ars.els-cdn.com · web view2016-02-04 · the overall probability that all of n nitrogen atoms in...
TRANSCRIPT

Supplementary Information
Absolute quantification of Corynebacterium glutamicum glycolytic and anaplerotic enzymes by QconCAT
Raphael Voges1, Stephanie Corsten1, Wolfgang Wiechert1, Stephan Noack*1
Table S1: Proteins, peptides, mass transitions and measurement parameters for C. glutamicum central carbon metabolism enzymes.
Protein Parent ionm/z [Da]
Measured product ions(m/z [Da], CE [V]) Sequence Charge
[-]DP[V]
PFK(cg1409)
740.9 y12 (1181.7, 35.0), y9 (865.6, 33.0), y7 (695.4, 37.0) WLSDNGIPVVGVPK 2 85.1517.3 y8 (934.5, 24.2), y7 (803.4, 21.2), y6 (690.4, 26.2) VMIVEVMGR 2 68.8372.7 y6 (673.4, 19.0), y5 (616.4, 19.0), y4 (503.3, 20.0) AGIDQIK 2 58.3
FBA(cg3068)
771.4 y9 (1115.5, 35.7), y8 (952.4, 37.7), y7 (851.4, 35.7) AGANLYTSPEDFEK 2 87.3618.3 y9 (993.6, 30.0), y8 (864.5, 27.0), y7 (793.5, 27.0) IEEALTYGVIK 2 76.2502.8 y8 (790.4, 21.4), y7 (675.4, 24.4), y6 (604.3, 24.4) TIDAIGTGEK 2 67.8
PGK(cg1790)
858.5 y13 (1315.7, 40.7), y10 (1018.6, 39.7), y7 (659.4, 39.7) IAESPEAPYVVVLGGSK 2 93.7492.8 y9 (871.5, 24.8), y7 (715.4, 23.8), y6 (602.4, 23.8) IGVIEALAAK 2 67.0533.3 y8 (804.4, 23.1), y7 (747.4, 24.1), y6 (618.4, 23.1) NFGEVLSTAK 2 70.0
PGM(cg0482)
769.9 y8 (1032.5, 42.6), y7 (885.5, 42.6), y6 (756.4, 42.6) FVPYFEEEILPR 2 87.2565.3 y7 (730.4, 24.0), y6 (659.3, 27.0), y5 (546.3, 25.0) TANIALNAADR 2 72.3524.3 y7 (813.4, 24.6), y6 (698.4, 27.6), y5 (585.3, 24.6) YADLDVVPR 2 69.3
ENO(cg1111)
594.8 y8 (860.5, 30.6), y7 (773.5, 27.6), y6 (702.4, 25.6) AAADSAGLPLFR 2 74.5408.3 y7 (744.5, 22.0), y6 (673.4, 19.0), y5 (559.4, 20.0) AANSILVK 2 60.9403.7 y5 (643.4, 22.7), y4 (529.3, 18.7), y3 (401.3, 20.7) YNQLLR 2 60.5
PK(cg2291)
591.3 y10 (1011.6, 29.4), y8 (841.5, 29.4), y7 (728.4, 24.4) AVGILADLQGPK 2 74.2426.7 y6 (696.4, 17.1), y5 (583.3, 19.1), y4 (496.3, 23.1) GVISYSAR 2 62.2425.2 y6 (736.3, 24.0), y5 (605.3, 19.0), y4 (490.2, 25.0) IMDEEGR 2 62.1
AceE(cg2466)
561.8 y8 (910.5, 24.8), y6 (700.4, 31.8), y5 (587.3, 31.8) LVPIIPDEAR 2 72.1481.8 y8 (758.5, 21.2), y7 (645.4, 23.2), y6 (532.3, 23.2) GFLLGATAGR 2 72.1436.2 y5 (701.4, 18.6), y4 (538.3, 23.6), y3 (425.2, 20.6) GIYLYSR 2 66.2
PEPCx(cg1787)
721.4 y6 (765.4, 26.4), y5 (636.4, 24.4), y3 (480.3, 24.4) ILATTAELIGEDAVEGVWFK 3 83.7447.8 y7 (781.4, 23.3), y6 (680.4, 19.3), y5 (593.3, 24.3) LTSFDIAK 2 63.8403.7 y5 (692.4, 18.7), y3 (416.3, 18.7) NYLLQR 2 60.5
PEPCk(cg3169)
695.4 y8 (830.5, 34.4), y7 (731.4, 31.4), y6 (602.4, 32.4) MAEDLVEAGTLIK 2 81.8648.3 y8 (849.4, 29.7), y7 (752.3, 37.7), y6 (695.3, 32.7) FLWPGFGDNSR 2 78.4438.7 y7 (745.4, 23.7), y5 (575.3, 22.7), y4 (446.3, 22.7) MGIEALDK 2 63.1
PCx(cg0761)
824.9 y12 (1202.6, 38.8), y9 (829.4, 37.8), y6 (584.3, 43.8) GLYLPFESGTPGPTGR 2 91.3
670.3 y7 (867.5, 28.9), y6 (766.4, 28.9), y5 (603.4, 29.9) SAEGQTYPIVFK 2 80.0433.7 y6 (632.3, 21.5), y5 (561.3, 22.5), y4 (474.3, 22.5) SFASEAVR 2 62.7
1

Table S2: Correction for isotopic impurities in heavy QconCAT and light sample peptides based on [1]. The overall probability that all of n nitrogen atoms in a QconCAT peptide are 15N-labeled is approximated as PQCC(0; n , pQCC
14 N ), depending on the probability pQCC14N =0.01 for
impurities of the tracer 15NH4SO4 (manufacturer instruction). In contrast, Psample (0 ;n , psample15N )
denotes the overall probability that all nitrogen atoms in the corresponding sample peptide are naturally 14N-labelled, depending on the probability psample
15N =0.00366 for naturally occurring 15N-isotopes. The correction factor ω is calculated as the quotient of both overall probabilities.
Peptide sequence n PQCC(0; n , pQCC14 N ) Psample (0 ;n , psample
15N ) ω
WLSDNGIPVVGVPK 17 0.8429 0.9396 0.8972VMIVEVMGR 10 0.9044 0.9640 0.9382AGIDQIK 9 0.9135 0.9675 0.9442AGANLYTSPEDFEK 16 0.8515 0.9430 0.9029IEEALTYGVIK 12 0.8864 0.9570 0.9263TIDAIGTGEK 11 0.8953 0.9605 0.9322IAESPEAPYVVVLGGSK 18 0.8345 0.9361 0.8915IGVIEALAAK 11 0.8953 0.9605 0.9322NFGEVLSTAK 12 0.8864 0.9570 0.9263FVPYFEEEILPR 13 0.8775 0.9535 0.9204TANIALNAADR 13 0.8775 0.9535 0.9204YADLDVVPR 10 0.9044 0.9640 0.9382AAADSAGLPLFR 13 0.8775 0.9535 0.9204AANSILVK 10 0.9044 0.9640 0.9382YNQLLR 9 0.9135 0.9675 0.9442AVGILADLQGPK 14 0.8687 0.9500 0.9145GVISYSAR 9 0.9135 0.9675 0.9442IMDEEGR 8 0.9227 0.9711 0.9502LVPIIPDEAR 11 0.8953 0.9605 0.9322GFLLGATAGR 11 0.8953 0.9605 0.9322GIYLYSR 8 0.9227 0.9711 0.9502ILATTAELIGEDAVEGVWFK 22 0.9225 0.8016 0.8690LTSFDIAK 9 0.9135 0.9675 0.9442NYLLQR 9 0.9135 0.9675 0.9442MAEDLVEAGTLIK 14 0.8687 0.9500 0.9145FLWPGFGDNSR 14 0.8687 0.9500 0.9145MGIEALDK 9 0.9135 0.9675 0.9442GLYLPFESGTPGPTGR 17 0.8429 0.9396 0.8972SAEGQTYPIVFK 14 0.8687 0.9500 0.9145SFASEAVR 9 0.9135 0.9675 0.9442
2

Table S3: Variables and constants for absolute protein quantification.
Symbol Denotation DimensionmX Biomass at sampling point mgCDW
N cell Cell number at sampling point counts
V cell Biovolume at sampling point µl
N A Avogadro constant (6.022 * 1023) mol-1
MW P Molecular weight of native protein (various) g mol-1
MW QCC Molecular weight of QCC protein (41225.1) g mol-1
V 1 Volume of crude extract (CE) in digest l
V 2 Volume of QCC stock solution in digest l
V 3 Total volume of digest/ LC-MS-sample l
V 4 Total volume of lysis buffer = total vol. of CE l
A14 N Native peptide peak area (14N) counts
A15N QCC peptide peak area (15N) counts
ω Correction factor for isotopic impurities -
cbradford15N Concentration of purified QCC stock solution µg ml-1
cstock15N Molar concentration of purified QCC stock solution nM
csample15N Concentration of QCC peptide in LCMS-sample (known) nM
csample14N Concentration of native peptide in LCMS-sample (unknown) nM
cCE14N Concentration of native peptide in CE nM
c P Biomass specific concentration of native protein mg gCDW-1
λP Native protein copy number per cell copies cell-1
c P,intra Intracellular concentration of native protein µM
3
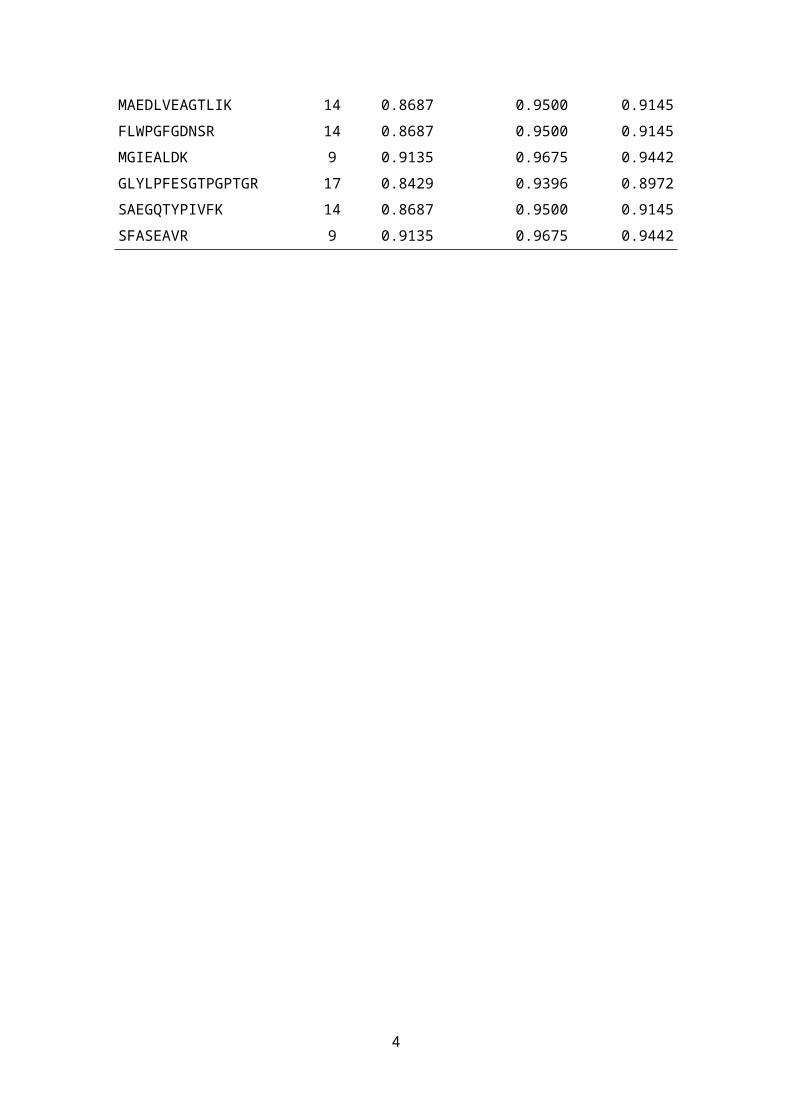
Figure S1: QconCat-workflow for absolute protein quantification. A-C: Schematic overview of sample preparation procedure from biomass sample to final LC-MS/MS sample. D: Equations for absolute protein quantification. For definition of variables see table S3.
4

Figure S2: Quantification of Q-peptides after prolonged lysis by bead beating. Relative peptide amounts were derived as described in [2]. Vertical straight lines indicate ± 2fold threshold for significant change of peptide amounts.
5

Figure S3: Final biomass (OD) and cultivation time for QconCAT expression strains E. coli BL21 pBAD::EAQCC and E. coli TOP10 pBAD::EAQCC in 7 defined media and complex LB medium.
Figure S4: Expression of QconCAT protein standard for C. glutamicum glycolytic and anaplerotic key enzymes in E. coli BL21 pBAD::EAQCC grown in 1 l 15N-labeled LR medium at 37°C, pH 7.0 (adjusted by the addition of 4 M sodium hydroxide) and 30 % dissolved oxygen (60 standard liter min-1, 400-1200 rpm) in a 1.8 l lab-scale bioreactor. Heterologous protein expression was induced at OD = 1.0 with 10 g l-1 L-arabinose and cells were harvested 3 h after induction.
6

Figure S5: Quantification of purified QconCAT using a modified Bradford protocol for low protein concentrations. Dotted horizontal line represents buffer extinction, dashed horizontal and vertical lines indicate concentration of purified QconCAT solution. 40 µl sample or BSA standard were mixed with 200 µl Bradford reagent (ratio of sample to Bradford solution increased from 1:40 to 1:5), incubated at room temperature for 5 min and measured at 595 nm.
7

Table S4: Prediction of missed proteolytic cleavages for the Q-peptides in the QconCAT and in their native sequence background, respectively. Following the approach of Lawless and Hubbard [3], all values represent the scores when applying the support vector machine method. Scores that are above a binary threshold value of 0.5 (shown in bold) are assumed to represent missed proteolytic cleavages.
QconCAT Native sequence background
Peptide sequence N-terminal C-terminal N-terminal C-terminalMGGSHHHHHHGMASMTGGQQMGR - 0.46 - -
DLYDDDDK 0.46 1.00 - -
WGSELER 0.93 0.51 - -
APHAELR 0.77 0.45 - -
WLSDNGIPVVGVPK 0.42 0.38 0.50 0.34VMIVEVMGR 0.38 0.43 0.45 0.42AGIDQIK 0.43 0.39 0.59 0.40AGANLYTSPEDFEK 0.39 0.68 0.37 0.40IEEALTYGVIK 0.68 0.39 1.00 0.39TIDAIGTGEK 0.39 0.43 0.40 0.46IAESPEAPYVVVLGGSK 0.43 0.40 0.40 0.46IGVIEALAAK 0.40 0.37 0.72 0.58NFGEVLSTAK 0.37 0.39 0.43 0.40FVPYFEEEILPR 0.39 0.42 0.41 0.74TANIALNAADR 0.42 0.47 0.65 0.60YADLDVVPR 0.47 0.41 0.39 0.45AAADSAGLPLFR 0.41 0.40 0.38 0.37AANSILVK 0.40 0.37 0.71 0.38YNQLLR 0.37 0.40 0.44 0.48AVGILADLQGPK 0.40 0.40 0.58 0.40GVISYSAR 0.40 0.43 0.95 0.50IMDEEGR 0.43 0.50 0.42 0.64LVPIIPDEAR 0.50 0.46 0.52 0.45GFLLGATAGR 0.46 0.44 0.43 0.42GIYLYSR 0.44 0.43 0.40 0.47ILATTAELIGEDAVEGVWFK 0.43 0.43 0.56 0.40LTSFDIAK 0.43 0.36 0.46 0.45NYLLQR 0.36 0.41 0.42 0.47MAEDLVEAGTLIK 0.41 0.41 0.52 0.44FLWPGFGDNSR 0.41 0.44 0.46 0.56MGIEALDK 0.44 0.49 0.48 0.54GLYLPFESGTPGPTGR 0.49 0.40 0.44 0.44SAEGQTYPIVFK 0.40 0.39 0.42 0.36SFASEAVR 0.39 0.51 0.42 0.46APHAELR 0.77 0.45 - -
WSHPQFEK 0.45 - - -
8

9

Figure S6: Determination of linear ranges for measurement of tryptic digested sample and QconCAT-peptides on a 4000 QTRAP LC-MS/MS device.
10

Table S5: Model-assisted evaluation of obtained proteomics data from QconCat approach. Published in vitro turnover numbers (k cat , Lit) and enzyme activities (vmax, Lit) were combined with actually determined intracellular protein concentrations (c P,intra) and metabolic fluxes (vFVA) to estimate a priori unknown turnover numbers (k cat ,QCC ) or maximal catalytic enzyme capacities (vmax,QCC).
k cat , Lit[s-1]
vmax, Lit[mM s-1]
ref. c P,intra[µM]
vFVA [mM s-1]
k cat ,QCC[s-1]
vmax ,QCC [mM s-1]
PFK - 0.04 [4] [5.20, 9.58] [0.31, 0.45] [4, 7] -
FBA [2.95, 21]a 0.11 [4-6] 32.50 [0.31, 0.43] - [0.10, 0.68]
PGK - 0.75 [7] [19.14, 41.77] [0.70, 0.93] [18, 39] -
PGM - 2.26 [8] [36.09, 119.65] [0.65, 0.87] [19, 63] -
ENO - - [68.47, 133.19] [0.65, 0.87] - -
PK 25401.64;1.81;3.29
[4, 9-11] [24.17, 208.16] [0.04, 0.27] - [61.47, 528.83]
AceE -0.14;0.16;0.24
[10, 12, 13] [48.52, 93.24] [0.36, 0.49] [1, 5] -
PEPCx -
0.05;0.33;0.54;0.55
[7, 10, 11, 14] [14.22, 18.18] [0.00, 0.20] [3, 38] -
PEPCk [6.1, 7.2]0.17;0.20;0.25
[11, 14-16] [9.00, 13.54] [0.00, 0.02] - [0.06, 0.10]
PCx -0.15;0.23;0.45
[17, 18] [12.28, 27.86] [0.00, 0.20] [5, 36] -
ataken from Bacillus cereus and Mycobacterium tuberculosis.
QconCAT sequence:
Molecule: pBADHisA_EMP_ANA_Expr, 5123 bps DNA CircularDescription: pBADHisA E. coli expression vector version C, His tag in frame A with
QconCAT for EMP and ANA of C. glutamicum
Details: QconCat Protein for EMP and ANA in C. glutamicumTranslation product 379 aaMol Wt 41225.1, Isoelectric Pt (pI) 5.01
Translation:
MGGSHHHHHHGMASMTGGQQMGRDLYDDDDKDRWGSELERAPHAELRWLSDNGIPVVGVPKVMIVEVMGRAGIDQIKAGANLYTSPEDFEKIEEALTYGVIKTIDAIGTGEKIAESPEAPYVVVLGGSKIGVIEALAAKNFGEVLSTAKFVPYFEEEILPRTANIALNAADRYADLDVVPRAAADSAGLPLFRAANSILVKYNQLLRAVGILADLQGPKGVISYSARIMDEEGRLVPIIPDEARGFLLGATAGRGIYLYSRILATTAELIGEDAVEGVWFKLTSFDIAKNYLLQRMAEDLVEAGTLIKFLWPGFGDNSRMGIEALDKGLYLPFESGTPGPTGRSAEGQTYPIVFKSFASEAVRRAPHAELRWSHPQFEK
11

References
[1] Eibl G, Bernardo K, Koal T, Ramsay SL, Weinberger KM, Graber A. Isotope correction of mass spectrometry profiles. Rapid Commun Mass Spectrom. 2008;22:2248-52.[2] Voges R, Noack S. Quantification of proteome dynamics in Corynebacterium glutamicum by 15N-labeling and selected reaction monitoring. J Proteomics. 2012;75:2660-9.[3] Lawless C, Hubbard SJ. Prediction of missed proteolytic cleavages for the selection of surrogate peptides for quantitative proteomics. Omics : a journal of integrative biology. 2012;16:449-56.[4] Engels V, Lindner SN, Wendisch VF. The global repressor SugR controls expression of genes of glycolysis and of the L-lactate dehydrogenase LdhA in Corynebacterium glutamicum. J Bacteriol. 2008;190:8033-44.[5] Ramsaywak PC, Labbe G, Siemann S, Dmitrienko GI, Guillemette JG. Molecular cloning, expression, purification, and characterization of fructose 1,6-bisphosphate aldolase from Mycobacterium tuberculosis - a novel Class II A tetramer. Protein expression and purification. 2004;37:220-8.[6] Labbe G, de Groot S, Rasmusson T, Milojevic G, Dmitrienko GI, Guillemette JG. Evaluation of four microbial Class II fructose 1,6-bisphosphate aldolase enzymes for use as biocatalysts. Protein expression and purification. 2011;80:224-33.[7] Inui M, Suda M, Okino S, Nonaka H, Puskas LG, Vertes AA, et al. Transcriptional profiling of Corynebacterium glutamicum metabolism during organic acid production under oxygen deprivation conditions. Microbiology. 2007;153:2491-504.[8] Seibold GM, Eikmanns BJ. Inactivation of the phosphoglucomutase gene pgm in Corynebacterium glutamicum affects cell shape and glycogen metabolism. Bioscience reports. 2013;33.[9] Jetten MS, Gubler ME, Lee SH, Sinskey AJ. Structural and functional analysis of pyruvate kinase from Corynebacterium glutamicum. Appl Environ Microbiol. 1994;60:2501-7.[10] Cocaign-Bousquet M, Guyonvarch A, Lindley ND. Growth Rate-Dependent Modulation of Carbon Flux through Central Metabolism and the Kinetic Consequences for Glucose-Limited Chemostat Cultures of Corynebacterium glutamicum. Appl Environ Microbiol. 1996;62:429-36.[11] Sawada K, Zen-in S, Wada M, Yokota A. Metabolic changes in a pyruvate kinase gene deletion mutant of Corynebacterium glutamicum ATCC 13032. Metab Eng. 2010;12:401-7.[12] Buchholz J, Schwentner A, Brunnenkan B, Gabris C, Grimm S, Gerstmeir R, et al. Platform engineering of Corynebacterium glutamicum with reduced pyruvate dehydrogenase complex activity for improved production of L-lysine, L-valine, and 2-ketoisovalerate. Appl Environ Microbiol. 2013;79:5566-75.[13] Schreiner ME, Fiur D, Holatko J, Patek M, Eikmanns BJ. E1 enzyme of the pyruvate dehydrogenase complex in Corynebacterium glutamicum: molecular analysis of the gene and phylogenetic aspects. J Bacteriol. 2005;187:6005-18.[14] Peters-Wendisch PG, Eikmanns BJ, Thierbach G, Bachmann B, Sahm H. Phosphoenolpyruvate carboxylase in Corynebacterium glutamicum is dispensable for growth and lysine production. FEMS Microbiol Lett. 1993;112:269-74.[15] Riedel C, Rittmann D, Dangel P, Mockel B, Petersen S, Sahm H, et al. Characterization of the phosphoenolpyruvate carboxykinase gene from Corynebacterium glutamicum and significance of the enzyme for growth and amino acid production. J Mol Microbiol Biotechnol. 2001;3:573-83.[16] Aich S, Imabayashi F, Delbaere LTJ. Expression, purification, and characterization of a bacterial GTP-dependent PEP carboxykinase. Protein expression and purification. 2003;31:298-304.[17] Peters-Wendisch PG, Schiel B, Wendisch VF, Katsoulidis E, Mockel B, Sahm H, et al. Pyruvate carboxylase is a major bottleneck for glutamate and lysine production by Corynebacterium glutamicum. J Mol Microbiol Biotechnol. 2001;3:295-300.[18] Koffas MA, Jung GY, Aon JC, Stephanopoulos G. Effect of pyruvate carboxylase overexpression on the physiology of Corynebacterium glutamicum. Appl Environ Microbiol. 2002;68:5422-8.
12