abstract jaime, xavier a. forest understory regeneration after
TRANSCRIPT

ABSTRACT
JAIME, XAVIER A. Forest Understory Regeneration after Anthropogenic Disturbances in a
Semi-deciduous Tropical Dry Forest in Puerto Rico. (Under the direction of Stacy A. C.
Nelson, Skip J. Van Bloem and Frank H. Koch).
Human disturbance stimulates changes in the community and canopy structure of
forests and reduces their capacity for resilience. There is limited understanding of how short-
term responses in understory regeneration after fire and bulldozing in dry forests can
influence colonization of pioneer generalist species, non-native invasive grasses or native
vegetation. There is an increasing demand for canopy restoration in dry forest ecosystems to
counter the biotic barriers that invasive vegetation can impose on native trees. The main
objectives of our study were to determine the following: 1) whether human disturbance
modifies the ecosystem of the deciduous dry forest, such that altered soil abiotic conditions
would favor the germination and growth of grasses over woody vegetation in the understory
after disturbances; and 2) with respect to post-disturbance regeneration, whether severe fire
or bulldozing will have distinctive impacts on seasonal seedlings, seedling size classes,
sprouts, species richness and invasion pools within disturbed sites. To answer these
questions, a total of 64 understory sampling subplots (1m2) for five treatments were located
in a mature forest site and a total of 36 subplots for two treatments in a secondary forest site.
Both sites had recently burned due to human activity. Data on soil moisture flux, vegetation
and plant invasion dispersal were collected in the study.
Results showed significant differences in fluctuations of soil moisture because of
seasonal rainfall among treatments in the mature site, but no difference among treatments in

the secondary site. Quicker understory seedling, sprout and species richness regeneration was
revealed in the burn treatment of the secondary site and the bulldozed treatment of the mature
site, while the burn treatment from the mature site had the lowest regeneration during the
year cycle. Recovery and control treatments maintained a stable high concentration of soil
moisture flux, seedling density, sprout density and species richness during the entire year
cycle. The open canopy treatment (Uniola virgata grass) of the mature site showed some
constant establishment of woody species seedlings, with no significant progress in size
classes and significant reduction in species richness during winter 2012, but a substantial
increase in species richness during summer 2013. However, the mature site old-burn
treatment and the secondary site burn treatment displayed strong seedling growth in terms of
both size class and species richness during the year cycle. Results of a post-bulldozing
species colonization study in the mature forest site showed strong dispersal of pioneer
species, both non-native invasive grasses and naturalized generalist trees, from the road into
the forest. These findings provide insight into how time and abiotic conditions in the soil
influence understory regeneration dynamics and invasion after wildfire and bulldozing
disturbances. Even though dry forests are capable of absorbing human activities that mimic
natural disturbances, the frequency and intensity of those disturbances will have major
impacts on the functionality of the system. Anthropogenic disturbances can impact forest
biodiversity and community assemblages by facilitating the intrusion of non-native
vegetation that outcompetes local flora for the open niche. Furthermore, community
assembly dynamics in such dry forest sites demand further long-term research on
community-level interactions between soil nutrient dynamics and plant community response

to such disturbances. This is particularly relevant since an alternative stable stage is
unreachable for this forest type, and the persistent damage caused by such anthropogenic
disturbances could degrade the integrity of the ecosystem to the point of shifting towards
other systems (i.e. chaparral, savanna or grassland).

© Copyright 2014 Xavier A. Jaime
All Rights Reserved

Forest Understory Regeneration after Anthropogenic Disturbances in a Semi-deciduous
Tropical Dry Forest in Puerto Rico
by
Xavier A. Jaime
A thesis submitted to the Graduate Faculty of
North Carolina State University
in partial fulfillment of the
requirements for the degree of
Master of Science
Forestry
Raleigh, North Carolina
2014
APPROVED BY:
_______________________________ ______________________________
Stacy A. C. Nelson, PhD. Frank H. Koch, PhD
Committee Co-Chair Committee Co-Chair
________________________________
Skip J. Van Bloem, PhD.
Committee Co-Chair

ii
DEDICATION
I would like to dedicate this thesis to my mother, Liana, my father Luis, my brother
Luis Armando, my grandfather Diego Jaime, to my advisers that kept me working hard and
many friends that kept me entertained in my spare time. I am greatly thankful for their moral
support and for everything I learned during this process. I especially feel in debt to those who
always believed in me, even if I didn’t believe in myself, and encouraged me to keep
working hard and never stop following my dreams.

iii
BIOGRAPHY
Xavier A. Jaime Dávila is originally from Humacao, Puerto Rico. He graduated cum
laude from the University of Puerto Rico at Humacao with a Bachelor of Science in General
Biology. During his undergraduate education, he found a special interest in courses such as
community ecology, taxonomy of vascular plants and botany. Those courses nourished his
passion for field research in tropical dry forest ecosystems, and lab work in plant phenology
and evolution. His performance as an undergraduate researcher and field assistant led to a
graduate level interview for a research opportunity in Forestry at North Carolina State
University, which concluded in this thesis. Once his Master’s degree is completed, he plans
to publish the chapters in peer-reviewed journals and continue his career in the ecology field,
with a particular emphasis in invasion ecology.

iv
ACKNOWLEDGEMENTS
First of all, I would like to thank Dr. Denny S. Fernandez Del Viso, Dr. Raymond
Tremblay Laland and Dr. Elvia Melendez-Ackerman for giving me my first scientific
experience in the fields of invasion ecology, ecosystem ecology and evolution. Because of
that opportunity during my undergraduate education, I have been able to gather the essential
tools to become a potential graduate student. In addition, they and Mrs. Emma Suarez
endorsed me for a potential graduate position at North Carolina State University. Special
thanks to Dr. Stacy A. C. Nelson from North Carolina State University for choosing me as
his graduate master’s student in the department of Forestry and Environmental Resources,
and for being able to find the funding from the USDA Forest Service to support my research,
living expenses and course load. Thanks to Dr. Skip J. Van Bloem for being my main
research adviser and for providing me important knowledge and guidance in the process of
studying ecosystem recovery after major disturbances. Special thanks to Dr. Frank Koch for
co-chairing my progress writing the chapters, and for his strong critical thinking in the
process of discussing the work done. I’m thankful for having Tristan Allerton as my field
partner during the brutal first field season and being able to share with him all the ups and
downs that come with the field season. Thank you to all my field technicians/volunteers from
field season II and III, James Rodriguez, Axel J. Acevedo, Luis A. Jaime and Diego Jaime.
Moreover, thanks to my dear pack friends from NCSTATE that became my second family
during this process.

v
TABLE OF CONTENTS
LIST OF TABLES.................................................................................................................vii
LIST OF FIGURES................................................................................................................ix
CHAPTER I
Literature Review........................................................................................................1
CHAPTER II
1.0 Introduction..........................................................................................................11
2.0 Materials and Methods........................................................................................14
2.1 Study sites..................................................................................................14
2.2 Experimental design and measurement of understory regeneration….....16
2.3 Statistical analysis.....................................................................................19
3.0 Results...................................................................................................................20
3.1 Soil abiotic conditions after disturbances………………………………..20
3.2 Woody vegetation coverage regeneration study........................................23
3.3 Native grass coverage regeneration study.................................................26
3.4 Seasonal understory regeneration.............................................................29
3.5 Open canopy vs. closed canopy study: seedling size classes.....................36
4.0 Discussion.............................................................................................................39
4.1 Forest cover regeneration..........................................................................39
4.2 Understory regeneration and species richness..........................................41
4.3 Seedling size class......................................................................................43
5.0 Conclusions...........................................................................................................44

vi
6.0 References...........................................................................................................46
CHAPTER III
Abstract....................................................................................................................56
1.0 Introduction................................................................................................................57
2.0 Materials and Methods..............................................................................................59
2.1 Study site.........................................................................................................59
2.2 Experimental design and measurement of bulldozed trail cross-
transects....................................................................................................60
2.3 Statistical analysis...........................................................................................61
3.0 Results.........................................................................................................................66
3.1 Study of invasive-native grasses and generalist tree species successional
niche colonization after bulldozing disturbance.......................................66
4.0 Discussion...................................................................................................................77
5.0 Conclusions.................................................................................................................78
6.0 References...................................................................................................................79

vii
LIST OF TABLES
CHAPTER II
Table Page
Table 1. Annual accumulation of soil moisture flux (m3/me VWC) from each
treatment following disturbances in Ensenada and Tallaboa-Castle sites as
explanatory variables for seasonality. Superscript letters represent Tukey’s
HSD test for multiple comparison procedure..................................................21
Table 2. Seedling size class (cm height) count during summer 2012 (S-12), winter
2012 (W-12) and summer 2013 (S-13) after disturbances in Ensenada,
Guánica mature site..........................................................................................37
Table 3. Seedling size class (cm height) count during summer 2012 (S-12), winter
2012 (W-12) and summer 2013 (S-13) after disturbances in Tallaboa-Castle,
Guayanilla secondary site................................................................................38
CHAPTER III
Table Page
Table 1. Descriptions and expected direction of a priori occurrence effect model for
invasive grasses from the 555-meter bulldozed cross-transect in the Guánica
Dry Forest reserve section of Ensenada, Guánica. All models are set under
95% confidence interval...................................................................................63
Table 2. Descriptions and expected direction of a priori occurrence effect model for
Uniola virgata and other native grasses from the 555-meter bulldozed cross-
transect in the Guánica Dry Forest reserve section of Ensenada, Guánica. All
models are set under 95% confidence interval.................................................64
Table 3. Descriptions and expected direction of a priori distribution effect model for
Leucaena leucocephala introduced tree species from the 555-meter bulldozed
cross-transect in the Guánica Dry Forest reserve section of Ensenada,
Guánica. All models are set under 95% confidence interval...........................65

viii
Table 4. Statistics for models of M. maximus probability of occurrence derived from
cross-transect surveys conducted along the 555-m bulldozed trail in the
Guánica Dry Forest reserve section of Ensenada. All models are set under
95% confidence interval set. (*) Preferred model.……………………………72
Table 5. Statistics for models of U. virgata probability of occurrence based on cross-
transect surveys conducted along the 555-m bulldozed trail in the Guánica
Dry Forest reserve section of Ensenada. All models are set under 95%
confidence interval set. (*) Preferred model......................................................74
Table 6. Statistics for models of L. leucocephala probability of occurrence based on
cross-transect surveys conducted along the 555- m bulldozed trail in the
Guánica Dry Forest reserve section of Ensenada. All models are set under
95% confidence interval set. (*) Preferred model.............................................76

ix
LIST OF FIGURES
CHAPTER II
Figure Page
Figure 1. Map of study area and treatments in Ensenada mature forest site, Guánica
PR.....................................................................................................................15
Figure 2. Map of study area and treatments in Tallaboa-Castle secondary forest site,
Guayanilla PR…..............................................................................................16
Figure 3. Cumulative concentrations of soil moisture flux (+SD) after disturbances over
time (April-12 to April-13) among all treatments in Ensenada site. P-value
applies to the soil moisture monthly variation.…………................................22
Figure 4. Cumulative concentrations of soil moisture flux (+SD) after disturbances over
time (June-12 to May-13) among all treatments in Tallaboa-Castle site. P-
value applies to the soil moisture monthly variation.……..............................23
Figure 5. Percentage of woody vegetation coverage (±SE) among all treatments in
Ensenada mature site during the year study. P-value applies to the difference
between treatments.………………………………..........................................25
Figure 6. Percentage of woody vegetation coverage (±SE) among all treatments in
Tallaboa-Castle secondary site during the year study. P-value applies to the
difference between treatments.……………….………...................................26
Figure 7. Percentage of Uniola grass coverage (±SE) among all treatments in Ensenada
mature site during the year study. P-value applies to the difference between
treatments.…………………………………………........................................27
Figure 8. Percentage of Uniola grass coverage (±SE) among all treatments in Tallaboa-
Castle secondary site during the year study. P-value applies to the difference
between treatments......................................……………………….…………28

x
Figure 9. Mean seedling regeneration (±SE) among all treatments in Ensenada mature
site during the year study. P-value applies to the difference between
treatments.........................................................................................................30
Figure 10. Mean seedling regeneration (±SE) among all treatments in Tallaboa-Castle
secondary site during the year study. P-value applies to the difference between
treatments.........................................…………………………………………32
Figure 11. Mean sprout regeneration (±SE) among all treatments in Tallaboa-Castle
secondary site during the year study. P-value applies to the difference between
treatments.........................................………………...……………………….34
Figure 12. Mean species richness density (±SE) among all treatments in Ensenada mature
site during the year study. P-value applies to the difference between
treatments.........................................................................................................35
Figure 13. Mean species richness density (±SE) among all treatments in Tallaboa-Castle
secondary site during the year study. P-value applies to the difference between
treatments.........................................………………………………….……...36
CHAPTER III
Figure Page
Figure 1. Map of the 555-meter long bulldozed trail (brown line) in Ensenada site with
mature forest stand nature, Guánica PR...........................................................60
Figure 2. Post-bulldozing dispersal pattern of M. maximus, L. leucocephala and U.
virgata along the bulldozed trail during (A) summer 2012, (B) winter 2012
and (C) summer 2013. Note: Locations where none of the three species
occurred were dominated only by sprouting trees other than L. leucocephala.
Gaps presence between distances represent areas dominated only by sprouting
trees..................................................................................................................67

xi
Figure 3. Logistic regression of invasive grasses occurrence model (β0+β1 (Dist_z),
β1< 0) of colonization from road side to bulldoze cross-transect end region at
600 meters range..............................................................................................70
Figure 4. Logistic regression model of native grass occurrence, based only on distance
(β0+β1 (Dist_z), β1>0)....................................................................................73
Figure 5. Logistic regression model of L. leucocephala occurrence, based only on
distance (β0+β1 (Dist_z),
β1<0)................................................................................................................75

1
CHAPTER I
Literature Review
Rates of ecosystem processes are frequently self-regulating with respect to
disturbances and successional changes in ecosystem dynamics, but are sensitive to the
severity, frequency and type of disturbance taking place (Chapin et al, 2002). With respect to
tropical forests, ecosystem restoration often benefits from the addition of positive inputs and
feedbacks between its components that provide full recovery or a new, more stable state for
the disturbed ecosystem (Chapin et al, 2002). Many anthropogenic and natural disturbances
such as soil erosion, wildfire, habitat fragmentation, invasive species introduction and land
degradation must be addressed if restoration practices are to recover natural composition,
structure and processes.
However, regeneration of native communities is highly dependent on resiliency,
species adaptability and composition. The species pool in an ecosystem dictates the
distribution and behavior of plant communities especially where non-native species have
altered fire regimes, creating dispersed and irregular effects on the landscape (Wolfe and Van
Bloem, 2012; D’Antonio and Vitousek, 1992). The interplay between species pool, plant
community structure, canopy, and soil quality determine the response to disturbance and re-
organization of the system, which can buffer changes and maintain crucial functions (Walker
et al, 2004; Folke, 2006). Even if the interaction between resilience and disturbance brings
opportunities of possible recombination of evolved structures and developmental processes in

2
the system (Folke, 2006), the continual occurrence of periodic anthropogenic disturbances
can push the system to a less desirable state where resilience is almost unapproachable.
With respect to ecological resilience, tropical forests represent a major area of focus
because of the increasing frequency of anthropogenic and natural disturbance due to climate
change and human population growth (Thompson et al., 2009; Gunderson, 2000; Molina and
Lugo, 2006). Over 70% of forest fires occur in tropical regions of the globe (Dwyer et al.,
2000; Dwyer et al., 1998), and climate change is expected to increase fire frequency, which
will have ecological and socio-economic repercussions, including for Caribbean ecosystems
(Robbins et al., 2008). Many studies of tropical forests have suggested that fragmentation,
deforestation, land-use change and periodic wildfires limit forest regeneration and ultimately
affect vulnerable species diversity and richness (Molina and Lugo, 2006; Ammondt and
Litton, 2012; Stork et al., 2009). Particularly in the understory succession process in tropical
dry forests, seed banks represent a major source of forest equilibrium from the dry season to
the rainy season (Ceccon et al., 2006), but since wildfires occur mostly in dry season periods,
seed banks can lose viability under extreme heat conditions (Robbins et al., 2008). Indeed, in
similar ecosystem scenarios such as Puerto Rico and Virgin Island’s dry forests, seed bank
viability is too low to drive regeneration under normal circumstances and seedling survival is
almost nonexistent after fire disturbances (Ray and Brown, 1994; Santiago-Garcia et al,
2008).
In recent studies regarding semi-deciduous dry forests and shrub lands in the tropics,
competition between invasive grasses and native vegetation appears to be a major factor
hindering the possibility of equilibrium in the system (J. Rojas-Sandoval et al., 2012). Non-

3
native species invasions in tropical dry forests around the equator have triggered significant
loss of biodiversity, economic problems and disastrous effects in semi-dry forested
ecosystems (Vitousek et al., 1997; Mack et al., 2000; Cabin et al., 2002). Native plant
communities in dry deciduous forests not only suffer niche competition by non-native grasses
(Gallagher et al., 2010), but during the dry season, these invasive grasses also increase fuel
loadings that are sensitive to fire ignition (Robbins et al., 2008; Brandeis and Woodall,
2008). Many initiatives involving reforestation and regeneration of dry deciduous forests that
have undergone fragmentation, wildfire and deforestation, are consistently disrupted by the
resulting synergetic effects of these disturbances, which promote changes in forest ecosystem
structure and microclimates (Cochrane, 2002). When large-scale recovery is constantly
interrupted by the combined effect of disturbances, species interactions and biological
constraints, it is important to consider abiotic parameters such as precipitation patterns when
developing restoration practices since environmental factors (i.e. soil moisture, temperature
and seasonality) can significantly influence plant community structure (Cabin et al., 2002; J.
Rojas-Sandoval et al., 2012).
The concept of alternative stable states as the result of perturbed communities shifting
from their original states is not entirely clear in experimental ecology (Beisner et al., 2003).
Essentially, when disturbances are highly frequent in previously disturbed forests, an
alternative stable state is practically an unreachable goal. For that reason, it is important to
identify the disturbances, the variables affected in the system and those changes that
eventually result in a progressive loss of resiliency (Beisner et al., 2003). The forest’s ability
to maintain resilience depends on tree canopy cover, vegetation vulnerability to hysteresis,

4
understory vegetation regeneration, precipitation-moisture variability, soil dynamics, human
disturbances, species pool and species richness (Hirota et al., 2011). On the perspective of
canopy cover, related studies in secondary subtropical and tropical dry forests, suggest that
tree species with dominant characteristics and that are well-adapted to disturbances (i.e., that
can withstand hysteresis) should be used for restoration purposes, and for natural inhibition
of invasive grass settlement (Lebrija-Trejos et al., 2008; Santiago et al, 2008; Wolfe et al.,
2012).
Since the deforestation of mature and secondary dry forest in Puerto Rico during the
1930's most of the island’s forest cover has been dominated by secondary forests (68%), with
a much smaller proportion (12%) in mature forest (Helmer et al., 2002). However, recent
forest inventories have found that forest cover increased from 364,000 ha to 552,000 ha, and
now comprises 62.2% (20% increase in the last 30 years) of total land cover in Puerto Rico
(Brandeis et al, 2007). In particular, dry forests in the tropics represent less than 42% of all
closed forest types, and only 12% of the live trees in this forest type surpass 10 cm diameter
at breast height or 5 m height, largely because these forests occur on abandoned agricultural
lands or lands clear cut for wood charcoal production in the 1970's (Murphy and Lugo, 1986;
Molina and Lugo, 2006; Wolfe and Van Bloem, 2012). Many of the native trees species are
limited to secondary forest and only 12.5% of the dry forest was left undisturbed, resulting in
the domination of secondary forests with mature forests fragmented into spread out patches
(Brandeis, 2003; Molina and Lugo, 2006). Compared to rainforests, dry forests have the
tendency to recover faster from large-scale natural disturbances such as hurricanes (Van
Bloem et al., 2005). However, continuous anthropogenic disturbances like arson fires are a

5
serious threat to fragile dry forest ecosystems such as in the Guánica Dry Forest Biosphere.
Subtropical semi-deciduous dry forests have slow decomposition of organic matter, low
natural fuel loads and no natural ignition source, which means this system essentially does
not possess a natural fire regime. Since the organic matter present aboveground is rapidly
used by the ecosystem, there is also limited presence of species that promote fire under
natural conditions (Murphy and Lugo 1986; Van Bloem et al. 2004).
High levels of human disturbance stimulate changes in forest composition and reduce
a forest’s capacity for resiliency (Hollings, 1973; Wolfe and Van Bloem, 2012), especially
when there is limited regeneration of native trees; this enables rapid population expansion
and spread of invasive grasses. In fact, dry forests in Puerto Rico are unable to reach an
alternative stable state, which leads to limited feasible solutions for management initiatives in
the implementation of canopy restoration to achieve secondary forests and revert the shift to
savannas or chaparrals caused by grass invasion. Especially when most wildfires are a
resulting feedback between the intrusion of fire-prone and invasive non-native grasses, which
colonize the disturbed areas, increasing fuel loads thereby increase the frequency and
severity of wildfires (Molina and Lugo, 2006; Thaxton et al, 2012).
In order to understand successional processes in post-disturbance regeneration, the
variations in plant community response and invasion need to be evaluated with the
consideration of abiotic factors influencing from both above- and below-ground (i.e. soil
moisture and temperatures). There is experimental evidence which indicates that a single
severe fire can turn an entire forest into chaparral, savanna and eventually grassland in Puerto
Rico, creating a cascade effect on the soil composition and plant community structure in the

6
long run (Litton et al., 2006; Shiels et al, 2008; Wolfe and Van Bloem, 2012). However,
short-term scientific studies need to be performed in order to assess the short-term vegetation
regeneration, as well as the plant community structure and potential invasion response to
wildfire and bulldozing disturbances within treatments in a mature forest site and secondary
forest site. There is a need to evaluate restoration practices for native tree recruitment in the
dry forest, since natural regeneration would not lead to closed canopy cover given a
persistent disturbance regime and grass invasion (Molina and Lugo, 2006; Santiago Garcia et
al, 2008; Ramjohn et al., 2012; Wolfe and Van Bloem, 2012; Garcia-Cancel, 2013).
Management efforts establishing intermediate facilitator pathways by using nursing species
that promote the regeneration process and growth of native tree seedlings in the understory
can be an effective mechanism to achieve regeneration of the ecosystem’s canopy cover. For
example, one tree species, Leucaena leucocephala (Lam.) de Wit, is capable of overcoming
hysteresis of the ecosystem, restoring nitrogen concentrations in soil as an n-fixator and
allocating moisture availability in the soil with its mycorrhizal roots (Parrotta, 2000). Such
species may be able to reverse canopy loss and outcompete the grasses to provide a more
feasible scenario for native tree seedlings to grow after major disturbances (Santiago-Garcia
et al, 2008; Wolfe and Van Bloem, 2012).
The continuous efforts to manage and prevent wildfires in the Guánica Dry Forest
Reserve have been partially successful in restraining the progress of uncontrolled fires from
the road sides to inside the protected forest lands (Canals, 2007). Even though a prescribed
burn initiative has been established among USFS and regional fire fighters since 2003, a
more effective forest management and restoration initiative is needed to control the invasion

7
of grasses and subsequently the wildfires. Nevertheless, before restoration initiatives and
other management actions are initiated, there needs to be better understanding of short-term
regeneration of plant communities in response to anthropogenic disturbances in both mature
and secondary dry forest sites. In particular, there should be detailed analyses of understory
dynamics, in order to clarify patterns of niche colonization by woody species or grasses.
Such analyses should include comparisons of sites and treatment responses, and should
examine the influence of soil abiotic conditions and time.
This thesis was designed to address and provide an understanding of the dynamics
and interactions involved in plant community regeneration in recently and previously
disturbed areas, and the role of soil moisture and temperature in ecosystem biodiversity and
resilience. In detail, this thesis will focus on short-term response of understory regeneration
after wildfires and bulldozing in a protected, mature, semi-deciduous forest stand and after a
wildfire in a previously disturbed secondary forest stand. An additional research objective
was to determine whether plant invasion is occurring after bulldozing disturbance in the
mature forest stand as a result of human activities.
Thesis layout
The thesis is comprised of this introductory chapter and two research chapters that are
projected for publication in peer-reviewed journals. General descriptive information about
the study sites is provided below so that it need not be repeated in each research chapter.
However, the Methods sections of each research chapter expands upon the site explanations
as necessary for each study.

8
Study Sites
The research was conducted in two sites. The first site is in the western section of the
Guánica Commonwealth Dry Forest and Biosphere Reserve near Ensenada, PR
(17°56'59.93" N 66°56'33.83" W elev. 176 ft.), and consists of ~713.69-ha of protected semi-
deciduous tropical dry forest (Ewel and Whitmore, 1973; Wolfe and Van Bloem, 2012). The
soil taxonomy of the site is Fine-loamy, mixed, superactive, isohyperthermic, Typic:
Haplocalcids from the order: Aridisols (Bulldozed entrance comprising 5.3% @ 3.5 acres
with Guayacan clay; 0 to 5% slopes) and Clayey-skeletal, mixed, superactive, nonacid,
isohyperthermic, shallow, Typic: Torriorthents from the order: Entisols (Plot sites comprising
94.7% @ 62.9 acres with Pitahaya-limestone and outcrop-seboruco complex; 40 to 60%
slopes) (USDA Soil survey).
The second site is located on private property in Tallaboa-Castle, Guayanilla, and
consists of ~ 35.69-ha of unprotected semi-deciduous subtropical dry forest, mainly
populated by Fabaceae and Euphorbiaceae shrubs and grasses (18°00'16.03"N 66°45'42.99"
W elev. 154 ft.). Its soil taxonomy is Coarse-loamy, carbonatic, isohyperthermic, aridic,
Typic: Calciustolls from the order: Mollisols (Plot sites comprising 100% @ 8.1 acres with
Aguilita stonyclayloam; 20 to 60% slopes) (USDA Soil survey). Also, the top soil is thin,
alkaline in pH and mostly composed of calcium carbonate and magnesium hydroxide
limestone (USDA, 2008).
Both sites have annual rainfall averaging 860 mm/year (Murphy and Lugo, 1986).
The annual rainfall is primarily distributed in two wet seasons from April-May and August-
November (Murphy and Lugo, 1986), and both sites have an average atmospheric

9
temperature of 25.1°C, which does not vary much throughout the year. The sites are located
~ 20.00 km apart and about 0.15 -1.70 km inland; the second study site is somewhat closer to
the coast than the first site.
Previous wildfires were observed in the Tallaboa-Castle site in the early 1990's,
whereas the Ensenada site had not burned previously except from the west side of the forest
trails (dated back to 2010) in which a 3-year post-burn recovery treatment is located (see
Chapter II). The conditions and nature of those wildfires are highly dependent on the grass
fuel loads (i.e., the amount of fire-prone biomass). The native grass species Uniola virgata
(Poir.) not only represents the most frequently encountered grass species in the ecosystem
under study, but it also tends to form densely clumped colonies in disturbed areas of the
forest with open canopy structure (Monsegur-Rivera, 2009). Additionally, this perennial C4
grass produces a good amount of dead biomass around its base that can represent a potential
source of fire fuel (García-Molinari, 2006). The species’ high germination rate under normal
circumstances, low germination rate after fire and wide distribution across the study sites
makes it useful as a study treatment. The Uniola grass treatment (see Chapter II) is a different
type of control in comparison to the closed canopy forest treatment, as well as a potential
variable to monitor after fire and for future ecosystem regeneration.
Both sites experienced relatively recent fires, Ensenada in March 2012 and Tallaboa-
Castle in April 2012. In addition to these incidents, the Puerto Rico Electric Power Authority
(PREPA) proceeded to bulldoze from the road PR-325 roadside to inside the forest (555.0
meters long by ~ 7.3 meters wide corridor) in the Ensenada site, resulting in further
disturbance. Since the forest is heavily threatened by a variety of invasive grasses across 500

10
– 5000m2 of forest land (Wolfe and Van Bloem, 2012), their percentage of coverage and
occurrence were assessed throughout the disturbed and undisturbed sections of the sites.

11
Chapter II
Case study of Caribbean subtropical semi-deciduous dry forests: Community
assemblage and vegetative understory regeneration after anthropogenic disturbances in
Puerto Rico
1.0 Introduction
Antillean dry forests have experienced high levels of anthropogenic disturbance since
the 16th century in some locations (Ramjohn et al., 2012). More importantly, these disturbed
forests are considered some of the world's hot spots of high endemic population densities of
tropical deciduous trees, but also have a high rate of biodiversity loss due to rapid
deforestation in the19th and early 20th centuries (Maunder et al., 2008; Ramjohn et al.,
2012). Additionally, land use changes are often driven by human activities, triggering
ecosystem alterations that are sometimes exacerbated by external factors like major
economic changes. For example, similar to other parts of the world, a collapse of the
agriculture-based economy promoted extensive land use change in Puerto Rico in the late
1930s, as abandoned agricultural lands experienced reforestation at astonishingly high rates
(Rudel et al., 2000; Brandeis et al., 2007). These sudden land use changes are major drivers
of secondary forest succession, which can have a significant impact on species distributions,
community assemblages and ecosystem functions (Verhoef and Morin, 2010). After the
downfall of agriculture in Puerto Rico (Rivera-Batiz and Santiago, 1996), novel forested
ecosystems became the main pattern of dry forest regeneration. Regardless of the progress

12
made in novel forest regeneration, the closed-canopy cover of these tropical dry forests has
been obstructed by the persistent threat of invasion, competition from shrub species, the
susceptibility of the system to altered fire regimes and additional human-mediated
disturbance (Griscom and Ashton, 2011; Ramjohn et al., 2012).
In tropical dry forests among other ecosystems, wildfires affect plant community
assembly to the extent that decreases in post-fire species richness and populations are often
seen (Hoffmann et al. 2003; Chase, 2003). Fire behavior over landscapes is heterogeneous
and patchy, since the distribution of aboveground biomass fuels and atmospheric conditions
determine the burn patterns (Thaxton et al. 2012; D’Antonio and Vitousek, 1992). As a result
of the historical deforestation that occurred in Puerto Rico during the 17th
through the 19th
centuries with aggressive agricultural practices (Murphy and Lugo, 1986; Franco et al., 1997;
Molina Colón and Lugo, 2006; Griscom and Ashton, 2011; Ramjohn, 2004), much of the
increasing secondary forest regeneration depends on adaptable species composition.
Additionally, land abandonment, land use history and increasing distance from the largest
undisturbed forest patches can have an impact on species richness, but not necessarily on
diversity or community structure (Chinea and Helmer, 2003). Native arboreal species that
are fire-adaptable are typically not found inside dry forests in Puerto Rico (Wolfe, 2008).
Thus, native shrubs and grass communities are capable of quickly colonizing niches left by
the decreasing woody species after frequent fires (Thaxton et al. 2012; Brandeis and
Woodall, 2008). Because of anthropogenic ignitions, vegetative adaptations to fire generally
rely on individuals to survive or regenerate from seed germinations, roots and basal sprouts,
and even resistance to aboveground low- to medium-intensity fires (Otterstrom and

13
Schwartz, 2006). During these types of fires, sprouting provides a much needed benefit to the
plant community, allowing subsistence of the main individual stem to enable post-fire
vegetative establishment. However, with the increasing intrusion of invasive non-native
grasses linked to ignition-prone biomass production, these types of ecosystems experience
constant fire regime alterations due to more intense and persistent fires (Thaxton et al. 2012;
Brooks et al. 2004).
The focus of this study was the quantification of understory vegetative regeneration
components in response to post-fire conditions. Our analyses accounted for differences
between unburned, and previously burned, open-canopy (i.e., grass-covered) and closed-
canopy forest plots by the documentation of short-term fluctuations in understory floristic
regeneration, species composition, and non-native grass invasion into once-burned dry forest
as well as an area of bulldoze disturbance. In order to understand the disturbance effect,
regeneration and resiliency of this system, we focused on the theory that strong species
interactions involved in community structures are either deterministic or historically
contingent (Fukami, 2005). To address these objectives, we examined the following question:
What are the effects that wildfire and bulldoze disturbances have on tropical deciduous dry
forest understory vegetative composition, changes in size class distribution across the
disturbed landscape and species richness within and between forest stands? Our research
hypotheses were as follows: 1) if human disturbance modifies the ecosystem of the
deciduous dry forest then altered soil abiotic conditions among treatment conditions would
favor the germination and growth of grasses over woody vegetation in the understory after
disturbances; and 2) if severe fire and bulldoze determine post-disturbance regeneration in

14
the system, then such disturbances will have distinctive impacts on seasonal seedlings,
seedling size classes, sprouts and species richness pools within disturbed sites.
2.0 Materials and Methods
2.1 Study sites
The research was conducted in two sites. The first site is in the western section of the
Guánica Commonwealth Dry Forest and Biosphere Reserve near Ensenada, PR
(17°56'59.93" N 66°56'33.83" W elev. 176 ft.), and consists of ~713.69-ha of protected semi-
deciduous tropical dry forest (Ewel and Whitmore, 1973; Wolfe and Van Bloem, 2012). The
soil taxonomy of the site is Fine-loamy, mixed, superactive, isohyperthermic, Typic:
Haplocalcids from the order: Aridisols (Bulldozed entrance comprising 5.3% @ 3.5 acres
with Guayacan clay; 0 to 5% slopes) and Clayey-skeletal, mixed, superactive, nonacid,
isohyperthermic, shallow, Typic: Torriorthents from the order: Entisols (Plot sites comprising
94.7% @ 62.9 acres with Pitahaya-limestone and outcrop-seboruco complex; 40 to 60%
slopes) (USDA Soil survey).
The second site is located on private property in Tallaboa-Castle, Guayanilla, and
consists of ~ 35.69-ha of unprotected semi-deciduous subtropical dry forest, mainly
populated by Fabaceae and Euphorbiaceae shrubs and grasses (18°00'16.03"N 66°45'42.99"
W elev. 154 ft.). The site is somewhat closer to the coast than the first study site. Its soil
taxonomy is Coarse-loamy, carbonatic, isohyperthermic, aridic, Typic: Calciustolls from the
order: Mollisols (Plot sites comprising 100% @ 8.1 acres with Aguilita stonyclayloam; 20 to

15
60% slopes) (USDA Soil survey). The top soil is thin, alkaline in pH and mostly composed
of calcium carbonate and magnesium hydroxide limestone (USDA, 2008).
Figure 1: Map of study area and treatments in Ensenada mature forest site, Guánica PR

16
Figure 2: Map of study area and treatments in Tallaboa-Castle secondary forest site,
Guayanilla PR
2.2 Experimental design and measurement of understory regeneration
The conditions in Ensenada’s mature forest site provided the opportunity to develop five
treatments that describe the different scenarios found in the site. The set of treatments is as
follows: 1) Burn - 3.46 acres of the site affected by the wildfire that happened on March 13,
2012, which was caused by a high-voltage power line catching fire after a lightning strike. 2)
Bulldoze – 20 meters long by 6.8 meters wide portion of a bulldozed trail used primarily for
comparison to the Burn treatment in terms of disturbance response. The bulldozed trail was
created by the Puerto Rico Electric Power Authority as a way to move machinery into the
forest to change the burned power line for a new, more efficient power line. This treatment
removed all vegetation as well as 10 cm of top soil. 3) Old-Burn – areas recovering from a

17
wildfire that occurred in 2010. The treatment area is mostly covered by a partially open
canopy structure dominated by shrubs in the understory. 4) Uniola grass open canopy control
treatment dominated by native grasses (i.e. Uniola virgata (Poir.) Griseb) in dense colonies
with trees spread out in areas not colonized by U. virgata and with patches dominated by tree
seedlings in the understory. 5) Forest - closed canopy control treatment that represents the
structure of a common mature forest site in the dry zones of Puerto Rico, dominated by
deciduous trees, matured shrubs and an understory heavily dominated by small woody
seedlings. The Tallaboa-Castle secondary forest site had only two treatments: a recent (April
22, 2012) post-fire scenario, which was labeled as the Castle-Burn treatment, and an
unburned forest control treatment that was labeled as Castle-Forest.
Pre-burn and post-burn understory regeneration were measured with four 1-m2
perpendicular sampling subplots located within a 5.6-m radius circular plot using compass
coordinated locations (1° North, 90° East, 180° South, 270° West). The circular plots were
located within each treatment using a stratified random approach that accounted for the
patchy effect of wildfires in soil, solar irradiation and heterogeneity of the landscape
(Thaxton et al., 2012; Bento-Gonçalves et al., 2012). For the Ensenada mature forest site,
there were six plots in the Burn treatment, three plots in the Old-burn treatment, three plots in
the Forest treatment, three plots in the Uniola grass treatment and four 1 m2 subplots located
in a portion of the Bulldoze trail treatment to maintain consistency with the subplot set-up for
the other treatments. The same sampling approach was employed for the Tallaboa-Castle site,
resulting in six plots in the Castle-Burn treatment and three plots in the Castle-Forest

18
treatment). The total number of understory sampling subplots for repeated measurements in
the Ensenada site was 64, versus 36 subplots in total for the Tallaboa-Castle site.
Grass cover and understory plant community successional behavior was measured by
quantifying the percentage of woody vegetation (trees and shrubs) and native grass coverage
inside the subplots, including any individual that touched a subplot boundary, densities of
emerging seedlings, sprouts (basal, stem or root) and species richness. Emerging seedling
density was determined by their physiological status during summer 2012 survey, winter
2012 survey and summer 2013 survey. All of those seedlings that reached adulthood by
producing inflorescence were disqualified as seedlings and labeled as mature individuals.
Since the majority of shrubs and pioneer species produce inflorescence within a few months,
they were considered mature individuals after the late August-December rainy season and in
summer 2013. We did not apply this same phenology standard for slow-growing hardwood
species, which required height measurement to study the growing process.
We recorded seedling count and composition within the following size classes (Jimu et
al, 2012) : <10cm, 10 – 30 cm, 30 – 50 cm, 50 – 70 cm, and 70 – 90 cm. The size class
distributions provided important information on how seedlings develop, react to biotic and
abiotic conditions present in each treatment and also served as a major component in the
interpretation of seedling mortality and maturity. A survivorship assessment in which
seedlings were monitored during survey seasons and year-round was managed by using
seedling tags (new emerging seedlings were tagged with plastic swizzle sticks color-coded
and documented to keep track of which individuals died, survived or matured). We also

19
measured dry-weight aboveground burned grass fuel biomass in each site and the diameter of
the area burned.
2.3 Statistical analysis
The data on fire consumption of aboveground biomass were used as a percentage index
of fuel load consumption in both the Ensenada and Tallaboa-Castle sites before and after
wildfire. Percentage of vegetation coverage for understory woody and grass vegetative
regeneration among all treatments within each site, between sites burned and forest
treatments, survey periods (summer 2012, winter 2012, and summer 2013) and soil abiotic
conditions were analyzed using two-way ANOVA and Tukey HSD with randomized
incomplete block design (Ensenada block (5 treatments) vs. Tallaboa-Castle (2 treatments)).
Multivariate ANOVA analysis was required for vegetative regeneration per each survey time
(summer 2012, winter 2012, and summer 2013) as categorical values. Vegetation data that
did not met ANOVA assumptions of normality and homogeneity of variance were log10-
transformed (Log10(X+C), C=1) (Howell, 2007; Tabachnick and Fidell, 2007). Shapiro-Wilk
(Royston, 1995) and Kruskal-Wallis (Kruskal and Wallis, 1952) non-parametric tests were
used to compare significance of potential differences found in vegetative maturity and
mortality data that did not have a normal distribution.
Soil moisture (m3/m
3 VWC) and temperature (°C) data were collected as continuous time
series (hourly data from 750 points/mo/probe); averages of all probes per treatment were
subjected to ANOVA analysis to examine vegetation regeneration (percentage of understory
coverage, seedling size classes and seedlings-sprouts-species richness densities) in response
to soil moisture and soil temperature fluxes.

20
3.0 Results
Pre-fire biomass was only collected for the Ensenada site (1.9343 kg/m2 dry weight),
and consisted entirely of Uniola virgata presence. Dry forest species composition between
Ensenada and Tallaboa-Castle were significantly different for species richness distribution (P
= 0.0369) after the fire took place. The percentage of burned grass fuel from the Ensenada
site (89.7%), on 3.46 acres of burned land, was slightly higher than the Tallaboa-Castle site
(86.7%) on 4.40 acres of burned landscape. However, the difference in fire consumption of
woody stem biomass between Ensenada and Tallaboa-Castle was not significantly different
(P = 0.16).
3.1 Soil abiotic conditions after disturbances
Cumulative mean soil moisture (m3/m
3 VWC) among treatments in the Ensenada site
significantly varied (P = 0.00621) during the year cycle (Table 1). The Old-Burn treatment
showed the highest moisture concentration in soil followed by Forest, Burn, Uniola grass and
Bulldoze with the lowest level. Seasonal rainfall and dry season (Figure 3) played an evident
role in soil moisture variation (P = 2.06e-13), showing significant increase in moisture during
the months of August-2012 (highest peak) through December-2012 and the lowest levels in
the dry period from January-2013 to April-2013. However, there was no significant
difference between the Castle-Burn and Castle-Forest treatments in cumulative
concentrations of soil moisture in the Tallaboa-Castle site. In addition, seasonal dry-wet
months (Figure 4) proved to be significantly different (P < 2.0e-16) with high concentration

21
of soil moisture from August-2012 through December-2012 and the lowest concentrations
during January-2013 through May-2013.
Table 1. Annual accumulation of soil moisture flux (m3/m
3 VWC) from each treatment
following disturbances in Ensenada and Tallaboa-Castle sites as explanatory variables for
seasonality. Superscript letters represent Tukey’s HSD test for multiple comparison
procedure.
Treatment Soil moisture flux
(m3/m
3 VWC)
(x̄ ±SD)
Burn 0.143±0.041AF
Bulldoze 0.082±0.021B
Old-Burn 0.186±0.044A
Forest 0.164±0.043AF
Uniola 0.132±0.032F
Castle-Burn 0.118±0.041G
Castle-Forest 0.129±0.045GF

22
Pr (>F) = 2.06e-13
Figure 3: Cumulative concentrations of soil moisture flux (+SD) after disturbances
over time (April-12 to April-13) among all treatments in Ensenada site. P-value
applies to the soil moisture monthly variation.

23
3.2 Woody vegetation coverage regeneration study
The mature forest in Ensenada (Figure 5) and secondary forest in Tallaboa-Castle
(Figure 6) were found to be significantly different (P = 0.0004) based on the percentage of
woody coverage within their respective burn treatments after wildfires. Also, the time effect
was significantly different (P < 0.00001) between the sites. Both sites were found
considerably different (P < 0.00001) between summer 2012 - winter 2012 and between
summer 2012 - summer 2013. Woody vegetation cover in both sites had a seasonal
fluctuation between 20% - 45% coverage in their burn treatments. However, when
evaluating treatment differences in post-disturbance recovery, the percentage of woody
Pr (>F) < 2e-16
Figure 4: Cumulative concentrations of soil moisture flux (+SD) after disturbances
over time (June-12 to May-13) among all treatments in Tallaboa-Castle site. P-
value applies to the soil moisture monthly variation.

24
coverage after wildfire and bulldoze disturbances changed significantly (P = 1.48e-10). The
Forest and the Old-Burn treatments exhibited much higher woody regeneration in the
understory than the Burn and Uniola grass treatments in the Ensenada mature site. These last
two treatments accounted for the lowest overall woody regeneration (below 40%). However,
the bulldoze treatment was not significantly different (P-adj > 0.05) from the other
treatments.
On the other hand, the summer 2012, winter 2012 and summer 2013 survey cycle
shows how rainfall in August-December 2012 played a major role, with winter 2012
displaying an approximately 45% higher rate of regeneration than summer 2012 and slightly
higher than summer 2013. The time effect promoted significant variability in woody
regeneration coverage among (P = 0.0002) all treatments in the Ensenada site. Contrast
analysis for time effect among treatments demonstrated significant variation in woody plant
coverage between summer 2012 – winter 2012 and summer 2012 – summer 2013, with no
significant variation in percentage of woody coverage regeneration during winter 2012 –
summer 2013 (Figure 5).

25
In the Tallaboa-Castle site, variations between forest and burn treatments were highly
significant, with the Castle-Burn treatment exhibiting an average of 40% higher woody
regeneration in the understory than the Castle-Forest control treatment (P = 1.74e-09). Time
effect responded in the same manner with the treatment difference (P = 3.30e-12) showing a
stable seasonal increase in percent of woody coverage in the burn treatment but not the forest
treatment (Figure 6).
-10
0
10
20
30
40
50
60
70
80
90
Burn Bulldoze Old-Burn Uniola Grass Forest
(%
cover
age/
4 m
2/t
reat
men
t)
Treatments from Ensenada mature site
Summer 2012 Winter 2012 Summer 2013
P = 1.48e-10
Figure 5: Percentage of woody vegetation coverage (±SE) among all treatments in
Ensenada mature site during the year study. P-value applies to the difference between
treatments.

26
3.3 Native grass coverage regeneration study
Forest stand comparison of the percentage of native grass coverage demonstrated a
significant difference (P = 0.00221) within the burn treatments between Ensenada (Figure 7)
and Tallaboa-Castle sites (Figure 8). Also, the time effect was significantly different (P<
3.42e-09) between the sites during the study. Among survey seasons, results showed a
significant difference (P-adj<0.0015) between summer 2012 - winter 2012 and (P-
adj<0.00001) between summer 2012 - summer 2013. Seasonal growth exhibited an increase
of an average of 10% during the winter season with some replicates showing outliers in the
30% range of grass regeneration and then, slightly declining to an average of 5% loss during
dry season 2013. When considering all treatments, grass regeneration was found to be
-20
0
20
40
60
80
100
Castle-Burn Castle-Forest
(% c
over
age/
4 m
2/t
reat
men
t)
Treatments from Tallaboa-Castle secondary site
Summer 2012 Winter 2012 Summer 2013
P = 1.74e-09
Figure 6: Percentage of woody vegetation coverage (±SE) among all treatments in
Tallaboa-Castle secondary site during the year study. P-value applies to the difference
between treatments.

27
significantly different (P < 0.00001) among them, with the Uniola grass treatment exhibiting
45% higher average regeneration and an upper level of 65%. Among the other treatments
including the Bulldozed, Burn and Forest treatments, there was no mean significant growth,
with some particular plots representing outliers with 15 to 60% of understory native grass
coverage. However, the Old-Burn treatment exhibited a significant average increase of 15%
with an upper level reaching 20%, which suggests successional progress after the wildfire
that took place two years prior the study. In contrast, seasonality did not represent a
significant influence in the regeneration process of native grasses among treatments in
Ensenada (P-adj > 0.05). Native grass colonization was facilitated by both burning and
bulldozing disturbances.
-20
-10
0
10
20
30
40
50
60
Burn Bulldoze Old-Burn Uniola Grass Forest (% c
over
age/
4 m
2/t
reat
men
t)
Treatments from Ensenada mature site
Summer 2012 Winter 2012 Summer 2013
Figure 7: Percentage of Uniola grass coverage (±SE) among all treatments in
Ensenada matured site during the year study. P-value applies to the difference
between treatments.
P < 0.00001

28
Time, however, played a significant role (P = 1.97e-06) in native grass extents across
the Castle-Burn treatment in the Tallaboa-Castle site, where grass presence increased from
0% in summer 2012 to an average of 10% in winter 2012 and to 20% in summer 2013. Only
one of the Castle-Forest plots displayed high density of Uniola virgata patches ranging from
35% to 70% presence, while the Castle-Burn treatment showed one outlier with 60% grass
coverage. Despite seasonal soil moisture and temperature, time played an important role (P-
adj < 0.05) in native grass colonization for the Castle-Burn treatment and not for the Castle-
Forest treatment, where a significant difference (P = 0.0042) was observed between dry
season summer 2012 and dry season summer 2013 (Figure 8).
-10
-5
0
5
10
15
20
25
30
Castle-Burn Castle-Forest (% c
over
age/
4 m
2/t
reat
men
t)
Treatments from Tallaboa-Castle secondary site
Summer 2012 Winter 2012 Summer 2013
P = 0.0042
Figure 8: Percentage of Uniola grass coverage (±SE) among all treatments in Tallaboa-
Castle secondary site during the year study. P-value applies to the difference between
treatments.

29
3.4 Seasonal understory regeneration
Temporal and spatial variations in the understory were assessed by monitoring the
progress of seedlings, sprouts, and species richness with the percentage of vegetative
coverage regeneration and grass regeneration. Multivariate analysis of variance was
conducted on a broader scope to understand the dynamics involved between abiotic
parameters and vegetative succession (seedlings-sprouts-species richness). Significant
temporal variation (P = 0.0019), variation among burn plots (P = 9.89e-06), and soil
temperature flux influence (P = 0.0142) were found for the Ensenada site. When same
procedure was conducted considering all treatments, significant difference (P = 8.89e-13)
was observed among them, while abiotic factors such as soil moisture and temperature fluxes
had an equally significant influence (P = 0.0091) on seedlings-sprouts-species richness
regeneration after disturbance.
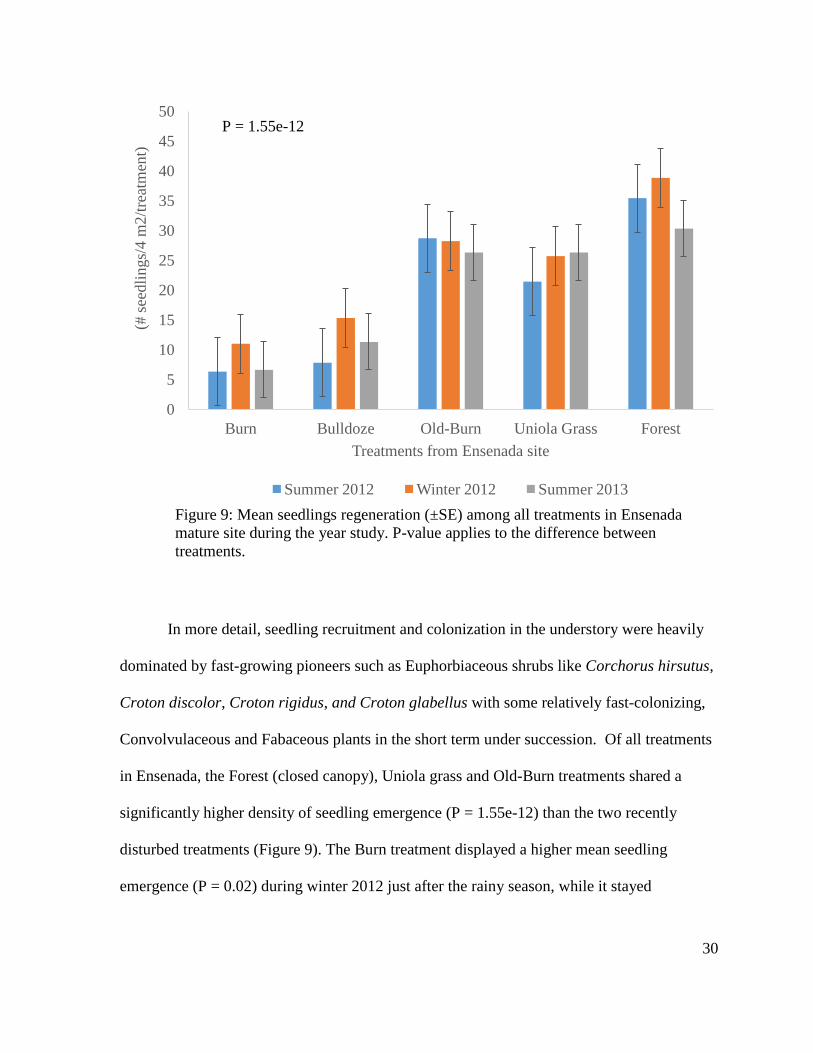
30
In more detail, seedling recruitment and colonization in the understory were heavily
dominated by fast-growing pioneers such as Euphorbiaceous shrubs like Corchorus hirsutus,
Croton discolor, Croton rigidus, and Croton glabellus with some relatively fast-colonizing,
Convolvulaceous and Fabaceous plants in the short term under succession. Of all treatments
in Ensenada, the Forest (closed canopy), Uniola grass and Old-Burn treatments shared a
significantly higher density of seedling emergence (P = 1.55e-12) than the two recently
disturbed treatments (Figure 9). The Burn treatment displayed a higher mean seedling
emergence (P = 0.02) during winter 2012 just after the rainy season, while it stayed
0
5
10
15
20
25
30
35
40
45
50
Burn Bulldoze Old-Burn Uniola Grass Forest
(# s
eedli
ngs/
4 m
2/t
reat
men
t)
Treatments from Ensenada site
Summer 2012 Winter 2012 Summer 2013
Figure 9: Mean seedlings regeneration (±SE) among all treatments in Ensenada
mature site during the year study. P-value applies to the difference between
treatments.
P = 1.55e-12

31
particularly low for summer 2012 and 2013. No significant time effect was observed on
seedling emergence among treatments. Soil moisture and temperature fluxes within
treatments reflected significant variation (P = 7.57e-10), with Old-Burn, Uniola grass and
Forest representing the most successful treatments in terms of establishment of seedlings. In
the disturbed treatments, the seedling recruitment process fluctuated with time (significant
growth in summer 2012, higher in winter 2012 and significantly lower mean growth in
summer 2013).
The pattern of seedling emergence in the Tallaboa-Castle site proved that soil
moisture flux played an important role in seedling-sprouts-species richness density variation
(P = 0.004), although soil temperature fluxes did not represent an important aspect of
changes in the Castle-Burn treatment. At the treatment level, neither soil moisture or soil
temperature flux had a significant influence on the burn and forest treatments. The
differences between the Castle-Forest and Castle-Burn treatments demonstrated an evident
higher recruitment activity for the Castle-Burn over the Castle-Forest treatment (P = 0.0002).
The time phenomenon had a slight influence (P = 0.0048) on mean seedling regeneration in
the Castle-Burn treatment as well as between both treatments (P = 0.0002) during the study
(Figure 10). In terms of seasonal temperature flux over the Castle-Burn and Castle-Forest
treatments in the Tallaboa-Castle site, slightly significant changes were observed (P =
0.0023), primarily because of the canopy cover effect in the Castle-Forest treatment lowering
the soil temperature and understory regeneration covering the burned landscape during

32
summer 2013 (showing lower soil temperature [27.26 °C] than the [29.00 °C] in winter
2012).
Basal and root sprout seasonal regeneration were dominated by the intensified
reestablishment of Euphorbiaceous species such as Gymnanthes lucida, Croton discolor,
Croton flavens and Corchorus hirsutus ,as well as Burseraceous (Bursera simaruba),
0
5
10
15
20
25
30
35
40
45
50
Castle-Burn Castle-Forest
(# s
eedli
ngs/
4 m
2/t
reat
men
t)
Treatments from Tallaboa-Castle secondary sites
Summer 2012 Winter 2012 Summer 2013
P = 0.0002
Figure 10: Mean seedling regeneration (±SE) among all treatments in Tallaboa-
Castle secondary site during the year study. P-value applies to the difference
between treatments.

33
Boraginaceous (Bourreria succulenta and Bourreria baccata), Malvaceous (Melochia
tomentosa and Waltheria indica), Rutaceous (Zanthoxylum sp.), Erythroxylaceous
(Erythroxylum sp.) and Rubiaceous (Randia portorricensis) species, among other woody
sprouts. Soil moisture and temperature fluxes and time within treatments in the Ensenada site
did not appear to cause any drastic shift in sprout density regeneration during the study.
The Tallaboa-Castle site, however, showed significant influence (P =0.002) of soil
moisture flux in sprout variation in the Castle-Burn treatment. Soil temperature flux was also
found significant (P = 0.00187) with a decrease in sprout density when reaching a high
temperature of 30.55°C. Time played a significant role in sprout density variation (P = 0.006)
in the Castle-Burn treatment, displaying an increase in sprout density. There was a
difference between treatments (P < 0.00001), as time induced an important vegetative
structural shift (P = 0.001) in which was observed higher sprout density (Figure 11) in the
understory of the Castle-Burn treatment, while such a dynamic was limited in the Castle-
Forest treatment.

34
Seasonal species richness after disturbances in Ensenada showed significant variation
(P = 1.27e-06) in the Burn treatment at the end of the year cycle. With respect to time
influence on species richness, winter 2012 presented a significant (P = 0.0023) increase in
species richness with a slight reduction during the following summer 2013. When soil
moisture and temperature flux parameters were examined, both appeared to have a limited
effect (P = 0.0403) on species richness density on the burn treatment. Time did not explain
significant variation in species richness between treatments. Among all treatments there were
significant differences (P = 5.95e-10), with Bulldoze, Burn, Old-Burn and Forest treatments
reaching an equivalent range of species richness density during winter 2012, while in
-40
-20
0
20
40
60
80
100
120
Castle-Burn Castle-Forest (# s
pro
uts
/4 m
2/t
reat
men
t)
Treatments from Tallaboa-Castle secondary site
Summer 2012 Winter 2012 Summer 2013
P < 0.00001
Figure 11: Mean sprouts regeneration (±SE) among all treatments in Tallaboa-Castle
secondary site during the year study. P-value applies to the difference between
treatments.

35
summer 2013 Bulldoze, Old-Burn and Forest treatments shared the highest species richness
density. The Burn treatment showed the lowest recorded density. The Uniola grass and Burn
treatments remained slightly low in species richness during part of winter 2012 and the entire
summer 2013. Soil moisture and temperature flux parameters played a significant role (P =
0.0005) in species richness density variation in the Ensenada site treatments.
At the end of the year cycle in the Tallaboa-Castle site (Table 4), time (P = 0.005) and
soil moisture flux (P = 0.004) significantly influenced the variation in species richness
0
1
2
3
4
5
6
7
8
9
Burn Bulldoze Old-Burn Uniola Grass Forest
(# s
pec
ies/
4 m
2/t
reat
men
t)
Treatments from Ensenada mature site
Summer 2012 Winter 2012 Summer 2013
P = 5.95e-10
Figure 12: Mean species richness density (±SE) among all treatments in Ensenada
mature site during the year study. P-value applies to the difference between
treatments.

36
density for the Castle-Burn and Castle-Forest treatments alike. According to the post-fire
treatment difference, the burn treatment maintained a slight increase in species richness, but
with different species composition (mostly shrubs and Fabaceae generalist trees) than the
Castle-Forest treatment with its higher population of native trees.
3.5 Open canopy vs. closed canopy study: seedling size classes
Multivariate analysis of variance for the 5 size class distribution for the Burn
treatment in the Ensenada site (Table 1) showed no significant differences, while time (P =
0.000187), soil moisture flux (P = 0.001533) and soil temperature flux (P = 0.00946) had
0
1
2
3
4
5
6
7
Castle-Burn Castle-Forest
(# s
pec
ies/
4 m
2/p
lot)
Treatments from Tallaboa-Castle secondary sites
Summer 2012 Winter 2012 Summer 2013
Figure 13: Mean species richness density (±SE) among all treatments in Tallaboa-
Castle secondary site during the year study. P-value applies to the difference between
treatments.
P = 0.005

37
significant influence on seedling size class distribution. In Tallaboa-Castle (Table 3), a
significant difference (P = 9.95e-07) was found in the Castle-Burn treatment with the same
effect of time (P<0.00001), soil moisture (P<0.00001) and soil temperature fluxes
(P<0.00001) on seedling size class distribution. Between treatments within each site, in
addition to time, soil moisture and soil temperature fluxes, strong significant differences were
found among all treatments in the Ensenada mature site (P <0.00001), as for the Tallaboa-
Castle secondary site with previous abiotic parameters (P <0.0003).
Table 2. Seedling size class (cm height) count during summer 2012 (S-12), winter 2012 (W-
12) and summer 2013 (S-13) after disturbances in Ensenada, Guánica mature forest site.
Size
Classes
S-12
Burn
(x̄ ±SD)
Bulldoze
(x̄ ±SD)
Old-Burn
(x̄ ±SD)
Grass
(x̄ ±SD)
Forest
(x̄ ±SD)
<10 cm 4.8±4.9 5.0±3.5 7.3±6.9 16.4±18.3 29.3±28.7
10-30cm 0.8±1.3 1.8±0.9 11.9±24.1 3.3±4.6 3.6±0.9
30-50cm 0.5±2.2 0.5±1.0 4.1±6.6 1.1±2.4 0.8±0.9
50-70cm 0.0±0.0 0.5±1.0 3.4±2.9 0.3±0.6 0.3±0.5
70-90cm 0.0±0.0 0.0±0.0 0.9±1.2 0.3±0.9 0.6±0.5
W-12 Burn Bulldoze Old-Burn Grass Forest
<10 cm 5.6±4.3 9.8±7.2 11.8±9.4 13.3±12.3 26.8±31.6
10-30cm 2.1±2.1 5.0±3.5 8.8±8.9 10.3±14.6 4.7±2.8
30-50cm 2.0±2.4 0.5±1.0 5.4±8.8 1.8±4.8 1.3±0.9
50-70cm 0.8±1.0 0.0±0.0 2.0±2.8 0.3±0.9 0.3±0.7
70-90cm 0.5±0.8 0.0±0.0 0.1±0.3 0.0±0.0 0.1±0.3
S-13 Burn Bulldoze Old-Burn Grass Forest
<10 cm 3.0±4.3 5.0±2.2 6.3±7.7 22.3±25.5 26.9±34.3
10-30cm 2.0±2.4 5.8±4.6 7.8±7.5 4.2±4.4 2.8±3.2
30-50cm 1.1±1.9 0.5±1.0 8.0±17.9 0.3±1.2 0.9±0.9
50-70cm 0.4±0.7 0.0±0.0 3.2±8.9 0.0±0.0 0.0±0.0
70-90cm 0.1±0.3 0.0±0.0 0.6±2.0 0.0±0.0 0.1±0.3

38
Table 3. Seedling size class (cm height) count during summer 2012 (S-12), winter 2012 (W-
12) and summer 2013 (S-13) after disturbances in Tallaboa-Castle, Guayanilla secondary
forest site.
Size Class
S-12
Burn
(x̄ ±SD)
Forest
(x̄ ±SD)
Size Class
S-13
Burn
(x̄ ±SD)
Forest
(x̄ ±SD)
<10 cm 31.8±30.9 8.3±6.0 <10 cm 8.8±9.9 7.1±6.8
10-30cm 0.3±1.4 6.5±14.7 10-30cm 12.9±20.1 6.4±11.7
30-50cm 0.0±0.0 0.5±0.9 30-50cm 2.9±7.3 0.8±1.1
50-70cm 0.0±0.0 0.2±0.4 50-70cm 0.1±0.6 0.0±0.0
70-90cm 0.0±0.0 0.1±0.3 70-90cm 0.0±0.0 0.0±0.0
Size Class
W-12
Burn
(x̄ ±SD)
Forest
(x̄ ±SD)
<10 cm 11.8±14.3 7.1±6.0
10-30cm 13.2±14.4 7.3±13.2
30-50cm 8.7±8.4 0.8±1.2
50-70cm 3.3±5.0 0.1±0.3
70-90cm 0.8±2.0 0.1±0.3
In Ensenada (Table 2), the Uniola grass, Old-Burn and Forest treatments were
evaluated for seedling-sapling growth based on seedling size class distribution. During
summer 2012, the lowest size class was dominated by the Forest treatment followed by the
Uniola grass treatment. The next size classes (10cm - 30 cm height and 30cm -50cm height)
did not show significant increases in general, with only the Old-Burn treatment having a few
subplots displaying high numbers of seedlings in the intermediate size class. In the bigger
size classes (50cm - 70cm height and 70cm - 90 cm height), the Bulldoze treatment exhibited
the first signs of seedling growth, while Old-Burn had the greatest number of seedlings in
both categories. The Forest treatment exhibited the lowest seedling growth. After winter
2012, the Forest treatment dominated the lowest size class followed by the Uniola grass and

39
Old-Burn treatments. In the 10cm - 30cm size class, the numbers of seedlings were higher in
the Uniola grass and Old-Burn treatments, whereas among disturbed treatments, the Bulldoze
treatment had a slight advantage in seedling presence over the Burn treatment. When
examining the next three bigger classes, Old-Burn dominated the seedling count in size
classes 30cm - 50cm and 50cm - 70cm, while the Burn treatment dominated the largest size
class 70cm - 90cm. In summer 2013, the smallest size class continued to be significantly
represented in the Uniola grass treatment followed closely by the Forest treatment. The next
size class of seedling growth showed a significant increase in seedling density for the Old-
Burn treatment, followed closely by the Bulldoze and Uniola grass treatments.
4.0 Discussion
4.1 Forest cover regeneration
Since few native woody species in Puerto Rican dry forest are fire-resistant or fire-
adapted (Wolfe, 2009), previously disturbed forest patches are susceptible to native and
exotic grass dominance, providing continuous fuel loads that can trigger more wildfires
(Thaxton et al., 2012). However, our fuel load index for both sites did not demonstrate a
difference between them in burned grass biomass as a result of wildfires occurring
spontaneously on separate occasions.
Furthermore, the percentage of woody vegetation and native grass regeneration
coverage after such disturbances proved to be different between the sites and influenced by
time and soil abiotic pressures. The Tallaboa-Castle site showed a higher percentage of
regeneration in both cases (woody vegetation and native grasses) than the Ensenada site.

40
Indeed, the Castle-Burn treatment in the Tallaboa-Castle site showed higher regeneration
density than the homologous Burn treatment as well as the Bulldoze treatment in the
Ensenada site. Nonetheless, it was also found that, beyond abiotic conditions and
disturbances, woody vegetation recolonized the disturbed areas faster than native grasses in
both sites. However, within the Ensenada site treatments it is important to denote that after
three years of regeneration, understory woody coverage (seedlings and saplings) was
reaching forest control levels in the previously burned treatment, whereas in disturbed and
grass treatments the regeneration was much lower at the end of the study. In the Tallaboa-
Castle site, woody regeneration occurred faster in the burn treatment and at a significantly
higher percentage than the control forest understory. These findings show how quickly
species are able to re-establish and colonize disturbed areas between a mature dry forest site
and a secondary dry forest site. Also, native grass coverage was significantly higher in the
Old-burn and Bulldoze treatments near undisturbed grass patches. At the end of the study,
Uniola virgata had colonized the disturbed areas ten times more than the forest plots. In
contrast, the Old-Burn recovery treatment showed essentially the same grass coverage with
minimal variation through time. With respect to the time effect, native grass regeneration in
Ensenada fluctuated depending on seasonal weather, while in Tallaboa-Castle, regeneration
was influenced more by time rather than seasonal wet months. It was also observed that
changes in soil moisture flux, which is an important indicator of groundwater dynamics
(York et al., 2002; Yeh and Eltahir, 2005; Soylu et al., 2014), had significant influence on
woody regeneration but not with native grass regeneration.

41
4.2 Understory regeneration and species richness
Since species are the main point connections of ecological networks and interactions,
many ecological community structures interact in response to abiotic-biotic conditions in a
synergic sequence (Petchey et al, 2010). Previous studies in this system have suggested
significant species recovery and resiliency after 45 years of undisturbed growth, yet not all
forested patches reached the same level of resilience (Molina Colón and Lugo, 2006). A
study based on post-fire vegetation succession in Mediterranean gorse shrublands (De Luis et
al., 2006), in which the authors tested the autosuccessional hypothesis, suggested that
communities dominated by a certain species can exhibit a change in species dominance after
fire. Understanding the severity of fire and its frequency of occurrence (Keeley, 2003;
Keeley et al., 2009) within tropical dry forest or shrubland systems, both systems can be
expected to display the same successional behavior: fast-growing seeder species will exploit
the opportunity that the fire disturbance provides and dominate the landscape, changing the
plant community structure (De Luis et al., 2006; Ojeda et al., 1996).
Our study of understory regeneration and species pools serves as a post-disturbance
survey of possible outcomes in the forthcoming development of the forest stands.
Throughout the seasons, the Old-Burn and Forest plots shared the same species richness
density. Bulldoze disturbance experienced an increase in its species pool, including the
establishment of invasive grasses, which made it the most species-rich (6.8±1.3) treatment at
the end of the study (Table 2). In contrast, the Uniola grass and Burn treatments carried the
lowest species richness throughout the study. In the Tallaboa-Castle site, burn (4.4±1.4) and
forest (4.5±1.4) treatments maintained equivalent species richness consistently throughout

42
the study with no significant changes per season (Table 3). In fact, our results showed that
the Ensenada site (protected mature forest nature prior to disturbances) had a larger species
pool than the Tallaboa-Castle site (unprotected secondary forest nature prior to disturbances).
Moreover, Ensenada’s seedling recruitment among burned and bulldozed treatments
displayed similar variation of gain and loss throughout the seasons, which suggests no
difference between these types of disturbance in terms of seasonal variation of seedling
density. Considering all five treatments, the Old-Burn, Forest and Uniola grass proved to be
similar among them in seedling density as well as with the Bulldoze disturbance treatment,
yet the Burn treatment had the lowest seedling density in general. Overall, the results suggest
that fire represented a more challenging disturbance for seedling establishment during the
successional process than the Bulldoze disturbance.
Sprout density only showed a slight difference between Old-Burn and Burn
treatments, whereas there were no differences among the other treatments. This evidence
suggests the recovery process from wildfires in the understory takes more time than recovery
from bulldoze disturbance. In addition, time, soil moisture and soil temperature fluxes were
pressure factors for seedling regeneration in disturbed treatments rather than in undisturbed
treatments. Canopy cover had a large influence on soil temperature and moisture fluctuations
and their impact on seedling establishment in disturbed areas in comparison to the forested
areas. However, in the Tallaboa-Castle site, the soil moisture and temperature did not show
an influence on seedling recruitment in disturbed areas in comparison to the forested areas.
Possible explanations for these differences between sites might depend on other factors such

43
as precipitation, evapotranspiration in exposed soil after disturbances and canopy structure
between the forested areas in each site.
4.3 Seedling size class
Guánica’s tropical dry forest, among other homologous dry forests, represents one of
the most endangered ecosystems in the tropics (Stoner and Azofeifa, 2009), which depends
greatly on understory resilience and recovery after major disturbances from human activities
resulting in habitat fragmentation. Disturbances are complex events that, depending on the
ecosystem and its species in the understory (Buma and Wessman, 2012), can have significant
impact on seedling growth and survival. Size class distribution with diameter and height
measurements of structural groups proved to be useful in understanding interspecific and/or
intraspecific interactions between trees within stands (Jimu et al., 2012). Although the
diameter at ground height did not represent a significant measurement with respect to post-
disturbance seedling establishment, seasonal height measurement of tagged seedlings
throughout the treatments provided significant evidence about how seedling development
varied seasonally. Seedling size classes between the sites’ analog treatments showed stronger
seedling growth overall in the Tallaboa-Castle burn treatment than Burn and Bulldoze
treatments in Ensenada. The conditions in the Tallaboa-Castle site landscape were
substantially more eroded and exposed than the Ensenada site, and even the soil moisture
flux was lower in Tallaboa-Castle than Ensenada. However, the Tallaboa-Castle secondary
site proved to be more active in the regeneration and growth of seedlings and sprouts than the
Ensenada site, although the Ensenada disturbed treatments showed higher species richness.

44
These findings provide an interesting perspective on mature versus secondary site post-
disturbance vegetation dynamics. According to Chinea and Helmer (2003), secondary forest
can be impacted in species richness after land use change or disturbance (as in this study),
but not necessarily in species composition of the forest. We observed strong input of
generalist Fabaceae trees, Euphorbiaceae shrubs and grasses from unburned areas in the
vicinity of the secondary forest region that was burned. Both the Ensenada and Tallaboa-
Castle sites are examples of tropical dry forest systems with resilient vegetation that can
respond quickly to disturbances (Van Bloem et al, 2007), but the community dynamics may
be influenced more by the site exposure to seed rain and intrusion of opportunist vegetation
that does not contribute to the local species richness.
Potential questions may arise regarding the seedling size class seasonal variations and
their implications for future management strategies and forest resilience. Our seedling
maturity and mortality density study provides important knowledge about those seasonal
variations. According to this study, seedling maturity and mortality through time seems to
conform to expectations for this dry forest succession.
5.0 Conclusion
Historically, the protected forests in the western region of Guánica’s dry forest
biosphere have been composed of large sections of open forest stands, and reduced, heavily
fragmented closed forest stands, in which a small number of large fragments represent most
of the closed forest areas (Ramjohn et al., 2012), including the Ensenada site. Despite the
different influence that large or small forest patches can have on biodiversity and species

45
composition (Murphy et al., 1995; Laurance et al., 2011; Ramjohn et al., 2012), periodic
anthropogenic disturbances can disrupt the process of regeneration and regrowth of forest
stands. Theoretically, regenerating forest stands should be resilient enough and follow the
historically contingent community assemblage regardless of environmental conditions.
However, this study was set for the purpose of examining short-term recovery and understory
regeneration after disturbance which makes it difficult to determine if either a historical
contingency or deterministic assembly scenario happens in the system in just a year cycle
(Fukami, 2010). Nevertheless, the questions answered in the study provide a first insight of
post-disturbance secondary succession. In fact, with available nutrient resources, open
landscape, limited inter- specific interaction and invasion, it seems more likely to observe a
historically contingent case scenario occurring in the system studied (Fukami, 2010). Since
community assembly begins with a disturbance, it will require future monitoring of the
system to evaluate if one or the other scenario applies. Additionally, patch size or
aggregation of forest patches, rates of population density and dispersal of native tree species
or invasive species into these forests can be signs of deterministic assembly in the future. The
major concern is the need for more native trees species dispersal from the undisturbed
patches of the forest adjacent to the disturbed areas, which could be difficult to achieve if
environmental or external conditions change the immigration history.

46
6.0 References
Brandeis, T. J. (2003). Puerto Rico’s forest inventory: adapting the forest inventory and
analysis program to a Caribbean island. J. For. 1011: 8–13.
Brandeis, T.J., Helmer, E.H. and Oswalt, S.N. (2007). The Status of Puerto Rico’s Forests,
2003. USDA Forest Service Southern Research Station, Asheville, North Carolina, 75
pp.
Brandeis, T. J., Woodall, C. W. (2008). Assessment of Forest Fuel Loadings in Puerto Rico
and the US Virgin Islands. Ambio. 37 (7–8). 557- 562.
Beisner, B. E., Haydon, D. T., Cuddington, K. (2003). Alternative stable states in ecology.
Frontiers in Ecology and the Environment. 1 (7): 376–382.
Brooks, M.L., C.M. D’Antonio, D.M. Richardson, J.B. Grace, J.E. Keeley, J.M. Ditomaso, R.J.
Hobbs, M. Pellant, and D. Pyke. (2004). Effects of invasive alien plants on fire regimes.
Bioscience 54 (7): 677-688.
Buma, B., Wessman, C.A. (2012). Differential species responses to compounded
pertubations and implications for landscape heterogeneity and resilience. Forest
Ecology and Management. 266: 25-33.
Cabin, R. J., Weller, S. G., Lorence, D. H., Flynn, T. W., Sakai, A. K., Sandquist, D.,
Hadway, L. J. (2000). Effects of long-term ungulate exclusion and recent alien
species control on the preservation and restoration of a Hawaiian tropical dry forest.
Conservation Biology. 14: 439–453.
Ceccon E., Huante P., Ricón E. (2006). Abiotic factors influencing Tropical Dry Forests
Regeneration. Brazilian Archives of Biology and Technology. 49 (2). 305-312.
Chase J. M. (2003). Community assembly: when does history matters? Oecologia. 136: 489-
495.

47
Chinea, J. D., Helmer, E. H. (2003). Diversity and composition of tropical secondary forests
recovering from large-scale clearing: results from the 1900 inventory in Puerto Rico.
Forest Ecology and Management. 180: 227-240.
D'Antonio C., Meyerson L.A. (2002). Exotic plant species as problems and solutions in
ecological restoration: a synthesis. Restor Ecol. 10:703–713.
De Luis, M., Baeza, M.J., Raventós, J., González-Hidalgo, J.C. (2004a). Fuel characteristics
and fire behavior in mature Mediterranean gorse shrublands. Int. J. Wildl. Fire 13:
79–87.
De Luis, M., Raventós, J., González-Hidalgo, J.C. (2006). Post-fire vegetation succession in
Mediterranean gorse shrublands. Acta Oecol. 30: 54-61.
Dickinson M.B., Johnson E.A. (2001). Fire effects on trees. In ‘Forest Fires: Behavior and
Ecological Effects’. (Eds EA Johnson, K Miyanishi): 477–525. (Academic Press: San
Francisco, CA)
Didham, R. K., Tylianakis, J. M., Hutshinson, M. A., Ewers, R. M., Gemmell, N. J. (2005)
Are invasive species the drivers of ecological change? Trends in Ecology and
Evolution. 20: 470–474.
Dodson, E. K., Peterson, D. W. (2010). Dry coniferous forest restoration and understory
plant diversity: The importance of community heterogeneity and the scale of
observation. Forest Ecology and Management. 260: 1702–1707.
Dunphy, B. K., Murphy, P. G., Lugo, A. E. (2000). The tendency for trees to be multiple-
stemmed in tropical and subtropical dry forests: studies of Guánica forest, Puerto
Rico. Trop. Ecol. 412. 161–167.
Dwyer, E., Gregoire, J.M. and Malingreau, J.P. (1998). A global analysis of vegetation fires
using satellite images: spatial and temporal dynamics. Ambio 27, 175–181.
Dwyer, E., Pinnock, S., Gregoire, J.M. and Pereira, J.M.C. (2000). Global spatial and
temporal distribution of vegetation fire as determined from satellite observations. Int.
J. Remote Sens. 21: 1289–1302.

48
Eaton, J. M., Lawrence, D. (2006). Woody debris stocks and fluxes during succession in a
dry tropical forest. 2006. Forest Ecology and Management. 232: 46–55.
Edwards, E. J., Benham, D. G., Marland, L. A., Fitter, A. H. (2004). Root production is
determined by radiation flux in a temperate grassland community. Global Change
Biology. 10: 209–227.
Ehrenfeld, J. G. (2003). Effects of exotic plant invasions on soil nutrient cycling processes.
Ecosystems. 6: 503–523.
Erickson, H.E., Davidson, E.A., Keller, M., (2002). Former land-use and tree species
affect nitrogen oxide emissions from a tropical dry forest. Oecologia. 130: 297–
308.
Ewel, J.J., Whitmore, J.L. (1973). The Ecological Life Zones of Puerto Rico and the US
Virgin Islands. USDA Forest Service, Institute of Tropical Forestry, Rio Piedras,
Puerto Rico.
Franco, P.A., Weaver, P.L., Eggen-McIntosh, S. (1997). Forest resources of Puerto Rico,
1990. Resource Bulletin SRS-22. USDA Forest Service, Southern Research Station.
Folke, C., Carpenter, S., Walker, B., Scheffer, M., Elmqvist, T., Gunderson, L., Holling, C.
S. (2004). Regime shifts, resilience, and biodiversity in ecosystem management.
Annual Review of Ecology, Evolution, and Systematics. 35: 557–581.
Foster, D. R., Fluet, M., Boose, E. R. (1999). Human or natural disturbance: landscape-
scale dynamics of the tropical forests of Puerto Rico. Ecol. Appl. 92: 555–572.
Freitas, S. R., Hawbaker, T.J., Metzger, J.P. (2010). Effects of roads, topography, and land use
on forest cover dynamics in the Brazilian Atlantic Forest. Forest Ecology and
Management. 259: 410-417.
Fukami, T. (2005). Integrating internal and external dispersal in metacommunity assembly:
preliminary theoretical analyses. Ecological Research. 20: 623-631.

49
Gallagher R. V., Beaumont L. J., Hughes L., Leishman M. R. (2010). Evidence of climatic
niche and biome shifts between native ranges in plant species introduced to Australia.
Journal of Ecology. 98: 790-799.
Garcia-Cancel, J. G. (2013). Effects of native and non-native grasses on woody species
regeneration in a Puerto Rican subtropical dry forest. M.S. Thesis. University of
Puerto Rico, Mayagüez.
Griscom, H. P., Ashton, M. S. (2011). Restoration of dry tropical forests in Central America:
A review of pattern and process. Forest Ecology and Management. 261: 1564-1579.
Holling, C. S. (1973). Resilience and stability of ecological systems. Annual Review of
Ecological Systems 4: 1–24.
Hoffmann, W.A., Jaconis,S.Y., Geiger, E.L., Gotsch, S.G., Franco, A.C. (2012).
Fuels or microclimate? Understanding the drivers of fire feedbacks at savanna-forest
boundaries. Austral Ecology, 37:634-643.
Hoffman, W.A., Orthen, B., Nascimento, P.K.V. (2003). Comparative fire ecology of
tropical savanna and forest trees. Functional Ecology. 17: 720-726.
Howell, D. C. (2007). Statistical methods for psychology (6th Ed.). Belmont, CA: Thomson
Wadsworth.
Jimu, L., Ngoroyemoto, N., Mujuru, L. (2012). Structural diversity and regeneration of the
endangered Prunus africana (Rosaceae) in Zimbabwe. Afr. J. Ecol. 51: 102-110.
Keeley J.E., Lubin D., Fotheringham C.J. (2003). Fire and grazing impacts on plant diversity
and alien plant invasions in the southern Sierra Nevada. Ecological Applications 13:
1355–1374.
Keeley, J. E. (2009). Fire intensity, fire severity and burn severity: a brief review and
suggested usage. International Journal of Wildland Fire. 18: 116-126.

50
Kruskal, W. H., Wallis, W. A. (1952). Use of Ranks in One-Criterion Variance Analysis.
Journal of the American Statistical Association. 47 (260): 583-621.
Laurance, W.F., Camargo, J.L.C., Luizão, R.C.C., Laurance, S.G., Pimm, S.L., Bruna, E.M.,
Stouffer, P.C., Williamson, G.B., Benítez-Malvido, J., Vasconcelos, H.L., Van
Houtan, K.S., Zartmank, C.E., Boyle, S.A., Didham, R.K., Andrade, A., Lovejoy,
T.E. (2011). The fate of Amazonian forest fragments: a 32-year investigation. Biol.
Conserv. 144: 56–67.
Lebrija-Trejos, E., Meave, J. A., Poorter, L., Pérez-García, E. A., Bongers, F. (2010).
Pathways, mechanisms and predictability of vegetation change during tropical dry
forest succession. Perspectives in Plant Ecology, Evolution and Systematics. 12: 267-
275.
Leibold, M. A., Holyoak, M., Mouquet, N., Amarasekare, P., Chase, J. M., Hoopes, M. F.,
Holt, R. D., Shurin, J. B., Law, R., Tilman, D., Loreau, M. and Gonzalez, A. (2004).
The metacommunity concept: a framework for multi-scale community ecology.
Ecology Letters, 7: 601-613.
Lewontin, R. C. (1969). The meaning of stability. Brookhaven Symposia in Biology 22: 13-
23.
Li, Y., Xu, M., Zou, X., Shi, P. and Zhang, Y. (2005). Comparing soil organic carbon
dynamics in plantation and secondary forest in wet tropics in Puerto Rico. Global
Change Biol. 11: 239–248.
Litton, C. M., Sandquist, D. R., Cordell, S. (2008). A non-native invasive grass increases
soil carbon flux in a Hawaiian tropical dry forest. Global Change Biology. 14: 726–
739.
Lugo, A. E., Domínguez Cristóbal, C., Santos, A., Torres Morales, E. (1999). Nutrient return
and accumulation in litter of a secondary forest in the coffee region of Puerto Rico.
Acta Cien. 131: 43–74.

51
Lugo, A.E., Medina, E., Trejo-Torres, J.C., Helmer, E. (2006). Botanical and ecological
basis for resilience of Antillean dry forests. In: Pennington, R.T., Lewis, G.P.,
Ratter, J.A. (Eds.), Neotropical Savannas and Seasonally Dry Forests: Plant
Diversity, Biogeography and Conservation. CRC Press, Boca Raton, pp. 359–381.
Maunder, M., Leiva, A., Santiago-Valentín, E., Stevenson, D. W., Acevedo-Rodríguez, P.,
Meerow, A. W., Mejía, M., Clubbe, C. Francisco-Ortega, J. (2008). Plant
Conservation in the Caribbean Island Biodiversity Hotspot. Botanical Review. 74:
197-207.
May, R. M. (1977). Thresholds and breakpoints in ecosystems with a multiplicity of states.
Nature. 267 (5628): 471–477.
Molina Colón S., Lugo A. E. (2006). Recovery of a Subtropical Dry Forest After
Abandonment of Different Land Uses. Biotropica. 38 (3): 354-364.
Monsegur Rivera, O.A. (2009). Vascular Flora of the Guánica Dry Forest. MS Thesis.
University of Puerto Rico, Mayagüez, Puerto Rico.
Murphy, P. G., Lugo, A. E. (1986). Structure and Biomass of a Subtropical Dry Forest in
Puerto Rico. Biotropica. 8 (2): 89-96.
Murphy, P.G., Lugo, A.E., Murphy, A.J., Nepstad, D.C. (1995). The dry forests of Puerto
Rico’s south coast. In: Lugo, A.E., Lowe, C. (Eds.), Tropical Forests: Management
and Ecology. Springer-Verlag, New York, pp. 178–209.
Ojeda, F., Maranon, T., Arroyo, J. (1996). Postfire regeneration of a Mediterranean
heathland in southern Spain. Int. J. Wildl. Fire. 6: 191–198.
Otterstrom, S. M., and Schwartz, M. W. (2006). Responses to fire in selected tropical dry
forest trees. Biotropica 38:592-598.
Padmanaba M., Sheil D. (2014). Spread of the invasive alien species Piper aduncum via
logging roads in Borneo. Tropical Conservation Science. 7 (1): 35-44.

52
Parrotta, J.A. 2000. Leucaena leucocephala (Lam.) de Wit Leucaena, tantan. In: J.K. Francis
and C.A. Lowe, eds. Bioecología de árboles nativos y exóticos de Puerto Rico y las
Indias Occidentales. General Technical Report IITF-15. U.S. Department of
Agriculture, Forest Service, International Institute of Tropical Forestry, Río Piedras,
PR. p. 308-316.
Petchey, O. L., Brose, U., Rall, B. C. (2010). Predicting the effects of temperature on food
web connectance. Philosophical Transactions of The Royal Society B. 365: 2081-
2091.
Quesada, M., Sanchez-Azofeifa, G.A., Alvarez-Anorve, M., Stoner, K.E., Avila-Cabadilla,
L., Calvo-Alvarado, J., Castillo, A., Espirito-Santo, M.M., Fagundes, M., Fernandes,
G.W., Gamon, J., Lopezaraiza-Mikel, M., Lawrence, D., Morellato, L.P.C., Powers,
J.S., Neves, F.D., Rosas-Guerrero, V., Sayago, R., Sanchez-Montoya,G. (2009).
Succession and management of tropical dry forests in the Americas: review and new
perspectives. For. Ecol. Manage. 258, 1014–1024.
Ramjohn, I.A. (2004). The Role of Disturbed Caribbean Dry Forest Fragments in the
Survival of Native Plant Diversity. PhD Dissertation. Michigan State University,
East Lansing, Michigan.
Ramjohn, I. A., Murphy, P. G., Burton, T. M., Lugo, A. E. (2012). Survival and rebound of
Antillean dry forests: Role of forest fragments. Forest Ecology and Management. 284:
124-132.
R Development Core Team. (2011). R: A Language and Environment for Statistical
Computing. R Foundation for Statistical Computing, Vienna, Austria, ISBN 3-
900051-07-0, <http://www.R-project.org/>.
Ray, G.J., Brown, B.J. (1994). Seed ecology of woody species in a Caribbean dry forest.
Restor. Ecol. 2, 156–163.
Ray, G.J., Brown, B. (1995). The structure of five successional stands in a subtropical
dry forest, St. John, US Virgin Islands. Caribb. J. Sci. 31: 212–222.

53
Ray, G.J. and B.J. Brown. (1995). Restoring Caribbean dry forests: evaluation of tree
propagation techniques. Restoration Ecology. 3(2):86-94.
Rivera-Batiz, F. and Santiago, C. (1996). Island Paradox: Puerto Rico in the 1990’s. Russel
Sage Foundation, New York.
Robbins, A. M., Eckelmann, C-M., Quiñones, M. (2008). Forest Fires in the Insular
Caribbean. Ambio. 8, (7–8): 526-534.
Royston, P. (1995). Remark AS R94: A remark on Algorithm AS 181: The W test for
normality. Applied Statistics, 44: 547–551.
Rudel, T. K., M. Perez-Lugo, and H. Zichal. 2000. When fields revert to forests:
development and spontaneous reforestation in post-war Puerto Rico. Professional
Geographer, 52: 386-397.
Saha, S., and H. F. Howe. 2003. Species composition and fire in a dry deciduous forest.
Ecology. 84:3118-3123.
Santiago Garcia, R.J., S. Molina Colón, P. Sollins, and S.J. Van Bloem. (2008). The role of
nurse trees in mitigating fire effects on tropical dry forest restoration: a case study.
Ambio 37: 604-608.
Shiels, A. B., West, C. A., Weiss, L., Klawinski, P. D., Walker, L. R. (2008). Soil factors
predict initial plant colonization on Puerto Rican landslides. Plant Ecol. 195:165–
178.
Shinneman, D. J., Cornett, M. W., Palik, B. J. (2010). Simulating restoration strategies for a
southern boreal forest landscape with complex land ownership patterns. Forest
Ecology and Management. 259: 446–458.

54
Soylu, M. E., Kucharik, C. J., Loheide II, S. P. (2014). Influence of groundwater on plant
water use and productivity: Development of an integrated ecosystem – Variably
saturated soil water flow model. Agricultural and Forest Meteorology. 189-190: 198-
210.
Stoner, K. E., Sánchez-Azofeifa, G. A. (2009). Ecology and regeneration of tropical dry
forests in the Americas: Implications for management. Forest Ecology and
Management. 258: 903-906.
Tabachnick, B. G., Fidell, L. S. (2007). Using multivariate statistics (5th Ed.). Boston: Allyn
and Bacon.
Thaxton, J.M., Platt, W.J., (2006). Small-scale fuel variation alters fire intensity and shrub
abundance in a pine savanna. Ecology 87: 1131–1337.
Thaxton, J. M., Van Bloem S. J., Whitmire, S. (2012). Fuel conditions associated with native
and exotic grasses in a subtropical dry forest in Puerto Rico. Fire Ecology. 8 (3): 9-
17.
Thompson, I., Mackey, B., McNulty, S., Mosseler, A. (2009). Forest Resilience,
Biodiversity, and Climate Change. A synthesis of the biodiversity/resilience/stability
relationship in forest ecosystems. Secretariat of the Convention on Biological
Diversity, Montreal. Technical Series No. 43. 67 pp.
Tolhurst, K.G. (1995). Fire from a flora, fauna and soil perspective: sensible heat
measurement. CALM Science. 4 (Suppl.): 45–88.
Van Bloem S. J., Lugo A. E., Murphy, P G. (2004). Tropical Dry Forests. Elsevier Ltd. 1-10.
Van Bloem, Skip J., Murphy, Peter G., Lugo, Ariel E., Ostertag, Rebecca, Rivera Costa,
Maria, Ruiz Bernard, Ivelisse, Molina Colón, Sandra, and Canals Mora, Miguel.
(2005). The Influence of Hurricane Winds on Caribbean Dry Forest Structure and
Nutrient Pools. Biotropica. 37 (4): 571-583.

55
Van Bloem S. J., Murphy P. G., Lugo A. E. (2007). A link between hurricane-induced tree
sprouting, high stem density and short canopy in tropical dry forest. Tree Physiology.
27: 475-480.
Verhoef, H. A., Morin, P. J. (2010). Community Ecology: Processes, Models, Applications
(1st Ed.). New York: Oxford University Press.
Wolfe, B.T., (2008). Post-Fire Regeneration in Subtropical Dry Forest of Puerto Rico. M.S.
Thesis. University of Puerto Rico, Mayagüez.
Wolfe B. T., Van Bloem S. J. (2012). Subtropical dry forest regeneration in grass-invaded
areas of Puerto Rico: Understanding why Leucaena leucocephala dominates and
native species fail. Forest Ecology and Management. 267: 253-261.
Weaver, P.L. (1998). Hurricane effects and long-term recovery in a subtropical rain forest.
In: Forest Biodiversity in North, Central and South America and the Caribbean.
Research and Monitoring. Dallmeier, F. and Comiskey, J.A. (eds). Parthenon
Publishing Group, Washington, DC, pp. 249–270.
Yeh, P.J.F., Eltahir, E.A.B. (2005). Representation of water table dynamics in a land surface
scheme, part I: model development. Climate 18:1861–1880.
York, J.P., Person, M., Gutowski, W.J., Winter, T.C.( 2002). Putting aquifers into
atmospheric simulation models: an example from Mill Creek Watershed, north
eastern Kansas. Adv. Water Resour. 25: 221–238.

56
Chapter III
Spread of a native grass Uniola virgata, an invasive grass Megathyrsus maximus and a
generalist tree Leucaena leucocephala along a bulldozed trail through subtropical semi-
deciduous dry forest.
Abstract
Tropical deciduous dry forests are among the most threatened terrestrial ecosystems
in the world. There is an increasing demand for canopy restoration in dry forest ecosystems
impacted by anthropogenic disturbances that can counter the biotic barriers exerted by
invasive vegetation on native tree development. A key question is the comparative speed at
which a non-native invasive grass (Megathyrsus maximus), naturalized generalist tree
(Leucaena leucocephala) and native grass (Uniola virgata) can colonize a disturbed (i.e.,
bulldozed and burned) tract of dry forest. The main objective of this study was to determine
whether bulldozing disturbance facilitates plant invasion that alters post-disturbance natural
regeneration of the system, or if the forest’s natural resilience provides resistance to those
biotic pressures.
Results of a post-bulldozing species colonization study in a mature dry forest site
showed strong dispersal of pioneer species, both non-native invasive grasses and naturalized
generalist trees, from the road (i.e. the starting point of the bulldozed path) into the forest.
These findings provide insight into how time influences understory regeneration dynamics
and invasion after wildfire and bulldozing disturbances. Even though dry forests are capable
of absorbing human activities that mimic natural disturbances, the frequency and intensity of
those disturbances will have major impacts on the functionality of the system. Anthropogenic

57
disturbances can impact forest biodiversity and community assemblages by facilitating the
intrusion of non-native vegetation that outcompetes local flora for the open niche.
Furthermore, community assembly dynamics in such dry forest sites demand further long-
term research on community-level interactions between soil nutrient dynamics and plant
community response to such disturbances. This is particularly relevant since an alternative
stable state is unreachable for this forest type, and the persistent damage caused by such
anthropogenic disturbances could degrade the integrity of the ecosystem to the point of
shifting towards less favorable systems (i.e. chaparral, savanna or grassland).
Keywords
Tropical Dry Forest · Grass invasions · Tree invasions · Anthropogenic disturbances ·
Occurrence probability · Plant community interactions · Understory regeneration
1.0 Introduction
An important goal of invasion ecology is to understand how biological invasions are
driven by anthropogenic alterations of native habitats (Vitousek et al., 1996; Wilcove et al.,
1998; Mack, 2003; Lockwood et al., 2007; Gooden and French, 2014). Anthropogenic
disturbances such as fires and bulldozing represent complex deterministic processes,
involving an extensive combination of factors such as species competition for available
resources, changes to ecosystem integrity that can shift community assemblages during
regeneration and environmental constraints that influence above- and belowground
dynamics. These deterministic processes can contribute to the propagation of invasive

58
species in the short-term, but if further disturbances are minimal, the spread of an invader
within an affected ecosystem may ultimately decline over the long term (Rew and Johnson,
2010). On the other hand, given our limited information about overlapping effects of niche
alteration and plant invasion on native communities (Didham et al., 2007; Gooden and
French, 2014), it is difficult to ascertain whether drivers of habitat modification might
interact in such a way as to further facilitate invasions during post-disturbance recovery in
the system (Didham et al., 2007).
In tropical dry forest systems, grass invasion and other habitat modifications driven
by anthropogenic disturbances can yield widely disparate outcomes depending on the degree
of degradation and frequency of perturbation. Since tropical dry forests are among the most
threatened ecosystems in the world (Janzen, 1988; Mooney et al., 1995; Molina and Lugo,
2006), ecological restoration and natural regeneration after disturbances represent primary
management goals for forest resilience and functionality (Griscom and Ashton, 2011). For
instance, in cases where the invasive species of interest are highly fire-prone grasses that
increase the frequency and intensity of wildfires in invaded dry forests, management
recommendations typically focus on controlling fuel loads with prescribed fires in
combination with a restoration of tree canopy cover (Thaxton et al., 2012). However, by
contributing to the spread of invasive grasses (Chinea 2002), additional disturbances such as
bulldozing may further exacerbate the threat of wildfire in dry forests.
Understanding the movement of dominant plant species after such disturbances is therefore
an important component of dry forest management. In Puerto Rico, there is a need to
visualize the distribution and movement of a native grass, Uniola virgata, and a naturalized

59
legume tree, Leucaena leucocephala, that each may be useful in promoting the regeneration
of native tree species and contrast their dispersal patterns with a fire-prone invasive grass,
Megathyrsus maximus, after recent disturbance.
The focus of the study was to examine the spread of these three species after a 7-m
wide track was bulldozed through a mature, native-dominated dry forest in the western
section of Guánica Commonwealth Forest (Puerto Rico). The aim of the study was to
determine whether or not invasion was occurring because of bulldozing, and if so, at what
rate, in order to understand how environmental and biological interactions affect each
species’ distribution throughout the disturbed area and inside the mature forest
2.0 Materials and Methods
2.1 Study site
The sites for this study were located near road PR-325 (~13- 69 meters above sea
level) in the Ensenada sector of Guánica Commonwealth Forest – a ~714-ha section of semi-
deciduous dry forest (Figure 1). Annual rainfall in Ensenada averages 860 mm/yr and is
highly seasonal with 6 dry months (December-March and June-July) split by two wet seasons
from April to May and August to November (Murphy and Lugo, 1986). Atmospheric
temperature averages 25.1°C with low variation annually.

60
Figure 1: Map of the 555-meter long bulldozed trail (brown line) in Ensenada site with
mature forest stand nature, Guánica PR
2.2 Experimental design and measurement of bulldozed trail cross-transects
A 555-m long x 7.3-m wide (on average) bulldozer trail was created by the Puerto
Rico Electric Power Authority in an attempt to extinguish a fire created by a downed high-
voltage power line inside the forest. The bulldozed trail begins at the roadside entrance on
PR-325 and ends 5 meters past the high voltage power-line that triggered the wildfire. The
trail has an uphill gradient. The first 401 m are through mature dry forest; the latter portion of

61
the trail crosses over mature forest (154 m) areas that had burned. Megathyrsus maximus had
already been established along the edge of PR-325 when the trail was created. The burned
area had been a mix of native tree species with a discontinuous canopy and U. virgata. A few
mature L. leucocephala were established in scattered locations within the mature forest. We
established transect lines across the trail at 15-m intervals (n=38). For each transect line, we
identified the species of sprouts or seedlings present at intercept points located every 20-cm
along the line.
2.3 Statistical analysis
To model the relationship between regeneration and invasion, we developed logistic
regression models under binary history of detection (detected = 1, not detected = 0) for each
species, whether there is any species interaction occurring among those species under study
in the bulldoze trail. . Each dependent variable was tested against a particular independent
variable or set of interacting variables that were considered to have a possible impact on
species distribution. This analysis produced a total of 7 models each for the two grasses, M.
maximus (Table 1) and U. virgata (Table 2), and 9 models for the generalist tree l.
leucocephala (Table 3). In all three cases, the set of models included a null model with zero
effect of covariates. For the analysis, we used the statistical software R-Gui 3.0.0 (R
Development Core Team, 2012).
We developed models, via logistic regression, that estimated the probability of
occurrence of M. maximus (Table 1), U. virgata (Table 2) and l. leucocephala (Table 3)
based on distance along the 555-m bulldozed trail. The models were intended to depict the

62
spread of each species from its presumed point of origin at either end of the trail; for the
invasive grass M. maximus and the naturalized tree species L. leucocephala, we measured
distance from PR-325 (i.e. the start of the bulldozed trail), while for the native grass U.
virgata, we measured distance from the trail’s end in the area of burned forest. Distance
from road (Quadratic effect) was designed to address the projected future movement of the
species after the year of study, seasons involved in the survey periods which represent
periods of ending dry months (Summer 2012), precipitation months (Winter 2012), and
ending in dry months cycle (Summer 2013). The parameter of time takes into account the
sampling periods without the influence of seasonality. The fire region parameter interacting
in 154 m of the bulldozed trail addressed the possible influence of fire and bulldozing
disturbances on species distribution.

63
Table 1. Descriptions and expected direction of a priori occurrence effect model for invasive
grasses from the 555-meter bulldozed cross-transect in the Guánica Dry Forest reserve
section of Ensenada, Guánica. All models are set under 95% confidence interval.
Hypothesis Model Structure of model Expected results
No detection or abiotic
influence
p (.) β0 -
Negative influence of trail
distance from roads
(Dist_z) β0+β1 (Dist_z) β1< 0
Positive influence of
distance quadratic effect
(Dist_z)2
β0+β1 (Dist_z)+ β2
(Dist_z)2
β1< 0, β2> 0
Negative influence of
distance and time
(Sea) β0+β1 (Dist_z)+ β2 (Sea-
S1)+ β3 (Sea-W)+ β4
(Sea-S2)
β1< 0, β2< 0, β3≤
0, β4≤ 0
Negative influence of
distance and wet season
(Wet) β0+β1 (Dist_z)+ β2 (Wet) β1< 0, β2≤ 0
Negative influence of
native grass presence
(NatG)
β0+ β1 (NatG) β1< 0
Negative influence of fire
region and time
(Fire) β0+ β1 (Fire) + β2 (Sea-
S1)+ β3 (Sea-W)+ β4
(Sea-S2)
β1< 0, β2< 0, β3≤
0, β4≤ 0

64
Table 2. Descriptions and expected direction of a priori occurrence effect model for Uniola
virgata and other native grasses from the 555-meter bulldozed cross-transect in the Guánica
Dry Forest reserve section of Ensenada, Guánica. All models are set under 95% confidence
interval.
Hypothesis Model Structure of model Expected results
No detection or abiotic
influence
p (.) β0 -
Positive influence of trail
distance from roads
(Dist_z) β0+β1 (Dist_z) β1> 0
Positive influence of
distance quadratic effect
(Dist_z)2
β0+β1 (Dist_z)+ β2
(Dist_z)2
β1> 0, β2< 0
Negative influence of
distance and time
(Sea) β0+β1 (Dist_z)+ β2 (Sea-
S1)+ β3 (Sea-W)+ β4
(Sea-S2)
β1> 0, β2< 0, β3≤
0, β4≤ 0
Positive influence of
distance and wet season
(Wet) β0+β1 (Dist_z)+ β2 (Wet) β1> 0, β2> 0
Negative influence of
invasive grass presence
(InvG)
β0+ β1 (InvG) β1≤ 0
Positive influence of fire
region and time
(Fire) β0+ β1 (Fire) + β2 (Sea-
S1)+ β3 (Sea-W)+ β4
(Sea-S2)
β1> 0, β2< 0, β3≤
0, β4≤ 0

65
Table 3. Descriptions and expected direction of a priori distribution effect model for
Leucaena leucocephala introduced tree species from the 555-meter bulldozed cross-transect
in the Guánica Dry Forest reserve section of Ensenada, Guánica. All models are set under
95% confidence interval.
Hypothesis Model Structure of model Expected results
No detection or abiotic
influence
p (.) β0 -
Negative influence of trail
distance from roads
(Dist_z) β0+β1 (Dist_z) β1< 0
Positive influence of
distance quadratic effect
(Dist_z)2
β0+β1 (Dist_z)+ β2 (Dist_z)2
β1< 0, β2> 0
Negative influence of
distance and time
(Sea) β0+β1 (Dist_z)+ β2 (Sea-
S1)+ β3 (Sea-W)+ β4 (Sea-
S2)
β1< 0, β2< 0, β3≥
0, β4≤ 0
Negative influence of
distance and dry season
(Dry) β0+β1 (Dist_z)+ β2 (Dry) β1< 0, β2< 0
Positive influence of
distance and wet season
(Wet) β0+β1 (Dist_z)+ β2 (Wet) β1> 0, β2> 0
Positive influence of
invasive grasses presence
(InvG)
β0+ β1 (InvG) β1> 0
Negative influence of
native grasses
(NatG) β0 + β1 (NatG) β1< 0
Negative influence of fire
region and time
(Fire) β0+ β1 (Fire) + β2 (Sea-S1)+
β3 (Sea-W)+ β4 (Sea-S2)
β1< 0, β2< 0, β3≥
0, β4≤ 0

66
3.0 Results
3.1 Study of invasive-native grasses and generalist tree species successional niche
colonization after bulldozing disturbance
Species establishment along the bulldozed trail in the Ensenada dry forest site showed
intense colonization of the non-native invasive grass M. maximus and the pioneer naturalized
tree L. leucocephala from the bulldozer entrance (0 m) to 255 m in patches and individuals
found as much as 435 m further along the bulldozed trail inside the forest after one year
(Figure 2). The native grass Uniola virgata spread from the end of the bulldozed trail cross-
transect (555 meters) to beyond the fire region and into the area where M. maximus and L.
leucocephala established (Figure 2).

67
Figure 2: Post-bulldozing dispersal pattern of M. maximus, L. leucocephala and U. virgata
along the bulldozed trail during (A) summer 2012, (B) winter 2012 and (C) summer 2013.
Note: Locations where none of the three species occurred were dominated only by sprouting
trees other than L. leucocephala. Gaps presence between distances represent areas dominated
only by sprouting trees.

68
R² = 0.3599 R² = 0.0848
R² = 0.4211
0
1
0
1
15 65 115 165 215 265 315 365 415 465 515 565 Pre
sen
ce o
f sp
eci
es
in b
ulld
oze
tra
il d
uri
ng
sum
me
r 2
01
2
Distance from Road (meters)
M. maximus U. virgata L. leucocephala Log. (M. maximus) Log. (U. virgata ) Log. (L. leucocephala )
R² = 0.209 R² = 0.4635
R² = 0.2311
0
1
0
1
15 65 115 165 215 265 315 365 415 465 515 565 Pre
sen
ce o
f sp
eci
es
in b
ulld
oze
tra
il d
uri
ng
w
inte
r 2
01
2
Distance from Road (meters)
M. maximus U. virgata L. leucocephala Log. (M. maximus) Log. (U. virgata ) Log. (L. leucocephala )
A
B

69
R² = 0.2532
R² = 0.5016 R² = 0.3464
0
1
0
1
15 65 115 165 215 265 315 365 415 465 515 565
Pre
sen
ce o
f sp
eci
es
in b
ulld
oze
d t
rail
du
rin
g su
mm
er
20
13
Distance from Roads (meters) M. maximus U. virgata L. leucocephala Log. (M. maximus) Log. (U. virgata ) Log. (L. leucocephala )
C

70
We developed occurrence probability models, via logistic regression, for invasive
grasses (Table 1), native grasses (Table 2) and introduced tree species (Table 3) along the
555-m bulldozed trail. With respect to M. maximus colonization, increasing distance (i.e.,
three-dimensional distance with elevation gradient embedded; β0+β1 (Dist_z), β1< 0) from
the roadside starting point to the end of the bulldozed trail had a negative influence on
occurrence probability (β = -1.0803, SE = 0.2788; P = 0.0001) during the year-long study
cycle (Figure 2).
Figure 3: Logistic regression of invasive grasses occurrence model (β0+β1 (Dist_z),
β1< 0) of colonization from road side to bulldoze cross-transect end region at 600
meters range.
(β = -1.0803, SE = 0.2788;
P<0.0001)

71
M. maximus presence decreased as the distance from road increased (β= -1.0071, SE
= 0.2872; P = 0.0004). Dry seasons of summer 2012 and distance effect in combination were
shown to have a negative influence on invasive grasses occurrence probability (β=-1.7721,
SE=0.6904; P = 0.0103), no significant effect of rainy season on winter 2012, and significant
negative influence again on summer 2013 (β=-0.8535, SE=0.3985; P = 0.0322). To examine
seasonal rainfall effects on invasive grass colonization, a distance and rainy season model
was developed excluding summer dry seasons influence and attested no significant influence.
The bulldozed trail crossed the region burned after the fire (β0+ β1 (Fire), β1< 0), fire region
(β=-1.7444, SE=0.7818; P = 0.0257) and dry season (β= -1.5539, SE= 0.6427; P = 0.0156)
effect on invasive grasses colonization proved to negatively influence invasive grasses
progress. If competition between native and invasive grasses influenced Megathyrsus
maximus spread, it could not be assessed after one year, because model combining the
presence of Uniola virgata (NatG) + distance from road was not significant (β= -0.7259, SE=
0.5124; P = 0.1565).

72
Table 4. Statistics for models of M. maximus probability of occurrence derived from cross-
transect surveys conducted along the 555-m bulldozed trail in the Guánica Dry Forest reserve
section of Ensenada. All models are set under 95% confidence interval set. (*) Preferred
model.
Models Number of
Parameter
AICc ΔAICc AICc
Weight
Cum.
Weight
Likelihood
(LL)
Null 1 129.14 21.08 0.00 1.00 -63.55
Distance 2 112.10 4.04 0.11 0.90 -53.99
Distance +
Distance2
3 113.84 5.79 0.04 1.00 -53.81
Distance + Wet
Season
3 113.51 5.45 0.05 0.95 -53.65
*Distance +
Time
4 108.05 0.00 0.80 0.80 -49.84
Fire + Time 4 121.44 13.39 0.00 1.00 -56.54
Native Presence 2 129.04 20.99 0.00 1.00 -62.47
The U. virgata occurrence model was based on the species’ colonization progress
from the end-point of the bulldozed trail (i.e., in the burned forest area) and toward its
starting point at PR-325.. Increasing distance from road side proved to be strongly significant
in positive detection of native grasses (β= 2.4745, SE= 0.4604; P = 7.68e-08) considering the
historical establishment of native grasses inside the forest prior to disturbance (Figure 3).

73
Distance from road increased (β = 3.9951, SE= 1.2652; P = 0.0015) native grasses
colonization, as Uniola re-entered or regenerated in the bulldozer trail in areas where it had
previously been established. The first dry season, in combination with the distance effect, had
a strong negative effect (β = -8.0797, SE = 2.1893; P = 0.00022) on Uniola presence, but
including the effects of subsequent seasons did not improve model predictions. However,
when dry seasons were excluded from the model, winter 2012 showed to have significant
Figure 4: Logistic regression model of native grass occurrence, based only on
distance (β0+β1 (Dist_z), β1>0).
β = 2.4745, SE = 0.4604;
P =7.68e-08

74
positive influence (β = 1.3927, SE = 0.6545; P = 0.0334) on native grass colonization,
increasing as well the effect of distance from roads (β = 2.6591, SE = 0.5028; P = 1.23e-07).
Table 5. Statistics for models of U. virgata probability of occurrence based on cross-transect
surveys conducted along the 555-m bulldozed trail in the Guánica Dry Forest reserve section
of Ensenada. All models are set under 95% confidence interval set. (*) Preferred model.
Models Number of
Parameter
AICc ΔAICc AICc
Weight
Cum.
Weight
Likelihood
(LL)
Null 1 145.74 102.73 0.00 1.00 -71.85
Distance 2 83.20 40.19 0.00 1.00 -39.55
Distance +
Distance2
3 81.96 38.95 0.00 1.00 -37.87
Distance + Wet
Season
3 80.35 37.34 0.00 1.00 -37.07
*Distance +
Time
4 43.01 0.00 1.00 1.00 -17.32
Fire + Time 4 87.80 44.79 0.00 1.00 -39.72
Invasive
Presence
2 145.64 102.63 0.00 1.00 70.77
Increasing distance from the road decreased (β = -1.1257, SE = 0.2607; P = 1.58e-05)
on L. leucocephala occurrence over the year (Figure 4).

75
The quadratic effect on the distance from road was slightly positive and significant
(β= 0.5982, SE = 0.2760; P = 0.0302) which implies that L. leucocephala projected
movement in the bulldoze trail will continue effectively in the long-run. A model
representing L. leucocephala sharing the same location as invasive grasses had a positive
influence on probability of occurrence (β = 1.2730, SE = 0.4545; P = 0.0051), while such
interactions with native grasses was showed to be slightly negative (β = -0.9514, SE = .4812;
P = 0.0480). Dry seasons effect in combination with distance from road appeared not to favor
the spread of Leucaena in the short-run (β = -1.1443, SE = 0.4899; P = 0.0195), having the
Figure 5: Logistic regression model of L. leucocephala occurrence, based only on
distance (β0+β1 (Dist_z), β1<0).
(β = -1.1257, SE = 0.2607;
P = 1.58e-05)

76
same influence than the bulldoze trail distance to travel. However, after the rainy and mild
temperatures in winter 2012, were shown to have a significant positive effect (β = 1.1443, SE
= 0.4819; P = 0.0195) on L. leucocephala movement deep into the forest through the
bulldozed trail. When dry and wet seasons were combined within distance from road model,
only distance from road and summer 2012 had a significant negative influence (β = -1.6867,
SE = 0.6611; P = 0.0107) on L. leucocephala dispersal. Preliminary observation of fire
region and time effect model on L. leucocephala occurrence probability showed a negative
effect of post-fire conditions (β = -1.2993, SE = 0.6035; P = 0.0313).
Table 6. Statistics for models of L. leucocephala probability of occurrence based on cross-
transect surveys conducted along the 555- m bulldozed trail in the Guánica Dry Forest
reserve section of Ensenada. All models are set under 95% confidence interval set. (*)
Preferred model.
Models Number of
Parameter
AICc ΔAICc AICc
Weight
Cum.
Weight
Likelihood
(LL)
Null 1 144.23 30.61 0.00 1.00 -71.10
Distance 2 122.33 8.72 0.01 1.00 -59.11
Distance +
Distance2
3 119.73 6.12 0.04 0.99 -56.76
Distance + Wet
Season
3 118.77 5.15 0.07 0.95 -56.28
*Distance +
Time
4 113.61 0.00 0.88 0.88 -52.62
Fire + Time 4 134.73 21.12 0.00 1.00 -67.14
Invasive grass
Presence
2 142.01 28.39 0.00 1.00 -68.95
Native grass
Presence
2 144.23 30.61 0.00 1.00 -71.10

77
4.0 Discussion
Historically, native grasses have thrived in this tropical dry forest without
aggressively competing with native woody vegetation inside the undisturbed forest. Once a
major disturbance occurs, the landscape is vulnerable to invasion by highly competitive non-
native grasses and introduced tree species. More importantly, we observed some of the
mechanisms that promote plant invasions. For example, the bulldozed trail started directly on
a local road with heavy traffic (Freitas et al., 2010) and substantial roadside presence of M.
maximus. This generated high propagule pressure with respect to this grass, which is known
to be an aggressive invader of tropical dry forests due to its tolerance of drought and fire,
abundant wind-dispersed seeds, and ability to establish quickly in a variety of conditions,
especially in disturbed sites. Indeed, the likelihood of invasion by M. maximus increases
greatly in disturbed areas where little competition is offered by the remaining local
vegetation (Santiago et al., 2008).
We assume that M. maximus colonized the bulldozed trail from road PR-325 since it
was absent inside the forest for the first few months after the fire caused by the downed
power-line. . At any rate, the probability of finding M. maximus at a location along the
bulldozed trail declined from 70% near the trail’s starting point to less than 20% in the forest
interior, when it appears to have been confronted by biotic constraints such as woody sprout
regeneration and native grasses. According to the AICc Weight analysis for M. maximus, U.
virgata and L. leucocephala movement, the [Distance + Time] model was the best-
performing model for estimating the probability of species occurrence along the bulldozed
trail. However, even if this model worked according to expectations, the study time window

78
was too short to observe significant presence of invasive grasses inside the fire region, of
Melinis repens, although M. maximus managed to spread 360 m along the bulldozed trail,
and into the mature forest, by the end of the study. Over the same timeframe, U. virgata
managed to colonize and spread evenly from the end of the fire region to the mid-section of
the bulldozed trail, just 165 m from the road. Since this native species was already
established in patches throughout the forest and alongside the bulldozed trail, U. virgata had
the opportunity to colonize or sprou from multiple locations along the disturbed area and
build new colonies quickly. In just about a year of study, U. virgata regenerated quickly from
stolon residues that survived the fire or from seed dispersal into the post-bulldozed trail, and
colonized more efficiently than M. maximus in the fire region due to its historical presence in
the surrounding undisturbed forest. However, U. virgata declined in presence abruptly near
the mid-section of the bulldozed trail, which was instead dominated by woody sprouts.
Leucaena leucocephala colonized the first 45 m of the bulldozed trail from the roadside
during summer 2012, and then spread at at the same rapid pace, and colonized the same
areas, as M. maximus, . By the end of the study, L. leucocephala individuals were present
even at 435 m away from the trail’s start.
5.0 Conclusions
The spread of Leucaena leucocephala this deep into the forest region demonstrated
how surprisingly quick an introduced generalist tree species could colonize the open niche in
just one year. This is particularly important since native tree species struggle to re-colonize
open niches dominated by grasses, since they tend to grow slowly and require more energy

79
from the system (Ramjohn et al., 2012). Our study was consistent with previous research,
which demonstrated that Leucaena leucocephala (Parrotta, 2000) individuals can establish
themselves relatively quickly in disturbed sites shared by invasive grasses such as M.
maximus, based on an outstanding growth rate and low mortality rate (Wolfe and Van Bloem,
2012). Thus, if this pioneer generalist tree manages to establish itself before grasses, then
native tree seedlings will have a better chance of regenerating in the same niche (Santiago-
Garcia et al., 2008). The value of L. leucocephala in Puerto Rican dry forest stands goes
beyond competing with grasses to overcome hysteresis and fixating nitrogen in the soil
(Erickson et al., 2002). This species can provide an important pathway in re-establishing
native trees effectively in future management purposes (Ramjohn et al., 2012). Effective
forest management practices can greatly facilitate conservation initiatives when plant
invasions, native plant regeneration and fire regime interactions are inter-connected aspects
of the overall ecosystem dynamics (Brooks et al., 2004).
6.0 References
Brooks, M.L., C.M. D’Antonio, D.M. Richardson, J.B. Grace, J.E. Keeley, J.M. Ditomaso, R.J.
Hobbs, M. Pellant, and D. Pyke. ( 2004). Effects of invasive alien plants on fire regimes.
Bioscience 54 (7): 677-688.
Buma, B., Wessman, C.A. (2012). Differential species responses to compounded
perturbations and implications for landscape heterogeneity and resilience. Forest
Ecology and Management. 266: 25-33.

80
Cabin, R. J., Weller, S. G., Lorence, D. H., Flynn, T. W., Sakai, A. K., Sandquist, D.,
Hadway, L. J. (2000). Effects of long-term ungulate exclusion and recent alien
species control on the preservation and restoration of a Hawaiian tropical dry forest.
Conservation Biology. 14: 439–453.
Cove, M. V., Niva, L. M., Jackson, V. L. (2012). Use of probability of detection when
conducting analyses of surveys of mesopredators: A case study from the Ozark
highlands of Missouri. The Southwestern Naturalist. 57 (3): 257-261.
D'Antonio C., Meyerson L.A. (2002). Exotic plant species as problems and solutions in
ecological restoration: a synthesis. Restor Ecol. 10:703–713.
D’Antonio, C.M., Vitousek, P.M. (1992). Biological invasions, the grass-fire cycle and
global change. Annual Review of Ecology and Systematics. 23: 63-87.
Didham, R.K., Tylianakis, J.M., Gemmell, N.J., Rand, T.A., Ewers, R.M. (2007). Interactive
effects of habitat modification and species invasion on native species decline. Trends
in Ecology & Evolution, 22, 489–496.
Erickson, H.E., Davidson, E.A., Keller, M. (2002). Former land-use and tree species affect
nitrogen oxide emissions from a tropical dry forest. Oecologia 130, 297–308.
Fajardo, L. (2007). Bases ecológicas para la restauración de bosques secos tropicales
en la Península de Macanao, Isla de Margarita. Centro de Ecología. Instituto
Venezolano de Investigaciones Científicas, Caracas, Venezuela, p. 183.
Folke, C., Carpenter, S., Walker, B., Scheffer, M., Elmqvist, T., Gunderson, L., Holling, C.
S. (2004). Regime shifts, resilience, and biodiversity in ecosystem management.
Annual Review of Ecology, Evolution, and Systematics. 35: 557–581.
Foster, D. R., Fluet, M., Boose, E. R. (1999). Human or natural disturbance: landscape-
scale dynamics of the tropical forests of Puerto Rico. Ecol. Appl. 92: 555–572.

81
Francis J.K. and Parrotta J.A. (2006). Vegetation response to grazing and planting of Leucaena
leucocephala in a Urochloa maximum-dominated grassland in Puerto Rico. Caribbean
Journal of Science, 42 (1): 67-74.
Fukami, T. and Nakajima, M. (2011). Community assembly: alternative stable states or
alternative transient states? Ecology Letters 14: 973-984.
Gallagher R. V., Beaumont L. J., Hughes L., Leishman M. R. (2010). Evidence of climatic
niche and biome shifts between native ranges in plant species introduced to Australia.
Journal of Ecology. 98: 790-799.
Garcia-Cancel, J. G. (2013). Effects of native and non-native grasses on woody species
regeneration in a Puerto Rican subtropical dry forest. M.S. Thesis. University of
Puerto Rico, Mayagüez.
Gooden, B., French, K. (2014). Non-interactive effects of plant invasion and landscape
modification on native communities. Diversity and Distributions. 20: 626-639.
Griscom, H.P., Ashton, M.S. (2011). Restoration of dry tropical forests in Central
America: a review of pattern and process. Forest Ecology and Management 261:
1564-1579.
Hoffmann, W.A., Jaconis,S.Y, Geiger, E.L., Gotsch, S.G., Franco, A.C. (2012).
Fuels or microclimate? Understanding the drivers of fire feedbacks at savanna-forest
boundaries. Austral Ecology, 37:634-643.
Hoffman, W.A., Orthen, B., Nascimento, P.K.V. (2003). Comparative fire ecology of
tropical savanna and forest trees. Functional Ecology. 17: 720-726.
Janzen, D.H. (1988b). Tropical dry forests: the most endangered major tropical
ecosystem. In: Wilson, E.O., Peter, F.M. (Eds.), Biodiversity. National Academy of
Science/Smithsonian Institution, Washington DC. pp 130 - 137.

82
Lebrija-Trejos, E., Bongers, F., Pérez-García, E.A., Meave, J.A. (2008). Successional
change and resilience of a very dry tropical deciduous forest following shifting
agriculture. Biotropica. 40: 422 - 431.
Lebrija-Trejos, E., Meave, J. A., Poorter, L., Pérez-García, E. A., Bongers, F. (2010).
Pathways, mechanisms and predictability of vegetation change during tropical dry
forest succession. Perspectives in Plant Ecology, Evolution and Systematics. 12: 267-
275.
Lugo A.E. 2004. The outcome of alien tree invasions in Puerto Rico. Frontiers in Ecology
Environment 2(5): 265- 273.
Mack, M. C., D’Antonio, C. M. (2003). Exotic grasses alter controls over soil nitrogen
dynamcis in a Hawaiian woodland. Ecological Applications. 13 (1): 154-166.
Molina Colón S., Lugo A. E. (2006). Recovery of a Subtropical Dry Forest After
Abandonment of Different Land Uses. Biotropica. 38 (3): 354-364.
Murphy, P.G., A.E. Lugo. (1986). Structure and biomass of a subtropical dry forest in
Puerto Rico. Biotropica 18: 89-96.
Parrotta, J. A. 1995. Influence of overstory composition on understory colonization by native
species in plantations on a degraded tropical site. Journal of Vegetation Science 6:
627-636.
Parrotta, J. A. (2000). Leucaena leucocephala (Lam.) de Wit. Leguminosae (Mimosoideae).
In J. K. Francis and C. A. Lowe, (Eds.). Silvics of native and exotic trees of Puerto
Rico and the Caribbean islands, pp. 306–316. USDA Forest Service, International
Institute of Tropical Forestry, Gen.Tech. Rep. IITF-15. R´ıo Piedras, Puerto Rico.
Quesada, M., Sanchez-Azofeifa, G.A., Alvarez-Anorve, M., Stoner, K.E., Avila-Cabadilla,
L., Calvo-Alvarado, J., Castillo, A., Espirito-Santo, M.M., Fagundes, M., Fernandes,
G.W., Gamon, J., Lopezaraiza-Mikel, M., Lawrence, D., Morellato, L.P.C., Powers,
J.S., Neves, F.D., Rosas-Guerrero, V., Sayago, R., Sanchez-Montoya,G. (2009).
Succession and management of tropical dry forests in the Americas: review and new
perspectives. For. Ecol. Manage. 258, 1014–1024.

83
Ray, G.J., Brown, B.J. (1994). Seed ecology of woody species in a Caribbean dry forest.
Restor. Ecol. 2, 156–163.
Ray, G.J., Brown, B. (1995). The structure of five successional stands in a subtropical
dry forest, St. John, US Virgin Islands. Caribbean Journal of Science. 31: 212–222.
Ramjohn, I.A. (2004). The Role of Disturbed Caribbean Dry Forest Fragments in the
Survival of Native Plant Diversity. PhD Dissertation. Michigan State University,
East Lansing, Michigan.
Ramjohn, I. A., Murphy, P. G., Burton, T. M., Lugo, A. E. (2012). Survival and rebound of
Antillean dry forests: Role of forest fragments. Forest Ecology and Management. 284:
124-132.
Rew, L. J., Johnson, M. P. (2010). Reviewing the Role of Wildfire on the Occurrence
and Spread of Invasive Plant Species in Wildland Areas of the Intermountain Western
United States. Invasive Plant Science and Management. 3 (4): 347-364.
R Development Core Team. (2011). R: A Language and Environment for Statistical
Computing. R Foundation for Statistical Computing, Vienna, Austria, ISBN 3-
900051-07-0, <http://www.R-project.org/>.
Santiago Garcia, R.J., S. Molina Colón, P. Sollins, and S.J. Van Bloem. 2008. The role of nurse
trees in mitigating fire effects on tropical dry forest restoration: a case study. Ambio. 37:
604-608.
Tanentzap, A. J., Bazely, D. R., Williams, P. A., Hoogensen, G. (2009). A Human Security
Framework for the Management of invasive Nonindigenous Plants. Invasive Plant
Science and Management. 2 (2): 99-109.
Thaxton, J. M., Van Bloem S. J., Whitmire, S. (2012). Fuel conditions associated with native
and exotic grasses in a subtropical dry forest in Puerto Rico. Fire Ecology. 8 (3): 9-
17.
Van Bloem S. J., Lugo A. E., Murphy, P G. (2004). Tropical Dry Forests. Elsevier Ltd. 1-10.

84
Van Bloem, Skip J., Murphy, Peter G., Lugo, Ariel E., Ostertag, Rebecca, Rivera Costa,
Maria, Ruiz Bernard, Ivelisse, Molina Colón, Sandra, and Canals Mora, Miguel.
(2005). The Influence of Hurricane Winds on Caribbean Dry Forest Structure and
Nutrient Pools. Biotropica. 37 (4): 571-583.
Van Bloem S. J., Murphy P. G., Lugo, A. E. (2007). A link between hurricane-induced tree
sprouting, high stem density and short canopy in tropical dry forest. Tree Physiology.
27: 475-480.
Vitousek, P.M., Dantonio, C.M., Loope, L.L., Westbrooks, R. (1996). Biological invasions
as global environmental change. American Scientist. 84: 468–478.
Wilcove, D.S., Rothstein, D., Dubow, J., Phillips, A., Losos, E. (1998). Quantifying threats
to imperiled species in the United States. BioScience. 48: 607–615.
Wolfe, B.T. (2008). Post-Fire Regeneration in Subtropical Dry Forest of Puerto Rico. M.S.
Thesis. University of Puerto Rico, Mayagüez.
Wolfe B. T., Van Bloem S. J. (2012). Subtropical dry forest regeneration in grass-invaded
areas of Puerto Rico: Understanding why Leucaena leucocephala dominates and
native species fail. Forest Ecology and Management. 267: 253-261.