action spectra for the generation of singlet oxygen from mitochondrial membranes from soybean...
TRANSCRIPT

Photochembtry and Photobiology Vol. 52, No. 3, pp. 561-566, 1990 Printed in Great Britain. All rights reserved
003 1-8655190 $03.00 +0.00 Copyright 0 1990 Pergamon Press plc
ACTION SPECTRA FOR THE GENERATION OF SINGLET OXYGEN FROM MITOCHONDRIAL MEMBRANES FROM
SOYBEAN (Glycine max) HYPOCOTYLS JIN JUNG,* HEE-JUNG KIM and MOONJAE CHO
Department of Agricultural Chemistry, Seoul National University, Suwon 440-744, Korea
(Received 28 August 1989; nccepted 8 February 1990)
Abstract-The action spectrum for the generation of singlet oxygen ('0,) from mitochondrial mem- branes under aerobic conditions was measured at wavelengths between 360 and 600 nm, using sub- mitochondrial particles (SMP) prepared from soybean hypocotyls. The spectrum, showing a peak at about 420 nm, remarkably resembles the absorption spectra of the Fe-S centers of nonheme iron proteins. Disruption of the Fe-S centers by treating SMP with mersalyl acid resulted in a substantial decrease in the efficiency of lo2 generation, leaving an action spectrum whose pattern is significantly similar to the absorption spectrum of flavins, at least in the region of near UV and blue light wavelengths. Estimating the contribution of the Fe-S centers to the generation of lo, from SMP, we suggest that the Fe-S centers act as very important endogenous photosensitizers in plant cells, in so far as the type I1 mechanism is concerned. Possible involvement of mitochondrial flavoproteins in the generation of '02 is also discussed.
INTRODUCTION
The exposure to near UV and visible light of mito- chondria can result in loss of respiration that is 0,- dependent and prevented by anaerobiosis (Epel, 1973; Aggarwal et a l . , 1976, 1978). The biological changes in living organisms induced by near UV and visible light in the presence of molecular oxygen and photosensitizers are believed to be the result of photodynamic reactions (Spikes, 1989; Elstner, 1982). Since mitochondrial membranes contain a substantial number of natural internal chromo- phores whose absorption maxima arise in the wave- length region from 350 to 600 nm, it can be said that the light-induced inhibition of mitochondrial respiratory activity may involve some of these chromophores acting as endogenous photo- sensitizers.
Photodynamic reactions proceed via either type I or type I1 mechanisms, according to whether they involve radical species such as superoxide (03 or singlet oxygen ('Ag 0,) as reactive intermediates (Spikes, 1989). Thus, it is natural to expect that irradiation of mitochondria with near UV and vis- ible light under aerobic conditions gives rise to the generation of at least one of these active oxygen species. Since singlet oxygen pathways are very common in photodynamic reactions, we first exam- ined the photogeneration of '02 from mitochondrial membranes, expecting to identify the internal chromophores which act as endogenous sensitizers.
*To whom correspondence should be addressed. tAbbreviarions: BSA, bovine serum albumin; DCIP, 2,6-
dichlorophenolindophenol; EDTA, ethylenediamine- tetraacetic acid; MA, mersalyl acid; MOPS, 3-N-mor- pholinopropane-sulfonic acid: NDH, NADH dehydro- genase; RNO, N,N-dimethyl-4-nitrosoaniline; SMP, submitochondrial particles: SOD, superoxide dismu- tase.
In this paper, we report that the action spectra for lo2 generation from submitochondrial particles (SMP)? prepared from soybean (Glycine mar) hypocotyls demonstrate that the Fe-S centers of nonheme iron proteins play a major role in photo- sensitization to generate lo2.
MATERIALS AND METHODS
Soybean hypocotyls were grown in the dark at about 25°C for 6-8 days. All chemicals, purchased from Sigma Chemical Co. (St. Louis, MO), were reagent grade and used without further purification.
The isolation of mitochondria from the etiolated tissue was based on the technique of Bonner (1967) as modified by Douce et al. (1972). The various aqueous media used for the isolation of mitochondria and the preparation of SMP were as follows: isolation medium; 0.3 M mannitol, 4 mM cysteine, 1 mM EDTA, 0.2% (wtlvol) defatted BSA, and 10 mM K-phosphate, adjusted to pH 7.2. Wash medium; 0.3 M mannitol, 1 mM EDTA, and 10 mM K- phosphate, adjusted to pH 7.2. Suspension medium: 0.3 M mannitol, 5 mM MgC12, 10 mM KCI, and 10 mM K-phos- phate, adjusted to pH 7.2. Mitochondria were purified by silica sol density gradient centrifugation as described by Jackson et al. (1979). The density gradient was prepared with final concentrations of 45% (vol/vol), 21% (vol/vol), and 13.5% (vollvol) Percoll, each containing 0.25 M sucrose, 0.2% (wtlvol) BSA, and 10 mM MOPS. The final pH was adjusted to 7.2 with KOH. Each gradient comprised 6 me of 45%, 12 me of 21%, and 12 me 13.5% Percoll mixtures in a 50 mE tube. Centrifugation was per- formed at 7500 g for 30 min, utilizing an angle rotor. After separation of the mitochondrial fraction, it was resus- pended in wash medium and pelleted at 11 OOO g for I5 min. Mitochondria1 pellets were suspended in suspension medium and sonicated at 23 kHz for 3 x 20 s at 4"C, using a Soniprep 150 Ultrasonic disintegrator (MSE Scientific, Sussex, England). The sonicated suspension was centri- fuged to remove unbroken mitochondria and was applied to Sepharose 4 B column equilibrated with suspension medium. The SMP fraction was collected from the void volume of the column. All procedures for SMP prep- aration were carried out at W C , minimizing the exposure of samples to room light. The SMP suspensions were kept under an N, atmosphere and in the dark in an ice bath
561
562 JIN JUNG et al.
before use. Protein was measured by the Lowry method. Assays of the enzymatic activities of SMP were
conducted at 25°C for NADH dehydrogenase, NADH-cytochrome c reductase and NADH oxidase: NADH dehydrogenase activity was determined as in Rao et nl. (1963), by measuring the decrease in absorption at 600 nm of 2,6-dichlorophenolindophenol (DCIP) in a reaction mixture containing SMP (0.5 mg proteinimt), NADH (0.2 mg). DCIP (20 pM) and KCN ( 5 mM) in the suspension medium. NADH-cytochrome c reductase activity was assayed as described in Yamaki and Uritani (1973), by following the reduction of cytochrome c spectrophotometrically in a reaction mixture containing SMP (0.5 mg protein/mZ), NADH (150 p M ) , ferricyto- chrome c (25 pM), EDTA (5 mM) and KCN (5 mM) in suspension medium. Assay of NADH oxidase activity was carried out using a polarograph constructed in this labora- tory (Jung et al . , 1985); the reaction mixture contained SMP (0.5 mg proteidme) and NADH (1 mM) in suspen- sion medium.
All irradiations, except for enzyme photoinactivation, were performed with monochromatic light. The source was a 750 W Xe lamp (Shanghai bulb factory No. 3, China) used with an f/3.4 grating monochromator (Applied Photo- physics, London, England) with slits set at 6.0 mm. A lamp housing equipped with a condensing mirror and a power supply unit was custom-made locally. In exper- iments where the photoinhibition of the respiratory system was examined, irradiations were done with blue light iso- lated from the Xe lamp by using an absorption filter with a maximum transmittance at 425 nm and an effective bandwidth of 98 nm. The light fluence rates were measured with a Macam quantum/radiometer/photometer model Q. 101 (Macam Photometrics. Livingston, Scotland). Reac- tion mixtures were irradiated at 15°C in a long necked cuvette (10 mm light path) with a water jacket. During irradiation, reaction mixtures were bubbled gently with air.
The imidazole plus RNO (imidazole-RNO) method, developed by Kraljif and El Mohsni (1978), was used to measure lo, generation from SMP upon near UV and visible light irradiation. The lo,-mediated oxidation reac- tion was monitored spectrophotometrically by measuring the decrease in absorption at 440 nm of N,N-dimethyl- 4-nitrosoaniline (RNO). The reaction mixtures for this experiment contained RNO (5 pM), imidazole (8 mM), and SMP (0.4 mg proteinhe) in suspension medium. (Hereafter, we will call these the SMP-imidazole-RNO systems.) For absorption spectra and kinetic measure- ments throughout the work, a Cary 118 spectrophotometer (Varian Assoc., Palo Alto, CA) was used.
RESULTS AND DISCUSSION
Detection of 'Or in an aqueous solution where it has a rather short lifetime is sometimes difficult, especially in cases where other strong reactive inter- mediates cause interference. The imidazole-RNO method, however, was used successfully in the selec- tive detection of '02 even in an aqueous system as y-radiolyzed water where the yields of interfering intermediates may be significantly higher than the yield of lo2 (KraljiC and El Mohsni, 1978). Although the y-radiation of air saturated water results in the formation of various primary and sec- ondary intermediates and products such as hydroxyl radicals (.OH), H atoms, hydrated electrons, super- oxide radicals (03, hydrogen peroxide, and mol- ecular hydrogen, only .OH radicals bleach RNO in
the imidazole-RNO system, which can be com- pletely eliminated by the addition of a certain .OH scavenger (KraljiC and Trumbore, 1965). Thus, if .OH radicals were generated in our experimental systems, the presence of mannitol, a .OH scavenger, at a relatively high concentration in the reaction mixtures should protect the systems against .OH attack.
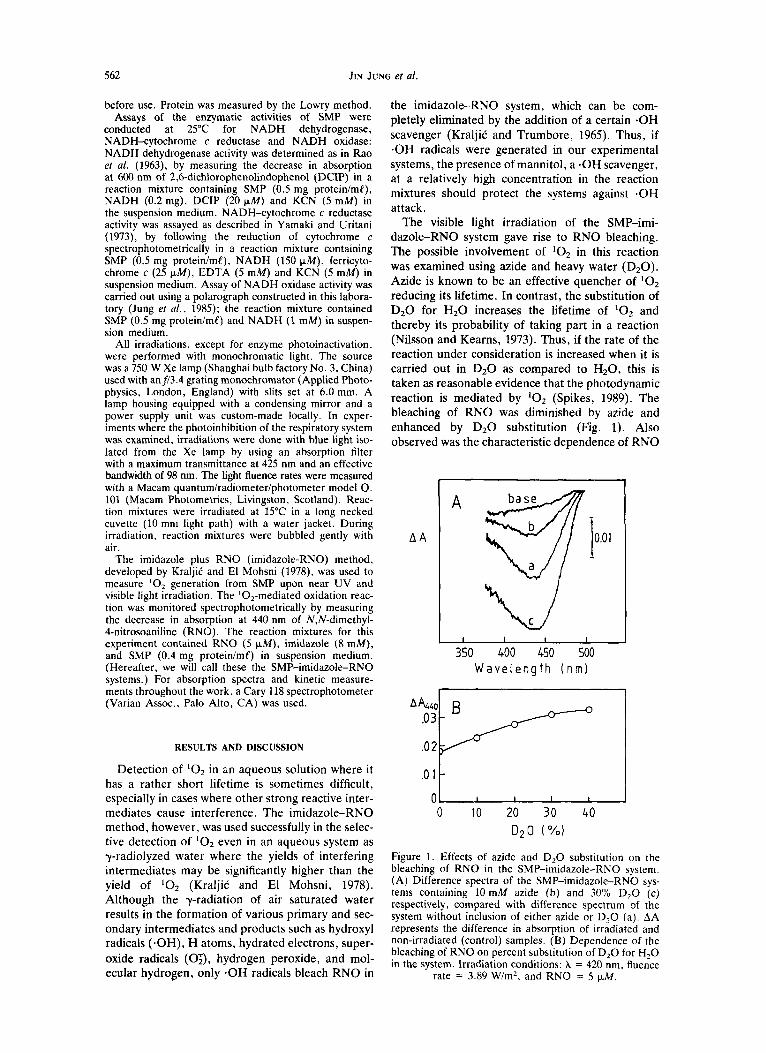
The visible light irradiation of the SMP-imi- dazole-RNO system gave rise to RNO bleaching. The possible involvement of lo2 in this reaction was examined using azide and heavy water (D20). Azide is known to be an effective quencher of lo2
reducing its lifetime. In contrast, the substitution of D 2 0 for H 2 0 increases the lifetime of '02 and thereby its probability of taking part in a reaction (Nilsson and Kearns, 1973). Thus, if the rate of the reaction under consideration is increased when it is carried out in D 2 0 as compared to HzO, this is taken as reasonable evidence that the photodynamic reaction is mediated by '02 (Spikes, 1989). The bleaching of RNO was diminished by a i d e and enhanced by D 2 0 substitution (Fig. 1). Also observed was the characteristic dependence of RNO
I
ba sf
350 400 450 500 W a v e l e n g t h ( n m )
.o 11 0 10 20 30 40
02 0
Figure 1. Effects of azide and D,O substitution on the bleaching of RNO in the SMP-imidazole-RNO system. (A) Difference spectra of the SMP-imidazole-RNO sys- tems containing 10 mM azide (b) and 30% D,O (c) respectively, compared with difference spectrum of the system without inclusion of either azide or D,O (a). AA represents the difference in absorption of irradiated and non-irradiated (control) samples. (B) Dependence of the bleaching of RNO on percent substitution of D,O for H,O in the system. Irradiation conditions: X = 420 nm, fluence
rate = 3.89 W/m2, and RNO = 5 w M .
Singlet oxygen from mitochondrial membranes S63
bleaching on the concentration of imidazole (Fig. 2), that is known to provide a specific test for the presence of lo2 in the system (KraljiC and El Mohsni, 1978; Krishna el al., 1987). These obser- vations indicate that '02 is generated from SMP by irradiation.
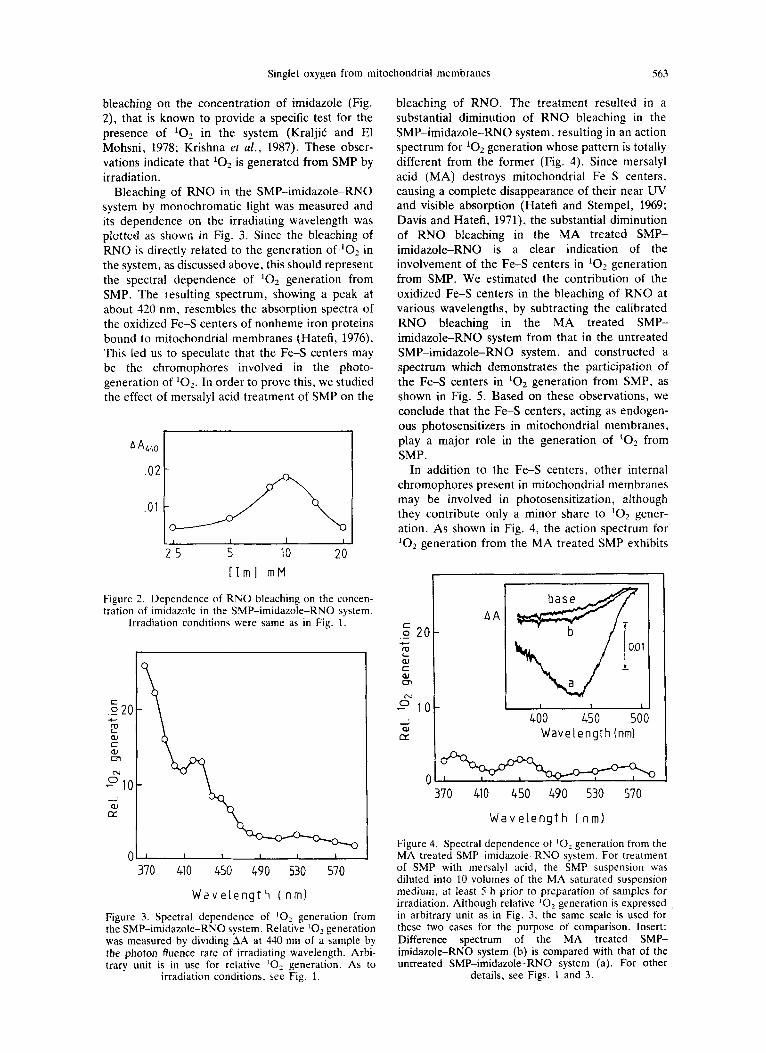
Bleaching of RNO in the SMP-imidazole-RNO system by monochromatic light was measured and its dependence on the irradiating wavelength was plotted as shown in Fig. 3. Since the bleaching of RNO is directly related to the generation of '02 in the system, as discussed above, this should represent the spectral dependence of '02 generation from SMP. The resulting spectrum, showing a peak at about 320 nm, resembles the absorption spectra of the oxidized Fe-S centers of nonheme iron proteins bound to mitochondrial membranes (Hatefi, 1976). This led us to speculate that the Fe-S centers may be the chromophores involved in the photo- generation of lo2. In order to prove this, we studied the effect of mersalyl acid treatment of SMP on the
I I I
2.5 5 10 20 [ I m l rnM
Figure 3. Dependence of RNO bleaching on the concen- tration of imidazole in the SMP-imidazole-RNO system.
Irradiation conditions were same as in Fig. 1.
9
c .o 20 2!
m
- .I-
W C W
cu
- 0 1 0 - d
W Dz
I I I I 1 1 370 410 450 490 530 570
W a v e l e n g t h ( n r n l
Figure 3. Spectral dependence of lo2 generation from the SMP-imidazole-RNO system. Relative IO, generation was measured by dividing AA at 440 nm of a sample by the photon fluence rate of irradiating wavelength. Arbi- trary unit is in use for relative '02 generation. As to
irradiation conditions, see Fig. 1.
bleaching of RNO. The treatment resulted in a substantial diminution of RNO bleaching in the SMP-imidazole-RNO system, resulting in an action spectrum for '02 generation whose pattern is totally different from the former (Fig. 4). Since mersalyl acid (MA) destroys mitochondrial Fe-S centers, causing a complete disappearance of their near UV and visible absorption (Hatefi and Stempel, 1969; Davis and Hatefi, 1971), the substantial diminution of RNO bleaching in the MA treated SMP- imidazole-RNO is a clear indication of the involvement of the Fe-S centers in '02 generation from SMP. We estimated the contribution of the oxidized Fe-S centers in the bleaching of RNO at various wavelengths, by subtracting the calibrated RNO bleaching in the MA treated SMP- imidazole-RNO system from that in the untreated SMP-imidazole-RNO system, and constructed a spectrum which demonstrates the participation of the Fe-S centers in '02 generation from SMP, as shown in Fig. 5. Based on these observations, we conclude that the Fe-S centers, acting as endogen- ous photosensitizers in mitochondrial membranes, play a major role in the generation of lo2 from SMP.
In addition to the Fe-S centers, other internal chromophores present in mitochondrial membranes may be involved in photosensitization, although they contribute only a minor share to '02 gener- ation. As shown in Fig. 4, the action spectrum for '02 generation from the MA treated SMP exhibits
-
A A
400 450 500 Wavelength (nrnl
370 410 450 490 530 570
W a v e l e n g t h (n rn )
Figure 4. Spectral dependence of lo2 generation from the MA treated SMP-imidazole-RNO system. For treatment of SMP with mersalyl acid, the SMP suspension was diluted into 10 volumes of the MA saturated suspension medium, at least 5 h prior to preparation of samples for irradiation. Although relative lo, generation is expressed in arbitrary unit as in Fig. 3, the same scale is used for these two cases for the purpose of comparison. Insert: Difference spectrum of the MA treated SMP- imidazole-RNO system (b) is compared with that of the untreated SMP-imidazole-RNO system (a). For other
details, see Figs. 1 and 3 .
564 JIN JUNG et al.
.- : 20 -I-
F aJ c aJ m N
,o 1 0 - a, CL
C 370 410 450 490 530 570
W a v e l e n g t h ( n r n )
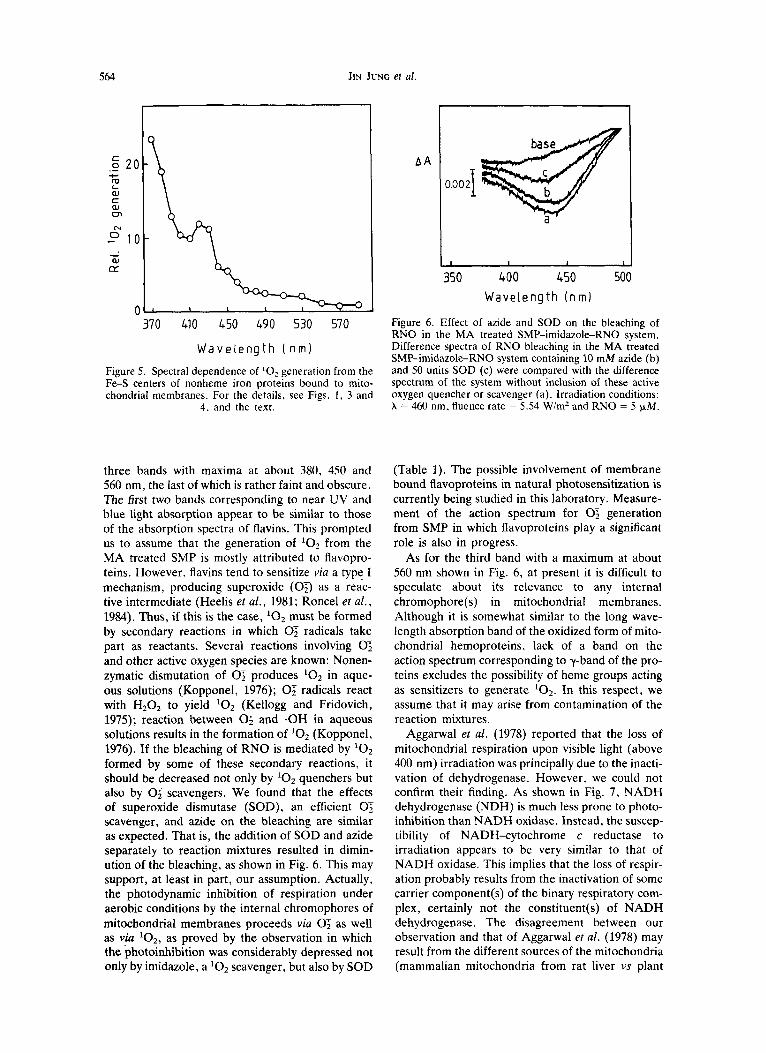
Figure 5. Spectral dependence of '0, generation from the Fe-S centers of nonheme iron proteins bound to mito- chondrial membranes. For the details, see Figs. 1 , 3 and
4. and the text.
three bands with maxima at about 380, 450 and 560 nm, the last of which is rather faint and obscure. The first two bands corresponding to near UV and blue light absorption appear to be similar to those of the absorption spectra of flavins. This prompted us to assume that the generation of '02 from the MA treated SMP is mostly attributed to flavopro- teins. However, flavins tend to sensitize via a type I mechanism, producing superoxide (03 as a reac- tive intermediate (Heelis et al., 1981; Roncel et al . , 1984). Thus, if this is the case, '02 must be formed by secondary reactions in which 0; radicals take part as reactants. Several reactions involving 0; and other active oxygen species are known: Nonen- zymatic dismutation of 0; produces '02 in aque- ous solutions (Kopponel, 1976); 0; radicals react with H202 to yield lo2 (Kellogg and Fridovich, 1975); reaction between 0; and .OH in aqueous solutions results in the formation of '02 (Kopponel, 1976). If the bleaching of RNO is mediated by '02 formed by some of these secondary reactions, it should be decreased not only by '02 quenchers but also by 0; scavengers. We found that the effects of superoxide dismutase (SOD), an efficient 0; scavenger, and azide on the bleaching are similar as expected. That is, the addition of SOD and azide separately to reaction mixtures resulted in dimin- ution of the bleaching, as shown in Fig. 6. This may support, at least in part, our assumption. Actually, the photodynamic inhibition of respiration under aerobic conditions by the internal chromophores of mitochondrial membranes proceeds via 0; as well as via lo2, as proved by the observation in which the photoinhibition was considerably depressed not only by imidazole, a '02 scavenger, but also by SOD
A A
350 400 450 500 W a v e l e n g t h (n m)
Figure 6 . Effect of azide and SOD on the bleaching of RNO in the MA treated SMP-imidazole-RNO system. Difference spectra of RNO bleaching in the MA treated SMP-imidazole-RNO system containing 10 mM azide (b) and 50 units SOD (c) were compared with the difference spectrum of the system without inclusion of these active oxygen quencher or scavenger (a). Irradiation conditions: A = 460 nm, fluence rate = 5.54 WinZ and RNO = 5 JLM.
(Table 1). The possible involvement of membrane bound flavoproteins in natural photosensitization is currently being studied in this laboratory. Measure- ment of the action spectrum for 0; generation from SMP in which flavoproteins play a significant role is also in progress.
As for the third band with a maximum at about 560 nm shown in Fig. 6, at present it is difficult to speculate about its relevance to any internal chromophore(s) in mitochondrial membranes. Although it is somewhat similar to the long wave- length absorption band of the oxidized form of mito- chondrial hemoproteins, lack of a band on the action spectrum corresponding to y-band of the pro- teins excludes the possibility of heme groups acting as sensitizers to generate '02. In this respect, we assume that it may arise from contamination of the reaction mixtures.
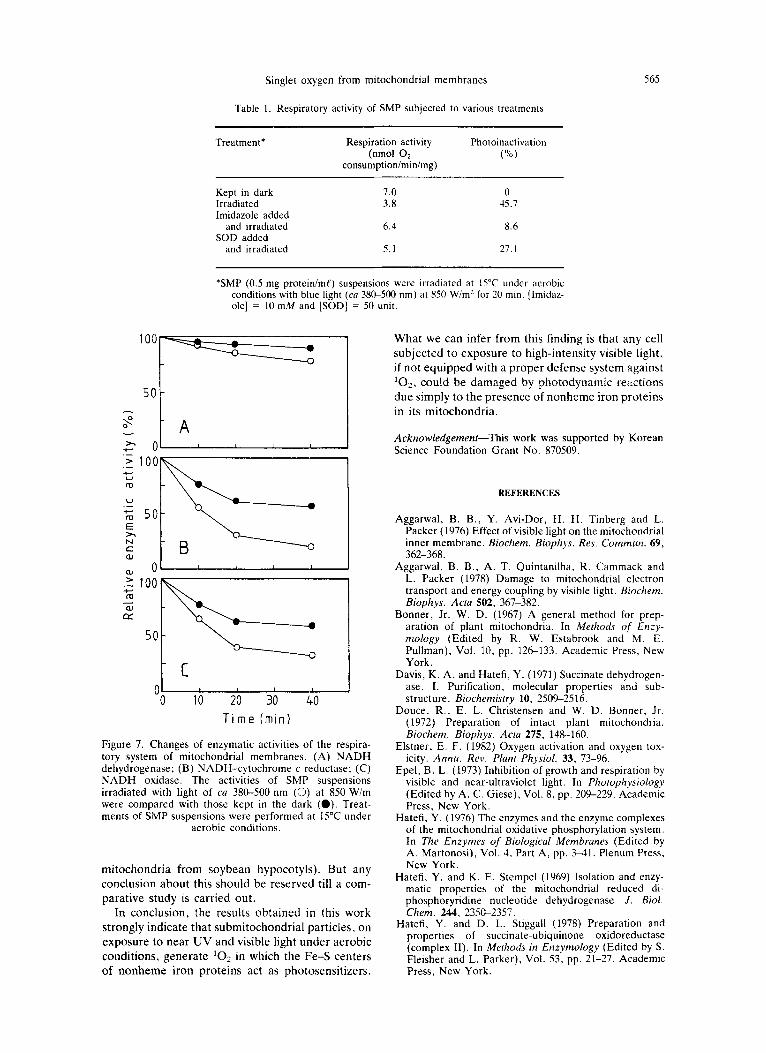
Aggarwal et al. (1978) reported that the loss of mitochondrial respiration upon visible light (above 400 nm) irradiation was principally due to the inacti- vation of dehydrogenase. However, we could not confirm their finding. As shown in Fig. 7, NADH dehydrogenase (NDH) is much less prone to photo- inhibition than NADH oxidase. Instead, the suscep- tibility of NADH-cytochrome c reductase to irradiation appears to be very similar to that of NADH oxidase. This implies that the loss of respir- ation probably results from the inactivation of some carrier component(s) of the binary respiratory com- plex, certainly not the constituent(s) of NADH dehydrogenase. The disagreement between our observation and that of Aggarwal et al. (1978) may result from the different sources of the mitochondria (mammalian mitochondria from rat liver vs plant
Singlet oxygen from mitochondrial membranes 565
Table 1. Respiratory activity of SMP subjected to various treatments
Treatment* Respiration activity Photoinaetivation (nmol O2 (YO)
consumptioniminlmg)
Kept in dark Irradiated Imidazole added
and irradiated SOD added
and irradiated
7.0 3.8
6.4
5.1
0 45.7
8.6
27.1
*SMP (0.5 mg proteinlmt) suspensions were irradiated at 15°C under aerobic conditions with blue light (ca 38&500 nm) at 850 Wim’ for 20 min. [Imidaz- ole] = 10 mM and [SOD] = 50 unit .
loo-
C I I I
0 10 20 30 40 0
T i m e (min)
Figure 7. Changes of enzymatic activities of the respira- tory system of mitochondrial membranes. (A) NADH dehydrogenase; (B) NADH-cytochrome c reductase; (C) NADH oxidase. The activities of SMP suspensions irradiated with light of ca 380-500 nm (0) at 850 Wim were compared with those kept in the dark (0). Treat- ments of SMP suspensions were performed at 15°C under
aerobic conditions.
mitochondria from soybean hypocotyls). But any conclusion about this should be reserved till a com- parative study is carried out.
In conclusion, the results obtained in this work strongly indicate that submitochondrial particles, on exposure to near UV and visible light under aerobic conditions, generate ‘02 in which the Fe-S centers of nonheme iron proteins act as photosensitizers.
What we can infer from this finding is that any cell subjected to exposure to high-intensity visible light, if not equipped with a proper defense system against ’02, could be damaged by photodynamic rext ions due simply to the presence of nonheme iron proteins in its mitochondria.
Acknowledgement-This work was supported by Korean Science Foundation Grant No. 870509.
REFERENCES
Aggarwal, B. B., Y. Avi-Dor, H. H. Tinberg and L. Packer (1976) Effect of visible light on the mitochondrial inner membrane. Biochem. Biophys. Res. Commun. 69,
Aggarwal, B. B . , A. T. Quintanilha, R. Cammack and L. Packer (1978) Damage to mitochondrial electron transport and energy coupling by visible light. Biochem. Biophys. Acta 502, 367-382.
Bonner, Jr. W. D. (1967) A general method for prep- aration of plant mitochondria. In Methods of Enzy- mology (Edited by R. W. Estabrook and M. E. Pullman), Vol. 10, pp. 126-133. Academic Press, New York.
Davis, K. A . and Hatefi, Y. (1971) Succinate dehydrogen- ase. I. Purification, molecular properties anJ sub- structure. Biochemistry 10, 2509-2516.
Douce, R., E . L. Christensen and W. D. Bonner, Jr . (1972) Preparation of intact plant mitochondria. Biochem. Biophys. Acta 275, 148-160.
Elstner, E . F. (1982) Oxygen activation and oxygen tox- icity. Annu. Rev. Plant Physiol. 33, 73-96.
Epel, B. L. (1973) Inhibition of growth and respiration by visible and near-ultraviolet light. In Photophysiology (Edited by A. C. Giese), Vol. 8, pp. 209-229. Academic Press, New York.
Hatefi, Y. (1976) The enzymes and the enzyme complexes of the mitochondrial oxidative phosphorylation system. In The Enzymes of Biological Membranes (Edited by A . Martonosi), Vol. 4, Part A , pp. 3-41. Plenum Press, New York.
Hatefi, Y. and K. E. Stempel (1969) Isolation and enzy- matic properties of the mitochondrial reduced di- phosphoryridine nucleotide dehydrogenase. 1. Biol. Chem. 2 4 , 2350-2357.
Hatefi, Y. and D. L. Stiggall (1978) Preparation and properties of succinate-ubiquinone oxidoreduetase (complex 11). In Methods in Enzymology (Edited by S. Fleisher and L. Parker), Vol. 53, pp. 21-27. Academic Press, New York.
362-368.

566 JIN JUNG et al.
Heelis, P. F . , B . J. Parson, G. 0. Phillips and J. F. Mckellar (1981) The flavin sensitized photooxidation of ascorbic acid: a continuous and flash photolysis study. Photochem. Photobiol. 33, 7-14.
Jackson, C., J. E. Dench, D. 0. Hall and A. L. Moore (1979) Separation of mitochondria from contamination subcellular structures utilizing silica sol gradient centri- fugation. Plant Physiol. 64, 150-153.
Jung, J., S. Park, S. Lee and S . Kim (1985) Determination of respiratory activity of mitochondria and sub- mitochondria1 particles by using dropping mercury elec- trode. J . Kor. Agric. Chem. SOC. 28, 271-277.
Kellogg, E. W. and I. Fridovich (1975) Superoxide, hydro- gen peroxide, and singlet oxygen in lipid peroxidation by a xanthine oxidase system. J . Biol. Chem. 250,
Koppenol, W. H. (1976) Reactions involving singlet oxy- gen and the superoxide anion. Nature 262, 420-421.
Kraljit, I . and S. El Mohsni (1978) A New Method for the detection of singlet oxygen in aqueous solutions. Photochem. Photobiol. 28, 577-581.
Kraljit, I. and C. N. Trumbore (1965) p-Nitrosodimethy- laniline as an OH radical scavenger in radiation chemis-
8812-8817.
try. J . Am. Chem. SOC. 87, 2547-2550. Krishna, C. M., Y. Lion and P. Riesz (1987) A study of
'0, production by immobilized sensitizer outside the solution. Measurement of '0, generation. Photochem. Photobiol. 45, 1-6.
Nilsson, R. and D. R. Kearns (1973) A remarkable deu- terium effect on the rate of photosensitized oxidation of alcohol dehydrogenase and trypsin. Photochem. Pho- tobiol. 17, 65-68.
Rao, N. A., S. P. Felton, F. M. Huennenkens and B. Mackler (1963) Flavin mononucleotide dehydrogenase. J . Biol. Chem. 238, 449455.
Roncel, M., J . A. Navarro, F. F. De La Rosa and M. A. De La Rosa (1984) Flavin-mediated production of hydrogen peroxide in photoelectrochemical cells. Photo- chem. Photobiol. 40, 395-398.
Spikes, J. D. (1989) Photosensitization. In The Science of Photobiology (Edited by K. C. Smith), 2nd edn, pp. 79-110. Plenum Press, New York.
Yamaki, S. and I. Uritani (1973) Mechanism of chilling injury in sweet potato, changes in lipid-protein interac- tion in mitochondria. Plant Physiol. 51, 883-890.