1 infectious endogenous retroviruses in cats and emergence of recombinant viruses yukari
TRANSCRIPT

1
Infectious Endogenous Retroviruses in Cats and Emergence of Recombinant 1
Viruses 2
Yukari Anai1, Haruyo Ochi1, Shinya Watanabe1, So Nakagawa2,3, Maki Kawamura1, 3
Takashi Gojobori2 and Kazuo Nishigaki1,* 4
1Laboratory of Molecular Immunology and Infectious Disease, Joint Faculty of 5
Veterinary Medicine, Yamaguchi University, 1677-1 Yoshida, Yamaguchi, Japan 6
2Center for Information Biology and DNA Data Bank of Japan, National Institute of 7
Genetics, 1111 Yata, Mishima, Shizuoka, Japan 8
3present address: Department of Organismic and Evolutionary Biology, Harvard 9
University, Cambridge, MA 02138, USA; Research Fellow of the Japan Society for the 10
Promotion of Science 11
Running Title: Infectious ERV and emergence of recombinant virus 12
13
*Corresponding author: Kazuo Nishigaki, DVM, Ph.D. 14
Joint Faculty of Veterinary Medicine, Yamaguchi University 15
1677-1 Yoshida, Yamaguchi, Japan 16
Phone: +81-83-933-5829 17
Fax: +81-83-933-5820 18
E-mail: [email protected] 19
20
21
22
Copyright © 2012, American Society for Microbiology. All Rights Reserved.J. Virol. doi:10.1128/JVI.00280-12 JVI Accepts, published online ahead of print on 6 June 2012
on April 10, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

2
Abstract 23
Endogenous retroviruses (ERVs) comprise a significant percentage of the mammalian 24
genome, and it is poorly understood whether they will remain as inactive genomes or 25
emerge as infectious retroviruses. Although several types of ERVs are present in 26
domestic cats, infectious ERVs have not been demonstrated. Here we report a 27
previously uncharacterized class of endogenous gammaretroviruses, termed ERV-DCs, 28
that is present and hereditary in the domestic cat genome. We have characterized a 29
subset of ERV-DC proviral clones, numbered according to their genomic insertions. 30
One of these, ERV-DC10 located in the q12-q21 region on chromosome C1, is an 31
infectious gammaretrovirus capable of infecting a broad range of cells including human. 32
Our studies indicate that ERV-DC10 entered the genome of domestic cats in the recent 33
past and appeared to translocate to or reintegrate at a distinct locus as an infectious 34
ERV-DC18. Insertional polymorphism analysis revealed that 92 of 244 domestic cats 35
had ERV-DC10 on homozygous or heterozygous locus. ERV-DC-like sequences were 36
found in primate and rodent genomes, suggesting that these ERVs and recombinant 37
viruses such as RD-114 and BaEV may have originated from an ancestor of ERV-DC. 38
We also found that a novel recombinant virus, FeLV-D, was generated by ERV-DC env 39
transduction into feline leukemia virus in domestic cats. Our results indicate that 40
ERV-DCs behave as donors and/or acceptors in the generation of infectious, 41
recombinant viruses. The presence of such infectious endogenous retroviruses, which 42
could be harmful or beneficial to the host, may impact veterinary medicine and public 43
health. 44
45
on April 10, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

3
Introduction 46
Retroviruses are classified as exogenous or endogenous according to their mode of 47
transmission. Endogenous retroviruses (ERVs) are present in all vertebrate genomes and 48
are thought to be the remnants of ancestral germline infections by exogenous 49
retroviruses. ERVs make up a significant fraction of the mammalian genome (for 50
example, 8–10% of the human or mouse genomes) and are transmitted according to 51
Mendelian fashion (4,19,42). Most ERVs contain deleterious mutations – large deletions 52
and insertions of repetitive elements that render them unable to make infectious viral 53
particles, but these proviruses often are transcribed and produce functional gene 54
products (4). Domestic cats (Felis silvestris catus) are descended from Felis silvestris 55
lybica, the Middle Eastern wildcat (10), and are sometimes infected with feline 56
leukemia virus (FeLV), which is a gammaretrovirus that is horizontally transmitted 57
among domestic cats during grooming or through bites (13,14). The endogenous 58
counterparts of exogenous FeLV, termed enFeLVs, are integrated into the genomes of 59
the Felis genus. Primary colonization of the feline germline by FeLVs is believed to 60
have occurred in an ancestor of the domestic cat after separation from the leopard 61
lineage (2,13,31). Like most endogenous retroviruses, enFeLVs are non-infectious and 62
non-pathogenic (4). However, recombination occurs between inherited enFeLVs and 63
exogenous FeLVs (exFeLVs) following infection, which can give rise to new infectious 64
agents that may be pathogenic (13,29). In addition to enFeLV, other feline ERVs are 65
found in the domestic cat. The RD-114 ERV, which was originally isolated from human 66
rhabdomyosarcoma cells transplanted into fetal kittens (23), is a replication competent 67
retrovirus which shows a high level of homology to the baboon endogenous retrovirus 68
(BaEV); however, an infectious RD-114 provirus has not been demonstrated in the cat 69
on April 10, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

4
genome. MAC-1, which is a genetically transmitted type-C virus of primates, has 70
been reported as a feline non-infectious ERV (5,39); however, no further studies have 71
been reported. Another class of feline ERVs with high homology to the pol region of 72
BaEV and RD-114 has also been described and partially characterized as ECE1 and 73
FcEV (3,41). In the present study, we have further characterized this later class of 74
feline ERVs, which we term ERV-DCs. Our studies indicate that while most 75
ERV-DCs contain deletions and mutations that render them non-infectious, several 76
ERV-DCs are infectious and mobile in the domestic cat, using re-infection, translocation 77
to another locus or recombination with an exogenous retrovirus to amplify their progeny. 78
These infectious ERV-DCs can infect a wide range of mammalian cells, including 79
human. Most ERV-DCs are not yet fixed in the cat genome and have the potential to 80
generate recombinant retroviruses which could be transmitted among cats and other 81
species. 82
83
Materials & Methods 84
Samples. Blood for peripheral blood mononuclear cells (PBMC) isolation and spleen 85
and muscle tissues from domestic cats and Prionailurus bengalensis euptilurus were 86
collected at private veterinary hospitals (27) and Tsushima Wildlife Conservation Center 87
(TWCC) in Japan. 88
89
Expression vectors and cell lines. The env of FeLV-A/Glasgow-1 (pFGA5) (38), 90
FeLV-B/Gardner-Arnstein (pFGB)(25), FeLV-C/Sarma (pFSC)(33), FeLV-D/Ty26, 91
on April 10, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

5
FeLV-D/ON-T and ERV-DC10 were PCR-amplified with specific primers and cloned 92
into pFUΔss expression plasmid, which was a modified pFUSE-hIgG1-Fc2 vector 93
(Invitrogen, Carlsbad, CA). The nβ-gal gene (kindly provided by Masaaki Nakaya, 94
Yokohama City University, Kanagawa) was introduced into the pMXs-IP vector (18) 95
(kindly provided by Toshio Kitamura, The University of Tokyo, Tokyo) to produce 96
pMXs-nLIP. HEK293T cells were persistently infected with FeLV-A (clone33)(28), 97
FeLV-B (pFGB), FeLV-C (pFSC), ERV-DC10 (pDC10) or ERV-DC18(pDC18). 293T 98
cells expressing FeLV-D/ON-T were established by transfection with a full-length 99
FeLV-D/ON-T plasmid and then were infected with FeLV-A/clone33 for viral 100
interference assays. An env-negative packaging cell line containing a LacZ-coding 101
retroviral vector, designated GPLac, was also established. 102
103
RNA isolation and reverse transcription. Total RNA was isolated using a FastPure 104
RNA Kit and cDNA was synthesized with a PrimeScript® II 1st strand cDNA Synthesis 105
Kit (TaKaRa, Tokyo, Japan). Viral RNA was isolated from infected cell culture 106
supernatants using the QIAamp Viral RNA mini kit (QIAGEN, Tokyo, Japan). 107
108
PCR. Primers used in this study are listed in Supplementary Table 4. PrimeSTAR GXL 109
polymerase (TaKaRa), KOD FX Neo (TOYOBO, Japan) and PrimeSTAR HS DNA 110
polymerase (TaKaRa) were used for various cloning procedures. The latter two 111
enzymes were also used to screen for FeLV-D and monitor viral infection. KOD FX 112
Neo and GoTaq (Promega, Madison, WI) were used for genotyping insertional 113
polymorphisms. 114
on April 10, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

6
115
Southern blotting. A total of 7.5 µg sample DNA was isolated from peripheral blood 116
mononuclear cells (PBMCs) or tissue samples (34), digested with EcoRI and BamHI 117
and hybridized to a digoxigenin (DIG)-labelled FeLV-D/ON-T env probe generated by 118
PCR with primers Fe-9S and Fe-3R (Supplementary Table 4). A CSPD Luminescent 119
detection kit was used to visualize the bound probe (Roche Molecular Biochemicals, 120
Mannheim, Germany). 121
122
Cloning of ERV-DC proviruses. Splenic DNA from a single cat (ON-T) was digested 123
with EcoRI or Sau3AI and ligated to Lamda DASH II vectors (Stratagene, La Jolla, CA). 124
DNA libraries were screened with a DIG-labelled FeLV-D env probe. ERV-DC 125
proviruses and flanking sequences were obtained from the NCBI cat genome database; 126
other sequence data were obtained by screening genomic libraries. We PCR-amplified 127
full-length ERV-DC and partial ERV-DC proviruses from feline chromosomal DNA 128
using specific primers and cloned amplicons into pCR4 blunt-TOPO (Invitrogen, 129
Carlsbad, CA). Some flanking sequences of ERV-DCs were obtained by gene walking 130
using the Right Walk® kit (BEX CO., LTD., Tokyo, Japan). Pre-integration sites and 131
ERV-DC provirus were detected by PCR. 132
FeLV-D screening and cloning. DNA from FeLV-positive PBMCs or tumour tissues 133
was PCR-screened using primers Fe-14S and Fe-3R. Gene walking was performed to 134
obtain the flanking DNA sequences of FeLV-D/ON-T provirus using each Fe-14S (first 135
PCR) and Fe-2S (second PCR) primer with each adaptor primer (Right Walk kit, BEX 136
on April 10, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

7
CO., LTD.). After obtaining one sequence of the pre-integration sites, the full length 137
FeLV-D/ON-T proviral genome was PCR-amplified with Prime STAR HS (TaKaRa) 138
DNA polymerase and Fe-63S and Fe-73R primers and cloned into pCR4 blunt-TOPO. 139
Partial FeLV-D/Ty26, FeLV-D/ON-C, FeLV/44B and FeLV-D/re-T sequences were 140
obtained by PCR. 141
142
Immunoblotting. Immunoblotting was performed as previously described (40). 143
Primary antibodies were goat anti-RD114 (National Cancer Institute (NCI), Frederick, 144
MD), goat anti-FeLV gp70 (NCI) and mouse anti-beta actin (Santa Cruz Biotechnology, 145
Santa Cruz, CA). Secondary antibodies were horseradish peroxidase (HRP)-conjugated 146
anti-mouse IgG antibody (GE Healthcare Japan, Tokyo, Japan) or anti-goat IgG 147
antibody (Santa Cruz Biotechnology). 148
149
Infection assay. HEK293T cells were transfected with 4 μg ERV-DC10 or -DC18 150
plasmid DNA using Lipofectamine 2000 (Invitrogen). Viruses in the culture supernatant 151
collected 72 h post-transfection were detected by PCR. Supernatants (300 μl, 152
0.22-μm-filtered) from persistently infected HEK293T cells were used to infect Molt-4, 153
K562, HepG2, HeLa, MCF7, THP-1, Vero, Cos7, L929, BHK-21 and NIH3T3 cells 154
(3.65 × 105 cells/well) in the presence of polybrene (8 μg/ml). KwDM cells established 155
from dog mammary tumour were also used for infection assay. Cellular DNA was 156
collected and ERV-DC proviral DNA was detected by PCR using primers Fe-13S and 157
Fe-18R. 158
on April 10, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

8
GFP-containing pMXs-IP vector with or without ERV-DC10 or ERV-DC18 sequences 159
was also transfected into HEK293T cells. At 72 h post-transfection, CRFK, AH927 and 160
G355 cell lines were infected with the HEK293T supernatant in the presence of 161
polybrene (8 μg/ml). GFP-positive cells were determined by microscopy or FACS. 162
Viral preparation from feline chromosomal DNA. ERV-DC8, ERV-DC10, 163
ERV-DC14 or ERV-DC18 amplified by PCR with primers Fe-57S and Fe-54R for 164
ERV-DC8, Fe-122S and Fe-38R for ERV-DC10, Fe-58S and Fe-56R for ERV-DC14 or 165
Fe-76S and Fe-81R for ERV-DC18 using feline chromosomal DNA was directly 166
transfected to HEK293T cells and the filtered supernatant was used to infect fresh 167
HEK293T cells. Viral infection was monitored by detecting proviral ERV-DC env and 168
LTR or immunoblot analysis using anti-RD114 antibody. Viral transmission was 169
confirmed by further infection. 170
Transmission electron microscopy. HEK293T cells persistently infected with 171
ERV-DC10 were washed with D-PBS, fixed with 2% glutaraldehyde at 4°C for at least 172
2 h, washed with D-PBS (2 × 15 min) and post-fixed with 2% osmium tetroxide for 1.5 173
h. Cells were dehydrated through a graded ethanol series and propylene oxide, then 174
embedded in EPON 812 for 48 h at 60°C. Ultrathin sections were stained with uranyl 175
acetate and lead citrate, and examined with an electron microscope (JEM-1200EX; 176
JEOL, Japan). 177
178
Viral interference assays. GPLac cells (3 × 106) were transfected with each 179
env-expressing plasmid to produce pseudotype viruses. After culturing with 200 μg/ml 180
on April 10, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

9
zeocin and 1 μg/ml puromycin for >2 weeks, supernatants were collected, filtered 181
through a 0.22-μm filter and stored at -80°C. Target cells (105) were infected with viral 182
supernatant in the presence of polybrene (2 μg/ml) for 2 days, then stained with X-gal 183
(5-bromo-4-chloro-3-indolyl-β-D-galactopyranoside). Single-cycle infectivity was 184
titrated by counting blue-stained nuclei using a microscope following incubation with 185
X-gal. X-gal-positive cells were considered infectious units. 186
187
Fluorescence in situ hybridization (FISH) analysis. PBMC cultures stimulated with 188
Concanavalin A were treated with 5-bromo-2′-deoxyuridine for 5.5 h and harvested 189
after colcemid treatment for 0.5 h. Replication-banded chromosomes were obtained by 190
exposure of the chromosome slides to UV light after staining with Hoechst 33258. The 191
pTKDC10L2.8 (2.8 kb of ERV-DC10 flanking sequences by PCR-amplified with 192
primers, Fe-137S (5'-ATTCCTTAGCTCATGGTCCTTTTCT-3') and Fe-38R 193
(5'-CACACATGCTCTAGACACAATACCC-3')) and pTKDC18L3.4 (3.4 kb of 194
ERV-DC18 flanking sequences by PCR-amplified with primers, Fe-146S 195
(5'-GTCAATTGGACTGCCTTGAATCTAT-3') and Fe-138R 196
(5'-AAAGTTCCCAGATAATCCTGATGGT-3')) plasmid DNAs were labelled with 197
digoxigenin-11-dUTP by nick translation for use as probes. After hybridization, the 198
slides were washed and probe signals were detected with Cy3-labelled anti-digoxigenin. 199
FISH images were captured with the CW4000 FISH application program (Leica 200
Microsystem, Wetzlar, Germany). 201
Phylogenetic and genomic analyses 202
Nucleotide sequences of ERV-DC regions and amino acid sequences of Pol and Env 203
on April 10, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

10
used in the phylogenetic analyses are summarized in Supplementary Table 5. For these 204
analyses, we generated multiple alignments for each gene using MAFFT (L-INS-i)(16). 205
Amino acid substitution models of LG (20) with gamma-distributed rate variation (+Γ, 206
α = 1.672557) and rtREV(8) with gamma-distributed rate variation and inferred 207
proportion of invariable sites (+Γ+I, α = 0.912332, invar = 0.000117) were selected for 208
Env and Pol alignments, respectively, using the Akaike information criterion 209
implemented in PROTTEST 3 (6). Phylogenetic trees were constructed using the 210
maximum-likelihood method (12) in RAxML v7.2.8 with robustness evaluated by rapid 211
bootstrapping (1000 times). To identify ERVs we used RetroTector©(35) and default 212
parameters for genomes of various species provided by Ensembl 213
(http://www.ensembl.org/). Four types of retroviral-like sequences in the genome were 214
identified and we conducted a homology search with BLASTP using the Pol or Env 215
sequences of ERV-DC as a query. 216
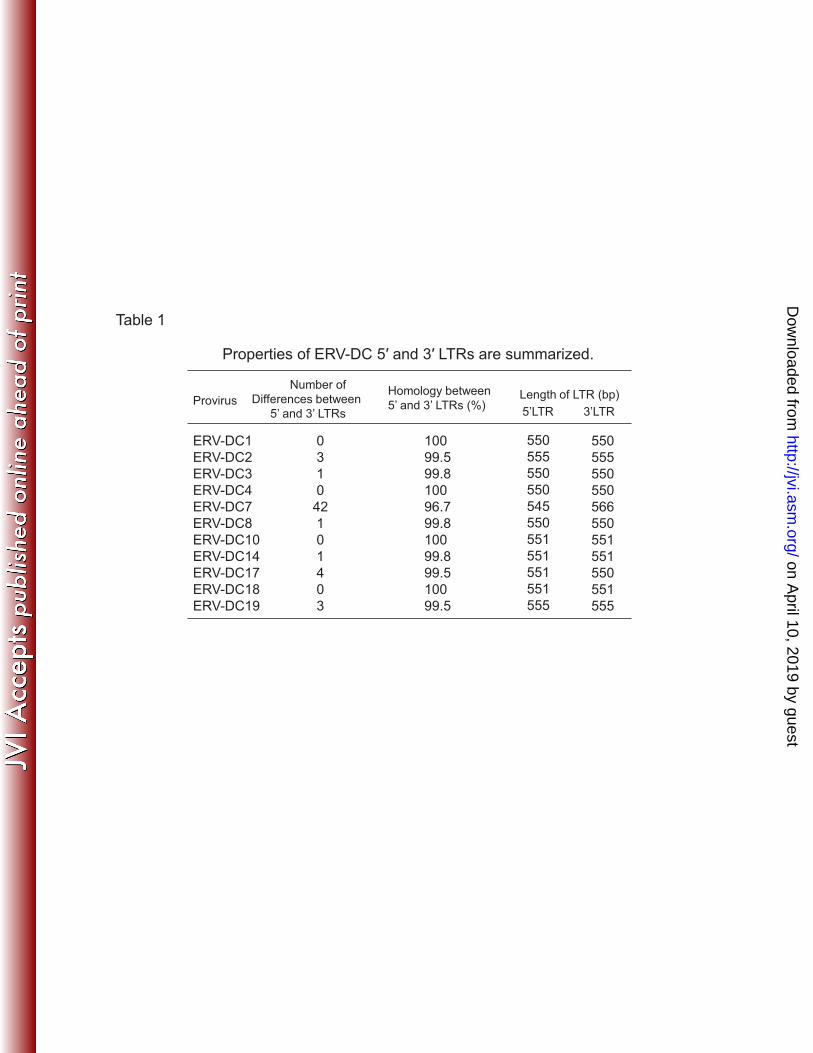
Time estimation of ERV-DC insertion. Because the mutation rate of LTRs in 217
ERV-DCs is unknown, we alternatively utilized a mean divergence rate of non-coding 218
regions in the domestic cat genome (1.2 × 10–8 substitutions/site/year)(21). Genetic 219
distances between LTRs were calculated using the Kimura two-parameter model (17). 220
221
on April 10, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

11
Results 222
Characterization of endogenous gammaretroviruses in the domestic cat 223
(ERV-DCs) 224
Ten full-length and three partial feline endogenous gammaretroviruses, which were 225
previously partially characterized as ECE1 and FcEV (3,41), were isolated by genomic 226
library screening and PCR using locus-specific primers. Sequence analysis revealed that 227
four of these proviruses (ERV-DC8, DC10, DC14 and DC18) have intact ORFs for gag, 228
pol and env (Fig. 1) and could potentially produce infectious retroviruses. Although 229
ERV-DC10 and 18 were isolated from different cats and had different integration loci, 230
their ERV-DC nucleotide sequences were almost identical. Only 1 bp out of 8863 in the 231
primer binding site was different between ERV-DC10 and 18. The gag gene had a 232
predicted size of 1644 bp and encoded a putative 547-aa protein. The pol gene had a 233
predicted size of 3570 bp and the deduced Pol protein encoded a putative protease, 234
reverse transcriptase, RNaseH and integrase. It is likely that the Gag and Pol proteins 235
are synthesized as a single large polypeptide precursor via termination suppression. The 236
env ORF of ERV-DC10/18 was located in a different reading frame than that of the 237
gag–pol gene. The env gene had a predicted size of 2016 bp and encoded a putative 238
671-aa protein, including a signal peptide, surface (SU) protein and transmembrane 239
(TM) protein. 240
Insertional polymorphic distribution of ERV-DC in cats 241
Multiple bands of ERV-DCs were detected in the healthy domestic cat genome by 242
Southern blot analysis with a specific env probe (Fig. 2A), indicating that ERV-DCs 243
on April 10, 2019 by guest
http://jvi.asm.org/
Dow
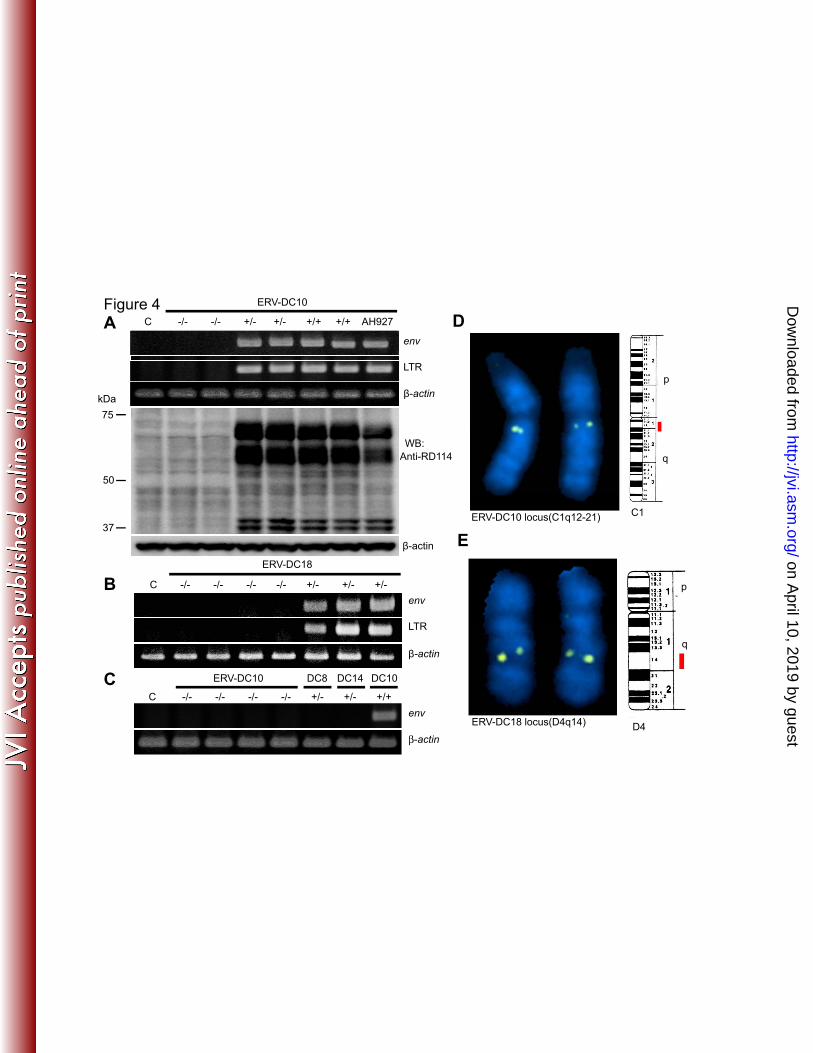
nloaded from

12
have infected cats and are vertically inherited. An insertional polymorphic distribution 244
of ERV-DC was determined among 244 cats in Japan using PCR primers based on the 245
genomic DNA flanking sequences unique to each ERV-DC, or with second primers 246
based on the proviral sequence (Fig. 2B). Randomly chosen cats from different genetic 247
backgrounds and geographic origins within Japan were screened for the presence of 10 248
full-length ERV-DC proviruses and three partial ERV-DC proviruses. All were 249
represented in the domestic cat genome at a frequency of 0.4–100% (Fig. 2C and Table 250
S1-1). We estimated that each cat had 7–17 copies of ERV-DC proviruses, with the 251
majority having 9 copies. The ERV-DC10 locus was heterozygous in 29.5% of the cats 252
examined and homozygous in 8.2%. The ERV-DC18 locus was detected only in one 253
cat (cat ON-T) and was heterozygous. When we examined ERV-DC provirus in siblings 254
of ON-T, the ERV-DC18 locus was heterozygous in three of four siblings which were 255
born to the same mother. Interestingly, these 3 cats also had ERV-DC10 and tested 256
positive for FeLV infection (Fig. 2D and Table S1-2). The sequences of ERV-DC18 are 257
almost identical to those of ERV-DC10; therefore, ERV-DC10 appears to have been 258
mobilized as an infectious virus in one of the parents of the three positive siblings, 259
generating provirus ERV-DC18. Although some ERV-DCs have become fixed in the cat 260
genome, others have not (Fig.2C). These results indicate that ERV-DC proviruses are 261
currently invading domestic cats. Sequence analysis verified that ERV-DC sequences 262
are integrated into the cat genome, and analysis of genomic sequences flanking the 263
proviruses, including identical target site duplication (TSD), identified unequivocally 264
the various ERV-DCs (Fig.1). Furthermore, 5′ and 3′ LTR sequences of each 265
ERV-DC were found to have high sequence identity or were identical with each other 266
(Table 1). There was no evidence of LTR recombination as determined by analyzing the 267
on April 10, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

13
4-bp identical TSD sequences (Fig. 1) and the phylognetic tree of LTR sequences 268
(Fig.5D). These results suggest that integration of ERV-DC occurred relatively 269
recently in the evolutionary history of cats. Supporting this idea is the absence of 270
ERV-DC proviruses and RD-114 in the Tsushima wild cat Prionailurus bengalensis 271
euptilurus (Fig. 2A and Fig. S1), which lives in close proximity to domestic cats. The 272
domestic cat is estimated to have separated from the leopard cat and the Tsushima wild 273
cat approximately 6.2–9.3 million years ago (MYA)(10,32). By comparing the 274
nucleotide sequences of the 5′- and 3′-LTR of ERV-DC7 (Table 1), which is fixed in the 275
cat population (Fig. 2C and Table S1-1), we estimated that the oldest time of ERV-DC 276
insertion was 2.8 MYA, assuming an average divergence rate of non-coding regions of 277
domestic cat genomes (21). Thus, we hypothesize that ERV-DC and RD-114 infected 278
domestic cats after separation from the leopard lineage. 279
280
Infectious viruses, ERV-DC10 and ERV-DC18 281
Since ERV-DC8, -DC10, -DC14 and -DC18 have intact gag/pol/env reading frames, we 282
carried out studies to determine if they were infectious. We transfected human 283
embryonic kidney 293T (HEK293T) cells with proviral plasmid DNA, and ~72 h later, 284
collected 0.22-μm-filtered cell-free supernatants that were used to infect HEK293T and 285
HeLa cells. ERV-DC10 and -DC18 were shown to be infectious by the presence of 286
proviral DNA and viral RNA (Fig. 3A,B), whereas ERV-DC8 and -DC14 were not. 287
Viral Gag and Env proteins were detected by immunoblotting (Fig. 3C). Surprisingly, 288
goat anti-FeLV gp70 detected ERV-DC Env as well as FeLV Env. Human cell lines 289
persistently infected with ERV-DC10 and -DC18 were established. Cell-free 290
on April 10, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

14
supernatants from HEK293T cells infected with ERV-DC10 and -DC18 could infect a 291
number of other human cancer cell lines, such as K562, Molt-4, HepG2, THP-1, dog 292
mammary tumour cells (KwDM) and monkey Vero cells (Fig. 3D and Fig. S2). The 293
viruses were also able to infect the feline kidney cell line CRFK, but not other feline 294
cell lines that can be infected with other feline retroviruses (e.g., AH927 fibroblasts that 295
are readily infected with FeLV; G355 astrocytes that can be infected with RD-114). 296
Transmission electron microscopy of HEK293T and HeLa cells persistently infected 297
with ERV-DC10 revealed 90–100-nm particles that were morphologically consistent 298
with a type C retrovirus (Fig. 3E). 299
We developed an alternative method to determine whether ERV-DC proviruses in 300
cat DNA are infectious. Full-length ERV-DC10 or ERV-DC18 proviral DNA from 301
primary cat PBMC was amplified by PCR with locus-specific primers and the PCR 302
products—without cloning of proviruses—were transfected into HEK293T cells. After 303
2-3 weeks, each filtered supernatant was used to infect HEK293T cells. As a result, 304
ERV-DC10 or -18 were shown to persistently produce infectious virus (Fig. 4A, B). 305
Even HEK293T cells transfected with DNA from the AH927 feline cell line, which has 306
a homozygous ERV-DC10 locus, produced infectious virus (Fig. 4A). Virus production 307
from DNA from cats that lacked ERV-DC10 or ERV-DC18 was never seen in cultures 308
(Fig. 4C), indicating that this method specifically assesses the activity of ERV-DC10 309
and -DC18 loci. Using this method, we also found that DNA from cats harbouring 310
ERV-DC8 and ERV-DC14 proviruses was unable to produce infectious virus (Fig. 4C), 311
consistent with data obtained using molecular clones of ERV-DC8 and -DC14. These 312
results demonstrate that ERV-DC10 and ERV-DC18 are potentially active and their 313
genomes encode infectious viruses. 314
on April 10, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

15
To define the active locus, we mapped two active loci by fluorescence in situ 315
hybridization (FISH) analysis: ERV-DC10 mapped to feline chromosome C1q12-21 and 316
ERV-DC18 to feline chromosome D4q14 (Fig. 4D,E). These results indicate that 317
ERV-DC10 and -DC18 loci are distinct from each other and we speculate that the 318
ERV-DC10 provirus on C1q12-21 mobilized to D4q14. We could not isolate ERV-DC10 319
viruses when PBMC from ERV-DC10–positive cats were cultured with HEK293T cells. 320
Thus, although infectious ERV-DC10 provirus is present in the genome of domestic cats, 321
PBMC and AH927 cells may not produce infectious viruses or ERV-DC10 may be 322
silenced in these cells. 323
Phylogenetic analyses of ERV-DCs 324
Phylogenetic analyses indicated that ERV-DC proviruses were genetically distinct from 325
exFeLVs, enFeLVs and RD-114, and comprised a separate group of gammaretroviruses 326
(Fig. 5A, B). ERV-DC proviruses obtained in this study were classified into three 327
groups based on their env genes (Fig. 5C). This classification was supported by the 328
phylogenetic tree of their LTR sequences (Fig. 5D), and suggests that independent 329
invasion of ERV-DCs may have occurred in the ancestry of domestic cats. 330
To examine if ERV-DCs interact with DNA of other species, we extracted 331
ERV-DC-like ERVs from the genome data of 12 different species (Table S2). As a result, 332
22, 19, 3, and 2 ERV loci were detected in the genomes of the house mouse, brown rat, 333
common chimpanzee and rhesus macaque, respectively (Table S3), although no 334
orthologous relationships were found among them. We then conducted phylogenetic 335
analyses of ERV-DCs using these ERV elements with various endogenous or exogenous 336
viruses (Fig. 5A, B). The Pol protein of ERV-DC was most closely related to that of 337
on April 10, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

16
RD-114 and BaEV (Fig. 5A), whereas the Env/TM protein of ERV-DC was more 338
closely related to that of the FeLV family (Fig. 5B). RD-114 is genetically related to 339
BaEV and both of these viruses are recombinant viruses that contain a type C 340
(gammaretrovirus group) ORF for gag-pol and a type D (betaretrovirus group) env gene, 341
and supposedly arose from cross-species transmission (1,15,36). The amino acid 342
identity was 98.6% for Pol proteins of ERV-DC18 and RD-114 SC3C, and 81.1% for 343
the TM regions of Env proteins of ERV-DC18 and FeLV-A clone 61E (55.8% for the 344
entire protein). In addition, the ERV sequences in the house mouse, brown rat, common 345
chimpanzee and rhesus macaque genomes were also found to be similar to ERV-DCs 346
(Fig. 5A, B). Pol from ERV-DC10/DC18 is most highly related to the Pol of 347
endogenous primate viruses, including chimpanzee, rhesus macaque, and baboon. In 348
contrast, Env/TM from ERV-DC10/18 is closely related to FeLV and rodent retroviruses. 349
These results indicate that ERV-DCs should be classified as a strain of 350
gammaretroviruses, and that ERV-DCs or ERV-DC-related viruses have passed through 351
several species. The lack of ERV-DC-like elements in the genomes of species other than 352
primates and rodents suggests that ERV-DCs have spread through interspecies 353
transmission among rodents, primates and cats. The interspecies transmission of 354
ERV-DCs may make it possible to generate recombinant viruses, and ERV-DCs could 355
be donor and/or acceptor viruses in domestic cats. 356
357
Evidence for transduction of ERV-DC env into FeLV 358
Recombination events occur in retroviruses by template switching during viral reverse 359
transcription (7). We discovered from the spleen of an FeLV-positive cat (ON-T; Fig. 360
on April 10, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

17
6A) a recombinant retrovirus, designated FeLV-D (strain FeLV-D/ON-T), containing the 361
env gene of ERV-DC and LTR, gag and pol sequences from FeLV. Sequence analysis 362
demonstrated that the env gene of FeLV-D was derived from ERV-DCs (Fig. S3A) and 363
the LTR of FeLV-D was derived from FeLV (Fig. S3B). The gag–pol gene contained an 364
ORF but also some deletions (Fig. 6A), suggesting that the virus is replication defective. 365
To determine if FeLV-D is expressed in the cat, spleen RNA from the cat ON-T was 366
amplified by RT-PCR using FeLV-D–specific primer pairs corresponding to FeLV 367
5′LTR and ERV-DC env sequences. Major bands corresponding to the genomic RNA 368
and spliced env mRNA were detected (Fig. S4) and confirmed by sequencing. To 369
determine if FeLV-D could be isolated from cells cultured from the spleen of cat ON-T, 370
we homogenized the spleen, filtered the sample using a 0.22-μm filter and used the 371
homogenate to inoculate HEK293T cells. FeLV-D, as well as prototype FeLV, was 372
detected at 4 weeks post-inoculation by PCR (Fig. 6B). The FeLV-D/ON-T strain was 373
successfully isolated in association with prototype FeLV, and FeLV-D was packaged as 374
a retrovirus. The supernatant from HEK293T cells infected with FeLV-D/ON-T for 6 375
weeks was used to inoculate fresh HEK293T and HeLa cells, and the virus was 376
successfully transmitted between the cells in culture. We re-cloned FeLV-D/ON-T 377
(designated FeLV-D/re-T) from the chromosomal DNA of HEK293T cells persistently 378
infected with FeLV-D and verified that the sequences of FeLV-D/re-T and 379
FeLV-D/ON-T are identical (Fig. S3A). When the DNA from peripheral blood or 380
tumour tissues from 283 other FeLV-positive cats was screened for the presence of 381
FeLV-D, three positive samples were detected (Fig. 6C). One cat (ON-C) was a sibling 382
of ON-T. FeLV-D was molecularly cloned from the three other FeLV-D–positive cats 383
(FeLV-D/ON-C, FeLV-D/Ty26 and FeLV-D/44B) in the region of env and 3′LTR (Fig. 384
on April 10, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

18
S3A,B,C). A schematic representation of the structures of the env genes of the various 385
FeLV-Ds is shown in Fig. 6D. Thus, FeLV-D appears to have been generated by 386
recombination between FeLV and ERV-DC, indicating that ERV-DC can serve as a 387
donor and/or acceptor in the generation of recombinant viruses. The recombination 388
junction between ERV-DC and exFeLV varied among FeLV-D strains, suggesting that 389
FeLV-D was generated de novo in each cat. It is not clear whether FeLV-D is an 390
infectious virus by itself or whether it is pathogenic. To determine if the FeLV-D env 391
gene plays a role in infection, FeLV-D env-expressing pseudoviruses were generated 392
that could transmit β-galactosidase (Fig. 7). The FeLV-D env-expressing pseudotype 393
viruses from FeLV-D/ON-T and FeLV-D/Ty26 could infect cells already infected with 394
FeLV-A, B or C, or cells persistently infected with ERV-DC10. Therefore, our data 395
show that FeLV-D belongs to a different interference group than ERV-DC10. These 396
results indicate that the receptor usage for both FeLV-D and ERV-DC10 to enter cells is 397
distinct from FeLV A, B, and C. 398
Discussion 399
This study characterizes 13 endogenous retroviral proviruses in the cat belonging 400
to a class of ERVs, ERV-DC, that has previously been only partially characterized. We 401
show that two of these ERV-DCs, ERV-DC10 and ERV-DC18, are novel feline 402
infectious gammaretroviruses that are inherited on chromosome C1q12-21 and D4q14, 403
respectively. Thus, our study is the first to demonstrate the existence of endogenous, 404
infectious retroviruses in the genome of the domestic cat. Approximately 38% of the 405
cats examined in Japan have heterozygous or homozygous ERV-DC10 loci. Unlike 406
ERV-DC7, which is fixed in the cat genome but contains deleterious mutations that 407
on April 10, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

19
make it non-infectious, ERV-DC10 has not yet been fixed in the cat genome. It is not 408
known whether ERV-DC10 will eventually disappear or become fixed in cats. Although 409
the open reading frames of ERV-DC8 and ERV-DC14 are intact, they are unable to 410
make infectious viral particles, most likely due to point mutations. It is unclear 411
whether ERV-DCs could be harmful or beneficial to the host. However, previous studies 412
have indicated that the presence of other ERVs in vertebrates is associated with 413
protection against infection of related exogenous retroviruses as well as with the 414
generation of pathogenic viruses or tumors (9,11,26). 415
Our studies further indicate that ERV-DC10 was not able to produce infectious 416
particles when it was present in some cell types, including feline PBMCs (ERV-DC10 417
+/+ genotype) and the feline fibroblast cell line AH927 (ERV-DC10 +/+), suggesting 418
that the ERV-DC10 promoter may be modified in a tissue-specific manner, similar to the 419
endogenous human ERV syncytin (24), as expression of retroviral genes can be 420
controlled by transcriptional modification (43). Moreover, an infectious provirus not 421
being expressed may be related to its methylation status in the cell, a common 422
mechanism by which endogenous retroviruses are silenced (22). Further investigation 423
will be necessary to understand the gene sequences in ERV-DCs controlling expression 424
and viral production. 425
Endogenous retroviral sequences have been known to exist in the domestic cat, 426
but until now they have not been identified as encoding infectious viruses. Although 427
RD-114, which was generated by xenotransplantation, is described as an infectious 428
feline ERV (23), an infectious RD-114 provirus has never been detected in the genome 429
of domestic cats. Recent work shows that novel, infectious mouse retroviruses can be 430
on April 10, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

20
generated through in vivo passage of human cells in rodents (30). It is, therefore, 431
possible to generate an infectious ERV through in vivo passage in other species. 432
Our phylogenetic analyses revealed that the pol gene of ERV-DCs has high 433
similarity to those of BaEV, RD-114 and primate ERVs, whereas the env/TM gene of 434
ERV-DCs is related to those of FeLV, MuLV and rodent ERVs. The fact that infectious 435
ERV-DCs are present in domestic cats and ERV-DC-like elements are detected in cats, 436
rodents and primates suggests that ERV-DCs may have been transmitted to primates or 437
rodents, or vice versa, becoming part of the DNA of the infected species. 438
FeLV-D was detected and characterized in this study as a recombinant virus 439
generated by transduction of the ERV-DC env gene into FeLV. This discovery supports 440
the idea that ERV-DCs may act as donor and/or acceptor viruses to generate new 441
recombinant viruses. We showed that FeLV-D belongs to a different interference group 442
from that of ERV-DC10, consistent with the fact that ERV-DC10 belongs to ERV-DC 443
group III, whereas the env genes in all 4 FeLV-D strains are similar to ERV-DC group I. 444
ERVs belonging to groups I and III may use distinct cell surface receptors and it will be 445
important to characterize these receptors and assess infectivity of the ERV-DC 446
recombinant viruses. 447
ERV-DC is present in the cat genome with different copy numbers and insertional 448
polymorphisms. Reinfection or infection with ERV-DCs may vary among individual 449
cats based on whether the host genes permit or restrict infection. ERV-DCs are present 450
in domestic cats, but not in the Japanese Tsushima leopard cat, Prionailurus bengalensis 451
euptilurus. Based on estimation of viral integration time and ERV-DC LTR sequences, 452
ERV-DCs infected domestic cats within the past 2.8 million years, after separation from 453
on April 10, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

21
the leopard lineage. It will be interesting to determine whether ERV-DC proviruses have 454
played a role in the evolution of cats or in the generation of polymorphisms in the cat 455
genome. 456
In conclusion, our findings provide evidence for the existence of infectious 457
ERV-DCs in the domestic cat. Because these ERV-DCs are currently being transmitted 458
among cats, and perhaps other species, as infectious ERVs or as recombinant viruses 459
containing ERV-DC sequences, future studies should follow their evolution and assess 460
the consequences of their presence for the health of domestic cats and other species. 461
462
463
464
465
466
467
468
469
470
471
472
on April 10, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

22
473
Data deposition The sequences reported here have been deposited in the 474
DDBJ/EMBL/GenBank and the accession numbers are AB673426-AB673432, 475
AB674572-AB674577 and AB674439-AB674452. 476
Acknowledgements We thank Drs. T. Kitamura and M. Nakaya for providing 477
packaging cells and expression plasmids. We also thank Drs. N. Oishi (Oishi Veterinary 478
Hospital) and H. Yamamoto (Tsushima Wildlife Conservation Center) for providing 479
blood and tissue samples. We are grateful to Sandra Ruscetti (National Cancer Institute, 480
Frederick) and Kenji Baba (Yamaguchi University) for helpful discussions with the 481
manuscript. This study was supported by grants from the Ministry of Education, 482
Culture, Sports, Science and Technology of Japan (K.N.). Y.A.,H.O. and S.W. 483
contributed equally to this work. 484
485
486
on April 10, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

23
References 487
1. Benveniste, R. E., and G. J. Todaro. 1974. Evolution of C-type viral genes: 488
inheritance of exogenously acquired viral genes. Nature 252:456-459. 489
2. Benveniste, R. E., C. J. Sherr, and G. J. Todaro. 1975. Evolution of type C viral 490
genes: origin of feline leukemia virus. Science 190:886-888. 491
3. Beyer, W., R. Mohring, B. Drescher, U. Notzel, and S. Rosenthal. 1987. Molecular 492
cloning of an endogenous cat retroviral element (ECE 1)--a recombinant between 493
RD-114 and FeLV-related sequences. Brief report. Arch. Virol. 96:297-301. 494
4. Boeke, J. D., and J. P. Stoye. 1997. Retrotransposons, endogenous retroviruses, and 495
the evolution of retroelements, p. 343-436. In J. M. Coffin, S. H. Hughes, and H. E. 496
Varmus (ed.), Retroviruses. Cold Spring Harbor Laboratory Press, Cold Spring 497
Harbor, N.Y. 498
5. Bonner, T. I., and G. J. Todaro. 1979. Carnivores have sequences in their cellular 499
DNA distantly related to the primate endogenous virus, MAC-1. Virology 500
94:224-227. 501
6. Darriba, D., G. L. Taboada, R. Doallo, and D. Posada. 2011. ProtTest 3: fast 502
selection of best-fit models of protein evolution. Bioinformatics 27:1164-1165. 503
7. Delviks-Frankenberry, K., A. Galli, O. Nikolaitchik, H. Mens, V. K. Pathak, and W. 504
S. Hu. 2011. Mechanisms and factors that influence high frequency retroviral 505
recombination. Viruses 3:1650-1680. 506
8. Dimmic, M. W., J. S. Rest, D. P. Mindell, and R. A. Goldstein. 2002. rtREV: an 507
amino acid substitution matrix for inference of retrovirus and reverse transcriptase 508
phylogeny. J. Mol. Evol. 55:65-73. 509
9. Dudley, J. P., J. A. Mertz, S. Bhadra, M. Palmarini, and C. A. Kozak. 2011. 510
Endogenous Retroviruses and Cancer, p.119-162. In J.Dudley (ed.), Retroviruses 511
and Insights into Cancer. Springer Press. 512
10. Johnson, W. E., E. Eizirik, J. Pecon-Slattery, W. J. Murphy, A. Antunes, E. Teeling, 513
and S. J. O'Brien. 2006. The late Miocene radiation of modern Felidae: a genetic 514
assessment. Science 311:73-77. 515
on April 10, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

24
11. Okeoma, C. M., and S. R. Ross. 2011. Genetics of Host Resistance to Retroviruses 516
and Cancer, p. 95-118. In J.Dudley (ed.), Retroviruses and Insights into Cancer. 517
Springer Press. 518
12. Felsenstein, J. 1981. Evolutionary trees from DNA sequences: a maximum 519
likelihood approach. J. Mol. Evol. 17:368-376. 520
13. Hardy, W. D., Jr. 1993. Feline oncoretroviruses, p. 109–180. In J. A. Levy (ed.), The 521
Retroviridae, vol. 2. Plenum Press, New York, N.Y. 522
14. Jarrett, W. F., E. M. Crawford, W. B. Martin, and F. Davie. 1964. A Virus-Like 523
Particle Associated with Leukemia (Lymphosarcoma). Nature 202:567-569. 524
15. Kato, S., K. Matsuo, N. Nishimura, N.Takahashi, and T.Takano. (1987). The entire 525
sequence of baboon endogenous virus DNA: A chimeric structure of murine type C 526
and simian type D retroviruses. Jpn. J. Genet. 62:127–137. 527
16. Katoh, K., K. Kuma, H. Toh, and T. Miyata. 2005. MAFFT version 5: improvement 528
in accuracy of multiple sequence alignment. Nucleic Acids Res. 33:511-518. 529
17. Kimura, M. 1980. A simple method for estimating evolutionary rates of base 530
substitutions through comparative studies of nucleotide sequences. J.Mol.Evol. 531
16:111-120. 532
18. Kitamura, T., Y. Koshino, F. Shibata, T. Oki, H. Nakajima, T. Nosaka, and H. 533
Kumagai. 2003. Retrovirus-mediated gene transfer and expression cloning: 534
powerful tools in functional genomics. Exp. Hematol 31:1007-1014. 535
19. Lander, E. S., L. M. Linton, B. Birren, C. Nusbaum, M. C. Zody, J. Baldwin, K. 536
Devon, K. Dewar, M. Doyle, W. FitzHugh, R. Funke, D. Gage, K. Harris, A. Heaford, 537
J. Howland, L. Kann, J. Lehoczky, R. LeVine, P. McEwan, K. McKernan, J. 538
Meldrim, J. P. Mesirov, C. Miranda, W. Morris, J. Naylor, C. Raymond, M. Rosetti, 539
R. Santos, A. Sheridan, C. Sougnez, N. Stange-Thomann, N. Stojanovic, A. 540
Subramanian, D. Wyman, J. Rogers, J. Sulston, R. Ainscough, S. Beck, D. Bentley, 541
J. Burton, C. Clee, N. Carter, A. Coulson, R. Deadman, P. Deloukas, A. Dunham, I. 542
Dunham, R. Durbin, L. French, D. Grafham, S. Gregory, T. Hubbard, S. Humphray, 543
A. Hunt, M. Jones, C. Lloyd, A. McMurray, L. Matthews, S. Mercer, S. Milne, J. C. 544
Mullikin, A. Mungall, R. Plumb, M. Ross, R. Shownkeen, S. Sims, R. H. Waterston, 545
R. K. Wilson, L. W. Hillier, J. D. McPherson, M. A. Marra, E. R. Mardis, L. A. 546
on April 10, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

25
Fulton, A. T. Chinwalla, K. H. Pepin, W. R. Gish, S. L. Chissoe, M. C. Wendl, K. D. 547
Delehaunty, T. L. Miner, A. Delehaunty, J. B. Kramer, L. L. Cook, R. S. Fulton, D. 548
L. Johnson, P. J. Minx, S. W. Clifton, T. Hawkins, E. Branscomb, P. Predki, P. 549
Richardson, S. Wenning, T. Slezak, N. Doggett, J. F. Cheng, A. Olsen, S. Lucas, C. 550
Elkin, E. Uberbacher, M. Frazier, et al. 2001. Initial sequencing and analysis of the 551
human genome. Nature 409:860-921. 552
20. Le, S. Q., and O. Gascuel. 2008. An improved general amino acid replacement 553
matrix. Mol.Biol.Evol. 25:1307-1320. 554
21. Lopez, J. V., M. Culver, J. C. Stephens, W. E. Johnson, and S. J. O'Brien. 1997. 555
Rates of nuclear and cytoplasmic mitochondrial DNA sequence divergence in 556
mammals. Mol. Biol. Evol. 14:277-286. 557
22. Maksakova, I. A., D. L. Mager, and D. Reiss. 2008. Keeping active endogenous 558
retroviral-like elements in check: the epigenetic perspective. Cell. Mol. Life Sci. 559
65:3329-3347. 65:3329-3347. 560
23. McAllister, R. M., M. Nicolson, M. B. Gardner, R. W. Rongey, S. Rasheed, P. S. 561
Sarma, R. J. Huebner, M. Hatanaka, S. Oroszlan, R. V. Gilden, A. Kabigting, and L. 562
Vernon. 1972. C-type virus released from cultured human rhabdomyosarcoma cells. 563
Nat. New biol. 235:3-6. 564
24. Mi, S., X. Lee, X. Li, G. M. Veldman, H. Finnerty, L. Racie, E. LaVallie, X. Y. Tang, 565
P. Edouard, S. Howes, J. C. Keith, Jr., and J. M. McCoy. 2000. Syncytin is a captive 566
retroviral envelope protein involved in human placental morphogenesis. Nature 567
403:785-789. 568
25. Mullins, J. I., J. W. Casey, M. O. Nicolson, K. B. Burck, and N. Davidson. 1981. 569
Sequence arrangement and biological activity of cloned feline leukemia virus 570
proviruses from a virus-productive human cell line. J.Virol. 38:688-703. 571
26. Mura, M., P. Murcia, M. Caporale, T. E. Spencer, K. Nagashima, A. Rein, and M. 572
Palmarini. 2004. Late viral interference induced by transdominant Gag of an 573
endogenous retrovirus. Proc. Natl. Acad. Sci. USA 101:11117-11122. 574
27. Nakamura, Y., A. Ura, M. Hirata, M. Sakuma, Y. Sakata, K. Nishigaki, H. 575
Tsujimoto, A. Setoguchi, and Y. Endo. 2010. An updated nation-wide 576
epidemiological survey of feline immunodeficiency virus (FIV) infection in Japan. J. 577
on April 10, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

26
Vet. Med. Sci. 72:1051-1056. 578
28. Nishigaki, K., C. Hanson, D. Thompson, T. Yugawa, M. Hisasue, H. Tsujimoto, and 579
S. Ruscetti. 2002. Analysis of the disease potential of a recombinant retrovirus 580
containing Friend murine leukemia virus sequences and a unique long terminal 581
repeat from feline leukemia virus. J.Virol. 76:1527-1532. 582
29. Overbaugh, J., N. Riedel, E. A. Hoover, and J. I. Mullins. 1988. Transduction of 583
endogenous envelope genes by feline leukaemia virus in vitro. Nature 332:731-734. 584
30. Paprotka, T., K. A. Delviks-Frankenberry, O. Cingoz, A. Martinez, H. J. Kung, C. G. 585
Tepper, W. S. Hu, M. J. Fivash, Jr., J. M. Coffin, and V. K. Pathak. 2011. 586
Recombinant origin of the retrovirus XMRV. Science 333:97-101. 587
31. Polani, S., A. L. Roca, B. B. Rosensteel, S. O. Kolokotronis, and G. K. Bar-Gal. 2010. 588
Evolutionary dynamics of endogenous feline leukemia virus proliferation among 589
species of the domestic cat lineage. Virology 405:397-407. 590
32. Rannala, B., and Z. Yang. 2007. Inferring speciation times under an episodic 591
molecular clock. Syst. Biol. 56:453-466. 592
33. Riedel, N., E. A. Hoover, P. W. Gasper, M. O. Nicolson, and J. I. Mullins. 1986. 593
Molecular analysis and pathogenesis of the feline aplastic anemia retrovirus, feline 594
leukemia virus C-Sarma. J.Virol. 60:242-250. 595
34. Sambrook, J., and D. W. Russell. 2001. Molecular cloning: a laboratory manual, 3rd 596
ed. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y. 597
35. Sperber, G. O., T. Airola, P. Jern, and J. Blomberg. 2007. Automated recognition of 598
retroviral sequences in genomic data--RetroTector. Nucleic Acids Res. 599
35:4964-4976. 600
36. Spodick, D. A., A. K. Ghosh, S. Parimoo, and P. Roy-Burman. 1988. The long 601
terminal repeat of feline endogenous RD-114 retroviral DNAs: analysis of 602
transcription regulatory activity and nucleotide sequence. Virus Res. 9:263-283. 603
37. Stamatakis, A. 2006. RAxML-VI-HPC: maximum likelihood-based phylogenetic 604
analyses with thousands of taxa and mixed models. Bioinformatics 22:2688-2690. 605
38. Stewart, M. A., M. Warnock, A. Wheeler, N. Wilkie, J. I. Mullins, D. E. Onions, and 606
on April 10, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

27
J. C. Neil. 1986. Nucleotide sequences of a feline leukemia virus subgroup A 607
envelope gene and long terminal repeat and evidence for the recombinational origin 608
of subgroup B viruses. J.Virol. 58:825-834. 609
39. Todaro, G. J., R. E. Benveniste, S. A. Sherwin, and C. J. Sherr. 1978. MAC-1, a new 610
genetically transmitted type C virus of primates: "low frequency" activation from 611
stumptail monkey cell cultures. Cell 13:775-782. 612
40. Umehara, D., S. Watanabe, H. Ochi, Y. Anai, N. Ahmed, M. Kannagi, C. Hanson, S. 613
Ruscetti, and K. Nishigaki. 2010. Role of phosphatidylinositol 3-kinase in friend 614
spleen focus-forming virus-induced erythroid disease. J.Virol. 84:7675-7682. 615
41. van der Kuyl, A. C., J. T. Dekker, and J. Goudsmit. 1999. Discovery of a new 616
endogenous type C retrovirus (FcEV) in cats: evidence for RD-114 being an 617
FcEV(Gag-Pol)/baboon endogenous virus BaEV(Env) recombinant. J.Virol. 618
73:7994-8002. 619
42. Waterston, R. H., K. Lindblad-Toh, E. Birney, J. Rogers, J. F. Abril, P. Agarwal, R. 620
Agarwala, R. Ainscough, M. Alexandersson, P. An, S. E. Antonarakis, J. Attwood, R. 621
Baertsch, J. Bailey, K. Barlow, S. Beck, E. Berry, B. Birren, T. Bloom, P. Bork, M. 622
Botcherby, N. Bray, M. R. Brent, D. G. Brown, S. D. Brown, C. Bult, J. Burton, J. 623
Butler, R. D. Campbell, P. Carninci, S. Cawley, F. Chiaromonte, A. T. Chinwalla, D. 624
M. Church, M. Clamp, C. Clee, F. S. Collins, L. L. Cook, R. R. Copley, A. Coulson, O. 625
Couronne, J. Cuff, V. Curwen, T. Cutts, M. Daly, R. David, J. Davies, K. D. 626
Delehaunty, J. Deri, E. T. Dermitzakis, C. Dewey, N. J. Dickens, M. Diekhans, S. 627
Dodge, I. Dubchak, D. M. Dunn, S. R. Eddy, L. Elnitski, R. D. Emes, P. Eswara, E. 628
Eyras, A. Felsenfeld, G. A. Fewell, P. Flicek, K. Foley, W. N. Frankel, L. A. Fulton, 629
R. S. Fulton, T. S. Furey, D. Gage, R. A. Gibbs, G. Glusman, S. Gnerre, N. Goldman, 630
L. Goodstadt, D. Grafham, T. A. Graves, E. D. Green, S. Gregory, R. Guigo, M. 631
Guyer, R. C. Hardison, D. Haussler, Y. Hayashizaki, L. W. Hillier, A. Hinrichs, W. 632
Hlavina, T. Holzer, F. Hsu, A. Hua, T. Hubbard, A. Hunt, I. Jackson, D. B. Jaffe, L. 633
S. Johnson, M. Jones, T. A. Jones, A. Joy, M. Kamal, E. K. Karlsson, et al. 2002. 634
Initial sequencing and comparative analysis of the mouse genome. Nature 635
420:520-562. 636
43. Wolf, D., and S. P. Goff. 2008. Host restriction factors blocking retroviral replication. 637
Annu. Rev. Genet. 42:143-163. 638
on April 10, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

28
44. Yoshinaka, Y., I. Katoh, T. D. Copeland, and S. Oroszlan. 1985. Murine leukemia 639
virus protease is encoded by the gag-pol gene and is synthesized through 640
suppression of an amber termination codon. Proc. Natl. Acad. Sci. USA 641
82:1618-1622. 642
643
644
645
646
on April 10, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

29
Figure legends 647
Fig. 1. ERV-DC structures. Structures of the genomes of 10 full length ERV-DC 648
proviruses and three partial ERV-DC proviruses. The gag, pol and env genes are 649
illustrated together with the 5′ and 3′ LTRs and positions of the gag and env 650
translational initiation codons (ATG). Asterisks indicate conserved stop codons. Gag 651
and Pol proteins may be synthesized as a large single polypeptide precursor via 652
termination suppression (44). Open triangle indicates a deletion of nucleotides and filled 653
triangle indicates an insertion of nucleotides. Flanking 4-bp TSD sequences are shown 654
for each provirus. 655
Fig. 2. Detection of ERV-DC proviruses in the domestic cat genome. (A) Southern 656
blotting of ERV-DC with an env probe derived from FeLV-D/ON-T in chromosomal 657
DNA from the domestic cat (F.s. catus) and P.b. euptilurus. (B) PCR for detection of 658
pre-integration sites (PISs) and proviruses. Red boxes: viral LTRs; yellow boxes: target 659
site duplications. Primers A–D are shown. The AC primer pair generated a PIS and/or 660
full-length provirus. (C) Insertional polymorphisms of 13 ERV-DCs in 244 domestic 661
cats. Green: provirus was detected on heterozygous loci; Red: provirus was detected on 662
homozygous loci; Yellow: provirus was detected (D) ERV-DC18 was only present in 3 663
of 4 siblings from one family. Each PIS, full-length ERV-DC10 or ERV-DC18 was 664
detected by primers Fe-122S and Fe-38R for ERV-DC10 or Fe-76S and Fe-81R for 665
ERV-DC18 (Table S4). 666
Fig. 3. ERV-DC10 and ERV-DC18 are infectious viruses. (A) env and pol were detected 667
by PCR at 2 and 8 days after infection of HEK293T cells. The supernatants from 8-day 668
cultures were used to infect HeLa cells. M: mock infection; 10: infected with 669
on April 10, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

30
ERV-DC10; 18: infected with ERV-DC18. (B) Viruses collected from supernatants were 670
used to detect env and pol genes by RT-PCR. (C) ERV-DC10 and ERV-DC18 proteins 671
were detected in HEK293T cells using goat antiserum against disrupted RD-114 virions 672
and anti-FeLV gp70 antibody. Anti-RD114 antibodies recognize many ERV-DC proteins, 673
indicated by arrows, but not ERV-DC Env. (D) Susceptibility of cells to infection by 674
ERV-DC10/ERV-DC18. (E) Transmission electron microscopy of retroviral particles in 675
HEK293T cells persistently infected with ERV-DC10. The bar under each panel 676
represents a 100-nm scale. 677
Fig. 4. Active loci of ERV-DC10 and ERV-DC18. (A) ERV-DC10; (B) ERV-DC18 or 678
(C) ERV-DC10, -DC8 or -DC14 DNA amplified by PCR with locus-specific primers 679
using feline chromosomal DNA was directly transfected to HEK293T cells and the 680
filtrated supernatant was used to infect fresh HEK293T cells. Viral infection was 681
monitored by detecting proviral DNAs or viral proteins. Each proviral genotype is 682
indicated as -/- (wild), +/- (heterozygous) and +/+ (homozygous). Independent DNA 683
samples were used in the figures. AH927 is a feline cell line with ERV-DC10 +/+. (D 684
and E) FISH analysis of ERV-DC10 and ERV-DC18 loci on feline chromosomes. 685
Fig. 5. Maximum-likelihood phylogenetic trees of Pol and Env TM proteins. We used 686
amino acid sequences of entire regions of (A) Pol proteins and (B) TM subunits of Env 687
proteins for 46 or 56 endogenous/exogenous viruses (Table S5). A monophyletic subtree 688
of the ERV elements obtained from the same species was compressed into a single 689
branch as indicated by arrows. Viral origin is indicated as blue (feline), orange (primate), 690
green (rodent), gray (avian) or black (other species). The percent values were 691
determined from 1000 repeats of fast bootstrapping using RAxML (37) and are 692
on April 10, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

31
indicated at the branch junctions. More than 70 bootstrap values are shown. 693
Maximum-likelihood phylogenic trees of ERV-DC env and LTR are indicated in Fig.5C 694
and Fig.5D. Nucleotide sequences of (C) entire ERV-DC env regions or (D) 5′ and 695
3′ LTRs in ERV-DCs obtained were used. The general time-reversible model with rate 696
heterogeneity among sites (GTR+Γ) was utilized. The percent values were determined 697
from 1000 repeats of fast bootstrapping using RAxML and are indicated at the branch 698
junctions. 3 or 5 indicates 3’ LTR or 5’ LTR. ECE1 (Accession No. X51929), Fc21 699
(AF155060), Fc41 (AF155061), SC3C (EU030001), CRT (AB559882) 700
701
Fig. 6. Transduction of ERV-DC env into FeLV. (A) Schematic representation of the 702
structure of FeLV-D provirus integrated in spleen DNA of cat ON-T and FeLV-A/61E. 703
White indicates FeLV sequences and gray indicates ERV-DC sequences. (B) Isolation of 704
FeLV-D in HEK293T cells inoculated with spleen extracts from cat ON-T. FeLV-D- and 705
FeLV env-specific PCR was carried out. (C) A total of 283 DNA samples were screened 706
for the presence of FeLV-D and four positive samples were detected. (D) Structure of 707
FeLV-D env. Recombination pattern of each FeLV-D env is indicated. White indicates 708
FeLV sequences and gray indicates ERV-DC sequences. 709
Fig. 7. Infection assay of LacZ pseudotype virus. FeLV-A/Glasgow-1 env, 710
FeLV-B/Gardner-Arnstein env, FeLV-C/Sarma env, FeLV-D/ON-T env, FeLV-D/Ty26 711
env and ERV-DC10 env were used for each pseudotype virus preparation. HEK293T 712
cells infected with FeLV-A/clone33, FeLV-B/Gardner-Arnstein, FeLV-C/Sarma, 713
ERV-DC10 or both FeLV-A/clone33 and FeLV-D/ON-T were used as target cells for an 714
interference assay. X-gal-positive cells were counted as infectious units (I.U.). Arrow 715
on April 10, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

32
indicates viral interference. 716
717
718
719
720
on April 10, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from
ERV-DC1 ERV-DC2 ERV-DC3 ERV-DC4 ERV-DC7 ERV-DC8 ERV-DC10 ERV-DC14 ERV-DC17 ERV-DC18 ERV-DC19
0 3 1 0 42 1 0 1 4 0 3
100 99.5 99.8 100 96.7 99.8 100 99.8 99.5 100 99.5
Provirus Number of
Differences between 5’ and 3’ LTRs
Homology between 5’ and 3’ LTRs (%)
Length of LTR (bp) 3’LTR 5’LTR
550 555 550 550 545 550 551 551 551 551 555
550 555 550 550 566 550 551 551 550 551 555
Table 1
Properties of ERV-DC 5′ and 3′ LTRs are summarized.
on April 10, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

Figure 1
ERV-DC1
*5’LTR
ATG * *ATG
3’LTRgag Δpol Δenvgag - Δpol Δenv
GTGG GTGG
250bp deletion1bp deletione
5’LTRATG *ATG
3’LTRenvenv
gag( ? )ERV-DC2
*ATG**3’LTR5’LTR
ATG
Δgag pol ΔenvΔ gagERV-DC3
GGAC GGAC
TAAC TAAC
1bp insertion71bp deletion
Δ gag Δenv
* *ATG*3’LTR
1bp insertion
5’LTRATG
gag pol Δenvgag - pol Δenv
ERV-DC4
*3’LTR
ATG
envERV DC6
TGAC TGAC
GGAG
* *ATG*3’LTR
1bp deletion
1bp insertion
5’LTRATG
Δgag Δpol ΔenvΔgag Δenv
GTG
1bp insertionERV-DC7
* *ATG*3’LTR5’LTR
ATG
gag pol env
envERV-DC6
ACAA ACAA
GGAG
3 LTR5 LTR gag pol envgag - pol env
ERV-DC8
* *ATG*3’LTR5’LTR
ATG
gag pol envgag - pol env
ERV-DC10
* *ATG*ATG
ATAC ATAC
TTGG TTGG
3’LTR5’LTR gag pol envgag - pol env
ERV-DC14
Δenv
*3’LTR
ATG
ΔenvERV-DC16
TATT TATT
AACC
* *ATG*3’LTR5’LTR
ATG
gag pol envgag - pol env
ERV-DC18
3’LTR*
1bp insertion
5’LTRATG * *ATG
gag Δpol envgag - Δpol env
ERV-DC17 AAAC AAAC
CATA CATA
3’LTR
2.4kbp deletion
5’LTRATG * *ATG
Δgag envΔgag - Δpol env
ΔpolERV-DC19 ATTG ATTG
1kb
on April 10, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

Figure 2
AC
kbp100
23
9.4
6 6
+/-+/++30
405060708090100
ts w
ith p
rovi
rus
6.6
2 3
+
0102030
1 2 3 4 6 7 8 10 14 16 17 18 19
% o
f cat
ERV-DC locus no.2.3
BamHIEcoRI
B D4bp
APIS
siblings
5’LTR 3’LTRgag pol env
IntegrationC
ERV-DC provirus B
PIS(ERV-DC18)
ERV-DC18
ERV-DC10CDC
p
A
C
PIS(ERV-DC10)
c-myc
on April 10, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

ACFigure 3 kDa
100
M 10 18
M 10 18 M 10 18 M 10 18HEK293T (2days) HEK293T (8days) HeLa (7days)
A
50
Anti-RD114 virus
M 10 18 M 10 18 M 10 18
env
pol
F LV
293T
/FeL
V-
BAnti-FeLV gp70β-actin
FeLV
HEK293T (2days) HEK293T (8days) HeLa (10days)B
Cell lines (species) ERV-DC10/18
Molt-4 (Human) +
K562 (Human) +
DAnti-β-actin
( y ) ( y ) ( y )
env
RT + + + - - - + + + - - - + + + - - -M 10 18 M 10 18 M 10 18 M 10 18 M 10 18 M 10 18
E
K562 (Human) +
HepG2 (Human) +
HeLa (Human) +
HEK293T (Human) +
MCF7 (Human) -
THP 1 (Human) +
pol
THP-1 (Human) +
CRFK (Cat) +
AH927 (Cat) -
G355 (Cat) -
Vero (Monkey) +
C 7 (M k )Cos7 (Monkey) -
NIH3T3 (Mouse) -
L929 (Mouse) -
BHK21 (Hamster) -
KwDM (Dog) +
on April 10, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from
DAFigure 4
C -/- -/- +/- +/- +/+ +/+ AH927
env
ERV-DC10
pLTR
β-actinkDa75
q
50
WB:Anti-RD114
C1ERV-DC10 locus(C1q12-21)
E37
β-actin
pB C -/- -/- -/- -/- +/- +/- +/-
ERV-DC18
env
LTR
q
C DC8 DC14 DC10ERV-DC10
LTR
β-actin
D4ERV-DC18 locus(D4q14)
C -/- -/- -/- -/- +/- +/- +/+env
β-actin
on April 10, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

A
Figure 5
PolB
Env/TM FERV-18100 ERV-DC18FERV-1876 ERV-DC18Pol Env/TM FERV-ECE1
FeLV-A clone33
FeLV-A 61E
enFeLV AGTT
Rat ERVs96
100
8581
ERV-DC ECE1
viru
s
FERV-ECE1
RD114 SC3C
BaEV M7
Macaque ERVs
Chimpanzee ERVs100
7699
9999
99
ERV-DC ECE1
Mouse ERVs and XMRV
Mouse ERV Chr.18
F-MuLV
M-MuLV
PERV C
Gamma
77
77
96
87
Gam
mar
etro
v
FeLV-A clone33
FeLV-A 61E
enFeLV AGTT
Rat ERVs
F-MuLV
Gamma
100
9099
87
100
8781 mar
etro
viru
s
PERV-C
KoRV
GaLV
BaEV M7
RD114 SC3C
REV-A Beta
82
95
89
viru
s
M-MuLV
XMRV
Mouse ERVs
PERV-C
KoRV
GaLV10090
100
88
100
Gam
m
MPMV
SRV-4
Beta
JSRV
HERV-K
MMTV
Beta
99
100
95
usB
etar
etro
vGaLV
REV-A
HERV-K
SRV-4
MPMV
JRSVBeta
998587
100
Bet
a-re
trovi
rus
JSRV ALV
MAVAlpha
100
0.5
Alph
a-re
trovi
r u MMTV
ALV
MAVAlpha
87
100
0.5
rAl
pha-
retro
viru
s
on April 10, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

Figure 5
ERV DC3 5
env LTRDC
ERV DC1
ERV-DC3 5
ERV-DC1 5
ERV-DC1 3
ERV-DC4 5
ERV-DC4 378
39
38 ERV-DC1
ERV-DC8
ERV-DC14
ERV-DC17
ERV-DC3
ERV-DC4Group I
100
98
ERV-DC14 5
ERV-DC14 3
ERV-DC3 3
ERV-DC17 5
ERV-DC17 3
62
38
50
54
98Group I
Group I
ERV DC4
Fc21
ERV-DC5
ERV-DC2
ERV-DC19
ERV-DC7
91
100
100
ERV-DC8 3
ERV-DC8 5
ERV-DC19 5
ERV-DC19 3
ERV-DC7 3
100
81
Fc41
ERV-DC16
Group II
ECE1
ERV-DC6
ERV-DC10
ERV DC18
Group III
100100
100
100
ERV-DC16 3
ERV-DC7 5
ERV-DC6 3
ECE1 3LTR
ERV-DC18 5
8497
95
Group III
Group II Group II
ERV-DC18100
0.01
ERV-DC10 3
ERV-DC10 5
ERV-DC18 3
SC3C 5LTR
CRT1 5LTR
100
100
95
RD114
Group III
CRT1 5LTR100
0.01
on April 10, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

Figure 6
A 5’LTR 3’LTRgag ATG env ATGpol stop env stopgag stop* **
gag - polFeLV-A/61E
FeLV-D/ON-T
envgag pol
1kb
gag stop5’LTR 3’LTRgag ATG env ATGpol stop env stop
env
* **
gag - Δpol
B FeLV-D positiveC 1632bp 720bp
FeLV-D env0 3 4 5 6
wks
c-myc
FeLV-D envβ-actinFeLV env
D FeLV-A/61E
F LV D/ON T
3’LTRSU TM
FeLV-D/ON-T
FeLV-D/re-T
FeLV-D/ON-C
FeLV-D/Ty26
FeLV-D/44B
ERV-DC8
on April 10, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

A (Glasgow1)
B
HEK293T/FeLV-A (clone33)
A (Glasgow1)
B
HEK293T/FeLV-BFigure 7
BCD (ON-T)D (Ty26)
ERV-DC10
FeLVBCD (ON-T)D (Ty26)
ERV-DC10
FeLV
ERV DC10LacZ Titer (I.U.)102 104 106
A (Glasgow1)
HEK293T/FeLV-C
ERV DC10LacZ Titer (I.U.)102 104 106
A (Glasgow1)
HEK293T/ERV-DC10
BCD (ON-T)D (Ty26)
FeLVBCD (ON-T)D (Ty26)
FeLV
ERV-DC10LacZ Titer (I.U.)102 104 106
ERV-DC10LacZ Titer (I.U.)102 104 106
HEK293T/FeLV-A (clone33)+FeLV-D (ON-T)
A (Glasgow1)
BCD (ON-T)
FeLV
D (Ty26)ERV-DC10
LacZ Titer (I.U.)102 104 106
on April 10, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from