2012 world federation of hemophilia rnai-mediated inhibition of … · introduction the hemostatic...
TRANSCRIPT

2012 World Federation of Hemophilia
RNAi-Mediated Inhibition of Natural
Anticoagulants for Treatment of Hemophilia
July 10, 2012

Introduction The hemostatic system has evolved to balance the need to control blood loss with the need to
prevent thrombosis. This intricate balance is achieved through a sophisticated coagulation network
consisting of both procoagulant and anticoagulant factors, interconnected through various regulatory
feedback loops. In hemophilia, the loss of certain procoagulant factors (Factor VIII (FVIII) and Factor
IX (FIX), in the case of hemophilia A and B, respectively) results in an imbalance of the hemostatic
system toward a bleeding phenotype. In contrast, in thrombophilia (e.g. Factor V Leiden, protein C
deficiency, protein S deficiency, antithrombin deficiency, prothrombin G20210A), certain mutations
result in an imbalance in the hemostatic system toward a thrombotic phenotype. Interestingly, there
have been reports suggesting that coinheritance of prothrombotic mutations may ameliorate the
clinical phenotype in hemophilia [1-4].
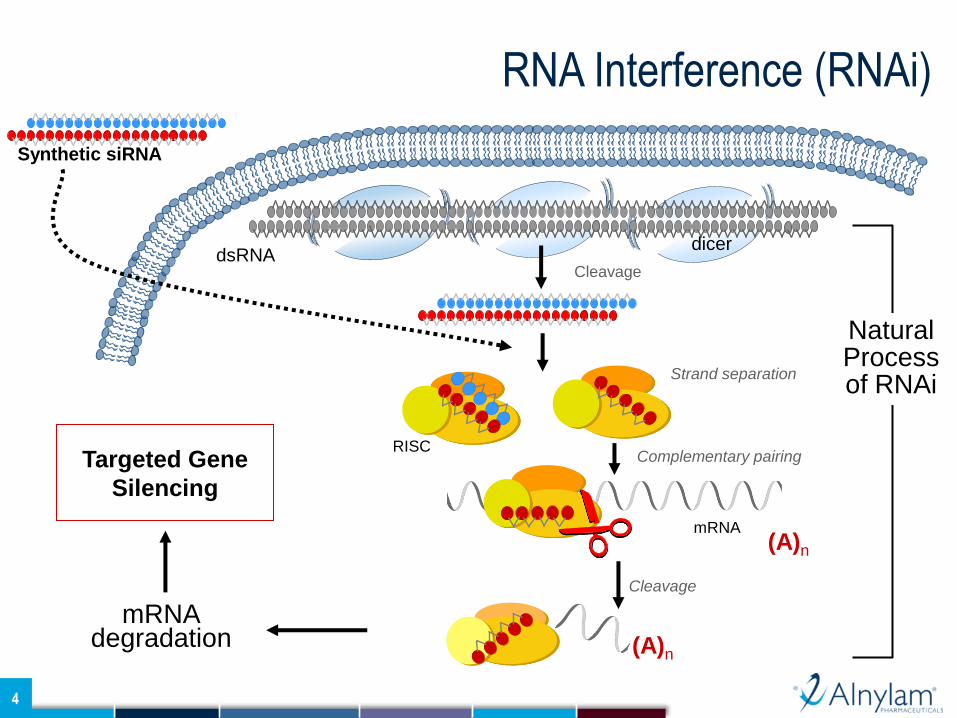
In this work, we investigated the use of RNA interference (RNAi) to target the natural anticoagulants
protein C (PC) and antithrombin (AT) as a strategy to rebalance the hemostatic system and improve
thrombin generation in hemophilia. We developed potent short interfering RNAs (siRNAs) against
both PC and AT and demonstrated effective silencing of both targets in vivo. In silico modeling
results, as well as human genetic data [5], suggested AT to be the more powerful anticoagulant, so
subsequent efforts focused on AT silencing. An siRNA employing a hepatocyte-targeting ligand was
developed against AT and demonstrated potent activity in both mice and non-human primates after
single subcutaneous (SC) administration (ED50 ~ 1 mg/kg). Thrombin generation assays (TGA)
demonstrated proof-of-concept that AT reduction in human plasma deficient in coagulation factor as
well as hemophilic mouse models leads to normalization of thrombin generation. These data
suggest that the use of a novel RNAi therapeutic targeting AT is a promising approach for the
treatment of hemophilia. Further, the SC route of administration, long duration of action, and
applicability to hemophiliacs with inhibitors, make this a particularly exciting potential therapy.
2
Therapeutic Hypothesis Rebalancing the Hemostatic System
PS
Intrinsic system
FIXa
FVIIa FVII
FVIIIa
Extrinsic system
FVa FV
FX
FXa
Fibrinogen Fibrin
Thrombin
Hemophilia B
Hemophilia A
Prothrombin
FIX
FVIII
Blood clot
TFPI
AT
APC PC
3
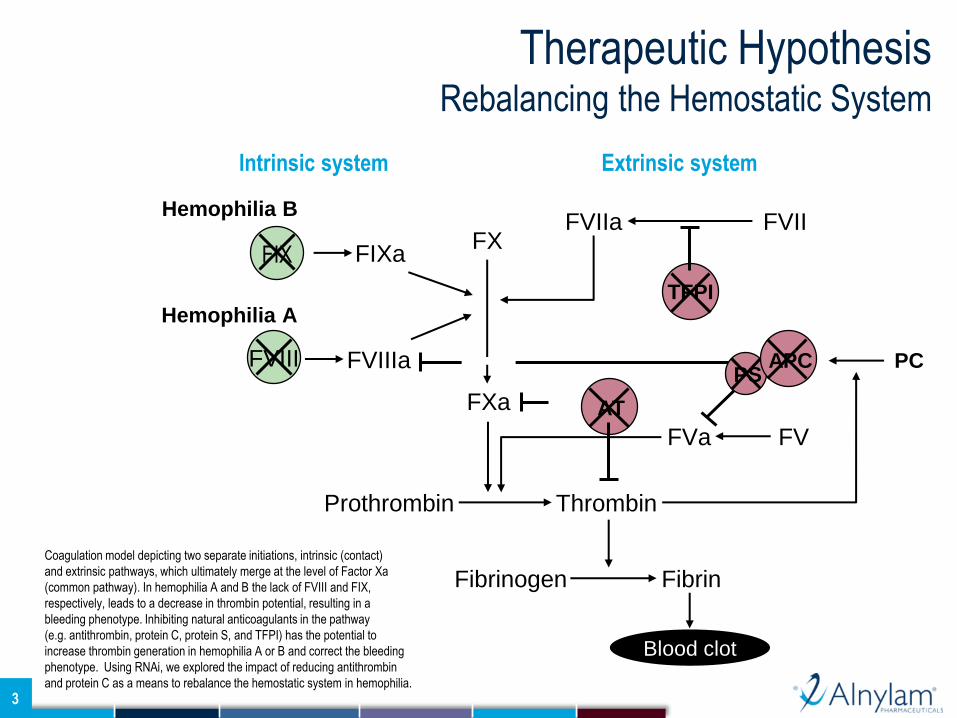
Coagulation model depicting two separate initiations, intrinsic (contact)
and extrinsic pathways, which ultimately merge at the level of Factor Xa
(common pathway). In hemophilia A and B the lack of FVIII and FIX,
respectively, leads to a decrease in thrombin potential, resulting in a
bleeding phenotype. Inhibiting natural anticoagulants in the pathway
(e.g. antithrombin, protein C, protein S, and TFPI) has the potential to
increase thrombin generation in hemophilia A or B and correct the bleeding
phenotype. Using RNAi, we explored the impact of reducing antithrombin
and protein C as a means to rebalance the hemostatic system in hemophilia.
RNA Interference (RNAi)
mRNA
mRNA degradation
dsRNA dicer
Cleavage
Strand separation
Complementary pairing
Cleavage
(A)n
(A)n
Natural Process of RNAi
Synthetic siRNA
Targeted Gene
Silencing
RISC
4

RNAi-Mediated In Vivo Silencing
of Natural Anticoagulants
5
Protein C Antithrombin
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.0 0.003 0.01 0.03 0.1 0.3 1.0
siCont
(mg/kg) siAPC
(mg/kg)
Rela
tiv
e P
rote
in C
mR
NA
(siC
on
t =
1)
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.0 0.003 0.01 0.03 0.1 0.3 1.0
siAT3
(mg/kg)
Rela
tiv
e A
nti
thro
mb
in m
RN
A (
siC
on
t =
1)
siCont
(mg/kg)
Using bioinformatics and in vitro screening approaches, siRNAs were developed against the natural anticoagulants protein C (siPC) and antithrombin (siAT). siPC,
siAT, as well as a control non-targeting siRNA (siCont), were formulated in lipid nanoparticles and administered to wild-type mice via tail vein injection (10 mL/kg).
Animals were sacrificed at 24 hours post-administration and livers were harvested for mRNA analysis by quantitative RT-PCR. Protein C and antithrombin mRNA
levels were normalized to GAPDH mRNA levels and the data were scaled to the control group. Administration of both (A) siPC and (B) siAT resulted in potent,
dose-dependent silencing of protein C and antithrombin, respectively. Both targets were found to be amenable to RNAi-mediated silencing in vivo. Bars represent
group mean ± standard deviation (N = 5).
In Silico Modeling of Thrombin Generation Modeling Parameter Values
Target Protein Protein C and antithrombin
Target Protein Levels 0, 5, 10, 20, 50, 80, and 100%
of normal
FVIII Levels 0, 0.2, and 100% of normal
Thrombomodulin Levels 0.1 and 10 nM
6
FVIII = 0%, TM = 0.1 nM
Normal
(FVIII = 100%)
Example Thrombin Generation Calculations Impact of Target Silencing on Max Thrombin
0
10
20
30
40
50
60
70
0 600 1200 1800 2400 3000 3600 4200 4800 5400 6000
Th
rom
bin
(n
M)
Time (sec)
0% PC
5% PC
10% PC
20% PC
50% PC
80% PC
100% PC
0
100
200
300
400
500
0 20 40 60 80 100
Ma
x T
hro
mb
in L
eve
l (n
M)
Protein Level (%)
FVIII = 0%, TM = 0.1 nM
Protein C
Antithrombin
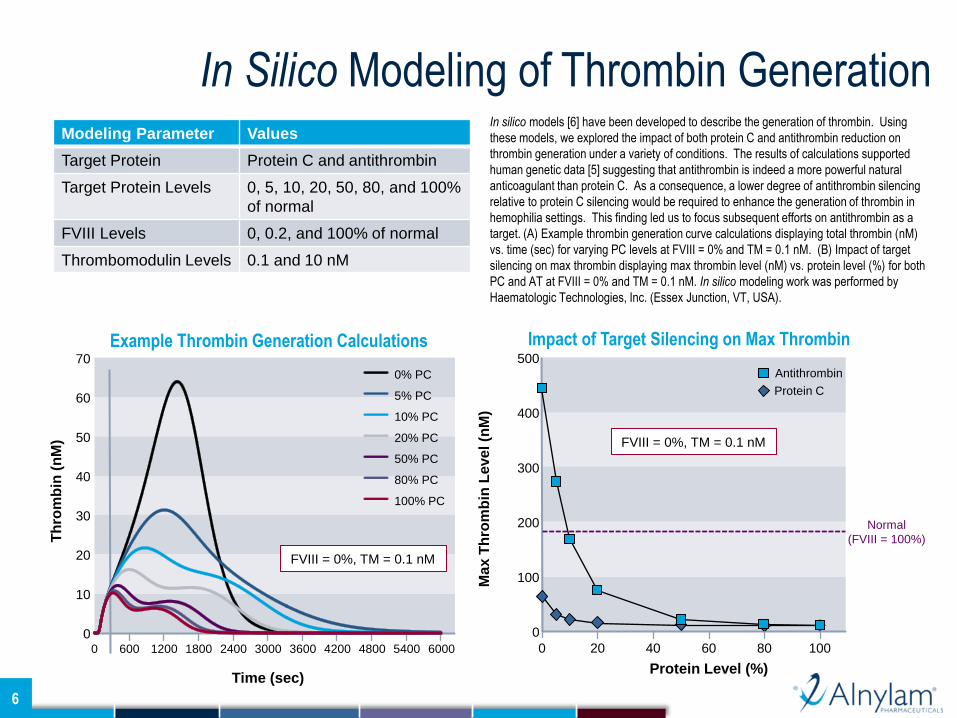
In silico models [6] have been developed to describe the generation of thrombin. Using
these models, we explored the impact of both protein C and antithrombin reduction on
thrombin generation under a variety of conditions. The results of calculations supported
human genetic data [5] suggesting that antithrombin is indeed a more powerful natural
anticoagulant than protein C. As a consequence, a lower degree of antithrombin silencing
relative to protein C silencing would be required to enhance the generation of thrombin in
hemophilia settings. This finding led us to focus subsequent efforts on antithrombin as a
target. (A) Example thrombin generation curve calculations displaying total thrombin (nM)
vs. time (sec) for varying PC levels at FVIII = 0% and TM = 0.1 nM. (B) Impact of target
silencing on max thrombin displaying max thrombin level (nM) vs. protein level (%) for both
PC and AT at FVIII = 0% and TM = 0.1 nM. In silico modeling work was performed by
Haematologic Technologies, Inc. (Essex Junction, VT, USA).

Antithrombin Reduction Increases Thrombin Generation in FIX-Depleted Human Plasma In Vitro
7
0
100
200
300
400
0 10 20 30 40 50 60
Th
rom
bin
(n
M)
Time (min)
0% FIX, 100% AT
0% FIX, 75% AT
0% FIX, 50% AT
0% FIX, 25% AT
0% FIX, 10% AT
0% FIX, 5% AT
0% FIX, 0% AT
100% FIX, 100% AT
0
100
200
300
400
500
600
100 75 50 25 10 5 0
Pe
ak
Th
rom
bin
(n
M)
% Antithrombin
100% FIX,
100% AT
Thrombin Generation in FIX- and AT-Depleted Human Plasma Peak Thrombin in FIX- and AT-Depleted Human Plasma
To investigate the impact of antithrombin reduction on thrombin generation in a hemophilia setting, thrombin generation studies were performed in FIX and AT
double-depleted human plasma. AT was then added back to the plasma at various levels (1 IU/mL = 100%) to generate FIX-depleted plasmas with different levels
of AT (0-100%). Control plasma was generated by adding back 1 IU/mL AT (100%) and 5 µg/mL FIX (100%) to the double-depleted plasma. (A) Thrombin
generation in FIX- and AT-depleted human plasma (tissue factor = 5 pM). (B) Peak thrombin in FIX- and AT-depleted human plasma (tissue factor = 5 pM).
Plasma depletion and TGA work was performed by Haematologic Technologies, Inc. (Essex Junction, VT, USA).
siRNA Conjugate Approach for Targeted Delivery to Hepatocytes
8
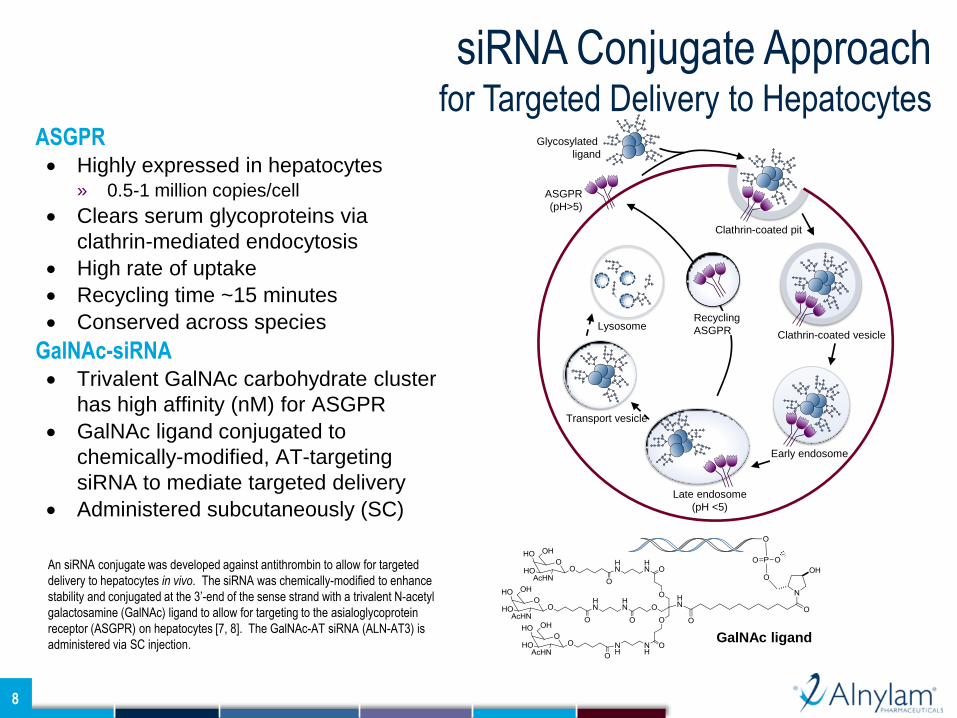
ASGPR Highly expressed in hepatocytes
» 0.5-1 million copies/cell
Clears serum glycoproteins via
clathrin-mediated endocytosis
High rate of uptake
Recycling time ~15 minutes
Conserved across species
GalNAc-siRNA Trivalent GalNAc carbohydrate cluster
has high affinity (nM) for ASGPR
GalNAc ligand conjugated to
chemically-modified, AT-targeting
siRNA to mediate targeted delivery
Administered subcutaneously (SC)
GalNAc ligand
ASGPR
(pH>5)
Glycosylated
ligand
Clathrin-coated pit
Clathrin-coated vesicle
Early endosome
Late endosome
(pH <5)
Lysosome
Transport vesicle
Recycling
ASGPR
An siRNA conjugate was developed against antithrombin to allow for targeted
delivery to hepatocytes in vivo. The siRNA was chemically-modified to enhance
stability and conjugated at the 3’-end of the sense strand with a trivalent N-acetyl
galactosamine (GalNAc) ligand to allow for targeting to the asialoglycoprotein
receptor (ASGPR) on hepatocytes [7, 8]. The GalNAc-AT siRNA (ALN-AT3) is
administered via SC injection.
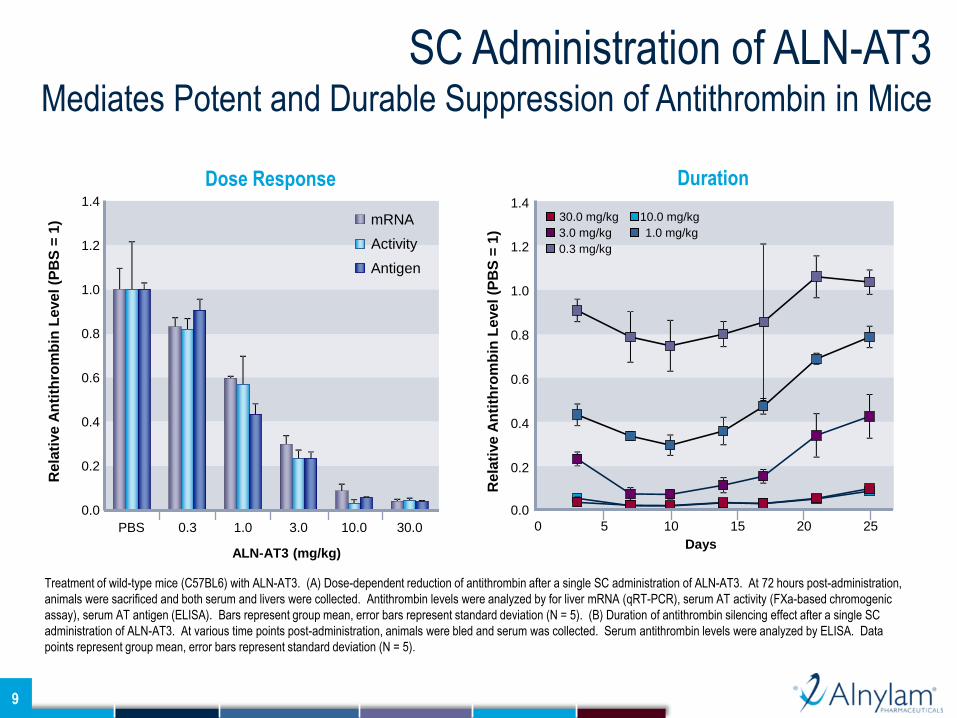
SC Administration of ALN-AT3 Mediates Potent and Durable Suppression of Antithrombin in Mice
9
Dose Response
mRNA
Activity
Antigen
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
0.3 1.0 3.0 10.0 30.0 PBS
ALN-AT3 (mg/kg)
Re
lati
ve
An
tith
rom
bin
Leve
l (P
BS
= 1
)
Duration
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
0 5 10 15 20 25
Re
lati
ve
An
tith
rom
bin
Leve
l (P
BS
= 1
)
Days
30.0 mg/kg 10.0 mg/kg
3.0 mg/kg 1.0 mg/kg
0.3 mg/kg
Treatment of wild-type mice (C57BL6) with ALN-AT3. (A) Dose-dependent reduction of antithrombin after a single SC administration of ALN-AT3. At 72 hours post-administration,
animals were sacrificed and both serum and livers were collected. Antithrombin levels were analyzed by for liver mRNA (qRT-PCR), serum AT activity (FXa-based chromogenic
assay), serum AT antigen (ELISA). Bars represent group mean, error bars represent standard deviation (N = 5). (B) Duration of antithrombin silencing effect after a single SC
administration of ALN-AT3. At various time points post-administration, animals were bled and serum was collected. Serum antithrombin levels were analyzed by ELISA. Data
points represent group mean, error bars represent standard deviation (N = 5).

Single SC Administration of ALN-AT3 in Non-Human Primates
10
Pre-Dose Level
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.0 3.0 10
ALN-AT3 (mg/kg)
Rela
tive S
eru
m A
T (
Pre
-do
se =
1) Activity of ALN-AT3 in non-human primates. Cynomolgus
monkeys were administered ALN-AT3 at the dose levels
indicated via single SC injection. Serum was collected
one week after administration and analyzed for
antithrombin protein level by ELISA. Antithrombin levels
are represented relative to the average of three pre-dose
measurements. Dose dependent AT silencing was
observed, with approximately 45, 55, and 70% silencing at
1, 3, and 10 mg/kg, respectively. Bars represent group
mean, error bars represent standard deviation (N = 3).

Thrombin Generation in HB Mice
0
10
20
30
40
50
0 10 20 30 40
Th
rom
bin
(n
M)
Time (min)
ALN-AT3 Activity in Hemophilia Mouse Models
11
Dose Response in HA Mice
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
1.0 3.0 10 30 PBS
ALN-AT3 (mg/kg)
Rela
tive S
eru
m A
T (
PB
S =
1)
Normalization of ETP in HB Mice
0
100
200
300
400
500
600
WT
Control
HB
Control
ALN-AT3 Treated
ET
P (
nM
min
)
WT Control
HB Control
ALN-AT3 treated #1
ALN-AT3 treated #2
ALN-AT3 treated #3
A) Dose-dependent reduction of antithrombin in Hemophilia A mice (B6;129S4-F8tm1Kaz/J) after a single SC administration of ALN-AT3. At 72 hours post-administration, animals (2 M/2 F)
were sacrificed and serum was collected. Antithrombin levels were analyzed for serum AT antigen (ELISA). Bars represent group mean, error bars represent standard deviation (N = 4).
(B), (C) Treatment of Hemophilia B mice (FIX < 1 IU/dL) with 30 mg/kg ALN-AT3. At 72 hours post-administration, animals (N = 3) were sacrificed and plasma was collected. (B) Thrombin
generation in HB mice (tissue factor = 0.5 pM). Curves depict data for representative WT mouse treated with PBS, representative HB mouse treated with PBS, and three HB mice treated
with ALN-AT3. (C) Endogenous thrombin potential (ETP) (AUC) for WT mice treated with PBS (N = 2), HB mice treated with PBS (N = 3), and HB mice treated with ALN-AT3 (N = 3). Bars
represent group mean, error bars represent standard deviation.

ALN-AT3: Hemophilia Program Conclusions
Subcutaneously administered RNAi therapeutic targeting
antithrombin for the treatment of hemophilia
ALN-AT3 treatment results in potent inhibition of AT in mice and
non-human primates (ED50 ~ 1 mg/kg)
Given long duration of action of ALN-AT3, once-weekly or twice-
monthly SC dosing is envisioned
Proof-of-concept has been achieved in human factor-depleted
plasma and in hemophilia mouse models demonstrating
normalization of thrombin generation after ALN-AT3 treatment
Hemostatic rebalancing approach utilizing ALN-AT3 represents
potentially new prophylaxis therapy option for hemophilia patients,
including those with inhibitors
12

References
Escuriola-Ettingshausen et al., Thromb Haemost; 85:218-220 (2001)
Kurnik et al., Hematologica; 92:982-985 (2007)
Lee et al., Thromb Haemost, 83:387-391 (2000)
Negrier et al., Blood; 81:690-695 (1993)
Bucciarelli et al., Arterioscler Thromb Vasc Biol; 19:1026-1033 (1999)
Hockin et al., J Biol Chem; 277:18322-18333 (2002)
Ashwell and Morell, Adv Enzymol Relat Areas Mol Biol; 41:99-128 (1974)
Cummings and McEver, C-Type Lectins. In Varki et al. (Eds), Essentials
of Glycobiology (2nd Edition, 2008)
13

Acknowledgements
Alnylam Pharmaceuticals A. Sehgal J. Brodsky I. Toudjarska T. Racie J. Qin S. Barros J. Hettinger D. Foster S. Milstein K. Charisse S. Kuchimanchi M. Maier B. Bettencourt Y. Jiang A. Akinc
Queen’s University, Kingston,
Canada D. Lillicrap
Edouard Herriot University
Hospital, Lyon, France Y. Dargaud
C. Negrier
14