algal flora of korea
TRANSCRIPT

National Institute of Biological ResourcesMinistry of Environment
ISBN 978-89-97462-58-2
Algal Flora of Korea
Vol. 5, No. 2
Freshwater Cyanoprokaryota
IIN
IBR
Algal Flora of Korea
Flora and Fauna of Korea
National Institute of Biological ResourcesMinistry of Environment
Volume 5, Number 2Cyanophyta: Cyanophyceae: Chroococcales, Oscillatoriales
Freshwater Cyanoprokaryota II
China
PB
JG
YG
HB
HN
PN
HWB
GW
GG
CB
CNGB
GB(Ulleung-do)
GN
JN
JJ
JB
HWN
Russia
East Sea
Yellow Sea
South Sea
CB Chungcheongbuk-doCN Chungcheongnam-doGB Gyeongsangbuk-doGG Gyeonggi-doGN Gyeongsangnam-doGW Gangwon-doHB Hamgyeongbuk-doHN Hamgyeongnam-doHWB Hwanghaebuk-doHWN Hwanghaenam-doJB Jeollabuk-doJG Jagang-doJJ Jeju-doJN Jeollanam-doPB Pyeonganbuk-doPN Pyeongannam-doYG Yanggang-do

Algal Flora of Korea
Volume 5, Number 2Cyanophyta: Cyanophyceae: Chroococcales, Oscillatoriales
Freshwater Cyanoprokaryota II
2012
National Institute of Biological ResourcesMinistry of Environment


Algal Flora of Korea
Jong-Geun ParkDaegu University
Volume 5, Number 2Cyanophyta: Cyanophyceae: Chroococcales, Oscillatoriales
Freshwater Cyanoprokaryota II

Copyright ⓒ 2012 by the National Institute of Biological Resources
Published by the National Institute of Biological ResourcesEnvironmental Research Complex, Hwangyeong-ro 42, Seo-guIncheon, 404-708, Republic of Koreawww.nibr.go.kr
All rights reserved. No part of this book may be reproduced, stored in a retrieval system, or transmitted, inany form or by any means, electronic, mechanical, photocopying, recording, or otherwise, without the priorpermission of the National Institute of Biological Resources.
ISBN : 9788997462582-96470Government Publications Registration Number 11-1480592-000366-01
Printed by Junghaengsa, Inc. in Korea on acid-free paper
Publisher : Sang Pal LeeProject Staff : Youn-Bong Ku, Ga Youn Cho, Jin Hee Kim
Published on December 24, 2012
The Flora and Fauna of Korea logo was designed to represent six major target groups of the project includingvertebrates, invertebrates, insects, algae, fungi, and bacteria. The book cover and the logo were designed byJee-Yeon Koo.

Preface
The adoption of the “Nagoya protocol on access to genetic resources and the fair and equitablesharing of benefits arising from their utilization” in 2010 led to the realization of international stan-dardization in the fulfillment of the biological sovereignty and the exploration and preservation ofindigenous biological organisms will play a critical role in enhancing the national development andthe international competitiveness. Most developed countries had already organized the informationof species inhabiting in their territories to claim their severeignty over those biological resources, butin this respect Korea was outpaced by these countries.
In order to effectively secure, use and manage the indigenous biological organisms it is imperativeto systematically understand them at the national level and to organize them to publish in the Floraand Fauna of Korea.
Recognizing the importance of its securement and management in taking the initiative in bio-industry in future, National Institute of Biological Resources of the Ministry of Environment hasbeen publishing the Flora and Fauna of Korea for systematic and efficient management of biologicalresources of our own.
For the last 5 years, professional research groups consisting of relevant professors and the likeconducted systematic surveys and organizations for a variety of and wide range of taxa. As a result,65 issues of Flora and Fauna of Korea, both in Korean and in English, covering 7,709 species and twoissues of world monograph covering 216 species were published and 25 issues of Flora and Fauna ofKorea, both in Korean and in English, covering 1,313 species are published this year.
These efforts serve not only to identify indigenous species living in Korea and to provide the sci-entific evidences and certifications to claim the sovereign rights over indigenous biological resourcesin Korea, but also provide the opportunity to prepare the framework for the biotechnological indus-trialization of biological resources.
Finally I would like to express sincere appreciation for Professor Jong-Geun Park of Daegu Univer-sity who did not spare his efforts to publish Biological Magazine.
Sang Pal LeePresidentNIBR


1
Contents
List of Taxa 3Introduction 5Materials and Methods 7Taxonomic Notes 9
1. Chroococcus limneticus Lemmermann 112. Chroococcus minimus (Keissler) Lemmermann 123. Chroococcus minor (Kützing) Nägeli 134. Chroococcus minutus (Kützing) Nägeli 155. Chroococcus obliteratus Richter 166. Chroococcus planctonicus Bethge 177. Chroococcus prescottii Drouet et Daily 188. Chroococcus subnudus (Hansgirg) Cronberg et Komárek 209. Chroococcus turgidus (Kützing) Nägeli 21
10. Chroococcus vacuolatus Skuja 2211. Aphanocapsa conferta (West et G.S. West) Komárková-Legnerová et Cronberg 2512. Aphanocapsa elachista West et G.S. West 2613. Aphanocapsa grevillei (Berkeley) Rabenhorst 2714. Aphanocapsa incerta (Lemmermann) Cronberg et Komárek 2915. Jaaginema minimum (Gicklhorn) Anagnostidis et Komárek 3216. Jaaginema profundum (Schröter et Kirchner) Anagnostidis et Komárek 3417. Leptolyngbya fontana (Kützing ex Hansgirg) Komárek 3718. Leptolyngbya lagerheimii (Gomont) Anagnostidis et Komárek 3919. Leptolyngbya subtilis (West) Anagnostidis 4120. Heteroleibleinia kossinskajae (Elenkin) Anagnostidis et Komárek 4321. Heteroleibleinia rigidula (Hansgirg) L. Hoffmann 4522. Planktothrix agardhii (Gomont) Anagnostidis et Komárek 4923. Planktothrix compressa (Utermöhl) Anagnostidis et Komárek 5124. Planktothrix isothrix (Skuja) Komárek et Komárková 5125. Phormidium aerugineo-caeruleum (Gomont) Anagnostidis et Komárek 5626. Phormidium amoenum Kützing 5827. Phormidium articulatum (Gardner) Anagnostidis et Komárek 5928. Phormidium autumnale Gomont 6129. Phormidium breve (Kützing ex Gomont) Anagnostidis et Komárek 6230. Phormidium chlorinum (Kützing ex Gomont) Umezaki et Watanabe 6331. Phormidium corium (C. Agardh) Kützing ex Gomont 6532. Phormidium crouaniorum Gomont 6633. Phormidium favosum (Bory) Gomont 6734. Phormidium granulatum (N.L. Gardner) Anagnostidis 6835. Phormidium inundatum Kützing ex Gomont 7036. Phormidium nigrum (Vaucher ex Gomont) Anagnostidis et Komárek 71

37. Phormidium okenii (C. Agardh) Anagnostidis et Komárek 7238. Phormidium taylori (Drouet et Strickland) Anagnostidis 7439. Phormidium terebriforme (C. Agardh ex Gomont) Anagnostidis et Komárek 7540. Phormidium tergestinum (Rabenhorst ex Gomont) Anagnostidis et Komárek 7741. Phormidium violaceum (Wallroth ex Gomont) Anagnostidis et Komárek 7942. Phormidiochaete balearica (Bornet et Flahault) Komárek 8243. Phormidiochaete fusca (Starmach) Komárek 8444. Phormidiochaete nordstedtii (Bornet et Flahault) Komárek 8645. Lyngbya birgei G.M. Smith 9146. Lyngbya hieronymusii Lemmermann 9247. Lyngbya major Meneghini ex Gomont 9348. Lyngbya martensiana Meneghini ex Gomont 9449. Lyngbya nigra C. Agardh ex Gomont 9650. Lyngbya stagnina Kützing ex Gomont 98
Literature Cited 101
Index to Scientific Names 105
2 Algal Flora of Korea∙Freshwater Cyanoprokaryota II

List of Taxa
Class CyanophyceaeOrder Chroococcales
Family Chroococcaceae Rabenhorst 1863Genus Chroococcus Nägeli 1849
Chroococcus limneticus Lemmermann 1898Chroococcus minimus (Keissler) Lemmermann 1904Chroococcus minor (Kützing) Nägeli 1849Chroococcus minutus (Kützing) Nägeli 1849Chroococcus obliteratus Richter 1885Chroococcus planctonicus Bethge 1935Chroococcus prescottii Drouet et Daily 1942Chroococcus subnudus (Hansgirg) Cronberg et Komárek 1994Chroococcus turgidus (Kützing) Nägeli 1849Chroococcus vacuolatus Skuja 1939
Family Merismopediaceae Elenkin 1933Genus Aphanocapsa Nageli 1849
Aphanocapsa conferta (West et G.S. West) Komárková-Legnerová et Cronberg 1993Aphanocapsa elachista West et G.S. West 1894Aphanocapsa grevillei (Berkeley) Rabenhorst 1865Aphanocapsa incerta (Lemmermann) Cronberg et Komárek 1994
Order OscillatorialesFamily Pseudanabaenaceae Anagnostidis et Komárek 1998
Genus Jaaginema Anagnostidis et Komárek 1988Jaaginema minimum (Gicklhorn) Anagnostidis et Komárek 1988Jaaginema profundum (Schröter et Kirchner) Anagnostidis et Komárek 1988
Genus Leptolyngbya Anagnostidis et Komárek 1988Leptolyngbya fontana (Kützing ex Hansgirg) Komárek 2001Leptolyngbya lagerheimii (Gomont) Anagnostidis et Komárek 1988Leptolyngbya subtilis (West) Anagnostidis 2001
Genus Heteroleibleinia (Geitler) L. Hoffmann 1985Heteroleibleinia kossinskajae (Elenkin) Anagnostidis et Komárek 1988Heteroleibleinia rigidula (Hansgirg) L. Hoffmann 1985
Family Phormidiaceae Anagnostidis et Komárek 1998Genus Planktothrix Anagnostidis et Komárek 1988
Planktothrix agardhii (Gomont) Anagnostidis et Komárek 1988Planktothrix compressa (Utermöhl) Anagnostidis et Komárek 1988Planktothrix isothrix (Skuja) Komárek et Komárková 2004
Genus Phormidium Kützing ex Gomont 1892Phormidium aerugineo-caeruleum (Gomont) Anagnostidis et Komárek 1988Phormidium amoenum Kützing 1843Phormidium articulatum (Gardner) Anagnostidis et Komárek 1988
3

Phormidium autumnale Gomont 1892Phormidium breve (Kützing ex Gomont) Anagnostidis et Komárek 1988Phormidium chlorinum (Kützing ex Gomont) Umezaki et Watanabe 1994Phormidium corium (C. Agardh) Kützing ex Gomont 1892Phormidium crouaniorum Gomont 1892Phormidium favosum (Bory) Gomont 1892Phormidium granulatum (N.L. Gardner) Anagnostidis 2001Phormidium inundatum Kützing ex Gomont 1892Phormidium nigrum (Vaucher ex Gomont) Anagnostidis et Komárek 1988Phormidium okenii (C. Agardh) Anagnostidis et Komárek 1988Phormidium taylori (Drouet et Strickland) Anagnostidis 2001Phormidium terebriforme (C. Agardh ex Gomont) Anagnostidis et Komárek 1988Phormidium tergestinum (Rabenhorst ex Gomont) Anagnostidis et Komárek 1988Phormidium violaceum (Wallroth ex Gomont) Anagnostidis et Komárek 1988Phormidiochaete balearica (Bornet et Flahault) Komárek 2001Phormidiochaete fusca (Starmach) Komárek 2001Phormidiochaete nordstedtii (Bornet et Flahault) Komárek 2001
Family Oscillatoriaceae Engler 1898Genus Lyngbya C. Agardh ex Gomont 1892
Lyngbya birgei G.M. SmithLyngbya hieronymusii Lemmermann 1905Lyngbya major Meneghini ex Gomont 1892Lyngbya martensiana Meneghini ex Gomont 1892Lyngbya nigra C. Agardh ex Gomont 1892Lyngbya stagnina Kützing ex Gomont 1892
4 Algal Flora of Korea∙Freshwater Cyanoprokaryota II

Introduction
The Cyanobacteria (blue-green algae) or Cyanoprokaryota (Cyanophyceae) are primitive unicellular,colonial or filamentous organisms. These organisms are characterized by the absence of a nucleusand membrane-bound organelles, and were the first photosynthetic organisms on earth 2.7 billionyears ago.
Two main classification systems of Cyanobacteria are currently used (Table 1); the botanical classi-fication system (Anagnostidis and Komárek 1988, 1990; Komárek and Anagnostidis 1989, 1999, 2005)and the bacteriological classification system (Castenholz 2001; Oren and Tindall 2005).Cyanobacteria were traditionally classified on the basis of their cytomorphological and ecologicalcharacters (Hoffmann 1988), and named according to the International Code of Botanical Nomen-clature (ICBN, Greuter et al. 2000). It is currently impossible to describe a new species of a genusunder the rules of the International Code of Nomenclature of Prokaryotes (ICNP), because of theabsence of ‘Approved Lists’ under the code of the ICNP (Oren et al. 2009).
The taxonomy in this monograph follows the revisions of Komárek and Anagnostidis (Anagnostidisand Komárek 1988, 1990; Komárek and Anagnostidis 1989, 1999, 2005).
5

6 Algal Flora of Korea∙Freshwater Cyanoprokaryota II
Table 1. Classification of cyanobacteria according to bacteriological and botanical systems.
Classification by Bergey’s Manual of Classification by Komárek and Anagnostidis
Systematic Bacteriology
(Castenholz 2001; Oren and Tindall 2005) (Anagnostidis and Komárek 1988, 1990;Komárek and Anagnostidis l989, 1999, 2005)
Subsection (Order) I Chroococcales:
- Unicellular or colonial - Unicellular or colonial- Division by binary fission or budding - 13 Families- 14 Form-genus: Microcystis, Synechococcus, Family Synechococcaceae: Aphanothece,
Synechocystis etc. Cyanobium, Synechococcus etc.- Not classified: Aphanocapsa, Merismopedia, Family Merismopediaea: Apanocapsa, Cyanotetras,
Woronichinia etc. Snowlla, Synechocystis etc.Family Microcystaceae: Eucapsis, Gloecocapsa,
Mycrocystis etc.
Subsection II
- Unicellular or colony- Division by multiple fission or multiple plus
binary fission- 6 Form-genus and Pleurocapsa-“group”
Subsection III Oscillatoriales
- Filament without heterocyst or akinete - Filament without heterocysts or akinete- 17 Form-genus: Lyngbya, Limnothrix, Oscillatoria etc. - 6 Families- Not classified: Planktolyngbya Family Pseudanabaenaceae
Family PhormidiaceaeFamily Oscillatoriaceae
Subsection IV Nostocales
- Filament with heterocyst and without branch - Filament with heterocyst, akinete and pseudobranch- 14 Form-genus: Anabaena, Aphnizomenon etc. Family Nostocaceae: Anabaena, Aphnizomenon etc.

Materials and Methods
Cyanobacteria from Korea were sampled from phytoplankton and periphytic or attached algae inlakes, ponds, reservoirs, streams and water ways. Phytoplankton or metaphytic algae were preparedby the sedimentation of a one liter water sample or netting (mesh size 20 μm). The samples werepreserved in formaldehyde solution to a final concentration of 2.5%.
Material was examined using a DIC (differential interference contrast) compound microscope(Nikon Eclipse 80i, Japan). Images were captured with digital camera (Nikon DS-5M, Japan) and acomputer. The resolution of digital image was 2560×1920 pixels. The terminology used followedKomárek and Anagnostidis (1999), Komárek and Anagnostidis (2005), McGregor (2007), Whitton(2011) and (Guiry and Guiry 2012).
7


Taxonomic Notes
Family Chroococcaceae Rabenhorst 1863: 57, 69.
Coccal, colonial, cells rarely solitary, usually microscopic, few-celled, rarely many-celled, irregular-spherical colonies, rarely in short rows, clustered densely together or distant from one another,mucilaginous colonies with irregularly or more or less packet-like shape, colonies sometimes aggre-gated into a macroscopic mass and layered. Modifications of cell arrangements in old colonies areusable for genus definitions and result from dominant type of division in later generations. Mucilagefine, diffluent or firm, stratified, gelatinous, mainly forming distinctly delimited envelopes or sheaths.Cells spherical, oval, hemispherical, rounded-polyhedral or irregular. Cell division successively inthree or more different planes, later forming small packet-like colonies, cells reaching more or lessthe original form and size before next division, but not into a spherical form, nanocytes lacking.Reproduction by disintegration of colonies.
GENERA AND SPECIES: There are 16 genera and 143 species are currently accepted as taxonomicallyvalid taxa (Guiry and Guiry 2012). Genera containing none or one species number 7 taxa. Majorgenera are Chroococcus (86 species), Asterocapsa (16 species), Cyanosarcina (8 species) and Gloeocapsopsis(6 species).
DISTRIBUTION: Worldwide, planktonic, epiphytic, aerophytic in freshwater or brackish water.KEY REFERENCE: Chung (1993), Cronberg and Komárek (1994), Komárek and Anagnostidis (1999),
Hindák (2008), Whitton (2011).
Genus Chroococcus Nägeli 1849: 45.
Colonies commonly microscopic, irregular or more or less spherical, gelatinous colonies, usuallywith a small cell number (2-16), many-celled agglomerations, never in rows or in regular arrange-ment, sometimes forming multicellular colonies sometimes in packet-like, 2-8 celled groups.Mucilage colorless or yellowish, usually concentrically lamellate, often delimited, well-marked atthe margin, only in certain species (subg. Limnococcus) irregularly disposed with homogeneous,amorphous, hyaline mucilaginous matrix; cells often surrounded by narrow individual envelopes,following the cell outline. Cells rarely spherical, at first subspherical, widely oval or spherical, laterhemispherical or in the form of a segment of the sphere, grey, grey-green, blue-green, olive-green,yellowish, orange, reddish or violet, homogeneous, granular, only in few planktonic species withaerotopes. Cell division in three perpendicular planes, later more or less irregular (in other direc-tions), cells grow more or less into the original size and shape before the next division, but notalways into the original shape, daughter cells form their own gelatinous envelopes. Widely distri-buted, free-living, epiphytic or in periphyton, freshwater in ponds, lakes, moorland waters, thermalwaters, soil biotopes and aerophytic. Many species are incorrectly identified and repeatedly misin-
9

terpreted. Ecology is particularly important for a correct determination.HOLOTYPE: Chroococcus rufescens (Kützing) Nägeli 1849.
SPECIES: Of the 135 species (infraspecific) reported up to date, 86 species are currently acceptedas taxonomically valid (Guiry and Guiry 2012).
DISTRIBUTION: Worldwide.KEY REFERENCE: Cronberg and Komárek (1994), Komárek and Anagnostidis (1999), Komárek (2003),
Hindák (2008), Whitton (2011).REMARKS: This genus is generally divided into two subgenus. Planktonic species from the sub-
genus Limnococcus are common in temperate and northern water bodies and include C. microscopicus,C. minimus, C. dispersus, C. distance and C. limneticus. Most species from the subgenus Chroococcusare metaphytic (Komárek 2003).
Key to the species of genus Chroococcus
1. Mucilaginous envelopes enlarged, sometimes diffluent, not lamellate, in outline spherical orirregular; cells grow more or less into the original spherical form before the next division (subg.Limnococcus)·····························································································································2
- Mucilaginous envelopes follow the outline of cells more or less closely or that of cell groups, thick-ened, diffluent or delimited at the surface, sometimes lamellate; cells do not grow the originalspherical form before the next division (subg. Chroococcus)·······················································6
2. Cells with facultative aerotopes, planktonic species··································································3- Cells always without aerotopes································································································43. 1.5-2.7 μm in diameter····························································································C. vacuolatus- 3.0-4.0 μm in diameter·························································································C. planctonicus4. Cells regularly arranged in more or less cubic formation, 5.0-9.0 μm in diameter, mucilaginous
envelopes lamellate···································································································C. prescottii- Cells irregularly arranged within colonies, usually attaining a spherical shape before next division,
mucilaginous envelopes not lamellate······················································································55. Cells 1.7-3.0 μm in diameter······················································································C. minimus- Cells 6.0-12.0 μm in diameter··················································································C. limneticus6. Cells up to 10.0 μm in diameter································································································7- Cells over 10.0 μm in diameter··································································································97. Cells 3.0-4.0 μm in diameter, thin and diffluent envelopes, metaphytic in swampy biotopes·············································································································································C. minor
- Cells over 4.0 μm in diameter···································································································88. Envelopes usually not distinctly delimited, not lamellate, metaphytic and often solitary, also in
plankton·····················································································································C. minutus- Envelopes usually distinctly delimited, slightly lamellate, not very wide, 4.0-10.0 μm in diameter·······························································································································C. obeliteratus
9. Envelopes not delimited and not lamellate, diffluent at the margin, grass-green or blue-green·····································································································································C. subnudus- Envelopes always delimited and usually slightly lamellate, up to 32.0 μm in diameter, bright blue-
green··························································································································C. turgidus
10 Algal Flora of Korea∙Freshwater Cyanoprokaryota II

1. Chroococcus limneticus Lemmermann 1898: 153 (Fig. 1A-D).
Chung 1993: 197. f. 26-a. f. 26-b. Komárek and Anagnostidis 1999: 290. f. 382. Komárek 2003: 95. f.14-A-c. Hu and Wei 2006: 88. pl. II-16-10. Hindák 2008: f. 267-270. Whitton 2011: 52. pl. 11D.
SYNONYM: Chroococcus limneticus var. carneus (Chodat) Lemmermann 1904.Anacystis thermalis f. major (Lagerheim) Drouet et Daily 1956.
Colonies free-floating, microscopic, with wide gelatinous slime, oval, rarely almost spherical orirregular, with up to 40 cells but usually fewer, usually with cells slightly distant from one another,irregularly arranged or within the homogeneous mucilage in 2-4 celled groups. Colonial mucilagecolorless, delicate, structureless, homogeneous, clearly delimited or diffluent at the margin, outermargin of colony usually distinct, sometimes scarcely visible, not lamellate, distant from the cells,individual cell envelopes developed, colorless, not lamellate, narrow, sometimes diffluent and indis-tinct envelopes. Cells spherical, after division hemispherical, bright blue-green or greyish blue-green, olive-green or yellowish, with finely granular protoplast, without aerotopes, 6.0-12.2 (-22.0)
Cyanophyceae: Chroococcales: Chroococcaceae 11
Fig. 1. A-D. Chroococcus limneticus.
25 μm
A B
C D

μm in diameter. Cell division in three perpendicular planes, later more or less irregular, cells growmore or less into the original size and shape before the next division, but not always into the origi-nal shape.
TYPE LOCALITY: Various in Germany.DISTRIBUTION: Cosmopolitan.KOREA: Han River (Chung et al. 1968); Nakdong River (Cho et al. 1993).SPECIMEN EXAMINED: GB, 01.x.2011.ECOLOGY: Planktonic in ponds and lakes, secondarily in rivers, usually maxima in summer and
in the autumn, present throughout the year, but never form bloom, characteristic of mesotrophicwater.
REMARKS: This species belongs to subgenus Limnococcus. Variable species, occurring in numerouslocal morphotypes and ecotypes, often different cell size in diameter (Komárek and Anagnostidis1999; Whitton 2011).
2. Chroococcus minimus (Keissler) Lemmermann 1904 (Fig. 2A-D)
Komárek and Anagnostidis 1999: 286. f. 371. Hu and Wei 2006: 88. pl. II-16-9. Whitton 2011: 54.
BASIONYM: Chroococcus minutus var. minimus Keissler 1901.SYNONYM: Gloeocapsa minima (Keissler) Hollerbach 1938.Chroococcus dispersus var. minor G.M. Smith 1920.Gloeocapsa minima f. smithii Hollerbach, Kosinskaja et Poljanskij 1953.
Colonies free floating, microscopic, with gelatinous slime, spherical or irregular, typically with 2-8 cells, more or less regularly arranged or in small groups. Colonial mucilage colorless, homogeneous,diffluent at the margin, individual cell envelopes present or absent, but sometimes recognizable withstaining, not lamellate. Cells spherical or hemispherical after division, pale blue-green or grayishblue-green, 1.7-3.0 μm in diameter, without aerotopes. Cell division in three perpendicular planes,later more or less irregular, cells grow more or less into the original size and shape before the nextdivision, but not always into the original shape.
TYPE LOCALITY: Austria.DISTRIBUTION: Temperate zones over the whole of the northern hemisphere, probably also in
south America and Africa.KOREA: Galgok stream and Incheon river in JN (Lee et al. 2008).SPECIMEN EXAMINED: GB, 01.x.2011; Haengsinkyo in GW (37°51′05′′N, 128°47′29′′E), 20.xi.2012.ECOLOGY: Planktonic in oligotrophic or mesotrophic reservoirs and lakes, sometimes loosely
associated with submerged plants, rarely among algae in metaphyton of stagnant waters.REMARKS: This species belongs to subgenus Limnococcus. Data reporting cells 8 μm in diameter
are clearly in error (Komárek and Anagnostidis 1999).
12 Algal Flora of Korea∙Freshwater Cyanoprokaryota II
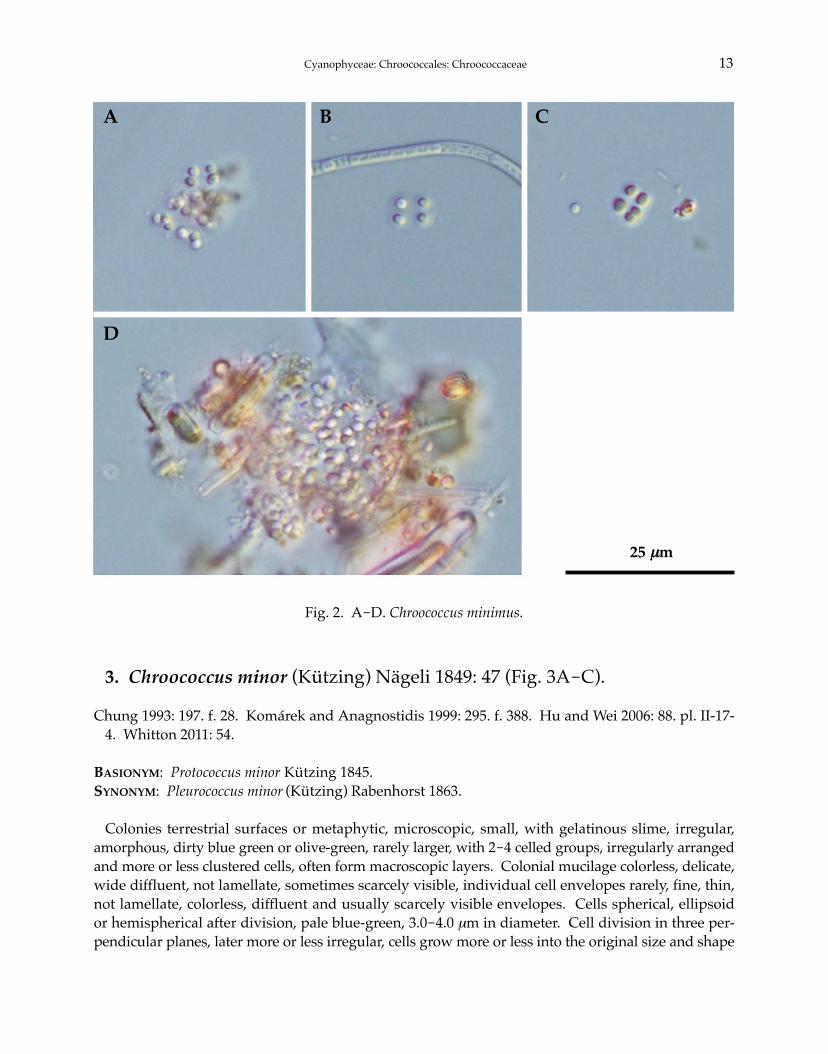
3. Chroococcus minor (Kützing) Nägeli 1849: 47 (Fig. 3A-C).
Chung 1993: 197. f. 28. Komárek and Anagnostidis 1999: 295. f. 388. Hu and Wei 2006: 88. pl. II-17-4. Whitton 2011: 54.
BASIONYM: Protococcus minor Kützing 1845.SYNONYM: Pleurococcus minor (Kützing) Rabenhorst 1863.
Colonies terrestrial surfaces or metaphytic, microscopic, small, with gelatinous slime, irregular,amorphous, dirty blue green or olive-green, rarely larger, with 2-4 celled groups, irregularly arrangedand more or less clustered cells, often form macroscopic layers. Colonial mucilage colorless, delicate,wide diffluent, not lamellate, sometimes scarcely visible, individual cell envelopes rarely, fine, thin,not lamellate, colorless, diffluent and usually scarcely visible envelopes. Cells spherical, ellipsoidor hemispherical after division, pale blue-green, 3.0-4.0 μm in diameter. Cell division in three per-pendicular planes, later more or less irregular, cells grow more or less into the original size and shape
Cyanophyceae: Chroococcales: Chroococcaceae 13
Fig. 2. A-D. Chroococcus minimus.
25 μm
A B C
D

14 Algal Flora of Korea∙Freshwater Cyanoprokaryota II
Fig. 3. A-C. Chroococcus minor.
25 μm
A
B C

before the next division, but not always into the original shape.
TYPE LOCALITY: An Brettern und Holz in und ausser dem Wasser (Nordhausen, Erfurt District,Germany fide Drouet and Moe 1956) (Silva et al. 1996).
DISTRIBUTION: Temperate zones and probably also tropical zones.KOREA: JB (Chung 1976); Paldang reservoir in GG (Chang and Jeon 1996); Lake Chungju (Kim et
al. 1998); Togyo reservoir in CW (Lee and Yoon 2003); Yeoju in GG (Lim and Lee 2008a).SPECIMEN EXAMINED: Jjokjibul in GN (35°32′41′′N, 128°23′49′′E), 17.ix.2010; Haengsinkyo in GW
(37°51′05′′N, 128°47′29′′E), 20.xi.2012; Duyangli in GN (35°04′49′′N, 127°53′43′′E), 02.xii.2012.ECOLOGY: On wet terrestrial surfaces, e.g., stones and wood near the water surface of streams
and swamps, metaphytic in bogs, swamps and ponds with submerged vegetation, in rice-fields.REMARKS: Data from thermal springs probably refer to another species (Komárek and Anagnostidis
1999).
4. Chroococcus minutus (Kützing) Nägeli 1849: 46 (Fig. 4A-D).
Chung 1993: 197. f. 29-a. f. 29-b. f. 29-c. Komárek and Anagnostidis 1999: 296. f. 391. Hindák 2008:f. 265-266. Whitton 2011: 54. pl. 11F.
BASIONYM: Protococcus minutus Kützing 1843.SYNONYM: Gloeocapsa minuta (Kützing) Hollerbach 1938.Chroococcus virescens Hantzsch 1865.
Cells free-floating, solitary or small, microscopic, more or less spherical or oval, irregular if theouter margin is more diffluent, with few-celled colonies, usually 2-8 cells. Colonial mucilage color-less, homogeneous or slightly lamellate, delimited, rarely somewhtat diffluent, sometimes withindistinct margin (required staining), individual cell envelopes rarely, fine, thin, colorless, notlamellate, diffluent and usually scarcely visible envelopes. Cells spherical or hemispherical afterdivision, pale blue-green or greyish blue-green, homogeneous or granulate content, (4.0-) 5.0-10.0(-12.0) μm in diameter. Cell division in three perpendicular planes, later more or less irregular, cellsgrow more or less into the original size and shape before the next division, but not always into theoriginal shape.
TYPE LOCALITY: In einem Wasserglase, (Nordhausen, Erfurt District, Germany) (Silva et al. 1996).Notes: This type locality was first recorded by Drouet and Daily (1956) (Silva et al. 1996).
DISTRIBUTION: Temperate zones but probably cosmopolitan distribution.KOREA: Han River (Chung et al. 1968); JB (Chung 1976); JN (Wui and Kim 1987); Lake Chungju in
CB (Kim et al. 1998); Yeoju in GG (Lim and Lee 2008a).SPECIMEN EXAMINED: Duyangli in GN (35°04′49′′N, 127°53 43′′E), 02.xii.2012.ECOLOGY: Planktonic, tychoplanktonic or metaphytic in oligotrophic up to slightly eutrophic
lakes, ponds, water reservoirs and swamps, often in littoral, but never in masses, mostly shallowerwater bodies and submerged macrophytes, often found in blackish waters.
REMARKS: Polymorphic species with numerous morphotypes and ecotypes (Komárek and Anag-nostidis 1999).
Cyanophyceae: Chroococcales: Chroococcaceae 15
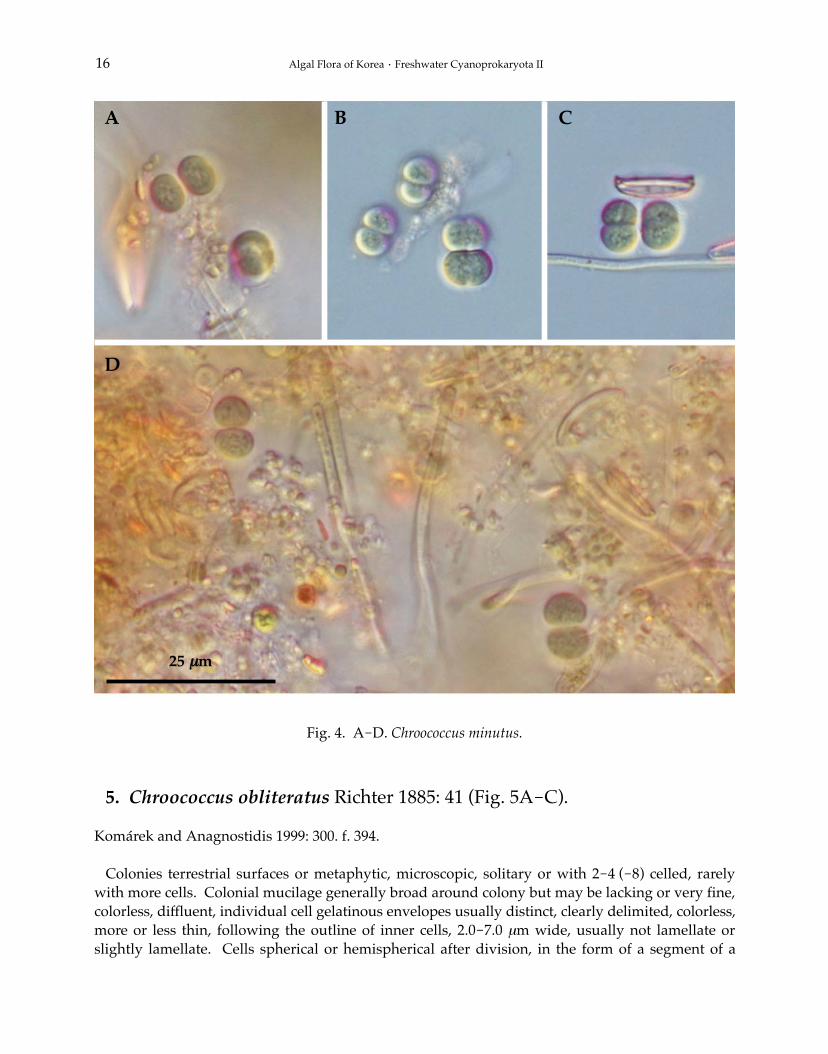
5. Chroococcus obliteratus Richter 1885: 41 (Fig. 5A-C).
Komárek and Anagnostidis 1999: 300. f. 394.
Colonies terrestrial surfaces or metaphytic, microscopic, solitary or with 2-4 (-8) celled, rarelywith more cells. Colonial mucilage generally broad around colony but may be lacking or very fine,colorless, diffluent, individual cell gelatinous envelopes usually distinct, clearly delimited, colorless,more or less thin, following the outline of inner cells, 2.0-7.0 μm wide, usually not lamellate orslightly lamellate. Cells spherical or hemispherical after division, in the form of a segment of a
16 Algal Flora of Korea∙Freshwater Cyanoprokaryota II
Fig. 4. A-D. Chroococcus minutus.
25 μm
A B C
D

sphere, pale blue-green or olive-green, yellowish, (4.0-) 6.0-10.0 μm in diameter. Cell division inthree perpendicular planes, later more or less irregular, cells grow more or less into the original sizeand shape before the next division, but not always into the original shape.
TYPE LOCALITY: Leipzig, Germany.DISTRIBUTION: North temperate zones but not abundance.KOREA: Previously unrecorded species.SPECIMEN EXAMINED: Nammaeji in GB (35°49′44′′N, 128°44′32′′E), 30.xiii.2010.ECOLOGY: Shallow waters with submerged vegetation, in the littoral of lakes and ponds, in
swamps, sometimes also in brackish waters, occasionally in plankton of shallow water bodies.REMARKS: Cosmopolitan distribution is likely (Komárek and Anagnostidis 1999). Often misinter-
preted as other species, e.g., the small type of Chroococcus turgidus (Kützing) Nägeli 1849.
6. Chroococcus planctonicus Bethge 1935 (Fig. 6A, B)
Komárek and Anagnostidis 1999: 286. f. 372.
Colonies free floating or metaphytic, with gelatinous slime, more or less irregular and flat, rarelyspherical, with several or many cells, to 100 μm in diameter, with randomly arranged cells, whichare sometimes in groups of 2-4 together. Colonial mucilage colorless, diffluent, usually almostinvisible, individual cell envelopes lacking. Cells spherical or slightly elongate, later hemisphericalor slightly irregular-rounded, pale yellowish, with aerotopes, 3.0-4.0 μm in diameter. Cell divisionusually in two perpendicular planes, resulting in flattened colonies, but sometimes in more planes,cells grow more or less into the original size and shape before the next division, but not alwaysinto the original shape.
Cyanophyceae: Chroococcales: Chroococcaceae 17
Fig. 5. A-C. Chroococcus obliteratus.
25 μm
A B C
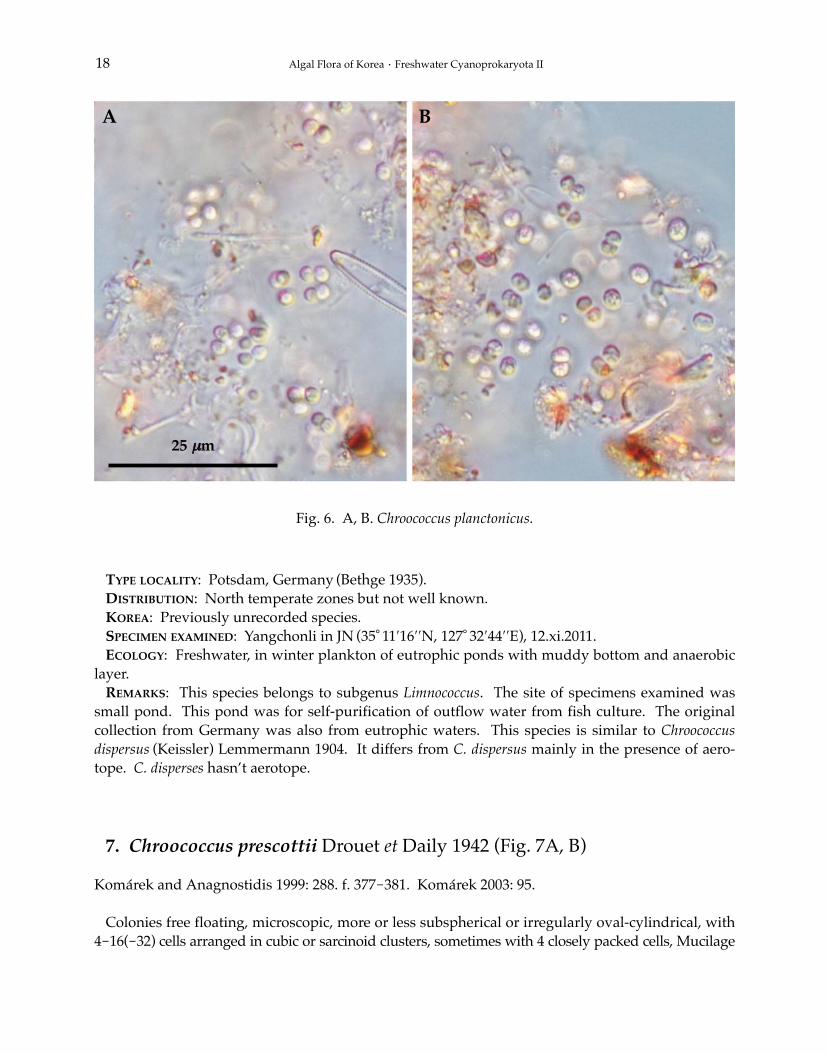
TYPE LOCALITY: Potsdam, Germany (Bethge 1935).DISTRIBUTION: North temperate zones but not well known.KOREA: Previously unrecorded species.SPECIMEN EXAMINED: Yangchonli in JN (35°11′16′′N, 127°32′44′′E), 12.xi.2011.ECOLOGY: Freshwater, in winter plankton of eutrophic ponds with muddy bottom and anaerobic
layer.REMARKS: This species belongs to subgenus Limnococcus. The site of specimens examined was
small pond. This pond was for self-purification of outflow water from fish culture. The originalcollection from Germany was also from eutrophic waters. This species is similar to Chroococcusdispersus (Keissler) Lemmermann 1904. It differs from C. dispersus mainly in the presence of aero-tope. C. disperses hasn’t aerotope.
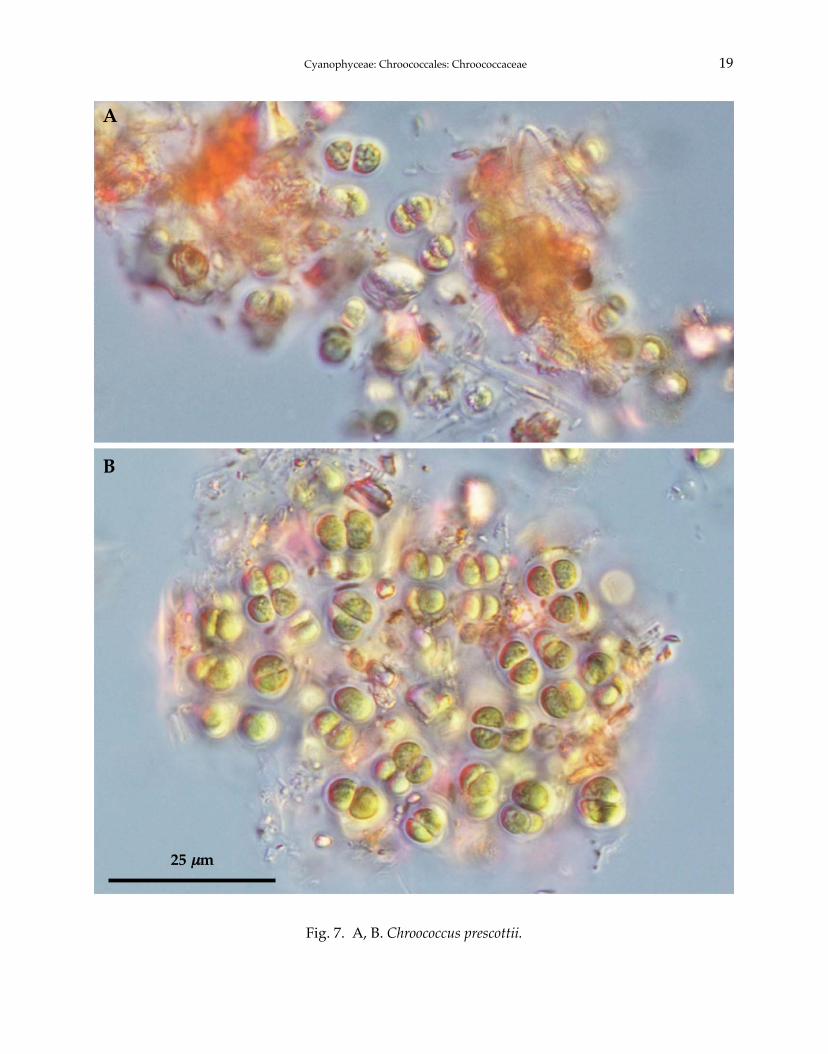
7. Chroococcus prescottii Drouet et Daily 1942 (Fig. 7A, B)
Komárek and Anagnostidis 1999: 288. f. 377-381. Komárek 2003: 95.
Colonies free floating, microscopic, more or less subspherical or irregularly oval-cylindrical, with4-16(-32) cells arranged in cubic or sarcinoid clusters, sometimes with 4 closely packed cells, Mucilage
18 Algal Flora of Korea∙Freshwater Cyanoprokaryota II
Fig. 6. A, B. Chroococcus planctonicus.
25 μm
A B
Cyanophyceae: Chroococcales: Chroococcaceae 19
Fig. 7. A, B. Chroococcus prescottii.
25 μm
A
B

generally broad around colony, clearly delimited, sometimes rectangular, hyaline, often slightlylamellate envelopes, individual cell envelopes colorless, not lamellate. Cells spherical or hemisph-erical after division, with bright blue-green, finely granular content, 5.0-8.0 μm in diameter. Celldivision in three perpendicular planes, cells grow more or less into the original size and shapebefore the next division.
TYPE LOCALITY: Massachusetts, USA.DISTRIBUTION: North temperate zones but not abundance.KOREA: Previously unrecorded species.SPECIMEN EXAMINED: Hwangjeji in GB (35°54′00′′N, 128°00′00′′E), 07.vii.2007.ECOLOGY: Freshwater, typchoplanktonic in clear acidic reservoirs.REMARKS: This species belongs to subgenus Limnococcus. This species is sometimes associated
with typical Chroococcus turgidus (Komárek and Anagnostidis 1999). Planktonic species from thesubgenus Limnococcus are common in temperate and northern water bodies. C. prescotti is morecommon in cold and slightly acidic waters (Komárek 2003). Relationship with the genus Eucapsisneed further study.
8. Chroococcus subnudus (Hansgirg) Cronberg et Komárek 1994(Fig. 8A, B)
Lukavsky 1994: 66. pl. I. f. 18. Komárek and Anagnostidis 1999: 306. f. 404.
SYNONYM: Chroococcus turgidus var. subnudus Hansgirg 1892.Gloeocapsa turgida f. subnuda (Hansgirg) Hollerbach.
Colonies benthic or metaphytic, microscopic, with solitary or usually 2-4 celled, rarely up to 32
20 Algal Flora of Korea∙Freshwater Cyanoprokaryota II
Fig. 8. A, B. Chroococcus subnudus.
20 μm
A B

cells. Colonial mucilage thin, diffluent, hyaline and not lamellate, slightly diffluent at the margin,but copying the cell outline, sometimes indistinct (required staining), envelops around cells thin,hyaline, not layered, Cells oval-spherical or subspherical-rounded, hemispherical after division, paleolive-green or greenish, to 32 μm in diameter. Cell division usually in two perpendicular planes,later more or less irregular, cells grow more or less into the original size and shape before the nextdivision, but not always into the original shape,
TYPE LOCALITY: Stechowic, Czech Republic.DISTRIBUTION: Temperate zone of northern hemisphere.KOREA: Previously unrecorded species.SPECIMEN EXAMINED: Hyogyeongri in JN (35°23′20′′N, 127°27′41′′E), 10.xi.2011.ECOLOGY: In moorland waters, bogs and swamps, on wet rocks, in the littoral of lakes, usually in
mountain region.REMARKS: The site of specimens examined was mountain valley. The bottom of valley was gravel.
This species belongs to subgenus Chroococcus. It is a large species and similar to Chroococcus turgidus(Kützing) Nägeli.
9. Chroococcus turgidus (Kützing) Nägeli 1849: 46 (Fig. 9A).
Yamaghshi and Akiyama 1985a: 3: 11. Chung 1993: 198. f. 31-a. f. 31-b. Komárek and Anagnostidis1999: 306. f. 407. Komárek 2003: 95. f. 14-A-h. Hu and Wei 2006: 85. pl. II-16-1. Hindák 2008: f.279-280. Whitton 2011: 54. pl. 6A, 11A, B.
BASIONYM: Protococcus turgidus Kützing 1846.SYNONYM: Gloeocapsa turgida (Kützing) Hollerbach 1938.Protococcus turgidus Kützing 1846.Trochiscia dimidiata Kützing 1834.Chroococcus dimidiatus (Kützing) Nägeli 1849.Anacystis dimidiata (Kützing) Drouet et Daily 1952.
Colonies benthic or metaphytic, microscopic, usually one or 2-8 celled, rarely solitary, occasionallyup to 32 cells. Colonial mucilage lacking, however, older envelopes usually surround the wholecolony, group mucilaginous envelopes usually clearly delimited, firm, replicating the cell outlineor that of group of cells, colorless, rarely pale brown in British population, generally striate, to 10μm wide, individual cells with hyaline, lamellate sheaths. Cells spherical or subspherical, ovoid orwidely oval, hemispherical or in the form of a segment of a sphere after divison, blue-green, rarelyolive-green, yellowish or slightly pinkish, homogeneous or finely granular, (6.0-) 8.0-32.0 (-45.0)μm in diameter. Cell division in three perpendicular planes, later more or less irregular, cells growmore or less into the original size and shape before the next division, but not always into the originalshape.
TYPE LOCALITY: “Zwischen Oscillarieen und Lyngbyeen”, (Freidburg im Breisgau, Baden-Württem-berg, Germany) (Silva et al. 1996). Notes: This type locality was first described by Drouet and Daily(1956) (Silva et al. 1996).
Cyanophyceae: Chroococcales: Chroococcaceae 21

DISTRIBUTION: Temperate zones but likely to be cosmopolitan distribution.KOREA: Han River (Chung et al. 1968); JB (Chung 1976); JN (Wui and Kim 1987); Gori in GN (Kim
et al. 1992); Lake Chungju (Kim et al. 1998); Pyeongtaek reservoir in GG (Shin 2003); Togyoreservoir in CW (Lee and Yoon 2003); Galgok stream and Incheon river in JN (Lee et al. 2008); Yeojuin GG (Lim and Lee 2008a, 2008b).
SPECIMEN EXAMINED: Gokgangkyo in GB (36°07′20′′N, 129°20′51′′E), 19.vi.2011.ECOLOGY: Common species in detritus on wet rocks, in bogs, swamps, most frequent in oligotro-
phic or mesotrophic waters with a lot of submerged vegetation, in the littoral of stagnant waters,ponds and lakes, absent from eutrophic waters.
REMARKS: This species belongs to subgenus Chroococcus. A wide spread and variable species inmoist terrestrial habitats. In general, the largest cell forms are characteristic of low nutrient waters(Whitton 2011). Data from thermal springs, endolithic or marine habitats etc. probably refer to otherspecies (Komárek and Anagnostidis 1999). Data from Gori in GN (Kim et al. 1992) is marine habitat.
10. Chroococcus vacuolatus Skuja 1939: 42 (Fig. 10A-C).
Komárek and Anagnostidis 1999: 284. f. 370.
SYNONYM: Gloeocapsa vacuolata (Skuja) Hollerbach, Kosinskaja et Poljanskij 1953.
Colonies benthic or metaphytic, tychoplanktonic or planktonic, microscopic, with gelatinous slime,with 4-16 (-32) cells, rarely small groups of 2-3 cells, irregular in outline, with more or less densely
22 Algal Flora of Korea∙Freshwater Cyanoprokaryota II
Fig. 9. A. Chroococcus turgidus.
25 μm
A
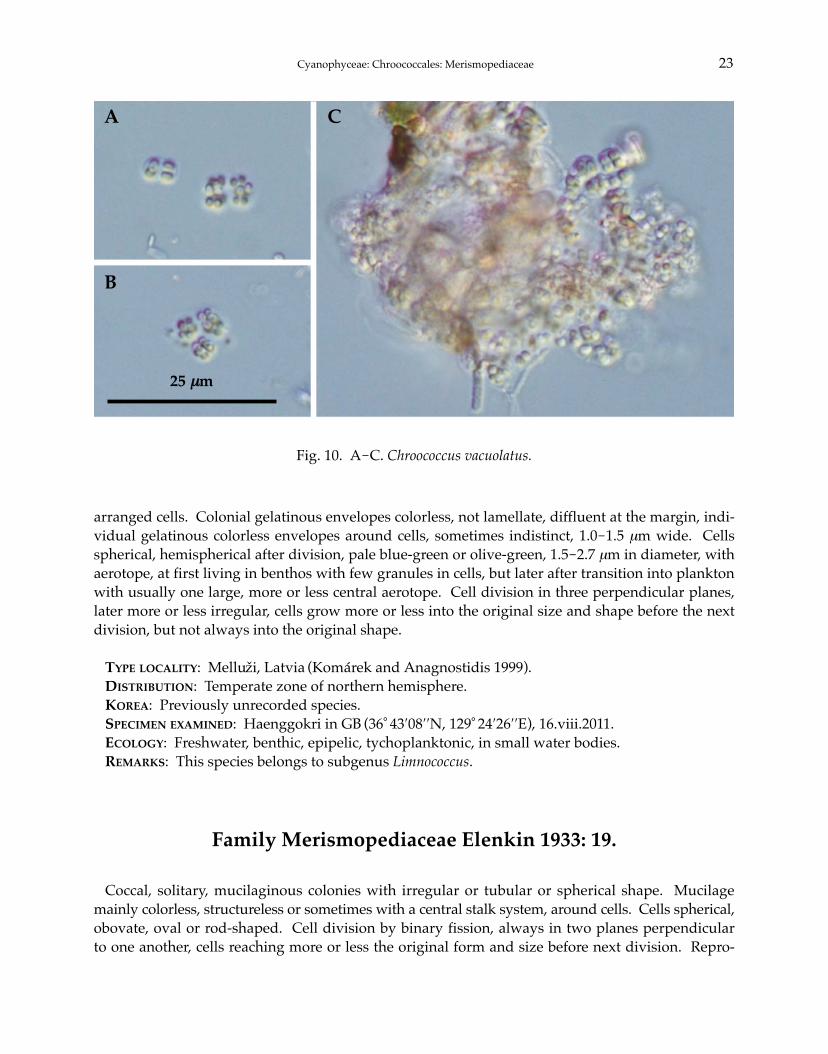
arranged cells. Colonial gelatinous envelopes colorless, not lamellate, diffluent at the margin, indi-vidual gelatinous colorless envelopes around cells, sometimes indistinct, 1.0-1.5 μm wide. Cellsspherical, hemispherical after division, pale blue-green or olive-green, 1.5-2.7 μm in diameter, withaerotope, at first living in benthos with few granules in cells, but later after transition into planktonwith usually one large, more or less central aerotope. Cell division in three perpendicular planes,later more or less irregular, cells grow more or less into the original size and shape before the nextdivision, but not always into the original shape.
TYPE LOCALITY: Melluzi, Latvia (Komárek and Anagnostidis 1999).DISTRIBUTION: Temperate zone of northern hemisphere.KOREA: Previously unrecorded species.SPECIMEN EXAMINED: Haenggokri in GB (36°43′08′′N, 129°24′26′′E), 16.viii.2011.ECOLOGY: Freshwater, benthic, epipelic, tychoplanktonic, in small water bodies.REMARKS: This species belongs to subgenus Limnococcus.
Family Merismopediaceae Elenkin 1933: 19.
Coccal, solitary, mucilaginous colonies with irregular or tubular or spherical shape. Mucilagemainly colorless, structureless or sometimes with a central stalk system, around cells. Cells spherical,obovate, oval or rod-shaped. Cell division by binary fission, always in two planes perpendicularto one another, cells reaching more or less the original form and size before next division. Repro-
Cyanophyceae: Chroococcales: Merismopediaceae 23
Fig. 10. A-C. Chroococcus vacuolatus.
A C
B
25 μm

duction by disintegration of colonies.
GENERA AND SPECIES: Some 23 genera and 178 species are currently accepted as taxonomicallyvalid taxa (Guiry and Guiry 2012). Genera containing none or one species number 11 taxa. Themajor genera are Aphanocapsa (53 species), Merismopedia (36 species), Synechocystis (29 species) andCoelosphaerium (15 species).
DISTRIBUTION: Worldwide, planktonic, epiphytic, aerophytic in freshwater or brackish water.KEY REFERENCE: Chung (1993), Cronberg and Komárek (1994), Komárek and Anagnostidis (1999).
Komárek and Komárková (2002), Sant’anna et al. (2004), Hindák (2008), Whitton (2011).REMARKS: This family is generally divided into two subfamiles. Subfamily Merismopedioideae:
Cells solitary or in irregular or tabular in one layer colonies. Subfamily Gomphosphaerioideae:Spherical colonies with cells arranged radially and more or less peripherally.
Key to the genera of family Merismopediaceae
1. Cells spherical and solitary, or in irregular or tabular colonies··················································2- Spherical colonies with cells arranged radially and more or less peripheral monolayer············52. Cells solitary and free-floating················································································Synechocystis- Spherical cells in colonies or in groups·····················································································33. Colonies irregular and mucilaginous with irregular arranged cells··························Aphanocapsa- Colonies tabular with regular arranged cells in perpendicular rows·········································44. Cells spherical or hemispherical after division·······················································Merismopedia- Cells elongate and slightly ovoid···············································································Cyanotetras5. Cells spherical and attached to the ends of thin pseudodichotomously divaricate stalks, usually
clearly visible without staining, colonies spherical and free-floating······························Snowella- Cells slightly elongated spherical or obovoid, attached to the ends of more or less wide mucilagi-
nous stalks, which are sometimes invisible without staining, colonies free-floating····················································································································································Woronichinia
Genus Aphanocapsa Nageli 1849: 52.
Colonies benthic or peiphytic, planktonic or metaphytic, microscopic or macroscopic, spherical,irregular or flat, cell arrangement irregular, sparsely or densely. Mucilage mainly colorless andmore or less homogeneous, usually diffuse with an indistinct margin, cells without individual enve-lopes or occasionally with narrow. Cells spherical or hemispherical after division, sometimes withgranular content, pale or bright blue-green without aerotopes. Freshwater planktonic species 0.5-3.0 μm in diameter. Cell division by binary fission in two perpendicular planes in successive gene-rations, and reach the original size and shape before the next division. Reproduction by disintegra-tion of colonies.
HOLOTYPE: Aphanocapsa parietina Nägeli 1849.
SPECIES: About 104 species are reported up to date, 53 species are currently accepted as taxono-mically valid taxa (Guiry and Guiry 2012).
DISTRIBUTION: Worldwide, planktonic in freshwater.
24 Algal Flora of Korea∙Freshwater Cyanoprokaryota II

KEY REFERENCE: Nägeli (1849). Cronberg and Komárek (1994), Komárek and Anagnostidis (1999),Komárek (2003), Whitton (2011).
Key to the species of Aphanocapsa
1. Planktonic species····················································································································2- In stagnant waters, periphytic or benthic, amorphous colonies, cells 3.0-6.0 μm in diameter,
bright blue-green········································································································A. grevillei2. Cells densely arranged in colonies····························································································3- Cells loosely distributed, the space between cells more than the diameter of one cell···············43. Cells 1.0−2.0 μm in diameter, joined into compact, more or less spherical colony··········A. incerta- Cells 1.0-1.5 μm in diameter, densely arranged in irregular colonies····················································································································································A. holsatica (See Vol 5. No.1)
4. Cells 0.8-1.2 μm in diameter and loosely arranged in irregular colonies········································································································································A. delicatissima (See Vol 5. No.1)
- Cells over 1.2 μm in diameter···································································································55. Cells 1.3-2.0 μm in diameter, very sparsely and irregular distributed within more or less spherical
colonies······················································································································A. elachista- Cells 1.6-2.0 (-2.4) μm in diameter, regularly distributed within irregular colonies, with diffluent
and scarcely visible margin·························································································A. conferta
11. Aphanocapsa conferta (West et G.S. West)Komárková-Legnerová et Cronberg 1994: 21 (Fig. 11A).
Chung 1993: 194. f. 13-a. f. 13-b. Yamaghshi and Akiyama 1985: 3: 3. Komárek and Anagnostidis1999: 155. f. 180, 183. Whitton 2011: 41.
BASIONYM: Aphanocapsa elachista var. conferta West et G.S. West 1912.SYNONYM: Microcystis pulverea f. conferta (West et G.S. West) Elenkin 1938.
Colonies free floating, planktonic, microscopic to 80 μm in diameter, more or less spherical orirregular, with scattered, more or less evenly densely arranged cells. Mucilage delicate, homoge-neous, colorless, structureless, indistinct at margin. Cells spherical, pale blue-green, or grayish,sometimes in twos, without aerotopes, 1.5-2.0 (-2.4) μm in diameter. Cell division by binary fissionin two perpendicular planes in successive generations, and reach the original size and shape beforethe next division. Reproduction by disintegration of colonies.
TYPE LOCALITY: Loch Katrine, Scotland.DISTRIBUTION: Temperate zones.KOREA: Galgok stream and Incheon river in JN (Lee et al. 2008).SPECIMEN EXAMINED: Sowolji in GB (35°56′29′′N, 128°50′08′′E), 31.viii.2010.ECOLOGY: Planktonic in eutrophic waters bodies.REMARKS: Reference records from Chung (1993) and Yamaghshi and Akiyama (1985) were reported
as Aphanocapsa elachista var. conferta West et G.S. West. Whitton (2011) recognize that A. conferta is
Cyanophyceae: Chroococcales: Merismopediaceae 25

the synonym of A. elachista West et G.S. West.
12. Aphanocapsa elachista West et G.S. West 1894: 276 (Fig. 12A, B).
Chung 1993: 194. f. 12. Komárek and Anagnostidis 1999: 154. f. 174. Hu and Wei 2006: 52. pl. II-6-1. Whitton 2011: 41. pl. 9A.
SYNONYM: Microcystis pulverea f. elachista (West et G.S. West) Elenkin 1938.Microcystis elachista (West et G.S. West) Compère 1967.Aphanocapsa elachista var. planctonica G.M. Smith 1920.Aphanocapsa planctonica (G.M. Smith) Komárek et Anagnostidis 1995.
Colonies free-floating, microscopic, spherical or oval, ellipsoid, to 100 μm in diameter, with looselyarranged solitary cells, after division in twos. Mucilage homogeneous, scarce, colorless, usuallydiffluent. Cells spherical, gray or pale blue-green, without aerotopes, 1.3-2.0 μm in diameter. Celldivision by binary fission in two perpendicular planes in successive generations, and reach theoriginal size and shape before the next division. Reproduction by disintegration of colonies.
TYPE LOCALITY: Dominica.
26 Algal Flora of Korea∙Freshwater Cyanoprokaryota II
Fig. 11. A. Aphanocapsa conferta; B. Aphanocapsa grevillei.
25 μm
A B

DISTRIBUTION: Common in tropical countries, rare in warm regions of the temperate zone.KOREA: Han River (Chung et al. 1968); Beopsu marsh in GN (Kim 2008); Yeoju in GG (Lim and
Lee 2008a).SPECIMEN EXAMINED: Yongdamho in JB (35°56′32′′N, 127°31′39′′E), 01.viii.2011.ECOLOGY: Planktonic in eutrophic water bodies, very common in tropical regions and usually
during the summer season in temperate regions.REMARKS: Data from other biotopes (e.g. bark of trees in W. Africa) probably belong to another
species (Komárek and Anagnostidis 1999). Whitton (2011) recognize that A. conferta (West et G.S.West) Komárková-Legnerová et Cronberg and A. planctonica (G.M. Smith) Komárek et Anagnostidisare the synonyms of this species.
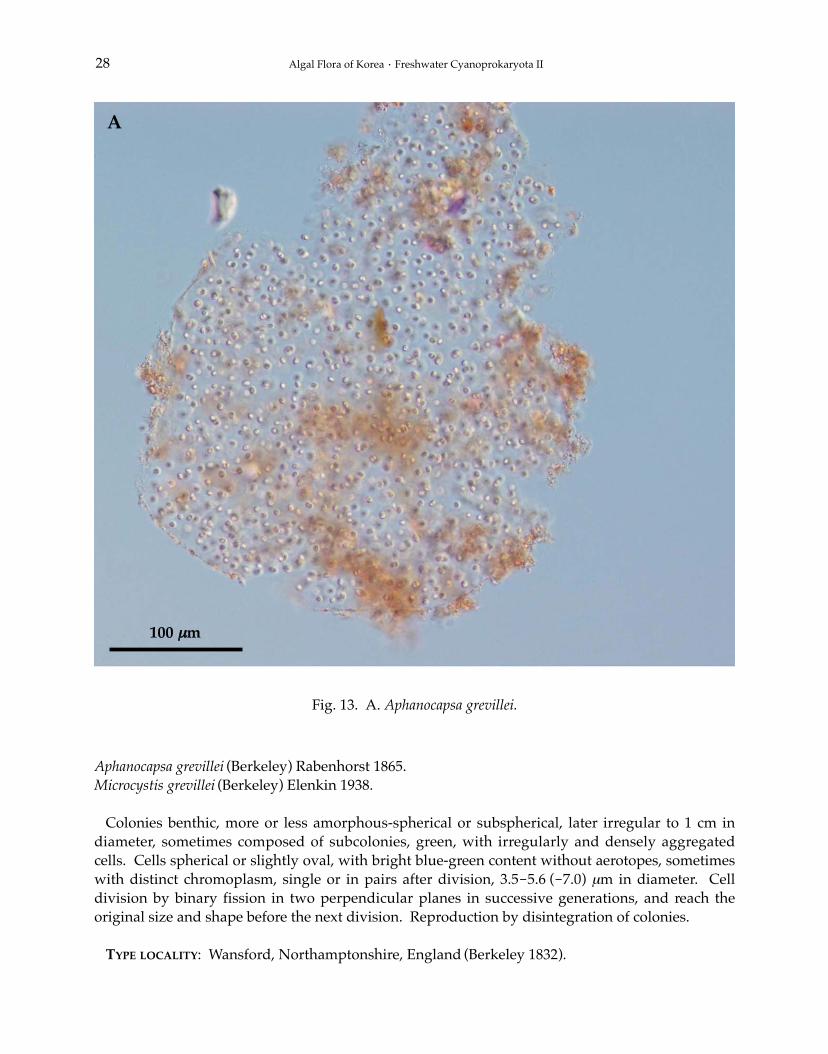
13. Aphanocapsa grevillei (Berkeley) Rabenhorst 1865: 50 (Figs. 11B, 13A).
Chung 1993: 195. f. 15-a. f. 15-b. Komárek and Anagnostidis 1999: 159. f. 194. Hu and Wei 2006: 55.pl. II-7-1. Whitton 2011: 41. pl. 9B.
BASIONYM: Palmella grevellei Berkeley 1832.SYNONYM: Coccochloris grevellei (Berkeley) Hassall 1845.Microcystis grevillei (Berkeley) Elenkin 1938.
Cyanophyceae: Chroococcales: Merismopediaceae 27
Fig. 12. A, B. Aphanocapsa elachista.
25 μm
A B
Aphanocapsa grevillei (Berkeley) Rabenhorst 1865.Microcystis grevillei (Berkeley) Elenkin 1938.
Colonies benthic, more or less amorphous-spherical or subspherical, later irregular to 1 cm indiameter, sometimes composed of subcolonies, green, with irregularly and densely aggregatedcells. Cells spherical or slightly oval, with bright blue-green content without aerotopes, sometimeswith distinct chromoplasm, single or in pairs after division, 3.5-5.6 (-7.0) μm in diameter. Celldivision by binary fission in two perpendicular planes in successive generations, and reach theoriginal size and shape before the next division. Reproduction by disintegration of colonies.
TYPE LOCALITY: Wansford, Northamptonshire, England (Berkeley 1832).
28 Algal Flora of Korea∙Freshwater Cyanoprokaryota II
Fig. 13. A. Aphanocapsa grevillei.
100 μm
A

DISTRIBUTION: Temperate zone.KOREA: Han River (Chung et al. 1968); Beopsu marsh in GN (Kim 2008).SPECIMEN EXAMINED: Yangchonli in GN (35°11′16′′N, 127°32′44′′E), 12.xi.2011; Nakdong river (36°
32′14′′N, 128°16′04′′E).ECOLOGY: Commonly benthic in moorland waters and acidic swamps, occasionally on peaty soil
and on granitic substrata.REMARKS: Planktonic records concern other species (mainly Microcystis smithii Komárek et Anag-
nostidis), records from thermal biotopes and tropical regions need revision (Komárek and Anagno-stidis 1999).
14. Aphanocapsa incerta (Lemmermann) Cronberg et Komárek 1994(Fig. 14A-C)
Yamaghshi and Akiyama 1984: 1: 60. Komárek and Anagnostidis 1999: 151. f. 172. Whitton 2011:41. pl. 15B.
BASIONYM: Polycystis incerta Lemmermann 1899.SYNONYM: Microcystis incerta (Lemmermann) Lemmermann 1907.Microcystis pulverea var. incerta (Lemmermann) Crow 1923.Microcystis pulverea f. incerta (Lemmermann) Elenkin 1938.Anacystis incerta (Lemmermann) F.E. Drouet et W.A. Daily 1952.
Colonies microscopic, spherical, sometimes slightly flattened, rarely somewhat irregular, with agreat number of irregularly and densely aggregated cells; the agglomerations of cells are yellowish-green. Mucilage colorless or slightly yellow, structureless, diffluent and indistinct at the margin (inculture completely diffluent and cells are solitary). Cells spherical with finely granulate yellowishgreen or pale blue-green content, without aerotopes. 0.5-2.0 (-2.7) μm in diameter. Cell divisionby binary fission in two perpendicular planes in successive generations, and reach the original sizeand shape before the next division. Reproduction by disintegration of colonies.
TYPE LOCALITY: Baselitz near Kamenz, Sachsen, Germany.DISTRIBUTION: Cosmopolitan.KOREA: Han River (Chung et al. 1968); JB (Chung 1976); Nakdong River (Lee et al. 1997; Cho et al.
1993); JN (Wui and Kim 1987).SPECIMEN EXAMINED: Yongdamho in JB (35°56′32′′N, 127°31′39′′E), 01.viii.2011.ECOLOGY: Planktonic in eutrophic water bodies.REMARKS: Data from other biotopes (e.g., subaerophytic) are probably erroneous (Komárek and
Anagnostidis 1999). Reference records in Korea: Han River (Chung et al. 1968); JB (Chung 1976);Nakdong River (Lee et al. 1997; Cho et al. 1993); JN (Wui and Kim 1987) as Microcystis incerta (Lem-mermann) Lemmermann. Reference record from Yamaghshi and Akiyama (1984) was reported asMicrocystis incerta (Lemmermann) Lemmermann.
Cyanophyceae: Chroococcales: Merismopediaceae 29

30 Algal Flora of Korea∙Freshwater Cyanoprokaryota II
Fig. 14. A-C. Aphanocapsa incerta.
25 μm
A B
C

Family Pseudanabaenaceae Anagnostidis et Komárek 1998: 379.
Trichomes solitary or forming mat-like colonies, without sheath or with fine and colorless sheath(Leptolyngbya, Leibleinia and Planktolyngbya), cylindrical, straight or slightly flexuous, unbranched,1.0-4.0 μm wide. Cells mainly longer than wide, rarely isodiametric or shorter than wide, with orwithout aerotopes. Heterocytes and akinetes absent. Cell division by binary fission in the perpen-dicular long axis of a trichome, cells reaching the original size before the next division. Reproduc-tion by disintegration of trichomes without help of necridic cells.
GENERA AND SPECIES: Some 17 genera and 413 species are currently recognized as taxonomicallyvalid taxa (Guiry and Guiry 2012). Genera containing none or one species are five taxa. Majorgenera are Leptolyngbya (151 species), Spirulina (59 species), Geitlerinema (37 species), Pseudanabaena(37 species) and Jaaginema (28 species), Heteroleibleinia (23 speces), Limnothrix (21 species), Tapinothrix(19 sepces) and Planktolyngbya (15 species).
DISTRIBUTION: Worldwide, planktonic, benthic, periphytic in freshwater, brackish and salinewaters.
KEY REFERENCE: Anagnostidis and Komárek (1988), Komárek and Anagnostidis (2005), Willame etal. (2006), McGregor (2007), Hindák (2008), Whitton (2011).
REMARKS: This family is generally divided into four subfamiles. Subfamily Pseudanabaenoideae:Trichomes without sheaths, straight or wavy. Subfamily Spirulinoideae: Trichomes without sheaths,regularly and usually screw-like coiled. Subfamily Leptolyngbyoideae: Trichomes with thin, fineor firm sheaths, isopolar, free-living, or solitary, or in clusters in mats. Subfamily Heteroleibleinio-deae: Trichomes with thin, fine or firm sheaths, heteropolar, with one end attached to the substate.
Key to the genera of family Pseudanabaenaceae
1. Trichomes without sheaths, or very fine diffluent mucilaginous envelopes·······························2- Trichomes with fine or firm sheaths in vegetative stage····························································52. Trichomes with or without constricted at cross-walls, cells with or without polar or central aero-
topes·········································································································································3- Trichomes usually cylindrical and not constricted at cross-walls, without aerotopes but sometimes
with granules in cells················································································································43. Trichomes solitary and straight, usually constricted at cross-walls, cells without aerotopes······································································································································Pseudanabaena
- Trichomes solitary, usually containing aerotopes, not constricted or slightly constricted at cross-walls··························································································································Limnothrix
4. Cell contents homogeneous and rarely granular, mainly in clusters but sometimes in mats·············································································································································Jagginema
- Cell contains granules, mainly in mats but sometimes in clusters·····························Geitlerinema5. Filaments planktonic, sheath extended beyond the both ends of trichomes··········Planktolyngbya- Filaments benthic, creeping on the substratum or forming mat, or attached to the substratum····66. Trichomes isopolar, benthic, creeping on the substratum or forming mat··································7- Trichomes cylindrical, never attenuated towards ends········································Heterobeibleinia7. Filaments usually epiphytic··························································································Leibleinia- Filaments forming clusters or mat···········································································Leptolyngbya
Cyanophyceae: Oscillatoriales: Pseudanabaenaceae 31

Genus Jaaginema Anagnostidis et Komárek 1988: 395.
Trichomes usually solitary or entangled in clusters into small colonies, forming thin, membranaeousthallus, rarely forming macroscopically visible mats, more or less flexuous, always without sheaths,cylindrical and isopolar, usually waved or coiled, always immotile. Trichomes up to 3 μm wide,uniseriate, usually not constricted at cross-walls, usually not narrowed to the ends, sometimes atten-uated at the ends, not capitate. Cells cylindrical, elongated, mostly longer than wide, up to 10 timeslonger than wide, rarely isodiametric, cell content homogeneous, sometimes with solitary granules,pale blue-green or grey, yellowish or olive-green, without aerotopes, thylakoids localized probablyparietally. Apical cells mostly rounded or narrowed, rarely pointed or conical, always withoutcalyptra, exceptionally with thickened outer cell wall. Cell division by the crosswise binary fission,perpendicular to the long axis of a trichome, daughter cells grow up to the original size before thenext division. Reproduction by fragmentation of trichomes, without necridic cells.
HOLOTYPE: Jaaginema subtilissimum (Kützing ex De Toni) Anagnostidis et Komárek
NUMBER OF SPECIES: Of the 32 species (infraspecific) reported to date, 28 species are currentlyaccepted as taxonomically valid taxa (Guiry and Guiry 2012).
DISTRIBUTION: Probably worldwide.KEY REFERENCE: Geitler (1932), Anagnostidis and Komárek (1988), Komárek and Anagnostidis
(2005), McGregor (2007).REMARKS: This genus belongs to subfamily Pseudanabaenoideae. The presence of immotile tri-
chomes is a generic character. Major species are benthic organisms, growing on the bottom ofdiverse water biotopes, pools, lakes, reservoirs with rich vegetation of water plants, common inmetaphyton. Several species are known from mineral, thermal or saline waters.
Key to the species of genes Jaaginema
1. Freshwater species, benthic and tychoplanktonic in lakes, cells more or less 3.0-4.0 μm×2.0 μm····································································································································J. profundum
- Freshwater species, benthic and metaphytic among water plant, cells yellow-green, 5.0-6.0 μm×2.0 μm····················································································································J. minimum
15. Jaaginema minimum (Gicklhorn) Anagnostidis et Komárek 1988: 396(Fig. 15A-D).
Komárek and Anagnostidis 2005: 114. f. 114.
BASIONYM: Oscillatoria minima Gicklhorn 1921.
Trichomes benthic or metaphytic, usually solitary, cylindrical and isopolar, waved or more or lessspirally coiled, yellow-green, with very delicate mucilaginous sheath, 2 μm wide, uniseriate, notconstricted at cross-walls, not attenuated at the ends, always immotile. Cells cylindrical and elon-gated, longer than wide, 5.0-6.0 μm long, cell content homogeneous, bright blue-green, without
32 Algal Flora of Korea∙Freshwater Cyanoprokaryota II
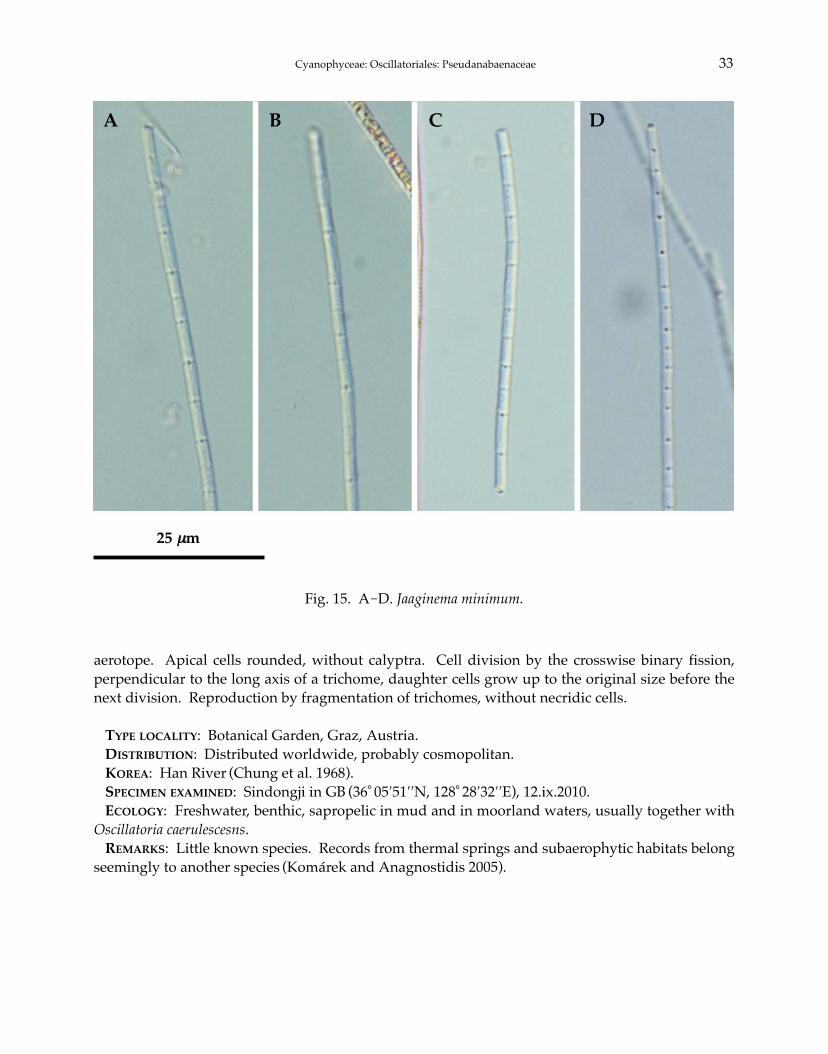
aerotope. Apical cells rounded, without calyptra. Cell division by the crosswise binary fission,perpendicular to the long axis of a trichome, daughter cells grow up to the original size before thenext division. Reproduction by fragmentation of trichomes, without necridic cells.
TYPE LOCALITY: Botanical Garden, Graz, Austria.DISTRIBUTION: Distributed worldwide, probably cosmopolitan.KOREA: Han River (Chung et al. 1968).SPECIMEN EXAMINED: Sindongji in GB (36°05′51′′N, 128°28′32′′E), 12.ix.2010.ECOLOGY: Freshwater, benthic, sapropelic in mud and in moorland waters, usually together with
Oscillatoria caerulescesns.REMARKS: Little known species. Records from thermal springs and subaerophytic habitats belong
seemingly to another species (Komárek and Anagnostidis 2005).
Cyanophyceae: Oscillatoriales: Pseudanabaenaceae 33
Fig. 15. A-D. Jaaginema minimum.
25 μm
A B C D
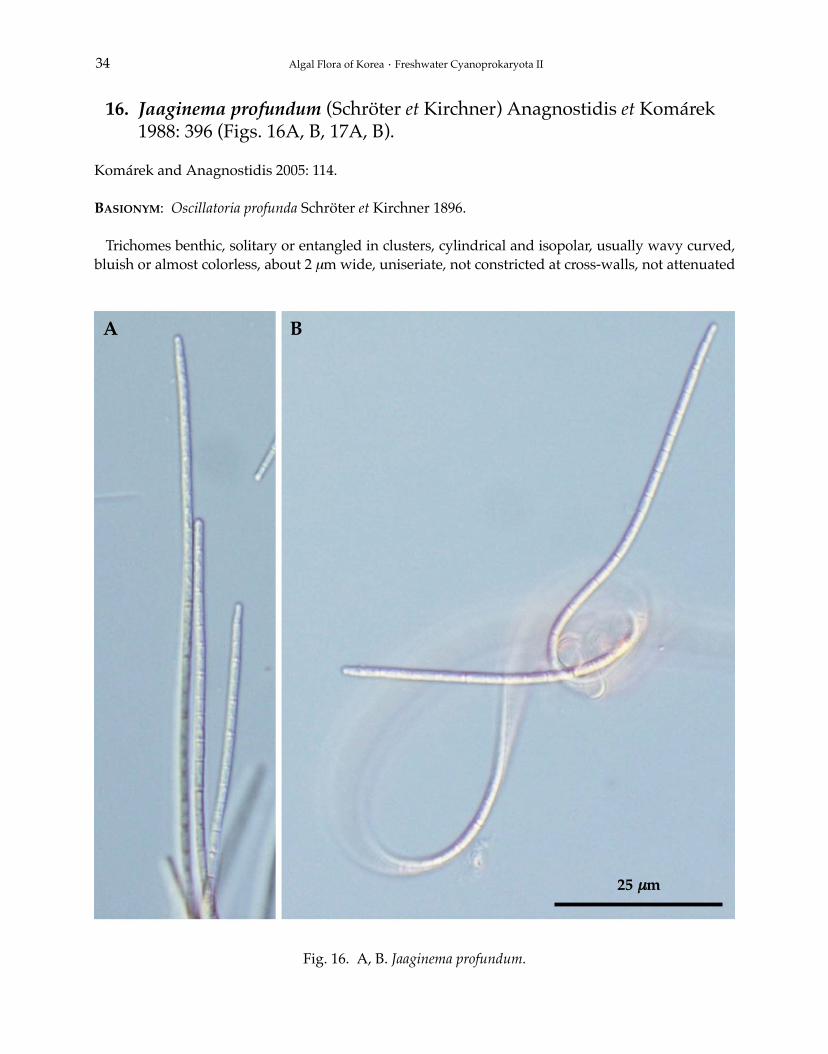
16. Jaaginema profundum (Schröter et Kirchner) Anagnostidis et Komárek1988: 396 (Figs. 16A, B, 17A, B).
Komárek and Anagnostidis 2005: 114.
BASIONYM: Oscillatoria profunda Schröter et Kirchner 1896.
Trichomes benthic, solitary or entangled in clusters, cylindrical and isopolar, usually wavy curved,bluish or almost colorless, about 2 μm wide, uniseriate, not constricted at cross-walls, not attenuated
34 Algal Flora of Korea∙Freshwater Cyanoprokaryota II
Fig. 16. A, B. Jaaginema profundum.
25 μm
A B

at the ends, immotile. Cells cylindrical and elongated, 1-3 times longer than wide, 3.0-6.0 μm long,cell content homogeneous, almost colorless, without aerotope. Apical cells widely rounded, with-out calyptra. Cell division by the crosswise binary fission, perpendicular to the long axis of a tric-
Cyanophyceae: Oscillatoriales: Pseudanabaenaceae 35
Fig. 17. A, B. Jaaginema profundum.
50 μm
100 μm
A
B

home, daughter cells grow up to the original size before the next division. Reproduction by frag-mentation of trichomes, without necridic cells.
TYPE LOCALITY: Langenargen, Bodensee, Germany.DISTRIBUTION: Worldwide.KOREA: Geumgok reservoir in GG (Park et al. 2006).SPECIMEN EXAMINED: Okjeongho in JB (35°32′27′′N, 127°06′35′′E), 01.viii.2011.ECOLOGY: Freshwater, benthic on mud in stagnant waters.REMARKS: This species was described from lake in Germany (Komárek and Anagnostidis 2005).
Reports from soil in India probably are of another species (Komárek and Anagnostidis 2005). Dis-tribution records from Korea is the records as Oscillatoria profunda Schröter et Kirchner.
Genus Leptolyngbya Anagnostidis et Komárek 1988: 390.
Filaments benthic or metaphytic, floating or more or less attached to the substratum, solitary orcoiled into clusters and macroscopic tangled mats, isopolar and long, more or less flexuous, finelywaved, rarely almost straight, 0.5-3.5 μm wide. Sheath firm and colorless, thin, enveloping usuallyone trichome and opened at the apex, unbranched or rarely false branching. Trichomes cylindrical,unconstricted or constricted at the cross-walls, usually not attenuated to the ends or slightly atten-uated, immotile or motile hormogonia. Cells cylindrical, isodiametric or longer than wide, sometimesshorter than wide, more or less homogeneous content, pale blue-green, grayish, olive-green, with-out aerotopes. Heterocytes and akinetes absent. Apical cells conical or rounded, without thickenedcell wall or calyptra. Cells division by symmetrical, crosswise binary fission, cells reaching theoriginal size before next division. Reproduction by fragmentation without the assistance of necridiccells.
HOLOTYPE: Leptolyngbya boryana (Gomont) Anagnostidis et Komárek 1988.
NUMBER OF SPECIES: Of the 169 species (infraspecific) are reported up to date, 151 species are cur-rently accepted as taxonomically valid taxa (Guiry and Guiry 2012).
DISTRIBUTION: Worldwide, freshwater or subaerophytic.KEY REFERENCE: Anagnostidis and Komárek (1988), Komárek and Anagnostidis (2005), McGregor
(2007).REMARKS: This genus belongs to subfamily Leptolyngbyoideae. Leptolyngbya species are very com-
mon in soils and in periphyton and metaphyton of freshwater and halophilous biotopes, severalspecies are known from thermal and mineral springs or from aerophytic rocky sites or walls; charac-teristic species grow endogloeically in mucilage and in the colonial slime of other algae (Guiry andGuiry 2012). Very common genus with many described species, but all need taxonomic reevaluationand definition of diacritical features; many species were described under the generic names Lyngbya,Phormidium and Plectonema (Guiry and Guiry 2012). This genus is generally divided into two sub-genus. Subgenus Leptolyngbya: cells more or less isodiametric or slightly shorter or 2 times longerthan wide. Trichomes disintegrate by the assistance of necridic cells. Subgenus Protolyngbya: Cellsalways distinctly longer than wide. Trichomes disintegrate without necridic cells.
36 Algal Flora of Korea∙Freshwater Cyanoprokaryota II

Key to the species of genes Leptolynbya
1. Freshwater species, benthic, periphytic, metaphytic, epiphytic, sheaths wider than trichomes,trichomes not constricted at cross-walls, 1.8-3.2 μm wide············································L. fontana
- Freshwater species, sheath attached to trichomes·····································································22. Usually spirally and single granules at the cross-walls (rarely missing), trichomes 1.2-1.8 μm
wide························································································································L. lagerheimii- Usually straight, without single granules at the cross-walls······················································33. Trichomes not constricted at cross-walls, without granules, benthic, metaphytic in mesotrophic
and eutrophic waters, often among other algae and water plants, more or less 1.8 μm wide············································································································································L. subtilis
- Trichomes constricted or sometimes indistinctly constricted at cross-walls, apical cells cylindricaland rounded, usually joined to stony substrata, in clusters or irregular mats, cells 0.8-2.2 μm×0.5-2.5 μm·······················································································L. foveloara (See Vol 5. No. 1)
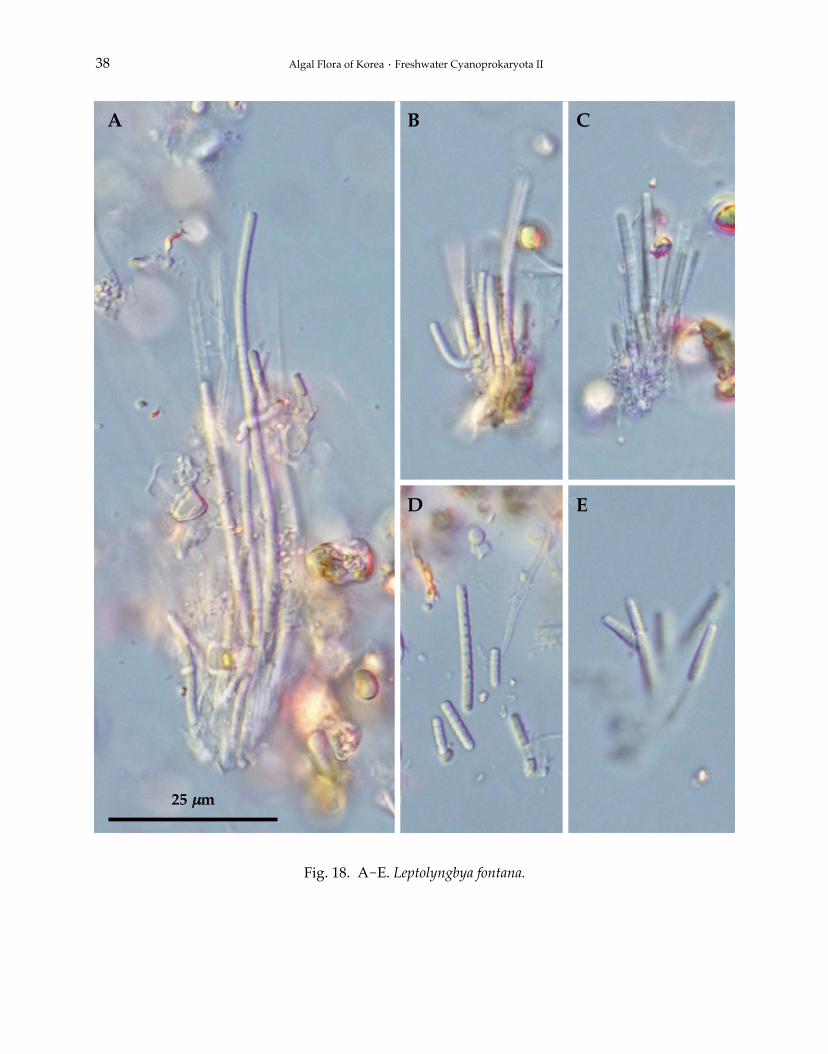
17. Leptolyngbya fontana (Kützing ex Hansgirg) Komárek 2001: 374(Fig. 18A-E).
Komárek and Anagnostidis 2005: 184.
BASIONYM: Lyngbya fontana Kützing ex Hansgirg 1892.SYNONYM: Leptothrix fontana Kützing 1843, nom. inval.Hypheothrix fontana (Kützing) Rabenhorst 1865.Lyngbya fontana (Kützing) Hansgirg 1891.Lyngbya fontana Hansgirg ex Hansgirg 1892.Heteroleibleinia fontana (Hansgirg ex Hansgirg) Anagnostidis et Komárek 1988.
Filaments benthic or metaphytic, more or less attached to the substratum, solitary or in clusters,intricate, rarely irregularly fasciculated and forming to 4.0-8.0 μm long, gelatinous, dark olive-green thallus. Sheaths thin, colorless, attached to trichomes, or slightly distant, one trichome andopened at the apex, unbranched. Trichomes cylindrical, pale blue-green or olive-green, unconstrictedat the cross-walls, not attenuated to the ends, 2.5-3.3 μm wide. Cells cylindrical, more or less isodia-metric, or slightly shorter than wide, homogeneous, without aerotopes, Heterocytes and akinetesabsent. Apical cells rounded, without calyptra. Cells division by symmetrical, crosswise binaryfission, cells reaching the original size before next division. Reproduction by fragmentation withoutthe assistance of necridic cells.
TYPE LOCALITY: Kölme near Halle, Germany (type of Leptothrix fontana Kützing, nom. nval.)DISTRIBUTION: Worldwide.KOREA: Previously unrecorded species.SPECIMEN EXAMINED: Haenggokri in GN (36°43′09′′N, 129°24′26′′), 19.xi.2011; Daesankyo in GN
(35°38′54′′N, 127°56′14′′), 02.xii.2011.ECOLOGY: Freshwater, on stones, wood, etc., usually in cold and oligotrophic, flowing waters, in
springs, wells, streams.
Cyanophyceae: Oscillatoriales: Pseudanabaenaceae 37
38 Algal Flora of Korea∙Freshwater Cyanoprokaryota II
Fig. 18. A-E. Leptolyngbya fontana.
A B C
D E
25 μm

REMARKS: This species belongs to Subgenus Leptolyngbya. Occurs in the older literature in twodifferent concept (cf. Heterleibleinia fontana), based on the same type. Both names must be taxono-mically and nomenclaturally solved (Komárek and Anagnostidis 2005).
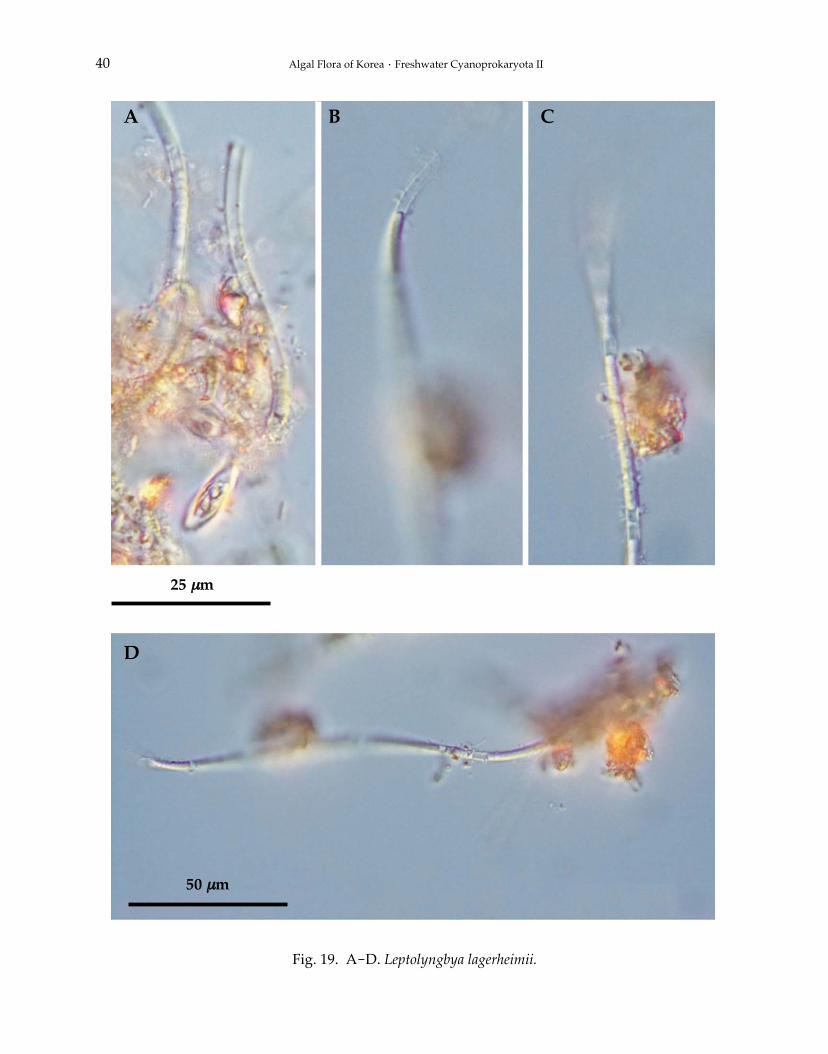
18. Leptolyngbya lagerheimii (Gomont) Anagnostidis et Komárek 1988:391 (Fig. 19A-D).
Komárek and Anagnostidis 2005: 187. f. 228. Hu and Wei 2006: 112. pl. II-21-17.
BASIONYM: Lyngbya lagerheimii Gomont 1892.SYNONYM: Spirocoleus lagerheimii Möbius 1889.Lyngbya lagerheimii (Möbius) Gomont 1890.Lyngbya lagerheimii f. minor (Voronichin et Chachina) Elenkin 1949.
Filaments planktonic, benthic or periphytic, solitary or rarely loosely entangled, sometimes irre-gularly spirally or screw-like coiled, rarely regularly spirally coiled, occasionally straight, 2.0-2.7μm wide. Sheaths thin, colorless, attached to trichomes, one trichome and opened at the apex,unbranched. Trichomes cylindrical, pale blue-green, unconstricted or slightly constricted at thecross-walls, not attenuated to the ends, with or without a single granule on either side. Spirals 6.0-8.0 μm wide, distance between spirals 20 μm. Cells cylindrical, more or less isodiametric, homoge-neous, without aerotopes, Heterocytes and akinetes absent. Apical cells rounded, without calyptra.Cells division by symmetrical, crosswise binary fission, cells reaching the original size before nextdivision. Reproduction by fragmentation without the assistance of necridic cells.
TYPE LOCALITY: Probably Brazil.DISTRIBUTION: Probably cosmopolitan.KOREA: JB (Chung 1976).SPECIMEN EXAMINED: Gucheonli in GN (34°48′50′′N, 128°38′08′′), 2011; Samdongdong in GN (35°
14′03′′N, 128°38′46′′), 02.viii.2011.ECOLOGY: Freshwater, planktonic, benthic or periphytic on hydrophyte and in mucilage of other
algae.REMARKS: This species belongs to Subgenus Leptolyngbya. Originally record was described from
Brazil. Distribution record from Korea is as Lyngbya lagerheimii Gomont.
Cyanophyceae: Oscillatoriales: Pseudanabaenaceae 39
40 Algal Flora of Korea∙Freshwater Cyanoprokaryota II
Fig. 19. A-D. Leptolyngbya lagerheimii.
25 μm
50 μm
A B C
D

19. Leptolyngbya subtilis (West) Anagnostidis 2001: 367 (Fig. 20A-C).
Komárek and Anagnostidis 2005: 186. f. 227. McGregor 2007: 30. f. 4F.
BASIONYM: Lyngbya subtilis West 1892.
Filaments benthic, initially attached by one end to the substratum, about 60 μm long, later free-floating, straight or slightly curved, 1.8 μm wide. Sheaths very thin, smooth, hyaline, one trichomeand opened at the apex, unbranched. Trichomes cylindrical, unconstricted at the cross-walls, notattenuated to the ends. Cells nearly as long as wide or slightly shorter or longer than wide, paleblue-green or blue-green, cell content delicately granular, without aerotopes. Heterocytes and aki-
Cyanophyceae: Oscillatoriales: Pseudanabaenaceae 41
Fig. 20. A-C. Leptolyngbya subtilis.
25 μm
A B C

netes absent. Apical cells rounded, without thickened cell wall or calyptra. Cells division by sym-metrical, crosswise binary fission, cells reaching the original size before next division. Reproductionby fragmentation without the assistance of necridic cells.
TYPE LOCALITY: Lake District, England.DISTRIBUTION: Probably cosmopolitan.KOREA: Previously unrecorded species.SPECIMEN EXAMINED: Kugdangkyo in GB (35°59′06′′N, 129°16′39′′), 01.viii.2011; Samdongdong in
GN (35°14′03′′N, 128°38′46′′), 02.viii.2011.ECOLOGY: Freshwater, initially benthic or metaphytic.REMARKS: This species belongs to Subgenus Leptolyngbya.
Genus Heteroleibleinia (Geitler) L. Hoffmann 1985: 76.
Filaments solitary or aggregated in groups, heteropolar, individually attached with one end tothe substratum. Sheaths firm or thin, usually colorless, enveloping the trichome, opened at theends, containing a single trichome. Trichomes straight, curved or slightly flexuous, constricted orunconstricted at the cross-walls, mainly to 4 μm wide, very rarely false branching with one lateralbranch. Cells more or less isodiametric, or slightly shorter or longer than wide, cylindrical or barrel-shaped, without aerotopes. Apical cell rounded or conically rounded, without calyptra or thickenedouter cell wall. Heterocytes and akinetes lacking. Cell division by the transverse fission, cells reachto the original size before the next division. Reproduction by disintegration of the trichome intomotile hormogonia at the apical parts.
HOLOTYPE: Heteroleibleinia rigidula (Hansgirg) L. Hoffmann 1985.
SPECIES: Of 31 species (infraspecific) reported up to date, 23 are currently accepted as taxonomi-cally valid taxa (Guiry and Guiry 2012).
DISTRIBUTION: Worldwide.KEY REFERENCE: Anagnostidis and Komárek (1988), Komárek and Anagnostidis (2005), McGregor
(2007), Whitton (2011).REMARKS: This genus belongs to subfamily Heteroleibleiniodeae. Most of species occur in aqua-
tic habitats, where they grow attached to different substrata, e.g., wood, stones, water plants, otheralgae, etc. But one species (H. thornensis (Hunting et Lund) Anagnostidis et Komárek) is epizoic onplanktonic crustaceans (Komárek and Anagnostidis 2005; Guiry and Guiry 2012). Not yet in culture.
Key to the species of genes Heteroleibleinia
1. Freshwater species, usually stagnant waters, filaments long and flexuous, sheaths thin and color-less, trichomes, pale blue-green, or olive green, 1.2-2.0 μm wide, cells longer than wide··········································································································································H. kossinskajae
- Freshwater species, usually stagnant waters, epiphytic, filaments to 150 μm long and flexuous,sheaths thin and colorless, trichomes indistinctly or slightly constricted at cross-walls, cells moreor less isodiametric of slightly longer than wide, 0.5-1.5 μm×0.5-2.0 μm··················H. rigidula
42 Algal Flora of Korea∙Freshwater Cyanoprokaryota II

20. Heteroleibleinia kossinskajae (Elenkin) Anagnostidis et Komárek 1988:434 (Figs. 21A, B, 22A, B).
Komárek and Anagnostidis 2005: 246. f. 321. Whitton 2011: 82. pl. 21K.
BASIONYM: Lyngbya kossinskajae Elenkin 1949.
Filaments flexuous, long, heteropolar, individually attached by one end to the substratum, arrangedloosely or densely in great number, (1.2-) 2.0-2.3 μm wide, Sheaths thin, narrow, colorless, envelop-ing the trichome, opened at the ends, containing a single trichome. Trichomes curved or flexuous,pale blue-green, constricted at the translucent cross-walls, not attenuated at the ends, (1.2) 1.8-2.0μm wide. Cells cylindrical, usually 2-3 times longer than wide, (2.4-) 3.4-5.7 μm long, with homo-geneous content, without aerotopes. Apical cells more or less attenuated, usually rounded-conica,without calyptra. Heterocytes and akinetes lacking. Cell division by the transverse fission, cellsreach to the original size before the next division. Reproduction by disintegration of the trichome
Cyanophyceae: Oscillatoriales: Pseudanabaenaceae 43
Fig. 21. A, B. Heteroleibleinia kossinskajae.
50 μm
A B

44 Algal Flora of Korea∙Freshwater Cyanoprokaryota II
Fig. 22. A, B. Heteroleibleinia kossinskajae.
50 μm
50 μm
A
B

into motile hormogonia at the apical parts.
TYPE LOCALITY: Near St. Petersburg, Russia.DISTRIBUTION: Worldwide.KOREA: Previously unrecorded species.SPECIMEN EXAMINED: Namgang in GN (35°23′01′′N, 128°25′21′′), 02.viii.2011.ECOLOGY: Freshwater, epiphytic on Cladophora in stagnant waters.REMARKS: The Lyngbya kossinskajae var. gracilis Claus 1961 differs by having slightly smaller dimen-
sions (cells=2.5-3.6 μm×1.2-1.3 μm), cell content with few granules and by its occurrence in thelittoral of a pond (Komárek and Anagnostidis 2005).
21. Heteroleibleinia rigidula (Hansgirg) L. Hoffmann 1985: 76 (Fig. 23A, B).
Komárek and Anagnostidis 2005: 248. f. 323. Whitton 2011: 83. pl. 21M.
BASIONYM: Lyngbya rigidula Hansgirg 1892.SYNONYM: Hygrocrocis rigidula Kützing 1843, nom. inval.Leptothrix rigidula (Kützing) Kützing 1845, nom. inval.Hypheothrix rigidula (Kützing) Grunow 1870, nom. inval.Leptotrichia rigidula (Kützing) Trevisan 1879, nom. inval.Lyngbya tenerrima var. rigidula 1884.
Filaments solitary or aggregated in irregular groups, individually attached by on end to the sub-stratum, heteropolar, 100-150 (-200) μm long, 1.5-2.0 (-2.5) μm wide, more or less straight or slightlycurved, not twisted. Sheaths thin, seemingly firm, distinct, colorless, enveloping the trichome,opened at the ends, containing a single trichome. Trichomes pale blue-green or olive-green, 0.8-1.5(-2.8) μm wide, indistinctly or very slightly constricted at the cross-walls, not attenuated at the ends.Cells 0.5-2 times as long as wide, 1.0-3.0 μm long, cylindrical, without aerotopes. Apical cells obtuserounded, without calyptra. Heterocytes and akinetes lacking. Cell division by the transverse fis-sion, cells reach to the original size before the next division. Reproduction by disintegration of thetrichome into motile hormogonia at the apical parts.
TYPE LOCALITY: Probably Germany (type of Hygrocrocis rigidula Kützing).DISTRIBUTION: Probably worldwide.KOREA: Previously unrecorded species.SPECIMEN EXAMINED: GB, 08.ix.2010.ECOLOGY: Freshwater, epiphytic on filamentous algae, also on submersed roots of water and
littoral plants, in swamps, old ponds, slow-flowing waters, etc.REMARKS: This is the type species (holotype) of the genus Heteroleibleinia. The twisted filaments
concern other species, probably the genus Leibleinia (Komárek and Anagnostidis 2005).
Cyanophyceae: Oscillatoriales: Pseudanabaenaceae 45

46 Algal Flora of Korea∙Freshwater Cyanoprokaryota II
Fig. 23. A, B. Heteroleibleinia rigidula.
50 μm
50 μm
A
B

Family Phormidiaceae Anagnostidis et Komárek 1998: 400.
Filaments benthic or metaphytic, unbranched, rarely solitary, usually forming gelatinous mat.Sheaths absent or present, fine or firm, thin or thickened, colorless, usually open at the ends, gener-ally containing one trichome. Trichomes cylindrical, straight or variously coiled, constricted orunconstricted at the cross-walls, motile, mainly 4.0-12.0 μm wide. Cells mainly isodiametric ormore or less slightly longer or shorter than wide. Apical cell rounded, attenuated or pointed, with-out or with calyptra. Heterocytes and akinetes absent. Cell division by the transverse fission, cellsreaching their original size before the next division. Reproduction by disintegration of the trichomeinto motile hormogonia with help of necridic cells.
GENERA AND SPECIES: 26 genera and 397 species are currently accepted as valid taxonomically(Guiry and Guiry 2012). Nine genera contain no or single species. Major genera are Phormidium(169 species), Microcoleus (37 species), Symploca (32 species) and Pseudophormidium (22 species).
DISTRIBUTION: Worldwide, benthic or metaphytic in freshwater, brackish and salty waters, includingextreme environments.
KEY REFERENCE: Gomont (1892), Anagnostidis and Komárek (1988), Chung (1993), Canter-lundand lund (1995), Komárek and Anagnostidis (2005), McGregor (2007), Hindák (2008), Whitton(2011).
REMARKS: This family is generally divided into three subfamilies. Subfamily Phormidioideae:Trichomes shape is more or less cylindrical and isopolar. There is one trichome in sheath. SubfamilyMicrocoleoideae: Trichomes shape is more or less cylindrical and isopolar. There are two up manytrichomes in sheath. Subfamily Ammatoideoideae: Trichomes shape is elongated into cellular hairsat the apical end.
Key to the genera of family Phormidiaceae
1. Filament and trichomes more or less cylindrical, isopolar·························································2- Filament and trichomes heteropolar, elongated into cellular hairs at the apical end, rounded at
the base···············································································································Phormidiochaete2. Cells with aerotopes within cells, usually planktonic species, trichomes without distinct sheaths
or occur facultatively················································································································3- Cells without gas vesicles and aerotopes, usually aggregated into colonies, mats, fascicles, crusts,
filamentous often with facultative sheaths················································································53. Trichomes usually in microscopic fascicles or clusters, only rarely disintegrating in solitary tri-
chomes·················································································································Trichodesmium- Trichomes usually solitary········································································································44. Trichomes more or less regularly screw-like coiled····················································Arthrospira- Trichomes straight or arcuated, rarely irregularly wavy, trichomes cylindrical or shortly attenuated
to the ends and sometimes with calyptra or thickened cell wall································Planktothrix5. Filaments solitary or in small groups, living endophytically; trichomes short, slightly narrowed
toward both ends·································································································Proterendothrix- Filaments free-living, solitary, or forming mats, fascicles, clusters or crusts coiled····················66. Filaments solitary in plankton or in fine mats, usually pale reddish···························Tychonema- Filaments usually forming colonies in mats, fascicles or crusts coiled·······································7
Cyanophyceae: Oscillatoriales: Phormidiaceae 47

7. Filaments with numerous false branching; trichomes often disintegrated into short segments····························································································································Pseudophormidium
- Filaments without or with very rare false branching, sheaths facultative, thin or thick, simple,not layered···············································································································Phormidium
Genus Planktothrix Anagnostidis et Komárek 1988
Filaments planktonic, usually without sheath or occur facultatively in several species under stresscondition, fine, colorless and diffluent, solitary, rarely in small irregular and disintegrating fascicles,more or less straight or slightly waved, isopolar, free living, immotile. Trichome with cylindricalcells, slightly constricted at the cross-walls, slightly attenuated or not attenuated at the ends, some-times with terminal calyptra, to 4 mm long, 3.5-12.0 (-15.0) μm wide, false branching absent. Cellscylindrical, rarely more or less slightly barrel-shaped, usually slightly shorter than wide to more orless isodiametric, rarely slightly longer than wide, thylakoids more or less radially arranged, aero-topes distributed the whole cell volume. Apical cells widely rounded when fully developed orslightly narrowed, sometimes with thickened outer cell wall or with calyptra. False branching,heterocytes and akinetes absent. Cell division crosswise, the cells grow to the original size beforethe next division. Reproduction by trichome disintegration into hormogonia, by help of necridiccells.
HOLOTYPE SPECIES: Planktothrix agardhii (Gomont) Anagnostidis et Komárek 1988.
SPECIES: Among 25 species (infraspecific) are reported up to date, 16 species are currently acceptedas taxonomically valid taxa (Guiry and Guiry 2012).
DISTRIBUTION: Worldwide, plankton in freshwater.KEY REFERENCE: Anagnostidis and Komárek (1988), Komárek and Komárková (2004), Komárek
and Anagnostidis (2005), McGregor (2007), Whitton (2011).REMARKS: Almost all species are planktonic, a few of them causing water blooms, mainly in meso-
trophic to eutrophic reservoirs. One species (Planktothrix cryptovaginata (Schkorbatov) Anagnostidiset Komárek) grows in metaphyton of unpolluted pools and ponds among submersed aquatic plants.Two species have cosmopolitan distribution, other ones are geographically limited, mainly occurringunder special ecological situations; two species are known from tropics only (Guiry and Guiry 2012).
Key to the species of genus Planktothrix
1. Filament usually straight or slightly curved, free-floating and planktonic, trichomes not attenuated,never capitate···························································································································2
- Filament usually straight or slightly curved, free-floating and planktonic, trichomes taperingtowards ends, apical cell sometimes capitate or with calyptra, blue-green, usually less than 6 μmwide····························································································································P. agardhii
2. Trichomes not constricted at cross-walls, cells usually more or less isodiametric, 6.0-10.0 μm wide,blue-green, apical cells cylindrical and widely rounded···············································P. isothrix
- Trichomes finely constricted at cross-walls, without calyptra, cells shorter than wide, 6.5-7.5 μmwide························································································································P. compresssa
48 Algal Flora of Korea∙Freshwater Cyanoprokaryota II

22. Planktothrix agardhii (Gomont) Anagnostidis et Komárek 1988: 416(Figs. 24A-C, 25A).
Suda et al. 2002: 1592. f. 6. Komárek and Komárková 2004: 7. Komárek and Anagnostidis 2005:359. f. 500. McGregor 2007: 63. Whitton 2011: 105. pl. 20A.
BASIONYM: Oscillatoria agardhii Gomont 1892.SYNONYM: Oscillatoria agardhii f. gomontii Elenkin 1949.Oscillatoria agardhii f. aequicrassa Elenkin 1949.Oscillatoria agardhii f. moebiusii Elenkin 1949.Oscillatoria agardhii f. lemmermannii Elenkin 1949.Oscillatoria agardhii f. wislouchii Elenkin 1949.
Cyanophyceae: Oscillatoriales: Phormidiaceae 49
Fig. 24. A-C. Planktothrix agardhii.
25 μm
A B C

Filaments usually planktonic, microscopic, mucilaginous envelopes or sheaths lacking, or veryrarely and facultatively, especially in young stages, with fine sheaths. Trichomes mostly solitary,with cylindrical cells, up to 300 μm long, straight or somewhat curved, sometimes joined to blue-green or olive green, loose fascicles, occasionally forming benthic membranous coat (thallus), notconstricted or very slightly constricted at the granulated cross-walls, more or less gradually attenuat-ed at the ends, (3.5-) 4.0-6.0 (-9.8) μm wide, 2.5-4.0 μm long, immotile. Cells mostly shorter thanwide or frequently isodiametric, up to 2 times longer than wide, blue-green, sometimes with gran-ules near cross-walls, with numerous aerotopes. Apical cells rounded or convex, sometimes bluntly-conical more or less pointed, sometimes with convex calyptra, rarely capitate. Cell division cross-wise, the cells grow to the original size before the next division. Reproduction by trichome disin-tegration into hormogonia, by help of necridic cells.
50 Algal Flora of Korea∙Freshwater Cyanoprokaryota II
Fig. 25. A. Planktothrix agardhii; B, C. Plalnktothirx compressa.
25 μm
A B C

SYNTYPE: Sweden (Lund); France (Croisic) (Gomont 1893).DISTRIBUTION: Widely distributed in temperate zone.KOREA: Lake Chungju in CB (Kim et al. 1988); Nakdong River (Cho et al. 1993); Geum River
(Chung and Lee 1979; Kim et al. 2000); Lake Daecheong in CB (Park and Lee 2005).SPECIMEN EXAMINED: Sajipo in GB (35°33′34′′N, 128°25′56′′E), 26.xi.2010.ECOLOGY: Freshwater and blackish waters, planktonic in lakes and ponds, often forming water
bloom.REMARKS: Harmful species (Guiry and Guiry 2012). Non-planktonic biotopes (cells without aero-
topes) probably belong to other species (Komárek and Anagnostidis 2005). Distribution records fromKorea are as Oscillatoria agardhii Gomont except for Park and Lee (2005).
23. Planktothrix compressa (Utermöhl) Anagnostidis et Komárek 1988: 416(Fig. 25B, C).
Komárek and Komárková 2004: 7. Komárek and Anagnostidis 2005: 358. f. 496.
BASIONYM: Lyngbya compressa Utermöhl 1925.SYNONYM: Oscillatoria compressa (Utermöhl) Geitler 1925.
Filaments planktonic, solitary, usually mucilaginous envelopes or sheaths lacking, very rarelywith fine and slightly visible sheaths, which develop well only in culture. Trichomes cylindrical,blue-green (5.0-) 6.5-7.5 (-9.5) μm wide, slightly constricted at cross-walls, not attenuated towardsthe ends, false branching absent. Cells usually shorter than wide, with numerous aerotopes, rarelyslightly longer than wide. Apical cells widely-rounded, sometimes with thickened outer cell wall,not capitate. Heterocytes and akinetes absent. Cell division crosswise, the cells grow to the originalsize before the next division. Reproduction by trichome disintegration into hormogonia, by help ofnecridic cells.
TYPE LOCALITY: Holstein, Germany.DISTRIBUTION: Widely distributed in temperate zone.KOREA: Previously unrecorded species.SPECIMEN EXAMINED: Pungrakji in GB (35°56′40′′N, 128°51′14′′E), 03.ix.2010.ECOLOGY: Freshwater, planktonic in lakes.REMARKS: This species was previously only from Germany (Komárek and Anagnostidis 2005).
Morphologically similar is Planktothrix mougeotii (Kützing ex Forti) Suda et al. (Komárek and Komár-ková 2004).
24. Planktothrix isothrix (Skuja) Komárek et Komárková 2004: 14(Fig. 26A-C).
Komárek and Anagnostidis 2005: 356. f. 459. McGregor 2007: 63. f. 11I. pl. 9J. Whitton 2011: 106.
Cyanophyceae: Oscillatoriales: Phormidiaceae 51

BASIONYM: Oscillatoria agardhii var. isothrix Skuja 1948.SYNONYM: Oscillatoria agardhii var. isothrix Skuja 1948.Planktothrix mougeotii (Bory ex Gomont) Anagnostidis et Komárek 1988 sine typo: non. Oscillatoria mougeotii Bory ex Gomont 1892.
Filaments at first benthic, attached, later free-floating, usually without sheath. Trichomes darkblue-green in masses, solitary often forming water blooms, rarely in small clusters without definitearrangement, usually straight or somewhat slightly curved, usually very long up to 3.5 mm, blue-green or grey-green, (5.0-) 6.0-9.5 (-10.0) μm wide, slowly motile usually inconspicuous cross-wall,but with a single row of prominent granules on each side, cylindrical and neither attenuated norcurved towards apex, false branching absent. Cells slightly shorter than wide or nearly isodiametric,(1.5-) 2.0-5.5 (-9.5) μm long, with plenty of irregular aerotopes causing a characteristic brown tobrown-blackish color of the trichomes. Apical cells cylindrical and widely-rounded or flat-rounded,
52 Algal Flora of Korea∙Freshwater Cyanoprokaryota II
Fig. 26. A-C. Planktothrix isothrix.
25 μm
A B C

rarely slightly conical, not capitate, without calyptra, sometimes thickened outer cell wall. Hetero-cytes and akinetes absent. Cell division crosswise, the cells grow to the original size before the nextdivision. Reproduction by trichome disintegration into hormogonia, by help of necridic cells.
TYPE LOCALITY: Lakes in Uppland, Sweden (Skuja 1948).DISTRIBUTION: Worldwide.KOREA: Previously unrecorded species.SPECIMEN EXAMINED: Yusuji in Gyeongin ARA waterway (37°33′56′′N, 126°37′03′′E), 23.vi.2010;
05.viii.2010.ECOLOGY: Freshwater, at first benthic, epipelic on mud, later planktonic forming water blooms in
eutrophic to hypertrophic stagnant waters and lakes.REMARKS: Data from thermal springs are considered debatable (Komárek and Anagnostidis 2005).
Genus Phormidium Kützing ex Gomont 1892: 156.
Filaments benthic or metaphytic, microscopic and later macroscopic, rarely solitary, usually forminggelatinous mat-like leathery strata, not pseudobranched, usually entangled. Sheaths present orabsent, tube-like, fine or firm, usually colorless, not lamellated, adherent to the trichome, opened atthe ends, containing always a single trichome. Trichomes isopolar and long, uniseriate, more orless straight, irregularly coiled or slightly waved, slightly constricted or unconstricted at the cross-walls, not attenuated and bent or screw-like twisted towards the ends, motile within and out ofsheaths, waving, creeping, oscillations, 4.0-12.0 (-15.0) μm wide. Cells cylindrical to slightly barrel-shaped cells, more or less isodiametric or slightly shorter or longer than wide, usually blue-green,rarely brownish, pinkish or violet, without aerotopes, or exceptionally with aerotopes under special,suboptimal conditions, sometimes with granular content or with prominent granules spread through-out the cell volume, or agglomerated at the cross-walls. Apical cell rounded, attenuated or pointed,without or with calyptra. Heterocytes and akinetes absent. Cell division by the transverse fission,cells reach to the original size before the next division, all cells capable of division with the excep-tion of apical ones. Reproduction by disintegration of the trichome into motile hormogonia withhelp of necridic cells or by fragmentation of whole trichomes.
LECTOTYPE: Phormidium lucidum Kützing ex Gomont 1892.
SPECIES: Of the 647 species (infraspecific) reported up to date, 169 are currently accepted as taxono-mically valid taxa (Guiry and Guiry 2012).
DISTRIBUTION: Worldwide, benthic or metaphytic in freshwater, brackish and marine waters.KEY REFERENCE: Anagnostidis and Komárek (1988), Komárek and Anagnostidis (2005), McGregor
(2007), Whitton (2011).REMARKS: Rarely solitary filaments, usually in mats on different substrata e.g., soil, wet rocks,
mud, aquatic plants, stones and woods in both stagnant water and streams, some species occur inthe marine littoral. Several species are known from extreme localities e.g., thermal springs, desertsoils, etc. (Guiry and Guiry 2012). Sheaths develop facultatively in different frequencies, only insuboptimal conditions (subgenus Gomontinema), in dependence on changing environmental factors(subgenus Phormidium), or regularly in all conditions (with exceptionally living free trichomes -subgenus Hansgirgia) (Komárek and Anagnostidis 2005; Guiry and Guiry 2012).
Cyanophyceae: Oscillatoriales: Phormidiaceae 53

Phormidium belongs taxonomically to the most difficult cyanoprokaryotic genera. It comprisesnumerous morphotypes with many transient forms. For easier identification, the genus is dividedinto 8 taxonomic groups, which differ by the morphology of their apical trichome ends (Komárekand Anagnostidis 2005; McGregor 2007).
Key to the species of genus Phormidium
Synopsis of species groupsGroup 1: Trichomes abruptly or gradually narrowed towards the ends; apical cells pointed to
sharply pointed, sometimes curved, without a calyptra or a thickened outer cell wall.Group 2: Trichomes gradually narrowed towards the ends; apical cells distinctly narrowed and
rounded, without a calyptra.Group 3: Trichomes abruptly narrowed towards the ends, often bent; apical cells slightly conical-
narrowed or almost cylindrical and rounded, without a calyptra, or with very slightly thickenedouter cell walls.
Group 4: Trichomes usually cylindrical along their whole length, only with distinctly conical apicalcells, without a calyptra or thickened cell walls.
Group 5: Trichomes cylindrical along their whole length; apical cells broadly rounded or obtuse,without a calyptra, but sometimes with thickened cell walls at the apex.
Group 6: Trichomes abruptly attenuated towards the ends; apical cells more or less sphericallycapitate.
Group 7: Trichomes abruptly or gradually narrowed towards the ends; apical cells capitate, some-times calyptrate.
Group 8: Trichomes cylindrical with cells always distinctly shoter than wide, abruptly attenuatedtowards the ends; apical cells calyptrate or with thickened outer cell walls.
Group 2: Trichome gradually narrowed, apical cells rounded, without calyptra.1. Trichomes blue-green, usually distinctly constricted at cross-walls, 5.0-8.0 (-9.0) μm wide in the
middle part·····················································································································P. okenii- Trichomes blue-green, finely or facultatively distinctly constricted at cross-walls, 4.0-6.8 μm wide
in the middle part··································································································P. terebriforme
Group 3: Trichome abruptly narrowed and bent, apical cells rounded, without calyptra.1. Trichomes blue-green, 3.8-6.5 μm wide····················································································2- Trichomes blue-green, 6.4-13.0 μm wide, slightly constricted at cross-walls, sheaths very rare···········································································································P. chaybeum (See Vol 5. No.1)
2. Trichomes in flat mats, cells usually shorter than wide, usually granulated at cross-walls, benthicmats in pools and ponds··································································································P. breve
- Trichomses benthic mats, all cells more or less isodiametric, 3.8-6.5 μm wide, sheaths very rare,facultative·······················································································P. formosum (See Vol 5. No.1)
Group 4: Trichome cylindrical, apical cells distinctly conical.1. In flowing water on limestone substratum, thallus often with calcareous precipitate, trichomes
3.0-6.5 μm wide····································································································P. incrustatum- Thallus without calcareous precipitate······················································································22. Trichomes not constricted at the cross-walls, 6.5-11.0 μm wide····························P. crouaniorum
54 Algal Flora of Korea∙Freshwater Cyanoprokaryota II

- Trichomes not constricted at the cross-walls, 3.0-5.0 μm, cells more less isodiametric to 2 timeslonger than wide, living submerged, periphytic and metaphytic····························P. inundatum
Group 5: Trichome cylindrical, apical cells rounded, without calyptra.1. Sheaths thick, colorless, trichomes 3.0-6.0 μm wide, blue-green, cells more or less isodiametric,
benthic, periphytic, sometimes epiphytic·················································P. aerugineo-caeruleum- Sheaths always thin················································································································22. Trichomes yellow-green, olive green or brownish···································································3- Trichomes pale or bright blue-green, or greyish blue-green·····················································43. Trichomes 3.0-4.0 (-6.0) μm wide, cells shorter than wide up to isodiametric, without prominent
granules·················································································································P. chlorinum- Trichomes 6.0-10.0 μm wide, cells shorter than wide··················································P. nigrum4. Trichomes distinctly constricted at the cross-walls··································································5- Trichomes unconstricted at the cross-walls·············································································65. Trichomes 3.5-6.0 μm wide, cells more less isodiametric to longer than wide, without granula-
tion, mainly in warmer regions·······················································P. hamelii (See Vol. 5. No. 1)- Trichomes 5.0-13.0 μm wide, common periphytic···························P. puteale (See Vol. 5. No. 1)6. Trichomes less than 4.0 μm wide·····························································································7- Trichomes more than 4.0 μm wide··························································································87. With distinct granulation at the cross-walls, benthic, periphytic···························P. granulatum- Without granulation at the cross-walls, benthic in warmer regions·······················P. articulatum8. Cells more less isodiametric····································································································9- Cells always shorter than wide, trichomes 9.5-12.0 μm wide, periphytic·····································································································································P. stagninum (See Vol. 5. No. 1)
9. Granulation scattered in cells and at the cross-walls, without prominent granules················10- Distinct granulation usually at the cross-walls·····················································P. tergestinum
10. Trichomes 2.5-5.0 μm wide, metaphytic and periphytic··············································P. corium- Trichomes 4.0-7.0 μm wide, benthic, epipelic·······························································P. taylori
Group 7: Trichome abruptly or gradually narrowed, apical cells calyptrate1. Cells blue green························································································································2- Cells reddish or violet, more or less isodiametric or longer than wide, subaerophytic on wet
rocks·························································································································P. violaceum2. Trihomes 3.0-9.0 μm wide, in mesotrophic swamps to eutrophic waters··································3- In cold stream, benthic mats, on calcareous substrate, cells (2.4-) 3.0-7.0 μm×4.0-9.0 μm, sheaths
facultative···················································································································P. favosum3. In mesotrophic swamps, littoral of stagnant waters, ponds with water vegetation, metaphytic
and periphytic, cells 2.5-4.2 μm×2.5-5.0 μm, sheaths very rarely·····························P. amoenum- In mesotrophic to eutrophic streams, cells 2.0-5.0 μm×3.5-7.0 μm, sheaths facultative, but com-
mon·························································································································P. autumnale
Group 8: Trichome cylindrical with cells always shoter than wide, apical cells calyptrate.1. Trichomes 4.0-11.0 μm wide, unconstricted at the cross-walls, cells alomost isodiametric, 4.0-11.0
μm×6.0-11.0 μm, greyish-blue or reddish, sheath absent··········································P. irriguum- Trichomes 4.0-9.5 μm wide, unconstricted at the cross-walls, calyptra more or less rounded,
periphytic on rocks and stones in stream·································································P. uncinatum
Cyanophyceae: Oscillatoriales: Phormidiaceae 55
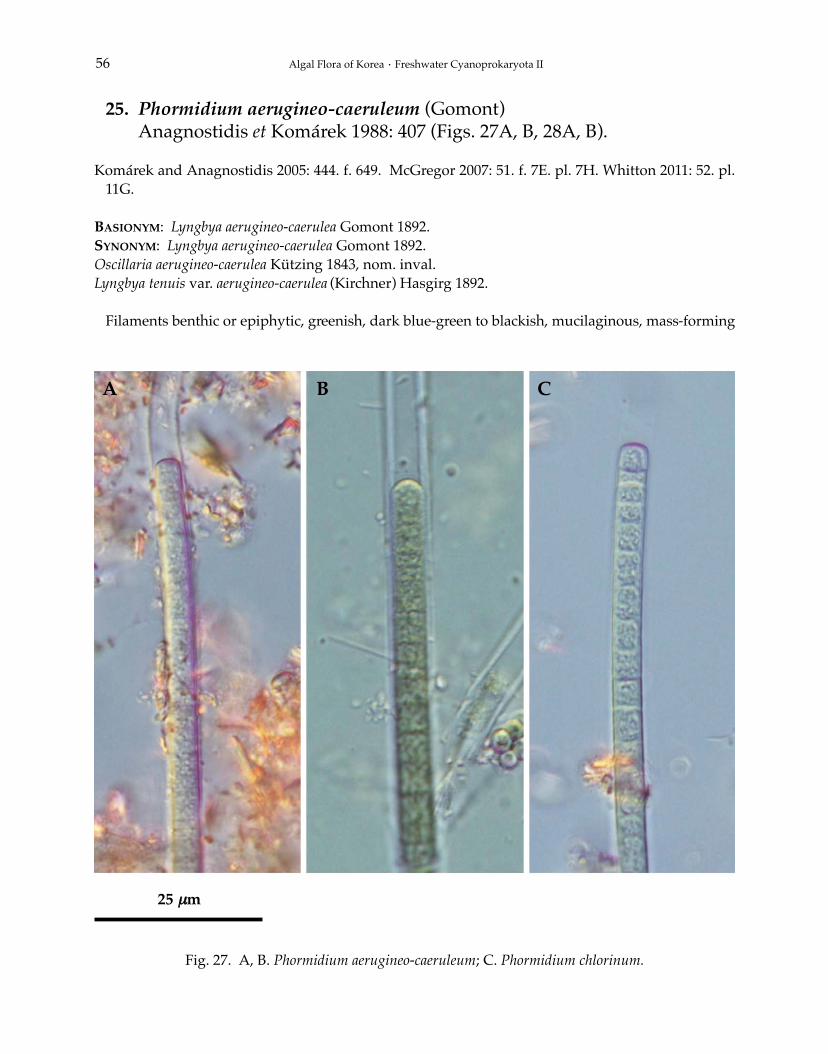
25. Phormidium aerugineo-caeruleum (Gomont)Anagnostidis et Komárek 1988: 407 (Figs. 27A, B, 28A, B).
Komárek and Anagnostidis 2005: 444. f. 649. McGregor 2007: 51. f. 7E. pl. 7H. Whitton 2011: 52. pl.11G.
BASIONYM: Lyngbya aerugineo-caerulea Gomont 1892.SYNONYM: Lyngbya aerugineo-caerulea Gomont 1892.Oscillaria aerugineo-caerulea Kützing 1843, nom. inval.Lyngbya tenuis var. aerugineo-caerulea (Kirchner) Hasgirg 1892.
Filaments benthic or epiphytic, greenish, dark blue-green to blackish, mucilaginous, mass-forming
56 Algal Flora of Korea∙Freshwater Cyanoprokaryota II
Fig. 27. A, B. Phormidium aerugineo-caeruleum; C. Phormidium chlorinum.
25 μm
A B C

with loosely tangled filaments or solitary scattered filaments, variously curved, 5.0-7.5 μm wide.Sheaths absent or present, thin, firm, or up to 1 μm thick, not lamellated, colorless, adherent to thetrichome, opened at the ends, containing always a single trichome. Trichomes isopolar and long,uniseriate, more or less straight, pale to bright blue-green, 3.0-7.0 μm wide, not constricted at thesometimes granulated, rarely at the apical parts slightly constricted, not attenuated at the ends, some-times capitate. Cells cylindrical, up to 0.5 shorter than wide to isodiametric, or a little elongated,2.0-7.8 μm long, contents homogeneous, sometimes with a few large granules. Apical cell cylindricalwith obtusely-conical or rounded tip, rarely weakly capitate, with slightly thickened outer cell wall.Cell division by the transverse fission, cells reach to the original size before the next division. Repro-duction by disintegration of the trichome into motile hormogonia with help of necridic cells .
TYPE LOCALITY: Probably Germany (type of Oscillatoria aerugino-caerulea Kützing, nom. inval.).DISTRIBUTION: Worldwide.KOREA: JB (Chung 1976).SPECIMEN EXAMINED: Suyeongang in Nakdong River (35°09′56′′N, 129°07′34′′E), 29.ix.2010; Yang-
baekli in GB (36°07′20′′N, 129°20′51′′E) 19.xi.2011; Daesankyo in GB (35°38′54′′N, 127°56′14′′E)02.xii.2011.
ECOLOGY: Freshwater, benthic and epiphytic in stagnant and flowing waters on mud.REMARKS: Distribution records from Korea are as Lyngbya aerugineo-coerulea Gomont 1892. Reports
Cyanophyceae: Oscillatoriales: Phormidiaceae 57
Fig. 28. A, B. Phormidium aerugineo-caeruleum.
A
B
25 μm
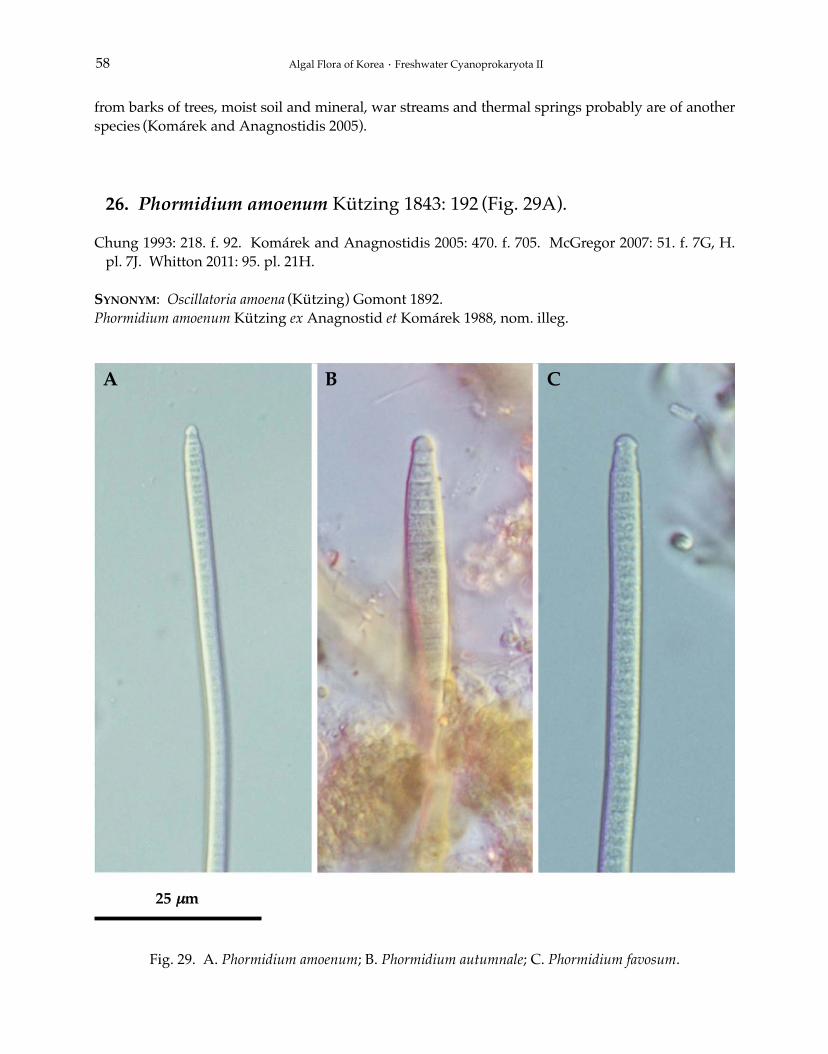
from barks of trees, moist soil and mineral, war streams and thermal springs probably are of anotherspecies (Komárek and Anagnostidis 2005).
26. Phormidium amoenum Kützing 1843: 192 (Fig. 29A).
Chung 1993: 218. f. 92. Komárek and Anagnostidis 2005: 470. f. 705. McGregor 2007: 51. f. 7G, H.pl. 7J. Whitton 2011: 95. pl. 21H.
SYNONYM: Oscillatoria amoena (Kützing) Gomont 1892.Phormidium amoenum Kützing ex Anagnostid et Komárek 1988, nom. illeg.
58 Algal Flora of Korea∙Freshwater Cyanoprokaryota II
Fig. 29. A. Phormidium amoenum; B. Phormidium autumnale; C. Phormidium favosum.
25 μm
A B C

Filaments benthic, microscopic, dark blue-green to dark bluish green, forming gelatinous tufts.Sheaths usually absent or very rarely fine sheath. Trichomes isopolar, straight or slightly flexuous,long, dark blue-green, 2.5-5.0 μm wide, slightly or not constricted at cross-walls, gradually attenu-ated and often hooked at the ends, motile within sheaths. Cells cylindrical, isodiametric or slightlylonger than wide, 2.5-6.0 μm long, granulated or ungranulated at the cross-walls. Apical cell longerthan others, capitate, broadly conical with depressed calyptra. Heterocytes and akinetes absent.Cell division by the transverse fission, cells reach to the original size before the next division, allcells capable of division with the exception of apical ones. Reproduction by disintegration of thetrichome into motile hormogonia with help of necridic cells.
TYPE LOCALITY: Padova, Italy (Drouet 1968). Lectotype: Meneghini; “in infusionibus algarumPatavii”. L (in herbarium Kützing) (Drouet 1968).
DISTRIBUTION: Temperate and tropical zones.KOREA: JB (Chung 1976, 1999); JN (Wui and Kim 1987).SPECIMEN EXAMINED: Sowolli in GB (35°56′29′′N, 128°50′08′′E), 31.viii.2010.ECOLOGY: Freshwater, in stagnant, unpolluted waters, muddy swamps, peaty pools, on water
plants.REMARKS: Known to produce the taste and odor compounds 2-methylisoborneol and geosmin
(McGregor 2007).The records from travertine, moist soil and flowing hot springs are revised (Komárekand Anagnostidis 2005). Reference records from Chung (1993) and Whitton (2011) are as Oscillatoriaamoena (Kützing) Gomont. And record of Komárek and Anagnostidis (2005) is as Phormidium amoe-num Kützing ex Anagnostid et Komárek. Distribution records from Korea are as Oscillatoria amoena(Kützing) Gomont.
27. Phormidium articulatum (Gardner) Anagnostidis et Komárek 1988:404 (Fig. 30A, B).
Komárek and Anagnostidis 2005: 439. f. 639.
BASIONYM: Oscillatoria articulata Gardner 1927.SYNONYM: Oscillatoria articulata var. circinata Gardner 1927 incl.Oscillatoria grunowiana var. articulata (Gardner) Drouet 1937.Oscillatoria articulata var. africana Welsh 1965 incl.
Filaments benthic, blue-green, usually solitary trichomes among other algae. Sheaths thin, usuallyindistinct, adherent to the trichome, opened at the ends, containing always a single trichome. Tri-chomes isopolar, long and straight or short and circinate, blue-green, 2.8-4.0 (-4.6) μm wide, uncon-stricted or slightly constricted at the cross-walls, not attenuated at the ends. Cells cylindrical, isodia-metric or up to two to three times shorter than wide, rarely longer than wide, without aerotopes,1.4-4.0 μm long. Apical cells rounded, without calyptra or thickened outer cell wall. Heterocytesand akinetes absent. Cell division by the transverse fission, cells reach to the original size beforethe next division. Reproduction by disintegration of the trichome into motile hormogonia with helpof necridic cells.
Cyanophyceae: Oscillatoriales: Phormidiaceae 59

TYPE LOCALITY: Porto Rico.DISTRIBUTION: Probably worldwide, mainly in tropical legions.KOREA: Lake Chungju in CB (Kim et al. 1988).SPECIMEN EXAMINED: Mokpo in GN (35°33′28′′N, 128°24′13′′E), 17.iv.2010.ECOLOGY: Freshwater, benthic later tychoplanktonic, in stagnant waters, also on moist rocks.REMARKS: Distribution record from Korea is as Oscillatoria articulata Gardner.
60 Algal Flora of Korea∙Freshwater Cyanoprokaryota II
Fig. 30. A, B. Phormidium articulatum; C. Phormidium chlorinum.
25 μm
A B C

28. Phormidium autumnale Gomont 1892: 187 (Fig. 29B).
Komárek and Anagnostidis 2005: 473. f. 707. Whitton 2011: 101. pl. 22B, C.
BASIONYM: Oscillatoria autumnalis C. Agardh 1812, nom. inval.SYNONYM: Oscillatoriella autumnalis (C. Agardh) Gaillon 1833, nom. inval.Phormidium autumnale (C. Agardh) Trevisan 1844, nom. inval.Microcoleus autumnalis (C. Agardh) Rabenhorst 1847, nom. inval.Phormidium autumnale Gomont 1892, nom. inval.Phormidium vulgare Kützing 1843, nom. inval.Phormidium membranaceum Kützing 1843, nom. inval.Microcoleus membranaceus Kützing ex Rabenhorst 1847, nom. inval.Oscillaria vulgaris (Kützing) P.L. Crouan et H.M. Crouan 1867, nom. inval.Lyngbya membranacea (Kützing) Thuret 1875, nom. inval.Lyngbya vulgaris (Kützing) Kirchner 1878, nom. inval.Oscillatoria limosa var. uncinata C. Agardh ex Mougéot et Rouméguère 1887.Phormidium membranaceum Kützing ex Gomont 1892.Lyngbya membranacea Thuret ex Hansgirg 1892.Phormidium subsalsum Gomont 1899.
Benthic. Thallus thick, leathery, velvety on the surface, externally blackish, black-brown in thelower layers, dark blue-green to brownish-green or olive green, sometimes yellowish. Filamentsmostly straight, or slightly curved at the ends, not pseudobranched, usually entangled. Sheathsthin, hyaline, distinct or diffluent, sometimes lacking. Trichomes bright blue-green or dirty green,(3.5-) 4.0-7.0 μm wide, not or sometimes slightly constricted at the granulated cross-walls, mostlyattenuated at the ends, straight or weakly hooked, motile with anticlockwise rotation and strongoscillation. Cells 0.5 shorter than wide or isodiametric, rarely longer than wide, 2.0-4.0 (-5.0) μmlong, contents homogeneous, sometimes granular at cross-walls. Apical cell somewhat elongated,capitate, with rounded or truncated calyptras. Heterocytes and akinetes absent. Cell division bythe transverse fission, cells reach to the original size before the next division, Reproduction bydisintegration of the trichome into motile hormogonia with help of necridic cells.
TYPE LOCALITY: “In nuda terra autumno”. Sweden (type of Oscillatoria autumnalis C. Agardh).DISTRIBUTION: Worldwide.KOREA: JB (Chung 1976); Galgok stream and Incheon river in JN (Lee et al. 2008).SPECIMEN EXAMINED: Hyogeongli in JB (35°23′20′′N, 127°27′41′′E), 10.vi.2011.ECOLOGY: Freshwater, periphytic on submersed substrata (stones, stony dams), less frequently
benthic on muddy banks of tanks, and mainly in streams, rivers, waterfalls.REMARKS: The records from moist soil, walls, marine coastal rocks, thermal springs concern pro-
bably other types from this taxonomic group (Komárek and Anagnostidis 2005). Reference recordfrom Whitton (2011) is as Phormidium autumnale (C. Agardh) Gomont. In Whitton’s record (2011),Phormidium autumnale (C. Agardh) Gomont and Phormidium uncinatum (C. Agardh) Gomont exGomont combined same species, Because of P. uncinatum and P. autumnale is similar on morphologi-cal properties except on width range of trichome.
Cyanophyceae: Oscillatoriales: Phormidiaceae 61

29. Phormidium breve (Kützing ex Gomont) Anagnostidis et Komárek 1988:405 (Fig. 31A-C).
Komárek and Anagnostidis 2005: 421. f. 599. McGregor 2007: 55. f. 9D. pl. 7D.
BASIONYM: Oscillatoria brevis Kützing ex Gomont 1892.SYNONYM: Oscillaria brevis Kützing 1833, nom. inval.Oscillaria neapolitana Kützing 1843, nom. inval.Lyngbya brevis (Kützing) Hansgirg 1887, nom. inval.Oscillatoria brevis var. neapolitana Gomont 1892.Oscillatoria neapolitana Kützing ex Gomont 1892.Oscillatoria brevis Kützing ex Gomont 1892.Lyngbya brevis Hansgirg 1892.
62 Algal Flora of Korea∙Freshwater Cyanoprokaryota II
Fig. 31. A-C. Phormidium breve.
25 μm
A B C

Lyngbya tenuis var. neapolitana Hansgirg 1893.Oscillatoria terebriformis f. brevis Kützing ex Schwabe 1944.Arthrospira brevis Drouet 1968.Arthrospira neapolitana (Kützing ex Gomont) Drouet 1969.
Filaments benthic, dark blue-green to olive-green, forming extensive mats, not pseudobranched,usually entnagled. Sheaths absent. Trichomes isopolar, straight or slightly curved, bright blue-green, 4.0-6.5 μm wide, not constricted at the cross-walls, not hooked, abruptly attenuated at theends, motile with intense oscillation. Cells cylindrical, two to three times shorter than wide, 1.5-3.0 (-3.5) μm long, contents finely granular, not granulated at the cross-walls. Apical cell obtuse-conical or rounded-conical, without calyptra or thickened outer cell wall. Heterocytes and akinetesabsent. Cell division by the transverse fission, cells reach to the original size before the next division,Reproduction by disintegration of the trichome into motile hormogonia with help of necridic cells.
TYPE LOCALITY: Germany (type of Oscillatoria brevis Kützing).DISTRIBUTION: Worldwide.KOREA: Gyenggi bay in the mid-west coast of Korea (Lee and Lee 1981); Muchangpo and Maryangri
in CN (Yoo and Kim 1990); Inchon dock (Yoo et al. 1996); Uljin in GB (Kim et al. 2004; Kim and Ahn2005); Wolseong in the east coast of Korean (Kim and Ahn 2006); Gori in Busan (Kim et al. 2007).
SPECIMEN EXAMINED: GB 08.ix.2010.ECOLOGY: Freshwater, benthic, mainly on mud, in shallow stagnant waters, road pools, ditches,
puddles, soil cultures, on mud of lakes, moist walls.REMARKS: Reference record from Whitton (2011) is as Oscillatoria brevis Kützing ex Gomont. The
records from thermal springs, brackish and marine waters, rocks and walls of marine concern pro-bably other types from this taxonomic group (Komárek and Anagnostidis 2005). Distribution recordsfrom Korea are Oscillatoria brevis Kützing ex Gomont and from marine habitat.
30. Phormidium chlorinum (Kützing ex Gomont) Umezaki et Watanabe1994: 194 (Figs. 27C, 30C, 32A, B).
Chung 1993: 219. f. 96. Komárek and Anagnostidis 2005: 436. f. 634. McGregor 2007: 55. f. 9E. pl. 7E.
BASIONYM: Oscillatoria chlorina Kützing ex Gomont 1892.SYNONYM: Oscillaria chlorina Kützing 1843, nom. inval.Lyngbya chlorina Hansgirg 1885, nom. inval.Lyngbya amoena var. chlorina (Kützing ex Gomont) Hansgirg ex Forti 1907.Phormidium chlorinum (Kützing ex Gomont) Anagnostidis 2001.
Filaments benthic, yellow-green, often in solitary filaments, rarely loosely fasciculate bundles grow-ing among algae. Sheaths indistinct or absent. Trichomes isopolar, straight to variously flexuous,yellow-green, 3.5-4.0 (-6.0) μm wide, not or slightly constricted at cross-walls. Cells more or lessisodiametric, (2.5-) 3.7-8.0 μm long, slightly granular and not granulated at cross-walls, withoutaerotopes. Apical cell widely-rounded to slightly flattened, without calyptra or thickened outer
Cyanophyceae: Oscillatoriales: Phormidiaceae 63

cell wall. Heterocytes and akinetes absent. Cell division by the transverse fission, cells reach to theoriginal size before the next division, Reproduction by disintegration of the trichome into motilehormogonia with help of necridic cells.
TYPE LOCALITY: Bennstädt, Halle, Germany (type of Oscillaria chlorina Kützing).DISTRIBUTION: Probably worldwide.KOREA: Lake Chungju in CB (Kim et al. 1988).SPECIMEN EXAMINED: Duyangli in GN (35°04′49′′N, 127°53′30′′E), 02.vii.2011.ECOLOGY: Benthic on organic mud, sometimes also in slight saline waters.REMARKS: Morphologically variable (Komárek and Anagnostidis 2005). Oscillatoria chlorina var.
64 Algal Flora of Korea∙Freshwater Cyanoprokaryota II
Fig. 32. A, B. Phormidium chlorinum.
A
B
100 μm
50 μm

perchlorina Lauterborn 1915 has slightly constricted cross-walls. Distribution record in Korea andreference records from Chung (1993) and Hu and Wei (2006) are as Oscillaria chlorina Kützing.
31. Phormidium corium (C. Agardh) Kützing ex Gomont 1892 (Fig. 33A)
Chung 1993: 225. f. 124. Komárek and Anagnostidis 2005: 450. f. 660. Whitton 2011: 103. pl. 22G.
BASIONYM: Oscillatoria corium C. Agardh 1812.SYNONYM: Phormidium corium (C. Agardh) Kützing 1843.Microcoleus corium (C. Agardh) Rabenhorst 1847.
Cyanophyceae: Oscillatoriales: Phormidiaceae 65
Fig. 33. A. Phorimidium corium; B, C. Phormidium granulatum.
A B C
25 μm

Lyngbya corium (C. Agardh) Cooke 1884.Lyngbya corium (C. Agardh) Cooke ex Forti 1907.
Filaments benthic, periphytic, blue-green to black-green or black-brown, forming thin, membranousor net-like expanded mats, densely entangled. Sheath thin, firm, colorless, sometimes diffluentand mucilaginous. Trichomes long, more or less variously curved, bright blue-green, dark-green,sometimes slightly violet, 3.0-5.5 (8.0) μm wide, not constricted or slightly constricted at the cross-walls, not or slightly attenuated at the ends, straight. Cells isodiametric up to 2 times longer thanwide, 3.4-6.4 (8.0) μm long, at the ends somewhat longer. Apical cells obtuse conical or rounded-conical, without calyptra, without or with thickened outer cell wall. Heterocytes and akinetes absent.Cell division by the transverse fission, cells reach to the original size before the next division, Repro-duction by disintegration of the trichome into motile hormogonia with help of necridic cells.
LETOTYPE: Not specified, presumably Germany (Silva et al. 1996). Notes: This type locality was firstdescribed by Drouet (1968) (Silva et al. 1996).
DISTRIBUTION: Probably Worldwide.KOREA: Uljin, Wolseong, Gori and Youngkwang coastal water (Kim and Kim 1991); Mid-western
coast of Korea (Kim et al. 1995); JB (Chung 1999).SPECIMEN EXAMINED: Seojui in GN (35°27′53′′N, 127°47′34′′E), 2011.ECOLOGY: Freshwater, periphytic, epilithic on stones and particularly epiphytic, in stagnant and
flowing waters, in rivers and cataracts, flowing cold waters.REMARKS: Data from soils, walls, green-house, mosses, decaying plants, thermal springs, brackish
coastal and littoral rock-pools are problematic. Populations with acute-conical terminal cells up to10 μm long, belong probably to another species (Komárek and Anagnostidis 2005).
32. Phormidium crouaniorum Gomont 1892: 175 (Fig. 34A, B).
Komárek and Anagnostidis 2005: 431. f. 233.
SYNONYM: Oscillatoria nigroviridis f. crassior Playfair 1913.
Filaments metaphytic, microscopic, pale blue-green. Sheaths absent or present, thin, diffluent,mucilaginous. Trichomes isopolar, straight or slightly curved, (6.5-) 7.5-10.5 (-11.0) μm wide, notor slightly constricted at the cross-wall. Cells cylindrical, shorter than wide or more or less isodia-metric, 4.0-8.0 μm long, contents finely granular, not granulated at the cross-walls. Apical cell obtuse-conical, without calyptra or thickened outer cell wall. Heterocytes and akinetes absent. Cell divi-sion by the transverse fission, cells reach to the original size before the next division, Reproductionby disintegration of the trichome into motile hormogonia with help of necridic cells.
TYPE LOCALITY: Guyana (Hab. Guyanam).DISTRIBUTION: Probably worldwide.KOREA: Rice field (Komárek and Anagnostidis 2005).SPECIMEN EXAMINED: Sicheonkyo in GG (37°34′14′′N, 126°40′45′′E), 07.vii.2011.ECOLOGY: Freshwater, described from ditches, rice field in Korea.
66 Algal Flora of Korea∙Freshwater Cyanoprokaryota II
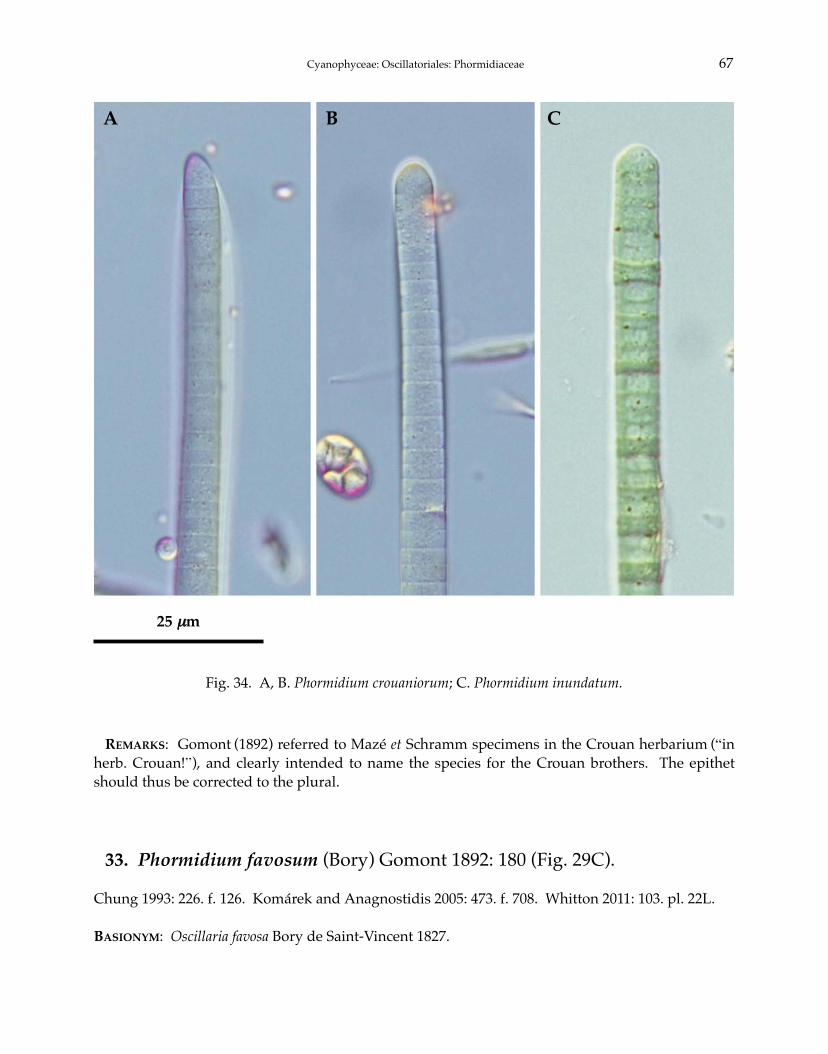
REMARKS: Gomont (1892) referred to Mazé et Schramm specimens in the Crouan herbarium (“inherb. Crouan!”), and clearly intended to name the species for the Crouan brothers. The epithetshould thus be corrected to the plural.
33. Phormidium favosum (Bory) Gomont 1892: 180 (Fig. 29C).
Chung 1993: 226. f. 126. Komárek and Anagnostidis 2005: 473. f. 708. Whitton 2011: 103. pl. 22L.
BASIONYM: Oscillaria favosa Bory de Saint-Vincent 1827.
Cyanophyceae: Oscillatoriales: Phormidiaceae 67
Fig. 34. A, B. Phormidium crouaniorum; C. Phormidium inundatum.
A B C
25 μm

Filaments benthic, dark blue-green or olive green, forming thickly attached layers to submersedsubstrata. Sheaths usually absent or fine, thin, diffluent, mucilaginous. Trichomes isopolar, long,more or less curved or straight, sometimes regularly loosely spirally-coiled, (4.0-) 4.5-7.5 (-9.0) μmwide, not constricted at the cross-walls, slightly gradually attenuated at the ends. Cells cylindrical,isodiametric to half times shorter than wide, rarely slightly longer, (2.4-) 3.0-7.0 μm long, contentshomogeneous, coarsely granulated at cross-walls. Apical cells capitate, with obtuse-conical to almosthemispherical calyptra. Heterocytes and akinetes absent. Cell division by the transverse fission,cells reach to the original size before the next division. Reproduction by disintegration of the trichomeinto motile hormogonia with help of necridic cells.
TYPE: Uncertain.DISTRIBUTION: Possibly wordwide.KOREA: JB (Chung 1999).SPECIMEN EXAMINED: Sugili in JN (34°20′51′′N, 127°27′33′′E), 12.xi.2011.ECOLOGY: Freshwater, mostly in cold, flowing waters on limestone substrata, benthic attached on
rocks, stones, on sand, and hydrophytes. Occasionally penetrating limestone surface (Komárek andAnagnostidis 2005; Whitton 2011).
REMARKS: The records from thermal springs probably are of another species (Komárek and Anag-nostidis 2005).
34. Phormidium granulatum (N.L. Gardner) Anagnostidis 2001: 370(Figs. 33B, C, 35A, B).
Komárek and Anagnostidis 2005: 439. f. 640. Hu and Wei 2006: 140. pl. II-27-10. McGregor 2007:56. f. 9G. pl. 7M. Whitton 2011: 99.
BASIONYM: Oscillatoria granulata N.L. Gardner 1927.SYNONYM: Tychonema granulatum (N.L. Gardner) Anagnostidis et Komárek 1988.
Filaments benthic, dark blue-green to olive-green, forming small mat, slimy flakes or solitary tri-chomes, rarely scattered among other algae. Sheaths absent. Trichomes cylindrical, more or lessstraight, slightly flexuous, particularly at the apex, (2.8-) 3.0-4.0 (-5.0) μm wide, not or very slightlyconstricted at cross-walls, not attenuated towards ends, motile. Cells more or less isodiametric, orsometimes shorter than wide, 2.0-5.0 (5.5) μm long, contents finely and distinct granules at the cross-walls (usually 2-5 in one cell). Apical cell rounded, without or with slightly thickened cell walls,calyptra absent. Heterocytes and akinetes absent. Cell division by the transverse fission, cells reachto the original size before the next division. Reproduction by disintegration of the trichome intomotile hormogonia with help of necridic cells.
TYPE LOCALITY: Santurce, Puerto Rico.DISTRIBUTION: Worldwide.KOREA: Lake Chungju in CB (Kim et al. 1988); Geum River (Kim et al. 2000).SPECIMEN EXAMINED: Suncheon in JN (34°59′56′′N, 127°29′11′′E); Daeeul-ri in JJ (33°26′56′′N, 126°
68 Algal Flora of Korea∙Freshwater Cyanoprokaryota II

Cyanophyceae: Oscillatoriales: Phormidiaceae 69
Fig. 35. A, B. Phormidium granulatum.
50 μm
50 μm
A
B

40′23′′E), 10.ii.2012.ECOLOGY: Freshwater, in stagnant, rarely slowly flowing, usually unpolluted waters with water
vegetation.REMARKS: An assessment of several similar species (P. tergestinum, P. nigrum etc.) is required and
the identity of all similar populations should be revised (Komárek and Anagnostidis 2005). Distri-bution Records in Korea and reference records from Hu and Wei (2006) and Whitton (2011) are asOscillatoria granulata Gardner.
35. Phormidium inundatum Kützing ex Gomont 1892: 172 (Fig. 34C).
Chung 1993: 226. f. 127. Komárek and Anagnostidis 2005: 430. f. 618. Whitton 2011: 104. pl. 22J.
SYNONYM: Phormidium inundatum Kützing 1843, nom. inval.Oscillaria inundata (Kützing) P.L. Crouan et H.M. Crouan 1867, nom. inval.Lyngbya inundata (Kützing) Kirchner 1880, nom. inval.Lyngbya inundata Kützing ex Gomont 1892.Hypheothrix inundata (Kützing ex Gomont) Hansgirg ex Dalla Torre et Sarnthein 1901.Oscillatoria inundata P.L. Crouan et H.M. Crouan ex Forti 1907.
Filaments benthic, dark blue-green, rarely pale green, forming thin membranaceous, usuallycoiled together or fasciculated. Sheaths thin and diffluent. Trichomes more or less straight, some-times curved, blue-green, 3.0-5.0 μm wide, not constricted at the cross-walls, straight and brieflyattenuated at the ends. Cells more or less isodiametric, (3.0-) 3.3-8.0 μm long, contents homogeneous,granulated at the cross-walls. Apical cell obtuse-conical or rounded-conical, without calyptra orthickened outer cell wall. Heterocytes and akinetes absent. Cell division by the transverse fission,cells reach to the original size before the next division. Reproduction by disintegration of the tri-chome into motile hormogonia with help of necridic cells.
TYPE LOCALITY: Germany (type of Phormidium inundatum Kützing).DISTRIBUTION: Worldwide.KOREA: JB (Chung 1976); JN (Wui and Kim 1987); Galgok stream and Incheon river in JN (Lee et
al. 2008).SPECIMEN EXAMINED: Nammaeji in GB (35°49′44′′N, 128°44′32′′E), 30.viii.2010.ECOLOGY: Freshwater, periphytic in oligotrophic to mesotrophic flowing and stagnant waters on
submersed vegetation, possibly also on moist and walls.REMARKS: Reports from subpolar regions, tropical lake and thermal springs are probably of another
species (Komárek and Anagnostidis 2005).
70 Algal Flora of Korea∙Freshwater Cyanoprokaryota II

36. Phormidium nigrum (Vaucher ex Gomont) Anagnostidis et Komárek1988: 405 (Fig. 36A, B).
Chung 1993: 221. f. 105. Komárek and Anagnostidis 2005: 449. f. 658. Whitton 2011: 99.
BASIONYM: Oscillatoria nigra Vaucher ex Gomont 1892.SYNONYM: Oscillatoria nigra Vaucher 1803, nom. inval.Trichophorus niger (Vaucher) Desvaux 1809, nom. inval.Oscillatoriella nigra (Vaucher) Gaillon 1833, nom. inval.Lyngbya nigra (Vaucher) Hansgirg 1884, nom. inval.
Filaments benthic, metaphytic, variably colored, blackish brown, olive-brown to red-violet, form-
Cyanophyceae: Oscillatoriales: Phormidiaceae 71
Fig. 36. A, B. Phormidium nigrum; C. Phormidium tergestinum.
25 μm
A B C

ing more or less leathery or membranous mats. Sheaths absent or present, rarely thin and colorless.Trichomes isopolar, straight or more or less curved, blue-green, olive-green, red-violet, 6.0-8.5 (-10.0)μm wide, not or somewhat constricted at the cross-walls, rarely shortly and slightly attenuated atthe ends, motile with usually slowly gliding, oscillation and rotation. Cells cylindrical, three timesshorter than wide or isodiametric, contents homogeneous, finely granular, mostly coarsely granu-lated at the cross-walls. Apical cells rounded-truncate or widened-conical, sometimes without calyp-tra, with thickened outer cell wall. Heterocytes and akinetes absent. Cell division by the transversefission, cells reach to the original size before the next division. Reproduction by disintegration ofthe trichome into motile hormogonia with help of necridic cells.
TYPE LOCALITY: “sur la surface des eaux peu courantes”; probably Switzerland (type of Oscillatorianigra Vaucher).
DISTRIBUTION: Probably worldwide.KOREA: JB (Chung 1976); Lake Chungju in CB (Kim et al. 1988); Geum River (Kim et al. 2000);
Galgok stream and Incheon river in JN (Lee et al. 2008).SPECIMEN EXAMINED: Koggangkyo in GB (36°07′20′′N, 129°20′51′′E), 21.xi.2011.ECOLOGY: Freshwater, benthic, attached to the substratum, later sometimes free floating, in stag-
nant and slow-flowing waters, also from low temperature thermal springs (Komárek and Anag-nostidis 2005).
REMARKS: Reference record from Chung (1993) is as Oscillatoria nigra Vaucher. Reference recordfrom Whitton (2011) is as Oscillatoria nigra Vaucher ex Gomont. Distribution records from Korea arethe records as Oscillatoria nigra Vaucher.
37. Phormidium okenii (C. Agardh) Anagnostidis et Komárek 1988: 405(Fig. 37A-C).
Chung 1993: 221. f. 107. Komárek and Anagnostidis 2005: 411. f. 583. McGregor 2007: 57. f. 10D.Whitton 2011: 100.
BASIONYM: Oscillatoria okenii C. Agardh 1827.SYNONYM: Lyngbya okenii (C. Agardh) Hansgirg 1884.Oscillatoria okenii C. Agardh ex Gomont 1892.
Filaments benthic, dark blue-green to blackish, forming mats. Sheath absent or thin, delicate ordiffluent. Trichomes isopolar, usually long, straight to slightly curved, bright blue-green, 5.0-9.0μm wide, slightly to distinctly constricted at the cross-walls, more or less gradually attenuated,undulating and slightly hooked at the ends. Cells cylindrical, mostly three times shorter than wide,2.5-5.0 (-6.0) μm long, at the ends 5.5-8.5 (-9.0) μm long and sometimes almost isodiametric, con-tents homogeneous or with minute granules, frequently translucent cross-walls. Apical cell obtu-sely-rounded to rounded-conical, almost pointed, without calyptra or thickened outer cell wall.Heterocytes and akinetes absent. Cell division by the transverse fission, cells reach to the originalsize before the next division. Reproduction by disintegration of the trichome into motile hormo-gonia with help of necridic cells.
72 Algal Flora of Korea∙Freshwater Cyanoprokaryota II
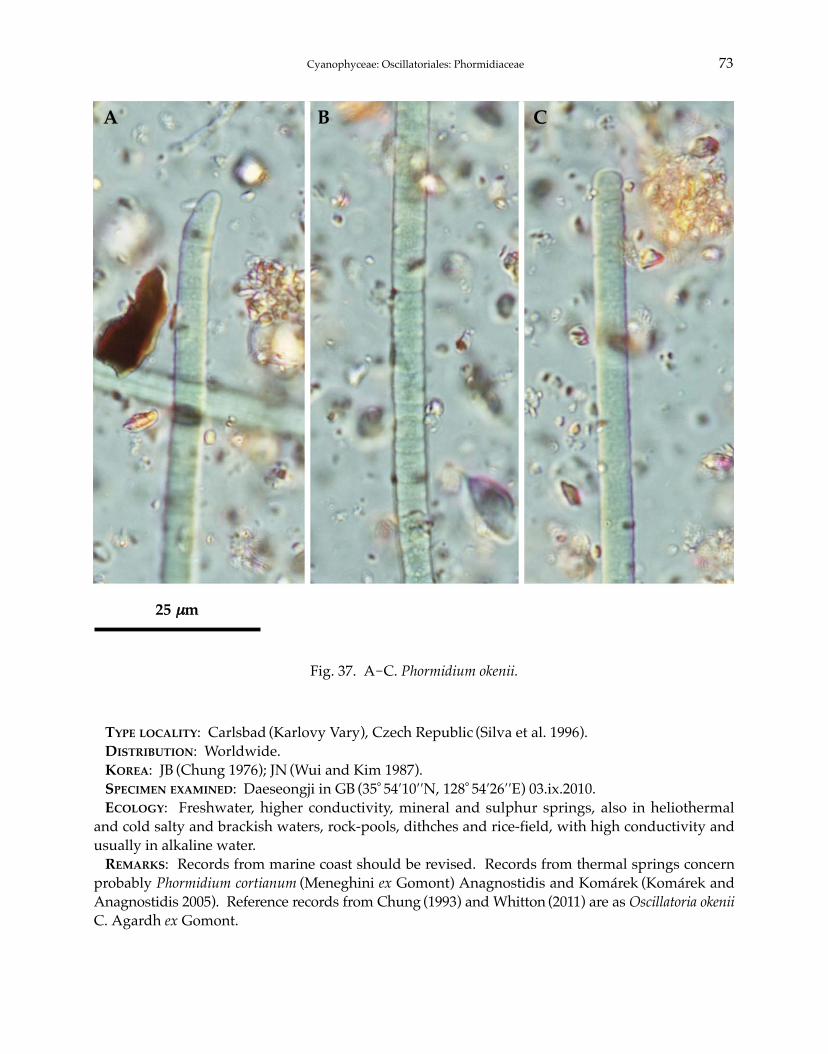
TYPE LOCALITY: Carlsbad (Karlovy Vary), Czech Republic (Silva et al. 1996).DISTRIBUTION: Worldwide.KOREA: JB (Chung 1976); JN (Wui and Kim 1987).SPECIMEN EXAMINED: Daeseongji in GB (35°54′10′′N, 128°54′26′′E) 03.ix.2010.ECOLOGY: Freshwater, higher conductivity, mineral and sulphur springs, also in heliothermal
and cold salty and brackish waters, rock-pools, dithches and rice-field, with high conductivity andusually in alkaline water.
REMARKS: Records from marine coast should be revised. Records from thermal springs concernprobably Phormidium cortianum (Meneghini ex Gomont) Anagnostidis and Komárek (Komárek andAnagnostidis 2005). Reference records from Chung (1993) and Whitton (2011) are as Oscillatoria okeniiC. Agardh ex Gomont.
Cyanophyceae: Oscillatoriales: Phormidiaceae 73
Fig. 37. A-C. Phormidium okenii.
A B C
25 μm

38. Phormidium taylori (Drouet et Strickland) Anagnostidis 2001: 371(Fig. 38A, B).
Komárek and Anagnostidis 2005: 451. f. 661.
74 Algal Flora of Korea∙Freshwater Cyanoprokaryota II
Fig. 38. A, B. Phormidium taylori.
50 μm
50 μm
A
B

BASIONYM: Lyngbya taylori Drouet et Strickland 1940.
Filaments benthic, blue-green to dark blue-green, usually more or less flexuous, very long fila-ments, forming tufts with parallel arranged, sometimes radiating and penicillate. Sheaths thin,firm, colorless, later thickened and lamellated, opened at the ends, containing always a single tric-home. Trichomes cylindrical, 4.0-7.0 μm wide, not or only slightly constricted at cross-walls, notattenuated at the ends. Cells more or less isodiametric, or little shorter than wide, 2.0-7.0 μm long,contents granulated but not granulated at the cross-walls. Apical cells broadly convex, sometimeswith thickened cell wall. Heterocytes and akinetes absent. Cell division by the transverse fission,cells reach to the original size before the next division. Reproduction by disintegration of the tric-home into motile hormogonia with help of necridic cells.
TYPE: Uncertain.DISTRIBUTION: Probably worldwide.KOREA: Geum River (Chung and Lee 1979).SPECIMEN EXAMINED: Gyeongsan in GB 26.viii.2010.ECOLOGY: Freshwater, benthic, epipelic, forming biofilm on submersed substrata, on plants,
rocks, and submersed wood, later sometimes free-floating, tychoplanktonic.REMARKS: Distribution record from Korea is the records as Lyngbya taylori Drouet et Strickland.
39. Phormidium terebriforme (C. Agardh ex Gomont)Anagnostidis et Komárek 1988: 406 (Fig. 39A-C).
Chung 1993: 224. f. 120. Komárek and Anagnostidis 2005: 412. f. 584. Hu and Wei 2006: 138. pl. II-27-3. McGregor 2007: 58. f. 10I. pl. 8I, J. Whitton 2011: 101.
BASIONYM: Oscillatoria terebriformis C. Agardh ex Gomont 1892.SYNONYM: Oscillatoria terebriformis C. Agardh 1827, nom. inval.
Filaments benthic, metaphytic, dark blue-green to gray-green, forming loosely tangled mat. Sheathsabsent. Trichomes long, straight or flexuous, bright blue-green, sometimes olive-green, 4.0-6.5 μmwide, not or sometimes slightly constricted at the cross-walls, usually loosely twisted and graduallyattenuated at the ends, motile with anticlockwise rotation and oscillation. Cells shorter than wideup to isodiametric, 2.0-6.0 (-7.0) μm long, contents usually finely granular, rarely large granulatedat the cross-walls. Apical cells rounded-hemispherical or slightly depressed and almost truncate toconical, without calyptra or thickened outer cell wall. Heterocytes and akinetes absent. Cell divisionby the transverse fission, cells reach to the original size before the next division. Reproduction bydisintegration of the trichome into motile hormogonia with help of necridic cells.
TYPE LOCALITY: Carlsbad, Czech Republic (type of Oscillatoria terebriformis C. Agardh).DISTRIBUTION: Worldwide.KOREA: JB (Chung 1976); Lake Chungju in CB (Kim et al. 1988); JN (Wui and Kim 1987); Beopsu
marsh GN (Kim 2008).
Cyanophyceae: Oscillatoriales: Phormidiaceae 75

SPECIMEN EXAMINED: Kimyoungri in GB (36°35′28′′N, 128°12′51′′E) 2011.ECOLOGY: Freshwater, periphytic and benthic in springs, littoral of ponds and lakes, particularly
in waters with submerged vegetation.REMARKS: This species is morphologically very variable (Komárek and Anagnostidis 2005). Refer-
ence records from Chung (1993), Hu and Wei (2006) and Whitton (2011) are as Oscillatoria terebriformisC. Agardh ex Gomont. Distribution records from Korea are the records as Oscillatoria terebriformisC. Agardh ex Gomont.
76 Algal Flora of Korea∙Freshwater Cyanoprokaryota II
Fig. 39. A-C. Phormidium terebriforme.
25 μm
A B C

40. Phormidium tergestinum (Rabenhorst ex Gomont)Anagnostidis et Komárek 1988: 406 (Figs. 36C, 40A-C, 41A, B).
Chung 1993: 224. f. 119. Komárek and Anagnostidis 2005: 451. f. 662. McGregor 2007: 60. f. 10H.pl. 8K.
BASIONYM: Oscillatoria tenuis var. tergestina Rabenhorst ex Gomont 1892.SYNONYM: Oscillatoria tenuis f. tergestina (Rabenhorst ex Gomont) Elenkin 1949.Oscillatoria tergestina Kützing 1836.Lyngbya phormidium Kützing 1847.
Cyanophyceae: Oscillatoriales: Phormidiaceae 77
Fig. 40. A-C. Phormidium tergestinum.
25 μm
A B C

78 Algal Flora of Korea∙Freshwater Cyanoprokaryota II
Fig. 41. A, B. Phormidium tergestinum.
100 μm
50 μm
A
B

Symploca phormidium (Kützing) Thuret 1875.Lyngbya phormidium Kützing ex Gomont 1892.
Filaments benthic, usually bright blue-green to olive-green, thin, mucilaginous. Sheaths presentor absent, occur very facultatively. Trichomes isopolar, usually straight, sometimes slightly andirregularly curved, varying length, color and width, 4.0-10.0 (-11.0) μm wide, not or slightly toclearly constricted at cross-walls, not attenuated bent at the ends, motile with oscillation and rota-tion. Cells cylindrical, usually shorter than wide, rarely isodiametric to longer than wide, (1.6-)2.4-6.5 (-10.5) μm long, contents homogeneous, sometimes with large granules, sometimes translu-cented and granulated at the cross-walls. Apical cells rounded or more or less hemispherical, withor without thickened cell wall. Heterocytes and akinetes absent. Cell division by the transversefission, cells reach to the original size before the next division. Reproduction by disintegration ofthe trichome into motile hormogonia with help of necridic cells.
TYPE LOCALITY: Trieste, Italy (type of Oscillatoria tergestina Kützing).DISTRIBUTION: Probably worldwide with the exception of the polar regions.KOREA: Previously unrecorded species.SPECIMEN EXAMINED: Yangbaekli in GB (36°07′20′′N, 129°20′51′′E), 19.xi.2011; Duyangli in GN
(35°04′49′′N, 127°53′30′′E), 02.xii.2011.ECOLOGY: Freshwater, benthic, common on the surface of mud and other substratum, in stagnant
and flowing waters, ditches wells, streams, sometimes rich in organic nutrients, also in pollutedwaters.
REMARKS: The records from saline waters and usually low-temperature thermal springs, moistsoils, bark of trees, are very problematic (Komárek and Anagnostidis 2005). Reference record fromChung (1993) is as Oscillatoria tenuis var. tergestina Rabenhorst.
41. Phormidium violaceum (Wallroth ex Gomont) Anagnostidis et Komárek1988: 406 (Fig. 42A-D).
Komárek and Anagnostidis 2005: 469. f. 697.
BASIONYM: Oscillatoria violacea Wallroth ex Gomont 1892.SYNONYM: Phormidium violaceum (Wallroth ex Gomont) Anagnostidis 2001.Oscillatoria violacea Wallroth 1833, nom. inval.Lyngbya violacea Hansgirg 1892.
Filaments submersed or subaerophytic, blue-green, greyish-blue or slightly violet, mat-formingor small bundles of filaments. Sheaths lacking or diffluent. Trichomes isopolar, straight or slightlyflexuous, reddish or violet-blue-green, not constricted at cross-walls, slightly and gradually attenuatedat the ends and rarely slightly curved, 4.0-5.7 μm wide. Cells cylindrical, shorter than wide toisodiametric, 2.5-4.3 μm long, contents finely granulated and distinct granulation at cross-walls.Apical cells narrow-elongated and with small more or less spherical calyptra. Heterocytes andakinetes absent. Cell division by the transverse fission, cells reach to the original size before thenext division. Reproduction by disintegration of the trichome into motile hormogonia with help of
Cyanophyceae: Oscillatoriales: Phormidiaceae 79

80 Algal Flora of Korea∙Freshwater Cyanoprokaryota II
Fig. 42. A-D. Phormidium violaceum.
20 μm
20 μm
A B
C
D

necridic cells.
TYPE LOCALITY: Germany (type of Oscillatoria violacea Wallroth).DISTRIBUTION: Probably worldwide.KOREA: Previously unrecorded species.SPECIMEN EXAMINED: Uljin in GB, 06.ix.2005.ECOLOGY: Submerged in small water bodies on stony substrata and subaerophytic on various wet
substrata.REMARKS: The site of specimens examined was sandfilter in Uljin water plant. Subaerophytic on
various wet substrata in glasshouses (Komárek and Anagnostidis 2005).
Genus Phormidiochaete Komárek in Anagnostidis 2001: 374.
Filaments heteropolar, distinctly widened or narrowed to wards ends, sometimes coiled and creep-ing on the substratum by their base, or more or less erect (at least at the ends), sometimes falselybranched and forming mat or fasciculated colonies, mat sometimes crusty. Sheaths distinct, openat the ends and usually slightly widened, sometimes slightly lamellated. Trichomes mainly hetero-polar, constricted or unconstricted at cross-walls, rounded at the base, continually narrowed towardsends, up into hair-like formations. Cells more or less isodiametric or slightly shorter or longer thanwide in the middle of the trichome. Heterocytes and akinetes absent. Reproduction by hormogonia.
HOLOTYPE: Phormidiochaete nordstedtii (Bornet et Flahault) Komárek 2001.
NUMBER OF SPECIES: Of the 4 species are reported up to date, 4 are currently accepted as taxono-mically valid taxa (Guiry and Guiry 2012).
DISTRIBUTION: Worldwide, benthic in freshwater.KEY REFERENCE: Anagnostidis and Komárek (1988), Komárek and Anagnostidis (2005), McGregor
(2007), Whitton (2011).REMARKS: This genus belongs to Subfamily Ammatoideoideae. Trichomes shape is elongated
into cellular hairs at the apical end. This genus belong to Homoeothrix (Thuret ex Bornet et Flahault)Kirchner 1898 until 2005. Komárek and Anagnostidis (2005) recognized two segregate genera, Tapi-nothrix Sauvageau 1892 and Phormidiochaete Komárek in Anagnostidis 2001, based on trichomemorphology and ultrastructure. The cells of Homoeothrix are always shorter than wide, it is the oneof characteristics Family Oscillatoriaceae.
Key to the species of genes Phormidiochaete
1. Filaments to 7.5 μm wide at the base, trichomes to 4.0 μm wide, not or very slightly constrictedat cross-walls, sheaths often yellow or violet····································································P. fusca
- Filaments usually 5.0-18.0 μm wide at the base, trichomes 4.0-11.0 μm wide, distinctly constrictedat cross-walls, sheaths yellow-brown at the base·······································································2
2. Cells mainly longer than wide, sheaths thick, lamellated, divergent, trichomes 4.0-9.0 μm wide·······································································································································P. balearica
- Cells mainly shorter than wide, up to more or less isodiametric, sheaths thin, not or slightly lamel-lated, rarely divergent, trichomes 4.0-11.0 μm wide················································P. nordstedtii
Cyanophyceae: Oscillatoriales: Phormidiaceae 81
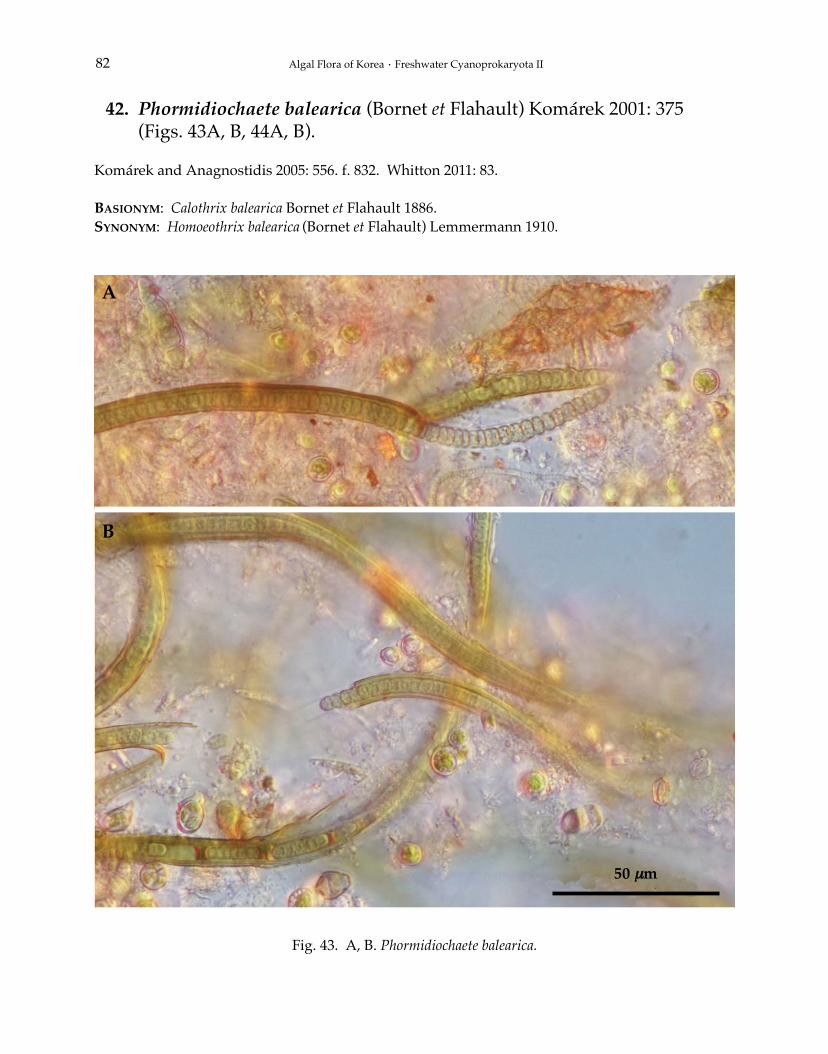
42. Phormidiochaete balearica (Bornet et Flahault) Komárek 2001: 375(Figs. 43A, B, 44A, B).
Komárek and Anagnostidis 2005: 556. f. 832. Whitton 2011: 83.
BASIONYM: Calothrix balearica Bornet et Flahault 1886.SYNONYM: Homoeothrix balearica (Bornet et Flahault) Lemmermann 1910.
82 Algal Flora of Korea∙Freshwater Cyanoprokaryota II
Fig. 43. A, B. Phormidiochaete balearica.
50 μm
A
B
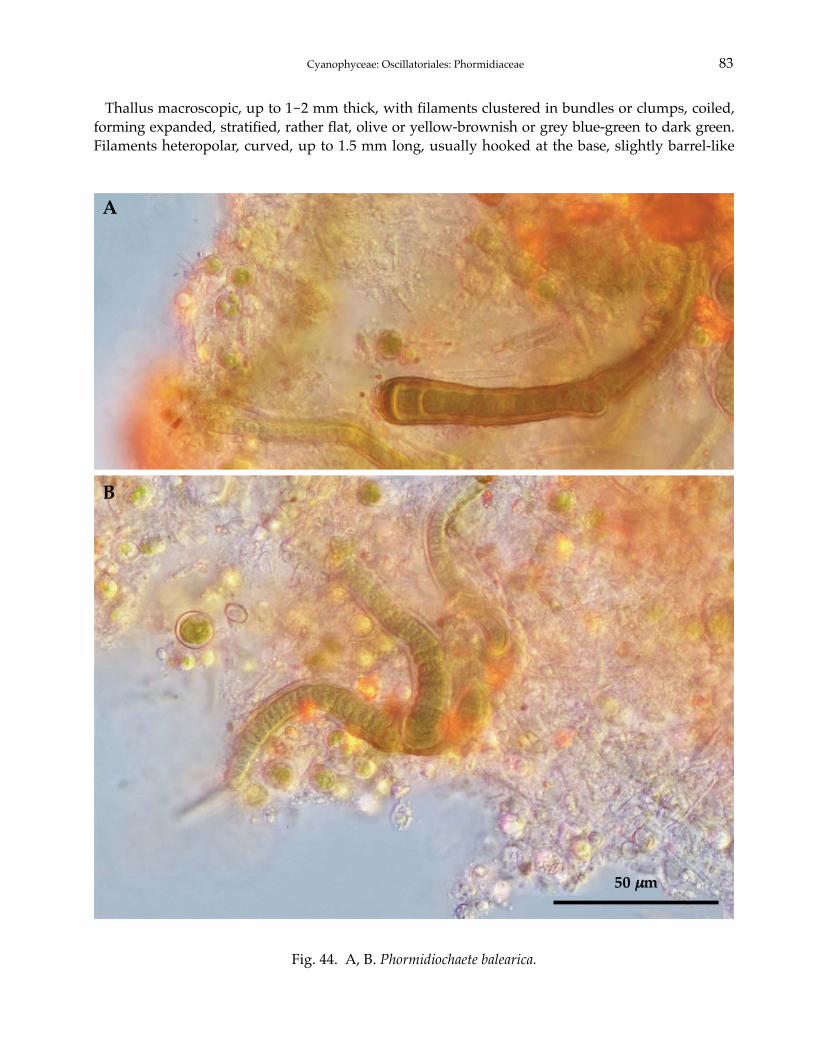
Thallus macroscopic, up to 1-2 mm thick, with filaments clustered in bundles or clumps, coiled,forming expanded, stratified, rather flat, olive or yellow-brownish or grey blue-green to dark green.Filaments heteropolar, curved, up to 1.5 mm long, usually hooked at the base, slightly barrel-like
Cyanophyceae: Oscillatoriales: Phormidiaceae 83
Fig. 44. A, B. Phormidiochaete balearica.
50 μm
A
B

widened or not widened, (3.5-) 5.0-15.0 μm wide, usually pseudobranched, creeping on the sub-stratum by their base. Sheaths thick, firm, yellow-brownish, less frequently yellowish or colorless,usually lamellated, smooth or frayed outside at the lower and upper parts. Trichomes pale or greyblue-green, olive or yellowish, (2.5-) 4.0-9.0 μm wide at the base, constricted at ungranulated cross-walls, variously long, gradually attenuated at the ends, ending in a long, colorless or glaucous long-celled hair. Cells barrel-shaped up to nearly spherical, shorter at the base, further above isodiametricto 2 times as long as wide; basal cells shorter. Heterocytes and akinetes absent. Reproduction byHormogonia with 3-8 celled.
TYPE LOCALITY: Minorca, Balearic Islands.DISTRIBUTION: Worldwide.KOREA: Previously unrecorded species.SPECIMEN EXAMINED: Bonghwali in GN (34°48′06′′N, 128°02′18′′E), 26.xi.2011.ECOLOGY: Subaerophytic, freshwater, epilithic, on moist calcareous rocks, on stony or peaty banks
of water reservoirs, also on old wood and in stony littoral of lakes and steep wet rocks, probablyonly in a limestone substratum.
REMARKS: Reference record from Whitton (2011) is as Homoeothrix balearica (Bornet et Flahault)Lemmermann.
43. Phormidiochaete fusca (Starmach) Komárek 2001: 375(Figs. 45A-C, 46A-C).
Komárek and Anagnostidis 2005: 554. f. 831. Whitton 2011: 84. pl. 17C.
BASIONYM: Homoeothrix fusca Starmach 1934.SYNONYM: Homoeothrix fusca f. minor Starmach 1934.Homoeothrix fusca f. elongata Starmach 1934.Homoeothrix fusca f. longissima Starmach 1968; incl.
Thallus macroscopic, up to 1 cm in diameter, spot-like, attached to the substratum, filaments soli-tary or in small groups, tufted or irregularly-arranged, with a somewhat uneven surface, sometimesmixed with other algae, rarely in monospecific stands. Filaments straight or flexuous, erect orirregularly curved, unbranched, sometimes slightly wide, gradually attenuated at the end, endingin a hair, 4.5-7.5 μm wide at the base, up to 85 (-160) μm long. Sheaths wide and thick, slightlylamellated, sometimes sharply limited or divergent or telescopic, violet to black-violet, or yellowand violet. Trichomes heteropolar, pale blue-green to grey-blue, 1.5-4.0 μm wide at the base, 1.5-2.5 μm in the middle, usually ending in hair, not or weakly constricted at the cross-walls. Cellsisodiametric or somewhat shorter at the base, longer at the end than wide. Heterocytes and akinetesabsent. Reproduction by hormogonia.
TYPE LOCALITY: Tatra Lakes, Poland.DISTRIBUTION: Northern temperate zones.KOREA: Previously unrecorded species.
84 Algal Flora of Korea∙Freshwater Cyanoprokaryota II

Cyanophyceae: Oscillatoriales: Phormidiaceae 85
Fig. 45. A-C. Phormidiochaete fusca.
50 μm
A
B
C

SPECIMEN EXAMINED: Haengsinkyo in GW (37°51′05′′N, 128°47′29′′E), 20.xi.2011.ECOLOGY: Submersed or subaerophytic, epilithic on moist calcareous and silicate substrata, very
rarely epiphytic on algae, in flowing and stagnant fresh waters, clear mountain waters.REMARKS: Reference record from Whitton (2011) is as Homoeothrix fusca Starmach 1934.
44. Phormidiochaete nordstedtii (Bornet et Flahault) Komárek 2001: 375(Figs. 47A, B, 48A, B).
Komárek and Anagnostidis 2005: 556. f. 833.
BASIONYM: Dichothrix nordstedtii Bornet et Flahault 1886.SYNONYM: Calothrix caespitosa Rabenhorst 1873.
86 Algal Flora of Korea∙Freshwater Cyanoprokaryota II
Fig. 46. A-C. Phormidiochaete fusca.
25 μm
A
B C
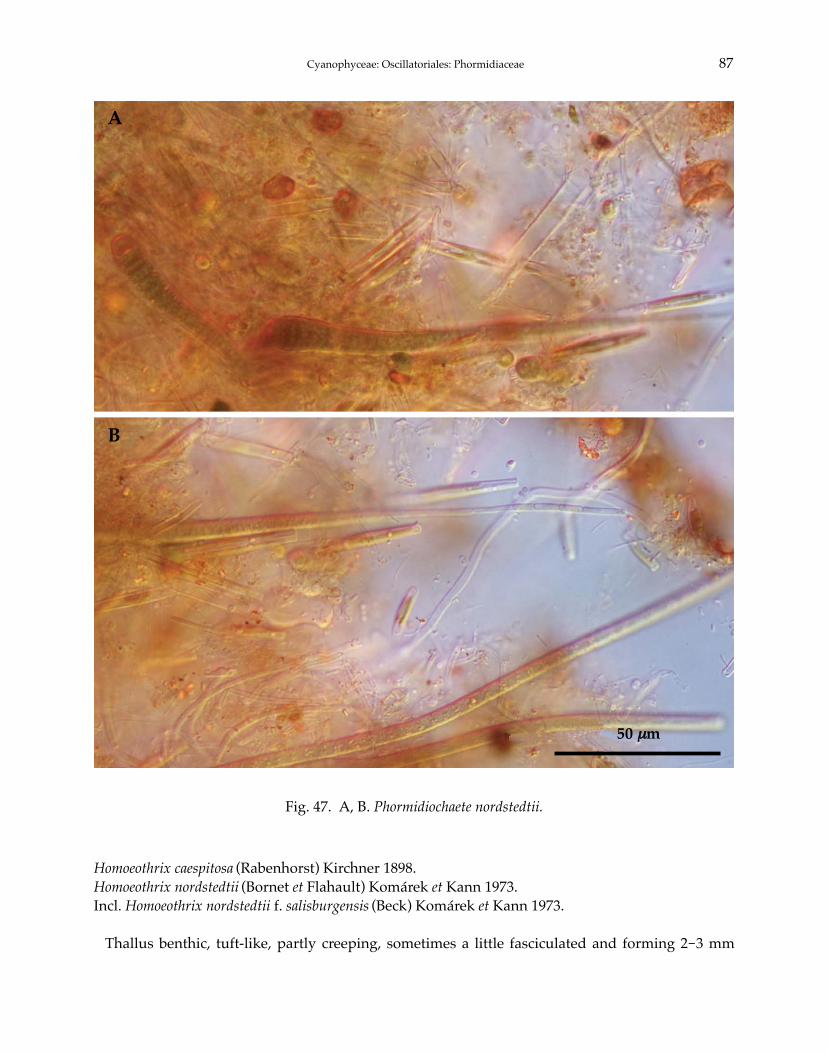
Homoeothrix caespitosa (Rabenhorst) Kirchner 1898.Homoeothrix nordstedtii (Bornet et Flahault) Komárek et Kann 1973.Incl. Homoeothrix nordstedtii f. salisburgensis (Beck) Komárek et Kann 1973.
Thallus benthic, tuft-like, partly creeping, sometimes a little fasciculated and forming 2-3 mm
Cyanophyceae: Oscillatoriales: Phormidiaceae 87
Fig. 47. A, B. Phormidiochaete nordstedtii.
50 μm
A
B

88 Algal Flora of Korea∙Freshwater Cyanoprokaryota II
Fig. 48. A, B. Phormidiochaete nordstedtii.
A
B
50 μm

high colonies. Filaments solitary or forming a small, microscopic, slightly curved, unbranched orbrush-like to ‘Dichothrix-like’ pseudobranched, slightly narrowed at the base (6.0-) 11.0-15.0 μmwide. Sheaths thin, open at the ends, not or slightly lamellated with a tendency to diverge, color-less, rarely to yellowish brown. Trichomes heteropolar, blue-green, 4.0-11.0 μm wide at the base,distinctly constricted at ungranulated cross-walls, gradually attenuated at the ends, ending in ahyaline, sometimes very long, usually long-celled hair, 100-200 μm long. Cells more or less barrel-shaped, isodiametric or shorter or longer than wide, cylindrical and longer in hair. Heterocytesand akinetes absent. Reproduction by hormogonia.
TYPE LOCALITY: Norway.DISTRIBUTION: Temperate zone.KOREA: Previously unrecorded species.SPECIMEN EXAMINED: Seojuli on Lim stream in GN (35°27′53′′N, 127°47′34′′E),01.xii.2010; Hoija
river in Ulsan (35°26′07′′N, 129°11′25′′E) 02.xii.2010.ECOLOGY: Freshwater, epilithic in flowing waters and waterfalls, rarely also in littoral of lakes,
among other algae, always on calcareous substrata, in xenosaprobic to slightly eutrophic water.REMARKS: This is the type species (holotype) of the genus Phormidiochaete.
Family Oscillatoriaceae (S.F. Gray) Harvey ex Kirchner 1989
Filaments isoploar or heteropolar, forming layered and prostrate mats, rarely in fascicles, solitaryor in clusters. Sheaths lacking (e.g., Genus Oscillatoria), or occurring (e.g., Genus Lyngbya), or onlyunder suboptimal conditions, firm and open, sometimes lamellated, generally containing one trichome,without false branching, or rarely with false branching. Trichomes cylindrical, straight or flexuous,6.0-20.0 (-80.0) μm wide, unconstricted or slightly constricted at the cross-walls, facultatively motile,gliding, oscillation, rotation, trembling. Cells discoid, always shorter than wide, usually severaltimes shorter than wide, without aerotopes. Thylakoids are short, waved, irregularly displayedmore or less throughout the whole cell content. Apical cell usually with a thickened outer cell wallor with a calyptra. Heterocytes and akinetes absent. Cell division transversal to the trichome axisin a rapid sequence, the cells not reaching their original size before the next division. Reproductionby disintegration of trichomes into short motile hormogonia, with the help of necridic cells.
GENERA AND SPECIES: 24 genera and 207 species are currently accepted as taxonomically validtaxa (Guiry and Guiry 2012). Genera containing none or one species number 17. Major genera areLyngbya (98 species), Oscillatoria (57 species), Blennothrix (20 species) and Plectonema (15 species).
DISTRIBUTION: Worldwide, planktonic or benthic in freshwater, brackish and salt waters.KEY REFERENCE: Gomont (1892), Chung (1993), Anagnostidis and Komárek (1988), Canter-lund and
lund (1995), Komárek and Anagnostidis (2005), McGregor (2007), Hindák (2008), Whitton (2011).REMARKS: This family is generally divided into two subfamilies. Subfamily Oscillatorioideae:
Trichomes isopolar, with morphologically the same ends in full-grown trichomes. Subfamily Homo-eotrichoideae: Trichomes heteropolar, with a widened basal end and narrowed. The major differ-ence of morphological characteristics between genera Lyngbya and Oscillatoria is sheath. The sheathof genus Oscillatoira is absent. The genus Lyngbya has thin sheath.
Cyanophyceae: Oscillatoriales: Oscillatoriaceae 89

Genus Lyngbya C. Agardh ex Gomont 1892: 118, nom. cons.
Filaments usually connected into thick, layered, sometimes leathery strata (mats), usually macro-scopic, to several centimeters or decimeters in diameter, with firm sheaths. Sheaths very rarely absent(usually only reproducing hormogonia), sometimes layered or stratified, colorless or slightly yellow-brown or reddish, opened at the ends, containing a single motile trichome. Trichomes isopolar,straight or slightly waved, uniseriate, composed of shortly cylindrical or barrel-shaped discoid cells,unconstricted or constricted at the cross-walls. Cells short, always shorter than wide and up toonly one-fifteenth times as long as wide, without aerotopes, or rarely in planktonic species withfacultative aerotopes, usually finely granulated, sometimes more at the cross-walls. Apical cellswith thickened outer cell wall or with prominent calyptra. Heterocytes and akinetes absent. Celldivision crosswise, perpendicular to the long axis of a trichome. Reproduction by more or lessshort, motile hormogonia by the help of necridia.
HOLOTYPE: Lyngbya confervoides C. Agardh 1824.
SPECIES: Of the 1043 species (infraspecific) reported up to date, 98 species are currently acceptedas taxonomically valid taxa (Guiry and Guiry 2012).
DISTRIBUTION: Worldwid, freshwater and marine species.KEY REFERENCE: Anagnostidis and Komárek (1988), Komárek and Anagnostidis (2005), McGregor
(2007), Whitton (2011).REMARKS: Hormogonia are formed as an ordered release of lengths of the trichome under condi-
tions favourable for growth. The release of a hormogonium often leaves a length of sheath project-ing beyond the remainder of the original trichome. The small length of sheath projecting beyondthe end provide a useful character to separate the genus from Oscillatoria (Whitton 2011).
Key to the species of genus Lyngbya
1. Freshwater, apical cells of developed trichomes with calyptra, trichomes 6.0-14.0 μm wide···············································································································································L. nigra
- Freshwater, trichomes without calyptra, but sometimes with thickened outer cell walls···········22. Filaments solitary, living in plankton························································································3- Filaments in colonies or mat, not planktonic·············································································43. Trihomes 11.0-13.0 μm wide, without thickened outer cell wall···························L. hieronymusii- Trihomes 18.0-23.0 μm wide, with thickened outer cell wall···········································L. birgei4. Sheaths always colorless···········································································································5- Sheahts in old filaments yellow up to yellow-brown, trihomes 8.0-13.0 μm wide······L. stagnina5. Apical cells always broadly-rounded, without thickened outer cell wall, trichomes 6.0-12.0 μm
wide······················································································································L. martensiana- Apical cells in developed trichomes with thickened outer cell wall, trichomes 11.0-17.0 μm wide···········································································································································L. major
90 Algal Flora of Korea∙Freshwater Cyanoprokaryota II

45. Lyngbya birgei G.M. Smith 1916: 482 (Fig. 49A, B).
Chung 1993: 230. f. 144. Komárek and Anagnostidis 2005: 625. f. 954. Hu and Wei 2006: 135. pl. II-26-6.
Filaments free-floating, straight or curved, solitary, 20.0-24.0 μm wide. Sheaths thin and colorless,opened at the ends, containing a single motile trichome. Trichomes isopolar, straight or slightlywaved, composed of shortly cylindrical cells, unconstricted at the cross-walls, not attenuated at theends. Cells short, always shorter than wide, 2.0-2.5 μm long, sometimes with aerotopes, finelygranulated and more at the cross-walls. Apical cells widely rounded, with thickened outer cell wall.Heterocytes and akinetes absent. Cell division crosswise, perpendicular to the long axis of a tri-chome. Reproduction by more or less short, motile hormogonia by the help of necridia.
TYPE LOCALITY: Various syntype localities in Wisconsin, U.S.A.DISTRIBUTION: Perhaps cosmopolitan.KOREA: Han River (Chung et al. 1968); JB (Chung 1976); JN (Wui and Kim 1987); JJ (Lee et al. 1990);
Galgok stream and Incheon river in JN (Lee et al. 2008).
Cyanophyceae: Oscillatoriales: Oscillatoriaceae 91
Fig. 49. A, B. Lyngbya birgei.
A B
25 μm

SPECIMEN EXAMINED: Nammaeji in GB (35°49′44′′N, 128°44′32′′E), 27.ii.2012.ECOLOGY: Freshwater, planktonic in lakes.REMARKS: This species is planktonic.
46. Lyngbya hieronymusii Lemmermann 1905: 146-147 (Fig. 50A-C).
Chung 1993: 231. f. 146. Komárek and Anagnostidis 2005: 614. f. 934. Hu and Wei 2006: 134. pl. II-26-2.
Filaments free-floating, microscopic, solitary, straight or slightly curved, 12.0-15.3 μm. Sheath wideand firm, homogeneous, colorless, outer rough, uneven, opened at the ends, containing a singlemotile trichome. Trichomes with shortly cylindrical cells, isopolar, straight, blue-green, 11.0-14.0μm wide, unconstricted at the granulated cross-walls, with aerotopes, not attenuated at the ends.Cells 2.5-4.0 μm long, always shorter than wide. Apical cells widely-rounded, without calyptra or
92 Algal Flora of Korea∙Freshwater Cyanoprokaryota II
Fig. 50. A-C. Lyngbya hieronymusii.
25 μm
A B C

thickened outer cell wall. Heterocytes and akinetes absent. Cell division crosswise, perpendicularto the long axis of a trichome. Reproduction by more or less short, motile hormogonia by the helpof necridia.
TYPE LOCALITY: Near Potsdam, Brandenburg, Germany (Silva et al. 1996: 37).DISTRIBUTION: Perhaps cosmopolitan.KOREA: JN (Wui and Kim 1987).SPECIMEN EXAMINED: Nammaeji in GB (35°49′44′′N, 128°44′32′′E), 30.xiii.2010.ECOLOGY: Freshwater, typical planktonic species, occurring in stagnant waters.REMARKS: The filament of Lyngbya hieronymusii var. crassi-vaginata in tropical and subtropical up
to 20 μm wide (Komárek and Anagnostidis 2005). The reports from other than planktonic habitatsare problematic (Komárek and Anagnostidis 2005).
47. Lyngbya major Meneghini ex Gomont 1892: 144 (Fig. 51A-C).
Chung 1993: 232. f. 149. Komárek and Anagnostidis 2005: 619. f. 941. McGregor 2007: 73. f. 14B. pl.10N, O. Whitton 2011: 91. pl. 19P.
SYNONYM: Lyngbya major Meneghini 1837.
Filaments benthic, among other algae, or clustered into cespitose thallus, or forming membranous,expanded, black-green layers, long, straight, or sometimes slightly curved, entangled, 22.0-26.0 μmwide. Sheaths thick, to 3.7 μm wide, usually lamellated, wrinkled, rough from outside, colorless,opened at the ends, containing a single motile trichome. Trichomes isopolar, straight or slightlycurved, dark blue-green. 11.0-17.0 μm wide, not or slightly constricted at the granulated cross-walls, sometimes slightly attenuated at the ends. Cells discoid, one-eighth to one-quarter times aslong as wide, (1.5-) 2.0-5.0 μm long, without aerotopes, often irregularly spaced granules. Apicalcells rounded, with slightly thickened outer cell wall. Heterocytes and akinetes absent. Cell divisioncrosswise, perpendicular to the long axis of a trichome. Reproduction by more or less short, motilehormogonia by the help of necridia.
TYPE LOCALITY: Italy.DISTRIBUTION: Perhaps cosmopolitan.KOREA: JB (Chung 1976); JN (Wui and Kim 1987); Pyeongtaek reservoir around Asan Bay (Shin
2003); Yeoju in GG (Lim and Lee 2008a, 2008b).SPECIMEN EXAMINED: Nakdong river at Ohori in GN (35°23′46′′N, 128°31′20′′E) 29.ix.2010.ECOLOGY: Benthic on mud, water plants and other submerged objects, in stagnant fresh (mostly
hard) and saline water, sometimes tychoplanktonic (liberating solitary filaments).REMARKS: Records from thermal springs are uncertain (Komárek and Anagnostidis 2005).
Cyanophyceae: Oscillatoriales: Oscillatoriaceae 93

48. Lyngbya martensiana Meneghini ex Gomont 1892: 145 (Fig. 52A-D).
Chung 1993: 232. f. 151. Komárek and Anagnostidis 2005: 612. f. 931. Hu and Wei 2006: 124. pl. II-24-4. McGregor 2007: 73. f. 14C. pl. 10P. Whitton 2011: 92. pl. 19G.
SYNONYM: Porphyrosiphon martensianus (Meneghini ex Gomont) Anagnostidis et Komárek 1988.Leibleinia martensiana Meneghini 1837.Leibleinia martensiana (Meneghini) Kützing 1847, nom. inval.
Benthic and metaphytic. Thallus caespitose, pale blue-green, violet when dried, composed ofentangled filamentous, usually arranged in more or less parallel fascicles, occasionally solitary andfree-floating. Filaments long, variously curved or flexuous, sometimes straight, 6.0-12.0 (-15.0) μm
94 Algal Flora of Korea∙Freshwater Cyanoprokaryota II
Fig. 51. A-C. Lyngbya major.
25 μm
A B C
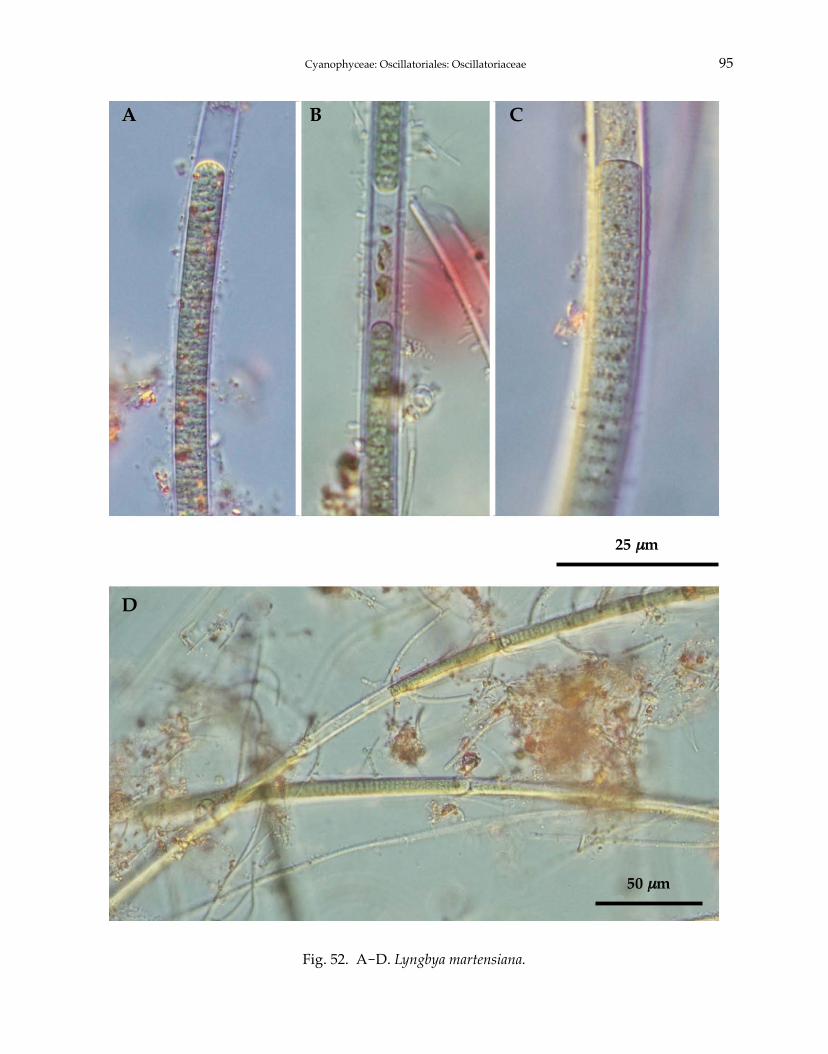
Cyanophyceae: Oscillatoriales: Oscillatoriaceae 95
Fig. 52. A-D. Lyngbya martensiana.
25 μm
50 μm
A B C
D

wide. Sheaths hyaline, colorless, thick, opened at the ends, containing a single motile trichome.Trichomes isopolar, cylindrical, pale blue-green to olive-green, 5.6-10.0 (-12.0) μm wide, not con-stricted at the cross-walls, not attenuated towards ends. Cells 0.2-0.5 times as long as wide, (1.2-)1.5-3.5 (-4.2) μm long, homogeneous, later granular, often granulated at cross-walls. Apical cellsbroadly-rounded, hemispherical, not capitate, without calyptra or thickened outer cell wall. Hetero-cytes and akinetes absent. Cell division crosswise, perpendicular to the long axis of a trichome.Reproduction by more or less short, motile hormogonia by the help of necridia.
TYPE LOCALITY: Probably Italy.DISTRIBUTION: Perhaps cosmopolitan.KOREA: Jeju island (Lee et al. 1990).SPECIMEN EXAMINED: Nam river at Jeongamli in GN (35°23′00′′N, 128°25′21′′E), 01.viii.2011;
Duyangli in GN (35°04′49′′N, 127°53′30′′E), 02.xii.2011.ECOLOGY: Freshwater, metaphytic and periphytic in stagnant and slow-flowing waters, among
littoral vegetation and woody debris.REMARKS: Perhaps cosmopolitan but not common, reports from soil, marine and thermal habitats
probably concern other species (Komárek and Anagnostidis 2005). Morphologically, but no usuallyecologically, L. martensiana resembles Lyngbya confervoides C. Agardh apart from its narrower width(6.0-10.0 μm) (Whitton 2011). Reference record from Hu and Wei (2006) is as Porphyrosiphon marten-sianus (Meneghini ex Gomont) Anagnostidis et Komárek 1988.
49. Lyngbya nigra C. Agardh ex Gomont 1892: 145 (Figs. 53A, B, 54A-C).
Komárek and Anagnostidis 2005: 614. f. 933. Hu and Wei 2006: 134. pl. II-26-4. McGregor 2007: 73.f. 13E. pl. 11A-C. Whitton 2011: 92.
SYNONYM: Lyngbya nigra C. Agardh 1824, nom. inval.
Benthic. Thallus flattened mat, tufty, extended, dark blue-green. Filaments long, generally straightor slightly flexuous, 10.0-14.5 μm wide, clustered. Sheaths thin, colorless, not lamellated, openedat the ends, containing a single motile trichome. Trichomes isopolar, straight, dark green, (6.0-)8.0-11.0 (-14.0) μm wide, not constricted at the ungranulated cross-walls, not or somewhat atten-uated at the ends. Cells 0.2-0.5 times as long as wide, 2.0-4.0 μm long, without aerotopes, withslightly granular content. Apical cells obtuse-rounded, slightly attenuated with rounded-conicalcalyptra. Heterocytes and akinetes absent. Cell division crosswise, perpendicular to the long axisof a trichome. Reproduction by more or less short, motile hormogonia by the help of necridia.
TYPE LOCALITY: Scåne, Sweden (type of Oscillatoria nigra C. Agardh).DISTRIBUTION: Perhaps worldwide.KOREA: JB (Chung 1999).SPECIMEN EXAMINED: Suyeongang in Nakdong River (35°09′56′′N, 129°07′34′′E), 09.ix.2010.ECOLOGY: Freshwater, stagnant and slow-flowing waters, periphytic, among other cyanoprokar-
yotes and aquatic marcophytes, rarely forming single species stands. Usually on alkaline substra-tum with a pH of about 8.
96 Algal Flora of Korea∙Freshwater Cyanoprokaryota II

Cyanophyceae: Oscillatoriales: Oscillatoriaceae 97
Fig. 53. A, B. Lyngbya nigra.
50 μm
100 μm
A
B

REMARKS: The recorded from thermal springs but this is questionably the same species (Komárekand Anagnostidis 2005). This species overlaps L. major and L. martensiana in many characteristics,but neither of latter two ever develop a typical calyptra (Whitton 2011).
50. Lyngbya stagnina Kützing ex Gomont 1892 (Figs. 55A, B, 56A-C).
Komárek and Anagnostidis 2005: 616. f. 936.
Benthic. Thallus dark blue-green, with clustered filaments. Filaments curved, 11.0-16.0 μm wide.Sheaths colorless or slightly yellowish to yellow-brown, lamellated, opened at the ends, containinga single motile trichome. Trichomes isopolar, flexuous, (8.0-) 9.5-12.0 (-13.0) μm wide, not con-
98 Algal Flora of Korea∙Freshwater Cyanoprokaryota II
Fig. 54. A-C. Lyngbya nigra.
25 μm
A B C

stricted at the distinctly granulated cross-walls, not attenuated at the ends. Cells short, shorter thanwide, 2.0-4.0 μm long, granulated near coss-walls. Apical cells widely-rounded, not narrowed,without thickened outer cell wall. Heterocytes and akinetes absent. Cell division crosswise, per-pendicular to the long axis of a trichome. Reproduction by more or less short, motile hormogoniaby the help of necridia.
TYPE LOCALITY: Probably Germany, but no locality cited by Kützing (1943).DISTRIBUTION: Perhaps cosmopolitan.KOREA: Previously unrecorded species.SPECIMEN EXAMINED: Woansa stream at Jakpalli in GN (35°06′51′′N, 127°59′05′′E), 01.viii.2011.ECOLOGY: Originally described from cold freshwaters in Europe. But reports also from mineral
and saline waters, also from tropical regions (Komárek and Anagnostidis 2005).REMARKS: Occurs in different concepts compare with Phormidum stagninum (Kützing ex Gomont)
Anagnostidis 2001.
Cyanophyceae: Oscillatoriales: Oscillatoriaceae 99
Fig. 55. A, B. Lyngbya stagnina.
50 μm
A
B

100 Algal Flora of Korea∙Freshwater Cyanoprokaryota II
Fig. 56. A-C. Lyngbya stagnina.
25 μm
A B C

Literature Cited
Anagnostidis, K., 2001. Nomenclatural changes in cyanoprokaryotic order Oscillatoriales. Preslia, Praha 73:359-375.
Anagnostidis, K. and J. Komárek, 1985. Modern approach to the classification system of cyanophytes, 1-Intro-duction. Archiv für Hydrobiologie, Algological Studies 38/39: 291-302.
Anagnostidis, K. and J. Komárek, 1988. Modern approach to the classification system of cyanophytes, 3-Oscilla-toriales. Archiv für Hydrobiologie, Algological Studies 50-53: 327-472.
Anagnostidis, K. and J. Komárek, 1990. Modern approach to the classification system of cyanophytes, 5-Stigo-nematales. Archiv für Hydrobiologie, Algological Studies 59: 1-73.
Berkeley, M.J., 1832. Gleanings of British algae; being an appendix to the supplement to English Botany. pls5-8. London: C.E. Sowerby.
Bostock, P.D. and A.E. Holland, (Eds.), 2010. Census of the Queensland Flora 2010. Queensland Herbarium,Department of Environment and Resource Management, Brisbane.
Canter-lund, H. and J.W.G. Lund, 1995. Freshwater algae: Their microscopic world explored. Biopress Ltd.,Bristol. 360 pp.
Castenholz, R.W., 2001. Phylum BX. Cyanobacteria. In: D.R. Boone and R.W. Castenholz, (Eds.), Bergey’smanual of systematic bacteriology, 2nd edition, volume 1. Springer-Verlag, NewYork. pp. 473-599.
Chang, Y.K. and S.L. Jeon, 1996. A study on the phytoplankton in the Paldang dam reservoir II. The changesof phytoplankton species composition. The Korean Journal of Phycology 11: 217-229.
Cho, K.J., I.K. Chung and J.A. Lee, 1993. Seasonal dynamics of phytoplankton community in the Naktong riverestuary, Korea. The Korean Journal of Phycology 8: 15-28.
Chung, J., 1976. A study on the cyanophyceae from Chun Ra Book Do area. Korean Journal of Botany 19: 19-30.
Chung, J., 1993. Illustration of the freshwater algae of Korea. Academy Publishing Company, Seoul. 496 pp.
Chung, Y.H., E.S. Kay and D.W. Park, 1968. A study on the microflora of the Han River (II). Journal of plantbiology 11: 41-70.
Chung, Y.H. and K. Lee, 1979. On the phytoplanktons of the Geum river near Gongju. Korean Journal ofBotany 22: 141-145.
Cronberg, G. and J. Komárek, 1994. Planktic cyanoprokaryotes found in south Swedish lakes during theXIIth International Symposium of Cyanophyte Research, 1992. Archiv für Hydrobiologie, AlgologicalStudies 75: 323-352.
Do Carmo Bittencourt-Oliveira, M., A. do Nascimento Moura, M.C. de Oliveira and N.S. Massola Jr., 2009.Geitlerinema species (Oscillatoriales, Cyanobacteria) revealed by cellular morphology, ultrastructureand DNA sequencing. Journal of Phycology 45: 716-725.
Geitler, L., 1932. Cyanophyceae. In: L. Rabenhorst, (Eds.), Kryptogammenflora von Deutschland, Osterreich,under de Sweitz. Leipzig: Akademische Verlagsgesellschaft. 14: 673-1056.
Gomont, M., 1892. Monographie des Oscillariées (Nostocacées homocystées). Annales des Sciences Naturelles,Botanique, Series 7 16: 91-264.
Greuter, W., J. McNeill, F.R. Barrie, H.M. Burdet, V. Demoulin, T.S. Filgueiras, D.H. Nicolson, P.C. Silva, J.E.Skog, P. Trehane, N.J. Turland and D.L. Hawksworth, 2000. International code of botanical nomencla-ture (St Louis code). Regnum Vegetabile 138. Koeltz Scientific Books, Königstein. 474 pp.
Guiry, M.D. and G.M. Guiry, 2012. AlgaeBase. World-wide electronic publication, National University ofIreland, Galway. http://www.algaebase.org; searched on December 2012.
101

Hindák, F., 2008. Colour Atlas of Cyanophytes. VEDA, Vydavatelel’stvo Slovenskej Akadémie Vied, Braislava.253 pp.
Hindák, F. and M.T. Moustaka, 1988. Planktic cyanophyte of Lake Vlovi, Greece. Archiv für Hydrobiologie,Algological Studies 50-53: 497-528.
Hoffmann, L., 1988. Criteria for the classification of blue-green algae (cyanobacteria) at the genus and at thespecies level. Archiv für Hydrobiologie, Algological Studies 50-53: 131-139.
Hu, H. and Y. Wei, 2006. The freshwater algae of China, systematics, taxonomy and ecology. Science Press,China. 1023 pp.
John, D.M., B.A. Whitton and A.J. Brook. 2011. The freshwater alga flora of the British Isles: An identificationguide to freshwater and terrestrial algae. Cambridge University Press, New York. 878 pp.
Kim, H.K. and Y.H. Kim, 1991. Marine algal communities around three nuclear power plants in Korea. TheKorean Journal of Phycology 6: 157-192.
Kim, J.H., Y.H. Kim and I.K. Lee, 1998. Dynamics of phytoplankton community in lake Chungju. Algae 13:339-354.
Kim, J.H., S.J. Lee and H.M. Oh, 2000. Dynamics of epilithic algal community in the Geum River, Korea. Algae15: 287-297.
Kim, J.I., J.H. Lim, J.W. Lee and H.B. Lee, 2002. Phylogenetic relationship of microcystics (cyanophceae) basedon partial 16S rRNA gene sequences in Korea. Algae 17: 153-159.
Kim, M.K., 2008. Patterns in phytoplankton community structure in the Beopsu marsh, Haman-gun, Gyeong-sangnam-do, Korea. Algae 23: 219-230.
Kim, M.K., S.H. Lee and C.H. Lee, 2007. Change of dominant species of phytoplanktons and hydrologicalcauses of water bloom in the Lake Unmun, Cheongdo-gun, Gyeonsangbuk-do. Algae 22: 261-271.
Kim, Y.H. and J.K. Ahn, 2005. Ecological characteristics of marine algal communities at the discharge canalsof three nuclear power plants on the east coast of Korea. Algae 20: 217-224.
Kim, Y.H. and J.K. Ahn, 2006. Effects of heated effluents on the intertidal macroalgal community nearWolseong, the east coast of Korea. Algae 21: 453-461.
Kim, Y.H., J.K. Ahn, J.I. Lee and H.M. Eum, 2004. Effects of heated effluents on the intertidal macroalgalcommunity near Uljin, the east coast of Korea. Algae 19: 257-270.
Kim, Y.H., H.M. Eum and Y.S. Kang, 1998. Qualitative and quantitative analysis of warm tolerant benthicmarine algae in Korea I. Kori nuclear power plant. Algae 13: 213-226.
Kim, Y.H., H.J. Yoon and J.S. Yoo, 1995. Species composition and biomass of marine algal community on themid-western coast of Korea. Journal of Plant Biology 38: 389-398.
Ko, S.R., C.Y. Ahn, Y.K. Lee and H.M. Oh, 2011. Dynamics of bacterial communities analyzed by DGGEduring cyanobacterial bloom in Daechung reservoir, Korea. Korean Journal of Environmetal Biology. 29:225-235.
Komárek, J., 1991. A review of water-bloom formning Microcystis species, with regard to populations fromJapan. Archiv für Hydrobiologie, Algological Studies 64: 115-127.
Komárek, J., 2003. Coccoid and colonial cyanobacteria. In: J.D. Wehr and R.G. Sheath, (Eds.), Freshwateralgae of North America, ecology and classification. Academic Press, Amsterdam Boston London NewYork Oxford Paris San Diego San Fransisco Singapore Sydney Tokyo. pp. 59-116.
Komárek, J. and K. Anagnostidis, 1989. Modern approach to the classification system of Cyanophytes. 4.Nostocales. Archiv für Hydrobiologie, Algological Studies 56: 247-345.
Komárek, J. and K. Anagnostidis, 1999. Cyanoprokaryota 1. Teil: Chroococcales. In: H. Ettl, G. Gärtner, H.Heynig and D. Mollenhauer, (Eds.), Süβwasserflora von Mitteleuropa, 19/1. Gustav Fischer, Jena-Stuttgart-Lübeck-Ulm. 548 pp.
102 Algal Flora of Korea∙Freshwater Cyanoprokaryota II

Komárek, J. and K. Anagnostidis, 2005. Cyanoprokaryota 2. Teil/ 2nd Part: Oscillatoriales. In: B. Büdel, L.Krienitz, G. Gärtner and M. Schagerl, (Eds.), Süβwasserflora von Mitteleuropa 19/2, Elsevier/Spektrum,Heidelberg. 759 pp.
Komárek, J., H. Kling and J. Komárková, 2003. Filamentous Cyanobacteria. In: J.D. Wehr and R.G. Sheath,(Eds.), Freshwater algae of North America, ecology and classification. Academic Press, AmsterdamBoston London New York Oxford Paris San Diego San Fransisco Singapore Sydney Tokyo. pp. 177-196.
Komárek, J. and J. Komárková, 2002. Review of the European Microcystis-morphospecies (Cyanoprokaryotes)from nature. Czech Phycology 2: 1-24.
Komárek, J. and J. Komárková, 2004. Taxonomic review of the cyanoprokaryotic genera Planktothrix andPlanktothricoides. Czech Phycology 4: 1-18.
Lee, H.B and I.K. Lee, 1981. Flora of benthic marine algae in Gyenggi bay, western coast of Korea. KoreanJournal Botany 24: 107-138.
Lee, J.A., A.R. Choi and M. Watanabe, 1997. Taxonomic implications of the genus Mirocystis (Cyanophyceae)from the Naktong River. The Korean Journal of Phycology 12: 167-176.
Lee, J.B., Y.B. Go and J.H. Choa, 1990. The structure and dynamics of phytoplankton communities aroundthe coastline of Cheju island, Korea. The Korean Journal of Phycology 5: 159-171.
Lee, K and S.K. Yoon, 2003. Ecological studies the Togyo reservoir in Chulwon, Korea VI. The list of phyto-plankton and periphyton. Algae 18: 263-272.
Lee, O.M., M.S. Yoo, B.I. Lee and A.S. Lim, 2008. The distribution and standing crop of phytoplankton at theestuaries of Galgok stream and Incheon river in Jeollanam-do. Algae 23: 257-268.
Lim, A.S. and O.M. Lee, 2008a. Distibution of aerial algae and biological classes in five stone cultural proper-ties of Korea. Algae 23: 63-69.
Lim, A.S. and O.M. Lee, 2008b. The distribution of aerial algae and the evaluation of algal inhabitation onfive stone cultural properties in Gyeonggi-do. Algae 23: 269-276.
Lukavsky, J., 1994. Algal flora of lakes in the high taxa mountains (Slovakia). Hydrobiologia 274: 65-74.
McGregor, G., 2007. Freshwater cyanoprokaryota of North-Eastern Australia 1: Oscillatoriales. AustraliaBiological Resources Study, Canberra. 123 pp.
Nägeli, C., 1849. Gattungen einzelliger Algen, physiologisch und systematisch bearbeitet. Neue Denkschriftender Allg. Schweizerischen Gesellschaft für die Gesammten Naturwissenschaften 10(7): i-viii, 139 pp.
Oren, A. and B.J. Tindall, 2005. Nomenclature of the cyanophyta/ cyanobacteria/ cyanoprokaryotes underthe International Code of Nomenclature of Prokaryotes. Archiv für Hydrobiologie, Algological Studies117: 39-52.
Oren, A., J. Komárek and L. Hoffmann, 2009. Nomenclature of the cyanophyta/ cyanobacteria/ cyanopro-karyotes - What has happened since IAC Luxembourg? Archiv für Hydrobiologie, Algological Studies 130,17-26.
Park, J.G. and J.J. Lee, 2005. Community dynamics of phytoplankton in lake Daecheong. Algae 30: 197-205.
Park, J.H., B.R. Moon and O.M. Lee, 2006. The phytoplankton compositions and trophic states at several lakesof Suwon-si, Korea. Algae 21: 217-228.
Park, Y.S. and Y.H. Kim, 1990. Phtogeographical study on the summer marine algal distribution in westerncoast in Korea. The Korean Journal of Phycology 5: 39-50.
Perkerson III, R.B., J.R. Johansen, L. Kovácik, J. Brand, J. Kastovsky and D.A. Casamatta, 2011. A unique pse-udanabaenalean (cyanobacteria) genus Nodosilinea gen. nov. based on morphological and moleculardata. Journal of Phycology 47: 1397-1412.
Rajaniemi-Wacklin, P., A. Rantala, M.A. Mugnai, S. Turicchia, S. Ventura, J. Komárková, L. Lepisto and K.Sivonen, 2005. Correspondence between phylogeny and morphology of Snowella spp. and Woronichinia
Literature Cited 103

naegeliana, cyanobacteria commonly occurring in lakes. Journal of Phycology 42: 226-232.
Romo, S., 1993. Geitlerinema amphibium (C. Agardh ex Gomont) Anagnostidis (Cyanophyceae): morphology,ultrasutructure and ecology. Archiv für Hydrobiologie, Algological Studies 69: 11-27.
Sant’anna, C.L., M.T.P. Azevedo, P.A.C. Senna, J. Komárek and J. Komárková, 2004. Planktic cyanobacteriafrom São Paulo State, Brazil: Chroococcales. Revista Brasileira de Botânica 27: 213-227.
Shin, J.K., 2003. Seasonal dynamics of aquatic environment and phytoplankton in Pyeongtaek reservoir, Korea.Algae 18: 145-156.
Shin, J.K. and K.J. Cho, 1997. Distribution and population dynamics of Mycrosystis (cyanophyta) in the Nack-dong river. The Korean Journal of Phycology 12: 283-290.
Silva, P.C., 1980. Names of classes and families of living algae: with special reference to their use in the IndexNominum Genericorum (Plantarum). Regnum Vegetabile 103: 156 pp.
Silva, P.C., P.W. Basson and R.L. Moe, 1996. Catalogue of the benthic marine algae of the Indian Ocean. Uni-versity of California Publications in Botany 79: 1259 pp.
Skuja, H., 1939. Beitrag zur Algenflora Lettlands II. Acta Horti Botanici Universitatis Latviensis 11/12: 41-169.
Song, C.B., 1986. An ecological study of the intertidal macroalgae on Kwangyang bay, southern coast ofKorea. The Korean Journal of Phycology 1: 203-223.
Suda, S., M. Watanabe, S. Otsuka, A. Mahakahant, W. Yongmanitchai, N. Nopartnaraporn, Y. Liu and J.G.Day, 2002. Taxonomic revision of water-bloom forming species of osillatorioid cyanobacteria. Interna-tional Journal of Systematic and Evolutionary Microbiology 52: 1577-1595.
Wacklin, P., 2006. Biodiversity and phylogeny of planktic cyanobacteria in temperate freshwater lakes. EditaPrima Oy, Helsinki. 69 pp.
Whitton, B.A., 2011. Phylum Cyanobacteria (Cyanophyta). In: D.M. John, B.A. Whitton and A.J. Brook, (Eds.),The freshwater algal flora of the British Isles. An identification guide to freshwater and terrestrial algae.Cambridge University Press, New York. pp. 31-158.
Willame, R., C. Boutte, S. Grubisic, A. Wilmotte, J. Komarek and L. Hoffmann, 2006. Morphological andmolecular characterization of planktonic cyanobacteria from Belgium and Luxembourg. Journal ofPhycology 42: 1312-1332.
Wui, I.S. and B.H. Kim, 1987. Flora of the fresh-water algae in Chol La Nam Do, Korea (II) cyanophyceae. TheKorean Journal of Phycology 2: 193-201.
Yamaghshi, T. and M. Akiyama, 1984. Photomicrographs of the fresh-water algae volume 1. Uchida RokakuhoPublishing Co., Ltd. Tokyo. pp. 1:1-1:100.
Yamaghshi, T. and M. Akiyama, 1985. Photomicrographs of the fresh-water algae volume 3. Uchida RokakuhoPublishing Co., Ltd. Tokyo. pp. 3:1-3:100.
Yoo, J.S., Y.H. Kim and I.K. Lee, 1996. Dynamice of the marine macroalgal community in Inchon dock eco-system, western coast of Korea. The Korean Journal of Phycology 11: 295-307.
104 Algal Flora of Korea∙Freshwater Cyanoprokaryota II

A
Ammatoideoideae 47, 81Anabaena 6Anacystis
dimidiata 21incerta 29
Aphanocapsa 6, 24conferta 25, 26, 27delicatissima 25elachista 25, 26, 27elachista var. conferta 25elachista var. planctonica 26grevillei 25, 26, 27, 28holsatica 25incerta 25, 29, 30parietina 24planctonica 26, 27
Aphanothece 6Aphnizomenon 6Arthrospira 47
brevis 63neapolitana 63
Asterocapsa 9
B
Blennothrix 89
C
Calothrixbalearica 82caespitosa 86
Chroococcaceae 9Chroococcus 9, 10
dimidiatus 21dispersus 10, 18dispersus var. minor 12distance 10
limneticus 10, 11limneticus var. carneus 11microscopicu 10minimus 10, 12, 13minor 10, 13, 14minutus 10, 15, 16minutus var. minimus 12obeliteratus 10, 16, 17planctonicus 10, 17, 18prescottii 10, 18, 19rufescens 10subnudus 10, 20thermalis f. major 11turgidus 10, 17, 20, 21, 22turgidus var. subnudus 20vacuolatus 10, 22, 23virescens 15
Coccochlorisgrevellei 27
Coelosphaerium 24Cyanosarcina 9Cyanotetras 6, 24
D
Dichothrixnordstedtii 86
E
Eucapsis 6, 20
G
Geitlerinema 31Gloecocapsa 6
minima 12minima f. smithii 12minuta 15
105
Index to Scientific Names

turgida 21turgida f. subnuda 20vacuolata 22
Gloeocapsopsis 9Gomontinema 53
H
Hansgirgia 53Heteroleibleinia 31, 42
fontana 37kossinskajae 42, 43, 44rigidula 42, 45, 46thornensis 42
Heteroleibleiniodeae 31, 42Homoeothrix 81
balearica 82, 84caespitosa 87fusca 84, 86fusca f. elongata 84fusca f. longissima 84fusca f. minor 84nordstedtii 87nordstedtii f. salisburgensis 87
Homoeotrichoideae 89Hygrocrocis
rigidula 45Hypheothrix
fontana 37inundata 70rigidula 45
J
Jaaginema 31, 32minimum 32, 33profundum 32, 34, 35subtilissimum 32
L
Leibleinia 31, 45martensiana 94
Leptolyngbya 31, 36boryana 36fontana 37, 38foveloara 37lagerheimii 37, 39, 40subtilis 37, 41
Leptolyngbyoideae 31, 36Leptothrix
fontana 37rigidula 45
Leptotrichiarigidula 45
Limnococcus 9, 10Limnothrix 6, 31Lyngbya 6, 36, 89, 90
aerugineo-caerulea 56, 57amoena var. chlorina 63birgei 90, 91brevis 62chlorina 63compresssa 51confervoides 90, 96corium 66fontana 37hieronymusii 90, 92hieronymusii var. crassi-vaginata 93inundata 70kossinskajae 43kossinskajae var. gracilis 45lagerheimii 39lagerheimii f. minor 39major 90, 93, 94, 98martensiana 90, 94, 95, 96, 98membranacea 61nigra 71, 90, 96, 97, 98okenii 72phormidium 77rigidula 45stagnina 90, 98, 99, 100subtilis 41taylori 75tenerrima var. rigidula 45tenuis var. aerugineo-caerulea 56tenuis var. neapolitana 63violacea 79vulgaris 61
106 Algal Flora of Korea∙Freshwater Cyanoprokaryota II

M
Merismopedia 6, 24Merismopediaceae 23Microcoleoideae 47Microcoleus 47
autumnalis 61corium 65
Microcystaceae 6Microcystis 6
elachista 26grevillei 27incerta 29pulverea f. conferta 25pulverea f. elachista 26pulverea f. incerta 29pulverea var. incerta 29smithii 29
N
Nostocaceae 6
O
Oscillatoria 6, 89aerugineo-caerulea 56, 57agardhii 49, 51agardhii f. aequicrassa 49agardhii f. gomontii 49agardhii f. lemmermannii 49agardhii f. moebiusii 49agardhii f. wislouchii 49agardhii var. isothrix 52amoena 58, 59articulata 59, 60articulata var. africana 59articulata var. circinata 59autumnalis 61brevis 62, 63brevis var. neapolitana 62caerulescesns 33chlorina 63, 64, 65chlorina var. perchlorina 64
compresssa 51corium 65favosa 67granulata 68, 70grunowiana var. articulata 59inundata 70limosa var. uncinata 61minima 32mougeotii 52neapolitana 62nigra 71, 72nigroviridis f. crassior 66okenii 72, 73profunda 34, 36tenuis f. tergestina 77tenuis var. tergestina 77, 79terebriformis 75, 76terebriformis f. brevis 63tergestina 77, 79violacea 79, 81vulgaris 61
Oscillatoriaceae 6, 81, 89Oscillatorioideae 89
P
Palmellagrevellei 27
Phormidiaceae 6, 47Phormidiochaete 47, 81, 89
balearica 81, 82, 83fusca 81, 84, 85, 86nordstedtii 81, 86, 87, 88
Phormidioideae 47Phormidium 36, 47, 48, 53
aerugineo-caeruleum 55, 56, 57amoenum 55, 58, 59articulatum 55, 59, 60autumnale 55, 61breve 54, 62chaybeum 54chlorinum 55, 56, 60, 63, 64corium 55, 65cortianum 73crouani 67
Index to Scientific Names 107

crouaniorum 54, 66, 67favosum 55, 67formosum 54granulatum 55, 65, 68, 69hamelii 55incrustatum 54inundatum 55, 67, 70irriguum 55lucidum 53membranaceum 61nigrum 55, 70, 71okenii 54, 72, 73puteale 55stagninum 55, 99subsalsum 61taylori 55, 74terebriforme 54, 75, 76tergestinum 55, 70, 71, 77, 78uncinatum 55, 61violaceum 55, 79, 80vulgare 61
Planktolyngbya 6, 31Planktothrix 47, 48
agardhii 48, 49, 50compresssa 48, 50, 51cryptovaginata 48isothrix 48, 51, 52mougeotii 51, 52
Plectonema 36, 89Pleurococcus
minor 13Polycystis
incerta 29Porphyrosiphon
martensianus 94, 96Proterendothrix 47Protococcus
minor 13
minutus 15turgidus 21
Protolyngbya 36Pseudanabaena 31Pseudanabaenaceae 6, 31Pseudanabaenoideae 31, 32Pseudophormidium 47, 48
S
Snowlla 6, 24Spirocoleus
lagerheimii 39Spirulina 31Spirulinoideae 31Symploca 47
phormidium 79Synechococcaceae 6Synechococcus 6Synechocystis 6, 24
T
Tapinothrix 31Trichodesmium 47Trichophorus
nigra 71Trochiscia
dimidiata 21Tychonema 47
granulatum 68
W
Woronichinia 6, 24
108 Algal Flora of Korea∙Freshwater Cyanoprokaryota II

China
PB
JG
YG
HB
HN
PN
HWB
GW
GG
CB
CNGB
GB(Ulleung-do)
GN
JN
JJ
JB
HWN
Russia
East Sea
Yellow Sea
South Sea
CB Chungcheongbuk-doCN Chungcheongnam-doGB Gyeongsangbuk-doGG Gyeonggi-doGN Gyeongsangnam-doGW Gangwon-doHB Hamgyeongbuk-doHN Hamgyeongnam-doHWB Hwanghaebuk-doHWN Hwanghaenam-doJB Jeollabuk-doJG Jagang-doJJ Jeju-doJN Jeollanam-doPB Pyeonganbuk-doPN Pyeongannam-doYG Yanggang-do

National Institute of Biological ResourcesMinistry of Environment
ISBN 978-89-97462-58-2
Algal Flora of Korea
Vol. 5, No. 2
Freshwater Cyanoprokaryota
IIN
IBR
Algal Flora of Korea
Flora and Fauna of Korea
National Institute of Biological ResourcesMinistry of Environment
Volume 5, Number 2Cyanophyta: Cyanophyceae: Chroococcales, Oscillatoriales
Freshwater Cyanoprokaryota II