arabidopsis kinesin kp1 specifically interacts with vdac3, a … · colombini, 1979; liu and...
TRANSCRIPT

Arabidopsis Kinesin KP1 Specifically Interacts with VDAC3, aMitochondrial Protein, and Regulates Respiration during SeedGermination at Low Temperature W OA
Xue-Yong Yang,1 Zi-Wei Chen,1 Tao Xu, Zhe Qu, Xiao-Di Pan, Xing-Hua Qin, Dong-Tao Ren,2 and Guo-Qin Liu2,3
State Key Laboratory of Plant Physiology and Biochemistry, College of Biological Sciences, China Agricultural University, Beijing
100193, China
The involvement of cytoskeleton-related proteins in regulating mitochondrial respiration has been revealed in mammalian
cells. However, it is unclear if there is a relationship between the microtubule-based motor protein kinesin and mitochon-
drial respiration. In this research, we demonstrate that a plant-specific kinesin, Kinesin-like protein 1 (KP1; At KIN14 h), is
involved in respiratory regulation during seed germination at a low temperature. Using in vitro biochemical methods and in
vivo transgenic cell observations, we demonstrate that KP1 is able to localize to mitochondria via its tail domain (C
terminus) and specifically interacts with a mitochondrial outer membrane protein, voltage-dependent anion channel 3
(VDAC3). Targeting of the KP1-tail to mitochondria is dependent on the presence of VDAC3. When grown at 48C, KP1dominant-negative mutants (TAILOEs) and vdac3 mutants exhibited a higher seed germination frequency. All germinating
seeds of the kp1 and vdac3 mutants had increased oxygen consumption; the respiration balance between the cytochrome
pathway and the alternative oxidase pathway was disrupted, and the ATP level was reduced. We conclude that the plant-
specific kinesin, KP1, specifically interacts with VDAC3 on the mitochondrial outer membrane and that both KP1 and VDAC3
regulate aerobic respiration during seed germination at low temperature.
INTRODUCTION
Much of the aerobic oxidation in eukaryotic cells takes place in
mitochondria. A number of studies have shown that microfila-
ments and microtubules function in mitochondrial movement
and positioning in eukaryotic cells (Hirokawa, 1998). Cytoskel-
etal proteins are also involved in regulating the permeability
of the mitochondrial outer membrane to ADP in animal cells
(Rappaport et al., 1998; Saks et al., 1995). It is well known that
the membrane permeability of mitochondria is mainly depen-
dent on the voltage-dependent anion channel (VDAC) (also
named as a porin), the most abundant integral membrane
protein in the mitochondrial outer membrane (Benz, 1994;
Colombini, 1979; Liu and Colombini, 1992). Recently, both
tubulin and actin from human and yeast (Saccharomyces
cerevisiae) cells were found to interact with VDACs (Carre
et al., 2002; Roman et al., 2006). In vitro reconstitution studies
demonstrated that fungal VDACs have two main conductance
states: an open state that allows the diffusion of large metab-
olites, including nucleotides, and a closed state that regulates
ATP flux through the membrane (Rostovtseva and Colombini,
1996). Based on patch-clamp and planar lipid bilayer tech-
niques, Rostovtseva et al. (2008) demonstrated that tubulin
could induce reversible blockage of VDACs and decrease ATP/
ADP permeability through the mitochondrial outer membrane
and that the addition of tubulin in the reaction system could
reduce the respiration rate of mitochondria (Rostovtseva et al.,
2008). Genomic sequence analysis revealed that there are
five VDAC isoforms in Arabidopsis thaliana, four of which have
been cloned and identified (Clausen et al., 2004). However, it is
still not known whether plant VDACs interact with cytoskeletal
proteins.
We identified a plant-specific kinesin member in Arabidopsis,
Kinesin-like protein 1 (KP1) (standardized nomenclature: At
KIN14h; see Malcos and Cyr, 2009), and found that it binds to
mitochondria based on immunoblot analysis (Ni et al., 2005). The
motor domain of KP1 has nucleotide-dependent microtubule
binding ability and microtubule-stimulated ATPase activity (Li
et al., 2007). Kinesins constitute a superfamily of microtubule
motor proteins and play critical roles in the transport of vesicles
and organelles, cytokinesis, morphogenesis, and signal trans-
duction (Reddy, 2001; Verhey et al., 2001; Lee and Liu, 2004;
Hirokawa et al., 2009). Several animal kinesins, such as KIF1B
and KIF5B in mouse cells (Nangaku et al., 1994; Tanaka et al.,
1998) and KLP67A in early Drosophila melanogaster embryos
(Pereira et al., 1997), have been implicated in the movement of
mitochondria. Green fluorescent protein (GFP) fusion and tran-
sient expression assays showed that two Arabidopsis kinesins,
MKRP1 and MKRP2, were expressed in mitochondria via their
N-terminal mitochondrial targeting signals (Itoh et al., 2001). It is
1 These authors contributed equally to this work.2Guo-Qin Liu and Dong-Tao Ren’s groups contributed equally to thiswork.3 Address correspondence to [email protected] author responsible for distribution of materials integral to thefindings presented in this article in accordance with the policy describedin the Instructions for Authors (www.plantcell.org) is: Guo-Qin Liu([email protected]).WOnline version contains Web-only data.OAOpen Access articles can be viewed online without a subscription.www.plantcell.org/cgi/doi/10.1105/tpc.110.082420
The Plant Cell, Vol. 23: 1093–1106, March 2011, www.plantcell.org ã 2011 American Society of Plant Biologists

not currently understood if kinesins are involved in regulating
mitochondrial functions in plant cells.
The membrane-associated electron transport chain of plant
mitochondria has unique features, such as the ubiquitous pres-
ence of a terminal alternative oxidase (AOX), an important
member of the cyanide (CN)-resistant pathway that competes
for electrons with the standard cytochrome pathway (Laties,
1982; Finnegan et al., 2004) and is able to reduce the levels of
reactive oxygen species (Maxwell et al., 1999; Umbach et al.,
2005). Therefore, the respiratory regulation of plant mitochondria
is expected to have unique characteristics. What is the interac-
tion protein of KP1 in the mitochondria? Does the microtubule-
based motor protein, KP1, function in mitochondrial respiration?
It is of crucial importance to elucidate these questions to reveal
the regulation mechanisms of plant mitochondrial respiration. In
this study, we found that KP1 specifically interacts with the
mitochondrial outer membrane protein VDAC3 and that both
KP1 and VDAC3 are involved in keeping the ATP levels stable
and balancing the aerobic respiration pathways during seed
germination at low temperature (48C).
RESULTS
The Tail Domain Is Responsible forMitochondrial Targeting
of KP1
According to our previous work, KP1 is found in isolated mito-
chondria (Ni et al., 2005). Based on the sequence alignment, we
know that the tail domain of KP1 (KP1-tail; 749 to 1087 amino
acids) is specific among all Arabidopsis kinesins. Tail domains
of many animal kinesins are responsible for cargo binding
(Hirokawa et al., 2009). To gain insight into the molecular mech-
anism underlying the interaction between KP1 and mitochon-
dria, GFP-KP1 and its truncated proteins, Dtail (1 to 748 amino
acids) with GFP at its N terminus, and tail (749 to 1087 amino
acids) with GFP at its C terminus (Figure 1A) were transiently
expressed in Arabidopsis protoplasts prepared from suspension
cells. By immunolabeling microtubules and microfilaments in
the transfected protoplasts and treating them with microtubule/
microfilament-depolymerizing drugs, oryzalin and latrunculin B,
respectively, we found that in addition to localizing to dot-like
Figure 1. The Tail Domain Is Responsible for Mitochondrial Targeting of KP1 in Arabidopsis Protoplasts.
(A) GFP fusion constructs of KP1 and its truncated variants (GFP-Dtail and tail-GFP). The domains of KP1 are indicated by head (head domain; 1 to 374
amino acids), motor (motor domain; 375 to 748 amino acids), and tail (tail domain; 749 to 1087 amino acids).
(B) Immunofluorescence labeling of microtubules and microfilaments in Arabidopsis suspension cell protoplasts transiently overexpressing GFP-KP1.
For microtubule labeling, anti-a-tubulin antibody and tetramethylrhodamine b-isothiocyanate–conjugated goat anti-mouse IgG were used. For
microfilaments, rhodamine-phalloidin was used. Bars = 5 mm.
(C) KP1 fusion proteins in suspension cell protoplasts are able to localize to mitochondria via their tail domain. From left to right, green channel
(GFP), red channel (MitoTracker, mitochondria selective reagent), merged images of green and red channel (Merge), and transmission images
(Bright). Bars = 5 mm.
1094 The Plant Cell

organelles, GFP-KP1 localized to microtubules (Figure 1B; see
Supplemental Figure 1 online). Costaining the protoplasts with
the mitochondrion-selective reagent MitoTracker Red revealed
that some dot-like signals of GFP-KP1 colocalized with mito-
chondria (Figure 1C, white arrows). Interestingly, tail-GFP was
located in mitochondria, but GFP-Dtail was distributed randomly
(Figure 1C). This indicates that KP1 is able to target to the
mitochondria via its tail domain.
To confirm the mitochondrial targeting of the tail domain
of KP1, the constructs for the stable expression of tail-GFP
were fused to the cauliflower mosaic virus 35S promoter (Fig-
ure 2A) and transformed into Arabidopsis. Two overexpression
lines (TAILOE1 and TAILOE2) were identified by immunoblot-
ting with a monoclonal antibody against GFP (Figure 2B). The
results indicate that tail-GFP indeed localizes to mitochondria
(Figure 2C).
KP1 Specifically Interacts with VDAC3
To identify if the tail of KP1 interacts with mitochondrial proteins,
we performed a yeast two-hybrid screen using the tail polypep-
tide as bait. A few polypeptides were identified as potential
interaction partners, of which we focused on a mitochondrial
outer membrane protein, VDAC3. The interaction between KP1
and VDAC3 was then further analyzed using the different do-
mains of KP1 in the yeast two-hybrid system (Figure 3A). Both
KP1-tail and KP1-tail200 (888 to 1087 amino acids) could interact
with VDAC3 to induce Trp1, Leu2,His3, Ade2, and LacZ reporter
genes in AH109 yeast lines, but the N-terminal domain (KP1-N;
1 to 373 amino acids) and the motor domain (KP1-M; 374 to 749
amino acids) of KP1 could not (Figure 3A). To determine if KP1
interacts with other members of the VDAC family in Arabidopsis,
VDAC1, VDAC2, and VDAC4 were cloned and transformed into
yeast cells. The results show that only VDAC3 interacts with KP1-
tail (Figure 3B).
The specific interaction between KP1-tail200 and VDAC3 in
vitrowas verified by aGSTpull down (Figure 3C) and a far-protein
gel blot (Figures 3D and 3E). As shown in Figure 3C, the 30-kD
KP1-tail200 associated with the bacterially expressed 60-kD
GST-VDAC3, while the control GST protein did not. For far-
protein gel blot analysis, two kinds of recombinant proteins, His-
VDAC3 (Figure 3D) and GST-VDAC3 (Figure 3E), were bacterially
expressed, isolated by SDS-PAGE, and transferred onto poly-
vinylidene difluoride (PVDF) membranes. After the membranes
were incubatedwith His-KP1-tail200, the anti-KP1 antibodies that
specifically label His-KP1-tail200 (Figure 3D, Immunoblot, lane 4)
not only recognized the His-KP1-tail200 (Figure 3D, far-western,
lane 4) but also the His-VDAC3 protein bands (Figure 3D, far-
western, lanes 2 and 3). Similarly, anti-His antibodies not only
specifically recognized His-KP1-tail200 (Figure 3E, Immunoblot,
lane 39; far-western, lane 39) but also the 60-kD protein band
of GST-VDAC3 (Figure 3E, far-western, lane 2’). Collectively,
we conclude that VDAC3 directly interacts with the KP1- tail200in vitro.
The specific interaction in vivo between KP1 and VDAC3 was
verified by two methods. Bimolecular fluorescence complemen-
tation (BiFC) showed that both KP1 and KP1-tail interact with
VDAC3 (Figure 4A). The interaction occurs specifically at mito-
chondria (Figure 4B), consistent with the localization of GFP-
VDAC3 (Figure 4C). To support these data, the firefly luciferase
complementation imaging (LCI) system was also performed.
When the constructed pairs of KP1-tail-NLuc/CLuc-VDAC3 and
KP1-NLuc/CLuc-VDAC3 were transformed into tobacco (Nico-
tiana tabacum) leaf cells, fluorescence signals were detected
(Figure 4D). All of these results demonstrate that KP1 specifically
interacts with VDAC3.
Figure 2. Mitochondrial Localization and Immunoblotting Test of KP1-Tail in Stem.
(A) The diagram represents the transgenic construct used to overexpress KP1-tail-GFP. GFP was fused to the tail domain of KP1.
(B) Immunoblot identification of tail-GFP in Col, TAILOE1, and TAILOE2 using anti-GFP antibody (1:10,000). Molecular marker is indicated in kilodaltons
in the left margin.
(C) KP1-tail-GFP in the stem of transgenic seedlings colocalized with mitochondria. From left to right, green channel (GFP), red channel (MitoTracker,
mitochondria selective reagent), merged images of green and red channel (Merge), and transmission images (Bright). Bars = 20 mm.
Kinesin and VDAC in Seed Respiration 1095

Mitochondrial Targeting of the KP1 Tail Is Dependent on Its
Interaction with VDAC3
To further analyze the localization of the KP1-tail via VDAC3
and to elucidate the function of KP1 and VDAC3, we isolated the
T-DNA insertion mutants of KP1 (kp1-1 and kp1-2) (Figure 5A)
and VDAC3 (vdac3-1 and vdac3-2) (Figure 5B) in the Columbia
(Col) background. In two kp1mutants, the T-DNA inserted within
the exon; in vdac3-1, the T-DNA inserted in the 59-untranslated
region; and in vdac3-2, the T-DNA inserted in the fourth intron
of the gene. The homozygous knockout mutants were isolated
and confirmed by PCR analysis, and their gene expression
was analyzed using RT-PCR. As shown in Figure 5C, the ex-
pression of KP1 was completely interrupted by the T-DNA
insertion in the kp1 mutants, and VDAC3 expression was signif-
icantly reduced in vdac3 mutants (Figure 5D).
We then tested the localization of tail-GFP in vdac3-1mutants
and GFP-VDAC3 in kp1-1 mutants to establish whether the
interaction between KP1 and VDAC3 confers the mitochondrial
targeting ability of the KP1 tail. As shown in Figure 6, the KP1
knockout did not inhibit the mitochondrial localization of GFP-
VDAC3; however, in vdac3-1 mutants, very little tail-GFP
Figure 3. Identification and Verification of a KP1-Interacting Protein, VDAC3.
(A) Yeast two-hybrid assay to analyze the interaction between KP1, KP1-N (1 to 373 amino acids), KP1-M (374 to 748 amino acids), KP1-tail (749 to
1087 amino acids), and KP1-tail200 (888 to 1087 amino acids) and VDAC3 on 4D agar medium (-Trp, -Leu, -His, -Ade). Only KP1-tail and KP1-tail200show binding activity with VDAC3.
(B) Yeast two-hybrid assay to analyze the interaction between four VDAC isoforms from Arabidopsis and KP1-tail and KP1-tail200. KP1-tail and KP1-
tail200 specifically interact with VDAC3.
(C) GST pull-down assay. GST (27 kD) or GST-VDAC3 (60 kD) protein was bound to beads and then incubated with 63His-tagged KP1-tail200 proteins
(30 kD). The eluted proteins were separated by SDS-PAGE. Anti-KP1 (1:5000) and anti-GST (1:100,000) antibodies were used for immunoblotting.
(D) Far-protein gel blot to verify the interaction between His-VDAC3 and His-KP1-tail200. Lane 1, unrelated His-tagged recombinant protein as negative
control; lane 2, purified His-VDAC3 (30 kD); lane 3, total extracts from Escherichia coli expressing His-VDAC3; lane 4, purified His-KP1-tail200 (30 kD).
The SDS-PAGE gel was stained with Coomassie blue and transformed onto PVDF membranes. The far-protein gel blot was performed with anti-KP1
antibody (1:5000) after the membrane was incubated with His-KP1-tail200. Two immunoblots were performed as positive controls with anti-KP1 (1:5000)
and anti-VDAC3 (1:5000) antibodies, respectively, to confirm the recombinant proteins. Molecular markers are indicated in kilodaltons in the left margin.
(E) Far-protein gel blot for the interaction assay between GST-VDAC3 and His-KP1-tail200. Lanes 1’ and 2’, the total extracts from E. coli expressing
GST and GST-VDAC3, respectively; lane 39, purified His-KP1-tail200. The far-protein gel blot was performed with an anti-His tag monoclonal antibody
after the membrane was incubated with His-KP1-tail200. Two immunoblots were performed as positive controls with anti-His (1:10,000) and anti-VDAC3
(1:5000) antibodies, respectively, to confirm the recombinant proteins. Molecular markers are indicated in kilodaltons in the left margin.
1096 The Plant Cell
colocalized with MitoTracker Red, suggesting that the mito-
chondrial localization of KP1 is dependent on its interaction
with VDAC3.
Both KP1 and VDAC3 Function during Seed Germination
at 48C
The phenotypes of all kp1 mutants and transgenic plants pre-
pared in this study were examined under normal growth condi-
tions throughout the plant’s life, but no visible phenotypes were
found, including regarding the distribution and morphology of
mitochondria (see Supplemental Figure 2 online). According to a
previous bioinformatics approach and RT-PCR analysis, KP1
expression is significantly reduced after cold treatment (Li et al.,
2008) and increased during seed germination (Schmid et al.,
2005). We designed real-time PCR experiments to understand
the gene transcription changes in detail. When wild-type seeds
were germinated at either 22 or 48C, the mRNA level of KP1
exhibited a sudden increase at day 2 or day 4 (Figures 7A and
7B), indicating the involvement of KP1 during seed germination.
Therefore, we decided to investigate the seed germination
phenotypes of the mutant lines resulting from the chilling
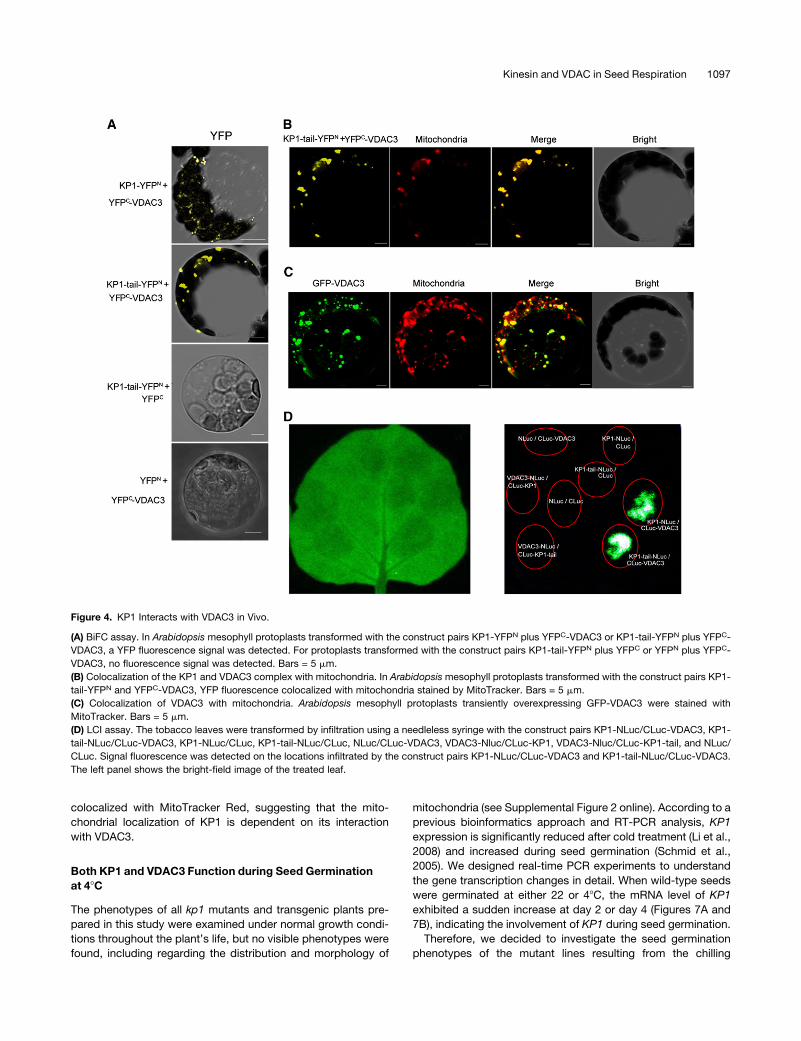
Figure 4. KP1 Interacts with VDAC3 in Vivo.
(A) BiFC assay. In Arabidopsis mesophyll protoplasts transformed with the construct pairs KP1-YFPN plus YFPC-VDAC3 or KP1-tail-YFPN plus YFPC-
VDAC3, a YFP fluorescence signal was detected. For protoplasts transformed with the construct pairs KP1-tail-YFPN plus YFPC or YFPN plus YFPC-
VDAC3, no fluorescence signal was detected. Bars = 5 mm.
(B) Colocalization of the KP1 and VDAC3 complex with mitochondria. In Arabidopsis mesophyll protoplasts transformed with the construct pairs KP1-
tail-YFPN and YFPC-VDAC3, YFP fluorescence colocalized with mitochondria stained by MitoTracker. Bars = 5 mm.
(C) Colocalization of VDAC3 with mitochondria. Arabidopsis mesophyll protoplasts transiently overexpressing GFP-VDAC3 were stained with
MitoTracker. Bars = 5 mm.
(D) LCI assay. The tobacco leaves were transformed by infiltration using a needleless syringe with the construct pairs KP1-NLuc/CLuc-VDAC3, KP1-
tail-NLuc/CLuc-VDAC3, KP1-NLuc/CLuc, KP1-tail-NLuc/CLuc, NLuc/CLuc-VDAC3, VDAC3-Nluc/CLuc-KP1, VDAC3-Nluc/CLuc-KP1-tail, and NLuc/
CLuc. Signal fluorescence was detected on the locations infiltrated by the construct pairs KP1-NLuc/CLuc-VDAC3 and KP1-tail-NLuc/CLuc-VDAC3.
The left panel shows the bright-field image of the treated leaf.
Kinesin and VDAC in Seed Respiration 1097

treatment. Interestingly, vdac3 T-DNA insertion lines exhibited
a higher seed germination frequency than the wild type at 48C,especially at day 10 (Figure 8A). After 14 d, all tested seeds
germinated completely, but the seedlings of the mutants were
stronger (Figure 8B) and the roots longer (Figure 8C). However,
no significant difference between kp1 mutants and the wild
type (Col) was observed at 48C, which may be due to gene
redundancy or the subtle function of KP1 during this process.
For this reason, transgenic lines overexpressing the KP1-tail,
TAILOE1 and TAILOE2, were generated and characterized as
dominant-negative mutants (Figures 2A and 2B). We inferred
that in dominant-negative mutants, the overexpressed KP1-
tail could compete with functional KP1 and its functionally redun-
dant proteins in plant cells. As expected, the transgenic lines
showed similar phenotypes to the vdac3 mutants at 48C (Figures
8A to 8C).
To determine if the visual phenotype of the mutants at
48C was due to quicker germination but not faster growth,
seeds were germinated at 228C for 2 d before being transferred
to 48C. Themutants did not present any obviousmorphological
phenotype. These results demonstrate that both KP1 and
VDAC3 function during seed germination at a low temperature.
KP1 and VDAC3 Regulate Respiration Pathways and
ATP Levels
The germination of seeds at the dormant stage requires efficient
energy from degradation metabolism. In a dry seed, the
Figure 5. Isolation of kp1 and vdac3 T-DNA Insertion Lines.
(A) Diagram of the KP1 gene structure. Introns are shown as lines, and exons are shown as black boxes. Triangles indicate the locations of the T-DNA
insertion sites for kp1-1 and kp1-2.
(B) The gene structure of VDAC3 and the T-DNA insertion sites of the vdac3 mutants. Introns are shown as lines, and exons are shown as black boxes.
Triangles indicate the locations of the T-DNA insertion sites.
(C) RT-PCR analysis of the wild type (Col) and kp1mutants. KP1 transcripts are absent in kp1mutants. TheUBQ5 transcript was amplified as an internal
control.
(D) RT-PCR analysis of the wild type (Col) and vdac3 mutants. VDAC3 transcripts are downregulated in vdac3 mutants. The UBQ5 gene was amplified
as an internal control. Similar results were obtained with at least three biological replicates of the experiments.
Figure 6. The KP1-Tail Localized to Mitochondria in a VDAC3-Dependent Manner.
GFP-VDAC3 merged with MitoTracker in Arabidopsismesophyll protoplasts from kp1-1, but the KP1-tail-GFP fusion expressed in vdac3-1 protoplasts
showed little colocalization with mitochondria.
1098 The Plant Cell

mitochondria barely function, and one of the initial changes
during the early stages of germination is the resumption of
respiratory activity (Bewley, 1997). For this reason, the respira-
tory rate of kp1 and vdac3mutantswas investigated. As shown in
Figure 9, oxygen consumption increased significantly in kp1 and
vdac3 mutants during the period of seed germination at 48C,especially in the plants germinated for 7 to 11 d. In higher plants,
the oxygen consumption usually results from the cytochrome
pathway and the AOX pathway in mitochondria; therefore, we
measured the oxygen consumption of seeds germinated at day
10 at 48C in the presence of the specific respiration inhibitors,
NaN3 (for inhibiting the cytochrome pathway) and salicylhy-
droxamic acid (SHAM; for inhibiting the AOX pathway). The
results showed that in kp1 mutants SHAM-resistant respiration
(similar to the cytochrome pathway) increased greatly, while CN-
resistant respiration (similar to the AOX pathway) decreased. The
ratio of oxygen consumption via the cytochrome pathway and
the AOX pathway changed from ;1:1 in the wild type to 6:1 in
kp1 mutants (Figure 10A). The vdac3-1 mutants showed similar
results, with a ratio of ;3:1. We obtained the VDAC3 over-
expression transgenic line (VOE) to confirm the above results
(Figures 10B and 10C). The VOE line restored the oxygen
consumption ratio of the cytochrome pathway to AOX pathway
(Figure 10A).
It is well known that the energy from the mitochondrial cyto-
chrome pathway is mainly used for ATP synthesis. Therefore, we
measured the ATP levels in 10-d-old seedlings. Interestingly, the
kp1 mutants grown at 48C produced ;0.6 nmol ATP/g fresh
weight, which is 31% lower than that in the wild type; the ATP
level in the vdac3-1 mutants also decreased, by 45% (Figure
11A). The restoration of the ATP level in transgenic Arabidopsis
plants overexpressing VDAC3 further confirmed this vdac3 mu-
tant phenotype. To eliminate the possibility that the observed
changes in ATP levels were due to a different quantity of mito-
chondria, the activity of citrate synthase, a marker for mitochon-
drial mass (Moraes et al., 1993), was assayed. No significant
difference between the wild type and the mutants was observed
(Figure 11B).
The SHAM/CN-resistant respiration and ATP levels were
also assayed in the dominant-negative mutants TAILOE1 and
TAILOE2. The ratio of the SHAM-resistant respiration to CN-
resistant respiration was found to be;6:1 (Figure 12A), and the
ATP level was 35% lower than that of Col (Figure 12B). This
is similar to our findings for kp1 mutants, indicating that the
overexpressed tail domain suppressed the function of KP1.
These results demonstrate that both KP1 and VDAC3 are in-
volved in regulating respiration pathways and ATP levels during
seed germination at a low temperature.
DISCUSSION
Kinesins constitute a superfamily of microtubule motor proteins
and are known to be essential for many cellular functions (Miki
et al., 2005). Studies on the kinesins of higher plants revealed
their participation inmicrotubule organization duringmeiosis and
mitosis (Chen et al., 2002; Marcus et al., 2003), cytokinesis (Lee
et al., 2007), cellulose microfibril deposition (Zhong et al., 2002),
and morphogenesis (Oppenheimer et al., 1997; Reddy et al.,
2004; Lu et al., 2005). In this study, a plant-specific kinesin KP1
(At KIN14h) was found to interact with the mitochondrial outer
membrane protein VDAC3 via its tail domain and to be involved in
the regulation of respiration during seed germination at low
temperature.
KP1 Specifically Interacts with the Mitochondrial Outer
Membrane Protein VDAC3
Systematic analysis of the sequences in theArabidopsis genome
with the conservative motor domain of kinesins revealed the
presence of 61 kinesins or kinesin-like proteins (Reddy and Day,
2001). KP1, a member of the subfamily kinesin-14, tightly binds
to mitochondria from Arabidopsis (Ni et al., 2005). Here, using
several biochemical and transgenic methods, we demonstrate
the specific interaction of KP1 with VDAC3, an isoform of
Arabidopsis VDACs that normally localizes to the outer mem-
brane of mitochondria (Colombini, 1979). With the exception of
VDAC5, whose gene has not been cloned and identified, all four
of the other VDAC isoforms from Arabidopsis were tested in
yeast two-hybrid experiments, and only VDAC3was identified as
interacting with KP1 (Figures 3A and 3B). To provide biochemical
evidence of this interaction, the VDAC3 and KP1 recombinant
Figure 7. KP1 Expression Increased during Seed Germination.
Real-time PCR analyses of KP1 expression in the wild type during seed imbibition and germination at 22 (A) or 48C (B). KP1 transcripts exhibit a sudden
increase during this period. The 18S rRNA gene was amplified as an internal reference. Data represent the mean 6 SE of three independent biological
determinations.
Kinesin and VDAC in Seed Respiration 1099
proteins were bacterially expressed, affinity purified, and ana-
lyzed by GST pull down and far-protein gel blot, respectively
(Figures 3C to 3E). For the direct observation of the interaction in
vivo, both BiFC and LCI assays were performed (Figures 4A and
4D). The results from these methods confirmed the specific
interaction between KP1 and VDAC3.
VDACs are the most abundant integral membrane proteins in
the mitochondrial outer membrane (Colombini, 1979; Benz,
1994). Dozens of cytosolic proteins have been reported to
interact with VDACs in mammalian and yeast cells (reviewed in
Rostovtseva and Bezrukov, 2008). Through affinity chromatog-
raphy analysis, VDAC was identified as a binding site for micro-
tubule-associated protein 2 (Linden andKarlsson, 1996). Purified
VDAC1 interacts with a cytoplasmic dynein light chain Tctex-1 in
vitro (Schwarzer et al., 2002). Recently, VDAC proteins were
shown to interact with tubulin and actin based on immunopre-
cipitation and surface plasmon resonance technology (Carre
et al., 2002; Roman et al., 2006). Interestingly, several kinesin
receptors were identified as binding to mitochondria, such as
syntabulin, kinectin, and milton (Stowers et al., 2002; Santama
et al., 2004; Cai et al., 2005). In plant cells, the relationship
between kinesin and mitochondrial proteins has not been re-
ported. The specific interaction between KP1 and VDAC3
reported here reveals novel roles for plant kinesin isoforms and
provides insight into the molecular mechanisms underlying mi-
tochondrial functions.
Although the transgenic cellular imaging study (Figure 1)
suggests that KP1 is able to target to mitochondria via its tail
domain, we remain puzzled about the regulation of the tail-cargo
binding. The binding affinity of full-length KP1 to microtubules
and mitochondria seems to be related to the regulation of the
KP1 protein domains. In Arabidopsis plants, KP1 is usually
expressed at a very low level and tightly localizes tomitochondria
(Ni et al., 2005). When transiently overexpressed in protoplasts,
GFP-KP1 fusion proteins accumulated on microtubules and,
to a small degree, on mitochondria (Figure 1C), whereas tail-
GFP preferentially localized to mitochondria (Figure 1C), sug-
gesting that the other domains of KP1 may negatively regulate
the tail-mitochondria association. Interactions between differ-
ent domains are abundant in animal kinesins. For example,
there is a direct interaction between the kinesin-1 head and
tail (Dietrich et al., 2008); a 65–amino acid C-terminal tail
domain is an inhibitory regulator of the ATPase and motor
activities of the head domains of the kinesin (Coy et al., 1999); in
the absence of bound cargo, the kinesin tail interacts with the
motor domains and inhibits their activity (Cross and Scholey,
1999). It will be interesting to establish how the head and
motor domains of KP1 influence the ability of its tail to bind to
mitochondria.
Figure 9. The kp1 and vdac3Mutants Have a Higher Aerobic Respiration
Rate during Seed Germination at 48C.
Oxygen consumption was measured in kp1 and vdac3 mutants during
the period of seed germination at 48C by a Clark-type electrode. Each
value is the mean 6 SE of three independent biological determinations.
Asterisks indicate a significant difference between mutants and Col: *P <
0.05; **P < 0.01. DW, dry weight.
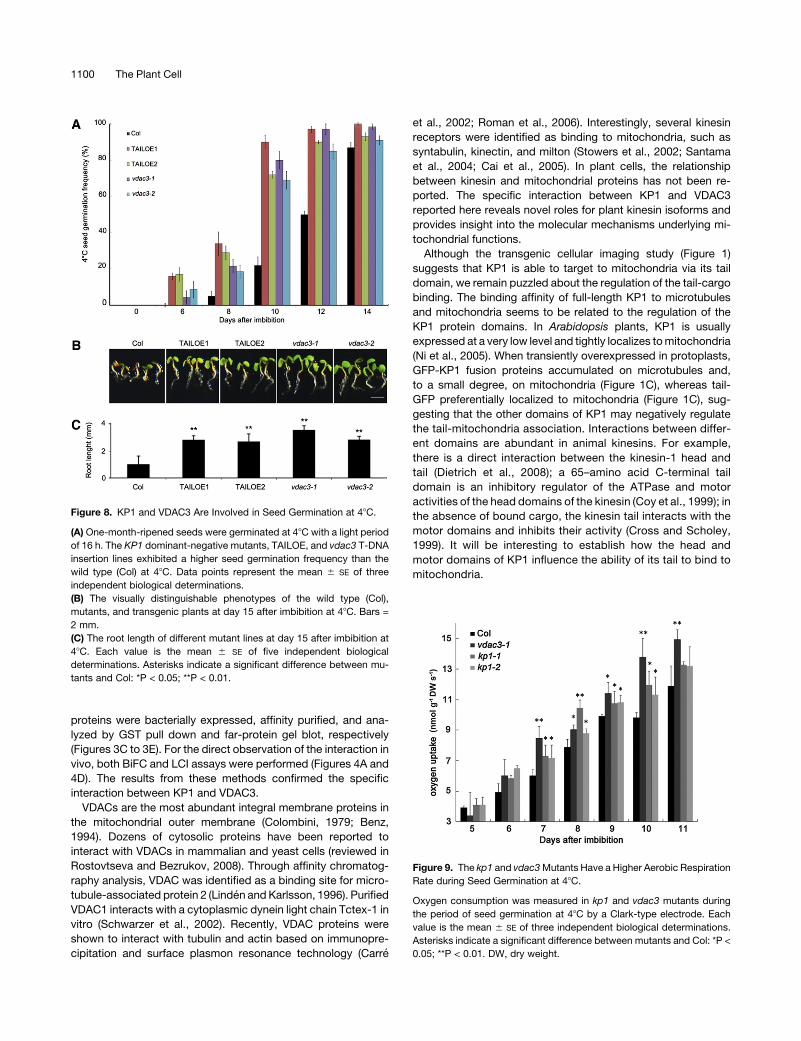
Figure 8. KP1 and VDAC3 Are Involved in Seed Germination at 48C.
(A) One-month-ripened seeds were germinated at 48C with a light period
of 16 h. The KP1 dominant-negative mutants, TAILOE, and vdac3 T-DNA
insertion lines exhibited a higher seed germination frequency than the
wild type (Col) at 48C. Data points represent the mean 6 SE of three
independent biological determinations.
(B) The visually distinguishable phenotypes of the wild type (Col),
mutants, and transgenic plants at day 15 after imbibition at 48C. Bars =
2 mm.
(C) The root length of different mutant lines at day 15 after imbibition at
48C. Each value is the mean 6 SE of five independent biological
determinations. Asterisks indicate a significant difference between mu-
tants and Col: *P < 0.05; **P < 0.01.
1100 The Plant Cell
KP1 and VDAC3 Participate in Mitochondrial Respiratory
Regulation during Seed Germination
Kinesins have many cellular functions, including the transport of
organelles (Hirokawa et al., 2009). The kinesin members, KIF1B
and KIF5B in mouse cells (Nangaku et al., 1994; Tanaka et al.,
1998) and KLP67A in early Drosophila embryos (Tanaka et al.,
1998), have been implicated inmitochondrial movement. Purified
KIF1B could drive mitochondria to move along microtubules to
the plus end (Nangaku et al., 1994). It is unclear if any kinesins are
involved in mitochondrial respiration. In this research, though no
significant seed germination phenotype was observed in kp1
T-DNA insertion lines, KP1 dominant-negative mutants, TAILOE,
exhibited a higher seed germination frequency at 48C (Figure 8A),
indicating that KP1 functions as a regulator during seed germi-
nation. Further analysis with specific respiration inhibitors
(SHAM and NaN3) showed that SHAM-resistant respiration (sim-
ilar to the cytochrome pathway) significantly increased in kp1
T-DNA insertion mutants and TAILOE lines at day 10 after
imbibition, while CN-resistant respiration decreased (Figures
10A and 12A). According to previous studies on several different
plants, CN-resistant respiration predominates during the early
stages of seed germination, but after 12 h of imbibition, SHAM-
and CN-resistant respiration achieve a balance (Yentur and
Leopold, 1976). Our data showed that at day 10 after imbibition,
the balance between the SHAM- and CN-resistant respiration
pathway was disrupted in kp1 T-DNA insertion mutants and
TAILOE lines, changing from a ratio of 1:1 in the wild type to 6:1 in
mutants (Figures 10A and 12A). This suggests that KP1 regulates
respiratory pathways during seed germination.
In animal cells, the mitochondrial channel VDACs form large
aqueous pores through membranes and play a crucial role in
regulating the transport of ATP/ADP, Ca2+, and other metabo-
lites between the cytosol and mitochondria (Rostovtseva and
Colombini, 1996; Lemasters and Holmuhamedov, 2006). The
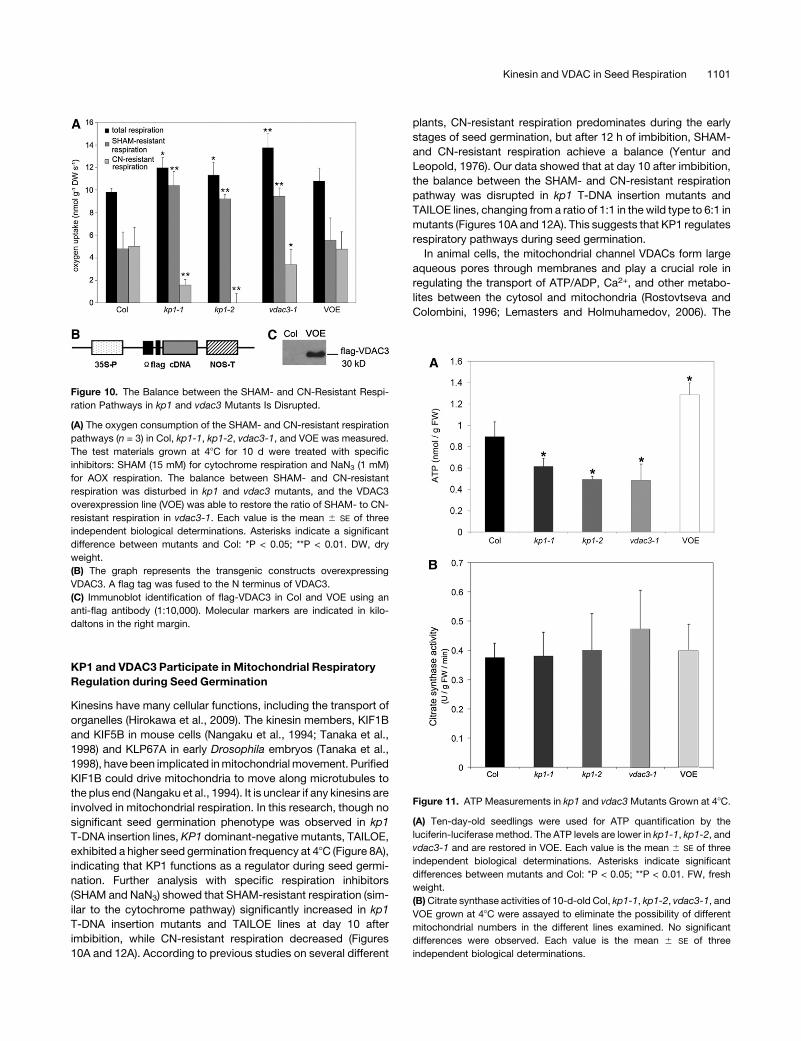
Figure 10. The Balance between the SHAM- and CN-Resistant Respi-
ration Pathways in kp1 and vdac3 Mutants Is Disrupted.
(A) The oxygen consumption of the SHAM- and CN-resistant respiration
pathways (n = 3) in Col, kp1-1, kp1-2, vdac3-1, and VOE was measured.
The test materials grown at 48C for 10 d were treated with specific
inhibitors: SHAM (15 mM) for cytochrome respiration and NaN3 (1 mM)
for AOX respiration. The balance between SHAM- and CN-resistant
respiration was disturbed in kp1 and vdac3 mutants, and the VDAC3
overexpression line (VOE) was able to restore the ratio of SHAM- to CN-
resistant respiration in vdac3-1. Each value is the mean 6 SE of three
independent biological determinations. Asterisks indicate a significant
difference between mutants and Col: *P < 0.05; **P < 0.01. DW, dry
weight.
(B) The graph represents the transgenic constructs overexpressing
VDAC3. A flag tag was fused to the N terminus of VDAC3.
(C) Immunoblot identification of flag-VDAC3 in Col and VOE using an
anti-flag antibody (1:10,000). Molecular markers are indicated in kilo-
daltons in the right margin.
Figure 11. ATP Measurements in kp1 and vdac3 Mutants Grown at 48C.
(A) Ten-day-old seedlings were used for ATP quantification by the
luciferin-luciferase method. The ATP levels are lower in kp1-1, kp1-2, and
vdac3-1 and are restored in VOE. Each value is the mean 6 SE of three
independent biological determinations. Asterisks indicate significant
differences between mutants and Col: *P < 0.05; **P < 0.01. FW, fresh
weight.
(B)Citrate synthase activities of 10-d-old Col, kp1-1, kp1-2, vdac3-1, and
VOE grown at 48C were assayed to eliminate the possibility of different
mitochondrial numbers in the different lines examined. No significant
differences were observed. Each value is the mean 6 SE of three
independent biological determinations.
Kinesin and VDAC in Seed Respiration 1101

reduction in the expression of VDAC1 in human cells leads to a
decrease in the ATP level (Abu-Hamad et al., 2006). In this study,
the vdac3mutants shared a similar phenotype to KP1 dominant-
negative mutants, TAILOEs, and kp1mutants, including a higher
cold germination frequency (Figure 8A), a higher respiration rate
(Figure 9), a higher ratio of SHAM- to CN-resistant respiration
(;3:1) (Figure 10A), and lower ATP levels (Figure 11A). This
indicates that VDAC3 also functions in regulating respiratory
pathways during seed germination at low temperature. Recently,
it was reported that in Trypanosoma brucei, a unicellular parasitic
protozoan, the downregulation of Tb VDAC reduces cellular
respiration via the AOX pathway, enhances respiration via the
cytochrome pathway, and exhibits a lower ATP level (Singha
et al., 2009).
What is the mechanism by which KP1 interacts with VDAC3?
What is the relationship between respiration pathway balance
and seed germination in both kp1 and vdac3 mutants at low
temperature? From our data, we could not answer these ques-
tions. We suggest that KP1 may positively regulate the opening
of VDAC3 and regulate the flux of ATP/ADP across the mito-
chondrial outer membrane; in mutants, the ATP/ADP flux may
be disturbed to a certain extent, resulting in a lower production
of ATP in mitochondria, which in turn stimulates respiration via
the cytochrome pathway in an attempt to compensate for the
energy crisis in the cells. It would be interesting to address the
above questions.
METHODS
Plant Material and Growth Conditions
Arabidopsis thaliana seeds were surface terilized. The seeds were strat-
ified at 48C for 2 d and then placed on half-strengthMurashige and Skoog
(MS)medium (1%Suc and 0.9%agar). For the transformation andmutant
screen, seedlings were transferred from plates to soil and grown at 228C
in a growth room with a photon flux density of 100 mEm22 s21 and a 16-h
photoperiod.
For the seed germination assays, seeds from the different genotypes
were harvested from plants grown simultaneously in glasshouse condi-
tions and then stored for 4 weeks. For the seed germination assay at 48C,
plates were kept in the incubator at 48C with a 16-h photoperiod at a
photon flux density of 70 mE/m22 s21. Germination was scored by radicle
emergence.
Plasmid Construction
The full-length KP1 cDNA was previously cloned (Li et al., 2007). The full-
length cDNA sequence of VDAC3 is available from GenBank (accession
number NM_121513). For protoplast transient expression assays, the full-
length cDNA of KP1 was amplified by PCR using the following two
primers: 59-TCTAGAATGGACCAAGGCGCGAT-39 (XbaI) and 59-GTC-
GACCTATGGTACCATGAACCTTG-39 (SalI). The KP1-Dtail (KP1 lacking
the C-terminal region; 1 to 751 amino acids) construct was made using
the following primers: 59-TCTAGAATGGACCAAGGCGCGAT-39 (XbaI)
and 59-GTCGACCTAGTTCCGAATGCTACCT-39 (SalI). For the KP1-tail
(749 to 1087 amino acids), the primers 59-TCTAGAATGAAGGAAA-
CCGGTGAAATTC-39 (XbaI) and 59-GTCGACTGGTACCATGAACCT-
TGC-39 (SalI) were used. For VDAC3, the primers 59-TCTAGAA-
TGGTTAAAGGTCCAGGACTCTAC-39 (XbaI) and 59-GAGCTCTCAGGG-
CTTGAGAGCGAGAG-39 (SacI) were used. The open reading frames
(ORFs) of GFP were amplified using the following primers: 59-GGATC-
CATGAGTAAAGGAGAAGAACT-39 (BamHI) and 59- TCTAGATTTGTA-
TAGTTCATCCATGCC-39 (XbaI) with no termination codon, and 59-GTC-
GACATGAGTAAAGGAGAAGAAC-39 (SalI) and 59-TTAGGTACCTTTG-
TATAGTTCATCCATG-39 (SacI) with the termination codon TAA. The
fusion constructs GFP-KP1, GFP-Dtail, and GFP-VDAC3 were made by
placing the coding region in frame with the C terminus of the GFP coding
region (digested with BamHI and XbaI). The tail-GFP construct was made
by fusing the target sequences to the N terminus of GFP (digested with
SalI and SacI). All of the fusion constructs above were cloned into a pUC
vector under the control of the 35S cauliflower mosaic virus promoter.
For the BiFC assay, the ORFs of VDAC3 were amplified by PCR using
the following primers: 59-TCTAGAATGGTTAAAGGTCCAGGACTCTAC-39
(XbaI) and 59-CTCGAGGGGCTTGAGAGCGAGAG-39 (XhoI). The DNA
fragments encoding full-length KP1 were amplified by PCR using the
following primers: 59-TCTAGAATGGACCAAGGCGCGAT-39 (XbaI) and
59-GTCGACTGGTACCATGAACCTTGC-39 (SalI). All of the DNA frag-
ments used were cloned into the plasmid pUC-SPYNE to form the fusion
proteins KP1-YFPN, KP1-tail-YFPN, and VDAC3-YFPN and into pUC-
SPYCE to form the fusion proteins KP1-YFPC, KP1-tail-YFPC, and
VDAC3-YFPC. For fusing YFPC to the N terminus of VDAC3, we amplified
Figure 12. The Ratio of SHAM- to CN-Resistant Respiration and ATP
Levels in TAILOE Dominant-Negative Mutants.
(A) The oxygen consumption of the SHAM/CN-resistant respiration
pathways (n = 3) in Col, TAILOE1, and TAILOE2 was measured. The
balance between SHAM/CN-resistant respiration is disturbed in TAILOE
lines. DW, dry weight.
(B) The ATP levels are lower in TAILOE lines. Each value is the mean6 SE
of three independent biological determinations. Asterisks indicate sig-
nificant differences between mutants and Col: *P < 0.05; **P < 0.01. FW,
fresh weight.
1102 The Plant Cell

the C-terminal half of YFP by PCR using the following primers:
59-TCTAGAATGTACGACGTACCAGATTA-39 (XbaI) and 59-ACTAGT-
CTTGTACAGCTCGTCCAT-39 (SpeI). We amplified the ORFs of VDAC3
using the following primers: 59-ACTAGTCGATCACTACAACGGGAAC-39
(SpeI) and 59-TCTAGAAGAAGAATCAGTGGAAACTTTG-39 (XbaI) with a
termination codon. TheseDNA fragments were inserted into pUC-SPYCE
to form YFPC-VDAC3. All of the constructs above were sequenced to
confirm their fidelity.
Screening of T-DNA Insertion Lines and Isolation of kp1 and
vdac3Mutants
The kp1-1 (SALK_056981), kp1-2 (SALK_117309), vdac3-1 (SALK_
127899), and vdac3-2 (SAIL_238_A01) mutant lines, carrying T-DNA
insertions in KP1 and VDAC3, were obtained from the ABRC. PCR-based
screening was used to test homozygosity. The locations of the T-DNA
insertion sites in the SALK and SAIL lines were determined by direct
sequencing of PCR products amplified by the T-DNA left border primer
LBa1, 59-TGGTTCACGTAGTGGGCCATCG-39 for SALK lines, the primer
LB3, 59-TAGCATCTGAATTTCATAACCAATCTCGATACAC-39 for SAIL
lines, and gene-specific primers. The gene-specific primers were as fol-
lows: for kp1-1 (SALK_056981), 59-CACACAGCACCATAAGCATTG-39;
for kp1-2 (SALK_117309), 59-GAAGCAGAGCTGGAACAATTG-39; for
vdac3-1 (SALK_127988), 59-AGACATTGTCAAAGACTCAACAAC-39; and
for vdac3-2 (SAIL_238_A01), 59-TGCCAGATTCGGTGTTATAGG-39.
Generation of Transgenic Plants
The plasmid constructs pBI121-35S--flag-VDAC3 and pBI121-35S-KP1-
tail-GFP were electroporated into Agrobacterium tumefaciens strain
C58C1. Stable transgenic Arabidopsis plants were generated using the
flower dipping method (Clough and Bent, 1998). Transgenic plants were
selected on 0.53MSplates with 50mg/L kanamycin. The T3 generations
of transgenic plants were used for experiments.
RT-PCR and Real-Time PCR
Total RNA was isolated from plant materials using Trizol reagent (Invi-
trogen) according to the manufacturer’s instructions. RNA isolation from
the dry, imbibed, and germinating seeds was performed using the RNA
extraction kit (Bioteke) according to themanufacturer’s instructions. RNA
concentration was determined using a NanoDrop ND-1000 photospec-
trometer. Reverse transcription was performed with 5 mg of total RNA
using M-MLV reverse transcriptase (Promega) according to the manu-
facturer’s instructions. PCR was performed for 35 cycles with the follow-
ing gene-specific primers: UBQ5, 59-CTCCTTCTTTCTGGTAAACGT-39
and 59-GGTGCTAAGAAGAGGAAGAAT-39; KP1, 59-CAGAAGCTACGA-
GACCAGAAGTTG-39 and 59-CTATGGTACCATGAACCTTGCATG-39;
and VDAC3, 59-TTTTTCCAGAGGCAATCATG-39 and 59-GCCCATTTGG-
TGGTATCTTC-39. Quantitative real-time PCR was performed following
the protocol of the Perfect Real-time PCR kit (TaKaRa) on the Applied
Biosystems 7500 Real-Time PCR system. Amplification products were
visualized by SYBR Green. Aliquots of the RT reaction products were
used as templates for real-time PCR reactions. For relative quantification,
the 18S rRNA gene was detected as an internal reference, and the 22DDCt
method (Livak and Schmittgen, 2001) was used.
Protoplast Transient Expression Assays and
Fluorescence Microscopy
Transient expression of various plasmids in Arabidopsis protoplasts was
performed as previously described (Sheen, 2002). The viability of proto-
plasts was determined using fluorescein diacetate staining (Larkin, 1976)
(see Supplemental Figure 3 online). For GFP-KP1, both bright and dim
protoplasts were chosen for observation. The BiFC assay was performed
according to a previous report (Walter et al., 2004). Fluorescence was
observed after protoplasts were incubated at 228C for 12 to 16 h.
Mitochondria were visualized by staining with MitoTracker Red (Molec-
ular Probes) according to the manufacturer’s protocol. To stain microtu-
bules and microfilaments, Arabidopsis suspension cell protoplasts were
attached to cover slips coated with 1 mg/mL poly-L-lysine (Mr >300,000;
Sigma-Aldrich) and fixed for 30 min at room temperature with 3% (w/v)
paraformaldehyde in PEMbuffer (50mMPIPES, pH 6.9, 5mMEGTA, and
1 mMMgSO4) supplemented with 1%DMSO, 0.3 mM PMSF, and 0.05%
Triton X-100. The fixed protoplast ghosts were thenwashedwith PBS, pH
7.4, and blocked in 1% (w/v) BSA (Sigma-Aldrich) for 10 min. For labeling
microtubules, the anti-a-tubulin monoclonal antibody (Sigma-Aldrich;
diluted 1:500) and the tetramethylrhodamine b-isothiocyanate–conju-
gated goat anti-mouse IgG (Jackson ImmunoResearch Laboratories;
diluted 1:200) were applied, and microfilaments were stained with 50 nM
rhodamine-phalloidin (Molecular Probes) at 258C for 1 h. After rinsing in
PBS, slides were observed under a Zeiss LSM 510 META confocal
microscope. The confocal settings were as follows: green images (GFP,
fluorescein diacetate staining) were obtained with a 488-nm argon laser
and Fset09 wf filter; red images (MitoTracker stain, rhodamine-phalloidin
stain) were obtained with a 543-nm HeNe laser and Fset15 wf filter. A
photomultiplier tube was used as the confocal detector.
Protein Expression and Antibody Production
His-VDAC3 proteins were expressed in the Escherichia coli BL21 (DE3)
strain, induced with 0.1 mM isopropyl thiogalactoside for 6 h at 228C, and
affinity purified using a Ni2+-chelating Sepharose Fast Flow (Amersham
Biosciences) column following the manufacturer’s instructions.
Polyclonal anti-VDAC3 antibodies were raised in rabbits using the
purified His-VDAC3 protein as the antigen. Antiserum was then affinity
purified using the AminoLink Plus kit (Pierce Chemical) with immobilized
VDAC3 according to the manufacturer’s instructions.
Yeast Two-Hybrid Screening
Arabidopsis cDNA clones encoding KP1-interacting proteins were
screened by a GAL4-based yeast two-hybrid system using the yeast
two-hybrid host strain AH109 as described by the manufacturer (Clon-
tech). KP1-tail was used in the bait construct to screen the cDNA library
for candidate interaction partners of KP1. To verify the interaction
between KP1 and VDAC3, truncated KP1 proteins, KP1-N (1 to 373
amino acids), KP1-M (374 to 749 amino acids), and KP1-tail200 (888 to
1087 amino acids) were used for bait constructs.
For the interactions between KP1 and VDAC family proteins, the ORF
cDNA of VDAC1 was amplified by PCR using the following two primers:
59-CCTCCAACTTTCTCAGATAAGCAAC-39 and 59-CTGAATATGCAA-
TTTTCATTATGACAC-39. For VDAC2, 59-CTCTCTCAATCTCCGATCA-
ACC-39 and 59-CTGCGGAACTATTTATTGATTCC-39 were used, and
for VDAC4, 59-GCATTTGTTTTCTATATCCGAAG-39 and 59-TCCCTTT-
TCTTTCACATCACA-39 were used. The full-length cDNAs of VDAC1,
VDAC2, and VDAC4 were then ligated into pGADT7 and transformed into
yeast cells with pGBKT7-KP1-tail and pGBKT7-KP1-tail200.
GST Pull Down and Far-Protein Blot Assays
GST, GST-VDAC3, His-VDAC3, and His-KP1-tail200 were expressed in E.
coli BL21 (DE3) and purified according to standard protocols (Sambrook
and Russell, 2001).
A GST pull-down assay was conducted. Aliquots of GST and GST-
VDAC3 beads (100 mL beads containing ;15 mg of protein) were
incubated for 2 h at 48C with the His-KP1-tail200 protein. After being
washed with PBS buffer, bound proteins were eluted from the beads with
Kinesin and VDAC in Seed Respiration 1103

50 mL of elution buffer (20 mM reduced glutathione in 50 mM Tris-Cl, pH
8.0), resolved on a 12.5% SDS-PAGE gel, and then immunoblotted with
anti-His antibody at a 1:10,000 dilution and with anti-GST antibody at a
1:100,000 dilution.
A far-protein gel blot was performed as previously described
(Schwarzer et al., 2002). The total proteins extracted from E. coli over-
expressing His-VDAC3 and GST-VDAC3 and the purified His-VDAC3,
GST-VDAC3, His-KP1-tail200, and unrelated His-tagged recombinant
protein were separated by SDS-PAGE and transferred electrophoretically
to PVDF membranes. After the blocking of nonspecific sites with 5%
nonfat dried milk in TBST (20 mM Tris-HCl, pH 7.5, 100 mM NaCl, and
0.05% Tween) for 2 h at room temperature, the membranes were washed
three times with TBST and incubated for 1 h at room temperature at 48C
with the recombinant His-KP1-tail200 protein (20mg/mL). Themembranes
were then washed three times in TBST and blocked again. Then, the
membranes were immunoblotted with anti-His, anti-GST, anti-KP1, and
anti-VDAC3 antibody, respectively.
LCI Assay
The LCI assay was performed as previously described (Chen et al., 2008).
To make KP1-/KP1-tail-NLuc, VDAC3-NLuc, CLuc-KP1/KP1-tail, and
CLuc-VDAC3 constructs for the LCI assay, the related cDNAs were
inserted into pCAMBIA-NLuc and pCAMBIA-CLuc vectors. All of the
resultant constructs were electroporated into Agrobacterium strain
C58C1. Bacterial suspensions were infiltrated into fully expanded leaves
of the 7-week-old Nicotiana benthamiana plants using a needleless
syringe. After that, plants were grown in darkness for 12 h and then with a
light period of 16 h for 60 h at 228C. The LUC activity was observed with a
CCD imaging apparatus (AndoriXon; Andor).
Measurements of Oxygen Consumption
Seeds (50 mg) germinated at 48C in liquid culture medium (0.53 MS
medium, 0.025% MES, and 1% Suc, pH 5.6) were used to measure
oxygen consumption at 48C using 1.5 mL medium in the dark, a Clark-
type electrode (Hansatech), and a 2-mL vessel. The oxygen consumption
in the seeds (germinated for 10 d) was detected in the presence or
absence of the specific respiratory inhibitors, 1 mM NaN3 and 15 mM
SHAM.
Measurements of ATP Level and Citrate Synthase Activity
The ATP concentration was measured as previously described (Meyer
et al., 2009). To extract ATP from the seedlings, ;250 mg frozen
Arabidopsis seedlings were ground and resuspended in 400 mL of
2.3% (v/v) trichloroacetic acid. A bioluminescent assay kit (Sigma-
Aldrich) was used to measure the ATP concentration. Citrate synthase
activity was assayed spectrophotometrically at 412 nm in a reaction with
0.2mMoxaloacetate, 0.1mMacetyl-CoA, and 0.2mM5, 59-dithiobis(2,4-
nitrobenzoic acid) (Bond et al., 2005). The change of OD in a unit
represents a U-activity unit.
Accession Numbers
Sequence data from this article can be found in the Arabidopsis Genome
Initiative or GenBank/EMBL databases under the following accession
numbers: KP1 (At3g44730), VDAC1 (At3g01280), VDAC2 (At5g67500),
VDAC3 (At5g15090), VDAC4 (At5g57490), and UBQ5 (At3g62250).
Supplemental Data
The following materials are available in the online version of this article.
Supplemental Figure 1. GFP-KP1 Decorated Microtubules in Arabi-
dopsis Suspension Cell Protoplasts.
Supplemental Figure 2. There Is No Significant Difference in Mito-
chondrial Organization and Morphology in Roots among 5-d-Old Col
and kp1-1 and vdac3-1 Mutants.
Supplemental Figure 3. The Viability of Protoplasts Was Determined
Using Fluorescein Diacetate.
ACKNOWLEDGMENTS
We thank Shao-Hua Li (Institute of Botany, Chinese Academy of Sci-
ences) for providing a Clark-type electrode for the measurement of
oxygen consumption. This work was supported by grants from the
National Natural Science Foundation of China (Project 31071259,
30770128, 30721062, and 31030010).
Received December 18, 2010; revised February 10, 2011; accepted
February 21, 2011; published March 15, 2011.
REFERENCES
Abu-Hamad, S., Sivan, S., and Shoshan-Barmatz, V. (2006). The
expression level of the voltage-dependent anion channel controls life
and death of the cell. Proc. Natl. Acad. Sci. USA 103: 5787–5792.
Benz, R. (1994). Permeation of hydrophilic solutes through mitochon-
drial outer membranes: Review on mitochondrial porins. Biochim.
Biophys. Acta 1197: 167–196.
Bewley, J.D. (1997). Seed germination and dormancy. Plant Cell 9:
1055–1066.
Bond, D.R., Mester, T., Nesbø, C.L., Izquierdo-Lopez, A.V., Collart,
F.L., and Lovley, D.R. (2005). Characterization of citrate synthase
from Geobacter sulfurreducens and evidence for a family of citrate
synthases similar to those of eukaryotes throughout the Geobacter-
aceae. Appl. Environ. Microbiol. 71: 3858–3865.
Cai, Q., Gerwin, C., and Sheng, Z.H. (2005). Syntabulin-mediated
anterograde transport of mitochondria along neuronal processes. J.
Cell Biol. 170: 959–969.
Carre, M., Andre, N., Carles, G., Borghi, H., Brichese, L., Briand, C.,
and Braguer, D. (2002). Tubulin is an inherent component of mito-
chondrial membranes that interacts with the voltage-dependent anion
channel. J. Biol. Chem. 277: 33664–33669.
Chen, C., Marcus, A., Li, W., Hu, Y., Calzada, J.P.V., Grossniklaus,
U., Cyr, R.J., and Ma, H. (2002). The Arabidopsis ATK1 gene is
required for spindle morphogenesis in male meiosis. Development
129: 2401–2409.
Chen, H., Zou, Y., Shang, Y., Lin, H., Wang, Y., Cai, R., Tang, X., and
Zhou, J.M. (2008). Firefly luciferase complementation imaging assay
for protein-protein interactions in plants. Plant Physiol. 146: 368–376.
Clausen, C., Ilkavets, I., Thomson, R., Philippar, K., Vojta, A.,
Mohlmann, T., Neuhaus, E., Fulgosi, H., and Soll, J. (2004). Intra-
cellular localization of VDAC proteins in plants. Planta 220: 30–37.
Clough, S.J., and Bent, A.F. (1998). Floral dip: A simplified method for
Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant
J. 16: 735–743.
Colombini, M. (1979). A candidate for the permeability pathway of the
outer mitochondrial membrane. Nature 279: 643–645.
Coy, D.L., Hancock, W.O., Wagenbach, M., and Howard, J. (1999).
Kinesin’s tail domain is an inhibitory regulator of the motor domain.
Nat. Cell Biol. 1: 288–292.
1104 The Plant Cell

Cross, R., and Scholey, J. (1999). Kinesin: The tail unfolds. Nat. Cell
Biol. 1: 119–121.
Dietrich, K.A., Sindelar, C.V., Brewer, P.D., Downing, K.H., Cremo,
C.R., and Rice, S.E. (2008). The kinesin-1 motor protein is regulated
by a direct interaction of its head and tail. Proc. Natl. Acad. Sci. USA
105: 8938–8943.
Finnegan, P.M., Soole, K.L., and Umbach, A.L. (2004). Alternative
mitochondrial electron transport proteins in higher plants. In Advances
in Photosynthesis and Respiration, Plant Mitochondria: From Genome
to Function, Vol. 17, D.A. Day, A.H. Millar, and J. Whelan, eds (Dor-
drecht, The Netherlands: Kluwer Academic Press), pp. 163–230.
Hirokawa, N. (1998). Kinesin and dynein superfamily proteins and the
mechanism of organelle transport. Science 279: 519–526.
Hirokawa, N., Noda, Y., Tanaka, Y., and Niwa, S. (2009). Kinesin
superfamily motor proteins and intracellular transport. Nat. Rev. Mol.
Cell Biol. 10: 682–696.
Itoh, R., Fujiwara, M., and Yoshida, S. (2001). Kinesin-related proteins
with a mitochondrial targeting signal. Plant Physiol. 127: 724–726.
Larkin, P.J. (1976). Purification and viability determinations of plant
protoplasts. Planta 128: 213–216.
Laties, G.G. (1982). The cyanide-resistant, alternative path in higher
plant respiration. Annu. Rev. Plant Physiol. 33: 519–555.
Lee, Y.R.J., Li, Y., and Liu, B. (2007). Two Arabidopsis phragmoplast-
associated kinesins play a critical role in cytokinesis during male
gametogenesis. Plant Cell 19: 2595–2605.
Lee, Y.R.J., and Liu, B. (2004). Cytoskeletal motors in Arabidopsis. Sixty-
one kinesins and seventeen myosins. Plant Physiol. 136: 3877–3883.
Lemasters, J.J., and Holmuhamedov, E. (2006). Voltage-dependent
anion channel (VDAC) as mitochondrial governator—Thinking outside
the box. Biochim. Biophys. Acta 1762: 181–190.
Li, X., Wang, H., Xu, T., Cao, Q., Ren, D., and Liu, G. (2007). Molecular
cloning, expression and biochemical property analysis of AtKP1, a
kinesin gene from Arabidopsis thaliana. Chin. Sci. Bull. 52: 1338–1346.
Li, Y., Zhu, Y., Liu, Y., Shu, Y., Meng, F., Lu, Y., Bai, X., Liu, B., and
Guo, D. (2008). Genome-wide identification of osmotic stress re-
sponse gene in Arabidopsis thaliana. Genomics 92: 488–493.
Linden, M., and Karlsson, G. (1996). Identification of porin as a binding
site for MAP2. Biochem. Biophys. Res. Commun. 218: 833–836.
Liu, M.Y., and Colombini, M. (1992). Regulation of mitochondrial
respiration by controlling the permeability of the outer membrane
through the mitochondrial channel, VDAC. Biochim. Biophys. Acta
1098: 255–260.
Livak, K.J., and Schmittgen, T.D. (2001). Analysis of relative gene
expression data using real-time quantitative PCR and the 2(-D D C(T))
Method. Methods 25: 402–408.
Lu, L., Lee, Y.R.J., Pan, R., Maloof, J.N., and Liu, B. (2005). An internal
motor kinesin is associated with the Golgi apparatus and plays a role
in trichome morphogenesis in Arabidopsis. Mol. Biol. Cell 16: 811–823.
Malcos, J.L., and Cyr, R.J. (2009). Domain complexity of plant kinesins.
In The Plant Cytoskeleton: A Key Tool for Agro-Biotechnology, Y.B.
Blume, W.V. Baird, A.I. Yemets, and D. Breviario, eds (Dordrecht, The
Netherlands: Springer), pp. 351–372.
Marcus, A.I., Li, W., Ma, H., and Cyr, R.J. (2003). A kinesin mutant with
an atypical bipolar spindle undergoes normal mitosis. Mol. Biol. Cell
14: 1717–1726.
Maxwell, D.P., Wang, Y., and McIntosh, L. (1999). The alternative
oxidase lowers mitochondrial reactive oxygen production in plant
cells. Proc. Natl. Acad. Sci. USA 96: 8271–8276.
Meyer, E.H., Tomaz, T., Carroll, A.J., Estavillo, G., Delannoy, E.,
Tanz, S.K., Small, I.D., Pogson, B.J., and Millar, A.H. (2009).
Remodeled respiration in ndufs4 with low phosphorylation efficiency
suppresses Arabidopsis germination and growth and alters control of
metabolism at night. Plant Physiol. 151: 603–619.
Miki, H., Okada, Y., and Hirokawa, N. (2005). Analysis of the kinesin
superfamily: Insights into structure and function. Trends Cell Biol. 15:
467–476.
Moraes, C.T., Ciacci, F., Bonilla, E., Jansen, C., Hirano, M., Rao, N.,
Lovelace, R.E., Rowland, L.P., Schon, E.A., and DiMauro, S.
(1993). Two novel pathogenic mitochondrial DNA mutations affecting
organelle number and protein synthesis. Is the tRNA(Leu(UUR)) gene
an etiologic hot spot? J. Clin. Invest. 92: 2906–2915.
Nangaku, M., Sato-Yoshitake, R., Okada, Y., Noda, Y., Takemura,
R., Yamazaki, H., and Hirokawa, N. (1994). KIF1B, a novel micro-
tubule plus end-directed monomeric motor protein for transport of
mitochondria. Cell 79: 1209–1220.
Ni, C.Z., Wang, H.Q., Xu, T., Qu, Z., and Liu, G.Q. (2005). AtKP1, a
kinesin-like protein, mainly localizes to mitochondria in Arabidopsis
thaliana. Cell Res. 15: 725–733.
Oppenheimer, D.G., Pollock, M.A., Vacik, J., Szymanski, D.B.,
Ericson, B., Feldmann, K., and Marks, M.D. (1997). Essential role
of a kinesin-like protein in Arabidopsis trichome morphogenesis. Proc.
Natl. Acad. Sci. USA 94: 6261–6266.
Pereira, A.J., Dalby, B., Stewart, R.J., Doxsey, S.J., and Goldstein,
L.S.B. (1997). Mitochondrial association of a plus end-directed mi-
crotubule motor expressed during mitosis in Drosophila. J. Cell Biol.
136: 1081–1090.
Rappaport, L., Oliviero, P., and Samuel, J.L. (1998). Cytoskeleton and
mitochondrial morphology and function. Mol. Cell. Biochem. 184:
101–105.
Reddy, A.S., and Day, I.S. (2001). Kinesins in the Arabidopsis genome:
A comparative analysis among eukaryotes. BMC Genomics 2: 2.
Reddy, A.S.N. (2001). Molecular motors and their functions in plants.
Int. Rev. Cytol. 204: 97–178.
Reddy, V.S., Day, I.S., Thomas, T., and Reddy, A.S.N. (2004). KIC, a
novel Ca2+ binding protein with one EF-hand motif, interacts with a
microtubule motor protein and regulates trichome morphogenesis.
Plant Cell 16: 185–200.
Roman, I., Figys, J., Steurs, G., and Zizi, M. (2006). Direct measure-
ment of VDAC-actin interaction by surface plasmon resonance.
Biochim. Biophys. Acta 1758: 479–486.
Rostovtseva, T., and Colombini, M. (1996). ATP flux is controlled by a
voltage-gated channel from the mitochondrial outer membrane. J.
Biol. Chem. 271: 28006–28008.
Rostovtseva, T.K., and Bezrukov, S.M. (2008). VDAC regulation: Role
of cytosolic proteins and mitochondrial lipids. J. Bioenerg. Biomembr.
40: 163–170.
Rostovtseva, T.K., Sheldon, K.L., Hassanzadeh, E., Monge, C.,
Saks, V., Bezrukov, S.M., and Sackett, D.L. (2008). Tubulin binding
blocks mitochondrial voltage-dependent anion channel and regulates
respiration. Proc. Natl. Acad. Sci. USA 105: 18746–18751.
Saks, V.A., Kuznetsov, A.V., Khuchua, Z.A., Vasilyeva, E.V., Belikova,
J.O., Kesvatera, T., and Tiivel, T. (1995). Control of cellular respiration
in vivo by mitochondrial outer membrane and by creatine kinase. A
new speculative hypothesis: possible involvement of mitochondrial-
cytoskeleton interactions. J. Mol. Cell. Cardiol. 27: 625–645.
Sambrook, J., and Russell, D.W. (2001). Molecular Cloning: A Labo-
ratory Manual, 3rd ed. (Cold Spring Harbor, NY: Cold Spring Harbor
Laboratory Press).
Santama, N., Er, C.P., Ong, L.L., and Yu, H. (2004). Distribution and
functions of kinectin isoforms. J. Cell Sci. 117: 4537–4549.
Schmid, M., Davison, T.S., Henz, S.R., Pape, U.J., Demar, M., Vingron,
M., Scholkopf, B., Weigel, D., and Lohmann, J.U. (2005). A gene
expression map of Arabidopsis thaliana development. Nat. Genet. 37:
501–506.
Schwarzer, C., Barnikol-Watanabe, S., Thinnes, F.P., and
Hilschmann, N. (2002). Voltage-dependent anion-selective channel
Kinesin and VDAC in Seed Respiration 1105

(VDAC) interacts with the dynein light chain Tctex1 and the heat-
shock protein PBP74. Int. J. Biochem. Cell Biol. 34: 1059–1070.
Sheen, J. (2002). A transient expression assay using Arabidopsis
mesophyll protoplasts. http://genetics.mgh.harvard.edu/sheenweb/.
(May 27, 2002).
Singha, U.K., Sharma, S., and Chaudhuri, M. (2009). Downregulation
of mitochondrial porin inhibits cell growth and alters respiratory
phenotype in Trypanosoma brucei. Eukaryot. Cell 8: 1418–1428.
Stowers, R.S., Megeath, L.J., Gorska-Andrzejak, J., Meinertzhagen,
I.A., and Schwarz, T.L. (2002). Axonal transport of mitochondria to
synapses depends on milton, a novel Drosophila protein. Neuron 36:
1063–1077.
Tanaka, Y., Kanai, Y., Okada, Y., Nonaka, S., Takeda, S., Harada, A.,
and Hirokawa, N. (1998). Targeted disruption of mouse conventional
kinesin heavy chain, kif5B, results in abnormal perinuclear clustering
of mitochondria. Cell 93: 1147–1158.
Umbach, A.L., Fiorani, F., and Siedow, J.N. (2005). Characterization of
transformed Arabidopsis with altered alternative oxidase levels and
analysis of effects on reactive oxygen species in tissue. Plant Physiol.
139: 1806–1820.
Verhey, K.J., Meyer, D., Deehan, R., Blenis, J., Schnapp, B.J.,
Rapoport, T.A., and Margolis, B. (2001). Cargo of kinesin identified
as JIP scaffolding proteins and associated signaling molecules. J. Cell
Biol. 152: 959–970.
Walter, M., Chaban, C., Schutze, K., Batistic, O., Weckermann, K.,
Nake, C., Blazevic, D., Grefen, C., Schumacher, K., Oecking, C.,
Harter, K., and Kudla, J. (2004). Visualization of protein interactions
in living plant cells using bimolecular fluorescence complementation.
Plant J. 40: 428–438.
Yentur, S., and Leopold, A.C. (1976). Respiratory transition during seed
germination. Plant Physiol. 57: 274–276.
Zhong, R., Burk, D.H., Morrison III, W.H., and Ye, Z.H. (2002).
A kinesin-like protein is essential for oriented deposition of cellulose
microfibrils and cell wall strength. Plant Cell 14: 3101–3117.
1106 The Plant Cell

DOI 10.1105/tpc.110.082420; originally published online March 15, 2011; 2011;23;1093-1106Plant CellGuo-Qin Liu
Xue-Yong Yang, Zi-Wei Chen, Tao Xu, Zhe Qu, Xiao-Di Pan, Xing-Hua Qin, Dong-Tao Ren andRegulates Respiration during Seed Germination at Low Temperature
Kinesin KP1 Specifically Interacts with VDAC3, a Mitochondrial Protein, andArabidopsis
This information is current as of March 28, 2020
Supplemental Data /content/suppl/2011/03/17/tpc.110.082420.DC1.html
References /content/23/3/1093.full.html#ref-list-1
This article cites 57 articles, 26 of which can be accessed free at:
Permissions https://www.copyright.com/ccc/openurl.do?sid=pd_hw1532298X&issn=1532298X&WT.mc_id=pd_hw1532298X
eTOCs http://www.plantcell.org/cgi/alerts/ctmain
Sign up for eTOCs at:
CiteTrack Alerts http://www.plantcell.org/cgi/alerts/ctmain
Sign up for CiteTrack Alerts at:
Subscription Information http://www.aspb.org/publications/subscriptions.cfm
is available at:Plant Physiology and The Plant CellSubscription Information for
ADVANCING THE SCIENCE OF PLANT BIOLOGY © American Society of Plant Biologists