virulence, immunity, and vaccine related to streptococcus pneumoniae
TRANSCRIPT

Critical Reviews in Microbiology, 18(2):89-114 (1991)
Virulence, Immunity, and Vaccine Related to Streptococcus pneumoniae
Chi-Jen Lee, Susan D. Banks, and Jean P. Li Center for Biologics Evaluation and Research, 8800 Rockville Pike, Bethesda, MD 20892
ABSTRACT: The pathogenesis of bacterial infection involves a series of interactions between the virulence determinants of the microorganisms and the immunity of the host. Studies on the molecular structure and immunological properties of pneumococcal virulence factors have provided general knowledge for the chemical basis of immunogenicity and prevention of bacterial infection. Antibody responses to PS and protein antigens can be greatly affected by their physicochemical properties, e.g., molecular size, specific determinants, con- formation, etc.
Characterization of group 19 pneumolysins and cloning of their ply genes were studied to examine the relationship of ply to virulence. Group 19 pneumococci all contained ply; the disease-isolated types of 19F and 19A appeared to show a higher specific hemolytic activity and yield than the nonpathogenic types, 19B and 19C. Genomic DNA that contained the ply gene from group 19 strains were analyzed by the polymerase chain reaction (PCR). Type 2 oligonucleotide primers recognized and initiated synthesis of an identical 1.5 kb DNA fragment in types 2, 19F, 19A, 19B, and 19C. Their sizes of restriction DNA fragments were also found to be homologous. Thus, group 19 ply genes showed remarkably similar characteristics. A difficult problem in the development of vaccines against bacterial diseases is the poor immune response of young children to purified PSs. The efficacy of pneumococcal vaccine might be improved by supplementation with inactivated pneumolysin in the form of a PS-protein conjugate.
KEY WORDS: virulence, immunity, pneumococcal vaccine, pneumococcus, immunity to polysaccharides, group 19 pneumolysins, pneumolysin genes.
1. INTRODUCTION
Many major contributions to the biochem- istry, immunology, and mechanisms of infec- tious diseases in the 20th century have resulted from the studies of the pneumococcal polysac- charides (PSs). These contributions include (1) an understanding of the specificity of polysac- charide antigens for antibody production, (2) the chemical basis of the immune response, and (3) host defense mechanisms against bacterial infec- tion. The development of a pneumococcal vac- cine represents a great achievement for basic medicine in promoting human health. Study of the pneumococcal vaccine provides an important method for characterizing virulence, disease pathogenesis, chemical structure, and regulation of the immune response to capsular polysaccharides.
Pneumococcal infection persists as a major cause of pneumonia, meningitis, and otitis me- dia. These diseases are caused by Streptococcus pneumoniae and remain some of the most prev- alent and serious infections in the U.S. and other areas of the world, particularly in young children, the elderly, and persons with certain high-risk conditions, e. g . , immunodeficiencies. I Pneu- mococcal pneumonia accounts for 10 to 25% of all pneumonias and an estimated 40,000 deaths annually.* Table 1 shows the epidemiology of pneumococcal diseases in the U.S. and Asia.
The course of pneumococcal bacteremia in patients treated either with type-specific anti- serum or with antibiotics has been found to be the same regardless of the therapy used in the first 5 days after onset of i l l n e ~ s . ~ . ~ These factors, including the high mortality and morbidity, en- demic and epidemic diseases, and emergence of
1040-84 1W9 I/. $50 0 1991 by CRC Press, Inc.
89
Cri
tical
Rev
iew
s in
Mic
robi
olog
y D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y C
DL
-UC
San
Die
go o
n 03
/15/
13Fo
r pe
rson
al u
se o
nly.

TABLE 1 Epidemiology of Pneumococcal Diseases
Number of
Mortality (%) Other
cases
4,137 (33.2%)
U.S. in 1989
Bacteremia Pneumonia
15-19 per 150,000-
40 5 >65 years: 50 c2 years: 160
100,000 570,000lyear
Asia 4.699 (37.8%)
3.610 (29.02)
47 83
bacterial strains resistant to various antibiotics and antimicrobial agents, have emphasized the importance of prevention and control of pneu- mococcal diseases.
There are 83 serologically defined pneumo- coccal types. Two nomenclature systems have been used to classify these types based on the PSs. The Danish system combined closely related types into groups. In the U.S. system, each type was chronologically assigned a number in the order of their recognition. In order to reduce the incidence of pneumococcal disease, a polyvalent pneumococcal vaccine was developed and li- censed in the U.S. The current 23-valent vaccine
Meningitis Otitis media
1-2 per >1 case in
55 0 100,000 first 5 years
was formulated based upon the prevalence of types isolated mainly in the U.S. and Europe. Re- cently, the distributions of pneumococcal disease isolate types in Asia have been investigated (Ta- ble 2). These studies in Asian populations showed that the types included in the 23-valent pneu- mococcal vaccine were considerably fewer in Asia than that observed in the U.S.: the proportion of these types was 62.9% in T a i ~ a n , ~ 72.9% in Japan,6 79.9% in China' as compared to 88.2% in the U.S.* A large number of pneumococcal types found in Asia, such as 15A, 23A, 24F, 27, and 34, are not present in the pneumococcal vac- cine. It may be necessary to develop a supple-
90
Cri
tical
Rev
iew
s in
Mic
robi
olog
y D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y C
DL
-UC
San
Die
go o
n 03
/15/
13Fo
r pe
rson
al u
se o
nly.

TABLE 2 Distribution of Pneumococcal Isolate Types in US. and Asia
Vaccine type (No. of Isolates)
Type
3 19F 23F 6B 14
11A 19A 9v
22F 18C 4
158 9N
1 OA 7F
12F 20 1 5
33F 8
17F 2
Japan
75 55 40 35 29 24 22 21 18 15 14 13 12 9 8 8 8 7 7 6 2 2 -
430 (72.9%)
Taiwan'
10 13 12 12 17 5 5 1
2 6
6 2 3
2 5 1
3 2
-
-
-
-
-
107 (62.9%)
China
25 41 33 26 34 4
7 1 8 5 8
5 11 12 3
44 63 11 9 1
35
385 (79.9%)
U.S. i? Europe (rank)
934 (5) 611 (10) 723 (8)
101 2b (4) 1282 (1) 131 (21) 347 (13) 438 (11) 222 (16) 703 (9)
1127 (2) 13gb (19) 296 (14) 132 (20)
413 (12) 191 (17)
1026 (3) 281 (15) 157 (18) 823 (6) 127 (22) 125 (23)
773 (7)
12,013 (88.2%)
Nonvaccine type (No. of isolates)
.Type
6A 15A 23A 34 24F 37 13 15C 31 16 28F 35c 35F 1 7A 38 36 180 21 29 35A 350 39 1 oc 1 OF 32F 25 32A 41 A 33c 28A Other 27 42
Japan
35 17 17 17 12 9 8 8 7 6 5 4 4 3 2
1 1 1 1 1
-
- - - - - - - - - -
160 (27.1 %)
Taiwan
4 5 3 5 2 1 5 1 4 1
- 3 2 .2 1
1 1 1 2 5 1 2 1
1 2 2 1
-
-
-
63 (37.1 )
China
26b 7b 4 1 1 2
3 7
2 1
6 4
2
1
9 16 3
63 (13.1%)
U.S. 81 Europe (rank)
- 105 (24) -
- - -
1154
1603 (1 1 .8%)
a The epidemiological survey of pneumococcal isolate types in Taiwan was performed by Drs. C. S. Yang, S. T. Chung, and C. J. Lee. The identification of types was done by Dr. R. Austrian. In China, 6A and 6B, 15A and 158 are divided, whereas in the US. these types are combined; in China 19F and 19A are combined.
mentary vaccine to include some additional types of pneumococcal PS antigens to be used for the Asian population.
Another population that is not fully protected by the vaccine are young children under 2 years of age. Their immune systems do not recognize
many PS antigens in the vaccine to allow pro- tective antibodies to be made against most en- capsulated b a ~ t e r i a . ~ Transplacental antibody from the mother is being explored to determine if it is protective and how early in gestation she should be immunized. New approaches to vac-
91
Cri
tical
Rev
iew
s in
Mic
robi
olog
y D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y C
DL
-UC
San
Die
go o
n 03
/15/
13Fo
r pe
rson
al u
se o
nly.

cine development should give rise to a vaccine that will be effective in immunizing young chil- dren against pneumococcal infection.
II. PNEUMOCOCCAL VIRULENCE FACTORS
A. Capsular Polysaccharide
Capsular PS is an essential virulence deter- minant of the pneumococcus. It was the first non- protein antigen found to induce antibody re- sponse in humans.’O The capsule is synthesized rapidly and extensively during the logarithmic phase of bacterial growth and PS antigens may be detected in the serum and urine of patients with pneumococcal disease. Pneumococcal vir- ulence and invasiveness vary among strains and depend on the composition and quantity of cap- sule produced. For example, type 3 and type 37 strains produce large amounts of capsule and show mucoid characteristics on a blood agar plate, but they differ in virulence to animals. Type 3, which is composed of a polymer of glucose and glu- curonic acid, is among the most invasive and virulent types, whereas type 37, with a homo- polymer of glucose, is rarely associated with pathogenesis. In experimental animals chal- lenged with capsular mutant strains of the same type but with different quantities of capsule, vir- ulence is found to be proportional to the amount of capsule produced.*
PSs are T-cell-independent antigens in that they can interact with B cells directly to initiate clonal expansion and antibody production. Stud- ies have shown that the predominant subclass of protective antibodies to PSs is IgG2 in humans and IgG3 in mice. IgG2 can cross the placenta and fix complement.
Although the capsule is a principal virulence factor, the mechanism of its activity in the host is not clearly understood. Capsular PS itself does not cause toxicity to animals and humans. Many capsular PSs are highly polar and hydrophilic and interfere with cell-to-cell interactions with phag- ocytes. The ability of host phagocytes to ingest and destroy invading bacteria requires coating of the organisms by antibodies and/or complement for opsonization. Antibodies bound to capsules
interfere with their charge and hydrophilicity and may act as bacteria-to-phagocyte ligands or com- plement activators. These PSs can interrupt the phagocytic activity elicited by activation of com- plement and opsonization by neutralizing the phagosome to prevent intracellular degrada- tion.12Js Although opsonization by antibodies and complement appears to be essential in host de- fense against encapsulated bacteria, under certain conditions both polymorphonuclear leukocytes (PMNs) and alveolar macrophages can use an opsonin-independent mechanism of phagocyto- sis. Furthermore, the opsonophagocytosis of type 1 pneumococci in vitro correlated with nei- ther antibody concentration nor with mouse pro- tection, whereas phagocytic killing of type 3 cells could be correlated with the concentration of an- tibody to type 3 PS (155). Complement receptors play a role in the B cell’s response to PSs. Results of a recent study indicate that PSs are able to activate complement via the alternative pathway and that C3d-bound PSs can interact with B cells. This mechanism is an important finding because infants have decreased cell surface expression of complement receptor 2 (CR2) on their B cells. This supports findings that a newborn’s B cells are too immature to respond to PSS.”~
Studies on the chemical structure and im- munological properties of pneumococcal PSs have provided the knowledge for the chemical basis of immunogenicity to many bacterial PSs. Most pneumococcal PSs are negatively charged and possess acidic components, including D-glucu- ronic acid (e.g. , types 1, 2, 3, 5, 8, 9A, and 9V), and phosphate in phosphodiester bonds (e.g., types 6A, 6B, 11A, 15F, 19F, 19A, and 23F). Type 4 PS contains pyruvate as its acidic com- ponent, while type 12F contains 2-acetamido-2- deoxy-D-mannuronic acid as the acidic compo- nent. The structures of all PS types contained in the 23-valent vaccine have been elucidated (Ta- ble 3).
Extensive immunologic cross-reactivity has been observed among pneumococcal capsular PSs. The biochemical basis of immunogenicity for pneumococcal group 9 and 19 PSs has been studied in animals as well as in Pneumococcal group 9 includes types 9N, 9A, 9L, and 9V. Types 9N and 9V comprise about 90% of the group 9 disease isolates. Only the
92
Cri
tical
Rev
iew
s in
Mic
robi
olog
y D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y C
DL
-UC
San
Die
go o
n 03
/15/
13Fo
r pe
rson
al u
se o
nly.

TABLE 3 Chemical Structures of Pneumococcal Polysaccharides
w Chemical structure Reference
AAT = 2-acetamido-4-amino-2,4,6,-trideoxy-D-galactose
4 +4)BD-ManpNAc(l~3)aGFucpNAc(l+3)o~alpNAc(1+4)aDCalp(l-. 3 x 2
H3C COOH (127)
5 +4)BD-Glcp(1+4)aL-~cpNAc(l+3)BD-Sugp(l+ 4 t 1
oGpnepNAc(l+4)&I+GlcpA
Sug - 2-acetamido-2,6,-dideoxy-D-xylohexos-4-ulose (128)
GpneuNAc - 2-acetamido-2,6,-dideoxy-Gtalooc (pneumosamine)
6A +P)aD-Galp( 1+3)aDClcp (1+3)oGRhap (1+3)Ribitol( 5-PO4- (129)
63 *2)oD-Calp(l+3)aD-Glcp( 1+3)oGRhap(l+4)Ribitol( 5-PO4- (130)
7F +6)oHalp (1+3)BGRhap (1+4)BIw;lcp (1+3)BDCalpNAc (l-. 2 2 4
t 1
t i 1 AcO
-alp aIbGlcpNAc(1+2)oGRhap (131)
8 44 ) BD-GlcpA ( 1+4) W l c p ( 144 ) oD-Glcp ( 1+4) oD4alp ( 14
9N ~4)aD-GlcpA(1+3)aD-Glcp(l+3)BD-ManpNAc(1+4)BD-Clcp(l+4)oD-
ClcpNAc (1+ (133)
93
Cri
tical
Rev
iew
s in
Mic
robi
olog
y D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y C
DL
-UC
San
Die
go o
n 03
/15/
13Fo
r pe
rson
al u
se o
nly.

TABLE 3 (continued) Chemical Structures of Pneumococcal Polysaccharides
94
Chemical structure Reference
9v 4 aDClcpA ( 1+3 ) aD-Calp ( 143) SbbnpNAc ( 1+4) BD-Glcp ( 1-4) aD-Clcp ( 1..
OAc OAc (134) I I
1OA ~S)~alf(l43)BDCalp(144)
BDCalp 1 t 6
3 t 1
~ a l p N A c ( 1 + 3 ) a ~ a l p ( 1 + 2 ) - D - R i b i t o l ( 5 - P 0 4 -
SD-Galf
OAc I 2/3
4 t
1I.A ~3)SD-Galp(l4)BD-Glcp(l+6)aD-Glcp (1+4)aD-Galp(l*
P04-l-Clycerol (136)
12F ~ 4 ) a G ~ c p N A c ( l + 3 ) B D a l p N A c ( l ~ ) ~ ~ n p N A c ( l ~ 3 3 t t 1 1
a M a l p aD-Glcp (1+2)aD-Clcp
14 -.4)BD-Clcp(l.*6)BlcpNAc(1+3)BD-Calp(l+ 4 t 1
M a l p
15B ~6)BDclcpNAc(l*3)SD4alp(l~4)&bGlcp(l-, 4 t 1
aPGalp(l~Z)BDCalp-3-P04-(CH2CHN+r(s3)0.2
OAc I 2
4 t 1
BbRhap (1+4)aCRhap (1+2)bArabinitol(l-P04-
aD-Galp (140)
Cri
tical
Rev
iew
s in
Mic
robi
olog
y D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y C
DL
-UC
San
Die
go o
n 03
/15/
13Fo
r pe
rson
al u
se o
nly.

TABLE 3 (continued) Chemical Structures of Pneumococcal Polysaccharides
IYJE Chemical structure Reference
18C oD-Glcp(l-4)aD-Galp(l+4)~lcp(l+3)aL-Rhap(l+l)-glycerol 2 (141 1 t 1
D-Glcp I OAf
19A - + 4 ) ~ ~ ( 1 + 4 ) a D - C l c p ( l + 2 ) a L A h a p ( 1 - P ~ - 2 3 t t PO4 p04 t t 1 1
BD-Gal a k h c 3 t 1
BD-G 1 cNAc
19F +4)SD-ManpNAc(l4)aDClcp(1+2)aGRhap(1-P~- (143)
20 + 6 ) a D 4 l c p ( l + 6 ) B D - G 1 c p ( l ~ 3 ) ~ a l f ( l ~ 3 ) & ~ l c p ( l + 3 )
PDGlcpNAc (1-PO4 4 t 1
BD-Galf (144)
BD-Glcp 1 t 3
2 22F -4)BIK;lcpA(14)&LAhap(l+4)aD-Glcp(l+3)aD-Galf(l+2)
I OAc
aGRhap ( 1+ (145)
95
Cri
tical
Rev
iew
s in
Mic
robi
olog
y D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y C
DL
-UC
San
Die
go o
n 03
/15/
13Fo
r pe
rson
al u
se o
nly.

TABLE 3 (continued) Chemical Structures of Pneumococcal Polysaccharides
ZYF Chemical structure
aGRhap 1 t 2
3 t P04-2-glycerol
23F 4)BDGlcp(1+4)BDCalp(1~4)BGRhap(l-.
33F +3)BDCalp(1+3)
Reference
o D - C a l p ( l - 3 ) ~ a l / ( 1 - 3 ) B D C l c p ( l + 5 ) B D - G a l f ( l ~ 2 2 t I 1 (OAc ) 0 .4
aD4alp
type 9N PS was contained in the 14-valent vac- cine. Type 9V is the most common group 9 dis- ease isolate among young children and also the most common group 9 isolate associated with vaccine failures. A clinical study on the antibody response to group 9 PSs in adults showed that type 9N PS stimulated a high homologous anti- body response against 9N PS but induced low antibody levels to 9V and 9L PSs. Type 9V PS monovalent vaccine also showed a poor response to induce cross-reactive antibodies. In contrast, a bivalent vaccine containing 9N and 9V PSs, or trivalent vaccine containing 9N, 9V, and 9L PSs, produced high antibody response to all of the group 9 types, and especially to type 9V.I6.l7 Studies on antibody response to group 19 PSs were also performed in humans. It was observed that immunization with one type of group 19 PS induced a poor antibody response to the other type. Immunization with both 19F and 19A PSs induced antibodies as high as that observed in the response to individuals receiving the homol- ogous PS.'8.'9 These results led to a recommen- dation to include both 9N and 9V, as well as 19F and 19A, PSs in the new 23-valent vaccine. Ex- tensive cross-reactions have also been found be- tween pneumococcal PSs and PSs from other bac- teria, such as Klebsiella and nongroupable streptococci. Klebsiella K2 PS produced cross-
(147)
reactive antibody to 19F PS and also enhanced the magnitude of the antibody response to 19F PS ,20.21 whereas streptococcal 14636174 strain showed an identical structure to that of 19F PS.22
B. C-Polysaccharide
C-polysaccharide (C-PS) is the major cell- wall component of the pneumococcus. It is co- valently linked to the peptidoglycan probably through the muramic acid r e s i d ~ e ~ ~ . ~ ~ and uni- formly distributed on both the inside and outside of the cell walls. The thickness of the C-PS varies with the strain. Encapsulated strains are covered by varied amounts of capsular PS concealing the C-PS of the bacteria so as to render it inaccessible to anti-C-PS antibodies." The structure of the repeating unit of type 1 C-PS is shown as: p-D-
GalNH p- 1 - 1 '-ribitol-5-phosphate, where AAT- Gal is 2-acetamido-4-amino-2,4,6-trideoxyga- l a c t o ~ e . ~ ~ . ~ ' C-PS can activate the alternative complement pathway, and its antibody response in mice and humans is dominated by the phos- phorylcholine portion of the molecule. Phos- phorylcholine is a growth factor for pneumo- cocci. The bacteria continue to grow when choline is substituted by ethanolamine; however, certain
Glup- 1 -ct-AAT-Galp 1 ~ c Y - D - G ~ N A c ~ 1 - 3 - p - ~
96
Cri
tical
Rev
iew
s in
Mic
robi
olog
y D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y C
DL
-UC
San
Die
go o
n 03
/15/
13Fo
r pe
rson
al u
se o
nly.

physiological defects, including inability to undergo genetic transformation and resistance to phages, soon occur. Furthermore, the inability for cell division arises from the resistance of the ethanolomine-containing teichoic acid to pneu- mococcal N-acetylmuramic acid-L-alanine ami- dase, so that long chains of cells are formed.28 Interaction with choline residues is essential for the enzymatic hydrolysis of the amide bond be- tween N-acetyl muramic acid and L-alanine. 29
Since C-PS is common to almost all pneu- ~ o c o c c ~ , ~ ~ , ~ ~ it has been considered that anti- bodies against this antigen might confer specific protection to pneumococcal infection. 32,33 Both monoclonal and polyclonal antibodies to phos- phorylcholine have been reported to protect against a lethal challenge with some types of p n e u m o c o ~ c i . ~ ~ ~ ~ ~ Among the different idiotypes of phosphorylcholine (PC) antibodies, T15 was shown to be more protective than the other M603 and M511 antibodies, and of these, IgG had higher protective activity than IgM antibodie~.’~ The mechanism of this protection was considered to involve enhancement of opsoni~ation.~~ Types 3 and 5 formed precipitation lines by immunodif- fusion when reacted with PC-binding IgA mon- oclonal antibodies (T15 and H8) in addition to their type-specific reactions. However, they did not react with PC-binding IgM antibody (HYPC1613) and no protection was conferred by this IgM antibody against types 1, 3, and 5. Both the isotype and reactivity with the cell wall C-PS of encapsulated pneumococci are important in inducing the protective antibody of anti-PC antibodies. Moreover, the protective activity of PC-binding antibodies is related both to their ad- ditional specificity for the sugar backbone of the C-PS and to their secondary biological activities such as complement activation. 34*157 Further- more, the antibodies to the C-PS induced by the C-PS conjugated to bovine serum albumin (BSA) also failed to protect mice against challenge with type 3 or type 6A pneurnoc~cci.~~
Antibody to PC appears in the bloodstream in the first few years of life,37 especially in re- sponse to pneumococcal infection, and is de- tectable in almost all healthy children and adults.38 However, the naturally present C-PS IgG anti- body does not protect against the evolution of acute pneumococcal infection from colonization
to acute purulent bronchitis, from bronchitis to pneumonia, or from pneumonia to bacter- emia.39v40 Thus, current evidence indicates that antibodies to C-PS are not protective against pneumococcal infection.
C. F Antigen
The F (Forssman) antigen is found to be uni- formly distributed in the pneumococcal plasma membranes and located with the C-PS parts of the molecules exposed on the surface and the lipid part anchored in the lipid bilayers of the plasma membrane^.^.^^.^' The lipid part is a mix- ture of eight fatty acids; one of which appears to be hydroxyacid. Lipid makes up 5.7% of the molecular weight of the antigen and 15% of cho- line incorporated into the organism may be found in the F antigen.” F antigen is the functional equivalent of the membrane lipoteichoic acid which contains a poly-glycerophosphate back- bone.42 It is a strong inhibitor of pneumococcal autolysin and a regulator of mureine hydrolase activity. All pneumococci are prone to lysis dur- ing the stationary phase of growth and in that state F antigen is released. Thus, the loss of this regulatory antigen results in the unrestrained ac- tivity of the N-acetylmuramic acid-L-alanine ami- dase causing cell wall degradati~n.~’
D. Protein Antigens and Toxins
1. Pneumolysin
Pneumolysin (ply), a thiol-activated cytolytic toxin produced by many pneumococcal disease isolates, is considered to be involved in the path- ogenicity of pneumococcal infection by evading the defense mechanism of the host resistance. It binds to eukaryotic membranes through the cho- lesterol receptor and is thought to oligomerize to form transmembrane channels that lead to the lysis of cell^.^,^^ Low doses of purified ply in- hibit the respiratory burst, chemotaxis, and an- timicrobial activities of human PMN leukocytes and macro phage^.^.^^ They also inhibit the pro- liferative response of human lymphocytes to mi- t ~ g e n . ~ ~ Higher toxin doses cause activation of
97
Cri
tical
Rev
iew
s in
Mic
robi
olog
y D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y C
DL
-UC
San
Die
go o
n 03
/15/
13Fo
r pe
rson
al u
se o
nly.

the classical complement pathway and depletion of serum opsonic activity.49 Furthermore, in- jecting mice with inactivated ply increased their survival time, after challenge with virulent pneu- mococci 2 weeks later.5o Thus, ply plays an im- portant role in the mechanism of pneumococcal virulence.
The sulfhydryl-activated toxins require re- duction for maximal a~tivity;~' this indicates that cysteine residues in the proteins are crucial for activity. These toxins are irreversibly inactivated by c h o l e ~ t e r o l . ~ ~ - ~ ~ Cholesterol has been consid- ered to serve as the membrane receptor for pneu- molysin because these toxins are lytic only for cells with cholesterol in their membranes and the inhibition by cholesterol shares many character- istics of the toxin-erythrocyte intera~tion.~~
Pneumolysin prepared from type 2 strain has a molecular weight of 53,000 with 471 amino acid residues. Its isoelectric point is 4.9 to 5.2. Pneumolysin has no obvious signal sequence as- sociated with the N-terminus of the protein. This may be the reason why pneumolysin is not se- creted from pneumococci into the culture medium.
The relationship between the structure and function of pneumolysin has been studied by the molecular cloning and expression of types 1 and 2 pneumolysin genes. -vS6 Moreover, the prepa- ration of pneumolysin gene-modified mutants and the site-directed mutagenesis on single cysteine residue of pneumolysin were carried out to ex- amine the changes in hemolytic activity and in- volvement of pneumolysin in virulence. The Cys- 428+Ser and Cys-4284Gly modified toxins showed reduced hemolytic activity on erythro- cytes and PMN leukocytes, 6 and 20 times less active than the wild type, respecti~ely.~~
Characterization of group 19 pneumolysins and cloning of these pneumolysin genes were studied to examine the relationship of pneumo- lysin to virulence. Group 19 pneumococci con- tain 4 types; types 19F and 19A account for 99% of all diseases caused by group 19 strains, whereas 19B and 19C are nonpathogenic for humans and mice. The pneumolysin protein was isolated from types 19F, 19A, 19B, and 19C strains. The so- dium deoxycholate lysate of pneumococcal cells was precipitated with 70% ammonium sulfate, chromatographed on DEAE-cellulose, and puri- fied by SDS polyacrylamide gel electrophoresis (Table 4A; Figure 1). Group 19 pneumococci all
contained pneumolysin; the disease isolate types 19F and 19A showed higher specific hemolytic activity and yield than the nonpathogenic types 19B and 19C (Table 4B).
The isolation and molecular cloning of pneu- molysin (ply) gene from 19F chromosomal DNA were performed, using clonal ply genes from two different pneumococcal strains as probes. A 5- kb fragment of 19F DNA was isolated and ligated into a ClaI site of a phagemid vector derived from pUC 19, pks( - ), and cloned into Escherichia coli DH5a. The transformed DH5a cells containing 19F 5-kb DNA expressed high pneumolysin ac- tivity." Genomic DNA that contained the ply gene from 19F, 19A, 19B, and 19C strains were analyzed by the polymerase chain reaction (PCR) technique. Oligonucleotides (30-mer) were pre- pared to the known DNA sequence of type 2's 5' and 3' termini. These oligonucleotides found homologous sequences and generated a 1.5-kb PCR product in all four type 19 isolates. They were also found to be similar in their restriction endonuclease digestion patterns. These group 19 ply genes showed remarkably similar character- istics (Figure 2). It is interesting to note the com- parison of ply genes and proteins from pneu- mococcal types 1 and 2 strains. The results showed that only 6 differences in the two genes were observed in 1650 bp of sequence, whereas only a single amino acid change was present.58
Pneumolysin is one of the leading proteins involved in the pneumococcal virulence. It also produces a strong protective antibody titer, in an inactivated form, in mice. Thus, the efficacy of the pneumococcal vaccine might be improved by supplement with inactivated pneumolysin in the form of a PS-protein ~onjugate."~
2. Protein A
Pneumococcal surface protein A (PspA) has been found on the cell surface of many strains. PspA from noncapsulated strain R36A has a mo- lecular weight of 84,000, with multiple antigenic determinants. PspA from different pneumococcal isolates varies both in its expression of these an- tigenic determinants and in its molecular weight. While it is not required for genetic transfonna- tion, its absence from the cell surface may affect the frequency of transformati~n.~~ Studies with
98
Cri
tical
Rev
iew
s in
Mic
robi
olog
y D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y C
DL
-UC
San
Die
go o
n 03
/15/
13Fo
r pe
rson
al u
se o
nly.

TABLE 4A Isolation and Purification of Type 19F Pneumolysin
Hemolytic activitv
Protein (HU/mg (Total Fold increase Purification (mg) protein)’ x 103U) (sp. activity)
Crude extract 21 8 340 79 too 1 Ammonium sulfate 72 820 59 80.0 2.4
DEAE chromatography, fr. 0.65 33,500 22 29.7 98.0
Polyacrylamide 0.28 52,000 15 20.3 153
precipitation
A
electrophoresis (0.13%)
Note: A lyophilized type 19F strain was reconstituted in sterile saline, inoculated on sheep blood agar and incubated at 3PC in humidified 5% CO, for 18 h. Colonies were transferred to 1 I of special brain heart infusion (BHI) medium (66) and incubated at 37°C for 18 h. The cells were lysed by sodium deoxycholate at the final concentration of 0.1% and extracted with pH 7 sodium phosphate buffer. The protein fraction of extract was concentrated by 70% ammonium sulfate precipitation. The precipitate was dissolved and dialyzed in 50 mM sodium phosphate solution. The protein fraction was then passed through DEAE- cellulose column (1.5 x 90 cm) and eluted with 50 mM phosphate pH 7 buffer, followed by 50 to 250 mM phosphate gradient. The fractions were assayed for hemolytic activity. In this assay procedure, 0.1 -ml sample was added to 30 mM of dithiothreitol and 1 ml of 2% washed sheep erythrocytes. The mixture was incubated for 20 min at 37% and centrifuged. The optical density of supernatant was measured at 541 nm. The fractions that exhibited the hemolytic activity, were concentrated by the ultrafiltration membrane and purified by SDS-polyacrylamide gel electrophoresis.’* The major protein band was eluted by an electroseparation system, carried out for 17 h at 4°C with a current of 100 V.
One hemolytic unit (HU) is defined as the amount of pneumolysin that causes 50% hemolysis on 1 ml of 1% sheep erythrocytes when incubated at 37°C for 20 min.
TABLE 4B Preparation of Group 19 Pneumolysins
Hemolvtic activitv Protein’ (HU/mg
Group (mg) protein) (Total x 10” U) (%)”
19F 0.65 33,500 22.0 29.7 19A 0.68 27,540 19.0 25.0 196 0.48 15,000 7.2 12.0 19c 0.12 19,160 2.3 4.0
Note: The experimental conditions for preparation of types 19A, 19B, and 19C pneumolysins were the same as that described in Table 4A.
Obtained from DEAE chromatography, fraction A. Proportion for the hemolytic activity of purified pneu- molysin from each group to their total hemolytic ac- tivity of crude extract.
monoclonal antibodies against this protein have shown that passive immunization may be pro-
tective. Furthermore, PspA is able to elicit a protective immunity in the partially immunode- ficient xid mouse that cannot respond to poly- saccharide antigens. PspA-mutant pneumococci resulted in a small but significant increase in the survival of challenged mice when compared with those challenged with the parental PspA + strain.6o These results indicate that PspA could be a contributing factor to the pneumococcal virulence.
3. Neuraminidase
Pneumococcal neuraminidase causes injury to the host by cleaving terminal sialic acid resi- dues from glycoproteins, glycolipids and oligo- saccharides on cell surfaces and in body fluids. The elevated concentrations of free N-acetyl-neu- raminic acid in the cerebrospinal fluid of patients
99
Cri
tical
Rev
iew
s in
Mic
robi
olog
y D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y C
DL
-UC
San
Die
go o
n 03
/15/
13Fo
r pe
rson
al u
se o
nly.

PAGE Electrophoresis of Group 19 Pneumolysin
FIGURE 1. Group 19 pneumolysin samples were analyzed by SDS-poly- acrylamide gel electroph~resis.'~' The purified types 19F, 19A, 198, and 19C pneumolysins showed single bands in electrophoretic pattern, each having the molecular weight of 53,000.
with pneumococcal meningitis correlate with coma and an adverse prognosis. Pneumococcal neuraminidase has a molecular weight of 107 kDa, which is predominantly associated with the cell wall. During various purification procedures, it can be degraded to the molecular weight of 86 kDa form. Degradation of neuraminidase was in- hibited by protease inhibitors.61 The significance of this enzyme in pneumococcal virulence has
not been well understood. It may be of less value than pneumolysin as a vaccine component.62
4. Aufolytic Enzyme
The major autolytic enzyme (autolysin) in pneumococcus is postulated as N-acetylmuramic acid-L-alanine amidase, which splits the bond be-
100
Cri
tical
Rev
iew
s in
Mic
robi
olog
y D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y C
DL
-UC
San
Die
go o
n 03
/15/
13Fo
r pe
rson
al u
se o
nly.

tween N-acetylmuramic acid and alanine in the mucopeptide portion of the cell wall. The lysis of pneumococci by sodium deoxycholate oc- curred through the hydrolysis of this enzyme.63 The autolytic enzyme seemed indispensable for the insertion of newly synthesized p-1,4,-N-ace- tyl glucosaminyl-N-acetyl muramyl peptide frag- ments into the cell wall peptidoglycan.a It might play a role in the bacteriolytic action of the an- tibiotics which inhibited cell wall bio~ynthesis .~~ Some functions of autolytic enzyme might de- pend on the specific enzymes, such as amidase and glycosidases contained in it;" muramidase would provide the receptor site for linking new disaccharide-peptide units into a growing mu- copeptide whereas a muramyl-L-alan- ine amidase might be required for the synthesis of cross linkage between muramic acid and peptide.68
Autolytic enzymes are reported to be in- volved in processes such as daughter cell sepa- ration, genetic transformation, and the inever- sible effects caused by treatment of the bacterial cells with p-lactam antibiotic^.^^ The deletion of the autolysin gene (lytA) in pneumococcal cells has resulted in the formation of small chains (six to eight cells), absence of lysis in the stationary phase of growth, and tolerant response to peni- illi in.^' Thus, autolytic enzymes have been thought to play important roles in several basic biological functions in bacteria.
5. Neutrophil Elastase Inhibitor
Pneumococci have produced two types of neutrophil elastase inhibitors. One is a negatively charged substance of low molecular weight that depends on an ionic interaction with the cationic elastase. The second inhibitor is located on the outside of the cell membrane, having a molecular weight of 140 kDa. The latter inhibitor inhibits elastase noncompetitively and specifically and is not inhibited by sodium chloride. It does not in- hibit other elastases or other neutrophil en- z y m e ~ . ~ ' It is sensitive to trypsin, and the cleav- age products are biologically active, protecting mice from alveolar damage by neutrophil-granule extracts .72
E. Others
Purpura-producing principle, a component derived from pneumococcal cell wall, is capable of producing purpura in mice, rabbits, and guinea pigs. It is a complex of peptidoglycan and tei- choic acid, having a molecular weight of 2.6 X
lo7. It is produced by the activity of autolysin on the cell wall.73
Many encapsulated pathogens, including pneumococcus, Haemophilus influenzae type b, and meningococcus, have produced a human IgA 1 -specific protease. 74-76 Hydrolysis of IgAl by the protease occurs in the hinge region of the immunoglobulin, at the site located in one half of the duplicated octapeptide r e g i ~ n . ~ ~ . ~ ~ IgAl protease may contribute to bacterial virulence through the activity as an aid to bacterial attach- ment to the tissue and helping the infecting agent to degrade IgA antibodies. Attempts have been made to clone the pneumococcal IgAl protease gene. 79
111. GENETIC REGULATION OF VIRULENCE
The pathogenic bacteria that cause death of the host have been considered as the most viru- lent. However, the death of the host exerts a negative effect on multiplication, dissemination, and survival of the microbes in nature. Thus, the regulation and expression of virulence determi- nants play a role in maintaining the life cycle of the pathogenic organisms to optimize the survival within the individual host, as well as within the population as a whole. The virulence genes in pathogenic bacteria are regulated and expressed in various stages so that organisms at one stage exhibit a phenotype different from that of organ- isms at another stage. Such genetic regulation of virulent determinants can provide benefits to the organisms by promoting survival within the ex- isting environment or prime the pathogens for exploring a new host environment.80
The respiratory pathogen Bordetella pertus- sis causes the disease whooping cough through the activities of several virulence factors includ- ing pertussis toxin, filamentous hemagglutinin,
101
Cri
tical
Rev
iew
s in
Mic
robi
olog
y D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y C
DL
-UC
San
Die
go o
n 03
/15/
13Fo
r pe
rson
al u
se o
nly.

A
FIGURE 2. Polymerase chain reaction (PCR) for generation of homologous DNA frag- ments were performed, using 30-mer oligonucleotides to the type 2 pneumolysin gene as primers. Approximately 1 ng of genomic DNA from types 19F, 19A, 19B, and 19C were combined with the type 2 oligonucleotides, dNTPs, pH 8.0, Tris-HCI buffer, and Taq polymerase (DNA amplification reagent kit, U.S. Biochemical Corporation, Cleve- land, OH).140 The PCR Unit used was a Perkin Elmer Thermocycler (Perkin Elmer, Norwalk, CT). The cycles were run with a melting temperature of 94°C for 4.5 min, an annealing temperature of 37°C for 5 min, and a polymerization temperature of 72°C for 15 min. The cycle was repeated 25 times to amplify the PCR product. The PCR product was run through a Sizeselect 400 Sephacryl Spun column (Pharmacia, Piscataway, NJ) to remove unincorporated dNTPs, oligonucleotides, and early termination sequences. The concentration was measured by UV spectrophotometer and approximately 0.5 p,g of the PCR products were run on a 1% agarose gel (Figure 2A). Of this DNA, 1 to 2 p,g was digested with the restriction enzyme EcoRl and 0.5 bg was loaded on a 1 % agarose gel (Figure 2B). The results showed that type 2 oligonucleotides recognized and initiated synthesis of an identical 1.5-kb DNA fragment in type 2,19F, 19A, 19B, and 19C (Figure 2A). Their sizes of restriction fragments, 0.5- and 0.9-kb DNA, were also found to be homologous (Figure 28).”
hemolysin, and other toxins.*’ Expression of vir- ulence-associated factors is regulated by the vir
locus which codes for two proteins BvgA and BvgS, of 209 and 1238 amino acids, respec-
102
Cri
tical
Rev
iew
s in
Mic
robi
olog
y D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y C
DL
-UC
San
Die
go o
n 03
/15/
13Fo
r pe
rson
al u
se o
nly.

FIGURE 2B
t i ~ e l y . ~ ~ . ’ ~ ~ These two proteins show significant sequence similarity to “two-component” regu- latory systems in many bacterial species. The vir locus has been shown to be the site of mutations leading to the phenomenon known as phase-var- i a t i ~ n . ~ ~ . ’ ~ ’ The vir locus also appears to be re- sponsible for the regulation of virulence potential in response to environmental conditions. This comes from the observation that regulation of FHA synthesis in E. coli under the control of the vir locus responds to the signals of reduced tem- perature, and increased magnesium sulfate or ni- cotinic acid concentrations, as it does in B. pertussis. 84~152~153
H. influenzae type b produces a capsule con- sisting of a polymer of a ribose-ribitol 5-phos- phate repeating unit; it serves as a major deter- minant of virulence in this organism.85 The genes necessary for capsule production are located in
a locus cap, which contains a duplication of 17 to 18 kb interrupted by a bridge region of 1.3 kb.86*87 Cap- variants of wild-type isolates occur at high frequency and are associated with reso- lution of the repeated DNA in a rec-dependent p r o c e s ~ . ~ ~ . ~ ~ Such deletions remove the bridge region, which encodes the bexA gene (b capsule expression A), whose product is required for transport of the cap~u le .~ ’ .~~
Recently, the localization and molecular cloning of genes encoding the capsule of pneu- mococcal type 3 have been studied. Using di- gested, size-fractionated type 3 DNA, the inves- tigators have localized the type 3 capsule genes to an approximately 30-kb Aval fragment and an approximate 25-kb BamHI fragment.g0 More- over, using insertion of transposon Tn916, a pu- tative capsulation sequence in the genome of pneumococcal type 3 was interrupted and re-
103
Cri
tical
Rev
iew
s in
Mic
robi
olog
y D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y C
DL
-UC
San
Die
go o
n 03
/15/
13Fo
r pe
rson
al u
se o
nly.

sulted in the formation of several noncapsulated strain^.^' These results indicated that capsule syn- thesis of pneumococci was subjected to the ge- netic regulation of a specific gene. Mutation of such a gene could result in the formation of non- capsulated strains, thus transforming from viru- lent to nonvirulent organisms.
Other pneumococcal genes expressing neur- aminidase, IgA1 protease, or surface protein A have also been cloned in E. cokm Furthermore, the isolation and molecular cloning of pneumo- lysin (ply) gene from pneumococcal type 19F chromosomal DNA were performed. The trans- formed DH5a cells containing 19F 5-kb DNA expressed high pneumolysin activity. In addition, aply mutant gene was transformed into R61 rough strain by the method of insertion-duplication me- tagenesis. Several erythromycin-resistant, rough pneumococci were obtained. None of these cells produced pneum01ysi.n.~~ These results reveal that the ply genes are separate from other virulence factors within its genome.
IV. VACCINE DEVELOPMENT AND CONTROL
A. Immunity and Antibody Response
Most adult animal sera contain antibodies against PS antigens of various pathogenic bac- teria. Antibodies to pneumococcal type 3 and other types were detected in the animals as they grew or in human adults without possible contact with these organism^.^^-^^ The possible antigenic sources for these serum protective antibodies have been identified and found to be derived from cross-reactive antigens among intestinal and pha- ryngeal bacteria .%sg7
Antibody to capsule PS is considered to be essential for the survival of an animal challenged with virulent pneumococci. Human capsular PS antibodies have been shown to be homogeneous and exhibited both IgM and IgG2, kappa light- chain molecules. Deficient IgG2 antibody for- mation has been associated with severe recurrent diseases due to encapsulated bacteria.98 After im- munization with PS antigens there is a significant rise in total IgG, predominantly of IgG1 and IgG2 antibodies. It has been reported, that after vac-
cination with pneumococcal type 3 PS, the geo- metric mean titer of IgG2 was produced higher than those of IgG 1, IgG3, or IgG4. Only the IgG2 antibody level was significantly higher than that of preimmune concentration.w The protective an- tibodies induced by the pneumococcal PSs could
.be present at one fifth to one half or more of their maximum peak values 2 to 3 years after a single immunization. High levels of antibodies per- sisted in some individuals for 8 years after in- jection.'m*lO' The level of specific antibodies, greater than 250 to 300 ng antibody nitrogen per milliliter serum, is considered to be protective against invasive pneumococcal diseases. lo* It has been proposed that antibody response to type 3 PS is regulated extensively by two types of T cells, suppressor (Ts) and contrasuppressor cells (T'CS).~~~*~O" The actions of these regulatory T cells are well characterized for type 3 PS, but are not generally applicable to all PSs. It was first pro- posed that the antibody response to type 3 PS was controlled by Ts and amplifier T cells (Ta). Ts limit the extent to which antibody-forming B cells proliferate in response to antigen, whereas Ta drive B cells to multiply further after antigenic stimulation. In most cases, the effects of Ts and Ta are not apparent unless the activity of one is either eliminated or altered. Although both Ta and helper T cells (Th) are CD4 + CD8 - and have a positive influence on the antibody re- sponse, they differ in several ways:
1.
2.
3.
4.
Th participate in antigen presentation and act during the first 24 h after immunization; by contrast, Ta activity is not evident until 2 days after immunization and is maximal 4 to 5 days after immunization. Th activity disappears within 28 weeks after thymectomy or treatment with antilympho- cyte serum ( A L S ) ; Ta activity is not af- fected by such treatment. Th activity is governed by genes encoded by the major (H-2) histocompatibility com- plex, whereas Ta activity is not associated with H-2 type. Th activities are eliminated by treatment with hydrocortisone; this steroid has no effect on Ta activities.
Th act mainly to initiate antibody formation,
1 04
Cri
tical
Rev
iew
s in
Mic
robi
olog
y D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y C
DL
-UC
San
Die
go o
n 03
/15/
13Fo
r pe
rson
al u
se o
nly.

whereas Ta expand an ongoing antibody re- sponse.1s8 Ts are activated in response to the idi- otypic determinants of B-cell-associated antibody specific for type 3 PS. In response to type 3 PS stimulation, Ts release a soluble factor that binds to determinants on B cells rather than antigen. This factor acts directly on antigen-stimulated B cells or inhibits the induction of Ta. Ta can be induced and activated by exposure to immune B cells, with a specificity due to the ability of Ta to recognize the idiotypic determinants of B-cell- associated antibody specific for 3 PS. 159-161
Contrasuppressor Tcs are induced in mice after injection of type 3 PS-coupled spleen cells or optimally immunogenic dose of 3 PS (0.5 to 5 pg). These Tcs belong to a unique T-cell sub- set, having antigen-binding receptors and the phenotype Lyt-1+2 - L3T4- I-J+ I-A + T cells that are adherent to the lectin Vicia villosu, and sensitive to cyclophosphamide and ALS. Tcs function by preventing the expression of Ts ac- tivities and do not show any activity in the ab- sence of Ts. Tcs are functionally distinct from Ta; Tcs are unable to augment the antibody re- sponse to 3 PS, that is, their activity is dependent on the presence of Ts. In contrast, Ta can mark- edly augment the antibody response in the pres- ence or absence of Ts. Furthermore, Ta are re- sistant to cyclophosphamide and ALS. Tcs apparently mediate their effects by releasing a soluble factor. The interaction of Tcs with B cells or Ts is through an antigen bridge rather than an idiotype-anti-idiotype interaction. Thus, it is likely that Tcs function either by making type-specific B cells resistant to Ts signals or directly interact with Ts to neutralize their activity.104*105
B. Pneumococcal Vaccine and Immunization
The current pneumococcal vaccine is com- posed of purified capsular PSs of 23 types, in- cluding 1, 2, 3, 4, 5 , 6B, 7F, 8, 9N, 9V, 10A, 11A, 12F, 14, 15B, 17F, 18C, 19F, 19A, 20, 22F, 23F, and 33F. It was licensed in the U.S. in 1983, replacing a 14-valent vaccine licensed in 1977. The important events during vaccine development are shown in Table 5. Most healthy adults and elders induce a twofold or greater rise in type-specific antibody within 2 to 3 weeks of
TABLE 5 Important Events during the Development of Pneumococcal Vaccine
1881 : Pasteur and Sternberg discovered pneumo- coccus and isolated it in a culture
191 1 : Wright and associates conducted immuniza- tion studies in South Africa, using whole cell pneumococcal vaccine
heat-killed whole-bacteria pneumococcal vaccine
1930: Tillet and Francis found that purified pneumo- coccal capsular PSs induced specific antibody in humans
1 938: Felton showed type-specific disease protection in individuals injected with type 1 and 2 PSs; Ekwurzel and associates conducted the first clinical trials of the capsular PSs and found significant reduction in episodes of pneumonia and its mortality
1945: MacLeod and Heidelberger demonstrated that
191 6: Lister demonstrated the protective value of
1946:
1947:
1964:
1968:
1977:
1981:
1983:
type-specific pneumocoical infection in man could be prevented by a tetravalent vaccine, including types 1, 2, 4, and 7 PSs Commercial hexavalent pneumococcal vac- cines were produced; however, the widely available antibiotics for treatment of pneumo- coccal infections led to the decreased interest in the vaccination; these vaccines were with- drawn from the market. Kauffman, using a trivalent vaccine, showed type-specific protection against pneumonia and bacteremia in elderly individuals Austrian and Gold found that the continuing high mortality of pneumococcal pneumonia was prevalent even in the patients treated with antibiotics National Institutes of Health and Merck, Sharp & Dohme conducted the efficacy and safety studies of 6- and 13-valent pneumococcal vaccines; these studies showed an 82.3% re- duction in pneumococcal bacteremia for types included in the vaccine 14-valent pneumococcal vaccine was licensed by FDA Compensation on pneumococcal vaccination for elderly person by Medicare was initiated 23-valent pneumococcal vaccine was licensed by FDA
vaccination. In immunocompromised patients, the antibody response may be less. In children less than 2 years of age, antibody response to most capsular types is generally poor. Further- more, the response to some pediatric pneumo- coccal types, such as 6A and 14, is decreased in
105
Cri
tical
Rev
iew
s in
Mic
robi
olog
y D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y C
DL
-UC
San
Die
go o
n 03
/15/
13Fo
r pe
rson
al u
se o
nly.

children less than 5 years old.’.’06.107 The anti- body levels for most pneumococcal vaccine types remain elevated in healthy adults at least 5 years after immunization. It was reported that a group of less than 40% of recipients showed antibody levels of several types that were significantly above their baseline values 10 years after vaccination. lo8
The pneumococcal vaccine has been shown to be safe and effective in preventing pneumo- coccal bacteremia in young adults and elderly persons, 60 to 65 years and over. The 23-valent vaccine has an efficacy of 60 to 70% in pre- venting pneumococcal bacteremia in the el- derly. 109.110 Immunization should be recom- mended for all elderly persons because of the high morbidity and mortality of pneumococcal infection in this group; it should also be rec- ommended to all persons in high risk populations. Revaccination should be considered for adults at high risk who received the 23-valent vaccine more than 6 years before and for those shown to have rapid decline in antibody levels.1J11
New PS-protein conjugate vaccines have been developed and may become available in the near future.112-115 They will have the capacity to in- duce an enhanced T-cell-dependent antibody re- sponse and long-term immunologic memory. They may help in solving problems of low im- munogenicity that occur in young children, cer- tain elderly and other high risk persons following immunization with pneumococcal PS vaccine.
C. Vaccine Control - Assurance of Safety and Efficacy
Pneumococcal vaccine is prepared from pur- ified PS that are chemically defined, so that they can be analyzed by physicochemical character- istics and their potency and safety estimated by various tests on their composition and molecular size. Control tests are carried out at two steps.
1. Polysaccharide Bulk Powders
- Each lot of PS shall be examined for (1) the chemical content, such as 0-acetyl group, uronic acid, methyl pentose, hexosamine and phospho-
rus etc; (2) contamination of cell components, such as protein and nucleic acids; (3) molecular size determination by gel filtration using Se- pharose CL4B or CL-2B; (4) serological spec- ificity and identity by countercurrent immunoe- lectrophoresis or comparable technique. The U.S. Food and Drug Administration specifications are shown in Table 6.
2. Final Container
The final vaccine contains 25 pg of each type of PS per 0.5-ml dose. The presence (identity test) and amount of each PS are determined ser- ologically by either rocket immunoelectro- phoresis1I6 or by rate nephel~metry.~~’ The vac- cine should be tested for pyrogenicity and pass the specification at a test dose of 2.5 p,g for each PS included in the vaccine per milliliter per kil- ogram body weight of rabbit. The vaccine must also pass the general safety and sterility test re- quirements. It should not contain blood group A- like antigen.
Manufacturers of pneumococcal vaccine were required to perform a clinical potency test on at least the first two consecutive lots of each vaccine formulation for consistency. The clinical test was done in groups of at least 25 adults using a dose of 25 pg per each type of PSs. At least 80% of the vaccine recipients should demonstrate at least a twofold increase in specific PS antibodies.
In 1986, during review of the license appli- cation of pneumococcal vaccine in Japan, several questions were posed by the Biological Products Review Committee of Japanese Ministry of Health regarding the safety and quality of the vaccine. These questions were sent to the U.S. FDA for response and reply:
1.
2.
3.
How were the specifications of pneumo- coccal PS bulk powders determined? What is the cause of pyrogenicity in this vaccine? Why was a “40-fold” dilution of vaccine employed in performing the pyro- gen test. Why was the animal potency test not used? What is the basis and justification for using molecular size determination to provide as- surance of potency?
106
Cri
tical
Rev
iew
s in
Mic
robi
olog
y D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y C
DL
-UC
San
Die
go o
n 03
/15/
13Fo
r pe
rson
al u
se o
nly.
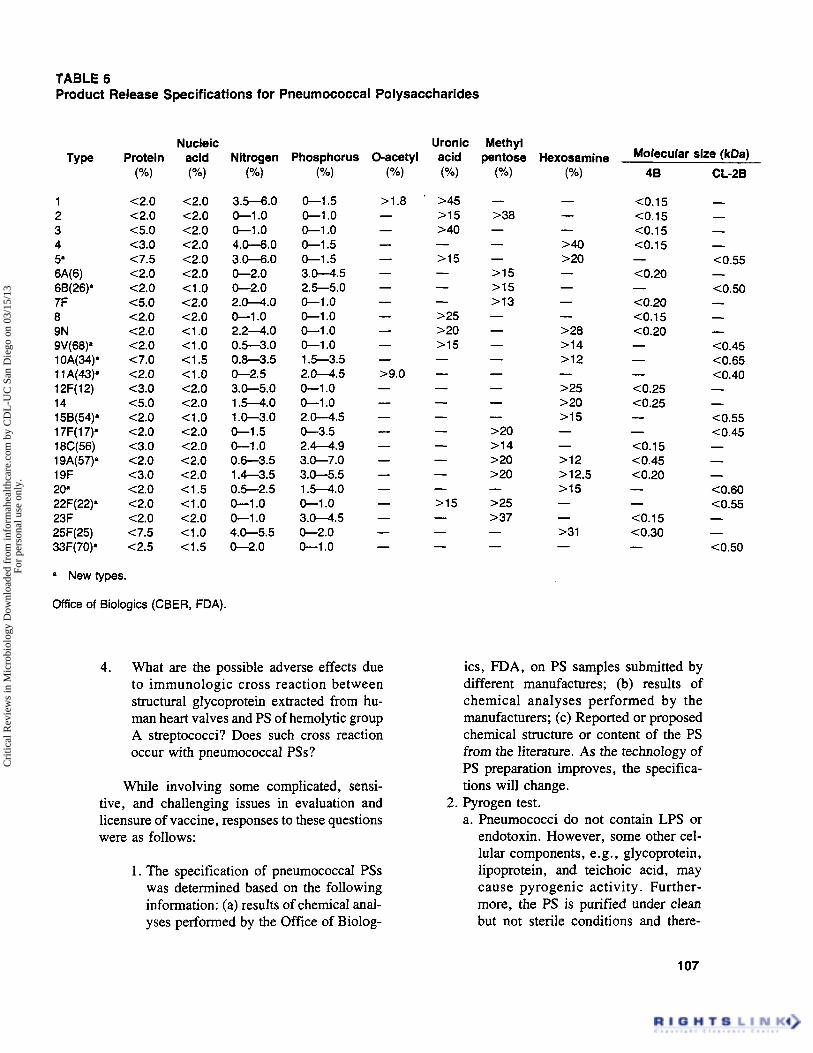
TABLE 6 Product Release Specifications for Pneumococcal Polysaccharides
Type
1 2 3 4 5’ 6A(6) 68(26)” 7F 8 9N 9V(68)‘ 1 OA(34)’ 1 1 A(43)’ 12F( 12) 14 15B(54)’ 1 7F( 1 7)’ 18C(56) 19A(57)* 19F 20’ 22F(22)’ 23F 25F(25) 33F(70)’
Protein ( O h )
<2.0 <2.0 <5.0 <3.0 <7.5 <2.0 c2.0 C5.0 <2.0 <2.0 <2.0 <7.0 c2.0 c3.0 <5.0 <2.0 <2.0 <3.0 <2.0 <3.0 <2.0 C2.0 <2.0 <7.5 <2.5
Nucleic acid (“A)
<2.0 c2.0 <2.0 <2.0 <2.0 <2.0 <1 .o <2.0 <2.0 <1 .o <1 .o <1.5 <1.0 <2.0 <2.0 <1 .o <2.0 <2.0 <2.0 <2.0 <1.5 <1 .o <2.0 <1 .o <1.5
a New types.
Office of Biologics (CBER, FDA).
Nitrogen (“w
3.5-6.0 0 - 1 .o 0 -1 .o 4.0-6.0 3.0-6.0 0-2.0 0-2.0 2.0-4.0 0 - 1 .o 2 .24 .0 0.5-3.0 0.8-3.5 0-2.5 3.0-5.0 1.5-4.0 1 .0-3.0 0-1.5 0-1 .o 0.6-3.5 1.4-3.5 0.5-2.5 0 -1 .o 0-1 .o 4.0-5.5 0-2.0
Phosphorus (”/.I
0-1.5 0-1 .o 0 -1 .o 0-1.5 0-1.5 3.0-4.5 2.5-5.0 0 -1 .o 0 -1 .o 0 -1 .o 0 -1 .o 1.5-3.5 2.0-4.5 0-1 .o 0 -1 .o 2.0-4.5 0-3.5 2.4-4.9 3.0-7.0 3.0-5.5 1.5-4.0 0 -1 .o 3.0-4.5 0-2.0 0 -1 .o
4. What are the possible adverse effects due to immunologic cross reaction between structural glycoprotein extracted from hu- man heart valves and PS of hemolytic group A streptococci? Does such cross reaction occur with pneumococcal PSs?
While involving some complicated, sensi- tive, and challenging issues in evaluation and licensure of vaccine, responses to these questions were as follows:
1. The specification of pneumococcal PSs was determined based on the following information: (a) results of chemical anal- yses performed by the Office of Biolog-
Molecular size (kDal
46
<0.15 C0.15 C0.15 <0.15
<0.20
(0.20 C0.15 C0.20
- -
- - - C0.25 c0.25 - - <0.15 <0.45 C0.20 - - C0.15 C0.30 -
its, FDA, on PS samples submitted by different manufactures; (b) results of chemical analyses performed by the manufacturers; (c) Reported or proposed chemical structure or content of the PS from the literature. As the technology of PS preparation improves, the specifica- tions will change.
a. Pneumococci do not contain LPS or endotoxin. However, some other cel- lular components, e.g., glycoprotein, lipoprotein, and teichoic acid, may cause pyrogenic activity. Further- more, the PS is purified under clean but not sterile conditions and there-
2. Pyrogen test.
107
Cri
tical
Rev
iew
s in
Mic
robi
olog
y D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y C
DL
-UC
San
Die
go o
n 03
/15/
13Fo
r pe
rson
al u
se o
nly.

fore, some Gram-negative endotoxin may be present. The required pyrogen test has limited the amount of possible contamination.
b. The pneumococcal 14-valent vaccine contains 100 kg PS/mVtype (50 kgl type/0.5 dose). Thus, 100 x 1/40 = 2.5 p,g/mVkg body weight dose of vaccine is injected into rabbit for the test. For the 23-valent vaccine, the di- lution should be 1/20 (50 kg/ml) rather than 1/40.
3. Potency test. a. Mice or rats have shown differences
in susceptibility to different types of pneumococci. For example, pneu- mococcal type 19F does not kill a mouse even at a very high concentra- tion, e.g., 1cT to lo6 organisms. In contrast, type 3 will kill a mouse with less than 10 organisms. Using the mouse, or another animal model, for potency test of pneumococcal PS in- troduced great experimental variance. Testing results are inconsistent and in- accurate. Furthermore, the antibody response of some pneumococcal PSs are different in rabbits and humans. The results on antibody response of pneumococcal PSs in animals can not directly be applied to humans.
b. The correlation between molecular size of bacterial capsular PS and the effi- cacy of vaccine or the human potency test is based on these studies in which the antigenicity and immunogenicity of PSs have been shown to be directly related to their molecular weights: b 1. The dextrans, having molecular
weight of 90,OOO and above, re- mained as excellent immunogens, whereas, at the molecular weight of 50,000 and below, they exhib- ited poor immunogenicity. ' I 8
b2.Fractions of different molecular weight of type 3 PS were injected into mice and the immunogenicity of each fraction was determined by plaque-forming cell (PFC) re- sponse. It was found that the num-
bers of PFC were directly related to the molecular weight of the frac- tion. The native PS was the most effective immunogen. 119
b3.The high molecular weight PSs of meningococcal groups A and C PSs showed high antibody response in humans. However, the group A PS isolated from cultures concentrated by rotary evaporator had lower molecular weights (C50,OOO) and proved to be nonimmunogenic in humans.Iz0 In clinical studies, the meningococcal groups A and C PSs degraded by a prolonged exposure to ambient temperatures were sig- nificantly less immunogenic. 1z1.1z2
4. Pneumococcal type 14 PS has a structure similar to blood group A hemagglutinin. Thus, the possibility of hemolytic reac- tion induced by such cross-immunologic reactions has been considered. The Of- fice for Biologics has investigated this problem through consulting with experts in National Institutes of Health and other investigators. It has been considered that the possibility of hemolytic activity due to such cross reactions between pneu- mococcal PSs and blood substances is more or less a theoretical concern. In- creases in blood group A antibodies in B + individuals have not been observed.
Since the pneumococcal vaccine was li- censed in 1977, more than 10 million doses have been distributed to the market. Reports of adverse reactions related to hemolytic reactions have not been received by FDA. The Office for Biologics is continuing to investigate whether the structural similarity between capsular PSs and cell-surface substances will cause any adverse reaction.
The above answers helped to resolve con- cerns of the reviewers regarding licensure of pneumococcal vaccine, and it was licensed in Japan shortly afterward.
V. CONCLUSIONS
S. pneumoniae is still one of the most prev-
108
Cri
tical
Rev
iew
s in
Mic
robi
olog
y D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y C
DL
-UC
San
Die
go o
n 03
/15/
13Fo
r pe
rson
al u
se o
nly.

alent and serious infections in humans. Molecular studies have been carried out to identify and char- acterize pneumococcal virulence factors, includ- ing capsular polysaccharide, pneumolysin, and several other cell-surface antigens. The infor- mation obtained from such studies can be used to investigate the immune responses of protective antigens for prevention of pneumococcal infec- tion. In the future, these studies will lead to the development of a more effective and safe pneu- mococcal vaccine.
ACKNOWLEDGMENT
We thank Dr. G. J. Boulnois for providing pJW208 for our studies.
REFERENCES
1. Immunization Practices Advisory Committee, Pneu- mococcal polysaccharide vaccine, MMWR. 38, 64, 1989.
2. Williams, W. W., Hickson, M. A., Kane, M. A., Kendal, A. P., Spika, J. S., and Hinman, A. R., Immunization policies and vaccine coverage among adults: the risk for missed opportunities, Ann. Intern. Med., 108, 616, 1988.
3. Austrian, R., Random gleanings from a life with the pneumococcus, J . Infect. Dis., 131, 474, 1975.
4. Austrian, R., Vaccines of pneumococcal capsular polysaccharides and the prevention of pneumococcal pneumonia, in The Role of Immunological Factors in Infectious. Allergic, and Autoimmune Processes, Beers, R. F.. Jr. and Bassett, E. G., Eds., Raven Press, New York, 1976, 79.
5. Lee, C. J., Bacterial capsular polysaccharide - bio- chemistry, immunity and vaccine, Mol. Immunol., 24, 1005, 1987. Fukumi, H., et al., Studies on clinical application of pneumococcal vaccine - distribution of Strep- tococcus pneumoniae in Japan, Jpn. Infect. Dis., 58, 39, 1984. Shen, S. and the Cooperation Group on Pneumococcal Serotyping, Serotypes and epide- miological study of pneumococcal infection in 18 provinces (cities) in China, Jiangxi Inst. Med. Sci., 10, 133, 1989. Robbins, J. B., Austrian, R., Lee, C. J., Rastogi, S. C., Schiffman, G., Henricksen, J., Makela, P. H., Broome, C. V., Facklam, R. R., Tiesjema, R. H., and Parke, J. C., Jr., Consideration for formulating the second-generation pneumococcd capsular polysaccharide vaccine with emphasis on the cross-reactive types, J . Infect. Dis., 148, 1136, 1983.
9. Briles, D. E. and Carroll, J., Natural selection and the response to polysaccharides, in Evolution and Vertebrate Immunity, Kelsoe, G . and Schulze, D. H., Eds., University ofTexas Press, Austin, 1987, 117.
10. Dochez, A. R. and Avery, 0. T., The elaboration of specific soluble substance by pneumococci during growth, J . Exp. Med., 26, 477, 1917.
* 11. Roitt, I. M., Brostoff, J., and Male, D., Immu- nology, 2nd ed., Gower Medical Publishing, London, 1989, 5.7.
12. Wood, W. B. and Smith, M. R., The inhibition of surface phagocytosis by the capsular slime layer of pneumococcus type III, J. Exp. Med., 90, 85, 1949.
13. Johnston, R. B., Klemperer, M. R., Alper, C. A., and Rosen, F. S., The enhancement of bacterial phagocytosis by serum, J . Exp. Med., 129, 1275, 1969.
14. Dhingra, R. K., Williams, J. C., andReed, W. P., Effect of pneumococcal mucopeptide and capsular polysaccharide on phagocytosis, Infect. Immun., 15, 169, 1977.
15. Schweinle, J. E., Pneumococcal intracellular killing is abolished by polysaccharides despite serum com- plement activity, Infect. Immun., 54, 876, 1986.
16. Lee, C. J., Biochemical aspects of the immunoge- nicity of pneumococcal polysaccharide vaccine, Memorial Symp. of Medical Sciences, College of Medicine, National Taiwan University, Taipei, Tai- wan, 1983, 4.
17. Szu, S., Lee, C. J., Parke, J. C., Jr., Schiffman, G., Henrichsen, J., Austrian, R., Rhtogi, S., and Robbins, J. B., Cross immunogenicity of pneumo- coccal group 9 capsular polysaccharides in adult vol- unteers, Infect. Immun., 35, 777, 1982.
18. Lee, C. J., Fraser, B. A., Szu, S., and Lin, K. T., Chemical structure of and the immune response to polysaccharide of Streptococcus pneumoniae, Rev. Infect. Dis., 3 , 322, 1981.
19. Penn, R. L., Lewin, E. B., Douglas, R. G., Jr., Schiffman, G., Lee, C. J., and Robbins, J. B., Antibody responses in adult volunteers to pneumo- coccal types 19F and 19A administered singly and in combination, Infect. Immun., 36, 1261, 1982.
20. Lee, C. J. and Koizumi, K., Immunochemical re- lations between pneumococcal group 19 and Kleb- siefla capsular polysaccharides, J . Immunol., 127, 1619, 1981.
21. Lee, C. J. and Wang, Z., Induction of increased antibody response to pneumococcal type 19F poly- saccharide by Klebsiella polysaccharide, J. Infect. Dis., 151, 658, 1985.
22. Lee, C. J., Koizumi, K., Henrichsen, J., Perch, B., Lin, C. S., and Egan, W., Capsular polysac- charides of non-groupable streptococci that cross-react with pneumococcal group 19, J. Immunol., 133,2706, 1984.
23. Fischer, H. and Tomasz, A., Peptidoglycan cross- linking and teichoic acid attachment in Streptococcus pneumoniae, J . Bacteriol., 163, 46, 1985.
1 09
Cri
tical
Rev
iew
s in
Mic
robi
olog
y D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y C
DL
-UC
San
Die
go o
n 03
/15/
13Fo
r pe
rson
al u
se o
nly.

24. Tomasz, A., Surface components of Streptococcus pneumoniae, Rev. Infect. Dis., 3, 190, 1981.
25. Sorensen, U. B., Blom, J., Birch-Andersen, A., and Henrichsen, J., Ultrastructural localization of capsules, cell wall polysaccharide, cell wall proteins, and F antigen in pneumococci, Infect. Immun., 56, 1890, 1988.
26. Jennings, H. J., Lugowski, C., and Young, N. M., Structure of the complex polysaccharide C-substance from Streptococcus pneumoniae type I, Biochemis- ny, 19, 4712, 1980.
27. Jennings, H. J., Capsular polysaccharides as vaccine candidates, Curr. Top. Microbiol. Immunol., 150, 98, 1990.
28. Briles, E. B. and Tomasz, A., Pneumococcal Forss- man antigen. A choline-containing lipoteichoic acid, J. Biol. Chem., 248, 6394, 1973.
29. Holtje, J.-V. and Tomasz, A., Specific recognition of choline residues in the cell wall teichoic acid by the N-acetylmuramyl-L-alanine amidase of pneumo- coccus, J. Biol. Chem., 250, 6072, 1975.
30. Sorensen, U. B. and Henrichsen, J., Cross-reac- tions between pneumococci and other streptococci due to C-polysaccharide and F-antigen, J. Clin. Mi - crobiol., 25, 1854, 1987.
31. Gray, B. M., Dillon, H. C., and Briles, D. E., Epidemiological studies of Streptococcus pneumon- iae in infants: development of antibody to phospho- choline, J. Clin. Microbiol., 18, 1102, 1983.
32. Szu, S. C., Schneerson, R., and Robbins, J. B., Rabbit antibodies to the cell wall polysaccharide of Streptococcus pneumoniae fail to protect mice from lethal challenge with encapsulated pneumococci, In- fect. Imrnun., 54, 448, 1986.
33. Briles, D. E., Forman, C., Hudak, S., and Claflin, J. L., Anti-phosphorylcholine antibodies of the T15 idiotype are optimally protective against Streptococ- cus pneumoniae, J . Exp. Med., 156, 1177, 1982.
34. Szu, S. C., Clark, S., and Robbins, J. B., Pro- tection against pneumococcal infection in mice con- ferred by phosphocholine binding antibodies. Spec- ificity of the phosphocholine binding and relation to several types, Infect. Immun., 39, 993, 1983.
35. Briles, D. E., Forman, D., Hudak, S., and Claflin, J. L., The effects of idiotypes on the ability of IgGl anti-phosphorylchole antibodies to protect mice from fatal infection with Streptococcus pneumoniae, Eur. J . Immunol., 14. 1027, 1984.
36. McDaniel, L. S., Benjamin, W. H., Forman, C., and Briles, D. E., Blood clearance by anti-phos- phocholine antibodies as a mechanism of protection in experimental pneumococcal bacteremia, J. Im- munol., 133, 3308, 1984.
37. Briles, D. E., Scott, G., Gray, B., Crain, M. J., Blaese, M., Nahm, M., Scott, V., and Haber, P.,
. _
110
Naturally occumng antibodies to phosphocholine as a potential index of antibody responsiveness to poly- saccharides, J. Infect. Dis., 155, 1307, 1987.
38.
39.
40.
41.
42.
43.
44.
45.
46.
47.
Holmberg, H., Krook, A., and Sjogren, A. M., Determination of antibodies to pneumococcal C poly- saccharide in patients with community-acquired pneumonia, J. Clin. Microbiol., 22, 808, 1985. Musher, D. M., Watson, D. A., and Baughn, R. E., Does naturally acquired IgG antibody to cell wall polysaccharide protect human subjects against pneumococcal infection?, J. Infect. Dis., 161, 736, 1990. Musher, D. M., Chapman, A. J., Goree, A., Jonsson, S., and Baughn, R. E., Natural and vac- cine-related immunity to Streptococcus pneumoniae, J. Infect. Dis., 154, 245, 1986. Sorensen, U. B. S. and J. Henrichsen, J., Cross- reactions between pneumococci and other strepto- cocci due to C-polysaccharide and F-antigen, J. Clin. Microbiol., 25, 1854, 1987. &ox, K. W. and Wicken, A. J., Immunological properties of teichoic acids, Bacteriol. Rev., 37,215, 1973. Horne, D. and Tomasz, A., Pneumococcal Forss- man antigen: enrichment in mesosomal membranes and specific binding to the autolytic enzyme of Strep- tococcus pneumoniae, J . Bacteriol., 161, 18, 1985. Bhakdi, S., Tranum-Jensen, J., and Szeigeleit, A., Mechanism of membrane damage by streptolysin 0, Znfect. Immun., 47, 52, 1985. Saunders, F. K., Michell, T. J., Walker, J. A., Andrew, P. W., and Boulnois, G. J., Pneumolysin, the thiol-activated toxin of Streptococcus pneumon- iae, does not require a thiol group for in vitro activity, Infect. Immun., 57, 2547, 1989. Nandoskar, M., Ferrante, A., Bates, E. J., Hurst, N., and Paton, J. C., Inhibition of human monocyte respiratory burst, degradation, phospholipid methy- lation and bactericidal activity by pneumolysin, Im- munology, 59, 515, 1986. Paton, J. C. and Ferrante, A., Inhibition of human polymorphonuclear leukocyte respiratory burst, bac- tericidal activity and migration by pneumolysin, In- fect. Immun., 41, 1212. 1983.
48. Ferrante, A., Rowan-Kelly, B., and Paton, J. C., Inhibition of in vitro human lymphocyte response by the pneumococcal toxin, pneumolysin, Infect. Im- mun., 46, 585, 1984.
49. Paton, J. C., Rowan-Kelly, B., and Ferrante, A., Activation of human complement by the pneumo- coccal toxin pneumolysin, Infect. Immun., 43, 1085, 1984.
50. Paton, J. C., Lock, R. A., and Hansman, D., Effect of immunization with pneumolysin on survival time of mice challenged with Streptococcus pneu- moniae, Infect. Immun., 40, 548, 1983.
51. Smyth, C. J. and Duncan, J. L., Thiol-activated (oxygen-labile) cytolysins, in Bacterial Toxin and CelI Membranes, Jeljaszewicz, J. and Wadstrom, T., Eds., Academic Press, New York, 1978, 129.
52. Johnson, M. K., Propexties of purified pneumococ- cal hemolysin, Infect. Immun., 6, 755, 1972.
Cri
tical
Rev
iew
s in
Mic
robi
olog
y D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y C
DL
-UC
San
Die
go o
n 03
/15/
13Fo
r pe
rson
al u
se o
nly.

53. Johnson, M. K., Geoffrey, C., And Alouf, J. E., Binding of cholesterol by sulfhydryl-activated cyto- lysins, Infect. Immun., 27, 97, 1980.
54. Walker, J. A., Allen, R. L., Falmagne, P., Johnson, M. K., and Boulnois, G. J., Molecular cloning, characterization, and complete nucleotide sequence of the gene for pneumolysin, the sulfhydryl- activated toxin of Streptococcus pneumoniae, Infect. Irnrnun., 55, 1184, 1987.
55. Oliver, D., Protein secretion in Escherichia coli, Annu. Rev. Microbiol., 39, 615, 1985.
56. Paton,J.C.,Beny,A.M.,Lock,R.A.,Hansman, D., and Manning, P. A., Cloning and expression in Escherichia coli of the Streptococcus pneumoniae gene encoding pneumolysin, Infect. Immun., 54, 50, 1986.
57. Li, J. P. and Lee, C. J., Characterization of group 19 pneumolysin and genetic analysis of pneumolysin gene, 3rd Int. Am. SOC. Microbiol. Conf. on Strep- tococcal Genetics, N43, Minneapolis, MN, 1990, 23.
58. Mitchell, T. J., Mendez, F., Paton, J. C., Andrew, P. W., and Boulnois, G. J., Comparison of pneu- molysin genes and proteins from Streptococcus pneu- moniae types 1 and 2, Nucleic Acids Res., 18, 4010, 1990.
59. McDaniel, L. S., Yother, J., Waltman, W. D., 11, and Briles, D. E., Insertionally inactivated mutants that lack pneumococcal surface protein A, in Strep- tococcal Genetics. Ferretti, J . J . and Curtiss, R., 111, Eds., American Society for Microbiology, Washing- ton, D.C., 1987, 172.
60. McDaniel, L. S., Yother, J., Viayakumar, M. N., McGarry, L., Guild, W. R., and Briles, D. E., Use of insertional inactivation to facilitate studies of biological properties of pneumococcal surface protein A (PspA), J . Exp. Med.. 165, 381, 1987.
61. Lock, R. A., Paton, J. C., and Hansman, D., Purification and immunological characterization of neuraminidase produced by Streptococcus pneumon- iae, Microb. Pathol., 4, 33, 1988.
62. Lock, R. A., Paton, J. C., and Hansman, D., Comparative efficacy of pneumococcal neuramini- dase and pneumolysin as immunogens protective against Streptococcus pneumoniae, Microb. Pathol., 5, 461, 1988.
63. Mosser, J. L. and Tomasz, A., Choline-containing teichoic acid as a structural component of pneumo- coccal cell wall and its role in sensitivity to lysis by an autolytic enzyme, J . Biol. Chem., 245,287, 1970.
64. Coyette, J. and Ghuysen, J. M., Wall autolysin of Lactobacillus acidophilus strain 63 AM Gasser, Bio- chemistry, 12, 2578, 1970.
65. Tomasz, A., The role of autolysins in cell death, Ann. N.Y. Acad. Sci., 235, 439, 1974.
66. Lee, C. J. and Liu, T. Y., The autolytic enzyme activity upon pneumococcal cell wall, Int. J . Biochem., 8, 573, 1977.
67. Shockman, G. D., Thompson, J. S., and Conover,
M. J., The autolytic enzyme system of Streptococcus faecalis II. Partial characterization of the autolysis and its substrate, Biochemistry, 6, 1054, 1967.
68. Ghuysen, J. M., Use of bacteriolytic enzymes in the determination of wall structure and their role in cell metabolism, Bacteriol. Rev., 32, 424, 1968.
69. Tomasz, A., Building and breaking of bonds in the cell wall of bacteria - the role of autolysins, in Microbial Cell Wall Synthesis and Autolysis, Nom- bela, C., ed., Elsevier, Amsterdam, 1984, 3.
70. Lopez, R., Sanchez-Puelles, J. M., Ronda, C., Garcia, P., Garcia, J. L., and Garcia, E., Bio- logical role(s) of the pneumococcal N-acetylmuramic acid-L-alanine amidase, in Streptococcal Genetics, Ferretti, J . J . and Curtiss, R., JJI, eds., American Society of Microbiology, Washington, D.C., 1987, 185.
71. Vered, M., Schutzbank, T., and Janoff, A., In- hibitors of human neutrophil elastase in extracts of Streptococcus pneumoniae. Am. Rev. Resp. Dis., 130, 1118, 1984.
72. Vered, M., Dearing, R., and Janoff, A., A new elastase inhibitor from Streptococcus pneumoniae protects against acute lung injury induced by neutro- phi1 granule, Am. Rev. Resp. Dis.. 131, 131, 1985.
73. Chetty, C. and Kreger, A., Generation of purpura- producing principle from pneumococcal cell walls, J . Bacteriol., 163, 389, 1985.
74. Male, C. J., Immunoglobulin A1 protease produc- tion by Haemophilus injluenzae and Streptococcus pneumoniae, Infect. Immun., 26, 254, 1979.
75. Mulks, M. H., Kornfeld, S. J., and Plaut, A. G., Specific proteolysis of human IgA by Streptococcus pneumoniae and Haemophilus injluenzae, J. Infect. Dis., 141, 450, 1980.
76. M u k , M. H., Plaut, A. G., Feldman, H. A., and Frangione, B., IgA proteases of two distinct spec- ificities are released by Neisseria rneningitidis, J . Exp. Med., 152, 1442, 1980.
77. Frangione, B. and Wolfenstein-Todel, C., Partial duplication in the “hinge” region of IgAl myeloma proteins, Proc. Natl. Acad. Sci. U.S.A., 69, 3673, 1972.
78. Liu, Y. S. V., Low, T. L. K., Infante, A., and Putnam, F. W., Complete covalent structure of a human IgAl immunoglobulin, Science, 193, 1017, 1976.
79. Pratt, S. A. and Bodnois, G. J., Molecular cloning of the immunoglobulin A1 protease gene from Strep- tococcus pneumoniae. in Streptococcal Genetics, Ferretti, J. J . and Curtiss, R., 111, Eds., American Society of Microbiology, Washington, D.C., 1987, 177.
80. DiRita, V. J. and Mekalanos, J. J., Genetic reg- ulation of bacterial virulence, Ann. Rev. Genet., 23, 455, 1989.
81. Weiss, A. A. and Hewlett, E. L., Virulence factors of Bordetella perpussis, Annu. Rev. Microbiol., 40, 661, 1986.
111
Cri
tical
Rev
iew
s in
Mic
robi
olog
y D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y C
DL
-UC
San
Die
go o
n 03
/15/
13Fo
r pe
rson
al u
se o
nly.

82.
83.
84.
85.
86.
87.
88.
89.
90.
91 I
92.
93.
94.
95.
96.
112
Stibitz, S. and Yang, M. S., personal communication. Stibitz, S., Aaronson, W., Monack, D., and Falkow, S. , Phase-variation in Bordetella perfussis by frameshift mutation in a gene for a novel two- component system, Nature, 338, 266, 1989. Lacey, B. W., Antigenic modulation of Borderella pertussis, J. Hyg., 58, 57, 1960. Moxon, E. R. and Vaughn, K. A., The type b capsular polysaccharide of Haemophilus influenzae: studies using clinical isolates and laboratory trans- formants, J . Infect. Dis., 143, 517, 1981. Hoiseth, S. K., Moxon, E. R., and Silver, R. P., Genes involved in Haemophilus influenzae type b capsule expression are part of an 18-kilobase tandem duplication, Proc. Natl. Acad. Sci. U.S.A., 83,1106, 1986. Kroll, J. S. and Moxon, E. R., Capsulation and gene copy number at the cap locus of Haemophilus injlwnzae type b, J . Bacteriol., 170, 859, 1988. Ely, S., Tippett, J., Kroll, J. S., and Moxon, E. R., Mutations affecting expression and maintenance of genes encoding the serotype b capsule of Haemo- philus injluenzae, J . Bacteriol., 167, 44, 1986. Kroll, J. S., Hopkins, I., and Moxon, E. R., Cap- sule loss in H. influenzae type b occurs by recom- bination-mediated disruption of a gene essential for polysaccharide export, Cell, 53, 347, 1988. Kelly, T., Caimano, M., and Yother, J., Locali- zation of genes encoding the type 3 capsules of Strep- tococcus pneumoniae, 3rd Int. Am. Soc. Microbiol. Conf. Streptococcal Genetics, N38, Minneapolis. MN, 1990, 22. Watson, D. A. and Musher, D. M., Molecular clon- ing of a sequence putatively required for encapsu- lation of Streptococcus pneumoniae serotype 3 , 3rd Int. Am. Soc. Microbiol. Conf. Streptococcal Ge- netics, N39, Minneapolis, MN, 1990, 22. Camara, M. M., Shepherd, A. M., Mitchell, T., Andrew, P. W., and Boulnois, G. J., Cloning and analysis of Streptococcus pneumoniae neuramini- dase, hyaluronidase and IgAl protease genes, 3rd Int. Am. Soc. Microbiol. Conf. Streptococcal Genetics, M40, Minneapolis, MN, 1990, 22. Robbins, J. B., Schneerson, R., Glode, M. P., Vann, W., Scheffer, M. S., Liu, T. Y., Parke, J. C., and Huntley, C., Cross-reactive antigens and immunity of disease caused by encapsulated bacteria, J. Clin. Immunol., 56, 141, 1975. Sutliff, W. D., Finland, M., and Jackson, H., Jr., Natural immunity of man to the Pneumococcus, J. Clin. Invest., 10, 660, 1931. S U M , W. D. and Davies, J. A. V., A study of the origin of naturally occumng immunity for pneumo- cocci in infants, J. Immunol., 32, 43, 1937. Robbins, J. B., Vaccines for the prevention of en- capsulated bacterial diseases: current status, problems and prospects for the future, Imrnunochernistry, 15, 839, 1978.
97. Heidelberger, M., Cross-reactions of polysaccha- rides of staphylococci and streptococci in antipneu- mococcal and other antisera, Mol. Immunol., 21, 1011, 1984.
98. Natig, J. B. and Kunkel, H. G., Human immu- noglobulins: classes, subclasses, genetic variants and idiotypes, Adv. Immunol., 16, 1, 1973.
99. Chudwin, D. S., Artrip, S. G., and Schiffman, G., Immunoglobulin G class and subclass antibodies to pneumococcal capsular polysaccharides, Clin. Im- munol. Immunopathol., 44, 114, 1987.
100. Heidelberger, M., Dilapi, M. M., Siegel, M., and Walter, A. W., Persistance of antibodies in human subjects injected with pneumococcal polysaccha- rides, J. Immunol., 65, 533, 1950.
101. Heidelberger, M., Persistance of antibodies in man after immunization, in The Nature of Signijicance of the Antibody Response, Pappenheimer, A. M., Jr., Ed., Columbia University Press, New York, 1953, 90.
102. Landesman, S. H. and Schiffman, G., Assessment of the antibody response to pneumococcal vaccine in high-risk populations, Rev. Infect. Dis., 3(Suppl.), S184, 1981.
103. Baker, P. J., Amsbaugh, D.F., Stashak, P. W., Caldes, G., and Prescott, B., Regulation of the antibody response to the pneumococcal polysaccha- ride by thymus-derived cells, Rev. Infect. Dis., 3, 332, 1981.
104. Braley-Mullen, H., Requirements for activation of contra-suppressor T cells by type-111 pneumococcal polysaccharide, J. Immunol., 136, 396, 1986.
105. Braley-Mullen, H., Characterization and activity of contra-suppressor T cells induced by type-III pneu- mococcal polysaccharide, J . Immunol.. 137, 2761, 1986.
106. Douglas, R. M., Paton, J. C., Duncan, and Hansman, D. J., Antibody response to pneumococ- cal vaccination in children younger than five years of age, J. Infect. Dis.. 148, 131, 1983.
107. Leinonen, M., Sakkinen, A., Kalliokoski, R., Luotenen, J., Tmonen, M., and Makela, P. H., Antibody response to 14-valent pneumococcal cap- sular polysaccharide vaccine in pre-school age chil- dren, Pediatr. Infect. Dis. , 5, 39, 1986.
108. Mufson, M. A., Krause, H. E., Schiffman, G.,
109.
110.
111.
and Hughey, D. F., Pneumococcal antibody levels one decade after immunization of healthy adults, Am. J. Med. Sci., 293, 279, 1987. Sims, R. V., McConville, J. H., Zwick, W., King, L., Steinman, W. C., and Schwartz, J. S., The clinical effectiveness of pneumococcal vaccine in the elderly, Clin. Res., 35, 491A, 1987. Shapiro, E. D., Austrian, R., Adair, A. K., and Clemens, J. D., The protective efficacy of pneu- mococcal vaccine, Clin. Res., 36, 470A, 1988. Fedson, D., Henrichsen, J., Makela, P. H., and Austrian, R., Immunization of elderly people with
Cri
tical
Rev
iew
s in
Mic
robi
olog
y D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y C
DL
-UC
San
Die
go o
n 03
/15/
13Fo
r pe
rson
al u
se o
nly.

polyvalent pneumococcal vaccine, Infection, 17, 437, 1989.
112. Robbins, J. R. and Schneerson, R., Polysaccha- ride-protein conjugates: a new generation of vaccines, J. Infect. Dis., 161, 821, 1990.
113. Paton, J. C., Lock, R. A., Lee, C. J., Li, J. P., Verry, A. M., Michell, T. J., Andrew, P. W., Hansman, D., and Boulnois, G. J., Purification and immunogenicity of genetically toxoided derivatives of pneumolysin and their conjugation to Streptococ- cus pneumoniae type 19F polysaccharide, Infect. Im- mun., 59, 2297, 1991.
1 14. Sarnaik, S., Kaplan, J., SchWman, G., Bryla, D., Robbins, J. B., and Schneerson, R., Studies on pneumococcus vaccine alone or mixed with DTP and on pneumococcus type 6B and Haemophilus influ- emae type b capsular polysaccharide-tetanus toxoid conjugates in 2 to 5-year old children with sickle cell anemia, Pediatr. Infect. Dis., 9, 18 1, 1990.
115. Dick, W. E., Jr. and Beurret, M., Glycoconjugates of bacterial carbohydrate antigens, in Conjugate Vuc- cines, Cruse, J. M. and Lewis, R. E., Eds., S. Kar- ger, Switzerland, 1989, 48.
116. Lee, C. J., Pearson, S. J., and Robbins, J. B., Quantitative determination of pneumococcal poly- saccharides in the polyvalent vaccine. I. Standard- ization of an immunoelectrophoretic method, J. Biol. Stand., 8, 271, 1980.
117. Lee, C. J., The quantitative immunochemical de- termination of pneumococcal and meningococcal capsular polysaccharides by light scattering rate ne- phelometry, J. Biol. Stand., 11, 55, 1983.
118. Kabat, E. S. and Benzer, A. E., The effect of variation in molecular weight on the antigenicity of dextran in man, Arch. Biochem. Biophys., 78, 306, 1958.
119. Howard, J. G., Zola, H., Christie, G. H., and Coutenay, B. M., Studies on immunological para- lyses. V. The influence of molecular weight on the immunogenicity, tolerogenicity, and antibody-neu- tralizing activity of type IU pneumococcal polysac- charide, Immunology, 21, 535, 1971.
120. Gotschlich, E. C., Liu, T. Y., and Artenstein, M. S., Human immunity to the meningococcus. III. Preparation and immunochemical properties of the group A and group C meningococcal polysaccha- rides, J. Exp. Med., 129, 1349, 1969.
121. Gotschlich, E. C., Rey, M., Trian, R., and Sparks, K. J., Quantitative determination of the human re- sponse to immunization with meningococcal vac- cines, J. Clin. Invest., 51, 89, 1972.
122. Gold, R., Lepow, M. L., Goldschneider, I., Drapu, T. L., and Gotschlich, E. C., Clinical evaluation of group A and group C meningococcal polysaccha- ride vaccines in infants, J. Clin. Invest., 56, 1536,
123. Fedson, D. S., Pneumococcal vaccine, in Vaccines, Plotkin, S . A. and Mortimer, E. A., Eds., W. B. Saunders, Philadelphia, 1988, 271.
, 1975.
124. Lindberg, B., Lindquist, B., Longren, J. and Powell, D. A., Structural studies of the capsular poly- saccharide from Streptococcus pneumoniae type 1, Carbohydr. Res., 78, 111, 1980.
125. Jansson, P. E., Lindberg, B., Anderson, M., Lindquist, U., and Henrichsen, J., Structural stud- ies of the capsular polysaccharide of Streptococcus pnewnoniae type 2, a reinvestigation, Carbohydr. Res., 182, 111, 1988.
126. Reeves, R. E. and G~ebel, W. F., Chemoimmu- nological studies on the soluble specific substance of pneumococcus. V. The structure of the type JII poly- saccharide, J. Biol. Chem., 139, 11, 1941.
127. Jansson, P. E., Lindberg, B., and Lindquist, U., structural studies of the capsular polysaccharide from Streptococcus pnewnoniae type 4, Carbohydr. Res., 95, 73, 1981.
128. Jansson, P. E., Lindberg, B., and Lindquist, U., Structural studies of the capsular polysaccharide from Streptococcus pneumoniae type 5, Carbohydr. Res., 140, 101, 1985.
129. Rebers, P. A. and Heidelberger, M., The specific polysaccharide of type VI pneumococcus. II. The repeating unit, J. Am. Chem. SOC., 83, 3056, 1961.
130. Kenne, L., Lindberg, B., and Madden, J. K., Structural studies on the capsular antigen from Strep- tococcus pneumoniae type 26, Carbohydr. Res., 73, 175, 1979.
131. Moreau, M., Richards, J. C., Perry, M. B., and Kniskern, P. J., Application of high resolution nu- clear magnetic resonance spectroscopy to the struc- tural elucidation of the specific capsular polysaccha- ride of Streptococcus pneumoniae type 7F, Carbohydr. Res., 182, 79, 1988.
132. Jones, J. K. N. and Perry, M. B., The structure of the type WI pneumococcus specific polysaccha- ride, J. Am. Chem. SOC., 79, 2787, 1957.
133. Jones, C., Mulloy, B., Wilson, A., Dell, A., and Oates, J. E., Structure of the capsular polysaccharide from Streptococcus pnewnoniae type 9, J. Chem. SOC. Perkin Trans., 1, 1665, 1985.
134. Perry, M. B., Daoust, V., and Carlo, D. J., The specific capsular polysaccharide of Streptococcus pneumoniae type 9V, Can. J. Biochem., 59, 524, 1981.
135. Richards, J. C. and Perry, M. B., Structure of the specific capsular polysaccharide of Streptococcus pnewnoniae type lOA, Carbohydr. Res.. in press.
136. Richards, J. C., Perry, M. B., and Moreau, M., Elucidation and comparison of the specific capsular polysaccharide of Streptococcus pneumoniae group 11 (11F, 11B, 11Cand 11A),Adv. Exp. Med. Biol., 228, 595, 1988.
137. Leontein, K., Lindberg, B., and Lonnpen, J., Structural studies of the capsular polysaccharide from Streptococcwpnewnoniae type 12F, Can. J . Chem., 59, 2081, 1981.
138. Lindberg, B., Lonngren, J., and Powell, D. A., Structural studies on the specific type 14 pneumo-
113
Cri
tical
Rev
iew
s in
Mic
robi
olog
y D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y C
DL
-UC
San
Die
go o
n 03
/15/
13Fo
r pe
rson
al u
se o
nly.

coccal polysaccharide, Carbohydr. Res., 58, 177, 1977.
139. Jansson, P. E., Lmdberg, B., Lindquist, U., and Lindberg, J., Structural studies of the capsular p l y - saccharide from Streptococcus pneumoniae types 15B and 15C, Carbohydr. Res. , 162, 11 1, 1987.
140. Perry, M. B., Bundle, D. R., Daoust, V., and Carlo, D. J., The structure of the specific polysac- charide of Streptococcuspneumoniae type 17F, Car- bohydr. Res., in press.
141. Phillips, L. R., Nishimura, O., and Fraser, B. A., The structure of the repeating oligosaccharide unit of the pneumococcal capsular polysaccharide 18C, Car- bohydr. Res., 121, 243, 1983.
142. Lugowski, C. and Jennings, H. J., Structural de- termination of the capsular polysaccharide of Strep- tococcus pnewnoniue type 18C (56), Carbohydr. Res., 131, 119, 1984.
143. Lee, C. J. and Fraser, B. A., The structure of the cross-reactive types 19(19F) and 57 (19A) pneumo- coccal capsularpolysaccharides, J. Bid . Chem., 255, 6847, 1980.
144. Richards, J. C., Perry, M. B., and Carlo, D. T., The specific capsular polysaccharide of Streptococ- cus pneumoniae type 20, Can. J. Biochem. Cell Biol., 61. 178, 1983.
145. Richards, J. C., Perry, M. B., and Kniskern, P. J., Structural analysis of the specific capsular polysac- charide of Streptococcus pneumoniae type 22F, Can. J. Chem., 67, 1038, 1989.
146. Richards, J. C. and Perry, M. B., Structure of the specific capsular polysaccharide of Streptococcus pneumoniae type 23F, Biochem. Cell Biol., 66,758, 1988.
147. Richards, J. G. and Perry, M. B., The specific capsular polysaccharide of Streptococcus pneumon- iae type 33F, Can. J. Biochem. Cell Biol.. 62, 666, 1984.
148. Laemmli, U. K., Cleavage of structural proteins dur- ing the assembly of the head of bacteriophage T4, Nature, 227, 680, 1970.
149. Saiki, R. K., Gelfand, D. H., Stoffel, S., Scharf, S. J., Higuchi, R., Horn, G. T., Mullis, K. B., and Erlich, H. A., Primer-directed enzymatic am- plification of DNA with a thermostable DNA poly- merase, Science, 239, 487, 1988.
150. Arico, B., Miller, J. F., Roy, C., Stibitz, S., Monadr, D., Falkow, S., Gross, R., and RappuoU, R., Sequences required for expression of Bordetella pertussis virulence factors share homology with pro- karyotic signal transduction proteins, Proc. Nutl. Acad. Sci. U.S.A., 86;6671, 1989.
151. Monack, D. M., Arico, B., Rappuoli, R., and Falkow, S., Phase variants of Bordetella bronchi- septica arise by spontaneous deletions in the vir locus, Mol. Microbiol., 3, 1719, 1989.
152. Stibitz, S., Weiss, A. A., and Falkow, S., Genetic analysis of a region of the Bordetella pertussis chro- mosome encoding filamentous hemagglutinin and the pleiotropic regulatory locus vir, J. Bacteriol., 170, 2004, 1988.
153. Roy, C. R., Miller, J. F., and Falkow, S. , The bvgA gene of Bordetella pertussis encodes a tran- scriptional activator required for coordinate regula- tion of several virulence genes, J. Bacteriol., 171, 6338, 1989.
154. Lee, D. H., Hoidal, J. R., Garkh, D. H., Clawson, C. C., Quie, P. G., and Peterson, P. K., Opsonin- independent phagocytosis of surface-adherent bac- teria by human alveolar macrophages, J. Leukocyte Biol.. 36, 689, 1984.
155. Fine, D. P., Kirk, J. L., schiffman, G., Schweinie, J. E., and Guckian, J. C., Analysis of humoral and phagocytic defense against Streptococcus pnewnon- iae serotypes 1 and 3, J. Lab. Clin. Med., 112,487, 1988.
156. Griffioen, A. W., Rijkers, G. T., Janssens- Korpela, P., and Zegers, B. J. M., Pneumococcal polysaccharides complexed with C3d bind to human B lymphocytes via complement receptor type 2, In- fect. Immun., 99, 1839, 1991.
157. Winkelstein, J. A. and Tomasz, A., Activation of the alternative complement pathway by pneumococ- cal cell wall teichoic acid, J. Immunol., 120, 174, 1978.
158. Baker, P. J., Regulation of magnitude of antibody response to bacterial polysaccharide antigens by thy- mus-derived lymphocytes, Infect. Immun., 58,3465, 1990.
159. Taylor, C. E., Stashak, P. W., Caldes, G., Prescott, B., Chused, T. E., Brooks, A., and Baker, P. J., Activation of antigen-specific sup- pressor T cells by B cells from mice immunized with type III pneumococcal polysaccharide, J. Exp. Med., 158, 703, 1983.
160. Taylor, C. E., Stashak, P. W., Chiang, J., Leiserson, W. M., Caldes, G., Prescott, B., and Baker, P. J., Characteristics of amplifier T cells involved in the antibody response to the capsular polysaccharide of type 111 Srreptococcuspneumoniae, J. Immunol., 132, 3103, 1984.
161. Taylor, C. E. and Baker, P. J., Production of sol- uble suppressor factor by spleen cells from mice im- munized with type 111 pneumococcal polysaccharide, J. Immunol., 135, 2551, 1985.
114
Cri
tical
Rev
iew
s in
Mic
robi
olog
y D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y C
DL
-UC
San
Die
go o
n 03
/15/
13Fo
r pe
rson
al u
se o
nly.