azotobacter vinelandii siderophore can provide nitrogen to support the culture of the green algae...
TRANSCRIPT

R E S EA RCH L E T T E R
Azotobacter vinelandii siderophore can provide nitrogen tosupport the culture of the green algae Neochloris oleoabundans
and Scenedesmus sp. BA032
Juan A. Villa1, Erin E. Ray2 & Brett M. Barney1,2
1Biotechnology Institute, University of Minnesota, St. Paul, MN, USA; and 2Department of Bioproducts and Biosystems Engineering, University of
Minnesota, St. Paul, MN, USA
Correspondence: Brett M. Barney,
Department of Bioproducts and Biosystems
Engineering, University of Minnesota, 1390
Eckles Avenue, St. Paul, MN 55108-6130,
USA. Tel.: +1 612 626 8751;
fax: +1 612 625 6286;
e-mail: [email protected]
Received 21 September 2013; revised 24
November 2013; accepted 24 November
2013. Final version published online 9
January 2014.
DOI: 10.1111/1574-6968.12347
Editor : Yaacov Okon
Keywords
Siderophore; Azotobacter vinelandii;
Neochloris oleoabundans; Scenedesmus;
nitrogen.
Abstract
Microalgae are viewed as a potential future agricultural and biofuel feedstock and
also provide an ideal biological means of carbon sequestration based on rapid
growth rates and high biomass yields. Any potential improvement using high-
yield microalgae to fix carbon will require additional fertilizer inputs to provide
the necessary nitrogen required for protein and nucleotide biosynthesis. The free-
living diazotroph Azotobacter vinelandii can fix nitrogen under aerobic conditions
in the presence of reduced carbon sources such as sucrose or glycerol and is also
known to produce a variety of siderophores to scavenge different metals from the
environment. In this study, we identified two strains of green algae, Neochloris
oleoabundans and Scenedesmus sp. BA032, that are able to utilize the A. vinelandii
siderophore azotobactin as a source of nitrogen to support growth. When grown
in a co-culture, S. sp. BA032 and N. oleoabundans obtained the nitrogen required
for growth through the association with A. vinelandii. These results, indicating a
commensalistic relationship, provide a proof of concept for developing a mutual-
istic or symbiotic relationship between these two species using siderophores as a
nitrogen shuttle and might further indicate an additional fate of siderophores in
the environment.
Introduction
Nitrogen is an important component of living systems. In
agriculture, conventional crops such as soybeans, alfalfa,
and clover meet their nitrogen requirements by forming
symbiotic relationships with diazotrophic soil bacteria
(Jones et al., 2007; Ikeda et al., 2010), while many other
agricultural crops such as corn require substantial nitro-
gen inputs from Haber-Bosch-based industrial processes.
Potential next-generation crops such as single-celled mic-
roalgae provide possible benefits in terms of total biomass
yield per square area, high density, and rapid growth vs.
current conventional agriculture crops, but would also
require substantial increases in specific nutrient inputs
such as nitrogen. Thus, a key concern in pursuing sus-
tainable next-generation oil crops should include specific
consideration of the sustainability of the nitrogen inputs.
Azotobacter vinelandii is a common Gram-negative soil
bacterium that can fix atmospheric nitrogen under
aerobic conditions. This characteristic differentiates it
from many other nitrogen-fixing bacteria that require
anaerobic or microaerobic conditions to protect the
oxygen-sensitive nitrogenase (Setubal et al., 2009). This
feature also makes it an ideal candidate strain for poten-
tial co-culture with oxygen-producing phototrophs such
as microalgae (Ortiz-Marquez et al., 2012). Additionally,
A. vinelandii is considered as an ideal strain in potential
biotechnology applications for the production of higher-
value bioproducts such as polyhydroxyalkanoates, which
could serve as potential bioplastics (Setubal et al., 2009).
Azotobacter strains produce a range of nitrogen com-
pounds which may be released into the extracellular space
under certain conditions. Extensive ammonia release was
reported two decades ago from A. vinelandii based on a
modification resulting in differential regulation of nitrogen
fixation genes, resulting in high concentrations of ammonia
accumulating in the growth media (Bali et al., 1992;
Brewin et al., 1999). This feature was recently utilized to
FEMS Microbiol Lett 351 (2014) 70–77ª 2013 Federation of European Microbiological Societies.Published by John Wiley & Sons Ltd. All rights reserved
MIC
ROBI
OLO
GY
LET
TER
S

demonstrate the potential to apply ammonia production to
the co-culture of various algae strains (Ortiz-Marquez
et al., 2012). A. vinelandii has also been reported to excrete
a range of additional nitrogen compounds to serve various
functions, including proteins involved in the production of
alginate (Gimmestad et al., 2006).
As a diazotroph, A. vinelandii requires substantial
quantities of iron to grow under optimal conditions.
Iron is important for central metabolism and is also
essential as a component of the two proteins that make
up the nitrogenase complex (Barney et al., 2006;
Wichard et al., 2009). A. vinelandii contains multiple sid-
erophore biosynthetic pathways and diverts a portion of
the nitrogen obtained through biological nitrogen fixa-
tion to the specialized nitrogen-rich pyoverdine and cate-
chol siderophores (Fig. 1), which are excreted into the
extracellular space, to assist in the uptake of required
metal ions (Tindale et al., 2000; Yoneyama et al., 2011).
Siderophore systems enable this bacterium to adapt to
metal limitation in the environment (Wichard et al.,
2009). The siderophores bind various metals with a high
affinity and are then taken up by the cell through mem-
brane-bound transport systems (Cornish & Page, 1995;
Palanch�e et al., 2004; Wichard et al., 2009). It is this lat-
ter class of siderophore compounds that were of particu-
lar interest in this study to determine the potential of
these compounds to serve as a nitrogen source to various
algal strains.
Materials and methods
Reagents, strains and cell counting
Azotobacter vinelandii DJ (trans) was obtained from Den-
nis Dean (Virginia Tech), while Scenedesmus sp. BA032 is
an environmental isolate collected from the Cache Valley
in northern Utah. Neochloris oleoabundans was obtained
from the UTEX culture collection of algae. All chemicals
and reagents were obtained from Sigma Aldrich (St.
Louis, MO) or Thermo Fisher Scientific (Pittsburgh, PA).
Cell counts of S. sp. BA032 were measured using a hemo-
cytometer following the directions of the manufacturer
(Hausser Scientific, Horsham, PA).
Bacterial strains and growth conditions
Cultures of A. vinelandii DJ (trans) and gene substitution
strains described below were grown aerobically in
modified Burk’s media for siderophore production (SPB
media), which lacks added iron and includes zinc
(58 mM sucrose, 0.34 mM CaCl∙2H2O, 0.41 mM
MgSO4∙7H2O, 40 lM Na2MoO4∙2H2O, 40 lM ZnCl,
Fig. 1. Examples of nitrogen-containing siderophores produced by Azotobacter vinelandii. Shown are the chemical structures of several nitrogen-
containing siderophores of the catechol class (aminochelin, azotochelin, and protochelin) and the pyoverdine siderophore azotobactin that
contain various percentages of nitrogen.
FEMS Microbiol Lett 351 (2014) 70–77 ª 2013 Federation of European Microbiological Societies.Published by John Wiley & Sons Ltd. All rights reserved
Siderophores support algal growth 71

0.73 mM K2HPO4, and 2.32 mM KH2PO4) at 30 °C with
agitation at 200 r.p.m. (Page & Sadoff, 1975; Huyer &
Page, 1988; Page et al., 2003).
Construction of A. vinelandii gene substitution
strains
The csbC gene of A. vinelandii codes for a key enzyme in
the biosynthesis of catechol-based siderophores (Tindale
et al., 2000). The csbC gene was cloned along with flank-
ing regions of c. 500 bp into a pUC19-derived plasmid
using primers BBP1301 (5′-CAGATAAG CTTGTCTG
GTCGATCA GGATCGCC ATG-3′) and BBP1302
(5′-GACAGGTA CCACGCTG TAGAAATA GTCGTCGT
G-3′) with HindIII and KpnI sites used to insert the gene
underlined. The csbC gene was then removed from the
plasmid using PCR with primers BBP1331 (5′-GACAGGAT CCTCTAGA TATGCATA TGGCCTCC TTACGGCT
AGAGGACG AG-3′) and BBP1332 (5′-CTGAGGAT CCG
AGCAC CTCGACCC CGACCTCT TC-3′) with BamHI
sites underlined. The spectinomycin resistance cassette
from pHP45Ω (Prentki & Krisch, 1984) was introduced
into the BamHI site to produce pPCRSIDK3. The strain
A. vinelandii AZBB040 was constructed by transforming
A. vinelandii DJ (trans) with pPCRSIDK3. Following a
double homologous recombination event, colonies were
selected for antibiotic resistance resulting in a strain
A. vinelandii AZBB040, which contains the DcsbC::spectr
substitution. Strains were confirmed to contain the geno-
mic modification by PCR with primers BBP1419 (5′-GAACAG CACGAA GCTCAG CATCAG C-3′) and BBP1420
(5′-CGAACA CCTGTT GCAGCT TGCAGC-3′) lying
outside of the region modified.
The gene coding for Avin_25580 of A. vinelandii encodes
a key enzyme in the biosynthesis of azotobactin sidero-
phore (Yoneyama et al., 2011). The gene for Avin_25580
was cloned along with flanking regions of c. 500 bp into a
pUC19-derived plasmid using primers BBP1502 (5′-GACTAAGC TTGAAGCG TTCCCGGC TGAAGGTC-3′) and
BBP1503 (5′-GTGACC CTGTTCAT GCTGCTGC TG-3′)with HindIII and EcoRI (downstream of primer) sites used
to insert the gene. The gene for Avin_25580 was then
removed from the plasmid using PCR with primers
BBP1562 (5′-NNNGGATC CACCCAGG TCAACGAC
CTGCTGCT G-3′) and BBP1563 (5′-NNNGGATC CG
TCCTCG CCACCTGC TCCTCGAT CAACTG-3′) with
BamHI sites. A tetracycline resistance cassette derived from
pRK415 (Mather et al., 1995) was introduced into the
BamHI site to produce pPCRSIDK9. A. vinelandii was
transformed as described above. Colonies were selected for
antibiotic resistance resulting in the strain A. vinelandii
AZBB041, which contains the substitution to the gene cod-
ing for Avin_25580. Strains were confirmed to contain the
genomic modification by PCR with primers BBP1623
(5′-CACTTG CTGGAC AAAGAG ACGGTC C-3′) and
BBP1624 (5′-CTGCGG TTCAGC TCTCCG TAGCTC
ATTTC-3′) lying outside of the region modified.
Optical characterization to confirm phenotype
Azotobacter vinelandii wild-type and gene substitution
strains were grown for 72 h in SPB media, and cell density
was determined by measuring the optical density (OD) at
600 nm to confirm similar rates of growth. Specific optical
features of the siderophores were used to monitor the
culture supernatant (Page & Huyer, 1984; Page et al.,
1991; Rodr�ıguez-L�opez et al., 1991). The absorbance spec-
tra were measured using a Varian Bio 50 UV/Vis spectro-
photometer. Levels of catechols were determined using an
approach similar to that used by Page and Huyer (Page &
Huyer, 1984) and the reported extinction coefficient for
dihydroxybenzoic acid (DHBA) at pH 3.0 (Rodr�ıguez-
L�opez et al., 1991). The absorbance at 380 nm was used
to estimate the levels of azotobactin in culture superna-
tants using the reported extinction coefficients
[kmax = 380 nm at pH 4.0, e = 23 500 M�1 cm�1 (Page
et al., 1991)] and catechols [kmax = 310 nm at pH 3.0,
e = 9200 M�1 cm�1 (Rodr�ıguez-L�opez et al., 1991)].
Algae growth medium and conditions
Algae were maintained on a derivative of Bold’s basal med-
ium (Bold, 1949) containing 25 mg L�1 NaCl, 75 mg L�1
MgSO4�7H2O, 25 mg L�1 CaCl2�2H2O, 100 mg L�1
Na2SO4, 300 mg L�1 K2HPO4, 600 mg L�1 NaNO3, and
5 mg L�1 FeSO4�7H2O, adjusted to pH 7.6. The medium
was further supplemented with 1 mL L�1 of trace metals
solution containing 1.0 g L�1 boric acid, 1 g L�1 sodium
EDTA, 200 mg L�1 MnCl2�4H2O, 20 mg L�1 ZnCl2,
15 mg L�1 CuCl2�2H2O, 15 mg L�1 Na2MoO4�2H2O, and
15 mg L�1 CoCl2�6H2O. Cultures of S. sp. BA032 and
N. oleoabundans were maintained on agar plates of the
modified medium listed above.
Siderophores isolation from A. vinelandii
Cultures of A. vinelandii wild-type cells were cultured on
SPB medium. Following growth, cells were centrifuged at
7000 g for 10 min. Supernatant was separated from the
cells, then filtered (0.22 lm pore size, Nalgene Filtration,
Thermo Scientific), and collected. The filtered supernatant
was added to a column containing Q-Sepharose (GE
Healthcare). The material retained by the column was
rinsed with distilled water and eluted with 500 mM NaCl
in distilled water. The sample of crude concentrated
siderophores was substituted for sodium nitrate in the
FEMS Microbiol Lett 351 (2014) 70–77ª 2013 Federation of European Microbiological Societies.Published by John Wiley & Sons Ltd. All rights reserved
72 J.A. Villa et al.

medium described above based on Bold’s basal medium
(Bold, 1949) to screen algae strains for potential growth
on agar plates.
Algae growth with siderophores
To determine the ability of specific algae strains to grow
on A. vinelandii siderophores, A. vinelandii wild-type and
gene substitution strains were first grown on SPB med-
ium for 3 days, then cells were removed by centrifuga-
tion, and the obtained supernatant was filtered as
described above into an autoclaved container. The super-
natant was transferred aseptically to a clean Erlenmeyer
flask, and an equivalent quantity of algal cells was inocu-
lated into each flask. Cells were grown under a bank of
fluorescent lights (c. 200 µmols s�1 m�2) with constant
agitation for c. 8 days while counting the algal cells using
a hemocytometer.
Results and discussion
Identification of algae capable of growing on
siderophores as a nitrogen source
A primary objective of this work was to determine
whether A. vinelandii-derived siderophores could serve as
a suitable shuttle of nitrogen from A. vinelandii to nondi-
azotrophic photosynthetic species such as algae. It has
been reported previously that certain species are known
to take advantage of siderophore-producing bacteria by
utilizing foreign siderophores for metal uptake (Amin
et al., 2009; D’Onofrio et al., 2010). To determine
whether this feature could also satisfy fixed nitrogen
requirements for a phototroph, we first isolated sidero-
phores from media of A. vinelandii using a simple ion-
exchange resin to collect the siderophores from spent
media as described in the methods. Then, 18 green algae
strains were screened on a simple Bold’s basal media
where sodium nitrate was replaced with the isolated sid-
erophores. Two strains were able to replicate on the sid-
erophore-containing media. The first was a strain of
Scenedesmus sp. BA032 isolated from the Cache Valley in
northern Utah, and the second was Neochloris oleoabun-
dans, a strain of interest for potential neutral lipid pro-
duction (Li et al., 2008; Gouveia et al., 2009). These two
strains were selected for further siderophore utilization
studies as described below.
Identification of specific class of siderophores
supporting algae growth
The siderophores produced by A. vinelandii fall into two
different classes called the catechol and pyoverdine
siderophores (Fig. 1). Others have recently reported the
successful disruption of siderophore biosynthetic genes in
A. vinelandii (Yoneyama et al., 2011). Here, we utilized a
similar approach to what was taken previously by target-
ing a key gene in each of the two pathways that produce
either class of siderophore (Tindale et al., 2000; Yoney-
ama et al., 2011). Gene substitution strains were con-
structed and the phenotype (deficient in the production
of either class of siderophore) was confirmed by measur-
ing the absorbance of spent culture media (Fig. 2), con-
firming the results reported previously by others
(Yoneyama et al., 2011). These gene substitution strains
are important for several reasons. First, there are two
classes of siderophores produced by A. vinelandii, and
disruption of either class should allow us to determine
whether algae are growing on either class specifically. Sec-
ond, siderophores are not the only extracellular nitrogen
compound produced by A. vinelandii. Azotobacter vine-
landii also produces other compounds, including ammo-
nia, urea, or extracellular proteins (Bali et al., 1992;
Brewin et al., 1999; Gimmestad et al., 2006), each of
which could serve as sources of nitrogen to support algae
growth. Thus, if the algae strain were able to grow on
both substitution strains, this might indicate that another
compound was responsible for providing the nitrogen, as
has been shown for alternative algae strains (Ortiz-Mar-
quez et al., 2012).
Fig. 2. Production of siderophores in Azotobacter vinelandii wild-type
and gene substitution strains AZBB040 and AZBB041. Bacterial cells
were grown for 72 h in SPB medium and the cultural supernatants
were collected. Specific absorbance of the siderophores at 310 and
380 nm is derived from the catechol and azotobactin, respectively.
Absorbance spectra were measured at pH 4.0 for AZBB041 and wild-
type strains and at pH 3.0 for AZBB040.
FEMS Microbiol Lett 351 (2014) 70–77 ª 2013 Federation of European Microbiological Societies.Published by John Wiley & Sons Ltd. All rights reserved
Siderophores support algal growth 73
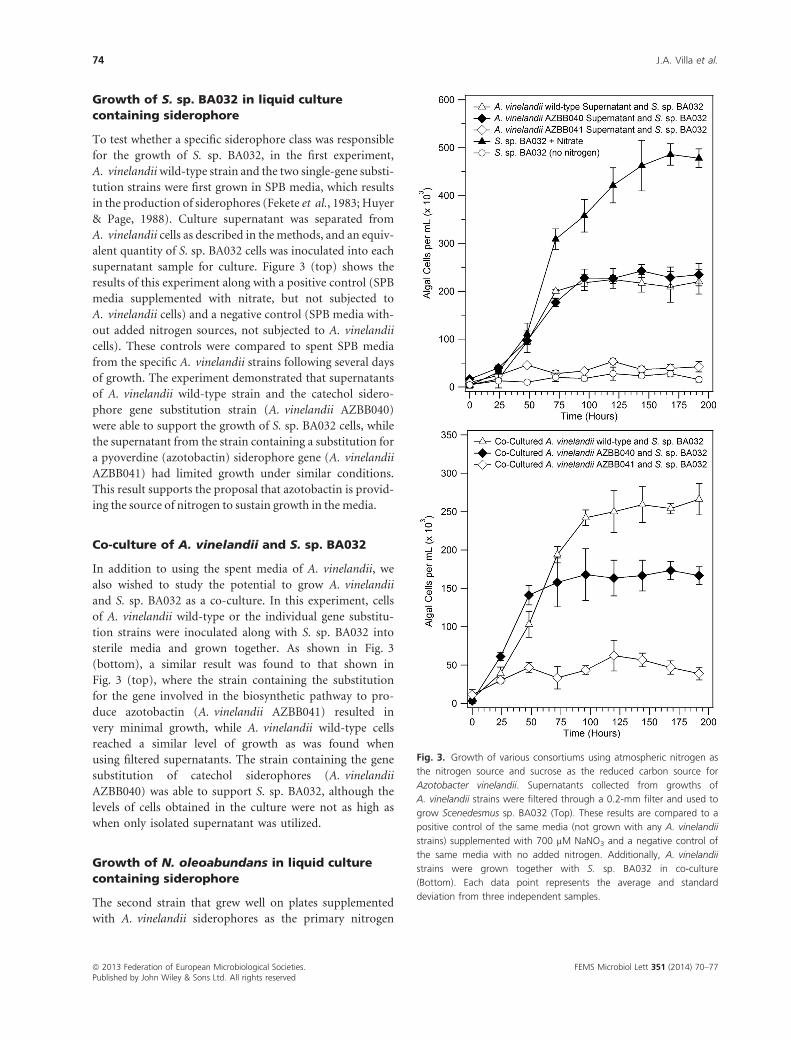
Growth of S. sp. BA032 in liquid culture
containing siderophore
To test whether a specific siderophore class was responsible
for the growth of S. sp. BA032, in the first experiment,
A. vinelandiiwild-type strain and the two single-gene substi-
tution strains were first grown in SPB media, which results
in the production of siderophores (Fekete et al., 1983; Huyer
& Page, 1988). Culture supernatant was separated from
A. vinelandii cells as described in the methods, and an equiv-
alent quantity of S. sp. BA032 cells was inoculated into each
supernatant sample for culture. Figure 3 (top) shows the
results of this experiment along with a positive control (SPB
media supplemented with nitrate, but not subjected to
A. vinelandii cells) and a negative control (SPB media with-
out added nitrogen sources, not subjected to A. vinelandii
cells). These controls were compared to spent SPB media
from the specific A. vinelandii strains following several days
of growth. The experiment demonstrated that supernatants
of A. vinelandii wild-type strain and the catechol sidero-
phore gene substitution strain (A. vinelandii AZBB040)
were able to support the growth of S. sp. BA032 cells, while
the supernatant from the strain containing a substitution for
a pyoverdine (azotobactin) siderophore gene (A. vinelandii
AZBB041) had limited growth under similar conditions.
This result supports the proposal that azotobactin is provid-
ing the source of nitrogen to sustain growth in the media.
Co-culture of A. vinelandii and S. sp. BA032
In addition to using the spent media of A. vinelandii, we
also wished to study the potential to grow A. vinelandii
and S. sp. BA032 as a co-culture. In this experiment, cells
of A. vinelandii wild-type or the individual gene substitu-
tion strains were inoculated along with S. sp. BA032 into
sterile media and grown together. As shown in Fig. 3
(bottom), a similar result was found to that shown in
Fig. 3 (top), where the strain containing the substitution
for the gene involved in the biosynthetic pathway to pro-
duce azotobactin (A. vinelandii AZBB041) resulted in
very minimal growth, while A. vinelandii wild-type cells
reached a similar level of growth as was found when
using filtered supernatants. The strain containing the gene
substitution of catechol siderophores (A. vinelandii
AZBB040) was able to support S. sp. BA032, although the
levels of cells obtained in the culture were not as high as
when only isolated supernatant was utilized.
Growth of N. oleoabundans in liquid culture
containing siderophore
The second strain that grew well on plates supplemented
with A. vinelandii siderophores as the primary nitrogen
Fig. 3. Growth of various consortiums using atmospheric nitrogen as
the nitrogen source and sucrose as the reduced carbon source for
Azotobacter vinelandii. Supernatants collected from growths of
A. vinelandii strains were filtered through a 0.2-mm filter and used to
grow Scenedesmus sp. BA032 (Top). These results are compared to a
positive control of the same media (not grown with any A. vinelandii
strains) supplemented with 700 µM NaNO3 and a negative control of
the same media with no added nitrogen. Additionally, A. vinelandii
strains were grown together with S. sp. BA032 in co-culture
(Bottom). Each data point represents the average and standard
deviation from three independent samples.
FEMS Microbiol Lett 351 (2014) 70–77ª 2013 Federation of European Microbiological Societies.Published by John Wiley & Sons Ltd. All rights reserved
74 J.A. Villa et al.

source was N. oleoabundans. As was found for S. sp.
BA032, supernatants of the wild-type and catechol sidero-
phore gene substitution strains (A. vinelandii AZBB040)
were able to support the growth of N. oleoabundans to
comparable cell densities, while the supernatant of the
strain carrying the azotobactin gene substitution resulted
in a limited growth (Fig. 4). These results further support
the likelihood that azotobactin is responsible for meeting
the nitrogen requirements in N. oleoabundans as well.
Quantities of siderophore available to culture
algae
The production of siderophores in A. vinelandii has been
studied for many years and is well characterized, includ-
ing the enzymatic pathways and genes involved in their
biosynthesis (Tindale et al., 2000; Page et al., 2003;
Wichard et al., 2009; Yoneyama et al., 2011). In many
reports, the production of specific siderophores under
laboratory culture conditions has been found to be very
high [40–60 mg L�1 of various azotobactin forms
(Demange et al., 1988)]. Based on the absorbance values
obtained in these studies (See Fig. 2 as an example) and a
molecular weight of c. 1370 g mol�1, our yields of azot-
obactin were calculated to be around 25–50 mg L�1 in
various cultures, which agrees well with previous reports
(Demange et al., 1988). Based on the molecular formula
of azotobactin 87 (C55H79N14O27), this constitutes a con-
siderable number of nitrogen equivalents. From the start-
ing quantities of azotobactin in supernatants used to
grow S. sp. BA032 (c. 25 µM for A. vinelandii wild-type
strain and AZBB040 gene substitution strain, Fig. 3 top)
and the positive nitrogen control containing 700 µM of
sodium nitrate, the number of cells obtained with S. sp.
BA032 correlated well with the amount of nitrogen pro-
vided from either source, indicating that S. sp. BA032
was able to utilize essentially all of the provided azotobac-
tin to support growth. Thus, siderophores constitute a
source of nitrogen for the proliferation of nondiazotroph-
ic microalgae organisms when excreted into the extracel-
lular space. Azotobactin is composed of a small peptide
chain (Fig. 1) synthesized via a nonribosomal peptide
synthetase (Yoneyama et al., 2011). Due to the extensive
number of nitrogen atoms found in an azotobactin mole-
cule, even smaller molar concentrations of this compound
can account for a significant mass amount of available
nitrogen when released to the environment. Based on the
potential for diffusion and previous reports of other
organisms hijacking foreign siderophores for their own
benefit (Amin et al., 2009; D’Onofrio et al., 2010), it was
of interest to demonstrate whether siderophores from
Azotobacter would also serve to satisfy nitrogen require-
ments to support other organisms in the environment.
Azotobacter is a bacterium predominantly associated
with soils, while Scenedesmus is a ubiquitous green algae
found in freshwater environments. Thus, there are some
differences between the conditions where these strains are
found. The light utilized to culture the algae strains could
also potentially lead to photodegradation of the sidero-
phores. Even if this were the case, the final fate of the
nitrogen appears to be suitable for the culture of the algae
strains selected here, which represent only a small sam-
pling of potential strains of phototrophs from the envi-
ronment that might make use of the excreted nitrogen
siderophore compounds provided by a diazotroph.
Implications of findings
Under the current experimental design, the cultures tested
here are best defined as commensal co-cultures and not
symbiotic, as A. vinelandii is providing a nitrogen source,
but is not getting anything in return from the algal strain.
Azotobacter vinelandii grows on simple media requiring
only sugar and minor amounts of additional minerals.
Because the sugar provided to support A. vinelandii is an
external source, this relationship is currently an
open-loop system. However, many strains of algae and
cyanobacteria are known to produce extracellular reduced
Fig. 4. Growth of various consortiums using atmospheric nitrogen as
the nitrogen source and sucrose as the reduced carbon source for
Azotobacter vinelandii. Supernatants collected from growths of
A. vinelandii strains were filtered through a 0.2-µm filter and used to
grow Neochloris oleoabundans. Each data point represents the
average and standard deviation from three independent samples.
FEMS Microbiol Lett 351 (2014) 70–77 ª 2013 Federation of European Microbiological Societies.Published by John Wiley & Sons Ltd. All rights reserved
Siderophores support algal growth 75

carbon compounds that could support the growth of
A. vinelandii (Brechignac & Schiller, 1992; Wolf, 1997; De
Philippis & Vincenzini, 1998; Corzo et al., 2000; Bar-Zeev
et al., 2009) and could be used as a secondary selection
pressure. Under such conditions, the two strains could be
grown in such a manner that a symbiotic relationship
might evolve. Examples of phototrophs producing extra-
cellular carbon that is released to the environment in the
form of polysaccharides and simple sugars are common,
although the exact reason for the energetically wasteful
excretion of extracellular reduced carbon is not clear
(Passow, 2002; Bar-Zeev et al., 2009). This research pro-
vides clear evidence that the nitrogen released to the envi-
ronment as siderophores can be utilized by other
organisms not only for obtaining metals from the envi-
ronment (D’Onofrio et al., 2010), but also to provide
requisite nitrogen required for growth and replication in
some strains of green algae.
Acknowledgements
This work is supported by a grant (RC-0007-12) from the
Initiative for Renewable Energy & the Environment (Insti-
tute on the Environment) to B.M.B and the Biotechnology
Institute at the University of Minnesota for fellowship
funding to J.A.V. We thank Jiashi Wei for assistance in
early isolation of siderophores and preliminary algal cul-
ture studies. We thank the kind suggestions of the anony-
mous reviewers.
References
Amin SA, Green DH, Hart MC, K€upper FC, Sunda WG &
Carrano CJ (2009) Photolysis of iron-siderophore chelates
promotes bacterial-algal mutualism. P Natl Acad Sci USA
106: 17071–17076.Bali A, Blanco G, Hill S & Kennedy C (1992) Excretion of
ammonium by a nifL mutant of Azotobacter vinelandii fixing
nitrogen. Appl Environ Microbiol 58: 1711–1718.Barney BM, Lee HI, Dos Santos PC, Hoffmann BM, Dean DR
& Seefeldt LC (2006) Breaking the N-2 triple bond: insights
into the nitrogenase mechanism. Dalton Trans 2277–2284.Bar-Zeev E, Berman-Frank I, Stambler N et al. (2009)
Transparent exopolymer particles (TEP) link phytoplankton
and bacterial production in the Gulf of Aqaba. Aquat
Microb Ecol 56: 217–225.Bold HC (1949) The morphology of Chlamydomonas
chlamydogama sp. nov. Bull Torrey Bot Club 76: 101–108.Brechignac F & Schiller P (1992) Pilot CELSS based on a
maltose-excreting Chlorella: concept and overview on the
technological developments. Adv Space Res 12: 33–36.Brewin B, Woodley P & Drummond M (1999) The basis of
ammonium release in nifL mutants of Azotobacter
vinelandii. J Bacteriol 181: 7356–7362.
Cornish AS & Page WJ (1995) Production of the
triacetecholate siderophore protochelin by Azotobacter
vinelandii. Biometals 8: 332–338.Corzo A, Morillo JA & Rodr�ıguez S (2000) Production of
transparent exopolymer particles (TEP) in cultures of
Chaetoceros calcitrans under nitrogen limitation. Aquat
Microb Ecol 23: 63–72.De Philippis R & Vincenzini M (1998) Exocellular
polysaccharides from cyanobacteria and their possible
applications. FEMS Microbiol Rev 22: 151–175.Demange P, Bateman A, Dell A & Abdallah MA (1988)
Structure of azotobactin D, a siderophore of Azotobacter
vinelandii strain D (CCM 289). Biochemistry 27: 2745–2752.D’Onofrio A, Crawford JM, Stewart EJ, Witt K, Gavrish E,
Epstein S, Clardy J & Lewis K (2010) Siderophores from
neighboring organisms promote the growth of uncultured
bacteria. Chem Biol 17: 254–264.Fekete FA, Spence JT & Emery T (1983) Siderophores
produced by nitrogen-fixing Azotobacter vinelandii OP in
iron-limited continuous culture. Appl Environ Microbiol 46:
1297–1300.Gimmestad M, Steigedal M, Ertesvag H, Moreno S,
Christensen BE, Esp�ın G & Valla S (2006) Identification and
characterization of an Azotobacter vinelandii type I secretion
system responsible for export of the AlgE-Type mannuronan
C-5-epimerases. J Bacteriol 188: 5551–5560.Gouveia L, Marques AE, da Silva TL & Reis A (2009)
Neochloris oleabundans UTEX #1185: a suitable renewable
lipid source for biofuel production. J Ind Microbiol
Biotechnol 36: 821–826.Huyer M & Page WJ (1988) Zn2+ increases siderophore
production in Azotobacter vinelandii. Appl Environ Microbiol
54: 2625–2631.Ikeda S, Okubo T, Anda M et al. (2010) Community- and
genome-based views of plant-associated bacteria:
plant-bacterial interactions in soybean and rice. Plant Cell
Physiol 51: 1398–1410.Jones KM, Kobayashi H, Davies BW, Taga ME & Walker GC
(2007) How rhizobial symbionts invade plants: the
Sinorhizobium-Medicago model. Nat Rev Microbiol 5:
619–633.Li Y, Horsman M, Wang B, Wu N & Lan CQ (2008) Effects of
nitrogen sources on cell growth and lipid accumulation of
green alga Neochloris oleoabundans. Appl Microbiol
Biotechnol 81: 629–636.Mather MW, McReynolds LM & Yu CA (1995) An enhanced
broad-host-range vector for Gram-negative bacteria:
avoiding tetracycline phototoxicity during the growth of
photosynthetic bacteria. Gene 156: 85–88.Ortiz-Marquez JCF, Nascimento MD, Dublan Mde L &
Curatti L (2012) Association with an ammonium-excreting
bacterium allows diazotrophic culture of oil-rich eukaryotic
microalgae. Appl Environ Microbiol 78: 2345–2352.Page WJ & Huyer M (1984) Derepression of the Azotobacter
vinelandii siderophore system, using iron-containing
minerals to limit iron repletion. J Bacteriol 158: 496–502.
FEMS Microbiol Lett 351 (2014) 70–77ª 2013 Federation of European Microbiological Societies.Published by John Wiley & Sons Ltd. All rights reserved
76 J.A. Villa et al.

Page WJ & Sadoff HL (1975) Relationship between calcium
and uroinic acids in the encystment of Azotobacter
vinelandii. J Bacteriol 122: 145–151.Page WJ, Collinson SK, Demange P, Dell A & Abdallah MA
(1991) Azotobacter vinelandii strains of disparate origin
produce azotobactin siderophores with identical structures.
Biol Met 4: 217–222.Page WJ, Kwon E, Cornish AS & Tindale AE (2003) The csbX
gene of Azotobacter vinelandii encodes an MFS efflux pump
required for catecholate siderophore export. FEMS Microbiol
Lett 228: 211–216.Palanch�e T, Blanc S, Hennard C, Abdallah MA &
Albrecht-Gary AM (2004) Bacterial iron transport:
coordination properties of azotobactin, the highly
fluorescent siderophore of Azotobacter vinelandii. Inorg
Chem 43: 1137–1152.Passow U (2002) Production of transparent exopolymer
particles (TEP) by phyto- and bacterioplankton. Mar Ecol
Prog Ser 236: 1–12.Prentki P & Krisch HM (1984) In vitro insertional mutagenesis
with a selectable DNA fragment. Gene 29: 303–313.Rodr�ıguez-L�opez JN, Serna-Rodr�ıguez P, Tudela J, Var�on R &
Garc�ıa-C�anovas F (1991) A continuous spectrophotometric
method for the determination of diphenolase activity of
tyrosinase using 3,4-dihydroxymandelic acid. Anal Biochem
195: 369–374.Setubal JC, dos Santos P, Goldman BS et al. (2009) Genome
sequence of Azotobacter vinelandii, an obligate aerobe
specialized to support diverse anaerobic metabolic processes.
J Bacteriol 191: 4534–4545.Tindale AE, Mehrotra M, Ottem D & Page WJ (2000) Dual
regulation of catecholate siderophore biosynthesis in
Azotobacter vinelandii by iron and oxidative stress.
Microbiology 146: 1617–1626.Wichard T, Bellenger JP, Morel FMM & Kraepiel AML (2009)
Role of the siderophore azotobactin in the bacterial
acquisition of nitrogenase metal cofactors. Environ Sci
Technol 43: 7218–7224.Wolf L (1997) Bioregeneration with maltose excreting
Chlorella: system concept, technological development, and
experiments. Adv Space Biol Med 6: 255–274.Yoneyama F, Yamamoto M, Hashimoto W & Murata K (2011)
Azotobacter vinelandii gene clusters for two types of peptidic
and catechol siderophores produced in response to
molybdenum. J Appl Microbiol 111: 932–938.
FEMS Microbiol Lett 351 (2014) 70–77 ª 2013 Federation of European Microbiological Societies.Published by John Wiley & Sons Ltd. All rights reserved
Siderophores support algal growth 77