uromodulin (tamm-horsfall … (tamm-horsfall glycoprotein/uromucoid) is a...
TRANSCRIPT

THE JOURNM. OF B~OLOGKAL. CHEMISTRY Q 1990 by The American Society for Biochemistry and Molecular Biology, Inc.
Vol. 265, No. 34, Issue of December 5, pp. 20764-20789, 1990 Printed in U.S. A.
Uromodulin (Tamm-Horsfall Glycoprotein/Uromucoid) Is a Phosphatidylinositol-linked Membrane Protein*
(Received for publication, April 23, 1990)
Michael J. Rindler@, Sharon S. Naik$, Nanxin Li$, Timothy C. Hoops#Yi~~, and Marie-Noelle PeraldiS: ** From the Departments of SCell Biology and llEnvironmenta1 Medicine, New York University Medical Center, New York, lieu: York 10016
-.
Uromodulin, originally identified as an immunosup- pressive glycoprotein in the urine of pregnant women, has been previously shown to be identical to human Tamm-Horsfall glycoprotein (THP). THP is synthe- sized by the kidney and localizes to the renal thick ascending limb and early distal tubule. It is released into the urine in large quantities and thus represents a potential candidate for a protein secreted in a polarized fashion from the apical plasma membrane of epithelial cells in ho. After introduction of the full-length cDNA encoding uromodulin/THP into HeLa, Caco-2, and Madin-Darby canine kidney cells by transfection, however, the expressed glycoprotein was almost exclu- sively cell-associated, as determined by immunoprecip- itation after radioactive labeling of the cells. By im- munofluorescence, THP was localized to the plasma membranes of transfected cells. In transfected cell ex- tracts, THP also remained primarily in the detergent phase in a Triton X-l 14 partitioning assay, indicating that it has a hydrophobic character, in contrast to its behavior after isolation from human urine. Triton X- 114 detergent-associated THP was redistributed to the aqueous phase after treatment of cell extracts with phosphatidylinositol-specific phospholipase C. Treat- ment of intact transfected HeLa cells with phosphati- dylinositol-specific phospholipase C also resulted in the release of THP into the medium, suggesting that it is a glycosylphosphatidylinositol (GPI)-linked membrane protein. Similar to other known GPI-linked proteins, uromodulin/THP contains a stretch of 16 hydrophobic amino acids at its extreme carboxyl terminus which could function as a GPI addition signal and was shown to label with [3H]ethanolamine. The results indicate that THP is a member of this class of lipid-linked membrane proteins and is released into the urine after the loss of its hydrophobic anchor, probably by the action of a phospholipase or protease.
Uromodulin, a glycoprotein purified from the urine of preg- nant women based on its ability to interfere with antigen-
* This work was supported in part by Grant AGO1461 from the National Institutes of Health and Grant BC-643 from the American Cancer Society. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “advertisement” in accordance with 18 USC. Section 1734 solely to indicate this fact.
§ Recipient of a career development award from the Irma T. Hirsch1 Charitable Trust. To whom correspondence should be ad- dressed.
I( Recipient of Physician Scientist Award NIEHS000136. ** Present address: Hopital Necker, 22 Rue de Sevres, 75015 Paris,
France.
induced T-cell proliferation and monocyte cytotoxicity in vitro (I), has been recently shown (2, 3) to be identical to Tamm-Horsfall urinary glycoprotein (THP).’ THP is a major protein of renal origin in human urine and forms a gel-like precipitate in the presence of NaCl(4-6). It is also the primary component of waxy nephron casts encountered in the urine of normal individuals, but more frequently in certain patho- logical states (cf Ref. 7). First isolated in 1950, the function of uromodulin/THP is still unknown. It may play a role in regulating the circulating activity of cytokines since it binds to recombinant interleukin-1 and -2 and tumor necrosis factor with high affinity (8-10). It has also been implicated as a possible inhibitor of certain types of bacterial infection in the bladder and urinary tract because THP can bind to the type 1 pilus of Escherichia coli and prevent its attachment to the surface epithelium (e.g. Refs. 11-15). Due to its physicochem- ical properties, other investigators (7, 16, 17) have suggested that THP plays a role in active NaCl reabsorption or in the regulation of water permeability in the nephron.
THP has been localized by immunoelectron microscopy to the thick ascending limb of the loop of Henle (TAL) and the early distal convoluted tubule (DCT) segments of the nephron (16, 17) and is released into the urine in quantities that average 50 mg/day in humans (18). The release presumably involves its specific secretion from the apical surface of the polarized epithelial cells of these renal tubules. Because the cDNA for this protein has been isolated (2, 3), uromodulin/ THP represents a potentially useful marker for the study of the signals responsible for sorting secretory proteins in epi- thelial cells.
In this study, we have used the cDNA encoding for uro- modulin/THP to study the release of the expressed protein from cultured cells. In transient expression experiments, we show that much of the uromodulin/THP synthesized by HeLa cells and other cell types is on the plasma membrane and that it is anchored to the membrane via GPI. As such, it joins a large group of proteins that possess a phosphatidylinositol group linked through a carbohydrate chain and ethanolamine to their carboxyl-terminal amino acid (19, 20).
EXPERIMENTAL PROCEDURES
Cell Lines and Vectors-HeLa, COS, and AtT-20 cells were ob- tained from the American Type Culture Collection and cultured in Dulbecco’s modified Eagle’s medium (Whitaker M. A. Bioproducts, Walkersville, MD) containing 5% FBS (with 10% horse serum for
* The abbreviations used are: THP, Tamm-Horsfall glycoprotein; PI, phosphatidylinositol; SDS-PAGE, sodium dodecyl sulfate-poly- acrylamide gel electrophoresis; DCT, distal convoluted tubule; GPI, glycosylphosphatidylinositol; FBS, fetal bovine serum; MDCK, Ma- din-Darby canine kidney; TAL, thick ascending limb of the loop of Henle.
20784
by guest on May 27, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Uromodulin Linked to Membrane via PI 20785
AtT-20 cells) (HyClone Laboratories, Logan, UT) in an 8% CO? incubator at 37 “C. MDCK and Caco-2 cells were cultured as previ- ously described (21, 22). Transfection of COS cells was performed by the DEAE-dextran procedure followed by a dimethyl sulfoxide shock and incubation with chloroquine (23). Transfection of the other cell lines was performed as outlined previously using CaPOl precipitation and, in most cases, induction with sodium butyrate (24). Identical results were obtained in HeLa cells in the absence of butyrate induc- tion. The cDNA for uromodulin was obtained from C. Hession (Biogen, Cambridge, MA) already inserted into a vector (pBG311) similar to pBG312Mo (3), but containing the SV40 early promoter (25). A plasmid with uromodulin under the control of the Rous sarcoma virus promoter was constructed by subcloning the cDNA into a pRSV-globin vector in place of globin (26). The resulting pRSVuro was utilized (without butyrate induction) in some experi- ments as indicated in the text.
Radioactiue Labeling and Triton X-l 14 Partitioning-Metabolic labeling was performed 48-54 h after transfection. In most experi- ments, cells in 12-well Corning dishes were preincubated in methio- nine-free minimal essential medium (GIBCO) containing 2% dialyzed FBS (GIBCO) for 30 min and labeled for 3-16 h with 125 &i/ml [ “Slmethionine (>lOOO Ci/mmol; Du Pont-New England Nuclear) in the same medium (for overnight labeling, 5% normal FBS was added). Medium was collected, and cell extracts were prepared by sonication in 0.2 ml of 1% SDS. [?H]Ethanolamine labeling (200 &i/ ml, 29.5 Ci/mmol; Amersham Corp.) was performed on COS cells in 6-well dishes in Dulbecco’s modified Eagle’s medium containing 5% dialyzed FBS and was 11 h in duration. In this case, [?S]methionine labeling was performed for 4 h in parallel dishes. Immunoprecipita- tion was as previously outlined (21) using goat anti-human uromucoid (Cappel Laboratories, Malvern, PA) followed by rabbit anti-goat IgG (Sigma) or protein G coupled to Sepharose (Pharmacia LKB Bio- technology Inc.). For Triton X-114 partitioning (27), cell extracts were solubilized by sonication in 0.2 ml of buffer A (1% Triton X- 114 in 5 mM EDTA, 50 mM Tris-HCl (pH 7.5), and 0.01% phenol red with a protease inhibitor mixture). After incubation at 4 “C for 30 min, the samples were loaded above 0.3 ml of 0.3 M sucrose, heated to 37 “C for 5 min, and centrifuged at 5000 x g for 5 min. The supernatant (aqueous phase) and pellet (detergent phase) were diluted in ice-cold 100 mM Tris-HCl, 6 mM EDTA, 190 mM NaCl, 2.5% Triton X-100 (pH 8.0) and immunoprecipitated. The resulting sam- ples were subjected to SDS-PAGE on 8.5% gels followed by fluorog- raphy using EN’HANCE (Du Pont-New England Nuclear). For PI- specific phospholipase C treatment, Triton X-114 pellets were resus- pended in 0.2 ml of buffer A and incubated with 3 units of the purified enzyme (from Bacillus thuringiensis; a generous gift of Drs. S. Uden- friend and J. Berger, Roche Institute for Molecular Biology, Nutley, NJ) for 3 h at 37 “C before a second round of Triton X-114 extraction was performed. In other experiments, PI-specific phospholipase C treatment was performed on cells detached from the culture dish by gentle scraping exactly as described by Berger et al. (28).
Isolation of Tamm-Horsfall Glycoprotein-THP was purified from human urine by two rounds of salt precipitation by the method of Tamm and Horsfall(5,6). Gel electrophoretic analysis and Coomassie Blue staining indicated that the protein was essentially pure. Triton X-114 extraction was performed using 25 pg as indicated above except that a preliminary centrifugation was conducted at 4 “C. One-fifth of each sample was taken for SDS-PAGE on 8.5% gels. Western blotting was performed by standard procedures after electrophoretic transfer of the separated proteins to nitrocellulose. Monoclonal antibody against THP (Axell, Westbury, NY) was applied as the first step followed by horseradish peroxidase-conjugated goat anti-mouse IgG (Cappel Laboratories).
Zmmunofluorescence Microscopy-Cells grown on glass coverslips were fixed in 4% paraformaldehyde 48 h after transfection and stained with either goat anti-uromucoid antibodies or a monoclonal antibody against THP (Axell) followed by rhodamine-conjugated rabbit anti- goat IgG or anti-mouse IgG (Cappel Laboratories), respectively, using procedures given in detail elsewhere (21). Depending on the cell line used, l-30% of the cells specifically expressed THP.
RESULTS
UromodulinlTHP Is a Membrane-associated Protein in Cells Transfected with cDNA-The cDNA encoding for uro- modulin/THP was introduced into HeLa cells by transfection. After labeling the cells for 3 h with [35S]methionine followed
A medium cells B 3h label + chase T--i-77-
-- Med. Cell Med. Cell
*
a b c-d a b c d FIG. 1. Uromodulin/THP remains cell-associated in trans-
fected HeLa cells. A, HeLa cells grown in 35-mm dishes were transfected with the cDNA for uromodulin in the pBG311 vector or mock-transfected as outlined under “Experimental Procedures.” 48 h after transfection, cells were labeled with [‘“Slmethionine for 3 h before the medium was collected, and cell extracts were prepared. Samples were subjected to immunoprecipitation with anti-THP an- tibodies, SDS-PAGE, and fluorography. Little protein corresponding to THP was found in the medium from transfected (T) cells (lane a) and none in nontransfected C (control) cells (lane b), even in this overexposed radiograph. A doublet corresponding to THP was ob- served, however, in the transfected cell extract (lane c) that was not detected in the control (lane d). B, in a separate experiment, HeLa cells were transfected with pRSVuro and analyzed as described for A. Little release of THP into the medium was observed during a 3-h pulse (lane a) or a subsequent 3-h chase period (lane c), yet the amount of THP in the cell extract was the same after the chase (lane d) as before (lane b). Thus, uromodulin/THP is a cellular protein and only poorly released into the medium. Arrowheads indicate the positions of the 97- and 68-kDa markers.
MDCK medium cells
T--E-T---c
> . w
ab cd
Caco-2 medium cells
F--i-G---L
e f 9 h FIG. 2. THP remains cell-associated in transfected MDCK
and Caco-2 cells. MDCK cells (lanes a-d) and Caco-2 cells (lanes e-h) were transfected and analyzed as described for Fig. 1A after a 5- h (MDCK) or 16-h (Caco-2) labeling period. As was true in the case of HeLa cells, the THP doublet was found in the cell extracts from transfected dishes (lanes c and g), but not from nontransfected dishes (lanes d and h). THP was virtually undetectable in the medium in either transfected dishes (lanes a and e) or control (lanes b and f).
by immunoprecipitation, THP was readily observed, migrat- ing on polyacylamide gels as a doublet of -87 and -97 kDa* in the extracts from transfected cells, but not from mock- transfected controls (Fig. IA). Interestingly, samples of the medium had almost no detectable THP (Fig. IA, lanes a and b), although a 97-kDa band was observed on prolonged ex- posure of the gel (data not shown). Failure to detect uromo- dulin/THP in the medium was not the result of slow kinetics of secretion. When cells were labeled for 3 h and chased for an additional 3 h, very little THP was released, yet the cell-
’ The distance of migration of the uromodulin/THP doublet on polyacrylamide gels varied depending on the cell type and the exper- iment. The lower band range was 87-92 kDa, whereas the higher molecular mass component migrated between 95 and 110 kDa. For simplicity, we refer to them as 87 and 97 kDa in the text.
by guest on May 27, 2018
http://ww
w.jbc.org/
Dow
nloaded from
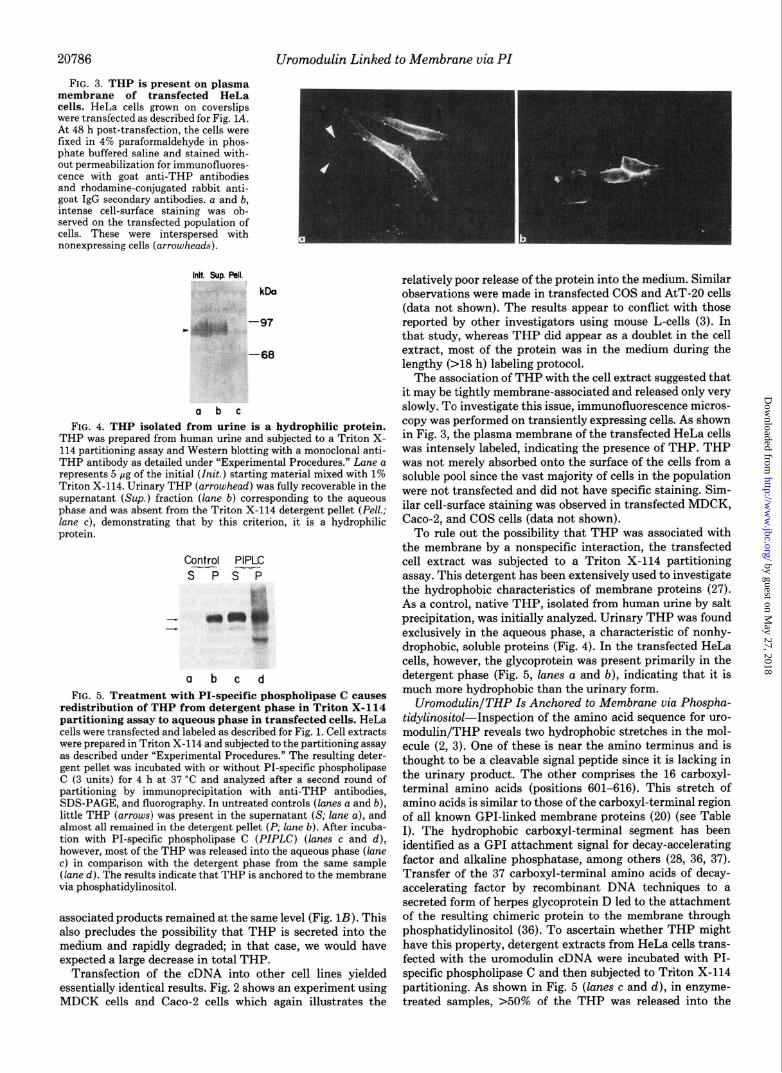
20786 urc FIG. 3. THP is present on plasma
membrane of transfected HeLa cells. HeLa cells grown on coverslips were transfected as described for Fig. 1A. At 48 h post-transfection, the cells were fixed in 4% paraformaldehyde in phos- phate buffered saline and stained with- out permeabilization for immunofluores- cence with goat anti-THP antibodies and rhodamine-conjugated rabbit anti- goat IgG secondary antibodies. a and b, intense cell-surface staining was ob- served on the transfected population of cells. These were interspersed with nonexpressing cells (arrowheads).
Ink. Sup RII.
kDa
-68
a b c
FIG. 4. THP isolated from urine is a hydrophilic protein. THP was prepared from human urine and subjected to a Triton X- 114 partitioning assay and Western blotting with a monoclonal anti- THP antibody as detailed under “Experimental Procedures.” Lane a represents 5 ig of the initial (init.) starting material mixed with 1% Triton X-114. Urinarv THP (arrowhead) was fullv recoverable in the supernatant (Sup.) fraction (lane b) correspondkg to the aqueous phase and was absent from the Triton X-114 detergent pellet (Pell.; lane c), demonstrating that by this criterion, it is a hydrophilic protein.
Control PIPLC s P ST
a b c d FIG. 5. Treatment with PI-specific phospholipase C causes
redistribution of THP from detergent phase in Triton X-114 partitioning assay to aqueous phase in transfected cells. HeLa cells were transfected and labeled as described for Fig. 1. Cell extracts were prepared in Triton X-114 and subjected to the partitioning assay as described under “Experimental Procedures.” The resulting deter- gent pellet was incubated with or without PI-specific phospholipase C (3 units) for 4 h at 37 “C and analyzed after a second round of partitioning by immunoprecipitation with anti-THP antibodies, SDS-PAGE, and fluorography. In untreated controls (lanes a and b), little THP (arrows) was present in the supernatant (S; lane a), and almost all remained in the detergent pellet (P; lane b). After incuba- tion with PI-specific phospholipase C (PIPLC) (lanes c and d), however, most of the THP was released into the aqueous phase (lane c) in comparison with the detergent phase from the same sample (lane d). The results indicate that THP is anchored to the membrane via phosphatidylinositol.
associated products remained at the same level (Fig. 1B). This also precludes the possibility that THP is secreted into the medium and rapidly degraded; in that case, we would have expected a large decrease in total THP.
Transfection of the cDNA into other cell lines yielded essentially identical results. Fig. 2 shows an experiment using MDCK cells and Caco-2 cells which again illustrates the
relatively poor release of the protein into the medium. Similar observations were made in transfected COS and AtT-20 cells (data not shown). The results appear to conflict with those reported by other investigators using mouse L-cells (3). In that study, whereas THP did appear as a doublet in the cell extract, most of the protein was in the medium during the lengthy (>18 h) labeling protocol.
The association of THP with the cell extract suggested that it may be tightly membrane-associated and released only very slowly. To investigate this issue, immunofluorescence micros- copy was performed on transiently expressing cells. As shown in Fig. 3, the plasma membrane of the transfected HeLa cells was intensely labeled, indicating the presence of THP. THP was not merely absorbed onto the surface of the cells from a soluble pool since the vast majority of cells in the population were not transfected and did not have specific staining. Sim- ilar cell-surface staining was observed in transfected MDCK, Caco-2, and COS cells (data not shown).
To rule out the possibility that THP was associated with the membrane by a nonspecific interaction, the transfected cell extract was subjected to a Triton X-114 partitioning assay. This detergent has been extensively used to investigate the hydrophobic characteristics of membrane proteins (27). As a control, native THP, isolated from human urine by salt precipitation, was initially analyzed. Urinary THP was found exclusively in the aqueous phase, a characteristic of nonhy- drophobic, soluble proteins (Fig. 4). In the transfected HeLa cells, however, the glycoprotein was present primarily in the detergent phase (Fig. 5, lanes a and b), indicating that it is much more hydrophobic than the urinary form.
UromodulinlTHP Is Anchored to Membrane via Phospha- tidylinositol-Inspection of the amino acid sequence for uro- modulin/THP reveals two hydrophobic stretches in the mol- ecule (2, 3). One of these is near the amino terminus and is thought to be a cleavable signal peptide since it is lacking in the urinary product. The other comprises the 16 carboxyl- terminal amino acids (positions 601-616). This stretch of amino acids is similar to those of the carboxyl-terminal region of all known GPI-linked membrane proteins (20) (see Table I). The hydrophobic carboxyl-terminal segment has been identified as a GPI attachment signal for decay-accelerating factor and alkaline phosphatase, among others (28, 36, 37). Transfer of the 37 carboxyl-terminal amino acids of decay- accelerating factor by recombinant DNA techniques to a secreted form of herpes glycoprotein D led to the attachment of the resulting chimeric protein to the membrane through phosphatidylinositol (36). To ascertain whether THP might have this property, detergent extracts from HeLa cells trans- fected with the uromodulin cDNA were incubated with PI- specific phospholipase C and then subjected to Triton X-114 partitioning. As shown in Fig. 5 (lanes c and d), in enzyme- treated samples, >50% of the THP was released into the
by guest on May 27, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Uromodulin Linked to Membrane via PI
TABLE I
20787
Carboxyl-terminal sequences of uromodulin/THP and known GPI-anchored proteins The extreme carboxyl-terminal amino acids of four members of the family of proteins linked to the plasma
membrane via GPI are compared to that of uromodulin/THP protein from its cDNA sequence. The terminal amino acids of the mature proteins, when known, are indicated by asterisks (for Torpedo marmorata acetylcholin- esterase, the carboxyl-terminal segment is assumed to be the same as that of the closely homologous Torpedo californica enzyme). Consecutive stretches of 10 or more hydrophobic residues are underlined.
Molecule Amino acid sequence Ref.
Uromodulin/THP KGVQATVSRAFSSLGLLKVWLPLLLSATLTLTFQ 2, 3 Trypanosoma brucei VSG” IL Tatl.1 TTNTTGSsNSFVIHKAPLFLAFLLF 29 Human thy-l RDKLVKC*EGISLLAQNTSWLLLLLLSLSLLQATDFMSL 30
Human placental APase PPAGTTD*AAHPGRSVVPALLPLLAGTLLLLETATAP 31-33 T. marmorata AChEase LLNATAC:DGELSSSGTSSSKGIIFYVLFSILYLIFY 34,35
a VSG, variant surface glycoprotein; APase, alkaline phosphatase; AChEase, acetylcholinesterase.
Control PIPLC 2FT-n
-aa@* * r3’
a b c d FIG. 6. THP is released from plasma membrane of trans-
fected cells by PI-specific phospholipase C. HeLa cells were transfected in this case with pRSVuro (see “Experimental Proce- dures”) and labeled as described for Fig. 1. Cells scraped from the dish were incubated in suspension with or without PI-specific phos- pholipase C (PZPLC) (3 units for 4 h at 37 “C). After centrifugation at 12,000 x g for 2 min, the supernatants (S; lanes a and c) and cell pellets (P; lanes b and d) were analyzed as outlined for Fig. 5. In untreated samples, as expected, there was little release of THP into the supernatant (lane a), with almost all appearing in the pellet (lane b). By contrast, PI-specific phospholipase C treatment caused most of the higher molecular mass form of THP to be released into the supernatant fraction (lane c) as compared with the cell extract (lane d). This indicates that THP on the cell surface is anchored to the plasma membrane by a phosphatidylinositol linkage.
+3 %i
r?‘:
a b FIG. 7. THP contains ethanolamine. COS cells were trans-
fected with pRSVuro (see “Experimental Procedures”) and metabol- ically labeled 48 h later with [8%]methionine for 4 h (lane a) or with [RH]ethanolamine for 11 h (lane b). Immunoprecipitates from the cell extracts were analyzed by SDS-PAGE and fluorography. Like other GPI-linked proteins, THP (arrowheads) was readily labeled with [3H] ethanolamine.
aqueous phase, indicating that THP fits the definition of a GPI-linked membrane protein. THP on the surface of the cell was also GPI-linked. When transfected cells were scraped from the culture dish and incubated directly with PI-specific phospholipase C, THP was released into the medium (Fig. 6). Interestingly, under these conditions, only the higher molec- ular mass form was released, suggesting that the faster mi- grating 87-kDa species was not accessible to the enzyme. By
contrast, both were detected in the aqueous phase when the PI-specific phospholipase C treatment was conducted after detergent solubilization of the cells (Fig. 5). This result indi- cates that the lower molecular mass form of the protein, which retains high mannose oligosaccharides sensitive to the action of endoglycosidase H (data not shown), is a lipid-linked intracellular precursor of surface THP.
To further confirm the existence of a GPI anchor in THP, transfected cells were labeled with [3H]ethanolamine. Etha- nolamine is a component of all known GPI linkages. In mammalian proteins, at least two ethanolamine molecules are known to be present per anchor, one of which is covalently attached to the carboxyl group of the terminal amino acid of the protein (reviewed in Refs. 19 and 20). As shown in Fig. 7, both the intracellular and mature forms of THP, in this case expressed in COS cells, were readily labeled with [“Hletha- nolamine.
DISCUSSION
Tamm-Horsfall glycoprotein, whereas previously consid- ered a secretory protein, has been shown, by immunoelectron microscopy, to be present on the plasma membranes of cells lining the renal DCT and TAL. In the epithelial cells of the human and rat DCTs, the protein was localized to the apical membrane (16, 38, 39). (Although label was seen on basal infoldings in some studies (17, 40, 41), this was likely due to misidentification of TAL cells as DCT cells at the TAL-DCT boundary (30).) Several investigators (16, 38) have argued, however, that the apical material represents secretory THP from the tubular lumen absorbed on the cell membrane, despite the fact that the neighboring collecting tubule and macula densa cells, which might also be expected to absorb the soluble protein, had no detectable THP on their surfaces. In the medullary TAL, THP is present on both the apical and basolateral surfaces (16, 17, 38-42). It is also prominent in clear vesicles in the apical cytoplasm (38, 39). Some of the label corresponding to THP was in the lumen of these vesicles, supporting the notion that in this location the protein is no longer membrane-associated (38). The apical vesicles appear to represent exocytotic intermediates, and thus, the TAL is considered a likely site of release of THP.
The data presented here demonstrate that uromodulin/ THP is a plasma membrane-associated protein and that its mode of attachment to the lipid bilayer is via phosphatidyli- nositol. The protein, when expressed from the cDNA, was located on the cell surface of transfected HeLa, Caco-2, and MDCK cells and also COS and AtT-20 cells (data not shown) and was released from the membrane by exogenously added PI-specific phospholipase C. [3H]Ethanolamine was also found to be incorporated into it. Moreover, the carboxyl terminus predicted from the cDNA sequence ends in a 16-
by guest on May 27, 2018
http://ww
w.jbc.org/
Dow
nloaded from

20788 Uromodulin Linked to Membrane via Pl
amino acid hydrophobic stretch typical of GPI-linked mem- brane proteins. A number of proteins that carry this lipid linkage are residents of the apical surface of polarized epithe- lial cells (19). Indeed, it has recently been shown (43,44) that proteins that either have no signal for polarized expression in epithelial cells or that normally appear on the basolateral surface are transported to the apical surface when a carboxyl- terminal GPI signal is attached to them. This implies that the GPI anchor itself is a signal specifying apical transport and would explain why THP is present primarily on the luminal cell surface of the DCT cells. The consistently ob- served appearance of THP also on the basal surface of cells of the TAL is more difficult to reconcile with this hypothesis and yet is perhaps more physiologically relevant if one of the functions of THP is to regulate the level of circulating cyto- kines in the circulation. It is conceivable that the steady-state distribution of THP on the cell surface does not reflect the polarity of its intracellular transport since newly synthesized THP destined for the apical membrane might be released rapidly into the tubular lumen and hence be underestimated in the immunolocalization. Nonetheless, it remains an open question at this time as to how THP, putatively synthesized as a GPI-anchored protein in the kidney itself, is to some extent routed to the basolateral membrane in this nephron segment.
One potential means for the secretion of THP into the urine would be the synthesis of forms that lack the carboxyl- terminal signal for GPI attachment altogether. In the case of decay-accelerating factor and acetylcholinesterase, it has been postulated (34, 35, 45) that secreted versions of the proteins are derived by alternative splicing of the mRNA, resulting in the elimination of the carboxyl-terminal hydrophobic segment of the translated proteins. This mechanism seems unlikely for THP since only one 2.3-2.4-kilobase mRNA species was observed by Northern blotting using the uromodulin cDNA as a probe, and no variants were detected in at least four independently isolated cDNAs reported by two different lab- oratories (2, 3). A more likely hypothesis is that the protein is released from the membrane by proteolytic or phospholi- polytic cleavage, as is apparently the case for forms of thy-l, N-CAM, and Qa, three naturally occurring GPI-linked pro- teins that are known to be shed from the surface of their host cells (46-48). In this regard, glycosylphosphatidylinositol- specific phospholipase C or D activities have been identified in plasma and liver (49-52), although none of these enzymes has been shown to act on native GPI-linked membrane pro- teins without prior solubilization. Similar activities might exist in the nephron, possibly even in the intracellular mem- branes of cells of the TAL.
Our observation that uromodulin/THP can exist as a mem- brane-associated protein appears to contradict the previously reported secretion of the protein from transfected cells (3). The discrepancy may in part be due to the lengthy labeling protocol used in that study (18 h) compared to the 3-4-h labeling period employed here. A slow rate of release coupled with turnover of the plasma membrane form would be ex- pected to result in a much higher proportion detectable in the medium after prolonged labeling periods. More important, the cells used previously (L-cells) have been reported to lack the machinery for efficient attachment of the GPI anchor to proteins (53), leading to the secretion of &a in cells transfected with the cDNA, in contrast to other cell types where the expressed Qa molecules remained tethered to the cell surface by the phosphatidylinositol group. Since the transfer of GPI to the carboxyl terminus in the endoplasmic reticulum must be a two-step process requiring both a proteolytic cleavage
event and a subsequent transfer of the lipid-glycan moiety to the newly revealed terminal amino acid (20), should only the first step occur, a secretory protein could result. The cells of the thick ascending limb of the nephron may also have a limited capacity for GPI attachment and secrete large amounts of newly synthesized THP.
Acknowledgments-We thank Catherine Hession for providing the cDNA, Min Dong Ren and Drs. Gary Beaudry, Joel Berger, Michael Davitz, and David Sabatini for assistance and advice, and Heidi Plesken, Jodi Culkin, and Frank Forcino for illustrations and pho- tography.
REFERENCES
1. Muchmore, A. V., and Decker, J. M. (1987) Science 229, 479- 481
2. Pennica, D., Kohr, W. J., Kuang, W.-J., Glaister, D., Aggarwal, B. B., Chen, E. Y., and Goeddel, D. V. (1987) Science 236,83- 88
3. H&ion, C., Decker, J. M., Sherhlom, A. P., Kumar, S., Yue, C. C.. Mattaliano, R. J.. Tizard, R., Kawashima. E.. Schmeisser, U.: Heletky, S., Chow, E. p., Burne, C. A.; Shaw, A., and Muchmore, A. V. (1987) Science 237, 1479-1484
4. Stevenson, F. K., Cleave, A. J., and Kent, P. W. (1971) Biochim. Biophys. Acta 236, 59-66
5. Tamm, I., and Horsfall, F. L. (1950) Proc. Sot. Exp. Biol. Med. 74,108-114
6. Tamm, I., and Horsfall, R. L., Jr. (1952) J. Exp. Med. 95, 71-97 7. Hover. J. R., and Seiler. M. W. (1979) Kidnev Znt. 16, 279-289 8. Muchmore, A. V., and Decker, J. M. (1986) 3. Biol. &em. 261,
13404-13407 9. Sherblom. A. P., Decker, J. M., and Muchmore. A. V. (1988) J.
Biol. Chem. 263,5418-5424 10. Sherblom, A. P., Sathyamoorthy, N., Decker, J. M., and Much-
more, A. V. (1989) J. Zmmunol. 143, 939-945 11. Orskov, I., Ferencz, A., and Orskov, F. (1980) Lancet 1,887-889 12. Ohanlev. P.. Lark. D.. Falkow. S.. and Schoolnik. G. (1985) J.
Clin. ?noedt. 75,‘347-360 13. Parkkinen, J., Virkola, R., and Korhonen, T. K. (1988) Inject.
Zmmun. 56, 2623-2630 14. Dulawa, J., Jain, K., Thomsen, M., Ranbausek, M., and Ritz, E.
(1988) J. Clin. Znuest. l&87-89 15. Duncan, J. L. (1988) J. Inject Dis. 158, 1379-1382 16. Hoyer, J. R., Sisson, S. P., and Vernier, R. L. (1979) Lab. Znuest.
41,168-173 17. Sikri, K. L., Foster, C. L., Bloomfield, F. J., and Marshall, R. D.
(1979) Biochem. J. 181, 525-532 18. Grant, A. M. S., and Neuberger, A. (1973) Clin. Sci. (Orf.) 44,
163-179 19. Low, M. G., and Saltiel, A. L. (1988) Science 239, 268-275 20. Ferzuson. M. A. J.. and Williams. A. F. (1988) Annu. Reu.
Biochem. 57, 285-320 21. Rindler, M. J., Ivanov, I. E., Plesken, H., Rodriguez-Boulan, E.,
and Sabatini, D. D. (1984) J. Cell Biol. 98,1304-1319 22. Rindler, M. J., and Traber, M. G. (1988) J. Cell Biol. 107, 471-
479 23. Kaufman, R. J. (1990) Methods Enzymol. 185,487-511 24. Gottlieb, T. A., Beaudry, G., Rizzolo, L., Colman, A., Rindler, M.
J., Adesnik, M., and Sabatini, D. D. (1986) Proc. Natl. Acad. Sci. U. S. A. 83,2100-2104
25. Cate, R. L., Mattaliano, R. J., Hession, C., Tizard, R., Farber, N. M., Cheung, A., Ninfa, E. G., Frey, A. Z., Gash, D. J., Chow, E. P., Fisher, R. A., Bertonis, J. M., Torres, G., Wallner, B. P., Ramachandran, K. L., Ragin, R. C., Mangarano, T. F., Mac- Laughlin, D. T., and Donahoe, P. K. (1986) Cell 46,685-698
26. Gorman, C. M., Padmanabhan, R., and Howard, B. H. (1983) Science 22 1,551-553
27. Bordier. C. (1981) J. Biol. Chem. 256, 1604-1607 28. Berger, ‘J., Howard, A. D., Brink, L., Gerber, L., Hauber, J.,
Cullen. B. R.. and Udenfriend. S. (1988) J. Biol. Chem. 263, 10016-10021
29. Rice-Ficht, A. C., Chen, K. K., and Donelson, J. E. (1982) Nature 298,676-679
30. Seki, T., Spurr, N., Obata, F., Goyert, S., Goodfellow, P., and Silver, J. (1985) Proc. Natl. Acad. Sci. U. S. A. 82, 6657-6661
31. Millan, J. L. (1986) J. Biol. Chem. 261, 3112-3115
by guest on May 27, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Uromodulin Linked to Membrane via PI 20789
32. Micanovic, R., Bailey, C. A., Brink, L., Gerber, L., Pan, Y. C., 28,441-452 Hulmes, J. D., and Udenfriend, S. (1988) hoc. N&l. Acad. Sci. 43. Brown, D. A., Crise, B., and Rose, J. K. (1989) Science 245, U. S. A. 85.1398-1402 1499-1501
33. Ogata, S., Hayashi, Y., Takami, N., and Ikehara, Y. (1988) J. Biol. Chem. 263,10489-10494
34. Gibney, G., MacPhee-Quigley, K., Thompson, B., Vedvick, T., Low, M. G., Taylor, S. S., and Taylor, P. (1988) J. Biol. Chem. 263,1140-1145
35. Sikorav, J.-L., Duval, N., Anselmet, A., Bon, S., Krejci, E., Legay, C., Osterlund, M., Reimund, B., and Massoulie, J. (1988) EMBO J. 7,2983-2993
36. Caras, I. W., Weddell, G. N., Davitz, M. A., Nussenzweig, V., and Martin, D. W., Jr. (1987) Science 238,1280-1283
37. Caras, I. W., and Weddell, G. N. (1989) Science 243, 1196-1200 38. Bachmann, S., Koeppen-Hagemann, I., and Kriz, W. (1985)
Histochemistry 83, 531-538 39. Peach, R. J., Day, W. A., Ellingsen, P. J., and McGiven, A. R.
(1988) H&o&em. J. 20,156-164 40. Sikri, K. L.. Foster, C. L., MacHueh. N.. and Marshall. R. D.
(1981) J. knat. 132, 597-605 - 41. Sikri. K. L.. Alexander. D. P.. and Foster. C. L. (1982) J. Anat.
135,29-45 I ~I
42. Sisson, S. P., and Vernier, R. L. (1980) J. Histoch-em. Cytochem.
44. Lisanti, M. P., Caras, I. W., Davitz, M. A., and Rodriguez-Boulan, E. (1989) J. Cell Biol. 109, 2145-2156
45. Caras, I. W., Davitz, M. A., Rhee, L., Weddell, G. N., Martin, D. W., Jr., and Nussenzweig, V. (1987) Nature 325,545-549
46. Freimuth. W. M.. Esselman. W. J.. and Miller. H. C. (1978) J. Zmmunbl. 120,‘1651-1658’
47. He, H. T., Finne, J., and Goridis, C. (1987) J. Cell Biol. 105, 2489-2500
48. Robinson, P. J. (1987) Proc. N&2. Acad. Sci. U. S. A. 84, 527- 531
49. Davitz, M. A., Hereld, D., Shak, S., Krakow, J., Englund, P. T., and Nussenzweig, V. (1987) Science 238,81-84
50. Low, M. G., and Prasad, A. R. (1988) Proc. N&l. Acad. Sci. U. S. A. 85,980-984
51. Davitz, M. A., Horn, J., and Schenkman, S. (1989) J. Biol. &em. 264,13760-13764
52. Fox, J. A., Soliz, M. N., and Saltiel, A. R. (1987) hoc. Natl. Acad. Sci. U. S. A. 87,2663-2667
53. Stroynowski, I., Soloski, M., Low, M. G., and Hood, L. (1987) Cell 50, 759-768
by guest on May 27, 2018
http://ww
w.jbc.org/
Dow
nloaded from

M J Rindler, S S Naik, N Li, T C Hoops and M N Peraldiphosphatidylinositol-linked membrane protein.
Uromodulin (Tamm-Horsfall glycoprotein/uromucoid) is a
1990, 265:20784-20789.J. Biol. Chem.
http://www.jbc.org/content/265/34/20784Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/265/34/20784.full.html#ref-list-1
This article cites 0 references, 0 of which can be accessed free at
by guest on May 27, 2018
http://ww
w.jbc.org/
Dow
nloaded from