university of adelaideabstract,,hls thesig covers the investigtion of non-lps antþene exp,rressed...
TRANSCRIPT

S.C-loç,É,
('
Non -Llpopolys aocharide Protecttve Antl gen s of Vi brio
Cholerae
Dharam Pal Sharma
I
ô*--rr-rçtr. I C".c. d å'¿, f .'ì,*¡
Department of Mic¡obiolog and Immu¡olo6l
Univenityof Adelaide
Adelaide 5000
Australia
A Thesis Submitted for the Degree of Doctor of Phibeophy
lYlarch 1990

. , - . , ., :,
', ,, .¡;'-.,¡5; ¡1¡1; .j
Non-Lipopolysaccharide Antigens
:'eof Vibrio cholero,e .i t..
"+.-t¡-,;+
ÐDhararrt PaI S lït

I dedicate this thesis to rywife and to my children for tùeir hue and patienoe. Thankyou
foryour loYal suPPort

Abstract
,,hls thesig covers the investigtion of non-LPS antþene exp,rressed byboú biotypeo (Ctassical
and El Tor) and serotypes (Ogrwa and Inaba) oî,l4bb døbæ and the determi¡ation of the
eúent to which these are shared e¡ 6fi'ain-specific, since rhic will be a criticat determinant of
theirvaccine potenriât. The established inf¡nt mou6e modelwao used to iludy both virulence
and protection The non-LPS antigens are eryressed both þ Claosical and El Tor drains tested
eæept for four old bolaæs of the El Torbiotype. The protective activitie¡of antibodies to the
Bhåred antþns correhted with their capacities to hhibit the å¿ vifro atræhment of vibrb
organisms to isolated murine enterccytes.
Suboequentsbrdies were di¡ected to*,a¡ds identi$ingtìe non-LPS antiget(s)- Su¡diee
from orreßeas suggested foxin<oregulatedpili GCP) is an important pilue colonþefien f¡ctor.
Considerable time wag spent in trying to find expressiea ef thi¡ laclor h víto elldthen witù the
aid of SDS-PACE, immunobloüing and electron microscopy techniques including im-
munogold-labetling itwas found to be eryreseed only by Classical strain&
The infant mouse cholera rcdel was used to evaluate the ¡el¿tive imporønce of TCP
as protectiveantþns of.V dpbæOl. Antibodies to TCPwere sufficþntto conferp¡otection
againet two Classical strains, and were mo¡e effrcient if the challengevibrios were culu¡red for
TCP ery,ression In cont¡ast, such antibodies did not protect mice aeainst challenge with any
of four st¡ai¡s of El Tbr biotype. Since two of the latær have previousþbeen shoq¡n to po*sess

non-Lps prot€ctive antrgenq the¡e results ouggest that TCP a¡e not the onþ such antigea il
thb model
A genrm containing antibodies to non-LPS protectiye antigels of V úolsac has been
use4 afær extensive absorption, to fac¡litate the cloningof genes involved in the rynthesis of
TCp A gene bankwa¡ conetructed fromV dptaae 2,17561 DNA using a mobiliz¡ble cæmll
vectorinE coli, a¡deubsequently transfer¡edþ conjugation i\lovúolqae O17. Thb shai¡
does not produce Tep ¿n vúto endlacts non-LPS protective antþns. E¡Ét pooitive clones
were isolate4 and of these, four producod TeP as deærmined by eþct¡on nicroecopic and
immunoblottinganaþs€& TCP positive O17 cloneswere 70-fold morevirulent^han fQp-¡sgp-
tive clones or O17 in the infant mouse cholera modet Only the former could remove p,rotective
anftodies by absorption and upon immunization elicit protective antibodieo specific for
non-LpS components. As a oorollary, serum containing antibodþe to rcP protoct€d mbe
from challenge with TCP-poeitive cloneo, but not with rcP-negative clones or O17,
demonstratingthatantibodies to TCP a¡e sufficient to mediate protectio¡ in thb rystem- The
data indicaæ that Ttxt ¡s both avinrlence determinant and a protective antþn in the i¡h¡t
mouse cholera model This çþrt þ the best of my knowtedge, hm demo¡etrated for the fnst
time the protective efrect of anti:TCP antibodies albeit i¡ the infa¡t lDouse cholera model

Thi¡ thesb ç6¡¡r¡ins no material which h¡¡ been accepted for the 8s'ad of any other degree or
diploma in any Univerrity ¡nd to the be¡t of Ey tûowledge a¡d belie[ tf,is thesis contains no
material psodoruþ prblbhed or wriüen þ another perso& ercePtuùere due reference i¡
made in the ûext of the tùeeis. The author oonsents to the thesb beiog made available for
photooopyiagandloan, if applicabþ and if accefled for the awad of the &g¡æ'
IÌù¡ram P¡l Sh¡ma

Acknowledgements
I wish to €fpress my fupest appreciation and gntitude to my supervísor, Dr' SÛephen R
Ãtt i¿ge for his excellent guidance and encouraçment throug[tout the cour¡e of tlis proiec¿
I am deepþ indebted to hofessor Derrick Ronley for giving me the opportunity þ conduct
thb research in the Department of Microbiologl and Immunologl and for valuable suggestions
and timeþ critbisn&
My hearúelt thanks go to: Eh. Connor Thomas for hb eryertire in eþctron micra-
co[y; Dr. Paul Manni¡g for his helpful discussions; Gafy Fenny and ct¡is cnrsaro fo¡
professional hetp in photognphy. My deep 8en8e of appreciation Pes Ûo Atbeft Horcn'oo4
Chief of the Medicat School 6¡imal House, for his ercelþ¡t mai¡ænance of tle i¡fa¡t mouse
colony without which thb proiect would not have been pæsible.
I c'uld also like 1e ¡[¡nk past and pres€nt colleagues in Labs 6 and Enætorra¡ Labo for
mâEng it such a pleasant envi¡onment in which towort I ac&nw'ledge also the receiptof a
university of Adelaide Postgraduate scholanhip duriag the period of Ey research'
Last, but by no meatrg leas! I am greatly indebted to my wife, Nen4 for her extreme
patience during the time'his thesis was being assembled.

Publications
Materiat contained in this thesis has been publbhed i¡ the following
Sharma D.p., Aüridge S,, HackeüJ. & Ror*'ley D. (19SÐ Non-lþpolyeaccharide protective
antþenssharedþClaseicålandElTorbiot¡rpes ofVibrío úobæ, Iotunalof Infætbns
Dísezrvs l55z 716-7?3..
Sharma D.P, Thomas Q.J., Hall R[L,I-er¡ine M.]vf. & Aüridgp S.R. (1989) Signiñcance of
toxinoregulated pili as protective antigeno of,V dplqw in tûe inf¡nt mouse model
Vacche. 7: 151456.
Sharma D.P., Stroeher U.H, Thom¡e CJ., Manning PÂ & Affiidgp SR (1989) The charac'
terization of a maior non-LPS protective antigen of.Víb¡bchobæ sEa¡n 21756l and
mobcu tar cloning of the TC? l¡: A dvwtø b tr^xath on dplaa ø¿ rclale¿ diarrhæt
(Eds. Y. Dkeda & R-B. SacL). Vol. VIIL Toþo: KTK publicatio¡s. In P¡es*
Sharma D.P., Stroeher U.H., Thomas CI., Manning P.A & Arridgp S.R (1990) The torin-
coregulated pilua (ICP) oî. Wbb úolawz molecular clqning of genes invoþed i¡
biosynthesb and waluation of TCP as a protective antigen in the infant mouse model
Mìøobíal Pathogencsí& In Pr€ss"

Abbrevlatlons
Ap
CFA
CFB
CP
CT
DIC
dNTP
DNA
E coli
ETISA
EM
E]IEÊ,
Gur
IIAg
ID
tDa
kn
LD
LPS
MFRIIA
MoAb
MSITA
NA
NB
ampicillin
oolonizati,on factor agar
colonization fætor broth
clone probing
cholera toxin
duodenal inocuhtbn with cecal lþtion
deoxy-nucleoeide triPhosPhaæ
deo:ryribonu cleic acid
Esrlwíc.hhcoli
enzyme-linked immunoeo¡bent assãy
elctron micrucoPY
enterotoxþnic
monosialoeYl gan glioeide
hemagglutinins
infectiou¡ dose
kilodalton
Lanauycia
leih¡l dæe
lipopolyt""charide
þ-m¡ nn e6e-frrcose-re¡ietant hemagglutinin
monoclonal antibodY
D-mannose+ensitive hemagglutinin
nutrient agar
nutrientbroth

OD
OMP
oRs
oRr
PAGE
PBS
PP
R
RBC
Sark-OMP
s
w
SDS
SE
SHA
Sm
SRBC
TBS
Tb
TCP
TSA
TTBS
ttg
pl
Vchobæ
optical deûsity
outer membrane Protein
oral rehydration solution
oral rehydration theraPY
poþacrylamide gel electrophoresie
phosphaæ buffered saline
Peyer'e patches
resistant
redblood cells
Sarkosyl-insoluble OMP
sensitive
subcuttneot¡s
¡odium dodecYl rulPhate
¡tanda¡d error
soluble hemagSlutinin
ctrepomycin
sheep red blood cell¡
Îis-bufiered salhe
tetracycline
f orin coregulated Piltts
lis-buffered sati¡e containing sodium a^&
Tlisåufiered saline 6paraining Tlreen Z)
microgram
nicrolite(t)
V¡bito ùolaac

Tbble of Contents
1 Introduction
1.1 HistoricalBacþround . . . . .'''''''
1.2 Etiologr
l.Ll Characteristics of the bacterium
l.Z2 Claseification
13 Fathogenes¡s andPathophysiologr 4
1.3.1 lngestionandinfectiousdose "" ""' 5
132 Penet¡ationoftlemucuslayer'''''''''' "' 5
1.3.3 Adherence to the epithelium of the smâll iltesti¡e
1.3.3.1 F'lagsllar Proteinr
1.3.1,2 Lipopoþsaccharide (LFS)
1.3.3.3 Hsmagglutinins (Htu)
1.3.3.4 Pili¿finbriae
1.3.4 hoduction of enterotoxin and fluid accumulation
1
1
3
3
3
6
7
I
8
10
11
1.3.5 Patholog ""'13
1.6 Ïleatment .... ""
1.4 ClinicalManiÍesations ....;... "" ""'13
1.5 Diagnoois 14
I
15

1.7 Epidemiology.. " " '16
1.7.1 lncidence " " '16
1.7.2 Aqrntic¡eservoi¡s ....17
1,7,3 Ihansmission "" "'19
1.8 Potential protective antþns of.V eholsac ' ' ' ' ' '21
1.8.1 LPS " "21
1.8.2 Non-I-PS antigens n
l.8.Zl OMPB B
I.&ZZ Flagellar (þ antþens - . . -24
l.8.Zg HAs. ............ ..U
l.&24 Pili ...25
1.&3 CT. .... .-..25
l.g Immunig ûo cholera . . .n
1.9.1 SystemicantibodyresPonsesto?clpbæ " ""n
1.9.2 Developoe¡tofintestinalslgAantibodyrcsponscs . .' .'D
1'9'3 sIgA anti-v cholqæ resPonses disPl^ty immunologiøl rcmory ' '32
1.9.4 Mechenism of sþA antbody-mediated protectbn eg¡'i¡st
Vetøbæ " "32
1.10 Vaccination asainst cholera 31
1.10.1 Pastapproachesûor¡accination . . . . . . . .34
1.10.1.1 Conventionalkilledvacci¡es ....31
1.10.1.2To¡oidvacci¡e¡.... ...35
u

l.lL,zGureotapproachestovaccination"' "'3ó
LlO.Zl Live attenuated oralvaccines ' ' ' '%
l.lO.Z2 Non-viableoralvaccines ' '38
1.lO.L3 Live oral Cholera/Ilphoid hybrid vaccine 39
40
4l
1.10.4 Conclrubn ....
1.11 Aims of Thesis
2 Materials and Methods 42
ZilBacterialstrains "" "" ""'42
22 lvlairtenence and growth conditions ' '42
Z3 Specidizæa growth media
21 Chemicak, reagents andbuffers
43
Z1,l Enz¡rme-linked immunoeo¡ùent assay (ELISA) Bufrer¡ 45
1z4.2 DetergenB """" ""46
1z1.3 Enzymes ¿¡d immunoconiugates ' ' ' 6
ZS d¡imal¡ "" "ß
u
47
2;5.l Adult mice
2,5.2 Infantmice ' ' ' '
z6 heparation of boiþd a¡d formalin-killed organisms
Z7 IsolationoflPSfuomKùol¡¡ræ.. "' ".. "'48
L8 Preparatio¡ of OMh 49
zg hoduction of a¡tisera towhole cells and to oMI)s olv úoleræ 49
2:g.l Preparation of mouse antisera
47
lu
49

I
úñ
29.2 hePa¡ation of rabbit¡ntisera 50
1ZIO Absorptionofantisera ".. ""'51
2:l}.l AbsorPtionwithlPSorOMP ' ' ' ' ' ' " " ' ' '51
ZlO,2 Absorptionwithbacteria ' ' ' ' ' ' ' ' "' ' ' "51
zll h ví¿.osssE¡B ol.V ùotsæadherence: ettachment to isolated murine intestinal
I
I
I
'tII
I
I
I
I
epithelial celb (enterocYtes).
Zll.lheparationofenteroc¡rûes "" "'51
2ill.z Invífroenterocyteadherenceassãy ""'52
Zill} Inhibiti'onofadhe¡ence" "" "53
1Zl2btwruassaysq¡itlinfantmioe "" ""'53
ZlLl VirulenceasEsyl "" ""51
51
I
I
Ll/2 Protectb¡ assrys
?Àl3 l¡olationo\VclPloæFlt. """55
?Àl4 btyito tihation of a¡tibodies toV ctpbæ ' ' ' ' ' 5ó
2ill.l Hemagglutination ' ' ' o ' ' ' ' ' ' ' ' ' ' ' ' '''' '' '56
214.2 ELISA 56
2¿IS SD5-PAQEandimmu¡oblotting .... "'57
zlS.lsDs-PACE "" "'s7
215.2 Immunobloü'ing(Westernblotting) " .. " " '5t
2|16 ColonYblotting .." ""'59
?Àl7 Constructbtof Vdplcrægenebank ' ' ' ' ' ' ' ' " "59
Ll7.t Prreporatio¡ of'V dtobæ genomic DNA
tv
59

f--
Lt7.2 DNAquantitetion . . . . . . . . . . . . . . . . . . .60
2,17.3 EndEllhgwithKþnoq,fragment ........ ..@
L17.1 Conetructbaof pPM2101 . . .60
2,17.5 Co¡¡tnrctbn of coemid gene b¡nt 6l
217.6 Mobilizationofcoomidbank .....61
2,17.7 Tlansformetb¡ """"'62
2l8 EM.... .... .."62
2,l&l ïhan¡micsion EM " ' '62
Ll&z Immunoelectronmicroecopy(IEM) .....63
3 Non-LPS Protective Antigens shaltd by Classical and
El Ïbr blotypes ofv cholerae g3.1 Introduction .... .... ........ ..""'U
3,2 Ç\anrcærization of.V døbæ gü'aine 65
33 The exbænce of non-LPS protective antiçnr in strai¡s C"{401 a¡d C4411 . . .67
33.1 hoduction of a¡tbera to 5698/165, CA1Ol and C4411 otraÍ¡s . .67
ï3.2 Protectiveactivitiesofantisera .... ....68
9.4 Disfibution of non-LFS pmotective aatþns among V clølqæ shain¡ . . 70
3.4.1 Oldlaboratory¡trains .... ........ ..70
3.1.2 RecentfeHbohte¡.... .... ...70
3.4.3 Conclusion.o.. ........ ...' ""72
3.5 Subseg of non-LPS protective antþns are serotype-restricted
Y
72

3.5.1 Demonstration þ protoctio¡ a888ys ' '72
3.5.2 Demo¡stratio¡ bY IEM ....73
3.6 Correlation between protective activities of antfue¡a and their capacities to
inhibittt víto attæ}¡rent ' ' '76
3.6.1 Invifreenterocyte adherence assry " " "76
3.6.2 Inhibitionof adhercnce " " '76
3.7 Discussion 78
4 Role of TCP as a Colonizatlon trbctor and a Non-LPS
Pmtective Antigen in the Infant Mouse Cholera Model &i
4.1 Introduction ........ "" "" "'83
4.2 Sûrdies ofVcløbæOMPs . . . . .. " " " " " " " "&5
85
85
4.23 OnPV .... "" "90
1.g F,rpressionofpiliþVcholqæstrai¡s ' ' ' ' ' ' ' '''' ' ' '''91
4.3.1 Preliminaryexperimente ' ' ' ' ' ' ' ' ' ' " ' ' ' '91
1.g2hoducti'onofTCP """" """'y2
4.3.3 Distributi'onofTCP .." ".."""'94
4.4 TCP is a vinrlence deærminant i¡ the infa¡t mouse model 98
45Auenputopuri$TGP ..""" "" ""101
4.6 TCP b a protective antþn i¡ the infaDt mouse model
4.Zl A¡at)'sb by SDS-PAGE a¡d imnu¡obloüing
1.22 Dete4ent e¡trætion of OMP preparations
vr
tuz

4.7 hotection þ anit-TCP a¡tibodies b biotype-restr¡cted 1(b
116
4.8 DiscNlssion .... .......' "'16
5 Molecular cloning of genes involved in bioqynthesis
Of TCP rtz5.1 Introduction .,,.112
5.2 heparationofcl,oneprobingeenun- ..... ... ' " " 113
5.3 p¡slimin¿¡ycbningeryeriments . . . . . . . . . o . . . . . .''' 113
5.4 MolecularclorringofçnesinvolvedinrcPpmduction ..... f15
5.4.1 Cloningstrateg . . . . . . . . . . . . . . . . .''''''' 115
5.4.2 ConstructionofthemobitizablecamidvectorpPM2l0l . . . . 116
5.4.3 Cloningthe TCP gstres ntoVcløbæ Ol7
5.4.4 Detectionofpositiveclones .... ..... ' " ' llt
5.5 Anatys¡softhecl,onesDsl-Dsl0 . . . . . . . . . . . . .''''' 118
55.r EMstudþs ................ .... ...llt
55.2 tmmunoblottingara$Pir IB
553 Moþcular analysis of cloned DI'IA ta
5.6 Confirnåtion of TCP a¡ avirulence determina¡tand non-LPS protecdive
antþen of,V dwbæ t?ß
5.6.1 TCP ¡ß aviruþncedeærmina¡t t?ß
5.6.2 rcP is a protective antiçn l?ß
5.63 rcP-producing O17 clones are imnunogenic
vu
r29

5.? Disct¡ssion . . . . . . . . . . . . . . . " " " " .. "' " "lgz
6 Genenal Discr¡ssion 135
6.1 Distributionof non-LPSprotectiveantigens ofVdplqæ . " ' 135
6.2 Role of TCP as a cobnization factor and a protective antigen t%
6.3 Molecular cloni¡g of genes invoþed i¡ the syÌthesis of TCP . . . 138
ó4 Is TCP the onþ non-LPS protective antigen of V ùolaæ il the ínfant mouse
model? .... .... ."' "139
6.5 Non-LFS protective antigeoo: ¡eler¡mt studies in otùer laboratorie¡ 139
6.6 Ftturestudig . . . . ' ' ' ' ' .. 142
6.6.1 Non-LPS proæctive antigens of Et Tor V døbae 142
6.ó.2 The nature of TCP-mediatedattachment 143
6.6.3 Vacci¡e Potenrizl of theTCP """" ""1ß
7 Bibtiography t47
vrlt

List of Ïhbles
3.1 Charæterieticsof Vdplsæstrains ........ .... "6
3,2 hotective ætivities of LPS-absorbed anti¡era to CA4OI a¡d C4411:
effect of ñrrîher abeorpti'ons with live homologous vibrioo 69
3,g Distr¡bution of shåred non-LPS protective antþne amongciÍeen streins of
g.1 The erbtenoe of eerotype-restricted non-LPS protective antigene inVùolqæ .71
4.1 DisffiutionofTCP . . . . . . . . . . . . . ..' ..' .. o'' "'n
4.2 Ant¡bodiestoTC?areprotectiveintheinf¡ntmousecholeramodel ......101
13 Antibodies to rcP are protective only ngainef challenge strains of
ClæsicalbiotyPe tgl
5.1 Prroductio¡ of TCP enìences thevirulence of Vúolqæ Ol7 tn
5.2 Absorption with TCPqrøbngOl7 clonee nemoves protective antibodiee . , 18
5.3 A¡tibodies to TCP are protective i¡ the infant mouse cholera model 130
5.1 TCP-pr,oducing cloner ate imrnunogeaic and protective 131
rI

List of Figunes
3.1 Demonstration of (Ogawa) sefotype-opeciûc non-LPS determi¡ant¡ 75
3.2 Antibody-mediated inhibition o|.V. dplaw adherence ¡n víOo 77
3.3 Capacitbs of th¡ee a¡tbera to inhibit the r¡ vútro aüachmeatof.V. cWæ
¡Eain¡ofClassicslorElTorbiot¡pe . . . .''''''''''''''''''79
g,1 Capacig of antberum to strain C4411 to inhibit the ø vùto atlæ\ment of
¡eoent%.ùfuæñeldi¡olatesofElTbrbiotype " " " '."80
1.1 SDS-PAOE and immunoblouing of V clpbræ OMPs 86
4.2 SDS-PACE aaaþeb of deærgent treeted V. ùfuac 5698 OMP preparationr . 88
l.g rmmunoblotanal¡nis ol? elplqæOMPswith a¡tiSark0MP a¡tisera . . . . .89
1.4 h,oductio¡ofTCPonCFA . . . . . - . . . . . . . .'''' "'''' " " "'yl
45 hoductionofTCPonT0G .... """'95
1.6 V chobæ 5698 grown o¡ CFA at?S C reacted with LPSobso¡öed a¡ti-O395 .96
1.7 Immunobbtting anaþb of V úolsac stfai¡s for TCP erpression 99
43 Immunoblottinganaþis ofTCP expression þvarious strains oî,V dPlaæ . 100
4.g Imm¡¡egoldlsbellingof%ctplqæ 5698 ' " o " " " " ' 1ß
5.1 heparationofcloneprobingoerum "" '114
I

5.2 ConstructionofpPM2l0l . . . . . . . . . . . . .''' "''''' 1f7
53 Colonyblotting ".. '119
5.1 Pr,oductbnofTC?þcloneDS2. ...... '. ' o.. "ln
55 Pr,oduction of TCP þ cloneo fF5 and N6
5.6 Imnunogold eþctron ntrccopy of TCP produced þ chne DSS ln,
5.7 rmmunoblot anal¡nis of whole celh ta
5.E Resr¡ctbn endonucleæe analyris of pPM2103 t25
6l hopæ"d organization of the gpDes associatedwith tùe production of TCP 145
tzt
Il

Introduction
1.1 Historical Backgrtund
-..bhæ lips, a únnl<øt fæ øúhotlou, ey,et, a løottcdbclly, wíîh limbs crønped øú ctPbd
æ tf by fvq breath clhgtts ø the wøtíor's face; f*W thA øe profu4 stalE aü eJav'ed; ød
h wìthíngs ihe palíenl qþes, thevìctím of Sìv*
Insrlptton atVldannusEer Ibmple (cited in Vougþt 18fB)
çholera (orrruneeas describedabove) ie a disease of antiquig recognieed in ancientlndian
nedical liærature. It probabþ originated in the deltaic regions of the Gaoges and the Brah-
maputra in the staæ of Bengal (non, knou'n as lvestBengal [ndia] and Bangþdesh) where for
cenürries it has beeq and still iq endemic, During the late fifteenth century Portuguese
explorers gare vivid descriptions of cholera outbreaks on tle l¡dian subcontinen$ hou'ever,
the disease did not spread wertwards until the nineteenth cenhrry (see Follitzer 1959).
In 1817, cholera broke outwith a hiÈ mortatity i¡ its endemic home; and in the next
fnre years, it spread over much of Asia and tle Middle East By 183\ the disease had spread
to Europe and America (Roeenberg 1962; Morris & Black 198Ð. From 1817 to l9/3, sa'
pandemic waves of cholera epread acnoss the world. Reliþue piþinagesr the mor¡ement of
I

troops and the sea t¡ade ¡oute centered around the endemic home all contributed to the
dissemination of the di¡ease (revien'ed by Pollitzer 1959).
During this period, two important obeervations we¡e made that greatly advanced the
understanding of the disease: one regarding transmission and the other etiologr. In 1855, Joh¡
Snow published an important and influential treati¡e on cholera epidemblogr (Snow 1855).
He clearþ established that the disease invoþed fecaloral transmission and discussed factors
promoting rhis mode of dissemination, higbliÉting the signiñcance of water (ûo be discussed
furtherinSection 1.7.3).Su@uentþ,Kochconfirmedthatcholera\r¡asindeed"anaffectation
of tle alimentary canaln (Snorv 1355). He identified the causative agent as a bacilluq
Wbrío cholaæ,which he cultured from the intestinal contents of choleravictims (Koch 1883'
1884 - cited by Polliøer 1959). This confirmed earlier obeervations made in 1854 þ Pacini of
the association of the diseåsewith vibrios (Hugh l%4).
In 1905, Gotschlich (cited þ Pollitzer 1959) isolated a new strain of.V cholcræ ftom
the cadavers of p,ilgrims rehrrning from Mecca to the quarantine cåmP6 at El Tor on the Sinai
Peninsuþ FryypL This strain proved to be different from tìe prenious lClassicaF strains and
was assigned a nelv biotype, El Tor. From ly8 tþ 1f)61, cholera was onoe again reduced to it¡
endemic form being confined mainþ to tìe B"og"l region, with small foci of endemicig
reported in countries to the east of India" I¡ 193? "paracholerC caueed by V cJølaae (El Tor
biotl'pe) was obeerved by de Moor (1933 - cited by l(anal ln4l in the Celebes from where it
gzve rise to sporadic câses in adjacent countries over the ¡ext 20year&
In 1f)61, the ser¡enth (and current) pandemic began. Unlike prer.ious pandemics, this
was caused by vibrioe of El Tor biotype @arua & CVjetanovic 190; tr(anal lnÐ.It begaû i¡
Sulawesi [Celebes] (I(anal ln4),the endemic focus described Ð d" Moor (de Moor 1938 -
cited by lç¿nal lnÐ and quickly spread to the Paciñc islands, across Asia into the Middle
East, the U.S.S.R and Africa, reachingthe North American continentby 19/3 (Weissman'd d
2

ln4).other than the united stateg no countty in Norú, South or central America reponed
cases until 19g3, when cholerawas detected in the car¡bbea¡ Coast of Mexico (Black 198ó).
The spread of the disease during this current pandemicwas facilitated by th" greater mobility
of people todây and B mnqs 6igation of refugees'
12 EiologrL2.L Ctaracterbtics of the bacterium
V cholqaeis a gram-negtive comma-shaped bacillus which possesses a sheathed polar
flaçllum of variable length. In addition, undercertai¡ culü¡ral conditions, the Presence of pili
(or fimbriae) on the organism has recently been desc¡ibed (freedy el aL 1968;Taylor et aL
lggT;trlall etøt lggg). The bacillus is a non-spore-forming aerobe or facultative anaerobe-
Biochemically, itis recognisedbybeingoxidase pooitive; itfermenb glucooe (withoutgas) and
sucrose, but not lactose; it utilizes citrate, þine and ornithine but not arginine; it is Voges-
proekauer (v") p*itiue a¡d b seDsitive rD ÙnD (Pterirtine) dito (150r¿g).
LLz Classiñcation
l¡ l71g,the çnus name Wbríowas given by Mäller. Su@uentþ, the species name'
choløæ,was given in honor of pacini¡ the fi¡st to give a vali<l description of the organism ("it"d
in Gallut lnq.The çnus Víbrbtw contains, in addition loV choløæ, otlervibrioo recently
upgraded to species level (see below)'
V chobæ strains are classified on the basis of the production of a cell wall potyrac-
charide antigen designated 01. Trro different classification systems for o gfouPs are used
(Safazafi ¿ $himada 1977;Smith lnÐ.In bolh systems O-goup 1 (Ol) strains afe respoD-
sible for true cholera infection The Ol vibrios can be divided into two biotypes, Classical and
3

El Ibr (Bauman d aL lgu), which can be differentiated in several ways: the latær can
aggfutinate chicken red blood cells (Finkelsæin & Mukeriee 19í3), and is resistant to
polymlmn B (Gangroo a 4 ú l%7) and to Mukeriees's type IV cholera Phage (Mukerjee
1963). Each biotype is further differentiated into antigenicalþ related sero$peq lnrh4 Ogawa
and, rareþ, Hikojima, based on the Prex¡ence of 3 antþenic factors (À g, & C)' Ao B and C are
foundinbothogvvlarandHikojimawhereaslnabaPossessesonlyAandCantþens@edmond
teTe).
Vibrioo thatproduce pot¡'saccharide antþens other than 01 are commonþ referred to
as NAG (non-agglutinable) vibrioo or NCIV (non+holera vibrioo; revierred by Finkelstein
lg1¡).NAG is a misnomer since, althougþ these organisms are not aggtutinated by 01 sera'
they do agglutinate in homologous antisera such strains produce diarrhea indistinguishable
from cholera (Morris & Black 1gs5). Some of the NAG vibrios causingenteritiswere upgraded
to species ler¡el on thebasb of DNAhþridization studies;in 1985, identifiedspecies included
v pøahaanotyticys,v nim¡aß,u ltilvialis an6v holTuae (Blake 1980; Morrb & Black 198Ð'
13 Eathogenesis and Fathophysiologr
Cholera is an enterotoxic enteropathy. The causative bacteria are strictly non-invasive,
remaining confined to the lumen and epithelial surface of ttre gut The essential pathogenic
events invoþe: (r) ingestion and entrance of viable v cholqæ into the sm¡ll intestine;
(2) colonization of the epithetial surface; (3) multiplication and release of enterotoxin and (4)
hypersecretion of isotonic fluid by the intestinal epithelium in resPonse to the enterotoxin'
4

1.31 Ingestion and infectious dæe
Following ingestion in either water or food V cholqæ must survive the acid environ-
ment of the stomach, the first line of defence in the host It is claimed that infectious dose (ID)
is high in healthy adults. Studies with Western volunþers have suggested that 108-1011
organisms are required to cause diarrhea n 50Vo aduls (IDso; Hornick et aL llTl;Cåshet aL
lyl4+). The sþificmce of gastric acidity as a barrier to choleraic infection is revealed by
studies which have shown that prior neutralization of gastric acid by ¿rlministration of sodium
bicarbonate dramaticalþ reduces the IDsoto td-fO6 orgaoisms (Hornicket øL llll;C-ashef al-
lf'l4c;Irvi¡e et al l98lb).
The IDso ¡motrg the inhabirants of endemic areas ¡spains unknovn, butmalnutrition
would be expected to reduce the efficacy of the gstric acid barrier. In such areas individuals
with achlorhydria are predisposed to this dise¿se (Gitelson l97l; Sackef aL 1fi2). The ID is
also lowered if vibrios are ingesæd in contaminated foo{ which presumabþ protecb the
organisms during their passage through ¡¡s stomech (I-evine A aL l98Lb)'
L3;Z hnehation of the nrrcus laYer
The viable organisms which reach the small intestine encounter two additional non-
specifichostdefence mechanisms; nemeþ, intestinal peristalsis (Dixon 1!)60) and a mucus layer
(Florey 1933; Freter & Jones 1976; Schr¡nl¡ & Verwey 1976) which the organisms mut
penetrate in order to reach the epithelial surface. V cholsae useE an arrãy of virulence
properties in overcoming these barriers. The flagetlum is responsible for motility and allows
the bacteria to respond to chemotaxins such as L-aminoacids and eimple Eugan¡which diffuse
away from the mucus surface (Freær & O'Brien 1981a, 1981b; Freter er aL l98l).
5

Several workers using animal or m yitro models have demonst¡ated that motile
V choløae can be raprdly detected in the intervillous spaces and crypts of the small intestine
(Guenrzel & Berrl' 1975; Nelson el aL 1976: Schrank & Verwey 1fi6; Freter d aL l98l).
Mutant strains that are non-motile but enterotoxigenic colonize these regions much less
efficientþ and tlerefore eúibit reduced virulence (Guentzel & Berry 1975; Schrank & Ver-
wey lgl1;pierce et aL 198f¡. Elect¡on microscopy (EM) studies have shou'n vibrios adherent
to enterocytes as well as associated with mucin (Nelson et aL lÍ1.6: Guentzel er 4I l98lr.
]b gAin entrance into the mucus gelcoveringthe epithelial surface,V clølerae releases
a potent mucinase @urnet & Stone 1947; Burnet 1949; Freter 1955b). In vìxo sh¡dies have
indicated that the forward movement of vibrios is impeded þ mucug although the organisms
can migrate through channels in the mucus gel (Jones et aI 1176). It is suggested lhat,invíw,
such channels result from the action of the mucinase. Various studies have suggested that a
soluble hemagglu':nin poosessing protease activity (see Section 1.3.3.3) mig[t cont¡ibute to
virulence of V cholqæ (revierved þ Booth &, Finkelstein 198ó), possibþ þ fulfilling a
mucinase function hvivo (Schneider & Parker lfi&;Finkelstein et aL L983).
L3.3 Adberere to the epithelim of the smll int€stine
Adhesion Becure6 tle bacteria against the effects of intestinal perisølsis and facilitates
delivery of the enterotoxin- The precise molecula¡ mechanism þ which tle organism attaches
to enûerocytes of the crlpts and villi is not entireþ clear. A number of cell surface components
(adhesins) could be invoþed These include flaçllar sheath proteins and otler flagellar
antigens @ubanlc et aI ll77; Attridge & Rorley 19334 1983b), lipopolysaccharide (Chitnis
et aL L982a,1982b), hemagglutinin sandcholqalecth (Finkelstena aL 1983), and pili (Taylor
et øL 1987; Herrington et aI 1988).
6

L33.1 ffagellar PFoteiß
The¡e is little doubt that the ftagellar structure plays an important role in the coloniza-
tion of V cholqae. Studies in the infant mouse model shoç,ed that motile strains were mone
virulent tlan non-motile derivatives obtained þ mutagenesis (GuenEel & Berry l9$, and
microscopic observations of intestinal sections shouted that this difference correlated with a
more effective colonization þ the former (Guentzel el aL lW). I¡ a laær etudy, mixed
suspensions of relatedmotilityvariantswere fed to infanf 6i66; and the motilebacteria sho$'n
to enjoy a significant advantage in terms of intestinal colonization (Attridæ & Row'ley 19834).
These reports are supported by numerous ìn vitro sh¡dies which show that motile vibrios attach
more efñciently to avariety of substrates (Guentzel & Berry lflS;Freter & Jones Ll16;Jones
& Freter 1976;Attridge & Rowley 1983a' 1983b).
Whetherthe invoþementof the flagellum extendsbeyondits roleas an agentof motility
remains uncertain, althougþ reports suggest that tìis stnicture also pla¡n a role in the Process
ofattachment Jones & Freter Qnqshowedthatf,agellatedvibrios couldbindtobrushborder
membranes h vítobutthat non-motile orqanbms could no! oren if they were compacted onto
the sub6trate þ ceatrifugation. Attridge & Rowþ (19S3a) reported that flaçllated vibrioo
which had been immobilized by preincubation with antibodies to LP$ retained the capacity
to adhere to seÊments of int€stinal tissug whereas non-flagellated orgnism.s were non-ad-
herenL The onþ aprparent difference between the two populations wÍts the presence of an
immobilized flagellun, suggesting the exisænce of a flagellar adhesin In this respect some
investþtors hãve described a flagellar sheath protein (Follet & Gordon 1fb3; HtaÃitzky eJ aL
1930). Antibody directed against rhis protein aPPears protective against chole¡a in two enimel
modele; and it has been suggested that thb protein msy serve as aD adhesin (Eubanh a al
lf17;yancey d aL lg7g). Nonetheless, EM sû¡dies indicate thatvibrios do not attach þ their
7

flagellum - the flagellum sticks into the lumen of the gut (Nelsotet al.ly76r.It could be that
the initial contact is made by th" flagellum and the organism then adheres horizontally to allo*'
greats sufface contacL
L33.2 LþPol¡naccbaride (LPS)
Some evidenc€ has accumulated to suggest that LPS may Play a role in adhesion of
Vcholsæ to intesti¡al mucæa Using the rabbit ileat-loop model Chitnis and co-workers
(1982b) have demonst¡ated that purified LPS prepared from an Inaba süah o1' V ùobae
(5698), but not from .Escå qíchia coli, car- signiñcantly reduce the attachment of other Inaba
strains to the mucosal epithelium. Furtlermore, serreral workers have shown the role of
antibodies to Lps (ogara and Inaba) in the inhibition of attachment of V ehobae strahs to
intestinal mucosa both nr vútro atd h víw (Freær & Jones 1fl6; Chitnis et aL 198241987b;
Auridge & Rovley 19g3c). Iåstly, Finkelstein and co-workers, using monoclonal antibodies
(MoAbs) direcæd against v ùolqae LFS (preumabþ directed against specific o-antiçn
determinants) were able to inhibit the hemagglutination reaction caused þ vibrios or LPS
(Booth er aL 198,6).
L333 He'mgglut¡nins (IIAs)
Alrhougþ hemaggfutination systems have been wideþ used in the study of Vcholaæ
adherence, the mechanisms of attachnent to erythrocytes (RBCs) and to intact mucoeal
su¡faces may differ (Attridge & Rowley 19s3b). Before usingRBCs aß a convenient subetituæ
for a more relevant substrate, it is therefore important to initialþ confirm that the charac-
teristics of aüachmentare simíla¡'in the two syntems (Jones el aL ll76l'Jones 1980)'
Vchfuae aggtutinate certain species of RBG ¿¡d rhis proPerty correlates with ad-
herence to brush border membranes (Jones & Freter ln6). Adhesion to the i¡testi¡ el
8

epithetium and hemagglutination may be inhibited þ different sugan (Jones & Freter lnq.
It is now known thatVcholqa¿ strains produce four distinct HAs which ca¡ be divided into
two classes: cell-associated and soluble (Hanne & Finkelsteh 1982). Three cell-associated
HAs have been described so fan a D-mannose+ensitive IIA (MSHA) produced by El Tor
strains; an L-fucose-eensitive HA (FSHA) e:ryressed þ Classical strains; and a mânno8e'
fucose-resistant HA (MFRIIA) expressed þ both El Tor and Classical strainß. The fourth IIA
is solublg po6se5ses protease activity (SHA/protease) and is found in all strains regardless of
biotype or serotYPe.
There is some evidence that cell-associated HAs of Vúobæ a¡e invoþed in ad-
herence and may represent adhesins (Guinée el aL 1985; Finn d 4I 1987) althougþ a direct
correlation between intestinal attachment and tIA e,ryression has not ahva¡rs been seen (Freter
&Jones 1976;Tep,pema daI1987).Morerecently, hvinoadherenceotudies olVchobaeto
formalin-frxed þ¡men villus cells (Yamamoto et aI 198ß) or human mucus coat (Yamamoto &
yokota lgSS) harre shon¡n that adherence did correlatewith cellular HA levels' irrespective of
bio$rye or serot¡pe. Similar inhibition patterns were obtained between adherence and hemag-
glutination reaction" indicatingthat cellula¡ [IAs -iÉt plr,y a role, at least i¡ Parl, in adherence
(Yamamoto et øL lgæ). The relerrance of these studies to the pathogenesis of the disease is
not knorvn. Finn d aL (1987) iÊolated an MSHA' muta¡t from a V ùolqae strain JBK70 (a
muta¡tþl Tor Inaba strai¡ lackingthe cholerafor gene),and the mutantwas much lets able to
colonize the ileal mucæa folloving oral administration to rabbin. This suggess that the El Tor
MSHA might be considered as a putative atlachment facÛor.
The SHAlproteåse, prwiousþ referred to ascholsaleãh (FinkelsteinA aL 198)' has
no*, been purified and charactqnzà (Finkelstein & Hanne 1982). Itwa¡ originalþ thougþt to
be an adhesive factor as it inhibited the attachment of vibrios of either biot¡pe to epithelial
ce[s both ínvitro and lr¡ urvo (Finkelstein d aL ln$). Hox,orer, there ie, at prcsent no con-
9

vincingevidence thatitcouldbe an adhesin.Althougþ tìeabove studiessugEp8tthate)rytression
of HAs could promote bacærial adhesion h vítro, it remai¡s to be seen whether any of tlese
HAs are operating as adheshs during infection. As di¡cussed belw, pili hære recently been
described onV choleræ(rrylor er øL 1987) "¡d
it is unclea¡ whetler these are associated with
HA propertiee as has been described for other bacteria pcseeeing adheeive pili on their surface
(Duguid & Otd 1980; Gaastra & de Graaf 1982}
1.33'4 PiVñdrhe
pili are non-fþellar, filamentous protein appendages proiecting outward from the
surface of many gram-negative bacteria and are aEsociated with the colonization of the natural
envi¡onment (Brinton J. 198; Smith & Linggood l9ll;Nagl d aL lffr' Theee appendages
u,er,e independentþ described and introduced as fimbriae þ Duguid el aI (195s) and as pili
by Brinton (1959). Evidence b accumulating to suggeot that the acüral adhesins of these
bacteria consist of minor protein molecules at the tip of surface Pili- This has been
demonst¡ated in both enterotoxigenic (ETEC) and uropathogenic strai¡s or' E coli
(Gaastra & de Graaf 1932). þ analog, it har therefore been assumed that pili would be
inrrohed in the pathogenesis of cholera but pili erpression by v ùolsa4 althougþ the subiect
of muchdiscussion,led to conflictingreports in thesifief ardseventies(Finkelntein & Muker-
iee 1fb3; Barua & Chatted æ1964;I-ankford & Lægsombunaûa 1%5; fteedycr øl 1!)618; Nel-
sonef aL lnq. But in the eightieg reports have appeared Gonfirmingpili expression þ
V cholsse(F.rir et aI 1982; Al-Kaissi & Mosl¡atos 1985; Eha¡a d al- 1986;kyh¡r el aL 1987)'
To date, four types of pili of V cholqae h¡ve been described" Ehara et aL (1986)
described flexible pili on V c)ølsae during the colonization of the upper emall intestine in
rabbits. This pilushasnou,been purifiedandthestnrcüualsubunitis aprotein of 16lDa (Ehara
e, aL lgg:¡, lggg) shosrn to be shared by 01 (both El Tor and Classicat strains) and non01
10

V cholsæ (Nakasone & Iwanag 1983). Antisera from convaleecent patienß contained an-
tibodies against the pili subunit whilst anti¡era raiced against pili subunib neutralized the
hemagglutinating activi$ of pff preparations @hara d aL l9æ).
Recent reports have described the discovery of a pilus stn¡chrre on the surface of
claseical ogawa 0395 (Taylor d aL lg87) confirmed by Hau d aL (1988). Thb pilus is now
refer¡ed to as a foxin co-regulatedpilus (rcÐ. In addition, Hell d aL (198) ha*'e descriH
thatVcholqæcan eryress two other morpholog."lly distinct pili besides the TCP; nameþ
Type B with a wavy morphologl and Tirpe C being more rigid" Whether or not ary of these two
t,¡pes a¡e rel,ated to or identical to that described by Eh* d aL (19%. 1988) remain¡ to be
established. The significanoe of theæ latter t)"es has yet to be elucidated" There is compelling
er¡idence that rcp may ble an inporønt coloniz¿tion factor oÍ,.V dtobæ. This significance is
discussed further in subsequent chapters'
L3.4 Pnoductbn of enterotodn 8nd f,tdd accumilation
Afær successfuþ attaching to the enterrrc¡rtes ,t\eV cløbæ organisms enter a phase
of active grcsrth, duringwhich they elaborate an enteroto.in^ This molecule comprises two
q¡pes of subunits, A a¡d B (see Section 1.8.3), the latter mediating bindiag to the enterccyte
membrane (Cuatrecasas lg73) via a receptor now identiñed as a eialidase-resistant
monosialosyl ganglrooide (GMt) (van Hgrningen et aL llll: King & van HeÏningen 1973)' It
is the otþsaccharide portion of the ganglioside to which the toxin binds and there is a
correlationbetween GMlcontentandbindingabilityof cells (Holmgren 4 aI1975æ,Haneson
et aL lgi'\. Once attachment has occurre4 the A eubuni! aD elu¿yme, then passe6 acroes the
cell membrane into the cytoplasm and activates the enzyme adenylate cyclase- This in turn
lea& to elerrated intracellular lerrels of cyclic 3'5'-adenosine monophcphate (cyclic AI\'ÍP)'
resulting in an irreve¡sible hypersecretory response @eærset d aL ln4 Holmgren ef øl
,l
{
11

l91Sa)by ".llr
i¡ the crypt and on the sides of the vifl The secreted fluid is low i¡ protein and
rich inelectrolyteincludingNât, Kr, Cf & HCol-. Althougþ thetoxin is rapidþboun4 there
is a lag period rangng from 10 minutes to t hour before changes in ion trnngPort are seen This
delaly probabty relates to tle -:me taken for the A subunit to penetrate the cell membrane
(Fishnan 1980).
h both clinical (Greenougù 1fb5; Banwell d aI lnq and experimental (Cår-
penter d aI lgæ) choþra, the fluid loss occurs from atl segments of the small bowel with a
decreasing gadient from duodenum to ileum. The resultant loss of large volumes of isotonb
fluid is responsible for the cha¡acteristic clinical Fofile of the disease. As mentioned below,
tlere is no æidence that any stnrchrral lesions ale producedby the toxin
It has been propced that the action of cåolera toxin (CT) promotes the intestinal
growth olVchobæ þ meking arr¡ilahle nut¡ients secreted þ epithelial celh (Mekalana
19SÐ. Construction of site+ piÍrcc9muta¡ts has provided tle moot convincingertidence rhet
toxin pnoduction i¡ beneficial for gron'th in the i¡¡6tinel environment (Mekatanæ 198$
These ¡on-to-inogpnic mutant strains colonize rabbit intestines less efficiently than their
parental stain¡ (Pierce d aI 19æ[
Althoug! CT is taræly responsible for the voluminous diarrhea cha¡acteristic of
choler4 two observations suggeet V dtolqae 01 produces another toxin(s) capable of causing
diarrhea Fint, there b widence that naurratþ occlu:ingCFnegative El Tor strai¡s - whetler
envi¡onmental in origin or associated with cases of diarrhea - cau¡¡e intes-:nal secretion when
inoculated int¡o certain animal model¡ (Nishibuchi d aI 1983: Sanyal a aL 1983). Secondty'
mild or moderate diarrhea wal¡ Been in about 30Vo of healthy adults fed with viable 01
orgpnismq of either biotype, that were CFnegative; theæ weÍe strains whqe CT genes had
been inactivated þ nutagenesis or d€teted þ recombinant DNA techniques (Iæ',dne et aL
19SSa). The exact nature of non-CT toxin is not known althougþ it could involve the shig-like
.t
{
ù'
I
¡
'l
{i
':
'ftI
I
I
II
t2

toxin said to be p,roduced by many V cholqæ ()1 organirms (O'Bri et el d 1984). In addition,
El Tor strains have been shoü'n ûo produce a soluble hemoþin which has cytotoxic and
ente¡otoxic activity (Yamamoto 4 aL L986),.
L35 nrthotogr
The hoot cellular rcsponse to infection is minimal, invoþing primarily a non+pecific
increase in mucus-producing goblet cells. There is no maior histological rlamage to the
gasgohtestinal tract in either mnn (Gan gutxiacf aL 1%0; Fresh el aL l9&) or in elperimental
animals (I-aBrec d aL 1965; Norris & Majno l9i8; Ghosh lnq. Biopsy specimens collected
from different regions of the small intestine re't eal an intactepitheliun There is edema of the
eubmucosq dilated cr¡¡pts and minimal numben ef infkmmatory cells in ¡[s lamine p¡oPria
The intercellular tigltjunctions and capillary endothelial cells arc also normal The brush
border remains intact and the organisms do not invade the mucosa
L.4 Clinicat Manifestations
Ctolera is a digease rangng fnom as¡impomatic infection with mild diarrhea, to a
firlminantfatal syndrome, inwhich form itis one of the moetrapidþ fatalinnesses known. The
incubation period nay vary from a fen, hours to a fen' da¡R with or without prodromal symptoms
Acutg potentialþfatat infection b comparatinely ra¡e; the large majorityof ca8€8 are mild and
usualþ indistinguishable from other forms of Eastroenteritb.
In its severest form, tle disease p¡ogresoes from tle first liquid stool to shock i\ 4-12
hours with death foll,owingwithi¡ 18 hours. It b characærizÆd by the sudden onsetof effortless
vomiting and profiue dia¡rhea (Greenougþ lnÐ which initialþ lacb the 'rice-water" aP
pearance- As the dia¡¡hea progresses the fecal fluid takes on the typical tice-watef appearance
t3

due to the preeence in the gtool of flecks of mucus discharged by th" goblet cells a8 a result of
the action of the cholera toxin Dehydration accounts for the maiority of the signs and sympomr
seen i¡ cholera These include thirst, muscular ç¡¡mpor sunken eyes, cold extremities, thrcaÙ
pulse a¡d sunken fonta¡elles i¡ babies. In untreated severe cases a patientcan lose up to twice
hiß body weigþt in liquid stools (Hirschhon d aL 1968)'
Complicatione of cholera can occur in both child¡en and adults. Theee include alæred
consciousness which in adult is manifesþd ry a detached mental statq while in children
unconsciousness or conwlsions can occur as awarningof impendinghypogþcemia Electroþte
imbalance leads to hypokalemia and hnernatremia as well as acidosig particularþ in childreo
Aspiration with fatal sequelae is a serious complication in patients with a combination of
atæred consciousness and vomiting (Greenough 199)'
15 Diagnosis
The moet rapid and effective wry of diagnroeing cholera in the laboratory is þ examin ing
a fresh fecal epecime¡ under dark-field illumination The vibriq can be recognized by their
characteris trcdæt¡¡gmotilityuåen presentin largp numbe¡s. Addition of homologous antisera
çritt immobilize thevibrios and immediaæ diagnosis can be made @enenron er aL 1964)' When
stools cannot be examined at once they are best placed in a transport medium such as Cary-Blair
medium (c"ty & Bhir 1964) or Monsur's medium (Monsur 1963).
In the laboratory, the specimens are inoculated into both (a) a¡ enrichnent medium
(alkatine-peptone water pH S.a-9.2) and (b) a selective medium CrcBS agar; Kobryashlia aL
1963). Growth from enrichment medium is sub+ulûued onto the selective medium. All
putative V cholqae coloniee that are oxidase pooitive on the selective medium a¡e then
subiected to aælutination with a poþalent 01 serum before being characterized biochemi-
t4

catry. The confirmed isolates o1 V cholqæ can then be phage tJ¡Ped for epidemiological
purposes (Gla6s 4 aL 1983>.
t6 lleahent
The treatment of cholera is simple, both iû concept a¡d execution' involving the
replacement of the water and salts lost in the cholera stools. Replacement therapy in the form
of intravenous infr¡sion of fluids was shown to prerrent deaths from severe cholera as earþ as
1831 (Latta 1831-1334 O'Shaughnessy 1831-1332). Improrement in rehydration therapy
based on accurate Deasu¡ements of salt8 lost in the cholera stool was suboequentry propupt"d
as a ooffect and totalty effective tfeahent (watten d aL lg59). Whe¡ promptly apflied this
replacement therapy reduced the adult mortality rate to less thil o'e penoent of those ca'€s
se\refe enougb to warrant hæpitalization (Hinchhon daL lnÐ' Howerrer' thic form of
therapy i8 technicalþ difficult and expensive'
successful rehydration caû be more convenþntþ achieved by ü" oral administration
of fluids 6e¡rrini¡g gfucooe and ions of sodium, potassium, chloride and bicaôonate (oral
Rehydration TherapyoRll Nalin & cåsh rnq.The gtucose facilitates the uptake of water
and electrolytes by th" small intestine and helpe to retolve hypoglycemia whiht potasaium
prevents the establishmentof hlpokare-ia (Natin & cash tyTr).The oral rehydratbn solution
(oRS) reco'rmendedby the world Health organization hee undergone severar clidcål triat'
and has been found to be safe aûd useful in the treatment of diarrhea fron all causes in all
patient golrpô (Mahel¡nabis 1981; Mahalanabb & Merson 1986)' In nrral 6pmmunities of
Bangladesh, the oR.S has significantly reduced diarrheat morbidity and mortality (Rahaman
et aL l(/1g: clen d 4L lgw). tn the last 10 years, er¡idence has accumulated sho*'ing the
importance of nutrition during diarhea oRs Í¡ccompanied with feeding ha^q been ¡hou¡n to
ra¡Édty etabilize and then rever¡e the weigþt loes aûd nitrogen imbala¡ce (Inærnational study
15

Group lgll;Bron¡n 4 aL tngr,Råbbani 1986). In the worst cåses, ho*'evet, such as severeþ
dehydrated children who a¡e comatoeed with high natee of purgng and/or uncontroll¡able
vomiting ingavenous replacement therapy is required (Greenoug! 1980)' As soon as tle
patient is stabilize4 ORS can be instib¡ted. With prompt and adequate fluid replacementthe
patient usually recovergwithin a fent daya
Antibiotics arc often adminisæred to reduce the duration andvolume of diarrhean the
most commonþ empl,oyed being tetracycline, which kills Iz. ùolqæ leading to bacteriologi-
calþ negative stools after Zlhours (Greenough d oI 196/¡: Carpenter 4 aL 196ó). Hon'er¡er,
widespread use of antibiotics has led to the emergenoe of plasmftt-D€dhtêd drugresistance in
V. chotqæ (Mhalu d aL lg7g: Glass et aL ß8f,: Tabtieng et al- l99o), making it imperative to
look for alærnative meatrs of t¡eatmenL One such ap,proæh would be pharmacological
inhibition of toxin-induced i¡teetinal secretion, the most promisiag inhibitor being
chlor¡rrom¡zine. Howeter, its attendant sedative effect would limit iu clinital usefulness
(Greenougþ & Rabbani 1986).
L:l þidemiologrL7,l IncHeree
A¡nualworldrvide incidence rates of cholera has b€€n estimated tobe nearþ 8 million
cases resulting in approxim atley l?A,t00 deaths (Black 1936). In endemic areas, cholera has
the higþest attack rate i¡ children, with a peak incidence in the 2 to 9-year old age gouP
(Glass et at- l9g2). Attack rates decline rapidþwith age and the dis€ese b ra¡e afær the age of
40 (Mccormack et aL lg69). Thb distribution pattern is thought to ref,ect the induction and
maintenance of activs imm¡aity as a result of repeated s¡virenmentd expoeurctoY dtobæ
(Mooley d aL 196¡¡). Thir is supported by obeervations that, wüen cholera outbreaks occur in
16

non-endemic regions, individuals of all age group are equaþ susceptible @aine d aL lnû),
a finding consistentwith the abce¡ce of (agedependent) naürral immunit)¡'
Since the onset of the seventh pandemic, the e¡Édemiologl of cholera has been
changing Althougþ either biotype of V clnleræ is capabb of causing cholera in iß severest
form, epidemiological shrdies revealed that the infection to e€ryere cåse ratiowas approximateþ
ten times higber for the El Tor bbt''pe (Bart d aI ln0: Shahid d aL lg84.). Th" more recent
predominance of the El Torbiotype could be due ûo the fâct that these strainß can suryive longer
in water and i¡ food (Felsenfeld 19ó3, 1t)ó5) as well as in nig[t+oil samples (B^fter aL lnq.
In Bangþd€6h, the Chssicat biotype was repla"ed by the El Tor biotype i¡ 193; but i¡
Septembe t l9g\Classical choþra reappeared as the cause of a maior epidemic a¡d h¡s since
remained as the predominantbiotype (Samadi ef aI 1983;Kha¡ er aL lg84.). In Indi4 howaner'
the El Tor biotype continues to be the principal G¡ute of di¡ea¡e-
In endemic a¡Eas the disease has a seasonal occu¡retroe which differs not only in
different countries but also in areas which are very clooe geographically (Martin d al1969\
In Bangladesh before lg12thepeak for cholera (C1assicat) was pæt-monsoonal (DecÆnber)
but in lnSftshifted to Ocrober as the ClassicEl biotlpewas replaced by th" El Tor. tilith the
reû¡rn of the classi€l biotype (sanadi d aI 1983), cholera peab twice enn¡¿ll¡r, in Decem-
ber/January and in Arril/May (Cohrell et at l98l). Yet not fa¡ nfay in Catcutt4 the peak is
p¡e-monsoonal (l(han & Greenoug! 1935). The underlyingbasb for the s€åsonal i¡cidence of
cholera remains unknoum, but one poesible contributing factor is discuesedbeloç'.
L,7.2 Aquatic nesrcrYoin¡
taditionalþ, cholera epÉdemiologists coneidered that the onþ natural reservoir of
V úobæwas the hurn¡n intesti¡e, with only a brief perid of survival poesible outside the
human body (Felsenfeld lnÐ,Honever, epidemiologcal studi€s undertaken in Australiaand
t7

southern u.s..À suggest the exbtence of aquatic reservoirs rot v clplaæ El rbr and thereby
ofrer an explanation for the perftrdic occu,,enoe of chorera cas€s in the*e areas (Blake er aL
1gg0; Bourke et aL lgfl'j). It is thougþt that, tike otlervibrioq vchobæ maybe an autoch-
thonous organism of esn¡arine a¡d brackish waters (coh*,ell a aL lgn,1987; Hood & Ness
lgg2),or at least that the organism is able to peniet and multipþ in such an envircnment for
long periods of t¡me (N"lio 1976; Blake 4 4L lg80)'
6lnS,the fi,,t indþnous case in the Norti American conti¡ent in over sixty years
occurred i¡ Texas (weissman d oL tn4). Since then cases ha'e been detected on tle Gulf
Coast of Texas and Louisiaûa (Blake e, at 1980; ghandera 4 aL 1983), a¡¿ indeed thh b nory
recognized as an endemic area The mæt recent cases occurred in Baltinore i¡ 19&{ (uo' d aL
1gs6) and in Louisiana and F.lorida in 19g6 (cD.c lgsó). Epideniologicar erudies ha'e
aü¡ibured outbreaks in us.À (Blarrc d aL rgæ) and sardinia (salna6o 4 aL 19æ) to the
ingestion of localþ caugþt conaminated seafood" In both l,ocations tlere had been no cholera
in the preceding fve yearg, arguing ag¡inst the notion tl'et the disease is Bai¡tained by regular
transmißsion hom infected indivi<tuars to water. I*tead, they support the hypothesis of the
existence of an aquatic reservoit of'V clpbæ'
laboratory shrdies hæe shown thttv clplqæin pure culture can su¡vive for prolonged
periods in warm wateß without nut¡ienß wherein survival is influenced primarity þ salinity'
pH and temperaûrre (Singteto t4 al- tgï2l'l98ã¡:colwell ø øl 1985b)' I¡ such an environ-
ment v chobæcan reach a viabre but non<urturabre stagp (colwen a aL l985a) yet regain
iß cutturable status on human Passage (Brayton cl aL lg87)' Thiß suggesb thatvchobæ cÀ'A
maintain its pathogenic potential during a period in an aqua6't s¡vironment
The seasonal incidence of choþra may be at lea¡t partty attributable 1o variations in
plankton populations. oppenheimer and colleagues (198) have sho$'tr t\atvclplqæ ca¡
associatewith planktonic oopepods. ohervations in Bangtadesh re'ealed thatthe zooplanllon
18

population decreases during the monsoon 8ea8on (May-Juþ) in responoe to a reduction of
nutrienb and alæration of calinig. SubsequentU, the poprlatbn increases duringAugustand
September, shordy before the appearance of the first cholera cases which initiate the annual
epidemic. V elptsøe hæe been shorn to produce chitinase (Dastidar & Narayanasr¡mi
196g), to adsorb to a chitin eubßtrate and multipþ on such a surface (Nalin 4 4L lng)' Since
copepods and cn¡stacea have chitin in their exookeletons (Huq et aL t9%), association with
sucb organisms rnãy therefore orplain the abunda nceoî.v cholaaebsurfacewaters ¿[ 6s¡tein
'im€s of the year (Colwell d al 1987).
L73 a¡rnnission
Hietoricalþ, moot e¡ridemiologistr have been proponentr of the concept of cholera a¡
an exclusiveþ or prinarity waterborne disease. This has been based on the 19th cæntury
ouöreals, ild in particular, as argued by Feachem (19821 on misitrterpretation of the
pioneeringworkof Snow. Snocs investigation of the ouôreak¡ in l-ondon rwealed that there
was a clusteringof cases li¡kedwith the route of one of the cit/s two mai¡ water supplies (Snon'
185Ð. As pointed out þ Feachem (1982), througþout hb writing, Snow (lssÐ maintained a
concept of fecaloral choþra tra¡smission, of whbh waterborne tran¡mi¡sion b but one special
case. But perrcn-to-person spread, as a mode of fecatoral transmission, was not overlooked
eve¡ in Snow's time (roden'ed þ Feachem 1982)'
Epidemiological evidence suggests that person-to-person transmiseion can occlu' Paf-
ticularþin situationswhere there b overcrowdingand poorhygþng such as hæpitals (Mhatu
et aL lgg4: ctiff¿f 4L lg%),prbons and refugee camp' (Monb d aL lgsz). l¡ f¿nzanin, this
node of transmi¡sion has been impticated in an ouóreak of cholera in a podiatric i¡fectbru
diseases wa¡d (tvfhalu er al" lg&4). Further opporurnity for person-to-penon t¡ansmi$ion
ooeur' atburial cerenoniegwhere local traditi'on dictate¡ the cþansingof the intesti¡es of ttre
19

deceased (erren of cholera victime) prior to burial After the ce¡emony, the participanb sharc
a meal served on a @mmunal plaæ and eating b done with fingers (Mandara & Mhalu 1980).
Recent case co¡t¡ol studþs ha\re confirmed thåt, i¡ addition to water, food can terve as
a vehicle fo¡ the transmission of cholera (fauæ d aL l9æ). Foodstuffs i¡criminated to date
include rffi' or pardally cooked shellfhh in Mataytia (Duü €/ aL lnl),Italy (Bain e 4 al- ln4)
and portug¡l (Bl"k" er aI tnT, rãuv 6sh i¡ the Gilbert Isla¡ds (Mclntyre d 4I lng), and
insufficiently cooked crabs in the United States (Bl"ke el aL 198lJ). AB al¡eådy dbcusse4 food
nay have the effectof loweringthe ID of.Vdptqæþ p,rotectingthe organbms from gætric
acidlty during their passags througþ the stomåch. Thru, an ID of 106 El Torvibrios produced
severie dianhea in volunteerswhen i¡gested in foo4 whereas the same number gven inwater
did not (Lrvine 4 al l98lb).
In the liglt of gro*'ing erridence for the existence of an aquatic reeervoir lot V dtobæ
(see Sectbn 1.7.2),Miller¿f a¿ (19SÐ have rccentty p"stuLted prinary and secondary nodes
of dbease ü.ensmi¡sion They sugøt that h prinary transmission vibrioe pass from the aquatb
reærvoir to hunans via drinking water or edible aquatic flora or huna Subæquentþ secondary
fecal-oral ü.ennmi¡sio¡ invoþes penon-to-penon epread, either directly or via cotrtaminated
food or water. This concep is in agreement with the studies of Deb ¿f 4¿ (198ó). [n areas such
as Alrstralia (Rogers cr a¿ 19S0) and the United States @lake a øl 1980), where tùere are hi$
standårds of hygiene a¡d eaniøtioq littþ or no eecondary ¡¡n¡miesion would be expected to
occur, andas a¡rsuhan outbreakmrycomprieeprimarycås€sonly. Thetra¡smission ofcholera
is therefore complex and it is not ahvays easy to determine the exact mode of dissemination of
the dbease.
n

18 Potenüal protecdrrc antigeru ofVibdo clrolerae
The identification and isolation of pnotective antþens offe¡¡ one aPProæ'h to the
problemof developingvaccinesaginrtbacærialpathogens.vinrlencedeterminantsidentifi{d
by ehrdies on the pathogenesis of infection are ob,vious candidates for protective antþens' For
examplg an ap,preciation of the role of K88 pili in the colonization of the piglet gut ry
enterotoxþe ,,ic,E col¡rúto the dorclopmentof an efrectivevaccine comprisingpurifiedKS8
(Ruüef & Jones 1973;Rutter et aL 196). Althougþ various comPonents arc thougþt to assist
the pathogeneeb of vcløbae (rce section 1.3.3) it remains unclear as to whether any car
fu¡ction as protective antigenc for human infection The erridence inpticating these vario¡u
factors as pmotective antþns isbrieny revfors'ed'
L&t tPs
The LpS of a1 serotypes oonsisß of th¡ee regions: the lipid A core oligæaccharides
and the repeating o-antþea polysaocharide subunits which represent the outermæt surface
of thevÍbrio. The lipid A b hydrophobic and b responsible for the endoto¡ic biological activity
of LFS. The O-antþen region is responribþ for antigenicvariability and its chemical compæi-
tion has been determined (Hieatsune d aL lng: IGn¡e et oL lng,l98l Redmond lnÐ'
Unlike the LpS of most gram-negÊtive bacæria, the 2-keto-3deo:ryoctonate which lints the
core sugaß to the lipidA is absent i¡VùolqæLFS; instead fn¡ctose is present (Janet aL
1973;I(abif 1932) a¡d thb presumabþ eerves the same linlÉgg functio¡'
Observations made in a varÞty of erperimental models have sho\pn that antäbodies
directed against the LpS deærminan ts olV cholcræ harre the potential to mediaæ protection
Thb fu the casewith "open" systeme euch as the infant mouse model (N"oh & Ro*'þ 1972) or
the DIC (duodenal inoculiarion with cecal tigation) model in adult rabbits (Jensen er 4L 19æ)'
2l

AntÍbodies to LPS are al¡o sufñcient to mediaæ protection in the moet commonþ employed
"clooed" E''stem, the rabbit ileal loop model (Svennerholm 195; Chitnis d aL 1982a),
As discussed beloç, (Section 1.&3), it is generalþ aocepted that resbta¡ce to clinical
disease is mediated by antibodies directed aginst antþns of the bacterium ¡ather rhrn tie
todtr The rehtive significance of antibodies to LPS determinanls ¡pm¡ins uncertain, althougþ
ci¡cumstantial erridence suggerb that LPS may be a protective antþn for human infection
First, field and clhical testing of cholera vaccines has occasionalþ rrcsulted i¡ serotype+pecific
protection (Mosley d aL ln}; Cash et al- 1974a); since atrtigeos associatedwith LPS are tìe
onþ ones tnovn to difier between the serotypeq thi¡ protection was presumabþ mediated þ
antibodies directed agnin"tlPs. Secon4 a p'urified LPS væci¡e conferred E¡nsient protection
upotr adult recipiene in a Bangtadeeh ñeld trial (Joo 194). Finally, epidemblogic sûrdies have
identifred inverse correlationsbetween inc¡eased levels of a¡ti-LPS antibodies anddecreased
incidence of disease (Mosky d aL 1969; Glass d aL 1985)'
L&2 Non-LPSantþre
Until recently, little interest had beer shorrn in the possible vaccine siÊnificance of the
non-LpS oomponentr of Vdøleræ. Studies in the infa¡t mou6e model had indicated that
antibodies to ouch oomponenür were sufñcient to mediate protection and that in thb ryastem at
least these antiibodies appeared to be more eflicient h this respect than antibodies specific for
LpS (Neoh & Rocil ey ITIO;Aüridge & Ron'ley 19S3c). The naûrc of the prtative non-LFS
protective antþens remai¡¡ undeñne4 but ca¡didateewould itrclude outer membrane proteinr
(OMPs), flagellar a¡trgenq tIAg a¡d ptt
n,

L&¿l oMk
Ctaracterization of the outer membranee of Vclplqæ begpn comparativeþ recentþ
(IGbif 1980; IGlly & Parker 1981; Manningcl aL lgsl Jonson d aI 1989). Slh- oMPs of h
vino grantn organisms are anabZed þ electrophoresis in poþacrylamide gek i¡ the Pnresenoe
of sodium dodecyl sulfate (SDS-PAGE), approximatety 8-10 maior proteins rangng in ap'
parent molecular weight from 27-94 kilodaltou (kDa) can be resolved" Maior proæine include
3-4 proteins in the ratrgp of 45-48 tD+ uÀirù p¡obably repreeent the porins (Kabir 198q
K.lly&Parker1981;Manningøf',L]lg94l(abir1983;Iang&Paþa1987;Jonsoner4I1989),
a 40 kDa prcteio' ompT and a 3g kDa proteiq ompu (petereon & Mekalanoe 1988; Jonson
er aL tggg). Other oMps include a 35 kDa OmpA-like protei¡ (Atm et al" 1986; Jonson ¿Ú a¿
19S9) and a 25 kDa prote¡n, OmpV (Stevenson d aI 1985; Fohlner et al l9ú41986b)'
Recentwork indicates that the oMP profile of V dtobae is significantly influenced þ
the gro$rth conditions empþed. The moet gtrikiog example of cr¡lu[edependent protein
s¡,athesb is the productioa of pili such as TCP (Section 1.3.3.4). Sciortino & Frnkehæin (1983)
reportedth atVdtolqaerecorrered f¡om the intesti¡es qf infantrabbiu expessednovelOMh
when oomparedwith h vito grotttorganisms. Cr¡lture in irondepleted medium regulted itr a
similar OMp proñle, suggesting that thb was a convenient means of sürdying Prote¡tr8 syn-
thesized in respons etohvir.p conditions. Ea¡lier etüdies had sho'c¡n thatV ùobæ OMh are
immunoggnic i¡ man (IGbir 1983; Seârs d oI lg8/). lVith the exception of TCP and the
flagellar stn¡cture, the cont¡ibution of any of these OMPo to pathogenesis (and thus the
potential protective signiñcançs) is unlnowa'
B

L&22 trlagenü fiI) antþere
St¡rdies in two differentcholera models rabe the poesibility lhar'V eholøae mi$tb€ar
a protectiveantþenassociatedwith the fl"ællretnrcture. Eubanla¿t aI (tg77)reportedthat
an antigen present in a crude flaçlta f¡action was able to elicit prrotective antibodies in a
modifrcation of the infant mouse model This antigen was not present on a noa-flagellated
mutant straiq nor did it seem to be ascociated with the fLællat core protein Subeequent
erperiments revealed that the sarne component was a protective antigen in the adult rabbit
tigated ileal loop õy6tem (Ya¡"ey et aL 1979). Althougb this antigen remaín¡ undefine4 tieee
¡ûrdies are of considerable interest in vien' of the poosibility ^hat the flagellum carries a¡
¿g¿phment factorwhich Playg a role in colonization (6ee Sectbn 133'1)'
L&2.3 IIAs
There is paucity of evide¡ce to suggest that anti-tlA antibodies are ¡xotective.
Chaicumpa & Atthâsfuhtha (199) reported that an anftody to a cell-associat€d HA from
Vdplqæ El Tbr Ol7 was marginalty protective i¡ the infant Douse c;hoþra nod€L Other
sû¡dies in the same systeq howorer, led to the conclusion that the O17 strain lacks non-LPS
protective antigens (Attridge & Rou,þ 19s3c). Finkelstein & Hanne (1982) found that Fab
fragments of antibodies againstpuriñed SHA/proteasecouldinhibitthe hvíto attachmentof
V ctplqæto infant rabþi1 sm¡Îl bwel Suboequent e.rperiment¡ failed to demonsEate protec-
tive activity associatedwith such antibodies (Booth & Finkelstein 19Só). A clearer appreciation
of the likeþvaccinesþificance of úevarior¡sHAs producedþ Vchfuæ mustarraitfurther
studies. lvith the cloningof the gene encodinga v dpleræ tIA (Franzon & Manning 1986) it
mãy 5. possible ro determine whether or not HAs possess protective activity in cholera
7A

1.&¿4 P¡n
At the co¡¡me¡¡oement of theee sûrdies, ¡orhing was knoq¡n of the prrrtective cig-
nificance of a¡tibodies directed against V dplaæ pili nâinly becar¡se the question of pilru
expression remained coDt¡o/enial Recently, howerrer, Taylor et aL (1987) described cultural
conditions which allorred TCp synthesis by Vctwbac and shoil,ed tìat these pli plty"d a
critical role in coloniz¿tion, at least in the infant rnouoe model Ð "n logwith other sy8tem$
one migþt speculate that anti-pih¡s antiHies could be protective againrt V drcbæ by
¡s¡ü.alizjng the colonþ¡tion capacity of the bacteda Thus, pilus vaccines prepared from
diarrheagsnic strains ol E coli and given parenterally to pregnmt so$'E and coctB harre sig-
nificantly reduced ¡eonaøl deaths ftom d¡arrheal disease (Rutter & Jones 1973;Moryatd aL
lflg;Nan, d aL lng). hotection was ehoq'n to be mediated by anti-pilus antibof passiveþ
transmiüed ir maternar colootnrm @uüer & Jone¡ lglS Morg¡n¿l ø1" t9l8¡ Nag et aL 198).
pilus vaccines have also been effective in preventingerperimentzlMorælh h,ús infectbn i¡
qos,B (Ilamont & Bcbgo 198Ð. There is, therefore, a rtrong possibili$ thet TeP win achier¡e
the stafirs of a Protective antþen'
L&3 CT
Theother maiorantþen producedbycholeravib¡ioe isthe CT(revieu'edbyFinkelst€itr
1fl3; Finkelsæin & Dorne¡ 19SÐ. It has been purified to homogeneity (Fhkelstei¡ &
Lospa¡uto 1969,tnqRichardson & Noftle lg0), and its subunitstrucü¡re and mechanism
of action elucidated (Lonn¡oth & Holmgre ¡ 1973;Hohgrea & Lonnroth lnq' cT i8 a¡ &4
kDa polyneric protein composed of two protomeÉ: a¡ A (ZI YDa) or aderylaæ cyclase
etimulating compotrent (chobragen) and a B (5S kDa) protomer (choleragenoid) associated
soleþ with binding (I-ospalluto & Finkels tÐln lnlPierce 193; van Heyningen 'gm'
The A
25

subunit comprisee two PoþpePtide chaing At(pkDa) and A2 (5 kDa) (Saüler 4 aL 197!'Ln
et aL 1976)while five identical subunits (11.6 kDa eaeh) make up the B prrotomer (Gill 196;
Nakashima 4 aL ln6). The r¡arious physiologicål manifesatbns of the toxi¡ on different
tirsues, including the hypersecretory r€sponse elicited in the intesti¡at e¡úthelium, have been
found to be based on a singte moþcular mechanism. Folbwing the binding of the B subunit to
the Gvt ganglioside of the cell rembra¡e (Eee section 1.3.4), the A subunit is cleaved and
ente* the celr, activating adenyrate cycrase and leading to elorated cytoplasmic levels of cyclic
AI\,íP @ierce 4 aL lnl: Oill lØr'
CT b hl#ly immunoÊÞnic with nearly all the antibodies directed 4gainst the B subunit
(svennerhotm 1980; Hotmgren 1981). Ninety percent of North American volunteers
developedrises in senrm þG antitorin afreranexperimentalcholera infection andlevebwere
etill detectåble two yeafs latff (I-evin e 4 d 1981c)' Approximaæþ 6t% o1' votunteers
manifested rises itr eIgA antitoxin in i¡teetinal flur4 but the r€sPotts€ tetrded to be relativeþ
ehort-lived with antibody levets dropping one month after challenge (cited by l-e'n^e et al-
1e83).
Sûdies in various animal modets have i¡dicated that, whether passiveþ acquired or
induced þ active immunization, antibodie to t[e toxin caa protect ¡gninst eryerimenøl
chobra (roriewed by Finketrtst\ lnl, tflS; Finkekteh & Dorner 1985; Holmgren cÚøL
1g5b; yoohiyama & Brown l9sÐ. In contrast, volunteer sû¡dies have consistently failed to
demonstrate the invoþeme¡t of such a¡tibodies in ¡esistsnce to clinical infection (c-ash d aL
tgl4a;:If,l/trle¿f al 1y7g,1981a). I¡dee4 a rece¡t report makes it cþar that immune re¡Ponses
to the toxin a¡e notnecessary for protection in man (Levine ef 4¿ 198sb)- onþ two of the three
choleravacci¡es currently Þing developed have the potential to generate a¡ti-toric immunity
(eee Section 1.10.2).
?ß

1.9 ImmunitY Ûo cholera
It is nos, clea¡ that cholera infectio¡ gives rise to suboø¡tial protective imnunity to
subeequent expæuru to the disease. This has been convincingþ demonstrated by the fact tlat
the i¡cidence of the disease raprdly declines with i¡creasing ag. (see Section 1'7'1), and that
recurrent i¡fecti,on¡ a¡e fare in the communities whe'te the disease ie endemic (Ñennerholm
1980; Glass d oL lgg2). In addition, I-e'ine 4 4L (lgSta) reported that, three years after an
inducedcholeraice¡risodgvolunteerswe¡ìe abletoreristchallengewitha strainof homologoue
or heterol,ogou eerotYPe.
The nahrre of the protective immune response remains unclear. Available erridence
suggests that anftodies directed aginet determinana of the bacærium rather than the toxi¡
are primariþ responsibh for immunity to clinical dbease (cash á aL l9l *a;l-€'v lü!le d aI lnÐ'
In addition, despiæ the ris€s in serum antibodiee elicitedþ parenteralvaccines, such formule-
tions hare prfled inefiective in conferring protection Thb i¡dicates that the location, and
poesibþ the isotype, of the a¡tibodies is aho cfidcal Evident! the IgG and lglvl antibodieg
inducedby parenæral inmunizstion do not reach the gut in sufficient concentration to make
an effective co¡tribution to enteric defence. It b now çneralþ accepted +hat atr effective
vaccine win need to be oratty administered (sectbn r.10.2) to generate antibodies in the
intestine to prevent col'onizatb n o1 V dPbaa
L9,1 SSnsúeuic antibody rcsponrcs toV cholerac
Infectbn with, Vcløtqæ is associated with a ri6€ in titer of a varie$ of circulating
antibodies, incrudingvibrocidar a¡tibodieo (Fint*tstein lg6e Mosley 1969) and antibodies to
cT (Pierce d aI lnq,IJ,S (Clements d aL tg$?4,svennerholm et aL l984b) a¡d oMP (Sears
ct aL lgu). Esrty studies demonstrated thatvibrioci¡tat antibodywas acquired eåriy in life i¡
n

endemic a¡eas; p1lels increase with ag. and remain elertated, presumably æ "
oonsequence of
repeated environmental exposure to Vùfuæ (Mosley 1969: Glass aaL l98S). SharP in-
cneases in tiæn of both vibriocidal and antitoxic antibodier have also been demonstrated in
volunteer studies and in infected individuals from non+ndemic areas (Cash cú aL tÍl4c;I-wine
et øL lgg¡c; Snyder d aI lgSl; Clemenb el øl^ 1982). Hoç'errer, in contrart to the situation ¡n
endemic a¡ìeas, kvels of such a¡tibodies in these individuals are not sustai¡ed (I-f5tin.e d aL
1981c; Clenenß 4 ølI1982).
As mentioned above (Section 1.9), the circuLating antibodies generated þ inactivated
parrenteral vaccines do not confer signiñcant protection against the non-invasfueV dpbæ.
Nevertheless, earþ epidemiological studies rwealed an inverse correlation between
vibriocidal antibof titen and the inci&¡ce of choþra in the field (Mosley 1969).Althougþ
such a relationrhip was not apparent in avolunteer study in which vibriocidsl antibodiee we¡e
elicited by parenæral immunization (Cash etaL lll4a), such antibody rcsPonses wele as-
¡ociated with resistanoe to challengp in a rccent evaluation of an o¡al cholera vaccine (hcket
et aL,I¡ pne$). Þidentþ, ifvibriocidal antibodiea arbe as a consequence of oral presentation
ol V ctplaæ antigenq they reflect the occurreace of a protective lq(al immune fesPonse-
Mbrioci¿al anti6y responses may therefore ofÍer a convenient and helpful index of im'
munogencity to ass¡st the developrment of improved choleravacci¡es (hcketel&L,h press).
Since V etpløæis a non-invasive pathogen producing dbease at the gut mucæa onþ, itwould
be likeþ that the secretory imnune system i¡ the mrior protective mechanism in limiting
infection and the severity of the di¡ease.
a

Lg.z Dwebprent of intestinat sIgA antibody rcIþßrcs
Induction of the intestinal reslþûse occurr primarily in the Peyer's patches (PP;
\uoodruff & clâfke lggT;Mestecþ & Mcchee 1gs7). These are dome-like ss'ellings on the
epithelial surface of the gUt which serve as a staging area for mþating þmphoid cells to come
in contact with luminal antigens (Husband & Gowans 198). This interaction begins with
uptake of antigen by specialized epithelial cells residing o'/er the pP (Bhalla & ou'en 1983)'
These cells were initially characterized by the presence of luminal surface microfolds rather
than microvini lendingthem the name M celrs for microfold cells (oren & Jones ln[\.\vhen
microrilli were observed on these cellsr the name persisted and now refers to imembranous"
epithelial cells because of the very thin rim of cytoplasm separating the intestinal lumen from
the þmphoid cells below (wotf & Rye 1gs4).In addition these cells are devoid of pnoteoþtic
enzymes and correred with scant mucus thus designed to sample and transport antigen without
degrading it (reviened by sneller & Strober 19g6). Absoôed subotances from the lumen are
pinocytosed by the¡e M cefls and are transmitted to underþing PP in an undenatured form' to
be suboequentþ preeented by accessory cells to T and B lynphocytes (sminia et aL 1982;
Wilders er aL 1983)-
The factors which deærmine whether bacteria present in the gutwill be "sampled" þ
M cells remain unclear. uptake þ abeorption may be facilitated by specific membrane
receptors or gþcoproteins shown þ lectin-binding experiments to be present on the surface
of M cells (orn en 1gg3). Further research is necessary to determine whether specific receptors
would enhance selective antigenic endocytoeis and transport onren et aL (1986) has recently
demonstrated that viable, but not inactivate4 V choteræ are indeed inæ.t"d by M cells and
trancferred apparentry intac! to mononuclear cells in underþing l¡'mphoid follicles. They
suggested that critical surface antþens or structures' apPafently mis¡iag from inactivated
29

bacæria, are required for uptake (ornen et øL 1986). Inman et aL (19%) have shown that the
nature of the surface poþaccharide can influence the rate of E' coli samplingby M cells'
Follouring antigen sensitization, the primed l¡rnphocyæs leave the PP via efferent
l¡rmphatics without further difierentiation (Craþ & Cebra t97l)' They then pass th¡ougþ
meeenteric nodes and enter the thoracic duct f¡om which they mþate entering the s¡ætemic
ci¡culation to home back to the gut lymph nodes. The concept of a common mucosal immune
system linked þ emigrating IgA Pfecun¡of cells has been supportedby e:perimental studies'
These harre sho*m thattùe circul,atingintestinalþ-Primedþmphoblasts notonþ return to the
sub-epithelial tissue6 of ûe gut, but migrate also to the lungs and genitourinary t¡act as well ae
to exocrine glands (salivary, lacrimal, mammary) (McDermott & Bienenstock 1fl9;
Bienenstock & Befi¡s 19g0; Bienenstock d aL r9g3). This ensures that the activation of
lymphoblasts in a particular mucosal sile can ¡esult in the deplo¡rmentof appropriate effector
cells th¡oug[out that mucosa as well as other mucosal surfaces'
Evidence has ¡ou, accumulated to indicate that humans also have a common mucooal
immune Bystem. primed þmphoblasts harre been detected in the peripheral blood afær earþ
intestinalexpooureto antþen; in addition, oralimmuniz¿tion resultsin thesecretion of specific
sIgA antibodies in saliva, tears, milk a¡d nasal secretions (roriewed by Mestecky 198Ð' l¡ this
contex! þcke et aL (lggÐfound thatSwedish volunteers immunized oralty with a combined
bacærial-toxoid vaccine generated peripheral blood l¡'mphocytes which differentiated into
specific cholera þA antitoxin and anti-I-PS antibody-producing cells during a period or'ín vino
culture. Other studies ha\¡e demonst¡ated that oral immunization of volunteefs or natural
disease in cholera induces ar¡n¡v chotsa¿ antibody respomes in sativa and breast milk
(Jertbom et 4L lg8fj) also reflecting the gut mucosal responses is part of the common mucosal-
associaæd immu¡e s)tstem.
30

Recent erridence shoun that cell surface molecules selectiveþ e:çressed in mucoeal
organs are required for lymphocyte honing to theee eites (Streeter e|øL 198ß)' To enter
intestinal tissues, þmphocytes must reach the pootcapillary venules in PR other þmphoid
aggregates or the muco6a, and must adhere to tìe endothelium to begin their migration- It is
nou, clear ft¿t this is due to seþctive and specifrc interactions between surface determinants
of þmphocytes a¡d the endothelial cells (referred to as hiÉ endothelialvenules, HEÐ of the
pootcapillary venules of the þmphoid tissues. Surface determinants that permit these interac-
tions ate found on both the tymphocytes and the endothelial cells. The determinants invohed
in mucosal tissues differ from those invoþed in peripheral lymph nodes (reviewed in Mõller
1e8e).
tl is beco,ming increasingþ errident that the þA resPonse is T cell dependent (revieu'ed
in Meetecþ & Mc6hee 19gz), particularþ in the induction and differentiation of IgA+ B cells'
They harre been ohourn to induce B cells to ss'itch from e:pression of surface IgM Ûo surface
IgAaswell ag inf,uencingthe proliferation and ærminal differentiation of B cells throu$ the
production of inærþukins IL4 and IL-5, respectiveþ (revien'ed þ Mestecky & McGhee
19sÐ. The dimeric IgA produced ry these cells in the lamina propa is transported into the gut
lumen through the columnar epithelial cellsvira a speciñc receptor, secretory component' which
is located on the basal surface of these cells (Mostov et aL l98/ù and produced by epitheliral
celts of avarieg of other secretory organs @randtzaeg 1gg5). The secletofy componenton the
sIgA nolecule confers some resistance to proteoþfu, il important characteristic because of
the environment in which sIgA exerts its activity (tindh 1975)- Recently, it has been
demonst¡ated that intes"îal antitoxin formatio¡ and pnotection against toxin challenge after
oral immunization with CT is thymuedependent and tikeþ to be under Tcell control (I-ycke
et al- 1987a).
31

1.9.3 sIgA ant¡-v c.holerae rcsPoßtes displ¡y irnrunobglcal nnlmry
The intestinal sIgA response to cholera is of relativeþ short duration, lasting for a fern¡
weeb to afeç,months (Sennerholmel aL lg8ø¡b).This contrastswith protective immunityin
cholerawhich appears to last for severalyears (Levine 1980; Glas6 et oL 1982). This paradox
has been recentþ resoþed by experimental and clinical studies which document long-term
sIgAmemory to cholera antigens (Ñennerholm¿f aL [9$4a;Holmgren & Lycke 1986; lq"b
etú 1987b). For example, women in Bangþdesh who had been gry"o 2 oral immunizations
with cholera B subunit 15 months eadier respondedwith intest¡"al IgA antitoxin production
Eofe rapldly, and to a lourer dose of antigen, than a concurrentþ teste4 p¡atioueþ unim-
munized gfoup of volunteers (þcke & Holmgren 1987). Similarly, 2years after initial oral
immunization of mice, a second dooe of antþn evoked a rapid intestinal antibody resPonse
which corretated with heigþtened resistance of gut loops to CT challenge (þcke & Holmgren
19s7).Inaddition,immunologcalmemofyhasbeenehol¡rntobecarriedþBcellsandthatit
is possible to adoptiveþ transfer long-term memoryþ these cells isolaædfrom donoranimals
1 year afær priming (þcke & Holmgrel 1989). Ttese studies all show that the intestinal
secretorylgAsyEtemhasaveryefflrcientimmunologicmemory.
Lg.4 Mechanismof sþa antibody-rediatcd Protectbn agdnstv choleme
Various studies hatre denonstrated that the biological function of sIgA is to prevent
initiation of infection þ organisms using the mucosal surface as their primary location of
attachment during deyelopment of disease. rilhile sþA is neither opeonic nor p*s€sses
bacteriocidal activities (re'iened by underdown &, schiff 198ó), it can efficbntly cross-link
antþns, prerrent adhesion and inactivate toxins. sIgA can render bacæria mucophilic (Mag-
32

trusson & Stiernslrom 1gg2). It is well knou¡n that vibrioo are associated with mucus ínvivo
(Jones d aI ln6; Nelson et aL 1976)'
The specific mechanism by which the mucooal immune system functions is poorþ
understood. purified sIgA antibodies har¡e been shoum to prorent attachment of bacteria to
mucosal surfaces. Intesthar loops prepared in orarþ immunized mice were partially prctected
from the effects of live cholera organbms in elperimental v ùobæ infection (Fubara &
Freter lg13).The protection appeared to be due to the action of specific antiMies excluding
the organisms from attaching to the gutwall. steele et aI (1974) reported that purified sIgA
di¡ected agúîÊtv e]þlqæLßwas prrotective in the inf¡nt mouse cholera model; subsequent
sûrdies in this sy'tem suggested that the antibody function necessary for protection is the
capacitytoinhibitbacterialattachment(Attfidge&Rou{ey1983c).
severat shrdies have shq¡n that þA antitoxi¡ antibodies induced by e:perimental
v chobøeinfection, orbyoraladministrationofcf areassociatedwith pmotection ef ialsstinal
loops asainst CT challengp (Pierce ef 4L lnE: Taman¡ & Broum 1985; Vaerman d aL 1985\'
sIgA can neutalize toxin (svennerùolm 19SO). I'.ryckeet al (l9s7a)have recentþ demonstr¿ted
that IgA antitoxin antibodie¡ qrnthesized h vito þ lamina propia cells have protective or
neutrarizing activity. Evidence suggests that the essentiar mechanism is þ preventing the B
subunits from binding to the crrar ænghoside recepto¡s on the epithelial cells; indeed moot
antitoxin antibodies a¡e directed ¡grinst the B subunit rather than the toxic A subunit after
both i¡fection and oral immu¡ization (Holmgren 1981; Lindholm eï4L L983)'
ILl,
Þ
I
I,1,
fl
II
I
I
i
33

f
I
li^
1.10 \äæination against cholera
L10.1 Ptst appmaclcs Ûo rraccinatbn
A century has ela@ since the first choleravaccine was adminisæred to human beings'
yet there is still no acceptable, effective vaccine for cholera (revietved by Finkelsæin 1984)'
The history of vaccination 4gaiast cholera began in 1885 when a vaccine composed of broth
cultures of tiving cholera vibrios was iniected subcutaneously by a spanish physician named
Jeime Ferrán y Clua (Bornside 19s1). In 1892, lValdeman Mordecai Haffkine, a Russian'
derreloped a resimen of two subcutaneous inoculati,ons of a vaccine which had been nat-
tenuatedn þ culture at 39 C Althougþ the vaccine caused malaise, ferter, pain and swelling
these side effects were considered acceptable and the vaccine euitable for admintst¡ation
(Bornside 1932). Honreveç due to ethical coneiderations, livevaccineswere then replaced by
rìe killed vaccines of Kolle and otherworkens (Pollitzer 1959).
1.10.L1 Conventionat Hlled vaccines
Theee vaccines $,e¡e prepared þ inactivation" either þ pheno[ formali¡ or heat
treatmen! of suspensions of Vchobæ groun on agar or in liquid media. only in the 19608t
with theadventof cont¡olled fieldtrialsasa meansof er¡aluatingvaccineefticacy, diditbecome
apparent that such vaccines were not eufficiently effective to have an impact on the incidence
and prwalence of cholera (reviened þ Finkelstein 19&4). Althougb some formul¡ations
conferred higþ lwels of protection, the immunity quickþ waned (Joo 194; Feeley & Gan-
garosa 19g0). Furthermore, killed parenteralvaccines were generatþ more effective in older
people, for whom the vaccines wene thougþt to booet environmentalþ acquired natural
immunity (Finkelste tr lg6}).More frequent admi¡ist¡ation of vaccine was required to observe
I
It(
I
I
v

simlar rates of protection in young children (Joo 194; Feeley & Gangarosa 1980)' the group
at highest risk of acquiring infection'
The oral administxation of killed vaccines had been considered ser¡eral time¡ prior to
1960 (Polli Eet 1959;Freter & Gangarosa 1963), but the impetus for such an app'roach was
provided þ a report that college students fed heat-killed vibrios mounted both serum and
intestinalantibodyresponses (Freter& Gangarosa 1g63). Howerrer, subsequenttestingshowed
that the oral route of administration was not as effective as parenteral vaccination in conferring
resistance to a clinical infection (Cash et al' 1974b)'
1.10.L2 Toxoid vaaines
The recognition of the fact that cholera is a toxin-mediated disease (De 1959;
Finkelstein e Lospalluto 1g6g), suggested the feasibility of developing protective toxoid
vaccines, an appfoach which had proved successful ia somþ¿tfing diphtheria and tetanus' As
discussed above (Section 1.g.3), data fromvarious animat moders indicated that antibodies to
the toxin could mediate protection againsterperimental cholera, promptingthe development
of toxoid vaccines for clinical apprication. A formarized toxoid (which was stabilized s'ith
gþcine to prevent partial rerrersion to toxicity) wÍ¡s prepared (Ohto mo 1977\ and tesæd in the
philippines (Noriki tgïl)but had no impact on the incidence of infection. A gluraraldehyde
toxoid prepared by Rappaport and colleagues (194) was tested in Banþdesh' Prolection was
observed up to 24 weeks, but the cumulative protection ovef a one-yeaf Periodwas marginal
(Curlin et at 197ó).
Volunteer studies suggest that resistance to clinical infection is prwided by antibacterial
ratherthanantitoxigimmunity(ca"hetaL|974a;I.er,inedaL|979).ofaladminist¡ationof
three doses of cholera toxoid failed to prorride any proteclie¡ against cholera challenge,
whereas previous clinical infection conferred enduring resistance to re-challengewith strains
i
35

of homologous or heterologous serotyPe (I-evine et aL l!79; 1981a)' More recentþ, lær'ine
et aL (l9ffjb) haye shon¡n unequivocalþ that resistance can be elpressed in the absence of
antibodies to cholera toxin.
L10.2 Cument approachcs Ûo vaccination
The disap,pointing results obtained from fietd trials designed to er¡aluate the efficacy of
inactivated parenteral vaccines contrasted with the realization that clinical infection is ibelf a
very effective immunizing pfooess. Epidemiologic frndin8s (Mosley er 4L Lgflf¡; Glas et aL
19g2) ande:perimentalchallenge studies involunteers (Cash er øL lg74a;l'erttneet aL l9Sla\
have demonstrated tùat cholera infection is followed þ poten! long'lasting immunity for at
least three year' (I-evine d ar lggla). current k¡o*'ledge about intestinal immunologl
(Section 1.9) indicates thatthe oral route is superior to the parenteral rr¡ute for stimulatingthe
formation of sIgA antibody in the gut mucoea Indeed, this knowledge, coupled with the
collective failure of parenteral vaccines, provided a strrong impetus for the development of
chorera vaccines suitåble for oral administ¡.ation Tr¡o main approaches have been followed
over the past 10-15 years, with a thiÌd option being develo@ mo¡e recently'
L10.¿1 üve attenuatcd oral mccines
The basic approach employed in the dei'elopment of these vaccines has been to
inaptivate the genes for toxin production, leaning i¡tact thoee bacterial comPonena required
for gut colonization. The prototype vaccine was t\ev choløae strain Texas star-sR which was
attenuated þ chemical mutagenesis (Honda & Finkerstein rg9). This mutant produces tle
B but not the A subunit (ATl* ) of cholera toxitr. Involunteer studies, itwas found to colonize
and proliferate in ttre gut and to elicit serum vibriocidal antibody responses comparable to
tiose observed followingnaûrral infection Production of slgAaminstsetreralvibrio antþens
%

was denonstr¿ted (I-evine et 4I lgf!|¡). Hou'ever, in spite of the Prcsence of the B subunit
antitoxin responses were poor (Iævi ne d øL l9g3, 19s4). Although the strain was well tolerated
by a maiority of volunteers, ?AVo e:rperienced mild dianheq a side effect which precluded
further work wittr this st¡ain (Lerrine et 4t. lg8p¡). It did Plry a role, however, in e:ploring the
feasibility of mimicking infectionderived immunity þ means of an oral attenuated vaccine
(I-evine d ar lgu), thus p*ing theway for the geneticalþ engineered st¡ains to be discussed
belou,
The ñ¡st geneticalþ engineered candidate vaccine strains were described in 1984
(Kaper d 4L lguíÐ19S4b). St¡ains olV choleræ knou to cause disease and to give rise to
protection in volunteen (Lerrine d 4L lng: Le'ine 1980; Levi¡e d 4L l98La) were used to
prepare a series of derivatives from which the genes encoding both the subunits of the toxin
(AT-) or just the A subunit (A-B* ) were deleted by oiædirected mutageneßß (Y:apr ef d
19844 184b).
Volunreer eh¡dieswith JBK70 (A B-) and CVD 101 (AB+) shou'ed that these strains
colonizedwellandeliciæd goodvibriocidal andantitoxic(cvD 101) reeponse& Furthefmo¡e'
JBK 70 (AB) vaccine strain gæe significant protection (89Vo efficacy), illustrating that
resistance is notdependent upon tie generation of antitoxic immunity- Howerrer, þeû strains
were reactogenic (r.evine er at. rgßb), causing mild diarrhea similar to that seen with the
prctot)?e Texas StarSR dercribed above. one poosibility is that the Prccess of colonization
pa sepnovides sufÍrcient irritation to the epithelial surface that mild diarrheawill inevitabþ
occuf in a propofion of recipients (snith & I-inggood l97t; Finkelstei¡ 19&4; Wanke &
Guerrant 1gs7). Alærnativ ety, v ehotøae mifit secrete multiple toxin molecules, so that
deletion of the genes encod.ing the well-recognized cholera toxin might not completeþ
abrogate diarrheagenic potential (Iørine d aL l98Á;b)'
37

I-errine et at (lgæar lgssb) have performed further volunteer trials to address this
issue. Deletion of the El Tor hemoþin/cytotoxin genes did not decrease reactogenicity of
JBK70 or CVD101 (Isrin e et al- 1988b). In addition, studies with (X/D103, an AB + derivative
of a Classicål Inaba strain 569B which lacb the geneß for the El Tor hemoþin, confirmed that
ttis molecule was not the cause of diarrheagenic activity (I-evine er^L l988z). Hou'ever,
althougþ CVD103 caused diarrhea i¡ l2To of volunteerõ, a mercury-resistant derivative
appears more p,romising CVD1O3-HgR was well tolerated by ?A of 25 recipients, while
retaining the immunogenicity and protective efficacy of cvDr03 (Irvine ef øl 1988a). Further
clinical evaluation of this strah is clearly warranted. At this stage the precise mechanbm
responsible for the reactogenicig of these derivatives remai¡s unclear (Kaper 1989)'
L.I|O.LZ Non-viable or¡l vaccines
Evidence thatantitoxic and antibacterial immunity actsynergisticalty to Protectagainst
e:rperimental cholera (Svennerholm & Holmgre n1976)prwided the impetus for the develop
ment of a combined bacterial-toxoid vacci¡e. Althougþ protective antitoxic immunity has been
shou,n to be directed ptrmarily egrinst determinanß of the B subunit (Holmgre¡et aL 1977:'
Holmgren & Svennerholm 198Ð, the targets of proæctive antibacterial antibodies remain to
be elucidated particularþ with respect to clinical infection (see Section 1.8). Accordingþ the
combined vaccine comprises killed whole bacteria a¡d the isolated B subunit of the cholera
toxin (Clemens d aL 1986).
This vaccines has now been extensiveþ evaluated. Administration has been shos¡n to
stimuliate local sþA antitoxin and anti-LPS reEponses in voluntee¡s in both endemic (Senner-
holm ef aL l9g4a)and non-endemic a¡eas (Jertborn 4 aL 1984). No side+frects were observed
in studies with several population groups (Black e! aI 1987; Svennerholn et øL l9ah;
Migasena et aL t9g9), or in a Bangladesh field trial (Clemens eÍ aI 198Ð. Vibriocidal serum
38

responses were lor' @lack et at. t987; Clemens ef 4t 1987; Migasena e¡øL 1989) when
comparedwith that inducedwith the attenuatedvaccine strains discussed above (I-evneet aL
19SSb). Nevertheless, it has proved to be prctective in volunteers (Black 4 aL 1987) and in a
recent field trial in Banþdesh (clemens d oL 1988, 1990). To confer a7o7o protection rate,
it had to be gryen in tno doses to adulte in Bangfadesh over a three year obsewation period
(Holmgren d 4L 1989: Clemens d aL ßm)'
The combined bacterial-toxoid vaccine clearly represents a maior advance over the
killed parenteral vaccines, in that the protection conferred is enduring. As a vaccine it is not
yet ideal, howwer; its main deficiency is that protection in young children waned rapidþ with
time aftervaccination (Clemens d aL lg8f,, 1990; Holmgren et al- 1989).It is also debatable
as to whether the inclusion of the B subunit is justifiable from the vieu'point of vaccine
production coat A comparison of the protective efficacies of the combined vaccine and a
vaccine conprisingiust the bacterial component indicates that the advantages of the former
are t¡ansient (KaPer 1989).
L10.23 üve oral CtolerqÍBrPhoid bùrid vaccine
In an att€mpt to combine the supe¡ie¡ immunogenicity of a livevaccinewith the eafety
of an inactivated formul,ation, an attempt is nou' underway to clone a protective antigen from
V chobæ into the attenuated, live oral typhoid vaccine Satmonella typhi Ty2la (Germanier
19g4). The latter is now acceptedas atr effective typhoidvaccine, thesafety of which has been
demonstrated h numerousvolunteerstudies and fþld trials (Germanier 19&4). Althougþ the
protective antigens of. V cholqac remained undefined with regard to clinical infection, cir-
cumstantiål evidence suggested that antibodies to LPs could confer immunity (Section 1'8'1)'
Accordingþ the genes responsible for O-antrgen synthesis nV dtolqæwefe cloned (Manning
et aL 19116) aod transferred hto T21a (Moronaet4¿ 1988)'
39

one of the resulting clones, EXó45, has nou' been shoutn to be safe and immunogenic
in man (Forrest d øL rggg). Io "
recent cholera challenge t¡iar¡ vorunteers immunized with
Ð(ó45 and then challengedwith pathogenic v cholcræ excreted sþificantly fenrer bacteria
and suffered significantly less diarrhea than unvaccinated cont¡ols (Tacket daL,In press)'
Althougþ this study showed that antibodies ûo the oantigen ol v cholqae are sufücient to
confer protection agninst clinical disease, a greatø consistency of immune responses lltiu be
required before such avaccine warrants extended evaluation'
LllI4 Conclusion
To achier¡e the greatest public health effect in endemic areas an ideal choleravaccine
should be (a) safe without side effects, (u) easy to administer, (c) inexpensive in vie$/ of the
target population and (d) higily protective preferably after one dooe. clearþ none of the
vaccine candidatesiustdiscussedmeetallof these criteria. Because of its safetyandlong-lasting
protective efficacy, the combined inactivated vaccine is the onþ suitable formulation for
widespread use in endemic areas. Unfortunateþ, hou'ever, althougþ safe' the cost of this
vaccine, and the fact that murtþle doses are require4 rimits its utility. A geat'" drawback is
the factthatitprorrides only shortterm protection inyoungchildren, the groupwith thegreatest
risk of infectioo AlthouÈ the attenuated live oral vaccine being derrel"p"d by l-evine's gouP
elicits stronger immune nesPonses and would be much cheaper to produce' two problems
remein. Fint, the issue of residual reactogenicity is notyet satisfactorily resohed" secon4 the
safety and efücacy of such a vaccine remains to be demonstrated in young child¡en. As poinæd
out þ lGper ( 1989), rapid progress in vaccine reæarch makes it unlikeþ that another 100 years
will pass without a successful vaccine against cholera
40

1.11 Aims of Thesis
Atthe commenoementof these ¡tudies theprotective antigens or.v ùolsøe remained
undefined. Altlougþ circumstantial er¡idence suggested that antibodies to LPS a¡e prrotective
against human disease, the inactivated whole cell vaccine possess€s these antigens yet remains
inadequate. The preoenoe of non-LPS protective antþns on at least one Vclølqæ strain
(5698) appeared to merit further invertigation, to at least ascertain.the distribution of these
components emong recent field isolates. Such a stuf represents the maior aim of Chapter 3
of this ttresis. Clearþ, if such strains share non-LPS protective antþne, frEther study of theee
antþns miÉt evenûralþ identi$ trew oomponents of vaccine sþificance- Duringthe oourse
of these initial e:perinents, ehrdies þ Mekalanos' group (T¡ylor d aL 1987) resolved the
cont¡oversy surrounding pilus production by V choløæ. The critical involvement of TCP in
infant mouse pathogenesis offered a ne,s' approach to choleravaccine development It seemed
important to confirm these finding¡ and to determine whether TCP represents a protective
antþn ¡¡ this model This, together with other attempts to identify non-LPS protective
ant' ns ofV dtobæ, is the focus of Chapter 4'
ffis ¡im of the ñnal eryerimental chapter of this thesis is to clone genes involved in the
bios¡rnthesis of TCp. The derivation of clones eryressing TCP would allow unequivocal
confirmation of initial findingp concerning the sþificance of these pnli * virulence deter-
minants and protective antigens in the infa¡t mouse model
4t

Materials and Methods
2.! Bacterial sfains
M*, of the st¡ains used are shor¡,n in Table 3.1, which lists biotype, serotype, virulence,
sounee andyear of isotation (wherever poosible). All are resistant to streptomycin (100Fg/nl);
when neoessa¡y, such ree istance was selected þ growth on st¡eptomycin+ontaining ( 100 pdml')
nut¡ient ag'r Otqr; Difco, Det¡oit) plat€e Other straiûs used but not listed in Table 3'1 are
detcribed and discussed in Chapters 4 and 5'
2.2 Malntenance and grcrvth condiüons
permanentetoclg were stored in the þophilized state in 107o skim milk at 4 Ç and also
frozen at -70c in a mixh¡re of 0.4 nI SOvo (viv) dycerol plus 0'6 nl lvo peptone (Difco'
Bacto-peptone). strains wefe reconstituted s'ith nutrient broth (NB; Difco, Detrcit) as
required. Storking stock culürres wefe 6¿iatrined on NA stabo stored at rr¡om temperature
for periods of up to eightweels. For rr¡utine uee, the stabs were streaked onto NA pl'ates which
were then incub atÃat3l CovernigþL scrreralof the resultingcolonieswere used to inoculiaæ
10-ml aliquots of NB, which u,ere then shakenwithvigorous aerationat3T c unless otherwise
42

specified" such culhrres t¡picalþ attained a concentfation of 1-2 x 109 bacteria per Dl afær 3-4
hours.
Asloppy 4gar orrerlay techniçe (Stocker lg4g)was employed to ensure thatthe st¡ains
remained hrghly motile because motile strains have been shou¡n to be more virulent in infant
mice (Guentzel & Berry 1975; Attridge & Rou,ley 1983a)- A NA platewith 100-200 isolaæd
colonies was overlaid \tith t}-lzmls ef sloppy aga¡ (2 parts NB to 1 part melted 2Vo NA¡
prewarmed to 45 C). The Plates $'efe left at room temperature for 15 minutes to allou' the
overlay to se! and then incubated a¡ 37 C. Those colonies comprised of motile bacteria
gradualþ derreloped a halo as the organisms swam into the sloppy agar' MicroscoPlc obser-
vation ofbroth cultures seededwith thaloed'ornnon-haloed" colonies confirmedthe reliability
of this technique.
23 Speciatizùgrowth media
Water used to prepare media was deionised and filtered, using the Milli RO60 system
(Millipore corporation, u.s-A,). Media used to sû¡dy the produclion of pili n v chol¿ræ
stlains were the follorving
typtone waÛer (T"e"dy el aL lgæ)t lgBacto tryptone and 0'5gNaClwere dissolved
in 100 ml of water.
Iþpticasesoybroth(Farisetal.1982\:obtainedfromBal¡moreBiological
I-aboratories @BL), Baltimore.
FepSone salim brcth @vans et øt.lglS):ZgBfictopeptone and 0.5g NaCl dissolved in
100 mlg of water.
hptone sallne agar (Ev¡ns et al. 1fl8)z Peptone saline broth to which 2gor' Agaf
(Difco) was added.
43

Colonlzation Í¡ctor bnoth (CfB; Evans et al. 1979): consisted of 10g casamino acids
(Difco), 1.5gyeast extract (Difco), 0.059 Mg$Oa and 0.0059 MnClz in 1 liter of water'
colonizatlon f¡ctor agar (c:M; Evans er at. t979): prepared by adding Ba cto ag'tr (27o
wft) to CFB medium-
'ICç Medium (Ehar a et aL 198ó): consisted of 10g Bacto try¡rtone, 29 yeast extract
(Difco), 59 NaC[ 39 NaHCOI 29 monosodium-L-gfutalnate' 0.2gthioproline (sigma)' lmM
EG?\ (Sigma) and 20g Bacto agar in 1 liter of water'
LB Medir¡m (Miller 1fl2): was Pfepared by adding 10g tr¡ptone (Difco), 59 yeast
extract (Difco) and l0gNaCl to l liter of waterwith adiustme¡tof pH to 6'5'
V cholqac strains urere inoculated into the above media a¡d incubated at different
temperatures forvarying periods (see Chapær 4)'
When ap,propriate, antibiotics were added to broth and solid media at the follou'ing
final concent¡ations: ampicillia (Ap; Sigpa) 100pg/nl; kanamycin (Ih; Sigma) 50 t'üml,
tetracycline CIc; Calbiochem), 10 rgûl for.E coli a¡d a pdñ fot V eholqae (Chapter $;
str,epomycin (Sm; Glaxo), 100 P g¡mt
2.4 ChemicalE neagents and buffers
Reaçnts were prepared with deionised and filæred Milli Qwater as described above.
Chemicals were anaþical gade. Pheno! sodium dodecyl sulphate (SDS) and sucrose were
from BDH chemicals. l}is was Thizma base from Signa EDTA was fiom Sigma Bovine
sefum albumin (BSA fraction Ð was fncm Flow l-aboratories, Roclville, Md). cesium
chloride (technical g¡ade) was from May & Baþr'
The follorrringelectrophoresis gaÃereagentswereobtainedfromthesou¡cesindicated:
acrylamide and ammonium persuþhaæ (Bio-rad), ultra pure N,N'-mettrylene-bisacrylamide
u

fron Boehringer Mannheim, lVestcermany. The subotraæ for immunoblottingwas 4-chloro-
l-naphthol (Siena).
Buffers and solutions were also prepared with Milli Q deionised water. Saline was
o.85ro 1wfu) NaCl in waler, while phosphate buffered saline (PBS; prl7'2) comprised of 8g
NaC[ 0.2g KCl' 1.159 NaZHPOa and 0.2g KHZPO¿ in 1 liær of water.
L4.L Enzyre-thlrrd irrnunosorbent assay (EtISA) Bufiers
2SmM tis-HCl þH ?.5): was made þ dissohing 3.039 TLis base in 900 ml of water
and adiustingthe pH tD7,'with lM HClbefore makingthevolume up to l liter'
Ihis-saline.Adde(TSAc-oatingbuffer): preparedþdissotvingT.T2gNaCl (132mM)'
3.039Tris (25 nM) pH 7.5 and 0.5gNaN¡ in 1 liter of water.
\ltrashing bufier wlth tlton X-1011 prepared þ dissolving43.83gNaCl 3'03gÎis' 5ml
titon x-100 in 4000 ml of water and adiusting the pH to7.6with ap'proximateþ 18 ml of lM
HCI before making the volume to 5 lite¡s'
O¿S M tiethanolamlne: 3T.Sgtriethanolaminewas neutralisedwith 2;S\ÁHCI (ca 60
mls) to pH7.6; 1g NaNf was added and the volume made up to 1 liter with water'
BSA[Ibeen: l6gNaC[ 2gNal|[r 0.4gBSA 100 mls of o.25 M triethanolemine' and 10
mls of lO% Tttæn 20 in 2liærs of water'
hzyme Dtluent (for dilutin gentymelattibody conjugate): 8g NaCl was dissolved in
g00 ml $ratet, follou,ed by the addition of 50 ml of 0.25M triethanolamine pH 7-ó and 2mls of
500XMg* * lZo+*(SOOnÀ,f MgClzll.zímMZnClz),0.2gBSAand0'2gNaN¡; thevolume .
was made uP to 1 liær.
45

2,42 IÞtergents
The follou¡ing detergents were used in an attempt to solubilise oute¡ membrane
preparatioñ of V choleraez
ionic:Sodiumdeo:rycholate(BDH)&sarkooyl(Ciba6eþ);
non-lonlc: Ti.iton x-100 (BDÐ, Tliton x-114 (serv4 Feinbiochemica, Heidelberg)'
Tneen Ð (Signa), Bfij 53 (Serva), Nonidet P40 (BDH), Urea (BDH-Analar), deærgent test
kit No ?ßYs}(Calbiochem Behring Diagnostics, La Jolla, California);
zwltterionlc Zwittergent detergent test kit No. 693030 (Calbiochem)'
L43 nnzlm and inrnrnorcoq¡qgatês
The following enzymes were obtained from gigme: deo:ryribonuclease I (DNAse I)'
ribonuclease I (RNAse I), þozyme and hyaluronidase. honase and ProÛeinase Kwere from
Boehringer-Mannheim.
Restriction endonucleaseo Banr{l[\ r/;oR:l, San3A aDld J(bal were purchased from
Boehringer-Ma¡nheim. DNAmodisingenzJmeswerepurchasedfromNen,EngfandBiolabs
(t4 DNAligase) andBoehringer-Mannheim (DNApolymeraseI, Klenow fragmentof DNA
polymerase I and moleculiarbiologr grade, calf intestinal alkeline phoophatase)'
Horseradish peroxidase-coniugated goat anti-¡abbit and goat anti-mouse IgG were
obtained from Nordic Immunological Reagents (Tilburg the Netherlands)'
25 Animals
2-S.l Adult uice
Adult female closed-colony inbred outbred I-{C mice urere obtained from tle Central
fl¡imal House of the university of Adelaide- These micewere bred under specific pathogen-
ß

free conditions and subcequent$ mored to a conventional animal room These micewere used
for preparation of antise¡a and as a source of enterocytes for h vìtro studies of bacterial
adherence.
L52 Inhntmice
Abreeding colony of conventionalized I-AC mice was maintained in the Medical School
Animal House of the University of Adelaide. Each breedingcage containedfour female mice
and one male, and the breeding stock was replaced at six-monthly inærvals. The infant mice
wereusedatabout5 da¡,sof age (weig[trange restricted toL4-275g) forstudieswiththeinfant
mous€ cholera model (see belov).
2.6 Preparation of hiled and formalin-killed organism s
(i) Boiling Bacteriawere gro\r,n in NB washed ¡n'ice in saline, then resuspended to a
concentration of approximatery 1010 per mL Aviable countwas performe4 followingwhich
the suspension was heated to 100 c for 2 hou¡s The organisms were then washed three times
in sali¡e and resuspended in såline to lìe original volume; aliquob wele Pl'ated onto NA to
ensure thatnoviableorganisms remained. o.LTo 1w&) Nal.I¡was added to thesuspeneionwhich
was then stored at 4 C.
(ii) Formalin-inactivaüon. An equal volume ol l7o (vfu) formalin was added to a
suspension of V cholqæ (at approximately 1010 Per ml ¡o PBS pH7.2). After incubation at
37 C for 60 minutes the bacteria were washed three times with PB.S and resuspended in saline
to the original volume; aliquon $'ere plated on NA to ensure that no viable oraanisms
remained. The suspension was stoned at 4 C with 0.1% (wfr) NaN}
47

2,7 Isolation of LPS fiom V cholerae
LpS was isolaæd from several strains during the course of these studies fo¡ the
production and absorption of antisera, and sensitization of RBCs for hemaʀlutination assays'
LFS was extracted from cells with hot 90% 1wft) phenolÄn ater using the procedure of Wesþhal
& Jann (1%5). Briefþ, a 100 mlbacterial saline suspension (20 ng/ml dryweigþt) was heated
to 6g q An equal volume or.9TVo phenol'rrater (pren'armed to 68 C) was mi:red into the cell
suspension and stirred continuously at 68 Cfor 30 min. The miÉure!\¡as allouted to cool and
the resultingphases reparated þ centrifugation in gþss buckets (1500 rp', 20 min' Coolspin'
MSE). The aqueous phase was collected and stored at 4 C The phenol phase was re-extracted
urith half the original volume of preurarmed water (68 C) for 30 min. with stining and the
phases aFin separated þ centrifugation. The aqueous phasee we¡e combined and dialysed
æainst 5 liters of water, overnight at 4 C Insoluble material was pelleted by centrifugation
(5009 for 5 min ¿1{ e i¡ an SS34 rotor, Sowall). The LPSwas precipitated from the supernatant
with 5ó votumes of cold ethanol containing 250 mg of sodium acetate powder. The precþitate
was collected þ centrifirgation (am0g for 30 min- at 4 C in a Sorvall GSA rotor) and
resuspended in 25 ml of distilled water containing 5 nl\d MgCIZ A small quantity (ap
proximately 1 ng) of DNAse I and RNAse I was added and incubated for 60 min' at room
temperature. This was folloured by the addition of pronase (1-2 mg) and further incubation
for 30 min The mixture was again centrifuged at low speed, (30009 for 10 min' at 4 C in an
ss34 rotor), before deposition of LPS þ ultracentrifugation (100,000g1or Zhr at 4 C; ó0 T:r
rotor in a Beckrnan I-SA0). Each pelletwas then suspended in 5 ml distilledwater, of which
0.25 mlwas used for dryweigbt and 0.1 ml for estimation of proæin þ l-ou'ry's method (I¡wry
e14t. LgSl)usingBSA as standard. Lps preparations obtained iû this manner usually contained
rnore t\a¡ lVo contaminating protein and were tìerefore su$ected to a second phenol
48

extraction. This then reduced the ler¡el of protein contamination to <lVo. All preparations
u,ere stored at 4 C in the presence ol.o.lTo lwrt) Nal'lr
2.8 PreParation of OMPs
preparation of oMI)s was car¡ied out as previousþ described by Mannin get øl' (1982)'
Bacteria u¡ere cultured in NB attl cfor 18 hr, han'ested by centrifugation in a sorvall GS3
nctor at 8,0009 for 15 min, and reeuspeaded in 50 ml of PBS, pH 7'L Af'¡e¡ disruption in a
French pressure cell (Aninco; siherspring MD), the þatewas clarifiedþ centrifrrgation at
10,0009 for 20 min, and the oMp in the supernatantwas recovered by ultracentrifugation at
100,üx)9 for 90 min at 4 C (45Tr rotor; Beckman)' Each of the resulting pellets wa's
resuspended in 5 ml of distilledwater; after pooling protein contentwas determined using the
nethod of l-o*'ry er al. (1951).
For eolubilization of oMPq 1 ml of oMP (2 mg/ml) and 1 ml of 50 nl\'f lis-Hcl (pH
?.5) with or without detergentwerc incubated at 37 cnot t hr. After incubatio¡' the mixture
w¿rs spun at 100,0009 for 30 min at 4 C The pe[etwas washed twice with rris-HCl and finaþ
resuspended in l ml of lxI-¡¡g(sample) buffer, aliquotted and frozen atâC
2.g Prtduc{ion of antisera to wlrole cells and to OMPs of
V cholerae
Lg.L heParation of muse antisera
Hyperimmune antisera to whole cells were prepared by intraperitoneal immunization
of mice with increasing doees (either rd-ro7 or to7-ro) of üve bacteria; animals were given
fortnigþtly iniections for 10weels. Bloodwas withdrau¡n from the retro-orbital plexus under
ether anesthesla lo_12 da¡n afær the last immunization and allw'ed to cloü 6era wefe
49

sepafateq fittered (pore sizæ,0.72 pm; Millipore Corp, Bedforrd, Mass), and stored at 4 C or
at -?nC in aliquots containing 0,O2Vo (w&) NaNa
For the preparation of antisera to OMPs mice were sequentially immunized in-
traperitoneally on d4¡s 0, !4,21and 28 with 10, 30, 100 and 120 pg of OMP extracted with
buffer alone (control) orwith ZVosf|rkæyl (SarkOmP, seeSection 4'2;2) respectiveþ' Mice
were bled 10 da¡rs afær the last immunization and sera collected as described aborte.
L92 heparation of rabbit antisera
Adult rabbits were obtained from the Central fl¡imal House of this University for
production of antise¡a towhole cells þ a combined subcutaneous/intravenous immunization
pr.tocol Equalvolu mæolH?osodiumalginate andwashedcholeravibrioo (101d in normal
saline)weremixedand0.25 mlof themiÉurewasdepositedsubcutaneousþ; withoutremoving
the needrg butusinga neu, syringe,0.r5 mrof.o.4%ocaclzwas injected intothe Bame site. This
pÍocedure was repeaæd a! three other sites. At the same time 0.5 ml of the washed bacterial
suspension was adminisæred intravenously. At 34 dap intervals thereafter, increasing doses
of live v chotsæ1r07-ro9nt in 0.5 ml saline)were given intravenousþ. The rabbitswere bled
by cardiac puncture under anesthesia 10 dãlts after the last immunization' The senrm was
collectd poole4 filtered and stored as described above.
The method used to prepare antisera to OMPs was essentialþ that of Boulard &
Lecroisey (19g2). 5698 oMp was elect¡ophoresed in a[ 16 tracts of an 11-207o SDS-PAGE
After destaining tle gelwae inmedi¿æþ cut into individual t¡acks containingall the proteins
and kept at 20 c The rabbit was immunized witlout aduvant þ sc injections of the gel
homogenates of one track on day 0, three tracks on day 10 and 6 t¡acks on day 17. The rabbit
was bled 10 days after the last iniection as described above.
50

2.lO AbsorPtion of antisena
Antisera were varior¡sþ aboorbed with LPS or oMP isol¡ated fuomV clplqæ' or with
intact bacteriq as described below'
Z1O1 Absorption ç'ith LPS or OMP
Antiserawere absorircd three or four times with LPS (0.1 mg/ml) or OMP (1'0 ng/nl)
as required. Alternate absorptions were incubated at3' Cfor 2 hr and at 4 C orernigþü the
latær ûo allow the remorral of less avid antibodies. After the first of each absorption series the
suspension \[ras slnrn at low speed to remorre the visible preciflAæ; with subeequent absorp
tions, LPS or OMP wefe remorred þ ultracentrifugation at 100,0009 (Bechan IJI80)'
2J:o;2 AbsorPtionwith bacteria
Unless specified otherwise, antisera were abno¡bed three or four times with live or
boiled vchobæ 1s x tolorhl) with alternating incubation conditions as descriH above-
After each abeorption, bacteriawere removed by low speed centrifugation (4,000Ð, "od
at the
end of the absorption series the serum was sterilized by Paseage througþ a0'72 pm fi'lter'
2.L! rn vitro assays ofv cholere adherencq attachment to
isolated murine inæscnat epithelial cell s (enteroc5Úes).
LLLI PreParation of enterocYtes
The method used for enterocyte preparation is a modification of that descriH bry
curman er øL (rylg). The mice wefe killed þ cervical dislocation, and the entire small
intestines excised and flushed$,ith 50-100 ml of ice<old PBS (PH 7'2)' A1ær trimmingaway
51

adhering mesentery and peyer,s patches, each intestine was slit longit'dinally and rinsed in
pBS before being cut into !-2 cmrdpu; these \\rere incubated in ice+old PBS (containing
smMEDTA)withmildagiøtionfor15min.ThestripswerethenwashedincoldPRSand
incubated in preurarmed PBS containing 1 mM GrC)IL 1 mM MgCIZ 1 mg BSA anrd? mg/nl
hyaluronidase for 50 min at37 Cwith intermittent shaking
The resulting enz¡¡matic digest was filæred througþ a nylon sieve to separate the
epithelial cells from tissue segments and large cell aggregtes' The filæred cells were sedi-
mented by centrifugation at 100gfor 3 min at 4 G washed twice in pBs and resuspended in 2
mro',*vo(vfu) percoll (pharmacia uppsala sr¡¡eden) conraining3 nlr{ dithiothreitol (signa)'
The suspension was then layered over a discontinuous percoll gradient (1 ml layers oî'70To,
6oTo ardsovo a,,da}-mfiayer or.4ovo1vfuD in screur+apped siliconized glass tubes. The cells
were centrifuged at 1009 for 10 min at 4 c in a bench centrifrrge (MSE) with a suringout rotor
for separation of enterocytes from þmphocyte* The purified enterocytes were washed twice
\rrith PBS and resuspended in 1 ml of RPMI 1ó40 medium (Flou',)' The total cell countwas
determined in a hemocytometer, and the concentration was adiusted to l-Zx 105 cells/ml by
addition of RpMI 1640 medium. Viability was assessed þ exclusion o10.2vo (w&) trypan blue
andwas generallY ffiTo.
LLLZ InYit¡o enÛeroc5úe adherence ass¡ay
For each a.ssay 0.1 ml of bacterial suspension (1-3 x tOT lmt in PBS) was added to 1 ml
of freshþ prepared enterocytes (r-2x105/hl in RpMI 1640 medium) in disposable plastic
centrifuge h¡be3 (sterilin, Middlese¿Engfand). heliminary e:rperimenß had indicated that a
ratio of 10 bacteria per enterocyte was optimal for assessment of bacterial adherence. Afær
incubation at37 Cfor 15 min, ttre enterocytes $'ere sedimenæd þ centrifugation at 1009 for
3 min, and nonadherent bacteria removed by three 10ml washing with PBS' The pelletwas
52

rcßuspended in 1 nt of incubation medium and homogenized at full speed for 15 sec (Ultra
Thrraa Janke & Kunkel, Staufen, Federal Republic of Germany). Suitable dilutions of the
homogenate were made in normal saline and plated on NA plates containing Sm. Percentage
adherence was calcul¡ated by relating the total number of bacæria adhering to the enterocytes
to the number added iûitirallY.
LLLS Inhibition of adherere.
An assay was developed to assess the capacities of various antisera to inhibit the
attachment of vibrioo to enterncytes. Bacteri¡a were pretreated with the test antisenrm at
various dilutions or with normal mouse serum at a final dilution of 1:10 for 10 min at 37 c
They u,ere then added to the enterrcc¡rte suspension for determination of residual adherence
capacity as described aborre'
2.Í2 In vivo assays rvith infant mict
The infant mouse cholera model was first described þ Ujüye d 4L (196) and sub-
requently eÉended þ Chaicumpa & Rowley (1972)who demonstrated thatspecific antisera
could protectmice from operimental cholera This model has been used i¡ these studies to
a'sess the vinrlence of different strains ol. V ehabæ and to determine the efñciencies with
which various antisera a¡e able to provide protection against e:perimental cholera (Attridge
& Rou,ley 1983a 1983c).
The infant mice used in tìese assays (Section 25.2)were removed from their parenß
about 5ó hours before use, to permit emptying of stomach contents. Unless stated otherwise,
the challenge strain u'as grorl,n in NB at37 C to a concentration of apprrcximatelt 109/mt
Depending upon the challenge dose, the organisnÁ¡ were either simply diluted or alternativeþ
53

spun out of growth medium and resuspended in PePtone saline (PS, a O'lVo YN solution of
proteose-peptone [Difco] in saline). After appropriate dilution' 0.1 nl aliçots wefe ad-
ministered orally to infant mice using a smooth-tipped hypodermic needle (nù' Afær
challenge the mice $,ere not rehrrned to their mothers but were kept in tissue-lined plastic
containers in a laboratory incubator at 25 G
LIIL,L Virulence asñ¡alt
Serial five-fold dilutions were prepared from a PS suspension of the Ûest strain of
vchobæ, ild each was used to feed one gouP of 8-12 mice. Forty-eigþt hours after
challenge, the survival of mice within each group was noted, and these data used to construct
a plot of cumul¡ative percentage mortality versus logro challenge dose (using the method of
Reed & Muench 1g3s). Ð inærpolation itwas then possible to determine tle (48 hour) LDso
dose for a given st¡ain - that is, the number of orgaisms capable of killingsovo olt\e mice to
which it is administered.
LL2,2 hotect¡on atxnlls
Five-fold dilutions of the test serum were used to pre-treat separate aliquots of a PS
suspension of the challenge o¡ganisms After incubation at 37 c for 15 minutesr these mixtures
were fed to different groups of infant mice (g-12 per group). The bacterial concentration in
the originar suspension had been arranged such that each mousewould receive 20 LDso doeee
in the 0.1 ml challenge inoculum. Control mice received organisms which had been incubated
in pS alone, or in the Presence of normal mous€ serufll The sunrival of all gouPß was then
followed until all of the control mice had died; generalþ this occurred,42-52 hours after
challenge (whereas mice given PS alone consistently survived fotT?hours)'
tå
54

ml.
Following the plotting of cumulative percentage mortality against logtO senrm dilution
(ReÆd & Muench 1938), the protective endpoint of the serum was obtained by interpolation
as that (theoretical) dilution which could protect 50% of the mice from death- The reciprocal
of this dilution was referred to as the protective titer of tùe serum, expressed in PDso units per
2.LS Isotation of V cholerae Pili
Glass tra)¡s (30.5x30.5 cm) containing5gg ml of CFAwere seededwith 109 (nonmotile)
v crwlqæ 5698 and incubated for 36 hours at ?5 C. The bacteria were han'ested and
resuspended rn lz|mM Tris-HCl, PH 7.0 containing 25mM NaCL 4mM MgCIZ and 4 mIVî
caclz (laylor et at.19SÐ or in 10 nM tis buffer, pH 7.5, (Korhonen et al' 1980) or in PBS pH
7.2 (/J'-lß rssi & Mostratoo 1985), to a final cell concentration of 0'14 g/nt Cells wefe sheared
by a 5 min fi.s¿tment in a Sorvall Omnimixer (at half speed) on ice' The suspension was clarified
bycentriñrgation at2í00gandthe pelletresuspendedto its originalvolumeandfurtlersheared
þ two passages throu fi a?jl-gtnge needle. After another clarising spin' the two supernatants
were pooled, treated with 507o saturated ammonium sulphate and left ovç¡night at 4 C The
resulting precipitaæ was centrifuged (10,0009 for 30 min at 4 C) and the free pili recovered
from the supernatartþ ultracentrifugation (100,0009 for t hr at 4 C) and suspended in 2 ml
of pBS This suspension was ap,plied to a self-generating isopycnic cesium chloride gradient
After ultracentrifugation ( 150,0009 for 1g hour¡ at 4 C) three distinctba¡ds were obseNed and
thesewere co[ecæd þ puncturing the side of the tube with a rg-gatgeneedle and aspirating
with a syrinæ. Afær removal of cesium chroride by dialysis against pBS, the three fractions
\f,'ere analysed by SDS-PAGE and þ transmission electron microscopy'
.t
I.ll
È"
I
I
\i
'l
'l
If,
i
i
ì
55

I
ì|*
2.L4 In vitro titration of antibodies to V dtolerae
LI.+l Hemggþtination
For effrcient sensitization of sheep RBCs (SRBG) with LPS, it was necessary to
pre-treat the LpSwith alkali. LPSwas diluæd to a final concentration of 2mf,mland tre¿ted
with 0.02N NaOH (usi¡g lN NaOH). After overnigbt stirring at room temperaûue the pH
was restored to neutrality (þ addition of concentrated HCI) and the material stored at 4 C
with 0.17o 1wft) Nal.ll
SRBC. were washed tlree times and resuspended to a concent¡aion oî.257o (v&) in
saline. Alkali-treated I-pS was added to a final concentration of 50 pdml and tle mixture
incubatedwith continuous rollin gat37 Cfor90 minute¡. The SRBCswere againwashedthree
tines, and finalþ resuspended to lVo (vN\ in saline. Sensitized SRBG were ahn'ays freshly
prepared on the daY of use-
Serial two-fold dilutions of the test sera or normal serum were made in saline in plastic
microtiter trays (Disposable hoductg Australia). A standard serum of knou'n hemagglutinat-
ing capacity was included to check the efüciency of erythrocyte sensitization. Following the
addition of an equal volume (2ti) of. a lVo (vr!) suspension of sensitized SRBG to each well'
the trays were shaken briefþ and incubated at37 Cfor 60 minutes. The hemagglutination tiær
wÍrs exprex¡sed as the reciprocal of the higþest serum dilution effecting an RBC settling pattern
clearþ different from that seen in control wells (lacking serum).
2.L+2 Í'I rSA
ELISA was performed in round-bottomed vinyl microtiter plates (Coßtar, Cambridge'
Mass) using buffers and solutions described in Section L[.L. Tays were sensitized with oMPs
of Vchaleræ (see section 28) at a @nceûtration of 10¡g¡ml in TSA buffer (coatingbuffer);
,l¡
;I
t:I
I
I
56

l00plof antigenwas addedto eachwelland incubatedat3T Cfor 2hours follow'edby afurther
incubation overnigþt at 4 C Afær washing th¡ee times with washing bufreç the wells were
blocked with lÐpl of BSASveen for 6-8 hours at 4 Ç and then washed agnin as before. To
assay antibodies in senrm, dupticate 4-fold serial dilutions were prepared using BSAfin'een
and incubarcd overnigþt at 4 C The trayr were then washed four times and bound antibody
was detected þ application of a goat anti-rabbit gfobulin alkaline-phoophatase conjugate
(kindty supplied by Dr. Peter Ey of this Deparmen! optimal dilution of 1:15ffi) in enzyme
diluent (100 pl per well). After overnigþt incubation at 4 Ç the trays were again washed four
times and 100pIof 1 mg/ml subotrate solution (pnitrophenyl phoophate disodium [sigma] itr
10Zo diethanolamine buffer) was added to each well After incubation for 3 hours at 37 C, the
resulting color derrelopment was read at 405nm using a Trtertek Multiscan automated
epectrophotometeç with reference to cont¡ol wells which had received diluent instead of
antibody dilution The antibody tiær of any g¡"en sample was defined as the recþocal of the
dilution which produced a meatr oD of 0.150. Normal and positive serawere ahvays included
as controls.
2.15 SDS-PAGE and immumbloüing'
z,Ls'I SDS.PAGB
sDs-pAGE of oMp, whole cell þates and other protein fractions wefe performed on
Í-n%polyacrylamide gradients or straigþt l2Vo potyacrylamide çls usinga modificati'on of
the procedure of Lügæ nbetgel al.(195) as described þ Achhan d al. (1978)' Samples rvere
resuspended in lx Lug (sample) buffer and heated at 100 C for J min Priot to loading Gels
were generally electrophoreæd at 100 V for 4 houre Ql-n% gradient gels) or 25 mA constant
current (l2To pAGE gels). hoæin stainingwas achieved þ incubation wernigþt with gsntle
agitation with Coomassie Brillia¡tBlue G250 hSoVo (v¡V) methenol and lÙVo (vN) acetic acid'
57

Destainingwas achier,red usingser¡eral changes oî.7.SVo (vrt) acetic acid, lÙVo (vlv) methanol
a¡d tOVo (vfu) ethanol over 24 hours'
Molecular weigþt markers (Signa) were a-lactalbumin (14,200), ß-lactoglobulin
(bovine milk; 18,400), trypsin inhibitor, soybean (20,100), tfyP6inoggn (24,000)' carbonic
anhydrase (29,000), gþceraldehyde-3-phosphatedehydrogenase (36,000), ovalbumin (45'000)'
bovineserumalbumin(óó,000),phosphorylaseB(97,400)'ß-galactosidase(116'000)and
myosin (Ð5,000).
L\5.2 lrnrnrnoblotting (Westem Hotting)
After SDS-PAGE the proteins Ìtere electrophoretically transferred to nitrocellulme
(schleicher & Schüll, Dassel) for 2 hours at?Ð}mA in a transblot cell (Bio-rad). The transfer
buffer used was 25mM Iiis-Hcl, pH 8.3, containing 192 nI\d gþcine altdS%o (vfu) methanol
After transfer, the nitrocellulose sheet was incubated for 30 minutes h SVo (wfu) skim milk
powder in Tpeen-Täs-buffered saline (TTBS; O.OSVo [vfo] ßveel-2f.,?Ã mM Täs-HCL0'97o
[wþ] NaCl) to block non-specific protein binding sites The nitrocelluloee was tlen reacted
with antiserum by the method of Towbin et aL (1979). This was then follou'ed þ overnigþt
incubation of the nitrocellulose at ¡oom temperature in a 1:21000 dilution of a horseradish
peroxidase+oniugated goat anti-mouse or anti-rabbit IgG' Binding of p'rimary antibody was
detected by 5-10 minutes incubation at room temperature with the substrate (10 mg 4-chloro-
l-napthol [Sigme] dissoþed in 3.3 mls 99.57o methanol and mixedwith 16'5 mls of Täs-buffered
saline (IBS; 20 mM Tris-HCl, O.9Vo fw9l Nacl) containing 30 pl H2o2) as described by
Hawkes et al. (1982).
58

2.L6 ColonY blotting
Anitrocellulose disc (9 cm diameter Schleicher& Schäll, Dassel)waspliacedonto agar
plates containing the colonies to be screened. once the colonies had adhered to the disc (1
min), itwas removed and placed, colony side up, on a piece of \l¡hahan 3 MM papef ; bacteria
u,ere lysed ín siruwith 0.5 M HCI and left in the dark for 30 min, according toHautkeÆ et al'
(19g2). The cell debris was removed from the nitrocelluloee with a iet of normal saline' The
method used for antigen detection was the Bame as that used follor¡'ing\Vestern transfer'
2.[l Construction of V cholerae gene bank
LIIZ.L Pneparatbn ofV clolerae çnonic DNA
Thiswas essentially as describedby Mannngel at. $9rß} cellswere groum overnigùt
q¡ith aeration at 37 c in 100 ml of brain heart infusion broth (BHI, Difco). Bacteria were
centrifuged at room temPefature and resuspended i¡ 2 mls cold2SVo 1wft) sucfose in 50 mM
T'is-Hcl (pH S.0). To this was added 1 nl þozyme (5 mg in 0.25 M EDTA pH 8'0) and the
miÍureincubatedonicefor 10min. Aftertheadditionof pronase (10ngin lmlof TEbuffen
10 ml\d rtis_HCl, 1 mM ED-,[A, pH g.0), the mixt.re was incubated for 1$ minutes at37 C
followed by the addition of 0.ã ml of lysis solution (svo sarkooyr in 50 nN'f rristrIcl, 0.25 M
EDTA, pH S.0). Thiswas i¡cubaæd at 56 C for 60 min with occasional gsntle su'irling' The
þate was then gently eiracted three times \ñ'ith TE buffer saturated with phenol (prepared
þ adding 1g phenol per ml of TE buffer). Residual phenol $'as removed by exÍracting twice
with diethyl ether, and the DNA solution was then dialysed orrernight against 5 liærs of rE
buffer at4C. The DNAyield was routinely S00 to 1,3t0 p/ml per 100 ml cultures'
59

LL7,2 DNAquantitation
The DNA concentration was determined þ measurement of absorption at 260 nm'
assumingan Azroof 1.0 conesponds to S0pgDNA/ml (Miller L972).
Lyl.3 End fillingwith l(lenow fragment
protruding ends creaþd þ the cleavage with EcoRl arrd&antHl were filled using the
Klenou, fragment of E. coli DNA poþmerase I. Typically 1 pg of digæted DNA 2pl l(h nick
translation buffer (Maniatis et al. t9g2), 1¡l of each dNTp at 2 mM concentration, and 1 unit
Klenou, fragmentwere mixed and incubated at37 Cfor 30 minutes. The reaction was stopped
þ heating at 60 C for 10 minuteg followed þ removal of unincorporated dNTP and enzyme
þcentrifugation througþ a l mlsepharose cLóB column-
Ln,4 C-onstmction of PPMÍ1101
plasmid ppM2l01 was constructed as follou's (Figure 5.2). Approximateþ 1pl of DNA
of rhe plasmid pHCyg (obt-ined fromDr. J. Qsllins; Hohn & C.ollins 19s0)was digested$'ith
the rest¡iction endonuclease fuoRj, The protruding ends produced by cleavage with EcoRl
were filled using Kleno*, frament ol E colí DNA poþmerase I. The plasmid P$I''P201-1
(obøined from Dr. d Pühler; Simon et 4L lg83) was digested witi the rest¡iction en-
donucleas e1mt1ltoyield a 1.6kb fragmentcontainingtle mobilization region of the plasmid
Rp4 from psup201-1. This digest was similarþ treated with the Klenou' fragment ol E' coli
DNA polymerase I. leated pnCTgwas ligated evsrnight to treated PsuP201-1 usinga 1:10
ratio and transformed into E. coli s1?-1 Qtro, hsdR, recA; and integrated plasmid RP4-2-
Tc::Mu-Km::Tn7) selecting for ApR and TcR. The cloningof the 1.64kb mobilization region
wa¡ selected for by looking for mobilization into E. cotiKl2 Sm10 (thí thr lat, SupE; KmR;
60

and integrated ptasmid RP4-2Tc::Mu), selecting for Ap& TcR and KnR Any positive clones
ufere further screened by making DNA preparations using the three-sæp alkali þis method
deecribed by Garger eJ at. (rgg3) and digested with the restriction endonucleases PsfI and
Bøntillto shou¡ 1.61ù increase in this fragment of positive clones'
L17.5 Construction of cosmkl gene bank
Genomic fragments of approximateþ 40 kb were generaæd by controlled partial
digestion ofv clprqæzrTsírgenomic DNAs'ith the restriction endonuclease sø¿3d The
coomid vector ppM2lol was clear/ed with BamHI and treated with molecul¡ar biologl grade
alkaline phoophatase (Boehringer Mannheim Biochemicaþ sydney) to prevent self-lþtion'
The two DNAs were mi-ed, lþted overnig[t and packaged ín vito ntobacteriophage lamMa
using a Promega Packagene kiL
The packaæd phaæ were then used to infect recombination deñcient SecA) E' coli
K12 517-1. Cells harboring cosmid clones were detected by plating onto nutrient media
containing ampicillin. Greater tha¡ govo of the colonies $'ere Tcs and could be assumed to
contain V dtobae DNA"
?.!7.6 Mobilization of osnid bank
The cosmid bank of Z l7 s6l n E cotíKl2 s 17 - 1 was mobilized into th e v chob æ strain
O17 (El Tor Ogana, S-R). This was done by replica-plating the cosmid bank onto a lawn of
o17 celrs er\ r¿rof log phase culture) on nutrient medium and incubating at 37 clor 4 hours.
These coniugation plates were tìen repricated onto nutrient medium cont^iningAp (Sopg/ml)
and Sm (200 pglnl). After overnigþt incubation the colonies were re-patched onto CEA agar
containin g Ap and scree¡ed-
61

L17.7 Ïhansf,omtion
This is basicalþ a modification of Brou'n et at. (1979b\. E colí K12 st¡ains were made
competent for transformation witl plasmid DNA as follows: an overnigþt shaking culture (in
NB) was diluted 1:20 into BHI a¡d incubated with shaking until the culture reached an ODeso
of 0.6. The cells wefe chilled on ice for 20 ltrh, pelleted at 4 c in a bench centrifuge,
resuspended in half volume of cold 100 mM Mgcl¿ centrifuged again, and resuspended in a
tenth volume of cold 100 mM C-¿clz. Thiswas allon'edto stand for 60 min. on ice, afterwhich
üe cells wefe deemed to be competent, and 0.2 mlwere mixedwith DNA (volume made to
0.1 mlwith 10 mlvf 1iis-Hcl, 1 mI{ EDTA, pH S.0) and left on ice for a frrrther 30 min' The
cell/DNA mixture was heated at 42C for 2 min, then 3 ml BHI was added and the mixture
incubatedwith shakingfor l-2hr atTl} The culürrewas plated onto selection plates directþ
or concentrated by centrifugation before plating Cells with sterile bufferwere run as a control
2.r8 ElVf
Z1&1 lhansrrÉssion EIVÍ
For negative staining strains of.V cholerøe u,ere suspended in PBS to 109 cells/ml One
drop of this suspension was applied to a Formrrar+oated coPPef g.ld (300 mesh, Graticules
IJd., Tonbridgg, Eog) for one minute, washed once with distiled $rater, then stained with tVo
uranyl acetate for 1 minute. Excess fluid was removed þ filter paPer' Grids were then
examined in a Jeol JEM 1005 electron micrrcscope operated at an acceleratingvoltåge of 80
KV
62

LI&2 Imnnnoplecüun nicrcscopy (IHV[)
Colloidal gold particles (ca. ß-?Ð nm) were prepared using the citrate method as
describedþ de Mey & Moermans (19s6) and coniugatedwith Protein A (Pharmacia).
V cholcræ cells grourn on NA or CEA plates $'ere harvested and washed once in PBS
(pH 7.2) conraining lvo BSA(PBS-BSA) A drop (35 pl) of cell suspension was placed on a
sheet of parafilm. A poly-L-lyshe tfeated formrar electron microecope grid was then placed
plastic side down on the surface of rhis drop for 2 min (Mazia et aL 1978). Th" grid was then
successiveþ transferred to drops of the following reagpnts placed on the same parafilm sheet:
pBS-BSA rwice (1 min each); anribody (1:10 dilution in PBS-BSA containing O.OSVo Tveen
20) for 15 nin; pBS-BSA rhree ':m€s (1 min *h); hoæin A-gold (1:50 dilution in PBS-BSA
containing o.117o5veen 20) for 15 min; distilledwater, three timec (1 min each). Excess fluid
was removed from the grid using filær paper and the grid allowed to air dry without negative
staining In some elperimentr, immuno-gold labelled preparations were negativeþ stained
with a solution of27ow¡! uranyl acetate for 1 min atroom temperature. Gridswere examined
in a Jeol JEM 100s transmission electron microocope operaæd with an acceleratingvoltage of
80 kv
63

Non-LPS PnotectiveClassical and El Tbr
3.1 Introduction
gnrdies in various model systems har¡e shou,n that antibodies to the LPS determinants of
Vcholqæcan mediate protection against experimental cholera (tilatanabe & Verwey 1%5;
Neoh & Rowþ lgzl svennerholm 1975; Holmgren et at.1977; Auridge & Rowley 1983c)'
In consequence the poosible prophylactic potential of the non-LPS components has been
largeþ ignored. As discussed in Section 1.10, conventional cholera vaccines generally com-
prise preserued suspensions of vibrios inactivatedþ treamentwith phenol' formalin or heat
despite tìe presence of LPS antþeng such vaccines have conferred onþ short-term protection
in the field (Joo lnÐ. The reatization tìat other bacterial enteropathogens po6sess labile
surface components that can function as protective antigens (Rutter & Jones 1973; Morsatr ef
at.lng)promptedtle evaluation of theprophylacticsignificanceof theno¡-LPSdeterminanß
oî.V cholqæ.
In 1970, Feeleyhadshown thatlivevibrioswere more effective than heatedones in the
active mouse protection test, raising the possibility of heat-labile protective determinants
(F"eley lnÐ. Subsequentþ, Neoh & Rowley ([gZz)showed tlata mouse antiserum prepared
asainst vcholqøesógB, and then repeatedly absorbed$rith higþty purified homologous LPS,
u

retained significant protective activi$. Bellamy et al. (1975) went further to show that an-
tibodies present in an IgG fraction of a similar rabbit antisenrm u'ere protective and direcæd
against heat_labile non-Lps determinants. Finally, more recent studies compared the protec-
tive activities of antibodies to the Lps or non-Lps antigens ofv cholqæ 5698 and concluded
that the liatter more efücientþ protected infant mice from challengewith this strain (Atfridge
&Rourþ1983c).Thesameworke¡salsoshowedúattheprotectiveactivitiesofvarious
antibody preparations correrated with their capacities to inhibit vibrio attachment in vitro,
suggeeting that at least some of the non-LPS protective antþens may Play a role in adherence'
This finding provides a¡r encouraging analogr with studies of enterotoxþe¡ic E- coli nwhich
isolated colonization factors harre been successfulþ used as prophylactic immunogens (Rutter
& Jones ll73; Morsan et al' tY78)'
The studies outlined above clearþ demonstrated the exisænce of non-LPS protective
antþens nV cholqæ, at least in the Classical Inaba strain, 5698' Further investigation of the
vaccine potentirat of such components seemed warranted. f[s immunoprophylactic sig-
nificance of the non-Lps antþens or.v chotqæobviousþ depends on the frequency of their
distribution and the extent to which they are shared or strain specific In a previous study,
Attridge & Ron ley (19g3c) were unable to detect such determinarts in the o17 and r50 strains,
both of which are of El ror biotype and ogærra sefotype. Itwas clearþ essential to begin by
examining other st¡ains for tle preËence of non-Lps protective antigens, as the existence of
shared compotrents of this type migþt offer a ne$' apProach to vaccine developmenL
3.2 Characterization ofv cholerae strains
For this study, a total oLlT strains was used (Table 3.1). v dtolqøe strains isolated in
l9g5 a¡e referred to as recent field isoliates, in contrast to the old laboratory strains which were
fi¡st isolate d m-nyears ago. The significance of t.his distinction will become apparent later'
65

Ibble 3.1 Cha¡acterbtics oî'V dtobæ etrains
Strain
5698
c4401
ztT56t
cA411
A414041
NIH4l
8æ3
v86
Aú{1399ß
BM69
358
ol7T50
A¡\14üt3
H-1
il56eB/165
Biot¡'Pe Serotype
Ctaßsical Inaba 5'5 t 0'66x ld
Classical Inaba ?-6 x'0'26 x 103
Clasßhal Inaba 9'3 t 0'a6x 1ú
Classical Ogær'a 1.1 t 0'01x ldClassical Ogawa 9'4 ¡0'08x 103
Classicat Ogawa 1'2 t 0'01x 103
ElTor Inaba 5'7 t 0'68r1d
El Tor Inabe 8'7 t 3'a0x 10Ê
ElTor Ineba 24 x'2'40xl}a
El Tor Inaba 1'0 t 0'23x 104
El Tor Inaba 5'5 r 0'65x 104
ElTbr Ogova 9'4 ¡0'18x10s
El Tor Ogma &0 t 280x ldElTor Ogrwa 4'1 t 12¿4xlú
El Tor Ogma 1'8 x'?'40x tdEl lbr Ogæva 1.8 t 0'31x ldClæsical Inaba x non01 V¡brb > 109
Vrnrlencel So*ce (year of isotation)2
a(1ea6)
b(1es3)
(leBs)
b(1es3)
(1e8s)
b(1e41)
b(pn -19Ð3
a(1e61)
c(1985)
d(1e8s)
d(re85)
a(p,re-1965)
a(pre-1962)
c(1e8Ð
d(le8s)
d(1e8Ð
a$e2)
1 Eoprored as LDSOvalues, calculated as descrig il Section 2'l2.l. Data a¡e mean tSE
values for two to four determinations
2
^ = Dr. K Bhaekaran (Central Dnrglnstiürte, Lucknou', hdi.)t b = fh. J. Berry (Univer-
sity of Teúa¡, Austin); c = Dr. B. I(ay (International center for Diarrheal Diseæe Re-
seafch, Dhak¡, Bangþdesh); d = Dr. S.C Pal (National Institute of chole¡a and Enæric
Diseases, Calcutta, hdia).
h.. Gu*tzel&BerY (195).
6

The 5698/165 hybrid lnbriowaf¡ preparedby coniugation between v cholerøe strain 5698 and
the non-cholera vibrio strain 165 (Bhaskaran 1971), and carries the LpS determinants of the
165 strain on the non-Lps antigenic bacþround of the 569B strain (Bhaskaran l97l; Attridge
& Rowþ 1983c).
Biotype was confirmed by assessing sensitivity to the antibiotic poþmixin B (50
units/ml) and þ typing with biotype-specific Phages. serotype was confirmed by slide ag-
gtutination using specific typing sera virulence war¡ determined by the infant mouse cholera
model (see Section zlzl). The data in Table 3.1 show that 16 of the 17 strains were pathogenic
in this model, althougb strains o17 and g233 had to be passaged in infantmice to increase tleir
virulence to the levels shourn. The 5698/165 hybridwas non-Pathogenic-
33 The existence of non-Lps pnot€c{¡ve antigens in strains
C4401 and C4411
33.1 hoduction of antisera to 569ry165' CA40l and ca4ll strains
To study the distribution of non-LPS protective antþns! it was decided to prepare
antisera to three strains of V cholqøez 56981165, C4401 and C4411' The first antiserum was
p,repared by immunizing mice with live vibrios of the hybrid strain 569F,1165 as described in
section zg.l.In a pro,,ious study, Arridge & Rowþ (19S3c) ha'e confirmed the antigenic
relationship between strain 5698/165 and its vcholerae parert, in particular the absence of
5698 LpS antigens from the hybrid strain. An anti-56gB/1ó5 serum was found to protect infant
mice from challenge with 5698, demonstrating the existence of non-LPS protective antigens
in this strain (Attridge & Rowþ 19S3c). Such a sefum' therefore, prwided a convenient
means of surveying other strains for the pfesence of such determinants'
67

Antjserawere also prepared against the C4401 (Classical Inaba) and C4411 (Classical
Ogawa) strainq but these sera had to be absorbed with homologous LFS (Section 210'1) so
that polentiatly protective anti-I-PS antibodis u'ere removed' fIA assays performed on the
LpS_absorbed antisera showed a titer of < 1:| indicatingthe efficacy of this procedure.
3.32 hotective activities of antlsera
The infant mouse model was used for assessment of the protective capacities of the
antisera. Initial assays shoured that tle anti-569H165 serum was indeed able to Protect against
challenge with the parentar 5698 strain. Furthermore, the Lps-absorbed anti-c4401 and
anti-CA41l serawere also protective againsthomologous challenge, suggestingthe existence
of non-LPS pfotective antþens in these two strains. To eliminaæ the possibility that in the
laüer instances protection was due to sub-aggtutinating concentrations of anti-LPS antibodies
in the absorbed sera, aliquots of each serum were further abeorbed with live or boiled
homologous vibrios. Aboorption with live orÊanisms reduced the protective tiærs of the
L'S_absorbed sera þ 97vo or ggvo,whereas absorption with boiledvibrios reduced the tiær
by only l4vo ot3.5Vo (Table 3.2). Itwa.s concluded that str¿ins c4401 and c4411 do pæsess
non-LPS Protective antigens'
To a^sse8s thevaccine significance of tleee comPonents, itwas importantto lookat their
distribution âmong other V cholqae strains, and to determine the eÍent to which üey were
sharedorstfain_specific. Atthis stage itseemedpossible thatposseesion of such antigens might
be restricæd to strains of the Classical biotype'
ó8

rhbre 3J hotective activities of Lps-absorbed antisera to cA40l and c4411: effect of
further absorptions with live homologous vibriosa
Protective Activity
Serum
I-PS-abso¡bed anti-C440 1
LPS-abso¡bed anti-C441 1
Before firther
abeorption
1980
2000
Afrer aboorPtionwith
Boiþdvibrbs Livevibrios
l7l0 (t4%o)b 55 (ÍtTo)
a0 QEVo)t*t0 Q.SVo\
" Anti¡erum to cA40l which had a protective tiær of 1:7480 (eee ßble 3'3) was diluted 3'7
times to grve an equivalent titer of 1i2000. Each antisenrmwas tested for its capacity to
mediaæ protection 4ginst challengewith the homlogor¡¡ stfain'
htgtto in parentheses referto Pefcentage reduction of protective tiærs
69

3.4 Distribution of non-LPS pmtectirrc antigens among
V cholerae strains
3.4.1 Old laboratorY shains
In the first series of experiments, the antiserum to strain 5698/165 and the LPS-ab-
sorbed antisera to strains CA401 and CA4ll were tested for their capacities to protect infant
mice from challenge with eight old laboratory strains of V dtolerøe. T}le same spectrum of
activig was seen witl each antiserum; protection was mediated against isolates of the Classical
biotype but not against those of the El Tor biotype (Table 3.3). The protection observedwas
not serotype specific These data were encouraging for they raised tle possibility that all
st¡ains of classical biotype might possess common non-Lps protective antigens. It seemed
importan! howerrer, to check this ñnding using v cholqøe strains recently isolated from the
ñeld.
!4,2 Rsoent ñeld isolates
A second series of protection elperiments was performed using eigþt isolates obtained
from Indira and BangÞdesh. In cont¡ast to the previous finding both of the sera tested
protected not onþ against the two Classical strains but also against the six of El Tor biotype
(Table 3.3). AgÊin, hourever, protection extended to isol,ates of heterologpus serotype, al-
thougþ the tiærs o6øined lvere consistently higher in cases of homologous challenç' These
results suggested a differencewith respect to the expression of non-LPS protective antigens by
old a¡d recent El Tor isolates. To confirm this apparent dichotomy, antisera were prepared
¡gainst two of the latter strains, abso¡bed with homologous LPS as before, and then surveyed
for protective capacities against a fange of challenge strains. The absorbed sera retained
significant protective activities aginst recently isolated strains of the four common biotype-
70

rhble 33 Distribution of shared non-LPS protective antigens a¡nong sixteen str¿i¡s of
Vclpbæ
Antiserum to
Strain 5698/165 cA401a c4411 4413993 H-1
Old l¡olates
5698
cA401
cA411
NIH4l823.3
\¡86
ot7T50
687 t 50
433 x,3l333 t 38
393 x,2<10<10<10<10
1780
2500
1310
tz50<10<10<10<10
NDb
ND
NDND<10ND<10
ND
ND
ND
NDND
<10I{D
<10
ND
t3?0 x,147
l2A0 x,121
2000 t 135
1690 t 190
<10<10<10<10
Rcccnt lsolatcg
zl756t4414041
AAl}w}BM69
358
4A.l4t73
H-1
u
ND
NDND
ND
ND
I{DND
I{D
34t0
2570
5590
4450
3990
1150
t290
r830
590 t 95
1630 t 130
573 x,4O
543t&05Ð x, t2
1ã)0 r 8ó
1390 t 8ó
LZTO x, tl?
25lO x,192
2190 t 3ó8
3770 x,ÐßNDNDND
1830 I 336
ND
1160 x,7
Ðß0 x,ZTI
13f) t l(DND
ND
ND
3560 t 88
ND
NOIE Data sho$, prrotective activities (mean tSE for multiple estinates) of a¡tisera, ex-
pr,essed as S0To protective endpoints as described in Section zlLZ Antbera to strains
56gBll65and cA4ll were titraæd three times against each straiD, antisera to strains
AA139g3 and H-l wene titrated twice, and the antiserum to strain CA40l was titrated onos
Teparaæ baæhes of a¡tben¡m to strain CA4O1 wer€ tested against the tu¡o sets of isolates.
The senrm assayed agpinßt the rccent isolatÊs had a prctective titer of t:7480 against
homol,ogous vibrios.
b l.ID = Not determined"
7l

serotype combinations (Table 3.3). As with the three sera tested earlier, no protection u'as
evident when the challenge strain were old isolates (8?313 and O1?) of El Tor biotype @ble
3.3).
3.43 Conclusbn
Collectiveþ, these data indicaæ that all of the Classical strains examined, as well as
recent isolateß of El Tor biotype, share comlþnents which function as protective antigens in
the infant mouse cholera model Analogous components were notdetected on any of four old
laboratory El Tor strains. The possibility that the liatter might possess the common antigens
but that the corresponding antibodies are not sufficient to confer protection against such strains
was eliminated by the finding that absorption of an anti- 56981165 serum with live o17 vibrioo
did not lead to a reduction in protective activity (data not shown, but see Section 5.2).
35 Subsets of non-LPS prctective antigens ane
semt¡rpe-restricted
3.5.1 Demnstration by proÚection assa¡t
The data in Table 3.3 shou, thag on average, tle protective titer of an antiserum was
two-fold higber when the challenge strain wa.s of homologous rather tlan heterologous
serotype. This finding suggested the existence of serotypicalþ restricted non-LFS protective
antigens in addition ûo the shared components discussed above. To examine this Point more
&otly, two serawere extensiveþ abeorbedwith sü'ains of heterologous serotype in an aüempt
to prepare sera which would protect mice against challenge with strains of homologous
serotype onlY.
The sera selected for these absorptions Ìl,ere rabbit antisera prepared against C4411
or 569B1165; the former w¿ts extensively absorbed with homologous LPS as described in
72

Section LlO.l. Both seratrrere protective against challenç with either serotype, althougþ as
noted prerriousþ, each was more active against homologous challengp (Tâble 3.4). Further
absorption of the LpS-absorbedanti-CA4ll serumwithvibrios of heterologous Inabaserotype
(strain 5698) effectiveþ removed protective activity against this serotype, but the residual
serum retained significant protective activity ¡grinst each of five Ogara challenge strains
(table 3.4).
Simila¡ absorptions of the anti-5698/165 serum with vibrios of heterologous Opwa
serotype (strain C4411) failed to remove all protective activity against challenge with the
absorbing strain, and this sih¡ation still pertained e\ren afær further absorptions with the seme
and subsequently a different ognra strain (4414041). However, the residual protective
potential against strains of ogawa serotlipe was considerabþ less than that observed against
homologous Inaba challengestrains (Ihble 3.4). Furthermore, these protectionassays revealed
a biotype-associated difference between the various ogawa challenge strains use4 a point
whichwa^s clarifiedby subequent studies (see section 4.6).
352 lÞmnstrationbY IEXVÍ
The¡e elperiments suggested the exi¡ænce of serotyPe-rest¡icted non-LPS protective
antþens. The extensively-absorbed anti-CA4ll serum was tested for its content of antibodiee
capable of binding to strains of homologous (C4411) or heterologous (5698) serotype þ IEM'
In line with the protection dat4 significant numbers of gold particles were observed in clooe
association with cA4ll vibrioo, whereas little bindingwas er¡identwith 5698 bacteria (Figure
3.1).
73

Table 3.4 The exbtence of serotyp+restrlcted non-LPS potective antþns lnv. choleætea
Challenç Strains
Ogawa serotype lnaba serotYPe
Antberum C4411 NlH41 AAl ¿tO41 H-l 64 5698 CA¿+01 217æ1 AA1399Íl BM69
LP9'ab6orb€d
anti-CA4l1
S6gBabsorbedb
4210
3320
3320
2500
ND
2950
ND
2810
1N<n
1080
<nND
<nND
<nND
<nND
2890
ND ND ND 6110 5000 ND ND NDUnabsorbed
antþ5698/16f, ',ß70 21ocA411/4414041
absorbed 26oe 4n zgoe <n <æ 3Í!rt0 25SO 2550 æ40 2780
a Antserawere frrded only once for protecüve actlvttþs.b Lps-absorbed ant-GA4i1 wasturttrerabsorbedñßümes (thrioe d.gr Q,Zhr,andhillce d4 c, ovemþÌrt) wlüì lh/e 5698'
" AnÞ569Ey16s tested unabsorbecl as lt contains only anüboclþs to non'LPS protective anügens (see sectk)n 3'2)'
d Absortring antt569B/16s soven times with live cA411 did not remove all protectlve ac-tivity agalnst sÙalrs of ogawa serotype; Ë was fur-
ther absorbed three times wlth llw AA14041
" pD¡o of 260 and 2g0 was obserræd when the ctnilenç strains cA411 and AA14o41 were growrì under rouüne ct¡lturalcondilions (NB d
97 c). Howew[ wtren they were grown on cFA ù.zsc Üìe PÈo values uære 970 and 1380, respactively (see sæüo-n 4'6 for detalb)'
74

Figure 3.1 Demonstration of (Ogawa) serotype+pecific non-LPS determinants . An exten-
siveþ absorbed anti-C4411 serum (see text) was used for IEM examinatiot of V dtoløøe
sf'ains C4411 (A) and 5698 (B). Following growth on NA plates (37 C for Vlh),bacteriawere
hanested, incubated with 1:10 dilution of absorbed anti-C4411 serum, follor¡'ed by 1:50
dilution of protein A+olloidal gold particles at room temperahrre. Bars rep,resent 200nm.
75

4.l
t -(t
.,
.t'
ta
a
)
lr "-
*-^
-
tl -,
.ì

3.6 Correlation between protectirrc activities of antisera and
their capacities Ûo inhibit in vitro attæhment
3.ú1 Invitro enterocXÚe adherence a$¡ay
Adhesion of Vcholqa¿ to the mucosa of the small intestine is now recognized as an
important earþ event in colonization. In an attempt to confirm the previousþ reported link
between ttre capacities of a serum to protect infant mice frr¡m choleraic infection and to inhibit
the ín vito atlachment of bacteria of the challenge strai¡ (Attridge & Rowley 1983c)' an
adherence assay war¡ developed using enterocytes isolaæd from the adult mouse snall intestine
(as described in section zll). phase+ontrast microscopy showed that the enterocyte isolation
procedure yielded populations of epithelial cells, which retain their characteristic columnar
morpholog and have clearþ deñned apical bnrsh borders (not shown)' On the average 'ffiVo
viabilitywas indic¿tedþ Trypan Blue exclusion. preliminary experiments demonst¡ated that
a ratio of 10 bacteria per enterrccyte was optimal for assessment of bacterial adherence'
3;6:2 Inhibition of adherence
Three of the antisera described above were tested for their capacities to inhibit the in
vito attachment of. V cholqae to isolaæd murine enterocytes. The antiserum to strain
56g8/165was initi;ally tested against the 56g8 parent st¡ain, and the LPS-absorbed antisera to
st¡ains cA401 and cA41l were tested against homologous vibrios. Each serum produced a
concentrationdependent inhibition of attachment (Figur e3.2), confirming a previous finding
þ Attridge & Rowþ (l9g3c) that antibodies to non-LPS determinants can inhibit theinvitro
binding of sub-aggtutinating concentrations of vibrioe. Each sefum was tìen tested fo¡ its
capacity to inhibit the adherence of the seven other old isolates used in tlis study (final serum
dilution, 1:10). All three antisera consistently inhibited the attachment of each of the four
i
76

;
,'{
¡ìl^'
IÞ
I{flrÌ
!
.'rl
tI
iI
Figure 32 Antibody-mediated inhibition ol V choløæ adherence h vito'' Antisera u'efe
tested for their capacities to inhibit bacterial attachment to isolated murine enterocytes, as
described in Material and Methods (section Lll.3). Residual percentage adherence after
pretreatme¡twitì antiserum is shourn relative to adherence observed after preincubationwith
a 1:10 dilution of normal mouse serum (rt}vo). Graphs show bindingof strain c4411 in the
prcsenc€ of LPS-abeorbed antiserum to strain c4411 ( o ), binding of strain c4401 in the
presence of LPS-absorbed antiserum to strain C4401 (r), and binding of st¡ain 5698 in the
presence of antiserum to strain 56gvl65 (v). Data are mean tSE (bans) attachmentvalues
f¡om three determinations.
I
77

100
60
IJJozl¡JÉ,l¡JrF¡to
UJol-zl¡lC)Eu¡o-
120
80
40
20
30
RECIPROCAL SERUM DILUTION
10010

strains of Classical biotype, whereas the binding of the El Tor strains was not reduced (Figure
3.3; results $,ith vs6 and T50 strains are not shown but were similiar to ttrose obtained with
st¡ains O17 and 8233).
Since therewas a consistent correlation between tle presence of the non-LPS protec-
tive antigens and susceptibility to antibody-mediated inhibition of adherence, itwas predicted
that tle binding of the recent field isolates would also be reduced in tle Presence of such
antise¡a. Figure 3.4 sho\rs that this is indeed the case. These resulß strongþ reinforce the
prwious demonst¡ation (Attridge & Rowley 1983c) of a correlation between tle capacities of
antisera to mediate protection and to inhibitôn viÞo attachment'
3J Discussion
The maior finding described in this chapter is ttrat each of eight recent field isolaæs of
V choleræbearcommon non-LPSprotectiveantigens, despitedifferences inbiotype' selotyPe'
and geographic origin Of the eigþt older isolates o|.V dtobæ, only those of Classical biotype
erpre's these antigens (table 3.3); the basis for this restricted distribution remains uncertain'
It seems unlikeþ that the old El Tor strains hat'e lost such components during prolonged storage
in the laboratory since similiar t¡eatment has not affected the old Classical v cholqae strains'
Perhaps the eryression of non-I-PS protective antigens þ recent ñeld El Tor isolates results
from inæractionbetween tle two biotypes in the environment Epidemiologicalstudies have
re,vealed that the biotype mGt commonþ associated with clinical cholera cases can change
over a period of time in a given location (Samadi ef aI 1983). Although the factorc responsible
for such biotype suritching remain undefind tìere would seem to be ample opporhrnity for
interaction between vibriæ of the two common biotypes in the field" It is ttrerefore conceivable
that vibrios of El Tor biotype harle, comparativeþ recentþ, acquired the genetic potential to
express non-LPS protective antþens. Since atleast some of these comPonents may play a role
78

Flgure 3.3 Capacities of tlree antisera to inhibit the ¡n vito attzchment of V dtolqae strains
of ClassicalorElTorbiotype. Residualadherencecapacities or.suV dtol.rz,aesüains (d5698;
B, C4401; Ç C4411; D, NIH41; F.r8?33; and E O17) are shovn afær preincubation witl
each of three antisera: (top) LPS-absorbed antiserum to strain C4411, (middle) LPS-absorbed
antiserum to strain CA40l, and (bottom) antiserum to st¡ain 5698/165. Percentage adherence
is relative to that dispþed by control orsanisms preincubaæd with a 1:10 dilution of normal
monse serum (l107o). Histograms show mean residual adherence and bars represent the SE
(fr om th¡ee determinatio ns).
79

J lo o z J I o Þ
(o oo) o
C^)
J N o Þ z { I o Þ o
J l\) oo Þ J I (tl
o) (o qt o) (,l
(oo o
o) o
PE
RC
EN
TA
GE
AD
HE
RE
NC
E
o) oo)
(oo
o
5698
cA40
1
cA41
1
NIH
41
8233 o1
7

Flgurc 3.4 Capacigof antis€rumtostrainCA4ll toinhibittheionvit¡p attachmentof recent
V e;løbæ ñeld isohtes of El Tor biot¡pe. Residusl adherence capacities of six V dtalaae
srraiDs (A 5698; B, O17; Ç 4413993; D, BM69; E H-1; and R 64) are shown after
preinorbationwith antieenrm to strain C4411 (absorbedwith homologous LPS). Percentage
adherence is relatir¡e to that shown þ contnol organisms preinorbated stith a 1:10 dilution of
normal mou¡e senrm (ltMo). Histogrems shos'meiatr residual adhe¡ence, andba¡s represent
tùe SE (from two determinationr).
80

PE
RC
EN
TA
GE
AD
HE
RE
NC
EJ ]u o
(o oo) o
(¡) o
o
Þ TD o (] m 'tl

in adherence (see belou,), it is possible that they contribuæ to pathogenic potential In this
context, it is interesting to note that the present pandemic is the first to have been caused by
vibrios of El TorbiotYPe.
In addition to the non-LpS protective antigens that are shared by strains of either
serotype, the data suggest the existence of additional serotype-restricted comPonenß' Thus
the protective titers of tle five antisera used in t.his study were ahrays greater aeainst strains
of homologous serotype (Table 3.3). The data presented in Table 3.4 provide more direct
er¡idence for the existence of serotype-rest¡icted compotrents. Abeorption of LPS-abao¡bed
anti-ogaura serum with vibrios of Inaba serot¡rpe effectively removed protective activity against
this serotype, but the residual serum retained significaat protective activity against ogawa
challenge strains" Similar datawere obtained in a second erperiment in which anti-5698/165
serum was absorbed with strains of ogawa serotype. C-ollectively these e:rperiments confirm
the existence of serotype-rest¡icted non-LPS protective antigens'
As noted previousþ by Attridge & Rou,ley (1983c), a consistent correlation was found
between the capacity of an antiserum to protect infant mice and its capacity to inhibit the i'l
vito attachmentof the challenge st¡ain The useof sub-agglutinatingconcent¡ations of vibrioe
in the attachment studies, and the fact that tnany sü'ain s are virulent when administered in lo¡'
doses (ie. sub-agglutinatingconcentrations),would argue ¡gainstantibody-mediatedbacterial
agglutination as a critical mechanism of protection in this model The blocking of vibrio
adhesins is a more likeþ mechanism (Freter & Jones 1976; Attridge & Rowley 1983c). The
correlation obsewed between these two activities of the non-LPS antibodies þrotective
capacity and inhibition of adherence) is consisænt with a role for at least some of these
componenß in colonization.
The failure to remove tie residual protective antibodies from l-PS-absorbed anti-
CA401 and anti-CA4ll sera by abeorption with boiled vibrios confirms the heat-labile nature
81

of these antigens (Watanabe et at.1969; Attridge lngr.Itseems unlikeþthatrelativeþ labile
non-LpS protective antigenswouldescape denahrration duringthe preparation of convention-
al killed whole cell vaccines. Studies W Cryzand colleagues (1932) have shown that phenol-
or heat-inactivation drasticaþ reduces the immunogenicity of cholera vibrios, althoug[ for-
malin treatment appeaß less deleterious. The recent demonst¡ation of the clinical sþificance
of at least one non-LpS component (see Chapter 5) will necessitate a more careful approach
to the problem of vaccine inactivation. Alternativeþ, should such components acquire the
status of protective antigens, itcouldbe argued thattheywouldbe more effectivelypresented
to the immune system þ an attenuated live oralvaccine (see Section 1.10.21).
82

Role of TCP as a Colonization Fbctor and aNon-LPS Protective Antigen in the InfantMouse CholeraModel
4.I Introduc{ion
,rh" results of Chapter 3 have clearly indicated thatcommon non-LPS protective antigens are
opressed by four old strains of Classical biotype and eigþt recent field isolates of both biotypes,
but that such determinants are not detectable on four old El Tor st¡ains. Althougþ these
antigens remain undefined, their relative lability (Attridge 1979) suggests that they migþt be
protein in nature (ùyz et aI lg82; Attridge & Rowþ 19S3c). Candidate proteinswould be
thoee associated with ftagelliar or pilus structures, or located elsewhere in tle outer membrane
(Section 1.8.2).
Several studies implicate the flagellum as a determinant of virulence ín V choløæ,
thougþ tlere has been debate as towhether this structure functions soleþ þ providing motility
(Guentzel & Berry lg75\, or whether it also plays a role in attachment (Attridge & Rowley
l9g3a). In the present contex! Eubanks et at. (1977) have described a non-LPS protective
antigen associaæd with the vibrio flagellum which was detecæd in both Inaba and Ogawa
st¡ains of the Classical biotype. Yancey et ø1. (1979) also described a simila¡ heat-labile
non-LPS flagellar-associated antigen.
83

V choleræinteracts intimateþwith themucosalsurface of the intestine, theinitiraler¡ent
in the patlogenesis of cholera. For some other bacterial enteropathogens, this colonization
has been shq¡rn to depend on the presence of pili which mediate adherence to receptors on
host cells (Jones 1980; Gaastra & de Graaf 1982). For example, K88 pili have been shown to
be critical for the ln vito attachment and rn vivo cnlo¡hation of ETBC strains of porcine orign
(Smith & Linggeod l97l;Jones & Rutter 1972). Subeequent studies showed tlat the (heat-
labile) K88 protein could be used as a protective immunogen, eliciting the production of
antibodies which confer protection bD'inhibiting colonization (Rutter & Jones 1973; Rutter
et aL 1976\.
As has already been pointed out (Section 1.3.3.4), earþ attempts to detect pili on
V chobæmetwith variable success (Finkelsæin & Mukeriee 1fb3; Barua & Chatteriee 19ó4;
Tneedy et aL 19ß: Adhikari & Chatterjee 1Í}69). Untilvety recent$, there \ilas no er¡idence
to suggest that such structures played any rnle in vibrio adherence (rerrian'ed by Jones 1980).
During the course of these studies, howerrer, reports from Ehara et al. (1986) andTzylor et øI
(19S7)revivedinterestinthepossibleinvoþementofpiliinthepathogenesisofcholera. There
is nou, no doubt that TCP are required for efficient colonization of both the humen (Herrington
et al. 1988) and infant mouse gut (Taylor et a|.198Ð.
The orperimenß to be described in this chapter u/ere aimed at elucidating the nature
of the non-LPS protective antigens of V c]øbae. C.omparisons of OMP preparations (by
SDS-PAGE and immunoblotting) failed to rer¡eal any consistent difference which could be
correliatedwith the presenceof non-LPS protectiveantigens, andsubsequenterperimentswith
detergent-t¡eated OMPE also failed to implicate any particular protein band" I-ater experi-
menß were desþed to er¡aluate the importance of TCP as a protective antiçn in the infant
mouse model
84

4.2 Studies ofV cholerae OMPs
+2.L Anat}'sis by SDS-PAGE and imrnrnoHoüing
heliminary eryeriments were carried outwith theV dnlerae 569B strain. Both OMP
and flagellar preparations of this strain u'ere compared" EM examination of the latter con-
firmed the presence of ftagellar structureg althougþ contamination with membrane vesicles
\\'as apparent (data not shown). SDS-PAGE anal¡nis (as described in Section 215. 1) of the two
preparations resulted in identical banding patterns (data not shou¡n). As there was no obvious
difference between theq OMP rather than flagellar preparations of otler V cholqøe strains
were use4 mainþ because of ease of preparation (section 28).
In o¡der to determine whether any consistent difference could be detected at the protein
level betw æn V choleræ strains possessing or lackin g non -LPS protective antigens, the protein
profrles of OMP preparations of 8 V choba¿ strains were examined by SDS-PAGE A
representative operiment is shown in Fþre 4.1, (left). None of the protein bands could be
correlated with the previousþ determined distribution of non-LPS protective antigens. To
determi¡e whetler such a correl¡ation migbt be revealed þ immunoblotting (Section 2L5.2),
OMp preparations of the 8 strains were run on SDS-PAGE transferred to nitrocellulose and
incubated with the protective serum (antiCA4ll; Section 3.3.2). As shown in Fþre 4.1,
(riÈÐ, ho*,ever, tlis approach also failed to implicate any protein as a candidate protective
antþen.
4;22 IÞtergent extr¡ction of OMP preparations
Becauseof thelarge numberofbands seen inSDS-PAGEandimmunoblottinganatyses,
it was decided to treat OMP preparations with a variety of detergents (see Section Z4-2) tþ
selectiveþ solubilize some of the proteins and thereby facilitate further experimentation. A
85

Figurr 4.1. SDS-PAGE and immunoblo tÃf¡golV choleræoMPs. I-ane d o17; lane B ,883;
lane Ç H-1; lane D, BM69; lane E, 5698; lane [, c4411; liane G,4A14041; andlane H 'Z1756l'
oMP, \r,e¡e prepared from various strains, arrdÐ pgnuantities were analJzed by SDS-PAGE'
protein staining (reft) or immunoblotting (with protective Lps-absorbed antiserum to strain
CA4ll; right) folloured"
86
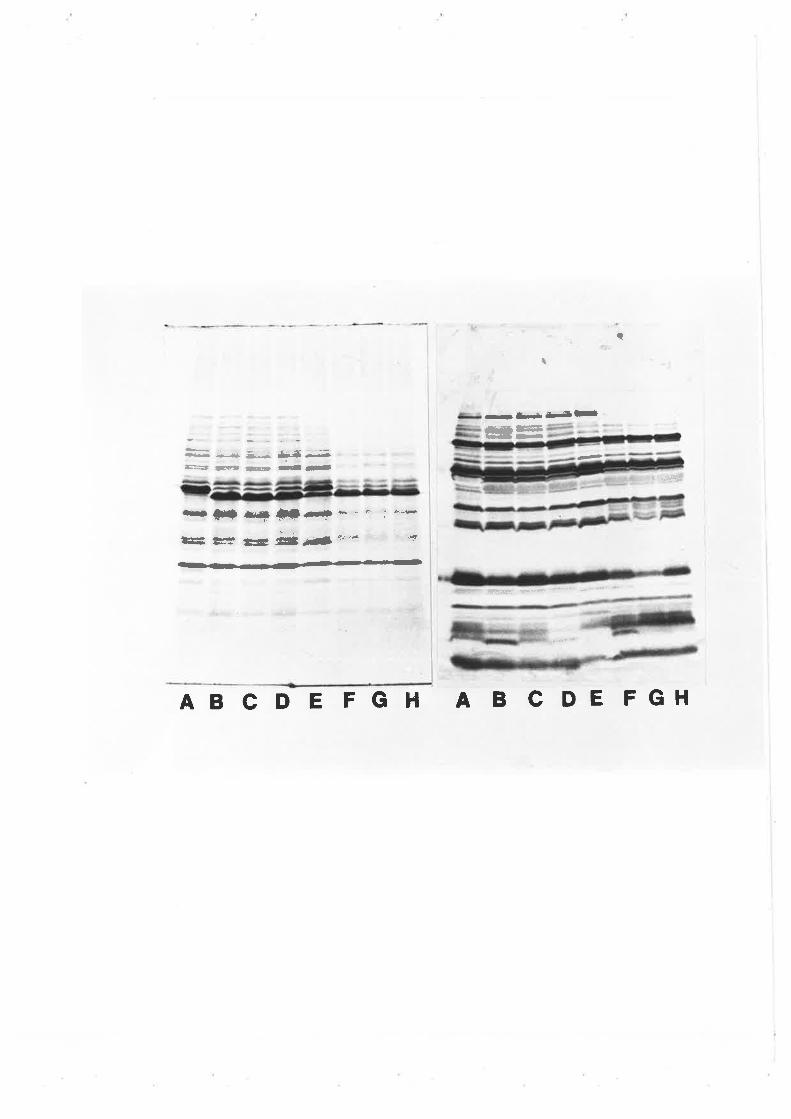
'lia
-¡r.-ÉIlIYÉ5Õts
t
-ék:F:= rtd æ
LZ;;;;Zâ t-,
r'r!å¡Ë
5lE¡tf tÇ--à-1-"*:--ài¿i¡¡-"o 'È
*. ü .æ'*'
ABCDEFGH ABCDEFGH

comparison of tle pfotein profiles by SDS-PAGE of the insoluble oMPs (pellets afær
ultracentrifugation, section zs) with tlose of the untreated oMk showed that sarkosyl
appeared the most promising of the ten detergents examined, in that it selectively extracted
proÞins from 5698 OMP (Frgot" 4.4 lanes c & D). The number of protein bands was reduced
to six (Mr values raßgng from 2145 kDa) (Fþre 4.2, lanes C & D)' By comparing the
capacities of detergent-treated or cont¡ol (treated with buffer only) OMP preparations to
remove prolective antibodies by absorption, itwas possibte to assess the protective potential
of antibodies directed against proteins presentin the insoluble pellet In one such etperiment,
Sarkosyl-insoluble 569B OMp (SarkoMP) rnmoved 797o protective activity from a rabbit
anti-5698/165 serum, whereas the cont¡ol OMP removed lt}To' This suggested that the
proteins solubilized þ this detergentwere not the major non-LPS protective antþens butwere
retained in the insoluble fraction.
This tentative conclusion was strengthened by a second experiment in which groups of
mice were immunized with Sark-oMP or control OMP prepared from the 5698 strain (see
section 29.1 ). The resulting sera were absorbed with 5ó98 LPS (as described in section
210.1) to remove potentiaþ protective antibodies specific for LPS determinants, and then
compared for protective activity aginst 5ó98 challenge. The similarity of the protective titers
obtained (pD50 2000 of the anti-Sark-OMP versus 2500 of the anti-control-OMP sera) indi-
cates that the maior non-LpS protective antigens ¿¡p ¡stained in the Sarkosyl-insoluble oMP
fraction.
Immunoblotting anal¡nis using the LPS-absorbed antiserum prepared against Sark-
oMP failed to reveal any difference in the protein p'rofiles of st¡ains possessing or liacking
non-LPS protective antigens (Fþre 4.3, lianes J, K & L). Itr addition, this serum was further
absorbedwittr boiled 5698 four times (to remove antibodies to heat-stable antþens) andwith
live Ol7 four times (to remove antibodies to non-protective heat-labile antigens) and anaþed
87

Figur.g 42 SDS-PAGE anaþsis of detergent treated V dtolqae 5698 OMP preparations
I-ane A,2Vo Nonidet P40; lane B, 3Vo Nonidet P40; lane Ç lVo Sarkosyl; liane D, 2To Sørkæyl;
lane E 2Vo soditm deorycholate; lane R lM urea; lane G, 2 M urea; lane H, ZVo Tween?ß;
lane I, control OMP; lane J, cont¡ol OMP; lane K ZVo Brq 58; lane l.lVo Brij 58; laneM, l%o
Ti.iton X-114; lane N, O.SVo Triton X-114; lane O, 20 mM EDTfu lane R 10 mM EDTA (Final
concentrations). Arrows on the left point to molecular weigbts of protein bands seen in lane
C after 1% Sarkoeyl treament These are, top to boüom, 45 kDa, 42kDa,38 kDq 32kDar?5
kDa and 2lkDa. Arrows on tle rigþt indicate standard moleculiar weigþt markers (lane M):
Ð5 lt)a, 116 kDa 97 .4 kDa,66 kDa, 45 kDa, 36 kDa, D kDa,Vl lÐa,20. 1 kDa and 14.2 kDa
88

\
-
-- -
a-<-<-
<-
d>
-
--b
.,É.','-|', æ
-.-æ-
I lrul mrr¿t-,r.' ffi
¿?-
åi¡¿ù - ù æ
il-
Ë':ry,&.--eG 1'
.rE-
:il
A B C II E F GH I J KL II il 0P0

Figure 43 Immunoblot anaþsis o[.V cholqac OMPs with anti-Sark-OMP antisera I-ane A
H-1; lane B, BM69; lane G 8ß3; lane D, AA14O41; lane E, 21756l; lane R CA411; lane G &
J,Ol/;lianes H e K 5698; lianes I & I,5698 Sarkosyl-treated OMP pelleÇ lane M, standard
moleculiarweigþt markers: arrows indicate positions - 116 kDa, 97.4kDar 65 kDa, 45 LDa 36
lÐq291ÐqZl kDa, Ð.1 l¡Da and l4.2bDa. Arrows on the leftindicate positions of specific
proteins. These are from top to bottom - 78 kDe, 58.5 kDa, 45 kDa, 38 kDa and 18 kDa l-anes
J-L tested with LPS-absorbed anti-Sark0MP while lanes A-I tested with LPS-absorbed
anti-Sark-OMP further abeorbedwith boiled 5698 (four times) and live O17 (four times).
89

.1
i,t
à,
¡I'
türÌ
't
I1,
),
lI
I
:i, W
r-+
-t-r+>
AB CD EF GH I J KL 1{

I
,t
iÞ-
again roy immunoblotting (Figure 4.3, ranes A-I). In both instances three proteins with Mr
values of 7g, 5g.5 and 1g lDa nor er¡ident by SDS-PAGE anaþis (Figure 4.2) rcacted with the
antisera (Fþre 4.3). After additional eúaustive absorptions with live O17, the antibodies
stillreactedwith o17 OMps (Fþre 4.3,lane G); Howerrer,when attemptswere made toexcise
the relevant protein bands fn¡m sDS-PAGE gels and to use tìe (denatured) proteins to
immunize mice, no protective antibodies utere generated; the antiser4 nevertheless reacted
with 5698 OMP in an ELISA (6400), showinglhe proteins to be immunogenic'
*2.3 OnPV
As mentioned in Section l.8.Zl,a?SkDaprotein (Omp$ is amaior outer membrane
prorein or.v cholqæsógB (stevenson et aL 1985: Pohlner et aL 19864198ób)- This proæin is
found in avariety olV cholcraestrains includingS 6gBtl6íand 1074 (Manning et aL L982)'t\e
ratte¡ a nontoxigenic Brazilian environmental isolate (L,evine et al- 1982). Since a 75 kDa
pfotein is present in 5698 OMP preparations (Figure 4.2), experinents were carried out to
determine whether this migþt represent a non-LpS protective antigen. The structural gene for
the OmpV protein had beên cloned (Stevenson et aL 1985) and an OmpV clone was kindþ
provided by Dr. P. Manning. 'lvhen aliquots of a (protective) anti-5698/165 sefum were
absorbed with the ompV-e:pressing clones or with the vector st¡ain as a @Dtrol no significant
reduction in protective activity were observed. In addition, antiserum raised against the clone
e:rpressing ompv was not protective. These eryerimenr indicaæd that ompv is not a
protective antigen in thi6 model
It'
tt:ri'l
ll
í
I
90

43 @nession of pili by v cholerae sûains
43.1 helininnrY elryerirents
During the cou¡se of these studies, Ehara et aL (1986) described culture conditions
suitable for the production of pili by v choleræ. These grounh conditions, &d others pre-
vionsþ reported to promote úe e:rpression of pili in vitro (Treedy et al' 1968; Faris et aI 1982;
Ehara et aL lgg6), were therefore used for an EM shrdy of pilus production by four strains:
5698 (tre strain most commonly used to cha[enge infant mice), 64 (a recent field isolate of El
Tor biorype), H-l (a recent field isolaæ of El Tor biotype) (see Table 3'1) and K23-7 (a
non-motile El Tor strain usedþ, and obtained from, Eha¡a et al^ 1986). Liquid cultures were
incubated at both 25 C and 37 C in shaking or stationary conditions, with varying incubation
pefids (16-72hrs). when the bacteriawere examinedry EM, a maximum ofl-Zpili per cell
were observed on a small minority (approximalley SVo) of vibrios
In an attemptto selectfor a piliated subpopulation of orpanisms' Eequential subcultures
urere performed; medium containing a sterile glass slide was inoculated wit\ V cholqae atö
alæt24hrs incubation, tle slidewas t¡ansfered to fresh medium and the procedure repeated
twice. However, no sþificant enrichment for piliated bacteria was achieved- Next, El Tor
st¡ains 64 and Y\B-lu,efe inoculaæd onto the TCG agar medium used þ Ehara et al' (1986)
and incubated at 3o c or 37 c overnighL A ferr flexible pili (5-7 nm in width) were seen oû
the surfaces of S-loVo of the cells, but agarn the degree of piliation was i¡adequate, not onþ
for an attempt to isolaæ these structures, but also for studies designed to examine their role in
pathogenesis.
91

+32 ProductionoflC?
In 1987, investigators from the united States reported the expression of TCP þ the
classical v chol¿raestrain o3g5 (lhylor et aL lggT). To evaluate ttre significance of such pili
in pathogenesis, these workers constructed a mutant of 0395 which carried a deletion in the
struchrrar gene for the pilin subunit (TcpA). This mutant showed a 100,000-fold greater LDso
than its parent st¡ain in the infant mouse model, and competition experiments indicated that
TCp promoæd colonization of the infant mouse intestine (Taylor et aL 1987). Althougþ the
pro*ctive activity of antibodies directed against TCP was not examined, these pili would be
obvious candidaæs for non-Lps protective antigens. Accordingfy, the classical strains 5698
and CA41l were cultured according to Taylor's protocol and examinedby EM' However' no
pili with the morphologr of TCP wefe observe4 althougþ other morphological gpes were
present (data not shorrn).
Nex! the coloníutiøt factormedia used extensively for tÏe expression of pili by ETEC
strains (Evans er aL lng) were tested for their capacities to suPport pilus production þ
v cholqæ. strain 5698 was inocurated onto cFA and into cFB and incubated at?S c ot 37
c for ?A to36 hrs. on examination by EM, thick aggregaæd bundles of pili similar to ttrose
described by Taylor et aL (1987) were observed on cells gfown at ?5 C' About 60vo ol the
orsanisms harvested from CFA expressed these pili, and about 4o%o of those grou'n in the
corfespondingliquid medium. Fþre 4.4 shows the morphologr of the pili produced þ two
v cholqæstrains when gro$'n on cEêr at 25 c rhe dimensions of the filaments were measured
by the laüice spacing of crystalline catalase according to the method of wrigley (1968)' Itr
support of the description given þ Taylor et aL (1987), individu¡l f¡laments were estimated to
be about 7-10 nm wide. These filaments aggregated closeþ in parallel to form bundles of
variable length and 0.1-0.3pm in fiemeter'
v2

Figune 4.4 hoduction of TCP on CEA V clølcræ 5698 (A B) and 217561 (C) grown on
CFA at ã C for 36 hours and negatively stained. F = flagellum; S = single TCP filament T
: bundles of TCP. Bar = 250 nm (A), 100 nm (8, C).
93

-
-
!s
T
V

Further studies showed that the TCG medium described by Ehara et aL (1986) could
also support rcP expression, althougþ piliation was inconsisænt (Figure 4'5)' The culture
conditions which most reliabty promoted TCP production invohed growth on NA at 37 C for
z h, followed by growth on cEA (pH 6.5) for vl-36hrs at 25 C At this stage itwas important
to confirm that the pili e4pressed ty these strains when gro!1¡r in this manner were indeed rcP.
To this end a serum containing antibodies to TCp was obtained from Dr. R. Hall (center for
Vaccine Developmen! Balttmore), with tle inæntion of examining the piliated bacteria þ
IErv[ This serum had been prepared by immunization with rcp+xpres sngv cholqae 0395
(Hall e\at.19ß) and upon receiptwas absorbed four times with ogau'a LPS (Section 210'1)
toremove anti-LpS antibodies,leavingbehind antibodies exclusiveþ to non-LPSdeterminants
including those ngainst rcp. IEM using this serum confirmed that the pili were indeed rcP
(Figure 4.6).
433 DistributionofTCP
A number of Vcholeræ st¡ains were examined for their capacities to produce TCP
under the growth conditions described above. In particular, a "o''elationwas
sougþtbetween
the potentialto expressTCp and the previousþ determined distribution of non-LPS protective
antþens. The results of an EM survey are presented in Thble 4.1. pili with ttre morphologl of
TCp were seen on all st¡ains of Classical biot¡'pe examined, regardless of serotype, with the
singfe exception of cA401; Taylor et ot (1987) also failed to deæctsigniñcantTCP production
by this strain. In contras! howe'er, TCp were not seen on any of 6 strains of El ror biotype'
Recent field isolates of the latter biotype, hou,erer, do possess non-LPS protective antigens
(Section 3.4.2).
Immunoblotting studies confirmed tle results of the EM suwey. The serum used in
theseerperimentswas an anti-S6gBll1'serumwhich hadbeen extensivelyabsorbed to remove
94

X'igurre 4.5 P¡oduction of TCP on TCG. V choløae 5698 grown in TCG agan at25 C 1o¡ %
hou¡s and negativeþ stained T = bundle of TCP. Bar = 5(Ð nm (A) 100 nm (B)
95

-
-
:úùt &
a'
a
Ì.
a
It ,
I1
ÒI
o
t
a
^a
ll.r
t'.a
\,
)t-
t
?
ot
A)

Figurc 4.6 Vdtobae 5698 grou¡n on CEA at?S Ç reacted with LPS-absorbed anti-O395
antiserum, follou,ed by hotein A colloidal gold conjugêþ, and negativeþ stained" T = bundle
of TCP pili Bar = TO nm (A) and 1ü) nm (B)
96

alT
?a
rË
d.'
ÒA
¡rÈ+
¡",t
J
ù
1t
l¡
a¡
l¡
IF
r\
J3
+t',
, ^ñ:ì
t ¡¡-,ù
ta atl
a't
titt È
t*a
"î
,¡-
,tt

Ihble 4.1 Disüibutionof TCP
Strain
Clasdcal
5ó98
c4401
cA411
ztTs6l
A414041
5698/165
0395b
Erpressiona
+
+
+
+
+
+
E lbr
BMó9
4413993
H-1
64
K?3,C
ol7
a Production of TCP on CFA at?S Cfor 3ó hrs'
b OggS is a Cla¡sical Ogma kindly prodd bD' Dr. RIt Hall' Center for Vaccine Derrelop
men! Baltimo¡e, US.A"
c K23isanElToreguastrainkindþp¡ovidedbyth.ìrf"Ehara,IngtihræofÏhopicalMedicine
Nag¡saki University' JaPan.
n

antibodies to irreler¡ant bacterial components as a prelude to the attempted cloningof non-LPS
protective antigens (see section 5.2). IEM had confirmed that this serum contained antibodies
to TCp (section 4.6). The absorbed senrm was initialry tested agninst two classical and four
El ror sü.ains (two recent fietd isolates and two old laboratory strains) grown under conditions
which prrcmoted (cFA ar?s C)or fePressed (NB at37 C)TCP production' Figure 4'7 shou/s
the results of subjectingthe bacteriar suspensions to SDS-pAGEand immunoblotting when
grown on cFA at?s cboth crassical st¡ains produced a protein which could be detected by
the antiserum. The molecular weight of this protein (20 lDa), and the the fact that itwas not
produced during gro\¡,th in NB at37 C (Fþre 4.7), is consistent with it being the st¡uctural
subunit of rcp. None of the El Tor strains synthesized this protein neither at37 c nor at 25 c
in any media
In a second experimen! nine V ctpbøest'nin s were cultured on CFA at 25 c and tested
fo¡ TCp eryression using the same antisenrm. A cnrde TCp preparation (section 213) was
used as a positive control while 5698 grown in NB at37 C served as a negative cont¡ol for
immunoblotting (see Figur e 4.7). As expecte4 the antiserum reactedwith a 20 kDa protein
expre*sed only by the classical strains (with the exception of c4401) whereas no reaction was
er¡ident with the El Tor strains The TCP preparatioû waÍ¡ strongþ positive, supporting the
conclusion that this protein represents the pilin subunit (Figure 4'8)'
4.4 TCp is a virulenoe determinant in the infant mouse model
Itwas important to confirm the conclusion that rcP is avirulence determinant in the
infant mouse model (Taylor et aI lgïi). To this end tle virulence of the non-motile variant of
5698 (selectedby the sloppy agar overlay technique as described in Sectiot2.2'¡ was assessed
followinggrowth under conditionswhich supported or repressed TCP production' Thisvariant
98

Figurt 4.7 Immunoblottingmaþis of,V clnleræstrains for TCP eryression. The follwoiag
strains were emamined after culnrne on CFA at 25 C for 36 hours, (lanee A-F) and NB at 37 C
(lanes G-L): H-l (El Tor biotype, lanes A & G); 4413993 (El Tor, B & H); CA411 (Ctassica[
c & D; 5698 (Classicâl D & Ð; O17 (Et Tor, E & K); and 8233 @l Tor, F & L). They u'ere
resuspended in electrophoresis sample buffer and su$ected to SDS-PAGE The pr,oteins were
t¡an¡ferred to nitrocellulose and tle blot¡ developed with 1:10ü) rafrùit anti-5698/165 CP
serum (knoum to contain antiiTCP antibodbs) fotloured by goat anti-¡abbit IgG coupled with
horse radish peroxidase and substrate. The arrorr indicates the pooition of the molecular
weigÞtmarker, ß-lactogfobulin (bovine -ilk), 18.4 kDa
99

a
A B C D E F G H I JK L

lTgure 4A Immunobloüing anal¡ruis of TC? e:pression þ varioru strains oî. V chobæ. A
crudepreparation ofTCP(tanesÀ L) orcellefiracts fromnine V dplcrae strainl (lanesB-K)
we¡e immunoblotted ruing the serum dÊscribed in the bgend to Fþre 4.7. St¡ains (biotype,
rerot¡ipe, lane position) teoted were: H-l (E¡ Tor, Ogau'q B); 4413993 (El Tor, I¡aba, C);
4414041 (C1assical Ogara D);217561 (Classical lnaba, E); O17 (El Tor, Ogaw4 Ð; E233
(El Tor, Inaba, G); C4411 (Ctassical, Og¡wa" H); C4401 (Classica[ Inaba, I); 5698 (Classical
Inabq J & K). Strains were cultured on CFA et?S C for 36 h an4 in the case of 5698, also in
NB at 37 C for purposes of comparison (Lane Ð. The a¡ros' indicates the position of the
moleculiarweig[t marker, &lactoglobulin (bovine mitk, 18.4 kDa).
100

+-
f¡rP
A B CDEF GH I JK L

11t
wa6 chosen in order to enmine the effect ot TCP in the abeence of the
flagellarstructure (Attridgs & Rowley 1983a).
Elpression of TCP by non-motile 5698 led to a 100-fold increase in virulence; or-
ganisms grc$,n on CEA atäSChad an LDso ol l.TI r 0.4 x 106, compared with L?5 x 0.11 x
108 for those grc$'n in NB at37 C.
45 Attempts to PurifY TCP
Given the pivotal role of TCP in colonization and virulence (tylor d aI 1987; Her-
rington et øL l9B8), and þ analogr with the pilus colonization factors of ETEG it seemed
probable thatTCpwould represent a protective antigen of.V choleræ. Accordingf¡ attempts
*,ere made to puriry these structures, in order to assess the protective activity of antibodies
directed ageinst them.
FollowinggrowthonCEAat25 Gbacteriawere resuspendedin differentsolutions (see
Section Zl3) prior to shearingto remove pili. UsingPBS (AlKaissi & Most¡atos 1985) ot ILS
mM 'Fiß HCI (Thylor et at- 1987), the pili couldbe subsequently precipitatedwitl am-onium
sulphate and pelleæd at low speed centrifugation, whereas with 10 mM tis buffer (Korhonen
d aI lgg1), the pili resisted ammonium sulphate precipitation and could onþ be recoveredby
centrifugation at 100,0009 for thr. Regardless of the method use4 such pili preparatiotrs were
always contaminated with other protein bands when examined bD' SDS-PAGE; when viewed
by EM, LpS vesicles and flagella were er¡ident (data not shourn). In an attempt to remove tìese
contaminants, the crude preparations $rene subjected to density gradient centrifugation follow-
ing the method of Ehara et aI (lgS7), but EM examination revealed that this method was of
liülevalue.
Attempts urene 1[s¡ made to puriS the crude pili by adsorption onto guinea pig RBG
(Al-Ikissi & Moetratos 1935) but this too was unsuccessful A final atfempt was made Ûo
RA
101

separate pili from otler cellular material using deorycholate and concentrated urea according
to the method of Korhon eneî al. (1980), but tlis also proved futile. Since efforts to puriff pili
ofV cholqæhad proved ineffective, ttris approachwas abandonedin favor of less directmeans
of assessing the protective activity of anti-TCP antibodies.
4.6 rcP is a prctective antigen in the infant mouse model
It was important to determine whetìer the protective anti-56981165 serum (Section
3.5.1) contained antibodies to TCP an{ if so, whether tlese antibodies contributed to the
protective capacity of the serum. Although this serum was generated by immunization with
organisms gfou/n under conditions unfavorable for TCP elpression (NB at37 C), it seemed
poesible that a feu, TCp might harre been produced under these conditions, or induced to do
so following injection into the rabbit The presence of anti-TCP antibodies was confirmed by
IEM as shou,n in Figure 4.9 (A & B), which shou¡s binding of antibody+oniugated colloidal
gold particles to TCP. Subsequent work confirmed that, when gfo$'n on CFA at25 G the
56gÐtl65 strain produces TCP as judæd by ENr, IEM and immunoblotting anaþses (not
shown).
Accordingþ, the capacity of the anti:TCP antibodies to msdiete protection against
challenge with TCp-poeitive or TCP-negative organisms of the 5698 strain (ie., those grow¡t
on CFA atZS Cor in NB at37 Çrespectivety) was assessed in the inf¡nt mouse model The
protective activity of the LpS-aboorbed anti:ICP serum obtained from Dr. Hall (Section 4.3'2)
was also assessed asainst both challenge inocula The data are presented in Table 4'Z
Anti-5698/165 sho¡,ed a greater tìan frvefold PDSO when the challenge strain was allowed to
express TCp pili in vito (cEA 25 C). A similar but greater effective rise in PDso (æn-folQ
rr/as seen in anti-O395 l-Ps-absorbed Eerum when the same challenge strain was allowed to
102

Figure 4.9 Immunogold labelling of.V dtolqæ 5698. V dtolqøe 5698 was growtr on CEA at
25 C for 3ó hrs and the resultingorganisms reactedwittr anti-5698/165 (ArB) or LPS-absorbed
anti{4411 (C,D) followed þ Protein A-colloidal gold conjugate, and negativeþ stained" T
= bundle of TCP pill Bar = 2N nm (A), 100 nm (B), 100 nm (C) and 100 nm (D).
I
103

I{
¿
I
Ça
¡a
¡Ye -t
!.r
ao
a
ra
f,r 'a a
,,.r'
o
at,
-{l>
¡4
"1'
"tÉ
à
).dt
Jtt
¡t
Cþ' ,t
I
t rt t'
ttl rì
r¡'l
(, r
¿
-'rr
¿d
e.ft
ît
-E-

I
,{
È-
It'
ì{
f;,i
'l
It,
i
Antiserum to
Ibble a¿ Antibodies to TCP are protective in the infant mouse choþra model
Protective activity aglinst 5ó9Ba
rcP.b rcP+b
5698/165
()395 abso¡bedwith
Ogawa LFS
6,000 t 330 34,m t !4¡g
B,W t 1,400 250,000 t 14000
I Protective activity in PD50 uniqthl
b Th" 5698 challenç st¡ain was grou'n under condition¡ uùich favo¡ed (cFA ar ?5 C) or
repreosed (NB at 37 C)the production of TCP CICP+ or TCP, respectiveþ'
104

:
I
I
f^-
express T.;p invitro. This clearry demonstrated that antiircp antibodies have protective
potential. l-at'9r studies (section 5.6.2) provided unequivocal confirmation tlat TCP is a
protective antigen in this model. The appreciation of the protective potential of antibodies to
TCp offered an e:rplanation for previous data which had hitherto remained inerylicable' In
Section 3.5.l itwasshowntha! despiteextensive aboorptionswithorganisms of ogawaserotype
(strain cA411 seven times, AA14041 three times) to remove antibodies to antigens common
to the two serotypes, the anti-5698/r65 serum retained some protective activity againstvibrioa
oftheogawasefotyPe.ThiswasindicatedbythePDsovaluesobtainedwhenmicewere
challenged with vibrios grou¡n in NB at 37 C (PDSæ of ?;60 and 290 against C4411 and
AA14041, respectiveþ . Table 3.4). Hor,er,er' as mentionedin footnote (e) of this table, when
these organisms \ilere grown on cEA at?s cand used as challenge strâins, the PDsovalues
increased about four-fold (cA411, g70; AA14041, 1380). The higþer protective endpoints
obtained when the challenge organism.s had been grown under conditions conducive to TCP
expression suggested that antibodies to TCP had not been completely remove4 and this was
confirmed bfy IEM (not shown). In retrospec! this is not surprising since the absorbing
bacteria had not been culn¡red under conditions favourable for TCP eryression'
In the fevefse eryeriment, absorption of an LPS-absorbed anti-c4411 serum with
bacteria of Inaba serotype (5698) successfully removed all protective activity against challenge
organisms of the Inaba serotype (Section 3.5.1). In this instance, the initial anti-cA4llserum
did not contain antibodies to TCp detect¿ble by IEM (Frgure 4.9, c &.D). Hence, this erylains
why (as shown in Table 3.4) there wa.s no residual activity left against the Inaba serotype (PDso
<2q.
I
ri
105

4.7 Pmtection by anti-TCP antibodies is biotype nestricted
EM and immunoblotting anaþses had suggesæd that the capacity to express TCPwas
restricted to strains of the Classical biotype (Section 4.3.3)' However, some strains of the El
Tor biotype czrrystnrct'rar genes for TCp (Tayror et aL r9ç8a) and so it is possible that strains
of this biotype can produce such pitinvivo. Therefore, itwas imperative to determine the
capacity of anti:TCp antibodies to protect infant mice from challenge with strains of classical
or El Tor biotYPe.
The anti-O395 supplied by Dr. Hall was chosen for the experiments to be described in
this Section because of its greater protective efficacy against TCP+xpressing challenge bac-
teria, compared to the anti-5698/165 serum (Table 4.2). This antiserumwa^s firstabsorbedfour
times with live o3g5 (gown in NB at37 C to inhibit expression of rcp) in order to remove
potentialþ protective antibodies directed against LPS or non:TCP, non-LPS protective an-
tigen".
Protection tests confirmed tìat the protective activity of the absorbed serum was
prinariþ due to antibodies to TCP (Table 4.3). Fint, tle serum was now onþ moderateþ
protective against 0395 or 5698 vibrios cultured under conditions inhibitory to TCP produc-
tion. In contras! if these strains Ìvere grown on cFA at?s Ç the protective tite¡s increased
2ß- or3O0-fold, respectiveþ (tble 4.3). Secon4 the absorbed serumu'as onþweakly protec-
liys agâinst C4401 (which, in this study, does not eryress TCP) challengg, despite the fact that
this strain e:rpre6ses non-LPS protective antigens (section 3'4)'
ConsisæntwiththeimmunoblottingandEManatyses,whenthechallengestrainswere
of the El Tor biotype grolvll on cFA or in NB, the residual serum was unable to mediate
protection (Table 4.3). These results clearþ demonstrate tlat there is a correlation between
i
106

rbble 4i Antibodies to TCp are protective onþ againstcha[enge strains of classi¡al
biotypea
TCP status of challengevibrioo
Challenge suain TCP rcP+
()395
5698
o17 (PA )b
8233(PA )
H-lGA+)
AA1393 (PA*)
c4401
3.3.
230 t 50
430 t 30
<n<n<n<n
70t5
5,000 t 5ü)
130,000 t 11,000
<n<n<n<n95t8
" Figures shos, protective activity of absoôed anti-TCP 0395 cerum in PDsodmt Challenge
straiü were gro.*a to be quantitativeþ TCp' or TCp+ a¡ descriH in legend to Dble 4'2
b pA-ÆA+ refer ûo prerence or aboence of non-LFS protective antigens as detcribed in Table
IUT

the distribution of pili amongst ll choleræstrains and prrrtective activity of and-rcP antibodies
and that it is biotYPe-restricæd'
4.8 Discussion
The experiments described in this chapter !\'ere performed in the hope of elucidating
the nafirre of the non-LpS protective antþens of v chobrøe. In an attempt to identis
differences between the fourstrainslackingnon-LpS protective antigens and thetwelve strains
which share such determinants (Section 3.4), OMP preparations were compared by SDS-
PAGE ¿¡d immunoblotting anaþses. This approach was not helpful hotilever, since the oMP
profires of the four old El ror strains were not consistentþ different (Figure 4.1). Given the
denahrring co¡ditions required for such anaþses, it is poñ'ible that none of tle bands detected
þ immunoblotting involves the binding of protective antibodies' A number of PAGE system's
and various detergent and solvent ext¡actions of the oMP preparations \Ã'efe tested in order
to obtain better resolution of the proteins but these did not pfove fruitfrtl'
Subsequentexperimentswere performed to testwhether any of the non-LPS protective
antþens might be associated \ilith pilus structures produced by v choleræ' Atthougþ experi-
ments described here focussed upon the expression of TCR other pili were seen by EM (data
not shown). oo" of the other t)rpes see& albeit in low numbers, was a flexibre ¡¡¡¡ filament
5-7 nm in width. Ibrahim (1934) found slender flexible pili (3a nm in diameter) present in 1
of 3 El ror strains tested as well as in one classical st¡ain. Al-Kaissi & Most¡atos (1985)
reported the presence of longtwisted pili on some strains of the cla.ssical biotypewhile Ehara
et aL (1986) described the pili they observed as flexible, long ñbres present on El Tor stains'
It is possible the flexible pili seen on El ror strains in this study are the same a.s those described
by orhers (Ibrahim 1984; Ehara eÎ aL l9tl6). Very recently, Hall et aL (1988) reported a
108

rystematic EM study of the pilus types produced by v dtoløa¿. TCP u'efe detected only on
classical strains, while two otler tlpes (B and C) were seen on all strains regardless of biotype'
production of TCp by V choleræ was first reported by Taylor et al- (1987) following
grovth of bacteria in LB medium at 30 c. subsequent experiments by ldall et øt (1988)
confirmed the existence of rcp and recommended growth on cEA at 25 c for optimal piliation.
Initial experiments were designed to identi$ cultural conditions which would consistentþ
support TCP production by v choleræ strains used in this shrdy' Inability to detect such
appendages following growth of bacteria in LB medium suggests some difference in medium
constituents. In agreement with Hall et at. (r9gg) howe'er, rcP were observed when the
bacteria wefe grown on cFA at 25 C The widtl of tle pilus filaments with rcP morphologr
was estimaædtobe 7-10 nm, simil¡ar to previous ssfimates (7nm, Taylot et aL L987;56nm, Hall
eJ ol. L988).
EM and immunobloning studies (Table 4.1, Figures 4.7 &' 4.8) confirmed Hall's finding
tha! underthegrowth conditionsadopted, productionof TCPis restrictedto st¡ains of Classical
biotype. In tota! TCp have now been detected on - classical strains (Taylor et al- 1987; Hall
et aL lgæ; this thesis) but on none of - El Torbiotype. The studies performed to date do not
eliminate the possibility that sü'ains of El ror biotype will qynthesize TCP under as yet
undefined sowth conditions invitto, or perhaps onþ in resPonse to environmenlal stimuli
present in the guL Infant mouse eryeriments designed to assess the capacity of anti:TCP
antibodies to mediate protection againstEl Tor challenge strains failed to provide support for
the latter (see below).
Various methods ha¡ebeen described forthe isol,ation andpurification of differentpilus
st¡uctures produced by v choterøe (Korhon en et a|.1980; Al-Ikissi & Mostratos 1985; Ehara
et at. 1987; Taylor et aL 1987). Horvever, i¡ my hands, trone of these provided adequate
purification of TCR as all resulæd in subst¡ntial protein and LPS contamination. It is knw'n
109

that pili are extremeþ hydrophobic, consisting of a higþ proportion of nonpolar amino acids
(Korhonen ef aL rggo;Gaastra & de Graaf 1gg2) and rcp aÍe no exception (Taylor et aL 1987).
Accordingly pili tend to associate strongþ with other bacterial surface components such as
outer membrane vesicles and LpS. Attempted fractionation of pili on the basis of size and
hydrophobicityusingasodium deoxychorate-ureabuffer (Korhonen et aL 1980) proved futile'
At least some of the protein contaminants seen in the TCp preparations could have been due
to the presence of pilus-associaæd proteins as have been described fot Neisseris gonorrhoeæ
(Muir et at. rggg). As time was limite4 further attempts at purification were abandoned in
favor of alternative approaches to the e'aruation of tre protective signiñcance of rcP.
pilus colonization factors have been recognized asvirulence determinants not onþ for
E. coli(satterwhite et aL lgTg: Gaast¡a & de Graaf 19s2) but also for serreral other gram-nega-
tivebacterial palhogens (Kellogg et aL 1963; Duguid & old 1980;Abraham & Beåchy 1985)'
During the course of these studies TCP were shown to be essential for gut colonization in both
infant mice (Taytor ef øL r9gl,¡and man (Herringto n et aL rgsg). In the former studies, mutant
strains unable to produce TCp were shown to be unable to efficiently colonize the intestines
of infant mice, and were consequently much less virulent tlan the wild-type v dtolqae (Taylor
et at. rggl). Heningto n et oL (1gSS) subsequentry used such murants to assess the importance
of TCp for the colonization potential and vaccine efficacy of non-toxigenic v cholqae' ln
cont¡ast to the vaccine strain which could produce TC! the mutants lf,'ere found to be
non-colontzilg¡g¡.imm¡nogenicandnon-protective.
Initial e:rperiments defined growth conditions which consistently resulæd in TcP
production by V choløae 569B,the strain most commonly used in protection studies' Afær
confirming the identity of the pili by IEM (Figure 4.6), infant mouse tests confirmed that TCP
production signifrcantly enhanced tie virulence of tle (non-motile) 5698 strain' Alttrougþ the
100-fold difference in virulence was less dramatic than that proriously reported by hylor et aL
110

(1937) for the 0395 strain, this probabþ results from reliance uPon a phenotypic rather than
genotypic difference in TCP production. Thus a minority of the bacteria cultured under
conditions unfavorable for TCp expression migþt have been TCP-positive; alærnatively, the
population might har¡e been uniformly TCP-negative at the time of challenge, but synthesized
these pili rapidþ ín vivo. In contrast, the mutants used þ Taylor et al' (1987) were genetically
incapable of TCP Production'
Since virulence determinants such as colonization factors are prime candidates for
protective antigens, itwas of great interest to determine whether TCP might represent one of
the non-Lps protective antigens of. v choreræ. In this respect it was encouraging to find that
the protective anti-S ilgÐlLilsserum (Section 3.5.r) contained antibodies to TCB as judged by
IEM (Figure 4.9) ¿¡¡d immunoblotting analyses (Figure 4.?). The first evidence in support of
the hlpothesis that TCp can function as a protective antigen came from infant mouse assays
designed to evaluate the protective potential of these anti:TCP antibodies- Sera containing
such antibodies were markedly more effective in mediating protection if the challenge or-
ganisms were expressing TCP at the time of administ¡ation (Table 4'2)'
Subsequentprotection tests indicated that antibodies to TCP could mediate protection
against challenge strains of Classical biotype only (Table 4'3), a finding consistent with the
inability to detect the production of rcp by El ror strains. This suggests that TCP is not the
only non-LpS protective antigen in this model, since recent field isolates of El Tor biotype
possess such components (Section 3.4). Attempts to identiS non-LPS protective antigens þ
gene cloning are described in the next chapter'
111

Molecular cloning of genes involved inbiosSmthesis of TCP
5.1 Intnoduclf,on
1¡ryrten attempts to isoLate non-LPS protective antþns from OMP preparations p¡oved
unsuccessful, it was neeessary to dwbe alternative approaches to further chatætet'ae tlese
componen6. To üb en4 the shrdies described in the previous chapÛer were performed 1o
examine the protective sþificance of pilus ¡tnrcürs produced by V cløleræ. Concurrcntly
attempts were beg¡n to isolate non-LPS protective antþns using the techniques of gene
cloning and the¡e studies form the bæis of the pnreseot chaser.
Initialeryerimentsweredesþedtoclonegenesencodingnon-LPsprotectiveantigens
of6 dtotsacsógg and to express them in an E æti tæipfut strain. As discl¡s8ed belw, this
approach proved unsuccessful At about that time, ?ylor 4 4L (1987) reported their s$dies
of TCR fr,omwhich itwæ clearthatsurfaoe expression of TCP andotherOMPswas dependent
upon a functional úoxR gene, Since the E coti rwi¡Åent strain used in rhi¡ 3f¡dy lacked this
getre an alærnative cl,oning strategl was devisedwhich allowed ctoning of genes inrroþed in
TCp production The availability of TCP-producingclones atlo\red unequivocal oonñrmatbn
of the conclusions that rcp acts as both a virule¡ce deterninant and protective antigen in the
infant mouse choþra model
ttz

52 Pnepffiation of done probing serum.
Aclone.probingoerumwasrequircdinordertodetectbyimmunoblottinganyclones
oryressing non-LFS protective antigens, and a protective rabbit anti-5698/165 serum (Section
3.S.1) was selected for this purpoße. Before u¡e the serumwas extensiveþ absoöed to remove
antibodiee to componente other than non-LPS protective antigeng thereþ reducing the
likelihood of detecting "false-positive" clones druing the screening pmocess- Since non-LPS
prrotective antigens a¡e heat-labile and absent from old El Tor st¡ains such as o17 (Section
3.4.1), the senrm was absorùed four times with boiled 569B,t165 (to remove antibodie¡ to
heat+tabþ antþns), three ''.nes with live 01? (to remove antibodies to non-protective
heat-labile antiçns), and ñnalþ two times with live E. coli K12 strain DHl (to remove
antibodies to the.E coli rccipterl strain).
Aliquots wer€ kept after each absorption and tested i¡ an ELISA using 5ó98/165 and
Ol7 OMps as antþns (Figure 5.1). The abeorbed Eerum had ELISA tiæ¡s of 7,300 and 49
agninst 569Fl¿65 and O17 OMh, respectiveþ. This represenls l47o and <0.lWo of the
orisinal activity against both antþens, respectiveþ (Fþre 5.1). In oo¡tras! the protective
activity of the residual serum rvas virtüatly unchanged (protective fi1sr ¡grinet 5698 challenge
fell from lZffi to 11f)), indicating selective retention of the antibodies of inærest The
abeo¡bed semm is heoceforth referred to as anti-5698/165 CP (clone-probing) serun.
53 PreIimfutsry donirry exlDeriments
Initial attemptr to clone non-LPS protective antigens invoþed construction of a 5698
gene bank iîE corietrain DHI ruingthe coemidvector pHCI/9. Details of this procedure are
not given because tùese etudþs were not successful A total of 4o0 colonies were screened þ
oolonyblotting(Soction Zt6),usinga 1:1000 dilution of anti-5698/165 CP seruE'" Sifeen of
113

Flgurr 5.1 preparation of clone probing serun An antfuenrm prepared in rabbits against live
5698/165 hybridvibrioq and lnom to contain protective anúodies specific for TCP (Section
4.6), was extensiveþ abeorbed to remove a¡ftodies of irrelevant specificities (Section 5.2).
Abeorptions 1-l were with boiled 5698/165 vibrioe, 5-? with live o17 and 8,9 with live E coli
K12 Strain DHl. The unabooñed and the aliçot of the absorbed sera at various stages were
tesred aginst56gB/165 oMp (tcfq ) and 01? oMP (dghq ) in an ELxsA assay. o represents
unabsorbed serum, ELISA tiær of u¡abeorbed serum against O17 OMP ßn,1N (not ehoum
in Fþre).
tt4

6
5
4
5
4(f)
Io
ulÉ,t--
F
U)
=1¡¡
t
3
Io
ulÉEt-
Ø
u¡
2
1
2
1
5
NO.
7 Io 1 2 34
ABSORPT¡ON

the clonee were positive and su$ecæd to further anaþie. Aliquots of unabsolbed anti-
silg}lrilsserum were absorbed (æ per Section zl0.z) 4 ':mes with each of the 16 positive
clone¡ orwith poeitive (5698) or negative (8. @tí)cont¡or strains and subeequentþ tested for
removal of antib@ activity þ both EusA and infant mouse protection test'. Althougþ there
waE no correr¡rtion between neduction in ELISA titer and loss of protective activity (data not
shom), 9 of the 16 clones reduced the protective titer by 75Vo oÍ mofe' Hou'ever' when mice
$,efeimmunizedwiththeseninecl.onesnoneoftheresultingantiserawereprotective(data
not shourn).
In an attempt to erplain these dbappointing results, the nine clones were again sub-
iected to immunoblottingwith cp senrm, some tweþe weeks after their original detection-
Despite etorage of the clones in gþcerol at -lOC (Section 22), all were now negative and thus
no l,onger opressing antþens of inærest Thb ap'parent in8tsbility was att¡ibuted to the f¡ct
that the E. colirecipient strain DHl lacLed areo4'bacþround required for clone stability'
However, cubsequent ¡tudiee by Metalanoe and oolleagues made it clear that surface exPfes-
sion of TCp and otùer oM¡¡ is dependentupon theV chobæ úoxR gene (Tylor et aI1987;
Mi'er & Mekalanoo 19ss). since no guch gene is present in, E coli, an alternative cloning
stratesrwas devised"
5.4 Molecular cloning of gerrcs invohad in TCP prcduction
S4.1 Clonhg stuat€' r
The innovation thatmade poosible the cloningof genee involved in rcP EÉthesiswas
the selection of avdtobæ strain, or7, as the recipient of a cosmid gene bank initially
constructed tn E. ætí. The O17 strain poss€ss€s thefaxR gene required for eryression of TCP
(Manning penonar communication), but does not produce these pili under cultural conditions
which prorcte TC? expression by strains of classical biot)"e (Table 4'1; Hall øaL 198ß)'
115

Moreover O17 tacks non-LPS protective antigens in general (Attridge & Rowley 1983c;
Section 3.4.1), facilitating detection of clones opressing such components'
V clplcræ z;llÍillwas selected as the donor st¡ain sinoe iteryresses TCPwellûn vúrro
Cfab1" a.1) and, being a recent field bolate, it also offered the poosibitity of cloning any other
non-LPS protective antþens it night PosEess (section 3.4.2\- Cosnid cloning an.d ínvütto
packagry of donor strain DNA into bacteriophage lambds (simon d aL t983) was used as a
meians of obtaining DNA fragments of sumcient size to clone the tc? oPeron'
5.42 Coretructionof the mobilizable coffiü vector pPM2llÌl
The cloning stratßg¡ adopted required a mobilizable vector, because of the need to
reintroduce cloned V cløbægenes via an E cdiintermediiaæ recipient back ntoV ùdøæ'
Accordindy, ¡[s 6obi lizable (Mob) reþn of the b¡oad host-range Plåsnid RP4 (from plasmid
psltp2gl-l) was introduced into the cosmid vector pHC?9. This constnrction is shou¡n in
Figure 5.4 de1¿ils of the construction afe g¡v.n in Section L17.1'
5.43 Cfoning tbe TC? genes into V clolerae O17
A gene bant wæ coDstructed by Paftiatty diæstiog t\e 217561 DNA (preparcd as
described in section zl7.l) with the restriction endonuclease san3L to obtain fragnents
approximately 40 lb in size, which were then cloned in to t\e BønHI siæ of the vector pPM2101
(see section 217.4). After overnigþt lþtion at 4 C the DNA miÚure was packaged into
bEcterbphage lamda (section Lll.Ð. The packaged pnaæ wefie then used to infect E coli
K12 strains 517-1, from which the cloned DNA was mobilized i¡to Vdtfuæ O17 by plate
mating (Section 2;17.6r. The resulting cosmid bank was stored on nutrient agu Plates
conøiningAP at 4 C
116

X'lgure 5.2 Constn¡ction of pPM2101. The cosmid pHC79 was cleaved with EcoRI and the
protrudingends filled usingKlenow fragments of DNApoþmerase I. Plasmid SUP201-1 was
cleaved with BantHI, the fragment containing the RP4 Mob region was isolated and tle
protruding ends filled as above. The cleaved pHCÍ/9 and the Mob fragment was ligated
overnight and the ligation transformed into ¡Eain 517-1 selecting for Teß. The correct.'
constructs were then selected by their ability to be mobilized into strain SM10 selecting for
fetR pnU ?r}l) KanR (counter-selection) agai¡ s¡ S 17- 1.
tt1

EcoRlPstl
EcoRl
Clol BômHtHñd
Soll
CoHnd
^Q:--è
BomHl
BE
Þ^
Pstl
ClolH¡ndlll
BomHlco9
IY
So
IV
Cleove with EcoRl.
End fill with Klenow frogment'
P3tl
Ligote.
Tronsform into stroin 51 7-1 ond select for Tet
Cleove with BomHl.
End fìll with Klenow frogment
R
RR
Select for mobilizotion into stroin SMlO screening for Tet ond Kon
(Ecoñl) MOB (EcoRl)
BomHl
Soll
let ñ
pHC79
ßl(ÞttQ
g\o
psuPz01- 1
cot (an R)
a'ofn,^
b t
ÉèE
oo
pPM2101
o¿ +
¿o'

144 IÞtectbn of Positive dones
About g{Ð oolonies were patched in duplicate onto CFA plates and incubated at25 C
for3óh topromoteTCp eryression (seeSection 4.3.2r. Oneeetof coloniee was transfered Ûo
nit¡ocellulose filters and þed în sín using the method of Henning el øL $r7\' Colony
blotfing (Section 216) with CP serum identifred 4 strrongþ- and 4 moderateþ-positive clones
from abankof abouts00 (Figure 5.3). These 8 clones atd2negative clones were deoignated
DS1-DS10 (containingplasnids pPM2lÛ2 to pPM2111, respectiveþ) and subiected tovarious
analyseg
55 Analysis of the clones DS1'DS10
55.1 nVf studiË
CIones DSl-DSlg, togetherwith O17 (recipient strai+ negative control) aúZl756l
(donor s[,ain, positive control) were inoculated onto CEA and incubated et 25 C for 3ó h¡*
EM examination of the hanrected bacteria rwealed that onty the four strongþ-positive clones
(DSZ DS4, DSs, DS6) eryreseed pili with the morphologr of TCP (Figuf€$ 5.4 e 5'Ð'
Indiyidual TCp filaments ?-10 nm in di¡meter were seen to aggrw tß closeþ in parallel to
form bundles 100-æ0 nm in width. The remaining clones and the O17 recipient strain failed
to pr,oduce euch pili under these growth conditions. The donor strain zllÍ6leho$'ed a higþer
level of TCP e:pression than the four O17 cl'ones'
IEM analysis confirmed that the CP senrm çp¡rainss antibodis which bound to the
pili produced by the four Ol? clones, as visuatized by the bindingof protein A-colloidal gold
particles (Figure 5.6 A & B). The pili produced þ clones DS| DS4, DSS and DS6 were ehoun
ro be TCp by IEM using LPS-abco¡bed anti0395 serum (Section 4.3.1 Figrre 5.6 C & D)'
Neither Eerum sþificantly labetled O17 or the other 6 clones.
118

Flgup 53 Colony blotting Appnoximately 800 colonies of a cosnid bank derived f¡om
Vc]øbæ /j756l (ClaEsical Inaba) were screened for tùe Presenoe of non-LPS antþn*
Figuf€ shor16 poeitiveþ reacting clones (indicated þ anowr) whilst * and - signs i¡dbaæ
poeitive (Zl7fil) andnegative (O17) oont¡ols.
119


I
I
!
I
i
.t
j¡
Flgure 5.4 hoduction of TCP þ clone DSZ Poeitive control (2,1756l) and clone DS2
(O17þPM21031) were grown on CEA (with Ap for clone) at25 C for 36 h, harvested in PRS
andappliedtogfidsforEMinspection(Section2l&1). AandB = Zl756t; CandD = DSL
The bars in A and C = ãX) nm and in B and þ = t(þ nm.
tm

È¡
Io
oa
,T
A {a
\t
'n
rt
Tr
"---
'
#

jtf-
IJl
tr
t't,,
:1
Ilgunc 55 hoduction of TCP þ clones DB5 and DSó. Clones DSs (OU[PPM2106D a¡d DS6
(O17üIPM21üID wer€ grour¡ on CEA (with Ap) at25 G harvested in PBS and tran¡fened to
grids for EM inspection (Section 213.1). A and B = DSS; C and D = DSó. The ba¡¡ in A
and C = 5ü) nm, and in B and þ = l(þ nm.
tzt

<T
AF
r+;ãl
F
V
T
V
--
¿
,ì
#o:
\ /, <'T
'¡tt'
--

Figurc 5.6 Immunogold electron microscopy of TCP produced by Clone DS5
(O17[IPM:¿1060. Clone DS5 was growa on CEA (with Ap) at 25 Ç and han'ested in PBS and
t¡ansfered to gdds for IEÌvl" Bacteriawere reactedwith the anti¡erum (anti-5698/165 CP for
A and B; LPS-abeorbed anti-O395 for C and D) and then protein A<olloidal gold as described
in Section Zl&Z The ba¡s oorrespond to 100 nm in eæh case.
tn

-oI
aI
o
.4.
a'/aa
t'ÈUota
oa
ç?aC;¡
¡ao
ti
it .:
t-'.a
O
a
a
I
a,
t
D a''a
o aao
a
a
ratl:-
a. Ia
aþl
TI
¡;, o
-
a

5.5¿ lrr¡mnobbttiryamlFb
For inmunoblottinganalysb, the ten o17 cloner, two positive cont¡ols (strains z1756l
and 5698) and one negative contror (o1?) were cultured on cFA at 25 c fo¡ 3ó h" The resulting
growth was ha¡rrested and the suspensions sorub'ized and subiected to Sffi-pAGE prior to
s/esterû brottingwith cp s€nrtrL In agreementwith the EM observations, onþ the Dsa Dgt'
DS5a¡dDs6cloneqtogetherwirh zl1s6latd56gB,wereposirivebyimmunoblotting(Figre
5.7). In each casg the presence of a 20 kDa pnotein' corfesPonding to the TcpA pilin subunit
(Þylot a aL 1987), w¿s detected'
153 Mohculnr ana[Ffs of cbned DNA
Inordertocha¡acterizetheDNAcariedbyth"spooitiveclones,cnrdeplasmid
preparationeweret¡ansformedinto.EætiK|2stfaitrsM10(seeSectiolZ|7.7,¡,whence
prasmid DNA sui'ble for restriction anarysis could be obtained (re. circumventing the
pr,oblemofDNAdegpdingenzymesproducedby V drcbæ)' This anaþb rwealedthateaph
of rhe 4 TCp-producingclones carried DNAwhich included theltbal fragmentr (Fþre 5'8)
characteristic of the region encoding TCp (D,yrot et d 19ssa). This suggested that moot of the
gpnes requiredforbioe¡nthesis of this pilus hadbeen clonedi¡to asingte coomid' Mobilization
of the plasmids back inlo 01? confirmed that the cloned DNA conferred all of the properties
originalþ associrated with the positive clones'
t8

Ftgurr 5.2 Inmunoblot anaþb of whole cell& llthole cells of the various strains wene grosin
on CEA (plus Ap for the clones) at}SCfor 3ó h. Theywere reruspended in electrophoresis
eample buffer and rubiected to SDS-PAGE The proteins were transferfêd to dtrocellulce
and the blots devel,oped by inorbatbn with 1:100 rabbit anti-5698/165 CP serum, foll'owed
by goat anti-rabbit Igg coupled with ho¡se¡adish peroxklase and ñnalþ eubstrat€ (Section
2.15.2). The sanplee are: A O17; 8,5698; Ç DS10; D DS9; E DS8; R DS7; G, DS6; H,
DS5; L DS4; J, DB3; K DSz; I, D81; M,z'17ft1. The arno$' indicates the poeition of the
molecul¡rr weigþt marker ß-lacrogfobulin (bovine nillç 13,400).
I?A

(-o
A BCD EFG HI JK L]'l
ü

Flgune S.t Restdction endonucleaoe analycb of pPM2lÛ3 (DS2 clone). 5ü) ngof DNAwas
digprtêdwithxbøI at37 Cfor t hr. Electlophoresis of the digest"d DNAwas carried outat
¡oom temperah¡e on horizontal 0.8% 4garoee gel The size of ¡estrictbn fragnents wel€
calculated by comparing their rel¡tive mobility wilh that of .&oRI digot"d fuíIltts st¿btilis
bacteriophage SPP1 DNA"
t25

oN=ô-o-
ÀÀat1
8.377 .20
6.05
4.90
3.55
2 .63
1 .731 .61
1.291.19
2\ kb
5kb
3kb
2kb
bÀÈ--.t

5.6 Confimúion of TC? as a úmlerre determirrant and
non-LPS prctective antigen ofV choletæ
5.61 IC" b a virulenæ deÛerninant
The availability of TCP-eryressing 01? clones provided an opportunity to confirm
prerrious conclugion¡ concerning the rrcle of TCP as a virulence determinant and protective
antþen in the i¡fant mouse model (Section 4.4 & 4.6). Three such clone$ we¡e found to be
o! average, seventy rimes more vinrlent than the O17 recipient strain and a TCP-negative O17
clone @ble 5.1), confirming that production of TCP is sufficient to enhance virulence- Itr
additiotr, protection tests indicated thatantibodies to rcP are suffrcient to confer protection
in this model
5,62 TCP b a protective antígen
The fi¡st eryeriment compared the capacities of TCP-pooitive and TCP-negative Ol7
clones to remove p,r,otective antibodie¡ þ absorption. Rabbit anti-5698Äó5 ¡erum was
abeorbed four times with TCP-negative Z1756l (grourn in NB ehaking culture at 37 C) in an
attempt 1q eþtain a serum whce protective activity was primariþ, if not exclnsiveþ, due to
anti:TCp antibodies. protection tests suggesæd that rhic aim had been achiwd since the
absorbedserumhadapDsoof l55again¡tTCP-negative ZlTÍúlbut4pg0agahstTCP-pooitive
7;1756l (gown on CEA at 25 C). Moreoveç the protective activity agrinst the CA4OI st¡ain"
which poßsesses non-I-pS protective antþns (Section 3.4.1) butdoes notproduce TCP Clhble
4.1), fell from 5,0(X) to ll0 as a result of these abnorptions. Five 2-mt aliçots of the abnorbed
serum were then firrther absorbed with one of the o17 clones or with 21756l as a pooitive
cont¡ol; the bacteria used for these abeorptions were groum on CFA at25 C to optimize TCP
erpression. Dble 5.2 shorrs that 2,17561 and each of three TCP-eryressing O17 clonet
t?ß

Ihble 5.1 hoduction of TCP enhances thevirulence oî'V dnleræOl7
Plâsmid TCPstaûsa VinrlencebSaain
ot7
DSz
DSs
DSó
DS9
pPM2103
pPM2106
pPM2tVt
pPM2110
+
+
+
2.82 x,0.10¡ ld5.66 t 0.40x ld4.38 t 0.13r ld3.Ð x,0.60 x ld3.61 t0.ßxld
" rcP stetns indicates the ability to produce TCP on CFA at 25 Cin?A\'
b Virulenoe is expreosed as the LD50 for infant mice; figur€s show mean t SE of 3 deter-
minations
tn

lbble 52 Absorpion s,ith TcP-expreseing o17 clones femoves pfotective antibodies
Abaorbingstrain
zl756t
DS2
DSs
DSó
DS9
TCP status hrotective activity vs Zl7 ftl
<n
<n
<n
<n
3980 t 130
+
+
+
+
' Th. antisemm uedwas raised in rabbite against the hybrid strah 5698/165 and had been
absoöed 4 times vtr\zllÍdrcr @roum in NB at 3? C) to learre residual protective an-
tibodieswhichwere directed primad¡y ngninstTcP' Alfo¡uots of thb eerumwhich han a
prrotective titer of 4290 t 3Ðwhen assõyedaginst Zl756lrcP+ organiemc,were then
fir¡ther absoröedwith the st¡ai¡s shou¡tr Residual protective acrivity against Zl756lrcP+
was then deærmined.
t8

r€movd all protective activity, wherea' the negative clonewas unable to do so. since the o17
host st¡ain has no non-Lps protective antigenq this result directly demonstrates the protective
efücacy of antibodies to TCP.
In a second experimen! the a¡ti:ICP sefum obtained above was tested for protective
activity "gainst O17 (negative contr ol), 277561 (positive control) and four of the clones'
challenge bacteria were gro!\,n on cFA to promote TCp production and tle results of the
protection assÍrys are shown in Thble 5.3. The absorbed serum did not protect mice from
challenge with o17 or the TCp-negativ eol7 clone, in support of prerrious results (section 4.7)
obtained using a different source of antibodies to TCP. Significantly, ho$'evef, protection was
oboervedagainstchallengewiththe TCP-producingOlT clones @ble5.3), shouringunequivo-
calþ that antibodies to TCP are sufficient for protection in this model'
5.6.3 ÏtCP-produßing Ot7 ctones ¡¡p irrrrunogenic
A final e4erimentwas designed to determinewhether immunization with TCP-produc-
',gorl clonee would elicit the production of antibodies to TCp. Rabbits were immunized with
three of rhe TCp-producingolT clones (DS¿ DS5 and Ds6), one negarive clone (DS9) or the
zr4ftillstrain as a positive control. The immunizingbacteriawere grou¡r on cEA at 25 C for
3ó h,,; afær confirmation of rcp production by Er.4 the grnwth was harvested, suspended in
PBS, and stored in aliquots at -?0C
Before assEnng the resulting aatisera for their capacities to protect infant mice from
challenge with rcP-producing z1756l, it was first necessary to absolb them to remcnre
potentialþ protective antibodies to ogau,a LPS. Thiswas achievedþ absorptionwith live o17
vibrios, and the success of the procedure is verified by the fact that the absorbed sera retained
no protective activity against o17 challenge (Iable 5.4). To determine whether the sera
t29

lhble 53 Antibodies to TCP are protective in the infent mouse chole¡a model
Cïallenge stlain TCP statusa hotective fte¡
zt156t
Z1756l (l.IB)
c4401
ot7
DS2
DSs
DSó
DSg
+ l?9O x,3fr
155t m
110 t 10
<n
4(þt4O
1030 t 94
510 t 45
<x)
+
+
+
stna¡cates the ability to produce TCP on CFA at25 C for Zh.
bff" protective tiær b the PDSO aginst tle various challenge strains' The anticerum uEed
was rabed in rabbig agninsl the hybrid strain 5@Wl65and hadbeen abßoöed 4 time¡ with
zflsa-I$p- Grown in NB atn crto leave residual protective antibodies which \f,'ere
directed PrinaritY ag¡in st TCP
130

Ibble5.4TCP-producingclonesareimmunogenicandprotective
Protective activity of Ol7-absorbed anti¡eraa vs
Immunizing
strain
zl756l
zn56tc
40,000 t 450
1,180 x. lN
1,740 x.VlO
1,570 t 110
<20
DS2
DS5
DSó
DSgb
ol7c
<m
<n
<n
<n
<n
u Rabbits \r,ere immunizedwith the strahs listed, and the resultingantisera ab'¡o¡bed four
timeswith live o17 (see text). lvhen the anti-Zl756lwas further abso¡ùedwith live
zl756lrcp- @ronn in NB at37 C) to remove antibodies to non-Lps protective antigens
other than TCR the PDsowas ¡educed to 25,9(Ð t 330'
b Th" pDso of the unaboorbed anti-DS9 againet o17 was ?.360 x, lgp,.
" challenge bacteria of the z1756lor o17 gtrains wefe cultured on cHl' at 25 G and each
protection test was performed twice'
t3t

contained protective antibodies directed against TC! protection tests were next performed
using TCP-pooitiv eZl756las the challenge bacteria'
The results of these assays @ble 5.a) indicate thatthe three TCP-producingOlT clones
generated protective antibodies to TCR whereas the TCP-negative clone did not Thus TCP
is imm'nogenic when preoented on the o17 clones and antibodies directed against tlese pili
are protective. The antiserum to zrlsîlcontained protective antibodies directed not onþ
against TCp but also against other non-LPS antigens. Further absorption of the ol7-absorbed
a,,¡_zl7s61 serumwith TCp-negative zlTsîlreduced the protective titerby 35To (to25,m)'
Even after this furtler abeorption, the protective activity of the serum was much higþer than
thoseoftheseraraisedagainsttheTcP-producingolTclones,presumabþreflectingthelouler
lerrel of TCP e:rpression by the l'atter'
5.7 Discussion
Initial attempts to clone non-LPS prrrtective antigens rrcmv dtolsæ lm;toa coliwere
unsuccessful In ret¡ospec! the approach adopæd was inappropriate for the cloning of rcP
since it is nou, known that production of TCP requires a functional tuR gene (Taylor er øL 1987)
which would notbe present n E. coli. eccordingly, a nell' cloning strategrwas de*isedwhich
involved the ch¡omosomal integration of a mobilizable plasmid" A cosmid gene bank prepared
fromv cttorqæ zrlsilrwas mobilized into v ehoreræ El ror o1z viln at E coli intermediate
recipientstrain. This was achieved usinga specialþ constructed mobilizable plasmid pPM2101
(Figure 5.2), which contains al.ilkb&antHl fragment encodin gthemob region of RP4' Miller
& Mekalanoo (19gs) recently employed a simila¡ method to introduce DNA into bacterial cells
in a nonreplicating form, to demonstrate úat úoxR regulates tle expression of cholera toxin'
TCP and some oMPs in response to certain environmental sþal* v cholqæ st¡ains z1756l
132

and olzwere selected as donor and recipient strains for the reasotrs e¡tlined above (Section
5.4.1).
The plasmids carriedby the four TCP-producingolT clones contain the 2kb, 3lb, 5tb
a¡dÐlú lftøI f.n,penß which characterize (Taylo r et al.19S8a) t\etcp oPeron (Figure 5'8)'
In addition, when present i\ E coli,these pl,asmids reactwith atcp,A-specific oligonucleotide
probe, confirming the presence of this structural gene (R. Faas! U.H. St¡oeher & P'A' Manning
unpublished data). Together these finding eliminate the possibility thatwhathas been cloned
is the foxR reguratory gene uponwhich TCp expreesion depends (Taylot et al- 1987). Moreover,
theþxR gene has recentlybeen clonedandshows adifferentrest¡iction pattern (P,ru Manning
personal comm unication).
Althougþ EM and immunoblottinganal¡rees har¡e failed to detect TCP expression þ El
Tor stfains (HaU et al- 1988; Section 4.3.3),Taylor et d (198fxù found that each of five clinical
isolates of this biotype carried DNA to which a TCp probe could hybridize. using pPm2103
as a probe for the fcp operon, it has recentþ been established lhatv choløae ol7 and three
ottrer El Tor s,fiains also poosess homologous DNA (U.H. Stroeher & P.A" Manning un-
published). Further shrdy of the plasmids cloned here will be aimed at elucidating the interpþ
between plasmid and chromosomal gene$ which results in TCP production by the o17 strain'
It remains feasible that the plasmids " *ya functional regulatory element other than þxtrl such
æ tcpT (Iaylor et øL lgßa) and that this complements a defective - or repliaces a missing -
controllinggene in v cholqaeol1. Alærnativeþ, the "defectnwhich prevents TCP production
by o17 (under grorvth conditio¡s examined to date) might lie in a structural ggtrê, thougþ a
simple gene mutation would not explain the failure of all El ror st¡ains to produce TCP Finally
it is pocsible that the TCp region can unde¡go rearr¿rngements of the type recently observed
with the genes speciffing pilus production n Morax¿lla bovfu (Marrs et aL 1988)' "nd
that El
Tor strains harre become locked into a non€xPressing mode.
133

The TCp-producingOlT clones permitted further evaluation of the significance of TCP
as avirulence determinant and protective antigen in the infantmouse model The demonst¡a-
tion that TCp expression sþificantly increases tle virulence of o17 (Table 5.1) confirms the
original demonstration of the critical involvement of these pili itr infant mouse pathogenesis
(raylor et d19s7). consistentwith tle role of TCP as a colonization factor (xaylor er aL 1987;
Herringto n et aL 19gs) is the finding that antibodies to these pili are protective, presumabþ
by inhibiting adherence to the intestinal epithelium (Attridge & Rowte'y 1983c)' Thus onþ
TCp-expressing o17 vibrios could remove protective antibodies þ absorption (Table 5'2) and
generate such antibodies upon immunization (Table 5.4). Antibodies to TCP are sufücient to
mediate protection against challenge with TCp-poeitiv ezl756lor o17 (Table 5.3), confirming
previous results with the 5698 and 0395 strains of v c]øteræ (Table 4.3). The protection
obeerved asainst the TCp-positive o17 clones is particularþ signiñcan! as earlier experiments
indicated that antibodies to TCp were not protective against challenge strains of El Tor biotype
(Table 4.3), which fail to express TCP under the growth conditions examined to date (Table
4.1; Hall et at. tg88). T'rme did not permit a study of tlose moderaæþ-positive o17 clones
detected þ coronyblottingwhich did not produce TCp Erryeriment' afe planned to determine
whether these clones eryress noniTCP non-LPS protective antigens'
L34

General Discussion
6.1 Distribution of non-LPS pmtective antigens ofv cholere
pretrious studies with the infant mouse chorera model have demonstrated the existence of
non-LPS protective antigens i¡ v cholqø¿. Antibodies to some of these ¿"lsrminants were
shown to poesess protective capacity (Neoh & Rowley lno, L972; Beltamy et øl' 1975) and
their protective efficacy was greater than that of anti-LpS antibodies (Attridge & Rowþ
19s3c). The presence of non-Lps protective antigens on at least one v dtolqae strain (5698;
Attridge & Rowley 1gg3c), made it critical to determine the distribution of these components
âmongfeoentfield isoliates. If such st¡ains sharednon-Lps proæctive antþens, itwas feltthat
further study of these antigens miÈt errenhrally identify ne$' componenB of vaccine sig-
nificance.
ItwasshowninChapter3thatVcholqacofeitherbiotypeandofbothcommon
sefotyPesexPfesscommonnon-LPSantigens,althougþfouroldlaboratoryElTorstrainslack
these determinants. The reason for this restricæd distribution is unclear' It is unlikely that
these determinanb have been lost from Er Tor strains during prolonged storage in the
laboratory since classical stfains that were stored in the sÍrme ¡nantrer still express tlem'
Perhaps expression of non-LPS protective antigens by the neu, El Tor strains resulß from
environmental interactions between tle two biotypes (Section 3.7). Nevertheless, further
135
I
',1
,ll
!
Ilà
ül1

studyoftheseantigenswasclearþwarranted.SDS-PAGEandimmunoblottinganal¡nesfailed
to identi$ a consistent difference in tle oMp profiles of strains expressingor lackingnon-LPS
protective antigens and attempts to charactenzæthese determinants were unsuccessful (sec-
tion 4.2).
6.2 Role of TCT as a colonization factor and a prctective
antigen
An earlier study of the antibacterial properties of poþclon al antt-V chobræ sera sougþt
to define a consiste ntin vito correliate of antibody-mediaæd protection in the infant mouse
cholera model It was found that the capacity to directly block the i' vitro attachnent of
v chorqæprwided such a correliate, whereas the capacities .q immobilize or agglutinaæ the
bacteria did not (Attridge & Rowley 1gs3c). Experimenß in this thesis have confirmed and
extended this ñnding Each of three sources of antibodies to non-LPS protective antigens
inhibited the in vírro adherence of strains poesessing such determinants (as deñned by proæc-
tion assa¡n). In contras! these sera did not protect infant mice from challenge with, nor block
the attachment of, old laboratory st¡ains of El Tor biotype (Section 3.6.2). The implications
from such data is that at least some of the non-LPS protective components may Play a role in
colonizatiot inviw.
Numerous studies attest to the significance of pili as colonization facto¡s for a variety
of bacterialpathogens, i¡cludingnoninvasive enteropathogenswhich establish toxin-mediaæd
diarrheal syndromee similar to chorera (section 1.3.3.4). Reports publishedduringthe couße
of this thesis resoþed the controversial issue of of wheth et v cholqæalso produces pili (Ehara
4 aL rgfr6; Taylor el aL rggl; Hall et aL lg8l iJ). The e:rperiments of Taylor d aL (1987) ìvefe
of particular relevance, identifying one pilus type - rcp - a virulence determinant in the infant
mouse chorera model Attention therefore hrrned to assessingthe significance of rcP.
.tI
,{
ùì-
It'
tt
''l
I
\t,
)
I
i
#
#
I
L%

Initial work involved optimizing gron'th conditions for TCP e:rpression, and performing
EMandimmunoblottinganaþef to determinewhetrertheorpressionof rcP correlatedwith
the expression of non-Lps protective antigens. These studies ¡ervealed that ín vítro TCP
production (folloving græ,th on cFA) was obsewed onþwith the classical strains (with the
exception of cA401) and notwith the El Tor strains (Table 4'1; Fþres 4'7 &' 4'8)' That is'
whereas non-LPS protective antigens were detected on the neu'El Tor strains or'v dtolqac'
this was not the case with TCp expression. This suggests that TcR arthougþ a major non-LPS
antigeno is not the only protective non-LPS antigen' This point will be discussed further
(Section 6.4).
Subeequent experiments confirmed the sþificance of TCP as avirulence determinant
in this model It was found that non-motile 5698 vibrios were over 100-fold more vinrlent if
culurred under conditions conducive to TCp expression (section 4.4). Eryeriments described
by Trylot d al. (1987\clearþ shou,ed that production of TCP conferred a significant advantage
in terms of persistence in the infant nouse guL Moreover, Herrington et øl' (1988)
demonstrated that TCp expression similarþ promotes coronization in humans. when volun-
teers were challenged witì virulent classical 0395 those who had been previously fed with a
TCP+ryressingstrainwereproæctedagaingli¡'esswhereasthosewhoreceivedthenonjTCP
e:rpressing st¡ain were not protected
þ analog with the pilus colonization factors of ETEC strains (Iærrine er al' 1983), it
was p,redicted that the pivotal role of rcP in vibrio colonization would similarþ confer uPon
these pili the status of protective antigens Although this remains to be demonst¡ated for
human infection, the experiments described in'his thesis show that this is indeed the case in
the infant mouse cholera model. The protective efficacies of antisera shown 1e çoatain
antibodies to TCp by IEM were dramatically increased if the challenge vibrios were TCP-posi-
tive (tble 4.2). one of these sefa was then absorbed to obtain a serum whose prolective
J
,{
ñ*
It'
I¡[.llrì
lrl
tt,
I
I
1
t37

If
f^-
activitywas prinariþ, if notexcrusivel¡ due to antibodies to TCp (section 4.7). This residual
sefum was still highly protective against challenge strains of classical biotype, but did not
protßct ag¡i¡st Er Tor vibrios which fail to produce TCp under the growth conditions tested to
date (Table 4.3).
These initial erçeriments provided the first evidence that rcP is a proæctive antigen
of v chorøae. unequivocal confirmation of this point carne from subsequent studies which
followed the cloning of genes invoþed in rcP production'
63 Molecr¡tar cloning of genes involved in the qynthesis of TcP
AnattemPttodefinethenon-LPSp,roæctiveantigensorVchoba¿involvedthecosmid
cloning of DNA prepared from a recent ñeld isolate, using a cloning stratery discussed earlier
(Section 5.4). This approach resulted in the cloning of genes involved in TCP production and
eryression of these g"Des hv cholcræ or7 , astrain which norma[y l,acks non-LPS protective
antigen* TCP-producing o17 clones lltere' on average' seventy times more virulent than the
unmodifiedrecipient strain (Iable 5.1) confirmingearlier findings' MostsþificanÙ' a sefum
containing antibodies to TCp was shown to protect mice from challenge with rcP-positive 017
clones thougþ not from challenge s'ith (rcp-negative) o17 (fable 5.3). This pmovided une-
quivocal confirmation of previous experiments which had suggested that rcP is a protective
antþen in this model Not surprisingly, the TCP-producing O17 clones' but not O17' could
remove protective antibodies by absorption (fable 5.2) and upoû immunization elicit pfotec-
tive antibodies specific for non-LPS compotretrs (Table 5'4)'
It'
I
I'i
138

6.4 Is TCP the orrly non-LPS prutectirrc antigen ofv drolerae
in the infant mouse model?
TheprotectiveefficacyofantibodiestoTCPraisedthequestionofwhelherthisantþn
is the onþ non-Lps protective component in this model. This was particularþ important
because of the finding that at least one of the antisera used to determine the distribution of
such components among v choreræstrains was found to contain antibodies to TCB despiæ the
fact that the bacteria used in the preparation of this serum had been grolvtr under conditions
unfarrorabre for TCp production (Section 3.5.1). Several rines of evidence suggests that
VchotøæpGs€ssadditionalnon.LPSprotectiveantigens,however.
First, the capacity to synth esizcTCP does not correlatewith the pattern of elpression
ofnon-LPSprotectiveantigensAlthougþthelattercomponentsarefoundonrecentfield
isotatesofElTbrbiotype,suchstrainsfailtoproduceTCPasjudggdbyEM,immunoblotting
and protection e:perimenß (Sections 4.3 & 4.ó). Theee results are consistentwith the presence
of non_TCp, non_Lps protective antþens, at lea^st on recently isorated Er Tor strains. Secon4
the existence of sefotype-fest¡icted non-Lps prctective components similarb¡ afgues against
TCP being the only such antþen, since TCP synthesis is not serotype related. Thir4 the C4401
strain possesses non-LPS protective antigens (Table 3'3) yet is atypical "molg strains of
classical biotype in that it fails to produce rcp rt viúro (section 4.3.3). Finatþ, t\e z1756l
st¡ain e'identþ possesses non:TC! non-LPS protective antigens (table 5'4)'
65 Non-LPS protec{¡Ye antigens: relerant shrdies in other
laboratories
Theerperimentsdescr¡bedintlisthesishaveidentifiedTCPasthefirstnon-LPS
protective antigen or. v cholqae, at least in the infant mouse model' workers in several
I
I
139

laboratories are shrdying the nature of vibrio colonization, as colonization factors would be
prime candidatee for protective antigens'
The important contributions of Tayloç Mekalanos and colleagues have been referred
to througþout ttris thesis, and represent a maior step forward in the quest for an improved
choleravaccine. This group has recentry puriñed rcp from a flagellum-negative derivative of
V chobæO395 and shorn n that a resulting antiTCP Eerum wÍ¡s higþly protective in tle infant
mouse model, confirmingresults obtained in this thesis (Iaylor ef øl 1988b)'
To date, TCP remains the onþ defined adhesin orV eholøøe (althougþ there is some
evidence to suggest that Lps can promote attachment - section 1.3.3.2). peterson & Mekalanos
(lgss) identified naccessory colonization factorsn in the infant rnouse model þ studying
TorR_regulated getres n v cholqae. Tiansposon insertions in any of a cluster of four genes
resulted in a much reduced capacity to colonize the i¡fant mouse gut thougþ the effectwas
less dramaticthan thataccompanyingdeletion of genes encodingTCp (peterson & Mekalanos
198s).Thenatureoftheaccessofycolonizationfactorsremainstobedefined.
Recentstudies have investigated the role of HAs hv chotqæ colofiwation' Ftn¡e| aL
(1gg7) constnrcted a mutant El Tor strain which l,acked the mannoee-sensitive cell-associaæd
fIA (Hanne & Finkelstein 1gg2) and found that this mutant showed a markedly impaired
capacity to cotonize the rabbit ileum. Nevertheless, oral immunization with the mutant
p¡otected rabbits from subsequent challenge with virulent v chorøae (Finn er ar 1987\'
AlthougÞ this would seem to indicate that antibodies to ^h i s rIA are not necessary for protection
h this qrstem, furtrer studies wourd be needed to eriminate the poosibility that the mutant
produces an inactive HAwhich retains the potentiar to elicit neutralizing antibodiec
Tepp"ma eJ øL (lggi) reported tlat a motile, (mannose-resistant) HApositive, El Tor
strain and an aflagellate HA-negative, classical st¡ain showed simila¡ capacities to colonize
ligated ileal loops constructed in adult rabbits. on this basis itwas concluded that factors other
140

than hemag€rutinating activity or motility are invoþed in colonization in this model. This
illustrated the importance of the erperimental s¡ætem in shrdies of vibrio pathogeneeis, as the
conclusion reached by Teppema e t at (Lgsi)conflicts with the results of several earlier shrdies
attesting to the significance of the flagellar st¡ucture in colonization (see section 1'3'3'1)'
Attachment to the intestinal surface evidentþ occurs more readily in a statig closed system
such as a ligated gutloop; indeed, some cholera moders require a drug-mediated inhibition of
peristalsis (Freter 19554; Finn ¿f aL 1987)'
Duringthepastdecade,sevetalgoupshaveadoptedtheRITARD(removableintes-
rinal tie - adult rabbit diarrhea) model developed þ spira et aL (1981) to shrdy infection by'
¡¡dimmunityto,Vcholsæ.Theoriginalprocedureinvolvedthetemporaryobotnrctionof
the smallbowel to promotebacterial colonization and the permanentligation of the cecum to
prevent fluid resorption, although modifications to this s¡rstem have been descrbed (Guinée
et aL LgËÐ. colonization facto¡s and protective antigens have not yet been defined for the
RT'ARD moder. Li,ev crølqæimmunize much more effectively aeainst suboequent chal-
lenge ¡¡¿¡ killed bacteria (Guinée et aL 1987; Riipkem ae,aL 198?; Pierce et al' 1988)' with
the capacity for mucooal association pnrviding the onþ correlate of protective effrcacy (Pierce
d aL rggs>. protection in this moder is associated with a marked inhibition of intestinal
colonization (Guinée 4 aL rggÍ; þcke et at- rgÍ36: pierce et øL tgSs), bur the speciñcity of
the protective antibodies remains unknoç'n'
Svennerholm et¿l have found that intestinal immunization with rough Vcholeræ
mutants can protect rabbits from subsequent challenge with fuþ virulent smooth cholera
vibrioe in the abeence of any anti-LFS antibody respome (cited by Jonson eJ aL 1989)' Thit
suggests a protective role for nol-LpS antibodies. This is in contrast to their earlier observa-
tions indicating that protective capacity in the rabbit model resided exclusiveþ in anti-LPS
antibody fraction (Svennerholm 1gg0), a finding that seemed to harre been corroborated by
141

Jansen et oL (lg8S). Nevertheless, this recent work þ Svennerholm (cited þ Jonson ef øl
l9s9) extends the notion that non-Lps protective antigens might be important ' o,¡"¡ ¿¡imal
models besides the infant mouse model'
In 19g3 Sciortino & Finkelstein reported that during growth in the intestines of infant
rabbits, v choleraedisplay a different ¡nttern of oMps from that characteristic of organisms
culhrred ìnvitro. This important finding focuses attention upon antigens expressed during
gro$,thín vivo,aptentiallyreu,ardingapproachforinvestigatorsinterestedinvaccinedevelop-
ment Recentry Jonson et øt. (rgEg) confirmed that novel proteins are producedby v choleræ
during gowth in the (adult) rabbit gut, and went on to shou' that at least some of tlese are
immunogenic" of interestwas the finding that several of the proteins induced antibodies that
were detected in convalescent phase sera from cholera patients (Jonson et aL 1989)' Howeveç
none of these novel proteins aPpeaf to be TCP as kvúto growth under conditions conducive
to Tcp erpression did not induce any of t\e invirao-specific proteins nor did anti:TCP serum
rcactwitl these proteins. clearþ it will be of great importance to define the antigens
qrnthesized fi V chobae during growth in the human gut
6.6 lhrhrrc studies
6ó1 Non-LFS proÛective antþns of El Tbrv cloþrae
The data presented in this thesis are consistentwith a biotype-associated expression of
TCP, but further studies in this anea ane required" Althougþ EM and immunoblotting ap
proaches failed to detect production of such pili by strains of El ror biotype, this migbt mereþ
reflect the fact that the s''nuli for TCP s¡rntheeis differ between the biotlpes. As discussed
prerriously (Section 5.7'¡ anumber of El Tor strains have now been shown to carry DNA
homologous to that encoding TCp. Taylor d aL (rgssb) have recently reported 807o sequence
homologr between fcpA genes present in the classical strain 5698 and the El ror F,1946'
142

potential antigenic epitopes were found to be hlghly conservd suggesting that protective
antigens niÉt be shared between the pilus stn¡ctures eryressed by the two biotypes (Taylor
eføl 1988b).
The protective efficacy of antibodies to TCP was evaluated against a panel of challenge
st¡ains to address the possibility that El ror strains synthesize rcP rn wvo. These experimenb
also shoured a clear, biotype-related dichotomy horve'er, in trat such antibodies onþ conferred
protection egrinst bacteria of classicar biotype (T"ble 4.3). It should be noted tlat proÞctive
titers against classical strains are low unless the challenç organisrN afe expressing TCP (as
is necessarily tre case with El ror strains). An independent means of assessing the in vivo
signiñcance of rcp for El ror v choleræwould be to construct strains carrying inactivated
tcp¡genæ(forexample) andtocomparethe colonization of suchstrainswith theircorresPotrd-
ingwildtypeparents.SucheryerimentsarecufÎetrtlyinprogress.
In addition, an attemptwillbe made to clonethe non-LPS protective antigens of recent
El ror isolates, using the approach which allowed cloning of genes involved in TCP erpression'
Any cloned components will be investigated for their capacities to promote in vito attachment
a¡d ínyira colonization. IEM will be used to examine their di¡tribution ovef the bacterial
surface, with particuliar interest in the poosibility that El fe¡ strains produce a biotype-as-
sociated plus stnrcture as suggesæd ry :Hail etal (19gg). As already discussed (Section 6'4)'
eryide¡ce suggests r\at v choraae erytess nonJTC! non-I-pS protective antþens and this
approach could lead to their identification In this context ttre four po'sitive 017 clones which
wereTCP-negative(Section5.5.1)meritfurtherstudy.
6;62 Th nahrc of TCP'rediatßd attachrent
The speciñcity of bacærial adherence is imparted þ the recognition of target cell
receptors by bacterial attachment factors. Evidence is accumulating to suggest that the actual
r43

adhesins of gram-negative bacteria consbt of minor protein molecubs at the tip of the surface
pili rhere is nor¡ ample eryidence that gþcolipirts can act as recepûo¡s for bacterial adhetins
(revieured þ I-effler & svanborg-Eden 1gs6). Adhesion of piliated E- øli to host cells has
been shown to be mediated by pilur-"ssociated adhesins of the Pap pilus not þ pilin subunit
pøffi(uhlin a al.1985). Th" pßotein associatedwith adhesion' has been found exclusiveþ at
the tip of the pilus and ie a 35 kDa protcin (PapG; Normarkø al' 19ú; Lindbefg¿ú al'1987)
that attaches to the gfoboeeries of gþcolipids with the recognition siæ being Gatcl4€alß in
human urinary tract epithelia (t-efiler & svanborg-Eden 1936)' Other enmples of lectin-like
tip adhesins produced þ E coti a¡e a r2tDa pnotein of s pili that recognizes sialic ackl (Moch
et aI. tg87) and a 29 kDa plotein produced by t',Pe 1 fimbriae which mediates attachment to
D-mannoce (Abrahan & Beachey 1988)'
It is not tnou¡n whether the attachment of ll clplsæ to hoet cells i8 mediated þ similar
adhesins,whichmãybveryming¡componentsofthepilue.ByanaloglwithltcolÍ,such
proteins nist harre a criti€t role in adheeion oî' v choløæ to host cells' Based on the
organization of genes in the Pap operon (Lindbergdll 198Ð, Taylor etaL (1988a) have
pnopoeea a model for the gene clusrer invoþed in the biosS¡ltheris of rcP (Figure 6'1)' The
tcpA geneproduces the naior ¡ilin subunit, witb lqG rcpænting a possible tip adhesin'
Furthe¡ molecul¡ar and biological an ryses invoMng the constnrction of defined mutant strains
ie required to elucidate the nechanism of rcp-mediated attachmenL such studies will alloril
the design of vaccines aimed at inhibitin g v cholaæ colonizatio¡ þ eliciting antibodies
capable ef ¡s¡t¡alizing the binding potential of TCP'
The nahrre of host receptors invoþed i¡ V cløteræ adherence ¡pmains undefi¡ed'
T¡ylor etal. (1987) hate shown rhatvchotqæ 0395 eryressing pili could hemagglutinate
nurine RBC8 thus demonstrating that the pili apparentþ bind to receptors on the surface of
hostcells. The nature of the receptorwas notidentified but the aggtutinationwas detected
144

Figune 6.1 hoposed organization of the genes associated with the production of the TCP
pilus; (Taylor er ø1. 1988a). T\e hal fraÊment containing /cpG does not appear to be
immediateþ contþous with the remainder of the genes shown here.
145

Xbo
rN)
lx-
LoI w () rl-
]-T
-l
Xbo
l
bol
X
C)
Xbo
l

in the presenoe of L-fucoee, a sugar that inhibits most of the hemaggfutination mediated þ
v clplqæo3gs (Jones & heær rg76; Jones 1gs0). one should be aware, hm'ever, that the
characteristicsofbindingtoRBCsnigþtdifferf¡omthenah¡reof thebacterialinæractionwith
moße relevantsubetrates (Attridge & Rowley 1983b)'
6.63 \haine Potentiat of the TC?
It seems probable that future studies wilt sho$' TCP to be a protective antigen for human
chole¡aic infection. Txybr da¿ (rgssa) harre úeady suggested the incorponation of rcP into
the oral ki¡ed whole cell vaccine devel,oped þ Hotagfen's gloup (Section l'10'2J2\'u'hich
would presentþ contain [ttle if any of thiß antigen. Thb night lead not onþ to an imP¡wement
in proæctive etñcecy but also to a reductioa in coeü if the preeence of TCP permin at ba¡t a
ûnnsþnt associ¡rtion of the kilþd bæteria with the intestinal surflce i1 migþt be pcsible to
¡educe the number of bacæria per dose'
otherwa¡n to eryloittheprtative protectivepotentialof TCP include thedevelopment
of a TCP-B subu¡¡it coniugrte (faylor et aL'tgïsa) or the delivery of TcP otr atr atlenuated
carrier etraitr such as s. typhirf2la. The latter could also eryress the pnotective o-antþens'
as p,revio'sly di¡cu$ed (Section 1.10.23)' hesumably rcP would be synthesizdhvivoby
attenuated live oral vaccine candidates, althougþ the onþ data ar¡ailable to date suggest that
suchstrainselicitanti:TCpresponsesinonþasmallminorityof recipients (I-evinea 41 1988b)'
Itwould be of interestto compare the immunogenicity, protective efñcacy and reactogencity
of va¡iou¡ doeeß of TCP-pooitive and TCP-negative non-toxþnicV ffi¡w'
The impact of incorporatingTCp into poæn*ial choleravacci¡e candidates is ãs'aited
with great interesL As alrcady discr¡sse4 it is poosible that any improvement in protective
efrcac,ywill be limited to infecti,ons causod by cra$ical v ùotqae. It would nerreÍìeless
eppear thatan efiectivecholeravacci¡e b æn¡ider¡bly clæer than itwasiusta fen'years ago
tß

BibliographY
Abraham s.N. & Beachey E H. (1gs5) Host defenses against adhesion of bacteria to mucosal
surfaces. Adu. Host Def' Mech 4: 63-88'
Abraham s.N. & Beachey E H. (lgss) Binding of bacteria to mucosal surfaces. Monogr
Allerg A'- ß43'
Achtman M., Schwuchou¡ S, Helmuth R, Morelli G' & Manning P"{ (19S) Cell-cell
interactions in conjuga rngbrchøichía cori: con- mutants and stabilization of mating
aggtegús. Mol¿¡. G4' &n4' lúf: 171-183'
Adhikari p.c & chatterjee s.N. (1969) Fimbriation and pellicle formation olwb¡úo El ror.
Indinttl- MeA 57: 1897-1901'
Al_Kaiesi E, & Moetratos A, (1gs5) .preparation and properties or.vibrío cholqæ aof'fimbnal
antibdy. J. AppL Bøctqiol Stz 2l-229'
Alm RA, Braun G., Morona R. & M¡nningP.A" (19S6) Detection of an ompA-like protein
nwbr¡ocholeræ.FnusMigobioLl-cfl.37;99-104.
Attridge SR. (199) Infection by, ¿¡d immunity to, Wbt¡o chobæ' PhD thesis' Adelaide'
Ar¡stralia: The University of Adelaide'
Attridge s.R. & Rowþ D. (19S3a) The role of the ftagellum in the adherence ollñhrio
chobae- J- InfeÃ- Dús' l.47z 8&A7Z
147

Attridge s.R. & Rowley D. (19s3b) The speciñcity of wbrio choleræ adherence and the
significance of the stime aggtutinin as asecond mediator of ínvtto attachment J' Infea
Dis. t4ilz 873-881.
Attridge s.R. & Rou,ley D. (19S3c) Prophylactic sþificance of the nonlipopoþsaccharide
antigensofWbríoeholcrac.Ilnfect.Dls.l4E:93|.939.
Baine lV.B., TampienA., Mazzotti M, AngioniG., Greco D', Di Gioia M"luþF^' Gangarosa
EJ. & Pocchiari E onq Epidemiolog of cholera in Itâly in 1973' Lqtcet' 2z
1370-1374.
Banwell J..G, Pierce N.E, Mitra R.C, Brigham I(L, Caranasoo GJ" KeimowiU RL' Fedson
D.S, Thomas J., Gorbach sL, Sack R.B. & Mondal ,{ (19?0) Intestinal fluid and
electrolyæ transport in human cholera. I. Cl¡r\^ Inve'Í. 4Ì 183-195'
Bart trCJ., HuqZ.,Khan M. & Mosley \il.H. (1970) seroepidemiologic studies during a
simultaneous epidemic of infection with El ror ogava and classical Inaba víbrío
elpbøe. J. Infed. Dús Lllt S17-SZ['
BaruaD.&Cbatteriees.N. (1964) Elect¡onmicroecopy of$lTotlnbriocholøaa Ind'IMed'
Rcs. 52:828S30.
Barua D. & C\ietanoic B. (1970) cholera during the period l%l-ln}' lrlz hh"ípl6 a"d
præticeof choløøcqúrol Public Health Papef No' 40' \il'H'OT Gener¡a pp' 15-21'
Bauman P., Furniss A.L & I-ee J.V. (1984) The genus whño. In: Bergey's manual of qrstemic
bacteriologl.BtN.R.KriegVolumel.,lsteditionpp.5lS-538.
Bellamy J.F-G, Ihop J., Steele E J., ChaicumPa \il. & Rorley D' (1975) Antibody cross-link-
ingasafactori¡immunitytocholeraininfantmice.t.Infed.Dis.|321181-188.
Benenson ,{.S., Islam M.R & Greenougþ Itr W.B. (1964) RaPrd identification of Víbrto
cholqaebydartûeldmicroscopy.BulLWldHtthorg.*'8n.83|.
148

Bhalla D.K & Or¡¡en R.L (1gs3) Migration of B and T þmphocytes to M cells in Peyer's
patches of the mouse after neonatal thymectomy andhydrocortisone iniectio¡' Am' J'
Attol. l¡SLz TZ7-88.
Bhaskaran IC (1g1) Isolation of hybridvibrio strains for immunological research in cholera
l'ù; hoceedinp of ø sympaium on búqìalvaccíne* Ed" B. Gusic Tagteb,Yugoolavia:
Yugoslav Academy of Arts and Science' pp' 2A7 -?ßl'
BienenstockJ. & Befus A.D. (1930) Mucosal immunologr. Immunologt' 4lz ?A9-Z7O'
Bienenstock J., Befus A.D., McDermott M., Mirski s., Rosential Ic & Tagliabue A" (1983)
The mucooal immunological network compartmentalization of þmphocyteq naürral
killer cells and mastcells. .Ann N.YAcad sci 4t9z 164-170'
BlackR.F^ (19Só) Theepidemiologrof choleraandenterotongeúcBæhøìdtíacolidianheal
disease.ln:DettelopnølsofvaccinesmddrugsøgainsdiøtheallthNobelCon-
ference, stockholm, 1935. Studentlitteratur, Ilr¡4 Sreden
Black R.F^, I¡rrine M.M., Clements M.L, Young CR', $ennerholm ^À-lvf" & Holmgren J'
(19g2) protective effrcacy in humans of killed whole-vibrio oral cholera vaccine with
and without the B subunit of cholera toxin. hfecL h¡vnun 55: 1116-1120'
Blake P.A (19S0) Diseases of humans (other than cholera) caused by víbríos' '$n Rcu'
Misobbl 342 Al-367'
Blake p.¡u, Allegra D.T, Synder J.D., Barrett TJ., McFarland L, Cararray cT, Feeley J'C'
craigJ.P.. Lee J.V, Puhr N.D. & Feldmen RA. (1930) cholera - a pæsible endemic
focus in the united States Nen'hg.I. Med 3ll2: 305-309'
Blake P.,A,., Rooenberg.IU-L, Coeta J.B, Ferreira P'S', Guimaraes CL & Gangarosa EJ'
(1y77') Cholera in Portuga\ ln4. I. Modes of transmission Am I' Epidøtiol 1:oSz
337-v3.
149

Booth B.A, & Finkelsæin RA, (19só) Presence of hemagglutinin/protease and other poten-
tial virulence factors in 01 and nono lvibrio chobae L InfæLDis' 154: 183-186'
Booth B.A, Sciortino Cv & Finkelstein R..¿r. (1986) Adhesins ol Víbrb eholqae. In:
Míqobíallcaíw øn¿øglutínírls- E¿ D' Mirelman' Neu' York: Johq lViley & Sons' pp'
169-l82
Bornside G.H. (1931) Jaime Ferran and preventive inoculation against cholera fulL lûst'
Md. 55: 516-532
Bornside G.H. (1gs2) waldemar Haflkine's choleravaccines and the Ferran-Haffkine priority
dispute. J- HisL Men- Ahied Scí 372 39942lz.
Bourard c & I-ecroisey ,{. (19g2) specfic antisera produced by direct immunization with
slices of poþacrylamide gel conaining small amounts of protein' I' hrannoL Meihods
sÈ, m-nß.
Bourke.ATC, Cossins Y.N., Gray B.R.W., I-unney T'I', Rostron N'4" Holmes RV' Gfiggs
ER,I.arsenD.J.&IGlkvR.(198ó)Investigationofcholeraacquiredfromthe
riverine environment in Queensland' Mel- I'Atts1, t4Él. 229-234'
Brandtzaegp (lgsÐ Rore of J chain and secretofy component in receptor-mediated glandular
and hepatic transport of immunoglobulins in man. scotd I ínunnol' 222 111-14ó'
Brayton P.R, C-ohtr'ell R.R, Tall 8.D., Herrrington D' & Leryine M'M' (1937) Recovery of
wfu¡o cttoløæ 01 from stools of human volunteers fed non-culturable cells' In:
Abstruds of 23rd II.S.-lrym Choba C-onføence, Wíllia nsbta. p,zL.
Brinton Jr. C.C (1959) Non-flagellar appendagø of bacteria Nature Lß: 782-786'
Brinton Jr. c:c. (lgs) The piliation phase syrdrome and the uses of purified pili in disease
control ltz hoceedhy of îhe 13th loínt c-:otfaatce on clwlera E(t c Miller'
PublicationNo.(MH)78-l5g0.DepartmentofHeålth'BlucationandWelfare'
lVashingtoû, D.C. PP' 34-70'
150

Bror¡n KH., parryL, Khatun M. & Ahmadc. (1979a) Lactooe malaboorption inBangladeshi
village children: relation dü ag", history of recent diarrhea, nutritional status and
breast feeding Amen I. Ctí't' Nutr' 32" 1962-1969'
Brourn M.CM., Weston À, Saunders J.R & Humphreys G'O' (199b) Ïì'ansformation of
E&het¡ctti\ cotí Koobyplasmid DNA at different phases of growtl' FEMS Mbobbl
Lú. 5z 219-2,22-
Burnet EM. (1g4g) ovomucin as a substrate for the mucinolytic enzymes ol V choleræ
filtraæs. Attsl- I E'x"- Bíol' Med tl: ?A5-25L
Burnet EM. & Stone J.D. (rg47) Desquamation of rhe inresrinar epithelium in víto w v
ciøIqaeñltrates: characterization of mucinase and tissue disinægrating enzyDe' AusL
l. EA. BioL Med Sci 25: 219-nß'
Carpentar CCJ., Barua D., \trhllace CK, Mit¡a P.P., Sack RB', Khanra S'R" Wells $A'' Dnns
pB & Chaudhuri R.N. (196ó) Clinical studies in Asiatic cholera. IV Antibiotic
therapyincholera.ButLloltt:a¡HopkfuHasp.|':az2l6.D.
Carpenter cGI., Sack RB., Feeley J.c & Steenberg R.lv. (1t}6s) Siæ and characteristics of
electrolyte loos and effect of intraluminal gfucose in eryerimental canine cholera' J
Clh. Inves- 472 L2l0-1Z20'
cafy s.G. & Blair E B" (1%4) Nec/ ü'lnsPort medium for shipment of clinical specimens' 1'
Fecal specimens. I' Baaerid EE: 9(í-98'
Cash R..{., Music SL, Libonati J.P, CraþJ.P., Pierce N'n' & Hornick R 'B' (1974a) Response
of man to infection with vibrio chotqae. II. Protection from illness afforded by previous
disease andvaccine. I' InfectDn 131Ì 325-333'
cash R.d, Music S.L, Libonati J.P., Schwartz AR & Hornick R'B' (194b) Li"" oral cholera
vaccine: evaluation of the clinical effectiveness of two strains ¡¡ ft¡mans ' InfecL
Intnun- Lù' 762:7&.
151

cash RA,, Music S.L, Libonati J.P., Synder MJ., lVenzel R.P. & Hornick RB' Gn4c)
Response of man to i¡fection with Vibrb choleræ. I. Clinical, se¡ologic and bac-
teriologic responses to a known inoculum. l. Infect. Dis' l;zy' 45-52
qD.q (1936) Toxigenic VíbrüocholqæOlinfections -LouisianaandFlotrrda- M'M'WR 35t
ffi.607.
chaicumpa rv. & Atthasishtha N. (1gg) Th" study of i¡testitral immunity against v e]rcbae:
purification olv elplqa¿ El ror haemaggfutinin and the protective role of its antibody
in erperimental cholera southeas Asian L Tlop. Med- PubL HhîIL l:Ù' 72-79'
Chaicumpa w. & Rourley D. (lgzzj F.ryerimental cholera in infant mice: protective effects
of antibody. J- InfecrDic lZ5: 480485'
Chen LC, Black R.B, Sardar 4.M., Merson M.H., Bhatia S', Yunus M' & Chahaborty J'
(1930) Villaæ based distribution of oral rehydration therapy packets in Bangþdesh'
An I. TroP- Med HYg. 2*' 285-290'
chitnis D.s., sharma KD. & Ibmat R.s. (19s2â) Role of bacterial adhesion in the
pathogenesisofchole¡a.].Med.Mbúiol15:43.51.
chitnis D.s., sharma I(D. & Ikmât Rs (19g2b) Role of somaric anrigen olwbrb choleræ
in adhesion to intestinal mucos u I. Med Microbiol 15: 53ó1'
Clemens J.D., Sack D..{., Harris J.R, Chakraborty J., Khan M'R, Stanton B'F" I(ay B'4" Khan
lvf.u., Yunus M., Alkinson lv., Svennerholm A-M. & Holmgren J' (198ó) Field trial
of oral cholera vaccine¡ in Bangþdesh. I-øtcd- 2: l?A-127.
Clemens J.D., Sack D.A., Harris J.R, van l-oon E, Chalrabofty J', Ahmed E'' Rao M'R" Khan
M.R,YunusM.,HudaN.,StantonB.E,,tr(ayB.A.,\\ålterS.,EeckelsR.,Svennerholm
A"-M. & Holmgren J. (1gg0) Field trial of oral choleravaccines in BangÞdesh: results
from tlree-year follow-up' I'ancel' Lz 270-/73'
r52

clemens J.D., Stanton" BH,, Chakraborty J., SackD.A" Khan M'R, Huda s., Ahmed E' Hafris
J.R., Yunus lyf", Khat lvLu., sennerholn'{.-M" Jertborn M' & Holngren J' (1987)
B subunit-whole cell and whole cellonly oral vaccines against cholera: shtdies on
reactogenicity and immunogenicity' I' InfeclDís 155: 7945'
Clemens J.D., Harri8 J.R., Sack D.*¿1", Chakraborty J', Ahmed n'' Stanton B'E' Khan M'U.' KÂy
BA,HudaN.,Khanlvf.R.,YunusM.,RaoM.R,SvennerholmA'-M'HolmgrenJ'
(19SS) FieldtriatoforalcholeravaccinesinBangladesh: Resultsofoneyearfollou'-up
I.InfectDís- 15& 60ó9'
clements M.L I-evine M.M., Young c.R, BLack R.E., Lim Y'-L, Robins-Broü'tre RM' &
craigJ.P.(1932)Magnitude,kineticsanddurationofvibriocidalantibodyfesPonses
in North Americans after ingestion of v,br¡o cholqæ' J' Infect"Dh 145: 465473'
clitr J.L, zinkin p. & Marrelli ,{ (19só) A hoepital outbreak of cholera in Map.to, Mozam-
bique. Trø¡la R Soy'Top' Med Hyg' *' 473-476'
colwell RR, Brayton P.R., Grimes D.J., RoezakP.R" HuqS*À & Palmerl,M' (1985a) Viable
but non+ulurrable lnbtío chobøeand relaæd pathogens in the environment implica-
tions for release of geneticalþ engineered organis ms' Bioædtttologt' 3z 817'8?f,'
colwell R.R., Ikper J. & Joseph s.lv. (197 7) wbtio chobøe v pørahaønolytiLas and other
vibrioo: occuffence and distribution in chesapeake Bay' sc¡qce 19E: 394'396'
C-olwell RR, Singfeton EL, Huq ,{., Xu H.S. & Roberts N. (19s5b) Etiologr of Vibrúo
c}øløøe,Vpøahættøtyticlßandrelatedvibrioointhenatufalenvi¡rcnment.In:
BaærialDúÚlæItDiseases.Ed&Y.Takeda&TMiwatantTokyo:KÏKScientific
hrblications . PP. 273-Ð5'
C.olwell RR, Tamplin lvLt-, Brayton P.R, Gauzens A'L, Tall BD" Herrington D" I-evine
lvf.M., Hall s., Huq r¿r. & sack D.A (1987) Environmental aspects oî'vtbrb choletæ
153

in transmission of cholera
tiamsbttrg. PP.33-34.
Abstracts of 23rd ILS--Iøpan Ctwlcra C';oferarce' Wíl'
craig Sw. & Cebra JJ. $nÐ Peyer,s patches: an enriched souroe of prec'ursors for IgA
producinginmunocytes in the ¡abbiL f' EtP'Met' t34z 188-æ0'
Cryo SJ. Jr., Fürer E. & Germanier R. (19s2) E,ffectof chemical andheatinactivation on the
antþenicity ¿¡| immunogenicity or'lftrb clwlqæ Infær Imtruln ß: 21.2ß.
cnatrecasas p (rg73,) Interaction or. víbrb crørerre enterotoxin with cell membranee'
Biochanisry- t2: 3547 -3558'
cnrlin G., I-evine R, Aziz lclvf,a, Rahman AS'M'M' & verwey lv'E (1976) Field trial of
cholera toxoid. Proc. of the |IthIr c-mtr. ctwlsa ([LS'-Ipn Coop' Med' sci ProgrmtL
pP.3L4-3D.
Crrrman B, Kampe o., Rask L & Peterson P.,â1. (199) Pre¡ence of alloreactive Ia antiçns
on murine intestine epithelial cells. scøú.I. Innuttol 10: 11-15'
Dastidar sG. & Narayanaswani (r%s) The occurrence of chitinase invibrios' h¿liml' Med
R¿s. 56: 654658'
DeS.N. (1959) Enterotoxicityofbacæria-freeculturefiltraæ ollöríocholeræ' Ndwe' !t3z
1533-1534.
Deb B.C, Sircar BjC, Sehgupta P.G., De S.P., Mondal s.trc, Gupta D'N., Saha N'C' Ghosh s.,
Mit¡aU.&PalS.c(19E6)StudbsoninterventionstopreventElTorcholera
transmissioninurbanslums.BulLWld-HltILor8.6:lZ7.l3l.
DeMeyJ.&MoemansM.(1986)heparationofcolloidalgoldprobesandtheiruseas
m¡rkers in elect¡on microecopy. l¡zAàvøtcedTechníweßhMological ElætonMiso*
coPY- E/.-J.K Koehler' 3: D-nL
Dixon J.M. (1960) The fate of bacteria in the small intesrine. I- pdhoLBøctqíol 7q 131-140'
154

Duguid J.p. & old D.c (1gs0) Adhesive properties or. Enterobæterùaceac. rrz Bætøí41
úlwance, ræEtbrs mdrecognirbn E(l. E M. Beachey' London: chapman & Hall'
SeriesB 6: 185-217.
Duguid J.P., Smith w., Dempster G. & Edmunds P.N. (1955) Non-flagellar filamentous
aprpendages ffrmbriae) and hemagglutinating activity in Búqíwt coli' I' PAhIþL
Baaqíol 7ù 335-348'
Dutt,{.trL, Alwi S. & Velauthan T (1971) A shelfish-borne cholera oubreak in Malaysia
Bløts-R Soe Tlop' Med Hyg' 6: 815{18'
Ehara rvr., Ishibashi M., Ichinooe y.,Iwanag lvr", shimodori s. & Naito T (19s7) Purification
andpartialcharacterizationofñmbriaeoî'Vîbrbcholqøeol.Væcíne.52283-?ß.
EharaM., Ichinoce Y., Naito T & Shimodoris. (19SS) Fimbriae or'v clnlera¿ 01' Abstacts
of24thlointC-onføanceUS.IapanC-æperatíveMedicatscíencehogram.Clwlcrø&
ReldedDiûTlßalDúseaæsPøteJ.Tokyo,Japan.p.48.
Ehara M.,Ishibashi M., Watanabe S., Iwanaga M., Shimodori s & Naito T (1936) Fimbriae
ol'lilbriocholqøeol..oboervationoffimbriaeontheorganismsadherenttolhe
intestinal epithelium and development of a ner¡' medium to enhance fimbriae produc-
nor. TIoP- Med, 2E: 21-33'
EubanbBR.,GuenlzelM.N.&Berry r-J. Qnr Evaluationofsurtacecomponents or,r4brb
ehoer/æasprotectiveimmunogelxlnfect.Immun15:533.538.
þyans D.G., Evans Jr. DJ., CleggS. & Pauley J.A" (199) Purification and characterization
oftheCFA/Iantiçnofenterotoxigeltc&scheñchilcotí.Infø.Irranun25z738-748.
Evans D.G., Evans Jr. DJ., Ïioa w.s. & Dupont H'L (198) Detection and characterization
of colonization factor of enterotoxigenic E$heñchia coli isolated from adults with
diarrhea. Infect- Intrutn ty' 7n'7%'
I
I
d,Ë
155

Faris ,4., Lindahl M. & wadstrom T (1gg2) Higþ surface hydrophobicity of hemagglutinating
Víbriocholqaeandotlervibrios.CunMicvobioLTz35T-362.
FeachemRG. (1gs2) Environmentalaspectsof choreraepidemiolog. III. Tiansmissionand
control TroP-Dis'BulL 79t t-47'
Feerey J.c. (1970) cholera vaccines. rn. principres and practices of chorera control- Public
HealthPaperNo'40' Geneva: W'H'O' pp' 87-93'
Feeley J.* & Gangarrcsa E_J. (r9s0) Field triat of cholera vaccine. rnz chorøa andRelæed
Diørheøs. Eds. o. ouchterlony & J. Holmgren. 43rd Nobel Symposium, Stockholm
1978. Karger, Basel' PP'204-210'
Felsenfeld o. (1963) some observations on the chorera (El ror) epidemic in 1961-196L BulL
Wld HlrlL Org' 2E:' ?ß9-296'
Felsenfeld o. (1%5) Notes on foo4 be'erages and fomites confaminatedwith víbrio choreræ-
BulL lltd HtttL OÌg' 33: 7?5-734'
Felsenfeldo.(|974)Thesurvivalofcholeravibrios.In:Cholera.Eds.D.Ba¡ua&w.
Burrows. l,ondon: W'B' Saunders' pp' 359-366'
Finkerstein R.A. (1962) vibriocidar antibody inhibition (vAI) analysis: A technique for the
identification of the predominantvibriocidal antibodies in serum and for the recogni-
tion and identification of víbio choleraeantigens' J' hnmunot' sÌ 264-271'
Finkelstein R.A. (1973) Choler t CRC Crit.Iteu. Microbíol 2: 553-623.
FinkelsteinR.A (1975) Immunoloryof cholera cunTop.MicrobioLlrvnunoL 69 137-796'
Finkelstein R.A (1934) Cholera In:
Academic Press' PP' 107-136'
BactqialVaccines- Ed. Rene Germanier' New York
Finkelstein RA, erita M., Clements J.D. & Nelson ET (197s) Isolation and purification of
an adhesive factor (cholera lectinn) rromvíbrio chorqøe- rn; hoceed,ings of the 13th
I.rt
rìì'
I
It
T¡
I
I
II
#
fl
156

i,t
È-
Jt.Cüf.Chotqa.(IS-Ipn.Cæp.Med^ScLÞogrøn.DHEÏVPubLno.78-1590.Na-
tionallnstitutes of Health, Bethesda' Md' pp'137'l5l'
Finkelstein R.,A,., Boesman-Finkelstein M. & Holt P' (19S3) víbrio choleræ hemag-
gtutininÄectin/proteasehydro\zesñbronectinandovomucin:F.M.Burnetrer¡isiæd.
hoc- NatL Acad' Scí' U'SA' &): 1092-1095'
Finkelstein R.A. & Dorner E (19s5) Cholera enterotoxin (choleragen)' Phmnac Thq' 27:
37-47.
Finkelstein R.A, & Hanne LE, (1gs2) purification and charactenzation of soluble hemag- ,,
gtutinin (cholera lectin) produced by víbrio chorcrae. Infect. hnmun. 36; 1199-1208' ì
Finkel¡tein R.A & LoSpalluto JJ. ( 1góg) pathogenesis of experimental cholera: preparation
and isolation of choleragen and choleragenoid' J' F-xp'Med L3O:' 185-2frZ-
Finkelstein R.A & I-ospalluto JJ. (1970) hoduction of higþly purified choleragen and ì
t
choleragenoid. J' InfecL Dß' 1121: 563-572'
Finkerstein R.A. & Mukerjee s. (1963) Hemaggtutination: arapidmethodfordifferentiating
Víbrio cholqaeand El Torvibrios . hoc. Soc. Exp Biol- Med 112: 355-359'
Finn TM., Reiser J., Germanier R. & cryz S.J. Jr. (19s7) cell-associated hemaggfutinindefi-
cientmutantolVtbríocholqæ.InfecLImmu¡t552942.946.
Fishman RH. (1gg0) Mechanisms of action of cholera toxin: studies in lag period. I. Mqn'
BioL 54 6l-7L
FloreyH.w.(1933)observationsonfunctionsofmucusandtheeaflystagesofbacterial
invasion of the intestinal mucosa J- Pathot. Bacteriol' 37: 2Í33-?ß9'
Fo'ett E"A.c & Gordon J. (1963) An electron microscope study of vibrio flagella J. Gen'
i¡
il
f.'
TtI
I
I
i
Microbiol. 32: ?35-239'
t57

I
{
h-
Forrest 8.D., I-aBrooy J.T, At¡.idgs S.R., Boehm G', Beyer L, Morona R' Shearman D'J'C'I
& Rowley D. (1939) Immunogenicity of a candidate live oral typhoidicholera hybrid jï
vaccine in humans ' I' Infect'DLs 159: L45-146' i
Franzon vL & Manning pd (1gs6) Molecular cloning and expression in Esctprichia coli i
K-l2ofthegeneforahemagglutininrromVíbriocholqøe.hfect.Immw.52z279-?ß4.
Fresh J.W., Versage P.M. & Reyes V (1964) Intes'inal morpholory in human and experimental
cholera. Arctt-Pah' 77: 5Ð-537'
Freter R. (1955a) The fatal cholera infection in the guinea pig achieved by inhibition of
normalentericflora' I' Infecf' Dis' llz 57{15'
Freter R. (1g55b) The serologic character of cholera vibrio mucinase. I- Infect- Dis. llz
88-245.
Freþr R., Alhveiss 8., o,Brien PC.M., Halstead S.A. & Macsai M'S' (19s1) Role of
chemotaxis in the association of motile bacteria with intestinal mucosa: in vito studies'
Infect- bnmun- 342 Vl[-Vl9'
Freter R. & Gangafosa E.J. (1963) oral immunization and production of coproantibody in
human volunteers ' J' bnrnwtol' 9fr: 72A-7D'
Freter R. & Jones G.W. (1976) Adhesive properties o1 Vibrio chorøaet nature of the
interaction with intact mucosal surfaces. Infect' hnmun' t4z 246-?56'
Freter R & o,Brien pcM. (lgsla) Role of chemotaxis in the association of motile bacteria
with intestinal mucosa: chemotactic responses ol.l1brb chol'qøe and description of
notile non-chemotactic mutants. Infecr. Immutt^ 342 215 -DL
Freter R. & o,Brien pcM. (lgslb) Role of chemotaxis in the asociation of motile bacteria
withintesitnalmucosæfitnessandvirulenceofnon-chemotacticVíbriocholeræ
mutants in infant mice' btþcL Immuru 34l. 2'2-733'
158

Fubar¿ E"S. & Freter R. (1973) protection against enteric bacterial infection by secretory IgA
antibodies. J- Imtnunol' l't.lz 395-403'
Gaastra w. & de Graaf F.IC (1gs2) Host specific fimbrial adhesins of non-invasive
enterotoxi geúc4,scherirhiacoü strains. MíctobioL Rat' 4e 129-t6l'
Gallut J. (1974\ The cholera vibrios. lltz cholera' Ed" D' Barua & w' Burrows' I-ondon:
W.B. Saunde¡s & Co' PP' 17-40'
Gangarosa EJ., Beisel W.R., Benyajati C., Sprinz H' & Piyaratn P' (1960) The nature of the
gastrointestinal lesion in Asiatic cholera and its relation to pathogenesis: a biopsy study'
Arn.I. TÌoP. Med HYg' 9: 125-135'
Gangarosa E.J., Sonati ,{., Saghari H. & Feeley J'C' (1967) Multiple serotypes of
Víbrioclwlqaefromacaseofcholeral.anceLt646-648.
GargersJ., Griffith o.M. & GrillLK (1gs3) Raptdpurification of prasmidDNAbfyasingle
centrifugtion in a two step cesium chloride-ethidium bromide gradienL Bioùq¡t
Biophys- Res Conmun' LL7" 835-842
GermanierR (1934) Tirphoidfever. l,.: Bacterialvaccines Ed'ReneGermanier' orliando:
Academic Press. PP' 137-165'
GhoshH.IC (19?0) Thepathogenesisofexperimentalcholera J'Med'Misobiol 3z 427440'
GiUD.M. (1976) The arrangemenrof subunis in choleratoxin. Bioùønistry- LSz lvl2-1248'
GillD.M. (1977) Mechanismofactionofcholeratoxin. Adu'cyclicNud'Res' E:85-118'
Giælson s. (1971) Gast¡ectomy, achlorhydria and cholera. Isaetl' Med sci 7z 663-6f|7'
Glass R.I., Becker s., Huq M.I., Stoll 8.J., Khan M.U., Merson M'H.' L'ee J'V' & Black R'E'
(1932) Endemic cholera in mral Bangtadesh, 1966-1980' Am' J' Epidemiol 116:
959-970.
Glass RL, Huq M.I., Alim A"R.M.A & Yunus M. (1930) Emergence of multþþ antibiotic-
resistant wbrio cholerain Bangladesh. I- Infect. Dis' 1r42z 939-942
159

Glass R.I., Iæe J.V., Huq lvf.I., Hossain trCM.B. & Khan M.R. (19s3) Phage types of
VíbriocholqaeolbiotypeEl]brisolatedfrompatientsandfaniþcontactsin
Bangtadesh:epidemiologicimplications.J.Infed.Dirl4t:998.1004.
Glass R.I., Svennerholm AM., Khan M.R., Huda S., Huq M'I' & Holmgren J' (1985)
SeroepidemiologicalshrdiesofElTorcholerainBangtadesh:associationofserum
antibodylevelswithprotection'J'Infect'Dis'151:2%-242
Greenougþ w.B.trI. (1965) pancreatic and hepatic h¡ipersecretion in cholera. Lørceî- 2z
Ðr-994.
Greenougb W.B.II[ (rg7g) wbrb ch.oreræ. rnz hittciptes mdhactíce of InfectíousDiseases
Eds. G.L Mandetl, R.G. Douglas Jr. & J.E BennetL New York: John rililey & Sons'
pP.l672-1ffi7.
Greenougþ w.B.Itr. (1930) Principles and prospects in the treahent of cholera and related
dianheas. lnz cholqa&RelntedDisrheas. Eds. o. ouchærlony&J' Holmgren' 43rd
Nobel Symposium, Stoclùolm lnï' Base} Krager' pp'2ll2l8'
Greenough w.B. IIL Gordon R.S., Rosenberg LS.' Darries B'I' & Benenson A'S' (19ó4)
Tetracyclineinthet¡eatmentofcholeral-ancelLz355-357.
Greenougb vÍ.B.Itr. & Rabbani G.H. (1936) Antisecretory and antimicrobiat drug for treating
diarrhea rrz Darclopntant of vaccines mdDntgs øgøinstDiorrtø. Eds. J. Holmgren'
,dUndberg&R'Möllb}''llthNobelConf"StockholmlgS5'S¡¡eden:Studentlit-
teratur. PP.27O-277'
Guentzel M.N., Amerine D., Guerrero D. & Gay TV (1931) Associatiot oî'Wbrio cholcræ
mutants with the intestinal muçosa of infant mice' Scmnhg Elecfron Micraçc' 4z
Llí-t2t4.
GuenEel M.N. & Berry LJ. (1975) Motility as a virulence factor lor Vibrio chorqae- Infect
hrunun. 11: 890-897.
160

GuenEel M.N., Field LH., Eubanks E"R. & Berry LJ. (1977) Use of fluorescent antitody in
studiesqfimmunitytocholeraininfa¡tmice.hþc|htpnun.15:539.548.
Guinée p.ÀM., Jansen w.H., Gielen H., Rijpkema S.G.T & Peters P.lvJ. (1987) hotective
immuni$againstwbliocholqaeinfectionintherabbitZbl.BkH.Hyg.A.266,..
552-562-
Guinée p.A.M., Jansen lV.H. & peters p.w. (1gg5) Wbtio chorerøe infection and acquired
immunityinanadultrabbitmodelZbt'Ba&''Hyg'A259:118-131'
Hall R.H., Vial P.4., Kaper J.8., Mekalanos JJ. & levine M'M' (1983) Morphological studies
onfimbriaeexPressedbyVíbríoc}pleraco|.MictobiatPathqøt.42757-2ß5.
Hanne LE & Finkelstein R.A, (19s2) characterization and distribution of the hemag-
gtutinins producedby víbrio clnlqae hfect- Imtnun 36:' ?fr9'214'
Hansson H._.d, Holmgren J. & svennerholm L (rgii) ultrastructural localization of cell
membraneGMrganeliosideþcholeratoxin.ProcNalLAcadScLUSA.T4z3TS2-
37K.
Hawkes R.B, Niday E. & Gordon J. (1982) A dot immunobinding assay for monoclonal and
other antibodies. Anal' Biodtem LL9 142-147'
HenningU.,Schwar1¿,H.&ChenR(lg7g)Radioimmunologicalscreeningmethodfor
specificmembraneProteins.Anat-Biochøt.9|lzlS3.|57.
HerringtonD.A,HallRH.,LooonsþG.,Mek¡lanosJJ',TaylorR'IC&LevineM'M'(1988)
Ïbxin, toxin-coregulated pili, and the foxR regulon a¡e essential for wbilo cholcræ
pathogenesis iû humans t' Exp' Med l:6Ez 1487-1492^
Hirschhorn N., Kinzie J.L, Sachan D.8., Northrup R'S', Taylor J'O" Ahmad S'Z' &' Phillips
R.A'(1%s)Decreaseinnetstooloutputinchildrenduringintestjnalperfusionwith
gfucose-containing solutions' New Eng' I' Med 2792 176-18l'
16r

Hirschhorn N., Pierce N.E, Kobari I( & carpenter ccJ' (1974) The treatment of cholera
hz clwlera Eds. D. Barua & w. Burrou¡s. I-ondon: lV'B' Saunders' pp' 23525:¿.
Hisatsune K, Kondo s., I(awata T & Kishimoto Y' onÐ Fatty acid composition of
lipopoþsaccha¡ides olVíbrìo cholqae35A3 (Inaba), NIH90 (ogaura) and 4715 (NAG)'
I. Bactøíol- 13E: 288-290'
Hohn B. & coilins J. (1gs0) A small coomid for efücient cloning of large DNA fragmenrs
Gqre Ll: Ðl-?9.8.
Holmgren J. (1gs1) Actions of cholera toxin and the prevention and t¡eatment of cholera
Naatre. 2922 413-417.
Holmgren J., Clemens J., Sack D.A, & svennerholm A'-M' (1989) Nerr cholera vacci¡es'
Vaccine. Tz 94'96.
Holmgren J. & l,önnroth I. (1976) Cholera toxin and the adenylate cyclase-activating signal
I. InfecLDi* 1i]3: Só4-S74'
Holmgren J., l_õnmoth I., Mansson J.-B & svennerholm L (1975a) Interaction of cholera
toxinandmembraneGMlgangtosideofthesmallintestine.hoc.NatLAcdSci.T2z
25m-2524.
Holmgren J. & rycke N. (1gs6) Immune mechanisms in entericinfections. hz Developmant
of vrccinæ øtd dntp again¡ diærhea Eds' J' Holmgren, A' Lindberg & R' Möllb5''
llth Nobel Conf., Stockholm 19s5. Sveden: Studentliterahtr. pp'9'ZZ
Holmgren J. & $ennerholm A.-M. (1gs5) Vaccine detelopment for the control of cholera
and related toxin-induced diarrhoeal diseases'
diseæe. L.ondon: Pitman PP' ?A2'?5,6'
In: Miqobiat tox¡n and diæthoeal
Ho}ngren J., Svennerholm rÁ\.-M., I-önnroth I', Fall-Persson M" Markman B' & Lundbeck H'
$nT Development of improved cholera vaccine based on subunit toxoid' Næute
269l. 6U2-604-
162

Holmgren J., Svennerholm À-M., OuchterlonY O', AnderssonA" Wallerström G' & lVeste¡-
berg.Berntssonu.(195b)Antitoxicimmunityinorperimentalcholeraprotection
and serum and local antibody responses in rabbits after enteric and parenteral im-
munizati,ot Infect Immun t2z 133l-1340'
HondaT & FinkelsteinR.A. (lg7g) selection andcharacteristics of.al/ìbrb eholaæ mutant
lacking the A (ADp-ribosytatinÐ portion of the cholera enterotoxin . Proe NatL Acad'
Sci U.SA- 76; Ð52-?Jùl56'
Hood M.A, & Ness G.E, (1gs2) suwival or,vibrío chorqæ andgscherüehía corí in estuarine
watersandsedimells.AppLfu'virØ.Miqobío|43:578-584.
Hornick R.8., Music S.I., Wenzel R., Cash R., Libonati J'P', Synder M'J' & lvood\ñ'ard TE
(191) The Broad St¡eet Pump revisited: resPome of volunteers to ingested cholera
vibrios. ButL N-Y. Acad- Med 47: 1181-1191'
Ilraniuþ I(W., Mulholland ,¿\., La¡son ,{D., Eubanks ER' & Hart LT (1980) charac-
terization of a flagellar sheath protein or'wbrio cholqæ' Infecr Im¡¡un Tl: 597ó03'
HuS R. (1%4) The proposed conservation of the generic name l/ibrío Pacini 1854 and
designation of the neotyPe strain ol Víbrúo cholqøe Pacini 1854' ht BulL Bacrqiol
Nomer Tax^ L4z 87-101'
Husband .ÀJ. & Gou,ans J.L (lgs) The origin and antþendependent distribution of
IgA-containingcellsintheintestine.r.Ery.Med.t4E:1146-1160.
Huq 4., Huq S.,{., Grimes DJ., O',Bfien lvL, chu trLH., C-ap]1tzzoJ'M' & C-otwell R'R' (1986)
c-olonization of tìe gut of the blue (.xab (cattineaes sryià/s)w wbtio cholcræ' AppL
Environ MbobbL 52: 586-588'
Ibrahim A,E.A, (19M) rmmunity to experimental cholera in the rabbit ph.D. thesis, utrecht
the Netherlands: The University of Ut¡echt
t63

Inman LR, Cantey J.R. & Formar s.B. (1gs6) Colonization, virulencæ and mucooal interac-
tion of an enteropathogenic,Erh qich¡ocoli (str. RDEC- 1) e:pressin g shiplla somatic
antþen in rabbit inæstine' I' InfectDrs' 154: 742-751'
International Study Group. (rg77) A positive effect on the nutrition of Phitippine children of
anoralgtucoeeælectroþæsolutiongivenathomeforthetreatmentofdianheaB¿ll
Wld- Htttl Org- 55: 87-94'
Jann 8., Jann trL & Beyert G.O. (193) 2-amino-2,6-dideory-D-gtucose (Dquinovosamine):
a constifirent of the lipopoþaccharide
531-534.
ol.Vtbrío clwlqae- European I' Bíochøl 372
Jansen lv.[L, Gieþn H, Riipkena s.G.T & Guinée Pfllvf (19SS) priming and boosting of
the rabbit intestinal immütr€ system with live and kitle4 smooth and rougþ V clwlcræ
ceÍs. MbobíatPatlogen 4: 2l-?ß'
Jertborn lvf,, Sennerholn A.-M. & Holmgren J' (1984) Gut mucooal' salivary and serum
anti_toxicandanti_bacterial antibody f€ßpons€s in svedes afterorarimmunizationwith
B subunit-whole cell choleravaccine. hrArc]æ'AltøgtAppL furntnol 75: 38-43'
Jertborn M., Svennerholm A,-M. & Holmgren J' (19Só) Saliva' breast milk and serum
antibody resPonses as indirect meiasufes of intestinal immunity afær oral'cholera
vaccintionornaturaldbease.I.ctínMigobiolAl?Ã3ffi.
Jones G.rv. (1gE0) The adhesive properties of wbrio chorsæ and other r4bì4j species-
RecqL Rerogtit- Set B' 6z 2Ð-?A9'
Jo¡es G.W., Abrams G.D. & Freter R (196) Adhesive p¡operties oî lörb c]øløaez
adhesion to isolated rabbit brush border membranes and hemagglutinating activity'
Infea- Im¡run. t4: 232-239'
tu

JonesG.w.&FreterR.(1976\Adhesivepropertiesor'Wbrioc}plerae:natureofthe
interaction with isolated rabbit brush border membranee and human erythrocytes'
Infect. Imman- t4z 2'40-?A5'
Jones G.w. & Rutter J .M. (rg72) Role of Kgg antigen in the pathogenesis of neonatardiarrhea
causedbyEschøichíøcotí|n,prdets.Infeclbnmun.6z9|8-127.
Jonson G., Svennerholm A._M. & Holmgren J. (1gSg) wbúo clprsae expresses cell surface
antigens during intestinal infection which are not expressed during invitro culture'
Inþa. Immtn- 57: 1809-1815'
Joo I. OnÐ cholera vaccines. l¡¡: cholera Eds" D' Barua & w' Burror¡'s' London: w'B'
Saunders& Co. PP' 333-355'
Kabir s. (1gg0) composition and immunochemical properties of outer membrane proteins
of.Vîbrb cholqæ' f' Buds¡oL t44z 382-389'
Ikbirs. (1gs2) characrerization of rhe ripoporysaccharide fromwbrb dplcræ395 (Ogau'a)'
Infea. bnnan ß: l?ß3-lZlL
IGbir s. (1933) Immunochemical properties of the m4or outer membrane protein or'víbrío
c]ølqae- hfect Inmun 39' 452455'
IKamatAM.$n4)Theseventhpandemicofcholeraln:CholgaEds.D.Ba¡ua&w.
Burrow¡- I-ondon: W'B' Saunders pp' 1-14'
IGper J.B. (1989'¡ l4brio elplsaev,accines. Rcu. Infea.Drs' 11: s568-S573'
Kaper J.B., L,ock6aû H, Baldini M.M. & I-evine M'lvf, (19Saa) A recombinant live and
choleravaccine. Bioæúnologt' 2¡' 345-349'
KaperJ.B.,I-ockman H., Baldini M.M. & I-evine M.lYf" (19s4b) Recombinantnontoxinogenic
Vtbrbc.holøæstrahs as attentuatedcholeravaccinecandidates' Nafrne' 3lB: 655ó58'
IGllog D.S, Peacock w.L, Deacon 1V.8, Brown L & Pirkle ct ( 1963) Ne,sdø goørrheoea'
I. Virulence geneticalþ li¡ked to clonalvariation' I' Baøìol E5: ln4'lng'
165

I(elly J.T & parker cD. (1gs1) Identification and p,reliminary characterization of wbtio
clpbaeoutermemb¡aneproteins.I.Bøcteríol145:1018-1024.
Kenne L, Lindberg 8., Unger P., Holme T & Holmgren J. $n9\' Structural studies of the
Wbrío cltolqaeO-antþn ' Øohyd'2R¿s' 6& l4-L6'
Khan M.U. & Greenough w.B. (1gs5) Epidemiolog of diarrhear diseases in Bangþdesh. In:
Bacterial d¡ûrhcnl dìseases' Eds' Y'
Publications. PP. 37 -5z-
Thkeda & T Miwatani. Toþo: KTK Scientific
Khan M.U., Samadi A.R, Huq M.I., Yunus M. & Eusof '{' (1934) Simultaneous classical and
El Tor cholera in Bangþdee h' I' Die' De* Rec 2: 13-18'
Kingc,A,. &van Heyningen w.E" (1973) Deactivation of cholera toxinby asiaridase-resistant
monosialosyl gangtioside ' J' hfect' Dís L2ilz 639-il7'
Kobayishi T, Enomotu s, sakazaki R. & Kuwahara s' (1%3) A new selective isolation
medium for the vifuiogroup: on a modified Nakanishi's medium (rcBS agar medium).
laP. I- BactqiaL LE¡' 387 -3lZ
Korhonen TIC, Nurmiaho E-L, Ranta H. & Edén c$ (1980) Nen'method for isolation of
immunologcally pure pili from E¡c]ærichia coli' Infea' Im¡n'utt' 272 569-575'
I-aBrec BH., Sprinz IL, Schneider H. & Formal sB' (1%Ð l-ocâlizåiton of vibrios in
experimental choleræ a fluorescent antibody study in guinea-pigs' Proc choL Rcc
Swp(Horclulu).U.S.PubLHlth.Serv.Pub.No.1328.pp,Z72.Z76.
I-aicy.,MendezE &changD. (ry76) Chemistryofcholeratoxin: treeubunitstructure'
I.InfecLDtu 133: S23-S30'
Lankford c.E & Legsomburana IJ. (1%5) Virulence factors of choleragenic vibrios. In:
ProcChot.M.Swp(Horcfuht).U.s.PubtHlth.Sew.Pub.No.1328.pp.109-120.
Lång H*4, & Paþa ET (19S?) A maior outer membrane protein or' lürío úolqæ ß
maltooe-inducible' Mioobiat Pæhogøt' 3z 143-147'
t6

r-ãtt^T (1331-1332) Malig'ant cholera: documents communicated by the central Board of
Health,I-ondon, rel¡ativetothe treatnentof cholerabythe copiousiniectionof aqueous
and saline fluids into theveins I'arcet' 2" 274-177'
r-efrrer H. & Svanborg_Edén c (1gg6) Gþcolipids as receptors rot Esdtqúùia corí lectins
or adhesins. In: Miqobiat lætitts md, øgþtkit's' Ed' D' Mirelman' New York: John
WileY & Sons. PP' 83-111'
I-er¡ine M.M. (1gg0) Immunity to cholera as evaluated in vorunteers. In: Chorerø mdreløed
dianheas. Bls. o. ouchterlony & J. Holmgren' 43rd Nobel $ropqiun' stockholm
1978. Basel S' Karger' pp' 195-203'
Irrrine M.M., Bl,ack R.F^, Clements M.L, Cisneros L, Nalin D'R & Young CR' (1981a)
Duration of infectionderived immunity to cholera I' Infø'Dfs' 143: 818{20'
I-eirine lvf.lvL, Btack R.E, Clements M.L, Cisneroa L, Saah A" Nalin D'R' Gill D'M" Cratg
J.P., YoungcR. & Ristaino P (1932) The pathogenecity of non+¡terotoxþenic wb¡o
chotøæserogfouPolbiotypeElTorisolatedfromseulagewaterinBrazilJ.Infecl
Dús. 145: 296-D9.
I-evine lvLM., BlackR.E , Clemenß M.L, Nalin D'R, Cbneros L & Finkelstein R'A' (1981b)
volunteer studies in development of vaccines egeinst cholera and enterotoxþenic
E*hqicttia cori: arerriew. rrz Aante enteñc hfeßtíons h chitùan- Ne*' prospecß fø
teøtmenl md præention FII& T Holme, J. Holmgren, M.H. Merson & R Mölby.
Amsærdam: ElsevierNorthHolland Biomedical Press pp 443459'
Irvine lvLM., Black RB, Clements M.L, I¡¡ata c, seals s', Honda T', Young cR' &
FinkelsteinR.A.(19s4)Evaluationinhumansofattenuatedwfuúoc}plgæElTor
ogawa strain Texas Star-sR as a live oral vaccin e' Infæt In¡runn 432 515-52
t67

I-evine M.M, Kaper J.8., Black R.E., & clements lvf'L (1933) New knou'ledge on
pathogeneoisofbacterialentericinfectionsasappliedtovaccinedevelopment.
Miqobiol Reu- 472 510-550'
I-evine M.M., Kaper J.8., Herrington D., Ketley J., I-osonsky G', Dcket co.' Dll B' e Ûyz
S. (19SSa) Safety, immunosgnicity a¡d efücacy of recombinant live oral cholera
vaccines, CDV 103 and CDV 103-Hg[.' Intø' 2: 46f'470'
I-evine M.M., KaperJ.B., Herringbn D., tosonsþG" Morris J'G.' Clements M'L' BlackRE '
Tall B & Hau R (19SSb) Volunteer studies of deletion mutants ofVîbrío clwlqae OL
prepared þ recombinanttechniqu æ' hfect' Innrun 56¡' 16l-167'
I-errine M.M., Nalin D.R., Craig J.P., Hoover D., Bergquist E J', Watenn¿rn D" Holley H'P"
HornickR.B.,PierceN.E&ubonatiJ.P.(19?9)Immunitytocholerainman:relative
role of antibacterialversus antitoxic immunity. Trans-R. Soc- Trop-Med Hyg' 732 3-9'
Levine lvf.ìá., Young cR., Hugþes TP., O'fbnnell S.' Black RB, Clements lvf,L' Robins-
Brou¡ne R. & Lim Y.-L (1981c) Du¡ation of serum antitoxin response follc,'Frfo¡gl4brfø.
chotsæ infection in North Americans:
EpËdmbL 114: 348-354'
relevance for seroepidemiologt' Am' J'
tin EY.C, Morris J.G. Jr., IbPer J.8., Gross T, Michalski J., Monison c' Libonati J'P' &
IsraelF^ (1ggó) persistenceof cholerainlheunitedstates: isolation or.víbríocholcræ
01 from a patient with diarrhea in Marylan d" I. ctin Mísobíol 232 6?A6?:6'
Lindberg¡, Lund 8., Johansso¡ L & Normark s (19sÐ I-ocalizåtion of the receptor-binding
p,rotein adhesin at the tip of the bacterial pilus Nøaue' 32E: 84-87'
ündh E. (1925) Increased resistâûce of immunogtobulin A dimers to proteolytic degradation
aftefbindingofsecretorycomponent.J.ÚnmwolL|422u.286^
168

I indholm L, Holmgren J., lVitst¡6m M., trkrlsson IJ., Andersson K & þcke N' (1983)
Monoclonal antibodies to cholera toxin with special reference to croes-reactions with
Eæhs¡IttiT æli heat-labile enterotoxin. Infed- bnmun' 4u 570-576'
Lonnroth I. & Hotmgren J. (rg3) subunit sürcture of cholera toxin. I. Gøt- MíqobbL 762
[fl4n.I-olvry O.H., Rooebrougþ N.J., Farr A.L & Randall RJ' (1951) Protein measureme¡twith
theFolinphenolreagenLl.BíoLC\eml!,322ß5.275.
Lospalluto J.J. & Finkelstein R.A. (Lg72) Chemical and physical propertiee of cholera
exoænterotoxin (choleragen) and its spontaneoueþ formed toxoid (choleragenoid)'
Bíochsn. Biophys- Acta 2572 158-166'
I-ugtenberg 8., Meiiers J., Peters R, van der Hoek P. & van Alphen L (195) Electropherctic
resolution of the lmaior outer menbrane protein" o1 Erchsichia colí KlZ into four
baûds. FønS l*fr- 5E; 254-258'
Þcke N., Eriben L & Holmgren J. (19S7a) Protection against cholera toxin after oral
immunization is thymus-dependent and associated with intestinal production of
neutralizinglgAantitoxin.ScødI.Invnwol25:413-419.
Þcke N., Hellstrom U. & Holmgren J. (1987b) Circulating cholera antitoxin memory cells
in the blood one year after oral choleravaccination in humans . Scand' I' ImmunoL 262
n7-21L.
I-y"k" N. & Holmgren J. (1987) Long-term cholera antitoxin memofy in the gut can be
triggered to antibody formation associated with protection within hours of an oral
challengeimmunizalto¡-Scørlú'I'ImtrutnolÉ:1A74lZ
þcke N. & Holmgren J. ( lggg) Adoptive transfer of gut mucoeal antitoxin memory þ isoLaþd
B cells l year afær oral immunization with cholera toxin . Infect - Imnu¿n 57 2 ll37 -ll4l'
t69

IyckeN.,IindþolmL&HolmgrenJ.(1985)Choleraantibodyproductionhvitroby
peripheralbloodþmphocytesfollowingq¡¿timmrrnizationofhumansandmice.dih
BE.Immu¡øL 62t 3947'
þcke N., Svennerholm.{.M. & Holmgren J. (198ó) strongbiotype andserotype cross-Protec-
tive antibacteriat and antitoxic immunity in rabbie after cholera infection. Mbobial
Paùogat L: 361-371'
Magnusson KE & stiernstrom I. (1gs2) Mucooar barrier mechanism¡. Interplay between
secretory IgA GIgA), IgG and mucins on the surface properties and association of
sabtonellacwith intestine and granulocytes . Immanotog¡t' ßz B9'?A8'
Mahalanabis D. (1gs1) Rehydration therapy in diarrhea. r¡z Aq¿tc entqíc ínfectíøs h
chilùan.Eds.THolme,J.Holmgren,M.H.Merson&RMollþ.Amsterdam:
Elsevier. PP. 303-318'
Mahalanabis D. & Merson rvr" (lgsó) Deveropement of an improrred formul,ation of orar
rehydration salts (oRs) with antidiarrheal and nutritional properties: a"su¡rer oRsn'
|nzDevebpttaúofVrcchæaúDntpøgahstDiarrheaEds.J.Holmgren,A"Iindbery
& R Moltby. llth Nobel conf., stockholm, 1935' $veden: studentlitæratur' Pp'
2n-2s6.
Mandara M.p. & Mhalu F.S (1gs0) cholera cont¡or in an inacceesible district in Dnzania:
importanceoftemporaryruralcenters.Med-I.Zmúia.15:10.13.
Manbtis T, Fritsch EF. & Sambrook J.c (1932) Motcotb ctonhg: a lúortury mannl
C-old Spring Harbor laboratory' C-old SpringHarbor' N'Y'
Manning P.,{., Heuzenrrceder lvL\\t, Yeadon J., Læavesle'y D'I.' Ree'ves P'R' & RO¡/ley D'
(19Só) Molecular cloning and eryression in Fscnq¡a¡a cotiKlzof the o antigens of
thel¡abaandogwaserotyPesofthewbríocholqæollþpoþsaccharirlesandtheir
poæntialforvaccinedorelopmeltlnfectlmmutlsSzTl2-T17.
170

Manning P.4., Imbesi F' & Ha¡rnes D'R. (1932) Cell envelope proteins inWbtúo cholqæ'
FEI¿IS Mbobiol l-en Lilz 159-166.'
Marrs CE, Ruehl lv.w., Schoolnik G.K & Falkow S. (19Es) Pitin gsne phase variation of
It[oræclhbovisiscausedbyaninversionofthepilinçnes.I.BadqíoLflÙ.3f/32-3['39.
Martin .{.R, Mqley w.H., Sau 8.8., Ahmad s. & Huq I' (1J}69) Epidemiologic ana$nis of
endemic cholera in urban EastPakista¡1964-lg6' Am'f' Epktcm¡ot Spt 572-582'
Mazia D., Schatten G. & sale w. (1g7s) Adhesion of cells to surfaces coatedu'ith poly-lysine'
l. CclI BioL 6z 198-200'
McCormack w.M., Mæley \il.H., Mohammed E & Benenson A'S (1J)69) Endemic cholera
in rural Fast Pakis ta¡' Am' I' Epídmiol' SÈ 393-404'
McDermottM.R.&BienenstockJ.Qng,)Evidenceforacommonmucosalimmunologic
syÉteD I. Migration of B immunoblasts into intestinal' respiratory and genital tissues'
J- In¡nnol L22: t8l2-1898'
MclntyreR.C,TiraT,FtoodT&BlakeP.A"(1979)Modesoft¡ansmissionofcholeraina
nady infected poprlation on an atoll: implications for control meaf¡ufe8' LmceÍ' lz
3ll-314.
Mekalanæ J.J. (1gs5) Cholera toxin: genetic anaþsis, regulation and role in pathogenesis'
Gut Top MisobioL bnmutPl 11E: 97-118'
Mestecþ J. (1gS7) The oommon mucosal immune system and current strategier for i¡duction
of immune resPonses i¡ efernalsecretions' I' ClÍn In¡nanol 7z ?ß5'T16'
Mestecþ J. & Mcchee J.R. (1987) Immunogtobulin A (Ig[): molecular and cellular
inte¡actions involved in IgA bioeynthesis and immune response' Adu' hwìtutpL 4Ù'
ts3-245.
t7l

Mhalu F,S., Mmafi P.lv. & Iiumba J. (199) Rapid emergence of El Tot lnbrb choleræ
¡çsi8tanttoantimicrobialagentsduringfirstsixmonthsofthefourthcholeraepidenic
in li¡nzania Larc*' lz 345.347'
Mharu ES., Mt^ngo ED.E & Msengj A,E. (1gg4) Hoopital outbreaks of cholera t¡ansmitted
througþclogepereon-to-Pefsoûco¡tactl-/,nc¿t.2282.84'.
Migasena S., Pitisuttitha- P., Suntharasamai P', haynra-HongB" Supanaranond lV" Desakorn
v.&BlackRE(19s9)Comparisonofthereactivitiesandimmunogenicitiesof
procholeragenoid and the B subunit of cholera toxin in Thai voluntee$' Infæt' Irunwt'
572 1942-1945.
Miller J.H OnÐ epqimørt hmoteailarganeÍícs. coldspringHarborLaboratory,cold
SpringHa¡bor, N'Y'
Miller vL & Mekeranos JJ. (lgss) A novel suicide vector and it use in construction of
insertionmutations:osmoregulationofoutermembraneproteinsandvinrlencedeter-
minants irl4brio ehobærequires tüR. J- BrctqioL L7Ù' 2575-2583'
Miller cJ., Feachem RG. & Drasar B.S. (1gs5) Cholera epidemiologl in developed and
derrelopingcountries: ne$, thougþs on transmission, seasonality and cont¡oL LanceL
l| ?ßl-2ßZ
Moch T, Hoechützky H., Hacker J', K¡öncke KD. & Jann trC (19ST) Isolation and charac-
terizationof thecr+iaþl-ß-1,3-garactoøyr-specificadhe¡in fromfrmbnarBscl.-.íchiacolí
hac NdLAcad- Sci U-SA U: 3462-%6'
Möller G. (1989) Lynphocyte homing Immunolog'Rsv' No' 108' Denmark: Munlsga^fi
International Publishers'
Monsur KA. (1%3) Bacteriotogical diagnoois of cholera under ñeld conditions. fulL 144d,
Htttt. Ory. 2E: 387-389.
172

Morgan R.L,Isaacson RB, Moon H.lv., Brinton cc & To C-C (19S) Immunization of
suckling pigs againet enterctoxigenic Fsenq¡A¡a coli-induced dianhe¿l disease þ
vaccinating dams with purified 987 or K99 pili: protection correlates with pilus homol-
ogt ofvaccine and challangs' Inþa' Imtrunt 222 771-777'
Morona R., Fonest 8., Attridge S.R., Tackett CO', Herrington D" Morris G" I-aBrooy J"
I.er¡ineM.M.&RowleyD.(198s)ConstnrctionandvaccineefficacyofEX64S,a
bivalentcholera-typhoidvaccine,Abstracßof24iht'L.S.-IøpøtC.ooperdíveMedicøl
Sciøtcehogrøtt, ChoteraandrelatedDíarrheatDúæasesPøteL bkyo, Japan' tr 58'
Morris J.G. & Btack RE (1gs5) Chorera a¡d other vibriooes in the united States. New Fng
I. Med StLz 343350.
Morris J.G., lvest G.R, Holck s.E, Blake P.r{., Echeverria P'D' & Karaulnik M' (1982)
Cholera amongrefugees in Rangsit, Thaitanô I. hrer|DÍs. 145: l3l.l3/¡,
Moeley lv.H. (1969) The role ef immunity in cholera A re'ien' of epidemiological and
serological studies' Tæ' FeP' BìoL n'Ied' 2ilz EI CAL
Moeley lv.fl, Benenson A-S. & Barui R. (1963) A serological suwey for cholera antibodies
in rural Fast pakistaû L The distribution of antibody in the cont¡ol population of a
cholera-vaccine ñeld-trial area and the reLation of antibody tit¡e to the pattern of
endemic cholera Bu& Wtd- HblL Otg' 3t¡' 377-33/.'
Mooley lV.H., McCormack w.lvL, Ahmed A-, Chowdhury A'KM'A" & Banri R'lL (1f)ó9)
Report of the 19ól.-1967 choleravaccine field trial in nrral Fast Pakistaû' II' ResultB
of the serological sufveys in the study population - the relationship of case rate to
antibody titre and an estimate of the inapparent infection ntewil\l/ibrio cholsæ' BulL
Wd- Httlv Org. 4u 187 -t97'
Mooley w.H., Woodward lv.E, Aaz KlvLA" Rahman A'S'M'M., Chowdhury A"trLM-A"
Ahmed A, & Feeþ J.c: (1g0) The lf}68-lf}6g cholera-vaccine field trial in rural East
173

pakistan. Effectiveness of monovalent ogawa and Inaba vaccines and a puriñed Inaba
antrgeq with comparative results of serological "¡¿
¿¡imal protection tests' I' Infed
Dú'c t;Zlz S1-S9.
Mootov K.E, Friedtander M. & Blohell G. (19&4) The receptor for transepithelial transport
of IgA and lgþl contains multiple immunogtobulin-like domains Naare' 3lE: 3743'
Mukeriee s. (1%3) The bacteriophage+usceptibility test in difre¡entiating whrío cholcræ
a¡ôVihriøEl Tor. ButL WId- HÃfrL Org' 8z 333-336'
Muir LL, Stnrgnell R.,{. & Davies J.K (19SS) hoæins that aPPear to be associatedwith pili
u.Ne/ssvlír ' hfect'hwruut- 56;1743-1747'
Nag 8., Moon H.w. & Isaacson R.E (|g77> Colonization of porcine intestine þ
enterotoxi genrc4sc)øichíacoJi: eelection of piliated forms in rauo, adhesion of piliated
forms to epithelial ce¡s h vítro, and incidence of a pilus antþn among porcine
enteropathogenic.&ch qict,¡4 colL Infect. Immun t6z 344-352
Nas¡ 8., Moon H.lv., Isaacson R.B, To CC & Brinton cc. (198) Immunization of suckling
pig against enteric enterotoxþe úc Eschøichi¿coJi infection þ vaccinating d¡ms with
purified PtlL hfea- Innun 2t 269-274'
Nakashima Y., Napiorkonski P., Schafer D.E & Konigsberg W'H' (1976) Primary structure
oftheBsubunitofcholeraentefotoxin.FEB9l.ú.6ft¡2n5.n8.
Nakasone¡.¿þtvanagalvL (19SS) Piliof genusWbri¿r.' Abwøasof24thU'S'-IqmCnopaa-
tíve Mdicat Scíance hogron, Choba oú rebted Dúû,,,e41 Dúseaæs Pøe]^ ftþo,
JaPan. P- 49.
NalinD.R.(1976)Choleracopepodsandchitinaæ'I-øtú'2:958'
Nalin D.R. & Cash R.^d Qnq oral therapy for cholera l!¡z clplcra E&' D' Barua & w'
Burrows.I.ondon:}V.B.Saunders&Co.pp.256.2ß1.
t74

Natin D.R., Daya V, Reid.au, Ladne M.M. & Cisneroe L (199) Adsorption and growth of
Víbrio clwlqaeon chitin' Infect' Im¡nun' 25: 768-770'
Nelson E T, Clemens J.D. & Finkelstein R..{. $nq wbrûo crøtsøeadherence and coloniza-
tion in experimental cholera: Elect¡on microscopic studies' Infecf' bnmwt' 14" 5n'
547
Neoh s.H. & Rowley D. (rg0) The antigens of wbrio chorqae involved in tre vibriocidal
actionofantibodyandcomplementl.InfeclDís.L212505-513.
NeohS.H.&RowþD.(1972)hotectionofinfantmiceagainstcholerabyantibodiesto
threeantigensolVibrbcholqael.InfecLDi's.1:2624147.
Nishibuchi M., Seidler RJ., Rollins D.M. & Joneph J'W' (1983) Vibrio factors ca.use rapid
fluid accumuliation in sucklingmice. Infect l¡nmwt 4û 1083-1091'
Noriki H. QnT Evaluation of toxoid field trial in the Philippines' Proc' of the 12th It hnr
Cholera(U.S..IapøttØopøativeMedic4lscíqt.cehqramLpp.32.3|.
Normarks, Båg¡ M., Göransson M., lindbergEP" I-'nd B.' Norgren M' & u-hlin B'E (1986)
Genetics and biogenesis of Eschabhiacoll adhesins. lliz Microbial lectins md ag-
glutinbt. Ed D. Mirelman, Nerr,York: John Wiley & Sons. pp. 113.143.
Norris H.T & Maino G. (1g6s) on the role of the ilear epitherium in the pathogenesis of
e4perimental cholera' Am' J' PatlL 532 ?ß3-279'
o,Brien AD., Chen M.F-, Holmes R.K, Kaper J. & Levine lYLM' (1984) Environmental and
human isolates oî. V cholcræ arrd V pøahaanotytiars produce a Shigella dysøtaiae I
(Shþ)-like cytotoxin' Iarcet' Lz 77 -78'
ohtomoN. (1972) safetyandpoæncytestsofchoreratoxoidl-otllinanimalsandvolunteers'
Proc. l2tlt Ioinf C-onføencc ø cholera' (tlS-Iapørt Cæpøative Medicgl Sciøce ho
gûn),úøPan PP' ?ß6-?96'
175

oppenheimer J.R, Ahmad IvLG., Huq d, Haque KA" Alam A.KM'A" Aziz Klvf's.' Ali s'
& Haque A,s.M. (1978) Limnological shrdies of three ponds in Dacca, Bangladesh' J'
Físhqics t l-?3-
o,shaugþnessy w.B. (1S31-1832) Proposal of a neu',method of treating the blue epidemic
cholera by the iniection of higþþ-o:rygenized salts into the venous syst'en LowÍ' lz
ffi-371.
orren IL L ( 1933) And noç, pathophysiolog of M cells - good neu's and bad ¡eu's from Peyer's
patches GaÑroantøotogt t5: 468-470'
onen RL, Pierce N.E, Apple R.T & Cray l\¡.CJr. (198ó) M cell transport of wbrío cholcræ
from the intestinal lumen into Peyer's patches: a mechanism for antigen sampling and
for microbial transepithelial migration. J- Infæt'Dís' 153: 1108-1118'
orren RL & Jones LL Qna) Epithelial cell specialization within humnn Peyer's patches:
an ultrastnrcûrral study o¡¡þsrinal þhoid follicles. Gøstoqúqobgt' lÍ¡t' 189-m3'
peterson IcM. & Mekalanos JJ. (198s) cha¡acterization of the lftrío e-Iølqæ ToxR regulon:
identification of novel ggnes invotved in intestinal colonizatiot Infe6' Immwt 562
28n 28Ð.
Peterson J.Sl., LoSpalluto JJ. & Finkebtein ILA" çnzìt Localization of cholera toxin hvövo'
I- Infect Dis- L2'62 617{128'
pierce N.E, (1g3) Differential inhibitory effects of cholera toxoids and gangliooide on the
enreroroxins of l4brb clpbræ atd&srhqúchia cotí. I. W- Me¿ 1i]7: 10(Þ-l0B.
Pierce N.E, Banwell J.G., Sack RB, Mitra R.C & Mondal A" (1970) Magnitude and duration
of antitoxic response to human infection with, l4brìo dtolqae. I- hfeá' D¡s' L2lz
s31-S35.
,t
È
*,
H
l,
176

Pierce N.E, Cray Jr. lv.c, trbper J.B. & Mekalanaos JJ. (1938) Determinants of im-
nunogenicity and mechanisms of protection by virulent and mutantwb¡o cholqæ'
Infee. Immun- 562 L42-148'
pierce N.E, Cray w.cJr. & Sircar BIC (1g7g) Induction of a mucosal antitoxin response and
its role in immunity to e:perimental canine cholera Infecl bnmun 21: 185-193'
Pierce N.E, Greenougþ \il.[ & Carpenter CCJ. (19?1) Wb¡o dpbæ enterotoxin and i¡g
modesof action. BacíRq' 35" t-13'
Pohlner J., Meyer TE, Jalaiakumari M.B. & M'nning P'A" (19Sóa) Nucleotide sequence of ii
ompv, the ggne for a m4 ot wbrio cttobæ oulr'I membrane protein' Moler' Gq^ GeneÍ'
Ð5: 494-500.
Pohlner J., Meyer TE & Manning P.,4,. (19S6b) Serological properties and processing in
E&hsíchío coti Kl2of OmpV fusion proteins of'Vîbrb úobæ' Mokt' Gq' Genet
ã)5: 501-506.
Pollitzer R. (1959) Chole¡a Monograph series No. 431\¡.H.O. Gener¡a'
Rabbanic.H. (198ó) Cholera Clh.Gastoqttqol LSz 507-528'
Rahanan M.M., A^ZKNLS., patwari y. & Munshi lvf"fl. QnÐ Diarrheal mortality in two
Baniladeshi vill4ges with and without community based oral rehydration therapy'
I-øcet.2: 8'09S12
Rappaport RS., Bonde G., McCann T, Rubin BA & Tint H. $nÐ Development of a
purified cholera toxoid" II. heparation of a stable, antþenic toxoid þ reaction of
purified toxinwith glutaraldehyde. Infect Immun u' 304-317.
Redmond J.!V. (1g7g) The structure of the o-antigenic side chain of the lipopoþsaccharide
olVíbrb clptqæ5698 (tnab"). B¡oehct L Bioplryx Acta. 5E/,: 346-352^
Re€dLJ.&MuenchH.A, (1g3S) Asimplemethodof estimatingfiftypercentendpoinß. Am
I. HYg. 272 493497.
I'
177

i¡I'i
Þ-
Richardson S.H. & Noftle trL,{. (1970) Purification and properties of permeability fac- i
tor/choleraenterotoxinfromcomplexandsyntheticmedia J.InfectDi^* L2lt s73-s?9' It
Riipkema s.G.T, Jansen .lV.H., Gielen H. & Guinée P.A'M. (1937) tmmunogtobulins in bile i
and serum of the rabbit associated with protection after Wbtúo eholsæinfection and i
vaccination Miøobiat Pælngan' 3: 365'375'
Rogers R.C, Cuffe R.G.CJ., Cossins Y.M, Murphy D.M. & Bourke A"TC (1980) The
eueensliand cholera incide¡t or.l977.II. The epidemiological investigation. BulI wld'
Hlth. Org. 5E: 6ó5-6ó9.
Rosenbergc.E (1%2) The chobøyeøt. chicago: university of chicago Press'
RutterJ.M.&JonesG.l\t(1973)ProtectionaginstentericdiseasecawdJ-byMærúchiacoli
- a model forvaccinationwith avirulence determinant? Nøture. A2z 531-532
Rutter J.M., Jones G.W., Brou¡n G.TH., Burrows M.R & Luther P'D' (1976) Antibacterial
activity in coloetrum and milk associated u'ith protection of piglets apinst enteric
disease caused by KSS-poe iwe Eç,clwíchia coli. hfecl Imtnt¿n t:3z 667 47 6'
Sack G.H., Pierce N.E, Henness€y KN., Mitra RC, Sack R'B' & Mazumdâf D'N'G ' (1972)
Gast¡ic acidity in cholera and non-cholera diarrhoea BulL Wld- HttlL otg' 472 3l'36'
sak^zaki R- & Shimada T (1y77') Serovars o|.Víbrb chobae. Jøp. J. Med sci BíoL 3lÌ
n9-28L
Salmaso S., Greco D., Bonfigtio 8., Castellani-Pastoris M., de Felip G', Bracciotti A"' Sitzia G"
congn 4., Piu G., Angion G., Barra L ,TamprenA" & Baine w'B' (1980) Recurrence
of pelec5rpod-aseociated cholera in Safdini¿ I'ûucd' 2t ll?A-llZ7'
samadi A,R, shahid N., Eusof 4., Yunus M., Huq lvLl, I(han M'U.' Rahman Às'M'lvf' &
Faruque A.SG. (1933) Classical lürio choleræ biotype displaces El Tor in
Banþdesh- I-anca t 805Sü/'
t78

sattler J., lViegandt H., Staerk J., Kranz T, Ronnebergef HJ., Schmidôerger R & zilgH'
(1975) Studies of the subunit stmcürre of choleragen' hropeot I BiochøL 572
309-316.
sanyal s.G, Alam K, Neogi P.KB., Huq M.I. & Al-Mahmud KA" (1983) A nerü cholera toxin'
I-Øcet 1z 1337.
Satterwhite TIC, Dupont H.L, Evans D.G. & Evans D'J' (1978) Role of Eschsichía colí
colonization factor antigen in acute diarrhea. Løtcet 2: 181-1M'
Schneider D.R. & parker c.D. (1g7g). Isolation and characterization of proteasedeficient
mutants o|'lhbrb cholqae t' InfæLDir lÍlt: 143-151'
schrank G.D. & verwey s/.E, (1976) Distribution of cholera orsaniôms in e:rperimeDltzlvíbrúo
chobæinfections: proposedmechanisms of pathogeneeis and antibacterial immunity'
Infect. hnmw- L3:' 195â3'
sciortino cv & Finkelstein RA. (1983) wbío clpbae exPresses iron-regulated outer
membraneproteinsinvívo.InfecLImmun42:99o-96.
Sears S.D., Richardson IG, YoungC, Parker CD. & Levine M'M' (1984) Evaluation of the
human immune responõe to outer memb¡ane proteins of lftrb cholqae Infecl
Irntnutt. 4: 439 444'
shahidN.S.,SamediA.R.,KhanM.U.&HuqM.I.(1984)Classicalvs'ElTorcholera:a
prospective famity study of a concurrent ouöreak I' Díû'Ds' R¿s' 2t 73-78'
Shandera \ryJC, Hefkin 8., Martin D.L, Taylor J'P', Maserang D'R' Wells J'G" Kelbt M.'
GhandhitrL,KaperJ.B.,[-eeJ.V&BlakePá'(1983)Persisænceofcholerainthe
United States. Am- I- Tlop' Med IIyg' 322 8l2Al7'
simon R, hiefer v & pührer,A. (1gs3) A broad hoet range mobilization qrctem lot h viw
genetic engineering transposon mutagenesis in gram negative bacteria Biotechnologt'
i
tz 784-791.
t79

singþton EL, Attwell Rl\t, Jang M.S. & Coì*'elt RR. (1982å) Influence of salinity and
nutrient concentration on survival and grcwrth oLl1brio dtobøein aquatic nüc¡ocosrns'
AppL Fnvirm- M¡qobíoL 43: 1081-1085'
Sing[eton F.L, Attwell R.W., Jangi M.S. & Cotwetl R.R' (1982b) Effects of temperature and
salinity ot vîbrío c]ølerægrosrth. Appt. hvírot. Miqobíol 44: 1O47-1058'
SminiaT, Janse E M. & lVilde¡s M.M. (1932) Antigen-trappingcells in Peyer',s patches of the
¡alu Scand I- immunol 16: 481-485'
smith H.L (199) Serotypingof non*,holeravibrios. L Clh. Micrabbl 10: 85-90'
smith H.W. & linggood M-a (19?1) Observations on tle pathogenic properties of the K88,
Hþ and Ent plasmids of Escl, erúchíacoli with particular reference to porcine diarrhea
I. Med Mísobiol 4; 467485'
sneller M.C. & Suobe¡ lv. (1936) M celts and host defense. I- InfæLDís' 154: 737:741'
Snou, J. (1S5Ð On ihc mofu of cønmtniætíot of choba M editbn London: John
Churchill
snyder J.D., Allega D.T, I-evine M.M., Craig J.P., Feeley J.c, Delvitt !v'E & Blake P'A'
(1931) Serologic shrdies of naturalþ acquired infectio¡ with'Víbrb e]øl¿rø serogrcuP
Ol in the Uniæd States. I- Infed'Dfu' 143: 182-187'
sprra \v.lvl, Sack RB. & Froehlich J.L (1981) Simple adult rabbit model lor víbrb choleræ
and enterotoxigenic Bc]1øi1hi4 coli dizrùea hfecr" hntnun- 32t 739-747'
Sæele F-J., chaicumpa rv. & Ronley D. (rg74) Isolation and biological properries of three
classes of rabbit antibodies to víbrb chotsæ. J. InfæLDts 13lh 93-103'
Stevenson G., Leavesly D.L, I-agnado C:4., Heuzenroeder M'W' & Mandng P'A (1985)
Purification of the 25 kDavibrb cholerae maior outer-membrane protein a¡d the
molecular cloningof is gene: ompv. European I. B¡ochstt 1¿ft: 385-390'
180

stocker B.A,D. (1g4g) Measurements of rate of mutation of flagellar antigenic phase in
Sabnotætla tphímühnt' J' Hye 472 398413'
st¡eeterP.R, BergF^L, Rouse B.TN., Batgalz.eR.E & Butcher E C (1983) Atissue-specific
endothelial cell molecule involved in þmphocyte homing Natute 33tz 4l'46'
svennerholm A.-M. (19?5) Experimental studies on cholera immunization. [v. The antibody
respo$e to formalized Wbrb cholqaeand purified endotoxinwith special reference to
protectivecapacity.Int.Archs.AtløgtApplhnmunoL4g:434452
sennerhorm A._M. (1gs0) The nahrre of protective immunity in cholera - r¡ crtorqa and
relared diûïtteAs E(ls. o. ouchterlony & J. Holmgren 43rd Nobel symposium'
Stockholm 1978. Basel: S' I(arger' pp' 171-184'
Sennerholm ,{ -M., Gothefors L, Sack D.,{., Bardhan P.IC & Holmgren J. ( 19S4a) I-ocal and
qntemic antibody responses and immunological memory in humans ¿fte¡ immunization
withcholeraBsubunitþdifferentroutes.Bulll44d'HlûLOrg'6'229Ú-918'
ñennerholm ^A.-M. & Holmgren J. $nq Synergistic protective effect in rabbits of im-
munization $t\wfuìo clplcræ lipopoþaccharide and toxinltoxoid" htfeã. I¡rvnun
t3z 735-740.
ñen¡erholm A--M., Jertborn MT Gothefors L, Iørim A"M'M'M', Sack D'À & Holmgren J'
(1gs4b) Mucosal antitoxic and antibacterial immunity after cholera disease and after
immunization with a combined B ¡ubunit-whole cell vaccine' l' hfeá' Dis' L49 :'
884S93.
TabtiengR., Wattanasri s., Echeverria P., seriwatana J., Bodhidaüâ L, Chatkaeomorakot '4"
& Rowe B. (lgsg) An epidemi c oi.víbrío chotsæ El lbr Inaba resistant to several
antibiotics with a coniugative Group C plasmid coding for Type II dihydrofolate
reductase in Thailan ð" AnL L TIop' Med Hyg' 41: 680ó86'
181

Tamaru T & Brou,n lV.R. (1985) IgA antibodies in rat bile inhibit cholera toxin-i¡duced
secretion in ilealtoops ¡n síûL Imrunologt 55: 579-583.
Þuxe R.V., Holmberg s.D., Dodin,{., wells J.G. & Blake P.A" (19SS) Epidemic cholera in
Mali: higþmortalityandmultiplercutesof transmissioninafamine arca. Epídem'Inf'
LW n9-2ß9.
Tøylor R.K, Miller vL, Furlong D.B. & Mekalanoo JJ. (19SÐ Use olphoA gene fusions to
identi$ a pilus colonization factor coordinateþ regulatedwith cholera toxin. hoc- NaL
Acod. Scû^ U-SA- Uz 7ß33-?ß37'
Taylor R., Shau, c, Peterson trL, Spears P & Mekalianoo J. (19SSa) Safe, live wbrto choleræ
vaccines? Vrccine 6: 151-154'
?ylor R.K Shaw cE, Sun D., Rhine J..A,., Peterson KM. & Mekalanos JJ' (1988b)
Molecular mechanisms or.Wbrío cholqaecolo¡ization and its prerrention Absrøcß of
24ih (,L1.-Iapø C-oopødive Mdicat Scíaæe hogrøn, Choba ød relatcd DiÛheal
Dúseaæs Pøtel Toþo, Japan' pp' 50-51'
Teppema J.S., Guinée PA.M., Ibrahim 4.4., Paques M. & Ruitenberg E J' (1937) In víw
adherence and colonization of Vibrb cløbæ strains that differ in hemagglutinating
activity and motility- Infecr.Immun 532 ÐI3-2IAZ
Tou¡bin H., staehelin T & Gordon J. (1979) Electrophoretic transfer of proteins from
poþacrylamide gels to nitrocelluloee sheets: procedure and some applicatio¡x hoc
NøtLAcd. ScLU-SA- 76: 435O-4354'
lamont E.C. & Boolego J.l\t (1935) Pilus vaccines' Væche' 3: 3-10'
Tveedy J.lvL, park Rlvá. & Hodgkiss w. (1%s) Evidence for the pfesence of fimbriae (pili)
on Vibrio species. J. GqL M¡súíoL 5t: B5'2M'
t82

uhlin B.E , Baga M., Goransson M., LindbergEP., Lund 8., Norgren M' & Normarks (198Ð
Genes determining adhesin formation in uropathogenic Escherichia coli Cun Top'
Microbíol hntnunol 1:1:&' 163-177'
{-[iiye ,4.., Nakatomi M., utsunomiya A., Mitsui K, sogne s., Iwanaga M' & Kobari K (1968)
Eryerimental cholera in mice. I. First report on the oral infection. Trop. med 1û
65-71.
underdown BJ. & Schiff M.J. (1936) Immunogtobulin A: strategic defense initiative at the
mucosal surface. AnnRev' bnmunoL 4z 3E94t7'
Vaerman J.-P., Deriik-I angendriês A, Rits M. & Delacroix D' (198Ð Neutralization of
cholera toxin þ ratbile secretory IgA antibodtç. Intrutnolo&t 54: 601603'
Van Heyningen s. (197) Choleratoxin. B¡oL Rct. 52: 509-549.
van Hgningen w.E, carpenter ccJ., Pierce N.E & Greenougþ w'B' (191) Deactivation
of cholera toxin þ gangtiooide. I.Infea. Dis- fi24z 415418.
vougþt llt (1s93) A Chrytø on cholqa ro, by reafurc. History swPø^+ hevøtiot and
Tleafinqt of thc Di,æase Phitadelphia: The EA. Davis company.
wanke cA, & Guerrant RL (1gS7) Small borvel colonization alone is a cauee of diarrhea
btfect. Imtru¿n. 552 197A-197;6'
watanabe Y. & Verwey lV.E (1%5) The preparation and properties of a purified mouse-
protective lþpoþsaccharide from the Ogawa subtype of the El Torvarie$ olVîbrío
chobæ. ln: hocetÅkgs ol fric Cholerø Raseodt Symposùnr' U'S' Deparment of
Health, Education and \trelfare publication SPHS 1328. \Vashington: Government
Printing Office. PP. 253-259'
lVata¡abe Y., Verwey W.H,, Guckian J.C, rili[iams H.R., Phillips PE & Rocha S'S' (1J]69)
Some of the properties of mouse protective antþns derived from Vibrio cholerae- T¿r'
ReP. BioL Med 272 n5-29E'
183

Watren R.H., Morgan EM, SonÉhla Y.N., Vanikiati B. & PhilliPs RA. (1959) Water a¡d
electrolyte sürdies in cholera r. cl¡r\^ hvest 3E: 1879-1889.
lVeissma¡ J.8., Dewiü W.E, Thompson J., Muchnick CN., Portnoy B.L, Feeley J.C &
Gangarosa F^J. (1974) A case of cholera in Texag ln3. Artt- I. Epífuníol. 10(}
487498.
lVestphal O. & Jann K (lf)65) Bacterial lipopolysaccharides. Extraction with phenol-water
andfurtherapplicationsof theprocedure. Metlødsíncøbolryùatechanístry- 5: 83-91.
Wilders lvf.M., Sminia T, Plesch B.E, Drexhage H..au, Welter¡reden F-E & Meuwissen S.G.
(19g3) Large mononuclear la-pooitiveveiled cells in Peyer's patchee. U. Localization
in ratPeyer's patches Intnwolog- 4Ez 461467'
Wolf J.L & Rye W.A" (1934) The membranous epithelial (M) cell and the mucooal immune
systen. Rev. Med 35: 95-112
\iloodruff JJ. & Clarke LM. (1987) Specific cell-adhesion mechanisms determining mi8fa-
tion pathways of recirculatinglymphocytes . Ann Rev. Immunol 5z Ðt'22'.
kid.y N.G. (1%S) The lattice spacingof cryrstalline catalase as an internal st^ndard of length
in elect¡on microscopy. J- Illtastrtrct- Res- A: 454'4&'
yamamoto T, tr(amano T, Uchimura M., Iwaaaga M. & Yokota T (19SS) Vbrb ùobae Ol
adherence to villi and þmphoid follicle epitheliumz in vìtro model using formalin-
treated human small intestine and correlation between adherence and cell-associated
hemaggtutinin lerrels' Infec| Dnmun 56z 32Al-3250'
yamamoto trC, Ichinoee Y., Nakasone N., Þnabe lvL, Nagahama M., Sakurai J. & Iwanaga M.
(1936) Identity of hemoþins produced by Vcholøæ non-Ol al'd V choløae o1,
biotype El Tor. Infeú- hnmwu SLz 9/7-93t'
184

yamemoto T & yokotaT (l9SS) Electron microecopicstuÛ ot.Víbrío e]øløaeOl adherence
to the mucus coat andvillus surface in the human small intestine. hfæt Imtrutn 56:'
n$-n5e.
yancey RJ., Willis D.L & Berry LJ. (199) Flagella-induced immunity against experimental
cholera in adult rabbits. Infæt bnmun 252 ffi'28'
ychiyama y. & Brou¡n W.R (1937) Specific antibodies to cholera toxin in rabbit milk are
pmotective egainst Wfu¡o choleræ-tnduced intc¡tinat secretion. Iwntnolog- 61: 543-
547.
185