william g. eberhard m - smithsonian institution
TRANSCRIPT

The Ecology and Behavior of aSubsocial Pentatomid Bugand Two Scelionid Wasps:
Strategy and Counterstrategy in aHost and Its Parasites
WILLIAM G. EBERHARD
m
wtu.
SMITHSONIAN CONTRIBUTIONS TO ZOOLOGY • NUMBER 205

SERIAL P U B L I C A T I O N S OF T H E S M I T H S O N I A N INSTITUTION
The emphasis upon publications as a means of diffusing knowledge was expressedby the first Secretary of the Smithsonian Institution. In his formal plan for the Insti-tution, Joseph Henry articulated a program that included the following statement:"It is proposed to publish a series of reports, giving an account of the new discoveriesin science, and of the changes made from year to year in all branches of knowledge."This keynote of basic research has been adhered to over the years in the issuanceof thousands of titles in serial publications under the Smithsonian imprint, com-mencing with Smithsonian Contributions to Knowledge in 1848 and continuing withthe following active series:
Smithsonian Annals of Flight
Smithsonian Contributions to Anthropology
Smithsonian Contributions to Astrophysics
Smithsonian Contributions to Botany
Smithsonian Contributions to the Earth Sciences
Smithsonian Contributions to Paleobiology
Smithsonian Contributions to Zoology
Smithsonian Studies in History and Technology
In these series, the Institution publishes original articles and monographs dealingwith the research and collections of its several museums and offices and of professionalcolleagues at other institutions of learning. These papers report newly acquired facts,synoptic interpretations of data, or original theory in specialized fields. These pub-lications are distributed by mailing lists to libraries, laboratories, and other interestedinstitutions and specialists throughout the world. Individual copies may be obtainedfrom the Smithsonian Institution Press as long as stocks are available.
S. DILLON RIPLEY
SecretarySmithsonian Institution

S M I T H S O N I A N C O N T R I B U T I O N S T O Z O O L O G Y • N U M B E R 2 0 5
The Ecology and Behavior of aSubsocial Pentatomid Bugand Two Scelionid Wasps:
Strategy and Counterstrategy in aHost and Its Parasites
William G. Eberhard
SMITHSONIAN INSTITUTION PRESS
City of Washington
1975

ABSTRACT
Eberhard, William G. The Ecology and Behavior of a Subsocial Pentatomid Bugand Two Scelionid Wasps: Strategy and Counterstrategy in a Host and Its Para-sites. Smithsonian Contributions to Zoology, number 205, 39 pages, 13 figures,24 tables, 1975.—The tropical bug Antiteuchus tripterus and two parasites of itseggs, Trissolcus bodkini and Phanuropsis semiflaviventris, were studied in urbanCali, Colombia. Female bugs guard both their eggs and first instar nymphs.Behavioral observations, combined with analyses of the patterns of parasitismwithin egg masses of a given form, showed that the details of the bug's defensivebehavior and the wasps' attacks strongly influence the survival rates of eggs indifferent positions in an egg mass.
By removing guarding female bugs from their eggs, it was shown that the neteffect of their defense against generalized egg predators is to greatly improve eggsurvival but that, paradoxically, the net effect against the parasitic wasps is todecrease survival. Although the bugs repel wasps that have found egg masses, thebug's presence over the mass betrays the location of the eggs to hunting wasps.A hypothetical evolutionary history suggests that as the bugs improved theirprotection of eggs, they produced thinner egg shells, and that they are nowtrapped into continuing their defense in spite of subsequent advances by thewasps in their hunting behavior. Experimental removal of bugs guarding firstinstar nymphs and calculations of the cost of guarding in terms of egg productionshowed that the net effect of this guarding behavior is positive.
Female P. semiflaviventris tend to stay and hunt in the tree where they emerge,while T. bodkini females tend to disperse. Parasitism rates by P. semiflaviventrisare strongly influenced by the species of tree where the bug eggs are laid, andboth species are influenced by the site of the egg mass (leaf, branch, sucker), butin different ways. Both species also wait near bugs about to oviposit and attackeggs as they are laid, apparently recognizing gravid bugs visually. Parasitismrates of eggs on naturally and experimentally isolated leaves showed that searchbehavior is accomplished in flight.
Natural selection with respect to oviposition site varies greatly with tree speciesand the site on a given tree. The balance of selection was evidently differentwhere the bugs evolved than in present-day Cali, where they commonly actagainst their own best interests.
Both wasp species agree in general with the behavior pattern for Scelionidae,but differ in several characteristics associated with oviposition. P. semiflaviventrisis consistently farther from the family norm than is T. bodkini. The two speciesof wasp compete at two stages in their life cycle, with neither species consistentlydominating. The males fight and exclude each other from egg masses wherefemales will emerge, and multiple ovipositions into single bug eggs produceonly single wasps. Both types of competition occur commonly in urban Cali.
Many other aspects of the natural history and ecology of the bugs and waspsare presented.
OFFICIAL PUBLICATION DATE is handstampcd in a limited number of initial copies and is recordedin the Institution's annual report, Smithsonian Year. SI PRESS NUMBER 5381. SERIES COVER DESIGN:The coral Montastrea cavernosa (Linnaeus).
Library of Congress Cataloging in Publication Data.Eberhard, William G.The ecology and behavior of a subsocial pentatomid bug and two scelionid wasps.(Smithsonian contributions to zoology ; no. 250)Supt. of Doc. no.: SI 1.27:2051. Antiteuchus tripterus. 2. Trissolcus bodkini. 3. Phanuropsis semiflaviventris. 4. Host-
parasite relationships. 5. Insects—Behavior. I. Title. II. Series: Smithsonian Institution.Smithsonian contributions to zoology ; no. 205.
QL1.S54 no. 205 no. 205 [QL523.P5] 591'.08s [595.7'54] 75-619087

Contents
Page
Introduction 1Materials and Methods 2The Natural History of Antitenchus tripterus limbativentris 2
Distribution and Host Plants of the Bugs 3Eggs 3First Instar Nymphs 6Second Instar Nymphs 6Older Nymphs and Adults 7Behavior of Females Guarding Eggs and Nymphs 10
The Natural History of the Wasps Trissolcus bodkini and Phanuropsissemiflaviventris 12
Hunting Behavior 13Oviposition 17Stages in the Eggs of A. tripterus 20Emergence from the Egg 21Sex Ratios 21Male Behavior 21Clusters of Females 22
Interactions between Wasps and Bugs 23Wasps vs. Bugs Guarding Eggs 23Wasps vs. Ovipositing Bugs 27
Competition between Species of Wasps 28Within Bug Eggs 28Fights between Males 29
Experimental Removal of Bugs Guarding Eggs 29Other Factors Affecting A. tripterus Survival 30
Unhatched Eggs 30Leaves and Egg Masses That Fell from Trees 31Losses of First Instar Nymphs 31Other Enemies of A. tripterus 33
Discussion 33Hunting Behavior of the Wasps 33Competition between the Wasps 34Evolutionary Relationships of the Wasps 35Selection Pressures on the Behavior of the Bugs 36
Literature Cited 38
in

FIGURE 1.—Female A. tripterus defending her eggs against the scelionid egg parasites Trissolcusbodkini (rear edge of egg mass) and Phanuropsis semiflaviventris (in front of bug). As theP. semiflaviventris female backs in with her abdomen extended, the bug lowers her antennae tothe leaf and scrapes the edge of the egg mass with her front foot, and the T. bodkini femaleoviposits in an egg at the poorly guarded rear edge.

The Ecology and Behavior of aSubsocial Pentatomid Bugand Two Scelionid Wasps:
Strategy and Gounterstrategy in aHost and Its Parasites
William G. Eberhard
Introduction
This paper describes the natural history of thetropical pentatomid bug Antiteuchus tripteruslimbativentris Ruckes and two parasitic wasps,Trissolcus bodkini and Phanuropsis semiflavi-ventris Girault, which attack the bug's eggs. Fe-male bugs guard their eggs from the wasps andalso guard their first instar nymphs. The wasps usespecial behavior patterns to overcome the femalebug's defensive behavior and oviposit in her eggs(Figure 1). The wasp larva kills the bug embryoand consumes the entire contents of the egg; thenit pupates and finally emerges as an adult from theegg (generally while the female bug is still standingguard over her nymphs) mates (usually with asibling), and then leaves to seek out new eggs toparasitize.
The quantitative results of the bug's selection ofoviposition site, of her defensive behavior, of com-petition between the two wasp species, and of thespecialized attack behavior of one of the wasps(P. semiflaviventris) were measured by analysesof variations in the patterns of parasitism betweenand within egg masses and by experimental re-
William G. Eberhard, Departmento de Biologia, Universidaddel Valle, Cali, Colombia, and Smithsonian Tropical ResearchInstitute, P. O. Box 2072, Balboa, Canal Zone.
moval of defending bugs from their eggs andnymphs. Parental protection of the young (sub-social behavior) has evolved many times in theinsects (see E. O. Wilson, 1971), and in theHemiptera is known in 14 species in 12 genera(Bequeart, 1935; Odhiambo, 1959), but this isapparently the first detailed study of the preciseeffects of parental care in subsocial insects.
ACKNOWLEDGMENTS.—I owe a large debt of grati-tude to the people who kindly identified the or-ganisms in this study: M. Becker {A. tripterus),W. Brown (ants), L. S. Espinal (plants), H. E.Evans (Bicyrtes), V. Gapud (other pentatomids),R. D. Gordon (coccinellids), B. Jimenez and D.Greer (fungus), and L. Masner and C. F. W.Muesebeck (parasitic wasps). I am especially grate-ful to James Slater and George Vogt for variousforms of help and encouragement. John Lawrencehelped obtain the sticky material used in variousexperiments, and the James Spain family allowedme to clutter their trees with sleeve cages. M. J. W.Eberhard, M. Emsley, D. Otte, J. Reiskind, G. W.Salt, N. G. Smith, and D. Strickland kindly criti-cized earlier drafts of this manuscript. Figure 1 wasdrawn by Dana Gardner.
This study was supported by the Comite de In-vestigaciones, Universidad del Valle, Cali,Colombia.

SMITHSONIAN CONTRIBUTIONS TO ZOOLOGY
Materials and Methods
Data used in this study were collected intermit-tently in and near Cali, Colombia, from November1969 through December 1972 and in Bucaramanga,Colombia, in October of 1972. Voucher speci-mens of A. tripterus, T. bodkini, P. semiflavi-ventris, and all the species listed in Table 22 havebeen deposited in the Museum of ComparativeZoology at Harvard University, Cambridge, Massa-chusetts 02138; other voucher specimens have beenplaced in the collection of the Universidad delValle in Cali, Colombia.
Unless otherwise noted, all tests of significancewere Chi2. Two statistical procedures used needspecial explanation. Sometimes, in order to obtainlarge numbers of data to test hypotheses, it wasnecessary to sum the data from several trees havingdifferent parasitism rates. In these cases, the me-dian was established for each tree and the datafrom that tree were classified as either above orbelow this median; finally, the data (above themedian, below the median) were summed overseveral trees.
In comparing total parasitism rates (betweentrees, for example), it was not possible to useChi2 because host eggs were laid in groups andtheir probabilities of being parasitized were thusnot independent. Instead, all masses in each samplewith the usual number of 28 eggs were ordered onthe basis of numbers of eggs parasitized, and thenthe two samples were compared using the Will-coxon Sum of Ranks Test.
The sticky material used to isolate trees and eggmasses from walking insects was "Tack Trap," anonvolatile, relatively inert substance manufac-tured by Animal Repellents, Inc., Griffin, Georgia,USA.
The reader should keep in mind that all of theobservations were made in the highly unnaturalenvironment of urban Cali and that great caremust be used in extracting conclusions about theevolutionary interactions between the organismsinvolved on the basis of this data. For instance,the fact that the bug's oviposition preferences inCali cause it to place most of its eggs in relativelyhigh- rather than low-risk sites does not imply thatnatural selection has somehow failed to operateon the bugs; the most likely explanation is thatone or several of the many causes of egg mortality
is so modified in the urban environment that therelative mortality rates between oviposition sitesare changed as compared to those in the environ-ment in which the bug evolved. On the other hand,since many aspects of the behavior of both bugsand wasps are presumably innate and would beperformed in the same way in any habitable en-vironment in which the insects found themselves,certain types of evolutionary conclusions can bedrawn. For instance, the fact that the bug's defenseof its eggs, which gives numerous signs of beingdesigned to combat the parasitic wasps, neverthe-less increases rather than decreases the chances ofparasitism by the wasps implies that the parasiticwasps have in some sense "out-evolved" their hosts.The behaviors involved—defense by the bug,searching and attack by the wasps—very likely arethe same in "natural" situations (those in whichthe animals originally evolved, if such places stillexist), and the net result of the bug's defensewould thus also be the same.
The Natural History ofAntiteuchus tripterus limbativentris
Antiteuchus tripterus limbativentris Ruckes, arace described from the Cauca Valley of Colombia(Ruckes, 1966) belongs to the principally neo-tropical subfamily Discocephalinae. This speciesranges from Panama to Paraguay. I have found itin Colombia in zones classified by Espinal andMontenegro (1963) as dry tropical forest (Cali andPalmira), humid semitropical forest (Bucara-manga), and humid tropical forest (Buena-ventura). In places in Colombia it is an importantpest of cultivated cocoa (cacao) and mango trees,and is apparently used as a folk cure for intestinalparasites (Kardonne, 1972). Naundorf (1954) andSepulveda (1955) showed that the bug can act as avector of the fungus Monilia, which attacks thefruit of cocoa. Lima (1940) notes that A. amplusalso damages cocoa in Brazil, and Callan (1944b)records A. piceus as a minor cocoa pest in Trini-dad. Rau (1918), Callan (1944a), and Kirkpat-rick (1957) have observed that A. tripterus femalesstand guard over eggs and first instar nymphs, andCallan, Kirk pa trick, and Sepulveda (1955) all de-duced from the pattern of parasitism of several eggmasses that the bug's defense was at least partiallyeffective against the scelionid egg parasite Phanu-

NUMBER 205
ropsis semiflaviventris. Callan (1944b) recordedmaternal care in several additional Antiteuchnsspecies, including A. (A.) piceus, mixtus, andvariolus, and A. (Neodine) macraspis.
DISTRIBUTION AND HOST PLANTS OF THE BUGS
The bug A. tripterns is extremely common intrees and bushes in urban Cali, Colombia. Thebugs were nonspecific in their host plant selectionand were found on the trees Pithecelobium dulce(Combretaceae—an exotic), Canagium odoratum(Anonaceae—also an exotic), Cassia sp. (Cesal-pinaceae), Lagerstromia speciosa (Litraceae), Cres-centia cujete (Bignoniaceae), cocoa (Theobromacacao—Sterculiaceae), and mango (Mangiferaindica—Anacardiaceae). They also occurred on theshrubs Malvavicus arboreus (Malvaceae), Hibiscussp. (Malvaceae), Heliconia sp. (Musaceae), andBouganvilla glabra (Mictaginaceae). In the coun-tryside near Cali the bugs were found on P. dulce,Calliandra sp., Croton sp. (Euphorbiaceae), andTabebuia chrysantha (Bignoniaceae). Sepulveda(1955) presents a similarly varied host list. Bugpopulations were generally larger on some speciesof plants (such as P. dulce, T. catappa, and Cassiasp.) than on others (such as B. glabra, M. indica,and Heliconia sp.), and the majority of my obser-vations were made on the first three species.
Bugs were much more common on plants insidethe city than outside. This difference was clear ontwo species of trees (P. dulce and Calliandra sp.)that occurred commonly in both areas. It also ap-peared that within the city the bugs tended to bemore common where there was heavy pedestrianand/or automobile traffic. For instance, nine smallP. dulce trees at the edge of a park near the centerof the city (along the Rio Cali), where manypeople walked, supported heavy bug populations,while other similar trees 10-20 m inside the parkhad fewer bugs. The reason for these tendencies isunknown.
The size and form of a plant also appeared toinfluence its attractiveness to A. tripterus. Trees orbushes of a given species having denser growthtended to have more bugs. In the case of P. dulce,many large and small trees were searched, and al-though the absolute bug populations could notusually be determined, it was certain that thedensest populations were in medium and small
trees. The bugs were very common. One particu-larly densely populated tree was about 2.5 m highat the center and about 7 m in diameter, and hadover 200 female bugs guarding eggs or nymphs atone time. Based on surveys in other trees, this rep-resents a total adult population of about 2400bugs.
The effects of both tree form and location onpopulation size were relatively weak within the city,however, and I was not able to correctly predicthow many bugs a given tree would have. Bug pop-ulations in individual trees sometimes changedsubstantially over time.
EGGS
DESCRIPTION.—Eggs were barrel-shaped, 1.47 mmtall and 0.97 mm in diameter, and lacked thespines (chorionic processes) found on the rims ofmany pentatomid eggs (Esselbaugh, 1947). Theircuticle was smooth and relatively thin and weakcompared with those of the eggs of other penta-tomids such as Nezara sp., Supputius sp., Podisuscloelia, and Loxa sp. The cap was the strongestpart and was reinforced by a fine network of smallridges on its inner surface.
Two distinct colors of unparasitized eggs couldoften be distinguished within the same mass. Usu-ally (perhaps always—some differences were diffi-cult to be sure of) a group of 28 had 14 of onecolor and 14 of another. Most of the eggs of a givencolor were usually grouped more or less together.In some cases where the order of oviposition couldbe deduced (see below), it was clear that the eggsof one color were all laid before any of the others.Dissection of a female killed just after laying the14th egg of a mass showed that each of her 14ovarioles had lost one egg and had one left. Thusthe differences in color of the eggs seem to be as-sociated with the position (basal or apical) of theeggs in the ovariole.
Almost all embryos in a given group of eggswere usually oriented in the same direction withintheir eggs (upward with their ventral surfaces alloriented in the same direction), but usually at
TABLE 1.—Sizes of egg masses (N = 230)
Number of eggs in the mass: 28 27 26 14Frequency: 90% 6% 3% 1%

SMITHSONIAN CONTRIBUTIONS TO ZOOLOGY
TABLE 2.—Frequency of masses of 28 eggs that werein the "compact" form (Figure 12) in different treespecies
Location
Leaves ...
Branches
CanagiumP. dulce Cassia sp. odoratum T. catappa
32%(N=766)
32%(N=236)
44%(N=406)
27%(N = 189)
42%N = 224)
least one or two embryos in each group were sub-stantially out of line dorsoventrally with the others.Since the female's bug orientation during oviposi-tion was constant, these eggs must have turned be-fore or as they emerged.
SITES AND ARRANGEMENTS.—As shown in Table 1,there were almost always 28 eggs in each egg mass.The eggs in masses were generally closely packed,and the most common arrangement was the "com-pact" form pictured in Figure 12, with minorvariations on this (such as that in Figure 4) alsorelatively common. The frequency of the compactarrangement varied with the species of tree, asshown in Table 2; both T. catappa and Cassia sp.differ significantly (p<0.05) from both C. odora-tum and P. dulce. Since compact egg masses were
TABLE 3.—Relationship between total number ofegg masses in a P. dulce tree and the frequency ofoviposition on leaves vs. branches (p <.O1)
Site
Leaves ..BranchesNumberNumber
of massesof trees
Trees with > 20guarded egg masses
88%12%
130813
Trees with < 20guarded egg masses
76%24%
1069
less heavily parasitized than others (below), thesedifferences may influence parasitism rates on dif-ferent trees.
As in other pentatomids (Esselbaugh, 1946), theeggs were usually laid on the undersurfaces ofleaves or branches. As shown in Table 3, there wasa positive correlation on P. dulce trees betweenthe relative numbers of masses on leaves vs.branches and the total number of egg masses onthe tree (probably an indirect measure of the adultpopulation). This does not necessarily mean thatthe bugs were reacting directly to the populationsize on a given tree, as an additional factor or fac-tors could influence both population size andchoice of egg sites.
Compact masses of eggs on leaves were usuallyoriented so that the group's longitudinal axis wasparallel to that of the leaf and its pointed end wastoward the leaf tip, but this varied somewhat indifferent species of tree (Table 4). The orienta-tion of masses on T. catappa was significantly dif-ferent from those on the other tree species: it maybe that the greater size of its leaves is somehowresponsible for the difference (Figure 2). Since theorientation of the defending bug is highly cor-related with the orientation of the egg mass, andthe orientation of the bug is probably in turn cor-related with the hunting behavior of egg parasites(below), the consistent orientation of the egg masseson leaves may be the result of selection pressureexercised by hunting wasps.
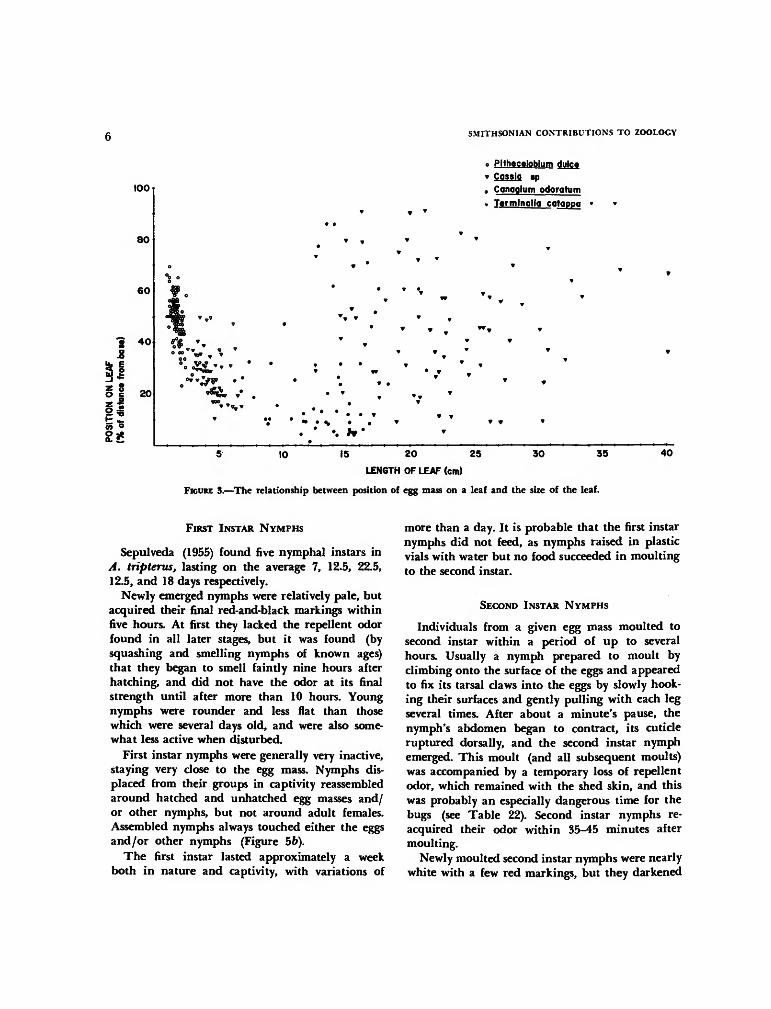
Figure 3 shows how oviposition sites varied withleaf size. On leaves from 3-10 cm long, the eggswere nearly always placed near the base. On verysmall leaves (less than 2 cm long) they tended tobe nearer to the tip (p<0.05), while on very largeleaves they were distributed randomly. The func-tion of the tendency to place eggs near leaf bases is
TABLE 4.—Orientation of "compact" egg masses on the leaves of different trees
Orientation
Longitudinal axis more nearly parallel thanperpendicular to leaf axis
Those so aligned which had point of grouptoward leaf tip
Total number of masses observed
P. dulce
100%
95%
163
Cassia sp.
97%
95%
132
Canagiumodoratum
90%
98%
50
T. catappa
65%
69%
87

NUMBER 205
not certain, but it may serve to protect them fromwater damage. On observing the undersides ofCassia sp. and P. dulce leaves just after a rain, Ifound that the water standing on them was con-centrated along the lower edge of the leaf, and inmost cases this was the tip of the leaf. Thus eggs atleaf bases are probably less likely to be coveredwith water during and after rain. Experimental im-mersal of eggs in water for various lengths of timeshowed that older eggs were killed more readily andthat appreciable reduction in hatching was firstseen after between 6 and 9 hours of immersion,when it fell from 70% to 10%.
The apparently random positioning of eggs onlarge leaves and their placement toward the tips ofsmall leaves do not at first seem to fit this explana-tion. Observations of large T. catappa leaves justafter rain indicate, however, that they seldom getwet at all on the underside where the bug eggs are
nearly always located; and placement of eggs nearerthe bases of leaves less than 2 cm long might bephysically difficult for the bug (length about 1 cm)since it lays the eggs nearest the leaf base last, andwould thus have to actually climb off the leaf tolay eggs nearer to the leaf base.
DEVELOPMENT.—No structure was visible withinthe eggs until 3.5-4 days after oviposition (thetimes varied somewhat, presumably due to temper-ature variations), when the embryo's eyes and eggburster became preceptible. They darkened gradu-ally (but did not grow in size) until the nymphshatched 130-170 hours (about 5.5-7 days) afterbeing laid. Sepulveda (1955), working in nearbyPalmira on cocoa (which is grown in the shadeand may have been cooler), reports the egg life as9-13 days. Eclosion times were spread over up to 12hours in a given egg mass, but no pattern in thesequence of hatching was discernible.
FIGURE 2.—Typical forms of the trees studied most intensively: a, Pithecelobium dulce (arrowmarks suckers); b, Cassia sp.; c, Terminalia catappa; d, leaves of (1) Cassia sp., (2) P. dulce,(3) T. catappa, and (4) Canagium odoratum (arrow marks bug guarding eggs); e, pattern ofleaf growth in P. dulce (arrow marks site where sticky substance was applied in isolation experi-ments); /, pattern of leaf growth in Cassia sp. (arrow marks site where sticky substance wasapplied in isolation experiments). (The scale bars in a, b, and c are approximately 2 m, thosein d and e are 5 cm, and that in / is 10 cm.)
SMITHSONIAN CONTRIBUTIONS TO ZOOLOGY
100
80
6 0
8 40
SI§ 1 20§1
PO
SIT
I'(%
of
d
f .
0 °° w» „ »
• •
* *
» » »
• » _ »• *
* • •• • • » » »
a Pithecelobium dulcev Cosslo sp
. Canagium odoratum, Terminolia catappa * *
V
w
»
T T »
10 3015 20 25
LENGTH OF LEAF (cm)
FIGURE 3.—The relationship between position of egg mass on a leaf and the size of the leaf.
35 40
FIRST INSTAR NYMPHS
Sepulveda (1955) found five nymphal instars inA. tripterus, lasting on the average 7, 12.5, 22.5,12.5, and 18 days respectively.
Newly emerged nymphs were relatively pale, butacquired their final red-and-black markings withinfive hours. At first they lacked the repellent odorfound in all later stages, but it was found (bysquashing and smelling nymphs of known ages)that they began to smell faintly nine hours afterhatching, and did not have the odor at its finalstrength until after more than 10 hours. Youngnymphs were rounder and less flat than thosewhich were several days old, and were also some-what less active when disturbed.
First instar nymphs were generally very inactive,staying very close to the egg mass. Nymphs dis-placed from their groups in captivity reassembledaround hatched and unhatched egg masses and/or other nymphs, but not around adult females.Assembled nymphs always touched either the eggsand/or other nymphs (Figure 5b).
The first instar lasted approximately a weekboth in nature and captivity, with variations of
more than a day. It is probable that the first instarnymphs did not feed, as nymphs raised in plasticvials with water but no food succeeded in moultingto the second instar.
SECOND INSTAR NYMPHS
Individuals from a given egg mass moulted tosecond instar within a period of up to severalhours. Usually a nymph prepared to moult byclimbing onto the surface of the eggs and appearedto fix its tarsal claws into the eggs by slowly hook-ing their surfaces and gently pulling with each legseveral times. After about a minute's pause, thenymph's abdomen began to contract, its cuticleruptured dorsally, and the second instar nymphemerged. This moult (and all subsequent moults)was accompanied by a temporary loss of repellentodor, which remained with the shed skin, and thiswas probably an especially dangerous time for thebugs (see Table 22). Second instar nymphs re-acquired their odor within 35-45 minutes aftermoulting.
Newly moulted second instar nymphs were nearlywhite with a few red markings, but they darkened

NUMBER 205
within a few hours to brown with black markings(see Sepulveda, 1955, for detailed descriptions ofall nymphs). They were longer and wider thanfirst instars (3.1 mm long X 2.4 mm wide vs. 2.4mm X 1.7 mm), but flatter and thinner, and weremuch more active. They dispersed readily whendisturbed and, if not disturbed, left the egg masstwo to three days after moulting. The nymphs ap-parently began to feed in this stage, and were foundalone on leaves and twigs far from the nearest eggmass. They showed little or no gregariousness afterleaving the vicinity of the egg mass, contrary to theobservations of Callan (1944b) on A. tripterus andA. picea.
OLDER NYMPHS AND ADULTS
Bugs which had left the egg mass were almostalways found on branches and trunks rather thanleaves, except on the very large leaves of T. catappawhere nymphs were often seen along the centralleaf veins. In general, the younger stages tended tostay on smaller branches. Outside the city mostbugs were found in hidden spots such as in crotchesand under tufts of lichen, but in urban trees, whichhad fewer epiphytes and thus fewer hiding places,they showed little tendency to concealment exceptthat they usually stayed on the lower rather thanthe upper surfaces of branches. In some trees theolder nymphs and adults clustered in groups of upto 40 or more; one sample group contained 7 males,10 females, and 11 penultimate nymphs.
OVIPOSITION.—Most adult bugs were found ontwigs and branches, but females with swollen ab-domens were not uncommonly found restingquietly on the undersides of leaves. These femaleswere evidently ready to oviposit, as they generallylaid eggs within several days without feeding whenbrought into captivity. In those trees on whoseleaves the female usually faced the tip while guard-ing her eggs (below), the swollen females also usu-ally faced the leaf tip. This immobile stage mayrepresent a period during which the female "mon-itors" the characteristics of a potential ovipositionsite before laying her eggs there. Females maychange sites several times before finally ovipositing;when I marked the leaves on which five such fe-males were resting (I tried not to disturb the bugas I marked the leaf), all were deserted several dayslater. These movements may serve as defense
against parasitic wasps which have located gravidbugs before they are ready to lay (below).
The act of oviposition was observed on sevenoccasions, and the movements were relatively con-stant. Figure 4 shows the sequence of the last 21eggs laid in one egg mass and illustrates a patternof placement also seen in other ovipositions. Thebug laid a line of three eggs (i.e., numbers 12, 13,and 14) on one side of the group, then two (i.e.,15 and 16) on the other side, and then repeatedthis sequence. This behavior appears to be de-signed to produce compact masses like that inFigure 12.
The bug began laying an egg by moving forwarduntil the tip of her abdomen was in line with thefront edge of the group of eggs already laid. Then,swinging her abdomen slightly from side to side,she positioned its tip in the groove between two ofthese eggs. The side-to-side movements probablyenabled her to feel the positions of already laideggs (which she could not see), possibly using thelaterotergites of segment nine. The bug loweredher abdomen as she positioned herself and touched
FIGURE 4.—The sequence of the last 21 eggs laid by a femaleA. tripterus.

SMITHSONIAN CONTRIBUTIONS TO ZOOLOGY
the emerging egg to the leaf. Then, moving slightlybackward and lifting her abdomen, she drew theegg out and tilted it back against the sides of theeggs already laid.
Females paused from 15 to 60 seconds betweenovipositions, some bugs moving backward and moreor less covering the egg mass during these pauses,others moving back only slightly. The backwardmovement may play a part in the defense of theeggs against parasitic wasps (below). Laying fe-males generally faced the base of the leaf, and aftercompleting a mass of eggs, they turned 180° andscraped the sides of the mass several times beforesettling down—probably another defense againstwasps.
The period of time between a female's abandon-ment of her second instar nymphs and the layingof her next egg mass was determined in the fol-lowing way. Bugs caught guarding second instarnymphs on P. dulce trees (i.e., females no morethan a day or two away from deserting theirnymphs) were placed in a sleeve cage on anotherP. dulce. The mean time (to the nearest day) tothe next oviposition in two such experiments wasabout 17 days (Table 5). Thus the normal timebetween ovipositions is probably on the order of30-35 days (7 days guarding eggs, plus 7 daysguarding first instar nymphs, plus 1-2 days guard-ing second instar nymphs, plus about 17 days offeeding to prepare for the next oviposition).
Since female bugs probably seldom if ever feedwhile guarding, and since females guarding eggshave more fat than those guarding second instarnymphs (below), it seemed likely that femalesfound guarding new eggs would be able to reovi-
TABLE 5.—Number of days required by female bugsto reoviposit
Treatment
Females removed from groupsof second instar nymphs
Experiment 1Experiment 2
Females removed fromnewly laid eggs
Experiment 1Females removed from groups of
newly hatched first instarsExperiment 2
Average
16.617 2
20
19
Range
12-2113-25
16-25
14-24
Numberof bugs
1335
21
22
posit sooner when removed from their eggs thanthose removed from second instar nymphs. Just theopposite was true, however, as 21 females whichwere found on new eggs and placed in the samesleeve cage, on the same day, with the females re-moved from second instar nymphs in the first ex-periment described above averaged 20 days to thenext oviposition, requiring significantly longer(p< 0.002, Willcoxon Sum of Ranks Test) thanthe others (Table 5). A similar experiment, duringsomewhat cooler weather and involving femalestaken from newly hatched first instar nymphs andothers taken from second instar nymphs, gave me-dian reoviposition time of 19 and 17 days respec-tively (Table 5), the females from second instarnymphs again laying eggs in significantly shortertime (p<0.01, Willcoxon Sum of Ranks Test).These data suggest that the female undergoes somesort of hormonal cycle controlling fat and/or eggsynthesis even when out of contact with her eggsand nymphs.
These data should perhaps be treated withcaution, at least concerning the absolute lengths ofinteroviposition periods, since of 35 masses laid byfemales removed from second instar nymphs in thesecond experiment 8 had only 14 eggs (differs fromTable I, (p<0.01). Only females taken from sec-ond instars showed this behavior, as none of 22masses laid during the same time by females fromfirst instar nymphs had 14 eggs (p<0.05). However,dissections of bugs found guarding newly laidmasses with less than 28 eggs showed that femalesprobably usually do have a full complement of 28eggs ready to lay, but may fail to lay some of them.Each of four females on masses of 14 eggs had 14more eggs of nearly identical size inside her ab-domen and a fifth female on a mass of 15 had 11more eggs inside.
MIGRATION.—The great majority of nymphs andadults on one P. dulce tree were removed by handon 11-13 September 1972, and the tree was thenrevisited and again cleaned of adults and largernymphs 16 and 25 days later. These repeat visitsrevealed a substantial rate of immigration; 214 fe-males and 213 males were found during the firstre-collection and 71 and 79 during the second, giv-ing a combined rate of 11.5 females and 11.7 malesarriving at the tree per day. This figure may beslightly high as a few adults were undoubtedlymissed during the first collection. The study tree

NUMBER 205
was not isolated, being part of a row of P. dulceand only about 2 m and 8 m from its nearest neigh-bors (trunk-to-trunk distances), both of which hadsubstantial bug populations. Nevertheless, thishigh rate of movement between trees was un-expected because (1) A. tripterus is not an espe-cially good flier and even dislodged bugs usuallysimply circled and flew back upwind to land on thetree where they had been, and (2) bugs were sel-dom seen flying, even in densely populated areas.
The original collection of adults from this treehad 518 males and 696 females or a ratio of 0.74male: female. Assuming that the male: female ratioin the trees from which the immigrants came wasthe same, the relatively large number of maleswhich actually arrived suggests that males are moremobile than females (p<0.01). This is perhapsnot surprising since females are immobile approxi-mately two weeks while guarding the eggs andnymphs of each brood. Since they live approxi-mately 80 days as adults (Sepulveda, 1955) andprobably lay 2-3 batches of eggs, this wouldamount to 35%-50% of their life spent immobile.In fact, assuming conservatively that females spendonly 35% of their time guarding, that both sexesmigrate equally, and that the total sex ratio in thepopulation is 0.74:1 male:female, the expectednumbers of arriving males and females would be306 and 269, not significantly different from theobserved 292 and 285.
Migrating females were apparently more likelyto lay eggs than others. Thus 22.2% of the 696 fe-males in the first collection were guarding eggs ornymphs; assuming a steady immigration rate (11.5females/day) and the frequency of ovipositioncalculated from the first collection (0.017 mass/female/day), only 33 of the immigrant femalesshould have been guarding offspring rather thanthe observed 58 (p<0.01).
The relative numbers of nymphs and adultsfound in this tree suggest the additional conclu-sion that, despite the high rate of arrivals, evenmore bugs left the tree than arrived. Relativelycareful counts of the numbers of unhatched eggspresent 16, 9, and 1 week (s) before the first col-lection indicated that the adult female populationwas approximately constant during this period.Since the total time from egg to adult is about 10weeks (Sepulveda, 1955), relatively constant num-bers of adults were being produced from nymphs
during at least the last 6 weeks before the first col-lection was made. The total number of last (5th)instar nymphs found on the tree in the first col-lection was 1108, or approximately 90% of thetotal number of adults. Since substantial move-ments between trees by nymphs are unlikely, andsince the duration of the 5th instar is only about20% that of the adult, it appears that a large por-tion of the bugs which grew up on the tree leftafter reaching maturity.
Several isolated observations indicated that adultbugs of both sexes leave in especially large numberswhen trees shed their leaves. The best studied casewas in one P. dulce, which had had weekly countsof 60-75 masses of unhatched bug eggs for fiveweeks; within three weeks during which the treewas shedding, the count fell to 5 masses. Observa-tions of the trunk and branches showed that theentire adult population in this tree had droppeddramatically.
Other more casual observations also associatedshedding with bug desertion in both P. dulce andT. catappa trees. None of the four tree specieswhich were observed most intensively seemed tohave an established season for shedding and, withthe exception of some individuals of T. catappa,these trees were never completely without leaves,new ones emerging before the last of the old onesfell. I saw no dramatic fluctuations in bug popula-tions during the erratic dry and wet seasons inCali, but did not make careful long-range counts.Eggs were found at all times of the year.
OTHER BEHAVIOR.—Mating, feeding, and defeca-tion were seen on trunks and branches. I foundsix mating pairs, each with the two bugs facingaway from each other. Feeding bugs often held thetips of their abdomens lifted away from the branch,and sometimes appeared to have difficulty with-drawing their probosces from the wood when dis-turbed. Although ants such as Ectatomma ruidumand Campanotus sp. often tended membracids andcoccids in trees harboring A. tripterus, I never sawan ant show any sign of interest in a bug. The bugslifted their abdomens when they defecated a clear,tasteless (to me) liquid, several drops at a time.
Both adults and nymphs gave characteristic re-sponses when they were disturbed by bugs or otherobjects which approached them. The most com-mon responses were to antennate toward the ob-ject, to give a brief "shudder" of the entire body,

10 SMITHSONIAN CONTRIBUTIONS TO ZOOLOGY
and to walk away. Upon more intense disturbance(pinching a foot, for example), both nymphs andadults kicked at the offending object with the mid-dle or hind legs and/or gave off an unpleasantodor. In favorable lighting one could sometimessee the adults squirt the substance several centi-meters laterally. Usually they buzzed their wingsfor several seconds a minute or so later, perhaps torid themselves of any liquid still clinging to theirbodies. This behavior is probably what Callan(1944b) took to be part of the defensive behaviorof females guarding eggs.
The repellent substance was poisonous and bothbugs and their eggs were killed by being placed in ajar where adults had recently given off odor. Ontwo occasions I saw a first instar nymph die withina minute after being hit directly by a spray (I hadpushed the female partway off her group ofnymphs). The substance was painful when placedon soft areas of my skin or near my eyes and alsoapparently corroded the plastic of a pen which Iused to nudge females off their groups of eggs andnymphs.
Both males and females had bright orange bandsalong the lateral edges of the ventral surface oftheir abdomens. When adult bugs were disturbedby the wasp Bicyrtes variegata (a predator onlarge nymphs—see Eberhard, in press), or by otherstimuli approaching them more or less dorsally,they sometimes tilted their bodies laterally, a move-ment probably designed to facilitate spraying theoffending object with repellent fluid (which wasejected from openings near the bases of the legs).The orange flanks of the bugs were also exposed bythis tilting of the body and the coloration maythus be aposematic in function.
Adult males were usually darker (dark brown)dorsally than females (tan to brown). I was able tocorrectly guess the sex of 71 of 75 bugs in the fieldsolely on the basis of their dorsal coloration, miss-ing two males and two females.
Bugs resting on the undersides of T. catappaleaves in bright sunlight extended their legs to lifttheir bodies away from the leaf surface, a move-ment probably serving in temperature regulation.Bugs in raised positions lowered themselves with-in a few minutes when their leaves were shaded,and rose up again when the shade was removed.Both bugs guarding eggs and others raised them-selves in this way, but guarding bugs immediately
lowered themselves when disturbed by anapproaching object or shadow.
BKHAVIOR OF FEMALES GUARDING EGGS AND NYMPHS
Female A. tripterus stood over their eggs andnymphs both night and day and responded to vari-ous types of disturbances. They kept the eggsunder the front part of their bodies (Figure 5) andwere not easily displaced by pushing or pulling.Their defensive movements included: tilting thebody toward the disturbance (thus forming a shieldbetween it and the eggs or nymphs); waving one orboth antennae toward it; shaking the entire bodywith quick shuddering movements; scraping theedge of the egg mass with the front legs (usuallydone alternately, with the bug swinging her bodylaterally so that she scraped along most or all ofthe entire front and lateral edge of the mass, al-though occasionally they also scraped with theirmiddle legs); and kicking backward with the hindand middle legs. Antennal and, to a lesser extent,leg movements were directed toward the disturbingstimulus. First instar nymphs clustered below thefemale did not generally respond to her defensivemovements, but second instars often dispersed.
When female defensive responses were tested bypresenting the bugs with a model parasitic wasp,females guarding eggs gave somewhat different re-sponses than those guarding nymphs. The modelconsisted of a small bit of black paper about thesize of a parasitic wasp glued to the end of a 15mm clear nylon thread that was fixed to the end ofa 25 cm glass tube. The stimulus was moved nearthe bug in approximately the pattern shown inFigure 6. Visual stimuli from my hand may havealso stimulated the bug and, since she usuallymoved her antennae toward the model as it ap-proached, presentations often involved tactile stim-ulation as well. It was clear from these tests andfrom observations of wasp-bug interactions, how-ever, that the sight alone of a wasplike object wouldrelease the bug's defensive behavior.
The frequencies of the various defensive re-sponses are presented in Table 6. Scraping behav-ior diminished significantly when the bugs wereguarding nymphs, but shuddering increased. It isreasonable that scraping should decrease since (1)the female might damage her nymphs or knockthem off the leaf, and (2) there is no longer any

NUMBER 205 11
danger of parasitism by the wasps against whichthis movement is particularly effective. The signifi-cance of the change in shuddering behavior is notknown.
Only very seldom, and only in moments of greatduress, did bugs battling wasps give off enoughodor for me to sense it. I also had the strong im-pression that bugs guarding eggs were much lesslikely to discharge the poisonous repellent odorwhen I disturbed them than were bugs over nymphsor nonguarding bugs.
Female bugs showed no sign of being able todistinguish their own eggs from those of others andit was possible to transfer females to new eggmasses if the bug was moved directly from one tothe other. The longer the bug was out of contactwith eggs, however, the less likely she was to acceptthe new group (even if she was placed back on heroriginal mass). On several occasions I saw a femalewhich had apparently changed groups spontane-ously, leaving a group of new eggs unguarded asshe stood over a nearby group of old, empty eggs.
Females guarding nymphs were apparently moreattracted by the nymphs than by the mass of empty
eggs, as several females whose nymphs had clus-tered a centimeter or so from the egg mass werestanding over the nymphs rather than the eggs.Females did not show any response to stray nymphs,however.
Female bugs probably seldom if ever fed whileguarding eggs or nymphs, since I never saw aguarding female feed and since adults were seenfeeding only on trunks and branches, never onleaves where most eggs were located. The defensivebehavior of a female with her proboscis stuck inthe plant would probably be less effective since shewould be less mobile, and this may explain thefemale's abstinence. Sepulveda (1955) noted punc-ture marks near egg masses on the stems of cocoapods and speculated that defending females mayfeed. The evidence is weak, however, since stemsare a preferred feeding site for eggless bugs (Naun-dorf, 1954). Dissections of ten females collectedguarding newly laid eggs and of ten others col-lected guarding second instar nymphs also indi-cated that defending females fed little, if at all.The females from new eggs all had moderate tolarge fat bodies, while the others, which had been
FIGURK 5.—a, Female A. tripterns guarding her eggs on a leaf of C.odoratum (arrow indicates rear, poorly defended side of egg mass);b, female bug guarding first instar nymphs on a branch.

12 SMITHSONIAN CONTRIBUTIONS TO ZOOLOGY
TABLE 6.—Responses of defending female bugs toartificial "wasp" stimuli
FIGURE 6.—Pattern of presentation of model wasp to defendingfemale bug.
guarding for about two weeks, had small or at mostmoderate fat bodies. Most of the fat was probablyused to maintain the bug itself, as the ovaries inboth groups of females were undeveloped and ap-proximately the same size.
The females' orientation as they guarded was notrandom. On all except very large leaves (i.e., T.catappa leaves), they tended to align themselveswith the leaf's axis, facing either its base or its tip.In a study of the position of female bugs on P.dulce leaves, it appeared that both the leaf itselfand, to a lesser extent, gravity had an orientingeffect on the bug. On nearly horizontal leaves,83% of 69 females faced toward the tip rather thanthe base; on leaves with their bases below their tips,this percentage was 73% (N = 66); and on leaveswith their tips below their bases, it was 94%(N = 522) (the last two figures differ significantly,p<0.01). This orientation, which would seem tomake the bug less able to sense dangers arriving atits leaf by walking along the stem, may be a resultof the parasitic wasp's tendency to hunt in flight.
Bug's response
Move antenna towardstimulus
Lean toward stimulusMove toward stimulusShudderScrapeKick
Bug guardingeggs (43 trials)
43
4238
17*28*16
Bug guardingnymphs (42 trials)
424141
36*8*
24
•Differ significantly (p < .05).
The orientation of females when guarding com-pact egg masses like that in Figure 12 was also farfrom random, even on T. catappa leaves where theegg mass itself was often not aligned with theleaf's axis (Table 4). Of 94 females found on suchgroups in T. catappa, 92% faced so their headswere over the pointed end of the group (the topof the group in Figure 12). This consistency prob-ably aids in the defense of the group by bringingthe bug's front legs to bear on the smooth sides ofthe mass where their scraping will be most effective.
The frequency with which female bugs desertedtheir offspring was lower in some trees than inothers. During weekly visits to one P. dulce tree ona very busy street where large trucks and busesoften passed close by (but where the bugs wereextremely common), females were missing from27% of 195 groups of first instar nymphs, while thecorresponding figure from a slightly smaller P.dulce at the edge of a small park in a residentialsection was 11% (N = 82) (p<0.05).
The Natural History of the Wasps Trissolcusbodkini and Phanuropsis semiflaviventris
The wasp Phanuropsis semiflaviventris Girualtis the only described species in the genus Phanu-ropsis (a second species, to be described by L.Masner, was raised from the eggs of Edessa sp.,another subsocial penatomid), and there is noliterature on its biology other than that it para-sitizes the eggs of A. tripterus and A. picea (Gir-ualt, 1916; Callan, 1944a). Wasps of the genusTrissolcus are parasites of pentatomid eggs (Ash-mead, 1893). Vaukassevitch (1925; cited in F.Wilson, 1961) describes several aspects of the ovi-

NUMBER 205 13
position behavior of T. simoni which conform tothe general pattern for the family Scelionidae.Both P. semiflaviventris and T. bodkini "werefound in A. tripterus eggs in Cali and Buenaven-tura, Colombia, while only P. semiflaviventriswas present in Bucaramanga, Colombia, and ap-parently in Trinidad (Callan, 1944a); T. bodkiniwas the only species in a small sample of four para-sitized masses collected in Guapi, Colombia.
HUNTING BEHAVIOR
GRAVID BUGS.—Females of both species of waspwere not infrequently found on leaves waitingnear swollen female bugs that were apparentlyabout to oviposit (Figure 7), and on several occa-sions they were observed parasitizing eggs as thebug oviposited (female wasps were distinguishedin the field by their more pointed abdomens andmore clubbed antennae). Several females were alsoseen waiting near mature male bugs and penulti-
FIGURF. 7.—Female T. bodkini wasp waiting near a gravidfemale bug.
mate nymphs which were resting immobile onleaves, suggesting that the important cues used torecognize gravid females were visual—their generalsize and the unusual sites (leaves) where theyrested. An experiment designed to test this, how-ever, indicated that additional factors were alsoinvolved.
About 40 mature bugs of both sexes were killedin alcohol, dried, and mounted with their legs re-moved on short insect pins. The bugs were thenstuck on the undersides of the leaves of a Cassiasp. tree, which had substantial populations of bugsand both species of wasp. When I returned thenext day, only one of the dead bugs had a femalewasp waiting near it; the behavior of this female(a P. semiflaviventris) suggested why wasps hadnot congregated around the models. She walked bythe base of the leaf with the model on it seven timeswithin about 15 minutes, each time starting outtoward the bug only to turn back before reachingit. This suggests that the sight of the bug was at-tractive to the wasp, but that the presence orabsence of some other cue (s) counteracted thisattraction. The wasp eventually flew away.
Gravid bugs probably change leaves from time totime before laying, but T. bodkini wasps whichwere waiting nearby did not seem able to followthem. When a gravid bug and two female T. bod-kini, which were found waiting at her side, wereplaced in a large glass jar along with the branchof C. odoratum on which they were found, the bugmoved periodically from one leaf to another, butthe wasps did not follow her as she moved. In onetypical case neither wasp moved until the bug was1.5-2.5 cm away, and then they both walked inapparently random directions rather than towardthe retreating bug. Nevertheless, within 30 min-utes the bug was quiet again and both wasps werewaiting near her. The bug's only response to thewasps in this situation was to antennate towardthem when they approached her. These observa-tions suggest that the immobility of the bug is animportant cue for the wasp, and that once the bugmoves it is no longer attractive until it stops again.
EGG MASSES.—Female wasps of both species be-haved as if they used the sight of female bugs tolocate egg masses as well as gravid females. Femalesof P. semiflaviventris demonstrated this most dra-matically. When a guarding female bug was dis-placed several centimeters to the side of her group

14 SMITHSONIAN CONTRIBUTIONS TO ZOOLOGY
TABLE 7.—Variations in numbers of wasps seen attacking bugs defending eggs<4 days old in 6 T. catappa trees during one day
Time
P. semiflaviventris7:00-9:00 AM ...1:45-3:00 PM9:00-9:30 PM
T. bodkini7:00-9:00 AM1:45-3:00 PM9:00-9:30 PM
Number of wasps Number of bugs Total number of bugs
seen attacking being attacked seen guarding eggs
306712
481
243211
481
875623
875623
of eggs, P. semiflaviventris females which hadbeen nearby preparatory to attacking (Figure 10a)would often ignore the undefended eggs and moveso as to realign themselves with the bug's new po-sition, sometimes passing very near the now un-guarded eggs (other times the wasps turned andbegan to oviposit in the unguarded eggs or movedaway). On one occasion a female wasp was seenattempting to oviposit (backing in with abdomenextended) below an adult female bug which didnot have any eggs but nevertheless scraped activelyin defense against the wasp.
Female T. bodkini also behaved as if they usedthe bug's presence to locate egg masses. On five dif-ferent occasions I saw a female walking on a trunkwhere there were large numbers of bugs and eachwasp turned to move toward motionless bugs (bothmales and females), approached to within about 1cm and paused for a moment. Then, sometimesafter moving away and approaching again fromanother angle, the wasp moved on. The bugs usu-ally remained quiet, but sometimes walked away orgave small shudders, kicked, or waved an antennatoward the wasp.
A further indication that females of both speciesof wasps hunted defending bugs rather than theireggs was that they were often seen attacking massesof nearly mature or already hatched eggs whichwere unsuitable for parasitization.
Females of P. semiflaviventris and perhaps alsoT. bodkini hunt mainly by day (Table 7); thenumbers of P. semiflaviventris found in the morn-ing and at night are both significantly lower thanthe number found in the afternoon (p<0.01), butthe difference between morning and evening countsis not significant. The T. bodkini numbers showthe same trends, but the differences are not sig-
nificant (0.1>p>0.05). One possible explanationfor these data is that the wasps begin hunting inthe morning and tend to stay near an egg massand battle with the defending bug once one isfound, thus "piling up" around the egg masses asthe day progresses, and then many wasps leave eggmasses in the evening. Most of the P. semiflavi-ventris females seen near eggs in the evening ap-peared not to be attacking. None were backing intoward the eggs with their abdomens extended andsome were facing partially away from the bug, aposition seldom seen during the day. Most hadtheir antennae quiet and directed toward thesubstrate.
EXPERIMENTAL AND NATURAL ISOLATION OF EGGMASSES IN TREES.—Repeated observations of walk-ing female wasps (especially T. bodkini) suggestedthat the wasps might walk while hunting theirhosts. This hypothesis was tested in two ways andboth types of evidence indicated that it was false,that the wasps fly rather than walk as they hunt.Young (<4 days old) egg masses (and the bugs
TABLE 8.—Lack of effect of experimental isolationof egg masses from xoalking insects on parasitismrates (rates from Cassia sp. are from three separateexperiments in the same tree using nearly equalnumbers of control and experimental egg masses ineach case; those from P. dulce are from three adja-cent trees observed simultaneously)
Wasp species
T. bodkini
P. semiflaviventris
Number of egg masses .
Cassia sp.
Isolated
18%44%51
Control
19%45%89
P. dulce
Isolated
12%2%
30
Control
13%1%
29
NUMBER 205 15
defending them) were experimentally isolated fromwalking wasps in both Cassia sp. and P. dulce treesby coating a small area at the base of the branchletbearing the leaf with the egg mass (see arrows inFigure 2) with Tack-Trap. The eggs were col-lected about a week later and the parasitism rateswere determined. Parasitism in isolated groups didnot differ from control rates in other groups of thesame age from the same trees (Table 8).
Cassia sp. branchlets had pinnately arrangedleaves (Figure 2/), and egg masses on leaves nearthe tips were thus naturally more isolated from thebases of the branchlets (where a wasp hunting onfoot would presumably first arrive). Again thedegree of isolation of the egg mass did not affectthe parasitism rate. Of 56 masses which had lessthan the median number of T. bodkini for thetree where they were found, 25 were in the basalhalf of the branchlet and 31 in the distal half; cor-responding numbers for P. semiflaviventris were29 and 36 and neither differed significantly fromthe totals of 57 masses in the basal half and 75 inthe distal half. Nor was there any correlation withthe absolute distance from the base of the branch-let: of 58 masses with more than the median num-ber of T. bodkini, 29 were more than four leavesfrom the base and 29 were on the basal fourleaves; corresponding figures for P. semiflaviventriswere 38 and 26 (total of 57 of 129 masses morethan four leaves from the base).
MOVEMENTS BETWEEN TREES.—The possibilitythat wasps tended to stay in the trees where theyemerged was tested by experimentally removing allor nearly all wasps before they emerged in alter-nate T. catappa trees growing in a row along theside of a small park (distances between the edgesof adjacent trees ranged from 0.9 to 3.1 m). Infour experimental trees I made biweekly collec-tions of all or nearly all egg masses after they weremore than approximately 5 days old (i.e., were nolonger susceptible to further parasitism, but be-fore the wasps emerged from parasitized eggs).Parasitism rates in these trees were monitored byexamining collected eggs (parasitized eggs were dis-tinguished by the membranes deposited next to thechorion by the wasp larvae; see Figure 11), whilethe rates in five control trees were determined fromweekly collections of eggs from which both bugsand wasps had already emerged. The numbers ofwasps emerging on the experimental trees were
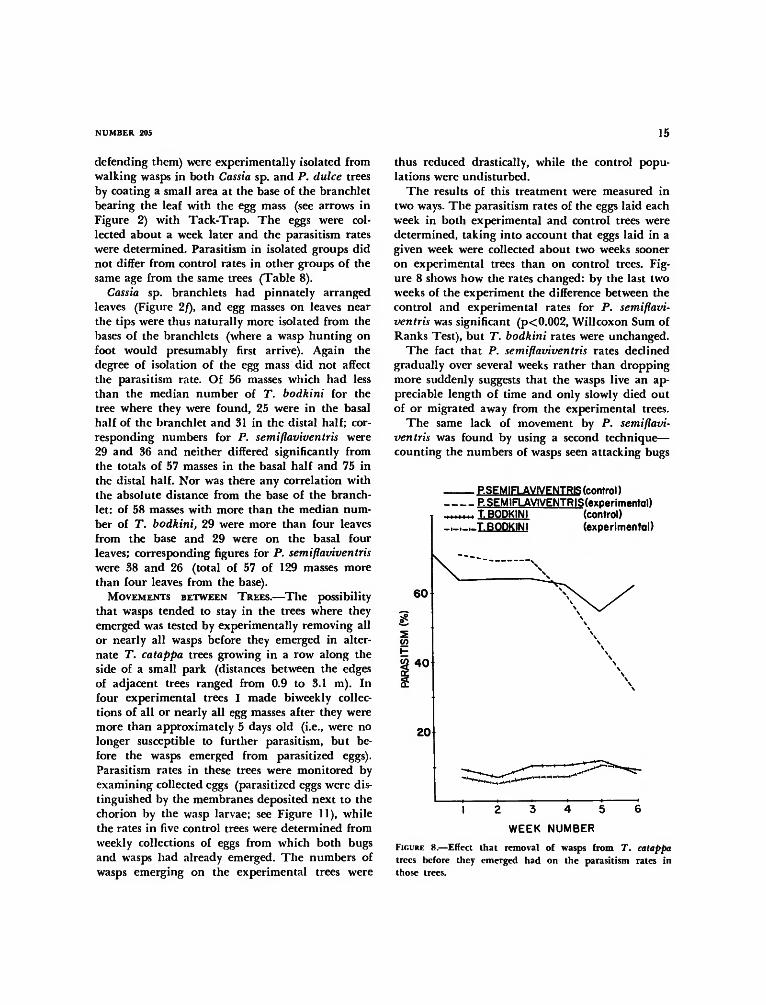
thus reduced drastically, while the control popu-lations were undisturbed.
The results of this treatment were measured intwo ways. The parasitism rates of the eggs laid eachweek in both experimental and control trees weredetermined, taking into account that eggs laid in agiven week were collected about two weeks sooneron experimental trees than on control trees. Fig-ure 8 shows how the rates changed: by the last twoweeks of the experiment the difference between thecontrol and experimental rates for P. semiflavi-ventris was significant (p< 0.002, Willcoxon Sum ofRanks Test), but T. bodkini rates were unchanged.
The fact that P. semiflaviventris rates declinedgradually over several weeks rather than droppingmore suddenly suggests that the wasps live an ap-preciable length of time and only slowly died outof or migrated away from the experimental trees.
The same lack of movement by P. semiflavi-ventris was found by using a second technique—counting the numbers of wasps seen attacking bugs
P.SEMIFLAVIVENTR1S (control)P.SEMIFLAVIVENTRIS(exDerimental)
,,,,,,, T.BODKINI (control)T. BODKIN I (experimental)
60
40
20
1 2 3 4 5 6
WEEK NUMBER
FIGURE 8.—Effect that removal of wasps from T. catappatrees before they emerged had on the parasitism rates inthose trees.
16 SMITHSONIAN CONTRIBUTIONS TO ZOOLOGY
(12)
(39)
(8)
(28) (15)(M)
(65)
(19)(42)
(26)(30)
I 1-2 2-3 3-4 4-5 5-6NUMBER OF DAYS ON TREEBEFORE COLLECTED
6 AND ALREADYHATCHED
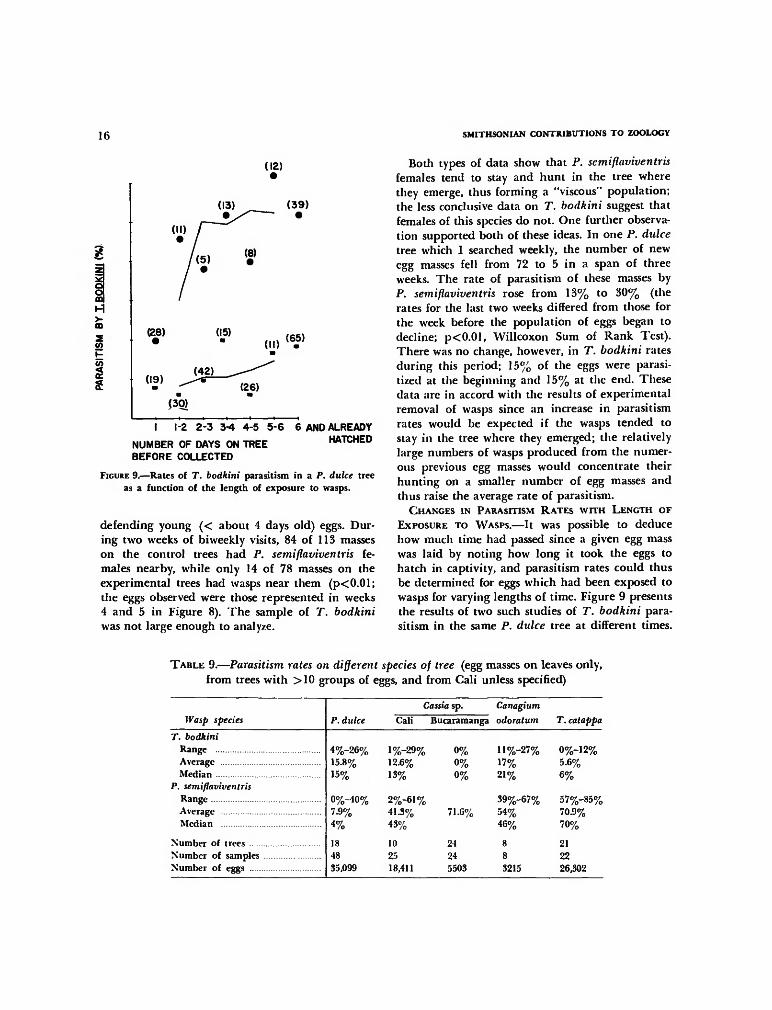
FIGURE 9.—Rates of T. bodkini parasitism in a P. dulce treeas a function of the length of exposure to wasps.
defending young (< about 4 days old) eggs. Dur-ing two weeks of biweekly visits, 84 of 113 masseson the control trees had P. semiflaviventris fe-males nearby, while only 14 of 78 masses on theexperimental trees had wasps near them (p<0.01;the eggs observed were those represented in weeks4 and 5 in Figure 8). The sample of T. bodkiniwas not large enough to analyze.
Both types of data show that P. semiflaviventrisfemales tend to stay and hunt in the tree wherethey emerge, thus forming a "viscous" population;the less conclusive data on T. bodkini suggest thatfemales of this species do not. One further observa-tion supported both of these ideas. In one P. dulcetree which I searched weekly, the number of newegg masses fell from 72 to 5 in a span of threeweeks. The rate of parasitism of these masses byP. semiflaviventris rose from 13% to 30% (therates for the last two weeks differed from those forthe week before the population of eggs began todecline; p<0.01, Willcoxon Sum of Rank Test).There was no change, however, in T. bodkini ratesduring this period; 15% of the eggs were parasi-tized at the beginning and 15% at the end. Thesedata are in accord with the results of experimentalremoval of wasps since an increase in parasitismrates would be expected if the wasps tended tostay in the tree where they emerged; the relativelylarge numbers of wasps produced from the numer-ous previous egg masses would concentrate theirhunting on a smaller number of egg masses andthus raise the average rate of parasitism.
CHANGES IN PARASITISM RATES WITH LENGTH OFEXPOSURE TO WASPS.—It was possible to deducehow much time had passed since a given egg masswas laid by noting how long it took the eggs tohatch in captivity, and parasitism rates could thusbe determined for eggs which had been exposed towasps for varying lengths of time. Figure 9 presentsthe results of two such studies of T. bodkini para-sitism in the same P. dulce tree at different times.
TABLE 9.—Parasitism rates on different species of tree (egg masses on leaves only,from trees with >10 groups of eggs, and from Cali unless specified)
Wasp species
T. bodkiniRangeAverageMedian
P. semiflaviventrisRangeAverageMedian
Number of treesNumber of samplesNumber of eggs
Cassia sp. Canagium
P. dulce Cali Bucaramanga odoratum T. catappa
4%-26% l%-29% 0% H%-27% 0%-12%15.8% 12.6% 0% 17% 5.6%15% 13% 0% 21% 6%
0%-40% 2%-61% 39%-67% 57%-85%7.9% 41.3% 71.6% 54% 70.9%4% 43% 46% 70%
18 10 24 8 2148 25 24 8 2235,099 18,411 5503 3215 26,302

NUMBER 205 17
TABLE 10.—Parasitism rates on branches and leaves of the same trees (trees with>10 groups on both leaves and branches in the case of P. dulce, and >5 in thecase of T. catappa)
Wasp species
T. bodkiniRangeAverageMedian
P. semiflaviventrisRangeAverageMedian
Number of trees ....Number of samplesNumber of eggs ...
P. dulce T. catappaBranches
31%-43%34.9%34%
0%-16%6.5%3 %
8117463
Leaves
15%-28%20.6%24%
0%-37%9.9%4 %
13.313
Branches
0%-17%9.1%7%
2%-62%373%37%
15273845
Leaves
0%-25%4.6%7%
40%-85%71.6%72%
15.735
Although the final rates of parasitism were quitedifferent, in both cases parasitism was essentiallycomplete two days after the eggs were laid.
VARIATION IN DIFFERENT TREE SPECIES.—Table 9shows that the rates of parasitism by P. semiflavi-ventris varied within different ranges in differentspecies of tree. It may be significant that higher P.semiflaviventris rates occurred in trees with largerleaves (see Figure 2). Cocoa, which has large leaves,also apparently has high P. semiflaviventris rates(Callan, 1944a; Sepulveda, 1955). Parasitism ratesby T. bodkini showed less correlation with treespecies, although rates on T. catappa were signi-ficantly lower than those on P. dulce (p<0.01,Willcoxon Sum of Ranks Test). Competition be-tween the two wasp species probably influencedthe rates on different tree species to some extent.
VARIATION AT DIFFERENT SITES IN GIVEN TREES.—Branches vs. Leaves: The parasitism rates by bothspecies of wasp differed for both P. dulce and T.catappa according to whether the bug eggs wereon leaves or branches, as shown in Table 10. TheT. bodkini rate was higher for egg masses onbranches than for those on leaves in both species oftrees, while the P. semiflaviventris rate was con-sistently lower.
Suckers vs. Trees: The rates of parasitism by T.bodkini were consistently lower in groups of eggson the leaves of suckers at the bases of P. dulcetrees (Figure 2a) than on the leaves of the sametrees. When the numbers of eggs parasitized ingroups of 26-28 eggs found on suckers around four
trees were compared with the median numbers ofeggs parasitized by T. bodkini on the leaves of eachtree and its suckers, 18 of the sucker masses werelower than the medians for their respective treesand 3 were higher (p<0.01). In another samplefrom one of these trees, the parasitism of massesfrom the suckers (N = 14) was also lower than thatof masses from the tree (N = 21) (p<0.01, Will-coxon Sum of Ranks Test).
OVIPOSITION
BEHAVIOR.—Oviposition behavior actually formsa part of the attack behavior described below, butis considered separately here in order to simplify thedescriptions. It should be kept in mind that someof the "ovipositions" I saw may have been onlyprobing behavior with the abdomen and/or theovipositor which did not result in eggs being laid.At least some "ovipositions," however, did resultin the production of wasps from otherwise virgin
T. bodkini: When recently laid A. tripterus eggswere placed in a jar with 2-day-old fertilized femaleT. bodkini wasps that had emerged in captivity,the wasps showed no signs of being attracted to theeggs, several times passing very close to them with-out touching them. But as soon as a wasp did en-counter eggs, she began to oviposit into them. Boththese wasps and others found attacking egg massesin nature and induced to oviposit in undefendedgroups showed rather consistent behavior. The

18 SMITHSONIAN CONTRIBUTIONS TO ZOOLOGY
TABLE 11.—Frequency of super parasitism by wasps attacking undefended eggs
Wasp species
T. bodkiniObservedRandom
P. semiflaviventrisObservedRandom
Number of ovipositions in each egg Number of Number of
3 or more egg masses female wasps
27320'.)
131132
185138
6265
1633
2219
17
wasp began by walking over the group, antennatingsuccessive eggs. After pausing and antennating oneegg for a second or more, she made a brisk 180°turn and placed the tip of her abdomen on theegg, then moved forward until the tip was againstthe near edge of this egg (or occasionally againstan adjacent one). She stayed motionless, crouchinglow with her antennae folded down (Figure 1) for10—30 seconds. Wasps in this position usually didnot respond to being pushed or nudged. Then, be-fore leaving, the wasp nearly always wiped the tipof her abdomen from side to side, one or moretimes, across the surface of the egg.
This "marking" behavior was similar to that
observed by F. Wilson (1901) in Asolcus basalisand appeared to have the same function—to pre-vent multiple ovipositions in single eggs. Thenumber of times females which were allowed toparasitize unguarded masses oviposited into eachegg in the mass was determined for 17 masses. Thenumbers of eggs which would have been attacked0, 1, 2, or 3 and more times if the wasps had beenchoosing host eggs randomly was calculated usingthe formula in Wilson's paper (the number of ovi-positions per group varied, so these numbers werecalculated for each egg mass and then summed)and compared with the observed numbers of ovi-positions (Table 11). The wasps reoviposited in
FIGURE 10.—Female P. semiflaviventris wasps attacking bugs defending their eggs: a, wasplooking in toward bug; b, wasp backing in with her abdomen extended; c, wasp ovipositing intoegg in exterior row of mass (arrow); d, wasp ovipositing into egg in interior of mass (arrow).

NUMBER 205 19
eggs less often than if they had chosen eggs at ran-dom, thus indicating that the marking did preventmultiple ovipositions.
Usually females laid several eggs at first, withonly very short pauses between ovipositions, butgradually they spent more and more time walkingover the mass antennating eggs between oviposi-tions and finally, after less than an hour, left. Inmost cases the number of eggs laid in the first burstseemed to depend on the age of the bug eggs,newer groups being more thoroughly attacked.The wasp usually left before parasitizing all theeggs in a mass and usually it flew rather thanwalked away.
On several occasions I saw more than one femaleT. bodkini waiting near a gravid bug or attackinga single egg mass, but in no case was there anysign of aggression between them. In one case inwhich both a T. bodkini and a P. semiflaviventriswere attacking an undefended mass, however, theT. bodkini moved toward the other female severaltimes and appeared to be responsible for her leav-ing the eggs.
P. semiflaviventris: It was more difficult to in-duce P. semiflaviventris to oviposit. Females whichemerged in captivity did not attack groups eitherwith or without bugs defending them. Thosefound attacking masses in nature would only sel-dom lay into undefended eggs. In order to conductstudies of development rate and interspecific com-petition within the egg, it was necessary to glueeggs with known histories (known virgin eggs, forexample) next to a bug whose eggs were beingattacked by a P. semiflaviventris female. By push-ing the bug over onto the other group (which she
then defended), the wasp's attention could bedrawn to them and she would oviposit in them.
Oviposition by P. semiflaviventris differed fromthat of T. bodkini in a number of respects. Ovi-position lasted for only approximately four secondsand was always performed with the abdomen ex-tended (Figure 10c). The female did not climbonto the egg mass, staying on the leaf instead. Shealso failed to antennate eggs before laying in them,moving directly from one to the next with her ex-tended abdomen. Marking behavior was neverperformed. Often a wasp laid a series of five to teneggs, backing along the base of the egg mass andparasitizing both edge and internal eggs; to reachthe latter, she inserted her abdomen between thebases of edge eggs (Figure \0d). The wasp's ex-tended abdomen was very flexible; when she cameto a corner of the egg mass, she curved the tiparound the corner and parasitized the first edge eggon the next side before bringing her body aroundthe corner.
Counts of the numbers of eggs reparasitized ineight different masses confirmed that P. semiflavi-ventris females do not differentiate eggs which theyhave already parasitized from others (Table 11).
Although I observed more than one P. semiflavi-ventris female attacking a single egg mass simul-taneously many times, there was never any sign ofaggression between them.
SUCCESS.—The results of ovipositions by bothwasp species are summarized in Table 12, whichshows that each species can displace the other andthat the winner in any given case is not predictable.Some of the eggs found in nature may have beenpreviously parasitized, but 15 of the eggs into
TABLE 12.—Results of ovipositions by wasps in bug eggs found in nature
Age of eggs
Eggs <4 days old ovipositedin by—
T. bodkiniP. semiflaviventrisBoth species
Eggs >4 days old ovipositedin by—
T. bodkiniP. semiflaviventris
Species
A. tripterus
19%
33%
12%
90%
97%
that emerged
T. bodkini
65%
o%24%
0%
0%
from the egg
P. semiflaviventris
4"'
53%
52%
0 %
0%
Fail to
develop
11%
15%
12%
10%
3 %
Number
of eggs
200
40
42
34
27

20 SMITHSONIAN CONTRIBUTIONS TO ZOOLOGY
which both species oviposited were known to havebeen virgin except for the observed ovipositions;these eggs produced six T. bodkini and nine P.semiflaviventris. Even within a given mass of vir-gin eggs where all ovipositions by each speciesoccurred at the same time, some eggs producedone species of wasp and others the other. The ratesof parasitism for doubly parasitized eggs differfrom those for singly parasitized eggs (p<0.01 forboth species), suggesting some form of competitionbetween the larvae inside the bug egg, with thewinner eating the loser as in other scelionids (e.g.,Kamal, 1937; Pickford, 1964). Older bug eggswere much less susceptible to parasitism (p<0.05for both species). Other scelionids such as Tele-nomns gifuensis also parasitize younger pentatomideggs more successfully than older ones (Hikada,1958).
It was clear, in addition, although no measure-ments were made, that the time between T. bod-kini ovipositions (i.e., the time the wasp spent in-vestigating eggs) was much greater on masses ofolder eggs than on new ones. Both species werefound attacking mostly newer masses in nature.
The first egg laid by T. bodkini females tendedto be a male. Of 24 masses in which only one eggwas parasitized by this species, 20 produced malesand 4 females. Assuming the frequency of males inTable 14 and a random assignment of sex to thefirst egg, these numbers should be 6 and 18; thedifference is significant (p<0.01).
STAGES IN THE EGGS OF A. triptems
Table 13 shows the developmental times of bothwasp species. The membranes referred to were de-posited against the chorion of the bug egg. Themembranes of the two species were distinct; that ofP. semiflaviventris was solid black on the bottomand sides (Figure lla), while that of T. bodkiniwas clear on the bottom and had alternating blackand transparent circular bands like barrel hoopsalong the sides (Figure lib). The membranes stayedintact after the wasps left the eggs, and rates ofparasitism by both wasp species could thus bedetermined by examining empty eggs. Althoughthe most common coloration of the membranes wasblack, both species rarely (especially in T. catappatrees) produced lighter, more brownish mem-branes, which were occasionally only slightlyclouded. Even in these cases, however, species
TABLE 13.—Developmental times of T. bodkini andP. semiflaviventris in days (measured from momentof oviposition by wasp; eggs checked at least onceevery 12 hours)
Stage T. bodkini P. semiflaviventris
Larva with membraneWhite pupaRed-eyed pupaBlack pupaEmergence
3.5- 6.65- 6.8
6.1- 8.275-11.810-12.3
3.8- 7.95.8- 8.96.8-10.27.9-12.79.7-13.4
aFIGURE 11.—Membranes left in bug eggs by: a, P. semiflaviventris; b, T. bodkini.

NUMBER 205 21
TABLE 14.—Sexes of wasps raised from A. tripterus eggs collected in the field(28 of the 49 groups were parasitized by both species of wasp; averages calculatedon the basis of the number of masses parasitized by a given species)
Average number of males/massAverage number of females/massRatio of male:femaleMaximum number of females/massNumber of masses with more than 1 maleNumber of masses with females but no malesRange of number of wasps/massTotal number of massesTotal number of wasps
T.bodkini P.semiflaviventris
identity could nearly always be distinguished bythe circular stripes and the paler bottoms of T.bodkini membranes. The function of the mem-branes and the reason for the variations in theircolor are not known. Species of the scelionid genusAsolcus also produce characteristic membranes(Cumber, 1964).
The wasp larvae did not defecate until they hadfinished feeding, and then deposited a slab of fecesalong one side of the egg. In all cases the feceswere placed ventrally with respect to the wasp pupaand adult, with the wasp nearly always facing thecap of the egg. In only one of several thousandeggs examined was there more than one wasp in asingle bug egg.
EMERGENCE FROM THE EGG
Males developed in less time than females andthe first wasp to emerge from the egg mass was amale in each of 40 cases for T. bodkini and in 44of 45 for P. semiflaviventris. The wasps emergedby cutting away the egg cap with their mandibles.They sometimes opened the egg in less than anhour, but other times paused for up to several hoursbefore completing the task. The two species differedslightly in the form of the cut made in the cap:T. bodkini cut mostly along the edge of the cap,while P. semiflaviventris tended to start in the cen-ter and cut in ever-widening arcs until a sufficientlylarge hole was produced. P. semiflaviventris tendedto emerge more in the night and early morningthan the day; of 214 wasps which emerged in cap-tivity over a period of a week, 149 emerged between7 PM and 9 AM (p<0.01).
1.54.8
1:3.2221331-2539245
SEX
3.113.31:4.3231961-2538527
RATIOS
Males of both species were less common thanfemales, but nearly every egg mass parasitized pro-duced at least one male wasp (Table 14), perhapsdue to tendencies to lay male eggs first. Onlyrarely were males as common as females in a givenmass of eggs (once in each species for the groups inTable 14), indicating, since unfertilized hymenop-teran eggs generally produced males (e.g., E. O.Wilson, 1971), that either unfertilized females wererare or that they do not oviposit.
MALE BEHAVIOR
FIGHTS.—Males of both wasp species fought forpossession of egg masses. Unfortunately, they wereusually disturbed when the leaf which the egg masswas on was placed under a microscope, so the de-scriptions below are only fragmentary.
Males stayed on the tops or at the sides of theegg masses from which they had emerged, walkingover the eggs and grooming periodically, but gen-erally remaining quiet. They could distinguisheggs from other objects, as they almost alwaysclimbed onto eggs and stayed on them when intro-duced into vials containing several egg masses onleaves. In this situation they mounted and stayedon completely hatched masses as well as on onesabout to produce female wasps.
I witnessed both inter- and intraspecific aggres-sive encounters, and the frequency with which morethan one male emerged from a given egg mass(Table 14) indicates that both types of fights arecommon in Cali.
The males' weapons were their mandibles and,

22 SMITHSONIAN CONTRIBUTIONS TO ZOOLOGY
although fights did not usually result in injuries,they were capable of biting off the antennae andlegs of opponents. Aggressive males often openedtheir mandibles as they looked toward other malesand, in at least some cases, it was possible to pre-dict the outcome of an encounter by observingwhich male opened his mandibles more. Less ag-gressive males were not as active as others, oftenkeeping their antennae lowered and still, andcrouching low on the egg mass or next to it.
When one male was much more aggressive thanthe other, interactions did not usually involvephysical contact, the loser running off the egg massas the other moved toward him. In these cases themore aggressive wasp did not pursue the other morethan 1-2 cm from the egg mass. In more evenlymatched pairs, the second wasp turned and loweredhis head toward his opponent's rush or even movedtoward him; the two wasps locked mandibles andapparently struggled, moving from side to side andbackward and forward, with one male finally ap-pearing to dominate, pushing the other backwardoff the egg mass.
MATING.—Mating probably normally occurs themoment the female leaves the egg, and I saw morethan ten matings of this type in each species. As thefemale began to climb out of the egg after cuttingopen the top, the male eased himself onto the topof her thorax. He immediately began to antennatethe front of her head and her antennae, and sheresponded by folding her antennae ventrally andstaying motionless. Then the male moved back-ward and bent his abdomen slightly downward tomake contact with the tip of her abdomen. Oftenthe male moved forward one or more times toreantennate the female's head before achievinggenital coupling. Both animals usually remainednearly motionless during the ten seconds while theywere coupled and then the male dismounted.
There were two small differences in the matingbehavior of the two species. Occasionally P. semi-flaviventris males rubbed their hind legs brisklyalong the dorsal and lateral surfaces of the fe-male's abdomen while antennating her head. Fe-male T. bodkini nearly always walked severalcentimeters away from the egg mass immediatelyafter mating, while P. semiftaviventris females didnot move away.
Males of T. bodkini copulated with virgin fe-males in plastic vials, and one male mated with
four females during his first minute in such a situa-tion. These matings were slightly different andsuggest that mating away from the egg mass is pos-sible in nature. The female remained nearly mo-tionless as the male approached, antennated herbriefly, and mounted her. In one case, the femalemoved away, and the male approached again, moreslowly and with more antennation before mount-ing. Several times a female began to walk as themale mounted on her attempted genital contact,and he responded by moving forward to antennateher antennae until she folded them and becamequiet again, then moved back to attempt genitalcontact.
Males could copulate several times and I sawone T. bodkini male mate with 12 females, andone P. semiftaviventris male mate with 7 females.The females mated only once and T. bodkini fe-males, kept in a vial where they could not escapefrom a male, avoided additional contacts by movingaway when another wasp came near. Males in thissituation approached both virgins and nonvirgins,but usually did not attempt to follow wasps whichmoved away. I saw one male P. semiftaviventrismount immobile nonvirgins at the edge of an eggmass three times; in each case the female stayedmotionless and the male dismounted without at-tempting to copulate.
I also saw two P. semiftaviventris females emergefrom an egg mass patrolled by a T. bodkini male.In both cases the male antennated the female asshe emerged, but did not mount her. In one casethe male did not allow the female to stay near theedge of the egg mass, chasing her away. In groupscollected in nature while the wasps were emerging,there was only a slight, insignificant tendency(0.25<p<0.1) for fewer female P. semiftaviventris(per empty egg parasitized by P. semiftaviventris)to cluster near masses patrolled by T. bodkinimales (N = 13) than near other groups (N=29),indicating that such behavior by T. bodkini malesis far from universal. Cumber (1964) found thatmales of two species of Asolcus would mount fe-males of the other species, but in neither case wasfertilization accomplished.
CLUSTERS OF FEMALES
Females of both species stayed near the egg massfor several hours after emerging and mating, al-

NUMBER 205
though they readily walked and flew away whendisturbed. Female T. bodkini were always dis-persed, each several centimeters from the egg mass;on P. dulce trees they were usually each on a dif-ferent leaf near the leaf with the egg mass. Theavoidance of other wasps by recently mated femalesis probably responsible for this pattern. Females ofP. semiflaviventris, in contrast, usually stayed incontact with the base of the egg mass, each one re-maining immobile amidst the bug nymphs as thefemale bug stood guard over them all.
As noted above, female bugs guarding nymphsscraped less often in response to artificial stimulithan those over eggs (Table 6). I never saw a fe-male bug drive off either female or male waspswhich had emerged from her egg mass. Bugsguarding masses from which wasps were emerginggave no response to immobile wasps and onlywaved their antennae toward moving wasps. Thebugs often raised themselves off the surface of theegg mass when touched ventrally by a wasp walk-ing over the eggs. This response probably serves togive moulting first instar nymphs room to emergefrom their old cuticles after they climb onto theeggs and hook their claws there (females withmoulting nymphs under them were seen in thisposition several times), but it also facilitated themovements of male wasps over the egg mass.
Interactions between Wasps and Bugs
WASPS VS. BUGS GUARDING EGGS
DIRECT OBSERVATIONS.—I observed parts of morethan 20 interactions between female wasps of eachspecies and bugs guarding eggs. Behavior of indi-vidual wasps varied somewhat, but the two specieswere quite distinct.
T. bodkini: Female T. bodkini waited near bugs,staying out of range of the bug's antennae andfacing in toward the eggs. One common tactic wasto rush to the egg mass and turn quickly to lay intoan egg in the edge of the mass. I had the strongimpression that wasps tended to make such rushesjust after I disturbed the bug by seizing the leafwith my hand; on several such occasions the bug,apparently distracted momentarily, failed to re-spond to the wasp. Another tactic was to waitseveral minutes on the other side of the leaf, thencome over the edge and quickly rush to the egg
mass. Usually the bug responded to rushes byscraping, kicking, waving her antennae, and oc-casionally shuddering as described above. Oftenthis behavior served to drive the wasp out of rangeof the bug's antennae and bugs occasionally suc-ceeded in knocking wasps completely away fromthe egg mass with a kick.
On several occasions a wasp appeared to hurryher oviposition in a way never seen in attacks onundefended eggs, omitting antennal inspection ofthe bug egg, and simply moving her abdomendirectly from one egg to the next.
The bugs were unable to effectively defend therear edge of the egg mass, i.e., the edge under thebug's abdomen (Figure 1, arrow in Figure 5a).They apparently could not bend their hind legs soas to scrape along that edge, and thus a wasp whichwas able to insinuate herself into this area was usu-ally able to parasitize several eggs along the backrow with little interference from the guarding bug.After ovipositing in the back row, T. bodkini fe-males often attempted to move stealthily along thelateral edge of the group. Usually, however, theseattempts were unsuccessful, as the bug respondedby kicking at the wasp with a hind leg or scraping.
It is interesting that even when a bug gave clearindications (kicks, shudders, etc.) that she sensedthat there was a wasp at the rear edge of her eggmass, she never turned to assume a new orienta-tion over the eggs. That is, the bugs never turnedso as to bring their front legs to bear on the un-protected back side of the mass. To do so of coursewould result in leaving another side of the massopen to attack, and the lack of turning implies asacrifice of the back eggs in order to insure therelative safety of the others. This failure to turnwas probably an important factor in keeping T.bodkini rates at relatively low levels in somesituations.
P. semiflaviventris: Attacks by P. semiflavi-ventris females were quite different. The generaltactic of this species was to move to about a bodylength from the edge of the egg mass (generallywithin range of the bug's antennae), quickly turn180°, and then extend the abdomen to abouttwice its normal length and back in slowly towardthe mass, attempting to contact an egg with the tipof her abdomen (Figures 1, 106, 10c). Upon reach-ing the mass, the wasp laid in the first bug egg shetouched, then moved to lay in an adjacent egg in

24 SMITHSONIAN CONTRIBUTIONS TO ZOOLOGY
the edge of the mass or to thread her abdomen be-tween two edge eggs and parasitize an interior egg(Figure lOd). Wasps attacked from all sides of thebug, not just the rear. This species also seemed toattack especially often just after I disturbed thebug.
DEDUCTIONS FROM PATTERNS OF PARASITISM WITH-IN EGG MASSES AND WITHIN TREES.—The patterns ofparasitism in empty (hatched) egg masses with thecommon "compact" shape (Figure 12) confirmedthe direct behavioral observations of wasp-buginteractions. Since defending female bugs nearlyalways positioned themselves over such groups in astandard way, detailed analyses of the directionsfrom which the wasps attacked could be made.
For instance, the eggs in position C in Figure126 formed the rear, more poorly protected edge
of the mass, and eggs in these positions were para-sitized by T. bodkini approximately four timesmore heavily than those in any other part of themass. Parasitism by P. semiflaviventris on theother hand was not confined to the rear edge, butwas distributed on all sides of the group. The cen-tral eggs were seldom parasitized by either species.
The patterns of parasitism of these groups alsobrought out several more subtle details of theinteractions between P. semiflaviventris and thedefending bugs. Excepting the back edge, edgeeggs (A, B, and D in Figure 126) were generallymore heavily attacked than eggs on the first inte-rior row (E, F, and G, but not H—see below)(p<0.05 excluding H). Since attacking wasps ar-rived first at edge eggs and only later at interioreggs, the difference between the two positions
aFIGURE 12.—Pattern of parasitism of "compact" egg masses: a, parasitism rate for each position(hatched = T. bodkini, black = P. semiflaviventris, white = no parasitism); b, positions groupedaccording to similarities in parasitism rates.
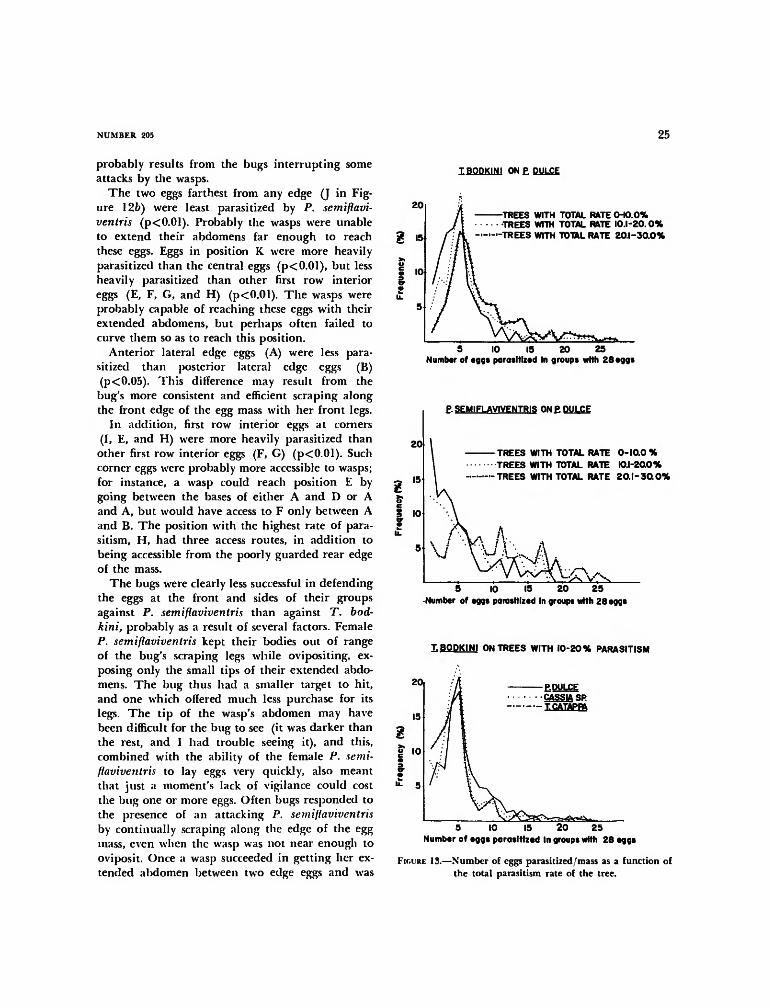
NUMBER 205 25
probably results from the bugs interrupting someattacks by the wasps.
The two eggs farthest from any edge (J in Fig-ure 12b) were least parasitized by P. semiflavi-ventris (p<0.01). Probably the wasps were unableto extend their abdomens far enough to reachthese eggs. Eggs in position K were more heavilyparasitized than the central eggs (p<0.01), but lessheavily parasitized than other first row interioreggs (E, F, G, and H) (p<0.01). The wasps wereprobably capable of reaching these eggs with theirextended abdomens, but perhaps often failed tocurve them so as to reach this position.
Anterior lateral edge eggs (A) were less para-sitized than posterior lateral edge eggs (B)(p<0.05). This difference may result from thebug's more consistent and efficient scraping alongthe front edge of the egg mass with her front legs.
In addition, first row interior eggs at corners(I, E, and H) were more heavily parasitized thanother first row interior eggs (F, G) (p<0.01). Suchcorner eggs were probably more accessible to wasps;for instance, a wasp could reach position E bygoing between the bases of either A and D or Aand A, but would have access to F only between Aand B. The position with the highest rate of para-sitism, H, had three access routes, in addition tobeing accessible from the poorly guarded rear edgeof the mass.
The bugs were clearly less successful in defendingthe eggs at the front and sides of their groupsagainst P. semiflaviventris than against T. bod-kini, probably as a result of several factors. FemaleP. semiflaviventris kept their bodies out of rangeof the bug's scraping legs while ovipositing, ex-posing only the small tips of their extended abdo-mens. The bug thus had a smaller target to hit,and one which offered much less purchase for itslegs. The tip of the wasp's abdomen may havebeen difficult for the bug to see (it was darker thanthe rest, and I had trouble seeing it), and this,combined with the ability of the female P. semi-flaviventris to lay eggs very quickly, also meantthat just a moment's lack of vigilance could costthe bug one or more eggs. Often bugs responded tothe presence of an attacking P. semiflaviventrisby continually scraping along the edge of the eggmass, even when the wasp was not near enough tooviposit. Once a wasp succeeded in getting her ex-tended abdomen between two edge eggs and was
TBODKINI ON £ DJiLCE
20
io
-TREES WITH TOTAL RATE OHO.0%TREES WITH TOTAL RATE 10.1-20.0%
—•—TREES WITH TOTAL RATE 20.1-30.0%
5 10 15 20 25Number of eggs parasitized in groups with 28 eggs
E.SEMIFLAVIVENTRIS 0 N £ DULCE
20
10
-TREES WITH TOTAL RATE 0-10.0%TREES WITH TOTAL RATE 10.1-20.0%TREES WITH TOTAL RATE 20.1-30.0%
5 10 15 20 25Number of eggs parasitized in groups with 28 eggs
T.BODKINI ON TREES WITH 10-20% PARASITISM
-P. DULCECASSIA SPTCATAPPA
20
15
£ 10I
S 10 15 20 25Number of eggs parasitized in groups with 28 eggs
FK.URE 13.—Number of eggs parasitized/mass as a function ofthe total parasitism rate of the tree.

26 SMITHSONIAN CONTRIBUTIONS TO ZOOLOGY
ovipositing in an interior egg, she was well bracedby the eggs and not easily moved; on several oc-casions I saw a bug scrape right over a wasp in thisposition without dislodging it.
Several additional details of bug-wasp inter-actions can be deduced from Figure 13, whichshows how the numbers of eggs parasitized inmasses of 28 eggs varied according to the totalparasitism rate in P. dulce trees. The total rate fora tree is probably a fair indication of the density ofwasps which were searching for eggs there and thuswhat is said below concerning total parasitismrates probably also applies to the density of search-ing wasps in a tree.
In both wasp species the number of egg massesmissed declined as the total parasitism rate for thetree rose (it should be kept in mind that my as-sumption that unparasitized egg masses were neverfound by wasps was undoubtedly incorrect in somecases in which the bug succeeded in repelling awasp before it laid a single egg; such interactionsmay have been more common in the case of P.semiflaviventris); at the same time, the number ofeggs parasitized per group rose for both species.The curves for T. bodkini show strong peaks inthe region of five eggs, almost certainly due to the
fact that the back edges of egg masses, the usualsite of T. bodkini parasitism, usually consisted ofabout five eggs; the curves for P. semiflaviventrisshow no such tendency.
The low numbers of eggs per group parasitizedby P. semiflaviventris in trees with low total ratesare surprising, indicating that the defense of thebugs against single attacks by P. semiflaviventriswas quite effective, perhaps more so than thatagainst single attacks by T. bodkini females. Thehigher rates of parasitism which P. semiflaviventrisachieves on some trees would thus seem to be dueto multiple attacks, a conclusion supported by theshift to the right of the P. semiflaviventris curvesin trees with higher total rates. The curves for T.bodkini also move to the right, but more slowly,probably because each subsequent female wasptends to attack the same five eggs in the rear edgeof the egg mass; P. semiflaviventris females, notbeing so restricted, probably attacked more effi-ciently, reparasitizing eggs much less often.
It was not uncommon to see more than one fe-male wasp attacking a single bug (up to 10 femaleP. semiflaviventris were seen around a single bugin T. catappa trees), and it seems possible that asort of "social facilitation" may occur in such situ-
TABLE 15.—Patterns of parasitism of "compact" egg masses by P. semiflaviventrisin different species of tree (the numbers are the number of wasps from the givenposition divided by the total number of wasps from eggs in that tree species notcounting those from position C; the positions are those indicated in Figure 126)
Position of egg P. dulce Cassia sp.Canagiumodora turn
Cali Bucaramanga
T. catappa
AB*CDEFGH»IJKTotal wasps (not counting those
from position C)Total number of egg massesTotal parasitism by T. bodkini
.20
.28
.23
.12
.08
.07
.08
.17.06.00.03
20887
.19
.22
.30
.16
.08
.05
.07
.16
.04.01.03
54573
.22
.21
.23
.17
.08
.07
.08
.11
.0.")
.01.03
1297
86
0%
.19
.22
.12
.15
.10
.07
.09
.11
.05.00.02
6825020%
.21-21.21.16.07.09.08.11.04.01.02
77350
* Difference between P. dulce and T. catappa significant (p <.O5).

NUMBER 205 27
ations, with a given wasp's presence aiding otherattacking females by distracting the bug's atten-tion.
EFFECT OF TREE SPECIES.—Battles between bugsand P. semiftaviventris wasps for "compact" eggmasses were only slightly influenced by the speciesof tree where they occurred. Since the rates of para-sitism by T. bodkini differed in different tree spe-cies, the eggs at the back edges of the masses(position C in Figure 12), where the great major-ity of T. bodkini parasitism occurred, were ex-cluded from the analysis to avoid the effects ofinterspecific competition. The percentage contri-bution from each position in the mass was calcu-lated by placing the total parasitism in a givenposition over the total number of P. semiflavi-ventris produced in compact masses in that par-ticular tree species minus those from position C.When the resulting patterns of parasitism in dif-ferent tree species are compared as in Table 15, itis clear that the general pattern was very similar.Two differences are significant, however; the num-bers of wasps from the two posterior lateral posi-tions B and H are higher in P. dulce than in theother trees. These differences may stem from awasp's being able to "surprise" the bug more easilyon small leaves by hiding for several minutes onthe other side of the leaf, then peeking over theedge and quickly beginning to lay before the bugsees her. This tactic might be more effective onsmaller leaves and in the rear part of the egg masswhere the bug's antennae and eyes would be lesslikely to sense the wasp.
A detailed analysis of this type was not possiblein the case of the T. bodkini, but Figure 13c indi-cates that the tree species also failed to substan-tially modify the bug's interactions with this speciesof wasp.
EFFECT OF SHAPE OF EGG MASS.—The effect of theshape of an egg mass on its susceptibility to waspattack was determined by counting the numbers ofeggs parasitized in already vacated egg masses ofdifferent shapes. Of 209 "compact" masses from 17P. dulce trees, 124 had fewer than the mediannumber of T. bodkini wasps for masses of 28 eggsin their respective trees (p<0.01, all trees had amedian parasitism greater than 0). Presumablythis reduction in parasitism resulted from the bugsbeing better able to defend a smooth-sided, com-pact group than more irregular, strung-out masses
which had more "edge" eggs and more nooks toprotect attacking wasps from the bug's feet. Theform of the mass did not have a significant effecton parasitism by P. semiflaviventris, however.
WASPS VS. OVIPOSITING BUGS
DIRECT OBSERVATIONS.—On three occasions Ifound wasps (three T. bodkini and one P. semi-ftaviventris) parasitizing eggs while the bug wasstill ovipositing. Except for kicking at the waspwhen it touched her, and perhaps moving fartherback over the eggs between ovipositions, none ofthe bugs appeared to modify her oviposition be-havior. The bugs were apparently unable to see thewasps (which were always behind them) and intwo cases did not appear to sense the wasp'spresence during the early part of the interaction.The kicks of the bugs at the wasps were essentiallyblind, evidently directed mainly by contact. Forinstance, on one occasion the bug moved forwardto lay an egg, then back, and kicked at the sidewhere she had touched a wasp just before movingforward, but meanwhile the wasp had changedsides; later the bug also kicked several times moreafter she succeeded in kicking the wasp clear of theeggs. Although the bugs finally kicked away all fourwasps before finishing ovipositing, their defensivebehavior was poorly organized; several times a bugbrushed or kicked an ovipositing wasp repeatedlyand, failing to dislodge it, ceased responding to it.
The behavior of the two species of wasp wasquite distinct in this situation. The P. semiflavi-ventris female behaved very much as if attacking agroup with a female bug in normal defensive po-sition; she stayed on the leaf, backed in and probedwith her extended abdomen, and laid several eggsin succession without inspecting them with herantennae. She showed no response to the bug'speriodic moves forward and back over the eggmass.
The behavior of the T. bodkini females differedfrom both that of P. semiftaviventris and from thatof T. bodkini females attacking eggs with a bug innormal guarding position. They stayed on the topsrather than the sides of the eggs, and moved overthem antennating the cap of each. All three waspsappeared to try to avoid touching the bug. Al-though each wasp spent the majority of her timeon the rearmost eggs (which were not covered by

28 SMITHSONIAN CONTRIBUTIONS TO ZOOLOGY
the bug's abdomen), she did not oviposit there—presumably because she had already parasitizedthose eggs. When the bug moved forward to layanother egg, the wasp also moved to the front partof the group, inspected, and then turned and ovi-posited in an egg there. Since the bug laid an eggin less time than it took T. bodkini to oviposit, thebug sometimes moved back over the mass while awasp was still ovipositing. The bugs often sensedthe wasps at this time, and two of the wasps werekicked clear of the eggs after being discovered atthis stage. In other cases, the wasp crouched lowand stayed very still under the bug's thorax, onlymoving again when the bug moved forward to layits next egg. On one occasion I was sure that thewasp marked the egg it had just parasitized, but onseveral other occasions the marking was either verylimited or did not occur. One of the T. bodkinifemales, kicked clear of the eggs by the bug, landedon my hand holding the branch, and after cleaningherself briefly, she flew away.
DEDUCTIONS FROM PATTERNS OF PARASITISM WITHIN
EGG MASSES.—Attacks made during the bug's ovi-position may explain one additional pattern ofparasitism of the "compact" group form. The in-terior eggs in the anterior part of the mass (posi-tions E, F, and I) were more heavily parasitized byT. bodkini than those in the posterior part (G, J,and K) (p<0.05, H is omitted since the high rateof parasitism by P. semiflaviventris may influencethe T. bodkini rate). Interior eggs are probablynearly inaccessible for T. bodkini once the bugfinishes ovipositing, and the eggs in the front partof the group are laid first (e.g., Figure 4) and arethus both exposed longer to attacks during ovi-
position and more poorly defended by the ovi-positing bug.
Competition between Species of Wasps
WITHIN BUG EGGS
Comparisons of the patterns of P. semiflavi-ventris parasitism of "compact" groups from siteswith different rates of T. bodkini parasitism showed(assuming that wasp-bug battles were essentiallythe same in all sites—see Table 15) that the twowasp species compete for eggs. The sites chosen fortheir different T. bodkini parasitism rates werethese: Cassia sp. trees in Bucaramanga (T. bodkiniabsent), P. dulce tree leaves in Cali (7\ bodkiniin moderate numbers), and P. dulce branches inCali (T. bodkini in high numbers). In all cases,the parasitism by T. bodkini was concentrated inthe rear edge of the egg mass (position C in Figure126). As shown in Table 16, parasitism by P. semi-flaviventris in these rear eggs was significantly re-duced where T. bodkini was more common.
The complementary comparison of patterns ofT. bodkini parasitism of "compact" masses in treespecies with high and low P. semiflaviventris rates(T. catappa vs. P. dulce) showed no difference.This probably results from the tendency notedabove for P. semiflaviventris to attack all sides ofthe mass more or less equally, and in turn indicatesthat the lower rates of P. semiflaviventris in reareggs on T. catappa and Cassia sp. (Figure 12a)probably also resulted from competition with T.bodkini.
In general, these data show conclusively that T.
TABLE 16.—Influence of interspecific competition on patterns of parasitism in"compact" egg masses (all samples from Cali, Colombia, unless otherwise indicated)
Wasp species
P. semiflaviventrisTotal parasitism ratePercent of total that came from
position CT. bodkini
Total parasitism rateRate of parasitism of eggs in
position CNumber of masses
•• b Pairs differ significantly (p<.01)
Cassia sp. in T. catappa
Bucaramanga
P. dulce trees
Leaves Branches
70%
23%»
0%
0%86
68%
17%-
9%
32%50
11%
18%'
15%
54%87
6%
9%'
41%
90%69

NUMBER 205 29
bodkini sometimes displaces P. semiflavivenlris,but not vice versa. The fact that neither speciesdominated consistently over the other when bothlaid into the same, otherwise virgin, eggs indicates,however, that each species sometimes displaces theother in nature.
FIGHTS BETWEEN MALES
Since the first wasps to emerge were nearlyalways males, it was possible to deduce the resultsof interspecific fights which had occurred in nature.In masses from which at least one wasp of bothspecies had emerged, but from which some wasps ofboth species had yet to emerge when the mass wascollected (all wasps and eggs were immediatelykilled with alcohol to prevent additional emergalsbefore I counted them), the numbers of males ofboth species were significantly reduced (p<0.01,Table 17). Thus, males of each species significantlydisplaced those of the other.
It appears that T. bodkini males may be supe-rior fighters. The total rates for those egg massesparasitized by both species of wasp were 18% T.bodkini and 45% P. semiflavivenhis. Assumingthe sex ratio in Table 5, this would imply an aver-age of 1.3 T. bodkini males/egg mass, and 2.4 P.semiflaviventris males/egg mass. If the two spe-cies were equally good fighters, the results of the20 cases in which only one male was left on the eggmass (Table 17) would have been 7 T. bodkiniand 13 P. semiflaviventris, rather than the ob-served 11 and 9 (0.05<p<0.1).
In addition, P. semiflaviventris males were moretolerant of the presence of conspecific males thanwere T. bodkini males. Of 48 masses which had T.bodkini males, only 4 had more than one, while9 of 33 masses had more than one P. semiflavi-ventris male (p<0.05). On several occasions I saw
a P. semiflaviventris male avoid an attack by anadvancing conspecific by behaving like a female,lowering his antennae to the substrate, and re-maining motionless. Such "feigning" behaviormight be more effective in P. semiflaviventristhan in T. bodkini since P. semiflaviventris fe-males usually stay clustered around egg massesafter emerging while T. bodkini females disperse.
There was no indication that males search outegg masses other than those where they emerge, asmales were never found on masses from which atleast one wasp of that species had not emerged(Table 17). Thus the exclusion of males of a given
species by the other eliminated the chances of fe-males of that species to mate near that egg mass,although the possibility remains that they mateaway from the eggs.
Experimental Removal of Bugs Guarding Eggs
Both the patterns of parasitism of egg masses bythe wasps and my repeated observations of bugsrepelling attacking wasps show that the defensivebehavior of the bugs lowers the number of eggsparasitized by wasp females which have found theegg mass. The observations reported in the sectionson the oviposition and hunting behavior of thewasps suggest, however, that both species, and espe-cially P. semiflaviventris, use the presence of guard-ing bugs to locate egg masses. The possibility thusarises that the net influence of the guarding bugon her eggs is negative—that her betrayal of theirposition by standing over them outweighs thebenefits she confers on them with her defensivebehavior.
This possibility was tested by removing bugsfrom some young egg masses (eggs without visibleembryonic eyes and thus less than about four daysold and still susceptible to parasitism), and leaving
TABLE 17.—Male wasps present on egg masses from which some but not all ofthe xoasps had emerged
Species
Masses from whichhad emerged
Masses from whichemerged
Masses from whichhad emerged
emerged
males of both species
only T. bodkini had
only P. semiflaviventris
T. bodkini
11
30
0
P. semiflaviventris
<)
0
38
Both
8
0
0
None
o
1
3
Total
30
31
41

30 SMITHSONIAN CONTRIBUTIONS TO ZOOLOGY
TABLE 18.—Results of removal of femaleA. tripterus guarding eggs on antless trees
Wasp species
Parasitism by T. bodkiniControlWithout female
Parasitism by P. semiflaviventrisControlWithout female
Number of control egg massesNumber of experimental egg masses
T. catappa Cassia sp.
4.5%72%
64.4%
2722
11.8%18.1%
61.4%46.4% *2339
• Experimental differs from controlcoxon's "Sum of Ranks Test").
(p <.O5, using Will-
or less excluding ants from one T. catappa treeand one Cassia sp. tree (twigs harboring Pseudo-myrmex colonies were removed and a ring of"Tack Trap" was laid around the trunk to preventother species from climbing into the tree) andthen repeating the experiment just described.
The results, presented in Table 18, show that thenet effect of the presence of the bugs is indeednegative; it aids rather than hinders P. semiflavi-ventris and has no significant effect on T. bodkini.Thus the bugs were "wasting their time" with re-spect to botli species of wasp, since they could havebeen investing it in feeding to produce more eggs.
the females guarding other similar masses in thesame trees to serve as controls. Both test and con-trol groups were identified by marks placed on ad-jacent leaves, and parasitism rates were determined1-2 weeks later. The first test of this kind was con-ducted in two P. dulce trees, and a total of 48 testand 41 control groups were recovered. Nearly allof the experimental egg masses had vanished, how-ever, when I recovered the leaves; in most casesthere was nothing but a greasy stain remaining onthe leaf. Of those few eggs recovered, not one pro-duced a bug (most failed to develop). I observedseveral species of insect feeding on undefended eggsin this species of tree (Table 22), and they (per-haps especially the ants) were probably responsiblefor the losses.
These results show that the defense of the bugsagainst generalized egg predators is highly effectiveand that the overall effect of their defense is un-doubtedly positive. The possibility remained, how-ever, that the effect of the bugs with respect to theparasitic wasps is negative. This was tested by more
Other Factors Affecting A. tripterus Survival
UNHATCHED EGGS
Often some eggs in a mass failed to produceeither bugs or wasps. In some cases it was possibleto determine whether a young bug or a wasp haddied before emerging; in others, where it was notpossible, the egg was termed "undeveloped." Somesuch eggs may have been infertile, while othersprobably contained dead bug embryos or parasiteswhich died before discoloring the egg's vitellinemembrane. The frequencies of both failures ofidentifiable bugs and wasps to emerge and of un-developed eggs were correlated with the site of theegg mass.
DIFFERENCES BETWEEN SPECIES OF TREE.—Eggs
laid during the same four-week period failed to de-velop more often on the leaves of a P. dulce treethan on those of two nearby Cassia sp. trees (Table19). The frequency of undeveloped eggs was alsocorrelated with tree species when data taken from
TABLE 19.—Frequencies of deaths within the bug eggs on leaves of different treespecies during the same four-week period (A) and during various periods (B)
Died within the egg (A)BugsT. bodkiniP. semiflaviventris"Undeveloped"
Frequencies of"undeveloped" (B) ...
P. dulce
5.2% (N = 3262)19.7% (N = 761)42.2% (N =491)
6% (N=4777)
4.9% (35,099 eggs,18 trees)
Cassia sp.
.9%(N = 1063)
.9%(N = 327)2.7% (N= 1251)1% (N=2778)
1.2% (18,411 eggs,10 trees)
Canagiumodor alum
2.1% (3,215 eggs,8 trees)
T. catappa
2.7% (26,302 eggs21 trees)

NUMBER 205 31
various trees at different periods were compared(Table 9). Hatching was consistently poorer onP. dulce than any other species of tree, but thelevel of significance cannot be determined since theeffects of parasitism on the production of "undevel-oped" eggs are unknown.
DIFFERENCES BETWEEN LEAVES AND BRANCHES.—InP. dulce trees, eggs laid on branches failed to de-velop more frequently than those on leaves. Thus,in 12 samples from 14 trees, the frequency ofundeveloped eggs in branch egg masses was higherthan that in leaf masses in 13 cases, lower in 7, andequal in 3 (p<0.1), and the total frequencies ofundeveloped eggs were 7.7% on branches (N =7463) and 4.3% on the leaves (N = 13,313) ofthese trees. Data from eggs on T. catappa trees didnot follow this pattern: 2.4% of the eggs onbranches (N = 3845) and 3.2% of those on leaves(N = 15,735) of six trees were undeveloped. Inthis case at least, the differences in developmentalfailures are not due to differences in parasitism,since the differences in parasitism rates on leavesand branches of both tree species are similar(Table 10).
LEAVES AND EGG MASSES THAT FELL FROM TREES
Substantial losses of entire egg masses were ob-served during weekly checks in several trees. Eachleaf which had an egg mass on it was marked (inthe case of P. dulce, an adjacent leaf was usuallymarked), and the mass was collected after all bugsand wasps had left it. In a span of five weeks, 290leaves marked on three P. dulce trees were re-covered; 31% had lost their eggs. On two Cassiasp. trees, 16% of 145 marked leaves recovered dur-ing the same period were without eggs, indicatingthat this type of egg loss was less common(p<0.01) in Cassia sp.
These figures show a clear difference betweenthe tree species, but they are only approximationsof the true rates of loss because of several compli-cations in taking the data: (1) marked leaves lack-ing eggs were probably less easily noticed, andtherefore underrepresented; (2) some egg massesprobably fell after the last bugs and wasps left, butbefore I returned to search the tree—these losseswould be biologically insignificant; and (3) in thecase of P. dulce, the entire leaf rather than just theegg mass may have fallen in some instances, leavingonly the adjacent marked leaf. (Repeated observa-tions of new eggs on yellow, ready-to-fall leaves onP. dulce and Cassia sp. trees [these trees continu-ally shed a few scattered leaves] suggest that thebugs did not distinguish such unsatisfactory sitesfrom others.) In balance, the data on losses by fall-ing are probably somewhat below the true rates.
One other factor appeared to influence the lossof egg masses by falling. In all four study species oftree (and especially in P. dulce), a greater pro-portion of the older masses were on branches thanon leaves (Table 20; guarded masses differ fromunguarded masses, p<0.01) indicating that eggsstayed stuck to branches longer before falling. Eggmasses also -appeared to stay glued to T. catappaand Canagium odoratum leaves longer than to P.dulce and Cassia sp. leaves, perhaps due to thegreater mechanical protection afforded by thelarger size of the leaves (Figure 2).
LOSSES OF FIRST INSTAR NYMPHS
VARIATION WITH SITE.—The numbers of first in-star nymphs lost from groups at different sites weredetermined in two ways. Direct measurements oflosses during the entire first instar period were de-termined in cases where the nymphs were foundjust moulting to the second instar by comparing
TABLE 20.—Distribution of young and old egg masses on branches and leaves ofP. dulce trees (total of 18 samples from 16 trees; the ages of unguarded masseswere judged by the accumulations of dirt and fungus on them)
Ages of eggs
Guarded egg massesUnguarded and empty.but relatively new eggs ..Old, weathered, empty eggs
Percent of masses found on— Total number of
Branches Leaves egg masses
18 82 83251 49 33458 42 283

32 SMITHSONIAN CONTRIBUTIONS TO ZOOLOGY
for each mass the numbers of eggs that had pro-duced bugs with the numbers of nymphs present;indirect measures were obtained by comparing thenumbers of hatched eggs and the numbers of firstinstar nymphs and then (assuming that on theaverage these nymphs were halfway through thefirst instar and that the rate of loss throughout theinstar was uniform) multiplying the losses by 2.The results of both types of counts are presentedin Table 21. It should be kept in mind that "losses"did not necessarily mean deaths of nymphs, butonly that the nymphs were no longer near the eggmass. The general low activity of the first instarnymphs suggests that vanished nymphs had notwalked away, but had either fallen or been eaten.Falling nymphs clung readily to surfaces whichthey struck but, even so, the chances of survival ofthe nymphs probably decreased greatly if they fell.
There were again significant differences in lossesfrom different species of tree and from differentsites within a given species (Table 21). The di-rectly measured rate of loss from P. didce leaveswas not quite significantly greater than that fromCassia sp. leaves (0.06>p>0.05), but was greaterthan that from T. catappa leaves (p<0.05). Theindirectly measured rate of loss from branches wasgreater (p<0.01) than that from leaves in P. dulce.Although none of the differences between directand indirect measurements were large enough tobe significant, the consistent tendency for indirectmeasurements to be larger suggests that the as-sumption that the rate of loss is constant through-out the instar may be incorrect and that youngernymphs may be more frequently lost.
It seemed that mechanical agitation, whichwould be greater on smaller-leafed trees such asP. dulce, might be a principal cause of loss, butcomparisons of losses (determined indirectly) onone P. dulce just after a period of heavy daily rainsand after periods of milder weather were not sig-nificantly different. First instar nymphs (as well asolder ones) were capable of clinging very stronglyto a substrate; when I tried to lift nymphs awayfrom their egg masses in captivity, some individualsclung to and suspended groups of eggs and nymphscorresponding to up to ol) times their own weight.
EXPERIMENTAL REMOVAL OF FEMALES GUARDINGFIRST INSTAR NYMPHS.—The effects of the bugs'defense of first instar nymphs were measured byremoving defending females. The number ofnymphs was counted when the female was removedand again two days later, and the total numberwhich would have been lost in seven days, the ap-proximate duration of the first instar, was thencalculated by multiplying the observed losses by3.5. The results of these observations (Table 21)were that losses rose significantly (p<0.01, calcu-lated on the basis of losses/day) on both P. dulceand Cassia sp. when the female was removed. Thedefense of the females, at least in P. dulce trees,had the effect of especially reducing the likelihoodof losses of large numbers of nymphs from anygiven group. Thus, of 39 unguarded groups on P.dulce which lost nymphs during the 48-hour ex-perimental period, 13 or 33% lost three or morenymphs; of 108 control groups which had lostnymphs (the average age of these nymphs was as-sumed to be 3.5 days), only 24% had lost three or
TABLE 21.—Losses of first instar A. tripterus nymphs from guarded and un-guarded groups in 7 days at different sites (methods used in arriving at thesenumbers are explained in the text; numbers in parentheses are number of eggmasses and total number of nymphs hatched)
Nymphs lostMeasured
Measured
Nymphs lostgroups
from defended groupsdirectly
indirectly
from undefended
P. dulce
Leaves
6.3%(41,808)11.7%(245,4802)
44.1%(51,1078)
Branches
32.2%(56, 726)
Cassia sp.
Leaves
3.4%(23,353)9.3%(146,1969)
40.2%(22, 332)
T. catappa
Leaves Branches
2%(10,99)3.5% 8%(79,665) (20,170)

NUMBER 205 33
more nymphs (p<0.05 after adjusting for differenttime spans).
reared from both adults and large nymphs of A.tripterus.
OTHER ENEMIES OF A. tripterus
The animals observed preying on various stagesof A. tripterus in nature and captivity are listed inTable 22. In addition to the species listed, a fungusof the genus Penicillium attacked and killed bothnymphs and adults in Palmira and Cali (Eberhard,in press). A parasitic wasp, Hexacladia sp. n. (?)(Encyrtidae) (det. C. M. Yoshimoto), was also
Discussion
HUNTING BEHAVIOR OF THE WASPS
Precise descriptions of the searching behavior ofparasitic insects are of great interest, but are rare(DeBach and Hagen, 1964). Several aspects of thehunting behavior of the two wasps are clear fromthe data of this study.
TABLE 22.—Other enemies of A. tripterus (all attacks on eggs and first instarnymphs occurred in unguarded groups unless otherwise noted; e=egg, n1=firstinstar nymph, n = other nymphs, a=adults, P.d.=Pithecelobium dulce, C.sp.=Cassia sp., T.c.= Terminalia catappa)
Predators
HymenopteraSphecidae
Bicrytes variegataVespidae
Polistes canadensisFonnicidae
Ectatomma ruidumCrematogaster spPseudomyrmex sppCampanotus sp
HemipteraReduviidac
Zelus spPentatomidae
Loxa spPodisus cloeliaSupputius sp. near typicus3
ColeopteraCoccinellidae
Olla abdominalis Say (?)Curinus colombianus Chapin
NeuropteraChrysopidae (1 species)
AraneaeTheridiidae
Achaearanea tesselata*Anelosimus sp
SalticidaeSpecies 1Species 2
Stage (s) of A. tripterus attacked Trees in which
In captivity In nature enemy was found
n P.d., Csp.
n1 P.d., Csp.
e P.d., Csp., Tx.e P.d., Tx.e Tx.e P.d.
«„ n P.d., Tx.
e P.d.e,nlTn iV.n.a P.d.e,n1(n e'.n^.a P.d., Tx.
e Pd.e P.d., Csp.
cnj P.d., Csp., Tx.
n,a P.d., Csp., T.c.n* P.d.
n Tx.a1 Tx.
1 Recently moulted.'Female A. tripterus present.3 Also preys on adult Phanuropsis semiflaviventris in nature.4 Also preys on Bicyrtes variegata in nature.

34 SMITHSONIAN CONTRIBUTIONS TO ZOOLOGY
Females of P. semiflaviventris, but not T. bod-kini, tend to stay and hunt in the tree where theyemerge; experimental predation on unemergedwasps in given trees significantly reduced the para-sitism by P. semiftaviventris, but not T. bodkini.Both wasps almost certainly use the presence of thedefending female bugs to locate host eggs, andtheir responses to male and dead bugs suggest thatthe location and immobility of the bug are impor-tant. Both wasps hunt their host visually, and P.semiflaviventris hunts during the day.
Both species seem to be primarily aerial hunters.The lack of correlation between the relative isola-tion (both natural and experimental) of the leafwith the eggs and the parasitism rates on the pin-nate leaves of Cassia sp. trees imply that mostsearching is not carried out on foot. A further, in-direct indication that the wasps find bug eggs fromthe air is the tendency of the bugs to face the leaftip rather than its base while guarding their eggs.
It is not easy to reconcile the tendency of thewasps to hunt in flight with the tendency of P.semiflaviventris to stay in the trees where theyemerge. The wasps are very small, and relativelyweak fliers, so that even moderate winds probablytend to wash them out of trees where they areflying. The wind in Cali is usually very weak in themornings, but is often moderate to strong in theafternoons, sometimes reaching 15-20 kmph asearly as 2:30-3:00 PM. Perhaps moderate orstronger winds cause P. semiflaviventris to stop hunt-ing, perhaps the wasps do not normally hunt laterthan about 2:00 PM, or perhaps hunting flights con-sist of many short hops.
It is possible that hunting for eggs on branchesinvolves behavior distinct from that of hunting onleaves, as on several occasions females of T. bodkiniwere seen walking along branches and respondingto bugs as if they were searching for eggs.
It is clear that the species of tree somehow in-fluences the hunting behavior of P. semiflavi-ventris. Parasitism by this wasp shows a generalcorrelation with leaf size, being highest in eggslaid on larger leaves. Even though the defensive be-havior of the bugs may be somewhat less effectiveagainst P. semiflaviventris on the small-leaved P.dulce, eggs on this tree are much less heavily para-sitized than those in other trees. The relativelyconstant rates of T. bodkini in different tree specieson the other hand suggest that its hunting be-
havior is not influenced by the tree species.The rate of parasitism in a given tree species
must be a function of both the facility with whicha wasp finds egg masses (hunting efficiency) andthe time it spends looking for them there (huntingeffort). The relative weights of these two factorsare unknown. It would seem that trees with largerand fewer leaves would, a priori, be easier tosearch; this would explain the variations in P.semiflaviventris rates, but would imply that T.bodkini hunts more in trees which are harder forit to search. Without more knowledge of thenatural habitats of the wasps and bugs and ofalternate hosts of the wasps, it is not possible toweigh these factors.
Although T. bodkini tends to disperse morerandomly among different species of tree than doesP. semiflaviventris, parasitism rates by T. bodkiniwere far from uniform, even in trees of the samespecies, and significant differences occurred ingiven trees at different times. These differencescould be explained by (a) more wasps arriving tohunt in some trees than in others, (b) individualwasps searching more in some trees than in others,and/or (c) some trees being more easily searchedthan others of the same species. The most likely ofthese is (a), at least in those cases in which a giventree's parasitism rates varied over time.
COMPETITION BETWEEN THE WASPS
The two species of wasps compete with eachother at two points in their life cycle; the malestend to exclude each other from egg masses and,probably as a result of larval competition withinthe egg, only one wasp grows from any egg para-sitized by both species. It appears that neitherspecies is completely dominant in either type ofcompetition, although T. bodkini males may besomewhat superior fighters. Both types of competi-tion are common in Cali.
The result of interspecific competition betweenmales is that, at least on some trees, substantialnumbers of females are left unfertilized. Analysesof the sexes of wasps from egg masses showed thatunfertilized females seldom lay eggs (assumingthat unfertilized eggs produce males, as is generallytrue for Hymenoptera). Whether unmated femalesmate away from the egg masses where they emerged(laboratory observations indicate that this is pos-

NUMBER 205 35
sible, at least in T. bodkini), or whether they neveroviposit is not known. In any case, interspecificcompetition among males probably causes appre-ciable reductions in the rates of parasitism by bothspecies in at least some situations.
It should be noted that this host-parasite systemis somewhat unusual in that the bug's defense ofits eggs probably buffers the competition betweenthe wasps to some extent. By more or less excludingT. bodkini from most parts of the egg mass, the bugmakes some eggs avilable to P. semiflaviventriseven in areas of relatively high T. bodkini density.The bug's defense also lowers the premium on thesearching ability of the wasps, since a wasp'sability to outmaneuver the bug once she has lo-cated an egg mass is also a factor in her competitivesuccess. Thus the superiority of one species inhunting might be such that it would displace theother under different circumstances, but would failto do so in this system because of its difficulty inovercoming the defenses of the bugs.
Net influence of the guarding behavior of thebugs on the total effectiveness of the wasp popu-lation's ability to parasitize eggs is not known. Thebugs certainly delayed and repelled some wasps andcaused many others to waste time trying to attackunsuitable eggs. But they also acted as beacons indi-cating the location of the eggs, and the balance ofthese factors is unknown.
EVOLUTIONARY RELATIONSHIPS OF THE WASPS
F. Wilson (1961) described various characteris-tics which are common to most wasps of the familyScelionidae. On comparing the two species of thisstudy against his scheme (Table 23), it is clear thatT. bodkini is less specialized (closer to the familynorm) than P. semiflaviventris and that the mainpoints of difference in both species are associatedwith oviposition.
The differences of both species from the familypattern are probably due to the unusual circum-stances they encounter while ovipositing. Theymust overcome the defensive behavior of the guard-ing bugs, while at least some of the scelionids con-sidered by Wilson attack unguarded eggs (Wilson,pers. comm.). It seems likely that characteristics#6, 7, and 8 would be disadvantageous duringbattles with defending bugs since they would slowthe wasp's oviposition and thus give the bug more
TABLE 23.—Characteristics of T. bodkini and P.semiflaviventris as compared with those of otherscelionids (F. Wilson 1961)
Family norm for Scelionidae
1. Males emerge beforefemales
2. Males stay on egg masswhere emerged, fight withother males there
3. Copulation occurs as femaleemerges from host egg...
4. More females than males...5. Each male can fertilize
many females6. Females fight each other
while attacking an eggmass
7. Female marks host eggafter ovipositing into it..
8. Female distinguishesalready parasitized eggsfrom others
Same characters found in—T. bodkini P
yes7
yes
yesyes
probably
almost never
sometimes
yes, at leastsometimes
. semiflaviventris
yesJ
yes
yesyes
probably
no
no
no
time to kick it away. This idea is supported by thefact that T. bodkini females perform more mark-ing and inspection when on undefended eggs thanwhen battling a bug.
F. Wilson (1961) also describes several othercharacteristics of Asolcus basalis, which he does notattribute to the rest of the family, and the speciesof this study share several of these, including themethod of cutting the cap of the bug's egg onemergence, and the predominance of emergencesin the morning (at least in P. semiflaviventris).They differ in that they oviposit much morequickly—approximately 4 seconds in P. semiflavi-ventris and 20-30 seconds in T. bodkini as against3-4 minutes in A. basalis. Oviposition times ofother scelionids which attack undefended eggs arealso relatively long: 2.5 minutes in Telenomusgifuensis (Hidaka, 1958), 6-8 minutes in T. fariei(Lima, 1928), 5-20 minutes in Hadronotus ajax(Schell, 1943), and 0.5-2 minutes in Platytelen-omus sp. (?) hylas (Moutia and Courtois, 1952).Egg marking behavior by T. bodkini is also muchshorter (1-2 seconds) than that of A. basalis (about20 seconds—F. Wilson, 1961). Again these differ-ences appear to represent adaptations to overcomethe bug's defense of her eggs. One other small dif-

36 SMITHSONIAN CONTRIBUTIONS TO ZOOLOGY
ference is the rubbing movement made occasion-ally by P. semifiaviventris males mounted on fe-males before mating.
P. semifiaviventris is more specialized to over-come the bug's defenses than is T. bodkini: in eachcase in which the two species differ from thescelionid norm, P. semifiaviventris is farther fromthe norm than T. bodkini and the ability of P.semifiaviventris to extend and curve its abdomen,an unusual feature for the family (Scelio pem-bertoni, a parasite of buried grasshopper eggs—Pemberton, 1933—also extends its abdomen whenovipositing), is also clearly an aid in penetratingthe bug's defense. P. semifiaviventris' combinationof "advanced" characters enables it to parasitizerelatively well-defended areas of the egg mass,while the "less specialized" T. bodkini is more orless restricted to one poorly defended edge. T.bodkini attacks appeared to be more efficient in onesituation, however—when the wasps parasitized eggsas they were being laid by the bug.
F. Wilson interpreted the various characteristicshe described as being adaptations favoring thespecies, but it is possible and thus preferable (seeWilliams, 1966, for example) to explain them asadaptations to benefit the individual. Each femalewould be favored by the ability to distinguish andrefrain from ovipositing into eggs already parasi-tized, and by her aggressiveness toward other fe-males on a given egg mass. The same would holdfor aggressiveness between males on egg masses.Aggressive males emerging early and staying tocopulate with females from the same mass would
avoid having to hunt for females, and thus benefitfrom the likely high concentration there, and theywould give their mates a maximum amount oftime to hunt for hosts. Females which mated assoon as they emerged would have more time tosearch for hosts. And within this framework, a fe-male would maximize the reproductive potential ofher offspring by producing more female than maleeggs, laying only the minimum number of maleeggs necessary to insure the fertilization of the fe-males in that egg mass.
SELECTION PRESSURES ON THE BEHAVIOROF THE BUGS
THE CHOICE OF SITES FOR EGG MASSES.—The datarelating the sites of egg masses to variations inparasitism, failure to hatch, losses from the tree byfalling, and losses of first instar nymphs reflect theworkings of natural selection on the bugs' choicesof oviposition sites. The balance of these factorswas not always the same in different trees, and thebugs did not always act in their own best interests.
Natural selection clearly favored laying eggs onbranches rather than leaves in T. catappa, whilelosses were nearly even in P. dulce (Table 24). Inboth species of trees, however, the bugs laid onleaves much more often than on branches, and thedifference between the tree species was the reverseof what survivorship data would predict, with T.catappa leaves being more heavily favored thanthose of P. dulce (Table 24). These data suggest
TABLE 24.—Percent losses of A. tripterus offspring in different trees, on leavesand branches of the same trees and frequency of oviposition in these sites
Cause of loss
Parasitism by—T. bodkiniP. semifiaviventris
Failure to developEggs fall from treeLoss of first instar nymphsTotal average survival
Frequency of oviposition .Number of egg masses ..Number of trees
T. catappa P. dulceLeaves
8%64%
5 %very small2-3%
3 1 %
92.5%519
15
Branches
13%32%
3 %very small8%
52%
7.5%
Leaves
2 1 %10%4%3 1 %10%
49%
87%
Branches
3 5 %
7%8%very small20%
4 5 %
13%1221
14

NUMBER 205 37
the hardly surprising conclusion that the bugsevolved in circumstances very different from thosein present-day urban Cali, and that they are prob-ably presently evolving new behavior to adjust tothis environment. Modified selective regimes inurban areas are probably common for many orga-nisms (e.g., Ford, 1955; Waldbauer and Stern-burg, 1967). Just what the original conditions werein which A. tripterus evolved a preference for ovi-positing on leaves is a matter of conjecture.
Survivorship data also permit comparisons be-tween species of tree. Thus the total survivorshipthrough the first instar on P. dulce was about 48%(49% of the 87% on leaves plus 45% of the 13%
on branches), that on T. catappa was about 33%(31% of 92.5% plus 52% of 7.5%), and that onCassia sp. was about 45% (branch masses were al-most nonexistent; average rates were 54% para-sitism, 1% failure to hatch, 16% falling from thetree, and 5% loss of first instar nymphs). Naturalselection on eggs and first instar nymphs favoredbugs ovipositing in Cassia sp. and P. dulce treesover those laying eggs in T. catappa, but in facttrees of the latter species commonly had large pop-ulations of bugs and eggs.
DEFENSE OF EGGS BY FEMALES.—The striking factthat the defense of their eggs by the female bugs,which in various respects appears to be specifi-cally designed to combat the parasitic wasps,nevertheless causes a net loss of eggs to at least oneof the wasp species, suggests the following pos-sible evolutionary history. The bugs are certainlydescended from an ancestor that did not guard itseggs, although at least four other Antiteuchusspecies are presently subsocial. This ancestor's eggswere probably strong enough to withstand attacksby generalized predators such as ants, but wererelatively defenseless against parasites such asscelionids, which were able to oviposit throughhard shells. Bugs began guarding their eggs becauseof the selective advantage of repelling parasites.Their existing responses to disturbances (as seenin present-day nymphs and males)—antennatingtoward the source of disturbance, shaking the body,and perhaps kicking toward the disturbance—com-bined with suppression of the tendency to flee,served to drive away enough of the parasites tomake guarding worthwhile, and incidentally droveoff generalized predators also.
Over time the defensive responses of the bugs
became more finely adjusted to the attack behaviorof the parasites: they acquired the defensive move-ments of scraping and leaning toward the source ofdisturbance, perhaps later suppressing scrapingafter the nymphs had hatched out; they sup-pressed squirting defensive liquid, feeding, and thetendency to turn and face disturbing stimuli; and,perhaps, they modified several details of oviposi-tion behavior, and began laying more compactlyarranged egg masses, and facing toward the tips ofsuch masses and toward the tip of the leaf. At thesame time, the egg cuticle became thinner, as theeggs were now guarded from attacks of generalizedpredators as well as from parasites, and this mate-rial could be more advantageously used in otherways.
The wasps, especially P. semiflaviventris, evolvedin response to the changes in the behavior of thebugs, modifying several aspects of their ovipositionbehavior to speed it up, and also beginning to usethe presence of the bug as a cue to the location ofegg masses. P. semiflaviventris also evolved a greatlyextendible abdomen. The advances of the waspshave been such that at the present time the bugloses more eggs to parasites than it saves from themby guarding. The production of thinner egg shells,however, has trapped the bugs into remaining ontheir eggs, which are now susceptible to many otherpredators in addition to the parasites.
DEFENSE OF FIRST INSTAR NYMPHS BY FEMALES.—Since losses of unguarded nymphs were not pro-hibitively high (Table 21), the possibility arosethat female bugs lost more than they gained byguarding nymphs. The cost of guarding behaviorwas calculated by comparing the lengths of timeuntil the next oviposition in two cases: femalestaken from newly hatched (<12 hours old) firstinstar nymphs, and females from second instarnymphs (these had spent about a week guardingfirst instar nymphs and 0.5-1 day guarding secondinstar nymphs). The females from the new firstinstars reoviposited on the average 19 days later(giving a rate of egg production of 28/19 or 1.47eggs/day), while the others averaged 17 days untilreoviposition (1.65 eggs/day). Comparing the twotactics—desertion or nondesertion of first instarnymphs—in terms of time, deserters would spend7+19 = 26 days between ovipositions, and non-deserters 7 + 7+17 = 31 days between ovipositions.The time difference multiplied by the rate of egg

SMITHSONIAN CONTRIBUTIONS TO ZOOLOGY
production by deserters (5 X 1.47) gives the actualcost in eggs of guarding behavior—7.4 eggs eachcycle.
The savings, in terms of nymphs, resulting fromguarding behavior amount to about 30% of thenymphs hatched in both P. dulce and Cassia sp.trees (Table 21). For unparasitized masses of 28eggs, this would amount to 8.4 nymphs per cycle.Thus, even without taking into account the advan-tages of protecting nymphs as they moult to thesecond instar (their bodies are soft and for a shortwhile they are without defensive odor) and the
possible dangers to the female before she ovipositedagain, the benefits outweigh the costs of guardingfirst instar nymphs. This should be equally true forsituations of high and low parasitism, unless para-sitism rates are declining steadily.
Desertion rates were higher in P. dulce than inCassia sp., even though the rates of loss of nymphsfrom unguarded groups were about the same inboth species. Thus, at least in urban Cali, thestimuli which cause the bugs to desert their nymphsdo not accurately reflect the likelihood of losses ofunguarded nymphs.
Literature Cited
Ashmead, W. H.1893. Monograph of the North American Proctotrypidae.
United States National Museum Bulletin, 45:5-472.Bequaert, J.
1935. Presocial Behavior among the Hemiptera. Bulletinof the Brooklyn Entomological Society, 30 (5):177—191.
Blatchley, W. S.1926. Hemiptera or True Bugs of Eastern North Amer-
ica. Indianapolis, Indiana: Native Publishing Co.Callan, E. McC.
1944a. A Note on Phanuropsis semiflaviventris Cirault(Hym., Scelionidae), an Egg-parasite of Cacao Stink-bugs. Proceedings of the Royal Entomological So-ciety of London (A), 19 (4-6):48-49.
1944b. Cacao Stink-bugs (Hem., Pentatomidae) in Trini-dad, B.W.I Revista de Entomologia (Rio deJaneiro), 15 (3):321-324.
Cumber, R. A.1964. The Egg-Parasite Complex (Scelionidae: Hymenop-
tera) of Shield Bugs (Pentatomidae, Acanthoso-midae: Heteroptera) in New Zealand. New ZealandJournal of Science, 7 (4):536-554.
DeBach, P., and K. S. Hagen
1964. Manipulation of Entomophagous Species. In P.DeBach (ed.) Biological Control of Insect Pests andWeeds. Pages 429-458. London: Chapman and Hall,Ltd.
Eberhard, W. G.
In press. Insectos y hongos que atacan al chinche delcacao, Antiteuchus tripterus. Revista de la Facul-tad Nacional de Agronomia, Medellin.
Espinal, L. S., and E. Montenegro1963. Formaciones vegetales de Colombia. Bogota: Insti-
tuto Geografico "Agustin Codazzi."Esselbaugh, C. O.
1946. A Study of the Eggs of the Pentatomidae (Hemip-tera). Annals of the Entomological Society of Amer-ica, 39(4):667-691.
Ford, E. B.1955. Moths. London: Collins.
Girault, A. A.1916. A New Genus of Scelionidae from the West Indies.
Entomological News, 49 (640): 198-199.Hidaka, T.
1958. Biological Investigation on Telenomus gifuensisAshmead (Hym.: Scelionidae), an Egg-parasite ofScotinophara lurida Burmeister (Hem.: Pentatomi-dae) in Japan. Ada Hymenoptera, 1 (1):75—93.
Kamal, M.1937. The Cotton Green Bug, Nezara viridula L., and
Its Important Egg-parasite Microphanurus mega-cephalus (Ashmead). Bulletin de la Societe Royaled'Entomologique d'Egypte, 1937:175-193.
Kardonne, G.
1972. La Ceniza del Cigarillo. Published by the author,Medellin, Colombia.
Kirkpatrick, T. W.1957. Insect Life in the Tropics. New York: Longmans,
Green, & Co.Lima, A. da Costa
1940. Insetos do Brasil. 2° Tomo, Hemipteros. Rio deJaneiro: Escola Nacional de Agronomia.
Moutia, L. A., and C. M. Courtois1952. Parasites of the Moth-borers of Sugar-cane in
Mauritius. Bulletin of Entomological Research,43(2):325-259.
Naundorf, G.
1954. Contribuciones al problema del la moniliasis encacao. Cacao en Colombia, 3:35-61.
Odhiambo, T.1959. An Account of Parental Care in Rhinocoris albo-
pilosus Signoret (Hemiptera-Heteroptera: Redu-vidae), with Notes on Its Life History. Proceedingsof the Royal Entomological Society of London (A),34 (10-12): 175-185.
Pickford, R.1964. Life History and Behavior of Scelio calopteni Riley
(Hymenoptera: Scelionidae), a Parasite of Grass-hopper Eggs. Canadian Entomologist 96(9):1167-1172.

NUMBER 205 39
Pcmbcrton, C. E.1933. Introduction to Hawaii of Malayan Parasites (Sce-
lionidac) of the Chinese Grasshopper Oxya chinensis(Thur.) with Life History Notes. Proceedings ofthe Hawaiian Entomological Society, 7 (2):253-264.
Rau, P.1918. Maternal Care in Dinocoris tripterus Fab. (Hemip-
tera). Entomological News, 29:75-76.Ruckes, H.
1966. The Genus Antiteuchus Dallas, with Descriptionsof New Species (Heteroptera, Pentatomidae, Disco-cephalinae). Bulletin of the American Museum ofNatural History, 127 (2):49-102.
Schell, S. C.1943. The Biology of Hadronotus ajax Girault (Hymen-
optera-Scelionidae), a Parasite in the Eggs ofSquash-bug (Anasa tristis De Greer). Annals of theEntomological Society of America, 36:625-635.
Sepulveda, R.1955. Biologfa del Mecistorhinus tripterus F. (Hem.
Pentatomidae) y su posible influencia en la trans-mision de la moniliasis de cacao. Cacao en Colom-bia, 4:15-42.
Waldbauer, G. P., and J. G. Sternburg1967. Differential Predation on Cocoons of Hyalophora
cecropia (Lepidoptera: Saturniidae) Spun on Shrubsand Trees. Ecology 48 (2):312-315.
Williams, G. C.1966. Adaptation and Natural Selection. Princeton, New
Jersey: Princeton University Press.Wilson, E. O.
1971. The Insect Societies. Cambridge, Massachusetts:Harvard University Press.
Wilson F.1961. Adult Reproductive Behavior in Asolcus basalts
(Hymenoptera: Scelionidae). Australian Journal ofZoology, 9 (5): 737-751.
U.S. GOVERNMENT PRINTING OFFICE: 1975 586—384/42


Publication in Smithsonian Contributions to Zoology
Manuscripts for serial publications are accepted by the Smithsonian Institution Press, sub-ject to substantive review, only through departments of the various Smithsonian museums. Non-Smithsonian authors should address inquiries to the appropriate department. If submission isinvited, the following format requirements of the Press will govern the preparation of copy.
Copy must be typewritten, double-spaced, on one side of standard white bond paper, withWz" top and left margins, submitted in ribbon copy with a carbon or duplicate, and accompa-nied by the original artwork. Duplicate copies of all material, including illustrations, should beretained by the author. There may be several paragraphs to a page, but each page should beginwith a new paragraph. Number consecutively all pages, including title page, abstract, text, litera-ture cited, legends, and tables. The minimum length is 30 pages, including typescript and illus-trations.
The title should be complete and clear for easy indexing by abstracting services. Taxonomictitles will carry a final line indicating the higher categories to which the taxon is referable:"(Hymenoptera: Sphecidae)." Include an abstract as an introductory part of the text. Identifythe author on the first page of text with an unnumbered footnote that includes his professionalmailing address. A table of contents is optional. An index, if required, may be supplied by theauthor when he returns page proof.
Two headings are used: (1) text heads (boldface in print) for major sections and chap-ters and (2) paragraph sideheads (caps and small caps in print) for subdivisions. Furtherheadings may be worked out with the editor.
In taxonomic keys, number only the first item of each couplet; if there is only one couplet,omit the number. For easy reference, number also the taxa and their corresponding headingsthroughout the text; do not incorporate page references in the key.
In synonymy, use the short form (taxon, author, date:page) with a full reference at theend of the paper under "Literature Cited." Begin each taxon at the left margin with subse-quent lines indented about three spaces. Within an entry, use a period-dash (.—) to separateeach reference. Enclose with square brackets any annotation in, or at the end of, the entry.For references within the text, use the author-date system: "(Jones, 1910)" and "Jones(1910)." If the reference is expanded, abbreviate the data: "Jones (1910:122, pi. 20: fig. 1)."
Simple tabulations in the text (e.g., columns of data) may carry headings or not, but theyshould not contain rules. Formal tables must be submitted as pages separate from the text, andeach table, no matter how large, should be pasted up as a single sheet of copy.
Use the metric system instead of, or in addition to, the English system.Illustrations (line drawings, maps, photographs, shaded drawings) can be intermixed
throughout the printed text. They will be termed Figures and should be numbered consecu-tively; however, if a group of figures is treated as a single figure, the components should beindicated by lowercase italic letters on the illustration, in the legend, and in text reference!:"Figure 96." If illustrations (usually tone photographs) are printed separately from the text asfull pages on a different stock of paper, they will be termed Plates, and individual componentsshould be lettered (Plate 9b) but may be numbered (Plate 9: figure 2). Never combine thenumbering system of text illustrations with that of plate illustrations. Submit all legends onpages separate from the text and not attached to the artwork. An instruction booklet for thepreparation of illustrations is available from the Press on request.
In the bibliography (usually called "Literature Cited"), spell out book, journal, andarticle titles, using initial caps with all words except minor terms such as "and, of, the." Forcapitalization of titles in foreign languages, follow the national practice of each language.Underscore (for italics) book and journal titles. Use the colon-parentheses system for volume,number, and page citations: "10(2) :5-9." Spell out such words as "figures," "plates," "pages."
For free copies of his own paper, a Smithsonian author should indicate his requirementson "Form 36" (submitted to the Press with the manuscript). A non-Smithsonian author willreceive 50 free copies; order forms for quantities above this amount with instructions for pay-ment will be supplied when page proof is forwarded.

i