universidade federal do cearÁ - ufccompreensão de minhas dificuldades. À profa. neuza márcia que...
TRANSCRIPT

UNIVERSIDADE FEDERAL DO CEARÁ
FACULDADE DE MEDICINA
DEPARTAMENTO DE FISIOLOGIA E FARMACOLOGIA
CURSO DE PÓS-GRADUAÇÃO EM FARMACOLOGIA
MODELO EXPERIMENTAL DE DISFUNÇÃO TEMPOROMANDIBULAR EM RATOS
KÁTIA DO NASCIMENTO GOMES
FORTALEZA-CE 2005

ii
KÁTIA DO NASCIMENTO GOMES
MODELO EXPERIMENTAL DE DISFUNÇÃO
TEMPOROMANDIBULAR EM RATOS
Fortaleza-Ce
2005

iii
KÁTIA DO NASCIMENTO GOMES
MODELO EXPERIMENTAL DE DISFUNÇÃO TEMPOROMANDIBULAR EM RATOS.
Dissertação apresentada à coordenação do Programa de Pós-graduação em Farmacologia da Universidade Federal do Ceará – UFC para a obtenção do título de Mestre.
Orientador: Prof. Dr. Carlos Maurício de Castro Costa
_________________________________
Fortaleza-Ce 2005

iv
G614m Gomes, Kátia do Nascimento.
Modelo experimental de disfunção temporomandibular em ratos/ Kátia
do Nascimento Gomes. – Fortaleza, 2005.
196f. : it.
Orientador : Prof. Dr. Carlos Mauricio de Castro Costa.
Dissertação (Mestrado) - Universidade Federal do Ceará, Faculdade de
Medicina.
1. Adjuvante de Freund. 2. Articulação temporomandibular. 3.
Medição da dor. 4. Dor orofacial.
CDD
616.0472

v
KÁTIA DO NASCIMENTO GOMES
MODELO EXPERIMENTAL DE DISFUNÇÃO TEMPOROMANDIBULAR EM RATOS.
Aprovada em: 27/06/2005
BANCA EXAMINADORA
Prof. Dr. Carlos Maurício de Castro Costa Orientador da Dissertação
Profa. Dra. Francisca Cléa Florenço de Sousa Departamento de Fisiologia e Farmacologia – UFC
Prof. Dr. Otoni Cardoso do Vale Departamento de Medicina Clínica– UFC

vi
Aos meus queridos filhos
Isabela, Mariana e Samuel, minha vida só tem sentido porque tenho vocês ao meu lado.

vii
Altissimo, onnipotente, bon Signore,
tue so' le laude, la gloria e l' honore et onne benedictione.
Ad te solo , Altissimo, se confano
e nullu homo ène dignu te mentovare.
Laudato sie, mi' Signore, cum tucte le tue criature,
spetialmente messer lo frate sole,
lo qual' è iorno, et allumini noi per lui
Et ellu è bellu e radiante cum grande splendore:
de te, Altissimo, porta significatione.
Laudato si', mi' Signore, per sora luna e le stelle:
in celu l' ài formate clarite e pretiose e belle.
Laudato si', mi' Signore, per frate vento
e per aere nubilo e sereno et onne tempo,
per lo quale a le tue creature dài sostentamento.
Laudato si', mi' Signore, per sor' acqua,
la quale è molto utile et humile et pretiosa et casta.
São Francisco de Assis (Cânticos das criaturas)

viii
Agradecimentos
A Deus, que ilumina e guia meus passos nesta caminhada... A Ele, toda honra e toda a glória.
Ao prof. Dr. Carlos Maurício de Castro Costa, orientador deste trabalho. Agradeço por todos as valiosos conhecimentos que me passou durante esse tempo em estive em seu laboratório. Toda a minha admiração e gratidão pela paciência, críticas, conselhos e a confiança em mim depositada. Ao prof. Dr Otoni e profa. Dra. Francisca Cléa Florenço de Sousa, que prontamente aceitaram participar da banca de Exame de Qualificação e Dissertação. Por todas as orientações que enriqueceram este trabalho, mas principalmente pela compreensão de minhas dificuldades. À profa. Neuza Márcia que incentivou e contribuiu com seus amplos conhecimentos em nosso trabalho. À profa. Régia Maria Evangelista pela realização da análise histopatológica necessária para enriquecer nosso trabalho. Meu sincero agradecimento aos amigos e colaboradores Carlos Câmara, Carlos Henrique, Débora, Fabiana, Hellíada e Neiberg pela compreensão e disponibilidade para a realização deste trabalho.
Agradeço especialmente `a minha amiga Ana Maria Monteiro pelo exemplo de pessoa que é e por ter as palavras certas para me ajudar.
Aos meus amigos Flávio Damasceno e Rivelilson Mendes, grandes estudantes da pós-graduação que me ajudaram com seus conhecimentos. À Virgínia pelo espírito de ânimo e partilha, além do interesse em ajudar oferecido a todos nós da pós-graduação. Ao Sílvio pelas orientações sobre informática e por ter colaborado com sua paciência e companhia durante os experimentos laboratoriais. À técnica de laboratório Vilani, Vandinha e Jaqueline que dispuseram material e disponibilizaram suas imprescindíveis habilidades laboratoriais. O meu sincero agradecimento à coordenação do Curso de Pós-graduação (Sílvia e Aura) e aos meus colegas de pós-graduação, em especial minhas colegas de profissão Ingrid Chaves e Raquel Carvalho.
À minha mãe, Maria do Nascimento, pelo aprendizado diário que tenho pelo convívio com ela e por ensinar-me o valor da persistência diante das dificuldades.

ix
Ao meu pai, Luiz Alves (in memoriam) que infelizmente não pode estar
presente para compartilhar desta vitória, mas sei que está feliz com essa conquista. Ao meu marido, José Gomes pela inestimável compreensão e companheirismo dedicados nos momentos em que mais precisei. Meu imenso agradecimento à minha irmã Karla Nascimento, que me ajudou a ingressar no Mestrado e que contribuiu com seus conhecimentos para conclusão do curso. Muito obrigada pelo apoio e conhecimentos transmitidos. Essa conquista é nossa!!! Aos meus familiares, especialmente Fernanda Amorim, Ehricka Gomes e Luiza de Marilac pelo apoio incondicional que me foi dedicado. Ao CNPq pela bolsa que permitiu a realização dessa dissertação de Mestrado e pelo apoio à pesquisa.

x
SUMÁRIO
RESUMO XVI
ABSTRACT XVIII
ABREVIATURAS XX
INTRODUÇÃO 1
I - REVISÃO DA LITERATURA 3
1.1- Conceito e epidemiologia 3
1.2- Manifestações clinicas 6
1.2.1- Dor aguda e crônica 6
1.2.2- Dor nociceptiva somática e visceral e dor neuropática 7
1.2.3- Fases da dor 8
2- Dor orofacial e as disfunções temporomandibulares 9
2.1- Tipos de disfunção e etiologia 10
2.2- Mecanismos implicados na fisiopatologia da dor relacionada a DTM. 12
3- Processamento da dor 14
3.1- Nociceptores 14
3.2- Fisiologia da nocicepção inflamatória 15
3.3-. Estruturas neurais relacionadas 17
4-. Mecanismo neural e caminhos da dor craniofacial 20
5- Contribuição da neuroplasticidade central na patologia da dor 23

xi
5.1- Hiperalgesia secundária 23
5.2- Dor referida e hiperalgesia 24
5.3- Sensibilização central 25
6- Mecanismos de modulação da dor 31
7- Modelos animais de dor orofacial e comportamentos associados. 34
8- Controle farmacológico da dor orofaciall 38
8.1 – Drogas antiinflamatórias não esteroidais (DAINES) 38
8.1.1 – Mecanismo de ação 39
8.1.2 – Efeitos indesejáveis 42
8.2 – Anestésicos locais 43
8.2.1 – Mecanismo de ação 43
8.2.2 – Efeitos indesejáveis 45
8.3 – Analgésicos opióides 46
8.3.1 – Mecanismo de ação 46
8.3.2 – Efeitos indesejáveis 48
9 - Relevância e justificativa 49
10 – Objetivos 50

xii
II – MATERIAL E MÉTODOS 51
2.1 – Animais 51
2.2 – Drogas e soluções 51
2.3 – Material e equipamentos 52
2.4 – Protocolo experimental 52
2.4.1 – Observações comportamentais 55
2.4.2 – Grupos experimentais 57
2.4.3 – Testes farmacológicos 58
2.5 – Análise histológica 62
2.6 – Análise estatística 62
III – RESULTADOS 63
3.1 – Curva temporal dos comportamentos 63
3.2 – Sumário das alterações comportamentais 96
3.3 – Análise Comparativa da interferência dos grupos controles sobre
os comportamentos avaliados 97
3.4 – Abordagem farmacológica dos comportamentos 102
3.5 - Sumário dos resultados farmacológicos 115
3.6 – Peso corporal dos animais 118
3.7 – Análise histológica 121
IV – DISCUSSÃO 123

xiii
V – CONSIDERAÇÕES FINAIS 142
VI – CONCLUSÃO 143
VII – REFERÊNCIAS BIBLIOGRÁFICAS 145

xiv
LISTA DE FIGURAS Figura 1 : Efeito da solução salina fisiológica 0,9%, formalina 2,5% e CFA sobre o comportamento snnifing (andar, cheirar e explorar)------------------------------------------64 Figura 2 : Efeito da solução salina fisiológica 0,9%, formalina 2,5% e CFA sobre o comportamento rearing (empinar-se)-------------------------------------------------------------67 Figura 3 : Efeito da solução salina fisiológica 0,9%, formalina 2,5% e CFA sobre o comportamento chewing like (movimento de rotação da mandíbula ou mastigação-70 Figura 4 : Efeito da solução salina fisiológica 0,9%, formalina 2,5% e CFA sobre o comportamento climbing (subir escadas)--------------------------------------------------------73 Figura 5 : Efeito da solução salina fisiológica 0,9%, formalina 2,5% e CFA sobre o comportamento freezing (congelar-se)-----------------------------------------------------------76 Figura 6 : Efeito da solução salina fisiológica 0,9%, formalina 2,5% e CFA sobre o comportamento sleeping (dormir /descansar)--------------------------------------------------79 Figura 7 : Efeito da solução salina fisiológica 0,9%, formalina 2,5% e CFA sobre o comportamento grooming (limpar-se)------------------------------------------------------------82 Figura 8 : Efeito da solução salina fisiológica 0,9%, formalina 2,5% e CFA sobre o comportamento licking (lamber-se)---------------------------------------------------------------85 Figura 9 : Efeito da solução salina fisiológica 0,9%, formalina 2,5% e CFA sobre o comportamento scratching (coçar-se)------------------------------------------------------------88 Figura 10 : Efeito da solução salina fisiológica 0,9%, formalina 2,5% e CFA sobre o comportamento biting (morder-se)----------------------------------------------------------------91 Figura 11 : Efeito da solução salina fisiológica 0,9%, formalina 2,5% e CFA sobre o comportamento head fliches (sacudir a cabeça)----------------------------------------------94 Figura 12: Análise comparativa da interferência dos grupos controle sobre o comportamento chewing like.-----------------------------------------------------------------------98 Figura 13 : Análise comparativa da interferência dos grupos controle sobre o comportamento res/sleeping.-----------------------------------------------------------------------99 Figura 14 : Análise comparativa da interferência dos grupos controle sobre o comportamento grooming.-------------------------------------------------------------------------100 Figura 15 : Análise comparativa da interferência dos grupos controle sobre o comportamento freezing.---------------------------------------------------------------------------101 Figura 16 : Efeito da morfina, indometacina e lidocaína sobre o comportamento chewing like no 3°dia após a injeção de CFA-------------------------------------------------104

xv
Figura 17 : Efeito da morfina, indometacina e lidocaína sobre o comportamento sleeping no 3°dia após a injeção de CFA-----------------------------------------------------106 Figura 18 : Efeito da morfina, indometacina e lidocaína sobre o comportamento freezing no 4°dia após a injeção de CFA------------------------------------------------------108 Figura 19 : Efeito da morfina, indometacina e lidocaína sobre o comportamento grooming no 4°dia após a injeção de CFA----------------------------------------------------110 Figura 20 : Efeito da morfina, indometacina e lidocaína sobre o comportamento grooming no 1°dia após a injeção de Formalina 2,5%.-------------------------------------112 Figura 21 : Efeito da morfina, indometacina e lidocaína sobre o comportamento scratching no 1°dia após a injeção de Formalina 2,5%.-----------------------------------114 Figura 22 : Efeito da solução fisiológica 0,9%, CFA e formalina 2,5% sobre o peso corporal dos animais--------------------------------------------------------------------------------119 Figura 23 : Avaliação histopatológica da região articular e periarticular da ATM de ratos ----------------------------------------------------------------------------------------------------122

xvi
LISTA DE TABELAS Tabela 1: Efeito da solução salina fisiológica 0,9%, formalina 2,5% e CFA sobre o comportamento snnifing (andar, cheirar e explorar)------------------------------------------65 Tabela 2: Efeito da solução salina fisiológica 0,9%, formalina 2,5% e CFA sobre o comportamento rearing (empinar-se)-------------------------------------------------------------68 Tabela 3: Efeito da solução salina fisiológica 0,9%, formalina 2,5% e CFA sobre o comportamento chewing like (movimento de rotação da mandíbula ou mastigação)71 Tabela 4: Efeito da solução salina fisiológica 0,9%, formalina 2,5% e CFA sobre o comportamento climbing (subir escadas)--------------------------------------------------------74 Tabela 5: Efeito da solução salina fisiológica 0,9%, formalina 2,5% e CFA sobre o comportamento freezing (congelar-se)-----------------------------------------------------------77 Tabela 6: Efeito da solução salina fisiológica 0,9%, formalina 2,5% e CFA sobre o comportamento sleeping (dormir /descansar)--------------------------------------------------80 Tabela 7: Efeito da solução salina fisiológica 0,9%, formalina 2,5% e CFA sobre o comportamento grooming (limpar-se)------------------------------------------------------------83 Tabela 8: Efeito da solução salina fisiológica 0,9%, formalina 2,5% e CFA sobre o comportamento licking (lamber-se)---------------------------------------------------------------86 Tabela 9: Efeito da solução salina fisiológica 0,9%, formalina 2,5% e CFA sobre o comportamento scratching (coçar-se)-----------------------------------------------------------89 Tabela 10: Efeito da solução salina fisiológica 0,9%, formalina 2,5% e CFA sobre o comportamento biting (morder-se)----------------------------------------------------------------92 Tabela 11: Efeito da solução salina fisiológica 0,9%, formalina 2,5% e CFA sobre o comportamento head fliches (sacudir a cabeça)----------------------------------------------95 Tabela 12:Efeito da morfina, indometacina e lidocaína sobre os comportamentos alterados pelo CFA----------------------------------------------------------------------------------116 Tabela 13:Efeito da morfina, indometacina e lidocaína sobre os comportamentos alterados pela formalina 2,5%--------------------------------------------------------------------117 Tabela 14 : Efeito da solução fisiológica 0,9%, CFA e formalina 2,5% sobre o peso corporal dos animais--------------------------------------------------------------------------------120
LISTA DE QUADROS
Quadro 1- protocolo experimental----------------------------------------------------------------54

xvii
RESUMO

xviii
Modelo experimental de disfunção temporomandibular em ratos. KÁTIA DO NASCIMENTO GOMES. Orientador: Carlos Maurício de Castro-Costa. Dissertação de Mestrado. Pós-graduação em Farmacologia. Departamento de Fisiologia e Farmacologia, UFC, 2005. Estudos de comportamentos espontâneos, atividade farmacológica sobre os comportamentos alterados, avaliação histopatológica e alteração de peso corporal foram realizados em ratos (180-300g), através da administração de Adjuvante completo de Freund (CFA) na região da articulação temporomandibular de ratos com a finalidade de investigar se um estímulo inflamatório crônico promove alteração de comportamento espontâneo comparado a estímulo agudo ou subagudo como a formalina 2,5%. Os animais após a injeção de CFA, formalina 2,5% e solução salina fisiológica 0,9% foram observados diariamente durante 30 minutos por 7 dias onde foram sacrificados para a remoção da região articular e periarticular para avaliação de alteração de parâmetros inflamatórios frente a um estímulo pelo CFA e pela formalina 2,5% comparando com o grupo controle. As principais alterações promovidas pelo CFA ocorreram no tempo de execução de alguns comportamentos como movimento de rotação da mandíbula ou mastigação (chewing like), tempo de descanso e sono (rest/sleeping), aparente congelamento (freezing) e auto cuidado como coçar a face com a pata anterior (grooming ou rubbing), enquanto que no grupo de animais da formalina 2,5%, os comportamentos alterados formam grooming e scratching. Os estudos comportamentais demonstraram que o CFA possui uma atividade tempo dependente sobre comportamentos, pois essas alterações ocorreram a partir do 2º dia e persistiu até o 7º dia após a administração do CFA, ao passo que a formalina apresentou uma atividade principal no 1º dia de injeção. A administração de drogas analgésicas (Morfina 4mg/Kg; Indometacina, 5mg/Kg e Lidocaína com fenilefrina) demonstrou que o comportamento chewing like foi sensível à ação da morfina e indometacina, o sleeping foi aumentado por administração de lidocaína e indometacina, o tempo de execução do comportamento grooming foi reduzido por atividade da morfina e lidocaína e o comportamento freezing não foi alterado por ação de nenhuma das drogas administradas. Comparativamente, os comportamentos alterados pela atividade nociceptiva da formalina 2,5% na região orofacial dos animais foram grooming e scratching. A morfina e a lidocaína foram efetivas em reduzir o grooming e o scratching desenvolvidos pela formalina. A atividade de drogas classicamente analgésicas testadas sobre esses comportamentos pode caracterizá-los como nociceptivos. Foi observado também que o peso corporal dos animais não foi alterado pela indução inflamatória do CFA. Comparando sessões histológicas da região articular e periarticular da ATM dos ratos, observamos que no grupo do CFA há presença de infiltrado inflamatório linfocitário intenso comparado ao grupo da formalina, além de alterações degenerativas e pequeno infiltrado inflamatório periarticular. Nós sugerimos que o fenômeno da neuroplasticidade está relacionado às alterações comportamentais desenvolvidas pelo CFA e a identificação de comportamentos relacionados a sua atividade inflamatória pode fornecer subsídios para a avaliação de novos e melhores agentes terapêuticos para o tratamento da dor orofacial como também avaliação dos benefícios dos tratamentos já existentes, em consonância com os avanços nos estudos de dores experimentais.

xix
ABSTRACT

xx
Experimental model of temporomandibular dysfunction in rats. KÁTIA DO NASCIMENTO GOMES. Promoter: Carlos Maurício de Castro Costa. Master of Science Dissertation. Post-Graduation in Pharmacology. Department of Physiology and Pharmacology, UFC, 2005. Studies on rats spontaneous behaviors and pharmacological effects of drugs on them, as well as their body weight and histopathological analysis of their joints were carried out to investigate the effect of chronic inflammation of the temporomandibular articulation induced by the administration of Complete Freund Adjuvant (CFA) compared with the acute and subacute stimulus of 2.5% formalin. After CFA, 2.5% formalin and 0.9% saline injection, the animals were daily observed during 30 minutes for 7 days; thereafter, they were killed for removal of the articular and periarticular region for evaluation of inflammatory changes induced by CFA and formalin compared with control rats. The main behavioral changes induced by CFA concerned chewing-like, rest/sleeping, freezing, grooming or rubbing whereas those induced by formalin were grooming and scratching. The CFA-induced behavioral changes were time-dependent from the 2nd up to the 7th day, while the formalin-induced changes occurred in the 1st day only. For pharmacological testing, morphine (4 mg/kg), indomethacin (5 mg/kg) and lidocaine with phenilephrine were used. In the CFA rats, morphine reduced significantly chewing-like and grooming, indomethacin decreased grooming and increased sleeping. Freezing was not changed by any drugs. In the formalin rats, grooming and scratching towards the orofacial region were increased and those behaviors were significantly decreased by morphine and formalin. The effects of the drugs on the different behaviors shown above show their nociceptive meaning. Moreover, the body weight of the animals was not changed. In both CFA and formalin groups, the histological analysis evidenced degenerative and small periarticular inflammatory infiltration. Neuroplasticity changes may be related to the CFA induction and the nociceptive meaning of the behaviors may be useful for experimental therapeutic studies.

xxi
ABREVIATURAS
5-HT- 5 hidroxitriptamina
AMP c- Monofosfato Cíclico de Adenosina
AMPA- A Metil Aspartato
ATM – Articulação Temporomandibular
CFA- Adjuvante Completo de Freund
COX- Ciclooxigenase
CPME- Corno Posterior da Medula Espinhal
DAINES- Drogas Antiinflamatórias não esteroidais
DNIC- Controle Inibitório Nocivo Difuso
DTM – Disfunção Temporomandibular
EMG - Eletromiografia
EPM – Erro Padrão da Média
i.m.- intramuscular
i.t. - intratecal
IASP –Associação Internacional para o Estudo da Dor
ICHD – Sociedade Internacional de Cefaléia
IL 1- Interleucina 1
IL-6- Interleucina 6
IL-8- Interleucina 8
KCl- Cloreto de Potássio
LDP – Limiar de Dor a Pressão
LTM- Neurônios Mecanossensitivo de Baixo Limiar
MDH- Corno Dorsal bulbar
NaCL- Cloreto de Sódio
Ne - Noradrenalina
NGF- Fator de Crescimento do Nervo
NKA – Neuroquinina A
NKB – Neuroquinina B
NMDA- N-Metil D-Aspartato
NRM- Núcleo da Rafe Mediana
NS- Neurônio Nociceptivo Específico

xxii
PAG- Região Periaquedutal Mesencefálica
PGE – Prostaglandina E
PGs- Prostaglandinas
CGRP- Peptídeo Relacionado ao Gene da Calcitonina
RNA- Acido Ribonucléico
RVM- Região Rostral Ventro Bulbar
SBED – Sociedade Brasileira para o Estudo da Dor
SNC – Sistema Nervoso Central
SNNVS- Sistema Nervoso Neurovegetativo Simpático
SNP – Sistema Nervoso Periférico
SP – Substância P
TNF – Fator de Necrose Tumoral
USA – Estados Unidos da América
v.o. via oral
VGSCs- Canais de Sódio Sensíveis a Voltagem
VLO- Região Ventro Lateral Orbital
WDR- Neurônio de Variação Dinâmica Ampla

1
I- INTRODUÇÃO
A Disfunção Temporomandibular (DTM) corresponde a ausência de
normalidade nas funções do aparelho mastigatório (Maciel,1998). Caracteriza-se por
um conjunto de sinais e sintomas, como dores na região temporomandibular (ATM),
nos músculos da mastigação e em regiões da cabeça e do pescoço.
A DTM apresenta etiologia multifatorial e normalmente envolve uma
abordagem interdisciplinar para seu tratamento (Rizzatti-Barbosa e colaboradores,
2000). Os fatores mais comumente relacionados são: oclusão, fatores emocionais,
ausências dentárias, mastigação unilateral, hábitos orais inadequados. Geralmente,
apenas um fator isolado não é desencadeante da disfunção, mas sim a associação
entre eles (Okeson, 1992).
Essa patologia orofacial é um mal que atinge parte da população e causa
impacto na qualidade de vida devido a dor provocada por ela. Além disso, a
população normalmente não recebe informações sobre o que é DTM e como tratá-la,
pois médicos e cirurgiões dentistas não investigam a causa e tratam somente o
sintoma, concorrendo assim para a não resolução definitiva do problema.
Além disso, a DTM é uma patologia pobremente estudada, devido em parte, a
limitados modelos experimentais com agente inflamatórios crônicos desenvolvidos
para estudar dor nessa região, bem como testar a eficácia farmacológica de drogas
que poderiam ser usadas para tratar esse distúrbio orofacial nociceptivo.
Mas o que se observa na literatura são modelos experimentais avaliando
comportamentos espontâneos envolvendo dor orofacial de origem inflamatória
aguda ou subaguda como a formalina (Roveroni e colaboradores, 2001), óleo de
mostarda (Ro e colaboradores, 2003) e a capsacina (Pelissier e colaboradores,
2002).
Por isso, considerando a necessidade de um modelo experimental avaliando
comportamentos espontâneos após um estímulo inflamatório crônico,
desenvolvemos um modelo experimental administrando o Adjuvante completo de

2
Freund (CFA) na região da ATM de ratos. Para isso, abordamos duas grandes
questões:
1. Identificação de comportamentos espontâneos alterados pela indução
inflamatória e nociceptiva na região orofacial de ratos;
2. Avaliação da interrelação entre ação analgésica de fármacos (DAINE,
agonista opióde e anestésico local) e comportamentos espontâneos sugestivos de
dor.

3
I- REVISÃO DA LITERATURA
1.1- CONCEITO E EPIDEMIOLOGIA
Os conceitos e as justificativas sobre as razões da ocorrência da dor sofreram
marcantes modificações ao longo do tempo em função das aquisições científicas e
dos aspectos culturais de cada povo.
Durante o século XX, acumularam-se conhecimentos básicos e critérios
característicos neuroanatômicos e neurofisiológicos do homem e dos animais, assim
como a compreensão da fisiologia da dor.
O comitê de taxonomia da Internacional Association for the Study of pain
(IASP), conceitua a dor como “experiência sensorial e emocional desagradável que é
associada ou descrita em termos de lesões teciduais”. Dessa forma parte da dor é
compreendida subjetivamente. Cada indivíduo utiliza a palavra dor de acordo com o
aprendizado frente a experiências prévias. É uma sensação desagradável localizada
em uma parte do corpo, constitui adicionalmente uma experiência emocional
(Merskey e colaboradores, 1979). A dor tem valor biológico fundamental de alertar o
indivíduo sobre a ocorrência de alguma forma de lesão orgânica instalada ou em
vias de se instalar.
Embora a dor seja reconhecida mais como uma experiência do que uma
sensação, ela realmente tem uma dimensão sensorial que registra a natureza do
estímulo iniciador, incluindo sua qualidade, intensidade, localização e duração. Mas
pode ter uma dimensão cognitiva, emocional e motivacional. Em humanos, a
experiência dolorosa ativamente envolve processos neocorticais relacionados com o
reconhecimento e a interpretação das conseqüências dessa experiência, exercendo
assim grande influência na dor e no sofrimento gerado por ela (Melzack, 1979).
A dor pode ser resultante de um ou mais fatores interno e/ou externos, sendo
por isto o seu diagnóstico mais complexo (SBED, 2004).

4
Alguns fatores podem influenciar a dor: biológicos (lesões nos tecidos,
condições físicas e efeito de medicações); sociais (suporte social, relação familiar,
influências culturais); psicológicos (comportamento, tipo de personalidade, grau de
conhecimento) (SBED, 2004).
Por tratar-se de uma experiência individual e subjetiva, a dor pode apresentar
diferentes características em função da: intensidade, qualidade, localização,
duração, etiologia, natureza e freqüência da dor (SBED, 2004).
A ocorrência de dor é crescente, talvez em decorrência dos novos hábitos de
vida, das modificações do meio ambiente, do prolongamento da sobrevida dos
indivíduos em geral e dos doentes com afecções clínicas naturalmente fatais, do
decréscimo da tolerância ao sofrimento do homem moderno e da aplicação dos
novos conceitos que traduzem seu significado (IASP, 1986). Von Korff e
colaboradores (1988) em pesquisa epidemiológica destacam como mais frequentes
as dores recorrentes abdominais, lombares, cranianas e na articulação
temporomandibular.
Bergman e colaboradores (2001) relatam que os estudos epidemiológicos têm
demonstrado o aumento das queixas de dores crônicas com a idade e que mais de
80% de relatos de pacientes com 80 anos de idade mostram dor significante.
Aproximadamente metade desses relatos é devida a desordens
musculoesqueléticas (Andersson e colaboradores,1999).
Bingefours e Isacson (2004), demonstram que a cefaléia e a dor
musculoesquelética são um dos maiores problemas da saúde pública e proporções
substanciais da população relatam de experiências com a dor afetando o trabalho,
vida diária e social. Segundo os autores, estudos epidemiológicos têm
consistentemente mostrado que a prevalência da maioria das condições dolorosas é
mais alta nas mulheres que nos homens, associada a problemas psicossomáticos e
psiquiátricos.
Bonica (1980) estima que 5 a 7 milhões de americanos sofrem de dor crônica
na face ou região bucal e entre 25 a 45% relatam que em alguma época da vida
sofreram dor nessa região (Lipton e colaboradores,1993; Von Korff, 1996).

5
Foi realizado um estudo do impacto da dor na vida de indivíduos portadores
de disfunção da articulação temporomandibular através de uma versão brasileira do
questionário McGill de dor que avalia questões específicas como: prejuízo social;
atividades da vida diária, percepção do outro, tolerância à dor, sensação de estar
doente e insatisfação com a vida (Oliveira e colaboradores, 2003). Os autores
concluíram que a dor da DTM tem um impacto negativo na qualidade de vida
afetando atividades do trabalho (59,64%), da escola (59,09%), no sono (68,18%) e
no apetite/alimentação (63,64%).
É importante salientar que a DTM atinge também indivíduos em idade pré-
escolar. Em estudo de prevalência de desordens em uma população de estudantes
com idade entre 9 e 12 anos revelam que há necessidade de examinação pré-
escolar na ordem de prevenir o progresso de estágios tardios dessa patologia que
pode ocorrer na infância. Nesse estudo se verificou que 11,7% dos indivíduos
apresentaram disfunção da articulação temporomandibular, 1,7% descreveram
limitação da abertura bucal e 5,4% têm um desvio ao realizar movimentos
mandibulares (Fariaby e Mohammadi, 2005).

6
1.2- MANIFESTAÇÕES CLÍNICAS
1.2.1- DOR AGUDA E CRÔNICA
De acordo com Braun e Braun (2004), a dor aguda se diferencia da dor
crônica não somente pelo aspecto temporal. Existem também diferenças quando
consideramos os mecanismos fisiológicos e, especialmente, os fisiopatológicos
envolvidos na cronificação das sensações dolorosas.
Os autores reportam que a dor aguda é uma consequência sensorial imediata
da ativação do sistema nociceptivo. É um sinal e alarme desencadeado pelos
sistemas protetores do indivíduo e geralmente possui uma relação entre lesão e dor;
manifesta-se como um padrão de comportamento. Nesse tipo de dor a remissão é
espontânea e coincide com a cura da lesão, sendo considerada mais como sintoma
que uma doença. Além disso, a dor aguda tem uma etiologia bem conhecida sendo
mais fácil o tratamento, tem como conseqüência o aparecimento de alterações
neurovegetativas e neuroendócrinas.
Por sua vez a dor crônica ultrapassa o período de cura do trauma; não possui
qualquer função biológica e freqüentemente desenvolve alterações físicas,
emocionais, sociais e econômicas; não se relaciona a lesão e a dor, levando a
processos adaptativos e é considerada uma doença de etiologia não conhecida, pois
pode advir de processos crônicos, como também de lesão ou disfunção do sistema
periférico (SNP) ou sistema nervoso central (SNC) e por isso pode levar a uma
dificuldade no tratamento.

7
1.2.2- DOR NOCICEPTIVA SOMÁTICA E VISCERAL E DOR NEUROPÁTICA
Braun e Braun (2004), destacam através de dados da literatura que a dor
nociceptiva é a forma de dor que surge em todos os indivíduos, como conseqüência
da aplicação de estímulos que produzem dano ou lesão nos órgãos somáticos ou
viscerais. Originam-se da ativação dos nociceptores, os quais poderão se encontrar
em estado normal ou sensibilizados.
A dor somática tem origem na pele, músculos, articulações, ligamentos e
ossos. Trata-se de uma dor bem localizada, circunscrita à área lesada e caracteriza-
se por sensações claras e precisas.
A dor visceral surge mais freqüentemente após enfermidades. É um sintoma
comum na maioria das síndromes dolorosas agudas e crônicas de interesse clínico.
É vaga, mal localizada e estende-se além do órgão lesionado. Vem acompanhada
de intensas reações reflexas motoras e vegetativas e pode ser referida em regiões
distantes da víscera que a originou.
A dor neuropática apresenta sintomatologias e muitas etiologias, incluindo
câncer, artrite, doenças vasculares, traumatismos, neuropatia diabética e alcoólica,
neuromas, causalgia (queimação), dor do membro fantasma, síndrome talâmica
(Kupers e colaboradores,1992). É circundada por um número variado de síndromes
de dor e tem em comum a presença de hiperalgesia e/ou alodinia, dor espontânea e
parestesia (Bennett, 1994), sugerindo que existem mecanismos fundamentais que
são comuns nas diferentes síndromes (Woolf e Manion, 1999).

8
1.2.3- FASES DA DOR
Cervero (1989), analisando os diversos estados das sensações dolorosas, os
dividiu em três fases diferentes:
A primeira fase é aquela conseqüente a um estímulo nocivo rápido. Sinaliza a
presença de uma lesão em potencial e é uma sensação necessária para
sobrevivência e o bem estar do indivíduo. É um mecanismo neurofisiológico simples
de transmissão central para o tálamo e o córtex cerebral, com possibilidade de
modulação sináptica dos núcleos centrais e periféricos.
A dor na segunda fase expressa a capacidade do sistema nociceptivo normal
de responder a estímulos prolongados, os quais tenham produzido lesão tissular e
iniciado um processo inflamatório, promovendo liberação de substâncias excitatórias
locais e pode levar a um envolvimento central.
Na terceira fase ocorrem estados dolorosos anormais, os quais se originam
freqüentemente de lesões de nervos periféricos ou SNC. A característica
fundamental é uma ausência da relação entre lesão e dor, que se manifesta como
dores espontâneas, provocadas por estímulos inócuos (alodínia) ou como respostas
exageradas a estímulos nocivos de baixa intensidade (hiperalgesia).

9
2- DOR OROFACIAL E AS DISFUNÇÕES TEMPOROMANDIBULARES
Dworkins e colaboradores, (1990) e Okeson & De Kanter, (1996) definem que
as disfunções temporomandibulares (DTMs) ou a ausência de normalidade das
funções do aparelho mastigatório representam um termo coletivo incluindo várias
condições patológicas relacionado a complexos sinais e sintomas envolvendo a
musculatura mastigatória, a articulação temporomandibular (ATM) e outras
estruturas orofaciais. Outros termos usados são síndrome de Costen (Costen,1934),
disfunção (Shore, 1959) e desordens craniomandibulares (McNeill e colaboradores,
1980).
Embora tenham sido sugeridos métodos para a padronização dos exames
diagnósticos em DTM (Le Resche e colaboradores, 1992) a fim de que se possam
avaliar e comparar os resultados de diferentes estudos com maior fidelidade, o que
se percebe na literatura é uma falta de padrões de definição, de métodos de
investigação e de apresentação de resultados. Esses fatores podem justificar a
grande variação de prevalência de sinais e sintomas da DTM encontrada em
diferentes estudos.
Cerca de 70% da população têm pelo menos um sinal de DTM leve ou
moderada. Entre os sintomas que a caracterizam podemos citar: otalgia, zumbidos,
dor nos olhos, na face e no pescoço, mudanças oclusais, mialgias, dor e ruído nas
articulações, cefaléias, entre outros. Das pessoas que apresentam um ou mais
sinais de disfunção temporomandibular, somente 5% procuram o tratamento. A
maioria dos que procuram tratamento são mulheres em proporção de 1:4 (Salonen &
Hellden, 1990).
Rocha (2002) e Salonen & Hellden (1990) referem que embora a DTM possa
ocorrer em qualquer idade, o paciente a apresenta mais comumente na fase adulta
jovem, entre os 20 e 40 anos.
Sarlani e Greenspan (2003), revelam que as discrepâncias entre os estudos
experimentais em humanos podem ser devido a numerosos critérios metodológicos,
incluindo regras de seleção de pacientes, idade, sexo, tamanho da amostra, tipo de

10
estimulação para diagnóstico diferencial, medidas de resposta à dor e sítio do corpo
em que foi feito o teste de sensibilidade.
Tomacheski e colaboradores (2004) verificaram através de um modelo de
questionário, que pouco mais da metade dos entrevistados eram portadores de DTM
leve ou moderada (50,56%), sendo a incidência maior em mulheres (67,76%) e as
queixas principais relatadas foram tensão emocional e o hábito de apertar ou ranger
os dentes.
2.1- TIPOS DE DISFUNÇÃO E ETIOLOGIA
A recente classificação das cefaléias realizada pela sociedade Internacional de
Cefaléias (ICHD, 2004), relatou que as disfunções temporomandibulares ocorrem
frequentemente nos músculos mastigatórios (dor miofacial; mialgia localizada nos
músculos mastigatórios) e nas articulações temporomandibulares (deslocamento do
disco e disfunções inflamatórias que incluem as capsulites, sinovites e as
osteoartrites).
Lobbezoo e colaboradores (2004) classificam as desordens
temporomandibulares em três grupos:
-Dor miofascial que é a forma mais comum de DTM, onde há um desconforto
ou dor nos músculos da maxila e pescoço;
-Desordens internas na articulação que ocorrem quando há deslocamento da
maxila, disco articular ou lesão no côndilo;
-Doença degenerativa da articulação que são DTMs associadas a
osteoartrites.
Para Silva (1991), as DTMs podem ser subdivididas de acordo com :
- Alterações internas como artrites, anquilose, subluxação, luxação.

11
-Alterações musculares que são mialgias (podendo causar trismo ou limitação
da abertura bucal), dor a palpação (trigger points) associada ao bruxismo (hábito de
ranger os dentes) ou apertamento dental.
-Disquinesia que é a incoordenação do movimento da mandibular com ou
sem tremor, podendo ocorrer sem episódios de dor, quando associados a problemas
neurológicos.
-Fatores etiológicos como fator hereditário (síndromes congênitas) e locais
(trauma na ATM, traumatismo craniano, causas dentárias iatrogênicas, invasão do
espaço de repouso fisiológico, desarmonia oclusal).
-Fatores gerais como fisiológicos (recente enfermidade sistêmica) e
psicológicos (estresse).
Há uma multiplicidade de fatores que contribuem para a etiologia da DTM,
dentre as quais se podem incluir: má oclusão, estresse emocional e hábitos
parafuncionais (Okeson, 2000), ausência de alguns elementos dentários e fatores
sistêmicos (Meira, 2002). Entretanto, Okeson, (1992) já havia relatado que muitas
vezes esses fatores estão associados e o que determina a instalação da doença é
tolerância fisiológica e resistência estrutural do indivíduo.
Dentre as manifestações clínicas das DTMs a mais comum é a presença de
dor e sensibilidade dos músculos da mastigação e da articulação temporomandibular
(Okeson, 1998).
Meira (2002) em pesquisa com pacientes relata que os sinais e sintomas
associados a DTM são dores intra-articulares, espasmo muscular, dor reflexa, dor à
abertura e ao fechamento da mandíbula, dor irradiada na área temporal,
masseteriana e infra-orbital, dor de cabeça crônica, dor ou zumbido no ouvido, dor
irradiada ao pescoço, sensação de tamponamento do ouvido, xerostomia (boca
seca), entre outros.
Em estudo de prevalência de disfunção temporomandibular em crianças com
dentição mista, Tuerlings e Limme (2004) demonstraram elevada ocorrência de
sensibilidade à palpação muscular (80,9%). Ruídos na articulação

12
temporomandibular estavam presentes em 35% das crianças, sendo maior parte
meninas.
Fujita e colaboradores (2003), relatam que 82,8% de pacientes com disfunção
temporomandibular têm hábitos parafuncionais como mastigação unilateral,
bruxismo, anormalidade na postura (desvios da coluna cervical), etc.
Além disso, um dado a ser considerado é a grande flutuação dos sinais e
sintomas de DTM com o tempo. São freqüentes os períodos de exacerbação e
remissão de sintomas e isto deve ser cosiderado na indicação do tipo de tratamento.
(Bruno, 2004).
Em resumo, DTMs são desordens das articulações e músculos da região
bucofacial e cervical (DTM miogênica), que têm entre seus principais sintomas a dor
facial, dor no ouvido, cefaléia, neuralgias, ruídos, etc. A etiologia da DTM é
multifatorial incluindo a oclusão bucal, fatores neuromusculares e psicogênicos,. mas
o determina a instalação da doença é a tolerância fisiológica e a resistência
estrutural do indivíduo.
2.2- MECANISMOS IMPLICADOS NA FISIOPATOLOGIA DA DOR RELACIONADA A DTM.
Várias linhas de estudo têm demonstrado que pacientes com DTM miogênica
exibiam significantemente baixo limiar na dor a pressão (LDP) dos músculos
mastigatórios que indivíduos saudáveis (Maixner e colaboradores, 1998). A
sensibilização de aferentes nociceptivos primários da área afetada, bem como a
hiperexcitabilidade do sistema trigeminal são a causa da hiperalgesia. Vários fatores
têm sido implicados para o início da dor muscular, como trauma tecidual,
sobrecarga, inflamação e isquemia, podendo sensibilizar esses aferentes da dor
(Mense, 1993). O aumento na atividade de nociceptores periféricos pode levar ao
desenvolvimento da sensibilização central, a saber, hiperexcitabilidade e expansão
do campo receptivo em neurônios de segunda ordem do sistema nervoso central

13
(SNC), contribuindo para a amplificação e persistência de sinais nociceptivos que
eventualmente evocam a dor (Sessle, 2000). O envolvimento da hiperexcitabiliade
de neurônios nociceptivos do tronco cerebral no aumento da sensibilidade de
pacientes com DTM à estimulação de regiões afetadas é reforçado por falha do
bloqueio anestésico de zonas desencadeadoras em alterar LDP nesses pacientes
(McMillan e Blasberg, 1994).
Além disso, pacientes com DTM relatam grande sensibilidade a estímulos
aplicados além da área indicada como dolorosa clinicamente, indicando uma
generalizada ou alta hiperexcitabilidade em neurônios nociceptivos de segunda
ordem no SNC. Como a sensibilização central tem sido implicada como importante
para o entendimento do mecanismo patofisiológico de diferentes tipos de dor crônica
(Dubner,1991), e pode contribuir para dor em DTM. Reid e colaboradores (1994)
demonstraram que pacientes com DTM têm uma significante redução de LDPs dos
músculos mastigatórios, como também exibem similar baixo LDPs em outros sítios
dolorosos. Estes achados foram interpretados como reforço da presença de dor
mediada centralmente. Além disso, pacientes que sofrem de DTM e que possuem
um componente miogênico, ou seja, que tem origem em algum músculo orofacial,
apresentavam uma somação temporal a estímulos térmicos repetidos aplicados na
palma da mão. (Maixner e colaboradores, 1998). Essa somação temporal é o
aumento da percepção da intensidade da dor quando estímulos nocivos, a uma
intensidade constante, são oferecidos a uma freqüência alta suficiente, refletindo
uma transitória up-regulation de neurônios nociceptivos do corno dorsal (Price e
colaboradores, 1977).
A amplificação da atividade neuronal em neurônios de segunda ordem do
corno dorsal da medula espinhal pode ser relacionada ao desequilíbrio entre
modulação descendente inibitória e caminhos facilitatórios que regulam a informação
nociceptiva processada. (Ren e Dubner, 2002). Um decréscimo na eficácia de
sistemas inibitórios endógenos tem sido implicado na fisiopatologia da dor crônica
em DTM (Maixner e colaboradores, 1995).

14
3- PROCESSAMENTO DA DOR
3.1- NOCICEPTORES
Segundo Coderre e colaboradores (1992), o sistema somatosensorial
normalmente funciona como um mecanismo de alerta individual ou dano potencial
do tecido. O fenômeno sensitivo doloroso é a transformação dos estímulos
ambientais em potenciais de ação que, das fibras nervosas periféricas, são
transferidos para o sistema nervoso central (SNC). Os receptores nociceptivos são
representados pelas terminações nervosas livres das fibras mielínicas finas A delta e
amielínicas C (Besson e Perl, 1969). Os nociceptores quimiomecanotérmicos do tipo
C evocam sensação de calor e os nociceptores relacionados com as fibras A delta
evocam sensação em pontada. Os nociceptores que veiculam informações pelas
vias A delta são classificados como tipo 1 e 2. Os do tipo 1 apresentam limiar
elevado para os estímulos térmicos, aumento progressivo da atividade com o
aumento da temperatura, latência prolongada à estimulação térmica, são sensíveis
aos estímulos mecânicos, veiculam informações pelas fibras do tipo A delta, são
sensilizados pelos estímulos térmicos e localizam-se na pele gabra e pele com
pêlos. Os do tipo 2 apresentam baixo limiar ao estímulo térmico, adaptam-se à
estimulação térmica intensa, apresentam pico de freqüência precoce, geralmente
são mecanoinsensíveis, veiculam informações pelas fibras do tipo A-delta, não são
sensibilizados pelos estímulos térmicos e localizam-se na pele com pêlos. A
estimulação dos aferentes A-Beta pode provocar sensação dolorosa quando há
sensibilização central (Jensen, 1996). A porção terminal do axônio é especializada
na transformação de eventos físicos e químicos que ocorrem nos tecidos em
potenciais de ação interpretados como dolorosos após o seu processamento no
SNC.

15
3.2- FISIOLOGIA DA NOCICEPÇÃO INFLAMATÓRIA
Os receptores nociceptivos são sensibilizados pela ação de substâncias
químicas, denominadas algogênicas, presentes no ambiente tecidual
(Webster,1971). Entre elas, destacam-se acetilcolina, bradicinina, histamina,
serotonina, leucotrieno, substância P(SP), fator de ativação plaquetário, radicais
ácidos, íons potássio, prostaglandinas (PGE2), tromboxana, interleucinas (IL1-beta,
IL6, IL8), fator de necrose tumoral (TNF-alfa), fator de crescimento do nervo (NGF) e
o monofosfato-cíclico de adenosina (AMPc) (Campbell e colaboradores, 1989).
As substâncias algogênicas são liberadas no ambiente tecidual do interior dos
mastócitos e de outros leucócitos, vasos sangüineos e células traumatizadas, sendo
responsáveis pela hiperalgesia mecânica e vasodilatação observada em lesões
traumáticas, inflamatórias e isquêmicas (Campbell e colaboradores, 1989).
A substância P, a neuroquinina A e B (NKA e NKB) e o peptídeo relacionado
ao gene da calcitonina (CGRP), entre outros, são liberados nos tecidos pelas
terminações nervosas dos aferentes nociceptivos e interagem com elementos
celulares envolvidos na inflamação (neutrófilos, linfócitos, plasmócitos, macrófagos,
fibroblastos, células de Schwann, etc) atraindo-os ou ativando-os, gerando
vasodilatação e instalação do processo inflamatório de origem neurogênica
(inflamação neurogênica) (Salt, 1992).
De acordo com Allen e colaboradores (1999) várias evidências suportam a
noção de que a substância P e a neuroquinina A (NKA), que são sintetizadas e
estão contidas em 20-30% dos neurônios do gânglio da raiz dorsal, estão envolvidas
na transmissão de informações nociceptivas das fibras aferentes para o corno
dorsal. Primeiro, a SP/NKA são co-liberadas pelas fibras aferentes primárias de
pequeno diâmetro em estímulos nocivos (Levine e colaboradores, 1993). Segundo,
os neurônios espinhais nociceptivos são excitados com administração de SP e de
NKA (Radhakrishnan e Henry 1991). Terceiro, a destruição dos estoques de SP em
fibras aferentes primárias não mielinizadas com aplicação de capsaicina atenua

16
resposta a estímulos nocivos (Buck e Burks, 1986). Quarto, Aimone e Yaksh (1989)
relataram que analgésicos opióides inibem a liberação de SP na medula espinhal.
Os aferentes primários não são apenas vias de condução de estímulos do
ambiente tecidual para o SNC, de vez que colaterais das fibras nervosas periféricas
parecem participar da regulação da atividade de gânglios da cadeia nervosa
neurovegetativa e da vasoatividade (Campbell e colaboradores, 1989).
Os neurotransmissores, liberados retrogradamente pelas terminações
nervosas em situações anormais, podem, indiretamente, participar da liberação de
vários mediadores químicos nos tecidos, entre eles a histamina (Barolat e
colaboradores, 1987).
As vias periféricas do sistema nervoso neurovegetativo simpático (SNNVS),
frente à ação da bradicinina ou por mecanismos reflexos, libera nos tecidos
noradrenalina (NE) e prostaglandina (PG), que sensibilizam os nociceptores.
Quando ocorrem processos inflamatórios, devido à ação das substâncias
algogênicas, muitos receptores em condições normais, especialmente os
relacionados com as fibras C, são recrutados. Esses receptores são chamados de
nociceptores silenciosos (Cervero, 1994). O recrutamento desses nociceptores
durante estados patológicos deve contribuir para somação temporal e espacial
aumentando o envio de informações das fibras C para o corno dorsal da medula
espinhal. Por exemplo, fibras aferentes primárias em articulações artríticas
desenvolvem tanto atividade espontânea como emitem sinais de resposta a
movimentos articulares antes inofensivos, e a inflamação de vísceras pode aumentar
enormemente sua sensibilidade para estímulos mecânicos inócuos.
A ativação de nociceptores silenciosos pode explicar a natureza distinta dos
estados inflamatórios dolorosos prolongados quando comparado à exposição aguda
a estímulos nocivos passageiros. De fato, o recrutamento de nociceptores
silenciosos promove uma importante contribuição para a sensibilização e as
mudanças de adaptação central que suportam a hiperalgesia primária causada por
inflamação tecidual (Millan, 1999).

17
As alterações inflamatórias e imunológicas estão relacionadas com ativação de
enzimas proteolíticas e colagenolíticas que causam degradação da matriz tecidual. A
liberação tecidual de neuropeptídeos participa da degeneração progressiva das
unidades funcionais estimulando a síntese de agentes inflamatórios (citocinas,
PGE2) e enzimas degradativas (proteases, colagenases). Como resultado direto e
indireto das interações químicas, podem ocorrer alterações secundárias nas
propriedades mecânicas que perpetuam o processo degenerativo (Calvino e
colaboradores, 1987).
3.3-. ESTRUTURAS NEURAIS RELACIONADAS
Cada receptor sensitivo é axônio de neurônio de primeira ordem ou neurônio
aferente primário que transmite o impulso para o SNC (Okeson, 1998).
Três classes de neurônios aferentes nociceptivos fornecem o impulso para o
encéfalo: 1- aferente mecanotérmicos que primariamente são fibras A–delta e
respondem a estímulos mecânicos e térmicos intensos; 2- aferentes polimodais que
são constituídos por fibras C e respondem a estímulos mecânicos, térmicos e
químicos e 3- aferentes mecanorreceptores com limiar alto, onde as fibras A-delta
respondem a estímulos mecânicos intensos (Dubner, 1985).
Estes neurônios de 1ª ordem fazem sinapse com neurônios de segunda
ordem e algumas vezes é chamado de neurônio de transmissão, uma vez que
transferem os impulsos para centros superiores (Okeson, 1998).
Os neurônios de segunda ordem são nomeados de acordo com o tipo de
impulso que ele predominantemente transmite. Os neurônios mecanossensitivos de
baixo limiar (LTM) transmitem informações de propriocepção e pressão e toque leve.
Os neurônios nociceptivos específicos (NS) transmitem exclusivamente impulsos
relacionados ao estímulo nocivo. Os neurônios de variação dinâmica ampla (WDR)
respondem a uma faixa de intensidade de estímulos inofensivos a nocivos (Okeson,
1998).

18
O corno dorsal da medula espinhal está subdividido em camadas ou lâminas.
Estas lâminas são numeradas de acordo com sua profundidade no corno dorsal de I
a VI. Estudos sugerem que os impulsos nociceptivos entrem no corno dorsal pela via
dos neurônios NS e WDR numa área das lâminas I, II e V. Os neurônios que não
transmitem nocicepção parecem estar mais concentrados nas lâminas III e IV
(Guyton, 1991).
Uma vez que os impulsos tenham sido transferidos dos aferentes primários,
os neurônios de segunda ordem transmitem o impulso, cruzando para o lado oposto
da medula espinhal formando o trato espinotalâmico antero-lateral, o qual ascende
para os centros superiores, transmitindo informações sensoriais de dor, calor, frio e
sensações táteis inespecíficas (Guyton ,1991).
O sistema Antero-lateral está dividido em dois tratos: trato neoespinotalâmico
e o trato paleoespinotalâmico. O trato neoespinotalâmico transmite os impulsos
nociceptivos A-delta diretamente para os centros superiores. O trato
paleoespinotalâmico predominantemente transmite a nocicepção das fibras C mais
lentas e percorre muitos outros centros antes de alcançar o encéfalo (Okeson,
1998).
Assim, segundo Kidd e Urban, (2001) tecidos cutâneos e somáticos profundos
são inervados por neurônios aferentes primários que fazem sinapses com neurônios
de segunda ordem no corno dorsal da medula espinhal. Os neurônios aferentes
primários têm três funções em relação a nocicepção: a) Transdução, que é a
propriedade do receptor em detectar ou diferenciar a intensidade do estímulo,
definindo-o como nocivo ou danoso. Receptores para estímulos não dolorosos são
caracterizados pela especificidade para um estímulo e possuem uma ampla faixa de
sensibilidade, onde poderá haver uma amplificação de sinais e rápida adaptação; b)
Condução, que seria a passagem do estímulo sensorial detectado para terminais
periféricos para corno dorsal da medula espinhal. Estudos neurofisiológicos
demonstram que, sob condições normais, as fibras A beta conduzem rapidamente
(velocidade de condução > 30 m -1) e são principalmente associadas com estímulos
não nocivos para receptores especializados. A maioria das fibras de pequeno
diâmetro ou A delta (velocidade de condução de 2,5-30 m s -1) e as fibras C
(velocidade de condução de <2.5 m s-1) presentes nos terminais periféricos

19
respondem a estímulos nocivos (Grigg e colaboradores, 1986). Embora, grande
parte das fibras C mostre uma resposta polimodal, algumas são quase que
exclusivamente quimiosensíveis sob condições normais e não respondem a
estímulos mecânicos ou térmicos. Esses receptores denominados “silenciosos”
foram primeiro descritos em articulações e mais tarde descobertos em outros tecidos
(Schaible e Grubb, 1993); c) A transmissão, em primeira instância, é a resposta da
medula espinhal a estímulos nocivos e são mediados por aminoácidos excitatórios
(glutamato) agindo em receptores AMPA e NMDA. A ativação desses receptores
produz uma seqüência de eventos levando a um incremento na excitabilidade e na
atividade de neurônios do corno dorsal (Dickenson e Sullivan, 1987).

20
4-. MECANISMO NEURAL E CAMINHOS DA DOR CRANIOFACIAL
O impulso sensitivo proveniente da face e da boca é transmitido através do V
nervo craniano, o trigêmio. Os corpos celulares dos neurônios aferentes trigeminais
estão localizados no gânglio trigeminal. Os impulsos transmitidos pelo nervo
trigeminal entram diretamente no tronco encefálico na região da ponte fazendo
sinapse no núcleo espinhal trigeminal. Esta região do tronco encefálico é
estruturalmente semelhante ao corno dorsal da medula espinhal.O complexo núcleo
trigeminal consiste do principal núcleo trigeminal sensitivo, localizado rostralmente e
recebe aferentes periodontais e pulpares; e do trato espinhal do núcleo trigeminal,
localizado mais caudalmente (Okeson, 1998).
O complexo trigeminal do tronco encefálico pode ainda ser subdividido em
núcleo sensorial principal e núcleos do trato espinhal, o qual compreende três
subnúcleos: oral, interpolar e caudal. Baseado em observações anatômicas, clínicas
e eletrofisiologicas, o subnucleo caudal é o principal sítio de informação nociceptiva
no tronco cerebral (Sessle, 1999).
Outro componente do complexo trigeminal do tronco encefálico é o núcleo
motor do V nervo craniano. Esta área do complexo é envolvida na interpretação de
impulsos que demandam respostas motoras da face (Okeson, 1998).
Estudos anatômicos têm revelado que aferentes de pequeno diâmetro levam
informações nociceptivas de vários tecidos craniofaciais terminando
predominantemente no núcleo caudal, lamina I, II, V e VI. Além disso, recentes
estudos histoquímicos da expressão de genes da proteína c-fos tem revelado o
incremento na sua expressão no neurônio caudal após estimulação de tecidos
craniofaciais incluindo pele, articulação temporomandibular, córnea e tecido cérebro
vascular (Hoskin e colaboradores, 1996).
Adicionalmente, estudos eletrofisiológicos têm demonstrado que muitos
neurônios de resposta nociceptiva cutânea orofacial são categorizados como
nociceptivos específicos (NS) e neurônios de variação dinâmica ampla (WRD)
(Sessle, 1999).

21
Esses neurônios (NS) respondem somente a estímulos nocivos (p.e. calor)
aplicados em campo receptivo craniofacial localizado e recebem aferentes de
pequeno diâmetro a partir das fibras A delta e/ou C; neurônios WDR são excitados
tanto por estímulos nocivos como também por não nocivos (p.e. tátil) e podem
receber fibras de grande e pequeno diâmetro (Sessle, 1999).
A maioria desses neurônios, NS e WDR, podem ser excitados por outros tipos
de aferentes periféricos como de tecido cerebrovascular, polpa dentária, ATM ou
tecidos musculares (Sessle, 1999).
O subnúcleo caudal trigeminal é uma extensão rostral do corno dorsal
espinhal cervical superior e também é referida como corno dorsal bulbar (MDH).
Existe uma similaridade anatômica e funcional entre corno dorsal espinhal e bulbar
(Gobel e colaboradores, 1981). Assim como no corno dorsal da medula espinhal, os
neurônios do tipo NS e WDR são referidos como neurônios de segunda ordem no
subnúcleodo caudal trigeminal (Dubner e Bennet, 1983). Broton e Sessle (1988)
relataram que neurônios excitados por estímulos na ATM são classificados como
WDR e NS e o campo receptivo desses neurônios estende-se para amplas áreas de
tecidos faciais e estruturas intraorais (dentes, ligamento periodontal).
Os neurônios trigeminais de segunda ordem projetam-se para o tálamo a
partir das junções sinápticas com aferentes primários no subnúcleo caudal. A
extensiva convergência de aferentes ativando neurônios NS e de WDR no
subnúcleo caudal podem explicar a dificuldade em localizar a dor, bem como
contribuir para dor difusa e referida que envolve a ATM e musculatura facial (Sessle,
1999).
Neurônios de todos os níveis do complexo trigeminal do tronco encefálico
projetam-se para outras regiões incluindo formação reticular e núcleo motor do nervo
craniano; sua conectividade para essas regiões particularmente provém de um
substrato central que compreende respostas autonômicas e reflexos musculares
para o estímulo craniano. Muitos neurônios do complexo trigeminal do tronco
encefálico podem se projetar para a região contralateral do tálamo diretamente ou
através de vias multisinápticas (Sessle, 1999).

22
As regiões do tálamo que são bem conhecidas por receberem e transduzirem
informações somatossensoriais da região craniofacial são o complexo ventrobasal
ou (núcleo ventro posterior) e região média do tálamo. Essas regiões talâmicas
contêm neurônios NS e WDR que têm propriedades que são geralmente
semelhantes aos neurônios do subnúcleo caudal. Neurônios nociceptivos têm
propriedades e conexões com córtex cerebral somatossensorial, indicando um papel
discriminador da dimensão dolorosa. Contrariamente, neurônios nociceptivos do
núcleo medial (núcleo intralaminar, núcleo parafascicular) têm propriedades e
conexões com a região anterior cingular, sugerindo um papel mais afetivo ou
motivacional da dor. Outros neurônios têm sido descritos no córtex somatossensorial
com propriedades que indicam uma função na codificação da localização e
intensidade do estímulo térmico facial ou pulpar dentário. Neurônios também
ocorrem em outras regiões corticais, como o córtex anterior cingular, o qual tem sido
implicado na dimensão afetiva da dor (Sessle, 1996; Iwata, e colaboradores, 1999).

23
5- CONTRIBUIÇÃO DA NEUROPLASTICIDADE CENTRAL NA PATOLOGIA DA DOR
5.1- HIPERALGESIA SECUNDÁRIA
Hardy e colaboradores (1950) propuseram que existem dois tipos de
hiperalgesia: primária e secundária. Hiperalgesia primária envolve incremento da
sensibilidade ao estímulo nocivo no sítio da lesão, enquanto que a hiperalgesia
secundária envolve aumento da sensibilidade extendendo-se além do sítio
lesionado, algumas vezes em locais distantes da lesão.
A hiperalgesia primária é mediada por mecanismos periféricos (inflamação
neurogênica) enquanto que a hiperalgesia secundária é relacionada a uma
hiperatividade central ou sensibilização.
Esta conclusão baseou-se nas observações de Hardy e colaboradores (1950)
de que a hiperalgesia se difundia para região além da área de resposta, o que é
conseqüente à inflamação neurogênica. O desenvolvimento da hiperalgesia após
lesões cutâneas foi prevenido por bloqueio anestésico antes da lesão no sítio
próximo ao nervo periférico.
La Motte e colaboradores (1991) evidenciaram que a hiperalgesia depende,
em parte da sensibilização central. Pois a hiperalgesia induzida por um estímulo
mecânico após injeção intradérmica de capsaicina é mantida mesmo quando se
anestesia essa região após a administração do agente algésico. Entretanto, se a
região da pele é anestesiada antes da injeção com capsaicina, hiperalgesia cutânea
não é desenvolvida.
Torebjörk e colaboradores (1992) têm mostrado que a injeção de capsaicina
causa dor intensa e pode resultar em uma pequena área de hiperalgesia ao calor e
uma ampla área circunvizinha com hiperalgesia a estímulos mecânicos. A
microestimulação intraneural elétrica que normalmente estimula sensações não

24
dolorosas foi acompanhada pela dor quando a sensação foi projetada para essas
áreas de hiperalgesia. Essa hiperalgesia mecânica também foi observada, mesmo
quando se anestesiava o território cutâneo, ou seja, as fibras aferentes cutâneas
daquela área, demonstrando que a capsaicina desenvolve mudanças no
processamento central para aferentes mecanorreceptivos dessas fibras que
normalmente evocam sensações não dolorosas.
Serra e colaboradores (2004) em estudo de microneurografia analisando a
ação hiperalgésica através da injeção intradérmica de capsaicina na pele de
humanos, verificaram que existem dois subtipos de nociceptores C com diferentes
propriedades em seus axônios.
Wasner e colaboradores (2004) em modelo humano utilizando aplicação
tópica de mentol sugerem que esse agente atua sensibilizando os nociceptores C e
as fibras A-delta frio específicas. A hiperalgesia em “pontadas” é devida a
sensibilização central baseada na continuidade da atividade em estimular os
nociceptores sensíveis ao frio.
5.2- DOR REFERIDA E HIPERALGESIA
Dor referida foi inicialmente descrita por Martyn, (1964) como uma condição
em que a dor não é relacionada com alguma lesão local, mas em sítio adjacente ou
distante que ocorre em conseqüência à lesão em tecidos profundos como músculo,
articulação ou vísceras.
Estudos mostram que há uma sensibilização ou a expansão do campo
receptivo do corno dorsal neuronal seguinte a um dano inflamatório em tecidos
viscerais como bexiga urinária (McMahon, 1988), inflamação aguda da articulação
(Doughert e colaboradores, 1992) ou estimulação elétrica de músculos aferentes
(Cook e colaboradores, 1987). A expansão do campo receptivo tem sido observada
em neurônios do tronco cerebral trigeminal após a estimulação química de aferentes
craniofaciais profundos (Hu e colaboradores, 1992). Após lesões inflamatórias na

25
articulação do joelho de ratos, neurônios do corno dorsal da medula espinhal
(Neugebauer e Schaible 1990) e talâmicos (Guilbaud e colaboradores, 1986)
exibiam uma responsividade acentuada não apenas a estimulação mecânica da
junção inflamada, mas também a estimulação dos músculos na região posterior
ipsilateral (mesmo lado) e contralateral (lado contrário) das patas.
5.3- SENSIBILIZAÇÃO CENTRAL
Segundo Coderre e colaboradores, (1992) prolongados distúrbios sensoriais
associada a lesões teciduais (hiperalgesia secundária e dor referida, bem como
alodinia e dor espontânea persistente) resultam da diminuição no limiar do
nociceptor ou incremento na excitabilidade do neurônio no SNC envolvido na
transmissão da dor.
A inflamação pode resultar em uma reorganização dos circuitos neurais
envolvidos na trasmissão nociceptiva, especialmente na inflamação crônica. Alguns
fenômenos podem contribuir para reestruturação central após lesões periféricas, por
exemplo: atividade anormal de aferentes primários, alteração na expressão de
neurotransmissores e receptores, degeneração de neurônios do gânglio da raiz
dorsal e processos regenerativos (Coderre e colaboradores, 1992).
Estudos mostram a sensibilização dos receptores periféricos após estímulos
nocivos (Koltzenburg e colaboradores, 1992), como um mecanismo periférico
responsável pela hiperalgesia que é desenvolvida após a lesão. Entretanto, outra
linha de experimento sugere que a sensibilização está relacionada com alterações
do sistema nervoso central que também contribui significantemente para este
fenômeno.
Outros dados na literatura indicam que lesões após estímulos nocivos, ou
estimulação elétricas de fibras C levam a uma sensibilização de neurônios do corno
dorsal da medula espinhal e em outras áreas no sistema somatosensorial (Coderre e
colaboradores, 1992).

26
Essa sensibilização é evidenciada por um incremento da atividade
espontânea, redução do limiar ou aumento na responsividade para entrada aferente,
mudanças com repetidas estimulações e expansão do campo receptivo periférico em
neurônios do corno dorsal (Coderre e colaboradores, 1992).
Expansão do campo receptivo tem sido observada em neurônios do tronco
encefálico trigeminal com uso de estimulação inflamatória em aferentes craniofaciais
profundos, como a ATM (Yu e colaboradores, 1993).
Várias linhas de evidências sugerem que neuropeptídeos liberados a partir da
ativação das fibras C são envolvidos no início da plasticidade após estímulos
nocivos (Coderre e colaboradores, 1992). Um papel para neuropeptídeos na
nocicepção é evidenciado uma vez que estímulos nocivos ou inflamação periférica
causam a liberação de substância P (SP) (Duggan e colaboradores, 1988),
neuroquinina A (Duggan e colaboradores, 1990), somatostatina (Morton e
colaboradores, 1988), peptídeo relacionado ao gene da calcitonina (CGRP) (Saria e
colaboradores, 1986) e galanina (Morton e Hutchinson 1990) no corno dorsal da
medula espinhal.
Além disso, citocinas como IL-1 beta, IL-6 e TNF alfa, que normalmente estão
presentes em concentração apenas constitutiva, apresentam um aumento após
lesão na medula espinhal (Bethea e colaboradores, 1999) e na dor neuropática (De
Leo e colaboradores, 1997).
Bradicinina tem atraído interesse por possuir a capacidade de reduzir o limiar
de comportamento nociceptivo (Taiwo & Levine, 1988), causar excitação direta de
nociceptores (Manning e colaboradores, 1991), sensibilizar mecanorreceptores
(Neugebauer e colaboradores, 1989) e receptores térmicos (Koltzenburg e
colaboradores, 1992). Incremento na sensitividade à bradicinina foi observado em
receptores em modelos animais de inflamação aguda. (Kirchhoff e colaboradores,
1990).
Há uma interação entre os neuropeptídeos liberados e aminoácidos
excitatórios no processamento nociceptivo central. Neuropeptídeos e aminoácidos
excitatórios são relacionados por localizarem-se no terminal central de neurônios

27
aferentes primários (De Biasi e Rustioni, 1988). Estudos demonstram que a co-
administração de N-metil-D-aspartarto (NMDA) e Substância P incrementa a
excitabilidade de neurônios na medula espinhal, propondo que há um mecanismo
envolvendo a liberação de aminoácidos excitatórios e peptídeos que podem levar a
hiperalgesia secundária. (Willcockson e colaboradores, 1984; Doughert e Willis,
1991).
O glutamato reconhece pelo menos quatro tipos de receptores que recebem
denominação de acordo com o agonista correspondente: os ionotrópicos (AMPA,
KAINATO E NMDA) e os metabotrópicos (Lerma, 1997).
O glutamato tem um papel importante no mecanismo periférico e na
nocicepção da articulação temporomandibular (ATM) (Cairns e colaboradores,1998).
Injeção de irritantes inflamatórios ou óleo de mostarda dentro da cápsula da ATM de
ratos resulta em aumento na atividade eletromiográfica dos músculos digástrico e
masséter (Bakke e colaboradores, 1998). Estudos demonstram que o óleo de
mostarda evoca atividade eletromiográfica que pode ser atenuada por aplicação
periférica de antagonista de receptor NMDA (Yu e colaboradores,1996).Outros
trabalhos descrevem que injeções de glutamato na região da ATM de ratos induzem
a uma similar estimulação de músculos mandibulares através de ativação de
receptores periféricos NMDA e não-NMDA (Cairns e colaboradores, 1998), ao passo
que injeção de GABA e de glicina, neurotransmissores inibidores, não levam a esse
aumento na atividade muscular mandibular (Cai e colaboradores, 1999).
Há um incremento na excitabilidade em neurônios do subnúcleo caudal
ocorrem após inflamação aguda ou crônica de tecidos craniofaciais profundos,
incluindo tecidos da ATM (Ren e Dubner, 1999) Adicionalmente, a duração da
atividade evocada em neurônios nociceptivos do subnúcleo caudal do tronco
encefálico e músculos mandibulares por aplicação de algésicos químicos na ATM
são similares (Hu e colaboradores,1992; Yu e colaboradores, 1996).
Essas mudanças neuroplásticas são marcantes uma vez que ocorre após
estímulos em forma de lesão ou inflamação, que levam horas, dias ou semanas e
são associadas com mudanças comportamentais na sensibilidade à dor. Parece
haver mecanismos que refletem uma plasticidade funcional ou sensibilização central

28
de neurônios nociceptivos e envolvimento de padrões extensivos de aferência para
esses neurônios (Sessle,1999).
Os aminoácidos excitatórios glutamato e aspartato estimulam o influxo de
cálcio através dos receptores dependentes de canais (MacDermott e colaboradores,
1986). SP promove uma elevação no cálcio intracelular por mobilizá-los para os
estoques intracelulares (Womack e colaboradores, 1988) e tanto SP (Womack e
colaboradores, 1989) quanto CGRP (Oku e colaboradores, 1988) aumentam o
influxo de cálcio através dos canais voltagem dependentes.
MacDermott e colaboradores (1986), sugerem que o desenvolvimento de
nocicepção persistente após irritação por formalina é influenciado pelo influxo de
cálcio, particularmente através de receptor NMDA.
Em adição às alterações celulares que provêm o mecanismo de plasticidade
estímulo-induzida que produzem essas mudanças na sua permeabilidade ao íon
cálcio, mecanismos moleculares levam a expressão de proto-oncogenes com a
proteína c-fos.
Hunt e colaboradores (1987) demonstraram que a proteína c-fos é expressa
em neurônios do corno dorsal pós-sináptico sucedendo estimulação térmica ou
química na pele. A expressão da fos tem sido demonstrada no corno dorsal espinhal
de ratos em resposta a compressão da pata (Bullitt, 1989),a injeção de formalina
(Kehl e colaboradores, 1991) ou carragenina (Draici e Ladarola 1989), na indução de
poliartrite com adjuvante de Freund (Menétrey e colaboradores, 1989), dentro da
articulação temporomandibular (Zhou e colaboradores, 1999). Foi também
observado o aumento dessa proteína na zona de transição do subnúcleo interpolar e
caudal do complexo trigeminal por inflamação do músculo masseter por injeção de
CFA (Imbe e colaboradores, 1999).
Estudos mostram uma relação entre a expressão da proteína fos estímulo-
induzida, neuroplasticidade central e comportamento de hiperalgesia.
Primeiro, o estímulo nocivo que produz a expressão da fos (calor, formalina e
agentes inflamatórios) também produzem hiperalgesia que é associada a
neuroplasticidade.(Coderre e colaboradores, 1990). Segundo, a curva temporal de

29
expressão da fos coincide com o desenvolvimento de comportamento hiperalgésico.
Por exemplo, inflamação periférica induzida por carragenina produz tanto incremento
de RNAm para c-fos como hiperalgesia com similar curva temporal (Draisci e
Ladarola, 1989).
A expressão da proteína fos estímulo induzida tem sido amplamente usada
para medir a ativação neuronal e é induzida nas vias trigeminais após estímulos
nocivos (Ren & Dubner, 1999).l
Uma das conseqüências desse aumento na expressão da proteína c-fos é a
sua participação na regulação de RNAm que codificam vários peptídeos na medula
espinhal de ratos. Inflamação periférica, estimulação do nervo trigêmio resultam em
incremento na expressão de RNAm codificador da dinorfina (Ruda e colaboradores,
1988), encefalina (Nishimori e colaboradores, 1989), SP (Minami e colaboradores,
1989) e peptídeo relacionado ao gene da calcitonina (Piehl e colaboradores, 1991)
no gânglio da raiz dorsal, corno dorsal ou no núcleo caudal.
Imbe e colaboradores (2001) comparando injeção de CFA na ATM e tecido
perioral de ratos, verificaram um aumento na expressão do RNAm para codificação
de preprodinorfina , um precussor da dinorfina, em tecidos inflamados na ATM
quando comparado com tecido perioral e relaciona essas mudanças como
colaboradoras para hiperexcitabilidade persistente e dor associada com desordens
temporomandibulares.
Laughlin e colaboradores (2000) relatam que citocinas têm sido associadas
com resposta inflamatória periférica, mostrando que IL1-beta contribui para o
desenvolvimento de alodinia dinorfina-induzida e o tratamento com citocinas
antiinflamatórias como IL-10 reverte esse processo.
Dinorfina tem sido mostrada tanto como uma inibidora (Kong e colaboradores,
1997) quanto estimuladora (Chao e colaboradores, 1995) na produção de citocinas.
A administração intratecal de dinorfina pode induzir alodinia crônica (Coderre e
colaboradores, 1992).
Encefalinas e outros agonistas delta opióides tipicamente produzem efeitos
inibitórios ou antinociceptivos (Vaught e colaboradores, 1982). Entretanto, esse

30
efeito pode prover um mecanismo onde a neuroplasticidade e a hiperalgesia são
mimetizadas (Dubner, 1991). Enquanto a dinorfina e outros kappa opióides são
classicamente conhecidos por favorecer efeito nociceptivo moderado (Millan e
Colpaert, 1991), eles apresentam um dual efeito no corno dorsal da medula
espinhal. Dinorfina, em particular, tem sido conhecida por aumentar o campo
receptivo e facilitar a resposta em aproximadamente um terço das células da
superfície do corno dorsal, enquanto produz inibição em outro terço das células
(Hylden e colaboradores,1991)

31
6- MECANISMOS DE MODULAÇÃO DA DOR
A transferência das informações nociceptivas do Corno Posterior da Medula
Espinhal (CPME) para segmentos rostrais do SNC sofreu profundas alterações
devido a atuação de influências excitatórias e inibitórias, em circuitos locais ou à
distância. Segundo Fields e Basbaum (1989) o tálamo é o centro da percepção da
dor e o neocortex, o centro da percepção discriminativa, que modula a resposta do
tálamo aos estímulos nociceptivos. As primeiras evidências concretas sobre a
existência de sistemas supraespinhais controladores das vias nociceptivas foram
apresentadas na década de 60 (Gordon e colaboradores, 1961). Um marco
importante na conceituação da existência de sistemas moduladores espelcíficos no
CPME foi apresentação da teoria de comporta por Melzack e Wall em 1965.
Segundos os autores, a inibição pré-sinaptica na medula espinhal decorreria da
colisão entre potenciais aferentes e potenciais antidrômicos negativos presentes na
raiz dorsal, originados na substância gelatinosa do CPME. A ativação prolongada de
interneurônios da substância gelatinosa pelos aferentes primários de grosso calibre
geraria potenciais nociceptivos. Segundo essa teoria, a ativação das fibras de
grosso calibre bloquearia a transflerência das informações dos aferentes primários
para os neurônios do CPME, assim como as influências hiperpolarizantes dos
aferentes de calibre fino. Entretanto, foi demonstrado que as aferências A-delta e C
também geram potenciais negativos na raiz sensitiva.
A via do segmento encefálico rostral ventromedial bulbar (RVM) representa um
importante mecanismo supraspinhal na modulação espinhal da transmissão
nociceptiva (Gjerstad e colaboradores, 2001). A estrutura da RVM inclui o núcleo
magno da rafe (NMR) e formação reticular lateral (Aimone e Gebhart,1986).
A transmissão nociceptiva trigeminal ativa a modulação descendente para
estruturas caudais encefálicas, incluindo a região ventro bulbar. Dubner e Ren
(2004) examinando a hipótese de que a atividade do subnúcleo caudal e interpolar
ter um papel importante na ativação do circuito modulatório da RVM, verificaram que
a injeção de CFA no músculo masseter leva a uma interação entre esses neurônios
trigeminais e modulação descendente na região ventrobulbar, indicando qlue lesões

32
nos tecidos profundos levam a ativação desses subnúcleos e alteram diversas
funções neuronais em tecidos craniofaciais e regiões orais.
O caminho modulatório descendente na antinocicepção mediada pelo núcleus
accumbens pode incluir a região periaquedutal (PAG) e região ventro bulbar (RVM).
Através de modelo reflexo de abertura da mandíbula em ratos, semelhante ao tail
flick, Gear e colaboradores (1999) verificaram que a capsaicina ativa circuitos
modulatórios que envolvem os sistemas opióide, dopaminérgico e gabaérgico.
Zhang e colaboradores (1998) analisando o efeito inibitório da estimulação
elétrica da região do córtex ventro lateral orbital (VLO) no reflexo de movimento de
abertura mandíbular em ratos, sugerem um papel importante dessa região na
modulação da nocicepção orofacial. Além disso, os autores propõem que o efeito
analgésico ocorre por estimulação da região VLO que, mediada pela PAG, ativa
sistemas inibitórios descendentes e deprime o processo nociceptivo trigeminal.
Recentes estudos indicam que a hiperalgesia em modelos animais de
inflamação e dor neuropática estão intimamente relacionados com circuitos
modulatórios descendentes envolvendo inibição e facilitação. Existem consideráveis
evidências de que a inibição descendente é acentuada após inflamação em locais
de hiperalgesia primária (Dubner & Ren, 2004).
Através de vários estudos enfocando a resposta de estímulos nocivos
transitórios (Willis, 1988), recentes evidências sugerem que a via descendente para
RVM também modula transmissão espinhal nociceptiva durante a dor inflamatória, e
tem um importante papel no desenvolvimento da dor persistente (Urban e
colaboradores, 1999; Terayama e colaboradores, 2000). Outros trabalhos reportam
que a modulação descendente da dor pode exibir mudanças na resposta a estímulos
nocivos persistentes sob várias condições (Dubner e Ren,1999; Hurley e Hammond,
2000). A atividade da RVM no circuito modulatório é aumentada durante a
inflamação e por isso acentua a inibição descendente da dor (Hurley e Hammond,
2000) bem como a facilitação (Terayama e colaboradores, 2000). Com todas essas
informações, sugere-se uma atividade modulatória na excitabilidade medular
espinhal e comportamento nociceptivo do circuito modulatório da dor.

33
Além dessas estruturas espinhais e supraespinhais destacadas, o controle
inibitório nocivo difuso (DNIC) também está envolvido na transmissão modulatória
nociceptiva. Danzinger e colaboradores (1999) investigando diferentes estágios
desenvolvidos durante a monoartrite induzida por adjuvante de Freund e verificando
a atividade dessa ação modulatória no contexto de dor aguda e crônica, observou
que em estágios agudos (24-48 horas) da inflamação há um aumento na atividade
da modulatória do DNIC. Em inflamação crônica (3-4 semanas), ocorre uma falha no
recrutamento desse controle modulatório. Essa diferença na atividade moduladora
de DNIC foi atribuída a uma sensibilização de neurônios durante a inflamação e uma
exacerbação paralela de DNIC durante o estímulo agudo, ao passo que a
cronicidade da lesão, leva a modificações nesse fenômeno modulatório como
resultado de mudanças neuroplásticas induzidas por dor prolongada.
Adicionalmente, estudos sugerem que aminoácidos excitatórios têm um papel
importante na modulação descendente na RVM (Urban e colaboradores, 1999). Em
animais normais, microinjeção de glutamato na RVM induz resposta bifásica
modulatória descendente na transmissão da dor, mostrando facilitação em baixas
doses e inibição em altas doses (Zhou e Gebhart, 1997).
Guan e colaboradores (2003) investigaram o papel dos aminoácidos
excitatórios da RVM na modulação descendente em modelo de hiperalgesia
inflamatória (CFA intraplantar) e demonstrou que a persistente hiperalgesia induz
acentuação tempo–dependente da neurotransmissão na RVM que envolvem
receptores NMDA e AMPA. Esses autores também demonstraram que há um
incremento da atividade de receptores AMPA do segmento RVM associado com
acentuação da inibição descendente após inflamação e hiperalgesia por ação de
CFA.
Essa progressão da modulação descendente da nocicepção foi também
estudada por Terayama e colaboradores (2002), constatando um aumento na
excitabilidade de neurônios RVM após inflamação da pata com CFA e inibição da
resposta nociceptiva. Esse estímulo nocivo induziu mudanças correlacionadas com
atividade dos receptores NMDA.

34
7- MODELOS ANIMAIS DE DOR OROFACIAL E COMPORTAMENTOS ASSOCIADOS.
Estudos em modelos animais de dor na região orofacial (músculo masseter,
lábio e região da ATM) através da indução por agentes pró-inflamatórios descrevem
um aumento na freqüência de comportamentos induzidos como hiperalgesia e
alodinia, correlaciona-os com eventos neurais em tecidos periféricos e neurônios do
SNC. Nesse mesmo contexto inflamatório, outros trabalhos mostram alterações
eletrofisiológicas e de comportamentos espontâneos (Yu e colaboradores, 1993;
Ren e Dubner, 1999, Svensson e colaboradores, 2003).
O fato de grandes distúrbios sensoriais ocorrerem na condição dolorosa
envolvendo tecidos mais profundos antes de incluir tecidos cutâneos (Sessle e Hu,
1991), levou a estudos comparando estímulos evocados por agentes algógenos em
tecidos orofaciais profundos versus orofaciais cutâneos. Os autores demonstraram
que há uma diferença na indução da neuroplasticidade (Yu e colaboradores, 1993) e
na expressão da proteína fos (Zhou e colaboradores, 1999) em neurônios do
sistema trigeminal.
Por injeção do Adjuvante Completo de Freund (CFA) no interior da ATM de rato
e na região do tecido perioral, desenvolveu-se uma persistente hiperalgesia
associada com inflamação orofacial (Zhou e colaboradores, 1999) semelhante ao
que foi visto em modelo de inflamação e hiperalgesia cutânea na pata (Hargreaves e
colaboradores, 1988). Nesse modelo também foi verificado por Imbe e
colaboradores (2001) a caracterização da resposta trigeminal na dor orofacial
profunda e no tecido cutâneo resultando em hiperalgesia e alodínia.
Outros trabalhos relacionando a estimulação de aferentes com algésicos (KCl,
histamina) na região orofacial e ATM demonstram um aumento na atividade reflexa
ou na eletromiografia (EMG) de músculos como: genioglosso, digástrico e temporal.
(Broton e Sessle, 1988).
Devido a necessidade de modelos experimentais que avaliem a dor orofacial
associada com alterações de comportamentos espontâneos, o teste da formalina,
originalmente descrito por Dubuisson e Dennis (1977), foi utilizado para testar a dor

35
nos tecidos superficiais da região orofacial por Clavelou e colaboradores (1989).
Neste modelo, a formalina é subcutâneamente injetada no lábio superior do rato e
respostas nociceptivas bifásicas são evocadas, principalmente fricção da área
injetada (rubbing ou grooming), descrita por ser um comportamento relacionado a
dor.
Clavelou e colaboradores (1995) testando o efeito de diferentes concentrações
de formalina observaram uma correlação positiva entre a concentração e a
amplitude da atividade rubbing. Foi observada uma atividade concentração-
dependente da formalina no aumento do tempo de execução da atividade rubbing,
mostrando uma atividade sobre esse comportamento até a concentração de 2,5%
que decresce em altas concentrações (5% a 10%). Esse comportamento foi
semelhante aos observados após estímulos elétrico, térmico e mecânico aplicados
na região orofacial de animais ou em neuropatia experimental como ligadura do
nervo infraorbital de ratos. Outros comportamentos variaram de acordo com a dose e
a fase do teste da formalina em que foram observados. Os autores relatam que um
único indicador como fricção da área da face ou lamber a pata, não podem ser
utilizados como um mensurador da intensidade de dor, pois alguns comportamentos
podem ser evidenciados de acordo com a concentração de formalina utilizada.
Essa observação também é relatada por Lee e colaboradores (2000) quando
descrevem que em altas concentrações de formalina (5% a 15%), alguns animais
apresentam considerável soma de tempo executando o comportamento de aparente
congelamento (freezing), que raramente é observado em concentrações
convencionais (por exemplo, 1,5% a 2,5%) e outros comportamentos como lamber
ou morder a pata são consideravelmente reduzidos na concentração de 5%.
Rosland e colaboradores (1990) relatam um efeito máximo no tempo de execução
de lamber a pata na concentração de 1,5% em ratos.
Vos e colaboradores (1998) avaliando comportamento de dor orofacial em
ratos após estimulos que variavam no grau de irritação (óleo mineral, compressão
das vibrissas, bupivacaína e formalina) verificaram o padrão de comportamento
coçar a face (grooming) no grupo injetado com formalina 5% e relatam que distúrbios
sensoriais como parestesias e anestesias produzem uma resposta transitória
assimétrica da face que permanece por alguns minutos.

36
Pelissier e colaboradores (2002) descrevem em modelo de dor orofacial
injetando capsaicina na região das vibrissas e relacionam como alteração
relacionada a dor, o comportamento de coçar a região. Os autores demonstram que
há uma correlação positiva entre a concentração de 0,25% a 1,5% e amplitude da
resposta de limpar-se ou coçar-se nessa região.
Ro e colaboradores (2003) injetando óleo de mostarda (20%), formalina (3%)
ou salina hipertônica (5%) na região média do músculo masseter verificaram uma
relação comportamental nociceptivo com o padrão de sacudir ou coçar com a pata
posterior na região injetada (scratching).
Hartwing e colaboradores (2003) demonstram através de modelo de
inflamação orofacial agudo descrito por Haas e colaboradores (1992), que o óleo de
mostarda desenvolve alteração de comportamentos caracterizados e relacionados a
nocicepção como: congelar-se, sacudir a cabeça, coçar-se com a pata posterior na
área injetada, movimento de rotação da mandibula.
Ro e colaboradores (2003) relatam que há uma diferença nos
comportamentos nociceptivos ou na ativação de musculos mastigatórios quando se
injeta óleo de mostarda no músculo masseter ou na ATM.Isso tem sido demostrado
por Roveroni e colaboradores (2001) que introduziram um modelo comportamental
de dor na região da ATM com a formalina. Os autores descrevem que os efeitos da
formalina variam de acordo com a concentração (0,5; 1,5; 2,5; 5,0%) e relacionam
alterações comportamentais nociceptivas como coçar a face na região injetada,
movimento de rotação da mandíbula ou mastigação e sacudir a cabeça como
respostas induzidas pela formalina.
Anderson e colaboradores (2003) descrevem que a constricção crônica do
nervo infraorbital acentua a resposta nociceptiva desencadeada pelo teste da
formalina e observaram que esse modelo de dor neuropática acentua a resposta de
fricção da região da vibrissa quando se injetava formalina 3%. Outros
comportamentos adicionais como sacudir a cabeça, vocalizações espontâneas,
trismo ou limitação da abertura bucal, vigorosas sacudidas seguidas por lambidas da
pata e movimentos de proteção da face enquanto fecha os olhos são sugestivos de
sofrimento moderado a severo e são indicativos de dor.

37
Além dos comportamentos como fricção da região, sacudir a cabeça para o
lado doloroso, movimento de rotação da mandíbula e coçar-se com a pata posterior,
devido a associação entre as manifestações dolorosas e os distúrbios do sono, um
fenômeno recentemente observado é o tempo de sono em ratos submetidos a
lesões da região orofacial com agentes flogísticos (Andersen e Tufik , 2000).
Schütz e colaboradores (2003) verificaram alterações no padrão de sono,
movimentos de limpeza e coceira do lado irritado na região orofacial, movimento de
rotação da mandíbula e sacudidas da cabeça em modelo de animais utilizando CFA
na ATM de ratos, relacionando uma redução na eficiência do sono e incremento nas
vezes em que o animal despertava como sinal indicativo de nocicepção.

38
8- CONTROLE FARMACOLÓGICO DA DOR OROFACIAL
Terapias analgésicas para condições dolorosas agudas e crônicas são
baseadas em três classes de drogas: drogas antiinflamatórias não esteroidais
(DAINES); analgésicos opióides e drogas com diversas ações farmacológicas
conhecidas como adjuvantes (antidepressivos, anticonvulsivantes, anestésicos
locais, agonista receptor alfa-2). Tanto DAINES quanto os opióides exibem uma
variedade de ações adversas, e em muitos estágios de dor crônica, particularmente
envolvendo lesões do nervo, são inadequadamente controlado por esses agentes,
os adjuvantes são freqüentemente necessários para adequar a dose antes
necessária para o alívio da dor ou diminuir os efeitos colaterais. A farmacoterapia na
dor crônica e dor neuropática tem sido descritas extensivamente em vários estudos
(MacPherson, 2000).
8.1- DROGAS ANTIINFLAMATÓRIAS NÃO-ESTEROIDAIS (DAINES)
As drogas antiinflamatórias não esteroidais (DAINES) caracterizam-se pela
propriedade comum analgésica, antipirética e antiinflamatória. Sua atividade reside
no decréscimo da inflamação periférica e depressão na dor por ela induzida. As
diferenças básicas entre os vários tipos de DAINES estão na sua farmacocinética,
potência antiinflamatória e analgésica e efeitos colaterais.

39
8.1.1- MECANISMO DE AÇÃO
Vane (1971) através de suas observações propôs a habilidade de DAINES
em suprimir a inflamação por inibir a enzima cicloxigenase, limitando a produção de
proinflamatórios (PGs) no sítio da lesão.
DAINES possuem um mecanismo de ação central e periférico por inibição das
cicloxigenases e subsequente inibição de endoperóxidos cíclicos e prostaglandinas,
mediadores da inflamação e da hiperalgesia primária (Barros, 2004).
Os DAINES de 1ª geração bloqueiam tanto a COX-1, constitutiva, presente na
maioria dos tecidos e relacionada à função renal, plaquetária e à proteção da
mucosa gástrica e intestinal, como a COX-2, constitutivamente presente no SNC e o
aparelho urogenital, e induzida pelo processo inflamatório e expressada no endotélio
e macrófagos. Recentemente, foi descrita uma nova forma de ciclooxigenase a
COX-3, presente no SNC e aparente alvo de ação da dipirona e do paracetamol
(Barros, 2004; Chandrasekharan e colaboradores, 2002).
Apesar de a importância da atividade da COX na produção de PGs ter sido
bem esclarecida, a inducibilidade dessa atividade e o papel central desta indução
na amplificação da inflamação ainda tem sido estudada. Evidências advindas de
modelos animais de artrite inflamatória sugerem que o aumento da expressão da
COX-2 é responsável por incremento na produção de PGs vista em tecidos
articulares inflamados (Anderson e colaboradores, 1996). O bloqueio periférico na
produção de prostaglandinas durante a inflamação reduz a continuação da dor e da
hiperalgesia e, ainda, interfere na inflamação neurogênica (Craig & Sorkin, 2001).
Danos locais e doenças inflamatórias como a osteoartrite são associadas com
incremento de PGs, e receptores nociceptivos são conhecidamente sensíveis a
baixos níveis de PGs (Dray & Urban, 1996). As PGs não ativam nociceptores, mas
causam sensibilização, decréscimo do limiar em que eles são ativados por estímulos
químicos, mecânicos e térmicos, onde normalmente são inócuos(Craig & Sorkin,
2001). Além disso, PGs atuam sobre a medula espinhal facilitando a transmissão de
resposta dolorosa e DAINES tem sua ação nesse sítio central (Cashman, 1996).

40
Durante estágios de dor crônica as PGs são liberadas na medula espinhal,
onde contribuem para a sensibilização central da mesma forma que ocorre em seus
efeitos periféricos, por exemplo, inibindo a liberação de neurotransmissores para
aferentes primários terminais. A administração na medula espinhal de pequenas
quantidades de DAINES elimina a acentuada liberação e, adicionalmente, reduz
inflamação neurogênica periférica por inibir estímulo antidrômico nos tecidos
periféricos. DAINES podem prover um efeito analgésico central por interagir com
mecanismo endógenos controladores de dor na região Periaquedutal Mesencefálica
(PAG) e suas ações podem ser sinérgicas com atividade de opióides (Vaughan e
colaboradores, 1997).
Pitcher e Henry (1999) descrevem que a administração de inibidores de
ciclooxigenase como a indometacina deprimem de forma dose-dependente a
resposta nociceptiva em neurônios do corno dorsal espinhal tanto em estímulos
cutâneos como elétricos do nervo ciático.
Drogas como aspirina, indometacina e cetoprofeno demonstram um efeito
antinociceptivo no plano medular espinhal (Bustamante e colaboradores, 1997).
Outros trabalhos têm demonstrado a ação de DAINES sobre as fibras C
(Bustamante e colaboradores, 1996); em neurônios WRD (Pitcher e Henry, 1999).
Os efeitos da administração intracerebroventricular (i.c.v.) e intratecal (i.t.) de
DAINES tem sido reportada por ter atividade em modelos animais de dor
experimental onde processo inflamatório e a hiperalgesia estão presentes.
Ferreira e colaboradores (1978) estudaram a ação da aspirina, indometacina
e do paracetamol e concluíram que esses DAINES agem diminuindo a hiperalgesia
desencadeada por injeção de carragenina em modelos de edema de pata. Okuyama
e Aihara (1984) em modelos de artrite experimental observaram que a indometacina
diminuiu a resposta de vocalização induzida eletricamente e o diclofenaco reduziu o
número de contorções abdominais (Björkman e colaboradores, 1992). Os salicilatos
mostraram-se efetivos analgésicos sobre a ação do adjuvante no edema de pata
(Higushi e colaboradores, 1986).

41
Malmberg e Yaksh (1992) usando o teste da formalina, relataram o efeito
antinociceptivo de DAINES e referiram sua atividade na inibição da formação de
prostanóides. A indometacina também reduziu a resposta de neurônios do corno
dorsal em teste da formalina.
Wang e colaboradores (1994) também reportam que a administração de
intratecal de DAINES inibe o reflexo tail-flick e a resposta nociceptiva da segunda
fase do teste da formalina (Malmberg e Yaksh, 1994) bem como a resposta de
retirada da pata em estímulos térmicos (Malmberg e Yaksh, 1992).
Outros trabalhos também reportam a ação da indometacina em atenuar a
resposta dolorosa na segunda fase do teste da formalina em ratos. Nesses estudos,
a segunda fase é um modelo válido de dor inflamatória (Hunskaar e Hole, 1987).
Tem sido demonstrado que indometacina reduz a produção de PGE2 que é
liberada por aplicação de capsaicina no corno dorsal da medula espinhal bem como
o incremento de PGE2 induzida por injeção de formalina. Contudo, a inibição de
COX pode favorecer ao incremento do substrato, o ácido araquidônico ou desviar o
metabolismo do ácido araquidônico através de outros caminhos derivados, como o
da lipoxigenase ou isoprostanos (Pitcher e Henry, 1999).
Vaughan e colaboradores (1997) especulam que inibidores de COX
aumentam a síntese de eicosanóides na PAG via lipoxigenase. Kirchner e
colaboradores (1997) reportam que a indometacina produz elevados níveis de
leucotrienos na mucosa gástrica de ratos. Por esta razão, não se deve excluir algum
envolvimento do ácido araquidônico, da via da lipooxigenase e de outros
componentes do ciclo lipídico no mecanismo excitatório de receptores nociceptivos.

42
8.1.2- EFEITOS INDESEJÁVEIS
Baseado em estudos sobre o mecanismo da cascata do ácido araquidônico,
onde se observou que existem duas isoformas da enzima ciclooxigenase, foi definido
que a forma constitutiva (COX-1) é importante nos processos fisiológicos modulados
pela produção de PGs de onde provém a homeostase do organismo, e que a
induzida (COX-2) é responsável pela lesão tissular e inflamação (Dubois e
colaboradores, 1998).
Dessa forma os maiores efeitos dos DAINES estão relacionados com
toxicidade gástrica (dispepsia, ulceração gastroduodenal e hemorragia
gastrointestinal), agregação plaquetária e alteração da função renal, aparentemente
causada pela inibição da COX-1 (Dubois e colaboradores, 1998). Outros efeitos
incluem: insuficiência e necrose hepática; rash cutâneo e prurido e problemas
respiratórios envolvendo rinite e piora de crises de asma.
Por outro lado, essas ações sobre a mucosa gástrica e sobre a atividade renal
levaram ao desenvolvimento de vários modelos experimentais com o intuito de
estudar os efeitos da indometacina sobre a fisiologia gástrica (Xiang e Huang, 2004)
e renal (Sokolova e colaboradores, 1977) através da administração crônica desse
DAINE em pequenas doses. Os autores observaram que a indometacina promovia
mudanças na resistência vascular periférica causando o desenvolvimento da
hipertensão arterial.

43
8.2.- ANESTÉSICOS LOCAIS
Os canais de sódio sensíveis a voltagem (VGSCs) têm um papel importante
no controle da excitabilidade neuronal e uma família de genes que codificam esses
canais tem sido identificada (Catteral, 2000). Neurônios sensoriais contendo canais
(VGSCs) sensíveis a tetrodotoxina , bem como alguns neurônios parcialmente
sensíveis ou resistentes a tetrodotoxina são expressos em terminais aferentes
sensoriais (McClesky e Gold,1999).
Alterações na expressão, distribuição e função desses neurônios podem
ocorrer após lesões ou inflamação crônica que tem um profundo efeito em neurônios
aferentes, contribuindo para expressão de comportamentos de dor (McCleskey e
Gold, 1999).
Anestésicos locais que bloqueiam VGSCs são usados para abolir a dor
temporariamente por bloquear a condução nervosa, mas também estão sendo
usados para o tratamento de muitas condições dolorosas crônicas (Sawynok, 2003).
8.2.1- MECANISMO DE AÇÃO
Os anestésicos locais bloqueiam o início e a propagação dos potenciais de
ação, impedindo o aumento da condutância aos íons Na+ através de canais
voltagem-dependentes. Apesar de exercerem uma variedade de efeitos
inespecíficos sobre a função da membrana, sua principal ação consiste em bloquear
os canais de Na+, o que fazem fisicamente ao fechar o poro transmembrana,
interagindo com radicais do domínio da hélice transmembrana S6 (Strichartz e
Ritchie, 1987).
A atividade anestésica local aumenta em pH alcalino. Isto decorre do fato de
que o composto precisa penetrar na bainha do nervo e a membrana axônica para,
então, alcançar a extremidade interna do canal de Na+ (onde se encontra o sítio de

44
ligação dos anestésicos locais). Como a forma ionizada não atravessa a membrana,
a sua presença é muito precária em pH ácido. Uma vez no interior do axônio, é a
forma ionizada da molécula anestésica local que se liga ao canal. Essa dependência
em relação ao pH pode ser clinicamente importante, visto que os tecidos inflamados
são freqüentemente ácidos e, portanto, levemente resistentes aos agentes
anestésicos locais (Rang e colaboradores, 1999a).
Em modelos de dor aguda os anestésicos mostram-se bastante eficazes.
Sakaue e colaboradores (2004) comparando os efeitos antinociceptivos de vários
bloqueadores de canais de sódio na dor aguda térmica e mecânica demonstraram
que esses agentes têm uma preferencial resposta antinociceptiva contra estimulação
térmica e possuem um papel importante na diferenciação entre nocicepção térmica e
mecânica.
Para compreensão da ação efetiva de anestésicos locais como um fator
diferenciador entre a estimulação aguda térmica da mecânica, é preciso conhecer os
distintos caminhos mediadores da geração e condução de sinais dolorosos térmicos
e mecânicos.
Na condução de sinal nociceptivo para neurônios do corno dorsal, Doucette e
colaboradores (1987), reportam que as fibras C são ativadas na resposta para
estímulos nocivos por calor. Além disso, Yeomans e colaboradores (1996),
demonstraram que os nociceptores do tipo C e/ou fibras A-delta ativados por
nocicepção mecânica depende da faixa de aquecimento da pele. Uma pequena
faixa de aquecimento preferencialmente ativa nociceptores das fibras C, enquanto
que altas faixas de calor na pele atinjem nociceptores das fibras A–delta. Em
contrapartida Hu e Sessle (1988), demonstram que tanto fibras A-delta e C são
ativadas em resposta para estímulos mecânicos. Adicionalmente, distintos
receptores medeiam diferentes sinais nociceptivos. Por exemplo, segundo Price e
colaboradores (2000) demonstram que sensações promovidas por substâncias
ácidas são relacionadas como estimulação mecânica nos nociceptores e os
receptores de capsaicina integram a nocicepção mediada por calor (Tominaga e
colaboradores, 1998).

45
Em geral os anestésicos locais bloqueiam a condução das fibras nervosas de
pequeno diâmetro mais facilmente que nas fibras grandes. Todavia, as fibras
menores nos nervos periféricos consistem em fibras não mielinizadas, que são bem,
menos suscetíveis do que as fibras mielizadas menores. Como os impulsos
nociceptivos são transmitidos por fibras mielinizadas e não mielinizadas, a sensação
da dor é bloqueada mais facilmente do que outras modalidades sensoriais (tato,
propriocepção). Os axônios motores, em virtude de seu grande diâmetro, também
são relativamente resistentes. As diferenças de sensibilidade entre diferentes fibras
nervosas, embora sejam facilmente medidas em condições experimentais, não tem
grande importância prática e raramente é possível produzir um bloqueio da
sensação dolorosa sem afetar outras modalidades e provocar paralisia local. (Rang
e colaboradores, 1999a).
8.2.2- EFEITOS INDESEJÁVEIS
Os principais efeitos indesejáveis dos anestésicos locais estão relacionados
ao sistema nervoso central e cardiovascular, constituindo uma fonte de risco quando
são utilizados clinicamente.
A utilização do anestésico local tem efeito estimulador sobre o sistema
nervoso central, resultando em tremor e agitação e sinais subjetivos que variam
desde confusão mental até agitação extrema. O tremor pode progredir para
verdadeiras convulsões, e o aumento adicional da dose provoca depressão do SNC.
A procaína tende particularmente a produzir efeitos centrais indesejáveis, que é
motivo de sua substituição por agentes como lidocaína e a prilocaína no uso clínico
em que os efeitos centrais são menos pronunciados (Rang e colaboradores, 1999a).
Os efeitos cardiovasculares decorrem, principalmente na depressão do
miocárdio e da vasodilatação. A redução da contratilidade miocárdica provavelmente
resulta, indiretamente, da inibição da corrente de Na+ no músculo cardíaco. A
conseqüente redução do Na+, por sua vez, diminui as reservas intracelulares de
Ca+, reduzindo a força da contração. O efeito antidisrítmico de alguns anestésicos

46
locais, particularmente lidocaína, tem utilidade clínica. (Rang e colaboradores,
1999a).
A vasodilação que afeta principalmente as arteríolas decorre, em parte, da
inibição do sistema nervoso simpático. A depressão miocárdica e a vasodilação
combinadas levam a uma queda da pressão arterial, que pode ser súbita e
potencialmente fatal.
8.3- ANALGÉSICOS OPIÓIDES
O termo opioíde aplica-se a qualquer substância seja endógena ou sintética,
que produz efeitos semelhantes ao da morfina, que são bloqueados por
antagonistas como a naloxona. O termo mais antigo, opiáceo, ficou restrito a drogas
semelhantes à morfina com estruturas não peptídicas (Herz, 1993).
Estudos farmacológicos (Lord e colaboradores, 1977) têm demonstrado a
existência de três subtipos de receptores: mu, lambda e kappa. Várias evidências
anatômicas e farmacológicas reforçam o papel de pelo menos dois desses
receptores em regular o processo nociceptivo. A expressão do receptor mu no
gânglio da raiz dorsal, bem como seus aferentes terminais centrais na medula
espinhal, foi demonstrado por Wang & Wessendorf (2001).
8.3.1- MECANISMO DE AÇÃO
Os opióides agem nesses receptores e cada um possui uma atividade e
desenvolve efeitos terapêuticos e/ou reações adversas. Essas ações dependem da
afinidade de ligação pelo receptor. Pelo menos dois tipos de receptores de opióides
(mu e kappa) medeiam a analgesia. Os receptores mu são amplamente distribuídos
através do SNC, especialmente no sistema límbico (córtex frontal, córtex temporal,

47
amígdala e hipocampo), tálamo, hipotálamo e mesencéfalo, assim como nas laminas
I, II, IV e V do corno dorsal e na medula espinhal. Os receptores mu são abundantes
na substância cinzenta periaquedutal do tronco cerebral e corno dorsal da medula
espinhal. Os receptores kappa estão presentes também nestas áreas, bem como
nas camadas mais profundas do córtex cerebral. (Bojrab, 1996).
Os opióides mostram-se eficazes como analgésicos quando administrados
por via intratecal em pequenas doses, implicando que uma ação central pode ser
responsável pelo seu efeito analgésico (Rang e colaboradores, 1999b).
A injeção de morfina na região da PAG produz analgesia acentuada, que
pode ser prevenida pela interrupção cirúrgica da via descendente para o NRM ou
pelo bloqueio de 5-HT com p-clorofenilalanina. A morfina aumenta a deflagação dos
neurônios associados às vias inibitórias descendentes, implicando a existência de
um componente supramedular significativo (Bojrab, 1996).
A analgesia opióide não é mediada exclusivamente pelo SNC. Aplicado
localmente os opióides têm ação antinociceptiva e antiinflamatória em tecidos
animais inflamados (Stein, 1993). A morfina intrarticular produz potente analgesia
após cirurgia da articulação do joelho e seus efeitos são mediados por receptores
opióides fora do SNC. Experimentos animais têm mostrado desta forma que
receptores opióides estão localizados em sistema nervoso sensorial terminal
(Hassan, 1993) e esses receptores podem ser ativados por agonistas mu, kappa e
delta exógenos (Stein e colaboradores, 1993), bem como por peptídeos opióides
endógenos expressos em células inflamatórias (Przewlocki e colaboradores, 1992).
Esses peptídeos podem ser liberados e produzirem potente inibição tônica da dor
clinica, com poder similar ao observado com a morfina. Além disso, esses efeitos
são revertidos por naloxona, sugerindo que a morfina e os peptídeos endógenos
agem na mesma população de receptores opióides periféricos (Sawynok, 2003).

48
8.3.2- EFEITOS INDESEJÁVEIS
Além da analgesia mediada pelo receptor mu, a morfina produz euforia
relacionada principalmente na atividade nesse receptor, que é equilibrada pela
disforia (ansiedade, alucinações) associada à ativação dos receptores kappa (Rang
e colaboradores, 1999b).
A depressão respiratória e do reflexo da tosse, náuseas, vômitos, constrição
pupilar, redução da motilidade gastriintestinal relacionam-se com a atividade sobre
os receptores opióides, variando de acordo com a sua afinidade (Rang e
colaboradores, 1999b).
O tratamento da dor central com analgésicos opióides é limitado
principalmente pela tolerância, que é o decréscimo do efeito da droga em
administração prolongada ou o composto relacionado agindo no mesmo receptor
(endógeno). Stein e colaboradores (1996) propuseram um mecanismo de tolerância
ao opióide e incluem: dowregulation de receptores, não acoplamento do receptor a
proteína G, inativação da proteína G e incremento da atividade da adenilato ciclase.
Powell e colaboradores (1999) investigaram o efeito de DAINES na tolerância
a morfina, associando-a com ibuprofeno, resultando em aumento da potência desse
opióide. Craig e Sorkin (2001) demonstram que a aspirina pode agir sinergicamente
com efeitos da morfina na região cinzenta periaquedutal e que a co-administração
de adjuvantes auxilia na prevenção do desenvolvimento da tolerância.

49
9- RELEVÂNCIA E JUSTIFICATIVA
Em face do exposto, ficou estabelecido que a patogênese das Disfunções
Temporomandibulares é baseada em um componente inflamatório originado da
resposta protetora dos tecidos que compõem a ATM, onde a sua persistência leva a
desenvolvimento e manutenção do estado hiperalgésico promovendo a dor na
região orofacial. Diante da necessidade de mais estudos experimentais envolvendo
um componente inflamatório crônico para esclarecer manipulações terapêuticas
farmacológicas propomos a elaboração de um modelo experimental de dor nessa
região.
É importante destacar que não existem trabalhos envolvendo ação
inflamatória persistente como o CFA na região orofacial e sua atividade sobre
movimentos espontâneos, bem como a caracterização nociceptiva de tais
comportamentos através da manipulação farmacológica dos mesmos.

50
10- OBJETIVOS
Portanto, o objetivo geral desse trabalho foi desenvolver um modelo
nociceptivo através de injeção de Adjuvante Completo de Freund (CFA) na região da
articulação temporomandibular em ratos.
Para isso osbjetivos específicos são os seguintes:
§ Verificar alterações de comportamentos espontâneos: 1- farejar e cheirar (sniffing),
2- empinar-se (rearing), 3- movimento de rotação da mandíbula (chewing like), 4-
subir escadas (climbing), 5- congelar-se (freezing), 6- dormir /descansar (rest/
sleeping), 7- limpar-se e fricção da região orofacial (grooming ou rubbing), 8- lamber-
se (licking), 9- coçar-se (scratching), 10- morder-se (biting), 11- sacudir a cabeça
(head fliches).
§ Determinar a curva temporal de cada comportamento, estabelecendo o período
onde as alterações se evidenciaram, correlacionando possíveis alterações com
padrões nociceptivos estudados na literatura;
§ Testar drogas classicamente analgésicas para caracterizar farmacologicamente os
comportamentos associados a nocicepção em nosso modelo.
§ Verificar se há alteração no peso dos animais devido a indução da disfunção
temporomandibular dos animais.
§ Avaliar o tipo de alteração celular na região da ATM dos animais através de
análise histológica comparando com alterações ocorridas entre os animais tratados
com solução salina 0,9%, CFA e formalina 2,5%.
§ Fazer uma análise comparativa entre o modelo da formalina e modelo com o
Adjuvante Completo de Freund (CFA) na região da ATM com o intuito de estabelecer
diferenças entre um modelo de inflamação subaguda ou aguda e o modelo de
inflamação crônica e dor persistentes respectivamente, estabelecendo quais
parâmetros são mais relevantes em cada modelo.

51
II - MATERIAL E MÉTODOS
2.1- ANIMAIS
Foram utilizados 90 ratos Wistar machos, pesando entre 180 e 300 gramas,
provenientes do Biotério Central do Campus do Pici – Universidade Federal do
Ceará e do Biotério Setorial do Departamento de Fisiologia e Farmacologia dessa
mesma universidade.
Os animais eram mantidos em caixas plásticas (grupo = 6) com água e
comida ad libitum, seguindo o ciclo claro-escuro. As caixas continham no solo cerca
de 5mm de raspa de madeira. As observações eram realizadas durante o dia,
sempre entre 8:00 e 17:00 horas.
Foram seguidas normas segundo Zimmerman (1983) que a partir do comitê
de estudo e ética da Associação Internacional para o Estudo da Dor-IASP (1983),
concernente aos aspectos de estudo em dor experimental e sofrimento que pode
causar no animal.
2.2- DROGAS E SOLUÇÕES
- Adjuvante completo de Freund (Sigma Chemical Co. St. Louis, USA)
-Cloridrato de Morfina (Dimorf, laboratório Cristália, Itapira, SP, Brasil) doada pelo
departamento farmacêutico do Hospital Walter Cantídio. Fortaleza – Ceará.
-Indometacina (Indocid, laboratório Merk Sharp & Dohme) doada pelo departamento
farmacêutico do Hospital Walter Cantídio. Fortaleza – Ceará.
-Cloridratos de lidocaína e de fenilefrina (Novocol 100 – SsWhite, Rio de Janeiro, RJ,
Brasil)
-Bicarbonato de sódio (Reagen, Rio de Janeiro, RJ, Brasil)

52
-Formaldeído 37% (Sigma-Aldrich Chemical Co. ST. Louis, USA)
-Éter etílico (DPF vetec, Rio de Janeiro, RJ, Brasil)
- Solução de bicarbonato de sódio utilizado a 5% (p/v) dissolvida em água destilada
- Solução de morfina em solução salina fisiológica 0,9% (10mg/5ml).
- Solução de indometacina em solução salina 0,9% (25mg/5ml)
-Solução salina fisiológica 0,9% estéril (endogen)
-Solução de formalina em água bidestilada (10%).
-Solução de formalina em água bidestilada (2,5%).
2.3. MATERIAL E EQUIPAMENTOS
- Computador IBM-PC e programa básico comporta
- balança analógica
- seringa descartável (1cc) (BD)
- agulha 30 G (BD)
- câmara para anestesia.
2.4- PROTOCOLO EXPERIMENTAL
No primeiro dia, os animais foram pesados e passaram por um período de
adaptação de 10 minutos em um observatório de madeira (100 cm x 50 cm x 50 cm)
iluminado com uma lâmpada (40 watts) em um dos cantos. A parede frontal do
observatório era de vidro de modo a permitir ao experimentador observar os
elementos comportamentais. O observatório era forrado com serragem de madeira.

53
Após o período de adaptação, os animais eram removidos do observatório,
anestesiados em uma câmara contendo éter etílico. Quando os animais se
encontravam anestesiados, eram removidos da câmara.
O sítio da injeção na região temporomadibular foi identificado por palpação do
arco zigomático e côndilo mandibular. Uma agulha 30G foi inserida no ponto
imediatamente inferior ao bordo postero inferior do arco zigomático. Após a
localização do ponto, a solução de acordo com cada grupo foi injetada (Zhou e
colaboradores, 1999) (Quadro 1).

54
Protocolo experimental: Quadro 1 – apresenta o resumo esquemático do
protocolo experimental utilizado para o estudo. Após 10 minutos de adaptação, os
animais foram divididos em três grupos de acordo com a solução a ser injetada,
após a palpação da região da ATM, em um volume de 50 microlitros.
NaCl 09,% Formalina 2,5%
CFA
10 minutos de adaptação
Observação por 30 minutos

55
2.4.1- OBSERVAÇÕES COMPORTAMENTAIS
Imediatamente após a injeção os animais eram recolocados na caixa para
observação comportamental. Esses dados eram registrados em um computador
IBM-PC, através de um programa Comporta de quantificação de comportamentos,
desenvolvido em nosso laboratório sob a orientação do Prof. Dr. Marcus Raimundo
Vale. No computador havia comando, onde cada tecla registrava os comportamentos
selecionados. Foram registrados os seguintes comportamentos abaixo descritos:
Congelar-se (freezing): o animal fixa o olhar em algum ponto mantendo o
corpo rígido, estático e com orelhas empinadas.
Empinar-se (rearing): o animal empina o corpo apoiando-se sobre as patas
posteriores ao mesmo tempo que mantém as patas anteriores livres ou recostadas
às paredes do observatório à medida que se desloca.
Farejar (sniffing): o animal fareja o solo, as paredes, os recôndidos do
ambiente através de seu focinho ao mesmo tempo em que movimenta suas vibrissas
de uma maneira rápida, ativa e deslocando-se por todos os locais do observatório.
Limpar-se (grooming ou rubbing): o animal através de movimentos de vai-e-
vem de suas patas anteriores em torno de seu focinho e região lateral da face e
girando para o dorso e patas, lambendo-os e às vezes, realizando pequenas
mordidas.
Repousar (rest/sleeping): o animal repousa quando mostra o relaxamento de
seu corpo dobrando-o num local cuidadosamente escolhido. O relaxamento também
se faz por flacidez e declínio de sua orelha. Toda essa série de atos culmina com o
fechamento dos olhos.
Coçar-se (scratching): esse comportamento é expresso por movimentos
rítmicos, rápidos das patas posteriores dirigidos à cabeça e ao dorso, tocando-os
levemente. Esse comportamento, em geral, faz parte de uma seqüência ao ato de

56
limpar-se (grooming). De forma rara e não comum coçar-se pode ocorrer de forma
isolada e consiste de uma vibração das patas ao ar, sem atingir o corpo.
Morder-se (biting): o animal realiza pequenas mordidas em toda região do seu
corpo além da pata anterior quando pode ser considerado grooming.
Lamber-se (licking): o animal limpa o seu corpo além a região anterior da face
(grooming).
Movimento de rotação da mandíbula ou mastigação (chewing like): o animal
inicia movimentos como estivesse mastigando ou rangendo os dentes, ou ainda,
tremor da musculatura facial.
Sacudir a cabeça (head fliches): o animal realiza movimentos aparentemente
com intuito de remover algo da cabeça ou região da face. Esse movimento é o
equivalente a sacudidas da pata anterior ou posterior em outros modelos.
Subir as escadas (climbing): verificava-se a capacidade do animal em subir os
degraus da escada. Era considerado o tempo (s) contabilizado a partir do momento
em que o animal colocava as patas anteriores no 1º degrau da escada.
Após cada observação, os animais foram recolocados em suas caixas e lhes
era oferecido água e comida à vontade.
O experimento terminou no 7º dia após a injeção das três soluções testadas,
quando foi observado o declínio da curva da resposta do CFA. Os animais foram
pesados para comparação com os valores obtidos no 1º dia do experimento e depois
sacrificados para análise histológica.

57
2.4.2- GRUPOS EXPERIMENTAIS
- Controle normal
Corresponde ao grupo de animais em que não foi injetada nenhuma solução,
mas que passou pelo período de adaptação e que foi realizada a avaliação
comportamental.
- Controle com agulha sem injeção de solução salina fisiológica
Compreende o grupo de animais em foi inserida a agulha na região da
articulação temporomandibular, mas sem injetar solução salina fisiológica 0,9% ou
agente irritante.
- Controle com agulha e com solução salina fisiológica
Corresponde aos animais em que foram injetados 50 microlitros de solução
salina fisiológica 0,9%.
- Grupo Adjuvante Completo de Freund
Corresponde aos animais em que foram injetados 50 microlitros de CFA.
- Grupo formalina
Corresponde aos animais em que foram injetados 50 microlitros de solução
de formalina 2,5%.

58
2.4.3 - TESTES FARMACOLÓGICOS
Nesses grupos tivemos como objetivo a caracterização farmacológica de nosso
modelo experimental. As drogas testadas foram: indometacina (5mg/Kg); Morfina
(4mg/Kg) e o anestésico local lidocaína com vasoconstrictor a 2% (0,4 ml).
Os grupos de animais para testes farmacológicos foram subdivididos de acordo
com os dias onde foram detectadas as principais alterações comportamentais
determinada estatisticamente significante na curva temporal de cada
comportamento. Assim, de acordo com a curva temporal de comportamento foram
determinados os principais comportamentos alterados pelo CFA foram: chewing like,
sleeping, grooming e freezing. Além disso, os comportamentos destacados pela
administração da formalina 2,5% foram o grooming e o scratching.
– INDOMETACINA (Indocid 25 mg)
Os animais, antes de se testar as drogas analgésicas, foram deixados 24 horas
em jejum devido a via de administração escolhida ser a oral.
Os animais eram pesados no dia da administração das drogas e a dose era
calculada de acordo com o seu peso.
As drogas foram administradas via oral, através de uma sonda orogástrica.
Decorridos 30 minutos após a administração do analgésico, se iniciavam as
observações.
- INDOMETACINA 3º dia após CFA (Indocid 25 mg)
A indometacina foi testada em grupos com oito animais no 3º dia após injeção
do CFA. A dose utilizada foi de 5mg/Kg (v.o) de solução (10mg/5ml) de indometacina
diluída em solução salina fisiológica 0,9% na proporção de 1:10.

59
- INDOMETACINA 4º dia após CFA (Indocid 25 mg)
A indometacina foi testada em grupos com oito animais no 4º dia após injeção
do CFA. A dose utilizada foi de 5mg/Kg (v.o) da solução (10mg/5ml) de indometacina
diluída em solução salina fisiológica 0,9% na proporção de 1/10.
- INDOMETACINA 1º dia após formalina 2,5% (Indocid 25 mg)
No grupo da formalina 2,5%, a indometacina foi testada em um grupo com seis
animais nos 30 minutos antes da injeção a solução de formalina. A dose utilizada foi
de 5 mg/Kg (v.o) da solução de bicarbonato de sódio diluído em de solução de
10mg/5ml de indometacina diluída em solução salina fisiológica 0,9% na proporção
de 1/10. Nesse grupo foram avaliados todos os comportamentos evidenciados ao
longo da determinação da curva temporal da formalina 2,5% em relação ao CFA.
- MORFINA (Dimorf 10mg)
Os animais, antes de se testar as drogas analgésicas, foram deixados 24 horas
em jejum devido a via de administração escolhida ser a oral,
Os animais eram pesados no dia da administração das drogas e a dose era
calculada de acordo com o seu peso.
As drogas foram administradas via oral, através de uma sonda orogástrica.
Decorridos 30 minutos após a administração do analgésico, se iniciavam as
observações.

60
- MORFINA 3º dia após administração do CFA
Seguindo-se o mesmo protocolo do grupo testado com indometacina 5mg/Kg,
a morfina foi administrada em um grupo com oito animais no 3º dia após injeção do
CFA, para avaliar a ação analgésica sobre todos os comportamentos, na dose de 4
mg/Kg em uma de 10 mg/5 ml de solução salina fisiológica 0,9%.
- MORFINA 4º dia após administração do CFA
Seguindo-se o mesmo protocolo do grupo testado com indometacina 5mg/Kg,
a morfina foi administrada em um grupo com oito animais no 3º dia após injeção do
CFA, para avaliar a ação analgésica sobre todos os comportamentos, na dose de
4mg/Kg.
- MORFINA 1º dia após administração do Formalina 2,5%.
A morfina foi testada 30 minutos antes da injeção do agente irritante, em um
grupo com seis animais, na dose de 4mg/Kg em uma diluição de 10mg/5ml de
solução salina fisiológica 0,9%, onde todos os parâmetros comportamentais foram
avaliados.
- LIDOCAÍNA COM FENILEFRINA 2% (NOVOCOL)
Nesse grupo a lidocaína foi administrada via intramuscula, na região do
músculo masseter, em um volume de 0,4 ml, dispensando o protocolo de jejum
necessário para os fármacos administrados por via oral.
Nesse grupo, os animais passavam por 10 minutos de adaptação ao
ambiente do observatório só então removidos para a câmara contendo éter etílico,
assim eram levemente anestesiados, para permitir a injeção de lidocaína na região

61
previamente sensibilizada por CFA e em seguida reconduzidos para caixa de
observação comportamental.
– LIDOCAÍNA no 3º dia após administração de CFA
Nesse grupo com oito animais foi testado o efeito da lidocaína sobre os
parâmetros comportamentais alterados no 3º dia após injeção de CFA.
– LIDOCÁINA no 4º dia após administração de CFA
Nesse grupo, com oito animais foi testado o efeito da lidocaína sobre os
parâmetros comportamentais alterados no 4º dia após injeção de CFA.
– LIDOCAÍNA no 1º dia após administração de Formalina 2,5%
Nesse grupo, com oito animais, a lidocaína foi administrada junto com a
solução de formalina 2,5% e todos os comportamentos foram imediatamente
analisados e registrados por 30 minutos.

62
2.5 -. ANÁLISE HISTOLÓGICA
Após a conclusão da curva temporal de cada comportamento, ou seja, no 7º
dia, os animais foram sacrificados, a região periarticular e articular foram removidas,
acondicionadas e fixados em formalina 10%, para avaliação do conteúdo
inflamatório dos grupos controle, CFA e formalina 2,5%.
2.6- ANÁLISE ESTATÍSTICA
Os resultados foram expressos como Média e Erro Padrão da Média (EPM).
As médias do tempo de execução de cada comportamento nos vários procedimentos
foram comparadas em teste paramétrico t – Student-Neuman-Keuls e o teste não
paramétrico Mann Whitney para análise dos dados comportamentais dos grupos
controles, formalina 2,5% e CFA.
O número de animais por grupo experimental está descrito nas figuras e a
significância mínima foi aceita com p<0,05.

63
III – RESULTADOS
3.1- CURVA TEMPORAL DOS COMPORTAMENTOS
Analisou-se neste trabalho 22 grupos de animais que foram observados
durante sete dias. Nos animais observados verificou-se que ao longo dos sete dias
houve mudanças significativas em alguns comportamentos.
- Curva temporal do comportamento sniffing avaliado durante 7 dia
O primeiro comportamento analisado no grupo de animais onde foi injetado o
CFA, o sniffing, não mostrou significativa (p<0,05) alteração no 1º (434,25+/- 41,91),
2º (468,25 +/- 65,67), 3º (720,13 +/- 75,23), 4º (817,13 +/- 65,43), 5º (644,25 +/-
112,49) e 7º (728 +/- 138,89) dia de observação em relação ao controle 1º (522,13
+/- 62,73), 2º (614,75), 3º (697,25 +/- 123,94), 4º (653,38+/-144,32), 5º (469,13 +/-
93,12), 7º (601,75 +/- 120,96). Entretanto, no 6º dia houve um aumento significativo
(p<0,05) do comportamento de farejar no grupo do CFA (705,38 +/-80,64) em
relação ao controle (318,75 +/- 58,58). Mas comparado a formalina 2,5%, não houve
alteração no tempo de execução desse comportamento no 1º (607,67 +/ 90,74), 3º
(675,67 +/- 89,1), 4º (658,5 +/-85,13), 5º (602,83 +/- 134,24), 6º (421,5 +/- 146,33),
7º (587,5 +/-203,92). Além disso, a formalina demonstrou uma atividade sobre esse
comportamento no 2º dia (1010+/- 113,65) em relação ao CFA (468,25 +/- 65,67) ao
grupo controle (614,75 +/- 120,68), ou seja, o tempo de execução desse
comportamento se apresentou reduzido no grupo do CFA em relação a formalina
2,5%.(fig. 1, tab.1)
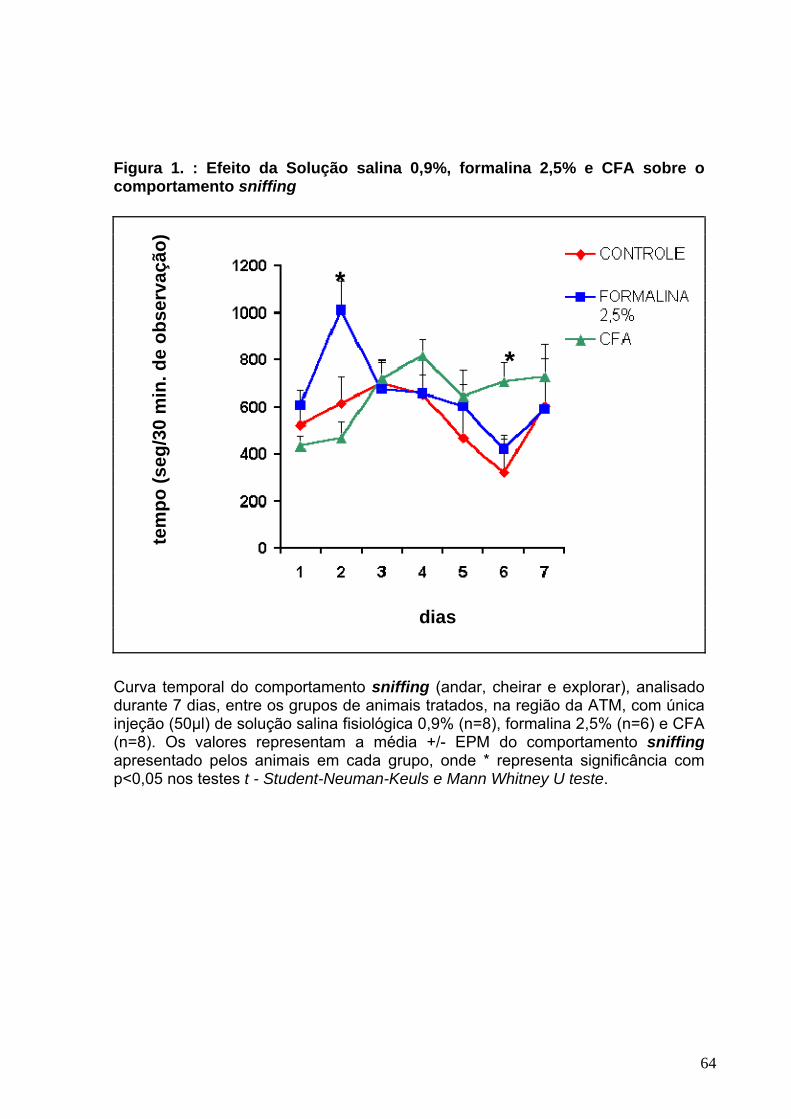
64
Figura 1. : Efeito da Solução salina 0,9%, formalina 2,5% e CFA sobre ocomportamento sniffing
tem
po (s
eg/3
0 m
in. d
e ob
serv
ação
)
dias
Curva temporal do comportamento sniffing (andar, cheirar e explorar), analisado durante 7 dias, entre os grupos de animais tratados, na região da ATM, com únicainjeção (50µl) de solução salina fisiológica 0,9% (n=8), formalina 2,5% (n=6) e CFA(n=8). Os valores representam a média +/- EPM do comportamento sniffingapresentado pelos animais em cada grupo, onde * representa significância comp<0,05 nos testes t - Student-Neuman-Keuls e Mann Whitney U teste.
*
*

65
Tabela 1 - Efeito da solução salina 0,9%, formalina 2,5% e CFA sobre o comportamento sniffing
DIA
GRUPO 1° 2° 3º 4° 5º 6º 7º
CONTROLE 522,13 ± 62,7(8) 614,75 ± 120,6 (8) 697,25 ± 123,9 (8) 653,38 ± 144,3(8) 469,13 ± 93,1(8) 318,75 ± 58,5 (8) 601,75 ±120,9 (8)
FORMALINA 2,5% 607,67 ± 90,7 (6) 1010 ±113,6 (6) * 675,67 ± 89,1 (6) 658,5 ± 85,1(6) 602,83 ±134,2 (6) 421,5 ± 146,3(6) 587,5 ± 203,9 (6)
CFA 434,25± 41,9 (8) 468,25 ± 65,6 (8) 720,13 ± 75,2 (8) 817,13 ± 65,4 (8) 644,25 ± 112,4 (8) 705,38 ± 80,6 (8)* 728 ± 138,8 (8)
Ratos Wistar (180 - 300g) foram tratados com uma única dose (50 µl) de solução salina 0,9%,formalina 2,5% e CFA. Os valores representam a média +/- EPM do tempo (s) de execução do comportamento sniffing apresentado pelo grupo de animais e o número de animais de cada grupo está entre parênteses, onde * representa significância estatística do CFA ou da formalina 2,5% em relação ao controle com p<0,05 nos testes t - Student-Neuman-Keuls e Mann Whitney U teste.

66
- Curva temporal do comportamento rearing analisado durante 7 dias
O comportamento rearing não foi alterado significativamente no grupo de
animais com CFA no 1º (3,25+/- 1,31), 2º (21,75 +/- 9,39), 3º (42,62 +/- 11,02), 4º
(34,75 +/- 6,65), 5º (36,75 +/- 10,76), 6º (44,12 +/- 10,48) e 7º dia (35,5 +/- 12,03) em
relação ao controle respectivamente no 1º (9,87 +/- 5,15), 2º (52,75 +/-18,01), 3º
(66,5 +/- 24,58), 4º (51,12 +/- 15,77), 5º (49,5 +/- 25,58), 6º (26,87 +/-11,68) e 7º dia
(36,12 +/- 11,42), Não houve alteração comportamental do grupo com CFA
comparado ao grupo de animais tratados com formalina 2,5% no 1º (11,33 +/- 5,32),
3º (32,83 +/- 14,11), 4º (33,83 +/- 11,52), 5 º (28,66 +/- 9,91) 7º dia (22 +/-6,3).
Contudo, as principais alterações significativamente (p<0,05) importantes foram, no
2º dia da formalina 2,5% (70,83+/- 15,93) em relação ao CFA (21,75 +/-9,39) e
superior e significativo tempo de execução do comportamento rearing no grupo do
CFA no 6º dia (44,12 +/-10,48) em relação a formalina (11,33 +/-6,38)(fig.2, tab.2).

67
Figura 2. : Efeito da Solução salina 0,9%, formalina 2,5% e CFA sobre ocomportamento rearing
tem
po (s
eg/3
0 m
in. d
e ob
serv
ação
)
dias
Curva temporal do comportamento rearing (empinar-se), analisado durante 7 dias, entre os grupos de animais tratados, na região da ATM, com única injeção (50µl)de solução salina fisiológica 0,9% (n=8), formalina 2,5% (n=6) e CFA (n=8). Os valores representam a média +/- EPM do comportamento rearing apresentado pelos animais em cada grupo, onde * representa significância com p<0,05 nostestes t - Student-Neuman-Keuls e Mann Whitney U teste.
*
*

68
Tabela 2 - Efeito da solução salina 0,9%, formalina 2,5% e CFA sobre o comportamento rearing
DIA
GRUPO 1° 2° 3º 4° 5º 6º 7º
CONTROLE 9,87 ± 5,1 (8) 52,75 ± 18 (8) 66,5 ± 24,5 (8) 51,12 ± 15,7 (8) 49,5 ± 25,5 (8) 26,87 ± 11,6 (8) 36,12 ± 11,4 (8)
FORMALINA 2,5% 11,33 ± 5,3 (6) 70,83 ± 15,9 (6)* 32,83 ± 14,1 (6) 33,83 ± 11,5 (6) 28,66 ± 9,9 (6) 11,33 ± 6,3 (6) 22 ± 6,3 (6)
CFA 3,25 ± 1,3 (8) 21,75 ± 9,3 (8) 42,62 ± 11 (8) 34,75 ± 6,6 (8) 36,75 ± 10,7 (8) 44,12 ± 10,4 (8)* 35,5 ± 12 (8)
Ratos Wistar (180 - 300g) foram tratados com uma única dose (50 µl) de solução salina 0,9%,formalina 2,5% e CFA. Os valores representam a média +/- EPM do tempo (s) de execução do comportamento rearing apresentado pelos animais em cada grupo e o número de animais de cada grupo entre parênteses, onde * representa significância estatística entre o grupo com CFA e o grupo da formalina 2,5% com p<0,05 nos testes t - Student-Neuman-Keuls e Mann Whitney U teste.

69
– Curva temporal do comportamento chewing like analisado durante 7 dias
O comportamento chewing like apresentou-se aumentado no grupo onde foi
administrado o CFA ao longo dos sete dias de observações comportamentais.
Entretanto, no 1º dia, não o fez significativamente (129,38 +/- 14,87) em relação ao
grupo controle (115,25 +/- 19,42) ou comparado a formalina 2,5% (114,83 +/-24,34),
como também no 2º (135,63 +/-18,95) e 4º dia (153,38 +/- 22,69) com o CFA em
relação ao grupo controle (105,25 +/- 22,72) e (99,75 +/- 32,79) respectivamente.
Entretanto, o CFA no 3º (142,25 +/- 14,94), 6º (190 +/- 33,3) e 7º dias (131,63 +/-
21,67) demonstrou um padrão significativamente maior em relação ao grupo de
animais controle no 3º (68,75 +/-9,42), 6º (59,62+/-14,66) e 7º dia (62,25+/-12,24)
respectivamente. Além disso, CFA apresenta tempo de execução significativamente
maior no 2º, 3º, 4º e 5º (124,13 +/- 20,69), 6º, 7º dia em relação ao grupo da
formalina com os respectivos valores para 2º (34 +/- 7,24), 3º (54,33 +/- 11,52), 4º
(53,33 +/- 12,09), 5º (25,66 +/- 9,26), 6º (16,83 +/- 7,49) e 7º dia (41,16 +/-
9,81).(fig.3, tab.3)

70
Figura 3. : Efeito da Solução salina 0,9%, formalina 2,5% e CFA sobre ocomportamento chewing-Like
tem
po (s
eg/3
0 m
in. d
e ob
serv
ação
)
dias
Curva temporal do comportamento chewing-like (movimento de rotação da mandibula ou mastigação), analisado durante 7 dias, entre os grupos de animaistratados, na região da ATM, com única injeção (50µl) de solução salina fisiológica0,9% (n=8), formalina 2,5% (n=6) e CFA (n=8). Os valores representam a média+/- EPM do comportamento chewing-like apresentado pelos animais em cada grupo, onde * representa significância em relação ao controle e # representasignificância em relação ao grupo da formalina, com p<0,05 nos testes t - Student-Neuman-Keuls e Mann Whitney U teste.
#
*
*# #
#
#
#

71
Tabela 3 - Efeito da solução salina 0,9%, formalina 2,5% e CFA sobre o comportamento chewing-Like
DIA
GRUPO 1º 2° 3º 4° 5º 6º 7º
CONTROLE 115,25 ± 19,4 (8) 105,25 ± 22,7 (8) 68,75 ± 9,4 (8) 99,75 ± 32,7(8) 77,75 ± 13,2 (8) 59,62 ± 14,6 (8) 62,25 ± 12,2 (8)
FORMALINA 2,5% 114,83 ± 24,3 (6) 34 ± 7,2 (6) 54,3 ± 11,5 (6) 53,33 ± 12 (6) 25,66 ± 9,2 (6) 16,83 ± 7,4 (6) 41,16 ± 9,8 (6)
CFA 129,38 ± 14,8 (8) 135,63 ±18,9 (8)# 142,25 ± 14,9(8)*# 153,38 ± 22,6 (8)# 124,13 ± 20,6 (8)# 190 ± 33,3 (8) *# 131,63 ±21,6 (8)*#
Ratos Wistar (180 - 300g) foram tratados com uma única dose (50 µl) de solução salina 0,9%,formalina 2,5% e CFA. Os valores representam a média +/- EPM do tempo (s) de execução do comportamento chewing like apresentado pelos animais em cada grupo, o número de animais de cada grupo entre parênteses, onde * representa significância estatística comparado ao grupo controle e # em relação ao grupo da formalina 2,5% com p<0,05 nos testes t - Student-Neuman-Keuls e Mann Whitney U teste.

72
– Curva temporal do comportamento climbing analisado durante 7 dias
De uma forma geral, o comportamento climbing apresentou-se reduzido no
grupo do CFA até o 3º (18,12 +/- 5,84) e 4º dia (14 +/- 3,85) em relação ao controle
3º (26,12 +/- 6,33) e 4º dia (19,25 +/- 4,26); ao passo que a partir do 5º dia (11,75 +/-
2,66) apresentou uma tendência de aumento no tempo de execução desse
comportamento, permanecendo aumentado no 6º (17,25 +/- 4,19) e 7º dia (16,12 +/-
6,32) em relação ao controle, tendo como valores para 5º (9,25 +/- 5,93), 6º (11,62
+/- 3,75) e 7º dia (12,5 +/-3,17). Mas, esse comportamento mostrou-se
significativamento reduzido (p<0,05) no 1º (4,37 +/-1,7) e 2º dia (7,75 +/-5,03) no
grupo do CFA tanto em relação ao grupo controle 1º (18,5 +/- 5,88), 2 º (22,37 +/-
8,02) e ao grupo da formalina 2,5% 1º (13,83 +/- 3,8) e 2º dia (46,33 +/- 6,72). É
importante destacar que, no grupo da formalina 2,5%, esse comportamento
apresentou uma diferença significativa (p<0,05) no 2º dia (46,33 +/- 6,72) quando
comparado ao grupo controle (22,37 +/- 8,02) (fig.4, tab.4).

73
Figura 4. : Efeito da Solução salina 0,9%, formalina 2,5% e CFA sobre ocomportamento climbing
tem
po (s
eg/3
0 m
in. d
e ob
serv
ação
)
dias
Curva temporal do comportamento climbing (subir em escada), analisado durante 7 dias, entre os grupos de animais tratados, na região da ATM, com única injeção (50µl) de solução salina fisiológica 0,9% (n=8), formalina 2,5% (n=6) e CFA (n=8).Os valores representam a média +/- EPM do comportamento climbingapresentado pelos animais em cada grupo, onde * representa significância emrelação ao controle e # representa significância em relação ao grupo da formalina,com p<0,05 nos testes t - Student-Neuman-Keuls e Mann Whitney U teste.
**
*
##

74
Tabela 4 - Efeito da solução salina 0,9%, formalina 2,5% e CFA sobre o comportamento climbing
DIA
GRUPO 1º 2° 3º 4° 5º 6º 7º
CONTROLE 18,5 ± 5,8 (8) 22,37 ± 8 (8) 26,12 ± 6,3 (8) 19,25 ± 4,2 (8) 9,25 ± 5,9 (8) 11,62 ± 3,7 (8) 12,5 ± 3,1 (8)
FORMALINA 2,5% 13,83 ± 3,8 (6) 46,33 ± 6,7 (6) * 34,33 ± 10 (6) 35,33 ± 10,5 (6) 17,5 ± 4,8 (6) 13,16 ± 6 (6) 33,66 ± 10,5 (6)
CFA 4,37 ± 1,7 (8) *# 7,7 ± 5 (8) *# 18,12 ± 5,8 (8) 14 ± 3,8 (8) 11,75 ± 2,6 (8) 17,25 ± 4,1 (8) 16,12 ± 6,3 (8)
Ratos Wistar (180 - 300g) foram tratados com uma única dose (50 µl) de solução salina 0,9%,formalina 2,5% e CFA. Os valores representam a média +/- EPM do tempo (s) de execução do comportamento climbing apresentado pelos animais em cada grupo, o número de animais de cada grupo entre parênteses, onde * representa significância estatística comparado ao grupo controle e # em relação ao grupo da formalina 2,5% com p<0,05 nos testes t - Student-Neuman-Keuls e Mann Whitney U teste.

75
–Curva temporal do comportamento freezing analisado durante 7 dias
O CFA apresentou em seu grupo de animais um tempo de execução superior
do comportamento de aparente congelamento, ou, freezing, destacado no 1º (812,38
+/- 100,07) e 2º dia (708,75 +/- 128,85) em relação grupo da formalina 2,5% 1º
(441,67) e 2º dia (202,33 +/- 66,86); no 4º dia o CFA (308,25 +/- 46,59) em relação
ao grupo controle (142 +/- 44,62) e 5º dia, O CFA apresentou um aumento
significativo (p<0,05) (380,38 +/- 81,77) em relação ao grupo controle (125,38 +/-
22,12) e a formalina 2,5% (121,17 +/- 22,63). No 3º, 6º e 7º dias não ocorreram
alterações comportamentais significativas (fig.5, tab.5).

76
Figura 5. : Efeito da Solução salina 0,9%, formalina 2,5% e CFA sobre ocomportamento freezing
tem
po (s
eg/3
0 m
in. d
e ob
serv
ação
)
dias
Curva temporal do comportamento freezing (congelar-se), analisado durante 7 dias, entre os grupos de animais tratados, na região da ATM, com única injeção(50µl) de solução salina fisiológica 0,9% (n=8), formalina 2,5% (n=6) e CFA (n=8).Os valores representam a média +/- EPM do comportamento freezing apresentado pelos animais em cada grupo, onde * representa significância em relação aocontrole e # representa significância em relação ao grupo da formalina, comp<0,05 nos testes t - Student-Neuman-Keuls e Mann Whitney U teste.
**
##
#

77
Tabela 5 - Efeito da solução salina 0,9%, formalina 2,5% e CFA sobre o comportamento freezing
DIA
GRUPO 1º 2° 3º 4° 5º 6º 7º
CONTROLE 772,88 ± 91,3(8) 538,25± 61,2 (8) 287,38 ± 86,4 (8) 142 ± 44,6 (8) 125,38 ± 22,1 (8) 213 ± 49,3 (8) 203,38 ± 48(8)
FORMALINA 2,5% 438,33 ± 97,4 (6) 202,33± 66,8 (6) 413,5± 84,2(6) 404,83 ± 33,4 (6) 121,17± 22,6 (6) 144,83 ± 42,8 (6) 211,17 ± 80,5(6)
CFA 812,38 ± 100 (8)# 708,75±128,8 (8)# 515,25± 79,4 (8) 308,25 ± 46,5 (8)* 380,38 ± 81,7(8)*# 271,75± 52,5 (8) 172,63 ± 32,3 (8)
Ratos Wistar (180 - 300g) foram tratados com uma única dose (50 µl) de solução salina 0,9%,formalina 2,5% e CFA. Os valores representam a média +/- EPM do tempo (s) de execução do comportamento freezing apresentado pelos animais em cada grupo, o número de animais de cada grupo entre parênteses, onde * representa significância estatística comparado ao grupo controle e # em relação ao grupo da formalina 2,5% com p<0,05 nos testes t - Student-Neuman-Keuls e Mann Whitney U teste.

78
– Curva temporal do comportamento rest/sleeping analisado durante 7 dias
O tempo de descanso e sono dos animais não apresentou mudanças
significativas no 1º dia nem no CFA (279,13 +/- 107,05), formalina 2,5% (323,5 +/-
162,86) e controle (168,88 +/- 44,17). No 2º dia, houve uma redução significativa do
sleeping no grupo da formalina (58,83 +/- 58,83) em relação ao grupo controle
(280,38 +/- 98,41). A partir do 3º dia de observação desse comportamento houve
uma redução significativa no grupo do CFA (69,37 +/- 33,62) tanto em relação ao
grupo controle (427,13 +/- 141,46) e a formalina 2,5% (317,83 +/- 97,91). No 4º não
houve alteração significante, porem reduzido com o CFA (180,75 +/- 64,81)
comparado ao controle (564,38 +/- 209,03) e a formalina (423,33 +/- 77,36). Ocorreu
ainda, no 5º dia, uma significativa redução no tempo de descanso e sono no grupo
de animais com CFA (332,38 +/- 161,51) em relação ao controle (703,75 +/- 164,49)
e ao grupo da formalina (902,83 +/- 178,4). No 6º dia, o sleeping destaca redução no
tempo de execução no grupo de animais do CFA (239,88 +/- 98,73) em relação a
formalina (1092,5 +/- 220,14) e ao controle (974,63 +/- 108,03). No último dia de
observação, no 7º dia, o comportamento permaneceu reduzido no grupo do CFA
(496,75 +/- 180,84) comparado ao grupo controle (554,38 +/- 166,39) e ao grupo da
formalina (733,33 +/- 244,35).(fig.6, tab.6).

79
Figura 6. : Efeito da Solução salina 0,9%, formalina 2,5% e CFA sobre ocomportamento sleeping
tem
po (s
eg/3
0 m
in. d
e ob
serv
ação
)
dias
Curva temporal do comportamento sleeping (dormir/descançar), analisado durante 7 dias, entre os grupos de animais tratados, na região da ATM, com única injeção(50µl) de solução salina fisiológica 0,9% (n=8), formalina 2,5% (n=6) e CFA (n=8).Os valores representam a média +/- EPM do comportamento sleepingapresentado pelos animais em cada grupo, onde * representa significância emrelação ao controle e # representa significância em relação ao grupo da formalina, com p<0,05 nos testes t - Student-Neuman-Keuls e Mann Whitney U teste.
*
*#
#
##
*

80
Tabela 6 - Efeito da solução salina 0,9%, formalina 2,5% e CFA sobre o comportamento sleeping
DIA
GRUPO 1° 2° 3º 4° 5º 6º 7º
CONTROLE 168,88 ± 44,1 (80 280,38 ± 98,4(8) 427,13 ±141,4(8) 564,38 ± 209(8) 703,75 ±164,4(8) 974,63 ± 108 (8) 554,38 ± 180,8(8)
FORMALINA 2,5% 323,5 ± 162,8 (6) 58,83 ± 58,8(6) 317,83 ± 97,9(6) 423,33 ± 77,3(6) 902,83 ±178,4(6) 1092,5 ± 220,1(6) 773,33 ± 244,3(8)
CFA 279,13 ± 107(8) 259,38 ± 91,5(8)# 69,37 ± 33,6 (8)*# 180,75 ± 64,8(8)* 332,38 ±161,5(8)# 239,88 ± 98,7(8)*# 496,75 ± 180,8(8)
Ratos Wistar (180 - 300g) foram tratados com uma única dose (50 µl) de solução salina 0,9%,formalina 2,5% e CFA. Os valores representam a média +/- EPM do tempo (s) de execução do comportamento sleeping apresentado pelos animais em cada grupo, o número de animais de cada grupo entre parênteses, onde * representa significância estatística comparado ao grupo controle e # em relação ao grupo da formalina 2,5% com p<0,05 nos testes t - Student-Neuman-Keuls e Mann Whitney U teste.

81
– Curva temporal do comportamento grooming analisado durante 7 dias
O comportamento de limpar-se ou grooming apresentou significativo (p<0,05)
aumento no seu tempo de execução no 1º (169,33 +/-29,5) e 2º (207 +/- 52,52) dia
no grupo da formalina 2,5% comparado ao grupo do CFA 1º (77 +/- 22,55) e 2º
(79,25 +/- 17,98) dia e ao grupo controle 1º (76,12 +/- 20,45) e 2º (77,62 +/- 23,03)
dia. A atividade do CFA em alterar o comportamento de limpar-se foi destacada
significativamente (p<0,05) no 4º dia (169,75 +/- 20,22) tanto em relação a formalina
2,5% (116,5 +/- 18,96) e ao grupo controle (101,5 +/- 17,57) e permanece
aumentado, mas sem significado estatístico no 5 º (122,25 +/- 26,33) e 6º dia
(110,25 +/- 15,77) (fig.7, tab.7).

82
Figura 7. : Efeito da Solução salina 0,9%, formalina 2,5% e CFA sobre ocomportamento grooming
tem
po (s
eg/3
0 m
in. d
e ob
serv
ação
)
dias
Curva temporal do comportamento grooming (limpar-se) entre os grupos de animais tratados, na região da ATM, com única injeção (50µl) de solução salinafisiológica 0,9% (n=8), formalina 2,5% (n=6) e CFA (n=8). Os valores representam a média +/- EPM do comportamento grooming apresentado pelos animais em cada grupo, onde * representa significância em relação ao controle, com p<0,05nos testes t - Student-Neuman-Keuls e Mann Whitney U teste.
*
*
*

83
Tabela 7 - Efeito da solução salina 0,9%, formalina 2,5% e CFA sobre o comportamento grooming
DIA
GRUPO 1° 2° 3º 4° 5º 6º 7º
CONTROLE 76,12 ±20,4 (8) 77,62 ± 23 (8) 135,63 ± 21,5 (8) 101,5 ± 17,5 (8) 94,87 ± 23,8 (8) 72,87 ±16,1 (8) 113 ± 20,8 (8)
FORMALINA 2,5% 169,3 ± 29,5(6) * 207 ± 52,5 (6) * 130,33 ± 29,7 (6) 116,5 ± 18,9 (6) 74 ± 28,2 (6) 67,5 ± 19,7 (6) 95,66 ± 33,9(6)
CFA 77 ± 22,5 (8) 79,25 ± 17,9 (8) 139,38 ± 17,7 (8) 169,75 ± 20,2 (8) * 122,25 ± 26,3 (8) 110,25 ± 15,7(8) 81,37 ± 19,7(8)
Ratos Wistar (180 - 300g) foram tratados com uma única dose (50 µl) de solução salina 0,9%,formalina 2,5% e CFA. Os valores representam a média +/- EPM do tempo (s) de execução do comportamento grooming apresentado pelos animais em cada grupo, o número de animais de cada grupo entre parênteses, onde * representa significância estatística comparado ao grupo controle e p<0,05 nos testes t - Student-Neuman-Keuls e Mann Whitney U teste.

84
– Curva temporal do comportamento licking analisado durante 7 dias
O comportamento de lamber-se ou licking, não apresentou alterações
comportamentais significantes até o 4º dia nos três grupos experimentais. No 5º dia,
o grupo do CFA (98,37 +/- 24,39) destacou alterações siginificativas (p<0,05) desse
comportamento em relação a formalina 2,5% (15 +/- 5,63). No 6º dia, o CFA também
evidenciou uma significativa (p<0,05) diferença (136,13 +/- 31,66) em relação ao
grupo da formalina (25,16 +/- 7,52) e ao controle (73,5 +/- 21,35). A formalina
demonstrou diferença significativa (p<0,05) na redução desse comportamento no 6º
dia (25,16 +/- 7,52) em relação ao grupo CFA (136,13 +/- 31,66) e no 7º dia, a
formalina também reduziu o tempo de execução (34,83 +/- 15,26) em relação ao
CFA (93,37 +/- 26,53) e ao controle (127,5 +/- 27,08) (fig.8, tab.8).

85
Figura 8. : Efeito da Solução salina 0,9%, formalina 2,5% e CFA sobre ocomportamento licking
tem
po (s
eg/3
0 m
in. d
e ob
serv
ação
)
dias
Curva temporal do comportamento licking (lamber-se), analisado durante 7 dias, entre os grupos de animais tratados, na região da ATM, com única injeção (50µl)de solução salina fisiológica 0,9% (n=8), formalina 2,5% (n=6) e CFA (n=8). Osvalores representam a média +/- EPM do comportamento licking apresentado pelos animais em cada grupo, onde * representa significância em relação aocontrole e # representa significância em relação ao grupo da formalina, comp<0,05 nos testes t - Student-Neuman-Keuls e Mann Whitney U teste.
*
#
#

86
Tabela 8 - Efeito da solução salina 0,9%, formalina 2,5% e CFA sobre o comportamento licking
DIA
GRUPO 1º 2° 3º 4° 5º 6º 7º
CONTROLE 54,62 ± 15 (8) 79,75 ± 30,8 (8) 92,5 ± 22,9 (8) 104,63 ± 27,7 (8) 109,5 ± 33,1 (8) 73,5 ± 21,3 (8) 127,5 ± 27 (8)
FORMALINA 2,5% 69,33 ± 27,2 (6) 30,83 ± 9 (6) 55,5 ± 19,2 (6) 38,5 ± 13,8 (6) 15 ± 5,6 (6) 25,16 ± 7,5 (6) 34,83 ± 15,2 (6)
CFA 53,12 ± 20,6 (8) 50,87 ± 18,3 (8) 96 ± 26,2(8) 70 ± 19,7 (8) 98,37 ± 24,3 (8)# 136,13 ± 31,6 (8)# 93,37 ± 26,5(8)
Ratos Wistar (180 - 300g) foram tratados com uma única dose (50 µl) de solução salina 0,9%,formalina 2,5% e CFA. Os valores representam a média +/- EPM do tempo (s) de execução do comportamento licking apresentado pelos animais em cada grupo, o número de animais de cada grupo entre parênteses, onde # representa significância estatística do grupo do CFA comparado ao grupo formalina 2,5% e p<0,05 nos testes t - Student-Neuman-Keuls e Mann Whitney U teste.

87
– Curva temporal do comportamento scratching analisado durante 7 dias
O comportamento scratching foi significativamente (p<0,05) aumentado no 1º
dia no grupo da formalina 2,5% (28 +/-11,14) comparado ao grupo do CFA (2,37 +/-
0,86) e ao grupo controle (2 +/- 0,9). Esse comportamento também apresentou um
significativo (p<0,05) aumento no grupo do CFA no 6º (13 +/- 3,15) e no 7 º dia (8,37
+/- 1,72) em relação formalina 2,5% no 6º (1 +/- 0,63) e 7º dia (1,33 +/- 0,84). Nos
outros dias de análise comportamental não houve alteração significativa importante
(fig.9, tab.9).

88
Figura 9. : Efeito da Solução salina 0,9%, formalina 2,5% e CFA sobre ocomportamento scratching
tem
po (s
eg/3
0 m
in. d
e ob
serv
ação
)
dias
Curva temporal do comportamento scratching (coçar-se), analisado durante 7 dias, entre os grupos de animais tratados, na região da ATM, com única injeção(50µl) de solução salina fisiológica 0,9% (n=8), formalina 2,5% (n=6) e CFA (n=8).Os valores representam a média +/- EPM do comportamento scratchingapresentado pelos animais em cada grupo, onde * representa significância emrelação ao controle e # representa significância em relação ao grupo da formalina,com p<0,05 nos testes t - Student-Neuman-Keuls e Mann Whitney U teste.
*
#

89
Tabela 9 - Efeito da solução salina 0,9%, formalina 2,5% e CFA sobre o comportamento scratching
DIA
GRUPO 1° 2° 3º 4° 5º 6º 7º
CONTROLE 2 ± 0,9 (8) 8,5 ± 2 (8) 9,12 ± 2,1 (8) 4,25 ± 1,2 (8) 11,75 ± 6,6 (8) 10,12 ± 5,7 (8) 7,87 ± 2,1 (8)
FORMALINA 2,5% 28 ± 11,1 (6) * 1,83 ± 0,8 (6) 6 ± 2,1 (6) 4,5 ± 1,3 (6) 1 ± 0,4 (6) 1 ± 0,6 (6) 1,33 ± 0,8 (6)
CFA 2,37 ± 0,8 (8) 2,37 ± 1,7 (8) 14,37 ± 6,8 (8) 7,62 ± 3,4 (8) 7,5 ± 3(8) 13 ± 3,1 (8) # 8,37 ± 1,7 (8)
Ratos Wistar (180 - 300g) foram tratados com uma única dose (50 µl) de solução salina 0,9%,formalina 2,5% e CFA. Os valores representam a média +/- EPM do tempo (s) de execução do comportamento scratching apresentado pelos animais em cada grupo, o número de animais de cada grupo entre parênteses, onde * representa significância estatística do grupo da formalina 2,5% comparado ao grupo do CFA e ao controle e # representa significância estatística do grupo com CFA em relação a formalina 2,5% com p<0,05 nos testes t - Student-Neuman-Keuls e Mann Whitney U teste.

90
– Curva temporal do comportamento biting analisado durante 7 dias
Avaliando a curva temporal do comportamento biting ou morder-se,
observamos que há uma ampla variação no padrão de execução desse
comportamento nos três grupos experimentais, sem destacá-lo significativamente
(p<0,05) em nenhum dos dias observados (fig.10, tab.10).

91
Figura 10. : Efeito da Solução salina 0,9%, formalina 2,5% e CFA sobre ocomportamento biting
tem
po (s
eg/3
0 m
in. d
e ob
serv
ação
)
dias
Curva temporal do comportamento biting (morder-se), analisado durante 7 dias, entre os grupos de animais tratados, na região da ATM, com única injeção (50µl)de solução salina fisiológica 0,9% (n=8), formalina 2,5% (n=6) e CFA (n=8). Os valores representam a média +/- EPM do comportamento biting apresentado pelos animais em cada grupo.

92
Tabela 10 - Efeito da solução salina 0,9%, formalina 2,5% e CFA sobre o comportamento biting
DIA
GRUPO 1° 2° 3º 4° 5º 6º 7º
CONTROLE 1,5 ± 1,1(8) 8,62 ± 4,4 (8) 14 ± 3,7 (8) 11,25 ± 5,4 (8) 13,12 ± 6,1 (8) 27,25 ± 11,6 (8) 15,25 ± 5,9 (8)
FORMALINA 2,5% 0,33 ± 0,33 (6) 6,83 ± 2,5 (6) 12 ± 8,7 (6) 21,33 ± 11,3 (6) 0,83 ± 0,83 (6) 1,83 ± 1,4 (6) 13,83 ± 9,4 (6)
CFA 1 ± 0,5 (8) 7,75 ± 3,3 (8) 11,5 ± 8,5 (8) 21,37 ± 7,8 (8) 18,87 ± 7,3 (8) 23,62 ± 10 (8) 16 ± 5,2 (8)
Ratos Wistar (180 - 300g) foram tratados com uma única dose (50 µl) de solução salina 0,9%,formalina 2,5% e CFA. Os valores representam a média +/- EPM do tempo (s) de execução do comportamento biting apresentado pelos animais em cada grupo, o número de animais de cada grupo entre parênteses.

93
– Curva temporal do comportamento head fliches analisado durante 7 dias
Assim como, no padrão de comportamentos biting, o movimento de sacudir a
cabeça ou head fliches também apresentou uma grande variação no seu tempo de
execução. Por isso, não houve alteração na apresentação desse comportamento no
1º, 2º, 3º, 4º, 5º e 7º dia de observação. Entretanto, no 6º dia houve um significativo
aumento (p<0,05) desse comportamento no grupo do CFA (6,5 +/- 2,98) em relação
a formalina 2,5% (0,33 +/- 0) (fig.11, tab.11).

94
Figura 11. : Efeito da Solução salina 0,9%, formalina 2,5% e CFA sobre ocomportamento head fliches
tem
po (s
eg/3
0 m
in. d
e ob
serv
ação
)
dias
Curva temporal do comportamento head fliches (sacudir cabeça), analisado durante 7 dias, entre os grupos de animais tratados, na região da ATM, com únicainjeção (50µl) de solução salina fisiológica 0,9% (n=8), formalina 2,5% (n=6) e CFA(n=8). Os valores representam a média +/- EPM do comportamento head flichesapresentado pelos animais em cada grupo, onde # representa significância emrelação ao grupo da formalina, com p<0,05 no teste Mann Whitney U teste.
#

95
Tabela 11 - Efeito da solução salina 0,9%, formalina 2,5% e CFA sobre o comportamento head fliches
DIA
GRUPO 1° 2° 3º 4° 5º 6º 7º
CONTROLE 6,62 ± 2,3 (8) 6,87 ± 2,87 (8) 4,25 ± 1,5 (8) 5,37 ± 1,9 (8) 3,5 ± 1,3 (8) 5,5 ± 2,3 (8) 7,87 ± 2,8 (8)
FORMALINA 2,5% 7,83 ± 2,9 (6) 9 ± 4,8 (6) 7,33 ± 4,9 (6) 3 ± 1,3 (6) 1 ± 0,8 (6) 0,33 ± 0,21 (6) 2,16 ± 1,2 (6)
CFA 7,83 ± 2,9 (8) 4,25 ± 1,5 (8) 2,62 ± 0,9 (8) 4,37 ± 2,3 (8) 7,12 ± 2,5 (8) 6,5 ± 2,9 (8) # 7 ± 2 (8)
Ratos Wistar (180- 300g) foram tratados com uma única dose (50 µl) de solução salina 0,9%, formalina 2,5% e CFA. Os valores representam a média +/- EPM do tempo (s) de execução do comportamento head fliches apresentado pelos animais em cada grupo, o número de animais de cada grupo entre parênteses, onde # representa significância estatística do grupo do CFA comparado ao grupo formalina 2,5% e p<0,05 nos testes t - Student-Neuman-Keuls e Mann Whitney U teste.

96
3.2 - SUMÁRIO DAS ALTERAÇÕES COMPORTAMENTAIS
Diante dos resultados obtidos pela avaliação de todos os comportamentos
dos animais, listamos quatro comportamentos se destacaram significativamente
estastístico (p< 0,05) no grupo de ratos tratados com o CFA, tanto em relação ao
grupo controle como também comparado ao grupo de animais tratados com
formalina 2,5%. A saber: chewing like (movimento de rotação da mandíbula ou
mastigação), rest/sleeping (descanso/sono), grooming (limpar-se) e freezing
(congelar-se). Contudo, além desses comportamentos, devemos listar alterações no
tempo de execução de outros comportamentos: sniffing (farejar), rearing (empinar-
se), climbing (subir escadas), licking (lamber-se), scratching (coçar-se) e head fliches
(sacudir a cabeça) em relação ao grupo controle ou em relação ao grupo formalina
2,5%. O comportamento biting (morder-se) não apresentou significância estatística
embora demonstre um aumento no tempo de execução em relação grupo da
formalina 2,5%. Devemos ressaltar também, que a formalina 2,5% injetada na região
da ATM dos ratos, alterou significativamente (p< 0,05) os comportamentos grooming
e scratching apresentados pelo seu grupo de animais tanto em relação ao CFA
quanto ao grupo salina fisiológica ou grupo controle.
Do ponto de vista temporal, as modificações significativas dos
comportamentos ocorreram no 3º e 6º dia (chewing like), no 3º dia (sleeping) e no 4º
dia após a inoculação do CFA na região da ATM (freezing e grooming). A formalina
destacou os comportamentos grooming e scratching ao 1 º dia avaliado após a
injeção desse agente algógeno.

97
3.3- ANÁLISE COMPARATIVA DA INTERFERÊNCIA DOS GRUPOS CONTROLES SOBRE OS COMPORTAMENTOS AVALIADOS.
Na figura 12 observamos que houve uma redução significativa no tempo de
execução do comportamento chewing like (p<0,05) no grupo normal (24,16 +/- 5,71)
comparado ao grupo controle com agulha sem solução salina fisiológica (86,33 +/-
17,54) e ao grupo controle com agulha e solução salina fisiológica 0,9%.
A figura 13 e 14 mostram que não houve interferência significativa (p<0,05)
dos grupos controles sobre o comportamento rest/sleeping.e grooming
O comportamento freezing mostrou umna sensibilidade sobre o tempo de
execução de seu comportamento comparando os grupos controles. Houve um
aumento significativo (p<0,05) desse comportamento no grupo controle agulha com
solução salina fisiológica (772,88 +/- 91,33) e grupo onde foi inserida apenas a
agulha na região da ATM dos ratos (554,5 +/- 62,64) em relação ao grupo normal
(150,83 +/- 59,31) (fig.15).

98
Figura 12.: Análise comparativa dos grupos controle experimentais emrelação ao comportamento chewing like alterado pela injeção de CFA na região da ATM de ratos.
Tempo de execução do comportamento chewing like (movimento de rotação da mandíbula) observado durante 30 minutos nos grupos controle com solução salina,controle normal e controle com inserção da agulha sem solução salina fisiológica0,9%. Os valores são expresso em média +/- EPM, * representa significância estatística, onde p<0,05 nos testes t-Student-Newman-Keuls e Mann Whitney U teste.
*

99
Figura 13.: Análise comparativa dos grupos controle experimentais emrelação ao comportamento sleeping alterado pela injeção de CFA na região da ATM de ratos.
Tempo de execução do comportamento sleeping (descanso/sono) observado durante 30 minutos nos grupos controle com solução salina, controle normal econtrole com inserção da agulha sem solução salina fisiológica 0,9%. Os valoressão expresso em média +/- EPM.

100
Figura 14.: Análise comparativa dos grupos controle experimentais emrelação ao comportamento grooming ou rubbing (coçar-se) alterado pela injeção de CFA na região da ATM de ratos.
Tempo de execução do comportamento grooming ou rubbing (coçar-se) observado durante 30 minutos nos grupos controle com solução salina, controlenormal e controle com inserção da agulha sem solução salina fisiológica 0,9%. Osvalores são expresso em média +/- EPM.
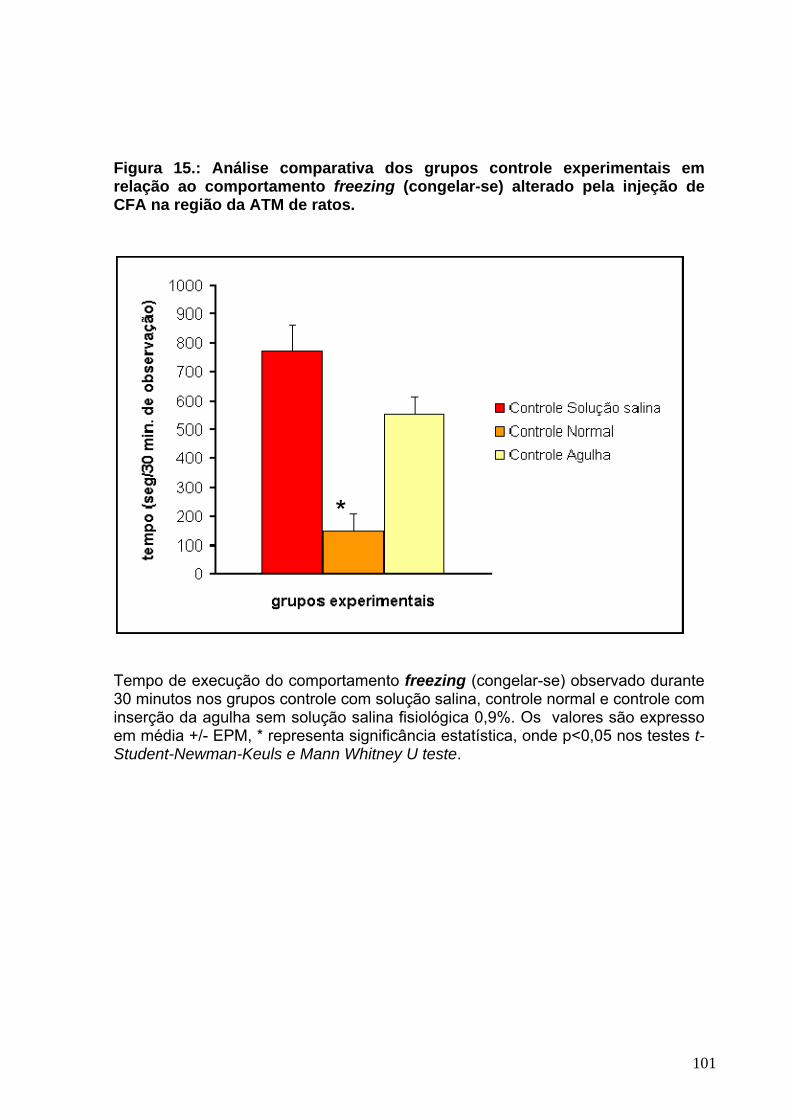
101
Figura 15.: Análise comparativa dos grupos controle experimentais emrelação ao comportamento freezing (congelar-se) alterado pela injeção de CFA na região da ATM de ratos.
Tempo de execução do comportamento freezing (congelar-se) observado durante 30 minutos nos grupos controle com solução salina, controle normal e controle cominserção da agulha sem solução salina fisiológica 0,9%. Os valores são expressoem média +/- EPM, * representa significância estatística, onde p<0,05 nos testes t-Student-Newman-Keuls e Mann Whitney U teste.
*

102
3.4 - ABORDAGEM FARMACOLÓGICA DOS COMPORTAMENTOS
Diante dos comportamentos destacados ao longo de sete dias de observação
comportamental foram definidos grupos experimentais para a abordagem
farmacológica do modelo experimental.
Esses grupos foram divididos de acordo com com o dia em que houve um
significativo aumento no tempo de execução após a injeção de CFA, formalina 2,5%
comparado a solução salina fisiológica.
Assim foram definidos os seguintes grupos experimentais:
§ Chewing like observado no 3° dia após a injeção de CFA
§ Rest/ Sleeping observado no 3° dia após a injeção de CFA
§ Freezing observado ao 4° dia após a injeção de CFA
§ Grooming observado no 4° dia após a injeção de CFA
§ Grooming observado no 1° dia após a injeção de formalina 2,5%.
§ Scratching observado no 1° dia após a injeção de formalina 2,5%.

103
– Efeito da administração de morfina, indometacina e lidocaína sobre o comportamento chewing like induzido pela CFA.
O comportamento chewing like (142,25 +/-14,94) (fig.12) induzido pelo CFA e
observado em nosso modelo ao 3º dia apresentou uma sensibilidade à ação da
morfina (4mg/Kg) (31 +/- 12,18) e a indometacina (5 mg/Kg) (77,37 +/- 18).
Entretanto, a lidocaína não causou nenhum efeito sobre o tempo de execução
chewing like (124,17 +/28,39) (fig.16, tab.12).
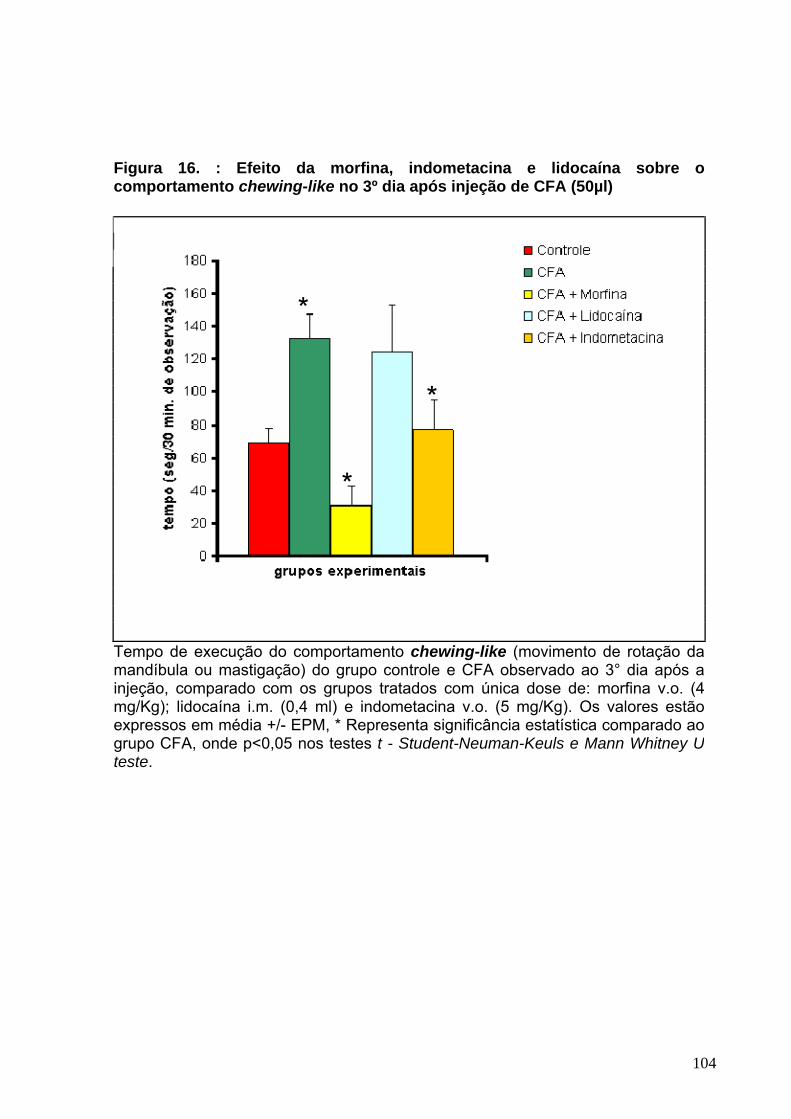
104
Figura 16. : Efeito da morfina, indometacina e lidocaína sobre ocomportamento chewing-like no 3º dia após injeção de CFA (50µl)
Tempo de execução do comportamento chewing-like (movimento de rotação da mandíbula ou mastigação) do grupo controle e CFA observado ao 3° dia após ainjeção, comparado com os grupos tratados com única dose de: morfina v.o. (4mg/Kg); lidocaína i.m. (0,4 ml) e indometacina v.o. (5 mg/Kg). Os valores estãoexpressos em média +/- EPM, * Representa significância estatística comparado aogrupo CFA, onde p<0,05 nos testes t - Student-Neuman-Keuls e Mann Whitney U teste.
*
*
*

105
- Efeito da administração de morfina, indometacina e lidocaína sobre o comportamento rest/sleeping induzido pela CFA.
A lidocaína aumentou significativamente o tempo de sono dos mesmos
animais desse grupo (428,67+/-129,91) (fig.13), assim como a indometacina
(5mg/Kg) (285,25 +/- 57,22) também apresentou significativo efeito analgésico
(fig.13) comparado ao grupo de animais onde foi injetado o CFA que não recebeu
tratamento analgésico (69,37 +/- 33,62). Porém, a morfina (4mg/Kg) nessa dose e
sobre esse comportamento e mesmo tendo aumentado o tempo de sono (170,67 +/-
111,14), não apresentou significativamente ação analgésica ou sedativa sobre o
tempo de sono dos animais (fig.17, tab.12).

106
Figura 17. : Efeito da morfina, indometacina e lidocaína sobre ocomportamento sleeping no 3º dia após injeção de CFA (50µl)
Tempo de execução do comportamento sleeping (dormir/descançar) do grupo controle e CFA observado ao 3° dia após a injeção, comparado com os grupostratados com única dose de: morfina v.o. (4 mg/Kg); lidocaína i.m. (0,4 ml) eindometacina v.o. (5 mg/Kg). Os valores estão expressos em média +/- EPM. * Representa significância estatística comparado ao grupo CFA, onde p<0,05 nostestes t - Student-Neuman-Keuls e Mann Whitney U teste.
*
*
*

107
- Efeito da administração de morfina, indometacina e lidocaína sobre o
comportamento freezing induzido pela CFA.
O comportamento freezing (309,5 +/- 46,66) (fig.14) não demonstrou
significativa sensibilidade a nenhum dos analgésicos testados, porém a morfina
(195,88 +/- 53,74) e indometacina (221,83 +/- 124,61) demonstraram a capacidade
de reduzir o tempo de execução desse comportamento avaliado, ao passo que a
lidocaína não reduziu o tempo de freezing nesse grupo de animais (399,83 +/- 71,63)
(fig.18, tab.12).

108
Figura 18. : Efeito da morfina, indometacina e lidocaína sobre ocomportamento freezing no 4º dia após injeção de CFA (50µl)
Tempo de execução do comportamento freezing (congelar-se) do grupo controle e CFA observado ao 4° dia após a injeção, comparado com os grupos tratados comúnica dose de: morfina v.o. (4 mg/Kg); lidocaína i.m. (0,4 ml) e indometacina v.o. (5mg/Kg). Os valores estão expressos em média +/- EPM. * Representa significância estatística comparado ao grupo controle, onde p<0,05 nos testes t - Student-Neuman-Keuls e Mann Whitney U teste.
*

109
- Efeito da administração de morfina, indometacina e lidocaína sobre o
comportamento grooming induzido pela CFA.
O grooming desenvolvido pelos animais submetidos à ação do CFA na ATM
avaliado ao 4 º dia (169,75 +/- 20,22), foi reduzido significativamente por todos os
analgésicos testados, a saber: indometacina (5 mg/Kg) (112,17 +/- 37,72) ; morfina
(4mg/Kg) (87,87 +/- 10,62); lidocaína (105,17 +/- 21,45), chegando ao de tempo de
execução desse comportamento apresentado no grupo de animais controle (101,5
+/- 17,57) (fig.19, tab.12).
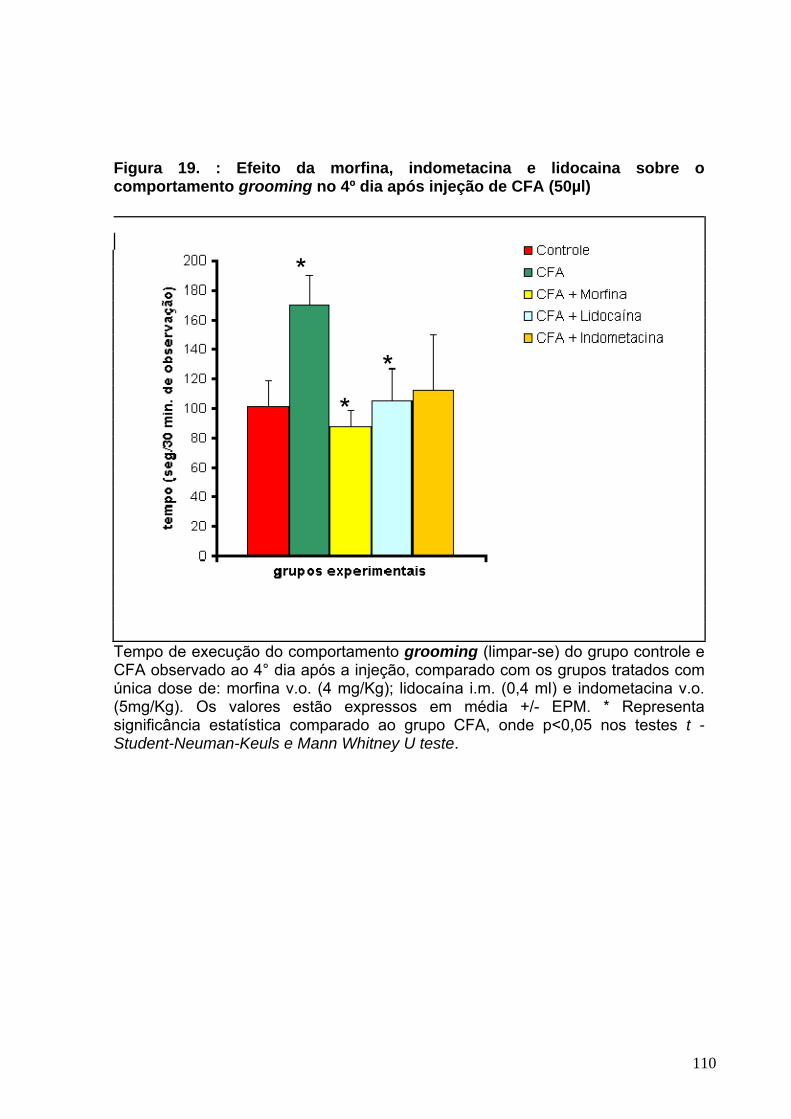
110
Figura 19. : Efeito da morfina, indometacina e lidocaina sobre o comportamento grooming no 4º dia após injeção de CFA (50µl)
Tempo de execução do comportamento grooming (limpar-se) do grupo controle e CFA observado ao 4° dia após a injeção, comparado com os grupos tratados com única dose de: morfina v.o. (4 mg/Kg); lidocaína i.m. (0,4 ml) e indometacina v.o.(5mg/Kg). Os valores estão expressos em média +/- EPM. * Representa significância estatística comparado ao grupo CFA, onde p<0,05 nos testes t -Student-Neuman-Keuls e Mann Whitney U teste.
**
*

111
- Efeito da administração de morfina, indometacina e lidocaína sobre o
comportamento grooming induzido pela formalina 2,5%.
Todos os analgésicos testados também apresentaram seus efeitos sobre o
comportamento grooming ou rubbing produzido pela atividade algógena da formalina
2,5% na região da ATM desse grupo de animais (fig.16) (169,33+/- 29,5). A
indometacina reduziu o tempo de execução desse comportamento induzido pela
formalina 2,5% (96,66 +/-35,96), mas signifivamente a morfina (4mg/Kg) (73 +/-
14,68) e lidocaína (64,33 +/- 11,19) destacaram seus efeitos (fig.20, tab.13).

112
Figura 20. : Efeito da morfina, indometacina e lidocaina sobre ocomportamento grooming no 1º dia após injeção de formalina 2,5% (50µl).
Tempo de execução do comportamento grooming (limpar-se) do grupo controle e formalina 2,5% observado por 30 minutos após a injeção, comparado com osgrupos tratados com única dose de : morfina v.o. (4 mg/Kg); lidocaína i.m. (0,4 ml)e indometacina v.o. (5 mg/Kg). Os valores estão expressos em média +/- EPM. * Representa significância estatística comparado ao grupo Formalina 2,5%, ondep<0,05 nos testes t - Student-Neuman-Keuls e Mann Whitney U teste.
**
*

113
- Efeito da administração de morfina, indometacina e lidocaína sobre o
comportamento scratching induzido pela formalina 2,5%.
O comportamento scratching desenvovido por ação da formalina 2,5% na
região orofacial (28 +/- 11,14), apresentou uma redução extremamente significativa
sob a ação da lidocaína (3,33 +/- 0) e da morfina (0,83 +/- 0,83) (fig.17), atingindo os
mesmos limites demonstrados pelo grupo controle (2 +/- 0,9). O inibidor não seletivo
de COX, a indometacina, nessa dose (5 mg/Kg) não apresentou atividade sobre
esse comportamento (31,83 +/- 15,45) (fig.21, tab.13).
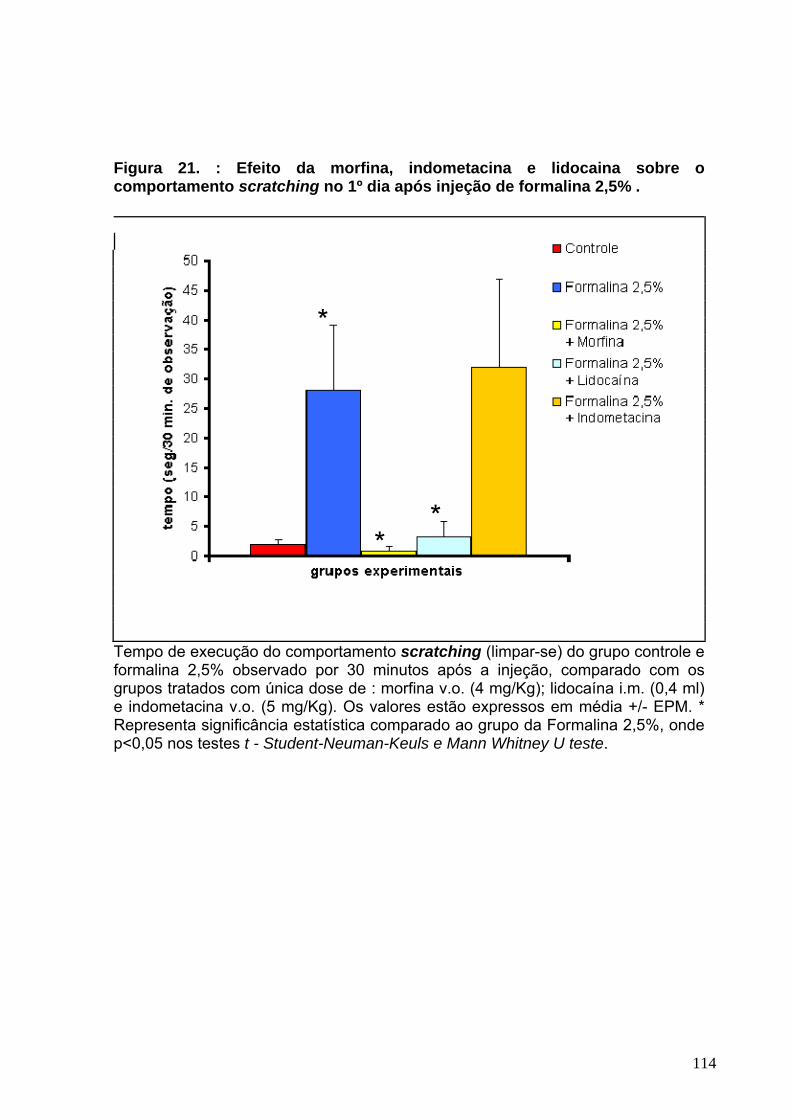
114
Figura 21. : Efeito da morfina, indometacina e lidocaina sobre ocomportamento scratching no 1º dia após injeção de formalina 2,5% .
Tempo de execução do comportamento scratching (limpar-se) do grupo controle e formalina 2,5% observado por 30 minutos após a injeção, comparado com osgrupos tratados com única dose de : morfina v.o. (4 mg/Kg); lidocaína i.m. (0,4 ml) e indometacina v.o. (5 mg/Kg). Os valores estão expressos em média +/- EPM. * Representa significância estatística comparado ao grupo da Formalina 2,5%, ondep<0,05 nos testes t - Student-Neuman-Keuls e Mann Whitney U teste.
**
*

115
3.5-SUMÁRIO DOS RESULTADOS FARMACOLÓGICOS
Em resumo, os comportamentos induzidos pela administração do CFA que
foram influenciados farmacologicamente foram: chewing like ((morfina (4mg/Kg, v.o.)
e indometacina (5mg/Kg, v.o.)); sleeping ((lidocaína i.m. e indometacina (5mg/Kg,
v.o.)) ao 3º após a injeção de CFA e grooming ((morfina (4mg/Kg, v.o.) e
indometacina (5 mg/Kg, v.o.)) no 4º dia após o tratamento com CFA (tab.12) O comportamento grooming induzido pela formalina 2,5% apresentou
sensibilidade à atividade analgésica da morfina (4mg/Kg, v.o.) e a lidocaína (i.m.),
enquanto que o scratching desenvolvido pela formalina 2,5% na ATM no 1º dia, foi
reduzido pela ação da morfina (4mg/Kg, v.o.) e pela lidocaína intramuscular (tabela
13).
É importante destacar que os outros comportamentos não foram alterados
pela atividade analgésica da indometacina (5mg/Kg, v.o.); morfina (4mg/Kg v.o.) ou
lidocaína nem no 1º, 3º ou 4º dia após a injeção dos agentes irritantes.

116
Tabela 12 - Efeito da morfina, indometacina e lidocaína sobre os comportamento alterados pela formalina
comportamento controle formalina morfina indometacina lidocaína
scratching 2,00 +/- 0,90 28,0 +/- 11,1 0,83 +/- 0,83 * 31,8 +/- 15,4 3,33 +/- 2,6 *
grooming 76,12 +/- 20,4 169,3 +/- 29,5 73,0 +/- 14,6 * 96,6 +/- 35,9 64,3 +/- 11,1 *
Ratos tratados com morfina (4mg/Kg), indometacina (5mg/Kg) administradas 30 minutos antes da observação, lidocaína (0,4 ml) administrada imediatamente antes da avaliação. O número de animais em cada grupo (n=6), os valores foram representados pela média +/- EPM do número de animais de cada grupo. Para análise estatística foram usados os testes t - Student-Neuman-Keuls e Mann Whitney U teste. * Representa diferença estatística (p<0,05) entre o grupo da formalina e grupo com tratamento analgésico.

117
Tabela 13 - Efeito da morfina, indometacina e lidocaina sobre os comportamento alterados pelo CFA
comportamento controle CFA morfina indometacina lidocaina
chewing like 68,75 +/- 9,4 142,25 +/- 14,94 31,00 +/- 12,18 * 77,37 +/ - 18,00 * 124,17 +/- 28,39
sleeping 427,13 +/- 141,4 69,37 +/- 33,60 170,67 +/- 111,10 285,2 +/- 57,20 * 428,6 +/- 129,90 *
freezing 142,00 +/- 44,6 308,20 +/- 46,50 195,8 +/- 53,70 221,8 +/- 124,60 399,8 +/- 71,60
grooming 101,50 +/- 17,5 179,70 +/- 20,20 87,8 +/- 10,60 * 112,1 +/- 37,70 105,1 +/- 21,40 *
Ratos tratados com morfina (4mg/Kg), indometacina (5mg/Kg) administradas 30 minutos antes da observação, lidocaína (0,4 ml) administrada imediatamente antes da avaliação. O número de animais em cada grupo (n=8), os valores foram representados pela média +/- EPM do número de animais de cada grupo. Para análise estatística foram usados os testes t - Student-Neuman-Keuls e Mann Whitney U teste. * Representa diferença estatística (p<0,05) entre o grupo CFA e o grupo que recebeu tratamento analgésico.

118
3.6- PESO CORPORAL DOS ANIMAIS
Fazendo uma análise comparativa da perda de peso dos animais que
sofreram lesões da região da ATM. Observamos que o grupo de animais submetidos
a solução salina e CFA na região orofacial não apresentaram uma diferença
significativa entre os valores de peso obtidos no 1º dia do experimento (332,33+/-
16,57) e os valores resultantes ao 7º dia após a lesão (336 +/- 14). Demonstrando
uma inabilidade do CFA em diminuir a freqüência de alimentação dos animais. Por
outro lado, a formalina apresentou capacidade (p ≤ 0,05) significativa em diminuir o
peso em seu grupo de animais, demonstrado pela diferença entre o peso inicial
registrado (314,5 +/- 4,06) e peso final (298,17 +/- 2,38) comparado com o grupo
controle que apresentaram um peso inicial (362+/- 17,48) e peso final (354+/- 17,48)
e com o grupo do CFA (fig.22, tab.14).

119
Figura 22. : Efeito da solução salina fisiológica 0,9%, CFA e formalina 2,5%sobre peso corporal dos animais.
Avaliação da alteração de peso em gramas no grupos experimentais após injeçãode 50µl de solução salina fisiológica, CFA e solução de formalina 2,5% na regiãoda ATM de ratos. * Representa diferença significante entre o peso inicial e pesofinal, onde p<0,05 nos testes t - Student-Neuman-Keuls e Mann Whitney U teste.
*

120
Tabela 14 - Efeito da solução salina fisiologica 0,9%, CFA e formalina 2,5% sobre peso corporal dos animais.
GRUPO peso inicial peso final
Controle 362 +/- 17,4 (8) 354 +/- 17,13 (8)
CFA 332,33 +/- 16,5 (8) 336 +/- 14 (8)
Formalina 2,5% 314,5 +/- 4 (6) 298,17+/- 2,3 (6) *
Avaliação da perda de peso (g) nos grupos experimentais após injeção de 50 μl de solução salina fisiológica (controle ), CFA e solução de formalina 2,5 % na região da ATM de ratos.Os valores representam a média +/- EPM entre parênteses. * Representa diferença significante entre o peso inicaial e o peso final, onde p< 0,05 nos testes t - Student-Neuman-Keuls e Mann Whitney U teste.

121
3.7- ANÁLISE HISTOLÓGICA
A avaliação histopatológica da região articular e periarticular da ATM dos
ratos onde foi injetada solução salina 0,9% demonstra sessões representadas por
músculo esquelético e tecido conjuntivo denso modelado (fig.23a e 23 b) e tecido
ósseo (fig.20) sem alterações inflamatórias ou em tecido hematopético
No grupo onde foi injetada formalina 2,5%, observa-se infiltrado inflamatório
linfocitário com focos no músculo esquelético (músculo masseter) (fig.23 c) e no
periósteo da ATM e tecido medular hematopoético sem alterações.
A ATM de ratos tratados com CFA apresenta alterações inflamatórias
linfomononuclear no intestício ou no citoplasma das fibras musculares (fig.23d e
23e), além de alterações degenerativas das mesmas e pequeno infiltrado
inflamatório periarticular. Vêem-se também trechos de tecido ósseo com medula
hematopoética sem alterações.

122
Figura 23: Avaliação histopatológica da região articular e periarticular da ATM de ratos.
a : Secção do músculo masseter (corado por HE) do grupo de animais tratados com solução
salina fisiológica 0,9%. Notar ausência de alterações inflamatórias nas fibras musculares.
b : Secção do ramo mandibular (corado por HE) do grupo de animais tratados com solução
salina fisiológica 0,9%. Notar ausência de alterações no periósteo e tecido hematopoetico.
c : Secção do músculo masseter (corado por HE) do grupo de animais tratados com solução
formalina 2,5%. Notar presençar de alterações inflamatórias nas fibras musculares.
d : Secção do músculo masseter (corado por HE) do grupo de animais tratados com CFA.
Notar presença de alterações inflamatórias degenerativas das fibras musculares.
e : Secção da mandíbula (corado por HE) do grupo de animais tratados com CFA. Notar
presença de alterações inflamatórias no periósteo, sem alterações na medula óssea.
b
c
d e
a

123
IV - DISCUSSÃO
O objetivo do presente trabalho foi elaborar um modelo de inflamação na
região da articulação temporomandibular de ratos, correlacionado-o com
padrões comportamentais alterados por injeção de Adjuvante Completo de
Freund na região periarticular.
Um dos principais aspectos relevantes em nosso trabalho foi o de
proprocionar um estudo experimental utilizando um estímulo nociceptivo com
características de inflamação crônica na região da ATM, pois as desordens da
ATM e região orofacial envolvem um componente inflamatório ou trauma
crônico (sessle, 2000). Além disso, não existem trabalhos envolvendo a relação
entre a dor orofacial crônica e comportamentos espontâneos.
Lesão na junção temporomandibular (ATM) pode causar reação
inflamatória nos tecidos circunvizinhos e região intracapsular. Esse processo
inflamatório envolve a mobilização de mecanismos de defesa, que ocorrem
tanto efeitos locais como sistêmicos e podem incluir mudanças
comportamentais e funcionais para regeneração e reparo dos tecidos. (Harper
e colaboradores, 2001).
As desordens temporomandibulares resultam em uma variedade de
sintomas incluindo dor persistente em áreas da mandíbula e região orofacial. A
severidade da dor pode ser resultado de alterações neuroplásticas no SNC
após lesões em tecidos periféricos (Dubner, 1991).
Estudo em modelo animal de dor persistente no plano medular espinhal
indicam que lesões em tecidos periféricos induzem a um estado de
hiperexcitabilidade que participa no desenvolvimento de dor persistente e
hiperalgesia (Dubner, 1991). Isso tem sido demonstrado que dor persistente na
região orofacial levam a um similar estado de hiperexcitabilidade no sistema
trigeminal.

124
O CFA tem sido utilizado como um agente indutor de inflamação e
hiperalgesia na pata de ratos. (Ladarola e colaboradores, 1988; Stein e
colaboradores,1988). Seguindo o mesmo princípio, Zhou e colaboradores
(1999) o utilizaram na região da ATM em modelo adaptado para região
orofacial (Imbe e colaboradores, 1999), para produzir inflamação e
hiperalgesia. Dor originada de tecidos em regiões como a ATM tem
características de hiperalgesia persistente e leva a forte hiperexcitabilidade de
neurônios do corno medular dorsal (MDR) (Iwata, 1999).
A injeção de CFA no espaço superior da ATM de ratos resulta em
profunda resposta inflamatória que pode se estende por 2 semanas (Imbe e
colaboradores, 2001); 6 semanas (Harper e colaboradores, 2001), seguindo um
padrão semalhante ao encontrado na estimulação com CFA e carragenina na
pata de ratos, onde o edema e hiperalgesia são evidenciados (Dubner e Ren,
2004).
Desordens na junção temporomandibular freqüentemente envolvem
componente inflamatório (Goulet e colaboradores, 1995). Mudanças na
articulação por indução de inflamação aguda têm sido extensivamente
estudadas e entre esses trabalhos Samsam e colaboradores (1999) relatam
que a inflamação na articulação temporomandibular leva a produção de
substancias pró-inflamatórias, incluindo taquicininas como a substancia P e
peptídeos como CGRP (Carleson e colaboradores, 1997; Spears e
colaboradores, 1998).
A citocina IL-1 beta tem sido implicada como o maior mediador
inflamatório em uma variedade de condições patológicas como artrite
reumatóide e doenças inflamatórias intestinais. Pacientes com desordens
temporomandibulares mostram uma alta incidência dos níveis de IL-1 beta no
fluido sinovial (Harper e colaboradores, 2000).
Essas citocinas demonstram uma atividade tempo e dose dependente,
pois Harper e colaboradores (2001) demonstraram, por injeção de CFA na ATM
de ratos, um incremento dose-dependente de IL-1 beta nas doses de 10, 25 e
50 microgramas, em que os níveis se apresentaram significativamente altos na

125
dose de 50 microgramas de CFA. Kerins e colaboradores (2003) verificaram
que 48 horas após injeção de CFA na ATM níveis de IL-1 beta foram mais
significativos que os níveis dessa citocina no grupo onde foi injetada solução
salina fisiológica.
Spears e colaboradores (2003) descrevem que a indução de inflamação
aguda na ATM de ratos resulta em incremento na concentração de TNF-ALFA.
Além disso, esse aumento promove o aparecimento de outras citocinas pró-
inflamatórias como IL-1 beta e Fator de crescimento do nervo (NGF).
Modelos experimentais de artrite têm avaliado os estágios do processo
inflamatório e estudos têm documentado mudanças morfológicas na membrana
sinovial (Nozawa-Inoue e colaboradores, 1998) ou no tecido periarticular
(Kapila & Xie, 1998) após indução de artrite experimental na ATM de animais.
Elvire e colaboradores (2001) investigando o efeito do processo
degenerativo na cartilagem demonstraram que a irritação por CFA nas fibras
nervosas promove mudanças na produção de PGEs e essa alteração na
síntese foi prevenida por antagonista de receptor para substância P, sugerindo
o envolvimento desse componente neurogênico nesse tipo de lesão.
Outros autores tentaram caracterizar o papel dos diferentes marcadores
neuroquímicos possivelmente associados com desordens
temporomandibulares dolorosas, como a serotonina, histamina,
prostaglandinas e fator de crescimento do nervo (NGF) na manutenção dessas
condições dolorosas (Kopp, 2001).
Em estudo avaliando a expressão do receptor da substância P sob ação
do CFA injetado em patas de ratos e sua internalização em dor aguda em curto
prazo (monoartrite)(3 dias) e longo prazo (poliartrite)(21 dias) observou-se uma
up-regulation do receptor para substância P em neurônios da lamina I da
medula espinhal no grupo tratado a longo prazo destacando a participação da
substância P no processo da nocicepção (Honoré e colaboradores, 1999).
Neuropeptídeos como CGRP e SP estão diretamente envolvidos na
inflamação neurogênica da articulação temporomandibular (Holmund e

126
colaboradores, 1991). Carleson e colaboradores (1997) relataram a função do
CGRP como mediador e modulador do processo inflamatório e verificaram um
incremento desse neuropeptídeo no fluido sinovial da ATM de ratos inoculados
com CFA. Estudos mostram que os neuropeptídeos estão amplamente
relacionados com a transmissão do estímulo doloroso das vias nociceptivas da
articulação temporomandibular.
A inflamação e hiperalgesia estão associadas com mudanças na medula
espinhal (Dubner & Ruda, 1992). Evidências indicam que o desenvolvimento da
hiperexcitabilidade espinhal e dor persistente envolvem a ativação de
receptores NMDA (Ren e colaboradores, 1992). O glutamato está colocalizado
com a substância P em aferentes terminais primários (De Biasi e Rustioni,
1988) e é liberado dentro da medula após ativação de receptores periféricos
(Sluka e colaboradores, 1994). Guo e colaboradores (2002) mostraram um
aumento dos níveis de receptor de NMDA (up-regulation) no corno dorsal da
medula espinhal em modelo de inflamação e correlacionaram este receptor
com o desenvolvimento da inflamação e hiperalgesia induzida por injeção
intraplantar de CFA.
Lawand e colaboradores (1997) estudaram o envolvimento desses
aminoácidos excitatórios na transmissão da nocicepção periférica em ratos
demonstrando o potencial papel dos receptores NMDA e não-NMDA na
transmissão nociceptiva. Uma vez que as respostas mediadas por receptores
NMDA são lentas e duráveis (Forsythe e Westbrook, 1988), uma conseqüência
da acentuação da resposta em neurônios de ratos tratados com CFA é um
longo potencial excitatório pós sináptico via medula espinhal. O receptor NMDA
ativado tem um papel importante no desenvolvimento e na manutenção da
hiperexcitabiliade central após lesão (Ren e colaboradores, 1992). A ativação
da proteína fos trigeminal após estímulos nocivos é atenuada por antagonistas
de receptor NMDA (Mitsikostas e colaboradores, 1998), sugerindo uma relação
do receptor NMDA com a sensibilização centrall trigeminal.
Zhou e colaboradores (1999) assim como Imbe e colaboradores (2001)
relatam a diferença entre a injeção de CFA em tecidos superficiais (tecido
perioral) e profundos (ATM) que resultam em comportamento de hiperalgesia e

127
alodinia (Dubner e Ren, 2004) com pico de 4-24 horas e persistência por duas
semanas. Comparando a injeção cutânea com a administração intra-articular
de CFA, foi relatado que a injeção intra-articulara) desenvolve uma inflamação
associada com seletiva up-regulation de preprodinorfina no complexo espinhal
trigeminal, além de uma acentuada hiperexcitabilidade no corno dorsal da
medula espinhal e grande expressão da proteína fos, um marcador de
atividade neuronal. Um incremento da atividade nas fibras C da ATM após a
inflamação e forte ativação neuronal central pode iniciar hiperexcitabilidade
central e contribuir para dor persistente associada com desordens
temporomandibulares. Ro (2003) e Guo e colaboradores (2002) relataram a
contribuição de receptores NMDA na nocicepção craniofacial e na formação do
edema após inflamação persistente com CFA.
Esses mediadores contribuem para um incremento na excitabilidade de
neurônios no corno dorsal da medula espinhal, levando ao que chamamos de
sensibilização central (Dubner e Ruda, 1992).
Esses estudos mostram que o CFA promove hiperalgesia através de
marcantes alterações que vão desde elevação das principais citocinas IL-beta,
TNF-alfa, NGF, substancia P e CGRP tanto localmente levando a uma
sensibilização periférica, tanto no SNC, passando por núcleo trigeminal,
aumentando a expressão da proteína fos também na medula espinhal e áreas
cerebrais.
Nosso trabalho sugere que há uma relação entre essas alterações
promovidas pelo potencial inflamatório do CFA intra-articular facial e baseada
em estudos prévios, correlaciona-o com as mudanças comportamentais
observadas ao longo do experimento.
De Castro-Costa e colaboradores (1987) em modelo de poliartrite com
CFA relata decréscimo no tempo de execução dos comportamentos: rearing
(empinar-se), running (correr) e climbing (subir escadas) e aumento nos
comportamentos: grooming (limpar-se), scratching (coçar-se), biting (morder-
se) e freezing (congelar-se) e ainda sugere que o comportamento scratching
(coçar-se) como marcador não invasivo de dor crônica. Carleson (2001) relata

128
o incremento da atividade scratching (coçar-se) em ratos submetidos a injeção
intra-articular de substância P.
O padrão de comportamento grooming (limpar-se) é relatado em vários
modelos experimentais (Clavelou e colaboradores, 1995; Einsenberg e
colaboradores, 1996) e mais recentemente por Roveroni e colaboradores
(2001), onde há incremento da atividade rubbing ou grooming (fricção da face
ou limpar-se), além de flinching ou head fliches (sacudir a cabeça) e chewing
like (mastigação ou movimento de rotação da mandíbula) e segundo Roveroni
e colaboradores (2001) a atividade máxima sobre esses comportamentos é
obtida na concentração de formalina 2,5%.
No presente trabalho foram observadas ao longo de uma semana,
alterações de comportamentos espontâneos (sniffing, rearing, chewing-like,
climbing, freezing, sleeping, grooming, licking, scratching, biting, head fliches)
através de uma única injeção de CFA na região da ATM de ratos.
No primeiro dia, não houve diferença significante nos comportamentos
entre os grupos de animais com o CFA e o grupo controle, possivelmente
houve interferência dos grupos controles (agulha sem solução e grupo controle
solução salina fisiológica) devido ao trauma proporcionado pela inserção da
agulha ou pelo volume da solução, desencadeando a ativação as fibras
nociceptivas, contrariando outros trabalhos (Roveroni e colaboradores, 2001)
onde a solução salina não apresentou nenhuma atividade ou não interferiu nos
resultados quando comparada no 1º dia de administração. Além disso, a
atividade inflamatória do CFA sobre comportamentos dos animais se revela
tempo-dependente (Zhou e colaboradores, 1999 e Imbe e colaboradores, 2001)
como foi esplanado anteriormente, as principais alterações inflamatórias,
mudanças neuroquímicas e ativação dos sistemas sensoriais nociceptivos
proporcionadas pela atividade do CFA, ocorrem a partir de 4 horas após a
injeção, podendo se estender por até 2 semanas. A 1ª observação foi realizada
nos primeiros 30 minutos após a injeção das soluções, não permitindo uma
diferenciação dos efeitos do CFA sobre a solução salina nessa etapa do
experimento.

129
Analisando a curva temporal de todos os comportamentos, notamos que
no grupo de animais onde injetamos o CFA, há um aumento de: chewing like
(movimento de rotação da mandíbula ou mastigação) (fig.3), grooming ou
rubbing (limpar-se ou fricção da ragião orofacial) (fig.7) e freezing (congelar-se)
(fig.5) e diminuição no tempo de sono (fig.6) dos animais tanto em relação a
formalina quanto ao controle.
Os comportamentos: sniffing (farejar) (fig.1), rearing (empinar-se) (fig.2) e
climbing (fig.4) mostraram algumas alterações significativas, mas de uma forma
geral, apresentaram-se diminuídos até o 5º dia aumentando o tempo de
execução a partir do 6º dia após a inoculação do CFA. O comportamento biting
(morder-se) (fig.10) não mostrou diferença significante durante toda a semana
de avaliação, pois esse comportamento pode não estar implicado com
alterações relacionadas a região orofacial ou o nosso modelo por um período
de 7 dias não permitiu a evidenciação desse comportamento.
Entretanto, esse experimento demonstrou um aumento do tempo de
execução dos comportamentos sniffing (farejar) (fig.1), scratching (coçar-se)
(fig.9) e licking (lamber-se) (fig.8) observado no 6º dia após a inoculação com
CFA, possivelmente relacionada a alterações comportamentais nociceptivas
ligadas a dor crônica (De Castro-Costa e colaboradores, 1987).
Há um incremento da imunoreatividade do canal de sódio em 24 horas
(Gould e colaboradores, 1998); IL-1 beta em 48 horas (Kerins e colaboradores,
2003), TNF-alfa em 48 horas (Spears e colaboradores, 2003), fenômeno de
internalização da substancia P no 3º dia após a administração intraarticular de
CFA. As principais alterações comportamentais ocorreram a partir do 3º dia
após a inoculação do CFA, corroborando e sugerindo que essas alterações no
plano neural refletem mudanças nos comportamentos que normalmente são
expressos em situações que não revelam ser de hiperalgesia.
De acordo com alguns autores (Harper e colaboradores, 2001; Kerins e
colaboradores, 2003; Kerins e colaboradores, 2004) em modelos utilizando o
CFA como agente indutor de dor na ATM, há uma alteração do comportamento
eating (comer ou alimentar-se) e que devido a dor gerada na ATM e região

130
orofacial durante a mastigação, o animal que necessita alimentar-se, promove
um alongamento no tempo de execução desse comportamento chewing ou
mastigação. Esse padrão de comportamento é observado com um
alongamento no padrão de duração desse comportamento de mastigação
(chewing) (movimento de rotação da mandíbula ou mastigação) (fig3), após
injeção bilateral de CFA na ATM de ratos machos e fêmeas (Kerins e
colaboradores. 2003) como discutido por Sternberg e Wachterman (2000) que
operacionalmente define o alongamento da média de duração de mastigação,
como um comportamento sugestivo de dor orofacial. Estudos clínicos de artrite
reumatóide juvenil com sintomas de doenças na ATM, revelam que a execução
de comportamento chewing é um hábito de suposta ‘defesa’ contra a dor
(Stohler e colaboradores, 1988).
Ogawa e colaboradores (2003) relatam por estudo em modelo animal
que comportamento mastigatório pode modular o processamento da dor com
relação a integração sensorial-motora via mecanismos corticais. E ainda,
relatam que o sistema sensorial trigeminal de alguma forma está envolvido na
modulação da dor crônica, mas o exato caminho neural ainda precisa ser
elucidado.
Por isso, movimento da mandíbula ou chewing like foi avaliado por todo
o experimento e mostrou significância a partir do 2º dia após a injeção do CFA,
mas destacou-se ao 3º e 6º dia comparando com os outros grupos
experimentais (fig.3), o que não ocorreu com solução salina nem com animais
injetados com formalina. Esse resultado demonstra a capacidade irritante
persistente do CFA, enquanto que a formalina não o fez. Hartwing e
colaboradores (2003) relatam episódios desse comportamento em modelo com
óleo de mostarda, relacionando esse comportamento à ação hiperalgésica em
tecidos como a ATM. No grupo da formalina observado em nosso modelo, no
3º e 6º dia, não houve alteração significativa desse comportamento, por isso,
nossos resultados corroboram ao que é relatado por Roveroni e colaboradores
(2001) que mostram em modelo de formalina na ATM de ratos que não há
diferença no tempo de execução do movimento de rotação da mandíbula
(chewing like) em relação ao controle com solução salina, mas contrariam

131
quando comparamos ao CFA, pois esse agente demonstrou uma atividade
sobre o chewing like desenvolvido pelos animais.
Estudos (Broton e Sessle, 1988; Yu e colaboradores, 1994) demonstram
que estímulos nocivos aplicados na região da ATM podem produzir sustentada
excitação de vários músculos mastigatórios incluindo o masséter e digástrico.
Ro e colaboradores (2003) relatam a contribuição dos receptores NMDA
(Cairns e colaboradores, 2001) na nocicepção do músculo craniofacial e na
formação do edema por óleo de mostarda. Essas alterações na musculatura
craniofacial podem contribuir para o aumento no tempo de execução desse
comportamento de mastigação ou rotação da mandíbula.
Tem sido demonstrado também que CFA injetado em patas de ratos
causa redução na eficiência do sono bem como na latência do primeiro
episódio de sono após o evento irritante (Andersen & Tufic, 2000).
O presente trabalho mostrou que o padrão de sono apresentado pelos
animais com CFA foi significativamente reduzido em relação ao grupo controle
e ao grupo da formalina (fig.6), podendo se estabelecer uma relação entre o
sono e a dor, semelhante aos resultados de Schutz e colaboradores (2003) que
observaram a ação do CFA na ATM de ratos por 2 dias e descrevendo haver
um decréscimo na eficiência do sono registrado nos animais, além de uma
fragmentação do sono e assim, demonstrado pelo número de vezes em que o
animal acordava. Nós observamos ao longo de uma semana que a partir do 3º
dia há manutenção do padrão de sono reduzido significativamente (p<0,05) em
relação ao controle e a formalina.
Rizzi e colaboradores (2004) relatam que dor músculo esqueletal
profunda ou a fibromialgia são freqüentemente associadas com queixas de
fadiga e má qualidade do sono. Foley e colaboradores (2004) e Stiefel (2004)
relacionam a qualidade do sono em humanos com a dor crônica.
A dor é um fator que tem sido reportado por levar a insônia na condição
clínica (Drewes e colaboradores, 1997). Estudos clínicos com artrite
reumatóide (Roizenblatt e colaboradores, 2001) e estudos animais com

132
modelos de poliartrite em ratos, confirmam a associação entre manifestações
dolorosas e distúrbios do sono.
De acordo com Zhang e colaboradores (2004) alguns
neurotransmissores como a substancia P, além de exercer funções biológicas,
contração muscular, excitação neuronal e transmissão de sinais dolososos, tem
sido implicada em uma ação modulatória na região ventro lateral pré-optica na
função do sono em ratos.
Um outro comportamento evidenciado pelo nosso trabalho foi grooming
ou rubbing (limpar-se ou fricção da região orofacial) (fig.7). Esse
comportamento é definido como uma atividade primária de importância para
sobrevida do animal que envolve formas de cuidado e de atenção com
superfície do corpo (McFarland, 1987). O animal utiliza o grooming ou ribbing
para defender-se de ectoparasitas e outros microorganismos (Svenson and
Randall, 1977). Em roedores, é essencial para termorregulação (Roberts e
colaboradores, 1974), e estende-se para fenômenos de sinalização social
(Melcer e Timberlake, 1985). Devida a multiplicidade de funções nesse
comportamento, o papel dessa resposta a irritantes tem importância para
estudos de dor experimental.
Estudos relatam que há uma diferença no padrão de grooming ou
rubbing evocado por irritantes locais ou estimulação dolorosa daquela
apresentada na manutenção da pelagem, termorregulação ou sinalização
social.
Prolongados grooming são observados após irritantes desde de
pequenos, moderados a intensos em regiões específicas do corpo (rubbing por
óleo mineral na pele), irritantes causados por fatores biológicos (ectoparasitas)
ou danos físicos na pele (Cohen e Price, 1979). Nesses casos, o grooming ou
rubbing corresponde a ações para tentar remover a causa da dor. (Vos e
colaboradores, 1998).
Em nosso modelo, o grooming ou rubbing apresentou significância no 4º
dia (fig.7) após inoculação com CFA, ao passo que a formalina apresentou sua

133
atividade sobre esse comportamento no 1º e 2º dia do experimento (fig.7),
sugerindo um comportamento tempo dependente relacionado a hiperalgesia
promovida pelas ações do CFA na ATM. Schutz e colaboradores (2003)
relatam a habilidade do CFA por injeção intra-articular mandibular de ratos e
Roveroni e colaboradores (2001) por injeção de formalina em promover
comportamentos nociceptivos como rubbing na região afetada.
Sherman e Kalin (1988) definem o comportamento freezing como uma
resposta adaptativa e freqüentemente induzida por estresse ou estímulo que
desenvolve o medo e associa a um modelo animal com ratos com a modulação
do sistema endógeno envolvendo a corticotropina como efetiva na alteração
desse comportamento. Em nosso trabalho, esse comportamento apresentou
alteração significante no 4º e 5º dia (fig.5) após a inoculação do CFA na ATM
dos animais comparada aos grupos controle e formalina. Também foi verificada
uma diferença significativa ao 1º, 2º e 5º dia em relação ao grupo da formalina
(fig.5). Harper e colaboradores (2001) identificaram o aumento de cortisona por
ação da IL-1 beta 48 horas após injeção de CFA na ATM de ratos e Van Der
Meer e colaboradores (1996) referem o aumento de corticosterona em
presença dessa citocina após injeções de CFA na ATM. A formalina em nossas
observações comportamentais apresentou uma redução significativa (p<0,05)
desse comportamento freezing em seu grupo de animais apresentado no 1º e
2° dia (fig.5).
Observando a curva temporal dos comportamentos, percebemos que há
uma tendência de restauração nos padrões de comportamentos entre os três
grupos experimentais a partir do 7 º dia. Nós sugerimos que possivelmente
ocorra uma ativação de áreas moduladoras, por exemplo, regiões como RVM
(Dubner e Ren, 2004), VLO (Zhang e colaboradores,1998) e DNIC (Danzinger
e colaboradores, 1999) com intuito de equilibrar os processos sensoriais e
comportamentais do animal. Vos e colaboradores (1998) relatam que o
estresse induzido no animal pode levar a ativação de mecanismo de controle
endógeno da dor e eixo pituitário adrenal, resultando em aparente aumento do
limiar de resposta dolorosa.

134
Como já foi esplanado anteriormente, a inflamação periférica tem sido
reconhecida por influenciar caminhos modulatórios, como por exemplo, uma
lesão inflamatória persistente pode acentuar inibição ou facilitação da
transmissão nociceptiva através da medula (Terayama e colaboradores, 2000).
Hurley e Hammond (2000) demonstraram que a potência de agonistas
opióides por microinjeções na região rostroventromedial bulbar (RVM) é
acentuada de maneira tempo dependente após a injeção de CFA na pata de
ratos. Guan e colaboradores (2003) relatam uma up-regulation na expressão
de receptores AMPA em circuitos modulatórios induzido por inflamação pelo
CFA e que a continuação desse fenômeno pode contribuir para a plasticidade
atividade-dependente.
Além disso, Honoré e colaboradores (1999) demonstram em modelos de
poliartrite que o CFA apresenta dois picos onde a substância P é evidenciada
com a hiperalgesia, a saber, o 3º dia e 21º dia. Por isso, provavelmente nosso
modelo poderia apresentar novas alterações comportamentais se houvesse um
alongamento dos dias em que foram feitas as análises comportamentais, por
exemplo, por 30 dias, nesse caso um modelo clássico de dor crônica.
Para comparar a curva temporal do modelo com CFA, injetamos
formalina 2,5% seguindo os experimentos feitos por Roveroni e colaboradores
(2001) que relatam aumento do tempo de execução do rubbing e não alteração
em chewing-like e head fliches, relacionando-o a atividade algógena da
formalina adaptada para tecidos da região orofacial por Clavelou e
colaboradores (1989).
Em nosso experimento, a formalina no 1º dia apresentou diferença
significante apenas no comportamento scratching (coçar-se) (fig. 9) e grooming
ou rubbing (limpar-se) (fig.7) em relação tanto ao CFA quanto ao controle. Esse
padrão de comportamento é provocado por irritação local e tem uma
organização diferente da relatada para manutenção da pelagem,
termorregulação ou sinalização social e neste caso, a atividade ocorre mais
especificamente na área do corpo relacionada (Vos e colaboradores,1998),
além de aparentemente tentar remover a causa da dor.

135
A ausência de comportamentos também relacionados como head fliches
e chewing like pode ser devido a concentração usada (2,5%), pois segundo
Lee e colaboradores (2000), movimento de sacudida da pata e freezing
observada nos testes com formalina é evidenciado quando utilizamos a
concentração de 5 %, enquanto que o grooming ou rubbing poderá ser melhor
evidenciado na concentração de 2,5%. Por isso, alguns comportamentos
poderão ser dependentes da concentração utilizada da formalina.
Pelo fato que, no 1º dia de observação, não ter havido diferença
significativa de comportamentos como: freezing, sleeping, grooming e chewing
like entre os grupos experimentais CFA e controles estudados, nós testamos se
poderia ter havido interferência do trauma da agulha ou do volume de solução
salina fisiológica sobre os resultados obtidos com grupos experimentais.
Por isso, noutro experimento fizemos uma análise comparativa dos
comportamentos em animais divididos em três grupos controle: grupo normal;
grupo com inserção da agulha, mas sem volume de solução e grupo com
injeção da solução salina fisiológica. Observamos que há uma diferença
significativa do grupo normal referente ao parâmetro chewing like e freezing
(fig.12, fig.15), sugerindo que pelo menos parcialmente, há uma interferência
do veículo e da injeção na evidenciação dos comportamentos elicitados pela
formalina 2,5%. Esse mesmo padrão de resposta da solução controle foi
observado por Roveroni e colaboradores (2001) no teste da formalina na ATM
de ratos. Nos outros comportamentos não foram observados interferências
significativas (fig.13, fig.14).
Do 2º ao 7º dia, com exceção do comportamento rearing (fig.2) e
climbing (fig.4) (2º dia) a formalina não apresentou nenhuma outra alteração
comportamental, concordando com relatos de que a formalina tem atividade
irritante geralmente avaliada por até 60 minutos (Capone & Aloisi, 2004) e esta
é dividida em duas fases: primeira (0-10 minutos) e a segunda (10-60 minutos).
Essa característica bifásica de resposta comportamental (Tjolsen e
colaboradores, 1992) é atribuída primeiro a atividade que reflete a ação direta
nos aferentes sensoriais (fibras A delta e C), enquanto que a segunda fase
reflete a continuação da ação nociceptiva periférica de mediadores

136
inflamatórios como: bradicinina, TNF-alfa, IL-1 BETA, IL-6 ,IL-8 (Chichorro e
colaboradores, 2004) e sensibilização central (Hunskaar e Hole, 1987).
A atividade de drogas classicamente analgésicas como a lidocaina,
indometacina e morfina, em bloquear esses comportamentos sugestivos de
dor, foi avaliada para caracterizar farmacologicamente o nosso modelo e
correlacionar os possíveis efeitos analgésicos aos comportamentos
nociceptivos.
A lidocaína possui efeito antinociceptivo tanto profilático quanto
terapêutico na dor induzida pela formalina em camundongos. (Chichorro e
colaboradores, 2004), atenua a resposta sacudir a cabeça por injeção de óleo
de mostarda na face (Ro e colaboradores, 2003), mas falha na prevenção do
edema produzida por esse irritante devido a não capacidade de inibir os
mediadores neurogênicos, além de reduzir o comportamento fliching e rubbing
induzido por formalina 2,5% na ATM (Roveroni e colaboradores, 2001). Em
nosso trabalho, a lidocaína foi efetiva em reduzir o efeito do CFA na atividade
grooming (fig.19) e na atividade sleeping (fig.17), mas não em reduzir o tempo
de execução de chewing like (fig.16) e freezing (fig.18). Esse anestésico
aplicado localmente na ATM dos ratos reduziu tanto o scratching (fig.21)
quanto o grooming (fig.20) produzido pela injeção da formalina 2,5%.
Essa inabilidade de a lidocaína bloquear completamente o efeito
nociceptivo do CFA em todos os comportamentos pode ser devido ao efeito da
lidocaína restrigir-se apenas perifericamente e alguns desses comportamentos
podem estar relacionados a alterações além do estímulo local possivelmente
nesse estágio do experimento já pode haver algum comprometimento ou
sensibilização na medula espinhal ou no SNC.
Além disso, segundo Sakaue e colaboradores (2004), os agentes
anestésicos locais, têm uma ação preferencial antinociceptiva contra
estimulação térmica e servem para diferenciar nocicepção térmica de
mecânica. Em nosso trabalho, a dor ou hiperalgesia foi estimulada por agente
pró-inflamatório persistente e portanto, não ofereceu uma situação adequada
para demonstrar ou testar o efeito analgésico da lidocaína.

137
Na ordem de investigar o papel do incremento de COX na resposta
comportamental, o efeito da indometacina foi testado na ação inflamatória do
CFA. A indometacina diminuiu significativamente o movimento de rotação da
mandíbula (fig.16), aumentou o tempo de sono (fig.17), embora tenha reduzido,
porém sem diferença significativa devido a grande variação comportamental
apresentada pelo grupo, o tempo de execução do comportamento grooming e
freezing estimulados pela inflamação provocada pelo CFA. A indometacina
também apresentou seu efeito analgésico, reduzindo o grooming estimulado
pela formalina, mas não o fez com significância estatística. Também se
mostrou ineficaz nos parâmetros scratching testados pela formalina intra-
articular. Esse efeito atribuído aos analgésicos inibidores da síntese de
prostraglandinas e muitos estudos têm implicado as prostaglandinas como uma
das mediadoras do ciclo sono-virgília em animais. (Ueno e colaboradores,
1983). Nossos resultados corroboram com os apresentados pelo trabalho de
Shultz e colaboradores (2003), onde a indometacina na dose de 5 mg/Kg
restabeleceu o padrão de sono alterado por injeção de CFA na ATM de ratos.
No teste com formalina, a indometacina intrigantemente não mostrou
atividade significativamente analgésica sobre o grooming (fig.19), embora
tenha reduzido o tempo de execução de tal comportamento. Em relação ao
comportamento scratching, a indometacina, nessa dose e via de administração
não apresentou nenhum efeito importante. Choi e colaboradores (2003) relatam
que a COX-2 tem um papel importante na hiperalgesia mediada por formalina
em tecidos orofaciais e nesse modelo, a indometacina reverteu o
comportamento scratching promovido pelo irritante.
A falha do efeito antinociceptivo da indometacina em alguns desses
parâmetros comportamentais pode ser atribuído a vários fatores.
A via de administração, tempo de administração (testamos o efeito
profilático e não terapêutico) e a dose utilizada (5mg/Kg) pode não ter sido
suficiente para suplantar o efeito nociceptivo provocado pela inflamação após
injeção de CFA. Além disso, outros trabalhos confirmam os efeitos da
indometacina em reverter a resposta dolorosa da formalina e de outros
irritantes como: carragenina e CFA em ratos (Tatsuo e colaboradores, 1994).

138
Pitcher e Henry (1999) citam que há uma relação dose dependente (4-8 mg/Kg)
no efeito analgésico da indometacina em resposta de neurônios do corno
dorsal por estimulação elétrica e cutânea. Abbott e Helleman (2000) destacam
uma diferença significativa da ação analgésica da indometacina entre as vias
sistêmicas e intracerebroventricular.
Uma segunda hipótese pressupõe que alguns desses comportamentos
podem não estar relacionados somente a via de produção das ciclooxigenases.
Além disso, a indometacina inibe a COX, mas deixa livre o ácido araquidônico
e via da lipooxigenase que poderá ser usada como um caminho alternativo
para aferentes nociceptivos, contribuindo para a persistência de dor como
também de comportamentos nociceptivos alterados. (Vaughan e
colaboradores, 1997; Pitcher e Henry, 1999).
O efeito central dos opióides têm sido estudado na transmissão da dor
por ações que envolvem o corno dorsal da medula espinhal e outros sítios
supraespinhais. É conhecido que o receptor opióide também está presente nos
terminais periféricos das fibras cutâneas mielinizadas e não mielinizadas
(Coggeshall e Carlton, 1997).
Os efeitos farmacológicos dos analgésicos opióides são derivados de
complexas alterações nos vários tipos de receptores opióides e a morfina é um
agonista mu receptor (Inturrisi, 2002). Esse opióde mostra um efeito sobre a
liberação de aminoácidos excitatórios, prostaglandinas (PGE2) na medula
espinhal evocado por injeção de formalina (Malmberg e Yaksh, 1992).
Roveroni e colaboradores (2001) mostram a redução da resposta fliching
e rubbing induzida por formalina sem afetar a atividade locomotora. Esses
resultados podem ser comparados a observações do teste da formalina
injetada no lábio superior em que a morfina reduz a atividade rubbing, sem
aboli-la.(Clavelou e colaboradores, 1989).
Doses de morfina sistêmica mostram-se efetivas em reduzir
comportamentos nociceptivos produzidos por óleo de mostarda em modelo de
dor muscular craniofacial (Ro e colaboradores, 2003), enquanto que Hartwing e

139
colaboradores (2003) relatam que a morfina administrada sistemicamente
resulta em atenuação dose dependente da inflamação e da resposta
nociceptiva (chewing like, grooming e head shaking) promovida por óleo de
mostarda na região da ATM de ratos além de reduzir a expressão da proteína
fos no subnúcleo caudal desses mesmos animais.
O efeito analgésico da morfina foi observado nos comportamentos
chewing like (fig.16) e grooming (fig.19), ao passo que o sleeping (fig.17) e
freezing (fig.18) estimulados pelo CFA na ATM de ratos, não foram alterados.
Portanto, não evidenciou efeito sedativo dos analgésicos opióides.
Provavelmente esses comportamentos poderiam ser abolidos com doses
maiores que as por nós utilizadas.
Os parâmetros scratching (fig.21) e grooming (fig.20) aumentados pela
formalina foram significativamente reduzidos sugerindo que há um componente
central envolvido no processamento desses comportamentos.
Harper e colaboradores (2001) relatam mudanças na freqüência de
alimentação e no peso corporal e mostram que a injeção de doses de CFA na
ATM diminui a freqüência de alimentação por até cinco dias, seguindo-se por
uma super consumo (rebote) que perdurou até o 7º dia. Os autores relatam
uma possível correlação com alterações no processo de mastigação. Shinoda
e colaboradores (2003) observaram um decréscimo no peso corporal a partir
do 6º dia após injeção de CFA na ATM relacionando-o com a dor devida a
artrite na ATM. Kerins e colaboradores (2003) descrevem também no mesmo
modelo que inicialmente há um aumento no tempo para o início de consumo de
alimentos (24 horas) retornando aos padrões normais até 48 horas. Os autores
revelam que esses padrões podem servir indiretamente para diferenciar a
magnitude da inflamação, pois a média do tempo de alimentar-se pode
alongar-se de acordo com a dose de CFA utilizada.
Em nosso experimento, não houve uma diferença significativa no peso
corporal final dos animais entre os grupos injetados com solução salina e
tratados com CFA. Essa aparente ausência de efeito do CFA sobre o peso final
nesse grupo de animais pode ser devido a três fatores: 1- efeito rebote descrito

140
por Harper e colaboradores (2001), levando a um aumento o consumo de
alimentos pelos animais após a injeção de CFA; 2- alongamento do tempo de
mastigação ou chewing like ocasionado pela dor e pelo aumento no consumo
de alimentos para promover a ativação de sistemas modulatórios endógenos
(Sternberg e Wachterman (2000) e (Kerins e colaboradores, 2004), levando ao
aumento no consumo de alimentos pelos animais; 3- o tempo de avaliação não
foi suficiente para evidenciar a perda de peso provocada pela inflamação e dor
nos tecido que compõe a ATM dos ratos. Zamma (1983) relata em modelo de
poliartrite com o CFA em ratos que apresentavam a doença na ATM, uma
redução do peso corporal ao longo de 5 semanas. Intrigantemente, a formalina
foi hábil em reduzir o peso corporal final em seu grupo de animais (fig.22).
Finalmente, sessões histológicas de tecidos da região da articulação
temporomandibular 48 horas após CFA nessa região, demonstram um infiltrado
inflamatório agudo incluindo a presença de neutrófilos e macrófagos com o
tecido intracapsular que varia a intensidade de acordo com a dose de CFA
utilizada. (Harper e colaboradores, 2001).
Comparando a evolução do quadro inflamatório da carragenina e da
formalina, observou-se a reação inflamatória de ambos os grupos
experimentais iniciada com um infiltrado inflamatório agudo, tornando-se misto
e depois crônico. Após 15 dias, um discreto infiltrado celular inflamatório foi
encontrado apenas no grupo da formalina. Sinais de hiperplasia da membrana
sinovial foram observados aos três dias, intensos aos 7 dias, estando presente
aos 15 dias somente no grupo da formalina (Goulart, 2002).
Para Zamma (1983) o aspecto histológico em seu modelo de resposta
inflamatória na ATM de ratos, caracteriza-se por média sinovite e hipertrofia
vilosa da cartilagem articular e do disco articular.
Nossos resultados histopatológicos mostram que há uma diferença entre
os tecidos articulares e periarticulares dos animais tratados com solução salina
fisiológica, CFA e formalina 2,5%. Nos tecidos dos animais do grupo controle
observa-se que no 7º dia, não houve alteração inflamatória no tecido muscular
esquelético ou tecido ósseo (fig.23). No entanto, na região muscular do tecido

141
tratado com formalina 2,5%, verificamos alterações inflamatórias apresentando
conteúdo linfocitário possivelmente promovido pela atividade irritante do formol
sobre tecidos celulares (fig.23). Entretanto, o tecido inoculado pelo CFA
apresentou o mesmo conteúdo linfocitário, porém mais acentuado em relação
ao grupo da formalina 2,5%, com invasão do interstício e citoplasma das fibras
musculares e fenômeno degenerativo das mesmas (fig.23), além de pequena
inflamação do periósteo, mas sem mudanças no tecido hematopoético (fig.23).
Esse resultado demonstra a capacidade do CFA em produzir inflamação
persistente e sugerimos portanto, ser colaboradora para alterações
comportamentais apresentadas pelo CFA comparado aos comportamentos
produzidos pela formalina ao longo de 1 semana.

142
V - CONSIDERAÇÕES FINAIS
Nossos achados sugerem que os a ação inflamatória do CFA sobre a
ATM ou região orofacial de ratos podem estar relacionadas à alterações
comportamentais originadas pela sua atividade pró-inflamatória que
determinam alterações locais como inflamação periférica e ativação
neurôgênica de vias aferentes nociceptivas. Essas alterações podem ter levado
a modificações nos padrões de neurotransmissão desses tecidos inflamados,
levando a mudanças no comportamento de ativação de regiões dol corno
dorsal medular e sistema nervoso central.
Nós sugerimos, o fenômeno da neuroplasticidade promovido pela
inflamação persistente devido a atividade inflamatória do CFA. Uma avaliação
ampla de nossos resultados demonstra que os animais em geral sob ação do
CFA na região orofacial apresentam uma tendência em diminuir movimentos
exploratórios como: sniffing, climbing, rearing, aumentando o tempo de
execução de comportamentos relacionados a auto cuidados como: freezing,
grooming, chewing-like, scratching, licking, biting e head fliches e redução do
tempo de sono e de descanso. Enquanto que a formalina foi hábil apenas para
destacar o scratching e grooming no seu grupo de animais no 1º dia,
relacionada a sua atividade nociceptiva sub aguda ao passo que em nosso
modelo há um conteúdo inflamatório crônico presente e portanto, com atividade
característica de um modelo crônico, mais semelhante ao contexto em que se
desenvolvem as disfunções temporomandibulares.
A identificação de comportamentos relacionados à atividade inflamatória
do CFA fornece subsídios para o desenvolvimento de novos e melhores
agentes terapêuticos para o tratamento de dor orofacial, como também
avaliação dos benefícios dos tratamentos já existentes, em consonância com
os avanços nos estudo de dores experimentais.

143
VI - CONCLUSÃO
1- Ao longo de 1 semana, os comportamentos alterados pela injeção de CFA
na região da ATM de ratos foram: chewing like, sleeping, grooming e freezing,
podendo servirem como marcadores não invasivos de análise comportamental
nociceptiva.
2- Os comportamentos destacados pelo uso da formalina foram: scratching e
grooming, significativos apenas no 1º dia, concordando com outros estudos
que relatam o tempo de atividade desse irritante.
3- É importante destacar que outras alterações comportamentais como:
aumento de scratching, licking e biting são observados como atividades
relacionadas ao CFA na ATM de ratos.
4- As alterações comportamentais provocadas pelo CFA tenderam a
normalizar-se ao longo de 1 semana, sugerido a ativação de um componente
modulatório endógeno.
5- Os testes farmacológicos evidenciaram uma redução significativa dos
comportamentos chewing like por morfina e indometacina e grooming por
morfina e lidocaína, além de um aumento no tempo de sono e descanso por
lidocaína e indometacina. O comportamento freezing não demonstrou
sensibilidade a nenhum dos analgésicos testados. Podemos sugerir há uma
correlação entre a analgesia promovida e os comportamentos induzidos pelo
CFA e pela formalina 2,5%.
6- As variações relacionadas à atividade analgésica das drogas são referidas
possivelmente devido a via de administração, dose e tempo utilizados e
variação nos componentes pró-inflamatórios indutores de cada atividade
comportamental, sendo por isso necessário mais estudos com dose, tipo e via
de administração de analgésicos.

144
7- Nesse modelo não foi identificado redução do peso corporal final do animal
frente ao estímulo inflamatório do CFA por 1 semana.
8- Histologicamente, o CFA mostrou-se mais efetivo em manter o componente
inflamatório mais intenso, valorizando esse modelo frente ao da formalina.
6- Esse modelo parece servir como um marcador nociceptivo não invasivo em
ratos e pode prover futuros estudos dentro da patogênese das disfunções
temporomandibulares, bem como testar novas terapias farmacológicas para o
tratamento dessa patologia.

145
VII – REFERÊNCIAS BIBLIOGRÁFICAS
ABBOTT, F.V.; HELLEMANS, K.G.C. Phenacetin, acetaminophen and
dypirone: analgesic and rewarding effects. Behav. Brain Res., v.112, p.177-
186, 2000.
AIMONE, L.D.; GEBHART, G.F. Stimulation-produced spinal inhibition from the
midbrain in the rat is mediated by an excitatory amino acid neurotransmitter in
the medial medulla. J Neurosci., v.66, p. 1803-1813, 1986.
AIMONE, L.D.; YAKSH, T.L. Opioid modulation of capsaicin-evoked release of
substance P from rat spinal cord in vivo. Peptides .v.10, p.1127-1131, 1989.
ALLEN, B. J.; LI J.; MENNIG, P.M.; ROGERS, S.D.; GHILARD, J.; MANTYH, P.
and SIMONE D.A. Primary afferent fibers that contribute to increased substance
P receptor internalization in the spinal cord after injury. J Neurophysiol., v. 81
(3), p. 1379-1390, 1999.
ANDERSEN, M.L.; TUFIK, S. Altered sleep and behavioral patterns of arthritic
rats. Sleep Res Online, v.3 (4), p. 161-167, 2000.
ANDERSON, L.C.; VAKOULA, A.; VEINOTE, R. Inflammatory hypersensitivity
in a rat model of trigeminal neuropathic pain. Arch of Oral Biol., v.48, p.161-
169, 2003.
ANDERSON, G.D.; HAUSER, S.D.; McGARITY, K.L.; BREMER, M.E.;
ISAKSON, P.C.; GREGORY, S.A. Selective inhibition of cycloxygenase (COX-
2) reverse inflammation and expression of COX-2 and interleukin 6 in rat
adjuvant arthritis.J Clin Invest., v. 97, p. 2672-2679, 1996.
ANDERSSON, H.I.; EJLERTSSON, G.; LEDEU, I.; SCHERSTEN, B.
Musculoskeletal chronic pain in general pratice. Studies of health care utilization
in comparison with pain prevalence. Scand J Prim Health Care, v. 17 (2), p.87-
92, 1999.
BAKKE, M.; HU, J.W.; SESSLE, B.J. Involvement of NK-1 and NK-2 tachykinin
receptor mechanisms in jaw muscle activity reflexly evoked by inflammatory

146
irritant application to the rat temporomandibular joint. Pain.v. 75, p. 219-227,
1998.
BAROLAT, G.; SCHWARTZMANN, R.; WOO, R. Epidural cord stimulation in
the management of reflex sympathetic dystrophy. Appl Neurophysiol., v.50,
p.2-443, 1987.
BARROS, N. Manifestações clínicas da dor crônica e princípios de tratamento. Dor diagnóstico e tratamento, v.1 (3), p. 3-10, 2004.
BENNETT, G.J.; NEUROPATHIC PAIN. In: WALL, P. D.; MELZACK, R. (Ed).
Textbook of pain. 3rd ed. New York: Churchill Livingstone,. p.494-510, 1994.
BERGMAN, S.; HERSTRON, P.; HOGSTRON, K.; PETERSSON, I.F.;
SVENSSON, B.; JACOBSON, L.T. Chronic musculoskeletal pain, prevalence
rates and sociodemographic associations in a Swedish populations study. J Rheumatol., v.28 (6), p.1369-1377, 2001.
BESSON, P; PERL, E.R. Responses of cutaneous sensory units with
unmyelinated fibers to noxious stimuli. J Neurophysiol., v. 39, p. 1025-1043,
1969.
BETHEA, J.R.; NAGASHIMA, H.; ACOSTA, M.; BRICENO, C; GÓMEZ, F.;
LOOR, K..; GREEN, J.; DIETRICH, W.D. Systemically administered interleukin-
10 (IL-10) reduces tumor necrosis factor alpha production and significantly
improves functional recovery following traumatic spinal cord injury in rats. J Neurotrama., in press, 1999.
BINGEFOURS, K; ISACSON D. Epidemiology, co-morbidity, and impact on
health-related quality of life of self-reported headache and musculoskeletal
pain-a gender perspective. Eur J Pain,. v.5, p.435-450, 2004.
BJORKMAN, R.L.; HALLMAN, K.M.; HEDNER, J.; HEDNER, T.,and HENNING
M. Localization of the central nociceptive effects of diclofenac in the rat. Brain Res., v.590, p. 66-73, 1992.

147
BOJRAB, M. Mecanismos da moléstia na cirurgia dos pequenos animais. 2ª ed.
p.1446, 1996.
BONICA JJ. Pain research therapy. Past and current issues and feature
needs. In: Bonica JJ (ed) Pain, discomfort and humanitarian care. Elsevier,
Amsterdam, p.1-46, 1980.
BRAUN, F.J.L; BRAUN, L. Dor aguda. In: J de Dor, diagnóstico e tratamento.
v.1 (2), p.3-13, 2004.
BROTON, J.G.; SESSLE, B.J. Reflex excitation of masticatory muscles induced
by analgesic chemical applied to the temporomandibular joint of the cat. Arch Oral Biol., v.33, p.741-747, 1988.
BRUNO, M.A.D. Disfunção temporomandibular. Aspectos clínicos de interesse
do cefaliatra. Migrâneas cefaléias, v.7, n.1, p.14-18, 2004.
BUCK, S.H.; BURKS, T.F. The neuropharmacology of capsaicin: review of
some recent observations. Pharmacol., v.38, p.179-250, 1986.
BULLIT, E. Induction of c-fos-like protein within the lumbar spinal cord and
thalamus of the rat following peripheral stimulation . Brain Res., v. 493, p. 391-
397, 1989.
BUSTAMANTEE, D.; PAEILE, C.; WILLER, J.; BARS, D.L. Effects of intrathecal
or intracerebroventricular administration of nonsteroidal anti-inflammatory drugs
on a C-fiber reflex in rats. Pharmacol., v.281, p.1381-1391, 1997.
BUSTAMANTEE, D.; PAEILE, C.; WILLER, J.; BARS, D.L. Effects of
intravenous nonsteroidal anti-inflammatory drugs on a C-fiber reflex elicited by a
wide range of stimulus intensities in the rat. J Pharmacol., v.276, p.1232-1243,
1996.
CAI, B.; CAIRNS, B.E.; SESSLE, B.J.; HU, J.W. Effects of application of
different amino acids to the rat temporomandibular joint (TMJ) region. In:
Abstract, 9 th World Congress on Pain. Vienna, Austria, p.519, 1999.

148
CAIRNS, B.E.; SESSLE, B.J.; HU J.W. Evidence that excitatory amino acid
receptors within the temporomandibular joint region are envolved in the reflex
activation of the jaw muscles. J. Neurosci., v.18, p. 8056-8064, 1998.
CAIRNS, B.E.; SESSLE, B.J.; HU,J.W. Characteristics of glutamate-evoked
temporomandibular joint afferent activity in the rat. J Neurophysiol., v.85(6),
p.7446-7454, 2001.
CALVINO, B.; VILLANUEVA, L.; BARS, D. Dorsal horn(convergent) neurons in
the intact anaesthesied arthritic rat. I. Segmental excitatory influences. Pain. v.28, p.81-98, 1987.
CAMPBELL, J.N.; RAJA, S.N.; COHEN, R.H.; MANNING, D.C.; KHAN, A.A.;
MEYER, R.A. Peripheral neural mechanisms of nociception. In: Wall PD ,
Melzack R (eds). Textbook of pain. Edinburgh, Churchill Livingstone, p.22-
45,1989.
CAPONE, F.; ALOISI, A.M. Refinement of pain evaluation techniques. The
formalin test. Ann Ist Super Sanitá, v. 40 (2), p. 223-229, 2004.
CARLESON, J.; KOGNER, P.; BILEVICIUTE, I.; THEODORSSON, E.;
APPELGREN, A.; APPELGREN, B.; KOPP, S.; YOUSEF, N.; LUNDEBERG, T.
Effects of capsaicin in temporomandibular joint arthritis in rats. Arch of Oral Biol., v. 42, p. 869-876, 1997.
CARLESON, J. Calcitonin gene-related peptide in temporomandibular
inflammation. Karolinska Inst. 2001.
CASHMAN, J.N. The mechanisms of actions of NSAIDS in analgesia. Drugs, v.52, p. 13-23, 1996.
CATTERALL, W.A. From ionic currents to molecular mechanisms: the structure
and function of voltage-gated sodium channels. Neuron., v.26, p. 13-25, 2000.
CERVERO, F. Sensory innervation of the viscera: peripheral basis of visceral
pain. Physiol Rev., v.74(1), p.95-138, 1994.

149
CERVERÓ, F. Neurophisiological aspects of pain and pain therapy. Swerdlow.
M (ed). The therapy of pain. Lacaster: MTP press.; p.-1-29, 1989.
CHANDRASEKHARAN, N.V.; DAÍ, H.; ROOS, K.L.; EVANSON, N.K.; TOMSIK,
J.; ELTON, T.S. and other. COX-3, a ciclooxigenase-1 variant inhibited by
acetaminophen and other analgesic/antipyretic drugs: cloning, structure and
expression. Proc Natl Acad SCI., v. 99 (21), p. 13926-13931, 2002.
CHAO, C.C.; GEKKER, G., HU, S.; SHENG, W.S.; PORTOGHESE, P.S.;
PETERSON, P.K. Upregulation of HIV-1 expression in cocultures of chronically
infected promonocytes and human brain cells by dynorphin. Biochem Pharmacol., v.50, p. 715- 722, 1995.
CHICHORRO, J.G.; LORENZETTI, B.B.; ZAMPRONIO, A. Involvement of
bradikinin, cytokines, sympathetic amines and prostaglandins in formalin-
induced orofacial nociception in rats. British J Pharmacol., v.141, p.1175-
1184, 2004.
CHOI, H.S.; LEE, H.J.; JUNG C.Y.; JU, J.S.; PARK, J.S; AHN, D.K. Central
cyclooxygenase-2 participates in interleukin-1 beta – induced hyperalgesia in
the orofacial formalin test of freely moving rats. Neurosci Let., v.352 (3), p.187-
190, 2003.
CLAVELOU, P.; PAJOT, J.; DALLEL, R.; RABOISSON, P. Application of the
formalin test to the study of orofacial pain in the rat. Neurosci Lett., v. 103, p.
349-353, 1989.
CLAVELOU, P.; DALLEL, R.; ORLIAGUET, T.; WODA, A.; RABOISSON, P.
The orofacial formalin test in rats: effects of different formalin concentrations.
Pain, v.62, p. 295-301, 1995.
CODERRE, T.J.; KATZ, J. Peripheral and central hyperexcitability: differential
sign. and symptoms in persistent pain. Behav and Brain Sci., v.20 (3), p. 404-
419, 1997.

150
CODERRE, T.J.; KATZ, J.; VACCARINO, A.L.; MELZACK, R. Contribuition of
central neuroplasticity to pathological pain: review of clinical and experimental
evidence. Pain. v.52 , p. 259-285, 1992.
CODERRE, T.J.; VACCARINO, A.L.; MELZACK, R. Central nervous system
plasticity in the tonic pain response to subcutaneous formalin injection. Brain Res., v. 535-, p. 155-158, 1990.
COGGESHALL, R.E.; CARLTON, S.M. Receptor localization in the mammalian
dorsal horn and primary afferent neurons. Brain Res , v. 24, p.28-66, 1997.
COHEN, J.A.; PRINCE, E.O. Grooming in the Norway rat : displacement activity
or ‘boudndary-shift’, Behav. Neural Biol., v.26, p.177-188, 1979.
COOK, A.J.; WOOLF, C.J.; WALL, P.D.; McMAHON, S.B. Dynamic receptive
field plasticity in rat spinal dorsal horn following C-primary afferent input. Nature. v. 325, p. 151-153, 1987.
COSTEN, J.B. Syndrome of ear and sinus symptoms dependent upon functions
of the temporomadibular joint. Ann Otol Rhinol Laryngol.,v.3, p.1-4, 1934.
CRAIG, A.D.; SORKIN, L.S. Pain and analgesia. Encyclopedia of life sciences/ Nature, p. 1-8, 2001.
DANZIGER, N.; WEIL-FUGAZZA, J.; Le BARS, D.; e outros autores. Alteration
of descending modulation of nociception during the course of monoarthritis in
the rat. The J of Neurosci., v.19 (6), p.2394-2400, 1999.
DE BIASI, S; RUSTIONI, A. Glutamate and substance P coexist in primary
afferents terminals in the superficial laminae of spinal cord. Proc Natl Acad Sci,
v.85, p.7820-7824, 1988.
DE CASTRO-COSTA, C.M.; GYBELS, J.; KUPERS, R.; HEES, J.V. Scratching
behavior in arthritic rats: a sign of chronic pain or itch? Pain, v.29, p.123-131,
1987.

151
DeLEO, J.A.; COLBURN, R.W.; RICKMAN, A.J. Cytokine and growth factor
immunohistochemical spinal profiles in two animal models of mononeuropathy.
Brain Res., v. 759, p. 50-57, 1997.
DICKENSON, A.H.; SULLIVAN, A.F. Evidence for a role of the NMDA receptor
in the frequency dependent potentiation of deep rat dorsal horn nociceptive
neurons following C-fibre stimulation. Neuropharm.,. v. 26, p. 1235-1238, 1987.
DOUCETTE, R.; THERIAULT, E.; DIAMOND, J. Regionally selective
elimination of cutaneous thermal nociception in rats by neonatal capsaicin. J Com Neurol., v.261, p. 583-591, 1987.
DOUGHERTY, P.M.; WILLIS, W.D. Enhancement of spinothalamic neuron
responses to chemical and mechanical stimuli following combined
microiontophoretic application of N- methyl-D-aspartic acid and substance P.
Pain, v.47, p.85-93, 1991.
DOUGHERT, P.M.; SLUKA, K.A.; SORKIN, K.N.;WESTLUND, K.N.; WILLIS,
W.D. Neural changes in acute arthritis in monkeys parallel enhancement of
responses of spinothalamic tract neurons to mechanical stimulation and
excitatory amino acids. Brain Res Reviews., . v. 17, p. 1-13, 1992.
DRAISCI, G.; IADAROLA, M.J. Temporal analysis of increases in c-fos ,
predrodynorphin and preproenkephalin mRNAs in rat spinal cord. Mol. Brain Res., v.6, p.31-37, 1989.
DRAY, A.; URBAN, L. New pharmacological strategies for pain relief. Annu. Rev. Pharmacol. Toxicol., v.36, p. 253-280, 1996.
DREWES, A.M.; NIELSEN, K.D; ARENDT-NIELSEN, L..; BIRKET-SMITH, L.;
HANSEN, L.M. The effect of cutaneuos and deep pain on the
electroencephalogram during sleep- an experimental study. Sleep, v.20, p. 623-
640, 1997.
DUBNER, R.; REN, K. Endogenous mechanisms of sensory modulation. Pain Suppl., v. 6, p. 45-53, 1999.

152
DUBNER, R.; REN, K. Braistem mechanisms of persistent pain following injury.
J Orofac Pain. v. 18, p-299-305, 2004.
DUBNER, R. Neuronal plasticity and pain following peripheral tissue
inflammation or nerve injury. In: Bond MR., Charlton J.E.,Woolf C.J. editors.
Proceedings of the VIth World Congress on Pain. Pain res and clinic manag.,. Amsterdam: Elsevier.v.4, p. 263-276, 1991.
DUBNER, R. Recent advances in our understanding of pain. In: Klineberg I,
Sessle, eds. Orofacial pain and neuromuscular dysfunction: Mechanisms and
Clinical Correlates. Oxford: Pergamon Press. p. 3-19, 1985.
DUBNER, R.; BENNETT, G.J. Spinal and trigeminal mechanisms of
nociception. Ann Rev Neurosci., v. 6, p. 381-418, 1983.
DUBNER, R.; RUDA, M.A. Activity-dependent neuronal plasticity following
tissue injury and inflammation . Trends Neurosci. v.15, p.96-102, 1992.
DUBNER, R. Basic mechanisms of pain associated with deep tissues. Can. J. Physiol. Pharmacol., v. 69, p. 607-609, 1991.
DUBOIS, R.N.; ARAMSON, S.B.; CROFFORD, L.; GUPTA, R.A.; SIMON, L.S.;
VAN DE PUTTE, L.B.A.; LIPSKY, P.E. Cyclooxygenase in biology and disease.
The Faseb J., v.12, p.1063-1073, 1998.
DUBUISSON, D.; DENNIS, S.G. The formalin test: a quantitative study of the
analgesic effects of morphine, meperidine , and brain stem stimulation in rats e
cats. Pain, v. 4, p.161-174, 1977.
DUGGAN, A. W.; HENDRY, I.A.; MORTON, C.R.; HUTCHINSON, W.D.; ZHAO,
Z.Q. Cutaneous stimuli releasing immunoreactive substance P in the dorsal
horn of the cat. Brain Res., p. 261-273, 1988.
DUGGAN, A.W.; HOPE, P.J.; JARROTT, B.; SCHAIBLE, H.G.; FLEET-WOOD-
WALKER, S.M. Release, spread and persistence of immunreactive neurokinin
A in the dorsal of the cat following noxious cutaneous stimulation. Studies with
antibody microprobes. Neurosci., v. 35, p. 195-202, 1990.

153
DWORKIN, S.F.; HUGGINS, K.H.; LE RESCH, L.; VON KORFF, M.;
TRUELOVE, E.; SOMMERS, E. Epidemiology of signs and symptoms in
temporomandibular disorders: clinical signs in cases and controls. J Am Dent Assoc., v.120, p.273-281, 1990.
EISENBERG, E.; VOS, B.P.; STRASSMAN, A.M. The peripheral
antinociceptive effect of morphine in a rat model of facial pain. Neurosci., v. 72,
p.519-525, 1996.
ELVIRE, G.; PHILIPPE, L.; MINN, A.; HAOUZI, P.; GILLET, P.; NETTER, P.;
TERLAI, B. Neurophysiological basis for neurogenic-mediated articular cartilage
anabolism alteration. AJP., v.280, p.115-122, 2001.
FARIABY J; MOHAMMADI M. Prevalence of temporomandibular joint disorders
in 9-12-old boy students in Kerman, Southeast of Iran. Iran J Med Sci., v. 30,
p.91-93, 2005.
FERREIRA, S.; BERENICE, H.; LORENZETTI, B.; CORREA, F.M.A. Central
and peripheral anti-algesic action of aspirin-like drugs. Eur. J. Pharmacol., v.
53, p.30-48, 1978.
FIELDS, H.L.; BASBAUM, A.I. Endogenous pain control mechanisms . In: Wall
PD, Melzack R (eds), Textbook of pain. Edinburg, Churchil Livingstone. p.
206-217, 1989.
FOLEY, D.; ANCOLI-ISRAEL, S.; BRITZ, P.; WALSH, J. Sep disturbances and
chronic disease in older adults; results of the 2003 National Sleep Foundation
Sleep in America Survey. J Psychsom., v. 56(5), p. 497-502, 2004.
FORSYTHE, I.D.; WESTBROOK, G.L. Slow excitatory postsynaptic currents
mediated by NMDA receptors on cultured mouse central neurons. J Physiol., v.
396, p.515-533, 1988.
FUJITA, Y.;MOTEGI, E.; NOMURA, M.; KAWAMURA, S.; YAMAGUCHI, D.;
YAMAGUCHI, H. Oral habits of temporomandibular disorders patients with
malocclusion. Bull Tokyo Dent Coll., v.44 (4), p. 201-207, 2003.

154
GEAR, R.W.; ALEY, K.O.; LEVINE, J.D. Pain-induced analgesia mediated by
mesolimbic reward circuits. J Neurosci., v.19, p.7175-7181, 1999.
GJESTAD, J.; TJOLSEN, A.; SVENDSEN, F.; HOLE, K. Induction of long-term
potentiation of single wide dynamic range neurons in the dorsal horn is inhibited
by descending pathways. Pain. v. 91, p. 263-268, 2001.
GOBEL, S.; HOCKFIELD, S.; RUDA, M.A. Anatomical similarities between
medullary and spinal dorsal horns. In: Oral-facial sensory and motor functions. Ed. Y. Kawamura and R. Dubner. Tokyo: Quintessence. p. 211-223,
1981.
GORDON, G.; LANDGREN, S.; SEED, W.A. The functional characteristics of
single cells in the caudal part of the spinal nucleous of the trigeminal nerve of
the cat. J Physiol., v. 158, p. 544-559, 1961.
GOULART, A.C. Study of the inflammatory process in temporomandibular joint induced by injection of carragenin or formalin. Dissertação de mestrado, São Paulo, 2002.
GOULD, H.J.; ENGLAND, J.D.; LIU, Z.P.; LEVINSON, S.R. Rapid sodium
channel augmentation in response to inflammation induced by complete
Freund’s adjuvant. Brain Res., v.802, p.69-74, 1998.
GOULET, J.P.; LAVIGNE, G.J.; LUND, J.P. Jaw positioning prevalence among
French-speaking Canadians in Québec and related symptoms of
temporomandibular disorders. J. Dent. Res., v.74, p.1738-1744, 1995.
GRIGG, P.; SCHAIBLE, H.; SCHMIDT, R.F. Mechanical sensitivity of group III
and IV afferents from posterior articular nerve in normal and inflammed cat
knee. J Neurophysiol.,. v. 55, p. 635-643, 1986.
GUAN, Y.; GUO, W.; ZOU, S.P.; DUBNER, R.; REN, K. Inflammation-induced
upregulation of AMPA receptor subunit expression in brain stem pain
modulatory circuitry. Pain. v.104, p. 401-413, 2003.

155
GUAN, Y.; TERAYAMA, R.; DUBNER, R.; REN, K. Plasticity in excitatory amino
acid receptor-mediated descending pain modulation after inflammation.
Pharmacol and Exp Therapeutics. v.300, p.513-520, 2002.
GUILBAUD, G.; KAYSER, V.; BENOIST, J.M.; GAUTRON, M. Modifications in
the responsiveness of rat ventrobasal thalamic neurons at different stages of
carragenin-produced inflammation. Brain Res.,. v. 385, p. 86-98, 1986.
GUO, W.; ZOU, S.; GUAN, Y.; IKEDA, T.; TAL, M.; DUBNER, R.; REN, K.
Tyrosine phosphorylation of the NR2B Subunit of the NMDA receptor in the
spinal cord during the development and maintenance of inflammatory
hyperalgesia. J Neurosci., v. 22 (14), p. 6208-6217, 2002.
GUYTON, A.C. Texbook of medical physiology (8 ed). Philadelphia: WB
Saunders, p. 499, 1991.
HAAS, D.A.; NAKANISHI, O.; MAcMILLAN, R.E.; JORDAN, R.C.; HU, J.W.
Development of an orofacial model of acute inflammation in the rat. Archs oral Biol., v.37, p. 417-422, 1992.
HARDY, J.D.; WOLFF, H.G.; GOODELL, H. Experimental evidence on the
nature of cutaneous hyperalgesia. J Clin Invest., v.29, p.115-140, 1950.
HARGREAVES, K.; DUBNER, R.; BROWN, F.; FLORES, C.; JORIS, J. A new
and sensitive method for measuring thermal nociception in cutaneous
hyperalgesia. Pain, v. 32, p. 77-88, 1988.
HARPER, R.P.; KERINS, C.A.; McINTOSH, J.E.; SPEARS, R.; BELLINGER,
L.L. Modulation of the inflammatory response in the rat TMJ with increasing
doses of complete Freund’s adjuvant. J OsteoArth Res.,. Osteoarthritis and
Cartilage , v.9, p.619-624, 2001.
HARPER, R.P.; KERINS, C.A.; TALWAR, R.; SPEARS, R.; CARLSON, D.S.;
McINTOSH, J.E., et al. Meal pattern analysis in response to temporomandibular
joint inflammation in the rat. J Dent Res.,. v. 79, p.1704-1711, 2000.

156
HARTWING, A.; MATHIAS, S.I.; LAW, A.S.; GEBHART, G.F. Characterization
and opioid modulation of inflammatory temporomandibular joint pain in the rat. J Oral Maxillofac., v.61, p. 1302-1309, 2003.
HASSAN, A.H.S.; ABLEITNER, A.; STEIN, C.; HERZ, A. Inflammation of the rat
paw enhances axonal transport of opioid receptors in the sciatic nerve and
increases their density in the inflamed tissue. Neurosci.,, v.55, p. 185-195,
1993.
HERZ, A. (ed). Definitive compendium of reviews on all aspects of opioid
pharmacology. Handbook of experimental pharmacology, 1993.
HIGUSHI, S.; TANAKA, N.; SHIOIRI, Y.; OTOMO, S.; AIHARA, H. Two modes
of analgesic action of aspirin , and the site of analgesic action of salicylic acid.
J. Tissue React.,. v.8, p.327-331, 1986.
HOLMUND, A; EKBLOM, A.; HANSSON, P.; LIND, J.; LUNDEBERG, T.;
THEODORSSON, E. Concentrations de neuropeptídes substance P, neurokinin
A, calcitonin gene-related peptide, neuropeptide Y and vasoactive intestinal
polypeptide in synovial fluid of the human temporomandibular joint. J Oral Maxillofac., v.20 (4), p.228-231, 1991.
HONORÉ, P.; MENNING, P.; ROGERS, S.; NICHOLS, M. et al. Spinal
substance P receptor expression and internalization in acute, short-term, and
long-term inflammatory pain states. The J Neurosci.,. , v.19 (17), p. 7670-7678,
1999.
HOSKIN, K.L.; KAUBE, H.; GOADSBY, P.J. Sumatriptan can inhibit trigeminal
afferents by an exclusively neural mechanisms. Brain. v. 119, p. 1419-1428,
1996.
HU, J.W.; SESSLE, B.J.; RABOISSON, P.; DALLEL, R.; WODA, A. Stimulation
of craniofacial muscle afferents induces prolonged facilitatory effects in
trigeminal nociceptive brain-stem neurons. Pain. v.48, p.53-60, 1992.

157
HU, J.W.; SESSLE, B.J.; Properties of functionally identified nociceptive and
nonnociceptive facial primary afferents and presynaptic excitability changes
induced in their brain stem endings by raphe and orofacial stimuli in cats. Exp Neurol.,. v. 101, p.385-399, 1988.
HUNSKAAR, S.; HOLE, K. The formalin test in mice: dissociation between
inflammatory and non-inflammatory pain. Pain, v. 30, p. 104-114, 1987.
HUNT, S.P.; PINI, A.; EVAN, G. Induction of c-fos-like protein in spinal cord
neurons following sensory stimulation. Nature, v.328, p.632-634, 1987.
HURLEY, R.W.; HAMMOND, D.L. The analgesic effects of supraspinal mu and
lambda opioid receptor agonists are potentiated during persistent inflammation. J Neurosci., .v. 20, p. 1249-1259, 2000.
HYLDEN, J.L.K.; NAHIN, R.L.; TRAUB, R.J.; DUBNER, R. Effects of spinal
kappa-opioid receptor agonists on the responsiveness of nociceptive superficial
dorsal horn neurons. Pain, v. 44, p.187-193, 1991.
LADAROLA, M.J.; BRADY, L.S.; DRASCI, G.; DUBNER, R. Enhancement of
dynorphin gene expression in spinal cord following experimental inflammation:
stimulus specificity behavioral parameters and opioid receptor biding. Pain, v.35, p. 313-326, 1988.
IASP (International Association for Study of Pain) Subcomittee on taxonomy.
Classification of cronic pain. Descritions of chronic pain syndromes and
definitions of pain terms. Pain (Suppl 3), 1986.
ICHD (The International classification of Headache Disorders), 2nd Ed. Cephal., v.24 (1), p.118, 2004.
IMBE, H.; DUBNER, R.; REN, K. Masseteric inflammation-induced fos protein
expression in the trigeminal interpolaris/caudalis transition zone: contribuition of
somatosensory-vagal-adrenal integration. Brain Res., v. 845 (2), p. 165-175,
1999.

158
IMBE, H.; IWATA, K.; ZHOU, Q.; ZOU, S.; DUBNER, R.; REN, K. Orofacial
deep and cutaneous tissue inflammation and trigeminal neuronal activation:
Implications for persistent temporomandibular pain. Cells Tissues Organs,. v.
169, p. 238-247, 2001.
INTURRISI, C.E. Clinical pharmacology of opioids for pain . Clin J Pain., v.18
(suppl.4): p.3-13, 2002.
IWATA, K.; TASHIRO, A.; TSUBOI, Y.; IMAI, T.; SUMINO, R.; MORIMOTO, T.;
DUBNER, R.; REN, K. Medullary dorsal horn neuronal activity in rats with
persistent temporomandibular joint and perioral inflammation. J Neurophysiol.,. v.82, p. 1244-1253, 1999.
JENSEN, T.S. Mechanisms of Neuropathic Pain. In : Campbell JN (ed) . IASP
Committee on Refresher Courses. Seattle IASP Press, p. 77-86, 1996.
KAPILA, S.; XIE, Y. Targeted induction of collagenase and stromelysin by
relaxin in unprimed and beta-estradiol –primed diarthrodial joint
fibrocartilaginous cells but not in synoviocytes. Lab Invest., v.78, p.925-938,
1998.
KEHL, L.J.; GOGAS, K.R.; LICHTBLAU, L.; POLLOCK, C.H.; MAYES, M.;
BASBAUM, A.I. and WILCOX, G.L. The NMDA antagonist MK-801 reduces
noxious stimulus-evoked Fos expression in the spinal dorsal horn. In: M.R.
Bond, J.E. Charlton and C.J. Wollf (eds). Pain Res and Clinical Management., . v.4, p.307-311, 1991.
KERINS, C.A.; CARLSON, D.S.; McINTOSH, J.E.; BELLINGER, L.L. A role for
cyclooxygenase II inhibitors in modulating temporomandibular joint inflammation
from a meal pattern analysis perspective. J Oral Maxillofac.,. v.62, p. 989-995,
2004.
KERINS, C.A.; CARLSON, D.S.; McINTOSH, J.E.; BELLINGER, LL. Meal
pattern changes associated with temporomandibular joint inflammation/pain in
rats; analgesics effects. Pharmacol Biochem and Behav., v. 75, p.181-189,
2003.

159
KIDD, B.L.; URBAN, L.A. Mechanisms of inflammatory pain. Br L Anaesth.,. v.87, p. 3-11, 2001.
KIRCHHOFF, C.; JUNG, S.; REEH, P.W.; HANDWERKER, H.O. Carrageenan
inflammation increases bradykinin sensitivity of rat cutaneous nociceptors.
Neurosci Lett.,. v. 111, p.206-210, 1990.
KIRCHNER, T.; APARICIO, B.; ARGENTIERI, D.C.; LAU, C. Y.; RITCHIE, D.M.
Effects of tepoxalin, a dual inhibitor of cyclooxynase/ 5-lipoxygenase, on events
associated with NSAID-induced gastrointestinal inflammation. Prostagl Leukot. Essent. Fatty Acids. v. 56, p.417-423, 1997.
KOLTZENBURG, M.; KRESS, M.; REEH, P.W. The nociceptor sensitization by
bradikinin does not depend on sympathetic neurons. Neurosci., . v.46, p.465-
473, 1992.
KONG, J.Q.; LEEDHAM, J.A.; TAYLOR, D.A.; FLEMING, W.W. Evidence that
tolerance and dependence of guinea pig myenteric neurons to opioids is a
function of altered eletrogenic sodium-potassium pumping. J Pharmacol Exp Ther.,. v. 280, p. 593-599, 1997.
KOPP, S. Neuroendocrine, immune, and local responses related to
temporomandibular disorders. J Orofac Pain, v.15, p.9-28, 2001.
KUPERS, R.C.; NUYTTEN, D; DE CASTRO-COSTA, M.; GYBELS, J.M. A time
course analisis of changes in spontaneous and evoked behavior in a rat model
of neuropathic pain. Pain., v.50, p.101-111.1992.
LaMOTTE, R.H.; SHAIN, C.N.; SIMONE, D.A.; TSAI, E.F.P. Neurogenic
hyperalgesia: psychophysical studies of underlying mechanisms. J Neurophysiol., v. 66, p. 190-211, 1991.
LAUGHLIN, T.M.; BETHEA, J.R.; YEZIERSKI, R.P.; WILCOX, G.L. Cytokine
involvement in dynorphin-induced allodynia. Pain. v.84, p. 159-167, 2000.

160
LAWAND, NADA, B, WILLIS, D.; WESLUND; KARIN, N. Excitatory amino acid
receptor involvement in peripheral nociceptive transmission in rats. Eur J Pharm., v.324, p.169-177, 1997.
LE RESCHE, L.; FRICTON, J.; MOHL, N. Research diagnostic criteria. Part II. J Cranio Dis Fac Oral Pain, v.6, p. 327-334, 1992.
LEE, I.; KONG, M.; KIM, N.; CHOI, Y.; LIM, S.; LEE, M. Effects of different
concentrations and volumes of formalin on pain response in rats. Acta Anaesth., v.38, p.59-64, 2000.
LERMA, J. Kainato reveals its targets. Neuron. v.19, p.1155-1158, 1997.
LEVINE, J.D.; FIELDS, H.L.; BASBAUN, A.I. Peptides and primary afferent
nociceptor . J Neuroci. v.13, p. 2273-2286, 1993.
LIPTON J. A.; SHIP J.A. & LARACH-ROBINSON D. Estimated prevalence and
distribuition of reported orofacial in the United States. J Am Dent Assoc.,
v.124, p.115-121, 1993.
LOBBEZOO, F.; DRANGSHOLT, M.; PECK, C.; SATO, H.; KOPP, S.;
SVENSSON, P. Topical review: New insights into the pathology and diagnosis
of disorders of the temporomandibular joint. J Orof Pain, v. 18 (3), p.181-191,
2004.
LORD, J.A.; WATERFIELD, A.A.; HUGHES, J.; KOSTERLITZ, H.W.
Endogenous opioid peptides: multiple agonists and receptors. Nature, v.267, p.
495-499, 1977.
MacDERMOTT, A.B.; MAYER, M.L.; WESTBROOK, G.L.SMITH, S.J.;
BARKER, J.L. NMDA-receptor activation increases cytoplasmic calcium
concentration in cultured spinal cord neurons. Nature, v.321, p.519-522, 1986.
MACIEL, R.N. Oclusão e ATM – Procedimentos clínicos. São Paulo, Editora
Santos, 1998.l

161
MacPHERSON, R.D. The pharmacological basis of contemporary pain
management. Pharmacol Ther, v. 88, p. 163-185, 2000.
MAIXNER, W.; FILLINGIM, R.; BROKER, D.; SIGURDSSON, A. sensitivity of
patients with painful temporomandibular disorders to experimentally evoked
pain. Pain. v. 63, p. 341-351, 1995.
MAIXNER, W.; FILLINGIM, R.; SIGURDSSON, A.; KINCAID, S.; SILVA, S.
Sensitivity of patients with painful temporomandibular disorders to
experimentally evoked pain: evidence for altered temporal summation of pain.
Pain, v. 76, p. 71-81, 1998.
MALMBERG, A.B.; YAKSH, T.L. Antinociceptive actions of spinal anti-
inflammatory agents on the formalin test in the rat. J Pharmacol. Exp. Ther. v.
263, p. 136-146, 1992.
MALMBERG, A.B.; RAFFERTY, M.F.; YAKSH T.L. Antinociceptive effect of
spinally delivered prostaglandin E receptor antagonist in the formalin test in the
rat. Neurosci, v.173, p. 193-196, 1994.
MANNING, D.C.; RAJA, S.N.; MEYER, R.A.; CAMPBELL, J.N. Pain and
hyperalgesia after intradermal injection of bradykinin in humans. Clin Pharmacol Ther. v. 50, p. 721-729, 1991.
MARTYN, S. On the physiological meaning of inframammary pain. Br Med J .
v. 2, p.296-298, 1964.
McCLESKY, E.W.; GOLD, M.S. Ion channels of nociception. Annu Rev Physiol, v.61, p.835-856, 1999.
McFARLAND, D. (ed). The Oxford Companion to animal Behavior, Oxford
University Press. p. 237-240, 1987.
McMAHON, S.B. Neuronal and behavioral consequences of chemical
inflammation of rat urinary bladder. Agents Actions. v. 25, p. 231-233, 1988.

162
McMILLAN, A.S.; BLASBERG, B. Pain-pressure threshold in painful jaw muscle
following trigger point injection. J Orofac Pain. v.8, p. 384-390, 1994.
McNEILL, C.; DANZIG, D.; FARRAR, W.; GELB, H.; LERMAN, M.D.;
MOFFETT, B.C. Craniomandibular (TMJ) disorders - state of the art. J Prosthet Dent. v.44, p.434-440, 1980.
MEIRA, G. S.P. DTM X Sintomas otológicos.
<http://www.aoonp.org.br/fso/revista7 /rev712a.htm>, 2002.
MELCER, T.; TIMBERLAKE, W. Poison avoidance and patch (location)
selection in rats. Anim learn and behav, v. 13, p. 60-68, 1985.
MELZACK, R. Psychological concepts and methods for the control of pain. In Bonica JJ (ed): Adv in neurology , v.4, p.275-280,1979.
MELZACK, R.; WALL, P.D. Pain mechanisms : a new theory. Science.v. 150,
p. 971-979, 1965.
MENÉTREY, D.; GANNON, A.; LEVINE, J.D.; BASBAUM, A.I. Expression of c-
fos protein in interneurons and projection neurons of the rat spinal dorsal in
response to noxious somatic, articular, and visceral stimulation. J Comp Neurol., v.285, p. 177-195, 1989.
MENSE, S. Nociception from skeletal muscle in relation to clinical muscle pain.
Pain. v.54, p. 241-289, 1993.
MERSKEY, H.; ALBE-FESSARD, D.G.; BONICA, J.J.; CARMON, A.; DUBNER,
R.; KERR, F.W.L.; LINDBLOM, U.; MUMFORD, J.M.; NATHAN, P.W.;
NOOORDENBOS, W.; PAGNICA; RENAER, M.J.; STEERBACH, R.A.;
SUNDERLANDS, S. Pain terms: a list with definitions and notes on usage.
Recommended by IASP subcommittee on taxonomy. Pain. v.6, p.249-252,
1979.
MILLAN, M.J. The induction of pain: an integrative review. Prog Neurobiol. v.57(1), p.161-164, 1999.

163
MILLAN, M.J.; COLPAERT, F.C. Opioids system in the response to
inflammatory pain: sustained blockage suggests role of k-but not d opioid
receptors in the modulation of nociception, behavior and pathology. Neurosci, v.42, p. 541-553, 1991.
MINAMI, M.; KURAISHI, Y.; KAWAMURA, M.; YAMAGUCHI, T.; MASU, Y.
Enhancement of preprotakhykinin A gene expression by adjuvant-induced
inflammation in the rat spinal cord: possible involvement of substance P-
containing neurons in nociception. Neurosci. Lett., v.98, p. 105-110, 1989.
MITSIKOSTAS, D.D.; SANCHEZ, DEL RIO, M.; WAEBER, C.; MOSKOWITZ,
M.A.; CUTRER, F.M. The NMDA receptor antagonists MK-801 reduces
capsaicin-induced c-fos expression within rat trigeminal nucleous caudalis.
Pain, v.76, p. 239-248, 1998.
MORTON, C.R.; HUTCHINSON, W.D. Release of sensory neuropeptides in the
spinal cord: studies with calcitonin gene-related peptide and galanin. Neurosci., v.31, p. 807-815, 1990.
MORTON, C.R.; HUTCHINSON, W.D.; HENDRY, I.A. Release of
immunoreactive somastostatin in the spinal dorsal horn of the cat. Neuropep, v.
12, p. 189-197, 1988.
NEUGEBAUER, V.; SCHAIBLE, H.G. Evidence for a central component in the
sensitization of spinal neurons with joint input during development of acute
arthritis in cat’s knee. J of Neurophysiol.. v. 64, p. 299-311, 1990.
NEUGEBAUER, V.; SCHAIBLE, H.G.; SCHMIDT, R.F. Sensitization of articular
afferents to mechanical stimuli by bradykinin. Pflügers Arch .v. 415, p. 330-
335, 1989.
NISHIMORI, T.; BUZZI, M.G.; MOSKOWITZ, M.A.; UHL, G.R.
Preproenkephalin mRNA expression in nucleous caudalis neurons is enhanced
by trigeminal stimulation. Mol. Brain Res., v.6, p.203-210, 1989.

164
NOZAWA-INOUE, K.; TAKAGI, R.; KOBAYASHI, T.; OHASHI, Y.; MAEDAT.
Immunocytochemical demonstration of the synovial membrane in
experimentally induced arthritis of the rat temporomandibular joint. Arch Histol Cytol, v.61, p.451-466, 1998.
OGAWA, A.; MORIMOTO, T.; HU, J.W.; TSUBOI, Y.; TASIRO, A.; NOGUSH,
K.; NAKAGAWA, H.; IWATA, K. Hard-food mastication suppresses complete
Freund’s adjuvant-induced nociception. Neurosci, v.120, p.1081-1092, 2003.
OKESON, J. P. Fundamentos da oclusão e desordens temporomandibulares. 2ª ed. São Paulo: Artes Médicas, 1992.
OKESON, J. P. Fundamentos da oclusão e desordens temporomandibulares. 7ª ed. São Paulo: Artes Médicas, 2000.
OKESON, J.P. Neuroanatomia da dor bucofacial. In: Dores bucofaciais de Bell. 5ª edição, Quintessence Publishing. p.18, 1998.
OKESON, J.P.; DE KANTER, R.J. Temporomandibular disorders in the medical
practice. In : J Fam Prat, v.43, p.347-356, 1996.
OKU, R.; NANAYAMA, T.; SATOH, M. Calcitonin gene-related peptide
modulates calcium mobilization in synaptosomes of rat spinal dorsal horn.
Brain Res., v.475, p.356-360, 1988.
OKUYAMA, S.; AIHARA, H. Effects of morphine and indomethacin on evoked
neuronal responses of ventro basal thalamic neurone: Site of action of
analgesic drugs in adjuvant arthritis rats. J Pharmacol. v.36, p.177-186, 1984.
OLIVEIRA A.S.; BERMUDEZ C.C.; SOUZA R.A. e outros autores. Impacto da
dor na vida de portadores de disfunção temporomandibular. J Appl Oral Sci. V.11,
PELISSIER, T.; PAJOT, J.; DALLEL, R. The orofacial capsaicin test in rats:
effects of different capsaicin concentrations and morphine. Pain, v.96, p.81-87,
2002.

165
PIEHL, F.; ARVIDSONN, U.; JOHNSON, H.; CULLHEIM, S.; VILLAR, M.;
DAGERLING, A.; TERENIUS, L.; HÖKFELT, T.; ULFHAKE, B. Calcitonin gene-
related peptide (CGRP)-like immunoreactivity and CGRP messenger-RNA in rat
spinal-cord motoneurons after different types of lesions. Eur. J. Neurosci. v. 3.
p.737-757, 1991.
PITCHER, G.M.;HENRY, J.L. Mediation and modulation by eicosanoids of
response of spinal dorsal horn neurons to glutamate and substance P receptor
agonists: results with indomethacin in the rat in vivo. Neurosci. v. 93, p.1109-
1120, 1999.
POWELL, K.J.; HOSOKAWA, A.; BELL, A.; SUTAK, M.; MILNE, B.; QUIRION,
R.; JHAMANDAS, K. Comparative effects of cyclo-oxygenase and nitric oxide
syntase inhibition on the development and reversal of spinal opioid tolerance.
Br. J. Pharmacol. v.127, p.631-644, 1999.
PRICE, M.P.; LEWIN, G.R.; McWRATH, S.L. The mammalian sodium channel
BNCL is required for normal touch sensation. Nature, v.497, p. 1007-1011,
2000.
PRICE, D.D.; HU, J.W.; DUBNER, R.; GRACELY, R.H. Peripheral suppression
of first pain and central summation of second pain evoked by noxious heat
pulses. Pain. v.3, p..57-68, 1977.
PRZEWLOCKI, R.; HASSAN, A.H.S.; LASON, W.; EPPLEN, C.; HERZ, A.;
STEIN, C. Gene expression and localization of opioid peptides in immune cells
of inflamed tissue. Functional role in antinociception. Neurosci. v. 48, p.491-
500, 1992.
RADHAKRISHNAN, V.; HENRY, J. L. Novel substance P antagonist, CP-
96,354 blocks responses of cat spinal dorsal horn neurons to noxious
cutaneous stimulation and to substance P. Neuroci. Lett. v.132, p.39-43, 1991.
RANG, H.P.; DALE, M.M.; RITTER, J.M. Anestésicos locais e outros fármacos
que afetam os canais iônicos. In: Farmacologia, 4ª ed, Guanabara Koogan,
cap.40, p.532-540, 1999a.

166
RANG, H.P.; DALE, M.M.; RITTER, J.M. Fármacos analgésicos. In:
Farmacologia, 4ª ed, Guanabara Koogan, cap.37, p.485-505, 1999b.
REID, K.L.; GRACELY, R.H.; DUBNER, R.A. The influence of time, facial side
and location on pain-pressure thresholds in chronic myogeneous
temporomandibular disorder. J Orofac Pain. v. 8, p. 258-265, 1994.
REN, K.; DUBNER, R. Central nervous system plasticity and persistent pain. J Orofac Pain. v.13, p. 155-163, 1999.
REN, K.; DUBNER, R. Descending modulation in persistent pain: an update.
Pain. v. 100, p.1-6, 2002.
REN, K.; HYLDEN, J.L.K.; WILLIAMS, G.M.; RUDA, M.A.; DUBNER, R. The
effects of a non-competitive nmda receptor antagonist, MK-801, on behavioral
hyperalgesia and dorsal horn neuronal activity in rats with unilateral
inflammation. Pain, v. 50, p.331-344, 1992.
RIZZATTI-BARBOSA, C.M.; ARANA, A.R.S.; CUNHA JR.,A.C. e outros
autores. Avaliação da dor na desordem temporomandibular: caso clínico. Rev ABO Nac., v.8, p. 171-175, 2000.
RIZZI, M.; SARZI-PUTTINI, P.; ATZENI, F.; CAPSONI, F.; ANDREOLI, A.;
PECIS, M.; COLOMBO, S.; CARRABBA, M.; SERGI, M. Cyclic alternating
pattern: a new marker of sleep alteration in patients with fibromyalgia? J Rheumatol, v.31 (6), p. 1193-1199, 2004.
RO, J.Y.; CAPRA, N.; MASRI, R. Development of a behavioral assessment of
craniofacial muscle pain in lightly anesthezided rats. Pain, v. 104, p. 179-185,
2003.
RO, J.Y. Contribution of peripheral NMDA receptors in craniofacial muscle
nociception and edema formation. Brain Res, v.979 (1-2), p.78-84, 2003.
ROBERTS, W.W.; MOONEY, R.D.; MARTIN. J.R. Thermoregulatory
behaviours of laboratory rodents, J Comp Physiol., v 86, p.693-699, 1974.

167
ROCHA, A.P.F. Epidemiologia das desordens temporomandibulares: Estudo de
prevalência da sintomatologia e sua interrelação com a idade e sexo dos
pacientes. Rev Serv ATM. v.2 (1), p. 5-10 (2002).
ROIZENBLATT, S.; MOLDOFSKY, H.; BENEDITO-SILVA, A.A.; TUFIK, S.
Alpha sleep characteristics in fibromyalgia. Arthritis Rheum., v. 44, p. 222-230,
2001.
ROSLAND, J.H.; TJOLSEN, A.; MACHLE, B.; HOLE, K. The formalin test in
mice: effect of formalin concentration. Pain, v.42, p. 235-242, 1990.
ROVERONI, R.; PARADA, C.A.; CECÍLIA, M.; VEIGA, F.A.; TAMBELI, C.H.
Development of behavioural model of TMJ pain in rats: the formalin test. Pain, v.94, p.185-191, 2001.
RUDA, M.A.; IADOROLA, M.J.; COHEN, L.V.; YOUNG, W.S., III. In situ
hybridization histochemistry and immunocytochemistry reveal an increase in
spinal dynorphin biosyntesis in a rat model of peripheral inflammation and
hyperalgesia. Proc. Natl. Acad. Sci. (USA), v.85, p.622-626, 1988.
SAKAUE, A.; HONDA, M.; TANABE, M.; ONO, H. Antinociceptive effects of
sodium channel-blocking agents on acute pain in mice. Advance Publication. J Pharmacol Scienc. 2004.
SALONEN, L.; HELLDEN, L. Prevalence of signs and symptoms of dysfunction
in the masticatory system: na epidemiological study in na adult Swedish
population. J Craniomand Disord Facial Oral Pain, v.4, p.241-250, 1990.
SALT, T.E. The possible involvement of excitatory amino acids and NMDA
receptors in thalamic pain mechanisms and central pain syndromes. Am Pain Soc J. v.1, p. 52-54, 1992.
SAMSAM, M.; COVEÑAS, R.; AHANGARI, R.; YAJEVA, J.; NARVAEZ,
J.A.;TRAMU, G. Alterations in neurokinin A- substance P and calcitonine gene-
related peptide immunoreactives in the caudal trigeminal nucleous of the rat

168
following electrical stimulation of trigeminal ganglion. Neurosci. v. 261, p. 179-
182, 1999.
SARIA, A.; GAMSE, R.; PETERMANN, J.; FISCHER, J.A.;
THEORODORSSON-NORHEIN, E.; LUNDEBERG, J.M. Simultaneous release
of several tachykins and calcitonin gene-related peptide from rat spinal cord.
Neurosci. v. 63, p. 310-314, 1986.
SARLANI, E.; GREENSPAN, J.D. Evidence for generalized hyperalgesia in
temporomandibular disorders patients. Pain, v. 102, p.221-226, 2003.
SAWYNOK, J. Topical and peripherally acting analgesics. Pharmacol Reviews, v.55, p.1-20, 2003.
SBED – Sociedade Brasileira para o Estudo da Dor. Disponível em
:<http://www.dor.org.br>. 2004.
SCHAIBLE, H.G.; GRUBB, B.D. Afferent and spinal mechanisms of joint pain.
Pain. v.55, p. 5-54, 1993.
SCHÜTZ, T.C.B.; ANDERSEN, M.L.; TUFIK, S. Sleep alterations in an
experimental orofacial pain model in rats. Brain Res, v. 993, p. 164-171, 2003.
SERRA, J.; CAMPERO, M.; BOSTOCK, H.; OCHOA, J. Two types of C
nociceptors in human skin and their behavior in areas of capsaicin-induced
secondary hyperalgesia. J Neurophysiol., v.91, p.2770-2781, 2004.
SESSLE, B.J. Acute and chronic craniofacial pain: brainstem mechanisms of
nociceptive transmission and neuroplasticity , and their clinical correlates. Crit Rev Oral Biol Med. v. 11, p. 57-91, 2000.
SESSLE, B.J. Mechanisms of trigeminal and occipital pain. Pain Rev., v. 3, p.
91-116, 1996.
SESSLE, B.J.; HU, J.W. Mechanisms of pain arising from articular tissues. Can J Physiol Pharmacol, v. 69, p. 617-626, 1991.

169
SESSLE, B.J. Neural mechanisms and pathways in craniofacial pain. Can J
Neurol, v .26, p. 3-11, 1999.
SESSLE, B.J. Acute and chronic craniofacial pain: brainstem mechanisms of
nociceptive transmission and neuroplasticity and their clinical correlates. Crit Rev Oral Biol & Med. In press, 1999.
SHERMAN, J.E.; KALIN, N.H. ICV-CRH alters stress-induced freezing behavior
without affecting pain sensitivity. Pharmacol Biochem Behav, v.30 (4), p. 801-
807, 1988.
SHINODA, M.; HONDA, T.; OZAKI, N.; HATTORI, H.; MIZUTANI, H.; UEDA,
M.; SUGIURA, Y. Nerve terminal into the temporomandibular joint of adjuvant
arthritic rats. Eur. J. Pain , v.7, p.493-505, 2003.
SHORE, N.A. Occlusal equilibration and temporomandibular joint dysfunction.
Philadelphia: J B Lippincott, 1959.
SILVA, A. Disfunção da ATM .c.17, p.355-366, 1991.
SLUKA, K.A.; JORDAN, H.H.; WILLIS, W.D.; WESTLUND, KN. Differential
effects of NMDA and non-NMDA receptor antagonists on spinal release of
amino acids after development of acute arthritis in rats. Brain Res, v. 664, p.
77-84, 1994.
SOKOLOVA, R.I.; NEKRASOVA, A.A.; LEVITSKAIA, Iu.V.; SPERANSKAIA,
N.V.; VOLKOV, V. 2 Models of indomethacin-induced hypertension in rats in
chronic experiments. Kardiologiia, v. 17(10), p.78-85, 1977.
SPEARS, R.; RAENIE, O.; BELLINGER, L.L.; HUTCHINS, B. Tumor necrosis
factor-alfa and apoptosis in the rat temporomandibular joint. Arch of Oral Biol, v.48, p.825-834, 2003.
SPEARS, R.; HUTCHINS, B.; HILTON, R.J. Capsaicin application to the
temporomandibular joint alters calcitonin gene-related peptide levels in the
trigeminal ganglion of the rat. J.Orofacial Pain, v. 12, p. 108-115, 1998.

170
STEIN, C. Peripheral mechanisms of opioid analgesia. Anesth. Anal. v.76,
p.182-191, 1993.
STEIN, C.; HASSAN, A.H.S.; LEHRBERGER, K.; GIEFING, J.; YASSOURIDIS,
A. Local analgesic effect of endogenous opioid peptides. Lancet. v. 342, p.
321-324, 1993.
STEIN, C.; MILLAN, M.J.; HERZ, A. Unilateral inflammation of the hindpaw in
rats a model of prolonged noxious stimulation: alterations in behavior and
nociceptive thresholds. Pharmacol Biochem Behav, v.31, p. 441-445, 1988.
STEIN, C.; PFLUGER, M.; YASSOURIDIS, A.; HOELZL, J.; LEHRBERGER, K.;
WELTE, C.; HASSAN, A. No tolerance to peripheral morphine analgesia in
presence of opioid expression in inflamed synovia. J. Clin. Invest. v.98, p.793-
799, 1996.
STERNBERG, W.F.; WACHTERMAN, M.W. Experimental studies of sex-
related factors influencing nociceptive responses: animal studies. In: Fillingim
rb, ed: Prog in Pain Res and Manag.. v. 17. Sex, gender and pain. p. 71-88,
2000.
STIEFEL, F.S.D. Management of insomnia in patients with chronic pain
conditions. CSN Drugs, v 18(5), p.285-296, 2004.
STOHLER, C.S.; ASHTON-MILLER, J.A.; CARLSON, D.S. The effects of pain
from the mandibular joint and muscles on masticatory motor behavior in man.
Arch Oral Biol, v.33, p.175-182, 1988.
STRICHARTZ, G.R.; RITCHIE, J.M. The action of local anesthesics on ion
channels of excitable tissues. Berlim, 1987.
SVENSON, RM.; RANDALL, W.J. Grooming behavior in cats with pontile
lesions and cats with tectal lesions, J Comp. Physiol, v.91, p.313-326, 1977.
SVENSSON, P.;CAIRNS, B.E.; WANG, K.; ARENDT-NIELSEN, L. Injection of
nerve growth factor into human masseter muscle evokes long-lasting
mechanical allodynia and hyperalgesia. Pain, v.104, p.241-247, 2003.

171
TAIWO, Y.O.; LEVINE, J.D. Characterization of the aracchidonic metabolites
mediating bradykinin and noradrenaline hyperalgesia. Brain Res. v. 458, p.402-
406, 1988.
TATSUO, M.A.; CARVALHO, W.M.; SILVA, C.V.; MIRANDA, A.E.; FERREIRA,
S.H.; FANCISCH, J.N. Analgesics and antiinflammatory effects of dypirone in a
rat adjuvant arthritis model. Inflam., v. 18, p.399-405, 1994.
TERAYAMA, R.; DUBNER, R.; REN, K. The roles of NMDA receptor activation
and nucleous reticularis gigantocellularis in time-dependent changes in
descending inhibition after inflammation. Pain. v. 97, p. 171-181, 2002.
TERAYAMA, R.; GUAN, Y.; DUBNER, R.; REN, K. Activity-induced plasticity in
brain stem pain modulatory circuitry after inflammation. Neuroreport. v.11, p.
1915-1919, 2000.
TJOLSEN, A.; BERGE, O.G.; HUNSKAAR, S.; ROSLAND, J.H.; HOLE, K. The
formalin test: an evaluation of the method. Pain, v.51(1), p. 5-17, 1992.
TOMACHESKI, D. F.; BARBOZA, V. L.; FERNANDES, M.R.; FERNANDES, F.
Disfunção temporomandibular: Estudo introdutório visando a estruturação de
prontuário odontológico. Plub. UEPG Biol. Health Sci, v. 10 (2), p.17-25, 2004.
TOMINAGA, M.; CATERINA, M.J.; MALMBERG, A.B. The cloned capsaicin
receptor integrates multiple pain-producing stimuli. Neuron. v.21, p.531-543,
1998.
TOREBJÖRK, H.E.; LUNDBERG, L.E.; LaMOTTE, R.H. Central changes in
processing of mechanoreceptive input in capsaicin-induced secondary
hyperalgesia in humans. J Physiol. v.448, p. 765-780, 1992.
TUERLINGS, V.; LIMME, M. The prevalence of temporomandibular joint in the
mixed dentition. Eur J Orthod. v. 26 (3), p. 311-320, 2004.
UENO, R.; HAYAISH, O.; INOUE, S.; NAKAYAMA, T. Prostaglandins D: a
cerebral sleep-regulating substance in rats. Adv. Prostaglandins,
Tromboxanane, Leukotriene Res. v.15, p.581-583, 1985.

172
ULFGREN, A.K.; GRONDAL, L.; LINDBLAD, S.; KHADEMI, M.; JOHNELL, O.;
KLARESKOG, L. In interindividual and intra-articular variation of
proinflammatory cytokines in patients with rheumatoid arthritis: potential
implications for treatment. Ann Rheum Dis. v. 59, p. 439-447, 2000.
URBAN, M.O.; COUTINHO, S.V.; GEBHART, G.F. Involvement of excitatory
amino acid receptors and nitric oxide in the rostral ventromedial medulla in
modulation secondary hyperalgesia produced by mustarda oil. Pain. v. 81, p.
45-55, 1999.
VANE, J.R. Inhibition of prostaglandin synthesis as a mechanism of action for
aspirin-like drugs. Nature, v. 231, p. 232-235, 1971.
VAN DER MEER, M.J.; SWEEP, C.G.; RIJNKELS, C.E.; PESMAN, G.J.;
TILDERS, F.J.; KLOPPENBORG, P.W. et al. Acute stimulation of the
hypothamic-pituitary-adrenal axis by IL-1 beta, TNF-alpha and IL-6: a dose
response study. J Endo Invest , v.19, p. 175-182, 1996.
VANEGAS, H.; SCHAIBLE, H. Prostaglandins and ciclooxygenases in the
spinal cord. Progr in Neurobiol, v.64, p. 327-363, 2001.
VAUGHAN, C.W.; INGRAM, S.L.; CONNOR, M.A.; CRISTIE, M.J. How opioids
inhibit GABA-mediated neurotransmission. Nature , v.390, p. 611-614, 1997.
VAUGHT, J.L.; ROTHMAN, R.B.; WESTFALL, T.C. Mu and delta receptors :
their role in analgesia and in the differential effects of opioid peptides on
analgesia. Life Sci., v.30, p. 1443-1455, 1982.
VON KORFF, A.; DWORKIN, S.F.; LE RESCHEL; KRUGER, A. An
epidemiologic comparison of pain complaints. Pain , v.32, p.173-183, 1988.
VON KORFF M. Health services research and temporomandibular pain . In;
Sessle BJ, Bryant PS, Dionne ra (eds). Progress in pain research and
management. IASP press, v.4, p.227, 1996.

173
VOS, B.P.; HANA, G.; ADRIAENSEN, H. Behavioral assesment of facial pain in
rats: face grooming patterns alter painful and non-painful sensory disturbance in
the territory of the rat’s infraorbital nerve. Pain, v. 76, p. 173-178, 1998.
WANG, B.C.; LI, D.; BUDZILOVICH, G.; HILLER, J.M.; ROSENBERG, C.;
HILLMAN, D.E.; TURNDORF, H. Antinociception without motor blockade after
subarachnoid administration of S-(+) ibuprofen in rats. Life Sci, v.54, p.715-
720, 1994.
WANG, H.; WESSENDORF, M.W. Equal proportions of small and large DRG
neurons express opioid receptor mRNAs. J Comp Neurol, v.429, p.590-600,
2001.
WASNER, G.; SCHATTSCHNIDER, J.; BINDER, A.; BARON, R. Topical
menthol- a human model for cold pain by activation and sensitization of C
nociceptors. Brain v.127, p. 1159-1171, 2004.
WEBSTER, K. E. Somaesthetic pathways. Br Med Bull, v.33, p.113-120, 1971.
WILLCOCKSON, W.S.; CHUNG, J.M.; HORI, Y.; LEE, K.H.; WILLIS, W.D.
Effects of iontophorectically release amino acids and amines on primate
spinothalamic tract cells. J Neurosci, v.4, p.732-740, 1984.
WILLIS, W.D. Anatomy and physiology of descending control of nociceptive
responses of dorsal horn neurons: comprehensive review. Brain Res. v. 77,
p.1-29, 1988.
WOMACK, M.D.; MacDERMOTT, A.B.; JESSELL, T.M. Sensory transmitters
regulate intracellular calcium in dorsal horn neurons. Nature, v.334, p.351-353,
1988.
WOMACK, M.D.; MacDERMOTT, A.B.; JESSELL, T.M. Substance P increase
calcium in dorsal horn neurons via two distinct mechanisms. Soc. Neurosci. Abs.15, p.184, 1989.
WOOLF,C. J.; MANNION, R.J. Neuropathic pain: a etiology, symptoms,
mechanisms and management. Lancet, v. 353, p. 1959-1964, 1999.

174
XIANG, Z.; SI, J.; HUANG, H. Chronic gastric rat model and role of inducing
factors. World J Gastroenterol, v.10 (21), p.3212-3214, 2004.
YEOMANS, D.C.; PIREC, V.; PRODFIT, H.K. Nociceptive responses to high
and low rates of noxious cutaneous heating are mediated by different
nociceptors in the rat: behavioral evidence. Pain, v.68, p.133-140, 1996.
YU, X.M.; SESSLE, B.J.; HU, J.W. Differential effects of cutaneous and deep
application of inflammatory irritant on mechanoreceptive field properties of
trigeminal brain stem nociceptive neurons. J. Neurophysiol. v. 70, p. 1704-
1707, 1993.
YU, X.M.; SESSLE, B.J.; HAAS, D.A.; IZZO, A.; VERNON, H.; HU, J.W.
Involvement of NMDA receptor mechanisms in jaw eletromyographic activity
and plasma extravasation induced by inflammatory irritant application to
temporomandibular joint region of rats. Pain. v. 68, p. 169-178, 1996.
YU, X.M.; SESSLE, B.J.; HAAS, D.A.; IZZO, A.; VERNON, H.; HU, JW. Effects
MK-801 on jaw muscle activity and plasma extravasation induced by mustard oil
injected into temporomandibular joint (TMJ). Abstr. Soc. Neurosci. v.20, p.763,
1994.
ZAMMA, T. Adjuvant-induced arthritis in the temporomandibular joint of rats.
Infec and Immun., v.39, p.1291-1299, 1983.
ZHANG, G.; WANG, L.; LIU, H.; ZHANG, J. Substance P promotes sleep in the
ventrolateral preoptic area of rats. Brain Res, v. 1028, p. 225-232, 2004.
ZHANG, S.; TANG, J.S.; YUAN, B.; JIA, H. Inhibitory effects of electrical
stimulation of ventrolateral orbital cortex on the rat jaw-opening reflex. Brain Res. v.813, p.359-366, 1998.
ZHOU, M.; GEBHART, G.F. Biphasic modulation of spinal nociceptive
transmission from the medullary raphe nuclei in the rat. J of Neurophysiol. v.
78, p.746-758, 1997.

175
ZHOU, Q.; HIROKI, I.; DUBNER, R.; REN, K. Persistent fos protein expression
after orofacial deep or cutaneous tissue inflammation in rats: Implications for
persistent orofacial pain. The J of Compar. Neurol. v.412, p. 276-291, 1999.
ZIMMERMANN, M. Ethical guidelines for investigations of experimental pain in
conscious animals. Pain, v.16, p.109-110, 1983.