2.evolutionary ecology, elite feasting, and the hohokam a case study from a southern arizona...
DESCRIPTION
EVOLUTIONARY ECOLOGY, ELITE FEASTING, AND THE HOHOKAM A CASE STUDYTRANSCRIPT

Society for American Archaeology
EVOLUTIONARY ECOLOGY, ELITE FEASTING, AND THE HOHOKAM: A CASE STUDY FROM ASOUTHERN ARIZONA PLATFORM MOUNDAuthor(s): Deanna N. Grimstead and Frank E. BayhamSource: American Antiquity, Vol. 75, No. 4 (October 2010), pp. 841-864Published by: Society for American ArchaeologyStable URL: http://www.jstor.org/stable/25766234 .
Accessed: 21/01/2015 07:15
Your use of the JSTOR archive indicates your acceptance of the Terms & Conditions of Use, available at .http://www.jstor.org/page/info/about/policies/terms.jsp
.JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range ofcontent in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new formsof scholarship. For more information about JSTOR, please contact [email protected].
.
Society for American Archaeology is collaborating with JSTOR to digitize, preserve and extend access toAmerican Antiquity.
http://www.jstor.org
This content downloaded from 202.41.10.30 on Wed, 21 Jan 2015 07:15:45 AMAll use subject to JSTOR Terms and Conditions

EVOLUTIONARY ECOLOGY, ELITE FEASTING, AND THE HOHOKAM: A CASE STUDY FROM A SOUTHERN ARIZONA PLATFORM MOUND
Deanna N. Grimstead and Frank E. Bayham
A number of researchers have shown that the abundance, diversity, and size of prey consumed or displayed at a feast can
be used by elites to solidify and/or aggrandize their social position. Expectations for archaeological signatures of elite
feasting?derived from ethnographic studies, archaeological research, and ecological theory?are used to assess the archae
ofaunal record from selected contexts of the Marana platform mound site, located in southern Arizona. The magnitude of work conducted in the region provides a unique opportunity to address the importance of feasting as a mechanism of power
consolidation among Hohokam elites. Here, we examine a hypothesized locus of elite feasting among the Classic period Hohokam (ca. A.D. 1250). A relatively high concentration of animal bone derived from a burned room adjacent to the Marana
platform mound was first thought to represent debris from elite feasting. Analysis reveals a proportionate taxonomic pro
file that is similar to the remainder of the community and an overwhelming abundance of small game relative to large prey.
Neither situation is consistent with elite feasting expectations. These results argue for a form of feasting among non-elites
that likely served to promote intragroup solidarity or political support within the community.
Un numero de investigadores han mostrado que la abundancia, la diversidad y el tamaho de presa son cosas que pueden ser
usadas por las elites para solidificar o agrandar su posicidn social. Aqui examinamos unfoco hipotetico delfestin de elites
entre los Hohokam del Periodo Cldsico (ca. 1250 D.C.) en el suroeste norteamericano. Dado la magnitud de trabajo hecho
en el area durante los ultimos 25 anos, existe una oportunidad unica para destacar la importancia delfestin como un mecan
ismo para la consolidacion de poder entre las elites del Hohokam. Expectativas del festin de elites se derivan de estudios etno
grdficos, investigaciones arqueoldgicas y teoria ecologica. Se usan las expectativas para comparar el registro de arqueo-fauna en unos contextos seleccionados del sitio Marana platform mound, que estd localizado en el sur de Arizona. Se encontro una
concentracion relativamente alta de huesos animates en una sala de quema al lado de platform mound. Inicialmente, se pre
sumia que esta concentracion era los restos prehistoricos delfestin de elites. Andlisis revela una semejanza taxonomica pro
porcionada al resto de la comunidad Hohokam y una abundancia significante de presa chica en relacion con la presa grande
que no es consistente con las expectativas delfestin de elites. Los resultados implican una forma delfestin de personas no
elites que servia para promover solidaridad entre grupos o como apoyo politico dentro de la comunidad.
lood consumption has long been recognized as a culturally prescribed phenomenon
JL imbued with social and political implications (van der Veen 2003). The study of feasting in the
archaeological record has been addressed by a num
ber of researchers in recent years (e.g., Bray 2003; Cameron 1995, 1998; Dietler and Hayden 2001; Fair 2000; Mills 2007; Potter 2000; Szuter 1989; Wiessner and Schiefenhovel 1996). The kinds of
behavior involved in feasting are highly variable
(Hayden 1995), but they generally involve the social
consumption of food, often in a ceremonial context,
and sometimes associated with the commemoration of an event (Dietler and Hay den 2001). Feasting is also associated with status and power. A substan
tial amount of research has explored how differen
tial access to and consumption of various animals
may reflect socioeconomic inequality and variation in status (e.g., Jackson and Scott 2003; Kelly 2001). The dynamics of a feast can also be used by elites
to solidify and aggrandize their social position (e.g.,
Hayden 1995, 1996; Pauketat et al. 2002). Apart from these general observations, however, there
remains a wide range of consumptive behaviors
Deanna N. Grimstead Department of Anthropology, University of Arizona, Emil W. Haury Building, P.O. Box 210030,
Tucson, AZ 85721-0030 ([email protected]) Frank E. Bayham Department of Anthropology, California State University, Chico, Chico, CA 95929-0400
(FB ayham @ csuchico.edu)
American Antiquity 75(4), 2010, pp. 841-864
Copyright ?2010 by the Society for American Archaeology
841
This content downloaded from 202.41.10.30 on Wed, 21 Jan 2015 07:15:45 AMAll use subject to JSTOR Terms and Conditions

842 AMERICAN ANTIQUITY [Vol. 75, No. 4,2010
associated with public and private social venues that
might qualify an event as a feast. Because of these
variations, there are few unambiguous or unique cor
relates that identify feasting in the archaeological record. This research brings together ethnographic observations of feasting, archaeological case stud ies from the Southwest, an uncontested feasting case study from Cahokia, and perspectives rooted in evolutionary ecology to derive a model of feast
ing at the Marana platform mound complex, then
applies this model to a hypothesized locus of feast
ing remnants and contexts.
The ethnographic and ethnohistoric literature record a wide range of variation linked to the wide
spread practice of feasting (e.g., Hayden 1995, 1996; Potter 2000). Elite feasting, for instance, can
be undertaken for different purposes: elites from one group may feast among themselves; they may invite elites from other groups to join; or they may hold feasts for their community. How might the
intentions, purposes, and subtleties of these differ ences be detected or understood in the absence of direct observations? Archaeological efforts to iden
tify these variables must also consider variability in space and over time. Despite these problems, detailed investigations of specific regions, time
periods, cultures, and environments associated with
feasting have begun to illuminate some common social and political dynamics that underlie this behavior (e.g., Cameron 1995; Mills 2007; Potter
1997). Because meat display and consumption is an integral part of many types of feasts, archaeo faunal data are particularly important for under
standing past behavior (e.g., Hockett 1998; Jackson and Scott 2003; Kelly 2001; Pauketat et al. 2002).
Here, we review some general expectations derived from Brian Hay den's (1995, 1996) work on feasting within prehistoric and traditional soci eties. Specific implications for faunal remains from
archaeological studies of feasting are examined as well from a case study using data from the site of Cahokia in the American Midwest (Kelly 2001; Pauketat et al. 2002). While unequivocal archaeo
logical indicators of elite feasting may be elusive, several studies have noted the disproportionate rep resentation of faunal remains as a measure of the
quantity and quality of meat consumed. Typically, the zooarchaeological perspective emphasizes the
presence or absence of higher meat-yielding species. Feasting activities that feature the use of
small animals are rarely considered. The subtleties of what food was chosen for a feast and the amount
provided, must be integral parts of our reconstruc
tion of the nuances of elite feasting in the archae
ological record. Human behavioral ecology (Smith and Winter
halder 1992) provides a complementary perspec tive to enhance archaeological inferences. Optimal foraging theory (Smith 1991; Winterhalder and Smith 1981) and signaling theory (Bliege-Bird and Smith 2005; Bliege-Bird et al. 2001; Hasson 1997; Smith 2004) offer a series of evolutionary-grounded models where the relative value of an animal, its cost of procurement, and the behavior of prestige seeking individuals are integrated to derive and outline some empirical expectations for feasting behavior. It may be possible to infuse motivational characterizations of feasting from the ethnographic record with behavioral ecological models to illu minate the roles that different animals have played in the complex theater of feasting.
This study identifies expectations derived from
ethnographic, archaeological, and evolutionary the
ory to evaluate faunal remains from an area that
may represent elite feasting among the Classic
period Hohokam in the Tucson Basin, Arizona. We
investigate whether a concentration of animal bone from a Hohokam platform mound compound rep resented elite feasting. We find that in many ways the faunal material from this unique archaeologi cal deposit is in fact similar to the archaeofaunal record from much of the surrounding community. There is one major exception however: this deposit contained an inordinate number of small mammal
bone, specifically jackrabbit and cottontails (Lago morpha). We interpret this deposit as representing an episode of feasting, but one that was not asso ciated with elite aggrandizement. Rather, the type of feasting represented here appears akin to what
Hayden (1995) has described as social solidifica tion feasting, political support feasting, and/or com
munal work party feasting. These kinds of social interactions among non-elites may have been par ticularly important in the social and political con text of the Classic period Hohokam society.
Feasting and Animal Consumption
Feasting is a cultural phenomenon deeply embed ded in the sociopolitical nature of human interac
This content downloaded from 202.41.10.30 on Wed, 21 Jan 2015 07:15:45 AMAll use subject to JSTOR Terms and Conditions

Grimstead & Bayham] EVOLUTIONARY ECOLOGY, ELITE FEASTING, AND THE HOHOKAM
Table 1. Categories and Subcategories of Feasting Types Defined by Hayden (1995, 1996).
843
Alliance and Cooperation Feasts_Economic Feasts_Diacritical Feasts
Solidarity feasts Competitive feasts Power display feasts Promotional feasts Work party feasts Status display feasts
_Reciprocal feasts_
tion. Anthropological indicators and definitions of feasts are evasive because feasts are held in a vari
ety of social contexts, take place at different scales, and the political meanings vary. Few archaeologi cal contexts have the temporal and spatial resolu tion necessary to identify ethnographically observed patterns of feasting. Hayden's (1995, 1996) review of feasting dynamics illustrates how
feasting has been investigated ethnographically. In an effort to synthesize ethnographic studies of feast
ing with previous archaeological inquiries, we
review archaeological evaluations of feasting from the perspective of Hayden's ethnographically grounded terminology.
Brian Hayden (1995,1996) has explored numer ous ethnographic examples of feasting, identifying three distinct categories of feasting, with a total of
eight subcategories (Table 1). The three major types of feasts, which are not mutually exclusive, include alliance and cooperation feasts, economic feasts, and diacritical feasts (Hayden 1996). Each category contains subtypes of feasting, each with various
sociopolitical implications (Hayden 1995:129). Alliance and cooperation feasting is a type of feast
ing in which feasting behavior reaffirms structural
solidarity along cultural, political, hierarchical, or
social boundaries. Examples include solidarity, pro motional, reciprocal, and political support feasts. Economic feasts, including competitive and work
party feasting, often involve the garnishing of non
elite labor by elites for the purpose of building large scale architectural features, collecting or harvest
ing surplus food managed or controlled by elites, etc. The results of such labor activities are often dis
played via a feast where a political competitor is in attendance; inherent within the invitation to this
adversary is the political expectation of a recipro cal feasting event. The final category, diacritical
feasting, may include whole communities or small
elite groups, but the message from the signaler is
clear: "My power and rank in this society is such that I can garner food, labor, and goods beyond your
ability to garner such items."
Hay den studied ethnographic and archaeologi cal data, revealing several archaeological indica tors that can be used to detect the presence of
competitive feasting by elites: the availability of food items that could be produced in surplus; the
presence of unique food items; the presence of spe cial or elite serving vessels and features; and the conversion of food into prestigious items could indicate feasting (Hayden 1995). In addition, spe cial feasting localities and the presence of elites are also indicators of competitive feasting (Hayden 1995). Hayden notes that not all these criteria will
always manifest themselves, especially given inher ent problems with the archaeological record, but when all these indicators are present it would cer
tainly be a strong indicator that such sociopolitical activities did occur (Hayden 1995:141).
Several Southwest researchers have attempted to identify the remnants of feasting (Cameron 1995; Dean 2001; Mills 2007; Potter 1997,2000; Szuter
1989). In southern Arizona, Dean (2001) suggested that evidence of a changing social and political landscape, where feasting served to solidify social structure and relations, may be found in several
archaeological indicators: a preponderance of high utility body parts from artiodactyls, a high fre
quency of large game compared to small game, clear visibility of the cooking or processing loca
tions, and unique ritual contexts. These indicators
align well with Hayden's (1995,1996) expectations for archaeologically visible elite competitive or
diacritical feasting: ritually linked contexts, and the association of high utility large mammal body parts and large mammals with elite contexts.
Another example of diacritical feasting comes
from the Four Corners Region of the United States. Potter (1997) evaluated the possibility that several
faunal assemblages among the Dolores Anasazi at
McPhee village, dating to A.D. 850-900, repre sented the remnants of communal ritualized feast
ing. Several of these assemblages: (1) were found
within ritually linked contexts; (2) contained a high degree of taxonomic richness; (3) displayed a high
This content downloaded from 202.41.10.30 on Wed, 21 Jan 2015 07:15:45 AMAll use subject to JSTOR Terms and Conditions

844 AMERICAN ANTIQUITY [Vol. 75, No. 4,2010
relative abundance of taxa susceptible to commu
nal hunting drives, and (4) represented a high fre
quency of fauna associated with ritual rather than
solely for consumption. This feasting example is consistent with Hayden's (1995,1996) category of diacritical feasting: (1) ritually linked fauna and
deposition of potential feasting remnants near rit ualized architecture; and (2) inferred sociopoliti cal power required to organize the labor necessary to obtain a high degree of richness or initiate a com
munal animal drive.
Elite feasting may be more readily identifiable
archaeologically because non-elite feasting may resemble day-to-day consumptive behavior. In a
comprehensive review of the ethnographic litera ture in the American Southwest, Szuter (1989) has shown agricultural activities like planting and har
vesting in the American Southwest are often asso
ciated with work party feasting. The selected animal of choice in these work party feasts appears to be
communally collected jackrabbits (Lepus sp.; Szuter 1989, 1991). Assemblages dominated by jackrabbits are common in the North American
Southwest, making the archaeological identifica tion of this type of feasting difficult, but with appro priate attention to context, associations, and
depositional history, this type of feasting may be identifiable.
In the Tonto Basin of Arizona, Cameron (1995) evaluated a potential locus of elite feasting. She identified social stratification among inhabitants of the platform mounds and surrounding commu
nity via the quality of food consumed and the rich ness of taxa consumed. Despite the visibility of social hierarchies, Cameron found it difficult to
clearly identify an elite feast from the archaeofau nal record she investigated. A feast can be a
dynamic sociopolitical event, in which the stakes and meanings may change intentionally or unin
tentionally throughout one single event or through time (Hayden 1995,1996). Cameron (1995) offered a similar explanation for a lack of archaeologically identifiable feasting events within the Tonto Basin.
A communal feast, meaning a feast in which the entire or a large section of the community partici pates, perhaps celebrating the yields of a success ful rabbit drive or harvest, may be deposited in the homes of the individual participant hunters, rather than in a large communal deposition, rendering it
archaeologically indistinguishable from day-to-day
household consumption. Similarly, elite consump tion depositions may mimic communal feasting or
feasts associated with a particular ritual or event,
making it difficult for archaeologists to extricate the two behaviors (Cameron 1995).
Feasting, in its essence, may be defined as any and all food consumption. Cameron (1995) and others (Hayden 1995, 1996; Mills 2007; Potter
2000; Szuter and Gillespie 1994) have warned about the complex and potentially confusing nature of feasting both in prehistory and modern society. This comes with a general warning: it is necessary to clearly define the type of feasting being investi
gated on a case-by-case basis and use multiple empirical indicators of the defined feast.
Elite Feasting Behavior
Among the food items displayed and consumed at,
any feast, perhaps none are laden with more sig nificance than meat. Large animals often have a pre eminent role among feasting foods. When coupled with a relatively high amount of fat, there are few food items that can match the high caloric content and palatability of meat (Speth and Spielmann 1983). Additionally, there is a rather robust ten
dency among foragers and horticulturalists to pref erentially select large prey items when there is a choice (Grayson 2001; Winterhalder and Smith
2000). Reviews of ethnographic literature and case studies have consistently found higher caloric return rates associated with the taking of large bod ied prey items, which explains this tendency (Broughton and Bayham 2003, Ugan 2005). In the
literature, these highly preferred prey items are ranked ordinally within a diet, where high prey ranks are associated with high caloric return?
generally large bodied animals, and low prey ranks are associated with low caloric returns?generally small bodied animals (Bayham 1979, Pyke et al.
1977; Ugan 2005). This reasoning is generally asso ciated with the prey choice model, under the rubric of optimal foraging theory (Broughton 1994). Under this reasoning large bodied prey are highly preferred, and within human society likely hold
high value measurable in several currencies. Thus, it is logical to ask: could the presence or absence of highly preferred animals be linked with prestige or used to understand the sociopolitical dynamics of a feast? These patterns are so compelling that it
may well be possible to use the presence or absence
This content downloaded from 202.41.10.30 on Wed, 21 Jan 2015 07:15:45 AMAll use subject to JSTOR Terms and Conditions

Grimstead & Bayham] EVOLUTIONARY ECOLOGY, ELITE FEASTING, AND THE HOHOKAM 845
of large taxa to discern the sociopolitical nature of a feast.
Certainly, large game is a highly valuable source of food, yet recent applications of the costly vari ant of signaling theory suggest that large game may be more valuable as a prestige item than as a con
sistent and economical source of food (e.g., Bliege Bird and Smith 2005; Hasson 1994,1997; Hawkes
1991; Hildebrandt and McGuire 2002, 2003; McGuire and Hildebrandt 2005; McGuire et al.
2007; Smith 2004; but see Broughton and Bayham 2003; Codding and Jones 2007; Grimstead 2010; Gurven and Hill 2009). This line of reasoning sug
gests that when large game is pursued to the exclu sion of smaller, more abundant prey items, the gross caloric returns decrease because higher daily yields could be garnered by intensively focusing on small, abundant, lower-caloric return foods. Under these
conditions, large game is costly because it does not
produce daily caloric returns as high as might be
expected from another foraging strategy. This sug
gests that large game hunters may have incurred inordinate costs in exchange for prestige and social
capital. Because this behavior is not optimal in a
caloric sense, only the highest quality individuals can "afford" a lower net return, making this behav ior both economically costly and prestigious because of its exclusionary tendency.
Although the question of the costliness of large bodied prey items remains,1 there is a significant amount of research to suggest large game produce the greatest returns when considering input versus
output (e.g., Bayham 1979; Broughton 1994,1999;
Grayson 2001; Grimstead 2010; Smith 1991; Ugan 2005). Consequently, from the perspective of for
aging theory, large game should have been pre ferred for feasting events based on its high economic value, and from the perspective of sig
naling theory, large game is preferred for feasts
where status could be garnered or lost because of
the tendency for large game to be associated with
status aggrandizement and prestige. Both optimal foraging theory and signaling the
ory come to the same conclusion: big game is high value and prestigious. It is the case that large mam
mals may be used as a prestige item both because
of their high rank as an economic food item and
because of their high value as a signaling item. Big
game may be costly under certain circumstances
and not others. If elites were signaling or compet
ing for prestige, one would expect a feast to be dominated by high caloric return prey, highly pres
tigious food items, and prey with the capacity to serve as a signal of high quality, such as big game. Thus, the prevalence of large game in an archaeo
logical feasting context may indicate status aggran dizement, particularly when other indicators corroborate that conclusion.
This conclusion, from a behavioral ecological perspective, is wholly consistent with Hayden's (1995, 1996) argument that an elite feast may include the conversion of food into prestige, because prestige may be garnered from a food item's exotic nature and associated high procure
ment cost, or from the food item's contribution as a high return food item. We therefore conclude, on
the basis of evolutionary and social theory and on
ethnographic observations that the proportionate representation of large game in a feasting context serves as a barometer of the degree of elite aggran dizement and status display. Notably, the visibility of this currency is increased in habitats where the
density of large-game resources compared to small or medium sized prey items is low, such as the North American Southwest.
The Cahokia Model and Feasting Expectations
Archaeofaunal remains have been used to detect
feasting events throughout the Southwest (e.g., Cameron 1995; Potter 1997; Szuter 1989), but a
recent analysis of a unique context at the prehis toric mound center of Cahokia in the Eastern Wood lands establishes a model for examining the faunal correlates of elite feasting (Pauketat et al. 2002). Our hypothesis seeks to evaluate elite feasting behavior. Therefore, despite the possible social
inequality among Cahokians and residents of the Marana platform mound, Cahokia serves as an ideal
model from which to derive expectations because elites were clearly signaling their prestige via food
and feasting (Kelly 2001; Pauketat et al. 2002). The
Cahokia research will be used to outline some
potential correlates of elite feasting behaviors, and serves as a model to identify several empirical indi
cators where elites were present and likely signal
ing their prestige. The proposed archaeological evidence of a feast
at Cahokia is compelling because of the strati
graphic isolation of a single event within a refuse
pit that was potentially only in use for one season
This content downloaded from 202.41.10.30 on Wed, 21 Jan 2015 07:15:45 AMAll use subject to JSTOR Terms and Conditions

846 AMERICAN ANTIQUITY [Vol. 75, No. 4,2010
(Pauketat et al. 2002:262). Archaeobotanical and
paleoentomological evidence in conjunction with intermittent deposition of geomorphologically dis tinct soil layers enabled archaeologists to narrow down the season of this specific feasting event to late summer or early fall (Kelly 2001:343; Pauke tat et al. 2002). In a single depositional context, Pauketat and colleagues found the following fau nal characteristics apparently associated with elite
feasting: (1) low taxonomic diversity; (2) a rela
tively large quantity of deposited bone; and (3) the
prevalence of high meat-yielding species. These three criteria may be particularly relevant because of their direct association with optimal foraging the
ory and social status. The large quantity of white-tailed deer
(Odocoileus virginianus) remains (99.7 percent of the identified mammal remains) and artifacts asso ciated with both elite and non-elite activities at Cahokia led Pauketat et al. (2002) and Kelly (2001) to conclude that this was a communal feasting event in which both elites and non-elites were in atten dance. The large quantity of deer remains was inferred to be representative of aggrandizing behav ior by the elites (Kelly 2001:354). The type of feast
ing described by Kelly (2001) could be categorized as promotional, political support, or status display feasting, using Hayden's (1995,1996) terminology. This interpretation was supported by ethnographic analogs of ritualized feasting behavior, and, as we
argue in this article, this interpretation is also con sistent with the high economic and social value of
artiodactyls as predicted under foraging theory and
signaling theory. To summarize, the results of archaeofaunal analyses from a discrete feasting locus at the complex mound center of Cahokia show: low taxonomic diversity; a prevalence of
high meat yielding species; and a relatively large quantity of deposited bone (Kelly 2001; Pauketat et al. 2002). These results, as well as nonarchaeo faunal evidence at Cahokia, are consistent with
Hayden's (1995) expectations of a competitive feasting event.
The archaeofaunal record may be one of the more critical ways to identify and evaluate the rit ual or sociopolitical meanings of prehistoric feast
ing. The above review of ethnographic, behavioral
ecological, and archaeological expectations for elite
feasting behavior provide a theoretical and method
ological basis for evaluating any hypothesized locus
of elite feasting. Elite feasting events comparable with that independently found at Cahokia, as observed by Hay den (1995,1996), and consistent with expectations derived from foraging and sig naling theory should exhibit the following charac teristics:
(1) there should be a large quantity of bone
deposited; (2) there should be a high frequency of large
prey items (i.e., preferentially ranked taxa or taxa that may be associated with high degrees of pres tige);
(3) the deposit should contain a low degree of taxonomic diversity; and
(4) there should be unique features, artifacts, and/or architecture associated with the loci.
Yet the ethnographically observed variation in
feasting clearly speaks to the underlying social
complexity behind this behavior, and foreordains the type of problems that archaeologists face when
attempting to identify feasting. Because of the moti vational and functional variability in the nature of
feasting events we would expect a commensurate
degree of complexity and variability in the archae
ological expression of feasting (Cameron 1995; Hockett 1998; Szuter 1989, 1991). Nevertheless, given the importance of this sociopolitical behav
ior, we are optimistic that an inquiry into this phe nomenon under certain archaeological conditions
may yield some insights into past human behavior. One such candidate is a unique locality at a Clas sic period Hohokam site: the Marana platform
mound complex.
The Hohokam and the Marana Platform Mound
The Hohokam were irrigation horticulturalists who
occupied extensive reaches of the deserts in south ern Arizona approximately A.D. 300-1350 (Fig ure 1; Crown 1990; Haury 1976; Plog 1998). Although they thrived on domesticates like maize, beans, and squash and on nondomesticates such as
mesquite, prickly pear cactus, and agave, the Hohokam routinely exploited the native fauna, including artiodactyls, lagomorphs (the dominant source of Hohokam animal protein), rodents, and carnivores (Dean 2007).
Extensive research conducted by Paul and Suzy
This content downloaded from 202.41.10.30 on Wed, 21 Jan 2015 07:15:45 AMAll use subject to JSTOR Terms and Conditions

Grimstead & Bayham] EVOLUTIONARY ECOLOGY, ELITE FEASTING, AND THE HOHOKAM 847
Figure 1. A watershed map of Arizona. This map shows the Hohokam cultural boundary (dashed line) and the Marana
study area. Hohokam boundaries and Marana study region based upon S. Fish et al. (1992a).
Fish and James Bayman (Bayman 1995, 1996, 2002; Bayman et al. 2010; Fish and Fish 2000; P. Fish et al. 1992; S. Fish et al. 1992a, 1992b) over the past 20 years has shown that the Marana plat form mound and the surrounding community is associated with the incipient stages of the Hohokam cultural tradition and extends into the post-Classic. The Marana platform mound was the central fea ture of a large Classic period (ca. A.D. 1150-1350)
community located within the Tucson Basin, Ari zona (Figure 2). The Classic period in the Marana
study area is marked by the abandonment of ball
courts, the construction of platform mounds, and the expansion of nonspecialized habitation sites from riverine and mountain localities to specialized sites in the riverine, montane, and middle bajada ecozones (S. Fish et al. 1992b). This period is marked by craft specialization, a significant varia
tion in social status, and apparently complex com
munity politics (Bayman 1995, 2002). Although pithouses were still occupied during the Classic
period, walled adobe compounds became common.
Archaeological researchers have concluded that the Marana platform mound is socially, politically, and economically distinct from other structures and activities within the broader community, and the mound itself was one of the few areas potentially associated with elite activities (e.g., Bayman 1995, 1996; Bayman et al. 2010; Fish and Fish 2000; S. Fish et al. 1992a; Harry 1997). Thus, the apparent centralization of power and marked social differ entiation within the Marana community sets the
sociopolitical arena for elite aggrandizement and
hegemonic behavior. The development of social complexity among
the Hohokam of southern Arizona has been
This content downloaded from 202.41.10.30 on Wed, 21 Jan 2015 07:15:45 AMAll use subject to JSTOR Terms and Conditions

848 AMERICAN ANTIQUITY [Vol. 75, No. 4, 2010
Figure 2. The Marana platform mound and Muchas Casas. The Marana platform mound complex and Muchas Casas are located at the northern reaches of the Early Classic and Classic period Hohokam sites. Adapted from Bayman (1996).
explored through archaeological study (Doyel et al.
2000; Reid and Whittlesey 1997) and the analysis of an impressive collection of archaeofauna (e.g., Cameron 1995, 1998; Szuter 1989, 1991). Exten sive survey and excavation of numerous middens, structures, compounds, and pithouses associated
with the Marana complex have resulted in an oppor tunity to investigate power and prestige among the
Hohokam (Bayman et al. 2010; S. Fish et al.
1992a). It was in the context of this larger study that a unique aggregation of artifacts and features was discovered at a locus that also produced an
inordinately large quantity of animal bone. During the 2002-2003 field seasons, a burned room was revealed adjacent to the Marana platform mound
yet within the larger compound wall. This area?
This content downloaded from 202.41.10.30 on Wed, 21 Jan 2015 07:15:45 AMAll use subject to JSTOR Terms and Conditions

Grimstead & Bayham] EVOLUTIONARY ECOLOGY, ELITE FEASTING, AND THE HOHOKAM 849
within the compound and in proximity to the plat form mound?had earlier been inferred to be asso ciated with higher-status individuals from the
Marana community (S. Fish et al. 1992b). The sub
sequent excavation and analyses of this room were
pursued with the intent of understanding the mech anism and structure of power within the Marana
community (Bayman et al. 2010). Three key observations by the project principals
spawned various questions about feasting and led to this research. First, previous research in the study area and ethnographic analogy suggest ceremonial
activity was one of the central functions of south ern Arizona platform mounds, and the centraliza tion of ceremony served as a political arena for
prestige-seeking individuals within Hohokam soci
ety (e.g., Elson 1998; Fish and Fish 2000; Rice
1987; Rice and Redman 1993). Second, previous research suggested that this room was associated with individuals of higher status (S. Fish et al.
1992b). Third, preliminary field analyses of site for mation processes suggested the room contents (incl. the faunal material) were deposited over a very brief period of time and were perhaps associated with the ritual closing of Room 9 (Suzanne and Paul
Fish, personal communication 2003). Furthermore, the discard patterns observed within Room 9 do not follow behaviors observed within the broader plat form mound community (Beck and Roos 2010). Archaeobotanical analyses across the Marana com
munity revealed high seed densities and low taxo nomic diversity within Room 9, supporting the idea that Room 9 was unique and perhaps associated
with some form of elite feasting event (Hansen
Speer 2010). In Hayden's (1995,1996) terms, the
project principals hypothesized that the faunal
deposit represented an elite feast?a diacritical feast where power and status were exclusively sig naled to the broader community.
Marana Sample Contexts
We bring together three lines of evidence to eval uate the locus of elite feasting at Marana: ethno
graphic observations of feasting, archaeological case studies, and expectations for elite feasting derived from evolutionary ecology. The review, case studies, and expectations collectively suggest that distinctive architecture, unique features, rare
artifacts, and the prevalence of large game are gen
erally associated with elite feasting. Predicated on
distinctions and interpretations made by project principals (Fish et al. 2010; Fish and Fish 2000), we constructed a comparative analysis using the
hypothesized elite feasting context (Room 9), a mundane habitation locality (Room 1), and a ritu alized elite context (James 1987; Rice 1987) to
explore how elite feasting activities may differ from
generalized food consumption in the Hohokam world.
The archaeofaunal remains from Marana used in this study were recovered during the spring of 2003 from two distinct localities in Compound 9 and Compound 1 (Figure 3). Room 9, from Com
pound 9, was adjacent to the platform mound itself and contained several unique features and artifacts. The sparse floor assemblage included a datura pot effigy (Figure 4), one Glycymeris sp. shell bracelet
fragment, several Conus sp. shell bead fragments, and was relatively devoid of consumptive faunal material. Two sets of floor features are notable in Room 9 when compared to the rest of the Marana
compound. First, there were 170 post-holes, none of which were large enough to support the roof beams and therefore suggesting that this room had a very large open floor arena. Additionally, two hearths were recorded; one was considered com
paratively normal in size, whereas the other was
uncharacteristically large for the site (Figure 5). The floor assemblage was covered by roof/wall fall,
creating a sterile layer on which the relatively large quantity of faunal remains, ceramic sherds, and lithic artifacts were deposited. This portion of the
assemblage was "capped" by a sterile layer of addi tional roof/wall fall.
The ceramic artifacts recovered from Room 9
suggest that the deposit was created sometime between A.D. 1150 and 1250 (Bayman et al. 2010). The unique features, artifacts, depositional attrib
utes, and spatial proximity to the platform mound
suggest Room 9 served a unique social and politi cal function, thought to be primarily attributable to
elites (S. Fish et al. 1992b). Project principals con
cluded that the Room 9 archaeofaunal deposit resulted from a single archaeofaunal deposition event, or several deposition events in rapid suc
cession, that was sealed by a likely deliberate
destruction or "closing" of the room. The strongest evidence of such a deliberate deposition of feast
ing remnants comes from the layers of culturally sterile roof and wall fall above and below the
This content downloaded from 202.41.10.30 on Wed, 21 Jan 2015 07:15:45 AMAll use subject to JSTOR Terms and Conditions

850 AMERICAN ANTIQUITY [Vol. 75, No. 4, 2010
I hc Marana Platform Mound Site
(AZ AA: 12:251)
Legend (om|H.?riii
| Identified C ompounds | Surface Mounds Contours (I in)
Compound Compound 1^
l ?H1l|MUItl(l
C> ̂ 5
o
o loo im 100 < (impound Ci > ( <Mlt|HIII!l(t ^
Figure 3. A map of the Marana platform mound site (Compound 9 and Compound 1 are in bold). Rendered by Phillip O. Leckman of the University of Arizona and adapted by the authors.
Figure 4. A picture of the fragmented datura pot effigy recovered from Compound 9, Room 9 of the Marana platform mound archaeological site. Photograph taken with permission from the Arizona State Museum, University of Arizona
photographer Jannelle Weakly.
This content downloaded from 202.41.10.30 on Wed, 21 Jan 2015 07:15:45 AMAll use subject to JSTOR Terms and Conditions

Grimstead & Bayham] EVOLUTIONARY ECOLOGY, ELITE FEASTING, AND THE HOHOKAM 851
Figure 5. Compound 9, Room 9 boundaries and excavations revealed during 1993 and 2003 excavations. Adapted by authors from original rendering by Rick Karl, M.A., Arizona State Museum.
archaeofaunal remains. There was little evidence
of deposit degradation associated with precipita tion or soil movement. The absence of such tapho nomic processes suggests that the faunal materials were deposited during a relatively brief period of
time, perhaps less than a season, and that the deposit came to be protected by the cap of roof/wall fall
shortly after deposition. Standing in contrast to this locus is Room 1 in
Compound 1, which is located 215m east of Room 9 (Figures 3 and 6). Room 1 was excavated using comparable procedures to those at Room 9, and the
quality of preservation is similar. Room 1 is a semi subterranean adobe structure, approximately 20
m2, characterized by an average-size hearth and
containing an artifact assemblage typically associ ated with domestic use (i.e., manos, cobbles, archaeofaunal remains, lithics, and broken
nonunique ceramic vessels). This room is thought to have been contemporaneously occupied with Room 9. Researchers studying the broader archae
ological manifestation of the Marana complex view Room 1 as representing day-to-day household activities typical of many domestic rooms at the
Marana site and illustrative of the more mundane social activities surrounding the platform mound
(references within Bayman et al. 2010). Room 1 serves as a comparative control to the hypothesized
This content downloaded from 202.41.10.30 on Wed, 21 Jan 2015 07:15:45 AMAll use subject to JSTOR Terms and Conditions

852 AMERICAN ANTIQUITY [Vol. 75, No. 4, 2010
activities in Room 9 because of its similarity to other domestic rooms, proximity to Compound 9, and temporal overlap with Room 9.
The Archaeofaunal Data Set
Archaeofaunal studies in southern Arizona (Bay ham 1982; Bayham and Hatch 1985a, 1985b; Cameron 1995, 1998; Dean 2007; Szuter 1989, 1991; Szuter and Bayham 1989, 1997) have doc umented the numerous ways that animal popula tions have been affected by the prehistoric sedentary horticulturalists known as the Hohokam and how animals have been fully integrated into the
complex social fabric of Hohokam society. Faunal studies at the Marana platform mound and sur
rounding community have also addressed issues of social complexity and the ritual use of animals (S. Fish et al. 1992a; James 1987).
The faunal material presented here anchors the remainder of this study and is based on the identi fication of 2,888 pieces of bone from two locali ties of the Marana site (Room 9, Compound 9 and Room 1, Compound 1). All bone recovered from Rooms 9 and 1 was comparatively examined and identified by both authors and California State Uni
versity, Chico zooarchaeology students while
This content downloaded from 202.41.10.30 on Wed, 21 Jan 2015 07:15:45 AMAll use subject to JSTOR Terms and Conditions

Grimstead & Bayham] EVOLUTIONARY ECOLOGY, ELITE FEASTING, AND THE HOHOKAM 853
supervised by the authors (Grimstead and Bayham 2010). Identifications were made to the most spe cific taxonomic level possible; nomenclature fol lows that of Whitaker (1996), the National
Geographic Society (2002), and Stebbins (2000). Room 9 of the Marana platform mound com
pound produced 2,567 pieces of bone, of which
1,091 (or 43 percent) were identifiable to at least the level of order; these are distributed among 21 different taxa. In total, Room 1 produced 321 spec imens, of which 155 (or 48 percent) were identifi able to the ordinal level. These specimens represented at least nine different taxa (Tables 2 and
3). Both rooms are clearly dominated by lago morphs, which constitute approximately 90 percent of the identifiable taxa in both cases.
One relatively clear faunal correlate of elite
feasting is the proportion of large-bodied prey in an assemblage, and it is reasonable to wonder a pri ori the extent to which they were available during the Classic period in the region. For this reason, we
have elected to include a relatively unique feature in this study for comparative purposes. Pithouse
496, Locus H, is an early Classic period room exca vated as part of a larger complex of structures referred to as Muchas Casas (Rice 1987) and situ ated only a few miles south of the Marana platform mound (Figure 2). The faunal material from Pit house 496 totaled 551 identifiable specimens, almost exclusively dominated by large-bodied high prestige food items (Table 4; James 1987). In fact, the large proportion of high utility body parts from the order Artiodactyla at this locus led James (Table 5; 1987) to interpret the deposition as an elite con text. It is unclear if this deposit was solely food refuse or included more ritually deposited archaeo fauna. However, because of its previous interpre
tation as an elite context and its ability to
demonstrate the availability of artiodactyls both on
the landscape and to elites during this time period, Pithouse 496 could well be important for the pur
pose of evaluating elite feasting. We assume that variation among these loci is,
to a large extent, a product of human behavior. At
this time, our examination of the faunal material
and our reading of the various reports do not sug
gest that taphonomic factors exerted considerable
influence over the degree of interassemblage fau nal variation. A broad spectrum of taphonomic issues could be examined to further buttress the
rationale for comparing these particular loci in the
future, but are not included here.
Comparative Analysis of Room 9, Room 1, and Pithouse 496
Despite the expected motivational and strategic variability in feasting behavior, the role of food is central during any feasting event. The unique loca
tion, contexts, and associations of Room 9 led to the hypothesis that this room represented the remains of an elite feasting event. Archaeofaunal remains can play an important role in testing any hypothesized locus of feasting. Using the previ ously identified archaeological signatures of elite
feasting we expect: (1) unique artifacts, features, and architecture; (2) low taxonomic diversity; (3) a high density of archaeofaunal material deposited within the feasting context compared to the mun dane loci; and (4) a high proportion of high meat
yielding species or highly prestigious taxa. Each
expectation is evaluated below.
Unique Artifacts, Features, and Architecture Expectations
If Room 9 was a locus of elite feasting, then the material culture should be indicative of such behav ior. Unique architecture, artifacts, or features may be indicators of an elite presence during the depo sition event(s). Room 9 was located adjacent to the
platform mound. Prior to the Classic period, plat form mounds did not exist, and ball courts were the common social arena (Crown 1990; S. Fish et al.
1992a). Mounds became the dominant large-scale community structures during the Classic period, and mounds were associated with unique social and political activities. Thus the location of Room
9, adjacent to the platform mound, strongly sug gests Room 9 was also associated with unique social and political meaning.
The architecture of Room 9 also stands in con
trast to other rooms within the Marana complex. There were no excavated roof support post-holes,
suggesting an architectural design that allowed for a large number of people to gather without visual
obstructions in the interior space. One hundred sev
enty relatively small post-holes were discovered, and were thought to have been associated with fur
niture (Bayman et al. 2010). Room 9 measured
approximately 35 m2, which was nearly twice the
This content downloaded from 202.41.10.30 on Wed, 21 Jan 2015 07:15:45 AMAll use subject to JSTOR Terms and Conditions

854 AMERICAN ANTIQUITY [Vol. 75, No. 4,2010
Table 2. NISP and Unidentifiable from Compound 9, Room 9 of the Marana Platform Mound Site.
Compound 9 Room 9 NE SE NW SW
Identifiable_Quad_Quad_Quad_Quad_Total
Reptilia 0 0 0 0 0 Testudinidae 0 2 0 0 2 Squamata 0 0 1 0 1
Aves 0 0 0 0 0 Callipepla spp. 0 16 0 7
Accipitridae 0 10 0 1 Buteo spp. 0 0 2 1 3
Mammalia 0 0 0 0 0 Lagomorpha 0 0 0 2 2 Leporidae 3 6 4 0 13
Lepus spp. 80 113 372 116 681 Lepus californicus 23 19 85 13 140
Lepus alleni 3 12 38 11 64 Sylvilagus spp. 10 19 32 24 85
Rodentia 2 3 3 1 9 Sciuridae 10 0 12
Ammospermophilus spp. 0 2 0 0 2
Spermophilus spp. 0 0 2 1 3
Geomyidae 0 0 0 0 0 Thomomys bottae 0 2 0 0 2
Cricetidae 0 12 0 3 Dipodomys spp. 1 3 2 0 6 Neotoma spp. 4 2 2 1 9
Perognathus spp. 0 0 1 0 1
Peromyscus spp. 0 0 2 0 2
Chaetodipus spp. 0 7 0 0 7 Mustelidae 0 0 0 0 0
Taxidea taxus 0 0 0 1 1 Carnivora 0 0 0 0 0 Canidae 1 1 1 2 5
Canis latrans 0 0 0 1 1 cf. Urocyon cinereoargenteus 0 0 0 0 0
Urocyon cinereoargenteus 0 0 1 0 1
Artiodactyla 0 4 16 3 23 cf. Antilocapra americana 110 0 2
Antilocapra americana 0 0 1 12 cf. Ovis canadensis 1 0 0 0 1 Ovis canadensis 0 12 0 3 Odocoileus spp. 1 0 5 0 6 Odocoileus hemionus 0 0 0 1 1
Sub-total 131 200 580 180 1,091
Non-identifiable
Aves 0 0 1 0 1 Mammal/indet 68 93 143 39 343
Sm mammal 114 133 490 128 865 Med mammal 0 0 0 0 0
M/L mammal 11 20 41 35 107 Indet/Unid 39 27 62 32 160
Sub-total 232 273 737 234 1,476
Total_363_473_1317_414_2,567 Note: Compound 9, Room 9 was excavated in four quadrants and that archaeological methodology is retained in this table for data retention and site formation visibility.
This content downloaded from 202.41.10.30 on Wed, 21 Jan 2015 07:15:45 AMAll use subject to JSTOR Terms and Conditions

Grimstead & Bayham] EVOLUTIONARY ECOLOGY, ELITE FEASTING, AND THE HO HO K AM
Table 3. NISP and Unidentifiable from Compound 1, Room 1 of the Marana Platform Mound Site.
855
Coumpound 1 Room 1 NE SE NW SW
Identifiable_Quad_Quad_Quad_Quad_Total
Reptilia 0 0 0 0 0 Testudinidae 0 0 0 0 0
Aves 0 0 0 0 0 Callipepla sp. 0 0 0 0 0
Accipitridae 0 0 0 0 0 Buteo spp. 0 0 0 0 0
Mammalia 0 0 0 0 0
Lagomorpha 0 10 0 1
Leporidae 10 1 2 4
Lepus spp. 12 14 24 50 100 Lepus californicus 0 1 0 10 11
Lepusalleni 0 0 0 12 12
Sylvilagus spp. 2 3 2 8 14
Rodentia 1 0 0 0 1 Sciuridae 0 0 0 1 1
Ammospermophilus spp. 0 0 0 0 0
Spermophilus spp. 0 10 0 1
Geomyidae 0 0 0 0 0 Thomomys bottae 0 0 0 0 0
Cricetidae 0 0 0 0 0
Dipodomys spp. 0 0 0 0 0
Neotoma spp. 0 0 0 2 2
Perognathus spp. 0 0 0 0 0
Peromyscus spp. 0 0 1 0 1
Chaetodipus spp. 0 0 0 0 0
Mustelidae 0 10 0 1
cf. Taxidea taxus 0 0 0 0 0
Carnivora 0 0 0 0 0
Canidae 3 0 0 0 3 cf. Urocyon cinereoargenteus 0 0 1 0 1
Urocyon cinereoargenteus 0 0 0 0 0
Artiodactyla 0 10 12
cf. Antilocapra americana 0 0 0 0 0
Antilocapra americana 0 0 0 0 0
cf. Ovis canadensis 0 0 0 0 0
Ovis canadensis 0 0 0 0 0
Odocoileus spp. 0 0 0 0 0
Odocoileus hemionus 0 0 0 0 0
Sub-total 19 22 29 86 155
Non-identifiable
Aves 0 0 0 0 0 Mammal/indet 19 18 6 14 57
Sm mammal 12 6 7 35 60 Med mammal 0 0 0 0 0 M/L mammal 0 0 3 2 5 Indet/Unid 2 0 10 33 45
Sub-total 33 24 26 84 167
Total_52_46_55_168_321 Note: Compound 1, Room 1 was excavated in four quadrants and that archaeological methodology is retained in this table
for data retention and site formation visibility.
This content downloaded from 202.41.10.30 on Wed, 21 Jan 2015 07:15:45 AMAll use subject to JSTOR Terms and Conditions

856 AMERICAN ANTIQUITY [Vol. 75, No. 4, 2010
Table 4. NISP and Non-NISP from Pit House 496, Locus H, Muchas Casas.
Pit House 496, Locus H, Muchas Casas
Identifiable_Total
Lagomorpha [Total] [3] Lepus sp. 1
Lepus californicus 1
Lepus alleni 1
Artiodactyla [Total] [548] Odocoileus sp./Ovis canadensis 10
Odocoileus sp./Ovis spJAntilocapra sp. 17
cf. Odocoileus sp. 132
Odocoileus sp. 23
Odocoileus hemionus 1
Ovis canadensis 368
Total NISP 551 Unidentifiable
Small Mammalia 1
Medium Mammalia 0
Large Mammalia 4
Total unidentifiable _ 5
Table 5. Body Part and MNE Data, Recorded by James
(1987), for Pithouse 496 and Ash Pit 497.
_NISP_MNE Ovis canadensis
Horn core 220 10
Skull [Total] [124] Occipital 5 5
Maxilla 4
Fragments 115
Pelvis 26 9 Odocoileus sp.
Antler 1 1
Pelvis 155 36
Distal Femur 1 1
Odocoileus sp./Ovis sp. Skull 2 1 Pelvis 21 4
Proximal Femur 3 3
Distal Metapodial 2 2
Total 555 72
size of Room 1 (20 m2), and relatively large com
pared to several other loci within the community. Thus, Room 9 appears to be unique in its size, architecture, and design.
The artifacts and features discovered within Room 9 also suggest a very unique function. No other room excavated to date has produced two hearth features. One of these hearths is much larger than all other hearths within the Marana study area,
and this large hearth is centrally located within the room. In addition to the hearths, the datura pot effigy and several other artifacts were atypical and
generally not found in day-to-day household con texts (Bayman et al. 2010).
The unique context expectation suggests elite
feasting should be associated with unique artifacts, features, and architecture. This expectation appears to be met by Room 9 but not Room 1. Room 9 is located adjacent to a platform mound, contains
unique features and artifacts, and is architecturally unique compared to other rooms within the Marana
study area. These results tend to support the pre
liminary interpretation that elites may have been associated with Room 9.
Diversity Expectation
If elite feasting produced the observed faunal
assemblage, then we would expect a low degree of taxonomic diversity. To evaluate the degree of
diversity represented in the identifiable faunal
remains, the inverse of the Simpson's index was
applied (Grayson 1984). This form of diversity index typically ranges between 1 and 18. A higher value from the reciprocal of the Simpson's index reveals a broader distribution of taxa. Results with lower numbers are consistent with low taxonomic
diversity, as would be expected with rooms heav
ily dominated by one or a few taxa. Grayson (1984) cautions that this index could be reflective of sam
ple size and suggests using Spearman's rho (rs) and the reciprocal of the Simpson's index in tandem to detect any correlation with sample size. Both equa tions were applied in this analysis.
The results show generally low degrees of diver
sity from all three rooms (Room 9 = 1.37; Room 1 = 1.45; Pithouse 496 = 1.74). These values are uni
formly low, and there is no statistically significant correlation with sample size (rs
= <.500; p - .667). All results are consistent with low taxonomic diver
sity, as would be expected with rooms heavily dom inated by lagomorphs or artiodactyls. At face value, these results tend to support the diversity expecta tion, but this interpretation must be tempered against the observation that all loci produced uni
formly low values.
Density Expectation
Our third expectation was that elite feasting would result in a large amount of faunal material, mean
This content downloaded from 202.41.10.30 on Wed, 21 Jan 2015 07:15:45 AMAll use subject to JSTOR Terms and Conditions

Grimstead & Bayham] EVOLUTIONARY ECOLOGY, ELITE FEASTING, AND THE HOHOKAM 857
Figure 7. Total identifiable (NISP) and unidentifiable specimens per m3 from Room 9 and Room 1. Room 9 contained a total of 209 specimens per m\ while Room 1 contained 39 specimens per m3.
ing that the density of archaeofaunal remains recov ered from Room 9 should be greater than the archaeofaunal remains recovered from other
Marana loci. The density of archaeofaunal remains
per unit volume of sediment was calculated by dividing the total number of specimens by the total amount of excavated soil for both Room 9 and Room 1 (Pithouse 496 was excluded from this
analysis, because volumetric excavation data was
not available). Figure 7 graphically depicts this cal culation by showing the total number of identifi able and nonidentifiable bone per m3 from each
room, whereas Table 6 provides the soil volume data used to derive these results.
Figure 7 shows a striking difference between both the identifiable and nonidentifiable specimens from Room 1 and Room 9. Room 9 has a total of 209 specimens per m3, while Room 1 only has 39
specimens per m3. These results show Room 9 had over five times the number of specimens per m3 when compared to Room 1. Room 9 also contained more floral and faunal material per m3 of exca vated soil than other loci within the Marana plat form mound site (Hansen-Speer 2010). This observation supports the expectation of a large quantity of deposited food during an elite feasting event.
Table 6. Soil Volume Data for Compound 9, Room 9 and Compound 1, Room 1 of the Marana Platform Mound Site.
Compound 9, Room 9
Level 1
Level 2
Level 3
Level 4
Level 5
Level 6
Level 7
Total Volume Per Quad
NE
Quad
1.29375
.7425
.7425
.7425
0
0
0
3.52125
SE
Quad
.81675
.7425
.7425
.51975
0
0
0
2.8215
NE
Quad
0
.50275
.90263
.9045
.9045
.9045
.9045
5.02338
SW
Quad
.55925
.55925
.61518
.39148
0
0
0
2.12515
Total volume
per level
2.66975
2.547
3.0028
2.55822
.9045
.9045
.9045
13.49128
Compound 1, Room 1
Level 1
Level 2
Level 3
Level 4
Level 5
Level 6
Level 7
Total Volume Per Quad
.40095
.4455
.4455
.4455
0
0
.4455
2.18295
.40205
.3655
.3655
.3655
0
0
.3655
1.86405
.6657
.317
.317
.2536
.2219
0
.5244
2.2996
.1782
.4455
.4455
.4455
0
0
.4455
1.9602
1.6469
1.5735
1.5735
1.5101
.2219
0
1.7809
8.3068
This content downloaded from 202.41.10.30 on Wed, 21 Jan 2015 07:15:45 AMAll use subject to JSTOR Terms and Conditions
858 AMERICAN ANTIQUITY [Vol. 75, No. 4, 2010
Lagomorpha Artiodacryla
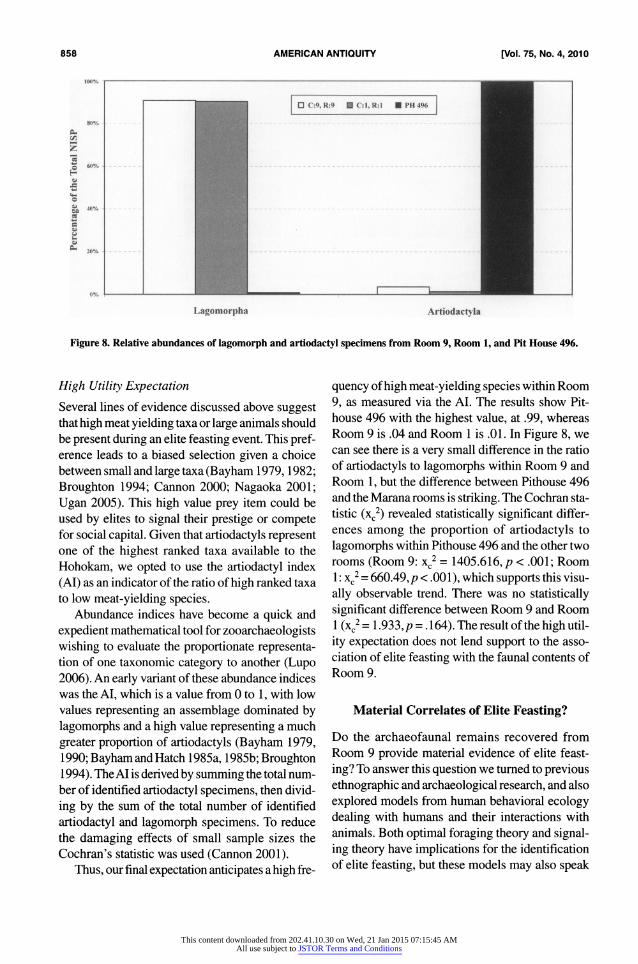
Figure 8. Relative abundances of lagomorph and artiodactyl specimens from Room 9, Room 1, and Pit House 496.
High Utility Expectation
Several lines of evidence discussed above suggest that high meat yielding taxa or large animals should be present during an elite feasting event. This pref erence leads to a biased selection given a choice between small and large taxa (Bayham 1979,1982;
Broughton 1994; Cannon 2000; Nagaoka 2001;
Ugan 2005). This high value prey item could be used by elites to signal their prestige or compete for social capital. Given that artiodactyls represent one of the highest ranked taxa available to the
Hohokam, we opted to use the artiodactyl index
(AI) as an indicator of the ratio of high ranked taxa to low meat-yielding species.
Abundance indices have become a quick and
expedient mathematical tool for zooarchaeologists wishing to evaluate the proportionate representa tion of one taxonomic category to another (Lupo 2006). An early variant of these abundance indices was the AI, which is a value from 0 to 1, with low values representing an assemblage dominated by lagomorphs and a high value representing a much
greater proportion of artiodactyls (Bayham 1979, 1990; Bayham and Hatch 1985a, 1985b; Broughton 1994). The AI is derived by summing the total num ber of identified artiodactyl specimens, then divid
ing by the sum of the total number of identified
artiodactyl and lagomorph specimens. To reduce the damaging effects of small sample sizes the Cochran's statistic was used (Cannon 2001).
Thus, our final expectation anticipates a high fre
quency of high meat-yielding species within Room
9, as measured via the AI. The results show Pit house 496 with the highest value, at .99, whereas Room 9 is .04 and Room 1 is .01. In Figure 8, we can see there is a very small difference in the ratio of artiodactyls to lagomorphs within Room 9 and Room 1, but the difference between Pithouse 496 and the Marana rooms is striking. The Cochran sta tistic (xc2) revealed statistically significant differ ences among the proportion of artiodactyls to
lagomorphs within Pithouse 496 and the other two rooms (Room 9: xc2
= 1405.616, p < .001; Room 1: xc2
= 660.49,/? < .001), which supports this visu
ally observable trend. There was no statistically significant difference between Room 9 and Room 1 (xc2= 1.933,/? = .164). The result ofthe high util
ity expectation does not lend support to the asso ciation of elite feasting with the faunal contents of Room 9.
Material Correlates of Elite Feasting?
Do the archaeofaunal remains recovered from Room 9 provide material evidence of elite feast
ing? To answer this question we turned to previous ethnographic and archaeological research, and also
explored models from human behavioral ecology dealing with humans and their interactions with animals. Both optimal foraging theory and signal ing theory have implications for the identification of elite feasting, but these models may also speak
This content downloaded from 202.41.10.30 on Wed, 21 Jan 2015 07:15:45 AMAll use subject to JSTOR Terms and Conditions

Grimstead & Bayham] EVOLUTIONARY ECOLOGY, ELITE FEASTING, AND THE HOHOKAM 859
to the broader social dynamics inherent within both elite and non-elite feasting behavior. From ethno
graphic evidence, archaeological research, and eco
logical theory, four faunal correlates of elite feasting were identified and served as our analytical expec tations.
In this study, our results were mixed. A low
degree of taxonomic diversity in Room 9 is con sistent with our expectation, but the relative sparse ness of meat resources in the desert Southwest
might be influencing this to a high degree. Essen
tially, most Classic period Hohokam faunal assem
blages from a variety of contexts show a low degree of taxonomic diversity in comparison to sites such as Cahokia. Therefore, we are not overly compelled to interpret this as evidence of feasting. At first
glance, this expectation is consistent with expec tations from both optimal foraging theory and sig naling theory, but on further examination it may also be contrary to those from signaling theory.
If elites were signaling their power or prestige, then we might expect them to have procured items that were uncommon, rare, exotic, or costly, which
may have the effect of increasing species richness,
although not necessarily evenness, and perhaps diversity. More ethnographic research into how tax onomic diversity may be associated with different
types of feasting is required, but it could well be the case that higher degrees of species richness should be associated with contexts where elites are
signaling their prestige to other elites. Where other
sociopolitical goals are at stake, such as political support or intercommunity cooperation, we might expect a lower degree of richness, leading to lower
degrees of diversity. Pithouse 496, which was pre
viously interpreted as an elite context, has a low
degree of richness when compared to the other con
texts, but the predominant taxon, Artiodactyla, is a
high prestige item with a high degree of signaling value. Thus an expectation derived from signaling theory that takes into consideration the unique eco
logical conditions of the desert Southwest antici
pates the following: an assemblage used by an elite to signal their power and prestige should have a
lower degree of taxonomic diversity, yet remain
dominated by high prestige food items. Is big game prestigious because it is costly or
because it is highly preferred? Again, the infusion
of criteria from human behavioral ecology may
provide an analytical tool that differs from Pauke
tat's and others' attention to "meat yield" alone
(Kelly 2001; Pauketat et al. 2002). It is the sparse ness of large-game resources in the Southwest that allows both optimal foraging theory and costly sig naling theory to provide relevant expectations of elite feasting. If large game was locally available, then it would be the highest-return prey item, the
most preferred, and it would have a high degree of associated prestige. On the other hand, if large game is locally unavailable, then the cost of procurement
may cause an otherwise high return prey item to diminish in economic value but not necessarily in social value. Despite a purely economic shift in actual value, large animals seem to have always been preferred and always pursued. A high pro curement cost for an otherwise highly preferred food item may make big game potentially presti gious and valuable as a signal of power among elites in the Southwest. Economically speaking, large game should always be preferred and may become particularly prestigious as the cost of pro curement increases. Deer, pronghorn, and bighorn sheep would have been some of the most presti gious fauna available to the Hohokam, and elite status individuals should be consuming these fauna to the exclusion of low prestige prey, especially when consumption is visible to other elites. Elites would have a considerable amount of social capi tal to lose if they did not provide high status meat when feasting with other elites. Therefore, the dom inance of this assemblage by lagomorphs may well be suggestive of a non-elite consumption event, where overt power and status displays by elites were not the primary motivation.
When the proportions of artiodactyls to lago morphs in Room 9 are compared to Pithouse 496, a nondomestic room associated with elites (James 1987), we see a stunning discrepancy. Pithouse 496
was dominated by artiodactyls, as was evident from an AI value of .99. Artiodactyls were nearly absent from Room 9, as was evident from an AI of .04. The high frequency of artiodactyls from Pithouse
496 shows that these large animals were poten
tially available and could have been used by elites
for aggrandizing behavior, regardless of the cost of
procurement. Pithouse 496 was previously shown to have been associated with elite activities, thus, the high AIs associated with Pithouse 496 provide support for using the frequency of high meat
yielding species as an indicator of elite activities?
This content downloaded from 202.41.10.30 on Wed, 21 Jan 2015 07:15:45 AMAll use subject to JSTOR Terms and Conditions

860 AMERICAN ANTIQUITY [Vol. 75, No. 4, 2010
an indicator that is absent from Room 9.
Perhaps the strongest evidence that the contents of Room 9 represent the results of a feast or feasts is found in the large quantity of faunal remains
deposited. Room 9 had more bone per m3 by a sig nificant order of magnitude than all other exca
vated deposits within the Marana complex. Although we cannot exclude use-related or func
tionally different activities contributing to this dif
ference, such as the sweeping and habitual cleaning of room floors (cf. Cameron 1995), the evidence at Room 9 does seem to represent an activity that is fundamentally different from day-to-day house hold consumption.
Returning to Hay den's (1995, 1996) expecta tions of competitive feasting, within Room 9 we find that there may have been sufficient resources to provide surpluses. In addition to the large quan
tity of lagomorphs, plant remains were also very
high in volumetric density, yet low diversity was
again observed via the high proportions of grass seeds (Hansen-Speer 2010). Although there was
only a single fragment of corn, the room contained a high proportion of corn pollen compared to other contexts in Marana (Hansen-Speer 2010). Unique features, artifacts, location, and architecture are
suggestive of a feasting locality, but there is not an
inordinate abundance of prestige items (food or
otherwise) within Room 9. The platform mound itself certainly suggests the presence of elite indi
viduals, but it is not clear if these elites directly par ticipated in the deposition of artifacts within Room 9.
The results of our study suggest that a type of food consumption occurred in Room 9 that is dif ferent from day-to-day household animal con
sumption, but it is not consistent with elite
aggrandizement or competition, where high pro portions of high quality prey items would pre dominate. The archaeofaunal remains may in fact
represent a feast, but they are not consistent with diacritical feasting or feasting that establishes divi sions between elites and non-elites, as defined by
Hayden (1996). Furthermore, promotional, recip rocal, and competitive feasting may all be excluded because of the link between these types of feasting and elite manipulation of social capital where other elites are the intended audience.
The archaeofaunal remains recovered from Room 9 are not consistent with elite feasting expec
tations, yet the unique context of Room 9 does sug gest that elites may have been present. In fact, we
have not excluded feasts that may involve signifi cant political strategizing, such as work party feast
ing, solidarity feasting, and feasting held for
intercommunity political support. These feasts most certainly involved a good portion of the com
munity, including elites, and these feasts may have been a forum in which intercommunity power
dynamics could unfold. On the basis of our ethno
graphic, archaeological, and theoretical reviews, we conclude that Room 9 represents a communal
feasting event that may have been the remnants of a work party feast, a social solidarity feast, or an
intercommunity political support feast in which both elites and non-elites participated. This con
clusion must be tempered with thoughts previously mentioned: feasting is more akin to a Gaussian dis tribution than a categorically discrete bar graph, and the location of a single event on the distribution of
feasting may change within the context of that sin
gle event. Simply stated, the identification of feast
ing is difficult, and its categorization is even more
problematic.
Conclusion
The goal of this article is to determine if the large faunal deposit within Room 9 at the Marana plat form mound site represented a locus of elite feast
ing. A review of anthropological literature, as well as evolutionary theory, produced four criteria that were potentially useful indicators of elite feasting. First, there should be a large quantity of bone
deposited. Second, there should be a high frequency of large prey items, i.e., preferentially ranked taxa or taxa that may be associated with high degrees of prestige. Third, the deposit should contain a low
degree of taxonomic diversity. Fourth, there should be unique features, artifacts, and/or architecture associated with the loci. Room 9, the hypothesized locus of Hohokam elite feasting, was compared to
Room 1, a domestic room, and Pithouse 496, a room with elite affiliation. Comparatively, Room 9 had low faunal diversity, but so did the other sam
ples. There was indeed a statistically significant high density of deposition in Room 9, but there was a very low frequency of preferentially ranked taxa. Room 9 does have unique architecture, artifacts, and contexts, which are important when interpret
This content downloaded from 202.41.10.30 on Wed, 21 Jan 2015 07:15:45 AMAll use subject to JSTOR Terms and Conditions

Grimstead & Bayham] EVOLUTIONARY ECOLOGY, ELITE FEASTING, AND THE HOHOKAM 861
ing the sociopolitical behavior that produced the archaeofaunal deposit. Furthermore, the strati
graphic isolation of the deposit supports this as one
single event or rapidly deposited series of events. This assemblage appears to represent a non
elite feasting event located within a room that is
very likely associated with elite individuals. This leads to the preliminary conclusion that Room 9 is
representative of a communal non-elite feasting event. The hypothesized locus of elite feasting does
appear to be feasting, but it does not seem to be an overt display of power or status. It is not consistent with a feast that had the sole purpose of enhancing an individual's stature within the community or
among several surrounding communities. The loca tion of Room 9 (i.e., adjacent to the platform
mound) and the unique features and artifacts asso
ciated with Room 9 suggest elites were present or had some hand in organizing the event. Yet because of the lack of large-bodied and highly prestigious prey items the primary consumers were most likely non-elite individuals. This pattern is characteristic of a feasting event that served to promote intragroup solidarity or political support for elites within the
community. To return to Hay den's (Table 1; 1995,
1996) categories, this is an example of alliance and
cooperation feasting. Elites may have been present and may have had
a significant political role, but the primary goal of this feast was probably not to display prestige for other potential allies or competitive elites. Although this conclusion does not support the original hypothesis, these results may enhance our appre ciation of how elites at the Marana platform mound and among the Hohokam may have manipulated power within their economy and community. The
type of feasting represented provides evidence of
strategic sociopolitical manipulation of power, where social solidarity and communal events were
the primary venues for signaling. This is not to sug
gest that overt power displays were not present within the community, but that the assemblage ana
lyzed here is more akin to a covert mechanism of
power consolidation within the Marana commu
nity. Further delineation of archaeological indica tors of distinct feasting situations will provide more
opportunities to extract the subtleties of sociopo litical interactions between elites and non-elites.
The phenomenon of feasting is a highly vari able and complex social event, and we do not expect
that the archaeological indices of the phenomenon would be any less complex. Ecological factors
undoubtedly have a significant affect on the man ifestation of empirical indicators of specific feast
ing events, and therefore, ecology must also be understood if archaeofaunal and archaeobotanical remains are to be used as indicators of feasting. Continued study of the empirical signatures of dif ferent types of feasting, and their archaeological and faunal correlates, will enhance our under
standing of prehistoric social and political com
plexity.
Acknowledgments. We thank Suzanne and Paul Fish of the
Arizona State Museum for access to the collection and fig ures, and for comments on drafts of this article. Participants of the 2006 Stanley J. Olsen Memorial Zooarchaeology Conference provided several suggestions and comments that
helped refine our thoughts and intentions. The logic and
readability of the final draft benefited from scrutiny by Barnet Pavao-Zuckerman, Dave Abbott, and several anony mous reviewers. Rick Carl, Phillip O. Leckman, Greg White, and Jannelle Weakly (ASM) were extremely generous with
graphical assistance. Students participating in several
California State University, Chico Zooarchaeology classes
(2003-2004) deserve our gratitude for their dutiful assis
tance in identification of the Marana assemblages. And
assuredly, we are responsible for any misrepresentations or
factual errors contained within.
References Cited
Bayham, Frank E. 1979 Factors Influencing the Archaic Pattern of Animal
Exploitation. The Kiva 44:219-235. 1982 A Diachronic Analysis of Prehistoric Animal Exploita
tion at Ventana Cave. Unpublished Ph.D. dissertation,
Department of Anthropology, Arizona State University, Tempe.
1990 Effects of a Sedentary Lifestyle on the Utilization of Animals in the Prehistoric Southwest. In Agriculture: Ori
gins and Impacts of a Technological Revolution, edited by C. Cameron, pp. 54-78. Occasional Papers of the Archae
ological Research Facility, CSU Fullerton No. 5. Museum of Anthropology, Fullerton.
Bayham, Frank E., and Pamela H. Hatch 1985a Archaeofaunal Remains From the New River Area.
In Hohokam Settlement and Economic System in the Cen
tral New River Drainage, Arizona, edited by David E.
Doyel and Mark D. Elson, pp. 405-^133. Soil Systems Pub
lications in Archaeology No. 4. Phoenix.
1985b Hohokam and Salado Animal Utilization in the Tonto
Basin. In Studies in the Hohokam and Solado of the Tonto
Basin, edited by Glen C. Rice, pp. 191-210. Office of Cul
tural Resource Management, Arizona State University,
Tempe. Bayman, James M.
1995 Rethinking "Redistribution" in the Archaeological Record: Obsidian Exchange at the Marana Platform Mound. Journal of Anthropological Research 51:37-63.
This content downloaded from 202.41.10.30 on Wed, 21 Jan 2015 07:15:45 AMAll use subject to JSTOR Terms and Conditions

862 AMERICAN ANTIQUITY [Vol. 75, No. 4, 2010
1996 Shell Ornament Consumption in a Classic Hohokam Platform Mound Community Center. Journal of Field
Archaeology 23:403^120. 2002 Hohokam Craft Economies and the Materialization
of Power. Journal of Archaeological Method and Theory 9:69-95.
Bayman, James M., Paul R. Fish, and Suzanne K. Fish (editors) 2010 Power and Economy in Early Classic Period
Hohokam Society: An Archaeological Perspective From the Marana Mound Site. Arizona State Museum Series, Uni
versity of Arizona Press, Tucson.
Beck, Margaret, and Christopher Roos 2010 From Households to Middens: Refuse Deposition Pat
terns in Two Communities. In Power and Economy in
Early Classic Period Hohokam Society: An Archaeologi cal Perspective From the Marana Mound Site, edited by James M. Bayman, Paul R. Fish, and Suzanne K. Fish, pp. 34-63. Arizona State Museum Series, University of Ari zona Press, Tucson.
Bliege-Bird, Rebecca, and Eric A. Smith 2005 Signaling Theory, Strategic Interaction, and Symbolic
Capital. Current Anthropology 46(2):221-248.
Bliege-Bird, Rebecca, Eric A. Smith, and Douglas W. Bird 2001 The Hunting Handicap: Costly Signaling in Male For
aging Strategies. Behavioral Ecology and Sociology 50:9-19.
Bray, Tamara L. (editor) 2003 The Archaeology and Politics of Food and Feasting
in Early States and Empires. Plenum Press, New York.
Broughton, Jack M. 1994 Declines in Mammalian Foraging Efficiency During
the Late Holocene, San Francisco Bay, California. Jour nal of Anthropological Archaeology 13:371^-01.
1999 Resource Depression and Intensification During the Late Holocene, San Francisco Bay: Evidence from the
Emeryville Shellmound Vertebrate Fauna. Anthropologi cal Records, Vol. 32. University of California Press, Berke
ley. Broughton, Jack M., and Frank E. Bayham
2003 Showing Off, Foraging Models, and the Ascendance of Large Game Hunting in the California Middle Archaic. American Antiquity 68:783-789.
Cameron, Judi L. 1995 Sociopolitical and Economic Roles of Faunal Resources in the Prehistory of the Tonto Basin, Arizona.
Unpublished Ph.D. dissertation, Department of Anthro
pology, University of Arizona. 1998 The Faunal Resource Use in the Prehistoric Tonto Basin. In Environment and Subsistence Change in the Classic Period Tonto Basin: The Roosevelt Archaeology Studies, 1989 to 1998, edited by Katherine A. Spielmann, pp. 133-188. The Roosevelt Monograph Series, Vol. 10,
Anthropological Field Studies 39. Arizona State Univer
sity, Tempe. Cannon, Michael D.
2000 Large Mammal Relative Abundance in Pithouse and Pueblo Period Archaeofaunas From Southwestern New Mexico: Resource Depression Among the Mimbres
Mogollan? Journal of Anthropological Archaeology 19:317-347.
2001 Archaeofaunal Relative Abundance, Sample Size, and Statistical Methods. Journal of Archaeological Science 28:185-195.
Codding, Brian E, and Terry L. Jones 2007 Man the Showoff? Or the Ascendance of a Just-So
Story: A Comment on Recent Applications of Costly Sig naling Theory in American Archaeology. American
Antiquity 72:349-358.
Crown, Patricia 1990 The Hohokam of the American Southwest. Journal
of World Prehistory 4:223-255.
Dean, Rebecca M. 2001 Social Change and Hunting During the Pueblo III to
Pueblo IV Transition, East-Central Arizona. Journal of Field Archaeology 28:271-285.
2007 Hunting Intensification and the Hohokam "Collapse." Journal of Anthropological Archaeology 26:109-132.
Dietler, Michael, and Brian Hayden (editors) 2001 Feasts: Archaeological and Ethnographic Perspec
tives. Smithsonian Institution Press, Washington, D.C.
Doyel, David E., Suzanne K. Fish, and Paul R. Fish (editors) 2000 The Hohokam Village Revisited. Southwestern and
Rocky Mountain Division of the American Association for
the Advancement of Science, Fort Collins, Colorado.
Elson, M. 1998 Expanding the View of Hohokam Platform Mounds: An Ethnographic Perspective. Anthropological Papers of the University of Arizona No. 63. University of Arizona
Press, Tucson.
Fair, Susan W. 2000 The Inupiag Eskimo Messenger Feast: Celebration,
Demise, and Possibility. The Journal of American Folk lore \tt'A6A-494.
Fish, Paul R., and Suzanne K. Fish 2000 The Marana Platform Mound Site: Patterns of Social
Differentiation in the Early Classic Period. In The Hohokam Village Revisited, edited by David E. Doyel, Suzanne K. Fish, and Paul R. Fish, pp. 245-275. South western and Rocky Mountain Division of the American Association for the Advancement of Science, Fort Collins, Colorado.
Fish, Paul R., Suzanne K. Fish, John H. Madsen, Charles H.
Miksicek, and Christine R. Szuter 1992 The Dairy Site Occupational Continuity on an Allu
vial Fan. In The Marana Community in the Hohokam World, edited by Suzanne K. Fish, Paul R. Fish, and John H. Mad
sen, pp. 64-72. Anthropological Papers of the University of Arizona No. 56. University of Arizona Press, Tucson.
Fish, Suzanne K., Paul R. Fish, and James M. Bayman 2010 Introduction: Power And Economy at the Marana Mound Site. In Power and Economy in Early Classic Period Hohokam Society: An Archaeological Perspective From the Marana Mound Site, edited by James M. Bay man, Paul R. Fish, and Suzanne K. Fish, pp. 15-33. Ari zona State Museum Series, University of Arizona Press, Tucson.
Fish, Suzanne K., Paul R. Fish, John H. Madsen (editors) 1992a The Marana Community in the Hohokam World.
Anthropological Papers of the University of Arizona No. 56. University of Arizona Press, Tucson.
Fish, Suzanne K., Paul R. Fish, and John H. Madsen 1992b An Introduction to Time, Place and Research. In The Marana Community in the Hohokam World, edited by Suzanne K. Fish, Paul R. Fish, and John H. Madsen, pp. 1-11. Anthropological Papers of the University of Ari zona No. 56. University of Arizona Press, Tucson.
Grayson, Donald K. 1984 Quantitative Zooarchaeology: Topics in the Analysis
of Archaeological Faunas. Academic Press, Orlando. 2001 The Archaeological Record of Human Impacts on
Animal Populations. Journal of World Prehistory 15:1-68.
Grimstead, Deanna N. 2010 Ethnographic and Modeled Costs of Big Game Pro
curement. American Antiquity 75:61-80.
This content downloaded from 202.41.10.30 on Wed, 21 Jan 2015 07:15:45 AMAll use subject to JSTOR Terms and Conditions

Grimstead & Bayham] EVOLUTIONARY ECOLOGY, ELITE FEASTING, AND THE HOHOKAM 863
Grimstead, Deanna N. and Frank E. Bayham 2010 Feasting at the Marana Platform Mound. In Power and
Economy in Early Classic Period Hohokam Society: An
Archaeological Perspective from the Marana Mound Site, edited by James M. Bayman, Paul R. Fish, and Suzanne K. Fish, pp. 152-190. Arizona State Museum Series, Tuc son.
Gurven, Michael, and Kim Hill 2009 Why Do Men Hunt: A Reevaluation of "Man the
Hunter" and the Sexual Division of Labor. Current Anthro
pology 50(l):51-7'5.
Hansen-Speer, Karla 2010 Analysis of Plant Remains From the Marana Mound
Site. In Power and Economy in Early Classic Period Hohokam Society: An Archaeological Perspective From the
Marana Mound Site, edited by James M. Bayman, Suzanne K. Fish, and Paul R. Fish, pp. 119-136. Arizona State Museum Series, University of Arizona Press, Tucson.
Harry, Karen G. 1997 Ceramic Production, Distribution, and Consumption
in Two Classic Period Hohokam Communities. Unpub lished Ph.D. dissertation, Department of Anthropology,
University of Arizona, Tucson.
Hasson, Oren 1994 Cheating Signals. Journal of Theoretical Biology
167(3):223-238. 1997 Towards a General Theory of Biological Signaling.
Journal of Theoretical Biology 185(2): 139-156.
Haury, Emil W. 1976 The Hohokam: Desert Farmers and Craftsmen. Uni
versity of Arizona Press, Tucson.
Hawkes, Kristen 1991 Showing Off: Tests of an Hypothesis About Men's
Hunting Goals. Ethology and Sociobiology 12:29-54.
Hayden, Brian 1995 Pathways to Power: Principles for Creating Socio
Economic Inequalities. In Foundations of Social Inequal ity, edited by T. Douglas Price and Gary M. Feinman, pp. 15-86. Plenum Press, New York.
1996 Feasting in Prehistoric and Traditional Societies. In
Food and the Status Quest: An Interdisciplinary Perspec tive, edited by Polly Wiessner and Wulf Schiefenhovel, pp. 129-147. Berghahn Books, Oxford.
Hildebrandt, William R., and Kelly R. McGuire 2002 The Ascendance of Hunting During the California
Middle Archaic: An Evolutionary Perspective. American
Antiquity 67:231-256. 2003 Large-Game Hunting, Gender-Differentiated Work
Organization, and the Role of Evolutionary Ecology in Cal
ifornia and Great Basin Prehistory: A Reply to Broughton and Bayham. American Antiquity 68:790-792.
Hockett, Brian S. 1998 Sociopolitical Meaning of Faunal Remains From
Baker Village. American Antiquity 63:289-302.
Jackson, H. Edwin, and Susan L. Scott
2003 Patterns of Elite Faunal Utilization at Moundville, Alabama. American Antiquity 68:552-572.
James, Steven R.
1987 Hohokam Patterns of Faunal Exploitation at Muchas
Casas. In Studies in the Hohokam Community of Marana, edited by Glen E. Rice, pp. 171-196. Arizona State Uni
versity Anthropological Field Studies No. 15. Arizona
State University, Department of Anthropology, Tempe.
Kelly, Lucretia S. 2001 A Case of Ritual Feasting at the Cahokia Site. In
Feasts: Archaeological and Ethnographic Perspectives on
Food, Politics, and Power, edited by Michael Dietler and
Brian Hayden, pp. 334-367. Smithsonian Institution Press,
Washington, D.C.
Lupo, Karen 2006 What Explains the Carcass Field Processing and
Transport Decisions of Contemporary Hunter-Gatherers? Measures of Economic Anatomy and Zooarchaeological Skeletal Part Representation. Journal of Archaeological
Method & Theory 13(l):19-66. McGuire, Kelly, and William Hildebrandt
2005 Re-Thinking Great Basin Foragers: Prestige Hunting and Costly Signaling During the Middle Archaic Period. American Antiquity 70:695-712.
McGuire, Kelly R., William R. Hildebrandt, and Kimberly L.
Carpenter 2007 Costly Signaling and the Ascendance of No-Can-Do
Archaeology: A Reply to Codding and Jones. American
Antiquity 72:358-366.
Mills, Barbara 2007 Performing the Feast: Visual Display and Supra
household Commensalism in the Puebloan Southwest. American Antiquity 72:210-240.
Nagaoko, Lisa 2001 Using Diversity Indices to Measure Changes in Prey
Choice at the Shag River Mouth Site, Southern New
Zealand. International Journal of Osteoarchaeology 11:101-111.
National Geographic Society 2002 Field Guide to the Birds of North America, 4th ed.
National Geographic Society, Washington D.C.
Pauketat, Timothy R., Lucretia S. Kelly, Gayle J. Fritz, Neal H.
Lopinot, Scott Elias, and Eve Hargrave 2002 The Residues of Feasting and Public Ritual at Early
Cahokia. American Antiquity 67:257-279.
Plog, Stephen 1998 Ancient Peoples of the American Southwest: Ancient
People and Places. Thames and Hudson, New York.
Potter, James M. 1997 Communal Ritual and Faunal Remains: An Example
from the Dolores Anasazi. Journal of Field Archaeology 24:353-364.
2000 Pots, Parties, and Politics: Communal Feasting in the
American Southwest. American Antiquity 65:471-492.
Pyke, G. H., H. R. Pulliam, and E. L. Charnov
1977 Optimal Foraging: A Selective Review of Theory and Tests. Quarterly Review of Biology 52:137-154.
Reid, James J., and Stephanie Whittlesey 1997 The Archaeology of Ancient Arizona. University of
Arizona Press, Tucson.
Rice, Glen R. (editor) 1987 Studies in the Hohokam Community of Marana. Ari
zona State University Anthropological Field Studies No.
15. Arizona State University, Department of Anthropol ogy, Tempe.
Rice, G., and C. Redman 1993 Platform Mounds of the Arizona Desert: An Experi ment in Organizational Complexity. Expedition 35:53-63.
Smith, Eric A.
1991 Inujjuamiut Foraging Strategies: Evolutionary Ecol
ogy of an Arctic Hunting Economy. Aldine de Gruyter, New
York. 2004 Why Do Good Hunters Have Higher Reproductive
Success? Human Nature 15(4):343-364. Smith, Eric A., and Bruce Winterhalder
1992 Evolutionary Ecology and Human Behavior. Aldine
de Gruyter, New York.
Speth, John D., and Katherine A. Spielmann 1983 Energy Source, Protein Metabolism, and Hunter
This content downloaded from 202.41.10.30 on Wed, 21 Jan 2015 07:15:45 AMAll use subject to JSTOR Terms and Conditions

864 AMERICAN ANTIQUITY [Vol. 75, No. 4, 2010]
Gatherer Subsistence Strategies. Journal of Anthropolog ical Archaeology 2:1-31.
Stebbins, Robert C. 2000 Peterson Field Guide to Western Reptiles and Amphib
ians, 3rd ed. Houghton Mifflin Company, New York.
Szuter, Christine R. 1989 Hunting by Prehistoric Horticulturalists in the Amer
ican Southwest. Unpublished Ph.D. dissertation, Depart ment of Anthropology, University of Arizona.
1991 Hunting by Prehistoric Horticulturalists in the Amer ican Southwest. Garland Publishing, New York.
Szuter, Christine R., and Frank E. Bayham 1989 Sedentism and Prehistoric Animal Procurement
Among Desert Horticulturalists of the North American Southwest. In Farmers as Hunters: The Implications of Sedentism, edited by Susan Kent, pp. 80-95. Cambridge University Press, Cambridge.
1997 Faunal Exploitation During the Late Archaic and Early Ceramic/Pioneer Periods in the South-Central Arizona. In
Early Formative Adaptations in the Southern Southwest, edited by Barbara J. Roth, pp. 65-72. Monographs in World
Archaeology No. 25. Prehistory Press, Madison.
Szuter, Christine R., and William B. Gillespie 1994 Interpreting the Use of Animal Resources at Prehis
toric American Southwest Communities. In The Ancient Southwestern Community: Models and Methods for the
study of Prehistoric Social Organization, edited by W. H. Wills and Robert D. Leonard, pp. 67-76. University of New Mexico Press, Albuquerque.
Ugan, Andrew 2005 Does Body Size Matter? Body Size, Mass Collect
ing, and Their Implications for Understanding Prehistoric
Foraging Behavior. American Antiquity 70:75-89.
van der Veen, Marijke 2003 When Is Food a Luxury? World Archaeology
34:405-427.
Wiessner, Polly, and Wulf Schiefenhovel (editors) 1996 Food and the Status Quest: An Interdisciplinary Per
spective. Berghahn Books, Oxford.
Winterhalder, Bruce, and Eric A. Smith 2000 Analyzing Adaptive Strategies: Human Behavioral
Ecology at Twenty-Five. Evolutionary Anthropology 9:51-72.
Winterhalder, Bruce, and Eric A. Smith (editors) 1981 Hunter Gatherer Foraging Strategies: Ethnographic
and Archaeological Analyses. University of Chicago Press,
Chicago. Whitaker, J. O.
1996 National Audubon Society Field Guide to North Amer ican Mammals. Alfred A. Knopf, New York.
Notes
1. For arguments on either side of this debate, see:
Broughton and Bayham (2003); Codding and Jones (2007); Gurven and Hill (2009); Hildebrandt and McGuire (2002,
2003), McGuire and Hildebrandt (2005); McGuire et al.
(2007); and Smith (2004).
Submitted April 4, 2009; Revised June 26, 2009; Accepted December 22, 2009.
This content downloaded from 202.41.10.30 on Wed, 21 Jan 2015 07:15:45 AMAll use subject to JSTOR Terms and Conditions