine addition compared to EGTA sensitive
strains (msbB or msbB pmrA
C-aminoarabinose mutants). MALDI-TOF mass spectrometry
was then used to analyze overall lipid A structure in the EGTA sensitive (EGTAS) and EGTA
ACCEPTED
on June 8, 2019 by guesthttp://jb.asm
.org/D
ownloaded from

11
resistant (EGTAR) strains. As seen in Figure 6A, msbB Salmonella ATCC 14028 (EGTA
S) have a
higher pentaacylated lipid A peak (at m/z 1588) whereas msbB pmrAC Salmonella ATCC 14028
(EGTAR, Figure 7B) have a higher hexaacylated lipid A peak (at m/z 1826), resulting from the
addition of palmitate, as well as phosphoethanolamine peaks (pentaacylated lipid A with
phosphoethanolamine at m/z 1711 and hexaacylated lipid A with phosphoethanolamine at m/z
1949). Our mass spectra suggest that there are differences in terms of relative amounts of lipid A 250
species between the strains. Since mass spectrometry is qualitative, we used thin-layer
chromatography and gas chromatography to demonstrate quantitative differences between the
amounts of lipid A species. Thin-layer chromatography of lipid A (Figure 6E) suggests msbB
pmrAC Salmonella ATCC 14028 have more hexaacylated (palmitoylated) lipid A (marked by
arrow; assignment based on previously published data) (38, 39) than msbB somA (Figure 6E) or
msbB Salmonella ATCC 14028 (not shown, msbB somA and msbB Salmonella ATCC 14028
have indistinguishable lipid A TLC profiles). Gas chromatography (Figure 6F, compare C16 bars
1 and 3) shows that the pmrAC mutation results in an ~7-fold increase in lipid A palmitate
incorporation in msbB Salmonella ATCC 14028. Thus, three independent techniques indicate
that the pmrAC mutation, in an msbB background, increases lipid A palmitoylation. We also 260
challenged msbB Salmonella ATCC 14028 with 1 mM EGTA to see if its lipid A structure became
modified. As shown in Figure 6F (C16 column, 2nd
bar), EGTA challenge results in an
approximately 2-fold increase in lipid A palmitoylation in EGTAS msbB Salmonella ATCC 14028.
Mass spectrometry of lipid A from EGTA-challenged msbB Salmonella ATCC 14028 did not
suggest any additional changes (not shown). Escherichia coli responds to EDTA similarly, by
palmitoylating its lipid A molecules (14).
The aminoarabinose genes are essential for lipid A phosphoethanolamine addition and
affect palmitate addition in msbB pmrAC Salmonella ATCC 14028. Since loss of the
aminoarabinose genes conferred EGTA sensitivity to msbB pmrAC Salmonella ATCC 14028 270
(Figure 3) and we concluded that aminoarabinose can not be added to msbB lipid A (compare
Figures 5C, 5D, and 6B), we investigated lipid A structure in msbB pmrAC strains with loss-of-
ACCEPTED
on June 8, 2019 by guesthttp://jb.asm
.org/D
ownloaded from

12
function mutations in pmrH, pmrI, pmrJ, pmrK, pmrL and pmrM to learn what affect these
mutations have on lipid A structure in the msbB pmrAC genetic background. The mutations in
pmrH, pmrF, pmrI, pmrJ, pmrK, and pmrL are non-polar deletions (11).
Figure 6C shows that a non-polar deletion mutation in pmrF, which restores EGTAS to
msbB pmrAC Salmonella ATCC 14028 (Figure 3), also restores pentaacylated lipid A (peak at m/z
1588) as the dominant form and loses lipid A phosphoethanolamine incorporation (loss of peaks
at m/z 1711 and m/z 1949). This decrease in palmitoylation and loss of phosphoethanolamine
incorporation was found consistently in the triple mutants. Lipid A from msbB pmrAC strains with 280
non-polar deletions in pmrH, pmrI, pmrJ, pmrK and pmrL reproducibly produce similar mass
spectra and the rpsL mutation in these triple mutants has no obvious effects on lipid A mass
spectra (not shown). pmrM, although part of the transcriptional regulon pmrHFIJKLM, is not
believed to be part of the aminoarabinose biosynthetic pathway and has a lipid A structural profile
similar to that of msbB pmrAC (not shown). Likewise, a loss-of-function mutation in pmrM does
not make msbB pmrAC EGTA
S (Figure 3).
Similar lipid A profiles were observed for these strains grown in MSB broth lacking
MgSO4 and in strains grown in this medium challenged with EGTA after entering exponential
growth. As mentioned above, the only difference noticed in EGTA-challenged cells was a 2-fold
increase in palmitoylation. Gas chromatography confirmed that loss-of-function mutations in 290
pmrH and pmrF (Figure 6F – under C16 column) and pmrIJKL (not shown) reduce the levels of
palmitate addition to those observed in unsuppressed msbB strains, despite the presence of the
pmrAC allele in the triple mutants. These data demonstrate that the aminoarabinose genes affect
palmitate incorporation (i.e. as described above, loss of the aminoarabinose biosynthetic genes
results in decreased palmitoylation of lipid A in msbB pmrAC Salmonella ATCC 14028).
Lipid A phosphoethanolamine addition is sufficient to confer EGTA and polymyxin
resistance to msbB Salmonella ATCC 14028. Lipid A from msbB pmrAC Salmonella ATCC
14028 has two distinct molecular differences from lipid A harvested from msbB Salmonella ATCC
14028: increased palmitoylation and phosphoethanolamine addition. Either or both could be 300
ACCEPTED
on June 8, 2019 by guesthttp://jb.asm
.org/D
ownloaded from

13
responsible for the EGTA and polymyxin resistance phenotypes. In order to address this, we
transduced a pagP loss-of-function mutation into msbB pmrAC Salmonella ATCC 14028. The
msbB pmrAC pagP transductants had EGTA (not shown) and polymyxin (Figure 4) resistant
phenotypes that were indistinguishable from msbB pmrAC strains, suggesting that palmitate
addition is not necessary for EGTA (not shown) or polymyxin resistance (Figure 4). To confirm
that lipid A phosphoethanolamine addition and no palmitoylation is occurring in these strains, we
performed lipid A structural analyses.
Figure 6D shows the lipid A profile from a msbB pmrAC pagP2::Tn10d strain. The
spectra in this sample were calibrated differently and are shifted to the left by approximately 18
m/z units. As seen in Figure 6D, the mutational block in pagP blocks hexaacylation (no peak at 310
m/z 1806) but not phosphoethanolamine (peak at m/z 1693) addition. The loss-of-function
mutation in pagP greatly reduces lipid A palmitoylation (Figure 6F, last bar). In the pagP mutant,
~1.5% of total lipid A fatty acids were C16. This suggests that another acyl transferase may be
catalyzing the addition of palmitate at a low level or that our lipid A samples were contaminated
with palmitate derived from phospholipids. In any case, our mass spectrometry data demonstrate
that the dominant form of lipid A in msbB pmrAC pagP strains is pentaacylated lipid A decorated
with phosphoethanolamine. Since this strain’s lipid A seems to be enriched for
phosphoethanolamine, it raises the possibility that palmitoylation may inhibit
phosphoethanolamine addition.
320
Summary. The msbB mutation is of clinical interest because it allows for Salmonella to be safely
administered to mammals, which is essential for live-vaccine or attenuated-bacterial-vector
development. Suppressor mutations for msbB allow for Salmonella to avoid the septic shock
response in a stable genetic background and thus are clinically useful. By choosing a suppressor
mutation that confers the desired characteristics of a given product, such as tumor-targeting
Salmonella VNP2009 (20, 23) being non-toxic and retaining tumor-targeting and tumor-inhibition,
attenuated bacterial delivery vectors or live-vaccine strains can be genetically optimized.
ACCEPTED
on June 8, 2019 by guesthttp://jb.asm
.org/D
ownloaded from

14
In this study, we have shown that the aminoarabinose biosynthetic genes are required for
the pmrAC mutation to suppress msbB growth defects. Lipid structural analysis of these mutants
provided an unexpected result, suggesting that the addition of phosphoethanolamine and 330
palmitate to lipid A, and not aminoarabinose, was responsible for the suppressed phenotype in
msbB pmrAC Salmonella ATCC 14028. To determine if the addition of phosphoethanolamine or
palmitate is responsible for the EGTA and polymyxin resistant phenotypes, we created a msbB
pmrAC pagP strain, which had EGTA and polymyxin-resistant phenotypes indistinguishable from
msbB pmrAC Salmonella ATCC 14028. This suggests that lipid A phosphoethanolamine, and not
palmitate, addition is sufficient and necessary for both EGTA and polymyxin B resistance.
Additional experiments will be needed to determine the mechanism of how the
aminoarabinose biosynthetic proteins affect lipid A phosphoethanolamine addition. One
possibility is that the aminoarabinose genes are multi-functional, affecting both the
aminoarabinose and phosphoethanolamine branches of the pmrA pathway. An alternative 340
hypothesis is that both aminoarabinose and phosphoethanolamine biosynthetic proteins form a
complex and when essential proteins of the complex are not present, the complex may
dissociate, resulting in a loss of proper protein localization and loss of activity for both pathways.
ACKNOWLEDGMENTS
We thank Hiroshi Nikaido, Pamela Obuchowski, and Karim Suwwan de Felipe for their critical
reading of the manuscript and Chris Murray for helping with the digital images. This work was
supported by a grant from Vion Pharmaceuticals, Inc. (K.B.L.), NIH SBIR grant 1 R43 CA97595-
01 (K.B.L.) and NIH grant AI30479 (S.I.M.). S.R.M. was supported by a National Institute of
Health predoctoral training grant in genetics (5 TM32 BM07499) and a Yale University 350
Fellowship.
ACCEPTED
on June 8, 2019 by guesthttp://jb.asm
.org/D
ownloaded from

15
REFERENCES
1. Bishop, R. E., H. S. Gibbons, T. Guina, M. S. Trent, S. I. Miller, and C. R.
Raetz. 2000. Transfer of palmitate from phospholipids to lipid A in outer
membranes of gram-negative bacteria. Embo J 19:5071-80.
2. Breazeale, S. D., A. A. Ribeiro, A. L. McClerren, and C. R. Raetz. 2005. A
formyltransferase required for polymyxin resistance in Escherichia coli and the
modification of lipid A with 4-Amino-4-deoxy-L-arabinose. Identification and
function oF UDP-4-deoxy-4-formamido-L-arabinose. J Biol Chem 280:14154-67.
3. Breazeale, S. D., A. A. Ribeiro, and C. R. Raetz. 2003. Origin of lipid A species 360
modified with 4-amino-4-deoxy-L-arabinose in polymyxin-resistant mutants of
Escherichia coli. An aminotransferase (ArnB) that generates UDP-4-deoxyl-L-
arabinose. J Biol Chem 278:24731-9.
4. Caroff, M., A. Tacken, and L. Szabo. 1988. Detergent-accelerated hydrolysis of
bacterial endotoxins and determination of the anomeric configuration of the
glycosyl phosphate present in the "isolated lipid A" fragment of the Bordetella
pertussis endotoxin. Carbohydr Res 175:273-82.
5. Darveau, R. P., and R. E. Hancock. 1983. Procedure for isolation of bacterial
lipopolysaccharides from both smooth and rough Pseudomonas aeruginosa and
Salmonella typhimurium strains. J Bacteriol 155:831-8. 370
6. Datsenko, K. A., and B. L. Wanner. 2000. One-step inactivation of
chromosomal genes in Escherichia coli K-12 using PCR products. Proc Natl Acad
Sci U S A 97:6640-5.
7. Davis, R. W., D. Botstein, and J. R. Roth. 1980. Advanced bacterial genetics.
Cold Spring Harbor, Cold Spring Harbor, N.Y.
8. Ernst, R. K., E. C. Yi, L. Guo, K. B. Lim, J. L. Burns, M. Hackett, and S. I.
Miller. 1999. Specific lipopolysaccharide found in cystic fibrosis airway
Pseudomonas aeruginosa. Science 286:1561-5.
9. Gunn, J. S., K. B. Lim, J. Krueger, K. Kim, L. Guo, M. Hackett, and S. I.
Miller. 1998. PmrA-PmrB-regulated genes necessary for 4-aminoarabinose lipid 380
A modification and polymyxin resistance. Mol Microbiol 27:1171-82.
10. Gunn, J. S., and S. I. Miller. 1996. PhoP-PhoQ activates transcription of
pmrAB, encoding a two-component regulatory system involved in Salmonella
typhimurium antimicrobial peptide resistance. J Bacteriol 178:6857-64.
11. Gunn, J. S., S. S. Ryan, J. C. Van Velkinburgh, R. K. Ernst, and S. I. Miller.
2000. Genetic and functional analysis of a PmrA-PmrB-regulated locus necessary
for lipopolysaccharide modification, antimicrobial peptide resistance, and oral
virulence of Salmonella enterica serovar typhimurium. Infect Immun 68:6139-46.
12. Guo, L., K. B. Lim, J. S. Gunn, B. Bainbridge, R. P. Darveau, M. Hackett,
and S. I. Miller. 1997. Regulation of lipid A modifications by Salmonella 390
typhimurium virulence genes phoP-phoQ. Science 276:250-3.
13. Helander, I. M., I. Kilpelainen, and M. Vaara. 1994. Increased substitution of
phosphate groups in lipopolysaccharides and lipid A of the polymyxin-resistant
pmrA mutants of Salmonella typhimurium: a 31P-NMR study. Mol Microbiol
11:481-7.
ACCEPTED
on June 8, 2019 by guesthttp://jb.asm
.org/D
ownloaded from

16
14. Jia, W., A. E. Zoeiby, T. N. Petruzziello, B. Jayabalasingham, S.
Seyedirashti, and R. E. Bishop. 2004. Lipid trafficking controls endotoxin
acylation in outer membranes of Escherichia coli. J Biol Chem 279:44966-75.
15. Kato, A., and E. A. Groisman. 2004. Connecting two-component regulatory
systems by a protein that protects a response regulator from dephosphorylation by 400
its cognate sensor. Genes Dev 18:2302-13.
16. Khan, S. A., P. Everest, S. Servos, N. Foxwell, U. Zahringer, H. Brade, E. T.
Rietschel, G. Dougan, I. G. Charles, and D. J. Maskell. 1998. A lethal role for
lipid A in Salmonella infections. Mol Microbiol 29:571-9.
17. Kolodkin, A. L., M. A. Capage, E. I. Golub, and K. B. Low. 1983. F sex factor
of Escherichia coli K-12 codes for a single-stranded DNA binding protein. Proc
Natl Acad Sci U S A 80:4422-6.
18. Lee, H., F. F. Hsu, J. Turk, and E. A. Groisman. 2004. The PmrA-regulated
pmrC gene mediates phosphoethanolamine modification of lipid A and
polymyxin resistance in Salmonella enterica. J Bacteriol 186:4124-33. 410
19. Low, K. B., M. Ittensohn, T. Le, J. Platt, S. Sodi, M. Amoss, O. Ash, E.
Carmichael, A. Chakraborty, J. Fischer, S. L. Lin, X. Luo, S. I. Miller, L.
Zheng, I. King, J. M. Pawelek, and D. Bermudes. 1999. Lipid A mutant
Salmonella with suppressed virulence and TNFalpha induction retain tumor-
targeting in vivo. Nat Biotechnol 17:37-41.
20. Low, K. B., M. Ittensohn, X. Luo, L. M. Zheng, I. King, J. M. Pawelek, and
D. Bermudes. 2004. Construction of VNP20009: a novel, genetically stable
antibiotic-sensitive strain of tumor-targeting Salmonella for parenteral
administration in humans. Methods Mol Med 90:47-60.
21. Miller, J. H. 1972. Experiments in molecular genetics. Cold Spring Harbor, Cold 420
Spring Harbor, N.Y.
22. Murray, S. R., D. Bermudes, K. S. de Felipe, and K. B. Low. 2001. Extragenic
suppressors of growth defects in msbB Salmonella. J Bacteriol 183:5554-61.
23. Murray, S. R., K. S. de Felipe, P. L. Obuchowski, J. Pike, D. Bermudes, and
K. B. Low. 2004. Hot spot for a large deletion in the 18- to 19-centisome region
confers a multiple phenotype in Salmonella enterica serovar Typhimurium strain
ATCC 14028. J Bacteriol 186:8516-23.
24. Nikaido, H. 1996. Outer membrane., p. 29-47. In F. Neidhardt, R. Curtis, J.
Ingraham, E. C. C. Lin, K. B. Low, B. Magasanik, W. Reznikoff, M. Riley, M.
Schaechter, and H. E. Umbarger (ed.), Escherichia coli and Salmonella: Cellular 430
and molecular biology. ASM Press, Washington, D.C.
25. O'Callaghan, D., and A. Charbit. 1990. High efficiency transformation of
Salmonella typhimurium and Salmonella typhi by electroporation. Mol Gen Genet
223:156-8.
26. Roland, K. L., L. E. Martin, C. R. Esther, and J. K. Spitznagel. 1993.
Spontaneous pmrA mutants of Salmonella typhimurium LT2 define a new two-
component regulatory system with a possible role in virulence. J Bacteriol
175:4154-64.
27. Somerville, J. E., Jr., L. Cassiano, B. Bainbridge, M. D. Cunningham, and R.
P. Darveau. 1996. A novel Escherichia coli lipid A mutant that produces an 440
antiinflammatory lipopolysaccharide. J Clin Invest 97:359-65.
ACCEPTED
on June 8, 2019 by guesthttp://jb.asm
.org/D
ownloaded from

17
28. Soncini, F. C., and E. A. Groisman. 1996. Two-component regulatory systems
can interact to process multiple environmental signals. J Bacteriol 178:6796-801.
29. Tamayo, R., B. Choudhury, A. Septer, M. Merighi, R. Carlson, and J. S.
Gunn. 2005. Identification of cptA, a PmrA-regulated locus required for
phosphoethanolamine modification of the Salmonella enterica serovar
typhimurium lipopolysaccharide core. J Bacteriol 187:3391-9.
30. Tamayo, R., S. S. Ryan, A. J. McCoy, and J. S. Gunn. 2002. Identification and
genetic characterization of PmrA-regulated genes and genes involved in
polymyxin B resistance in Salmonella enterica serovar typhimurium. Infect 450
Immun 70:6770-8.
31. Tran, A. X., M. E. Lester, C. M. Stead, C. R. Raetz, D. J. Maskell, S. C.
McGrath, R. J. Cotter, and M. S. Trent. 2005. Resistance to the antimicrobial
peptide polymyxin requires myristoylation of Escherichia coli and Salmonella
typhimurium lipid A. J Biol Chem 280:28186-94.
32. Trent, M. S., A. A. Ribeiro, W. T. Doerrler, S. Lin, R. J. Cotter, and C. R.
Raetz. 2001. Accumulation of a polyisoprene-linked amino sugar in polymyxin-
resistant Salmonella typhimurium and Escherichia coli: structural characterization
and transfer to lipid A in the periplasm. J Biol Chem 276:43132-44.
33. Trent, M. S., A. A. Ribeiro, S. Lin, R. J. Cotter, and C. R. Raetz. 2001. An 460
inner membrane enzyme in Salmonella and Escherichia coli that transfers 4-
amino-4-deoxy-L-arabinose to lipid A: induction on polymyxin-resistant mutants
and role of a novel lipid-linked donor. J Biol Chem 276:43122-31.
34. Vaara, M. 1981. Increased outer membrane resistance to
ethylenediaminetetraacetate and cations in novel lipid A mutants. J Bacteriol
148:426-34.
35. Vaara, M., T. Vaara, M. Jensen, I. Helander, M. Nurminen, E. T. Rietschel,
and P. H. Makela. 1981. Characterization of the lipopolysaccharide from the
polymyxin-resistant pmrA mutants of Salmonella typhimurium. FEBS Lett
129:145-9. 470
36. Williams, G. J., S. D. Breazeale, C. R. Raetz, and J. H. Naismith. 2005.
Structure and function of both domains of ArnA, a dual function decarboxylase
and a formyltransferase, involved in 4-amino-4-deoxy-L-arabinose biosynthesis. J
Biol Chem 280:23000-8.
37. Wosten, M. M., L. F. Kox, S. Chamnongpol, F. C. Soncini, and E. A.
Groisman. 2000. A signal transduction system that responds to extracellular iron.
Cell 103:113-25.
38. Zhou, Z., S. Lin, R. J. Cotter, and C. R. Raetz. 1999. Lipid A modifications
characteristic of Salmonella typhimurium are induced by NH4VO3 in Escherichia
coli K12. Detection of 4-amino-4-deoxy-L-arabinose, phosphoethanolamine and 480
palmitate. J Biol Chem 274:18503-14.
39. Zhou, Z., A. A. Ribeiro, S. Lin, R. J. Cotter, S. I. Miller, and C. R. Raetz.
2001. Lipid A modifications in polymyxin-resistant Salmonella typhimurium:
PMRA-dependent 4-amino-4-deoxy-L-arabinose, and phosphoethanolamine
incorporation. J Biol Chem 276:43111-21.
ACCEPTED
on June 8, 2019 by guesthttp://jb.asm
.org/D
ownloaded from

18
Figure and Table Legends
Figure 1. Lipid A structure. Core lipid A structure is shown in black; in msbB strains, no myristoyl 490
group is added (marked with filled arrow); PmrA/B - and PhoP/Q - regulated modifications are
shown in grey. Activation of PmrA/B results in modification of the phosphates of lipid A with
phosphoethanolamine and aminoarabinose. The PmrA/B two-component system can be turned
on by PhoP via the PmrD protein (15). Activation of PhoP/Q results in lipid A lacking one acyl
group (marked by open arrow; this only occurs on lipid A molecules lacking aminoarabinose),
decorated with palmitate (grey 16 carbon chain) and hydroxylated at the myristoyl residue
(marked by a grey X).
Figure 2. Replica plate series showing the effect of PmrA/B mutations on the growth of msbB
Salmonella ATCC 14028. Single colonies were patched onto an LB-no salt agar plate (not 500
shown) and incubated overnight at 37°C. Replica plates were incubated for 10 hours at 37°C.
(A) MSB agar; (B) EGTA agar; (C) Galactose MacConkey agar. 14028 grows well on all 3
media. msbB is sensitive to EGTA and Galactose MacConkey agar. msbB pmrA has a similar
phenotype to unsuppressed msbB Salmonella ATCC 14028. msbB pmrAC Salmonella ATCC
14028 is EGTA resistant but Galactose MacConkey sensitive. All strains grew patches on the
master plate (not shown).
Figure 3. Aminoarabinose biosynthetic genes are required for pmrAC to suppress msbB growth
defects. Replica plates were incubated for 10 hours at 37°C. All strains in this figure carry the
rpsL mutation (for streptomycin resistance). The pmrAC allele confers EGTA resistance to msbB 510
Salmonella ATCC 14028. Non-polar deletions in pmrHFIJKL restore EGTA sensitivity to msbB
pmrAC Salmonella ATCC 14028. However, a loss-of-function mutation in pmrM (which is not
believed to play a role in aminoarabinose biosynthesis) did not restore EGTA sensitivity to msbB
pmrAC Salmonella ATCC 14028.
ACCEPTED
on June 8, 2019 by guesthttp://jb.asm
.org/D
ownloaded from

19
Figure 4. The effect of loss-of-function mutations in the aminoarabinose biosynthetic genes on
polymyxin resistance in an msbB pmrAC genetic background. Polymyxin sensitivity was
measured in terms of percent survival. Cells were exposed to 0.1 µg/ml polymyxin B sulfate for
1.5 hours at 37°C and then were plated to determine colony forming units (CFUs). Y-error bars
show variation in CFUs between 3 independent clones for each strain. The pmrAC mutation 520
confers polymyxin resistance and loss-of-function mutations in pmrHFIJKL restore polymyxin
sensitivity to msbB pmrAC Salmonella ATCC 14028. Loss-of-function mutations in pagP and
pmrM do not restore polymyxin sensitivity. PagP and PmrM are not believed to play a role in
aminoarabinose biosynthesis.
Figure 5. Mass spectra and gas chromatography of lipid A from ATCC 14028 and ATCC 14028
pmrAC show increased palmitate addition when grown in the presence of salt. Lipid A was
purified from at least two independent cultures for each strain and the resulting mass spectra
were nearly identical. Y-axis units represent relative intensity and X-axis units (m/z) represent the
mass-to-charge ratio of different lipid A species. (A) ATCC 14028 grown in LB broth; (B) ATCC 530
14028 grown in LB-no-salt broth; (C) 14028 pmrAC (SM2206) grown in LB broth; (D) ATCC
14028 pmrAC (SM2206) grown in LB-no-salt broth. Hexaacylated lipid A (m/z 1797),
heptaacylated lipid A (m/z 2036), hexaacylated lipid A with phosphoethanolamine (m/z 1920) or
aminoarabinose (m/z 1928), and heptaacylated lipid A with phosphoethanolamine (m/z 2159) or
aminoarabinose (m/z 2167) peaks, are shown. (E) Gas chromatography results for lipid A from
these strains grown in LB and LB-no-salt broth.
Figure 6. Lipid A structure in msbB, msbB pmrAC, msbB pmrA
C pmrF, and msbB pmrA
C
pagP strains reveals pmrAC and pmrHFIJKL-dependent phosphoethanolamine addition
and increased palmitoylation. Lipid A was purified from at least two independent 540
cultures for each strain and the resulting mass spectra were nearly identical. Y-axis units
represent relative intensity and X-axis units (m/z) represent the mass-to-charge ratio of
ACCEPTED
on June 8, 2019 by guesthttp://jb.asm
.org/D
ownloaded from

20
different lipid A species. (A) msbB (YS1) in MSB broth; (B) msbB pmrAC rpsL
(SM2192) in MSB broth; (C) msbB pmrAC pmrF rpsL (SM2103) in MSB broth; (D)
msbB pmrAC pagP (SM2035) in MSB broth lacking MgSO4. Peaks representing
pentaacylated lipid A (m/z 1588 [A, B, C] and m/z 1570 [D]), hexaacylated lipid A (m/z
1826), pentaacylated lipid A with phosphoethanolamine (m/z 1711 [A,B, C] or m/z 1693
[D]), and hexaacylated lipid A with phosphoethanolamine (m/z 1949) are shown. (E)
Thin-layer chromatography confirms that msbB pmrAC
has a modified lipid A structure.
msbB and msbB somA have indistinguishable lipid A TLC profiles (data not shown) but 550
msbB pmrAC has a distinct profile which includes phosphoethanolamine and
palmitoylated lipid A. Palmitoylated lipid A has been putatively marked with an arrow
and the additional spots near the bottom of the TLC plate likely represent different forms
of lipid A containing phosphoethanolamine. (F) Gas chromatographic analysis of LPS
fatty acids from msbB, msbB pmrAC, msbB pmrA
C pmrH and msbB pmrA
C pmrF
Salmonella ATCC 14028 grown in MSB broth without MgSO4. msbB strains have
greatly reduced amounts of C14 incorporation compared to wild type (as shown in Figure
5E). msbB pmrAC strains have increased levels of palmitate (C16) addition and EGTA
challenge also leads to increased palmitate addition.
560
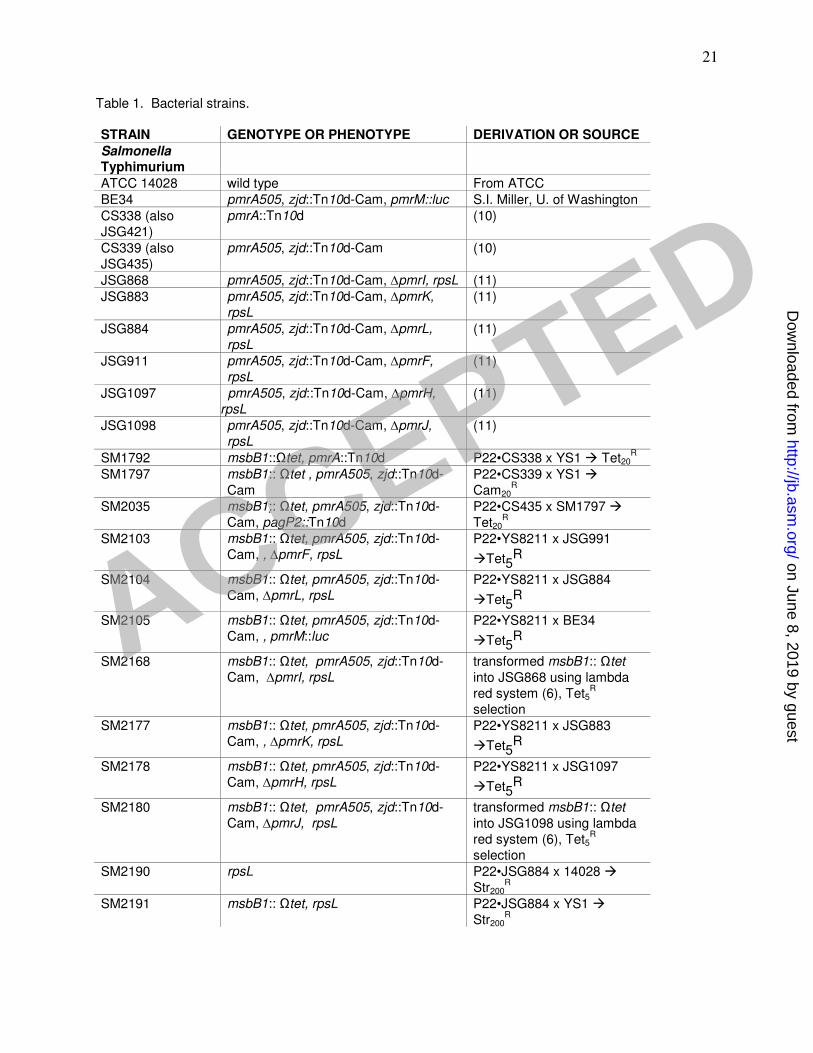
Table 1: Bacterial Strains
Genotypes/phenotypes and derivation/source are shown for each strain of Salmonella
Typhimurium ATCC 14028 used in this study.
ACCEPTED
on June 8, 2019 by guesthttp://jb.asm
.org/D
ownloaded from
21
Table 1. Bacterial strains.
STRAIN GENOTYPE OR PHENOTYPE DERIVATION OR SOURCE
Salmonella Typhimurium
ATCC 14028 wild type From ATCC
BE34 pmrA505, zjd::Tn10d-Cam, pmrM::luc S.I. Miller, U. of Washington
CS338 (also JSG421)
pmrA::Tn10d (10)
CS339 (also JSG435)
pmrA505, zjd::Tn10d-Cam (10)
JSG868 pmrA505, zjd::Tn10d-Cam, ∆pmrI, rpsL (11)
JSG883 pmrA505, zjd::Tn10d-Cam, ∆pmrK,
rpsL (11)
JSG884 pmrA505, zjd::Tn10d-Cam, ∆pmrL,
rpsL (11)
JSG911 pmrA505, zjd::Tn10d-Cam, ∆pmrF,
rpsL (11)
JSG1097 pmrA505, zjd::Tn10d-Cam, ∆pmrH,
rpsL (11)
JSG1098 pmrA505, zjd::Tn10d-Cam, ∆pmrJ,
rpsL (11)
SM1792 msbB1::Ωtet, pmrA::Tn10d P22•CS338 x YS1 Tet20R
SM1797 msbB1:: Ωtet , pmrA505, zjd::Tn10d-Cam
P22•CS339 x YS1 Cam20
R
SM2035 msbB1:: Ωtet, pmrA505, zjd::Tn10d-Cam, pagP2::Tn10d
P22•CS435 x SM1797 Tet20
R
SM2103 msbB1:: Ωtet, pmrA505, zjd::Tn10d-Cam, , ∆pmrF, rpsL
P22•YS8211 x JSG991
Tet5R
SM2104 msbB1:: Ωtet, pmrA505, zjd::Tn10d-Cam, ∆pmrL, rpsL
P22•YS8211 x JSG884
Tet5R
SM2105 msbB1:: Ωtet, pmrA505, zjd::Tn10d-Cam, , pmrM::luc
P22•YS8211 x BE34
Tet5R
SM2168 msbB1:: Ωtet, pmrA505, zjd::Tn10d-Cam, ∆pmrI, rpsL
transformed msbB1:: Ωtet into JSG868 using lambda red system (6), Tet5
R
selection
SM2177 msbB1:: Ωtet, pmrA505, zjd::Tn10d-Cam, , ∆pmrK, rpsL
P22•YS8211 x JSG883
Tet5R
SM2178 msbB1:: Ωtet, pmrA505, zjd::Tn10d-Cam, ∆pmrH, rpsL
P22•YS8211 x JSG1097
Tet5R
SM2180 msbB1:: Ωtet, pmrA505, zjd::Tn10d-Cam, ∆pmrJ, rpsL
transformed msbB1:: Ωtet into JSG1098 using lambda red system (6), Tet5
R
selection
SM2190 rpsL P22•JSG884 x 14028 Str200
R
SM2191 msbB1:: Ωtet, rpsL P22•JSG884 x YS1 Str200
R
ACCEPTED
on June 8, 2019 by guesthttp://jb.asm
.org/D
ownloaded from

22
SM2192 msbB1:: Ωtet, pmrA505, zjd::Tn10d-Cam, rpsL
P22•JSG884 x SM1797 Str200
R
SM2193 msbB1:: Ωtet, pmrA505, zjd::Tn10d-Cam, pmrM::luc, rpsL
P22•JSG884 x SM2105 Str200
R
SM2206 pmrA505, zjd::Tn10d-Cam P22•CS339 x 14028 Cam20
R
YS1 msbB1:: Ωtet (22)
ACCEPTED
on June 8, 2019 by guesthttp://jb.asm
.org/D
ownloaded from

Figure 1.
aminoarabinose
phosphoethanolamine
O
OO
O
OH
O
OH
O
orHO
NH2
H2NP
OH
OH
OH
HOHO
HO
HO
HO
O
O
OO
O
O
O
O
O
or
NH2
NH2
P
OH
OHHO
P
HOO
O
OO
OO
O O
O
O
O
O
XX OH
or H
O
O
HO
OH
HO
NH O
ONH
OP
OH
O
ACCEPTED
on June 8, 2019 by guesthttp://jb.asm
.org/D
ownloaded from

Figure 2.
ATCC 14028
msbB (YS1)
msbB pmrA::Tn10 (SM1792)
msbB pmrAC (SM1797)
A. MSB B. EGTA C. gal.-Mac.
ACCEPTED
on June 8, 2019 by guesthttp://jb.asm
.org/D
ownloaded from

Figure 3.
ATCC 14028 rpsL (SM2190)
msbB rpsL (SM2191)
msbB pmrAC rpsL (SM2192)
msbB pmrAC pmrH rpsL(SM2178)
msbB pmrAC pmrF rpsL (SM2103)
msbB pmrAC pmrI rpsL (SM2168)
msbB pmrAC pmrJ rpsL (SM2180)
msbB pmrAC pmrK rpsL (SM2177)
msbB pmrAC pmrL rpsL (SM2104)
msbB pmrAC pmrM rpsL (SM2193)
A. MSB B. EGTA
ACCEPTED
on June 8, 2019 by guesthttp://jb.asm
.org/D
ownloaded from


Figure 5.
E.
ACCEPTED
on June 8, 2019 by guesthttp://jb.asm
.org/D
ownloaded from

Figure 6.
E. F. ACCEPTED
on June 8, 2019 by guesthttp://jb.asm
.org/D
ownloaded from