,382 jlggc
TRANSCRIPT

Supplementary Methods
Bioinformatics. The literature, FlyBase50 and an in situ hybridisation database41 were searched for genes
expressed selectively or exclusively in oenocytes. In addition, 24 lacZ and GAL4 enhancer traps reported to
be expressed in oenocytes were rescreened and the insertion sites of 15 showing bona fide oenocyte
expression were sequenced. This combined approach yielded 51 oenocyte genes (Supplementary Table 1).
The predicted functions of Drosophila genes/proteins expressed in oenocytes but with no detailed annotated
function available from Flybase were deduced from the highest-scoring matches obtained by TBLASTN and
PSI-BLAST searches of mammalian genomes. Reciprocal TBLASTN searches of the Drosophila genome
were also performed, using human protein sequences obtained from the NCBI database. This also revealed
that the Drosophila genome encodes at least one orthologue of each core enzyme in the mitochondrial and
peroxisomal β–oxidation pathways.
Drosophila strains and larval analysis. All crosses were raised at 25ºC using a standard cornmeal/agar diet
(1% autolysed yeast, 5.8% cornmeal, 5% glucose, 0.6% agar) unless stated otherwise. UAS-slifanti, UAS-
PTEN, UAS-TSC1+2, UAS-bmm, UAS-Lsd2, ppl-GAL4 and Lsp2-GAL4 were as described19-21, 31, 51, 52. ppl-
GAL4 activity is not detected in larval oenocytes. For GAL80ts experiments, specimens were raised at 25°C
until 96 hr AEL and then switched to 29°C. Cyp4g1 mutants were generated by imprecise excision of two
viable P elements: 5-SZ-3132 (insertion at X:326709, release 4.2.1, from Szeged Stock Centre) and NP2199
(insertion at X:327565, from GAL4 Enhancer Trap Insertion Database) using P-element transposase. DNA
sequencing indicates that 419 bp (X:326282..326701) are deleted in Δ1-3, ∼5.8 kb (X:321802..327565) in
Cyp4g1Δ4 and ∼1.8 kb (X:325767..327565) in Cyp4g1Δ14-9. Cyp4g1Δ4 and Cyp4g1Δ14-9 retain ∼3.3 kb and
∼300 bp of P-element respectively and both complement Exp68-36-2, sc8 and Df(1)svr indicating that flanking
genes are not deleted. Both alleles are protein-null as they remove the transcription start site and either some
or all of the Cyp4g1 coding sequence. For phenotypic analysis, Cyp4g1 alleles were balanced with FM7c,
P{GAL4-Kr.C}DC1, P{UAS-GFP.S65T}DC5.
In fasting experiments, larvae were transferred from grapejuice/agar (25% red grape juice, 1.25%
sucrose, 2.5% agar) supplemented with live yeast paste and placed onto PBS-moistened filter paper for 12-
14hr. For BO>rpr larvae, L1 animals hatching during a 3hr window were raised at 25 ºC and staged
according to elapsed time and morphological criteria53. Ecdysone and 20-hydroxyecdysone were diluted in
yeast paste54 and added to grapejuice/agar (<60hr AEL) or cornmeal/agar (>60hr AEL) with daily
SUPPLEMENTARY INFORMATION
doi: 10.1038/nature05382
1www.nature.com/naturewww.nature.com/nature 1

replacement for continuous exposure, or added to grapejuice/agar for a 6hr period prior to the L2/L3 moult
for transient exposure54. Mouth hook contractions in solid agar were measured as described32.
Construction of BO-Gal4 and svp-Gal80 transgenes. A variant of pGawB (Ref. 55) with the EcoRI site
destroyed, was digested with BamHI, Klenow filled and then digested with BstZ17 I. The resulting 3.5 kb
fragment containing the P-transposase basal promoter and GAL4 was inserted into pCaSpeR-4 (Ref. 56) cut
with Xho I, 2-base pair filled-in with Klenow and then HpaI cut to form pC4G4, an enhancer vector with
unique cloning sites for EcoR I, Kpn I, Sac II, Sfi I and BamH I. To make pBO-GAL4, the 7.6 kb EcoRI
fragment from pBO-C4PLZ containing an enhancer active, in embryos, exclusively in oenocytes57, was
inserted into the EcoRI site of pC4G4. To construct svp-GAL80, a ~150 bp PstI-BamHI fragment of hsp70
basal promoter from pWHZ vector was cloned into pCaSpeR-3 (Ref. 56) to produce pC3hsp70. A PstI
fragment from ey-Gal80 containing hsp70, Gal80 and a polyA site58 was inserted into pC3hsp70 to create
pC3G80 vector, that contains a Sac II unique site appropriate for cloning enhancer elements. A 3 kb EcoRI
Klenow-filled fragment from svp containing the oenocyte enhancer (3R: 8099915..8102523, release 4.2.1, in
pCHAB, gift from R. Schultz) was then inserted into SacII linearised, T4 DNA polymerase blunted pC3G80,
forming svp-GAL80.
In situ hybridization, histochemical and antibody staining. Embryo and larval in situ hybridizations,
immunostaining and ß-galactosidase detection were performed as described previously59. Hnf4 and CG11151
probes were prepared from SD08077 and LP03652 respectively and Alas probe was synthesized as
described60. Mouse anti-Cyp4g1 (433-43-2e, gift of S. Kennel) was used at 1:1000. Methylene Blue-yeast
was prepared as described61. For unstained fat body, tissue was dissected from larvae in PBS and
photographed immediately using Differential Interference Contrast microscopy.
2www.nature.com/nature
doi: 10.1038/nature05382 SUPPLEMENTARY INFORMATION
www.nature.com/nature 2

Supplementary Figures
Supplementary Figure 1 | Oenocyte-specific ablation using the BO>rpr system. a-d, ß-galactosidase
expression pattern of sal[BO,7.6kb]>lacZ in the embryo (a) and in L3 larval tissues (b-d) indicating that
GAL4 driver activity is strong in oenocytes (a,b), weak in two symmetrical cell clusters (arrowheads) in the
CNS (c) and not detectable in the ring gland (d). e,f, The BO>rpr system specifically deletes all oenocytes,
labelled with svp-lacZ, but not lateral chordotonal organs (arrowhead in f inset). g-i, svp-GAL80 expression
is sufficient to repress sal[BO,7.6kb]GAL4 activity in oenocytes (g) but not in the eye disc (h) nor in the cell
clusters (arrowheads) of the CNS (i). j, svp-GAL80 rescues BO>rpr animals to viable adults with undersized
rough eyes (see inset).
2www.nature.com/nature
doi: 10.1038/nature05382 SUPPLEMENTARY INFORMATION
www.nature.com/nature 3

Supplementary Figure 2 | Oenocyte ablation disrupts feeding behaviour. a, Petri dish with central yeast
source. Control L2 larvae (n=35, left half) tend to remain in the yeast but BO>rpr L2 larvae (n=35, right half)
are dispersed. b, L2 BO>rpr animals undergo fewer mouth hook contractions (per 30 sec) than controls at
72 hr AEL. ** p<0.001 for BO>rpr versus UAS-rpr, *p<0.01 for fed versus starved controls.
2www.nature.com/nature
doi: 10.1038/nature05382 SUPPLEMENTARY INFORMATION
www.nature.com/nature 4
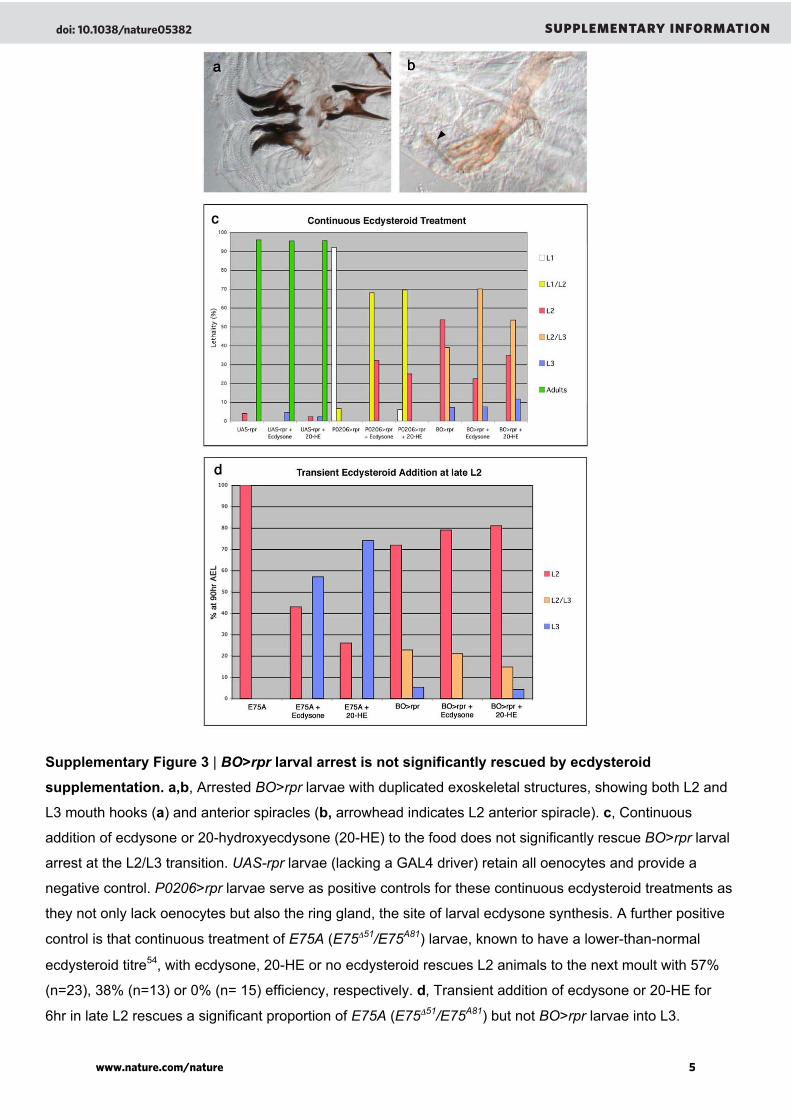
Supplementary Figure 3 | BO>rpr larval arrest is not significantly rescued by ecdysteroid
supplementation. a,b, Arrested BO>rpr larvae with duplicated exoskeletal structures, showing both L2 and
L3 mouth hooks (a) and anterior spiracles (b, arrowhead indicates L2 anterior spiracle). c, Continuous
addition of ecdysone or 20-hydroxyecdysone (20-HE) to the food does not significantly rescue BO>rpr larval
arrest at the L2/L3 transition. UAS-rpr larvae (lacking a GAL4 driver) retain all oenocytes and provide a
negative control. P0206>rpr larvae serve as positive controls for these continuous ecdysteroid treatments as
they not only lack oenocytes but also the ring gland, the site of larval ecdysone synthesis. A further positive
control is that continuous treatment of E75A (E75Δ51/E75A81) larvae, known to have a lower-than-normal
ecdysteroid titre54, with ecdysone, 20-HE or no ecdysteroid rescues L2 animals to the next moult with 57%
(n=23), 38% (n=13) or 0% (n= 15) efficiency, respectively. d, Transient addition of ecdysone or 20-HE for
6hr in late L2 rescues a significant proportion of E75A (E75Δ51/E75A81) but not BO>rpr larvae into L3.
2www.nature.com/nature
doi: 10.1038/nature05382 SUPPLEMENTARY INFORMATION
www.nature.com/nature 5

Supplementary Figure 4 | Oenocytes express many lipid-metabolising genes. Panels show the
embryonic expression patterns of 16 genes involved in lipid metabolism. All genes are expressed selectively
or exclusively in the seven segmental clusters of oenocytes (see Supplementary Table 1 for details).
2www.nature.com/nature
doi: 10.1038/nature05382 SUPPLEMENTARY INFORMATION
www.nature.com/nature 6

Supplementary Table 1 | Genes expressed in oenocytes
Gene FB/Gut expression* Evidence
• LDL Receptor familyLipophorin receptor 1 (LpR1) -(E) ALipophorin receptor 2 (LpR2) -(E) A
• Lipid-modifying Microsomal EnzymesCytochrome P450-4g1 (Cyp4g1) -(E),-(L) A, BCG6921 Fatty Acid Elongase Gut(E) ACG18609 Fatty Acid Elongase -(E) ACG9743 Stearoyl-CoA-14-desaturase FB+Gut(E) AStearoyl-CoA-9-desaturase 1 (desat1) -(E) ACG17292 Triacylglycerol lipase (TGL) Gut(E) AFatty aldehyde dehydrogenase (FALDH) Gut(E),-(L) CJuvenile hormone epoxide hydrolase 2 (Jheh2) Gut(E) A
• Other Microsomal EnzymesCytochrome P450 reductase (Cpr) -(E) ACytochrome b5 (Cyt-b5) FB+Gut(E) A
• Peroxisomal ß-Oxidation ProteinsCatalase (Cat) FB+Gut(E) ACG9527 Pristanoyl-CoA oxidase Gut(E) ACG11151, like Sterol Carrier Protein 2 Gut(E) A, CCG12428 Carnitine-O-octanoyltransferase (COT) Gut(E) A
• Other Peroxisomal ProteinsCG1444 [17ß-hydroxysteroid dehydrogenase-like] -(E) ACG17562 [3ß-hydroxysteroid dehydrogenase-like] -(E) ACG18031 [3ß-hydroxysteroid dehydrogenase-like] -(E) A
• Heme Synthetic Enzymes∂-Aminolevulinate synthase (Alas) -(E) DPorphobilinogen synthase (Pbgs) Gut(E) Al(3)02640 [Porphobilinogen deaminase] -(L) C
• Other Lipid-modifying EnzymesCG7920 4-hydroxybutyrate CoA-transferase FB+Gut(E) ACG12262 Acyl-CoA-dehydrogenase FB(E) ACG14615 Acyl-CoA-N-acyltransferase -(E) ABaldspot -(E), FB(L) C
• Nuclear Hormone ReceptorsEcdysone receptor isoform A (EcR-A) Gut(E), FB+Gut(L) C, EEcdysone-induced protein 75B (Eip75B) -(E), FB(L) CHepatocyte nuclear factor 4 (Hnf-4) Gut(E), FB+Gut(L) C, Fseven-up (svp) -(E), FB(L) C, G
• Other Transcription FactorsCG4427 [C2H2 Zinc-finger] -(E),-(L) CCG12424 [Pointed related] Gut(E) Cdiminutive (dm) Gut(E) Chindsight (hnt) Gut(E) C, HKruppel homolog 1 (Kr-h1) -(E),Gut(L) Cmirror (mirr) Gut(E),-(L) Cpipsqueak (psq) Gut(E),-(L) Cspalt (sal) Gut(E),Gut(L) C, I
• Miscellaneous ProteinsActivin-like protein 23B (Alp23B) FB(E) Aamnesiac (amn) -(E),-(L) CCG3132 ß-galactosidase -(E) A
2www.nature.com/nature
doi: 10.1038/nature05382 SUPPLEMENTARY INFORMATION
www.nature.com/nature 7

CG3328 Gut(E) ACG31764 -(E) ADopamine N acetyltransferase (Dat) Gut(E) JFerritin 2 light chain homologue (Fer2LCH) -(E),Gut(L) CFKBP59 -(E) KFurin 1 (Fur1) Gut(E) LLamin C (LamC) Gut(E) MMSP-300/nesprin -(E) ANa+ H+ exchanger 2 (NHE2) FB+Gut(E) Apericardin (prc) -(E) N
------------------------------------------------------------------------------------------------------------------------------------------------*Gene expression in the fat body (FB) and/or intestine (Gut) of embryos (E) and L3 larvae (L), where known, isindicated. Absence of detected expression in fat body and gut, where known, is also indicated (-). Gene products wereassigned to peroxisomes based on the presence of a Peroxisomal Targeting Sequence and/or data from mammalianorthologues.
A- in situ hybridization41; B- Antibody staining30 and this study; C- Enhancer trap (this study), may not recapitulate thefull endogenous expression pattern of the corresponding gene; D- in situ hybridization60; E- Antibody staining (thisstudy); F- In situ hybridization62, 63; G- Enhancer trap28; H- Antibody staining64; I- Antibody staining57; J- in situhybridization65; K- in situ hybridization66; L- in situ hybridization67; M- in situ hybridization68; N- Antibody staining69
Supplementary Notes
50. Drysdale, R. A. & Crosby, M. A. FlyBase: genes and gene models. Nucleic Acids Res 33, D390-5(2005).
51. Huang, H. et al. PTEN affects cell size, cell proliferation and apoptosis during Drosophila eyedevelopment. Development 126, 5365-72 (1999).
52. Tapon, N., Ito, N., Dickson, B. J., Treisman, J. E. & Hariharan, I. K. The Drosophila tuberoussclerosis complex gene homologs restrict cell growth and cell proliferation. Cell 105, 345-55 (2001).
53. Bodenstein, D. in Biology of Drosophila (ed. Demerec, M.) (John Wiley & Sons, New York, 1950).54. Bialecki, M., Shilton, A., Fichtenberg, C., Segraves, W. A. & Thummel, C. S. Loss of the ecdysteroid-
inducible E75A orphan nuclear receptor uncouples molting from metamorphosis in Drosophila. DevCell 3, 209-20 (2002).
55. Brand, A. H. & Perrimon, N. Targeted gene expression as a means of altering cell fates andgenerating dominant phenotypes. Development 118, 401-15 (1993).
56. Thummel, C. S. & Pirrotta, V. Technical Notes: New pCaSpeR P-element vectors. D. I. N. (1991).57. Barrio, R., de Celis, J. F., Bolshakov, S. & Kafatos, F. C. Identification of regulatory regions driving
the expression of the Drosophila spalt complex at different developmental stages. Dev Biol 215, 33-47 (1999).
58. Chotard, C., Leung, W. & Salecker, I. glial cells missing and gcm2 Cell Autonomously Regulate BothGlial and Neuronal Development in the Visual System of Drosophila. Neuron 48, 237-51 (2005).
59. Brodu, V., Elstob, P. R. & Gould, A. P. abdominal A specifies one cell type in Drosophila byregulating one principal target gene. Development 129, 2957-63 (2002).
60. Ruiz de Mena, I., Fernandez-Moreno, M. A., Bornstein, B., Kaguni, L. S. & Garesse, R. Structureand regulated expression of the delta-aminolevulinate synthase gene from Drosophila melanogaster.J Biol Chem 274, 37321-8. (1999).
61. Layalle, S., Coessens, E., Ghysen, A. & Dambly-Chaudiere, C. Smooth, a hnRNP encoding gene,controls axonal navigation in Drosophila. Genes Cells 10, 119-25 (2005).
62. Hoshizaki, D. K. et al. Embryonic fat-cell lineage in Drosophila melanogaster. Development 120,2489-99 (1994).
63. Zhong, W., Sladek, F. M. & Darnell, J. E., Jr. The expression pattern of a Drosophila homolog to themouse transcription factor HNF-4 suggests a determinative role in gut formation. Embo J 12, 537-44(1993).
64. Wilk, R., Reed, B. H., Tepass, U. & Lipshitz, H. D. The hindsight gene is required for epithelialmaintenance and differentiation of the tracheal system in Drosophila. Dev Biol 219, 183-96 (2000).
65. Hintermann, E., Grieder, N. C., Amherd, R., Brodbeck, D. & Meyer, U. A. Cloning of anarylalkylamine N-acetyltransferase (aaNAT1) from Drosophila melanogaster expressed in thenervous system and the gut. Proc Natl Acad Sci U S A 93, 12315-20 (1996).
2www.nature.com/nature
doi: 10.1038/nature05382 SUPPLEMENTARY INFORMATION
www.nature.com/nature 8

66. Zaffran, S. Molecular cloning and embryonic expression of dFKBP59, a novel Drosophila FK506-binding protein. Gene 246, 103-9 (2000).
67. Roebroek, A. J., Creemers, J. W., Pauli, I. G., Bogaert, T. & Van de Ven, W. J. Generation ofstructural and functional diversity in furin-like proteins in Drosophila melanogaster by alternativesplicing of the Dfur1 gene. Embo J 12, 1853-70 (1993).
68. Riemer, D. et al. Expression of Drosophila lamin C is developmentally regulated: analogies withvertebrate A-type lamins. J Cell Sci 108 ( Pt 10), 3189-98 (1995).
69. Chartier, A., Zaffran, S., Astier, M., Semeriva, M. & Gratecos, D. Pericardin, a Drosophila type IVcollagen-like protein is involved in the morphogenesis and maintenance of the heart epitheliumduring dorsal ectoderm closure. Development 129, 3241-53 (2002).
2www.nature.com/nature
doi: 10.1038/nature05382 SUPPLEMENTARY INFORMATION
www.nature.com/nature 9