virtual free radical school iron chelation in...
TRANSCRIPT

Iron in Biology Society For Free Radical Biology and Medicine Crumbliss 1
Virtual Free Radical School
Iron Chelation in BiologyIron Chelation in Biology
Alvin L. CrumblissDepartment of Chemistry
Duke UniversityBox 90346
Durham, NC 27708-0346
Telephone: (919) 660-1540Fax: (919) 660-1605
E-mail: [email protected]: http://www.chem.duke.edu/%7Ealc/labgroup/

Iron in Biology Society For Free Radical Biology and Medicine Crumbliss 2
Iron Chelation in BiologyIron Chelation in BiologyTutorial Guide
Introduction: Biological Iron Coordination ChemistryPanels 3, 4 & 5
Chelation and SolubilityPanel 6
Chelation and Redox PotentialPanel 7
Common Iron Ligands in BiologyPanel 8
Chelate Stability DefinitionsPanel 9
Chelation and Redox ControlPanels 10, 11 & 12
Oxidation State Influence on Chelate StabilityPanel 13
Iron Chelation and TransportPanels 14, 15 &16
Influence of pH on Chelate StabilityPanel 17
Influence of Chelate Stability on E0
Panel 18 Influence of Chelation on Kinetics
Panel 19

Iron in Biology Society For Free Radical Biology and Medicine Crumbliss 3
Introduction: Introduction: Biological Iron Coordination ChemistryBiological Iron Coordination Chemistry Iron is the second most abundant metal on the earth’s surface, falling closelybehind aluminum and in near equivalent concentration to calcium andsodium. It is an essential element for virtually every living cell.
The biochemistry of iron is controlled to a large extent by its coordination chemistry; i.e. the immediate chemical environment in the first coordinationshell. This first coordination shell controls iron’s biological activity in smallmolecule storage (e.g.O2), electron transport, and catalysis.
Fe
1st coordination shell;immediate chemical environment
Common oxidation states: +2, +3
Common coordination numbers: 4, 5, 6
3 References [1,2]

Iron in Biology Society For Free Radical Biology and Medicine Crumbliss 4
Introduction: Introduction: Biological Iron Coordination ChemistryBiological Iron Coordination Chemistry
Examples of the extensive use of iron in biological systems, all of which are controlled or mediated by chelation, are as follows:
redox chemistry involved in simple electron-transfer reactions;
redox chemistry involved in reactions with O2, ranging from O2 transport and storage to O2 reduction by cytochrome oxidase, and O atom insertion catalyzed by cytochrome P450; and
substrate activation by the electrophilic behavior of iron; for example, hydrolase enzymes such as purple acid phosphatase.
References [1,2]4

Iron in Biology Society For Free Radical Biology and Medicine Crumbliss 5
Introduction: Introduction: Biological Iron Coordination ChemistryBiological Iron Coordination Chemistry
The first coordination shellPrevents hydrolysis/precipitationInfluences molecular recognitionControls redox potentialControls mobility
Fe
5

Iron in Biology Society For Free Radical Biology and Medicine Crumbliss 6
Iron Chelation and SolubilityIron Chelation and SolubilityFe insoluble due to hydrolysis
FeH2O OH2
OH2H2O
OH2
OH2
FeH2O OH
OH2H2O
OH2
OH2
3+ 2+
FeH2O O
OH2OOH2
OH2
FeOH2
OH2
OH2
OH2H
H 4+
H+
H+
Higher insoluble polymers
Fe
L:
[Feaq3+]tot = 10-10 M @ pH 7
Strong chelators prevent hydrolysis and precipitation
6
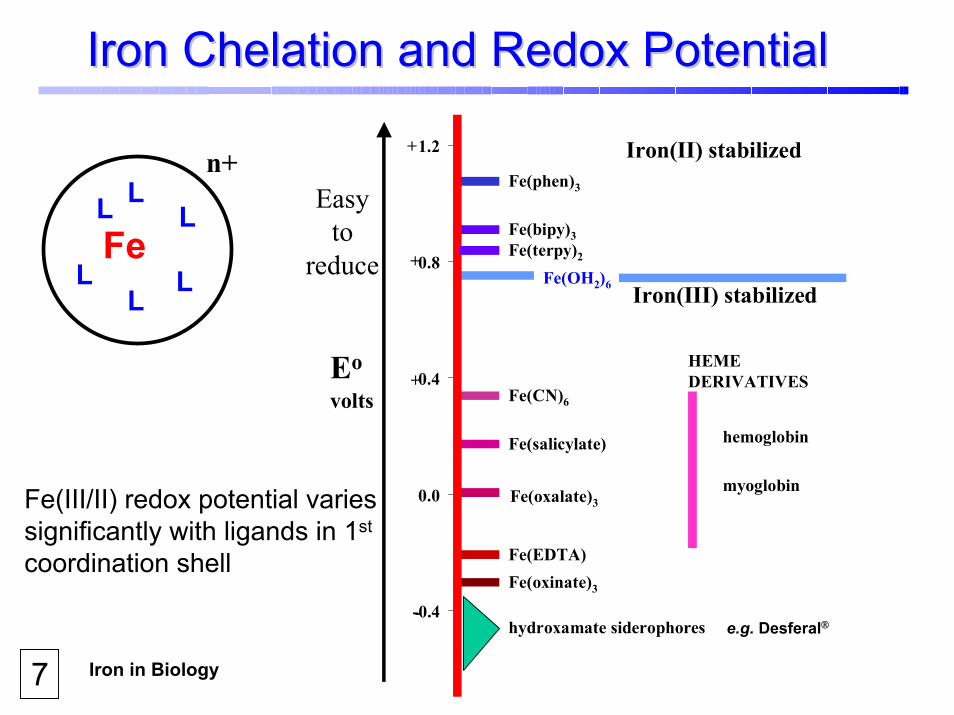
Iron in Biology Society For Free Radical Biology and Medicine Crumbliss 7
-0.4
0.0
0.4
0.8
1.2
Fe(OH2)6
Iron(II) stabilized
Iron(III) stabilized
Fe(terpy)2
Fe(phen)3
Fe(bipy)3
Fe(salicylate)
Fe(CN)6 -
Fe(EDTA)Fe(oxinate)3
HEMEDERIVATIVES
hemoglobin
myoglobin
+
+
+
-
Easy to
reduce
Eo
volts
Fe(oxalate)3
hydroxamate siderophores
FeL L
LL L
L
n+
Iron Chelation and Redox PotentialIron Chelation and Redox Potential
Fe(III/II) redox potential varies significantly with ligands in 1st
coordination shell
7e.g. Desferal

Iron in Biology Society For Free Radical Biology and Medicine Crumbliss 8
Common Iron Ligands in BiologyCommon Iron Ligands in Biology
Iron(III) is a hard Lewis acid and prefers ligation to hardLewis base donors (e.g. O, amine N) and iron(II) is aborderline soft Lewis acid and prefers ligation to soft Lewisbase donors (e.g. S, pyrrole N).
N
N
N
N
Fe
Fe
OO
Fe
OO
R1
NR2
Fe
OO
O
Fe
O
O
Fe
OFe
H2N
Fe
N
NH
Fe
S
Common iron ligand donor groups in biology include amino acid side chains, suchas amine (I), carboxylate (II), imidazole (III), phenol (IV), and thiol (V). Otherligating groups include α-hydroxy carboxylate (VI), catecholate (VII), hydroxamate(VIII) and porphyrin (IX).
(I)
(II)(III)
(IV) (V) (VI)
(VII)(VIII)
(IX)8

Iron in Biology Society For Free Radical Biology and Medicine Crumbliss 9
Iron Chelate Stability DefinitionsIron Chelate Stability DefinitionsCompilations of metal-ligand complex stabilities, such as that edited by Martell andSmith, use pH independent equilibrium constants, βFeLH, as defined below for the reaction between Fe(III) and a hexadentate triprotic ligand, LH3, in aqueous solution. Fe(OH2)6
3+ + L3- FeL β110 = However, in an in vivo or in vitro situation protons compete for the Fe(III) binding sitesand the degree of complexation of the metal will be influenced by the ligand pKa values and the pH of the medium. Fe(OH2)6
3+ + H3L FeL + 3 H+ K = Since stability constants β and K are determined as concentration quotients, their unitsdiffer on changing the denticity of the ligand. Consequently, β110 for a hexandentate ligand and β130 for a bidentate ligand cannot be directly compared. A pFe scalecircumvents this problem and the problem of H+ competition due to different ligand pKavalues. The pFe value for a particular ligand is the negative log of the free Fe(III)concentration at a fixed set of conditions: [total ligand] = 10 µM, [total Fe(III)] = 1 µM, and pH = 7.4. A high pFe value denotes a stable chelate complex. Panel 17 illustrates the influence of pH on Fe(III)-siderophore complex stability, usingpFe values to express the stability of the complex.
[FeL][Fe(OH2)6
3+][L3-]
[FeL] [H+]
[Fe(OH2)63+][H3L]
9 References [3,4,5,6]

Iron in Biology Society For Free Radical Biology and Medicine Crumbliss 10
Iron Chelation and RedoxIron Chelation and Redox Control Control Why is it important?Why is it important?A mechanism for preventing iron from participating in a catalytic cycle to producetoxic hydroxyl radicals and/or reactive oxygen species (ROS) (e.g. via the Fenton reaction or Haber Weiss cycle) is to control its redox potential by selectivechelation. Through chelation, the redox potential for iron may be removed fromthe region where it can undergo redox cycling and produce hydroxyl radicals andROS. This is illustrated in Panel 12. From the following thermochemical cycle, Equation (1) can be derived whichrelates the redox potential of an Fe complex to the chelator’s ability to discriminatebetween Fe(III) and Fe(II), as expressed by βIII and βII. This relationship illustratesthat the selectivity of a chelator for Fe(III) over Fe(II) increases with decreasingredox potential. Fe(H2O)6
3+ + L Fe3+L
Fe(H2O)62+ + L Fe2+L
βIII
βII
E0aq E0
complex E0aq – E0
complex = 59 log(βIII/βII) [1]
10 Reference [6]

Iron in Biology Society For Free Radical Biology and Medicine Crumbliss 11
Iron Chelation and RedoxIron Chelation and Redox ControlControlWhy is it important?
From Equation (1) it is evident that the redox potential and stability of an ironcomplex are inter-related. These inter-relationships are important in characterizing the biologicalchemistry of iron because controlling the oxidation state of iron is a method ofcontrolling both the thermodynamic and kinetic stability of a coordinationcompound. This is illustrated in Panel 13. As a result, the redox potential of acomplex may be viewed as a measure of the sensitivity of a molecular levelswitch for changing the chemical environment of the iron (1st coordination shell). Data in Panel 13 show that for high spin complexes, changing theoxidation state of iron from +2 to +3 changes both the kinetic lability andthermodynamic stability of an iron chelate complex.
Why is it important?
11 Reference [6]
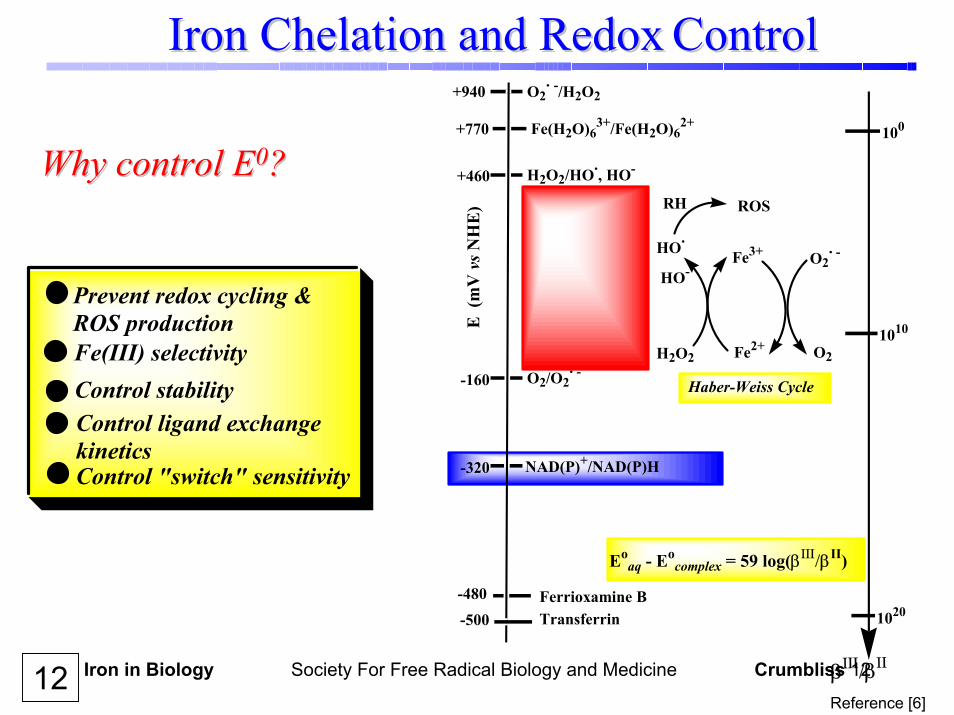
Iron in Biology Society For Free Radical Biology and Medicine Crumbliss 12
Iron Chelation and RedoxIron Chelation and Redox ControlControl
-480
-320E
(m
V v
s NH
E)
NAD(P)+/NAD(P)H
-160 O2/O2. -
+940 O2. -/H2O2
+460 H2O2/HO., HO-
Fe3+
Fe2+ O2
O2. -
H2O2
HO.
HO-
ROSRH
Haber-Weiss Cycle
Eoaq - Eo
complex = 59 log(βΙΙΙ/βII)
Fe(H2O)63+/Fe(H2O)6
2++770
1020
1010
100
βΙΙΙ/βΙΙ
-500Ferrioxamine BTransferrin
Why control EWhy control E00??
Prevent redox cycling & ROS productionFe(III) selectivityControl stabilityControl ligand exchange kineticsControl "switch" sensitivity
12Reference [6]
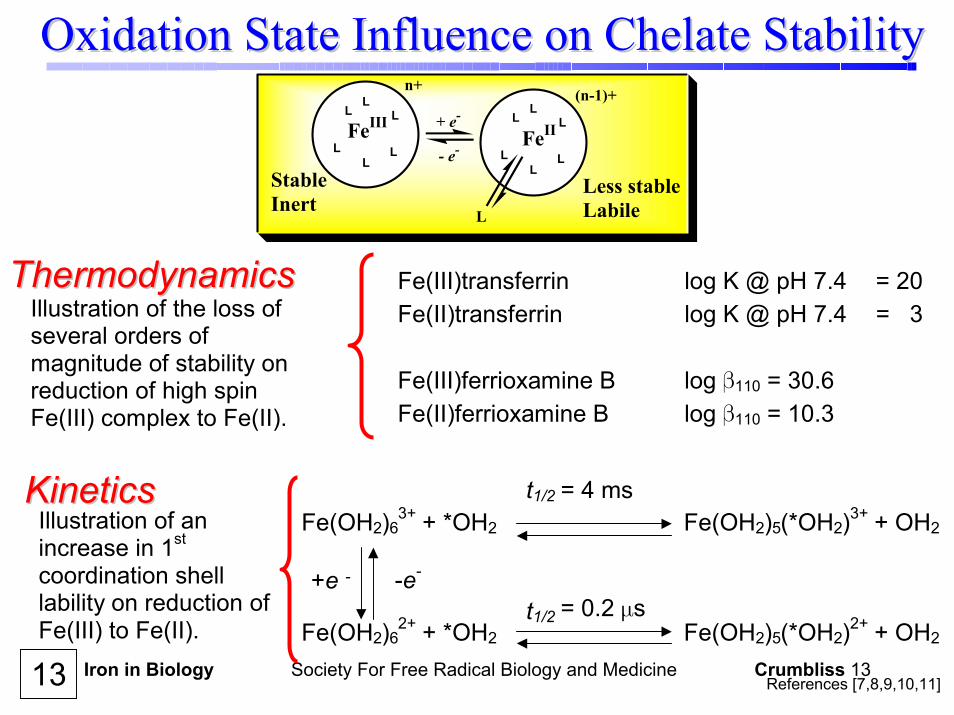
Iron in Biology Society For Free Radical Biology and Medicine Crumbliss 13
FeIIIL L
LLL
L L L
LLL
L
L
n+ (n-1)+
+ e-
- e- FeII
StableInert
Less stableLabile
Fe(III)transferrin log K @ pH 7.4 = 20 Fe(II)transferrin log K @ pH 7.4 = 3 Fe(III)ferrioxamine B log β110 = 30.6 Fe(II)ferrioxamine B log β110 = 10.3
ThermodynamicsThermodynamicsIllustration of the loss of several orders of magnitude of stability on reduction of high spin Fe(III) complex to Fe(II).
KineticsKineticsIllustration of an increase in 1st coordination shell lability on reduction of Fe(III) to Fe(II).
Fe(OH2)63+ + *OH2 Fe(OH2)5(*OH2)3+ + OH2
Fe(OH2)62+ + *OH2 Fe(OH2)5(*OH2)2+ + OH2
-e-+e -
t1/2 = 4 ms
t1/2 = 0.2 µs
13 References [7,8,9,10,11]
Oxidation State Influence on Chelate StabilityOxidation State Influence on Chelate Stability
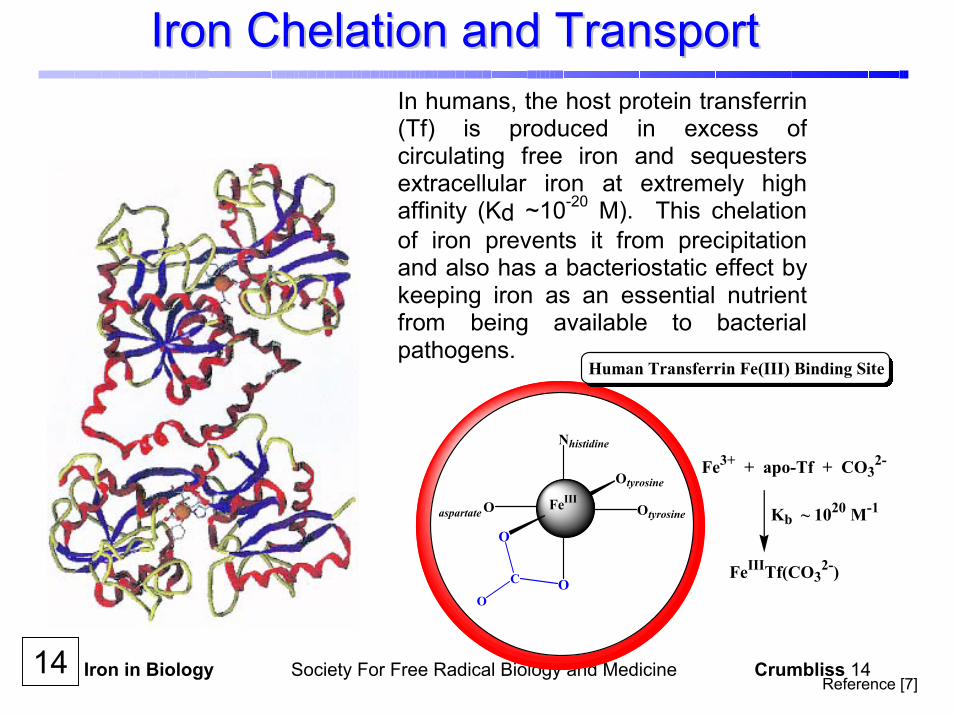
Iron in Biology Society For Free Radical Biology and Medicine Crumbliss 14
Iron Chelation and TransportIron Chelation and TransportIn humans, the host protein transferrin(Tf) is produced in excess ofcirculating free iron and sequestersextracellular iron at extremely highaffinity (Kd ~10-20 M). This chelationof iron prevents it from precipitationand also has a bacteriostatic effect by keeping iron as an essential nutrientfrom being available to bacterialpathogens.
FeIII
O
Otyrosine
OtyrosineO
Nhistidine
O
aspartate
C
O
Human Transferrin Fe(III) Binding Site
Fe3+ + apo-Tf + CO32-
Kb ~ 1020 M-1
FeIIITf(CO32-)
14Reference [7]
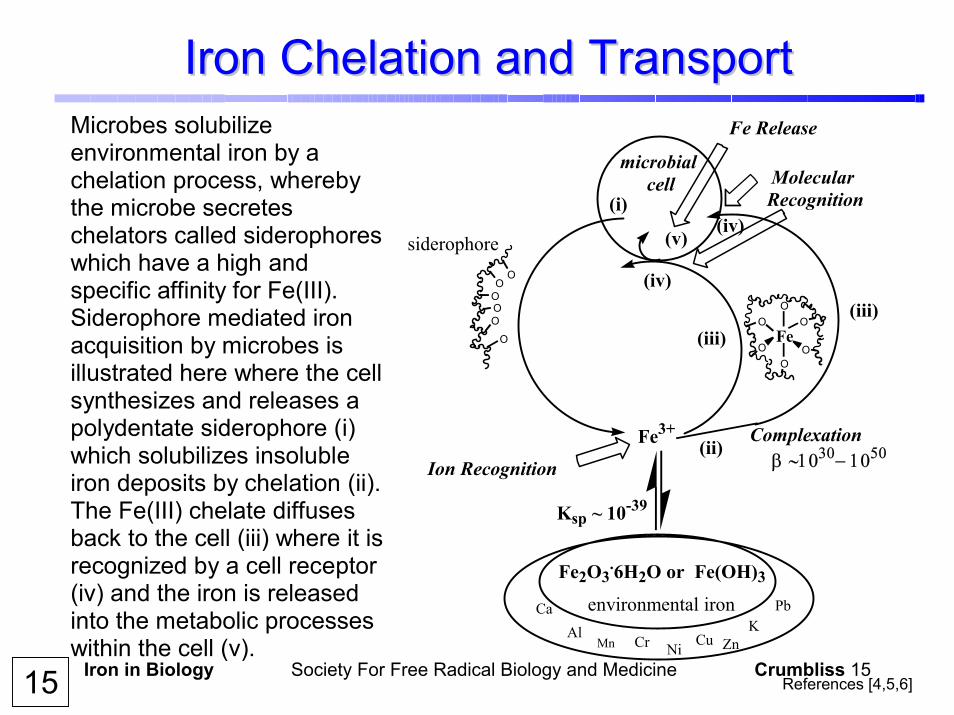
Iron in Biology Society For Free Radical Biology and Medicine Crumbliss 15
Iron Chelation and TransportIron Chelation and TransportMicrobes solubilize environmental iron by a chelation process, whereby the microbe secretes chelators called siderophores which have a high and specific affinity for Fe(III). Siderophore mediated iron acquisition by microbes is illustrated here where the cell synthesizes and releases a polydentate siderophore (i) which solubilizes insoluble iron deposits by chelation (ii). The Fe(III) chelate diffuses back to the cell (iii) where it is recognized by a cell receptor (iv) and the iron is released into the metabolic processes within the cell (v).
OOOO
O
O
Ion Recognition
microbial cell
O
O
OO
OO
Fe2O3.6H2O or Fe(OH)3
Fe3+
Molecular Recognition
Ksp ~ 10-39
environmental iron
siderophore
Fe Release
Complexationβ ∼1030− 1050
AlZnCr CuMn Ni
KCa Pb
Fe
(i)
(ii)
(iii)(iii)
(iv)
(iv)(v)
15 References [4,5,6]

Iron in Biology Society For Free Radical Biology and Medicine Crumbliss 16
Iron Chelation and TransportIron Chelation and TransportSiderophores, microbially synthesized Fe(III) specific chelators, are low molecularweight molecules that usually incorporate bidentate catechol, hydroxamic acid, and/orα-hydroxy carboxylic acid donor groups. These chelators exhibit high Fe(III) complexstabilities (high β and pFe) to enhance delivery of iron to the cell, and large negativeredox potentials (Panels 7, 12 and 18) for Fe(III) complexing specificity and toprevent redox cycling leading to the production of toxic hydroxyl radicals and ROS(Panels 10, 11 and 12). Shown below are the structures of two hexadentate siderophores; enterobactin, a tris catecholate (in red), and ferrioxamine B, a trishydroxamate (in blue).
enterobactin
Fe(III)-enterobactin complexβ110 = 1049 ; pFe = 35.5
HOO
OH HOO
NHN
O
N
HN
O
N
NH3+
O
NH
O
O
O
OHN
O O
NH
O
O
OH
OH
O
OH
HOOH
HO
O
O
O
OOO
NHN
O
N
HN
O
N
NH3+
Fe
desferrioxamine B
ferrioxamine B complexβ110 = 1030.2 ; pFe = 26.6
O
O
O
O
O O
Fe
O
OOO
O OHN
O
OHN
O
NH
References [4,5,6] 16

Iron in Biology Society For Free Radical Biology and Medicine Crumbliss 17pH
0 2 4 6 8 10 12
pFe
0
10
20
30
40
Exochelin MN (I)Ferrioxamine B (II)
Plot of Fe(III) complex stability,expressed as pFe (Panel 9), as afunction of pH for two siderophores,exochelin MN (I) and ferrioxamine B (II).Although they have approximately thesame stability at pH 6.0, above this pHexochelin MN has a higher affinity forFe(III) and below this value ferrioxamineB exhibits a higher affinity. This is dueto different levels of competition from H+
for the Fe(III) binding sites, due todifferent pKa values for the donor groups (shown in blue) in these two siderophorechelators.
NNH
O
NHN
NH
ONH
NH2
ON
O
OHOH
NHO
NH
OOHNH2
(I)
(II)
HOO
OH HOO
NHN
O
NHN
O
N
NH3+
O
17
Influence of pH on Fe(III)Influence of pH on Fe(III)--Chelate StabilityChelate Stability
Reference [9,12]
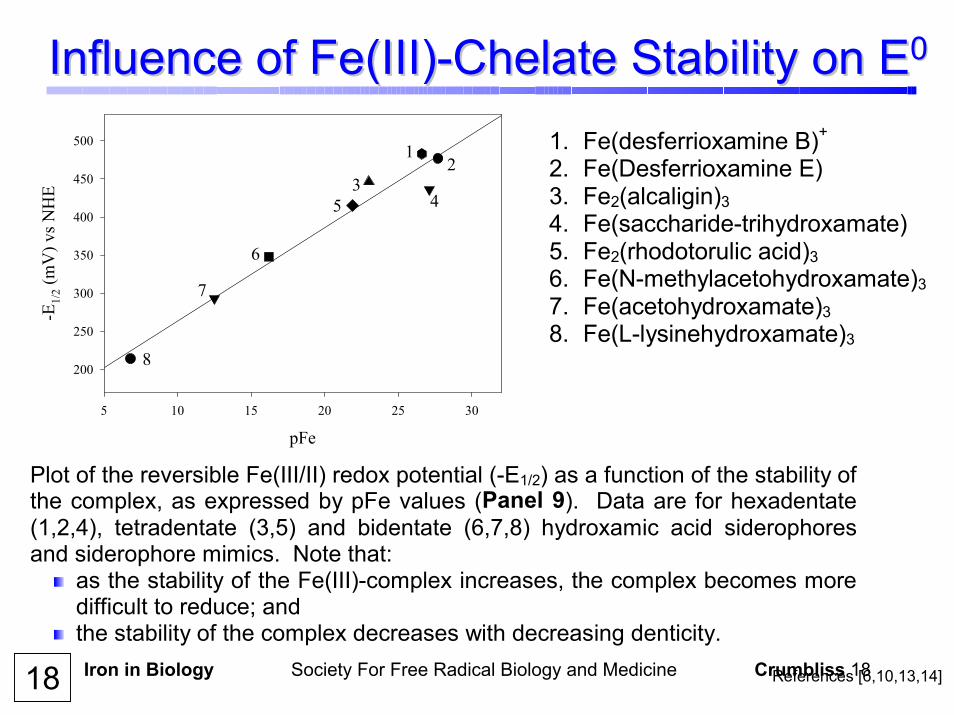
Iron in Biology Society For Free Radical Biology and Medicine Crumbliss 18
pFe5 10 15 20 25 30
-E1/
2 (m
V) v
s NH
E
200
250
300
350
400
450
500
8
6
53
12
7
4
Influence of Fe(III)Influence of Fe(III)--Chelate Stability on EChelate Stability on E00
Plot of the reversible Fe(III/II) redox potential (-E1/2) as a function of the stability of the complex, as expressed by pFe values (Panel 9). Data are for hexadentate(1,2,4), tetradentate (3,5) and bidentate (6,7,8) hydroxamic acid siderophoresand siderophore mimics. Note that:
as the stability of the Fe(III)-complex increases, the complex becomes moredifficult to reduce; and
the stability of the complex decreases with decreasing denticity.
1. Fe(desferrioxamine B)+ 2. Fe(Desferrioxamine E) 3. Fe2(alcaligin)3 4. Fe(saccharide-trihydroxamate) 5. Fe2(rhodotorulic acid)3 6. Fe(N-methylacetohydroxamate)37. Fe(acetohydroxamate)3 8. Fe(L-lysinehydroxamate)3
18 References [6,10,13,14]

Iron in Biology Society For Free Radical Biology and Medicine Crumbliss 19
Influence of Fe(III)Influence of Fe(III)--Chelation on KineticsChelation on Kinetics
Fe
OH2
OOO
O OH2
H3C
NCH3
Fe
OH2
OH2
OH2H2O
H2O OH2
Fe
OH2
OOH2O
H2O OH2
H3C
NCH3
N
H3C
H3C
OHO
H3C
NCH3
O
OHN
H3C
H3C
H+
H+
3+ 2+
1+
k1 = 1.8 M-1s-1
k2 = 8.1 x 102 M-1s-1
Iron chelate formation places a strong electron donor in the first coordination shell, which labilizes the remaining aquated coordination sites. This is illustrated here for the reaction of hexa(aquo)iron(III) with N-methylacetohydroxamic acid, a siderophore mimic. Incorporation of the bidentate hydroxamate group in the first coordination shell labilizes the remaining aquo ligands by a factor of ~500 (k2/k1).
19 Reference [5,15]

Iron in Biology Society For Free Radical Biology and Medicine Crumbliss 20
ReferencesReferences1. Crichton, R. (2001) Inorganic Biochemistry of Iron Metabolism, John
Wiley & Sons, Ltd, New York. 2. Harris, W. R. (2002) in Molecular and Cellular Iron Transport
(Templeton, D. M., Ed.) pp 1-40, Marcel Dekker, Inc., New York. 3. Martell, A. E. and Smith, R. M., Eds. (1974, 1975, 1976, 1977, 1982,
1989) Critical Stability Constants, Plenum Press, New York. 4. Raymond, K. N. and Stintzi, A. (2002) in Molecular and Cellular Iron
Transport (Templeton, D. M., Ed.) pp. 273-320, Marcel Dekker, New York.
5. Albrecht-Gary, A.-M., and Crumbliss, A. L. (1998) in Metal Ions in Biological Systems Vol. 35, Iron Transport and Storage in Microoganisms, Plants and Animals (Sigel, A. and Sigel, H., Ed.) pp. 239-327, Marcel Dekker,New York.
6. Crumbliss, A. L. and Boukhalfa, H. (2002) BioMetals 15, 325-339. 7. Aisen, P. (1998) in Metal Ions in Biological Systems Vol. 35, Iron
Transport and Storage in Microoganisms, Plants and Animals (Sigel, A. and Sigel, H., Ed.) pp. 585-632, Marcel Dekker,New York.
8. Harris, W. R. (1986) J. Inorg. Biochem. 27, 41-52. 9. Schwarzenbach, G., and Schwarzenbach, K. (1963) Helv. Chem. Acta
46, 1390-1400.
20

Iron in Biology Society For Free Radical Biology and Medicine Crumbliss 21
ReferencesReferences10. Spasojevic, I., Armstrong, S. K., Brickman, T. J., and Crumbliss, A. L. (1999)
Inorg. Chem. 38, 449-454. 11. Helm, L. and Merbach, A. E. (1999) Coord. Chem. Rev. 187, 151-181. 12. Dhungana, S., Miller, M.J., Dong, L., Ratledge, C. and Crumbliss, A. L.
(2002) manuscript in preparation. 13. Wirgau, J. I., Spasojevic, I., Boukhalfa, H., Batinic-Haberle, I., and
Crumbliss, A. L. (2002) Inorg. Chem. 41, 1464-1473. 14. Dhungana, S., Heggemann, S., H., Gebhardt, P. Möllmann, U. and
Crumbliss, A.L. (2002) Inorg. Chem. 41, submitted for publication. 15. Caudle, M. T., and Crumbliss, A. L. (1994) Inorg. Chem. 33, 4077-4085.
21
AcknowledgementsAcknowledgementsI thank my co-workers, some of whose names appear in the References, fortheir hard work, questions, ideas, and intellectual stimulation. Our work in thisarea is supported by the National Science Foundation, the National Instutitesof Health, and the American Chemical Society Petroleum Research Fund.