validation of virologic cut-offs for preemptive therapy and im
TRANSCRIPT

NEW MICROBIOLOGICA, 34, 229-254, 2011
Management of human cytomegalovirus infectionin transplantation: validation of virologic cut-offsfor preemptive therapy and immunological cut-offs
for protection
Giuseppe Gerna1, Daniele Lilleri1, Milena Furione2, Fausto Baldanti21Laboratori Sperimentali di Ricerca, Area Trapiantologica, Fondazione IRCCS Policlinico San Matteo, Pavia, Italy;2S.S. Virologia Molecolare, S.C. Virologia e Microbiologia, Fondazione IRCCS Policlinico San Matteo, Pavia, Italy
INTRODUCTION
Human cytomegalovirus (HCMV) infection(Figure 1A to D) represents a major infectiouscomplication in the post-transplant period forboth solid-organ transplant recipients (SOTR)(Fishman and Rubin, 1998) and hematopoieticstem cell transplant recipients (HSCTR) (Boeckhand Ljungman, 1998). In seronegative recipientsof transplants from seropositive donors, HCMV
Corresponding authorProf. Giuseppe GernaLaboratori Sperimentali di Ricerca Area TrapiantologicaFondazione IRCCS Policlinico San Matteo27100 Pavia, ItalyE-mail: [email protected]; [email protected]
can be transmitted through the transplanted or-gan or HSC (primary HCMV infections), or theadministration of non-leukocyte-depleted bloodunits or blood products. This event occurs in aminor number of cases in HSCTR, whereas it isquite frequent in SOTR. In the great majority oftransplant recipients, reactivation of latent HCMVin HCMV-seropositive patients does occur. In ad-dition, in transplant recipients undergoing im-munosuppressive regimens, besides reactivationof latent HCMV, a reinfection by a new HCMVstrain may occur, thus associating reactivation oflatent virus with a new HCMV infection.Clinically, HCMV infection can be either asymp-tomatic or symptomatic. The presence of clinicalmanifestations is directly related to the inabilityof the HCMV-specific T-cell immune response to
Human cytomegalovirus (HCMV) still causes major viral complications in the post-transplant period of both solid-or-gan (SO) and hematopoietic stem cell (HSC) transplant (T) recipients (R). Diagnosis of HCMV infection is mostlymade by real-time PCR-based methodologies, which allow quantification of viral DNA in both blood and, if required,organ tissues or local secretions. HCMV infection/disease can be prevented by either universal prophylaxis or pre-emptive therapy. The latter approach has mostly been used in European Transplantation Centers upon reaching pre-determined cut-off levels of viral load, predictive of high risk for HCMV disease. In our Department, these cut-offs arehigher for SOTR (3x105 DNA copies/ml whole blood) and lower for HSCTR (3x104 DNA copies/ml). Antiviral therapyis continued until viral DNA disappearance from blood or tissues. However, the authentic long-term control of HCMVinfection is achieved when HCMV-specific CD4+ and CD8+ T-cells are detected in blood or tissues. Proposed im-munological cut-off levels conferring protection are: one HCMV-specific CD4+ and three CD8+ T-cells/ml blood forHSCTR, and 0.4 HCMV-specific T-cells/ml for both CD4+ and CD8+ in SOTR. However, anti-rejection in SOTR and an-ti-GvHD in HSCTR steroid therapies make patients susceptible to HCMV infection, even in the presence of protectivelevels of specific T-cells.
KEY WORDS: Human cytomegalovirus, Solid organ transplant, Hematopoietic stem cell transplant, CD4+ T-cells,CD8+ T-cells, Viral load cut-off, Pre-emptive therapy, Immunological cut-off
SUMMARY
Received April 06, 2011 Accepted May 19, 2011

control and limit the infection. The balance be-tween virus replication and host defense immunemechanisms is the major pathogenetic factor un-derlying the clinical presentation of HCMV in-fection in different transplantation settings.When asymptomatic, HCMV infection consistsof detection of virus and virus products in bloodduring programmed routine examinations, orfrom local organ sites, in which collection of bodysecretions or tissue biopsies are performed blind-ly (Table 1). Monitoring of HCMV infection inthese cases, must be associated with monitoringof immunological response in order to adopt themost timely and efficacious preventive approach.Sequential testing for HCMV load in blood is cur-rently performed when using preemptive therapy
strategies, which are aimed at detecting HCMVinfection prior to appearance of clinical symp-toms (see below).The alternative approach to prevention of HCMVinfection is HCMV prophylaxis, i.e. administra-tion of antiviral drugs to all patients in the post-transplant period starting from the first day oftransplant for a period of 3-6 months (see below).A third approach to treat HCMV infection in thepost-transplant period consists of delaying anytype of intervention until appearance of HCMVdisease (deferred therapy). This may consist of asystemic syndrome or end-organ disease or both(Ljungman et al., 2002).HCMV systemic syndrome is characterized byfever greater than 38°C for at least two days,
230 G. Gerna, D. Lilleri, M. Furione, F. Baldanti
FIGURE 1 - HCMV replication in hu-man embryonic lung fibroblast cell cul-tures. (A) HCMV-infected human fi-broblast 120h post-infection, showingintranuclear (IN) and intracytoplasmic(IC) inclusion bodies. (B to D) Electronmicroscopy of HCMV-infected humanfibroblast. (B) Horseshoe-shaped in-tranuclear inclusion (IN). (C) Densebodies (arrows). (D) Maturing virusparticles at the level of the nuclear mem-brane (From Revello MG and Gerna G,p. 681, Clin Microbiol Rev 2002; 15:680-715).
TABLE 1 - Spectrum of HCMV infection/disease in humans.
HCMV infection Clinical symptoms Diagnosis
Asymptomatic None Detection of low level of virus/virus products in blood or tissue biopsies
Systemic (HCMV syndrome) Fever, leukopenia, thrombocytopenia, Detection of high level of virus/virus> hepatic transaminases, malaise, products in blood (see below)
headache
Localized (end-organ disease) Symptoms/signs of organ Detection of virus and virus productsinvolvement (gastrointestinal, in organ biopsies or local secretions
pulmonary, hepatic, neural or other) in association or not with virus detection in blood

malaise, leukopenia, thrombocytopenia, increasein hepatic transaminases greater than twofold thenormal upper limit (with exclusion of liver trans-plant recipients), and presence of HCMV or itsproducts in blood, in the absence of other possi-ble causes. HCMV end-organ disease is charac-terized by symptoms and signs of organ involve-ment (gastrointestinal, pulmonary, hepatic, neu-ral, or, less frequently, involving other organ sites)in association with virus detection in biopsy tis-sues or local body secretions (independently ofvirus presence in blood), and in the absence ofother possible causes of organ disease (Table 1).However, in most patients, systemic and local in-fections are associated.
Diagnosis of HCMV infectionApart from transplantation centers using pro-phylaxis as an HCMV preventive measure, whichare not routinely involved in diagnosing HCMVinfection (at least until prophylaxis is continued),in all the other centers early diagnosis of HCMVinfection is mandatory in view of deciding themost adequate intervention measures aimed atpreventing or treating HCMV disease.A major preliminary consideration in the devel-opment of guidelines for diagnosing HCMV in-fection in transplanted patients concerns the typeof specimen to be examined for virus detection.While virus detection in clinical samples such asurine, saliva, cervical secretions or breast milk,may be useful for other purposes, in patientstransplant recipients, only two major types ofspecimens must be taken into consideration:whole blood samples for diagnosis of dissemi-nated infection, and biopsy samples or local se-cretion samples for diagnosis of end-organHCMV infection/disease.Until the mid 1980s, the only available assay wasconventional virus isolation in human embryon-ic lung fibroblasts (HELF). However, this assaywas cumbersome and time-consuming, provid-ing results in 2-3 weeks when infectious virus waslow in titer. At that time, the availability of mon-oclonal antibodies (MAbs) led to the developmentof rapid virus isolation methods based upon useof the shell vial technique (Gleaves et al., 1984),which allowed detection of HCMV-infected cellsprior to the appearance of a cytopathic effect(Figure 1A). Virus strain recovery, however, is stillof critical importance for studies of cell tropism
and susceptibility to antiviral drugs (Erice et al.,1998). The shell-vial assay, which provides resultsin 24-48 h post-inoculation, gained popularity inthe transplantation setting as the “viremia” assay(Gerna et al., 1990). This assay allowed rapid virusdetection in blood samples by staining an inocu-lated HELF monolayer with anti-p72 MAb 24-48h p.i. and visualization of infected cell nuclei bythe immunofluorescence (IFA) or the im-munoperoxidase antibody (IPA) technique.However, this assay does not allow virus strainrecovery.A major advancement in the diagnosis of dissem-inated HCMV infection was made at the end of1980s with the introduction of the antigenemiaassay (Figure 2B) developed simultaneously bytwo groups of researchers in Groningen, theNetherlands, and in Pavia, Italy (Van der Bij et al.,1988; Revello et al., 1989). This assay detected thepresence of HCMV pp65 in nuclei of peripheralblood leukocytes (neutrophils and monocytes) byusing a pool of pp65-specific MAbs (Gerna et al.,1992) and IFA or IPA. Compared to viremia, antigenemia displayedhigher sensitivity and a shorter turnaround time,requiring about two hours to complete. The testperformance does not require expensive equip-ment and is fairly easy to perform. However, itmay not be possible with neutrophil counts lessthan 1,000 cells/ml, and blood samples should beprocessed within 6-8 hours of collection to pre-vent decrease in sensitivity.One of the major achievements of the last decadeshas been the development of the polymerasechain reaction (PCR) and PCR-based moleculartechniques for detection of HCMV genomic DNAand RNA transcripts in clinical samples followingamplification of target sequences by a cyclic en-zymatic procedure. Besides PCR-based target am-plification techniques, methods for detectingHCMV DNA through signal amplification havebeen developed using either branched DNAprobes (Kolberg et al., 1996) or RNA probes(Baldanti et al., 1997; Hebart et al., 1998).A different amplification technique (nucleic acidsequence-based amplification, NASBA) was alsodeveloped for detection of HCMV transcripts inblood of immunocompromised patients (Gernaet al., 1999).More recently, automatic extractionprocedures and a real-time readout format haveallowed better standardization of PCR conditions.
HCMV management in transplant recipients 231

Viral organ localization, either in the presence orabsence of virus in blood, can be diagnosed byexamining organ biopsies or local secretions byone of the above reported methods, showing lo-cal presence of infectious virus or viral DNA.
Quantification of HCMV load in blood or tissuesQuantitative determination of viral load in bloodwas shown to possess a high positive predictivevalue for the development of HCMV disease(Grossi et al., 1995). Among the different methods reported above,rapid virus isolation by the shell vial assay wasfound to provide quantitative results correlating
with HCMV replication (Figure 2A). However, themethod was found to lack sensitivity (Gerna etal., 2001). The possibility of using the viremia as-say to quantify viral load was based on the ob-servation that a single PBL was able to infect asingle HELF (Gerna et al., 1990). In other words,the number of p72-positive HELF nuclei corre-lated with the number of PBL carrying infectiousvirus. Measurement of viral load in patients with dis-seminated HCMV infection allowed the responseto antiviral therapy to be monitored and repre-sented the basis for the adoption of pre-emptivetherapy strategies (Grossi et al., 1995; Locatelli etal., 1994). In addition, the shell vial assay allowed
232 G. Gerna, D. Lilleri, M. Furione, F. Baldanti
FIGURE 2 - (A) Viremia, indicated by the presence, in a shell vial monolayer, of HCMV p72-positive fibroblast nu-clei following cocultivation with peripheral blood leukocytes carrying infectious virus, and immunostaining withfluorescein-conjugated p72-specific monoclonal antibody (Gerna et al., 1990) (B) Antigenemia ex-vivo, showing im-munofluorescent staining, with a pool of pp65-specific monoclonal antibodies, of pp65-positive peripheral bloodpolymorphonuclear leukocytes from a patient with AIDS and disseminated HCMV infection (Van der Bij et al., 1988;Revello et al., 1989; Gerna et al., 1992). (C) Antigenemia in vitro, showing pp65-positive polymorphonuclear leuko-cytes from a healthy blood donor following cocultivation with HCMV-infected human umbilical vein endothelialcells and immunofluorescent staining with the same pool of pp65-specific monoclonal antibodies used in B (Revelloet al., 1998). (D) Circulating cytomegalic endothelial cell with a pp65-positive leukocyte (arrow) (From Revello MGand Gerna G, p. 690, Clin Microbiol Rev 2002; 15: 680-715).

quantification of viral load in different biologicalmaterials, such as CSF, BAL and tissue biopsies.However, a major advance in the quantification ofHCMV load was the introduction of the antigen-emia assay (Figure 2B). Threshold values associ-ated with the appearance of HCMV disease wereidentified following weekly monitoring of anti-genemia values in SOTR (Grossi et al., 1995).Thus, pre-emptive therapy of disseminatedHCMV infection could be initiated upon reach-ing cut-off antigenemia values preceding the ap-pearance of clinical symptoms in both SOTR andHSCTR (Locatelli et al., 1994; Grossi et al., 1995;Boeckh, 1999). The antigenemia assay was also ofgreat help in monitoring the response to antiviraltreatment, including the emergence of drug-re-sistant HCMV strains (Baldanti et al., 1998, 2004).In the late 1990s, the in vitro transfer of HCMVpp65 into PBL allowed standardization of the as-say (Figure 2C) (Revello et al., 1998; Gerna et al.,1998a), even though some pitfalls of the assaypersist:1) in HSCTR it cannot be used during engraft-ment (Locatelli et al., 1994; Limaye et al.,1997);
2) a paradoxical rise in antigenemia values mayoccur during ganciclovir (GCV) treatment(Gerna et al., 1998c, 2003c, 2005a), and thusantigenemia quantification does not correlatedirectly with viral replication (Gerna et al.,1998b). During the performance of the anti-genemia assay, circulating cytomegalic en-dothelial cells were detected in peripheralblood of immunocompromised patients andfetuses with congenital HCMV infection(Figure 2D).
These limitations were overcome by the intro-duction of PCR-based molecular assays (Gernaet al., 1991; Humar et al., 1999; Emery et al., 2000;Caliendo et al., 2002; Razonable et al., 2003). Mostlaboratories performing viral load quantificationhave been moving to real-time PCR techniques.These techniques are more precise and rapid, pos-sess a broader linear range and a higher output,and have a lower risk of carryover contamination(Cortez et al., 2003; Mengelle et al., 2003;Piiparinen et al., 2004). Viral DNA can be quan-tified in different blood fractions (leukocytes orplasma) or whole blood (Gerna et al., 1994b).Although no general consensus has been reachedand plasma and whole blood both provide prog-
nostic and diagnostic information (Hamprecht etal., 1998; Razonable et al., 2002), whole blood isnow accepted as the specimen of choice, since itallows detection of HCMV DNA earlier and ingreater amounts compared to plasma.However, in order to make results obtained bydifferent laboratories comparable, a standardizedPCR methodology is warranted, as well as refer-ence to an International Standard or to externalquality control panels by all laboratories. Inter-institutional comparison of quantitative DNA val-ues through the exchange of specimens and useof external quality control samples has been ini-tiated among laboratories (Lilleri et al., 2009b;Pang et al., 2009).Given the general consensus that higher viral loadvalues correlate with an increased risk for devel-opment of HCMV disease, it must be specifiedthat viral load changes less than 5-fold (0.7 log10copies/ml) may not be considered significant(Kotton et al., 2010). In general, there is a good but not uniform cor-relation between antigenemia and DNA viral loadlevels in blood (Caliendo et al., 2002). One recentstudy comparing viral load in blood by real-timePCR (DNAemia) and antigenemia in SOTRshowed that use of DNAemia versus antigenemiafor initiation of pre-emptive therapy significant-ly reduced the number of patients requiring treat-ment without an increase in HCMV disease(Gerna et al., 2007).Diagnosis of end-organ disease such as hepatitisor gastrointestinal infection must be performedby immunohistochemistry or in situ DNA hy-bridization (Lautenschlager et al., 2006).However, quantification of viral DNA in tissuebiopsies as well as BAL samples is preferable dueto greater sensitivity without loss of specificity.As for BAL samples, in the absence of random-ized studies, quantification of viral DNA has beensuggested as helpful in predicting pneumonitis(Westall et al., 2004; Chemaly et al., 2005; Gernaet al., 2009). In particular, the HCMV DNA con-centration in the epithelial lining fluid diluted inBAL would be critical for differentiating betweensymptomatic and asymptomatic infections(Zedtwitz-Liebenstein et al., 2004). Diagnosis ofother organ syndromes, such as central nervoussystem disease or retinitis would require detec-tion of viral DNA in CSF or aqueous humour(Gerna et al., 1994a).
HCMV management in transplant recipients 233

As a final consideration, it is important to recallthat an International Reference Standard (con-taining 5x106 DNA copies/mL) has become re-cently available through WHO (Freyer et al.,2010), whereas a consensus has yet to be reachedon the type of blood sample to be used.
Prevention of HCMV infection/diseaseTwo major prevention strategies have been usedin the last two decades to avoid HCMV infec-tion/disease in the post-transplant period: uni-versal prophylaxis and pre-emptive therapy. Sometransplantation centers have also been using hy-brid methods based on the combined use of bothstrategies. Use of deferred (symptomatic) therapy, i.e. initi-ation of antiviral treatment upon appearance ofclinical symptoms, is now restricted to a fewtransplantation centers. Antiviral drugs used fordifferent strategies have been ganciclovir (GCV)and foscarnet (PFA) initially, and more recentlyvalganciclovir (VGCV), and, as a second line ther-apy, cidofovir (CDV). There were no differences inlong-term outcomes of HCMV disease betweeniv GCV and VGCV (Åsberg et al., 2009), whileGCV was found to be equally effective in univer-sal prophylaxis vs pre-emptive therapy (Small etal., 2006).The major advantage of universal prophylaxis(which is currently based on antiviral drug ad-ministration for 3-6 months after transplantation)is its daily administration in the absence of viraltesting. However, problems related to this strate-gy are mostly due to the toxic effects of prolongedantiviral drug administration (myelotoxicity-neu-tropenia for GCV and VGCV, electrolyte distur-bances for PFA, and nefrotoxicity for CDV). Inaddition, potential drawbacks of universal pro-phylaxis may include the following:1) late HCMV disease may affect up to 29% ofprophylactically treated patients (Limaye et al.,2004, 2006; Eid et al., 2008; Sun et al., 2008);
2) prophylaxis may interfere with and delayHCMV-specific T-cell reconstitution due toabortive stimulation of viral antigens (Hakki etal., 2003). However, a recent study reports thatVGCV prophylaxis does not appear to impairthe development of HCMV-specific immunityin lung transplantation (Humar et al., 2009;Snyder et al., 2011);
3) suboptimal drug dosage used according to the
prophylaxis schedule may induce the emer-gence of drug-resistant strains.
On the other hand, pre-emptive therapy refers toadministration of antiviral drugs when viral loadin blood reaches levels predictive of HCMV dis-ease, but prior to onset of clinical symptoms. Themajor advantage of preemptive therapy with re-spect to universal prophylaxis is treatment of asmaller proportion of patients for a shorter peri-od of time. However, the pre-emptive therapy ap-proach requires continuous virological monitor-ing. Additional advantages of pre-emptive thera-py include:1) savings in terms of drug toxicity;2) savings in terms of patient management costs(Kusne et al., 1999);
3) according to some authors, an efficacy com-parable to that of prophylaxis in preventingthe indirect effects of HCMV infection, such asgraft failure/rejection and fungal/bacterial in-fections (Singh et al., 2005, 2006a, 2006b;Khoury et al., 2006). However, although in theabsence of statistical support, Snydman et al.(2009) stated a superiority of prophylaxis onthe indirect effects of HCMV infection, name-ly mortality and opportunistic infections.
Initiation of pre-emptive therapy in SOTRSince the beginning of the 1990s, at our Institute(Fondazione IRCCS Policlinico San Matteo) therecommended preventive strategy for SOTR hasbeen pre-emptive therapy with GCV until disap-pearance of virus from blood. Initially, the anti-genemia assay was used for deciding the onset ofpre-emptive therapy. It was observed that HCMVdisease in D+/R+ and D-/R+ SOTR started in thepresence of antigenemia levels ≥400 pp65-posi-tive PBL/2x105 examined (Grossi et al., 1995).Therefore, an arbitrary cut-off threshold of 100pp65-positive PBL/2x105 examined was estab-lished for initiating antiviral treatment (Table 2).In addition, due to the reported higher risk forHCMV disease, a much lower antigenemia cut-off was adopted for D+/R- SOTR (≥2 pp65-posi-tive PBL/2x105 examined, or 1 pp65-positivePBL/2x105 confirmed twice in a week). This strat-egy was used for about a decade and was flankedby viremia and DNAemia testing as complemen-tary assays. During this period, it was observedthat antigenemia did not correlate closely andconsistently with virus replication. The molecular
234 G. Gerna, D. Lilleri, M. Furione, F. Baldanti

basis of this discrepancy was later clarified by invitro studies (Gerna et al., 1998c, 2003c, 2005a). To overcome these problems, efforts were direct-ed towards the development of molecular assaysdetecting virus products more directly related tovirus replication. One of these new approacheswas the detection of HCMV late transcripts, i.e.the development of the NASBA assay, allowingspecific detection of unspliced mRNAs in a DNAbackground (Gerna et al., 1999). A prospective randomized open-label trial inves-tigated whether HCMV pp67 (late) mRNA couldrepresent a valuable alternative to quantitativeantigenemia in a group of 82 HTR and LTR di-vided into two arms. In the NASBA arm, both pri-mary and recurrent infections were treated uponthe first confirmed positive NASBA result, whilein the antigenemia arm, primary infections weretreated upon the first confirmed positive resultand recurrent infections upon a cut-off of 100pp65-positive leukocytes. Results showed that thenumber of treated/infected patients was signifi-cantly higher in the NASBA arm, as was the num-ber of treated/relapsing patients. In addition, inthe NASBA arm the overall number of days oftherapy was significantly higher. It was concluded that pp67 NASBA could safelyreplace antigenemia, with some disadvantages interms of overtreatment of patients and greaterduration of overall treatment (Gerna et al.,2003a). In the same study, the retrospective de-
termination of quantitative HCMV DNAemiaopened the door for subsequent prospective stud-ies aimed at defining better pre-emptive therapy-based control of HCMV infections in SOTR, ascompared to antigenemia.A retrospective analysis of DNAemia and anti-genemia in SOTR performed at our Instituteshowed that a DNAemia cut-off of 300,000 DNAcopies/ml whole blood could provide positive andnegative predictive values as high as those givenby antigenemia, i.e. >90% with reference to pa-tients requiring pre-emptive therapy (Lilleri et al.,2004; Gerna and Lilleri, 2006). On this basis, atrial was designed to compare the DNAemia cut-off of 300,000 DNA copies/ml with the antigene-mia cut-off of 100 pp65-positive/2x105 leukocytesfor pre-emptive treatment of both primary andreactivated HCMV infections (Table 2) (Gerna etal., 2007). Results showed that, compared to anti-genemia, the selected DNAemia cut-off:1) significantly reduced the number of patientsreceiving treatment (20% vs 40%);
2) was capable of safely guiding pre-emptivetherapy of both primary and reactivatedHCMV infections in SOTR;
3) did not significantly modify the overall dura-tion of treatment.
Furthermore, no case of HCMV disease occurredin patients treated after reaching the cut-off.However, four patients (three in the antigenemia,and one in the DNAemia arm) suffered from
HCMV management in transplant recipients 235
TABLE 2 - Levels of HCMV viral load detected in blood in association with disease or selected for preemptive therapy.
HCMV clinical condition Viral load in blood
pp65-antigenemia DNAemia
Disease in SOTR 400 pp65-pos PBL/2x105 PBL 1,000,000 DNA copies/ml bloodexamined (Grossi et al., 1995) (Gerna et al., 1998b)
Preemptive therapy in SOTR* 100 pp65-pos PBL/2x105 300,000 DNA copies/ml blood examined (historical) (Gerna et al., 2007)(Gerna et al., 2007)
Disease in HSCTR ? ?
Preemptive therapy in HSCTR** 2 pp65-pos PBL/2x105 10,000 DNA copies/ml bloodexamined (historical) (Lilleri et al., 2007a; Gerna et al., 2008b)
*Indicated DNAemia cut-offs were found in randomized trials to be safe for both primary and reactivated infections. Many SOT Centers still start treatment ofprimary infection (high-risk patients) upon first confirmed virus detection by either antigenemia or DNAemia. **Most HSCT Centers still start treatment uponfirst confirmed detection of virus in blood by either assay.

HCMV disease prior to reaching the relevant cut-off. In these patients, organ localization in lungs(two patients) and gastrointestinal tract (two pa-tients) occurred prior to virus dissemination inblood (Einsele et al., 1995; Parente et al., 1998;Sanchez et al., 2001; Humar et al., 2009). Thus,in the presence of local clinical symptoms andabsence of virus in blood, local samples (biopsytissues or secretions) must be taken for exami-nation.This inconvenience could be avoided by pro-phylaxis, which however presents the majorproblem of late-onset HCMV disease after pro-phylaxis discontinuation. Although it was shownin one study in kidney transplant recipients thatprophylaxis reduced HCMV infection, while graftsurvival was improved at four years post-trans-plant, thus suggesting a beneficial effect of pro-phylaxis (Kliem et al., 2008), late-onset diseaseappears to be associated with higher rates ofmortality (Limaye et al., 2006) and graft loss(Arthurs et al., 2008).A major advantage of the use of a single DNAemiacut-off for both primary and reactivated infec-tions was the lack of a greater risk of HCMV dis-ease or recurrence in primary infections com-pared with treatment started upon first virus de-tection in blood (Gerna et al., 2007). This ap-proach allowed a greater and more sustainedantigenic stimulus during the development of aprimary T-cell-mediated immune response. Still,in several transplantation centers, a differentialapproach to preventive treatment of primary andreactivated infections is maintained, and recent-ly guidelines promulgated by the TTS of Americasuggested prolonging prophylaxis from three tosix months in D+/R- patients (Kotton et al., 2010).In order to export a pre-determined DNAemiacut-off to different centers, different commercialand in-house-developed methods using anInternational Standard should be standardized.As mentioned above, in a recent multi-centerstudy guided by our Institution, we observed anacceptable range of variation (≤0.7 log10 or five-fold) among different centers (Lilleri et al.,2009b). Thus, it was recommended that at par-ticipating centers, pre-emptive therapy should bestarted when reaching levels of ≥5.5 log10(300,000) copies/ml blood in SOTR.Besides the cut-off of 300,000 DNA copies/mlwhole blood for systemic infections, a study on
pre-emptive therapy for systemic and pulmonaryHCMV infection in lung transplant recipients test-ed a tentative cut-off of 5.0 log10 (100,000) DNAcopies/ml BAL fluid (Gerna et al., 2009). This cut-off was intermediate between that reported byWestall et al. (2004) and that found by Chemalyet al. (2005). The conclusion of this study was thatmonitoring of HCMV infection in both blood andBAL as well as pre-emptive therapy for both types(sites) of infection when reaching the pre-deter-mined HCMV cut-offs, should be recommendedin LTR to prevent HCMV disease. Although de-finitive cut-offs for intervention have not reachedconsensus, other centers have used cut-offs in therange of 5,000 DNA copies/mL plasma (Humaret al., 1999) or 1,000-10,000 copies/mL wholeblood (Li et al., 2003; Khoury et al., 2006; Reischiget al., 2008). Since our cut-offs were determinedon several hundreds of SOT recipients and we didnot encounter problems of HCMV disease, wecontinue using our cut-offs higher than thoseadopted by other transplantation centers.Prophylaxis and pre-emptive therapy have beenonly partially investigated in the pediatric trans-planted patient population, (Seu et al., 1997; Kelly,2006; Spivey et al., 2007; Lilleri et al., 2007a). Astudy comparing prophylaxis followed by pre-emptive therapy with pre-emptive therapy alonein pediatric patients undergoing liver transplan-tation found that, although numbers of both in-fected and treated patients were comparable inthe two arms, the median number of total days ofantiviral therapy was significantly higher in theprophylaxis arm. Therefore, the trial was inter-rupted and prophylaxis replaced with pre-emp-tive therapy alone (Gerna et al., 2008a).
Initiation of pre-emptive therapy in HSCTRStarting in the 1990s, several European HSCTcenters employed confirmed qualitative PCR orantigenemia results to guide pre-emptive therapy(Ljungman et al., 1998a; Hebart et al., 2001;Reusser et al., 2002). Then, in a prospective ran-domized trial, qualitative determination of HCMVimmediate-early transcripts (IE-mRNA) by NAS-BA was compared to antigenemia and retrospec-tively to DNAemia as a potential guiding param-eter for pre-emptive therapy in HSCTR (Gerna etal., 2003b). Results showed that IEmRNA is moresensitive than antigenemia and DNAemia in de-tecting active HCMV infection. However, many
236 G. Gerna, D. Lilleri, M. Furione, F. Baldanti

patients had to be submitted to treatment withrespect to antigenemia.Thus, efforts were directed towards selection of asafe DNAemia cut-off for pre-emptive therapy ofHCMV infections in HSCTR. To this aim, a ret-rospective analysis of DNAemia levels observedin transplant recipients at the time of initiation ofantigenemia-guided pre-emptive therapy was per-formed (Lilleri et al., 2004). It was found thatamong HSCTR only 19.6% of patients treated up-on first confirmed antigenemia positivity reachedDNAemia levels above a cut-off of 10,000 DNAcopies/ml whole blood, whereas 94.7% of un-treated antigenemia patients had DNAemia levelsbelow this cut-off. Thus, a cut-off of 10,000 DNAcopies/ml whole blood showed a positive predic-tive value of 91.7%, and a negative predictive val-ue of 28.6% with respect to HSCTR requiringantigenemia-based pre-emptive therapy.Therefore in HSCTR, use of a quantitative, ratherthan a qualitative, approach to pre-emptive ther-apy appeared suitable to increase the negativepredictive value without modifying the positivepredictive value with respect to qualitative anti-genemia (Lilleri et al., 2004).This hypothesis was verified in a single-centerrandomized prospective open-label study aimedat defining the effectiveness of the real-timeDNAemia versus antigenemia cut-off as the guid-ing parameter for pre-emptive treatment in pe-diatric HSCTR (Table 2) (Lilleri et al., 2007a).Using a cut-off of 10,000 DNA copies/ml wholeblood in the DNAemia arm, and first positive anti-genemia in the antigenemia arm in a cohort of178 HSCTR (89 assigned to the DNAemia arm,and 89 in the antigenemia arm), it was found thatin the presence of a comparable number ofHCMV infections in the two arms a significantlylower number of patients required treatment inthe DNAemia arm (18%) with respect to the anti-genemia arm (31%) (p=0.026). Thus, a DNAemiacut-off-guided pre-emptive treatment proved safeand effective in controlling HCMV infection inthe HSCT setting, thereby avoiding unnecessaryantiviral treatment in a significant proportion ofpatients. Comparable results were obtained in anadult HSCTR population, where the major dif-ference with respect to the pediatric populationwas that the HCMV infection rate was almostdouble (80% vs 40%) (Table 2) (Gerna et al.,2008b).
In addition, in agreement with a previous report(Verkruyse et al., 2006) an observational studyshowed that adoption of a cut-off level of 4 log10(10,000) HCMV DNA copies/ml blood for start-ing pre-emptive therapy is safe and more cost-ef-fective (significantly reducing the number of pa-tients receiving pre-emptive therapy) than start-ing treatment upon the first positive result.Furthermore, the utility of the progressive re-placement of antigenemia with DNAemia as aguiding method for pre-emptive therapy was sup-ported by the frequent rise in antigenemia levelsdetected after initiation of pre-emptive therapyin allogeneic HSCTR, in the absence of steroidtreatment for GvHD or antiviral drug resistance(Gerna et al., 2005a). In addition, the incidence of HCMV infection andpre-emptive treatment in patients with refracto-ry solid tumors receiving nonmyeloablative allo-geneic SCT were shown to be significantly re-duced compared to recipients of standard SCTfor hematologic malignancies (Zambelli et al.,2005).
Immunological controlImmunological control of HCMV infection in-volves both the innate immune and the adaptiveimmune response to HCMV.
a) Innate immunityInnate immune response seems to play an im-portant role both in the defense against HCMVinfection as well as in priming the adaptive im-mune response. HCMV stimulates toll-like re-ceptors (TLRs), thus activating signal transduc-tion pathways. These cause secretion of inflammatory cytokinesand recruitment of cells of the innate immunesystem, as well as upregulation of costimulatorymolecules important for the activation of adaptiveimmunity (Boehme and Compton, 2004; Boehmeet al., 2006). In the murine model, TLR-mediatedsignal transduction pathways result in the pro-duction of interferon by dendritic cells (DC) andmacrophages, and activation of natural killer(NK) cells (Tabeta et al., 2004; Delale et al., 2005).NK cells have been shown to contribute to virusclearance in the experimental model of murineCMV infection (Polic’ et al., 1998) and to provideprotection against murine CMV infection(Bukowski et al., 1985), while some strains of
HCMV management in transplant recipients 237

mice resistant to murine CMV became suscepti-ble upon NK cell depletion (Scalzo et al., 1992).In humans, although the role of NK cells in thedefense against HCMV infection has not been in-vestigated in depth, NK activity was shown to in-crease in the control of both primary and recur-rent HCMV infection (Venema et al., 1994;Hadaya et al., 2008; Stern et al., 2008).
b) Adaptive immunityAdaptive immune responses consist of both hu-moral and T-cell-mediated immune responses.HCMV is a potent immunogen, eliciting a strongimmune response from both arms of the immunesystem.
Humoral immune responseAlthough the role of the humoral immune re-sponse is debated, there are indications thatCMV-specific antibodies play a somewhat pro-tective role against both HCMV and murine CMV,namely in limiting dissemination and diseaseseverity (Jonjic’ et al., 1994; Boppana and Britt,1995). Thus far, viral glycoproteins gB and gHhave been considered the major targets of theHCMV neutralizing antibody response (Britt etal., 1990; Rasmussen et al., 1991). Recently, how-ever, a major target of the HCMV neutralizing an-tibody response was identified in a glycoproteincomplex present in the virus envelope of wild-type HCMV strains cultured in endothelial andepithelial cells.This pentamer contains three gene products ofthe UL131-128 locus of the HCMV genome,which were identified in 2004 as indispensablefor HCMV infection of endothelial cells and trans-fer of virus and viral products from infected cellsto PBL (Hahn et al., 2004). Subsequently, pUL130and pUL128 were shown to form a complex withgH and gL (in the absence of pUL131), which wasrequired for epithelial and endothelial cell tro-pism (Wang and Shenk, 2005). Finally,gH/gL/pUL131/pUL130/pUL128 were shown toform a protein pentamer complex mediating en-try into epithelial and endothelial cells (Ryckmanet al., 2008). As mentioned above, this complexhas been preliminarily shown to be the target ofa number of potent neutralizing human mono-clonal antibodies elicited during both primaryand reactivated HCMV infection in both the im-munocompetent and the immunocompromised
host (Macagno et al., 2010). The protective effectof these neutralizing antibodies remains to be de-termined.In the mouse model, memory B-cell transfer pro-vided long-term protection from the lethal courseof infection that is invariably seen in immune-de-ficient animals (Klenovsek et al., 2007). Transferof memory B-cells was also effective in protectinganimals from an already ongoing viral infection.T-cells were not involved in this process. In addi-tion, in the guinea pig model, both passive(Chatterjee et al., 2001) and active immunization(Schleiss et al., 2004) have been shown to reduceboth fetal HCMV infection and disease in guineapigs. In humans, it is well known that transfer ofHCMV-specific antibodies from a seropositivemother to a newborn infant is able to protectagainst HCMV infection transmitted by seropos-itive blood transfusion units (Yeager et al., 1981).Furthermore, mothers with preconceptionalHCMV immunity are known to transmit the in-fection to the fetus at a much lower rate thanmothers experiencing primary HCMV infection(Stagno et al., 1982; Fowler et al., 1992). However,the role of HCMV hyperimmune globulin orHCMV-specific monoclonal antibody in prevent-ing or curing HCMV disease in transplant recip-ients is controversial (Ljungman et al., 1992,1998b; Machado et al., 2000; Boeckh et al., 2001).The role of hypogammaglobulinemia as a riskfactor for HCMV infection in the post-transplantperiod remains to be defined.
Cell-mediated immune responseHowever, based on the observation that HCMVdisease usually occurs in immunocompromisedpatients with profound cellular immunodefi-ciency, it is well accepted that the cell-mediatedimmune response is the major mechanism ofcontrol for HCMV infection/disease. Within theT-cell immune response, HCMV-specific CD8+ T-cells, CD4+ T-cells, and γδ T-cells all seem to playan important role in the control of HCMV reac-tivation.As for CD8+ T-cells, using the murine CMV mod-el, it was shown that the selective depletion oflymphocyte subsets revealed that CD8+ T-cellswere the most important factor in the immunecontrol of murine CMV infection (Polic’ et al.,1998). In humans, in the HSCT setting, the de-velopment of CD8+ cytotoxic T lymphocytes
238 G. Gerna, D. Lilleri, M. Furione, F. Baldanti

(CTL) was shown to correlate with protection andrecovery from HCMV disease (Quinnan et al.,1982; Li et al., 1994),whereas lack of a CD8+ T-cellresponse was associated with development ofHCMV disease (Reusser et al., 1991). Pioneerstudies by Riddell et al. (1992) and Walter et al.(1995) definitively showed that adoptive transferof CD8+ T-cell clones expanded in vitro resulted inimmune reconstitution in the post-transplant andfinally in protection from HCMV disease in therecipients. Similarly, in the SOT setting, CD8+ T-cell re-sponses, in addition to the CD4+ T-cell immuneresponse, were repeatedly reported to protectagainst HCMV disease in the post-transplant pe-riod (Reusser et al., 1999; Sester et al., 2002;Radha et al., 2005; Shlobin et al., 2006).However,in the absence of CD4+ T-cell help, CD8+ T-cellsare not able per se to confer effective long-termprotection from HCMV infection (see below).Within the CD8+ arm of the T-cell immune re-sponse, the proportion of CD8+ T-cells commit-ted to develop an anti-HCMV response, is sur-prisingly large, ranging from 10% in healthy in-dividuals up to 40% in elderly individuals(Gillespie et al., 2000; Khan et al., 2004; Crough etal., 2005; Sylvester et al., 2005). The biologicalreason for such an extended T-cell immune re-sponse is not yet known.The fine specificity of CD8+ T-cell responses andthe viral proteins to which they are directed havebeen extensively investigated. By taking advan-tage of using overlapping 15-mer peptides fromall 213 ORF of the HCMV genome in ex-vivo T-cell assays, it was found that CD8+ (and/or CD4+)T-cells are directed towards more than 70%ORFs. The most immunodominant antigens represent-ing targets for CD8+ T-cells are pUL123 (IE-1),pUL122 (IE-2) and pUL83 (pp65). The differentantigens acting as targets for CD8+ T-cells are re-ported in a recently published review (Croughand Khanna, 2009). It must be borne in mind thatthe largest response does not necessarily repre-sent the most effective in limiting HCMV repli-cation (Crough and Khanna, 2009).The two major HCMV-specific T-cell subpopula-tions (CD4+, CD8+) can be investigated for boththeir functional markers, such as the expressionof IFN-γ, IL-2, granzyme, and perforin, and theirphenotypic markers, including CD45RA,
CD45RO, CCR7, CD27, CD28, and so on. Highperforin expression has been claimed as an easy-to-measure prognostic marker of severe viral re-activation soon after HSCT (Pietersma et al.,2010). Different sets of T-cell subpopulations canbe recognized by analyzing surface expressionmarkers. In particular, the analysis of the chemokine re-ceptor CCR7 (which enables cells to migrate tothe lymph nodes) and the different isoforms ofthe tyrosine phosphatase CD45 allows the studyof T-cell differentiation (Sallusto et al., 1999).Naïve T-cells (CD45RA+ CCR7+) switch from theCD45RA isoform to CD45RO, and memory T-cells, according to the differential CCR7 expres-sion, are divided into central memory T-cells (TCMcells; CD45RA- CCR7+), which are able to migrateto lymph nodes and display a high proliferationpotential, and effector memory T-cells (TEM cells;CD45RA- CCR7-) which exert effector functionsin peripheral tissue. However, it has been shown that a proportion ofTEM cells, in case of persistent infection, can re-vert to the RA isoform of CD45 (TEMRA cells;CD45RA+ CCR7-) after the acute phase of infec-tion (Callan et al., 1998; Wills et al., 1999). In par-ticular, this event was recently found to occur inan elevated percentage of HCMV-specific CD8+
(51%) and CD4+ (33%) T-cells during the first yearafter primary infection, thus reaching levels sim-ilar to those of subjects with remote HCMV in-fection (Lilleri et al., 2008b). In addition, CD45RA expression correlated withHCMV disappearance from blood, while the lev-el of specific CD45RA+ T-cells during the firstmonths after primary infection in pregnantwomen was significantly lower in mothers whotransmitted the infection to the fetus than inmothers who did not transmit. These results sup-ported the conclusion that HCMV-specific effec-tor T-cells reverting to the CD45RA+ phenotypemay represent true long-lived memory lympho-cytes in the HCMV-specific pool. Two typical features of the HCMV-specific CD8+
T-cell response include the accumulation of anoligoclonal T-cell repertoire (HCMV-specificclonotypes) (Price et al., 2005; Day et al., 2007)and the accumulation of HCMV-specific CD8+ T-cells occurring with age (Ouyang et al., 2003;Khan et al., 2004). This event, termed “memoryinflation” has been shown to extend to murine
HCMV management in transplant recipients 239

CMV (Karrer et al., 2003) and to the HCMV-spe-cific CD4+ T-cell response (Pourgheysari et al.,2007). Expansion of HCMV-specific CD8+ T-cellsin the elderly is invariably oligoclonal or evenmonoclonal and is considered to be a contribut-ing factor in immune senescence (Khan et al.,2002; Pawelec et al., 2005). In addition, it isthought that the HCMV immunodominancemight impair immunological responses to otherpathogens (Trzonkowski et al., 2003). On the oth-er hand, it seems possible that a “bystander” ef-fect of infection with heterologous stimulation ofT-cell populations of other specificities may oc-cur (Crough et al., 2005).In addition to MHC class I-restricted CD8+ T-cells,evidence has accumulated indicating that alsoCD4+ T-cells are essential for control of HCMV in-fection. This conclusion has been reached bothin mice (Jonjic’ et al., 1990; Polic’ et al., 1998) andhumans, with special reference to the transplan-tation setting (Kumar et al., 2009; BenMarzouk-Hidalgo et al., 2011). In SOT recipients, it was ob-served that low levels of HCMV-specific CD4+ T-cells correlated with emergence of infectiouscomplications (Sester et al., 2001, 2005), while ef-fector-memory T-cells were required for recoveryfrom infection (Gamadia et al., 2003). Similarly,in HSCT recipients CD4+ T-cells were found to beassociated with protection from HCMV disease(Li et al., 1994; Hebart et al., 2002; Tormo et al.,2010). In addition, HCMV-specific CD4+ T-cellswere required for expansion of CD8+ CTLs(Einsele et al., 2002).As with HCMV-specific CD8+ T-cells, a very highfrequency of CD4+ T-cells was observed in someblood donors (Sylwester et al., 2005). The mostfrequent targets of HCMV-specific CD4+ T-cellswere products of the TLR14, UL16, UL55, andUL83 genes (Crough and Khanna, 2009). WhileHCMV-specific CD4+ T-cells have been consid-ered to display only an indirect role (help in main-taining the antibody response and expandingCD8+ T-cells), more recently gB-specific andpp65-specific CD4+ T-cells with cytolytic activityhave been reported ((Elkington et al., 2004;Casazza et al., 2006).Similar to CD8+ T-cells, an oligoclonal or mono-clonal expansion of CD4+ T-cells has been re-ported in some hematologic clinical conditions(Garrido et al., 2007; Crompton et al., 2008). Inaddition, regulatory T-cells (Tregs) have been
shown to correlate with recovery of HCMV-spe-cific CD8+ T-cells after allogeneic HSCT, prevent-ing GvHD and promoting immune recovery(Pastore et al., 2011).Finally, the γδ T-cell subset, representing less than6% of T-cells in blood of healthy people, but amuch larger fraction in body surfaces exposed tothe contact with external material, seems to pos-sess an important role in protection againstHCMV disease. Both findings in the mouse mod-el (Ninomiya et al., 2000) and in transplanted pa-tients (Lafarge et al., 2001) indicate that γδ T-cellsare involved in the anti-HCMV immune response.Their expansion has been associated with reso-lution of HCMV infection in HSCT recipients, asconfirmed by a recent report (Knight et al., 2010).
Methods for immunological monitoring of HCMV infectionWhile virologic monitoring is critical for evalu-ating the severity of HCMV infection, immuno-logic monitoring, with special reference to the T-cell immune response, is mandatory for drawingprognostic indications. A number of ex-vivo T-cellassays have been developed for measuring theHCMV-specific cellular response. An ideal assayshould be able to determine both the specificityof CD4+ and CD8+ T-cells as well as function.Among assays determining HCMV-specific T-cells, the most widely used have been peptide-conjugated MHC class I tetramers. These assayscan measure virus-specific CD8+ T-cell frequency,but are epitope-specific and require determina-tion of patients’ HLA type. While some reportshave found a correlation between CD8+ T-cellsmeasured by tetramer-based technology and pro-tection (Gratama et al., 2001, 2010), other studieshave questioned this conclusion (Crough et al.,2007).Among functional assays, intracellular cy-tokine staining (ICS) for IFN-γ (or other cy-tokines), following T-cell stimulation with HCMVpeptide mixtures or whole antigen lysate, is notHLA-restricted, and requires, like the tetramer-based method, the availability of a flow cytome-ter. Both CD4+ and CD8+ can be determined, andseveral clinical studies have shown a correlationbetween the presence of HCMV-specific cytokine-producing T-cells and absence of HCMV diseaseand viremia, or viceversa (Sester et al., 2001, 2005;Shlobin et al., 2006; Kumar et al., 2009; Gerna etal., 2006; Mattes et al., 2008).
240 G. Gerna, D. Lilleri, M. Furione, F. Baldanti

The other widely used functional assay is the en-zyme-linked immunospot (ELISPOT) assay thatdetermines T-cells producing IFN-γ followingstimulation of T-cells with peptide pools or anti-gen lysates, without differentiating between CD4+
and CD8+ T-cells. IFN-γ released by activated T-cells is captured by a bound antibody and de-tected by a labelled anti-IFN-γ antibody. ELISPOTis not standardized; however, it seems able to pre-dict HCMV disease (Mattes et al., 2008).All the above-mentioned assays are also quanti-tative. Our laboratory developed a novel assay tomeasure both CD4+ and CD8+ T-cells followingstimulation with a number of HCMV antigens(Lozza et al., 2005). This assay was based on dif-ferentiation of monocyte-derived autologous den-dritic cells (DCs) in 5-7 days in the presence ofGM-CSF and IL-4 (Sallusto et al., 1994), followed
by DC infection with an endotheliotropic (HU-VEC+) and leukotropic (Leuk+) HCMV strain(Gerna et al., 2005b) and subsequent 18-24 h in-cubation with PBMC for stimulation (Figure 3).ICS and flow cytometry analysis allowed thequantification of functional HCMV-specific CD4+
and CD8+ T-cells following stimulation by multi-ple HCMV antigens. Using this assay, it becameevident how clinically relevant it was to quantifyT-cells for careful immunologic monitoring ofHCMV infection. Previous studies often drewconflicting conclusions, i.e. while one study re-ported that protection in transplanted patientscorrelated with IE-1, but not pp65-specific CD8+
T-cells (Bunde et al., 2005), another study con-cluded that pp65-specific T-cells were also im-portant for protection (Lacey et al., 2006; Croughet al., 2007; Lilleri et al., 2007b).
HCMV management in transplant recipients 241
FIGURE 3 - Flow chart of a novel method for simultaneous quantification of HCMV-specific CD4+ and CD8+ T-cellsby using autologous monocyte-derived HCMV-infected immature dendritic cells (Lozza et al., 2005). FACS analysisof CD4+ and CD8+ cytokine-producing HCMV-specific T-cells.

It has become evident that the analysis of a sin-gle antigen, as done in most studies, may be in-sufficient to predict the clinical course of HCMVinfection (Sylwester et al., 2005; Lilleri et al.,2007b). The determination of phenotypic markers incombination with functional assays, such as thatdeveloped in our laboratory, has allowed the iden-tification of functionally special T-cell popula-tions at different stages of differentiation. For in-stance, the reversion of effector memory T-cells tothe CD45RA+ phenotype has been shown to cor-relate with virus clearance from blood, while ahigh percentage of these CD45RA+ CD4+ T-cellswas found to protect against HCMV transmissionto the fetus (Lilleri et al., 2008b).The quantiFERON-CMV assay is a commercial-ly available ELISA-based assay measuring IFN-γlevels released in blood following stimulationwith a range of CD8+ HCMV-specific T-cell epi-topes from a number of HCMV viral proteins spe-cific for a wide range of HLA class I alleles(Walker et al., 2007; Westall et al., 2008). This as-say is being evaluated in clinical trials (Kumar etal., 2009; Fleming et al., 2010).
Definition of protective threshold levels of the HCMV-specific T-cell response in transplant recipients: validation in clinical trialsNotwithstanding the different methodological ap-proaches used, it is now agreed that early recon-stitution of the T-cell immune response preventsor limits HCMV infection, whereas a delay or lackin the development of the cellular immune re-sponse is the pathogenetic basis for severe infec-tion, often leading to a systemic syndrome and/ororgan disease in the absence of antiviral treat-ment. PBMC stimulation with pp65 and/or IE-1peptide mixtures for rapid monitoring of HCMV-specific T-cell responses in both the immuno-competent and the immunocompromised hostwas found to underestimate real T-cell responsesagainst HCMV (Lilleri et al., 2007b, 2009c). Dueto the partial response to single HCMV proteinsor epitopes, the above-mentioned novel assaybased on T-cell stimulation by autologous DCs in-fected with an endotheliotropic HCMV strain wasdeveloped in our laboratory to allow a more com-prehensive evaluation of the T-cell immune re-sponse (Lozza et al., 2005; Gerna et al., 2005b).
Based on results obtained by testing a series ofHCMV-seropositive and HCMV-seronegativehealthy blood donors, subjects with virus-specif-ic cellular immunity were those with more than0.4 HCMV-specific CD4+ and CD8+ T-cells/mlblood (Lozza et al., 2005).A prospective clinical study evaluating HCMV-specific CD4+ and CD8+ T-cell reconstitution inyoung allogeneic HSCT recipients found that re-covery of both HCMV-specific CD4+ and CD8+ T-cell immunity occurred in all 39 HCMV-seropos-itive (R+) patients within six months, and in 6/18(33%) HCMV-seronegative (R-) patients within 12months (Lilleri et al., 2006). In addition, the re-ceiver-operator characteristics (ROC) analysisshowed that levels of HCMV-specific CD4+ lessthan 1/ml and CD8+ less than 3 cells/ml (Table 3)are not protective against recurrent infection(Figure 4A). Similar cut-off levels were reportedby others (Tormo et al., 2010). Patients without prompt recovery of HCMV-spe-cific cellular immunity underwent repeatedepisodes of recurrent infection, as previously re-ported by others (Krause et al., 1997; Hebart et al.,2002; Boeckh et al., 2003). Another study onHCMV reconstitution in young patients receivingT-cell-depleted allogeneic HSCT showed that, us-ing the same cut-offs as above, immune reconsti-tution was delayed in these patient groups com-pared to patients receiving unmanipulated HSCTs(Lilleri et al., 2009a). A parallel study on immunecontrol of HCMV infection and HCMV immunereconstitution in adult allogeneic HSCT recipientsreported that levels of three CD8+ and one CD4+
242 G. Gerna, D. Lilleri, M. Furione, F. Baldanti
TABLE 3 - Protective threshold levels of HCMV-specific CD4+ and CD8+ T-cell responses in
transplant recipients.
Patients Protective levels* of HCMV-specific T-cells/µl
CD4+ CD8+
Immunocompetent 0.4 0.4Immunocompromised- SOTR 0.4 0.4
- HSCTR 1.0 3.0
*Upon steroid treatment for rejection episodes in SOTR or GvHD in HSCTR,the reported cut-offs may change.
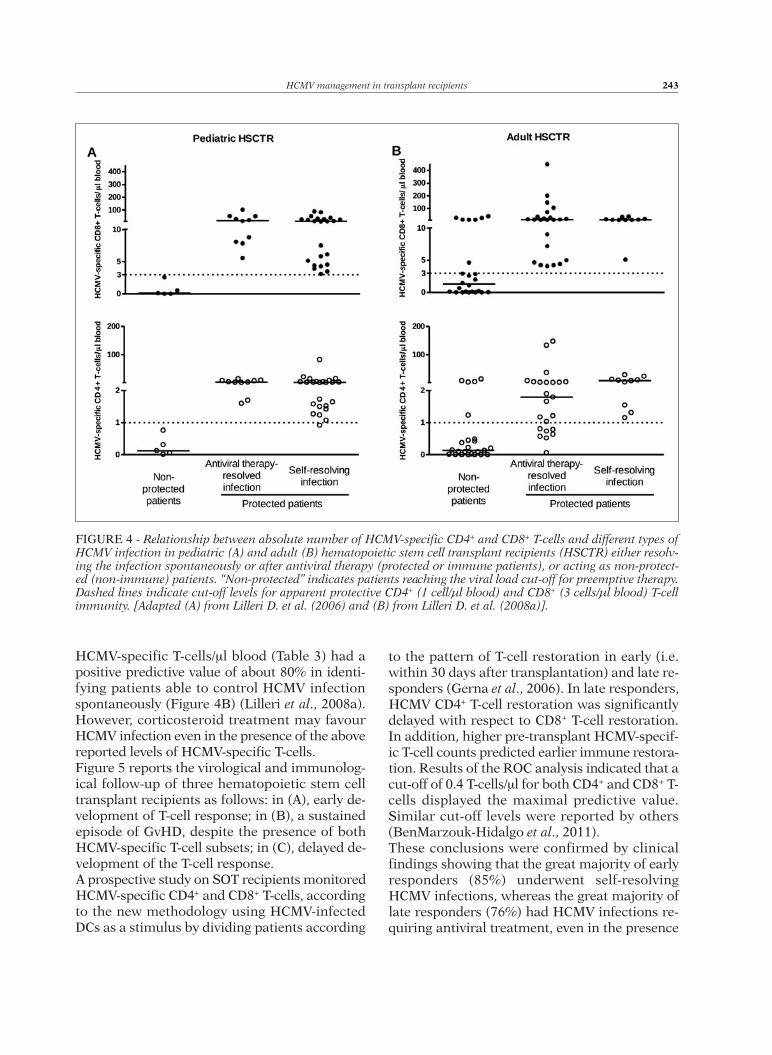
HCMV-specific T-cells/ml blood (Table 3) had apositive predictive value of about 80% in identi-fying patients able to control HCMV infectionspontaneously (Figure 4B) (Lilleri et al., 2008a).However, corticosteroid treatment may favourHCMV infection even in the presence of the abovereported levels of HCMV-specific T-cells.Figure 5 reports the virological and immunolog-ical follow-up of three hematopoietic stem celltransplant recipients as follows: in (A), early de-velopment of T-cell response; in (B), a sustainedepisode of GvHD, despite the presence of bothHCMV-specific T-cell subsets; in (C), delayed de-velopment of the T-cell response.A prospective study on SOT recipients monitoredHCMV-specific CD4+ and CD8+ T-cells, accordingto the new methodology using HCMV-infectedDCs as a stimulus by dividing patients according
to the pattern of T-cell restoration in early (i.e.within 30 days after transplantation) and late re-sponders (Gerna et al., 2006). In late responders,HCMV CD4+ T-cell restoration was significantlydelayed with respect to CD8+ T-cell restoration.In addition, higher pre-transplant HCMV-specif-ic T-cell counts predicted earlier immune restora-tion. Results of the ROC analysis indicated that acut-off of 0.4 T-cells/ml for both CD4+ and CD8+ T-cells displayed the maximal predictive value.Similar cut-off levels were reported by others(BenMarzouk-Hidalgo et al., 2011). These conclusions were confirmed by clinicalfindings showing that the great majority of earlyresponders (85%) underwent self-resolvingHCMV infections, whereas the great majority oflate responders (76%) had HCMV infections re-quiring antiviral treatment, even in the presence
HCMV management in transplant recipients 243
FIGURE 4 - Relationship between absolute number of HCMV-specific CD4+ and CD8+ T-cells and different types ofHCMV infection in pediatric (A) and adult (B) hematopoietic stem cell transplant recipients (HSCTR) either resolv-ing the infection spontaneously or after antiviral therapy (protected or immune patients), or acting as non-protect-ed (non-immune) patients. “Non-protected” indicates patients reaching the viral load cut-off for preemptive therapy.Dashed lines indicate cut-off levels for apparent protective CD4+ (1 cell/µl blood) and CD8+ (3 cells/µl blood) T-cellimmunity. [Adapted (A) from Lilleri D. et al. (2006) and (B) from Lilleri D. et al. (2008a)].
of specific CD8+ T-cells. In the past, predominantlong-term protection conferred by HCMV-specif-ic CD4+ T-cells has been repeatedly reported(Sester et al., 2001, 2002). In addition, it wasshown in renal transplant recipients with primaryinfection that asymptomatic patients had a CD4+
T-cell response preceding the CD8+ T-cell re-sponse, whereas in symptomatic patients the spe-cific CD4+ T-cell response was delayed in the pres-ence of a CD8+ T-cell response that was not pro-
tective (Gamadia et al., 2003). Thus, HCMV-spe-cific CD4+ T-cells appear to play a major role inlong-term protection from HCMV infection/dis-ease.Figure 6 reports the virological and immunolog-ical follow-up of three solid organ transplant re-cipients showing: in (A) early development, in (C)delayed development of the T-cell response, whilein (B) steroid therapy due to a rejection episodecaused loss of T-cell immunity.
244 G. Gerna, D. Lilleri, M. Furione, F. Baldanti
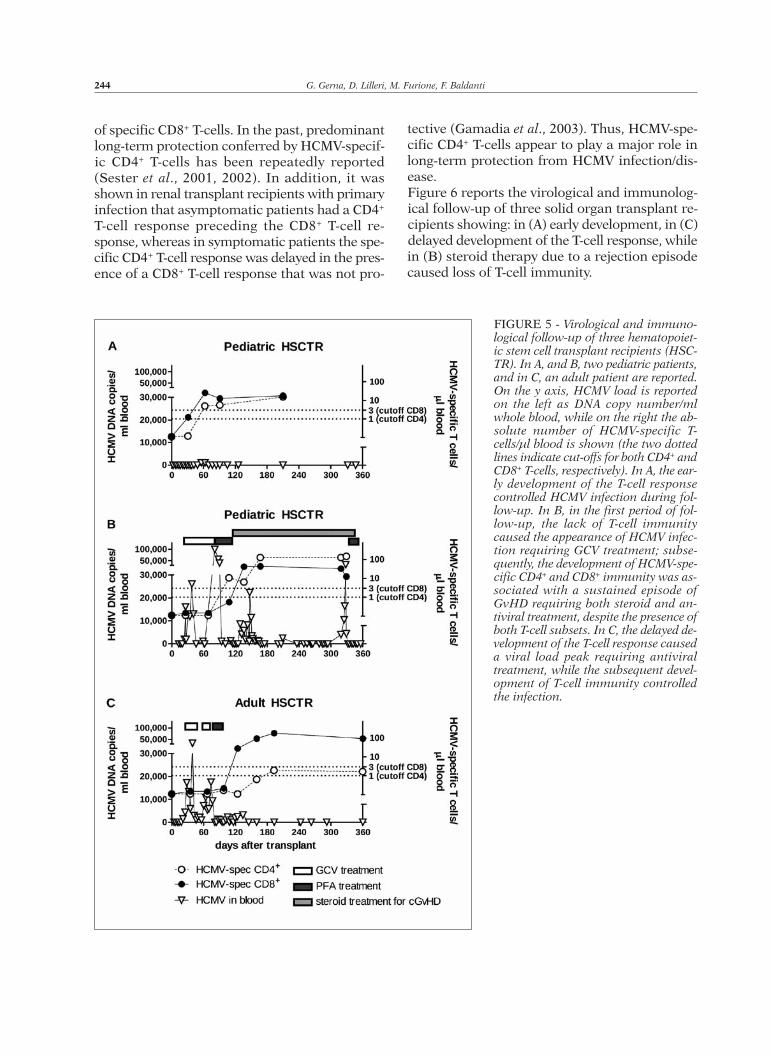
FIGURE 5 - Virological and immuno-logical follow-up of three hematopoiet-ic stem cell transplant recipients (HSC-TR). In A, and B, two pediatric patients,and in C, an adult patient are reported.On the y axis, HCMV load is reportedon the left as DNA copy number/mlwhole blood, while on the right the ab-solute number of HCMV-specific T-cells/µl blood is shown (the two dottedlines indicate cut-offs for both CD4+ andCD8+ T-cells, respectively). In A, the ear-ly development of the T-cell responsecontrolled HCMV infection during fol-low-up. In B, in the first period of fol-low-up, the lack of T-cell immunitycaused the appearance of HCMV infec-tion requiring GCV treatment; subse-quently, the development of HCMV-spe-cific CD4+ and CD8+ immunity was as-sociated with a sustained episode ofGvHD requiring both steroid and an-tiviral treatment, despite the presence ofboth T-cell subsets. In C, the delayed de-velopment of the T-cell response causeda viral load peak requiring antiviraltreatment, while the subsequent devel-opment of T-cell immunity controlledthe infection.

CONCLUDING REMARKS
From the clinical standpoint, it appears reason-able to conclude that simultaneous immunolog-ical and virologic follow-up of individual patients
may improve the management of HCMV infec-tion in transplanted patients (Radha et al., 2005;Sester et al., 2005), thereby avoiding treatment ofpatients with an apparently efficient T-cell im-mune response. Given the ongoing debate be-
HCMV management in transplant recipients 245
FIGURE 6 -HCMV virological and im-munological follow-up of three solid or-gan transplant recipients. In A, and C,two heart transplant recipients (HTRs)are reported. In B, a lung transplant re-cipient (LTR) is shown. On the y axis,viral load expressed as copy number/mlwhole blood, is reported on the left,while on the right the absolute numberof HCMV-specific T cells/µl blood isshown (dotted line indicates predeter-mined cut-off for both CD4+ and CD8+
T-cells). In A, the maintenance of T-cellimmunity during the entire follow-upperiod prevented HCMV infection. In B,steroid therapy due to a rejectionepisode caused loss of T-cell immunityand appearance of a peak of HCMV in-fection in blood, which was treated withganciclovir (GCV); a high level ofHCMV load was also detected in bron-choalveolar lavage (BAL) fluid.However, the subsequent developmentof HCMV-specific CD4+ (first) and(then) CD8+ T-cell immunity controlledthe infection at both sites. In C, lack ofT-cell immunity caused the appearanceof multiple peaks of HCMV infection inblood, which required two courses ofGCV treatment; subsequently, develop-ment of HCMV-specific CD8+ (first) and(then) CD4+ T-cell response controlledthe infection.

tween prophylaxis and pre-emptive therapy inview of reducing costs of HCMV infection intransplant recipients, the identification of a co-hort of patients who are less likely to developHCMV reactivation may suggest discontinuationof HCMV monitoring. On the other hand, theidentification of another cohort of patients whoare more likely to develop HCMV reactivationmay prompt a prophylactic or therapeutic inter-vention.
ACKNOWLEDGMENTSThe authors are indebted to all the technical staff ofthe Servizio di Virologia and the medical staff ofdifferent transplantation centers of the FondazioneIstituto di Ricovero e Cura a Carattere ScientificoPoliclinico San Matteo. We are grateful to DanielaSartori for carefully preparing and checking the ma-nuscript. We also thank Laurene Kelly for revisionof the English.This work was partially supported by the Ministerodella Salute, Ricerca Corrente grant no. 80221, and the Italia-USA Project grant no. 28C5/3;and the Fondazione Carlo Denegri (FCD), Torino,Italy.
REFERENCES
ÅSBERG A., HUMAR A., JARDINE A.G., ROLLAG H.,PESCOVITZM.D., MOUAS H., ET AL. (2009). Long-termoutcomes of CMV disease treatment with valgan-ciclovir versus IV ganciclovir in solid organ trans-plant recipients. Am. J. Transplant. 9, 1205-1213.
ARTHURS S.K., EID A.J., PEDERSEN R.A., KREMERS W.K.,COSIO F.G., PATEL R., ET AL. (2008). Delayed-onsetprimary cytomegalovirus disease and the risk of al-lograft failure and mortality after kidney trans-plantation. Clin. Infect. Dis. 46, 840-846.
BALDANTI F., ZAVATTONI M., SARASINI A., GATTI M., CHEZZIL., GERNA G. (1997). Comparative quantification ofhuman cytomegalovirus DNA in blood of immuno-compromised patients by PCR and Murex HybridCapture System. Clin. Diagn. Virol. 8, 159-165.
BALDANTI F., SIMONCINI L., SARASINI A., ZAVATTONI M.,GROSSI P., REVELLOM.G., ET AL. (1998). Ganciclovirresistance as a result of oral ganciclovir in a hearttransplant recipient with multiple human cy-tomegalovirus strains in blood. Transplantation. 66,324-329.
BALDANTI F., LILLERI D., CAMPANINI G., COMOLLI G.,RIDOLFO A.L., RUSCONI S., ET AL. (2004). Human cy-tomegalovirus double resistance in a donor-posi-tive/recipient-negative lung transplant patient with
an impaired CD4-mediated specific immune re-sponse. J. Antimicrob. Chemother. 53, 536-539.
BENMARZOUK-HIDALGO O.J., CISNEROS J.M., CORDEROE., MARTÍN-PEÑA A., SANCHEZ B., MARTIN-GANDULC., ET AL. (2011). Therapeutic effect of the acquisi-tion of cytomegalovirus-specific immune responseduring preemptive treatment. Transplantation. 91,927-933.
BOECKHM., LJUNGMAN P.L. (1998). Cytomegalovirus in-fection after bone marrow transplantation, InTransplant Infection, Paya C. (Ed.) Lippincot-RavenPhiladelphia, PA, USA. 215-227.
BOECKHM. (1999). Current antiviral strategies for con-trolling cytomegalovirus in hematopoietic stem celltransplant recipients: prevention and therapy.Transpl. Infect. Dis. 1, 165-178.
BOECKH M., BOWDEN R.A., STORER B., CHAO N.J.,SPIELBERGER R., TIERNEY D.K., ET AL. (2001).Randomized, placebo-controlled, double-blindstudy of a cytomegalovirus-specific monoclonal an-tibody (MSL-109) for prevention of cy-tomegalovirus infection after allogeneic hematopoi-etic stem cell transplantation. Biol. Blood MarrowTransplant. 7, 343-351.
BOECKHM., LEISENRINGW., RIDDELL S.R., BOWDEN R.A.,HUANG M.L., MYERSON D., ET AL. (2003). Late cy-tomegalovirus disease and mortality in recipients ofallogeneic hematopoietic stem cell transplants: im-portance of viral load and T-cell immunity. Blood.101, 407-414.
BOEHME K.W., COMPTON T. (2004). Innate sensing ofviruses by toll-like receptors. J. Virol. 78, 7867-7873.
BOEHME K.W., GUERRERO M., COMPTON T. (2006).Human cytomegalovirus envelope glycoproteins Band H are necessary for TLR2 activation in per-missive cells. J. Immunol. 177, 7094-7102.
BOPPANA S.B., BRITT W.J. (1995). Antiviral antibody re-sponses and intrauterine transmission after pri-mary maternal cytomegalovirus infection. J. Infect.Dis. 171, 1115-1121.
BRITT W.J., VUGLER L., BUTFILOSKI E.J., STEPHENS E.B.(1990). Cell surface expression of human cy-tomegalovirus (HCMV) gp55-116 (gB): use ofHCMV-recombinant vaccinia virus-infected cells inanalysis of the human neutralizing antibody re-sponse. J. Virol. 6, 1079-1085.
BUKOWSKI J.F., WARNER J.F., DENNERT G., WELSH R.M.(1985). Adoptive transfer studies demonstrating theantiviral effect of natural killer cells in vivo. J. Exp.Med. 161, 40-52.
BUNDE T., KIRCHNER A., HOFFMEISTER B., HABEDANK D.,HETZER R., CHEREPNEV G., ET AL. (2005). Protectionfrom cytomegalovirus after transplantation is cor-related with immediate early 1-specific CD8 T cells.J. Exp. Med. 201, 1031-1036.
CALIENDO A.M., ST GEORGE K., ALLEGA J., BULLOTTA A.C.,GILBANE L., RINALDO C.R. (2002). Distinguishing cy-tomegalovirus (CMV) infection and disease with
246 G. Gerna, D. Lilleri, M. Furione, F. Baldanti

CMV nucleic acid assays. J. Clin. Microbiol. 40,1581-1586.
CALLANM.F., TAN L., ANNELS N., OGG G.S., WILSON J.D.,O’CALLAGHAN C.A., ET AL. (1998). Direct visualiza-tion of antigen-specific CD8+ T cells during the pri-mary immune response to Epstein-Barr virus in vi-vo. J. Exp. Med. 187, 1395-1402.
CASAZZA J.P., BETTS M.R., PRICE D.A., PRECOPIO M.L.,RUFF L.E., BRENCHLEY J.M., ET AL. (2006).Acquisition of direct antiviral effector functions byCMV-specific CD4+ T lymphocytes with cellularmaturation. J. Exp. Med. 203, 2865-2877.
CHATTERJEE A., HARRISON C.J., BRITT W.J., BEWTRA C.(2001). Modification of maternal and congenitalcytomegalovirus infection by anti-glycoprotein Bantibody transfer in guinea pigs. J. Infect. Dis. 183,1547-1553.
CHEMALY R.F., YEN-LIEBERMAN B., CHAPMAN J., REILLYA., BEKELE B.N., GORDON S.M., ET AL. (2005).Clinical utility of cytomegalovirus viral load inbronchoalveolar lavage in lung transplant recipi-ents. Am. J. Transplant. 5, 544-548.
CORTEZ K.J., FISCHER S.H., FAHLE G.A., CALHOUN L.B.,CHILDS R.W., BARRETT A.J., ET AL. (2003). Clinicaltrial of quantitative real-time polymerase chainreaction for detection of cytomegalovirus in pe-ripheral blood of allogeneic hematopoietic stem-cell transplant recipients. J. Infect. Dis. 188, 967-972.
CROMPTON L., KHAN N., KHANNA R., NAYAK L., MOSS P.A.(2008). CD4+ T cells specific for glycoprotein Bfrom cytomegalovirus exhibit extreme conservationof T-cell receptor usage between different individ-uals. Blood. 111, 2053-2061.
CROUGH T., BURROWS J.M., FAZOU C., WALKER S.,DAVENPORT M.P., KHANNA R. (2005).Contemporaneous fluctuations in T cell responsesto persistent herpes virus infections. Eur. J.Immunol. 35, 139-149.
CROUGH T., FAZOU C., WEISS J., CAMPBELL S., DAVENPORTM.P., BELL S.C., ET AL. (2007). Symptomatic andasymptomatic viral recrudescence in solid-organtransplant recipients and its relationship with theantigen-specific CD8(+) T-cell response. J. Virol. 81,11538-11542.
CROUGH T., KHANNA R. (2009). Immunobiology of hu-man cytomegalovirus: from bench to bedside. Clin.Microbiol. Rev. 22, 76-98.
DAY E.K., CARMICHAEL A.J., TEN BERGE I.J., WALLER E.C.,SISSONS J.G., WILLS M.R. (2007). Rapid CD8+ T cellrepertoire focusing and selection of high-affinityclones into memory following primary infectionwith a persistent human virus: human cy-tomegalovirus. J. Immunol. 179, 3203-3213.
DELALE T., PAQUIN A., ASSELIN-PATUREL C., DALOD M.,BRIZARD G., BATES E.E., ET AL. (2005). MyD88-de-pendent and -independent murine cytomegalovirussensing for IFN-alpha release and initiation of im-
mune responses in vivo. J. Immunol. 175, 6723-6732.
EID A.J., BAKRI S.J., KIJPITTAYARIT S., RAZONABLE R.R.(2008). Clinical features and outcomes of cy-tomegalovirus retinitis after transplantation.Transpl. Infect. Dis. 10, 13-18.
EINSELE H., EHNINGER G., HEBART H., WITTKOWSKI K.M.,SCHULER U., JAHN G., ET AL. (1995). Polymerasechain reaction monitoring reduces the incidence ofcytomegalovirus disease and the duration and sideeffects of antiviral therapy after bone marrow trans-plantation. Blood. 86, 2815-2820.
EINSELE H., ROOSNEK E., RUFER N., SINZGER C., RIEGLERS., LÖFFLER J., ET AL. (2002). Infusion of cy-tomegalovirus (CMV)-specific T cells for the treat-ment of CMV infection not responding to antiviralchemotherapy. Blood. 99, 3916-3922.
ELKINGTON R., SHOUKRY N.H., WALKER S., CROUGH T.,FAZOU C., KAUR A., ET AL. (2004). Cross-reactiverecognition of human and primate cytomegalovirussequences by human CD4 cytotoxic T lymphocytesspecific for glycoprotein B and H. Eur. J. Immunol.34, 3216-3226.
EMERY V.C., SABIN C.A., COPE A.V., GOR D., HASSAN-WALKER A.F., GRIFFITHS P.D. (2000). Application ofviral-load kinetics to identify patients who developcytomegalovirus disease after transplantation.Lancet. 355, 2032-2036.
ERICE A., BORRELL N., LI W., MILLERW.J., BALFOUR H.H.JR. (1998). Ganciclovir susceptibilities and analy-sis of UL97 region in cytomegalovirus (CMV) iso-lates from bone marrow recipients with CMV dis-ease after antiviral prophylaxis. J. Infect. Dis. 178,531-534.
FISHMAN J.A., RUBIN R.H. (1998). Infection in organ-transplant recipients. N. Engl. J. Med. 338, 1741-1751.
FLEMING T., DUNNE J., CROWLEY B. (2010). Ex vivo mon-itoring of human cytomegalovirus-specific CD8+ T-cell responses using the QuantiFERON®-CMV as-say in allogeneic hematopoietic stem cell transplantrecipients attending an Irish hospital. J. Med. Virol.82, 433-440.
FOWLER K.B., STAGNO S., PASS R.F., BRITT W.J., BOLLT.J., ALFORD C.A. (1992). The outcome of congeni-tal cytomegalovirus infection in relation to mater-nal antibody status. N. Engl. J. Med. 326, 663-667.
FREYER J.F., HEATH A.B., ANDERSON R., MINOR P.D., ANDthe Collaborative Study Group. (2010).Collaborative study to evaluate the proposed 1st
WHO International Standard for human cy-tomegalovirus (HCMV) for nucleic acid amplifica-tion (NAT)-based assays. WHO/BS/10.2138, pp. 1-40. Geneva, 18-22 Oct. 2010.
GAMADIA L.E., REMMERSWAAL E.B., WEEL J.F., BEMELMANF., VAN LIER R.A., TEN BERGE I.J. (2003). Primaryimmune responses to human CMV: a critical rolefor IFN-gamma-producing CD4+ T cells in protec-
HCMV management in transplant recipients 247

tion against CMV disease. Blood. 101, 2686-2692.GARRIDO P., RUIZ-CABELLO F., BÁRCENA P., SANDBERG Y.,CANTÓN J., LIMAM., ET AL. (2007). Monoclonal TCR-Vbeta13.1+/CD4+/NKa+/CD8-/+dim T-LGL lympho-cytosis: evidence for an antigen-driven chronic T-cell stimulation origin. Blood. 109, 4890-4898.
GERNA G., REVELLO M.G., PERCIVALLE E., ZAVATTONI M.,PAREA M., BATTAGLIA M. (1990). Quantification ofhuman cytomegalovirus viremia by using mono-clonal antibodies to different viral proteins. J. Clin.Microbiol. 28, 2681-2688.
GERNA G., ZIPETO D., PAREA M., REVELLO M.G., SILINIE., PERCIVALLE E., ET AL. (1991). Monitoring of hu-man cytomegalovirus infections and ganciclovirtreatment in heart transplant recipients by deter-mination of viremia, antigenemia and DNAemia.J. Infect. Dis. 164, 488-498.
GERNA G., REVELLO M.G., PERCIVALLE E., MORINI F.(1992). Comparison of different immunostainingtechniques and monoclonal antibodies to the low-er matrix phosphoprotein (pp65) for optimal quan-titation of human cytomegalovirus antigenemia. J.Clin. Microbiol. 30, 1232-1237.
GERNA G., BALDANTI F., SARASINI A., FURIONE M.,PERCIVALLE E., REVELLOM.G., ET AL. (1994a). Effectof foscarnet induction treatment on quantitationof human cytomegalovirus (HCMV) DNA in pe-ripheral blood polymorphonuclear leukocytes andaqueous humor of AIDS patients with HCMV re-tinitis. Antimicrob. Agents Chemother. 38, 38-44.
GERNA G., FURIONEM., BALDANTI F., SARASINI A. (1994b).Comparative quantitation of human cy-tomegalovirus DNA in blood leukocytes and plasmaof transplant and AIDS patients. J. Clin. Microbiol.32, 2709-2717.
GERNA G., PERCIVALLE E., TORSELLINI M., REVELLOM.G.(1998a) Standardization of the human cy-tomegalovirus antigenemia assay by means of invitro-generated pp65-positive peripheral blood poly-morphonuclear leukocytes. J. Clin. Microbiol. 36,3585-3589.
GERNA G., ZAVATTONIM., BALDANTI F., SARASINI A., CHEZZIL., GROSSI P., ET AL. (1998b). Human cy-tomegalovirus (HCMV) leukoDNAemia correlatesmore closely with clinical symptoms than antigen-emia and viremia in heart and heart-lung trans-plant recipients with primary HCMV infection.Transplantation. 65, 1378-1385.
GERNA G., ZAVATTONI M., PERCIVALLE E., GROSSI P.,TORSELLINI M., REVELLO M.G. (1998c). Rising lev-els of human cytomegalovirus (HCMV) antigene-mia during initial antiviral treatment of solid-or-gan transplant recipients with primary HCMV in-fection. J. Clin. Microbiol. 36, 1113-1116.
GERNA G., BALDANTI F., MIDDELDORP J.M., FURIONE M.,ZAVATTONI M., LILLERI D., ET AL. (1999). Clinical sig-nificance of expression of human cytomegaloviruspp67 late transcript in heart, lung, and bone mar-
row transplant recipients as determined by nucle-ic acid sequence-based amplification. J. Clin.Microbiol. 37, 902-911.
GERNA G., BALDANTI F., GROSSI P., LOCATELLI F., COLOMBOP., VIGANÒ M., ET AL. (2001). Diagnosis and moni-toring of human cytomegalovirus infection in trans-plant recipients. Rev. Med. Microbiol. 12, 155-175.
GERNA G., BALDANTI F., LILLERI D., PAREAM., TORSELLINIM., CASTIGLIONI B., ET AL. (2003a). Human cy-tomegalovirus pp67 mRNAemia Vs. pp65 antigen-emia in heart and lung transplant recipients: aprospective randomized controlled open-label trial.Transplantation. 75, 1012-1019.
GERNA G., LILLERI D., BALDANTI F., TORSELLINI M.,GIORGIANI G., ZECCA M., ET AL. (2003b). Human cy-tomegalovirus immediate-early mRNAemia versuspp65 antigenemia for guiding pre-emptive therapyin children and young adults undergoing hemato-poietic stem cell transplantation: a prospective,randomized, open-label trial. Blood. 101, 5053-5060.
GERNA G., SARASINI A., LILLERI D., PERCIVALLE E.,TORSELLINI M., BALDANTI F., ET AL. (2003c). In vitromodel for the study of the dissociation of increas-ing antigenemia and decreasing DNAemia andviremia during treatment of human cy-tomegalovirus infection with ganciclovir in trans-plant recipients. J. Infect. Dis. 188, 1639-1647.
GERNA G., LILLERI D., ZECCA M., ALESSANDRINO E.P.,BALDANTI F., REVELLO M.G., ET AL. (2005a). Risingantigenemia levels may be misleading in pre-emp-tive therapy of human cytomegalovirus infectionin allogeneic hematopoietic stem cell transplant re-cipients. Haematologica. 90, 526-533.
GERNA G., PERCIVALLE E., LILLERI D., LOZZA L., FORNARAC., HAHN G., ET AL. (2005b). Dendritic-cell infectionby human cytomegalovirus is restricted to strainscarrying functional UL131-128 genes and mediatesefficient viral antigen presentation to CD8+ T cells.J. Gen. Virol. 86, 275-284.
GERNA G., LILLERI D. (2006). Monitoring transplant pa-tients for human cytomegalovirus: diagnostic up-date. Herpes. 13, 4-11.
GERNA G., LILLERI D., FORNARA C., COMOLLI G., LOZZA L.,CAMPANA C., ET AL. (2006). Monitoring of human cy-tomegalovirus-specific CD4+ and CD8+ T-cell im-munity in patients receiving solid organ transplan-tation. Am. J. Transplant. 6, 2356-2364.
GERNA G., BALDANTI F., TORSELLINI M., MINOLI L.,VIGANÒM., OGGIONNI T., ET AL. (2007). Evaluation ofhuman cytomegalovirus DNAemia vs pp65-anti-genemia cutoff for guiding preemptive therapy intransplant recipients: A randomized study. Antivir.Ther. 12, 63-72.
GERNA G., LILLERI D., CALLEGARO A., GOGLIO A., CORTESES., STROPPA P., ET AL. (2008a). Prophylaxis followedby preemptive therapy vs preemptive therapy forprevention of human cytomegalovirus disease in
248 G. Gerna, D. Lilleri, M. Furione, F. Baldanti

pediatric patients undergoing liver transplantation.Transplantation. 86, 163-166.
GERNA G., LILLERI D., CALDERA D., FURIONEM., ZENONEBRAGOTTI L., ALESSANDRINO E.P. (2008b). Validationof a DNAemia cut-off for pre-emptive therapy ofcytomegalovirus infection in adult hematopoieticstem cell transplant recipients. Bone MarrowTransplant. 41, 873-879.
GERNA G., LILLERI D., ROGNONI V., AGOZZINOM., MELONIF., OGGIONNI T., ET AL. (2009). Preemptive therapy ofsystemic and pulmonary human cytomegalovirusinfection in lung transplant recipients. Am. J.Transplant. 9, 1142-1150.
GILLESPIE G.M., WILLS M.R., APPAY V., O’CALLAGHAN C.,MURPHYM., SMITH N., ET AL. (2000). Functional het-erogeneity and high frequencies of cy-tomegalovirus-specific CD8(+) T lymphocytes inhealthy seropositive donors. J. Virol. 74, 8140-8150.
GLEAVES C.A., SMITH T.F., SHUSTER E.A., PEARSON G.R.(1984). Rapid detection of cytomegalovirus inMRC-5 cells inoculated with urine specimens byusing low-speed centrifugation and monoclonal an-tibody to an early antigen. J. Clin. Microbiol. 19,917-919.
GRATAMA J.W., VAN ESSER J.W., LAMERS C.H., TOURNAYC., LÖWENBERG B., BOLHUIS R.L., ET AL. (2001).Tetramer-based quantification of cytomegalovirus(CMV)-specific CD8+ T lymphocytes in T-cell-de-pleted stem cell grafts and after transplantationmay identify patients at risk for progressive CMVinfection. Blood. 98, 1358-1364.
GRATAMA J.W., BOECKH M., NAKAMURA R., CORNELISSENJ.J., BROOIMANS R.A., ZAIA J.A., ET AL. (2010).Immune monitoring with iTAg MHC Tetramers forprediction of recurrent or persistent cy-tomegalovirus infection or disease in allogeneichematopoietic stem cell transplant recipients: aprospective multicenter study. Blood. 116, 1655-1662.
GROSSI P., MINOLI L., PERCIVALLE E., IRISH W., VIGANÒM., GERNA G. (1995). Clinical and virological mon-itoring of human cytomegalovirus infection in 294heart transplant recipients. Transplantation. 59,847-851.
HADAYA K., DE RHAM C., BANDELIER C., BANDELIER C.,FERRARI-LACRAZ S., JENDLY S., ET AL. (2008). Naturalkiller cell receptor repertoire and their ligands, andthe risk of CMV infection after kidney transplanta-tion. Am. J. Transplant. 8, 2674-2683.
HAHN G., REVELLO M.G., PATRONE M., PERCIVALLE E.,CAMPANINI G., SARASINI A., ET AL. (2004). Human cy-tomegalovirus UL131-128 genes are indispensablefor virus growth in endothelial cells and virus trans-fer to leukocytes. J. Virol. 78, 10023-10033.
HAKKI M., RIDDELL S.R., STOREK J., CARTER R.A.,STEVENS-AYERS T., SUDOUR P., ET AL. (2003). Immunereconstitution to cytomegalovirus after allogeneichematopoietic stem cell transplantation: impact of
host factors, drug therapy, and subclinical reacti-vation. Blood. 102, 3060-3067.
HAMPRECHT K., STEINMASSL M., EINSELE H., JAHN G.(1998). Discordant detection of human cy-tomegalovirus DNA from peripheral bloodmononuclear cells, granulocytes and plasma: cor-relation to viremia and HCMV infection. J. Clin.Virol. 11, 125-136.
HEBART H., GAMER D., LOEFFLER J., MUELLER C.,SINZGER C., JAHN G., ET AL. (1998). Evaluation ofMurex CMV DNA hybrid capture assay for detec-tion and quantitation of cytomegalovirus infectionin patients following allogeneic stem cell trans-plantation. J. Clin. Microbiol. 36, 1333-1337.
HEBART H., BRUGGER W., GRIGOLEIT U., GSCHEIDLE B.,LOEFFLER J., SCHÄFER H., ET AL. (2001). Risk for cy-tomegalovirus disease in patients receiving poly-merase chain reaction-based preemptive antiviraltherapy after allogeneic stem cell transplantationdepends on transplantation modality. Blood. 97,2183-2185.
HEBART H., DAGINIK S., STEVANOVIC S., GRIGOLEIT U.,DOBLER A., BAUR M., ET AL. (2002). Sensitive dete-ction of human cytomegalovirus peptide-specificcytotoxic T-lymphocyte responses by interferon-gamma-enzyme-linked immunospot assay and flowcytometry in healthy individuals and in patients af-ter allogeneic stem cell transplantation. Blood. 99,3830-3837.
HUMAR A., GREGSON D., CALIENDO A.M., MCGEER A.,MALKAN G., KRAJDEN M., ET AL. (1999). Clinical uti-lity of quantitative cytomegalovirus viral load de-termination for predicting cytomegalovirus diseasein liver transplant recipients. Transplantation. 68,1305-1311.
HUMAR A., SNYDMAN D., and the AST. (2009).Cytomegalovirus in solid organ transplant recipi-ents. Am. J. Transplant. 9 (Suppl. 4), S78-86.
JONJIC’ S., PAVIC’ I., LUCIN P., RUKAVINA D., KOSZINOWSKIU.H. (1990). Efficacious control of cytomegalovirusinfection after long-term depletion of CD8+ T lym-phocytes. J. Virol. 64, 5457-5464.
JONJIC’ S., PAVIC’ I., POLIC’ B., CRNKOVIC’ I., LUCIN P.,KOSZINOWSKI U.H. (1994). Antibodies are not es-sential for the resolution of primary cy-tomegalovirus infection but limit dissemination ofrecurrent virus. J. Exp. Med. 179, 1713-1717.
KARRER U., SIERRO S., WAGNERM., OXENIUS A., HENGELH., KOSZINOWSKI U.H., ET AL. (2003). Memory in-flation: continuous accumulation of antiviral CD8+
T cells over time. J. Immunol. 170, 2022-2029. KELLY D.A. (2006). Current issues in pediatric trans-plantation. Pediatr. Transplant. 10, 712-720.
KHAN N., SHARIFF N., COBBOLD M., BRUTON R.,AINSWORTH J.A., SINCLAIR A.J., ET AL. (2002).Cytomegalovirus seropositivity drives the CD8 Tcell repertoire toward greater clonality in healthyelderly individuals. J. Immunol. 169, 1984-1992.
HCMV management in transplant recipients 249

KHAN N., HISLOP A., GUDGEON N., COBBOLDM., KHANNAR., NAYAK L., ET AL. (2004). Herpesvirus-specific CD8T cell immunity in old age: cytomegalovirus im-pairs the response to a coresident EBV infection.J. Immunol. 173, 7481-7489.
KHOURY J.A., STORCH G.A., BOHL D.L., SCHUESSLER R.M.,TORRENCE S.M., LOCKWOOD M., ET AL. (2006).Prophylactic versus preemptive oral valganciclovirfor the management of cytomegalovirus infection inadult renal transplant recipients. Am. J. Transplant.6, 2134-2142.
KLENOVSEKK., WEISEL F., SCHNEIDER A., APPELT U., JONJICS., MESSERLEM., ET AL. (2007). Protection from CMVinfection in immunodeficient hosts by adoptivetransfer of memory B cells. Blood. 110, 3472-3479.
KLIEM V., FRICKE L., WOLLBRINK T., BURG M.,RADERMACHER J., ROHDE F. (2008). Improvement inlong-term renal graft survival due to CMV prophy-laxis with oral ganciclovir: results of a randomizedclinical trial. Am. J. Transplant. 8, 975-983.
KNIGHT A., MADRIGAL A.J., GRACE S., SIVAKUMARAN J.,KOTTARIDIS P., MACKINNON S., ET AL. (2010). The roleof Vδ2-negative γδ T cells during cytomegalovirusreactivation in recipients of allogeneic stem celltransplantation. Blood. 116, 2164-2172.
KOLBERG J., MINER R., HOO B., ET AL. (1996).Development of a branched DNA signal amplifica-tion assay to monitor patients on anti-CMV thera-pies. Biologicals. 24, 216.
KOTTON C.N., KUMAR D., CALIENDO A.M., ÅSBERG A.,CHOU S., SNYDMAN D.R., ALLEN U., HUMAR A.; on be-half of the Transplantation Society InternationalCMV Consensus Group. (2010). InternationalConsensus Guideliness on the Management of cy-tomegalovirus in solid organ transplantation.Transplantation. 89, 779-795.
KRAUSE H., HEBART H., JAHN G., MÜLLER C.A., EINSELEH. (1997). Screening for CMV-specific T cell pro-liferation to identify patients at risk of developinglate onset CMV disease. Bone Marrow Transplant.19, 1111-1116.
KUMAR D., CHERNENKO S., MOUSSA G., COBOS I., MANUELO., PREIKSAITIS J., ET AL. (2009). Cell-mediated im-munity to predict cytomegalovirus disease in high-risk solid organ transplant recipients. Am. J.Transplant. 9, 1214-1222.
KUSNE S., GROSSI P., IRISH W., ST GEORGE K., RINALDOC., RAKELA J., ET AL. (1999). Cytomegalovirus pp65antigenemia monitoring as a guide for preemptivetherapy: a cost effective strategy for prevention ofcytomegalovirus disease in adult liver transplantrecipients. Transplantation. 68, 1125-1131.
LACEY S.F., LA ROSA C., ZHOU W., SHARMA M.C.,MARTINEZ J., KRISHNAN A., ET AL. (2006). Functionalcomparison of T cells recognizing cytomegaloviruspp65 and immediate-early antigen polypeptides inhematopoietic stem-cell transplant and solid organtransplant recipients. J. Infect. Dis. 194, 1410-1421.
LAFARGE X., MERVILLE P., CAZIN M.C., BERGÉ F., POTAUXL., MOREAU J.F., ET AL. (2001). Cytomegalovirus in-fection in transplant recipients resolves when cir-culating gammadelta T lymphocytes expand, sug-gesting a protective antiviral role. J. Infect. Dis. 184,533-541.
LAUTENSCHLAGER I., HALME L., HÖCKERSTEDT K.,KROGERUS L., TASKINEN E. (2006). Cytomegalovirusinfection of the liver transplant: virological, histo-logical, immunological, and clinical observations.Transpl. Infect. Dis. 8, 21-30.
LI C.R., GREENBERG P.D., GILBERT M.J., GOODRICH J.M.,RIDDELL S.R. (1994). Recovery of HLA-restrictedcytomegalovirus (CMV)-specific T-cell responses af-ter allogeneic bone marrow transplant: correlationwith CMV disease and effect of ganciclovir pro-phylaxis. Blood. 83, 1971-1979.
LI H., DUMMER J.S., ESTES W.R., MENG S., WRIGHT P.F.,TANG Y.W. (2003). Measurement of human cy-tomegalovirus loads by quantitative real-time PCRfor monitoring clinical intervention in transplantrecipients. J. Clin. Microbiol. 41, 187-191.
LILLERI D., BALDANTI F., GATTI M., ROVIDA F., DOSSENAL., DE GRAZIA S., ET AL. (2004). Clinically-based de-termination of safe DNAemia cutoff levels for pre-emptive therapy of human cytomegalovirus in-fections in solid organ and hematopoietic stemcell transplant recipients. J. Med. Virol. 73, 412-418.
LILLERI D., GERNA G., FORNARA C., LOZZA L., MACCARIOR., LOCATELLI F. (2006). Prospective simultaneousquantification of human cytomegalovirus-specificCD4+ and CD8+ T-cell reconstitution in young re-cipients of allogeneic hematopoietic stem cell trans-plants. Blood. 108, 1406-1412.
LILLERI D., GERNA G., FURIONE M., BERNARDO M.E.,GIORGIANI G., TELLI S., ET AL. (2007a). Use of aDNAemia cut-off for monitoring human cy-tomegalovirus infection reduces the number of pre-emptively treated children and young adults re-ceiving hematopoietic stem cell transplantationcompared with qualitative pp65 antigenemia.Blood. 110, 2757-2760.
LILLERI D., ZELINI P., FORNARA C., COMOLLI G., GERNAG. (2007b). Inconsistent responses of cy-tomegalovirus-specific T cells to pp65 and IE-1 ver-sus infected dendritic cells in organ transplant re-cipients. Am. J. Transplant. 7, 1997-2005.
LILLERI D., FORNARA C., CHIESA A., CALDERA D.,ALESSANDRINO E.P., GERNA G. (2008a). Human cy-tomegalovirus-specific CD4+ and CD8+ T-cell re-constitution in adult allogeneic hematopoietic stemcell transplant recipients and immune control ofviral infection. Haematologica. 93, 248-256.
LILLERI D., FORNARA C., REVELLO M.G., GERNA G.(2008b). Human cytomegalovirus-specific memo-ry CD8+ and CD4+ T cell differentiation after pri-mary infection. J. Infect. Dis. 198, 536-543.
250 G. Gerna, D. Lilleri, M. Furione, F. Baldanti

LILLERI D., GERNA G., FORNARA C., CHIESA A., COMOLLIG., ZECCA M., ET AL. (2009a). Human cy-tomegalovirus-specific T-cell reconstitution inyoung patients receiving T-cell-depleted, allogene-ic hematopoietic stem cell transplantation. J. Infect.Dis. 199, 829-836.
LILLERI D., LAZZAROTTO T., GHISETTI V., RAVANINI P.,CAPOBIANCHI M.R., BALDANTI F., ET AL. (2009b).Multicenter quality control study for human cy-tomegalovirus DNAemia quantification. NewMicrobiol. 32, 245-253.
LILLERI D., ZELINI P., FORNARA C., COMOLLI G., REVELLOM.G., GERNA G. (2009c). Human cytomegalovirus-specific CD4+ and CD8+ T-cell responses in primaryinfection of the immunocompetent and immuno-compromised host. Clin. Immunol. 131, 395-403.
LIMAYE A.P., BOWDEN R.A., MYERSON D., BOECKH M.(1997). Cytomegalovirus disease occurring beforeengraftment in marrow transplant recipients. Clin.Infect. Dis. 24, 830-835.
LIMAYE A.P., BAKTHAVATSALAM R., KIM H.W., KUHR C.S.,HALLDORSON J.B., HEALEY P.J., ET AL. (2004). Late-onset cytomegalovirus disease in liver transplantrecipients despite antiviral prophylaxis.Transplantation. 78, 1390-1396.
LIMAYE A.P., BAKTHAVATSALAM R., KIM H.W., RANDOLPHS.E., HALLDORSON J.B., HEALEY P.J., ET AL. (2006).Impact of cytomegalovirus in organ transplant re-cipients in the era of antiviral prophylaxis.Transplantation. 81, 1645-1652.
LJUNGMAN P., ENGELHARD D., LINK H., BIRON P., BRANDTL., BRUNET S., ET AL. (1992). Treatment of intersti-tial pneumonitis due to cytomegalovirus with gan-ciclovir and intravenous immune globulin: experi-ence of European Bone Marrow Transplant Group.Clin. Infect. Dis. 14, 831-835.
LJUNGMAN P., ASCHAN J., LEWENSOHN-FUCHS I., CARLENSS., LARSSON K., LÖNNQVIST B., ET AL. (1998a). Resultsof different strategies for reducing cytomegalovirus-associated mortality in allogeneic stem cell trans-plant recipients. Transplantation. 66, 1330-1334.
LJUNGMAN P., CORDONNIER C., EINSELE H., BENDER-GÖTZE C., BOSI A., DEKKER A., ET AL. (1998b). Use ofintravenous immune globulin in addition to an-tiviral therapy in the treatment of CMV gastroin-testinal disease in allogeneic bone marrow trans-plant patients: a report from the European Groupfor Blood and Marrow Transplantation (EBMT).Infectious Diseases Working Party of the EBMT.Bone Marrow Transplant. 21, 473-476.
LJUNGMAN P., GRIFFITHS P., PAYA C. (2002). Definitions ofcytomegalovirus infection and disease in transplantrecipients. Clin. Infect. Dis. 34, 1094-1097.
LOCATELLI F., PERCIVALLE E., COMOLI P., MACCARIO R.,ZECCA M., GIORGIANI G., ET AL. (1994). Human cyto-megalovirus (HCMV) infection in paediatric pa-tients given allogeneic bone marrow transplanta-tion: role of early antiviral treatment for HCMV an-
tigenemia on patients’ outcome. Brit. J. Haematol.88, 64-71.
LOZZA L., LILLERI D., PERCIVALLE E., FORNARA C.,COMOLLI G., REVELLO M.G., ET AL. (2005).Simultaneous quantification of human cy-tomegalovirus (HCMV)-specific CD4+ and CD8+ Tcells by a novel method using monocyte-derivedHCMV-infected immature dendritic cells. Eur. J.Immunol. 35, 1795-1804.
MACAGNO A., BERNASCONI N., VANZETTA F., DANDER E.,SARASINI A., REVELLOM.G., ET AL. (2010). Isolationof human monoclonal antibodies that potently neu-tralize HCMV infection by targeting different epi-topes on the gH/gL/UL128-131A complex. J. Virol.84, 1005-1013.
MACHADO C.M., DULLEY F.L., BOAS L.S., CASTELLI J.B.,MACEDO M.C., SILVA R.L., ET AL. (2000). CMV pneu-monia in allogeneic BMT recipients undergoingearly treatment of pre-emptive ganciclovir therapy.Bone Marrow Transplant. 26, 413-417.
MATTES F.M, VARGAS A., KOPYCINSKI J., HAINSWORTHE.G., SWENY P., NEBBIA G., ET AL. (2008). Functionalimpairment of cytomegalovirus specific CD8 T cellspredicts high-level replication after renal trans-plantation. Am. J. Transplant. 8, 990-999.
MENGELLE C., SANDRES-SAUNÉ K., PASQUIER C., ROSTAINGL., MANSUY J.M., MARTY M., ET AL. (2003).Automated extraction and quantification of humancytomegalovirus DNA in whole blood by real-timePCR assay. J. Clin. Microbiol. 41, 3840-3845.
NINOMIYA T., TAKIMOTO H., MATSUZAKI G., HAMANO S.,YOSHIDA H., YOSHIKAI Y., ET AL. (2000). Vgamma1+
gammadelta T cells play protective roles at an ear-ly phase of murine cytomegalovirus infectionthrough production of interferon-gamma.Immunology. 99, 187-194.
OUYANG Q., WAGNERW.M., WIKBY A., WALTER S., AUBERTG., DODI A.I., ET AL. (2003). Large numbers of dys-functional CD8+ T lymphocytes bearing receptorsfor a single dominant CMV epitope in the very old.J. Clin. Immunol. 23, 247-257.
PANG X.L., FOX J.D., FENTON J.M., MILLER G.G.,CALIENDO A.M., PREIKSAITIS J.K. (2009).Interlaboratory comparison of cytomegalovirus vi-ral load assays. Am. J. Transplant. 9, 258-268.
PARENTE F., BIANCHI PORRO G., and The ItalianCytomegalovirus Study Group. (1998). Treatment ofcytomegalovirus esophagitis in patients with ac-quired immune deficiency syndrome: a random-ized controlled study of foscarnet versus ganci-clovir. Am. J. Gastroenterol. 93, 317-322.
PASTORE D., DELIA M., MESTICE A., PERRONE T.,CARLUCCIO P., GAUDIO F., ET AL. (2011). Recovery ofCMV-specific CD8+ T cells and Tregs after allo-geneic peripheral blood stem cell transplantation.Biol. Blood Marrow Transplant. 17, 550-557.
PAWELEC G., AKBAR A., CARUSO C., SOLANA R, GRUBECK-LOEBENSTEIN B, WIKBY A. (2005). Human im-
HCMV management in transplant recipients 251

munosenescence: is it infectious? Immunol. Rev.205, 257-268.
PIETERSMA F.L., VAN DORP S., JACOBI R., RAN L., NANLOHYN.M., SCHUURMAN R., ET AL. (2010). High level ofperforin expression in T cells: an early prognosticmarker of the severity of herpesvirus reactivationafter allogeneic stem cell transplantation in adults.Clin. Infect. Dis. 50, 717-725.
PIIPARINEN H., HÖCKERSTEDT K., GRÖNHAGEN-RISKA C.,LAUTENSCHLAGER I. (2004). Comparison of twoquantitative CMV PCR tests, Cobas Amplicor CMVMonitor and TaqMan assay, and pp65-antigenemiaassay in the determination of viral loads from pe-ripheral blood of organ transplant patients. J. Clin.Virol. 30, 258-266.
POLIC’ B., HENGEL H., KRMPOTIC’ A., TRGOVCICH J., PAVIC’I., LUCCARONIN P., ET AL. (1998). Hierarchical andredundant lymphocyte subset control precludes cy-tomegalovirus replication during latent infection.J. Exp. Med. 188, 1047-1054.
POURGHEYSARI B., KHAN N., BEST D., BRUTON R., NAYAKL., MOSS P.A. (2007). The cytomegalovirus-specificCD4+ T-cell response expands with age and marked-ly alters the CD4+ T-cell repertoire. J. Virol. 81, 7759-7765.
PRICE D.A., BRENCHLEY J.M., RUFF L.E., BETTS M.R.,HILL B.J., ROEDERER M., ET AL. (2005). Avidity forantigen shapes clonal dominance in CD8+ T cellpopulations specific for persistent DNA viruses. J.Exp. Med. 202, 1349-1361.
QUINNAN JR G.V., KIRMANI N., ROOK A.H., MANISCHEWITZJ.F., JACKSON L., MORESCHI G., ET AL. (1982).Cytotoxic T cells in cytomegalovirus infection: HLA-restricted T-lymphocyte and non-T-lymphocyte cy-totoxic responses correlate with recovery from cy-tomegalovirus infection in bone-marrow-transplantrecipients. N. Engl. J. Med. 307, 7-13.
RADHA R., JORDAN S., PULIYANDA D., BUNNAPRADIST S.,PETROSYAN A., AMET N., ET AL. (2005). Cellular im-mune responses to cytomegalovirus in renal trans-plant recipients. Am. J. Transplant. 5, 110-117.
RASMUSSEN L., MATKIN C., SPAETE R., PACHL C., MERIGANT.C. (1991). Antibody response to human cy-tomegalovirus glycoproteins gB and gH after natu-ral infection in humans. J. Infect. Dis. 164, 835-842.
RAZONABLE R.R., BROWN R.A., WILSON J., GROETTUM C.,KREMERS W., ESPY M., ET AL. (2002). The clinicaluse of various blood compartments for cy-tomegalovirus (CMV) DNA quantitation in trans-plant recipients with CMV disease. Transplantation.73, 968-973.
RAZONABLE R.R., VAN CRUIJSEN H., BROWN R.A., WILSONJ.A., HARMSEN W.S., WIESNER R.H., ET AL. (2003).Dynamics of cytomegalovirus replication duringpreemptive therapy with oral ganciclovir. J. Infect.Dis. 187, 1801-1808.
REISCHIG T., JINDRA P., HES O., SVECOVÁ M., KLABOCH J.,TRESKA V. (2008). Valacyclovir prophylaxis versus
preemptive valganciclovir therapy to prevent cyto-megalovirus disease after renal transplantation.Am. J. Transplant. 8, 69-77.
REUSSER P., RIDDELL S.R., MEYERS J.D., GREENBERG P.D.(1991). Cytotoxic T-lymphocyte response to cyto-megalovirus after human allogeneic bone marrowtransplantation: pattern of recovery and correla-tion with cytomegalovirus infection and disease.Blood. 78, 1373-1380.
REUSSER P., CATHOMAS G., ATTENHOFER R., TAMM M.,THIEL G. (1999). Cytomegalovirus (CMV)-specificT cell immunity after renal transplantation medi-ates protection from CMV disease by limiting thesystemic virus load. J. Infect. Dis. 180, 247-253.
REUSSER P., EINSELE H., LEE J., VOLIN L., ROVIRA M.,ENGELHARD D., ET AL. (2002). Randomized multi-center trial of foscarnet versus ganciclovir for pre-emptive therapy of cytomegalovirus infection afterallogeneic stem cell transplantation. Blood. 99,1159-1164.
REVELLO M.G., PERCIVALLE E., ZAVATTONI M., PAREA M.,GROSSI P., GERNA G. (1989). Detection of human cy-tomegalovirus immediate early antigen in leuko-cytes as a marker of viremia in immunocompro-mised patients. J. Med. Virol. 29, 88-93.
REVELLO M.G., PERCIVALLE E., ARBUSTINI E., PARDI R.,SOZZANI S., GERNA G. (1998). In vitro generation ofhuman cytomegalovirus pp65 antigenemia, vire-mia, and leukoDNAemia. J. Clin. Invest. 101, 2686-2692.
RIDDELL S.R., WATANABE K.S., GOODRICH J.M., LI C.R.,AGHA M.E., GREENBERG P.D. (1992). Restoration ofviral immunity in immunodeficient humans by theadoptive transfer of T cell clones. Science. 257, 238-241.
RYCKMAN B.J., RAINISH B.L., CHASE M.C., BORTON J.A.,NELSON J.A., JARVIS M.A., ET AL. (2008).Characterization of the human cytomegalovirusgH/gL/UL128-131 complex that mediates entry in-to epithelial and endothelial cells. J. Virol. 82, 60-70.
SALLUSTO F., LANZAVECCHIA A. (1994). Efficient presen-tation of soluble antigen by cultured human den-dritic cells is maintained by granulocyte/macrophage colony-stimulating factor plus inter-leukin 4 and downregulated by tumor necrosis fac-tor alpha. J. Exp. Med. 179, 1109-1118.
SALLUSTO F., LENIG D., FÖRSTER R., LIPP M.,LANZAVECCHIA A. (1999). Two subsets of memory Tlymphocytes with distinct homing potentials andeffector functions. Nature. 401, 708-712.
SANCHEZ J.L., KRUGER R.M., PARANJOTHI S., TRULOCKE.P., LYNCH J.P., HICKS C., ET AL. (2001). Relationshipof cytomegalovirus viral load in blood to pneu-monitis in lung transplant recipients. Transplan -tation. 72, 733-735.
SCALZO A.A., FITZGERALD N.A., WALLACE C.R., GIBBONSA.E., SMART Y.C., BURTON R.C., ET AL. (1992). Theeffect of the Cmv-1 resistance gene, which is linked
252 G. Gerna, D. Lilleri, M. Furione, F. Baldanti

to the natural killer cell gene complex, is mediatedby natural killer cells. J. Immunol. 149, 581-589.
SCHLEISS M.R., BOURNE N., STROUP G., BRAVO F.J.,JENSEN N.J., BERNSTEIN D.I. (2004). Protectionagainst congenital cytomegalovirus infection anddisease in guinea pigs, conferred by a purified re-combinant glycoprotein B vaccine. J. Infect. Dis.189, 1374-1381.
SESTER M., SESTER U., GÄRTNER B., HEINE G., GIRNDTM., MUELLER-LANTZSCH N., ET AL. (2001). Levels ofvirus-specific CD4 T cells correlate with cy-tomegalovirus control and predict virus-induceddisease after renal transplantation. Transplantation.71, 1287-1294.
SESTER M., SESTER U., GÄRTNER B.C., GIRNDT M.,MEYERHANS A., KÖHLER H. (2002). Dominance ofvirus-specific CD8 T cells in human primary cy-tomegalovirus infection. J. Am. Soc. Nephrol. 13,2577-2584.
SESTER U., GÄRTNER B.C., WILKENS H., SCHWAAB B.,WÖSSNER R., KINDERMANN I., ET AL. (2005).Differences in CMV-specific T-cell levels and long-term susceptibility to CMV infection after kidney,heart and lung transplantation. Am. J. Transplant.5, 1483-1489.
SEU P., WINSTON D.J., HOLT C.D., KALDAS F., BUSUTTILR.W. (1997). Long-term ganciclovir prophylaxis forsuccessful prevention of primary cytomegalovirus(CMV) disease in CMV-seronegative liver transplantrecipients with CMV-seropositive donors.Transplantation. 64, 1614-1617.
SHLOBIN O.A., WEST E.E., LECHTZIN N., MILLER S.M.,BORJA M., ORENS J.B., ET AL. (2006). Persistent cy-tomegalovirus-specific memory responses in thelung allograft and blood following primary infec-tion in lung transplant recipients. J. Immunol. 176,2625-2634.
SINGH N., WANNSTEDT C., KEYES L., WAGENER M.M.,GAYOWSKI T., CACCIARELLI T.V. (2005). Indirect out-comes associated with cytomegalovirus (oppor-tunistic infections, hepatitis C virus sequelae, andmortality) in liver-transplant recipients with the useof preemptive therapy for 13 years. Transplantation.79, 1428-1434.
SINGH N. (2006a). Cytomegalovirus infection in solidorgan transplant recipients: new challenges andtheir implications for preventive strategies. J. Clin.Virol. 35, 474-477.
SINGH N. (2006b). Antiviral drugs for cytomegalovirusin transplant recipients: advantages of preemptivetherapy. Rev. Med. Virol. 16, 281-287.
SMALL L.N., LAU J., SNYDMAN D.R. (2006). Preventingpost-organ transplantation cytomegalovirus diseasewith ganciclovir: a meta-analysis comparing pro-phylactic and preemptive therapies. Clin. Infect.Dis. 43, 869-880.
SNYDER L.D., MEDINAS R., CHAN C., SPARKS S., DAVISW.A., PALMER S.M., ET AL. (2011). Polyfunctional cy-
tomegalovirus-specific immunity in lung transplantrecipients receiving valganciclovir prophylaxis. Am.J. Transplant. 11, 553-560.
SNYDMAN D.R. (2009). Cytomegalovirus preventionstrategies: the case for prophylaxis. Am. J.Transplant. 9, 1254.
SPIVEY J.F., SINGLETON D., SWEET S., STORCH G.A.,HAYASHI R.J., HUDDLESTON C.B., ET AL. (2007). Safetyand efficacy of prolonged cytomegalovirus pro-phylaxis with intravenous ganciclovir in pediatricand young adult lung transplant recipients. Pediatr.Transplant. 11, 312-318.
STAGNO S., PASS R.F., DWORSKY M.E., HENDERSON R.E.,MOORE E.G., WALTON P.D., ET AL. (1982). Congenitalcytomegalovirus infection: the relative importanceof primary and recurrent maternal infection. N.Engl. J. Med. 306, 945-949.
STERNM., ELSÄSSER H., HÖNGER G., STEIGER J., SCHAUBS., HESS C. (2008). The number of activating KIRgenes inversely correlates with the rate of CMV in-fection/reactivation in kidney transplant recipients.Am. J. Transplant. 8, 1312-1317.
SUN H.Y., WAGENERM.M., SINGH N. (2008). Preventionof posttransplant cytomegalovirus disease and re-lated outcome with valganciclovir: a systematic re-view. Am. J. Transplant. 8, 2111-2118.
SYLWESTER A.W., MITCHELL B.L., EDGAR J.B., TAORMINAC., PELTE C., RUCHTI F., ET AL. (2005). Broadly tar-geted human cytomegalovirus-specific CD4+ andCD8+ T cells dominate the memory compartmentsof exposed subjects. J. Exp. Med. 202, 673-685.
TABETA K., GEORGEL P., JANSSEN E., DU X., HOEBE K.,CROZAT K., ET AL. (2004). Toll-like receptors 9 and 3as essential components of innate immune defenseagainst mouse cytomegalovirus infection. Proc.Natl. Acad. Sci. USA. 101, 3516-3521.
TORMO N., SOLANO C., BENET I., NIETO J., DE LA CÁMARAR., GARCIA-NOBLEJAS A., ET AL. (2010). Kinetics ofcytomegalovirus (CMV) pp65 and IE-1-specificIFNγ CD8+ and CD4+ T cells during episodes of vi-ral DNAemia in allogeneic stem cell transplant re-cipients: potential implications for the managementof active CMV infection. J. Med. Virol. 82, 1208-1215.
TRZONKOWSKI P., MYSLIWSKA J., SZMIT E., WIECKIEWICZJ., LUKASZUK K., BRYDAK L.B., ET AL. (2003).Association between cytomegalovirus infection, en-hanced proinflammatory response and low level ofanti-hemagglutinins during the anti-influenza vac-cination - an impact of immunosenescence.Vaccine. 21, 3826-3836.
VAN DER BIJ W., TORENSMA R., VAN SON W.J., ANEMA J.,SCHIRM J., TEGZESS A.M., ET AL. (1988). Rapid im-munodiagnosis of active cytomegalovirus infectionby monoclonal antibody staining of blood leuco-cytes. J. Med. Virol. 25, 179-188.
VENEMAH., VAN DEN BERG A.P., VAN ZANTEN C., VAN SONW.J., VAN DER GIESSEN M., THE T.H. (1994). Natural
HCMV management in transplant recipients 253

killer cell responses in renal transplant patients withcytomegalovirus infection. J. Med. Virol. 42, 188-192.
VERKRUYSE L.A., STORCH G.A., DEVINE S.M., DIPERSIOJ.F., VIJ R. (2006). Once daily ganciclovir as initialpre-emptive therapy delayed until threshold CMVload > or =10000 copies/ml: a safe and effectivestrategy for allogeneic stem cell transplant patients.Bone Marrow Transplant. 37, 51-56.
WALKER S., FAZOU C., CROUGH T., HOLDSWORTH R., KIELYP., VEALE M., ET AL. (2007). Ex vivo monitoring ofhuman cytomegalovirus-specific CD8+ T-cell re-sponses using QuantiFERON-CMV. Transpl. Infect.Dis. 9, 165-170.
WALTER E.A., GREENBERG P.D., GILBERTM.J., FINCH R.J.,WATANABE K.S., THOMAS E.D., ET AL. (1995).Reconstitution of cellular immunity against cy-tomegalovirus in recipients of allogeneic bone mar-row by transfer of T-cell clones from the donor. N.Engl. J. Med. 333, 1038-1044.
WANG D., SHENK T. (2005). Human cytomegalovirusvirion protein complex required for epithelial andendothelial cell tropism. Proc. Natl. Acad. Sci. USA.102, 18153-18158.
WESTALL G.P., MICHAELIDES A., WILLIAMS T.J., SNELL G.I.,KOTSIMBOS T.C. (2004). Human cytomegalovirusload in plasma and bronchoalveolar lavage fluid: alongitudinal study of lung transplant recipients. J.Infect. Dis. 190, 1076-1083.
WESTALL G.P., MIFSUD N.A., KOTSIMBOS T. (2008).
Linking CMV serostatus to episodes of CMV re-activation following lung transplantation by meas-uring CMV-specific CD8+ T-cell immunity. Am. J.Transplant. 8, 1749-1754.
WILLS M.R., CARMICHAEL A.J., WEEKES M.P., MYNARDK., OKECHA G., HICKS R., ET AL. (1999). Humanvirus-specific CD8+ CTL clones revert fromCD45ROhigh to CD45RAhigh in vivo:CD45RAhighCD8+ T cells comprise both naive andmemory cells. J. Immunol. 162, 7080-7087.
YEAGER A.S., GRUMET F.C., HAFLEIGH E.B., ARVIN A.M.,BRADLEY J.S., PROBER C.G. (1981). Prevention oftransfusion-acquired cytomegalovirus infectionsin newborn infants. J. Pediatr. 98, 281-287.
ZAMBELLI A., LILLERI D., PEDRAZZOLI P., PECCATORI J.,BALDANTI F., FREGONI V., ET AL. (2005). Incidenceof human cytomegalovirus infection in patientswith refractory solid tumors receiving nonmye-loablative allogeneic stem cell transplants versusrecipients of standard SCT for hematologic ma-lignancies. Biol. Blood Marrow Transplant. 11,423-428.
ZEDTWITZ-LIEBENSTEIN K., JAKSCH P., BAUER C., POPOWT., KLEPETKO W., HOFMANN H., PUCHHAMMER-STOCKL E. (2004). Association of cytomegalovirusDNA concentration in epithelial lining fluid andsymptomatic cytomegalovirus infection in lungtransplant recipients. Transplantation. 77, 1897-1899.
254 G. Gerna, D. Lilleri, M. Furione, F. Baldanti