voluntary exercise followed by chronic stress strikingly increases mature adult-born hippocampal...
TRANSCRIPT

Neurobiology of Learning and Memory 109 (2014) 62–73
Contents lists available at ScienceDirect
Neurobiology of Learning and Memory
journal homepage: www.elsevier .com/ locate /ynlme
Voluntary exercise followed by chronic stress strikingly increasesmature adult-born hippocampal neurons and prevents stress-induceddeficits in ‘what–when–where’ memory
1074-7427/$ - see front matter � 2013 Elsevier Inc. All rights reserved.http://dx.doi.org/10.1016/j.nlm.2013.12.001
⇑ Corresponding authors. Address: Laboratorio de Medicina Regenerativa, Hos-pital Regional Universitario Carlos Haya de Málaga, Pabellón de Gobierno, sótano,Avenida Carlos Haya 82, 29010 Málaga, Spain. Fax: +34 952 614 102 (E. Castilla-Ortega). Address: Departamento de Psicobiología y Metodología de las CC, Facultadde Psicología, Universidad de Málaga, Campus de Teatinos S/N, 29071 Málaga,Spain. Fax: +34 952 134 142 (L.J. Santín).
E-mail addresses: [email protected] (E. Castilla-Ortega),[email protected] (L.J. Santín).
Estela Castilla-Ortega a,⇑, Cristina Rosell-Valle b, Carmen Pedraza b, Fernando Rodríguez de Fonseca a,Guillermo Estivill-Torrús c, Luis J. Santín b,⇑a Unidad de Gestión Clínica de Salud Mental, Hospital Regional Universitario Carlos Haya, Instituto de Investigación Biomédica de Málaga (IBIMA), Spainb Departamento de Psicobiología y Metodología de las Ciencias del Comportamiento, Universidad de Málaga, and Instituto de Investigación Biomédica de Málaga (IBIMA), E-29071Málaga, Spainc Unidad de Microscopía and Unidad de Gestión Clínica de Neurociencias, Hospital Regional Universitario Carlos Haya, Instituto de Investigación Biomédica de Málaga (IBIMA),E-29009 Málaga, Spain
a r t i c l e i n f o
Article history:Received 10 June 2013Revised 22 November 2013Accepted 3 December 2013Available online 11 December 2013
Keywords:Object recognitionEpisodic-like memoryAdult hippocampal neurogenesisIntermittent restraint stressc-FosFunctional networks
a b s t r a c t
We investigated whether voluntary exercise prevents the deleterious effects of chronic stress on epi-sodic-like memory and adult hippocampal neurogenesis. After bromodeoxyuridine (BrdU) administra-tion, mice were assigned to receive standard housing, chronic intermittent restraint stress, voluntaryexercise or a combination of both (stress starting on the seventh day of exercise). Twenty-four days later,mice were tested in a ‘what–when–where’ object recognition memory task. Adult hippocampal neuro-genesis (proliferation, differentiation, survival and apoptosis) and c-Fos expression in the hippocampusand extra-hippocampal areas (medial prefrontal cortex, amygdala, paraventricular hypothalamic nucleus,accumbens and perirhinal cortex) were assessed after behavior. Chronic intermittent restraint stressimpaired neurogenesis and the ‘when’ memory, while exercise promoted neurogenesis and improvedthe ‘where’ memory. The ‘when’ and ‘where’ memories correlated with c-Fos expression in CA1 andthe dentate gyrus, respectively. Furthermore, analysis suggested that each treatment induced a distinctpattern of functional connectivity among the areas analyzed for c-Fos. In the animals in which stressand exercise were combined, stress notably reduced the amount of voluntary exercise performed. Nev-ertheless, exercise still improved memory and counteracted the stress induced-deficits in neurogenesisand behavior. Interestingly, compared with the other three treatments, the stressed exercising animalsshowed a larger increase in cell survival, the maturation of new neurons and apoptosis in the dentategyrus, with a considerable increase in the number of 24-day-old BrdU + cells that differentiated intomature neurons. The interaction between exercise and stress in enhancing the number of adult-born hip-pocampal neurons supports a role of exercise-induced neurogenesis in stressful conditions.
� 2013 Elsevier Inc. All rights reserved.
1. Introduction
New cells are constantly generated in the adult hippocampus.The cells mature into neurons that are functionally integrated intothe circuits of the dentate gyrus and have a potential role in behav-ior (Castilla-Ortega, Pedraza, Estivill-Torrus, & Santin, 2011; Deng,Aimone, & Gage, 2010; Jessberger & Kempermann, 2003). These
adult-born hippocampal neurons show an enhanced susceptibilityto modulation by experience (Castilla-Ortega, Pedraza et al., 2011;Deng et al., 2010), and are highly sensitive to the effects of chronicstress and voluntary exercise. Therefore, while chronic stressdownregulates the proliferation, differentiation and survival ofnew neurons and impairs many forms of hippocampal-dependentmemory, voluntary exercise has the opposite effect, potentiatingboth hippocampal neurogenesis and cognition (Conrad, 2010;Klaus & Amrein, 2012; Schoenfeld & Gould, 2012). Furthermore,voluntary exercise has been shown to be a critical factor in theimprovement of neurogenesis and memory that occurs followingthe broadly used environmental enrichment protocols thatcombine novel stimulation with free access to a running wheel(Mustroph et al., 2012). Despite the reported ability of voluntaryexercise to prevent or enable recovery from stress-induced deficits

E. Castilla-Ortega et al. / Neurobiology of Learning and Memory 109 (2014) 62–73 63
in hippocampal plasticity and behavior (Head, Singh, & Bugg, 2012;Zheng et al., 2006), few studies that combine these two treatmentshave focused on adult hippocampal neurogenesis (Kiuchi, Lee, &Mikami, 2012; Nakajima, Ohsawa, Ohta, Ohno, & Mikami, 2010;Yau, Lau, Tong et al., 2011; Yau et al., 2012). Those studies revealedthat exercise counteracts the suppressive effects of chronic stressor corticosterone administration on hippocampal cell proliferation(Kiuchi et al., 2012; Nakajima et al., 2010; Yau, Lau, Tong et al.,2011; Yau et al., 2012), but other aspects of neurogenesis such ascell survival have been less frequently assessed and may not ben-efit from this protective effect (Nakajima et al., 2010). On the otherhand, the assessment of adult hippocampal neurogenesis underconditions of stress and voluntary exercise is of great interest be-cause the adult-born neurons may be part of the mechanism bywhich exercise counteracts the deleterious effects of stress(Snyder, Soumier, Brewer, Pickel, & Cameron, 2011; Yau, Lau, &So, 2011; Yau, Lau, Tong et al., 2011).
Here, we studied the impact of chronic intermittent restraintstress and voluntary exercise, both separately and in combination,on adult hippocampal neurogenesis. In addition, hippocampaldependent memory was evaluated in the three-trial ‘what–when–where’ object recognition task (Www-Task) developed byDere, Huston, and De Souza Silva (2005). This task requires miceto remember both the temporal order in which a familiar objectwas presented (‘when’ memory) and the place it was presented(‘where’ memory) because the integrated memory for an event, atime and a place constitutes the basis of episodic-like memory(Dere et al., 2005). The ‘when’ and ‘where’ memories are morecomplex than recognition memories for familiar objects and loca-tions and involve greater hippocampal demand (Albasser, Amin,Lin, Iordanova, & Aggleton, 2012; Castilla-Ortega et al., 2012). Arole for the medial prefrontal cortex (mPFC) has also been estab-lished, as the mPFC interacts with the hippocampus to solve the‘where’ memory (DeVito & Eichenbaum, 2010). On the other hand,functional imaging has revealed that, compared to resting levels,performing the Www-Task increases functional activity in the hip-pocampus, mPFC, basolateral amygdala (BLA), paraventricularhypothalamic nucleus (PVN), accumbens (Acb) and perirhinal cor-tex (PRh) (Castilla-Ortega et al., 2012 and unpublished observa-tions). In addition to their potential role in the Www-Task, theseextrahippocampal areas are relevant because of their involvementin stress and reward processing (Herman et al., 2003; Tzschentke &Schmidt, 2000). Ninety minutes after the completion of the Www-Task, animals were sacrificed and perfused. The hippocampal andextrahippocampal (mFPC, BLA, PVN, Acb and PRh) c-Fos expressionwas then assessed as a measure of the functional activity inducedby behavior (Barry & Commins, 2011). Moreover, following eachexperimental treatment, c-Fos expression in the newly born cellswas examined to explore their potential involvement in the behav-ioral task.
2. Experimental procedures
2.1. Animals
Experiments were performed on 3-month-old male mice with ahybrid C57BL/6J�129X1/SvJ background. Six mice were used foreach experimental condition, excluding the control group, whichincluded 10 animals for behavioral analysis. Six of these controlsanimals were then randomly selected for subsequent histology.In all cases, mice were maintained on a 12-h light/dark cycle (lightson at 08:00 a.m), with water and food provided ad libitum. All pro-cedures were performed in accordance with European animal re-search laws (European Communities Council Directives 86/609/EEC, 98/81/CEE and 2003/65/CE; Commission Recommendation
2007/526/EC) and the Spanish National Guidelines for AnimalExperimentation and the Use of Genetically Modified Organisms(Real Decreto 1205/2005 and 178/2004; Ley 32/2007 and 9/2003).
Please note that, as it is reported through the manuscript, theC57BL/6J�129X1/SvJ mice seem to differ from frequently usedmouse strains in both their adult hippocampal neurogenesis andcognitive ability (please see the last paragraph of the Discussionfor a summary), so some caution is needed to directly extrapolateour findings to mice with a different genetic background.
2.2. BrdU administration
BrdU administration was performed on the first day of theexperiment. Mice received four doses of 75 mg/kg BrdU (Sigma,St. Louis, USA) dissolved in saline, which were administered intra-peritoneally at 2-h intervals.
2.3. Environmental treatments
The day after BrdU administration, mice were assigned to one offour experimental treatments: standard housing (Control), chronicintermittent and uncontrollable restraint stress (Stress), voluntaryexercise (EX) or chronic intermittent restraint stress combinedwith voluntary exercise (Stress + EX) (Fig. 1A).
The Control condition consisted of individually housing mice instandard cages (11 � 30 cm and 13 cm high) provided with nestingmaterial. The Stress mice also received standard housing condi-tions, but, beginning 7 days after the administration of BrdU, theywere restrained for 3.5 h per day (10:00–13:30 a.m.), excluding theweekends, in 50 ml clear polystyrene conical centrifuge tubesmodified with air holes for ventilation. For EX mice, the exercisetreatment began 1 day after the BrdU injections, and mice wereindividually housed in exercise cages consisting of two-floor cages(20 � 26 cm and 27 cm high) provided with nesting material, a lad-der and a running wheel equipped with a magnetic counter (Day-ang Pet Products, Foshan City, China). For the Stress + EX group,both the stress and exercise treatments were applied, so mice werehoused in the exercise cages and, after 6 days of running-wheeladaptation, were exposed to chronic intermittent restraint(Fig. 1A). This experimental desing emphasizes the use of exerciseas a ‘preventive’ approach (Wright & Conrad, 2008), because exer-cise is allowed to exert its effects for 6 days prior to the onset ofchronic stress. Because 6 days seem enough for exercise to induceplastic changes within the hippocampus (Adlard, Perreau, &Cotman, 2005; Fabel et al., 2003), exercise may establish aprotective environment that could counteract the deleteriousconsequences of the subsequently applied stressor.
As a measure of voluntary exercise, wheel running was moni-tored daily in both the EX and Stress + EX groups. The distancerun (number of rotations multiplied by the wheel perimeter) wasaveraged every three days for analysis across the exercise protocol.In addition, the total distance run was calculated as the mean dis-tance run per day.
2.4. Behavioral analysis in the ‘what–when–where’ object recognitiontask
On day 24, all mice were tested using the behavioral protocoldepicted in Fig. 1B. Mice received 5 min of habituation to anopen-field arena (40 � 40 cm). After 60 min, four copies of an ob-ject were positioned in the apparatus in a triangular shape andmice allowed to explore for 10 min (Sample 1). Ninety minutes la-ter, four copies of a new object were placed in the corners and themice allowed to explore for 10 min (Sample 2). The Test Trial began90 min later and lasted another 10 min. Two objects from thesecond sample (recent objects) were placed in their previous

Fig. 1. (A) Experimental design. Voluntary exercise began the day after the administration of BrdU in the EX and Stress + EX groups, while chronic stress started on day 7 forthe Stress and Stress + EX animals. On day 24, all animals were tested on the Www-Task and perfused 90 min later. (B) Behavioral design of the Www-Task.
64 E. Castilla-Ortega et al. / Neurobiology of Learning and Memory 109 (2014) 62–73
positions. Two objects from the first sample (old objects) were alsoincluded, but one was placed in its previous position (old-staticobject) and the other displaced to a location that was previouslyoccupied by a recent object (old-displaced object). The type ofobject designated old or recent was counterbalanced across mice,and objects were replaced by identical copies in each trial. A descrip-tion of the objects used is included in Supplementary Material 1.
The Habituation Trial was analyzed for exploration (rearing andlocomotion) and anxiety-like behaviors (defecation and thigmo-taxis) within a novel context (Castilla-Ortega et al., 2010). Rearing(the mouse standing up on its back paws) and defecation were ana-lyzed observationally, while locomotion (cm travelled) and thig-motaxis (percentage of time spent in the periphery of the maze,defined as the area within 8 cm of the walls) were analyzed withEthovision XT 5.0. software (Noldus, Wageningen, The Nether-lands). During the Sample and Test trials, locomotion, thigmotaxisand the total time the mice spent exploring objects (i.e., touchingthem with the nose or forepaws, analyzed observationally) wererecorded.
Memory was analyzed during the Test Trial. First, an objectexploration ratio was calculated for each object, defined as thetime spent exploring the specific object divided by the total timespent exploring all objects (Barbosa, Pontes, Ribeiro, & Silva,2010). The ‘when’ memory was defined as the preference forexploring old objects over recent objects (Dere et al., 2005) andwas determined using two procedures: by comparing the meanof the exploration ratios for both old objects to the mean of theexploration ratios for both recent objects (‘when’ memory-1);and by comparing the exploration ratio of the old-static objectto the mean of the exploration ratios for both recent objects(‘when’ memory-2), to remove the potential influence of theold-displaced object. The ‘where’ memory was measured basedon the preference for the old-displaced over the old-static object(Dere et al., 2005) after comparing their respective exploration ra-tios. Each memory was also calculated as a preference ratio,which was evaluated for correlation with the other variables as-sessed in this study: ‘when’ ratio-1: the mean of the explorationratios for both old objects minus the mean of the exploration ra-tios for both recent objects; ‘when’ ratio-2: the exploration ratioof the old-static object minus the mean of the exploration ratiosfor both recent objects; ‘where’ ratio: the exploration ratio of theold-displaced object minus the exploration ratio of the old-staticobject.
2.5. Histology
2.5.1. Immunohistochemistry and determination of apoptosisNinety minutes after the completion of the Www-task, mice
were intracardially perfused with 0.1 M phosphate-buffered saline,pH 7.4 (PBS), and a periodate-lysine-paraformaldehyde solution(PLP) in PBS (McLean & Nakane, 1974). Brains were post-fixed for48 h in PLP and cut through the interhemispheric fissure intotwo hemispheres. Each hemisphere was then cut in coronal sec-tions (50 lm) using a vibratome. Sections of the left hemispherewere processed for immunohistochemistry with the peroxidase-conjugated ExtrAvidin method using diaminobenzidine (DAB)as the chromogen, as described previously (Castilla-Ortega,Hoyo-Becerra et al., 2011; Matas-Rico et al., 2008). The primaryantibodies used were rabbit anti-c-Fos (Santa Cruz Biotechnology,Santa Cruz, USA; diluted 1:2500) to detect neuronal activation,mouse anti-Proliferating Cell Nuclear Antigen (PCNA; Sigma;1:1000) to mark cells undergoing proliferation, goat anti-double-cortin (DCX; Santa Cruz; 1:200) to detect immature neurons, andmouse anti-BrdU (Developmental Studies Hybridoma Bank, Iowa,USA; 1:1000) to reveal the 24-day-old surviving cells. Secondaryantibodies were biotin-conjugated swine anti-rabbit, rabbit anti-mouse or rabbit anti-goat (1:500, Dako, Glostrup, Danmark), asappropriate.
Sections of the right hemisphere were used for fluorescenceimmunohistochemistry. Sections were first subjected to aheat-induced antigen retrieval with EnVision buffer (Dako) in amicrowave for 1 min. To detect neurogenesis, the neuronalphenotype of the BrdU + cells was studied using double immuno-histochemistry for BrdU in conjunction with the mature neuronalmarker calbindin (rabbit anti-calbindin; CB; Swant, Bellinzoa,Switzerland) or the immature neuronal marker DCX. Sections wereincubated overnight in either the CB (1:500) or the DCX (1:200)antibody, then incubated in the corresponding secondary antibodybefore being submitted to DNA denaturation in 2 N HCl for 30 minat 37 �C, followed by 10 min in 0.1 M boric acid (pH 8.5) at roomtemperature. Sections were then incubated overnight in BrdUantibody (1:500), followed by incubation with the appropriatesecondary antibody. Double immunohistochemistry was alsoperformed to study c-Fos expression within the population ofsurviving cells (c-Fos/BrdU) and immature neurons (c-Fos/DCX).For c-Fos/BrdU labeling, acid DNA denaturation and subsequentneutralization were carried as described above. Sections were then

E. Castilla-Ortega et al. / Neurobiology of Learning and Memory 109 (2014) 62–73 65
incubated overnight in a mixture of c-Fos and BrdU antibodies(1:500), followed by a mixture of the secondary antibodies. Finally,for the c-Fos/DCX staining, sections were first processed for DCXimmunohistochemistry, followed by c-Fos inmmunolabeling.Secondary antibodies were donkey Alexafluor� 488 or donkeyAlexafluor� 568 anti-mouse, anti-goat, or anti-rabbit (Invitrogen,Carlsbad, USA), as appropriate, diluted at 1:800 in PBS for a 2 hincubation. In all cases, rinses with PBS or Tris–PBS (for BrdU) wereperformed after each step. Apoptosis was evaluated in theright hippocampus using an in situ apoptosis detection kit(NeuroTACS-II, Trevigen, Gaithersburg, USA) according to themanufacturer’s instructions.
2.5.2. Cell countingCell counting was carried out in the dorsal hippocampus be-
tween �1.22 and �2.54 mm from bregma (Paxinos & Franklin,2001). The dentate gyrus (DG) was analyzed for the neurogene-sis-related markers, while the whole hippocampus (DG, CA3 andCA1) was analyzed for apoptosis and c-Fos expression. For c-Fosanalysis, the DG was separated into the suprapyramidal (SupraDG)and infrapyramidal (InfraDG) blades because each blade shows adistinct c-Fos response following the Www-Task (Castilla-Ortegaet al., 2012). In addition, the mPFC (including both the infralimbicand prelimbic cortices), BLA, PVN, Acb (both the core and shell) andPRh were also quantified for c-Fos.
For the quantification of DAB staining and apoptosis, high mag-nification photos (4080 � 3072 pixels using a 10� lens) were takenwith an Olympus BX51 microscope equipped with an OlympusDP70 digital camera (Olympus, Glostrup, Denmark). The softwareImageJ (US National Institutes of Health, Maryland, USA) with a cellcounter plug-in was used for quantification. The boundaries of thestructures were delineated by hand with the ImageJ software andthe guide of a mouse brain atlas (Paxinos & Franklin, 2001). Thepixels included within each quantification zone (i.e. the surface de-fined by the boundary) were measured, and we did not find anydifference in the analyzed areas that could account for the numberof cells quantified in each environmental treatment. Following pre-viously reported criteria (Plumpe et al., 2006), DAB-stainedDCX + cells were classified as belonging to a less mature popula-tion (with absent or short dendritic processes, referred to as A–Dcells) or a more mature population (with at least one vertical apicaldendrite penetrating the granule cell layer, referred to as E–F cells).To quantify fluorescence, sections were examined with an epifluo-rescence microscope (Olympus BX51) using a 40� objective. Po-tential double-labeling was examined further using a confocalmicroscope (Leica SP5 II; Leica, Solms, Germany) to confirm co-localization of the two labels.
One out of every 4 (for DAB immunolabeling) or 1 out of every 8(for fluorescent immunolabeling and apoptosis) sections werequantified. The total number of labeled cells was then multipliedby 4 or 8, as appropriate, to estimate the total number of positivelystained cells per structure (Kempermann, Gast, Kronenberg,Yamaguchi, & Gage, 2003). Please note that c-Fos data in ourprevious study (Castilla-Ortega et al., 2012) were expressed as amean per section, so both studies are not directly comparableone to the other in terms of the total c-Fos counts.
2.6. Statistical analysis
The threshold for statistical significance was set at P 6 0.05 inall cases. Wheel running in the EX and Stress + EX groups was ana-lyzed throughout the experiment using a repeated measures ANO-VA (‘stress � day’, with ‘day’ as a repeated measure) and post hocFisher’s least significant difference (LSD) analysis. The total dis-tance run was analyzed using Student’s t test. Memory was ana-lyzed by comparing the objects’ exploration ratios using repeated
measures ANOVAS (‘stress � exercise � object’, with ‘object’ as arepeated measure), followed by LSD for intragroup comparisons.The rest of the measures were analyzed using factorial ANOVAS(‘stress � exercise’) followed by LSD when appropriate. The poten-tial relationships among the assessed variables were exploredusing Pearson’s correlations, carried out on the whole sample ofdata as well as separately for each experimental treatment. Thus,we tested the correlation between the c-Fos activity in the ana-lyzed brain areas in each experimental group. This approach hasbeen recently used to research the functional neuronal networksthat are activated after a behavioral task (Leger et al., 2012; Sietteet al., 2012). The significant c-Fos correlations between areas weregraphically represented for each group, with each brain area drawnas a node and a line connecting two nodes (indicating functionalconnectivity) if the c-Fos expression between these two brain areaswas correlated (Siette et al., 2012). Because several brain areaswere tested for c-Fos correlations, we applied the False DiscoveryRate method of Storey (Storey, 2002; Storey & Tibshirani, 2003)to control for the alpha inflation that results from multiple com-parisons, as has been done previously (D’Anna and Gammie,2006; Rhodes, Ryabinin, & Crabbe, 2005). This method allows thecalculation of q-values, which indicate the proportion of significantresults attributed to chance (i.e., false discoveries) expected for acertain P value in a set of multiple tests. Calculations were carriedout with the open source Q-value Software [http://genom-ics.princeton.edu/storeylab/qvalue/] (Storey, 2002).
3. Results
3.1. Voluntary exercise is reduced by restraint stress
The analysis of voluntary exercise revealed that the EX miceprogressively increased the distance they ran across successivedays. However, in the Stress + EX mice, restraint stress strongly re-duced voluntary exercise, and mice did not escalate their runningbehavior in the stress condition (‘stress’: F(1,10) = 15.544,P = 0.002; ‘day’: F(7,70) = 4.460, P = 0.000; ‘stress � day’:F(7,70) = 7.833, P = 0.000; LSD is shown in Fig. 2A). Accordingly,the total distance run throughout the experiment was notably lessin the Stress + EX mice (Student’s t test result: t(10) = 3,943,P = 0.003; Fig. 2B). It is interesting to note that the reduction inexercise was evident from the first day restraint was applied, indi-cating an effect of acute stress. Moreover, the Stress + EX animalsdid not increase their running behavior on weekends, when theywere not submitted to restraint and therefore had the same acces-sibility to the wheel than the EX mice (individual data from eachday are not shown).
3.2. Voluntary exercise prevents stress-induced deficits in ‘when’memory and improves ‘where’ memory on the Www-Task
3.2.1. Exploratory and anxiety-like behaviorOn the Habituation Trial, no between-group differences were
found on the exploratory measures of locomotion and rearing, oron defecation. An effect of ‘exercise’ was found on thigmotaxis(F(1,20) = 7.421, P = 0.013), indicating EX and Stress + EX animalsspent less time in the periphery (LSD: P < 0.05). On the Sample tri-als and Test trial, averaged as a unique measure, no between-groupdifferences were found in the total locomotion or in the total timespent in object exploration. However, regarding thigmotaxis, theStress group spent more time in the periphery than the othergroups (LSD: P < 0.05). Data are shown in Supplementary Material2.

Fig. 2. Voluntary wheel running in stressed and non-stressed mice. (A) Mean daily distance run, averaged over three days. Arrow indicates the onset of stress on day 7 in theStress + EX group. (B) Total distance run (mean per day) throughout the whole experiment. Mean ± SEM. Difference between groups: �P < 0.05; within-group difference fromthe previous day: $P < 0.05.
66 E. Castilla-Ortega et al. / Neurobiology of Learning and Memory 109 (2014) 62–73
3.2.2. ‘When’ and ‘where’ memoriesAnalysis of the Test Trial revealed that both Control and EX mice
exhibited ‘when’ memory (LSD: P < 0.001 for ‘when’ memory-1;P < 0.05 for ‘when’ memory-2; Fig. 3A and B). However,‘when’ memory was impaired by chronic stress, because the Stressgroup did not discriminate between objects (LSD: P > 0.05) in any‘when’ memory ratio (‘when’ memory-1: ‘exercise’: F(1,24) =12.375, P = 0.002; ‘object’: F(1,24) = 18.694, P = 0.000; ‘object �stress’: F(1,24) = 8.415, P = 0.008; ‘object � exercise � stress’:F(1,24) = 4.418, P = 0.047; Fig. 3A; ‘when’ memory-2: ‘object’:F(1,24) = 7.813, P = 0.011; ‘object � stress’: F(1,24) = 8.512, P =0.009; ‘object � exercise � stress’: F(1,24) = 5.155, P = 0.034;Fig. 3B). When chronic stress was applied to exercising animals,the mice (Stress + Ex group) preferred old objects over recent ob-jects (LSD: P < 0.05; ‘when’ memory-1; Fig. 3A), but this preferencewas not significant when only the old-static object was included inthe analysis (LSD: P > 0.05; ‘when’ memory-2; Fig. 3B). However, itis noteworthy that the object exploration ratios of the Stress + EXgroup differed significantly from the ratios of the Stress group(LSD: P < 0.05), but not from the ratios of the EX group (LSD:P > 0.05). Furthermore, the interaction between ‘object � exer-cise � stress’ was significant, suggesting that the negativeeffects of stress on the ‘when’ memory were mostly blocked byexercise.
Regarding the spatial component of the task, the ‘where’memory, it was absent in both the Control and Stress conditionsbecause animals did not discriminate the old-displaced objectfrom the old-static object (LSD: P > 0.05). However, exerciseestablished this memory in both the EX and Stress + EX animals(LSD: P < 0.05; ‘stress’: F(1,24) = 5.010, P = 0.035; ‘object’:F(1,24) = 8.967, P = 0.006; ‘object � exercise’: F(1,24) = 4.483,P = 0.045; Fig. 3C).
Fig. 3. Memory on the Www-Task. (A and B) Temporal order (‘when’) memory assessed aobject over recent objects (B). (C) Object location (‘where’) memory assessed as the prWithin-group difference between object exploration ratios, $P < 0.05, $$P < 0.001.
3.3. Environmental treatments modulate hippocampal c-Fosexpression, which is correlated with memory, and result in distinctpatterns of hippocampal functional connectivity
3.3.1. Total c-Fos expressionAll environmental treatments induced changes in the
hippocampal c-Fos expression elicited by the Www-Task(SupraDG: ‘exercise’: F(1,20) = 23.265, P = 0.001; InfraDG: ‘exer-cise � stress’: F(1,20) = 6.447, P = 0.020; CA3: ‘exercise � stress’:F(1,20) = 10.104, P = 0.005; Fig. 4A). The Stress animals showedmore c-Fos + cells in area CA3 than the Control and EX + Stress mice(LSD: P < 0.05). The EX mice also showed increased c-Fos expressionin CA3 (LSD: P < 0.05 vs. Control and EX + Stress), but it was also in-creased in the SupraDG (LSD: P < 0.05 vs. Control and Stress) and re-duced in the InfraDG (LSD: P < 0.05 vs. Control and EX + Stress).Finally, the Stress + EX group only showed increased numbers ofc-Fos + cells in the SupraDG compared to Control mice (LSD:P < 0.05). No significant changes in c-Fos expression in CA1 werefound among treatments. Of the extra-hippocampal areas(Fig. 4B), the treatments did not change the total c-Fos expressionin the mPFC, BLA or PRh, although increased c-Fos was found inthe PVN of EX mice (LSD: P < 0.05 vs. all other groups; ‘stress’:F(1,20) = 7.125, P = 0.015; ‘exercise � stress’: F(1,20) = 4.360,P = 0.050) and Acb (LSD: P < 0.05 vs. Control and EX + Stress; ‘exer-cise � stress’: F(1,20) = 4.563, P = 0.045). Photomicrographs show-ing c-Fos labeling are included in Supplementary Material 3, whilethe total c-Fos counts are graphically represented in Fig. 4A and Band shown in Supplementary Material 4.
3.3.2. Correlation of c-Fos measuresWhen the whole sample of data (i.e., all groups merged) was
analyzed for correlations between c-Fos expression and the
s the preference for old over recent objects (A), or as the preference for the old-staticeference for the old-displaced object over the old-static object. Mean ± SEM. LSD:
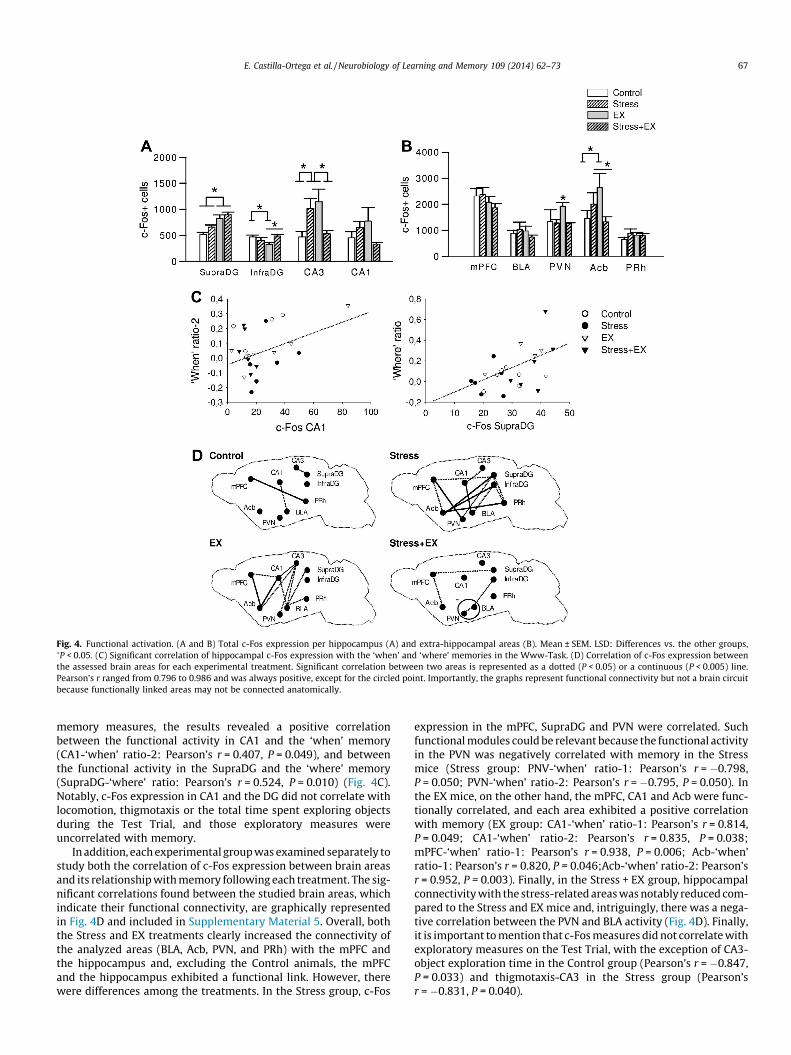
Fig. 4. Functional activation. (A and B) Total c-Fos expression per hippocampus (A) and extra-hippocampal areas (B). Mean ± SEM. LSD: Differences vs. the other groups,�P < 0.05. (C) Significant correlation of hippocampal c-Fos expression with the ‘when’ and ‘where’ memories in the Www-Task. (D) Correlation of c-Fos expression betweenthe assessed brain areas for each experimental treatment. Significant correlation between two areas is represented as a dotted (P < 0.05) or a continuous (P < 0.005) line.Pearson’s r ranged from 0.796 to 0.986 and was always positive, except for the circled point. Importantly, the graphs represent functional connectivity but not a brain circuitbecause functionally linked areas may not be connected anatomically.
E. Castilla-Ortega et al. / Neurobiology of Learning and Memory 109 (2014) 62–73 67
memory measures, the results revealed a positive correlationbetween the functional activity in CA1 and the ‘when’ memory(CA1-‘when’ ratio-2: Pearson’s r = 0.407, P = 0.049), and betweenthe functional activity in the SupraDG and the ‘where’ memory(SupraDG-‘where’ ratio: Pearson’s r = 0.524, P = 0.010) (Fig. 4C).Notably, c-Fos expression in CA1 and the DG did not correlate withlocomotion, thigmotaxis or the total time spent exploring objectsduring the Test Trial, and those exploratory measures wereuncorrelated with memory.
In addition, each experimental group was examined separately tostudy both the correlation of c-Fos expression between brain areasand its relationship with memory following each treatment. The sig-nificant correlations found between the studied brain areas, whichindicate their functional connectivity, are graphically representedin Fig. 4D and included in Supplementary Material 5. Overall, boththe Stress and EX treatments clearly increased the connectivity ofthe analyzed areas (BLA, Acb, PVN, and PRh) with the mPFC andthe hippocampus and, excluding the Control animals, the mPFCand the hippocampus exhibited a functional link. However, therewere differences among the treatments. In the Stress group, c-Fos
expression in the mPFC, SupraDG and PVN were correlated. Suchfunctional modules could be relevant because the functional activityin the PVN was negatively correlated with memory in the Stressmice (Stress group: PNV-‘when’ ratio-1: Pearson’s r = �0.798,P = 0.050; PVN-‘when’ ratio-2: Pearson’s r = �0.795, P = 0.050). Inthe EX mice, on the other hand, the mPFC, CA1 and Acb were func-tionally correlated, and each area exhibited a positive correlationwith memory (EX group: CA1-‘when’ ratio-1: Pearson’s r = 0.814,P = 0.049; CA1-‘when’ ratio-2: Pearson’s r = 0.835, P = 0.038;mPFC-‘when’ ratio-1: Pearson’s r = 0.938, P = 0.006; Acb-‘when’ratio-1: Pearson’s r = 0.820, P = 0.046;Acb-‘when’ ratio-2: Pearson’sr = 0.952, P = 0.003). Finally, in the Stress + EX group, hippocampalconnectivity with the stress-related areas was notably reduced com-pared to the Stress and EX mice and, intriguingly, there was a nega-tive correlation between the PVN and BLA activity (Fig. 4D). Finally,it is important to mention that c-Fos measures did not correlate withexploratory measures on the Test Trial, with the exception of CA3-object exploration time in the Control group (Pearson’s r = �0.847,P = 0.033) and thigmotaxis-CA3 in the Stress group (Pearson’sr = �0.831, P = 0.040).

68 E. Castilla-Ortega et al. / Neurobiology of Learning and Memory 109 (2014) 62–73
The c-Fos correlations among brain areas (Fig. 4D) were ana-lyzed for False Discovery Rates. This analysis revealed that thestandard P = 0.05 cut-off would correspond to a q-value of 0.042in the EX group and of 0.031 in the Stress group (false discoveryrates of 4.2% and 3.1%, respectively). Therefore, less than 5 out of100 significant correlations found in these groups would be falsepositives. In contrast, high q-values were found for the Control(0.610) and EX + Stress (0.374) groups, indicating that as much as61% and 37.4% of the significant correlations found in the Controland EX + Stress groups, respectively, could be false positives. Theseresults suggested that the functional correlations found in the EXand Stress groups were highly reliable and highlighted the differ-ence of these functional networks with those activated duringthe Control and EX + Stress conditions.
3.4. The combination of voluntary exercise with chronic stress inducesthe biggest increase in the number of mature BrdU+/CB+ hippocampalneurons
3.4.1. Expression of neurogenesis-related markers and apoptosis in thehippocampus
The results confirmed that the Stress and EX treatments in-duced opposite effects on adult hippocampal neurogenesis. Com-pared to Control mice, Stress animals exhibited a significantreduction in both cell proliferation, labeled by PCNA (LSD:P < 0.001; Fig. 5A), and young DCX + neurons with mature E-F mor-phology (LSD: P < 0.001; ‘stress’: F(1,20) = 5.760, P = 0.027; Fig. 5B).Although the number of 24-day-old BrdU + cells was not reducedin the Stress mice (only a non-significant tendency toward reduc-tion was found; Fig. 5C), these cells were less frequently differen-tiated into DCX + neurons (LSD: P < 0.05; ‘stress’: F(1,20) = 15.459,P = 0.000; Fig. 5D), and the total number of young BrdU+/DCX + neurons was reduced (LSD: P < 0.05; ‘stress’:F(1,20) = 8.938, P = 0.007; Fig. 5E). In contrast, compared to theControl condition, EX mice increased the population of E-FDCX + cells (LSD: P < 0.05; ‘exercise’: F(1,20) = 35.737, P = 0.000;Fig. 5E) and the rate of cell survival (LSD: P < 0.001; ‘exercise’:F(1,20) = 84.215, P = 0.000; Fig. 5C). Exercise did not change thedifferentiation of the surviving cells into neurons, but the totalnumber of BrdU+/DCX + neurons increased (LSD: P < 0.001; ‘exer-cise’: F(1,20) = 11.208, P = 0.003; Fig. 5E) as a result of the aug-mented cell survival in this group.
When stress and exercise were combined together, voluntaryexercise counteracted the neurogenic damage elicited by thestressful conditions, because the Stress + EX mice showed noreduction in cell proliferation (LSD: P > 0.05 vs. Control and EX,P < 0.001 vs. Stress; ‘exercise’: F(1,20) = 6.360, P = 0.020; ‘exer-cise � stress’: F(1,20) = 19.474, P = 0.011; Fig. 5A), and their num-bers of E-F DCX + cells increased to levels similar to those of theEX animals (LSD: P > 0.05 vs. EX, P < 0.05, 0.001 vs. Control andStress; ‘exercise � stress’: F(1,20) = 4.713, P = 0.043; Fig. 5B). Inaddition, the Stress + EX group showed the greatest cell survivalout of all four of the treatments (LSD: P < 0.05 vs. EX, P < 0.001vs. Control and Stress; ‘exercise � stress’: F(1,20) = 7.763,P = 0.011; Fig. 5C) and a notable increase in the percentage ofBrdU + cells that differentiated into CB+, mature neurons (LSD:P < 0.001 vs. all the other groups; ‘exercise’: F(1,20) = 18.594,P = 0.000; ‘stress’: F(1,20) = 9.700, P = 0.005; exercise � stress’:F(1,20) = 9.559, P = 0.005; Fig. 5D). The Stress + EX group alsoexhibited a reduction in the percentage of BrdU + cells that ex-pressed DCX (LSD: P < 0.05 vs. Control and EX; Fig. 5D), resultingin a striking increase in the total number of BrdU+/CB + cells inthe EX + Stress mice (LSD: P < 0.001 vs. all the other groups; ‘exer-cise’: F(1,20) = 30.261, P = 0.000; ‘stress’: F(1,20) = 12.869,P = 0.002; ‘exercise � stress’: F(1,20) = 14.951, P = 0.001; Fig. 5E).Nevertheless, in both DG blades, c-Fos expression was absent in
BrdU + and DCX + cells following all of the experimental treat-ments (Fig. 5F). The Stress + EX mice also showed increased hippo-campal apoptosis, specifically in the DG (LSD: P < 0.05 vs. all theother groups; ‘exercise � stress’: F(1,20) = 9.858, P = 0.005;Fig. 5G).
3.4.2. Correlation between exercise, memory and adult hippocampalneurogenesis
The potential relationship of neurogenesis with exercise andmemory was explored. The total distance run did not correlatewith behavior in the Www-Task or with any of the previouslymentioned neurogenesis-related measures. However, the amountof exercise achieved by the animals at the end of the experiment(i.e., mean distance run on days 22–23) was strongly correlatedwith the number of generated BrdU+/CB + neurons, separately forthe EX and the Stress + EX conditions (EX: Pearson’s r = 0.885,P = 0.019; Stress + EX: Pearson’s r = 0.967, P = 0.002; Fig. 5H). Neu-rogenesis-related measures were not correlated with behavior ineither the whole sample of data or separately for each treatment.
4. Discussion
This study showed that voluntary exercise is an effective ap-proach for preventing the deficits induced by chronic intermittentstress on both hippocampal-dependent memory in a Www-Taskand adult neurogenesis. Regarding memory, the effects of exerciseclearly surpassed those of chronic stress because the Stress + EXmice performed nearly as well as the EX animals. However, nota-bly, the functional brain networks recruited to perform the taskwere different for each environmental treatment, as suggested bythe c-Fos analysis. On the other hand, the combination of the stressand exercise treatments resulted in a significant enhancement ofadult hippocampal neurogenesis, supporting the idea that adult-born hippocampal neurons could be involved in the compensatorymechanisms driven by exercise in stressful situations.
Voluntary wheel running is considered a valid measure ofexercise in rodents, which escalate this behavior over severalweeks because they find voluntary exercise rewarding (Greenwoodet al., 2011; Novak, Burghardt, & Levine, 2012). Nevertheless, it isimportant to note that voluntary running was quickly andstrongly reduced by restraint stress in the Stress + EX mice. Tothe best of our knowledge, research on the impact of stress onvoluntary exercise is scarce. One previous study suggestedchanges in exercise in chronically stressed mice (Nakajimaet al., 2010), but the results were inconclusive due to the lackof a non-stressed exercise group for comparison. In any case,the reduced amount of exercise performed by the Stress + EXmice resembles the anhedonic effect of stress in inhibitingengagement in naturally rewarding behaviors, such as sexualbehavior and sucrose consumption (Gronli et al., 2005). The func-tional implications of a reduction in the quantity of voluntaryexercise are still unclear. In agreement with existing evidence,we found that the amount of wheel running was not correlatedwith hippocampal-dependent behavior (Greenwood et al., 2003;Pietropaolo et al., 2008; Rhodes, Garland, & Gammie, 2003), butwas correlated with adult hippocampal neurogenesis (Clark,Kohman et al., 2011; Holmes, Galea, Mistlberger, & Kempermann,2004; Mustroph et al., 2012; Rhodes, van Praag et al., 2003).Because neurogenesis specifically correlated with the plateau inthe amount of voluntary exercise performed by the animals atthe end of the experiment, the late phase of exercise could havesome unique effects on adult neurogenesis (Clark et al., 2010) or,conversely, new hippocampal neurons could play a role in laterunning behavior (Clark, Bhattacharya, et al., 2011).
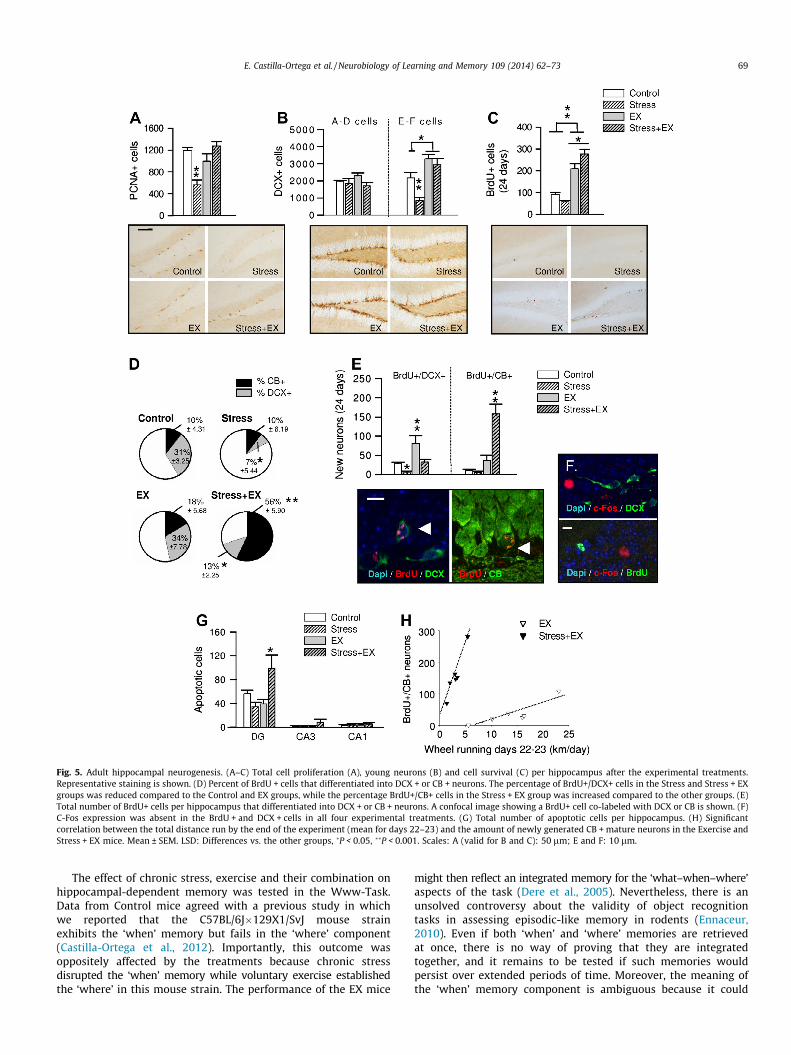
Fig. 5. Adult hippocampal neurogenesis. (A–C) Total cell proliferation (A), young neurons (B) and cell survival (C) per hippocampus after the experimental treatments.Representative staining is shown. (D) Percent of BrdU + cells that differentiated into DCX + or CB + neurons. The percentage of BrdU+/DCX+ cells in the Stress and Stress + EXgroups was reduced compared to the Control and EX groups, while the percentage BrdU+/CB+ cells in the Stress + EX group was increased compared to the other groups. (E)Total number of BrdU+ cells per hippocampus that differentiated into DCX + or CB + neurons. A confocal image showing a BrdU+ cell co-labeled with DCX or CB is shown. (F)C-Fos expression was absent in the BrdU + and DCX + cells in all four experimental treatments. (G) Total number of apoptotic cells per hippocampus. (H) Significantcorrelation between the total distance run by the end of the experiment (mean for days 22–23) and the amount of newly generated CB + mature neurons in the Exercise andStress + EX mice. Mean ± SEM. LSD: Differences vs. the other groups, �P < 0.05, ��P < 0.001. Scales: A (valid for B and C): 50 lm; E and F: 10 lm.
E. Castilla-Ortega et al. / Neurobiology of Learning and Memory 109 (2014) 62–73 69
The effect of chronic stress, exercise and their combination onhippocampal-dependent memory was tested in the Www-Task.Data from Control mice agreed with a previous study in whichwe reported that the C57BL/6J�129X1/SvJ mouse strainexhibits the ‘when’ memory but fails in the ‘where’ component(Castilla-Ortega et al., 2012). Importantly, this outcome wasoppositely affected by the treatments because chronic stressdisrupted the ‘when’ memory while voluntary exercise establishedthe ‘where’ in this mouse strain. The performance of the EX mice
might then reflect an integrated memory for the ‘what–when–where’aspects of the task (Dere et al., 2005). Nevertheless, there is anunsolved controversy about the validity of object recognitiontasks in assessing episodic-like memory in rodents (Ennaceur,2010). Even if both ‘when’ and ‘where’ memories are retrievedat once, there is no way of proving that they are integratedtogether, and it remains to be tested if such memories wouldpersist over extended periods of time. Moreover, the meaning ofthe ‘when’ memory component is ambiguous because it could

70 E. Castilla-Ortega et al. / Neurobiology of Learning and Memory 109 (2014) 62–73
reflect recency memory instead of memory for the temporal orderin which objects were presented (Ennaceur, 2010). In spite ofthese issues, it is important to note that the Www-Task stillallows to establish strong conclusions about hippocampalfunctioning. Lesion and inactivation studies have shown thatthe ‘when’ and ‘where’ memories are independent of one another,but both require the integrity of the hippocampus (Barbosa,Pontes, Ribeiro, Ribeiro, & Silva, 2012; DeVito & Eichenbaum,2010). Additionally, the SupraDG and CA1 areas have been shownto increase their functional activation after the Www-Task in ac-Fos imaging experiment, and mutant mice with hippocampalpathology fail the Www-task in the absence of deficits indiscriminating novel objects and locations from familiar ones(Castilla-Ortega et al., 2012). Therefore, a remarkable result of thisstudy is that the Stress + EX mice showed a mostly preserved‘when’ memory and a correct ‘where’ memory in the Www-Task.Although some caution should be taken to interpret this outcomein terms of episodic-like memory, it clearly shows that the effectsof exercise surpassed those of chronic stress to improve andpreserve complex forms of hippocampal-dependent memory,such as the ones assessed in the Www-Task.
Interestingly, memory was linked to functional activation inthe hippocampus. Each treatment resulted in unique patterns ofhippocampal c-Fos expression following the Www-Task, butwhen data from all experimental conditions were merged andanalyzed as a whole, the amount of c-Fos expression in CA1and the SupraDG could predict the ‘when’ and ‘where’ perfor-mance, respectively. This finding supports a functional dissocia-tion of the hippocampal subfields, with CA1 forming temporalrelationships and the DG processing complex spatial information(Gilbert, Kesner, & Lee, 2001; Hoge & Kesner, 2007). It also agreeswith a recent inactivation study in which CA1 and the DG/CA3area were required for the ‘when’ and ‘where’ memories, respec-tively, in the Www-Task (Barbosa et al., 2012). Neuronal activitywas also studied in several extra-hippocampal areas (mPFC, BLA,PVN, Acb and PRh) that regulate stress and/or reward processing(Herman et al., 2003; Tzschentke & Schmidt, 2000) and are re-ported to exhibit changes in c-Fos expression after stress andexercise (Herman et al., 2003; Rhodes, Garland, et al., 2003).However, we found few differences among the treatments inthe total number of c-Fos + cells. This could be explained becausewe assessed the c-Fos response induced by the Www-Task per-formance instead of the basal activation resulting after the treat-ments. However, as no animals were assessed in basal conditions,we can’t rule out that a potential dysregulation in the basal activ-ity could contribute to our c-Fos results.
In addition to the c-Fos quantification, we assessed whetherc-Fos expression correlated between the analyzed areas, as ameasure of functional connectivity. Such functional relationshipsare likely relevant to the behavioral outcome, given that theinteractions among the hippocampus, mPFC, BLA and PRh haveimportant contributions to object recognition memory (Aggleton,Blindt, & Rawlins, 1989; Leger et al., 2012; Warburton & Brown,2010). In both the Stress and EX mice, functional connectivity be-tween the hippocampus and the extra-hippocampal areas in-creased after behavior, although these connections may havedifferent implications for each treatment. For example, the EXmice showed connections among areas whose individual c-Fosactivity was positively correlated with memory (CA1, Acb andmPFC) but, in the chronically stressed mice, the hippocampusand mPFC were linked to the PVN that negatively correlated withmemory instead. On the other hand, the functional connectivityin the Stress + EX mice was scarce, suggesting that exercise dur-ing chronic stress conditions could inhibit the deleterious influ-ence of the stress-related areas on the hippocampus and mPFC.In any case, the methodology used here is not enough to describe
the complex brain circuits involved in the Www-Task nor themeaning of the functional connections involved. What can beconcluded is that each of the four environmental treatmentsyielded a different functional brain network reorganization. It isdescribed that both stress and exercise induce profound dendriticand synaptic changes (Eadie, Redila, & Christie, 2005; McEwen,2010), so unique morphological and functional restructuringcould take place when these treatments are combined together,which should be investigated further in future research.
Finally, we assessed how the environmental treatments im-pacted adult hippocampal neurogenesis. Chronic stress and vol-untary exercise exerted opposite effects, so exercise enhancedcell survival and DCX + neurons without affecting cell prolifera-tion (Clark et al., 2010; Fuss, Ben Abdallah, & Vogt, 2010; Snyder,Glover, Sanzone, Kamhi, & Cameron, 2009) but stress reducedboth proliferating and DCX + cells (Joels, Karst, Krugers, &Lucassen, 2007; Yun et al., 2010). In the Stress + EX mice, exercisedid prevent the stress-induced impairments and, importantly, themost notable potentiation of neurogenesis was found in thiscondition. Therefore, the Stress + EX mice generated more newmature neurons than mice that underwent the other three treat-ments, and also showed increased apoptosis in the DG, suggestinga structural remodeling due to the increased neurogenesis(Chambers, Potenza, Hoffman, & Miranker, 2004). Although adulthippocampal neurogenesis may be enhanced by certain forms ofstress (Lyons et al., 2010; Parihar, Hattiangady, Kuruba, Shuai, &Shetty, 2011; Snyder, Glover, et al., 2009), this work is the firstto report that a deleterious stressor can interact with voluntaryexercise to promote neurogenesis. Accordingly, the Stress + EXmice performed less exercise than the EX animals, but theamount of exercise performed correlated with greater increasesin new mature neurons. The little literature that has researchedthe combined effect of repeated stress (or corticosterone adminis-tration) and exercise on adult hippocampal neurogenesis has re-ported a protective effect of exercise for the stress-induceddeficits on cell proliferation, but has failed to show the neuro-genic interaction reported here (Kiuchi et al., 2012; Nakajimaet al., 2010; Yau, Lau, Tong, et al., 2011; Yau et al., 2012). It couldbe possibly explained because those studies were performed inrats or in a different mouse strain. Furthermore, they were mainlyfocused on hippocampal cell proliferation and rarely assessedother aspects of neurogenesis, and they also differed in the stressand exercise protocols used. Accordingly, a key factor could bethat here, unlike in previous studies, we submitted the cells thatincorporated BrdU to six days of voluntary exercise alone untilthe stress started, which likely modified the micro-environmentwhere these cells matured. Voluntary exercise elevates glucocor-ticoid levels, but buffers their effect through the concomitantliberation of endogenous molecules such as neurotrophic factors,opioid neuropeptides and dopamine (Schoenfeld & Gould, 2012).That enriched medium thoroughly ‘sculpts’ the adult-born neurons,especially those that are immature (Castilla-Ortega, Pedrazaet al., 2011; Eadie et al., 2005), and could modulate the effectof the deleterious stressor such that the new neurons respondedby increasing their survival and maturation.
Taking these results into account, a main question is whether theenhanced number of adult-born hippocampal neurons is a mecha-nism by which exercise counteracts the effects of stress. It has beenreported that neurogenesis could be more necessary for memorywhen the hippocampus is compromised by a deleterious factor, suchas a chronic stressor (Yau, Lau, Tong et al., 2011). In this study, how-ever, neurogenesis was not functionally related to performance onthe Www-Task, as indicated by its lack of a correlation with memoryand by the absence of c-Fos expression in the BrdU + and DCX + cells.It is possible that performance on the Www-Task, like certain formsof hippocampal memory, may not be neurogenesis-dependent

E. Castilla-Ortega et al. / Neurobiology of Learning and Memory 109 (2014) 62–73 71
(Clark et al., 2008). Nevertheless, this seems unlikely consideringthat the Www-Task involves spatial demands and engages the DG.A stronger possibility is that we did not assess the involvement ofnewly born neurons in memory when they were at an appropriateage. As early as 2–4 weeks of age, new hippocampal neurons canbe functionally integrated in the DG and respond to experience byexpressing immediate early genes (Agis-Balboa et al., 2011;Castilla-Ortega, Pedraza et al., 2011; Snyder, Choe et al., 2009), butsuch studies are usually performed in rats or in C57/BL6 mice.Functional integration of adult-born neurons may be delayed inour hybrid C57BL/6J�129X1/SvJ mouse strain because only �10%of 24-day-old BrdU + cells differentiated into mature CB + neuronsin our Control mice, when at least �60% differentiation is reportedin cells of C57/BL6 mice at 28 days of age (Kempermann, Kuhn, &Gage, 1997). Therefore, new neurons in a later stage of developmentcould contribute to the Www-Task. On the other hand, it is possiblethat the increased neurogenesis in the Stress + EX animals may notbe directly involved in memory, but in stress compensation mecha-nisms instead. A tempting hypothesis is thus that these neuronscould have been integrated into the DG circuits with the specific roleof buffering the stress response to restraint (Snyder et al., 2011). Inany case, considering the overwhelming evidence supporting theinvolvement of adult hippocampal neurogenesis in hippocampal-dependent function, the question that should remain is not ‘if’ theadult born-hippocampal neurons have a functional role, but ‘when’their role takes place.
Ultimately, it is important to mention that, as suggested byour results, the C57BL/6J�129X1/SvJ mice apparently have someunique characteristics that differ from frequently used mousestrains, probably as a consequence of their hybrid background.For example, their voluntary running activity shown in this studyand other (Castilla-Ortega et al., 2013) seems notably higher com-pared to what it is reported for the C57/BL6 mice (Nakajima et al.,2010). Also, hippocampal-dependent cognition is blunted in ourCtrl mice, considering that the C57/BL6 mice are able to performboth memories in the Www-Task (Dere et al., 2005) withoutexposure to any environmental treatment to potentiate their hip-pocampal function. Interestingly, such cognitive impairment islikely related to the unusually low number of new mature hippo-campal neurons in our Ctrl mice, because an impaired memory isa common feature of those mouse strains with reduced neurogen-esis (Kempermann & Gage, 2002). Taking all this into account,this study would benefit from additional experiments performedin rats or in additional mouse strains, as all of our findings maynot be directly comparable.
Funding
This work was supported by grants from Spanish Ministry ofEconomy and Competitiveness, (MEC SEJ2007-61187 -co-fundedby ERDF- and MICINN PSI2010-16160, to L.J.S.; PI10/02514 fromCarlos III Health Institute, co-funded by ERDF, to G.E.-T.; Red deTrastornos Adictivos RD012/0028/0001, to F.R.F.; and ‘‘Sara Bor-rell’’ research contract from Carlos III Health Institute, to E.C.-O.),Andalusian Ministry of Economy, Innovation, Science and Employ-ment (SEJ-4515, to L.J.S; CTS643 and CTS433 research group grantsto G.E.-T. and F.R.F., respectively; SAF2010-20521, to F.R.F; FPUGrant FPDI 2010, to C.R.-V.) and Andalusian Ministry of Health(‘‘Nicolás Monardes’’ Programme, to G.E.-T.). The confocal micros-copy was adquired by the University of Malaga with ERDF funds.
Acknowledgments
We are grateful to Juan Gómez Repiso for his technical assis-tance and for the application of the environmental treatments, toElena Baixeras Llano for accesibility to epifluorescence, to CarmeloMillón Peñuela for his advice on the immunofluorescence labelingtechniques, to David Navas Fernández for his assistance in the useof the confocal microscopy, to University of Málaga for mainte-nance of mice and to the members of the ‘Neuropsicofarmacologíade los Transmisores Lipídicos’ research group for their support.
Appendix A. Supplementary material
Supplementary data associated with this article can be found, inthe online version, at http://dx.doi.org/10.1016/j.nlm.2013.12.001.
References
Adlard, P. A., Perreau, V. M., & Cotman, C. W. (2005). The exercise-inducedexpression of BDNF within the hippocampus varies across life-span.Neurobiology of Aging, 26, 511–520.
Aggleton, J. P., Blindt, H. S., & Rawlins, J. N. (1989). Effects of amygdaloid andamygdaloid-hippocampal lesions on object recognition and spatial workingmemory in rats. Behavioral Neuroscience, 103, 962–974.
Agis-Balboa, R. C., Arcos-Diaz, D., Wittnam, J., Govindarajan, N., Blom, K., Burkhardt,S., et al. (2011). A hippocampal insulin-growth factor 2 pathway regulates theextinction of fear memories. EMBO Journal, 30, 4071–4083.
Albasser, M. M., Amin, E., Lin, T. C., Iordanova, M. D., & Aggleton, J. P. (2012).Evidence that the rat hippocampus has contrasting roles in object recognitionmemory and object recency memory. Behavioral Neuroscience, 126, 659–669.
Barbosa, F. F., Pontes, I. M., Ribeiro, S., Ribeiro, A. M., & Silva, R. H. (2012). Differentialroles of the dorsal hippocampal regions in the acquisition of spatial andtemporal aspects of episodic-like memory. Behavioural Brain Research, 232,269–277.
Barbosa, F. F., Pontes, I. M., Ribeiro, A. M., & Silva, R. H. (2010). Extending possibleapplications of an episodic-like memory task in rats. Behavioural Brain Research,215, 326–331.
Barry, D. N., & Commins, S. (2011). Imaging spatial learning in the brain usingimmediate early genes: Insights, opportunities and limitations. Reviews in theNeurosciences, 22, 131–142.
Castilla-Ortega, E., Hoyo-Becerra, C., Pedraza, C., Chun, J., Rodriguez De Fonseca, F.,Estivill-Torrus, G., et al. (2011). Aggravation of chronic stress effects onhippocampal neurogenesis and spatial memory in LPA(1) receptor knockoutmice. PLoS ONE, 6, e25522.
Castilla-Ortega, E., Pedraza, C., Chun, J., de Fonseca, F. R., Estivill-Torrus, G., & Santin,L. J. (2012). Hippocampal c-Fos activation in normal and LPA(1)-null mice aftertwo object recognition tasks with different memory demands. Behavioural BrainResearch, 232, 400–405.
Castilla-Ortega, E., Pedraza, C., Estivill-Torrus, G., & Santin, L. J. (2011). When is adulthippocampal neurogenesis necessary for learning? Evidence from animalresearch. Reviews in the Neurosciences, 22, 267–283.
Castilla-Ortega, E., Rosell-Valle, C., Blanco, E., Pedraza, C., Chun, J., Rodriguez deFonseca, F., et al. (2013). Reduced wheel running and blunted effects ofvoluntary exercise in LPA-null mice. The importance of assessing the amount ofrunning in transgenic mice studies. Neuroscience Research, 77, 170–179.
Castilla-Ortega, E., Sanchez-Lopez, J., Hoyo-Becerra, C., Matas-Rico, E., Zambrana-Infantes, E., Chun, J., et al. (2010). Exploratory, anxiety and spatial memoryimpairments are dissociated in mice lacking the LPA1 receptor. Neurobiology ofLearning and Memory, 94, 73–82.
Chambers, R. A., Potenza, M. N., Hoffman, R. E., & Miranker, W. (2004). Simulatedapoptosis/neurogenesis regulates learning and memory capabilities of adaptiveneural networks. Neuropsychopharmacology, 29, 747–758.
Clark, P. J., Bhattacharya, T. K., Miller, D. S., & Rhodes, J. S. (2011a). Induction of c-Fos, Zif268, and Arc from acute bouts of voluntary wheel running in new andpre-existing adult mouse hippocampal granule neurons. Neuroscience, 184,16–27.
Clark, P. J., Brzezinska, W. J., Thomas, M. W., Ryzhenko, N. A., Toshkov, S. A., &Rhodes, J. S. (2008). Intact neurogenesis is required for benefits of exercise onspatial memory but not motor performance or contextual fear conditioning inC57BL/6J mice. Neuroscience, 155, 1048–1058.
Clark, P. J., Kohman, R. A., Miller, D. S., Bhattacharya, T. K., Brzezinska, W. J., &Rhodes, J. S. (2011b). Genetic influences on exercise-induced adult hippocampalneurogenesis across 12 divergent mouse strains. Genes Brain Behav, 10,345–353.
Clark, P. J., Kohman, R. A., Miller, D. S., Bhattacharya, T. K., Haferkamp, E. H., &Rhodes, J. S. (2010). Adult hippocampal neurogenesis and c-Fos inductionduring escalation of voluntary wheel running in C57BL/6J mice. BehaviouralBrain Research, 213, 246–252.

72 E. Castilla-Ortega et al. / Neurobiology of Learning and Memory 109 (2014) 62–73
Conrad, C. D. (2010). A critical review of chronic stress effects on spatial learningand memory. Progress in Neuro-Psychopharmacology and Biological Psychiatry,34, 742–755.
D’Anna, K. L., & Gammie, S. C. (2006). Hypocretin-1 dose-dependently modulatesmaternal behaviour in mice. Journal of Neuroendocrinology, 18, 553–566.
Deng, W., Aimone, J. B., & Gage, F. H. (2010). New neurons and new memories: Howdoes adult hippocampal neurogenesis affect learning and memory? NatureReviews Neuroscience, 11, 339–350.
Dere, E., Huston, J. P., & De Souza Silva, M. A. (2005). Integrated memory for objects,places, and temporal order: Evidence for episodic-like memory in mice.Neurobiology of Learning and Memory, 84, 214–221.
DeVito, L. M., & Eichenbaum, H. (2010). Distinct contributions of thehippocampus and medial prefrontal cortex to the ‘‘what–where–when’’components of episodic-like memory in mice. Behavioural Brain Research,215, 318–325.
Eadie, B. D., Redila, V. A., & Christie, B. R. (2005). Voluntary exercise alters thecytoarchitecture of the adult dentate gyrus by increasing cellular proliferation,dendritic complexity, and spine density. J. Comp. Neurol., 486, 39–47.
Ennaceur, A. (2010). One-trial object recognition in rats and mice: Methodologicaland theoretical issues. Behavioural Brain Research, 215, 244–254.
Fabel, K., Tam, B., Kaufer, D., Baiker, A., Simmons, N., Kuo, C. J., et al. (2003). VEGF isnecessary for exercise-induced adult hippocampal neurogenesis. EuropeanJournal of Neuroscience, 18, 2803–2812.
Fuss, J., Ben Abdallah, N. M., Vogt, M. A., Touma, C., Pacifici, P. G., Palme, R., et al.(2010). Voluntary exercise induces anxiety-like behavior in adult C57BL/6J micecorrelating with hippocampal neurogenesis. Hippocampus, 20, 364–376.
Gilbert, P. E., Kesner, R. P., & Lee, I. (2001). Dissociating hippocampal subregions:Double dissociation between dentate gyrus and CA1. Hippocampus, 11,626–636.
Greenwood, B. N., Foley, T. E., Day, H. E., Campisi, J., Hammack, S. H., Campeau, S.,et al. (2003). Freewheel running prevents learned helplessness/behavioraldepression: Role of dorsal raphe serotonergic neurons. Journal of Neuroscience,23, 2889–2898.
Greenwood, B. N., Foley, T. E., Le, T. V., Strong, P. V., Loughridge, A. B., Day, H. E., et al.(2011). Long-term voluntary wheel running is rewarding and producesplasticity in the mesolimbic reward pathway. Behavioural Brain Research, 217,354–362.
Gronli, J., Murison, R., Fiske, E., Bjorvatn, B., Sorensen, E., Portas, C. M., et al. (2005).Effects of chronic mild stress on sexual behavior, locomotor activity andconsumption of sucrose and saccharine solutions. Physiology & Behavior, 84,571–577.
Head, D., Singh, T., & Bugg, J. M. (2012). The moderating role of exercise on stress-related effects on the hippocampus and memory in later adulthood.Neuropsychology, 26, 133–143.
Herman, J. P., Figueiredo, H., Mueller, N. K., Ulrich-Lai, Y., Ostrander, M. M., Choi, D.C., et al. (2003). Central mechanisms of stress integration: Hierarchical circuitrycontrolling hypothalamo-pituitary-adrenocortical responsiveness. Frontiers inNeuroendocrinology, 24, 151–180.
Hoge, J., & Kesner, R. P. (2007). Role of CA3 and CA1 subregions of the dorsalhippocampus on temporal processing of objects. Neurobiology of Learning andMemory, 88, 225–231.
Holmes, M. M., Galea, L. A., Mistlberger, R. E., & Kempermann, G. (2004). Adulthippocampal neurogenesis and voluntary running activity: Circadian and dose-dependent effects. Journal of Neuroscience Research, 76, 216–222.
Jessberger, S., & Kempermann, G. (2003). Adult-born hippocampal neurons matureinto activity-dependent responsiveness. European Journal of Neuroscience, 18,2707–2712.
Joels, M., Karst, H., Krugers, H. J., & Lucassen, P. J. (2007). Chronic stress:Implications for neuronal morphology, function and neurogenesis. Frontiers inNeuroendocrinology, 28, 72–96.
Kempermann, G., & Gage, F. H. (2002). Genetic determinants of adult hippocampalneurogenesis correlate with acquisition, but not probe trial performance, in thewater maze task. European Journal of Neuroscience, 16, 129–136.
Kempermann, G., Gast, D., Kronenberg, G., Yamaguchi, M., & Gage, F. H. (2003). Earlydetermination and long-term persistence of adult-generated new neurons inthe hippocampus of mice. Development, 130, 391–399.
Kempermann, G., Kuhn, H. G., & Gage, F. H. (1997). More hippocampal neurons inadult mice living in an enriched environment. Nature, 386, 493–495.
Kiuchi, T., Lee, H., & Mikami, T. (2012). Regular exercise cures depression-likebehavior via VEGF-Flk-1 signaling in chronically stressed mice. Neuroscience,207, 208–217.
Klaus, F., & Amrein, I. (2012). Running in laboratory and wild rodents: Differences incontext sensitivity and plasticity of hippocampal neurogenesis. BehaviouralBrain Research, 227, 363–370.
Leger, M., Quiedeville, A., Paizanis, E., Natkunarajah, S., Freret, T., Boulouard, M.,et al. (2012). Environmental enrichment enhances episodic-like memory inassociation with a modified neuronal activation profile in adult mice. PLoS ONE,7, e48043.
Lyons, D. M., Buckmaster, P. S., Lee, A. G., Wu, C., Mitra, R., Duffey, L. M., et al. (2010).Stress coping stimulates hippocampal neurogenesis in adult monkeys.Proceedings of the National Academy of Sciences of the United States of America,107, 14823–14827.
Matas-Rico, E., Garcia-Diaz, B., Llebrez-Zayas, P., Lopez-Barroso, D., Santin, L.,Pedraza, C., et al. (2008). Deletion of lysophosphatidic acid receptor LPA1reduces neurogenesis in the mouse dentate gyrus. Molecular and CellularNeuroscience, 39, 342–355.
McEwen, B. S. (2010). Stress, sex, and neural adaptation to a changing environment:Mechanisms of neuronal remodeling. Annals of the New York Academy ofSciences, 1204(Suppl.), E38–59.
McLean, I. W., & Nakane, P. K. (1974). Periodate-lysine-paraformaldehyde fixative. Anew fixation for immunoelectron microscopy. Journal of Histochemistry andCytochemistry, 22, 1077–1083.
Mustroph, M. L., Chen, S., Desai, S. C., Cay, E. B., DeYoung, E. K., & Rhodes, J. S. (2012).Aerobic exercise is the critical variable in an enriched environment thatincreases hippocampal neurogenesis and water maze learning in male C57BL/6Jmice. Neuroscience, 219, 62–71.
Nakajima, S., Ohsawa, I., Ohta, S., Ohno, M., & Mikami, T. (2010). Regular voluntaryexercise cures stress-induced impairment of cognitive function and cellproliferation accompanied by increases in cerebral IGF-1 and GST activity inmice. Behavioural Brain Research, 211, 178–184.
Novak, C. M., Burghardt, P. R., & Levine, J. A. (2012). The use of a running wheel tomeasure activity in rodents: Relationship to energy balance, general activity,and reward. Neuroscience and Biobehavioral Reviews, 36, 1001–1014.
Parihar, V. K., Hattiangady, B., Kuruba, R., Shuai, B., & Shetty, A. K. (2011).Predictable chronic mild stress improves mood, hippocampal neurogenesis andmemory. Molecular Psychiatry, 16, 171–183.
Paxinos, G., & Franklin, K. (2001). The mouse brain in stereotaxic coordinates. SanDiego: Academic Press.
Pietropaolo, S., Sun, Y., Li, R., Brana, C., Feldon, J., & Yee, B. K. (2008). The impact ofvoluntary exercise on mental health in rodents: A neuroplasticity perspective.Behavioural Brain Research, 192, 42–60.
Plumpe, T., Ehninger, D., Steiner, B., Klempin, F., Jessberger, S., Brandt, M., et al.(2006). Variability of doublecortin-associated dendrite maturation in adulthippocampal neurogenesis is independent of the regulation of precursor cellproliferation. BMC Neuroscience, 7, 77.
Rhodes, J. S., Garland, T., Jr., & Gammie, S. C. (2003). Patterns of brain activityassociated with variation in voluntary wheel-running behavior. BehavioralNeuroscience, 117, 1243–1256.
Rhodes, J. S., Ryabinin, A. E., & Crabbe, J. C. (2005). Patterns of brain activationassociated with contextual conditioning to methamphetamine in mice.Behavioral Neuroscience, 119, 759–771.
Rhodes, J. S., van Praag, H., Jeffrey, S., Girard, I., Mitchell, G. S., Garland, T. Jr.,, et al.(2003). Exercise increases hippocampal neurogenesis to high levels but doesnot improve spatial learning in mice bred for increased voluntary wheelrunning. Behavioral Neuroscience, 117, 1006–1016.
Schoenfeld, T. J., & Gould, E. (2012). Stress, stress hormones, and adult neurogenesis.Experimental Neurology, 233, 12–21.
Siette, J., Westbrook, R. F., Cotman, C., Sidhu, K., Zhu, W., Sachdev, P., & Valenzuela,M. J. (2012). Age-Specific Effects of Voluntary Exercise on Memory and theOlder Brain. Biological Psychiatry.
Snyder, J. S., Choe, J. S., Clifford, M. A., Jeurling, S. I., Hurley, P., Brown, A., et al.(2009a). Adult-born hippocampal neurons are more numerous, faster maturing,and more involved in behavior in rats than in mice. Journal of Neuroscience, 29,14484–14495.
Snyder, J. S., Glover, L. R., Sanzone, K. M., Kamhi, J. F., & Cameron, H. A. (2009b). Theeffects of exercise and stress on the survival and maturation of adult-generatedgranule cells. Hippocampus, 19, 898–906.
Snyder, J. S., Soumier, A., Brewer, M., Pickel, J., & Cameron, H. A. (2011). Adulthippocampal neurogenesis buffers stress responses and depressive behaviour.Nature, 476, 458–461.
Storey, J. D. (2002). A direct approach to false discovery rates. Journal of the RoyalStatistical Society, Series B, 64, 479–498.
Storey, J. D., & Tibshirani, R. (2003). Statistical significance for genomewide studies.Proceedings of the National Academy of Sciences of the United States of America,100, 9440–9445.
Tzschentke, T. M., & Schmidt, W. J. (2000). Functional relationship among medialprefrontal cortex, nucleus accumbens, and ventral tegmental area inlocomotion and reward. Critical Reviews in Neurobiology, 14, 131–142.
Warburton, E. C., & Brown, M. W. (2010). Findings from animals concerning wheninteractions between perirhinal cortex, hippocampus and medial prefrontalcortex are necessary for recognition memory. Neuropsychologia, 48, 2262–2272.
Wright, R. L., & Conrad, C. D. (2008). Enriched environment prevents chronic stress-induced spatial learning and memory deficits. Behavioural Brain Research, 187,41–47.
Yau, S. Y., Lau, B. W., & So, K. F. (2011a). Adult hippocampal neurogenesis: A possibleway how physical exercise counteracts stress. Cell Transplantation, 20, 99–111.
Yau, S. Y., Lau, B. W., Tong, J. B., Wong, R., Ching, Y. P., Qiu, G., et al. (2011b).Hippocampal neurogenesis and dendritic plasticity support running-improvedspatial learning and depression-like behaviour in stressed rats. PLoS ONE, 6,e24263.
Yau, S. Y., Lau, B. W., Zhang, E. D., Lee, J. C., Li, A., Lee, T. M., et al. (2012). Effects ofvoluntary running on plasma levels of neurotrophins, hippocampal cellproliferation and learning and memory in stressed rats. Neuroscience, 222,289–301.

E. Castilla-Ortega et al. / Neurobiology of Learning and Memory 109 (2014) 62–73 73
Yun, J., Koike, H., Ibi, D., Toth, E., Mizoguchi, H., Nitta, A., et al. (2010). Chronicrestraint stress impairs neurogenesis and hippocampus-dependent fearmemory in mice: Possible involvement of a brain-specific transcription factorNpas4. Journal of Neurochemistry, 114, 1840–1851.
Zheng, H., Liu, Y., Li, W., Yang, B., Chen, D., Wang, X., et al. (2006). Beneficial effectsof exercise and its molecular mechanisms on depression in rats. BehaviouralBrain Research, 168, 47–55.