university of veterinary medicine hannover thesis doctor of
TRANSCRIPT

University of Veterinary Medicine Hannover
Department of Neurosurgery, Hannover Medical School Center for Systems Neuroscience, Hannover
Deep brain stimulation in rats with deficient sensorimotor gating: behavioral and electrophysiological studies
THESIS
Submitted in partial fulfillment of the requirements for the degree
DOCTOR OF PHILOSOPHY
(PhD)
awarded by the University of Veterinary Medicine Hannover
by
Svilen Angelov
Gabrovo, Bulgaria
Hannover, Germany 2014

Supervisor: Prof. Dr. Joachim K. Krauss
Co-Supervisor: Prof. Dr. Kerstin Schwabe
Supervision Group: Prof. Dr. Andrej Kral Prof. Dr. Joachim K. Krauss Prof. Dr. Stefan Bleich
1st Evaluation: 1st Evaluation: Prof. Dr. Joachim K. Krauss
(Department of Neurosurgery, Hannover Medical School)
Prof. Dr. Andrej Kral (Institute for AudioNeurotechnology, Hannover Medical School)
Prof. Dr. Stefan Bleich (Department of Psychiatry, Hannover Medical School)
2nd Evaluation: Prof. Dr. Michael Koch (Brain Research Institute,
Department of Neuropharmacology, Center for Cognitive Sciences, Bremen)
Date of final exam: 28.03.2014
Sponsorship: Tourette Syndrome Association

To my friend and colleague Edita


Table of contents 1 Introduction ................................................................................................................... 1
2 Tourette’s syndrome ..................................................................................................... 3
3 Basal ganglia ................................................................................................................ 6
4 Deep brain stimulation ................................................................................................ 10
5 Animal Models for neuropsychiatric disorders ............................................................ 14
5.1 Pharmacological models for deficient PPI .......................................................... 16
5.2 Developmental models for deficient PPI ............................................................ 18
5.3 Genetic models for deficient PPI......................................................................... 18
5.4 Selective breeding for deficient PPI .................................................................... 19
6 Objectives ................................................................................................................... 22
7 Manuscript one: Effect of deep brain stimulation in rats selectively bred for reduced
prepulse inhibition ........................................................................................................... 23
8 Manuscript two: Neuronal activity of the prefrontal cortex is reduced in rats selectively
bred for deficient sensorimotor gating ............................................................................ 25
9 Discussion. ................................................................................................................. 47
10 Summary (Zusammenfassung) ................................................................................ 50
11 References ............................................................................................................... 54
12 Appendix ................................................................................................................... 62

Frequently used abbreviations
5-HT Serotonin (5-hydroxytryptamine)
ADHD Attention deficit hyperactivity disorder
ASR Acoustic startle reaction
BG Basal ganglia
CM-Pf Centromedian-parafasciculus
CSPT Cortico-striato-pallido-thalamic
DA Dopamine
DBS Deep brain stimulation
EPN Entopeduncular nucleus
GPi Globus pallidus internus
HD Huntington’s disease
mPFC Medial prefrontal cortex
NAC Nucleus accumbens
NMDA N-methyl-D-aspartate
OCD Obsessive compulsive disorder
PD Parkinson’s disease
PPI Prepulse inhibition
PPTg Pedunculopontine tegmental nucleus
SNr Substantia nigra pars reticulata
STN Subthalamic nucleus
TS Tourette’s syndrome

Introduction
1
1 Introduction
Deficiencies in the mechanisms that enable healthy individuals to suppress and “gate”
irrelevant sensory, cognitive and motor information are found in several neuropsychiatric
disorders, such as schizophrenia, obsessive-compulsive disorder (OCD), Tourette’s
syndrome (TS) and Huntington’s disease (Perry and Braff, 1994; Swerdlow et al., 1995,
2000, 2001a; Castellanos et al., 1996; Karper et al., 1996; Miguel et al., 2000; Perry et
al., 2001; Braff et al., 2001a). This disturbed inhibitory mechanism and ineffective
information filtering could be linked to the inability to filter or “gate” irrelevant or inferring
information of motor, cognitive and emotional domains with consecutive sensory
overload and respective clinical symptoms, e.g., in TS to the “urge” to tic patients cannot
enduringly suppress. It is still not clear how exactly sensorimotor gating is regulated, but
the cortico-striato-palido-thalamic (CSPT) network appears to play an important role in
this process (DeLong and Wichmann, 2007; Swerdlow et al., 2001b).
High frequency deep brain stimulation (DBS) in the basal ganglia (BG) and other brain
regions is currently under investigation for treatment of pharmacoresistant
neuropsychiatric disorders, including TS. While in movement disorders the procedure is
fairly standardized e.g., in Parkinson`s disease (PD) the main targets are the
subthalamic nucleus (STN) and the globus pallidus internus (GPi), for DBS in TS several
different areas of the brain have been targeted to treat tics (Hariz and Robertson, 2010;
Viswanathan et al., 2012). The Nucleus ventralis oralis/centromedian-parafascicular
complex (Voi/CM-Pf) in the thalamus has been used in the majority of those cases, but
the GPi and the nucleus accumbens (NAC) have also been targeted (Houeto et al.,
2005; Müller-Vahl et al., 2012; Shahed et al., 2007; Porta et al., 2012). Initially, DBS was
thought to reduce neuronal activity at the stimulation site, thus acting as a “functional
lesion”. However, recent studies support more complex modulation of neuronal networks
by DBS, such as alterations in oscillatory behavior of neuronal networks (Deniau et al.,
2010; DeLong and Wichmann, 2012; Lozano et al., 2010; Modolo et al., 2011; Vitek,
2008).

Introduction
2
The deficient sensorimotor gating can be operationally measured by the laboratory
paradigm “prepulse inhibition (PPI) of the acoustic startle response (ASR)” i.e. the
reduction of the ASR when the startling noise is shortly preceded by a weak prepulse
(Koch and Fendt, 2000; Swerdlow et al., 2001b). This reduction of the ASR is found
attenuated in patients and experimental animals with disturbed sensorimotor gating. PPI
can be measured in humans and animals by using virtually the same approaches (Braff
et al., 2001b). PPI is regulated by a neuronal circuitry that includes portions of the basal
ganglia (BG) also implicated in the pathophysiology of disorders with deficient
sensorimotor gating. Experimentally induced PPI-deficits in rodents are therefore
regarded a reliable and quantifiable cross-species endophenotype to study the
pathophysiology of disorders with deficient sensorimotor gating and the effect of DBS
thereof. In animals deficient PPI can be experimentally induced by a variety of
manipulations, including structural, pharmacological and genetical approaches.
Our main goal is to investigate the pathophysiological mechanisms underlying
neuropsychiatric disorders with deficient information processing, such as TS, and their
modulation by DBS. For our experiments we use rats, which are selectively bred for high
and low PPI (Schwabe et al., 2007). Behavior of rats with low PPI corroborate a number
of clinical findings in neuropsychiatric disorders with reduced sensorimotor gating
(Dieckmann et al., 2007, Freudenberg et al., 2007). We first evaluated the effect of DBS
in targets used to treat tics in TS, i.e., in the entopeduncular nucleus EPN (equivalent to
the human GPi), the CM-Pf and the NAC. Since PPI is regulated by a neuronal circuitry
that includes portions of the basal ganglia (BG) also implicated in the pathophysiology of
neuropsychiatric disorders with deficient sensorimotor gating, we additionally tested the
neuronal activity in key regions of this circuitry, i.e., the medial prefrontal cortex (mPFC),
the NAC and the EPN in rats with high and low PPI. With these experiments we wanted
to gain insight into the compromised neuronal network activity related to deficient
sensorimotor gating, to investigate in future studies the effect of DBS thereof.

Tourette’s syndrome
3
2 Tourette’s syndrome
Tourette`s syndrome is a neuropsychiatric disorder with a strong heritable component,
which is clinically characterized by the presence of motor and vocal tics, which usually
begin between the ages of 6 and 8 (Cohen SC et al., 2012). It is a quite common
disorder, with prevalence of about 1% of the population almost worldwide (Hariz and
Robertson, 2010). Diagnosis of TS requires the presence of multiple motor and at least
one vocal tic, which wax and wane in severity, but last longer than one year. Tics are
rapid, non rhythmic, stereotyped motor movements (motor tics) or vocalizations (vocal
tics). They can affect single muscle or small groups of muscles (simple tics, such as eye
blinking, throat clearing), or also more muscles acting in a coordinated pattern (complex
tics, such as gestures or uttering phrases). In contrast to the abnormal movements of
other movement disorders (choreas, dyskinesias), the tics of TS are temporarily
suppressible and often preceded by a premonitory urge. Patients describe the need to
tic as a tension or pressure which they need to release in order to relieve the unpleasant
sensation.
The etiology of TS is still unclear, but it is assumed that both genetic and environmental
factors are involved. It is currently estimated that the first-degree relatives of a TS
patient have a 5-15% risk of developing the disease themselves and a 10-20% risk of
developing any sort of tic (Am J Hum Genet, 2007). Genetic studies have shown that the
majority of cases are inherited, but to date a gene or a common variant with major effect
in TS etiology has not been identified (Bertelsen et al., 2013). The presence of genetic
heterogeneity, comorbidity, incomplete penetrance, involvement of both genetic and
environment factors makes the identification of candidate genes for TS very difficult.
Nevertheless, several genes including SLITRK1, IMMP2L, CNTNAP2, NLGN4, and
HDC are reported as involved in TS, but they are likely to represent rare susceptibility
variants (Bertelsen et al., 2013; Paschou P. 2013). Environmental factors such as
perinatal hypoxic events, prenatal maternal smoking, exposure to androgens and heat
are also believed to be involved in the TS etiology (Swain et al., 2007).

Tourette’s syndrome
4
The exact pathophysiological mechanisms of TS are also unknown. One possible
reason for the tics is a dysfunction in different cortical and subcortical regions.
Functional neuroimaging of TS patients reveals abnormal activation of the striatolimbic
and premotor area and the metabolic rate of the striato-sensorimotor region correlates
positively with tic expression (Alfredo Berardelli, 2003). The fact that dopamine receptor
antagonists (e.g. haloperidol) are effective in the treatment of tics in many patients with
TS suggests a major pathophysiological role for the dopaminergic systems. Indeed, an
increased dopaminergic receptors expression and increased dopamine levels have been
found in the striatum and prefrontal cortex (Singer HS., 2005). Exaggerated startle reflex
response is found in patients with TS, which can be a result from dopaminergic
hyperactivity (Gironell A, Berthier M, 2000). Another report describes abnormal PPI of
the ASR in TS patients (Swerdlow NR et al., 2001a), which is linked to the patient‘s
inability to suppress the response to internal stimuli and which can be very likely related
to a pathology within the CSPT network. Additionally, impaired functioning of the
serotoninergic system is also presumed (Mueller-Vahl et al., 2005).
Tourette’s syndrome is a spectrum disorder, so its severity ranges from light to severe.
While the overall prognosis is positive and the condition in most cases improves with
maturity, still some individuals with TS show symptoms in adulthood that can prevent
them from holding a job or having a satisfactory social life despite normal life span and
intelligence. The onset of tics is usually between 5 and 10 years of age and boys are
affected three to four times more commonly than girls (Arzimanoglou AA, 1998).
Comorbid conditions such as attention-deficit hyperactivity disorder (ADHD) and OCD
are present in aproximately 80-90% of clinical cohorts (Hariz and Robertson, 2010) and
these conditions often cause even more functional impairment to the individuals than the
TS itself, so they must be also always correctly identified and treated.
Most of the cases do not require pharmacological treatment at all, instead -
psychobehavioral therapy and education may be effective. In other, more severe cases,
when tics are functionally and socially impairing, or even painful, tic suppressive
medication is necessary. Standard therapy for TS includes anti-dopaminergic (e.g.
haloperidol), and alpha2-adrenergic drugs (e.g. clonidine). Most of the medically treated
patients nowadays receive treatment with atypical neuroleptic drugs - risperidone,

Tourette’s syndrome
5
pimozide, sulpiride, clonidine. However, the efficacy of the pharmacotherapy for TS is
limited and serious adverse effects, such fatigue, weight gain, sexual dysfunction,
akathisia and tardive dyskinesia are frequent (Roessner V. et al., 2011). In severe,
pharmacoresistant TS, DBS in different BG regions - NAC, GPi, STN and some
associated regions e.g. CM-Pf, bed nucleus of stria terminalis have been described to
be beneficial (Houeto et al., 2005; Ackermans et al., 2006; Kuhn et al., 2007; Maciunas
et al., 2007; Larson 2008; Welter et al., 2008; Neuner et al., 2009;)

Basal ganglia
6
3 Basal ganglia
There is general agreement that the BG circuitry and its frontocortical connections are
implicated in the pathophysiology of TS (Swerdlow NR and Young AB., 2001; Mink,
2001). Basal ganglia also play an important role in the regulation of sensorimotor gating,
which is disturbed in patients with TS is linked to the unwanted behaviors and thoughts
(Swerdlow et al., 2000). The targets where DBS is effectively applied for TS are also
parts of this circuitry. Therefore, better understanding of BG anatomy, physiology and
their role in sensorimotor gating could be useful to investigate the pathophysiology of
TS.
The BG are several richly interconnected nuclei in the brain. They receive major input
from the cerebral cortex and thalamus, and send their output back to the cortex (through
the thalamus) and to the brainstem. Basal ganglia take part in a variety of behaviors,
including emotional, cognitive and motor functions. They integrate multimodal sensory
information with preprogrammed behavioral routines and are consequently involved in
action selection and reinforcement learning.
The basal ganglia include the striatum, the globus pallidus (consisting of the pars
externa and pars interna), the substantia nigra (pars reticulata and pars compacta), and
the STN.
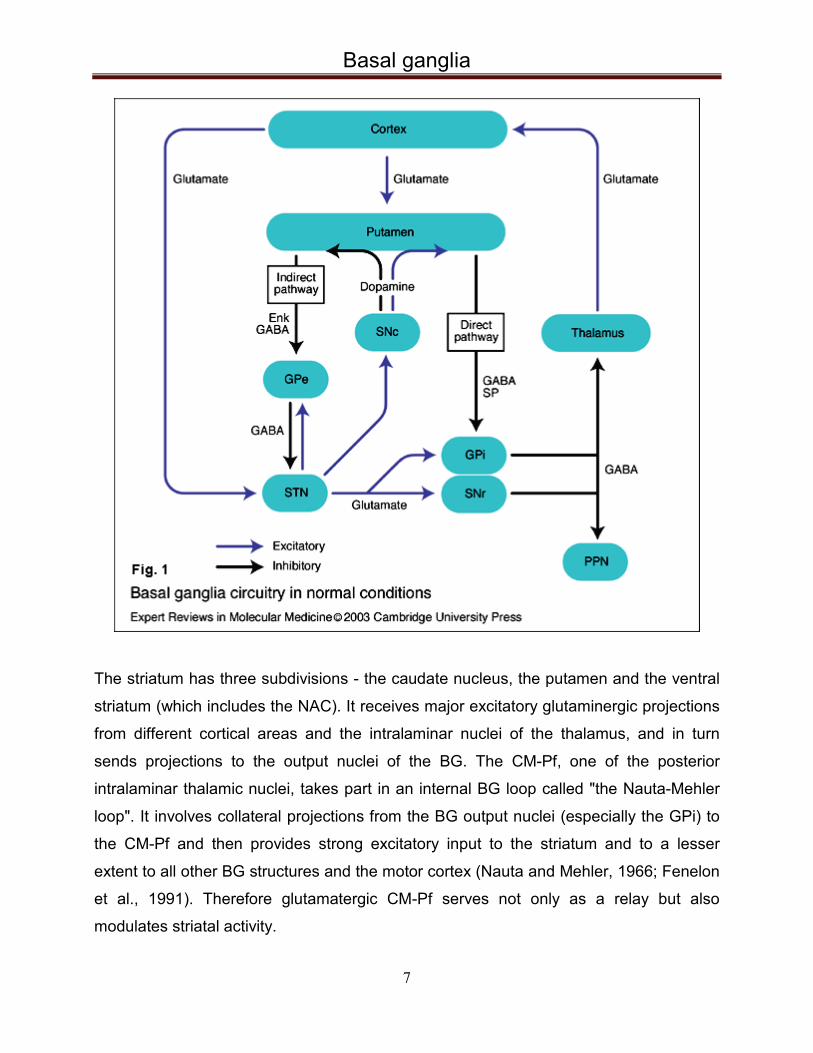
Basal ganglia
7
The striatum has three subdivisions - the caudate nucleus, the putamen and the ventral
striatum (which includes the NAC). It receives major excitatory glutaminergic projections
from different cortical areas and the intralaminar nuclei of the thalamus, and in turn
sends projections to the output nuclei of the BG. The CM-Pf, one of the posterior
intralaminar thalamic nuclei, takes part in an internal BG loop called "the Nauta-Mehler
loop". It involves collateral projections from the BG output nuclei (especially the GPi) to
the CM-Pf and then provides strong excitatory input to the striatum and to a lesser
extent to all other BG structures and the motor cortex (Nauta and Mehler, 1966; Fenelon
et al., 1991). Therefore glutamatergic CM-Pf serves not only as a relay but also
modulates striatal activity.

Basal ganglia
8
The two output nuclei of the basal ganglia, the GPi and substantia nigra pars reticulata
(SNr), keep their targets in the thalamus and brainstem under tonic inhibition. This
inhibitory output is modulated mainly by two parallel pathways – “direct” and “indirect”. In
the direct pathway, striatal neurons project directly to the GPi via a monosynaptic
inhibitory connection. The indirect pathway passes from the striatum first to the globus
pallidus externus (GPe), then to the STN, and finally from the STN to the output nuclei.
The STN is connected with both segments of the globus pallidus and the substantia
nigra and its glutamatergic projections are the only excitatory internal BG connections.
Activation of the direct pathway supresses the activity of GPi and SNr, which in turn
disinhibits the thalamus, and thereby leads to increased cortical activity and facilitates
movement. In contrast, activation of the indirect pathway further inhibits thalamocortical
neurons due to the increased GPi/SNr activity in result of the STN disinhibition, which
inhibits movement. An imbalance between the direct and indirect pathway as a result
from pathologically overactive dorsal striatum, which triggers repetitive, involuntary,
stereotyped movements through inhibition of GPi, could play an important role in the tic
generation in Tourette syndrome (Mink, 2001). Another hypothesis states that when
voluntary movement is generated by cerebral and cerebellar mechanisms, the basal
ganglia act to inhibit competing motor programs that would otherwise interfere with the
desired movement. A focused facilitation and surround inhibition of motor mechanisms
in thalamocortical and brainstem circuits may occur, which has been summarized as the
“central surround inhibition model” (Mink, 1996; 2003).
The BG system is often subdivided into three parallel functional circuits, although there
is significant level of overlapping between them. Best studied and maybe mostly affected
in movement disorders is the sensorimotor loop. It begins in the precentral motor and
postcentral somatosensory fields, which project mainly to the putamen and then through
the BG output nuclei and the thalamus back to the cortical areas. The associative loop
starts with the prefrontal cortex, which mainly projects to the caudate nucleus.
Thereafter, the information is directed through the GPi and SNr to the ventral anterior
and centromedian nuclei of the thalamus, which in turn project back to the cortical
zones. In the limbic loop, the input from the anterior cingulate cortex and medial
orbitofrontal cortex goes primarily to the ventral striatum, which includes NAC. The

Basal ganglia
9
ventral striatum projects through the ventral pallidum to the mediodorsal nucleus of the
thalamus and then again back to the cortex. These loops process motor, cognitive, and
emotional or motivational information, respectively (Parent and Hazrati 1995; Nakano et
al., 2000; Yelnik, 2002).

Deep brain stimulation
10
4 Deep brain stimulation
Deep brain stimulation (DBS), i.e., high frequency electrical stimulation of brain regions
involved in the pathophysiology of a particular neurological disease, is a functional
neurosurgical method. It is a developing, but potent treatment option for a number of
neurological and psychiatric disorders including TS. In comparisson with ablative
procedures, DBS is reversible, can be safely bilaterally performed and the stimulation
parameters can be precisely adjusted (Breit S. et al., 2004).
Fig. 2: X-Ray image of the DBS electrodes, implanted in the brain of a TS patient (2 in GPi and 2 in thalamus; Neurosurgery, MHH)
Deep brain stimulation is achieved by stereotactic surgical implantation of stimulation
electrodes in the targeted brain area and subsequent connecting of the electrodes to an
implantable pulse generator. The exact position of the target is accurately defined and
reached using a three dimensional Cartesian coordinate system based on a stereotactic
head ring (Krauss JK, Volkmann J, 2004)

Deep brain stimulation
11
Stimulation parameters i. e. waveform, frequency and the contact configuration, can be
precisely adjusted in the post-operative period via transcutaneous programming unit.
Electrophysiological investigation of the target can be performed prior to implantation in
order to increase targeting accuracy (Krauss JK, Volkmann J, 2004).
Fig. 3: Electrode and pacemaker implantation for DBS
(Picture from N Engl J Med. 2012, Okun MS.)
Stereotactic surgery in animals was first time established by Horsley and Clark (1908).
In humans, Spiegel and Wycis (1947) performed stereotactic lesions in the globus
pallidus in a patient with Huntington’s disease. The lesions were made by using different
techniques, including alcohol (Spiegel and Wycis, 1947), cryotherapy (Cooper and Lee,
1961) and temperature-controlled high frequency ablation (Cosman et al., 1983). The

Deep brain stimulation
12
indications included pain, movement disorders and neuropsychiatric conditions. First
attempts to treat TS patients by stereotactic procedures (thalamotomy) were made in the
early 1960s (Hassler and Dieckmann, 1970; Temel and Visser-Vandewalle, 2004).
While electrical stimulation was used to verify the localization prior to the stereotactic
lesions, it was found that the stimulation itself improved symptoms such as tremor or
pain, achieving the same effect as in the subsequently performed lesion. These
observations led to the idea that electrodes could be stereotactically implanted to
permanently apply high frequency electrical stimulation in the selected brain area. Brice
and McLellan (1980) are between the establishers of the DBS in targets within the BG
and associated brain regions as a method for neuromodulation. In 1987 it was first time
used in the treatment of PD by Benabid and colleagues. Despite the positive clinical
results, little is known about the exact mechanism of action of DBS. While it was initially
assumed that DBS leads primarily to a neuronal inhibition, which interrupts the
information flow, there is also data that contradicts this hypothesis. For example, in
contrast to what would be expected if stimulation inhibits output from the stimulated
structure, stimulation in GPe improves bradykinesia (Vitek JL, 2002). Furthermore,
stimulation of GPi improves both hypokinetic and hyperkinetic movement disorders, i.e.,
PD and dystonia. This data leads to the assumption, that DBS exerts not just a local
inhibitory, but more likely a network-wide modulatory effect, possibly by disruption of
abnormal circuit activity (Hashimoto et al., 2003; Lozano et al., 2002; Vitek, 2002;
McIntyre et al, 2004). It has been shown that DBS in STN can suppress pathologically
synchronised oscillation in the vicinity of stimulation in patients with PD, (Eusebio A. et
al., 2011). Since the clinical effect from DBS is in some patients delayed for days or
even months (DBS in GPi for dystonia) and there are also carryover effects like slowly
returning hypokinesia after DBS cessation, presumably the stimulation leads also to
neuronal plasticity, network reorganization or affects the gene expression (Nowak K. et
al., 2011; Quartarone and Hallet, 2013).
After reviewing the stereotactically performed thalamic lesions by Hassler and
Dieckmann (1970), in 1999 Vandewalle and colleagues stereotactically implanted
quadripolar DBS electrodes in the thalamus of pharmacoresistant TS patients
(Vandewalle et al., 1999). Significant improvement of symptoms was reported. In 2002

Deep brain stimulation
13
DBS in the GPi was first time performed in order to treat tics in TS patients (Van der
Linden et al., 2002) and again considerable effect was reported. In 2007 Kuhn et al.
described a case report on a patient with TS and OCD, who received DBS in the NAC
and the internal capsule. The group reports significant reduction in both the frequency
and magnitude of the tics during the two and half year observation period (Kuhn J. et al.,
2007).
After these pioneering trials, a growing number of groups continued to work on the
development DBS as a therapy for refractory TS. Although most of the patients are
reported to benefit from surgery, a lot of future research is still needed, because so far
most of the data comes from case reports as the above-mentioned and open
uncontrolled trials (reviewed in Hariz and Robertson, 2010). The effectiveness of DBS
for TS, its safety, best indications and best stimulation targets/parameters remain to be
investigated.

Animal models for neuropsychiatric disorders
14
5 Animal models for neuropsychiatric disorders
To validate an animal model usually three main criteria are applied - face, predictive and
construct validity. The "face validity" requires similar behavioral features between the
animal model and the human clinical condition. The „predictive validity” is fulfilled when
clinically effective treatment has also a corresponding effect in the animal model of the
disorder, i.e. the model could be used to test and predict the potential efficacy of new
interventions or substances. The third criterion, the so called “construct validity” requires
the animal model to be consistent with what is known about the pathophysiology of the
disorder in humans.
The neuropsychiatric disorders are difficult to model in animals, because they are not
directly linked to certain neuropathological processes in the brain, usually typical human
features are affected and often the symptoms can be evaluated only by self report.
Therefore models for neuropsychiatric disorders use so called endophenotypes -
measurable physiological or behavioral markers, which are closely linked to the
biological origin of the symptoms. An endophenotype represent one aspect of the
complex pathophysiology of the disease and may be biochemical, endocrinological,
neuroanatomical, cognitive, neuropsychological etc. in nature. Heritability and stability
(state independency) are key components of any useful endophenotype (Gould and
Gottesman, 2006).
Deficient sensorimotor gating is found in patients with several neuropsychiatric disorders
- schizophrenia, OCD, HD, TS (Perry and Braff, 1994; Swerdlow et al., 1995, 2000,
2001; Castellanos et al., 1996; Karper et al., 1996; Miguel et al., 2000; Perry et al., 2001;
Braff et al., 2001) and can be used as an endophenotype to study their etiology,
pathophysiology and possible treatment srategies.
An operational measure of deficient sensorimotor gating is the so called PPI of ASR.
Startling stimuli (loud sound, air puff, bright light) are used to evoke reflexive startle
response in the test subject. Electromyography of musculus orbicularis oculi is used to
measure the blink reflex in humans, while in rats the whole body startle reaction is

Animal models for neuropsychiatric disorders
15
measured in a stabilimetric chamber. When the intense startling stimulus is shortly (up to
500ms) preceeded by a weaker, nonstartling prestimulus, the subsequent startle
response in healthy individuals is generally diminished. In patients and experimental
animals with disturbed sensorimotor gating, this reduction of the startle reaction is found
attenuated (Perry and Braff, 1994; Swerdlow et al., 1995, 2000, 2001; Castellanos et al.,
1996; Karper et al., 1996; Miguel et al., 2000; Perry et al., 2001; Braff et al., 2001).
The role of the “gating” period may be to protect the information in the weak initial
stimulus, so it can be normally processed regardless of the disruptive influence of the
stronger subsequent stimulus (Swerdlow et al., 2000). Studies in rats showed that PPI is
regulated by projections between the limbic cortex (including the temporal and medial
prefrontal cortex), the ventral striatum (NAC), the ventral pallidum and the
pedunculopontine tegmental nucleus (PPTg, Koch and Schnitzler, 1997; Swerdlow et
al., 1999; Swerdlow et al., 2000).
Fig. 4: Prepulse inhibition of the acoustic startle response.
A) Startle response to a loud sudden noise
B) Reduction of startle
reaction by a weak non-startling stimulus, preceding the startling one (prepulse inhibition)
C) No startle-reduction by the
prepulse (prepulse inhibition deficit)

Animal models for neuropsychiatric disorders
16
This limbic cortico-striato-pallido-pontine circuitry connects with the primary startle
circuitry (linking the auditory nerve with the spinal motor neurons) at the level of the
nucleus reticularis pontis caudalis (Koch et al., 1993; Swerdlow et al., 2000).
Studies show that the deficient sensorimotor gating is heritable and can also be found in
phenotypically healthy first-degree relatives of the affected individuals (Braff et al.,
2007). All these findings, together with the facts that PPI of ASR is a conserved
phenomenon among vertebrates and the PPI deficit improves from antipsychotics both
in animals and humans (Swerdlow et al., 1993), makes the sensorimotor gating
deficiency a promising candidate endophenotype to explore.
5.1 Pharmacological models for deficient PPI Dopaminergic, glutamatergic and serotonergic systems appear to play an important role
in the pathophysiology of the neuropsychiatric disorders with deficient processing of
sensorimotor information. Pharmacologically, PPI of ASR is reduced in rodent animal
models mainly via three approaches: stimulation of the D2 dopamine (DA) receptors,
blockade of N-methyl-D-aspartate (NMDA) receptors and activation of serotonergic (5-
HT) systems (Geyer et al., 2001).
The earliest and most frequently studied model of PPI deficiency is the dopaminergic
model. Systemic administrations of the direct DA receptor agonist apomorphine or the
indirect DA receptor agonist amphetamine lead to disruption of PPI in intact rats
(Mansbach et al., 1988). Same group also showed that this disruptive effect of the DA
agonists can be prevented by haloperidol injection, which is a standart medication for
TS. Additionally, in our group we have shown that excitotoxic lesions (Lütjens et al.,
2011) and DBS (Posch et al., 2012) of the EPN prevent PPI-deficits induced by the DA
receptor agonist apomorphine.
Kretschmer and Koch (2000) show that lesions in substantia nigra pars reticulata also
reduced PPI and prevented the enhancement of the ASR induced by amphetamine.
Swerdlow et al. (1990) show that 6-hydroxydopamine lesions that deplete dopamine
from the nucleus accumbens, olfactory tubercles and anterior striatum reversed the
disruption of PPI caused by amphetamine, but did not disrupt the amphetamine
potentiation of ASR. These findings suggest that increased mesolimbic DA activity is

Animal models for neuropsychiatric disorders
17
one of the substrates of the amphetamine-induced disruption of PPI. Same group
showed also that lesions of medial prefrontal cortex or ventral hippocampus in rats
increases their sensitivity to the PPI-disruptive effects of apomorphine, which is
consistent with the hypothesis that cell damage in frontal and temporal cortex increases
the sensitivity to the sensorimotor gating-disruptive effects of dopamine receptor
activation (Swerdlow et al., 1995; 2000).
Experiments with selctive DA, D1 and D2 receptor agonists show that PPI is primarily
disrupted by activation of the D2 receptors and not by the D1, but nevertheless, it seems
that there is synergism between them (Peng et al., 1990; Geyer et al., 2001).
NMDA receptor antagonists such as phencyclidine and dizocilpine (MK-801) are also
reported to disrupt PPI robustly in rats (Mansbach and Geyer, 1989). Like the
amphetamine, they can also induce schizotypal symptoms in humans. At the same time,
there is controversial data about the effect of ketamine on PPI including even a study
reporting an increased PPI in humans after ketamine application (Duncan et al., 2001;
Geyer et al, 2001). These data compromises the predictive validity of the glutamatergic
PPI disruption model.
Nonselective indirect 5-HT agonists like fenfluramine, methylendioxyethylamphetamine
and methylenedioxymethamphetamine, and selective 5-HT agonists, such as
dimethoxyiodoamphetamine were shown to disrupt PPI in rats (Mansbach et al., 1989;
Sipes and Geyer, 1994). Nevertheless, since amphetamine in humans appears to
increase PPI, rather than disrupting it as in rats, the predictive validity of the serotonergic
model of PPI deficiency is also compromised (Vollenweider et al., 1999; Geyer et al.,
2001).
Pharmacologically induced PPI deficit is only temporary and usually limited to a special
transmitter system. These models only imitate the genetically determined, much more
complex deficits in humans. Therefore, non-pharmacological models can be an option to
avoid these disadvantages.

Animal models for neuropsychiatric disorders
18
5.2 Developmental models for deficient PPI There is general agreement that external factors, such as prenatal infections, play an
important role in the development and course of many neuropsychiatric disorders,
including schizophrenia, TS and OCD (Adams et al., 1993; Brown et al., 2004; Martino
et al., 2009). Developmental models are therefore introduced to study environmental
influences on neuroanatomy and behavior.
Peripheral administration of bacterial endotoxin lipopolysaccharide to pregnant female
rats leads to reduced PPI in the offspring, which can be reversed by the antipsychotics
haloperidol and clozapine (Borrell et al., 2002). In another model, neonatal excitotoxic
hippocampal lesions with Ibotenic acid, leads to postpubertal deficient PPI related to
excessive DA transmission in the mesolimbic/nigrostriatal systems (Lipska et al., 1995).
One other study shows that after brief interruption of brain cellular proliferation with
methylazoxymethanol during late gestation at embryonic day 17, the litters show PPI
deficits at adulthood, as well as hyperactivity, social interaction deficits, spatial working
memory disturbance and may be used as a model for schizophrenia (Le Pen et al.,
2006).
Another developmental approach is isolation rearing. Rats reared in isolation from the
time of weaning exhibit deficits in PPI compared to socially reared rats, when both
groups are tested in adulthood. DA antagonist raclopride normalizes the PPI deficiency
(Geyer et al., 1993).
Klein et al. (2013) studied the effect of high frequency DBS in two other developmental
models, characterizied with reduced PPI - the maternal immune stimulation with poly I:C
and the pubertal cannabinoid (WIN) administration rat model. They found that DBS in
the medial prefrontal cortex and dorsomedial thalamus normalizes the PPI deficits
induced in both of the models.
5.3 Genetic models for deficient PPI Mutant mouse models with PPI deficiency have been based primarily on the known
effects of antipsychotic drugs and deletion of genes that encode different
neurotransmitter systems.

Animal models for neuropsychiatric disorders
19
Dopamine transporter knock-out mice DAT (−/−) show disrupted PPI, which is reversed
significantly by pretreatment with the preferential D2 receptor antagonist raclopride, but
not by the selective D1 receptor antagonist SCH23390, which also shows the important
role of the D2 receptors in the dopaminergic modulation of PPI (Ralph et al., 2001).
Except involvement of the dopaminergic system, glutamate system dysfunction is also a
possible cause of PPI deficite. Transgenic hypomorph mice NRG+/- that express 50% of
normal levels of the NR1 subunit common to all NMDA receptor isoforms (NR1 null is
lethal), display increased motor activity and deficient PPI (Harrison and Weinberger,
2005).
The phosphatase calcineurin plays an important role in the central nervous system and
calcineurin signaling is thought to be implicated in schizophrenia pathogenesis.
Forebrain-specific calcineurin knockout mice have been generated and these animals
showed increased locomotor activity, decreased social interaction, impaired PPI
(Miyakawa et al., 2003).
Genetically modified mice were furthermore developed for a variety of other transmitter
systems, risk genes and other factors leading to hyperlocomotion and disturbed PPI.
Despite being a powerful tool for research, these models have also their disadvantages.
The main limitation of the genetic mouse models is that they do not reproduce the broad
pathology of the human diseases, which are complex and involve the interaction of
many genes and inveroinmental factors. They also show either no or only a modest
behavioral phenotype.
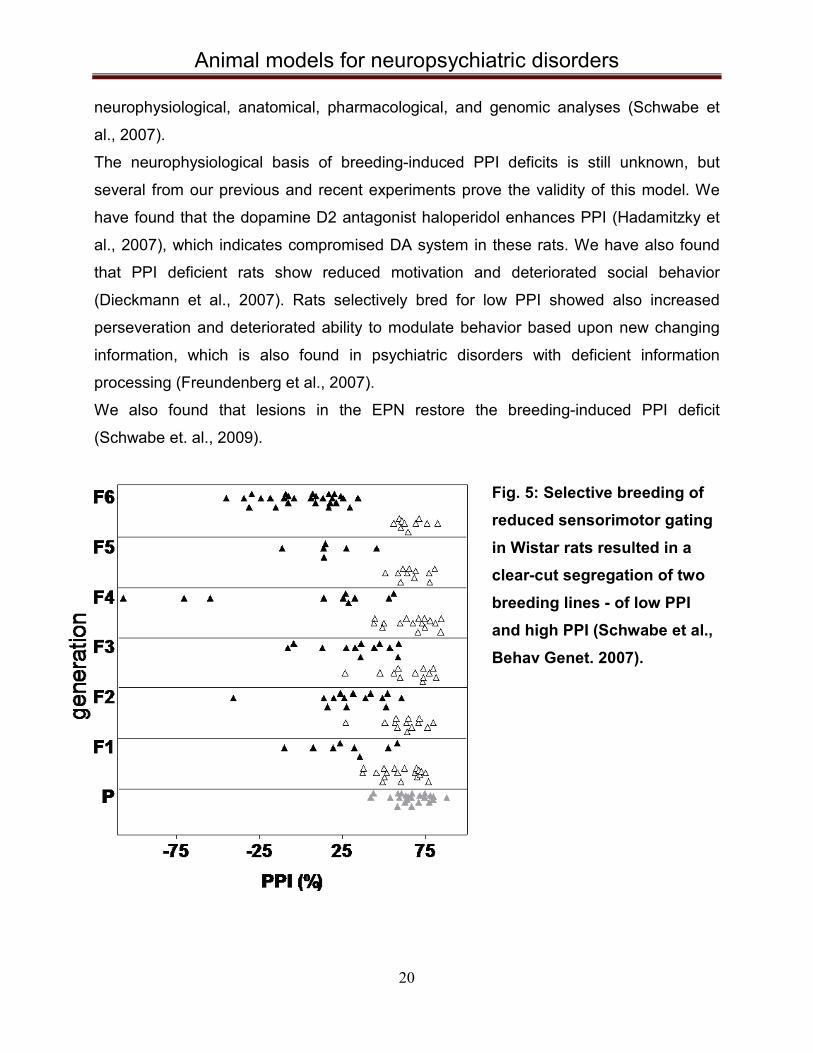
5.4 Selective breeding for deficient PPI In our group we have selectively bred Wistar rats for high (PPI high) and low (PPI low)
expression of PPI for 19 generations by now. Initially, 23 male and 27 female rats were
tested and two animals with the highest or lowest PPI of each sex were used for
breeding (Schwabe et al., 2007). Same selection was continued in the next filial
generations, which resulted in two clearly segregated groups of animals, one with high
and another one with low PPI of ASR, after the 6th generation.
The stable phenotype of breeding-induced PPI-deficits indicates that PPI has strong
genetic determinants and that selectively bred rats can be used for future
Animal models for neuropsychiatric disorders
20
neurophysiological, anatomical, pharmacological, and genomic analyses (Schwabe et
al., 2007).
The neurophysiological basis of breeding-induced PPI deficits is still unknown, but
several from our previous and recent experiments prove the validity of this model. We
have found that the dopamine D2 antagonist haloperidol enhances PPI (Hadamitzky et
al., 2007), which indicates compromised DA system in these rats. We have also found
that PPI deficient rats show reduced motivation and deteriorated social behavior
(Dieckmann et al., 2007). Rats selectively bred for low PPI showed also increased
perseveration and deteriorated ability to modulate behavior based upon new changing
information, which is also found in psychiatric disorders with deficient information
processing (Freundenberg et al., 2007).
We also found that lesions in the EPN restore the breeding-induced PPI deficit
(Schwabe et. al., 2009).
Fig. 5: Selective breeding of reduced sensorimotor gating in Wistar rats resulted in a clear-cut segregation of two breeding lines - of low PPI and high PPI (Schwabe et al., Behav Genet. 2007).

Animal models for neuropsychiatric disorders
21
In this model the PPI deficit is chronic, so there is no need to repeatedly induce it like in
the pharmacological models. The rats can be used for behavioral studies and also being
an inbred model, they have lower genetic variability, which makes the results more
consistent.

Objectives
22
6 Objectives
The neuronal circuitries that regulate PPI, appear to overlap with those implicated in the
pathophysiology of TS. EPN, CM-Pf and NAC play important roles in these networks
and are also found to be effective DBS targets for TS patients. Therefore, our aim was to
investigate the effect of chronic DBS in these three nuclei on our endophenotype model
of deficient PPI. As a second goal we wanted to examine the neuronal activity in the
NAC, the EPN and the mPFC of the PPI low rats in comparison with the PPI high group.

Manuscript one
23
7 Manuscript one
Title:
Effect of deep brain stimulation in rats selectively bred for reduced prepulse inhibition
Order of Authors:
Svilen Angelov, Carolin Dietrich, Joachim K Krauss, Kerstin Schwabe
Contribution:
Authors Angelov and Schwabe designed the study and wrote the protocol. Stimulation of
the entopeduncular nucleus i.e. the first part of the project, was performed by author
Dietrich, while all the other experiments - by author Angelov. Authors Schwabe and
Angelov undertook the statistical analysis of the data. Author Angelov wrote the first
draft of the manuscript. All authors contributed to and have approved the final version of
the manuscript. Critical revision was done by authors Schwabe and Krauss.

Manuscript one
24
Brain Stimul. 2014 Apr 4. pii: S1935-861X(14)00128-4. doi: 10.1016/j.brs.2014.03.013.
Effect of Deep Brain Stimulation in Rats Selectively Bred for Reduced Prepulse Inhibition Svilen D. Angelov, Carolin Dietrich, Joachim K. Krauss, Kerstin Schwabe Department of Neurosurgery, Medical University Hannover, Carl-Neuberg-Str. 1, D-30625 Hannover, Germany
Abstract
BACKGROUND: Sensorimotor gating, measured as prepulse inhibition (PPI) of the acoustic startle reaction (ASR), is disturbed in certain neuropsychiatric disorders, such as schizophrenia, obsessive compulsive disorder, and Tourette's syndrome (TS). Deep brain stimulation (DBS) of the centromedian-parafascicular complex (CM-Pf), globus pallidus internus (in rats the entopeduncular nucleus - EPN), and the ventral striatum (in rats the nucleus accumbens - NAC) has been used for treatment in TS.
OBJECTIVE: We tested whether DBS of these regions would alleviate breeding-induced low PPI in rats.
METHODS: Rats with breeding-induced low and high PPI were bilaterally implanted with electrodes in the CM-Pf, the EPN, or the NAC. After two weeks, they were stimulated or sham stimulated for epochs of 6 days (in the EPN with a current of 20% below the individual threshold for stimulation-induced side effects, in the NAC or CM-Pf with 100 μA and 150 μA). On the 6th day the rats were tested for PPI of ASR.
RESULTS: Stimulation in the CM-Pf with 150 μA significantly alleviated PPI, while NAC stimulation was less effective. In PPI low rats electrode implantation in the EPN already improved PPI, while subsequent stimulation had no additional effect. Startle reaction of PPI low rats was not affected by stimulation of either region.
CONCLUSION: The CM-Pf and the EPN are important for the modulation of sensorimotor gating in rats with breeding-induced low PPI. These rats may therefore be useful to further investigate the pathophysiological mechanisms of deficient sensorimotor gating and also mechanisms of action of DBS in these circumstances.
Copyright © 2014 Elsevier Inc. All rights reserved.
KEYWORDS: Centromedian parafascicular nucleus, Entopeduncular nucleus, Globus pallidus internus, Sensorimotor gating, Tourette's syndrome

Manuscript two
25
8 Manuscript two
Title:
Neuronal activity of the prefrontal cortex is reduced in rats selectively bred for deficient
sensorimotor gating
Order of Authors:
Mesbah Alam, Svilen Angelov, Meike Stemmler, Christof von Wrangel, Joachim K
Krauss, Kerstin Schwabe
Contribution:
Authors Alam and Schwabe designed the study, wrote the protocol and managed the
literature searches and analyses. Acquisition of data was performed by authors Alam,
Stemmler and Angelov. Author Alam undertook the statistical analysis, and author
Schwabe wrote the first draft of the manuscript. All authors contributed to and have
approved the final manuscript. Critical revision of the manuscript for important
intellectual content was done by authors Schwabe and Krauss.

Manuscript two
26
Neuronal activity of the prefrontal cortex is reduced in rats selectively bred for deficient sensorimotor gating Mesbah Alam, Svilen Angelov, Meike Stemmler, Christof von Wrangel, Joachim K.
Krauss, Kerstin Schwabe
Department of Neurosurgery, Hannover Medical School, Carl-Neuberg-Str.1, D-
30625 Hannover, Germany
This work was supported by the Tourette Syndrome Association.
Abstract Rats selectively bred for deficient prepulse inhibition (PPI), an operant measure of
sensorimotor gating, may be used to study certain pathophysiological mechanisms and
therapeutic strategies for neuropsychiatric disorders with abnormalities in information
processing, such as schizophrenia and Tourette`s syndrome (TS). The medial prefrontal
cortex (mPFC) and the nucleus accumbens (NAC) are involved in the modulation of PPI.
Additionally, lesions of the entopeduncular nucleus (EPN) alleviated PPI in rats with
breeding-induced low PPI. We here examined the neuronal activity in these regions.
Male rats with breeding-induced high and low expression of PPI (n=7, each) were
anesthetized with urethane (1.4 g/kg). Single-unit (SU) activity and local field potentials
(LFPs) were recorded in the mPFC, the NAC and in the EPN. In the mPFC discharge
rate, measures of irregularity and burst activity were significantly reduced in PPI low
compared to PPI high rats (p<0.05), while analysis in the NAC showed approximately
inverse behavior. In the EPN no difference between groups was found. Additionally, the
oscillatory theta band activity (4-8 Hz) was enhanced and the beta band (13-30 Hz) and
gamma band (30-100 Hz) activity was reduced in the NAC in PPI low rats.
Reduced neuronal activity in the mPFC and enhanced activity in the NAC of PPI low
rats, together with altered oscillatory behavior are clearly associated with reduced PPI.

Manuscript two
27
PPI low rats may be used to study the pathophysiology and therapeutic strategies for
neuropsychiatric disorders accompanied by deficient sensorimotor gating.
Keywords: prepulse inhibition, nucleus accumbens, entopeduncular nucleus, local
field potentials, neuropsychiatric disorders
Objectives of the study Neuropsychiatric disorders are increasingly recognized as circuit disorders.
Understanding the disturbances in the firing patterns and synchrony of neuronal activity
throughout cortico-subcortical loops would be useful to develop and improve
therapeutics to attenuate such pathological processes (Carlson et al., 2006; Kopell &
Greenberg, 2008).
Sensorimotor gating mechanisms, which allow the nervous system to suppress or ”gate”
responding to most external stimuli and internally generated signals or impulses, are
disturbed in certain neuropsychiatric disorders (Swerdlow & Geyer, 1998; Braff et al.,
2001). Such gating mechanisms have been operationalized in measures of prepulse
inhibition (PPI) of the acoustic startle response (ASR; Koch et al., 2000; Swerdlow et al.,
2001). Deficient PPI has been demonstrated in Tourette’s syndrome (TS) and
schizophrenia (Braff et al., 2001; Swerdlow & Sutherland, 2006), and experimentally-
induced PPI deficits in rodents are used as an endophenotype for these disorders
(Cadenhead et al., 2002; Braff & Light, 2005). Selective breeding in Wistar rats for high
and low PPI leads to a segregation of two rat lines with significantly different PPI
(Schwabe et al., 2007). The antipsychotic dopamine (DA) receptor antagonist
haloperidol alleviated the breeding-induced PPI-deficit (Hadamitzky et al., 2007).
Additionally, behavioral deficits in PPI low rats corroborate clinical findings of a number
of neuropsychiatric disorders (Dieckmann et al., 2007; Freudenberg et al., 2007).
Within the neuronal circuitry that regulates PPI, the medial prefrontal cortex (mPFC) and
the NAC play key roles (Swerdlow et al., 2001; Pothuizen et al., 2005). Lesions or deep
brain stimulation of the entopeduncular nucleus (EPN), i.e., the equivalent to the human
GPi, alleviate breeding- or apomorphine-induced deficient PPI in rats, indicating that
dysfunction of neuronal activity within this region may be important (Schwabe et al.,

Manuscript two
28
2009; Lütjens et al., 2011; Posch et al., 2012). Notably, deep brain stimulation of the GPi
is clinically used to improve tics in TS (Houeto et al., 2005; Shaded et al., 2007; Servello
et al., 2008). We here examined the spontaneous neuronal activity in the mPFC, the
NAC and the EPN of PPI high and low rats.
Material and Methods Subjects Rats with either breeding-induced reduced (PPI low) or increased PPI (PPI high) were
housed in groups of four in standard Macrolon Type IVS cages (Techniplast,
Hohenpeissenberg, Germany) under a 14-h light/10-h dark cycle (on at 07:00 h) at a
room temperature of 22 ± 2 °C, with food and water ad libitum. All experiments were
carried out in accordance with the EU Directive 2010/63/EU and were approved by the
local animal ethic committee.
Breeding selection for PPI high or low The parental generation for our PPI high and low lines consisted of 23 male and 27
female rats (outbred adult Hannover-strain Wistar rats from Harlan-Winkelmann,
Borchen, Germany). The TSE Startle Response SystemTM (Bad Homburg, Germany)
was used to test rats for PPI, i.e., the percent decrease of the startle response in pulse-
alone (20 ms white noise pulse at 105 dB sound pressure level (SPL)) compared to the
startle response in prepulse-pulse trials (80 dB SPL, 10 kHz pure tone pulse, 20 ms
duration followed by pulse 100 ms after prepulse onset). Two females and males with
the highest and the lowest level of PPI, respectively, were chosen for selective breeding
of two lines with either high or low level of PPI. After the 11th generation the startle
response system of San Diego instruments was used for testing of PPI as described
before, but with 68 dB white noise as prepulse. For this study rats from the 11th and
12th generation were used, i.e., seven PPI low rats with a mean PPI of 8.38 % and a
mean ASR of 1945 arbitrary units (AU), and seven PPI high rats with a mean PPI of 66.7
% and a mean ASR of 1762 AU.

Manuscript two
29
Single-unit and local field potential recording procedures Rats were anaesthetized with urethane (1.4 g/kg, i.p. ethyl carbamate, Sigma; with
additional doses as needed, depth of anaesthesia was checked by the foot pinch). Body
temperature was kept at 37.5 ± 0.5°C with a heating pad. Electrocardiographic (ECG)
activity was monitored constantly to ensure the animals’ well-being. Rats were placed in
a stereotaxic frame and craniotomies were made over the target coordinates, relative to
bregma (flat skull position). For all regions we used two trajectories within the following
coordinates in millimeter scale; for the mPFC: anterior-posterior (AP), +3.2 and +2.2;
mediolateral (ML), ±0.5 and ±0.8; ventral (V), - 3.2 and 4.5; for the NAC: AP +1.7 and
+1.2, lateral ±1.5 and ±1.7, V 6.5 and 7.8, and for the EPN: AP -2.3 and -2.8, lateral ±2.4
and ±2.6, V 7.4 and 8.0. At the end of all recordings the electrode tip was used to
coagulate the tissue along each of the trajectories in 200 μm steps with bipolar current
of 10 μA for 10 s to verify recordings in the targets after sacrifice of the animals (Fig. 1).
A single microelectrode for extracellular recordings (quartz coated electrode pulled with
a ground platinum-tungsten alloy core (95%–5%), diameter 80 μm, impedance 1–2 MΩ)
was connected to the Mini Matrix 2 channel version drives headstage (Thomas
Recording, Germany). The electrode was guided stereotactically through the guide
cannula towards the target coordinates in the mPFC, NAC or EPN, respectively. The
microelectrode signal was passed through a headstage with unit gain and then split to
separately extract the single unit (SU) and the local field potentials (LFPs) components.
For SU recording signals were bandpass-filtered between 500 and 5000 Hz and
amplified from × 9,500 to19,000. The LFP signals were filtered to pass frequencies
between 0.5 and 140 Hz, before being amplified and digitized at 1 kHz. Data were
acquired using the CED 1401 A/D interface (Cambridge Electronic Design, Cambridge,
UK).
Data and statistics analysis Action potentials arising from a single neuron were discriminated by the
templatematching function of the spike-sorting software (Spike2; Cambridge Electronic
Design, Cambridge, UK). For analyses of spontaneous activity, one 300 s epoch of
simultaneously recorded spiking and LFP activities that was free of artefacts was used

Manuscript two
30
from every recorded neuron. Data are represented as means ± standard error of the
means (SEM). Data was analysed between PPI high and low rats by Mann-Whitney U
test with a p<0.05 representing significance.
Firing rates and burst detection
The firing rate was calculated with the firing rate histograms produced in NeuroExplorer
version 4 (NEX Technologies, NC). The burst activity (burst in percentage) of the
neurons, percentage of spikes in bursts and mean spikes in bursts were analysed by the
criteria of surprise methods.
Asymmetry index
Variations to the Gaussian distribution were evaluated by determining the asymmetry
index, which is the ratio of the mode to the mean ISI. An asymmetry index close to 1
reveals a relatively regular firing pattern, whereas the more the index differs from unity,
the more irregular the spike trains. A ratio of less than 1 reflects an asymmetrical shape,
indicating a larger fraction of short interspike intervals (positively skewed), as is
expected when there is bursting activity.
Complexity analysis
The non-linear signal-processing metric cLZ was used to evaluate occurrence and
recurrence of patterns along electrophysiological data series. This index quantifies
dynamic features of time series especially for highly correlated and short sequences
(Lesne et al., 2009). The algorithm to compute cLZ is explained in detail in Hu et al,
(2006). In the present paper the spike trains were binned by using a 1 millisecond bin
size. The number 0 or 1 is assigned to each bin, according to whether it contains no
spike (0) or at least one spike (1) (Szczepański et al., 2004). The binary representation
of electrophysiological data was finally analyzed for cLZ by using the original algorithm
proposed by Lempel and Ziv (1976). The resulting cLZ was then normalized to the
length of spike train, to overcome known complexity dependence from block size; as a
rule of thumb, the higher the complexity the more irregular and higher the firing patterns.

Manuscript two
31
cLZ calculations were performed by in-house developed procedures using C++ and Mat
lab software.
Regularity of spikes events
To determine three distinct patterns: regular, irregular and bursting activity we used a
modified approach of the discharge density histogram of Kaneoke et al., (1996). The
estimated density histograms d (λ) were compared to reference probability density
function px(λ) by means of a mathematical distance. The discharge density of a regular
neuron is expected to follow a Gaussian distribution PG(λ) with mean equal to 1 and
variance equal to 0.5. An irregular neuron is expected to follow a Poisson distribution
PP(1)(λ) with mean equal to 1, a bursting neuron with mean equal to 0.2. Because of the
reduced number of samples forming d(λ) and the possibility to take values very close to
zero, we chose the 2-norm distance to determine the goodness-of-fit. The distribution
with the smallest distance determines to which class the neuron is assigned (Labarre et
al., 2008). Differences in the incidence of discharge patterns were evaluated by Chi2-
Test.
Spike train spectral analysis
The modulating function of neuronal spike trains and the different spectral bands of
oscillatory activities in the spike trains are important to understand the characteristics of
neuronal activity. Oscillatory activity was quantified by using the local spike shuffling
method. The ISI local shuffling is computed with confidence limits that are based on the
first-order statistics of the spike trains, thus providing a reliable estimation of auto- and
cross-spectra of spike trains (Rivlin-Etzion et al., 2006). Peaks above the significance
level (P=0.05) were considered to represent oscillatory activity. The percentages of
neurons showing a peak over the significance level as defined within standard frequency
bands (0.1-4, 4-8, 7-12, 12-30, 30-100 Hz) were determined and considered as ”ON”
oscillatory modulating spikes if the power spectrum had one or more significant peaks
between 0.1-100 Hz. When neuronal oscillatory modulating spikes had no such
significant peaks, they were termed as non-oscillatory firing, i.e., ”OFF”.

Manuscript two
32
Spectral power analysis of local field potentials
For analysis of LFPs, each recording segment was detrended to remove any slow DC
components and padded with zeros to increase frequency resolution. Spectra were
determined for the total 300 s recordings and the signal notch (50 Hz) and low pass
filtered (100 Hz). Autospectra of LFPs was derived by discrete Fourier transformation
with 1024 blocks. The relative power of epochs of 300 s, from the same recording
section like the spike trains, was analyzed (θ=4-8 Hz, α=8-13 Hz, β=13-30 Hz, γ = 30-
100 Hz), averaged, and compared between PPI high and PPI low rats.
Results Spontaneous neuronal activity in the mPFC, the NAC and the EPN were analysed after
histological verification of recording sites in the different regions (see Fig. 1).
The total number of neurons in PPI high rats in the mPFC was n=58, in the NAC n=67,
and in the EPN n=38. In the PPI low rats the number of neurons in the mPFC was n=84,
in the NAC n=60, and in the EPN n=47.
Manuscript two
33
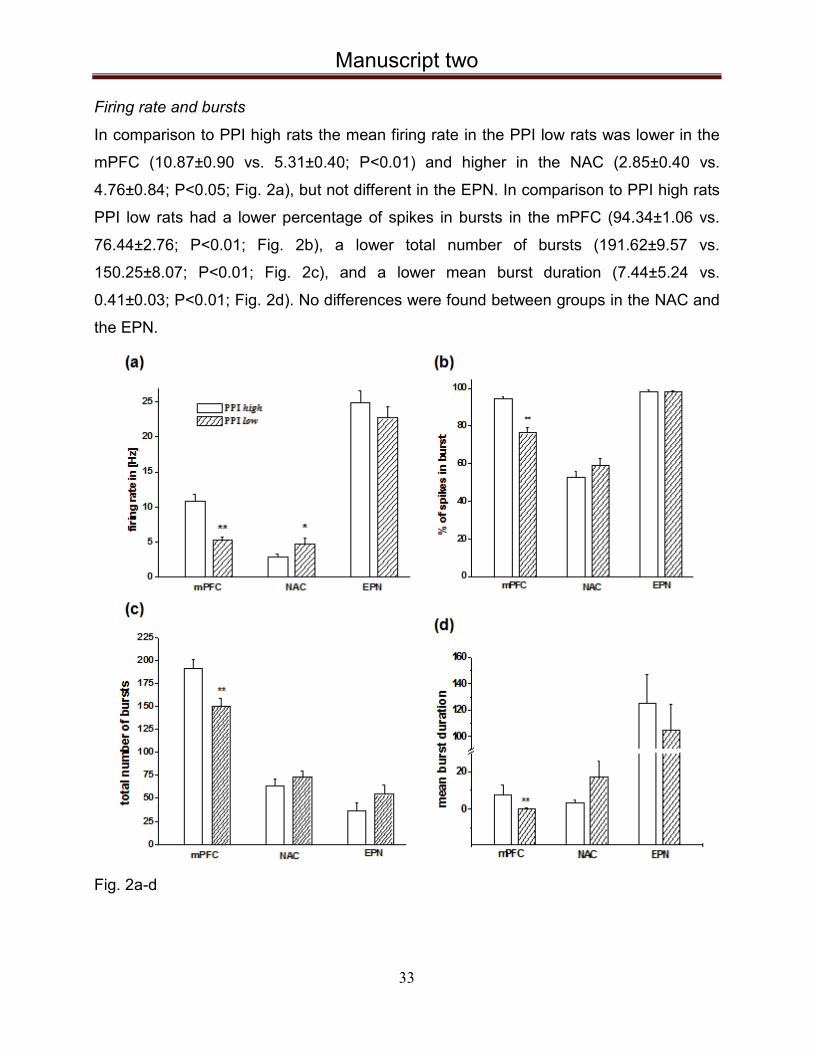
Firing rate and bursts
In comparison to PPI high rats the mean firing rate in the PPI low rats was lower in the
mPFC (10.87±0.90 vs. 5.31±0.40; P<0.01) and higher in the NAC (2.85±0.40 vs.
4.76±0.84; P<0.05; Fig. 2a), but not different in the EPN. In comparison to PPI high rats
PPI low rats had a lower percentage of spikes in bursts in the mPFC (94.34±1.06 vs.
76.44±2.76; P<0.01; Fig. 2b), a lower total number of bursts (191.62±9.57 vs.
150.25±8.07; P<0.01; Fig. 2c), and a lower mean burst duration (7.44±5.24 vs.
0.41±0.03; P<0.01; Fig. 2d). No differences were found between groups in the NAC and
the EPN.
Fig. 2a-d

Manuscript two
34
Asymmetry index
In the mPFC the asymmetry index was enhanced in PPI high rats as compared to PPI
low rats (0.32±0.03 vs. 0.124±0.01; P<0.01), which is compatible with the reduced burst
activity in PPI low rats. In contrast, in the NAC the asymmetry index was reduced in PPI
high as compared to PPI low rats (0.11±0.01 vs. 0.16±0.02; P<0.05), which indicates
higher burst behavior of PPI low rats in this region (Fig. 2e).
Estimation of complexity
The averaged cLZ complexity estimation showed low complexity values in the PPI low
rats in the mPFC and high complexity values in the NAC as compared to PPI high rats
(0.04±0.02 vs. 0.07±0.01; P<0.01 and 0.04±0.01; P<0.01). In the EPN, no difference
was found between groups (Fig. 2f).
Fig. 2e-f
Firing patterns
Examples of regular, irregular and bursty firing patterns are shown in Fig. 3a-c. The
percentage of firing patterns in the mPFC, the NAC and the EPN for PPI high and low
groups are shown in Fig. 3d. Analyses showed a lower percentage of irregular spikes
and a higher percentage of burst patterns in the PPI low rats as compared to PPI high
rats in the mPFC (74% vs. 84% and 26% vs. 16%), which did not reach the level of
significance. No regular firing pattern was observed in the mPFC in both groups. In the

Manuscript two
35
NAC the percentage of irregular firing of PPI high and low rats was 85% vs. 88%, of
burst firing 7% vs. 11%, and of regular firing 8% vs. 3%, which was not statistically
significant. Further, in the EPN of PPI high rats the percentage of irregular and burst
firing was enhanced (59% vs. 15% and 12% vs. 8%, respectively), while the regular
pattern was reduced (29% vs.77%; all P<0.05; Chi-square test).
Fig. 3a-d

Manuscript two
36
Spike train spectral analysis
In the mPFC the average percentage of spikes involved in oscillatory modulatory ON
neurons were 7.14% in the delta band of PPI high rats, while PPI low rats showed only
1.28% ON neurons in gamma oscillatory range. In the NAC the total ON neurons were
6.06% in PPI high animals, while in PPI low rats only 1.82% ON neurons were found in
the beta band. In the EPN the total percentage of modulatory activity was higher in both
groups of animals as compared to mPFC and NAC. The total ON neurons of PPI high
rats in the EPN was 23.53% and 31.71% in PPI low rats respectively (Table 1).
Table 1
Local field potentials
Theta (4–8 Hz): The relative power of theta bands in the NAC and EPN were higher in
PPI low as compared to PPI high rats (P<0.01 and P<0.01, respectively; Fig. 4a), while
in the mPFC no difference was observed between groups.
Alpha (8–13 Hz): In the PPI low rats in the EPN the percentage of relative power of
alpha band activity was lower as compared to PPI high rats (P<0.01; Fig. 4b), while no
difference was observed in the mPFC or in the NAC in both groups.
Beta (13–30 Hz): In PPI low rats the percentage of relative power of beta band activity
was decreased in all regions (mPFC: P<0.05, NAC: P<0.01 and EPN: P<0.01; 4c).

Manuscript two
37
Gamma (30–100 Hz): The relative power of gamma band activity was decreased in the
PPI low rats in the NAC (P<0.01; Fig. 4d), while no difference was found in the mPFC
and EPN (P=0.55 and P=0.64) respectively.
Fig. 4a-d

Manuscript two
38
Discussion Our data sheds new light on the neuronal activity in the basal ganglia-cortex interplay in
disturbed sensorimotor gating. In PPI low rats the firing rate and the burst behavior of
mPFC neurons is reduced as compared to PPI high rats. This is corroborated by a
reduced asymmetry index, which also indicates reduced burst behavior in PPI low rats,
as well as a reduced cLZ, which indicates less functional complexity and dynamic
features in the firing pattern of mPFC neurons.
The PFC cortex is involved in higher-order executive tasks, such as learning, working
memory, and behavioral flexibility. In humans, dysfunction of prefrontal cortical areas,
especially its dorsolateral part, contributes to a decline in cognitive performance in
neuropsychiatric disorders (Heidbreder & Groenewegen, 2003). In patients with
schizophrenia 'hypofrontality' has been reported (Sabri et al., 1997). Also, functional
brain imaging found reduced perfusion within the dorsolateral PFC and the anterior
cingulate cortex in TS patients (Moriarty et al., 1995). Although the question, whether
rodents have a region analogous to the primate dorsolateral PFC is still disputed,
anatomical, electrophysiological and behavioral evidence supports the view that the
rat medial PFC, although at a rudimentary level, combines elements of the primate
anterior cingulate cortex and dorsolateral PFC. The rat prelimbic PFC, which has been
targeted for electrophysiological recordings in the present study, is regarded a
developmental homologue to the dorsolateral region of the primate PFC, although
anatomically more closely related to the primate medial PFC (Heidbreder &
Groenewegen, 2003; Seamans et al., 2008).
Overall, the firing behavior of NAC neurons of PPI low rats was quite the opposite of that
found in the mPFC, which may reflect a dysregulation in the mPFC feedback loop in the
NAC. In the NAC of PPI low rats the firing rate was increased, together with enhanced
asymmetry index and cLZ estimation, indicating more regular or tonic activity, and a
more complex firing pattern as compared to that of PPI high rats (Chen et al., 2011).
Several neuroanatomical and neurophysiological studies suggest a functional
relationship between the mPFC and the NAC (Tzschentke, 2001; Sesack et al., 2003).
Experimental manipulations that decrease mPFC DA ”tone” in rats lead to deficient PPI,
probably via disinhibition of descending glutamatergic fibers that causes increased NAC

Manuscript two
39
DA transmission (Koch & Bubser, 1994; Ellenbroek et al., 1996). However, although this
well-known relationship between cortical and subcortical DA seems to be an attractive
potential explanation of the decreased activity in the PFC and the increased activity in
the NAC, it refers to the DA inputs, not the output neurons, which mainly have inhibitory
D2 receptors. An increased DA tone in the NAC should thus decrease firing in the NAC.
Nevertheless, how DA contributes to information processing within the NAC is still
debated since the actions of DA depend on a complex interplay of cellular and synaptic
properties. It has been reported that DA can either excite of inhibit NAC neurons (Nicola
et al., 2000; Bennay et al., 2004).
On the other hand, although disordered dopaminergic signalling in the striatum and
NAC, together with abnormal neuronal activation through basal ganglia structures, are
thought to be critical determinants in several neuropsychiatric disorders, including TS
and schizophrenia (Albin et al., 2003; Wang et al., 2011), there is also clinical and
preclinical evidence that other transmitter systems, e.g., noradrenergic transmission, are
involved (cf. Leckmann et al., 2010; Swerdlow et al., 2012). Notably, although
haloperidol restored the breeding-induced PPI deficit to a level similar to that of the PPI
high group, indicating disturbed DA function in these rats (Hadamitzky et al.,2007), the
atypical antipsychotics clozapine and risperidone had similar, albeit nonsignificant
effects, suggesting that other transmitter systems may be affected as well. The NAC
exerts its modulating effect on PPI by way of a projection from the ventral pallidum to the
pedunculopontine tegmental nucleus (PPTg), which modulates the primary startle
circuitry in brainstem nuclei (Kretschmer & Koch, 1998; Alam et al., 2011). Additionally,
the NAC projects to the EPN, which is one of the output regions of the BG circuitries that
also projects to the PPTg (Groenewegen & Russchen, 1984; Zahm & Heimer, 1993;
Alam et al., 2011). The EPN is considered the equivalent to the human GPi, which has
been used for functional neurosurgery to treat movement disorders, e.g., Parkinson’s
disease and dystonia (Krauss et al., 2004; Andrade et al., 2009; Fasano et al., 2012),
and which is currently under investigation for neurosurgical treatment of TS (Houeto et
al., 2005; Shahed et al., 2007). We recently showed that lesions or DBS of the EPN
alleviate PPI in PPI low rats and counteract deficient PPI induced by the dopamine
receptor agonist apomorphine (Schwabe et al., 2009; Lütjens et al., 2011; Posch et al.,

Manuscript two
40
2012). Unanticipated with regard to these findings, in the present study the firing rate
and burst features were unchanged in the EPN. With that regard, however, it should be
noted that the major projection from the NAC is to the ventral pallidum, while the major
input into the EPN comes from the dorsal striatum and may thus be part of a different
circuitry than the mPFC and the NAC.
Oscillatory activity in the LFPs is regarded essential for information transfer and the
binding of electrical signals from distant structures (Uhlhaas & Singer, 2010). In PPI low
rats we found an enhanced level of theta band oscillatory activity in the NAC and EPN,
which was accompanied by reduced beta band activity in both regions and reduced
alpha band activity in the EPN and reduced gamma band activity in the NAC. In the PFC
only beta band activity was reduced in PPI low rats. In human studies, current evidence
points to a crucial role for altered neural oscillatory activity and synchrony in the
pathophysiology of neuropsychiatric disorders. Relative to healthy controls, in patients
with schizophrenia gamma oscillations were markedly reduced after transcranial
magnetic stimulation, particularly over fronto-central regions and the right temporal lobe,
which was accompanied by abnormal high resting-state theta activity (Uhlhaas & Singer,
2010; Hanslmayr et al., 2012). Reductions of beta and gamma oscillations and their
synchronization have also been demonstrated during cognitive tasks (Spencer et al.,
2003; Whittington, 2008). Additionally, it has been shown in TS that LFP spectra in
certain regions of the thalamus in TS patients have a prominent oscillatory activity at low
frequencies and in the alpha band, while beta activity was virtually absent (Marceglia et
al., 2010). Similar findings were recently demonstrated in the GPi of TS patients (Kühn
et al. 2011).
With regard to decreased beta band activity in the PPI low rats, it is conceivable that it
might be an epiphenomen of overflow of high dopamine concentration in the brain. Both
animal and clinical studies have shown that the changes in beta activity are supported
by net dopamine levels in the cortico-basal ganglia loop, since DA antagonists increase
beta activity and dopamine agonists decrease beta activity. In line with this, increased
beta activity has been related to hypokinetic symptoms in PD (Hammond et al., 2007;
Dejean et al., 2009).

Manuscript two
41
With regard to oscillatory activity of spike trains, mPFC neurons in PPI low rats showed
no slow oscillatory modulatory activity in the delta and theta band range, while neurons
in PPI high rats showed modulatory activity in these ranges, giving further evidence for
hypofrontality. Additionally, in the NAC modulatory activity of spike trains was enhanced
in PPI low rats, which is in line with the enhanced neuronal postsynaptic activity in this
region. Similar to the results of single unit activity in the EPN, no obvious differences
were observed between PPI high and low rats.
Whether the altered neuronal activity found in PPI low rats is one of the
pathophysiological mechanisms leading to reduced PPI or whether it is subsequent to its
occurrence, is unclear. Also, several disorders that are accompanied by deficient
sensorimotor gating are associated with specific patterns of abnormal neural oscillations
and synchrony. They may thus be non-specific features of diverse pathophysiological
processes (Swerdlow et al., 1996). However, evidence from pharmacologic,
neurochemical and volumetric imaging studies, as well as emerging therapeutic reports,
suggest an overlap between the many reported (but potentially mechanistically
unrelated) brain disturbances in neuropsychiatric disorders, such as TS, and the
neurobiology of PPI. The experimental association of reduced PPI and TS has thus
been regarded useful for testing hypotheses and potential etiologies, and even for
developing novel therapeutics (Swerdlow, 2012).
In conclusion our study reveals that neuronal activity in a rat model with breeding-
induced PPI deficit shows findings similar to those seen in neuropsychiatric disorders
accompanied by disturbed sensorimotor gating. This model may be used to further
investigate and characterize the neuronal basis activity in these circuits to better
understand underlying mechanisms of these disorders.
References Alam M, Schwabe K, Krauss JK. The pedunculopontine nucleus area: critical evaluation
ofinterspecies differences relevant for its use as a target for deep brain stimulation. Brain 2011;
134:11–23.
Andrade P, Carrillo-Ruiz JD, Jiménez F. A systematic review of the efficacy of globus pallidus
stimulation in the treatment of Parkinson’s disease. J Clin Neurosci 2009; 16:877–881.

Manuscript two
42
Bennay M, Gernert M, Schwabe K, Enkel T, Koch M. Neonatal medial prefrontal cortex lesion
enhances the sensitivity of the mesoaccumbal dopamine system. Eur J
Neurosci 2004;19(12):3277-90.
Benabid AL, Koudsie A, Benazzouz A, Le Bas J-F, Pollak P. Imaging of subthalamic nucleus
and ventralis intermedius of the thalamus. Mov Disord 2002; 17 Suppl 3:S123–129.
Braff DL, Geyer MA, Swerdlow NR. Human studies of prepulse inhibition of startle: normal
subjects, patient groups, and pharmacological studies. Psychopharmacology (Berl) 2001;
156:234–258.
Braff DL, Light GA. The use of neurophysiological endophenotypes to understand the genetic
basis of schizophrenia. Dialogues Clin Neurosci 2005; 7:125–135.
Cadenhead KS, Light GA, Geyer MA, McDowell JE, Braff DL. Neurobiological measures of
schizotypal personality disorder: defining an inhibitory endophenotype? Am J Psychiatry 2002;
159:869–871.
Carlson PJ, Singh JB, Zarate CA Jr, Drevets WC, Manji HK. Neural circuitry and neuroplasticity
in mood disorders: insights for novel therapeutic targets. NeuroRx2006; 3:22–41.
Chen TY, Zhang D, Dragomir A, Akay YM, Akay M. Complexity of VTA DA neural activities in
response to PFC transection in nicotine treated rats. J Neuroeng Rehabil 2011; 8:13.
Dejean C, Hyland B, Arbuthnott G. Cortical effects of subthalamic stimulation correlate with
behavioral recovery from dopamine antagonist induced akinesia. Cereb Cortex 2009; 19:1055–
1063.
Dieckmann M, Freudenberg F, Klein S, Koch M, Schwabe K (2007) Disturbed social behavior
and motivation in rats selectively bred for deficient sensorimotor gating. Schizophr Res 97:250–
253.
Ellenbroek BA, Budde S, Cools AR. Prepulse inhibition and latent inhibition: the role of
dopamine in the medial prefrontal cortex. Neuroscience 1996; 75:535–542.
Fasano A, Daniele A, Albanese A. Treatment of motor and non-motor features of Parkinson’s
disease with deep brain stimulation. Lancet Neurol 2012; 11:429–442.
Freudenberg F, Dieckmann M, Winter S, Koch M, Schwabe K. Selective breeding for deficient
sensorimotor gating is accompanied by increased perseveration in rats. Neuroscience 2007;
148:612–622.
Groenewegen HJ, Russchen FT. Organization of the efferent projections of the nucleus
accumbens to pallidal, hypothalamic, and mesencephalic structures: a tracing and
immunohistochemical study in the cat. J Comp Neurol 1984; 223:347–367.

Manuscript two
43
Hadamitzky M, Harich S, Koch M, Schwabe K. Deficient prepulse inhibition induced by selective
breeding of rats can be restored by the dopamine D2 antagonist haloperidol.Behav Brain Res
2007; 177:364–367.
Hammond C, Bergman H, Brown P. Pathological synchronization in Parkinson’s disease:
networks, models and treatments. Trends Neurosci 2007; 30:357–364.
Hanslmayr S, Backes H, Straub S, Popov T, Langguth B, Hajak G, Bäuml K-HT, Landgrebe M.
Enhanced resting-state oscillations in schizophrenia are associated with decreased
synchronization during inattentional blindness. Human brain mapping 2012 [Epub ahead of
print].
Heidbreder CA, Groenewegen HJ. The medial prefrontal cortex in the rat: evidence for a dorso-
ventral distinction based upon functional and anatomical characteristics. Neurosci Biobehav Rev
2003; 27:555–579.
Houeto JL, Karachi C, Mallet L, Pillon B, Yelnik J, Mesnage V, Welter ML, Navarro S, Pelissolo
A, Damier P, Pidoux B, Dormont D, Cornu P, Agid Y. Tourette’s syndrome and deep brain
stimulation. J Neurol Neurosurg Psychiatr 2005; 76:992–995.
Hu J, Gao J, Principe JC. Analysis of biomedical signals by the lempel-Ziv complexity: the effect
of finite data size. IEEE Trans Biomed Eng. 2006; 53:2606-9.
Jahanshahi M, Czernecki V, Zurowski AM. Neuropsychological, neuropsychiatric, and quality of
life issues in DBS for dystonia. Mov Disord 2006; 26 Suppl 1:S63–78.
Kaneoke Y, Vitek JL. Burst and oscillation as disparate neuronal properties. Journal of
neuroscience methods 1996; 68:211–223.
Koch M, Bubser M. Deficient sensorimotor gating after 6-hydroxydopamine lesion of the rat
medial prefrontal cortex is reversed by haloperidol. Eur J Neurosci 1994; 6:1837– 1845.
Koch M, Fendt M, Kretschmer BD. Role of the substantia nigra pars reticulata in sensorimotor
gating, measured by prepulse inhibition of startle in rats. Behav Brain Res 2000; 117:153–162.
Kopell BH, Greenberg BD. Anatomy and physiology of the basal ganglia: implications for DBS in
psychiatry. Neurosci Biobehav Rev 2008; 32:408–422.
Krauss JK, Yianni J, Loher TJ, Aziz TZ. Deep brain stimulation for dystonia. J Clin Neurophysiol
2004; 21:18–30.
Kretschmer BD, Koch M. The ventral pallidum mediates disruption of prepulse inhibition of the
acoustic startle response induced by dopamine agonists, but not by NMDA antagonists. Brain
Res 1998; 798:204–210.

Manuscript two
44
Kühn AA, Neumann WJ, Hübl J, Brücke C, Capelle H.H, Krauss JK. Pallidal and thalamic local
field potentials in patients with Tourette-Syndrome. Movement Disorders 2011; Vol 26 (Suppl
S2) S1-S385
Labarre D, Meissner W, Boraud T. Measure of the regularity of events in stochastic point
processes, application to neuron activity analysis. In: Acoustics, Speech and Signal Processing,
2008. ICASSP 2008. IEEE International Conference on, pp 489 –492.
Leckman JF, Bloch MH, Smith ME, Larabi D, Hampson M. Neurobiological substrates of
Tourette’s disorder. Journal of Child and Adolescent Psychopharmacology 2010; 20:237–247.
Lempel A, Ziv J. On the Complexity of Finite Sequences. Information Theory, IEEE Transactions
on 1976; 22:75 – 81.
Lesne A, Blanc J-L, Pezard L. Entropy estimation of very short symbolic sequences. Phys Rev E
Stat Nonlin Soft Matter Phys 2009; 79:046208.
Lütjens G, Krauss JK, Schwabe K. Lesions of the entopeduncular nucleus in rats prevent
apomorphine-induced deficient sensorimotor gating. Behav Brain Res 2011; 220:281–287.
Marceglia S, Servello D, Foffani G, Porta M, Sassi M, Mrakic-Sposta S, Rosa M, Barbieri S,Priori
A. Thalamic single-unit and local field potential activity in Tourette syndrome. Mov Disord 2010;
25:300–308.
Moriarty J, Costa DC, Schmitz B, Trimble MR, Ell PJ, Robertson MM. Brain perfusion
abnormalities in Gilles de la Tourette's syndrome. Br J Psychiatry. 1995;167(2):249-54
Neuner I, Podoll K, Janouschek H, Michel TM, Sheldrick AJ, Schneider F. From psychosurgery
to neuromodulation: deep brain stimulation for intractable Tourette syndrome. World J Biol
Psychiatry 2009; 10:366–376.
Nicola SM, Surmeier J, Malenka RC. Dopaminergic modulation of neuronal excitability in the
striatum and nucleus accumbens. Annu Rev Neurosci. 2000;23:185-215. Review.
Posch DK, Schwabe K, Krauss JK, Lütjens G. Deep brain stimulation of the entopeduncular
nucleus in rats prevents apomorphine-induced deficient sensorimotor gating. Behav Brain Res
2012; 232:130–136.
Pothuizen HHJ, Jongen-Rêlo AL, Feldon J. The effects of temporary inactivation of the core and
the shell subregions of the nucleus accumbens on prepulse inhibition of the acoustic startle
reflex and activity in rats. Neuropsychopharmacology 2005; 30:683–696.
Rivlin-Etzion M, Ritov Y, Heimer G, Bergman H, Bar-Gad I. Local shuffling of spike trains boosts
the accuracy of spike train spectral analysis. J Neurophysiol 2006; 95:3245–3256.

Manuscript two
45
Sabri O, Erkwoh R, Schreckenberger M, Cremerius U, Owega A, Dickmann C, Schulz G, Zimny
M, Sass H, Buell U. Altered relationships between rCBF in different brain regions of never-
treated schizophrenics. Nuklearmedizin 1997; 36:194–201.
Schwabe K, Freudenberg F, Koch M. Selective breeding of reduced sensorimotor gating in
Wistar rats. Behav Genet 2007; 37:706-712.
Schwabe K, Polikashvili N, Krauss JK. Deficient sensorimotor gating induced by selective
breeding in rats is improved by entopeduncular nucleus lesions. Neurobiol Dis 2009; 34:351–
356.
Seamans JK, Lapish CC, Durstewitz D. Comparing the prefrontal cortex of rats and primates:
insights from electrophysiology. Neurotox Res. 2008; 14(2-3:249-62. Review.
Servello D, Porta M, Sassi M, Brambilla A, Robertson MM. Deep brain stimulation in 18 patients
with severe Gilles de la Tourette syndrome refractory to treatment: the surgery and stimulation. J
Neurol Neurosurg Psychiatr 2008; 79:136–142.
Sesack SR, Carr DB, Omelchenko N, Pinto A. Anatomical substrates for glutamatedopamine
interactions: evidence for specificity of connections and extrasynaptic actions. Ann N Y Acad Sci
2009; 1003:36–52.
Shahed J, Poysky J, Kenney C, Simpson R, Jankovic J. GPi deep brain stimulation for Tourette
syndrome improves tics and psychiatric comorbidities. Neurology 2007;
68:159–160.
Spencer KM, Nestor PG, Niznikiewicz MA, Salisbury DF, Shenton ME, McCarley RW. Abnormal
neural synchrony in schizophrenia. J Neurosci 2003; 23:7407–7411.
Swerdlow NR. Update: Studies of prepulse inhibition of startle, with particular relevance to the
pathophysiology or treatment of Tourette Syndrome. Neurosci Biobehav Rev. 2012 [Epub ahead
of print]
Swerdlow NR, Bakshi V, Geyer MA. Seroquel restores sensorimotor gating in
phencyclidinetreated rats. J Pharmacol Exp Ther 1996; 279:1290–1299.
Swerdlow NR, Geyer MA. Using an animal model of deficient sensorimotor gating to study the
pathophysiology and new treatments of schizophrenia. Schizophr Bull 1998; 24:285–301.
Swerdlow NR, Geyer MA, Braff DL. Neural circuit regulation of prepulse inhibition of startle in the
rat: current knowledge and future challenges. Psychopharmacology (Berl) 2001; 156:194–215.
Swerdlow NR, Sutherland AN. Preclinical models relevant to Tourette syndrome. Adv Neurol
2006; 99:69–88.
Szczepański J, Amigó JM, Wajnryb E, Sanchez-Vives MV. Characterizing spike trains with
Lempel–Ziv complexity. Neurocomputing 2004; 58–60:79–84.

Manuscript two
46
Tzschentke TM. Pharmacology and behavioral pharmacology of the mesocortical dopamine
system. Prog Neurobiol 2001; 63:241–320.
Uhlhaas PJ, Singer W. Abnormal neural oscillations and synchrony in schizophrenia. Nat Rev
Neurosci 2010; 11:100–113.
Wang Z, Maia TV, Marsh R, Colibazzi T, Gerber A, Peterson BS. The neural circuits that
generate tics in Tourette’s syndrome. American Journal of Psychiatry
2011;168:1326–1337.
Whittington MA. Can brain rhythms inform on underlying pathology in schizophrenia? Biol
Psychiatry 2008; 63:728–729.
Zahm DS, Heimer L. Specificity in the efferent projections of the nucleus accumbens in the rat:
comparison of the rostral pole projection patterns with those of the core and shell. J Comp
Neurol 1993; 327:220–232.

Discussion
47
9 Discussion
We used rats with breeding-induced deficient sensorimotor gating to examine whether
DBS in brain regions targeted for TS would alleviate the inbred deficiency. Additionally
we studied the neuronal activity in EPN, NAC and mPFC. In the first study we tested the
effect of chronic DBS in EPN, NAC and CM-Pf on the PPI of ASR in our PPI low and PPI
high rats. Examining the data from EPN stimulation, we found that the PPI deficit in the
PPI low group is already significantly improved after the implantation with no further
effect from DBS, which indicates that the electrode itself induced lesion effect in the
implantation site. The effect of implantation in CM-PF on the PPI was also nearly
significant. It seems that our PPI low rats are very sensitive to the microlesion caused by
the electrode implantation in EPN and CM-PF, so in any case, one should be aware of
the possible „settle effect“, when implanting in nuclei small in size. These results still
indicate a role of EPN in the regulation of the sensorimotor gating, but unfortunately
prevented us from testing the DBS effect. What about CM-Pf Additionaly, we implanted
monopolar electrodes with ~25% reduced diameter, but the settle effect remained.
Stimulation in the NAC did not lead to a significant improvement of the PPI in the low
group. Significantly alleviated PPI deficiency was found after stimulation with 150µA in
the CM-Pf. Our result, which is also in line with the results from the clinical studies (Hariz
et al., 2010; Savica et al., 2012), supports the assumed important role of CM-Pf in the
modulation of the sensorimotor gating.
Improved PPI is found also after DBS in several other brain regions e.g. mPFC,
dorsomedial thalamus (Klein et al., 2013) and STN (Lindemann et al., 2012).
We found that DBS did not affect the PPI high groups in any of the 3 different
implantation sites (EPN, NAC and CM-Pf). Not affecting the “normal” condition could
mean less side effects of the treatment.
The results we achieved by this experiment also prove the validity of our selectively bred
endophenotype model of disturbed sensorimotor gating. The predictive validity of our
model is additionaly supported by the results from our second project, where we

Discussion
48
recorded and compared the neuronal activity in mPFC, NAC and EPN of the PPI low
animals to the activity of the PPI high ones. After evaluating the data, we found reduced
firing rate, burst behavior and asymmetry index of mPFC neurons in PPI low rats, which
corresponds to the hypofrontality found in patients suffering from schizophrenia (Sabri et
al., 1997; Knyazeva et al., 2008). Functional brain imaging also found reduced perfusion
within the dorsolateral prefrontal cortex and the anterior cingulate cortex in TS patients
(Moriarty et al., 1995).
In contrast, the firing rate and the measures of regularity and complexity in NAC of the
PPI low animals were enhanced, when compared with the PPI high group. Possible
dysregulation in the mPFC feedback loop in the NAC could be a reason, since there is a
known functional relationship between the NAC and mPFC. NAC exerts PPI modulation
via projection to PPTg and also projects to the major BG output nucleus – EPN (GPi in
humans), which is effectively used for functional neurosurgery to treat movement and
neuropsychiatric disorders. We also have shown that lesions in this nucleus alleviate
PPI in PPI low rats and counteract apomorphine induced PPI deficit, but nevertheless in
the present study we found the firing rate and burst activity in EPN unchanged.
Since altered neural oscillations and synchrony seem to play a crucial role in the
pathophysiology of neuropsychiatric disorders accompanied by sensorimotor gating
deficits, we also recorded and analyzed the local field potentials in the mPFC, the NAC
and in the EPN. In PPI low rats we found an enhanced level of theta band oscillatory
activity in the NAC and EPN, which was accompanied by reduced beta band activity in
all the regions studied and also reduced alpha band activity in the EPN. This
corroborates with the results from TS patients, which show that LFP spectra in some
intralaminar thalamic nuclei and GPi have a prominent oscillatory activity at low
frequencies (2-7 Hz) and in the alpha band (8-13 Hz), and virtually absent beta activity
(Marceglia et al., 2010; Kühn et al. 2011).
In the near future, we plan to use PPI low and high rats to evaluate the effect of DBS (or
lesions) of the EPN and the CM-Pf on compromised neuronal activity in the mPFC and
NAC. We also plan to do the recordings of network activity in both anaesthetized and
awake, behaving rats.

Discussion
49
Since blockade of D2 dopamine receptors in TS patients most effectively alleviates tics,
and the DA D2 receptor antagonist haloperidol has been shown to alleviate deficient PPI
in PPI low rats, we also aim to investigate the effect of DA D2 receptor blockade on
neuronal activity. Further, we would like to simultaneously record neuronal activity from
the mPFC and NAC to assess their interplay, because we expect the concept of
simultaneous multichannel stimulation and recording will advance our understanding of
how DBS affects compromised neuronal circuit activity thought to be involved in
neuropsychiatric disorders with deficient sensorimotor gating.
However, we have to keep in mind also some criticism and limitations of the PPI
paradigm. It remains unclear, whether the altered neuronal activity found in PPI deficient
rats and patients causes the PPI deficit or whether it results from it. Since, PPI deficits
are found in several different neuropsychiatric disorders such as schizophrenia, TS,
OCD and HD - it rather indicates a specific dysregulation of central inhibitory
mechanisms, but not a clinically diagnosable condition (Swerdlow et al., 2008).
No clear qualitative value of “normal” or “deficient” can be applied to any level of PPI,
since there is significant overlap in PPI between clinically normal and disordered
individuals. It seems that low PPI levels can result from normal variations in the CSPT
circuitry and at the same time pathological processes can be the reason. This makes
PPI neither diagnostically nor physiologically specific. It is also quite sensitive to different
stimulus conditions and parameters, so same test population can exhibit either relatively
reduced or increased PPI, depending on the experimental conditions (Swerdlow et al.,
2006, 2008). In this context, however, we have actually tested our rats with prepulse
intensities ranging from 62-86 dB and with interstimulus intervals ranging between 30-
1000 ms and the PPI deficits remained evident (Schwabe et al., 2007).
There is also no proof, that neurobiological abnormalities, resulting in PPI deficiency are
necessary for the expression of TS, schizophrenia or any other neuropsychiatric
disorder. Clearly, the rodent brain and the human brain are not the same, but
furthermore there are even significant inter-strain differences in PPI, which could reflect
involvement of different PPI regulatory circuitries. Thus, findings about the neural
substrates regulating PPI in rodents are not always directly applicable to the human
brain (reviewed in Swerdlow et al., 2008).

Summary
50
10 Summary
Deep brain stimulation in rats with deficient sensorimotor gating: behavioral and electrophysiological studies Svilen Angelov
In certain neuropsychiatric disorders, such as Tourette’s syndrome (TS), obsessive
compulsive disorder (OCD) and schizophrenia, disturbed sensorimotor gating has been
described, which is linked to the inability of these patients to filter or “gate” irrelevant or
inferring information of motor, cognitive and emotional domains with consecutive
sensory overload and respective clinical symptoms. Pathophysiologically, a
dysregulation of neuronal network activity between cortex and associative-limbic basal
ganglia loops is suspected. In cases of pharmacoresistant TS and OCD high frequency
deep brain stimulation (DBS) is successfully used to relieve symptoms. Despite the
positive results, the exact mechanism of action of DBS remains unclear.
The use of experimental animals allows investigation of specific neuronal network
activity of functional and behavioral dysregulation. One operational measure of
sensorimotor gating processes in a cross species approach is the “prepulse inhibition
(PPI) of the acoustic startle response (ASR)”. Deficient PPI has been described in
patients with the aforementioned neuropsychiatric disorders and experimentally induced
PPI deficiency in rodents is therefore regarded as an endophenotype for deficient
sensorimotor gating in these disorders. Using rats, selectively bred for deficient
sensorimotor gating, we showed that neuromodulatory approaches in targets used for
DBS in human patients, alleviate the inbred PPI deficit. This proves the important role of
EPN and CM-Pf in the modulation of the sensorimotor gating and the reliability of our
endophenotpe model. Additionally, we found similarities between the neuronal activity
of our rats and the activity shown in neuropsychiatric disorders accompanied by
sensorimotor gating deficits i.e., hypofrontality and enhanced oscillatory theta band
activity in different coritico-basal ganglia regions. Considering the data we got from
these recent and our previous studies, we can conclude that rats, selectively bred for

Summary
51
deficient sensorimotor gating, may be used to further investigate the pathophysiology of
the disorders characterized with this deficiency, including TS. This model may also be
used to study the neuronal activity of the affected circuits, the mechanisms of DBS and
to develop and improve the therapeutic and preventive strategies.

Zusammenfassung
52
10 Zusammenfassung Tiefe Hirnstimulation bei Ratten mit sensorimotorischem Bahnungsdefizit: Verhaltenstechnische und elektrophysiologische Untersuchungen
Svilen Angelov
Bei bestimmten neuropsychiatrischen Störungen wie dem Tourette-Syndrom (TS), den
Zwangsstörungen und der Schizophrenie wird eine gestörte sensorimotorische Bahnung
beschrieben. Diese geht mit der Unfähigkeit der Patienten einher irrelevante oder
interferierende Informationen aus motorischen, kognitiven oder emotionalen Bereichen
zu filtern. Die daraus resultierende Reizüberflutung wird mit den entsprechenden
klinischen Symptomen in Verbindung gebracht. Pathophysiologisch wird eine
Fehlregulation der neuronalen Netzwerkaktivität zwischen Kortex und assoziativ-
limbischen Basalganglienschleifen vermutet. Bei Zwangsstörungen und
pharmakoresistentem TS wird die hochfrequente tiefe Hirnstimulation (THS) erfolgreich
eingesetzt, um die Symptome zu lindern. Trotz der positiven Ergebnisse, ist der genaue
Wirkmechanismus der THS weitgehend unbekannt.
In tierexperimentellen Ansätzen kann die neuronale Netzwerkaktivität von Funktions-
und Verhaltensstörungen gezielt untersucht werden. Eine operante Methode zur
Messung sensorimotorischer Bahnungsprozesse, die speziesübergreifend angewandt
werden kann, ist die Prepulsinhibition (PPI) der akustischen Schreckreaktion. Eine
verminderte PPI wurde bei Patienten mit den oben erwähnten neuropsychiatrischen
Erkrankungen beschrieben. Experimentell induzierte PPI-Defizite bei Nagern gelten
daher als Endophenotyp für eine gestörte sensorimotorische Bahnung.
Wir haben bei Ratten mit einer zuchtinduzierten verminderten sensorimotorischen
Bahnung gezeigt, dass neuromodulatorische Eingriffe im entopedunkulären Nukleus
(EPN, das Äquivalent zum Globus pallidus internus beim Menschen) und im
zentromediären parafaszikulären Nukleus (CM-Pf), also in Zielgebieten, die für die THS
bei Patienten verwendet werden, das zuchtinduzierte PPI-Defizit verbessern. Dies
beweist die wichtige Rolle des EPN und CM-Pf bei der Modulation sensorimotorischer

Zusammenfassung
53
Bahnungsprozesse, sowie die Validität unseres Zuchtmodells. Des Weiteren ähnelt die
neuronale Aktivität der untersuchten Ratten der Gehirnaktivität von Patienten mit
neuropsychiatrischen Erkrankungen, die mit einer gestörten sensorimotorischen
Bahnung einhergehen, d.h. eine Hypofrontalität und eine verstärkte oszillatorische
Theta-Band-Aktivität in verschiedenen Regionen der Basalganglien. Ratten, die selektiv
auf eine gestörte sensorimotorische Bahnung gezüchtet werden, eignen sich daher zur
weiteren Untersuchung der Pathophysiologie von neuropsychiatrischen Erkrankungen
mit dieser Störung, einschließlich TS. Dieses Modell kann auch genutzt werden, um den
Einfluss der THS auf die neuronale Aktivität der betroffenen Netzwerke zu untersuchen,
um gegebenenfalls neuartige therapeutische und präventive Strategien zu entwickeln
und zu verbessern.

References
54
11 References Ackermans L, Temel Y, Cath D, van der Linden C, Bruggeman R, Kleijer M, Nederveen P, Schruers K, Colle H, Tijssen MA, Visser-Vandewalle V; Dutch Flemish Tourette Surgery Study Group "Deep brain stimulation in Tourette's syndrome: two targets?", Mov Disord. 2006 May;21(5):709-13. Adams W, Kendell RE, Hare EH, Munk-Jørgensen P. "Epidemiological evidence that maternal influenza contributes to the aetiology of schizophrenia. An analysis of Scottish, English, and Danish data", Br J Psychiatry. 1993 Oct;163:522-34. Alfredo Berardelli, "Pathophysiology of tics and Tourette syndrome", J Neurol (2003) 250:781–787 Am J Hum Genet "Genome scan for Tourette disorder in affected-sibling-pair and multigenerational families", Tourette Syndrome Association International Consortium for Genetics, 2007 Arzimanoglou AA. "Gilles de la Tourette syndrome", J Neurol. 1998 Dec;245(12):761-5, PMID: 9840347 Benabid AL, Pollak P, Louveau A, Henry S, de Rougemont J. “Combined (thalamotomy and stimulation) stereotactic surgery of the VIM thalamic nucleus for bilateral Parkinson disease”, Appl Neurophysiol 1987;50:344-6. Bertelsen B, Debes NM, Hjermind LE, Skov L, Brøndum-Nielsen K, Tümer Z. "Chromosomal rearrangements in Tourette syndrome: implications for identification of candidate susceptibility genes and review of the literature", 2013, Neurogenetics. 2013 Nov;14(3-4):197-203. doi: 10.1007/s10048-013-0372-y. Epub 2013 Aug 29. Borrell J, Vela JM, Arévalo-Martin A, Molina-Holgado E, Guaza C. "Prenatal immune challenge disrupts sensorimotor gating in adult rats. Implications for the etiopathogenesis of schizophrenia", Neuropsychopharmacology. 2002 Feb;26(2):204-15. Braff DL, Freedman R, Schork NJ, Gottesman II. "Deconstructing schizophrenia: an overview of the use of endophenotypes in order to understand a complex disorder", Schizophr Bull. 2007 Jan;33(1):21-32. Epub 2006 Nov 6. Braff DL, Geyer MA, Swerdlow NR “Human studies of prepulse inhibition of startle: normal subjects, patient groups, and pharmacological studies”, Psychopharmacology 156:234-258, 2001b. Braff, D.L., Geyer, M.A., Light, G.A., Sprock, J., Perry, W., Cadenhead, K.S. and Swerdlow, N.R., 2001a. “Impact of prepulse characteristics on the detection of sensorimotor gating deficits in schizophrenia”, Schizophrenia research, 49(1-2), pp. 171-178. Breit S, Schulz JB, Benabid AL."Deep brain stimulation", Cell Tissue Res. 2004 Oct;318(1):275-88. Epub 2004 Aug 19. Brice J, McLellan L. “Suppression of intention tremor by contingent deep-brain stimulation”, Lancet 1980;1:1221-2. Brown AS, Begg MD, Gravenstein S, Schaefer CA, Wyatt RJ, Bresnahan M, Babulas VP, Susser ES."Serologic evidence of prenatal influenza in the etiology of schizophrenia", Arch Gen Psychiatry. 2004 Aug;61(8):774-80.

References
55
Castellanos, F.X., Fine, E.J., Kaysen, D., Marsh, W.L., Rapoport, J.L. and Hallett, m., 1996 “Sensorimotor gating in boys with Tourette's syndrome and ADHD: Preliminary results”, Biological psychiatry, 39(1), pp. 33-41. Cohen SC, Leckman JF, Bloch MH "Clinical assessment of Tourette syndrome and tic disorders", Neurosience & Behavioral reviews (2012) Cooper IS, Lee AJ.” Cryostatic congelation: a system for producing a limited, controlled region of cooling or freezing of biologic tissues”, J Nerv Ment Dis 1961;133:259-63. Cosman ER, Nashold BS, Bedenbaugh P. “Stereotactic radiofrequency lesion making”, Appl Neurophysiol 1983;46:160-6. Delong M, Wichmann T. “Deep brain stimulation for movement and other neurologic disorders”, Ann N Y Acad Sci. 1265:1-8, 2012. Delong MR, Wichmann T “Circuits and circuit disorders of the basal ganglia”, Arch Neurol 64:20-24, 2007. Deniau JM, Degos B, Bosch C, Maurice N “Deep brain stimulation mechanisms: beyond the concept of local functional inhibition”, Eur J Neurosci. 32(7):1080-91, 2010. Dieckmann M, Freudenberg F, Klein S, Koch M, Schwabe K. "Disturbed social behavior and motivation in rats selectively bred for deficient sensorimotor gating", Schizophr Res. 2007 Dec;97(1-3):250-3. Epub 2007 Sep 12. Duncan EJ, Madonick SH, Parwani A, Angrist B, Rajan R, Chakravorty S, Efferen TR, Szilagyi S, Stephanides M, Chappell PB, Gonzenbach S, Ko GN, Rotrosen JP. "Clinical and sensorimotor gating effects of ketamine in normals", Neuropsychopharmacology. 2001 Jul;25(1):72-83. Eusebio A, Thevathasan W, Doyle Gaynor L, Pogosyan A, Bye E, Foltynie T, Zrinzo L, Ashkan K, Aziz T, Brown P. "Deep brain stimulation can suppress pathological synchronisation in parkinsonian patients", J Neurol Neurosurg Psychiatry. 2011 May;82(5):569-73. doi: 10.1136/jnnp.2010.217489. Epub 2010 Oct 9. Fenelon G, Francois C, Percheron G, Yelnik J. "Topographic distribution of the neurons of the central complex (centre médian-parafascicular complex) and of other thalamic neurons projecting to the striatum in macaques", Neuroscience. 1991;45(2):495-510 Freudenberg F, Dieckmann M, Winter S, Koch M, Schwabe K. "Selective breeding for deficient sensorimotor gating is accompanied by increased perseveration in rats", Neuroscience. 2007 Sep 7;148(3):612-22. Epub 2007 Aug 10. Geyer MA, Krebs-Thomson K, Braff DL, Swerdlow NR. "Pharmacological studies of prepulse inhibition models of sensorimotor gating deficits in schizophrenia: a decade in review", Psychopharmacology (Berl). 2001 Jul;156(2-3):117-54. Geyer MA, Wilkinson LS, Humby T, Robbins TW. "Isolation rearing of rats produces a deficit in prepulse inhibition of acoustic startle similar to that in schizophrenia", Biol Psychiatry. 1993 Sep 15;34(6):361-72. Gironell A, Rodriguez-Fornells A,Kulisevsky J, Pascual B, Riba J, Barbanoj M, Berthier M (2000) "Abnormalities of the acoustic startle reflex and reaction time in Gilles de la Tourette syndrome", Clin Neurophysiol 111:1366–1371 Gould TD, Gottesman II "Psychiatric endophenotypes and the development of valid animal models", Genes Brain Behav. 2006 Mar;5(2):113-9.

References
56
Hadamitzky M, Harich S, Koch M, Schwabe K."Deficient prepulse inhibition induced by selective breeding of rats can be restored by the dopamine D2 antagonist haloperidol", Behav Brain Res. 2007 Feb 27;177(2):364-7. Epub 2006 Dec 19. Hariz MI, Robertson MM. “Gilles de la Tourette syndrome and deep brain stimulation”, Eur J Neurosci. 32:1128-34, 2010 Harrison PJ, Weinberger DR. "Schizophrenia genes, gene expression, and neuropathology: on the matter of their convergence", Mol Psychiatry. 2005 Jan;10(1):40-68; image 5. Hashimoto T, Elder CM, Okun MS, Patrick SK, Vitek JL. “Stimulation of the subthalamic nucleus changes the firing pattern of pallidal neurons”, J Neurosci 2003,23:1916-23. Hassler R, Dieckmann G. “Stereotaxic treatment of tics and inarticulate cries or coprolalia considered as motor obsessional phenomena in Gilles de la Tourette’s disease”, Rev Neurol 1970;123:89-100. Horsley V, Clark RH. “The structure and functions of the cerebellum examined by a new method.”, Brain 1908;31:45-124. Houeto JL, Karachi C, Mallet L, Pillon B, Yelnik J, Mesnage V, Welter ML, Navarro S, Pelissolo A, Damier P, Pidoux B, Dormont D, Cornu P, Agid Y “Tourette`s syndrome and deep brain stimulation”, J Neural Neurosurg Psychiatry 76:992-995, 2005. Karper, L.P., Freeman, G.K., Grillon, C., Morgan, C.A.,3rd, Charney, D.S. and Krystal, J.H., 1996 “Preliminary evidence of an association between sensorimotor gating and distractibility in psychosis”, The Journal of neuropsychiatry and clinical neurosciences, 8(1), pp. 60-66. Klein J, Hadar R, Götz T, Männer A, Eberhardt C, Baldassarri J, Schmidt TT, Kupsch A, Heinz A, Morgenstern R, Schneider M, Weiner I, Winter C. "Mapping brain regions in which deep brain stimulation affects schizophrenia-like behavior in two rat models of schizophrenia", Brain Stimul. 2013 Jul;6(4):490-9. doi: 10.1016/j.brs.2012.09.004. Epub 2012 Oct 8. Knyazeva MG, Jalili M, Meuli R, Hasler M, De Feo O, Do KQ “Alpha rhythm and hypofrontality in schizophrenia”, Acta Psychiatr Scand. 2008 Sep;118(3):188-99. Koch M, Fendt M, Kretschmer BD. "Role of the substantia nigra pars reticulata in sensorimotor gating, measured by prepulse inhibition of startle in rats", Behav Brain Res. 2000 Dec 20;117(1-2):153-62. Koch M, Fendt M. “Startle response modulation as a behavioral tool in neuropharmacology”, Curr Neuropharmacol 2003;1:175-85. Koch M, Kungel M, Herbert H."Cholinergic neurons in the pedunculopontine tegmental nucleus are involved in the mediation of prepulse inhibition of the acoustic startle response in the rat", Exp Brain Res. 1993;97(1):71-82 Koch M, Schnitzler HU. "The acoustic startle response in rats-circuits mediating evocation, inhibition and potentiation", Behav Brain Res. 1997 Dec;89(1-2):35-49. Krauss JK, Pohle T, Weber S, Ozdoba C, Burgunder JM. „Bilateral stimulation of globus pallidus internus for treatment of cervical dystonia”, Lancet 1999;354:837-8. Krauss JK, Volkmann J. „Tiefe Hirnstimulation. 1st edition“, Darmstadt: Steinkopf; 2004. Kühn AA, Neumann WJ, Hübl J, Brücke C, Capelle H.H, Krauss JK. „Pallidal and thalamic local field potentials in patients with Tourette-Syndrome”, Movement Disorders 2011; Vol 26 (Suppl S2) S1-S385

References
57
Kuhn J, Lenartz D, Mai JK, Huff W, Lee SH, Koulousakis A, Klosterkoetter J, Sturm V. "Deep brain stimulation of the nucleus accumbens and the internal capsule in therapeutically refractory Tourette-syndrome", J Neurol. 2007 Jul;254(7):963-5. Epub 2007 Apr 6. Larson PS. "Deep brain stimulation for psychiatric disorders", Neurotherapeutics. 2008 Jan;5(1):50-8. doi: 10.1016/j.nurt.2007.11.006 Le Pen G, Gourevitch R, Hazane F, Hoareau C, Jay TM, Krebs MO."Peri-pubertal maturation after developmental disturbance: a model for psychosis onset in the rat", Neuroscience. 2006 Dec 1;143(2):395-405. Epub 2006 Sep 14. Lindemann C, Krauss JK, Schwabe K. "Deep brain stimulation of the subthalamic nucleus in the 6-hydroxydopamine rat model of Parkinson's disease: effects on sensorimotor gating", Behav Brain Res. 2012 Apr 21;230(1):243-50. doi: 10.1016/j.bbr.2012.02.009. Epub 2012 Feb 11. Lipska BK, Swerdlow NR, Geyer MA, Jaskiw GE, Braff DL, Weinberger DR. "Neonatal excitotoxic hippocampal damage in rats causes post-pubertal changes in prepulse inhibition of startle and its disruption by apomorphine", Psychopharmacology (Berl). 1995 Nov;122(1):35-43. Lozano AM, Dostrovsky J, Chen R, Ashby P. “Deep brain stimulation for Parkinson's disease: disrupting the disruption”, Lancet Neurol 2002,1:225-31. Lozano AM, Snyder BJ, Hamani C, Hutchison WD, Dostrovsky JO “Basal ganglia physiology and deep brain stimulation”, Mov Disord. 25 Suppl 1:S71-5. Review, 2010. Lütjens G, Krauss JK, Schwabe K: Lesions of the entopeduncular nucleus in rats prevent apomorphine-induced deficient sensorimotor gating. Behav Brain Res. 2011 Jul 7;220(2):281-7. Maciunas RJ, Maddux BN, Riley DE, Whitney CM, Schoenberg MR, Ogrocki PJ, Albert JM, Gould DJ. "Prospective randomized double-blind trial of bilateral thalamic deep brain stimulation in adults with Tourette syndrome", J Neurosurg. 2007 Nov;107(5):1004-14. Mansbach RS, Braff DL, Geyer MA. "Prepulse inhibition of the acoustic startle response is disrupted by N-ethyl-3,4-methylenedioxyamphetamine (MDEA) in the rat", Eur J Pharmacol. 1989 Aug 11;167(1):49-55. Mansbach RS, Geyer MA, Braff DL. "Dopaminergic stimulation disrupts sensorimotor gating in the rat", Psychopharmacology (Berl). 1988;94(4):507-14. Mansbach RS, Geyer MA."Effects of phencyclidine and phencyclidine biologs on sensorimotor gating in the rat", Neuropsychopharmacology. 1989 Dec;2(4):299-308. Marceglia S, Servello D, Foffani G, Porta M, Sassi M, Mrakic-Sposta S, Rosa M, Barbieri S,Priori A. “Thalamic single-unit and local field potential activity in Tourette syndrome”, Mov Disord 2010; 25:300–308. Martino D, Dale RC, Gilbert DL, Giovannoni G, Leckman JF. "Immunopathogenic mechanisms in tourette syndrome: A critical review", Mov Disord. 2009 Jul 15;24(9):1267-79. doi: 10.1002/mds.22504. McIntyre CC, Savasta M, Kerkerian-Le Goff L, Vitek JL. “Uncovering the mechanism(s) of action of deep brain stimulation: activation, inhibition, or both”, Clin Neurophysiol 2004;115:1239-48. Miguel EC, De Rosario-Campos MC, Prado HS, Do Valle R, Rauch SL, Coffey BJ, Baer L, Savage CR, O`Sullivan RL, Jenike MA, Leckman JF “Sensory phenomena in obsessive-compulsive disorder and Tourette`s disorder”, J Clin Psychiatry 61: 150-156, 2000.

References
58
Mink JW. "The basal ganglia: focused selection and inhibition of competing motor programs", Prog Neurobiol. 1996 Nov;50(4):381-425. Mink JW."The Basal Ganglia and involuntary movements: impaired inhibition of competing motor patterns", Arch Neurol. 2003 Oct;60(10):1365-8. Mink, J.W. "Basal ganglia dysfunction in Tourette’s syndrome: a new hypothesis", Pediatric neurology, 2001 Miyakawa T, Leiter LM, Gerber DJ, Gainetdinov RR, Sotnikova TD, Zeng H, Caron MG, Tonegawa S."Conditional calcineurin knockout mice exhibit multiple abnormal behaviors related to schizophrenia", Proc Natl Acad Sci U S A. 2003 Jul 22;100(15):8987-92. Epub 2003 Jul 8. Modolo J, Legros A, Thomas AW, Beuter A “Model-driven therapeutic treatment of neurological disorders: reshaping brain rhythms with neuromodulation”, Interface Focus. 1:61-74, 2011. Moriarty J, Costa DC, Schmitz B, Trimble MR, Ell PJ, Robertson MM. "Brain perfusion abnormalities in Gilles de la Tourette's syndrome", Br J Psychiatry. 1995 Aug;167(2):249-54. Mueller-Vahl KR, Meyer GJ, Knapp WH, Emrich HM, Gielow P, BrГјcke T, et al. "Serotonin transporter binding in Tourette Syndrome", Neurosci Lett. 2005 Müller-Vahl KR. “Surgical treatment of Tourette syndrome”, Neurosci Biobehav Rev. 2013 Jul;37(6):1178-85. doi: 10.1016/j.neubiorev.2012.09.012. Epub 2012 Oct 4. Nakano K, Kayahara T, Tsutsumi T, Ushiro H. "Neural circuits and functional organization of the striatum", J Neurol. 2000 Sep;247 Suppl 5:V1-15. Nauta WJ, Mehler WR. "Projections of the lentiform nucleus in the monkey", Brain Res. 1966 Jan;1(1):3-42. Neuner I, Podoll K, Janouschek H, Michel TM, Sheldrick AJ, Schneider F. "From psychosurgery to neuromodulation: deep brain stimulation for intractable Tourette syndrome", World J Biol Psychiatry. 2009;10(4 Pt 2):366-76. doi: 10.1080/15622970802513317 Nowak K, Mix E, Gimsa J, Strauss U, Sriperumbudur KK, Benecke R, Gimsa U "Optimizing a rodent model of Parkinson's disease for exploring the effects and mechanisms of deep brain stimulation", Parkinsons Dis. 2011;2011:414682. doi: 10.4061/2011/414682. Epub 2011 Apr 5. Parent A, Hazrati LN."Functional anatomy of the basal ganglia. I. The cortico-basal ganglia-thalamo-cortical loop", Brain Res Brain Res Rev. 1995 Jan;20(1):91-127. Paschou P. "The genetic basis of Gilles de la Tourette Syndrome", Neurosci Biobehav Rev. 2013 Jul;37(6):1026-39. doi: 10.1016/j.neubiorev.2013.01.016. Epub 2013 Jan 17. Peng RY, Mansbach RS, Braff DL, Geyer MA. "A D2 dopamine receptor agonist disrupts sensorimotor gating in rats. Implications for dopaminergic abnormalities in schizophrenia", Neuropsychopharmacology. 1990 Jun;3(3):211-8. Perry, W. and Braff, D.L., 1994. “Information-processing deficits and thought disorder in schizophrenia”, The American Journal of Psychiatry, 151(3), pp. 363-367. Perry, W., Minassian, A., Feifel, D. and Braff, D.L., 2001 “Sensorimotor gating deficits in bipolar disorder patients with acute psychotic mania”, Biological psychiatry, 50(6), pp. 418-424.

References
59
Porta M, Servello D, Zanaboni C, Anasetti F, Menghetti C, Sassi M, Robertson MM. “Deep brain stimulation for treatment of refractory Tourette syndrome: long-term follow-up”, Acta Neurochir (Wien). 154:2029-41, 2012. Posch DK, Schwabe K, Krauss JK, Lütjens G: Deep brain stimulation of the entopeduncular nucleus in rats prevents apomorphine-induced deficient sensorimotor gating. Behav Brain Res. 2012 Jun 15;232(1):130-6. Quartarone A, Hallett M. "Emerging concepts in the physiological basis of dystonia", Mov Disord. 2013 Jun 15;28(7):958-67. doi: 10.1002/mds.25532. Ralph RJ, Paulus MP, Fumagalli F, Caron MG, Geyer MA. "Prepulse Inhibition Deficits and Perseverative Motor Patterns in Dopamine Transporter Knock-Out Mice: Differential Effects of D1 and D2 Receptor Antagonists", J Neurosci. 2001 Jan 1;21(1):305-13. Roessner V, Plessen KJ, Rothenberger A, Ludolph AG, Rizzo R, Skov L, et al. "European clinical guidelines for Tourette syndrome and other tic disorders. Part II: pharmacological treatment", Eur Child Adolesc Psychiatry. 2011 Sabri O, Erkwoh R, Schreckenberger M, Cremerius U, Owega A, Dickmann C, Schulz G, Zimny M, Sass H, Buell U “Altered relationships between rCBF in different brain regions of never-treated schizophrenics”, Nuklearmedizin. 1997 Sep;36(6):194-201 Savica R, Stead M, Mack KJ, Lee KH, Klassen BT. "Deep brain stimulation in tourette syndrome: a description of 3 patients with excellent outcome", Mayo Clin Proc. 2012 Jan;87(1):59-62. doi: 10.1016/j.mayocp.2011.08.005. Schwabe K, Freudenberg F, Koch M. "Selective breeding of reduced sensorimotor gating in Wistar rats", Behav Genet. 2007 Sep;37(5):706-12. Schwabe K, Polikashvili N, Krauss JK. "Deficient sensorimotor gating induced by selective breeding in rats is improved by entopeduncular nucleus lesions", Neurobiol Dis. 2009 May;34(2):351-6. doi: 10.1016/j.nbd.2009.02.004. Epub 2009 Feb 20. Shadet J, Poysky J, Kenney C, Simpson R, Jankovic J “GPi deep brain stimulation for Tourette syndrome improves tics and psychiatric comorbidities”, Neurology 68: 159-160, 2007. Singer HS. "Tourette's syndrome: from behaviour to biology.", Lancet Neurol. 2005 Mar;4(3):149-59. Sipes TA, Geyer MA."Multiple serotonin receptor subtypes modulate prepulse inhibition of the startle response in rats", Neuropharmacology. 1994 Mar-Apr;33(3-4):441-8. Spiegel EA, Wycis HT, Marks M, Lee AJ. “Stereotaxic Apparatus for operations on the human brain”, Science 1947;106:349-50. Swain JE, Scahill L, Lombroso PJ, King RA, Leckman JF. "Tourette syndrome and tic disorders: a decade of progress", J Am Acad Child Adolesc Psychiatry. 2007 Aug;46(8):947-68. Swerdlow NR, Braff DL, Geyer MA. "Animal models of deficient sensorimotor gating: what we know, what we think we know, and what we hope to know soon", Behav Pharmacol. 2000 Jun;11(3-4):185-204. Swerdlow NR, Braff DL, Geyer MA."Cross-species studies of sensorimotor gating of the startle reflex", Ann N Y Acad Sci. 1999 Jun 29;877:202-16. Swerdlow NR, Geyer MA, Braff DL “Neural circuit regulation of prepulse inhibition of startle in the rat: current knowledge and future challenge”, Psychopharmacology 156:194-215, 2001b.

References
60
Swerdlow NR, Geyer MA. "Clozapine and haloperidol in an animal model of sensorimotor gating deficits in schizophrenia", Pharmacol Biochem Behav. 1993 Mar;44(3):741-4. Swerdlow NR, Lipska BK, Weinberger DR, Braff DL, Jaskiw GE, Geyer MA. "Increased sensitivity to the sensorimotor gating-disruptive effects of apomorphine after lesions of medial prefrontal cortex or ventral hippocampus in adult rats", Psychopharmacology (Berl). 1995 Nov;122(1):27-34. Swerdlow NR, Mansbach RS, Geyer MA, Pulvirenti L, Koob GF, Braff DL. "Amphetamine disruption of prepulse inhibition of acoustic startle is reversed by depletion of mesolimbic dopamine.", Psychopharmacology (Berl). 1990;100(3):413-6. Swerdlow NR, Taaid N, Halim N, Randolph E, Kim YK, Auerbach P. "Hippocampal lesions enhance startle gating-disruptive effects of apomorphine in rats: a parametric assessment", Neuroscience. 2000;96(3):523-36 Swerdlow NR, Talledo J, Sutherland AN, Nagy D, Shoemaker JM. "Antipsychotic effects on prepulse inhibition in normal 'low gating' humans and rats", Neuropsychopharmacology. 2006 Sep;31(9):2011-21. Epub 2006 Feb 8. Swerdlow NR, Weber M, Qu Y, Light GA, Braff DL. "Realistic expectations of prepulse inhibition in translational models for schizophrenia research", Psychopharmacology (Berl). 2008 Aug;199(3):331-88. doi: 10.1007/s00213-008-1072-4. Epub 2008 Jun 21. Swerdlow NR, Young AB. "Neuropathology in Tourette syndrome: an update", Adv Neurol. 2001 Swerdlow, N.R., Braff, D.L. and Geyer, M.A., 2000 “Animal models of deficient sensorimotor gating: what we know, what we think we know, and what we hope to know soon”, Behavioural pharmacology, 11(3-4), pp. 185-204. Swerdlow, N.R., Karban, B., Ploum, Y., Sharp, R., Geyer, M.A. and Eastvold, A., 2001a “Tactile prepuff inhibition of startle in children with Tourette’s syndrome: in search of an “fMRI-friendly” startle paradigm”, Biological psychiatry, 50(8), pp. 578-585. Swerdlow, N.R., Paulsen, J., Braff, D.L., Butters, N., Geyer, M.A. and Swenson, M.R., 1995 “Impaired prepulse inhibition of acoustic and tactile startle response in patients with Huntington's disease”, Journal of neurology, neurosurgery, and psychiatry, 58(2), pp. 192-200. Temel Y, Visser-Vandewalle V. “Surgery in Tourette syndrome”, Mov Disord 2004;19:3-14. Van der Linden C, Colle H, Vandewalle V, Alessi G, Rijckaert D, de Waele L. “Successful treatment of Tics with bilateral internal pallidum (GPi) stimulation in a 27-year-old male patient with Gilles de la Tourette’s syndrome (GTS)”, Mov Disord 2002;17:1130. Vandewalle V, van der Linden C, Groenewegen HJ, Caemaert J. “Stereotactic treatment of Gilles de la Tourette syndrome by high frequency stimulation of thalamus”, Lancet 1999;353:724. Viswanathan A, Jimenez-Shahed J, Carvallo JF, Jankovic J. “Deep brain stimulation for Tourette syndrome: target selection”, Stereotact Funct Neurosurg. 90:213-24, 2012. Vitek JL. "Mechanisms of deep brain stimulation: excitation or inhibition", Mov Disord. 2002;17 Suppl 3:S69-72. Vitek JL. “Deep brain stimulation: how does it work?”, Cleve Clin J Med. 75 Suppl 2:S59-65, 2008.

References
61
Vollenweider FX, Remensberger S, Hell D, Geyer MA."Opposite effects of 3,4-methylenedioxymethamphetamine (MDMA) on sensorimotor gating in rats versus healthy humans", Psychopharmacology (Berl). 1999 Apr;143(4):365-72. Welter ML, Mallet L, Houeto JL, Karachi C, Czernecki V, Cornu P, Navarro S, Pidoux B, Dormont D, Bardinet E, Yelnik J, Damier P, Agid Y. "Internal pallidal and thalamic stimulation in patients with Tourette syndrome." , Arch Neurol. 2008 Jul;65(7):952-7. doi: 10.1001/archneur.65.7.952. Yelnik J. "Functional anatomy of the basal ganglia", Mov Disord. 2002;17 Suppl 3:S15-21.

Appendix
62
12 Appendix
A. Thionine staining
1 | 100% ethanol 3 min 2 | 90% ethanol 3 min 3 | 80% ethanol 3 min 4 | 70% ethanol 3 min 5 | 50% ethanol 3 min 6 | H2O bidest 2-3 min 7 | Thionine solution 75-90 sec 8 | H2O bidest 2-3x dipping 9 | 70% ethanol 3 min 10 | 80% ethanol 3 min 11 | 90% ethanol 3 min 12 | 100% ethanol 3 min 13 | Roti-Histol®/100% ethanol (1:2) 5 min 14 | Roti-Histol® 5 min 15 | Roti-Histol® 5 min 16 | cover with Vitro-clud® Thionine solution: 100 mL 1 M acetic acid
(5.7 mL 100% acetic acid ad 100 mL H2O bidest) 36 mL 1 M NaOH (4 g NaOH platelets ad 100 mL H2O bidest) ad 1 L H2O bidest heat to 60-70 °C 1.25 g thionine (Sigma-Aldrich) mix 1 h, filtrate hot store at 60 °C, cool down and filtrate before usage

Appendix
63
B. Hemalum/Eosin staining according to Mayer (20 μm cryosections) 1 | 100% ethanol 5 min 2 | 100% ethanol 5 min 3 | 95% ethanol 2 min 4 | 70% ethanol 2 min 5 | H2O bidest 3 min 6 | Hemalum solution 8 min 7 | warm running tap water 10 min 8 | H2O bidest 2-3x dipping 9 | 96% ethanol 10x dipping 10 | Eosin Y solution 1 min 11 | 96% ethanol 5 min 12 | 100% ethanol 5 min 13 | 100% ethanol 5 min 14 | Roti-Histol®/100% ethanol 5 min 15 | Roti-Histol® 5 min 16 | Roti-Histol® 5 min 17 | cover with Vitro-clud® Hemalum solution: dilute “Mayers Hämalaun” (Merck) 1:2 with PBS Eosin Y stock solution (1%): 10 g Eosin Y
200 mL H2O bidest 800 mL 95% ethanol
Eosin Y solution (0.25%): 350 mL stock solution
750 mL 80% ethanol 5 mL glacial acetic acid

Appendix
64
C. Solutions for histological preparations Phosphate buffer saline 0.01 M (PBS): 9.55 g PBS powder (w/o Ca/Mg; Dulbecco)
ad 1 L H2O bidest adjust pH to 7.6
Gelatin for slide coating: 3.5 g gelatin powder
ad 500 mL H2O bidest 1.4 mL chrome alum few crumbs thymol (inhibition of bacteria) mix at 50 – 60 °C cool down and filter before usage
Antifreeze solution for brain slice storage (2 L): 600 mL glycerol
600 mL ethylene glycol 800 mL PBS
Paraformaldehyde for brain perfusion (4%): 10 g Paraformaldehyde
ad 250 mL PBS mix at 60-70 °C wait for clarity of the solution cool down and filter before usage check pH-value (7.4)

65
Acknowledgements
It would not have been possible to write this doctoral thesis without the support of many kind people around me. Although for reason of space I can’t give everyone's name here, I will mention at least some of them. Above all, I would like to thank my mother and my father for their unconditional support all these years. My whole education would not be possible without them and I know they have given up many things for me to get it. I would like to thank Prof. Dr. Joachim K. Krauss for giving me the opportunity to do research at the Neurosurgical Clinic of the Hannover Medical School and for his continuous support, expert scientific advice, and kindness. I would like to express my deepest gratitude to my supervisor Prof. Dr. Kerstin Schwabe for the patient guidance and advice throughout my time as her student. I have been very lucky to have a supervisor who cared so much about my work, and who responded to my queries so promptly. I am thankful to my co-supervisors Prof. Dr. Stefan Bleich and Prof. Dr. Andrej Kral for their support and scientific comments. Special thanks to Dr. Mesbah Alam for teaching me electrophysiology, supporting me whenever I needed it, for his great patience all the time and at last, but by no means least, being a very kind friend of mine. Here is the place to say: Thank you Linda for the great Indian food you cooked many times for me! I would like to thank Assoc. Prof. Dr. Dobri Yarkov from the Trakia University in Bulgaria for his support throughout my study years there. Many thanks to my special friends Martina Toncheva and Dr. Elvis Hermann, without you both, I guess I would not go to Germany at all. I would like to thank Juergen Wittek for his great assistance in the laboratory during my PhD study, but most of all for being a friend, without who my stay in Germany would not be even half the pleasure. I would also like to acknowledge the valuable technical assistance I always get from Monika van Iterson, Heike Achilles and Hans Heissler.

66
I would like to thank all the professors from the Center for Systems Neuroscience – I have learned something from every one of you. I am thankful to Christoph Lindeman, PhD and Carolin Dietrich for establishing some of the devices and methods I readily used afterwards. With their help I also made my first steps in science. Thanks to Dr. Goetz Luetjens for his assistance and to Nadine Polascheck, PhD for the advices. Thanks to Prof. Dr. Beatrice Grummer, Dr. Tina Selle and Saskia Guenter for their kind assistance at the HGNI coordination office. Thanks to all the little Bulgarian patients treated in the Children's Clinic and to their loving parents for making the non-scientific part of my life so meaningful. I wish you good health and happiness! Thanks to all the PhD students I worked with and to all my dear friends in Hannover for their kindness and support.