ultraviolet-b induced inflammation of human skin: characterisation and comparison with traditional...
TRANSCRIPT

European Journal of Pain 13 (2009) 524–532
Contents lists available at ScienceDirect
European Journal of Pain
journal homepage: www.EuropeanJournalPain.com
Ultraviolet-B induced inflammation of human skin: Characterisationand comparison with traditional models of hyperlagesia
Thomas Bishop a, Angela Ballard a, Helen Holmes a, Antony R. Young b, Stephen B. McMahon a,*
a Neurorestoration Group, The Wolfson Centre for Age Related Disease, King’s College London, The Wolfson Wing, Hodgkin Building, Guy’s Campus, London Bridge,London SE1 1UL, United Kingdomb King’s College London, St. John’s Institute of Dermatology, Division of Genetics & Molecular Medicine, Tower Wing, Guy’s Hospital, London SE1 9RT, United Kingdom
a r t i c l e i n f o a b s t r a c t
Article history:Received 29 February 2008Received in revised form 27 May 2008Accepted 7 June 2008Available online 8 August 2008
Keywords:Primary hyperalgesiaSecondary hyperalgesiaSunburnUltraviolet-BNociceptor sensitisation
1090-3801/$36.00 � 2008 Published by Elsevier Ltd odoi:10.1016/j.ejpain.2008.06.006
* Corresponding author. Tel.: +44 (0)20 7848 6270E-mail address: [email protected] (S.B.
The effect on human skin of over-exposure to solar ultraviolet radiation (UVR) has been well described.The erythema produced is commonly referred to as ‘sunburn’. Recently UVR induced inflammation hasbeen utilised as a human model of sub-acute pain. Our aim was to characterise the sensory phenotypeof UVB inflammation in human volunteers. We delivered UVB to small areas of volar forearm skin inhealthy volunteers and found that the degree of inflammation and concomitant increase in sensitivityto cutaneous stimuli were UVB dose and time dependant. We directly compared UVB induced inflamma-tion and the more established thermal burn and topical capsaicin pain models. UVB inflammation pro-duced precisely demarcated erythematous lesions without secondary flare. Both thermal burns andtopical capsaicin produced large areas of flare, indistinguishable in character from the primary lesions.Moreover, UVB inflammation induced large reductions in mechanical pain threshold restricted to the pri-mary lesion site, whereas the more established inflammatory pain models produced not only primaryhypersensitivity but also significant areas of secondary mechanical hypersensitivity. Taken together thesefindings suggest that UVB inflammation, at least using moderate doses produces sensory changes primar-ily by sensitising peripheral pain processing in the relative absence of alterations in central painprocessing.
� 2008 Published by Elsevier Ltd on behalf of European Federation of Chapters of the InternationalAssociation for the Study of Pain.
1. Introduction
Inflammatory pain is a common clinical state but there are fewhuman experimental models available (see Staahl and Drewes,2004 for review). Recently several groups have begun to assessthe effects of UVR on human skin. UVR, especially UVB (280–315 nm), produces a well described erythema (‘‘sunburn”), maximalafter approximately 24 h with concomitant reductions in thermaland mechanical pain thresholds (allodynia) and increased re-sponses to suprathreshold stimulation (hyperalgesia) at the siteof UVR injury (Harrison et al., 2004; Benrath et al., 2001; Bickelet al., 1998; Hoffmann and Schmelz, 1999; Gustorff et al., 2004a).
It is well recognised that tissue trauma can trigger erythemathat extends into adjacent uninjured tissues (Lewis, 1927). In someinflammatory pain states (including thermal burns and topicalcapsaicin application) this flare response is accompanied by an in-creased sensitivity to mechanical stimulation in areas surroundingthe injury (Lewis, 1935; Hardy et al., 1950; Raja et al., 1984; Ali
n behalf of European Federation o
; fax: +44 (0)20 7848 6165.McMahon).
et al., 1996; Meyer et al., 2006); so-called secondary hyperalgesia.The changes in mechanical sensitivity in secondary areas are nowthought to be largely due to activity-dependent plasticity of centralneuronal pathways, prominent in the spinal cord (LaMotte et al.,1991; Torebjork et al., 1992; Koltzenburg, 2000; Klede et al.,2003; Woolf and Salter, 2006). Brush-evoked allodynia and pin-prick hyperalgesia appear to represent circuits activated by Ab-fi-bres (Torebjork et al., 1992; Koltzenburg et al., 1994) and Ad-fibres(Ziegler et al., 1999; Magerl et al., 2001; Fuchs et al., 2000),respectively.
Previous experiments from our laboratory (Harrison et al.,2004) have suggested that changes in mechanical pain thresholdsare restricted to the primary UVR induced lesion site with nonein secondary skin regions, supporting the hypothesis that periphe-ral sensitisation predominates as the underlying mechanism. How-ever this finding is disputed by the work of another group whoobserved a large secondary area of pin-prick hyperalgesia follow-ing UVB induced inflammation (Sycha et al., 2003, 2005; Gustorffet al., 2004a,b). This latter claim was taken as evidence supportingthe hypothesis that central sensitisation contributes to theUV-induced sensory changes.
f Chapters of the International Association for the Study of Pain.

600 Naïve1MED
T. Bishop et al. / European Journal of Pain 13 (2009) 524–532 525
Our aim here was to characterise the time course and dosedependence of inflammation and sensory changes induced byUVB irradiation and to compare these to sensory changes producedusing more established paradigms [thermal burn (Pedersen andKehlet, 1998; Dirks et al., 2003; Norbury et al., 2007) and topical1% capsaicin application (Kilo et al., 1994; Mohammadian et al.,1998; Liu et al., 1998)]. UVB inflammation produced well demar-cated erythematous lesions without secondary flare. Both thermalburns and topical capsaicin produced large areas of flare indistin-guishable from the primary lesions. Moreover, UVB inflammationinduced reductions in mechanical pain threshold restricted to theprimary lesion site, whereas the other models produced both pri-mary hypersensitivity and significant areas of secondary mechan-ical hypersensitivity. Together these findings suggest that UVBinflammation produces sensory changes primarily by alteringperipheral pain processing and do so by sensitizing a subgroup ofprimary sensory neurones.
0 6 24 48 72 96
0
100
200
300
400
500 2MED3MED
Flu
x (
AU
)
Time Post Irradiation (hrs)
Naïve 1MED 2MED 3MED
0
500
1000
1500
2000
2500
3000
UVB Dose
% Incre
ase In
Cuta
neous B
lood F
low
*
**
Fig. 1. Acute cutaneous UVB exposure produces dose-dependent erythema. (A)Shows the experimental UVB burn 24 h post-irradiation induced to investigate thetime course and dose–response of erythema and sensory changes. From left to rightdoses of 1, 2 and 3 MED can be seen on the volar aspect of the forearm. (B) Showsthe time course of the erythema following exposure of the skin to UVB (naïve [shamirradiated], 1, 2 and 3 MED) which is dose-dependent. (C) Shows the percentageincrease in cutaneous blood flow at the peak of erythemal changes, 24 h post-irradiation, with doses of 1, 2 and 3 MED producing significant changes compared tosham irradiated skin (*p < 0.05, one-way ANOVA followed by a Tukey’s post hoctest; n = 12 for all groups).
2. Methods
2.1. Volunteers
A total of 27 healthy adult volunteers of skin types I/II wereinvestigated, none was taking medication. They gave informedconsent to take part in the studies that were approved by theEthics Committee of King’s College London and conformed to Dec-laration of Helsinki guidelines and IASPs guidelines for experimen-tation in humans (Charlton, 1995). All test sites were on previouslyun-irradiated volar forearm skin. Experiments were performedduring the autumn and winter to limit the effect of environmentalUVR exposure. The subjects each received remuneration for partic-ipating in the studies and could withdraw at anytime withoutjeopardising this remuneration.
2.2. Study protocols
Experiments were carried out in three blocks.
2.2.1. Study I: time course and dose dependence of UVB inducedsensory changes
In study I, we assessed the time- and dose-dependent effects ofUVB irradiation in 12 volunteers (#/$ 8/4; age 27.3 ± 2.4; MED476 ± 20.6 mJ/cm2). Volunteers had three 10 � 10 mm patches ofvolar forearm skin irradiated with 1, 2 and 3 MED of UVB, whichproduced the characteristic changes in skin appearance illustratedin Fig. 1A. The endpoints assessed were changes in cutaneousblood flow as well as alterations in the pain sensitivity to thermaland mechanical stimuli of the primary lesion site. Measurementswere taken at 2, 4, 6, 24, 48, 72 and 96 h after irradiation. Measure-ments were also taken at the same time from a naïve (un-irradi-ated) patch of skin.
2.2.2. Study II: comparison of sensory changes in the primary andsecondary skin areas induced by UVB inflammation, topical 1%capsaicin and thermal burn models of pain
In study II, we assessed the emergence of changes in both the pri-mary lesion site and in adjacent but uninjured skin (secondary area).We compared inflammation induced by 3 MED of UVB with twomore established models of pain: cutaneous thermal burn and top-ical application of 1% capsaicin. In both these cases, lesions of32 � 32 mm and made, located approximately equidistant betweenthe elbow and wrist. The outcome measures taken within the pri-mary lesion were alterations in cutaneous blood flow and thermaland mechanical pain thresholds. The outcome measures taken with-in the secondary area were measurements of flare (erythema in sec-
ondary areas of skin), brush-evoked allodynia and pin-prickhyperalgesia. Measurements were taken: 24 h after induction ofUVB inflammation; following 30 min of capsaicin application; and15 min after the induction of the thermal burn. These time pointsrepresent the peak of sensory changes as defined by study I in thecase of UVB inflammation (see Section 3); and by previous authorswith regards to topical capsaicin and by Norbury et al. (2007) withregards to the thermal burn. These experiments were performedon a cohort of 12 volunteers (#/$ 8/4; age 26.6 ± 2.4 years; MED439 ± 28.3 mJ/cm2) some of whom had been used in study I.

526 T. Bishop et al. / European Journal of Pain 13 (2009) 524–532
2.2.3. Study III: distribution of changes in sensitivity to mechanicalstimulation around annular lesions induced with UVB inflammationand topical capsaicin application
In study III, we determined if changes in skin mechanical painthreshold were localized to the site of UVB irradiation in compar-ison with those provoked by topical capsaicin application. In 12volunteers (#/$ 9/3; age 24.4 ± 1.0 years; MED 454 ± 25.7 mJ/cm2) an annulus-shaped lesion of the skin (see Fig. 4B and C) wereinduced either with 3 MED of UVB or application 1% topical capsa-icin, approximately equidistant between the elbow and wrist. Theannulus was 50 mm in diameter and had a 20 mm central un-stim-ulated zone. Twenty-four hours after irradiation with UVB and30 min after topical capsaicin application (the respective peaks ofsensory changes), we tested the mechanical sensitivity at sites inand around the lesions sites.
2.3. Induction of inflammatory models
2.3.1. UVB irradiation protocolThe UVB source used for all experiments was the TL01 (Phillips,
UK, kmax = 311 nm), the irradiance produced by the tubes duringeach irradiation was determined using a calibrated meter(IL1400A with SEL240/UVB-1/TD filter [ABLE Instruments &Controls Ltd., UK] calibrated by St Thomas’ Medical Physics Depart-ment, London, UK) and the duration of exposure altered accord-ingly. The irradiation room was air conditioned at 20 �C and a fanwas positioned to blow over the irradiated site to minimize any in-creases in skin temperature. All volunteers had their Individualsensitivity to UVR assessed by the determination of the minimalerythemal dose (MED) defined as the dose required to produce ery-thema with a clear border 24 h after exposure. This was done byexposing six 10 � 10 mm areas of volar forearm skin to a UVR doseseries with
p2 increments. With MED identified for each subject in
this preliminary trial, multiples of the MED were used to generatethe lesions described above for studies I–III using exactly the samemethod as described above.
2.3.2. Topical capsaicin protocolThe topical capsaicin pain model was induced using 1% capsai-
cin (Sigma, UK) made up in 70% alcohol (Kilo et al., 1994; Moham-madian et al., 1998; Liu et al., 1998). Filter paper was soaked in thissolution and applied under an occlusion dressing (Tegaderm, 3 M)to the volar forearm for 30 min. The filter paper was a square32 � 32 mm in study II and an annulus (50 mm in diameter witha 20 mm hole in the centre) in study III. The dressing was then re-moved with care taken not to dislodge the filter paper and an ace-tate template used to delineate the edge of capsaicin application.Outwardly radiating spokes were marked along the skin with, onwhich testing of mechanical sensitivity was performed.
2.3.3. Thermal burn protocolThermal burn injuries in study II were created with a servo con-
trolled peltier device (TSA-II, Medoc, Israel) using a 32 � 32 mmprobe. The probe was placed on the volar surface of the forearm,approximately equidistant from the elbow and wrist, and securedwith a fabric strap. A manual sphygmomanometer cuff waswrapped over the probe and inflated to a pressure of 20 mmHgto standardise the contact pressure of the probe on the skin and en-sure the probe did not move during the creation of the burn. Theprobe was held at an adaptation temperature of 32 �C, before beingheated to 45 �C at a rate of 0.5 �C/s and maintained at this temper-ature for 330 s. Once the thermode was removed the edges of theprimary burn site were marked on the skin with a pen and an ace-tate template was used to mark spokes radiating out from the pri-mary burn area. This model was adapted from Pedersen and Kehlet(1998) by Norbury et al. (2007). Assessment of the changes in skin
blood flow and sensitivity to thermal and mechanical stimuli werecarried out 15 min after the removal of the thermode; this time-point represents the peak of the changes in this model.
2.4. Measurement of superficial blood flow within primary lesion sites
Superficial blood flow at the test sites was measured by laserDopplar flowmetry. A laser Dopplar probe (MoorLDI, Moor Instru-ments Ltd., UK) was held against the skin using a ring of doublesided adhesive tape and data collected for at least 10 s, from whichan average flux was calculated.
2.5. Sensory testing
Prior to experimentation, the volunteers had a training sessionto familiarise themselves with the sensations caused by the testsystems and define their baseline heat and mechanical painthresholds. Throughout sensory testing volunteers were instructedto keep their eyes shut.
2.6. Heat pain thresholds
Sensitivity to thermal challenge was assessed with a TSA-2001Thermal Sensory Analyzer (Medoc, Israel). A 10 � 10 mm probewas held against the subject’s skin. The initial holding temperatureof the probe was 30 �C. Skin temperature was then increased at arate of 0.5 �C s�1 until the volunteer judged the heat stimulus pain-ful, which was signaled by the subject with an electronic push but-ton. A computer logged the temperature which was taken as thethermal pain threshold. Immediately after signaling the painthreshold, the thermal probe is rapidly cooled back to a tempera-ture of 30 �C. For each measurement site the average of three stim-uli was taken.
2.7. Mechanical pain thresholds
Sensitivity to mechanical challenge was assessed with an elec-tronic von Frey system (Somedic, Hörby, Sweden). This devicehas a plastic monofilament (1 mm in diameter) attached to a trans-ducer, which records applied pressure. The probe is held by theexperimenter and pushed against the skin to increase pressure ata rate of 4 g/s. The pressure increase was monitored on a computerscreen, and the experimenter matches the applied force against atemplate of required force. Pressure was increased until subjectsjudged the stimulus to be painful, at which time they indicatedwith an electronic push button. The force exerted at this timewas recorded as the mechanical pain threshold.
2.8. Changes in secondary skin areas
2.8.1. FlareFollowing the induction of inflammatory lesions the edges of
the primary burn site were marked on the skin and an acetate tem-plate was used to mark points at 10 mm increments along eightspokes radiating out from the primary burn area.
The extent of the flare along each spoke was measured by eye tothe nearest 5 mm.
2.8.2. Brush-evoked allodyniaThe degree of brush-evoked allodynia was assessed with a No. 2
sable paintbrush (Justbrushes, UK). The brush stimulus consisted offour 10 mm long brush strokes applied perpendicularly to eachpoint along the spoke, drawn previously to assess flare, at a fre-quency of approximately 0.5 Hz. The brush was initially appliedto the outermost point of each spoke, to serve as a reference pointfor normal sensation, then applied to each point in turn working

T. Bishop et al. / European Journal of Pain 13 (2009) 524–532 527
towards the primary burn area until a change from a ‘soft or tickly’to a ‘scratchy or prickly’ sensation was reported and recorded.
2.8.3. Pin-prick hyperalgesiaThe degree of pin-prick hyperalgesia was assessed with a
retractable 10 g von Frey filament (Bailey Instruments, UK). Thetesting procedure was similar to that used to assess brush-evokedallodynia. The von Frey stimulus was applied once to each point,again starting from the outer most point, with pressure beingmaintained for approximately 1 s. The point at which a sensationchange described by subjects as a shift from ‘an innocuous prod-ding’ to a ‘sharp pricking’ was recorded.
The area of secondary (not including the primary lesion area)flare, brush-evoked allodynia and pin-prick hyperalgesia was sub-sequently calculated.
2.9. Statistical analysis
All values, including percentage changes, are expressed asmean ± standard error of the mean (SEM). All statistical analysiswas carried out using SigmaStat (SPSS, IL, USA). Details of the testsperformed can be found in Section 3 and in the figure legends. In allcases a p value of 0.05 or less was taken as significant. Dose-depen-dency was tested statistically by undertaking linear regressionanalysis using GraphPad Prism v4.00 (GraphPad Software, San Die-go, CA, USA) and testing whether the slope of this line was signif-icantly greater than zero.
3. Results
3.1. UVB irradiation produces a dose-dependent increase in cutaneousblood flow (Fig. 1)
Sites irradiated with one or more MED of UV showed highlyconsistent and statistically significant increases in cutaneous bloodflow at 24 h (p < 0.05, one-way ANOVA followed by a Tukey’s posthoc test; n = 12 for all groups; Fig. 1). The peak of erythema for allUVB doses tested was seen at 24 h, and at this time, there was aclear dose–response between increased cutaneous blood flowand UVB irradiation as demonstrated by a positive slopes of the lin-ear regression analysis on the data in Fig. 1C (slope significantlygreater than zero, p < 0.01, t-test). The percentage increases inblood flow compared to baseline for doses 1, 2 and 3 MED at24 h were 513 ± 213%, 1959 ± 287% and 2487 ± 247%, respectively.Though the increased blood flow produced by UVB peaked at 24 hit was evident (and statistically significant) from 2 h after irradia-tion with all doses tested (p < 0.05, one-way ANOVA followed bya Tukey’s post hoc test; n = 12 for all groups; data not shown). Skinexposed to 2 and 3 MED also sometimes showed oedema. The ery-thema gradually faded over the next few days. The only apparentlonger-term consequence of irradiation was tanning of the skinat irradiated sites. None of the volunteers irradiated with thesedoses (up to 3 MED) developed blisters or other signs of loss of skinbarrier function which would preclude the use of these doses as asterile model of cutaneous inflammatory pain.
3.2. UVB inflammation produces dose-dependent decreases in thermaland mechanical thresholds without spontaneous pain sensations(Fig. 2)
Using 1, 2 and 3 MED we irradiated the volar skin of the forearmand examined changes in sensory function. Volunteers did not re-port ongoing (unprovoked) pain at any of the sites at any timepoint. Significant decreases in thermal and mechanical painthresholds emerged following treatment with all UVB doses
(p < 0.05, one-way ANOVA followed by a Tukey’s post hoc test;n = 12 for all groups; Fig. 2). The peak of sensory changes for alldoses correlated with the peak in increased blood flow and wasseen at 24 h. At this time there is a clear UVB dose-dependencyfor the changes in both thermal and mechanical pain thresholds,as demonstrated by a positive slopes of the linear regression anal-ysis on the data in Fig. 2B and D (slope significantly greater thanzero in both cases, p < 0.01, t-test).
The mean drop in thermal pain thresholds at the peak of sen-sory changes compared to naïve un-irradiated skin (the percentagedrop is shown in brackets) were 2.7 ± 0.7 �C (5.8 ± 1.5%),5.4 ± 0.5 �C (11.5 ± 1.0%) and 6.2 ± 0.5 �C (13.2 ± 1.1%) for UVBdoses of 1, 2 and 3 MED, respectively. The mean drops in mechan-ical pain thresholds at the peak of sensory changes compared tobaseline were 28.8 ± 7.1 g (16.6 ± 7.6%), 52.4 ± 9.9 g (32.4 ± 7.5%)and 73.1 ± 3.6 g (44.4 ± 6.4%), respectively, for UVB doses of 1, 2and 3 MED. All of these changes were statistically significant(p < 0.05, ANOVA followed by Tukey’s post hoc test; n = 12 for allgroups).
The emergence of significant decreases in thermal pain thresh-olds was seen at 4 h post-irradiation with doses of 1 and 2 MED,whereas 3 MED produces significant decreases at 2 h post-irradia-tion (p < 0.05, ANOVA followed by Tukey’s post hoc test; n = 12 forall groups; data not shown). The emergence of significant de-creases in mechanical pain thresholds was seen at 4 h post-irradi-ation with all UVB doses tested [1, 2 and 3 MED (p < 0.05, ANOVAfollowed by Tukey’s post hoc test; n = 12 for all groups; data notshown)].
The duration and resolution of sensory changes was UVB dose-dependent (p < 0.05, ANOVA followed by Tukey’s post hoc test) andclosely paralleled the time course of erythema and increased bloodflow observed following irradiation.
3.3. UVB inflammation does not produce secondary areas of flare,brush-evoked allodynia or pin-prick hyperalgesia
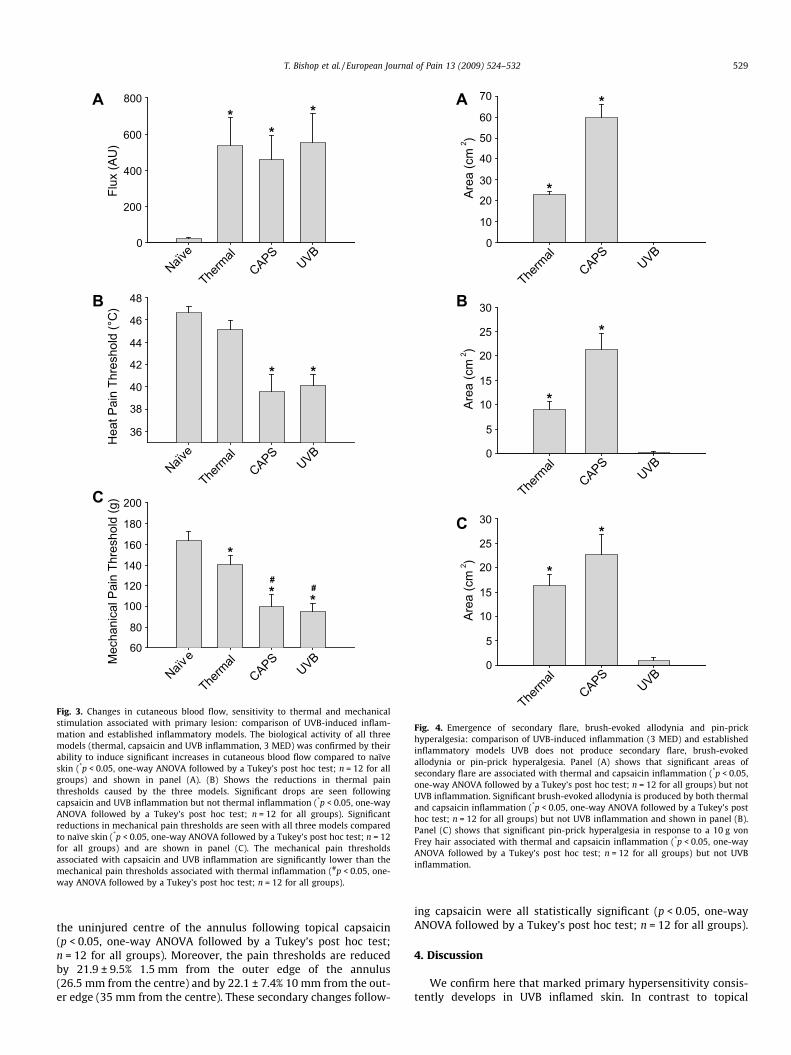
Despite producing significant alterations in blood flow, heatpain threshold and mechanical pain threshold in the primary le-sion site (p < 0.05, ANOVA followed by Tukey’s post hoc test;n = 12 for all groups; Fig. 3), UVB did not produce areas of flare,brush-evoked allodynia or pin-prick hyperalgesia outside of theirradiated site (Fig. 4). This is in contrast to topical capsaicin appli-cation and thermal burn lesions, both of which produce significantareas of flare, brush-evoked allodynia and pin-prick hyperalgesia(p < 0.05, ANOVA followed by Tukey’s post hoc test; n = 12 for allgroups; Fig. 4) as well as altering pain thresholds associated withthe primary lesion site (p < 0.05, ANOVA followed by Tukey’s posthoc test; n = 12 for all groups; Fig. 3).
The areas of secondary flare, brush-evoked allodynia and pin-prick hyperalgesia produced by topical capsaicin were significantlylarger than those produced by thermal burn (p < 0.05, ANOVA fol-lowed by Tukey’s post hoc test; n = 12 for all groups; Fig. 4). Thearea of flare produced by capsaicin was 59.6 ± 6.4 cm2 comparedto 23 ± 1.5 cm2 produced by the thermal burn. The areas ofbrush-evoked allodynia produced by capsaicin and the thermalburn were 21.3 ± 3.4 cm2 and 9.0 ± 1.6 cm2, respectively. The areasof pin-prick hyperalgesia produced by capsaicin and the thermalburn were 22.7 ± 4.1 cm2 and 16.3 ± 2.3 cm2, respectively.
All three models produced biological effects as indicated by theincreased blood flow induced in the primary lesion site in all cases.These increases in blood flow were not significantly differentbetween models, although the changes in pain thresholds were.Both UVB inflammation and topical capsaicin produced largerreductions in the thermal and mechanical pain thresholds com-pared with the rather modest drops produced by thermal lesions(p < 0.05, ANOVA followed by Tukey’s post hoc test; n = 12 for all

Naïve1MED2MED3MED
Heat
Pa
in T
hre
shold
(°C
)
0 6 24 48 72 96
Time Post Irradiation (hrs)
38
40
42
44
46
48
50
Naïve 1MED 2MED 3MED
UVB Dose
0
4
8
12
16
Heat
Pa
in T
hre
shold
% D
ecre
ase In
Naïve1MED2MED3MED
Mechanic
al P
ain
Th
reshold
(g)
0 6 24 48 72 96
Time Post Irradiation (hrs)
60
80
100
120
140
160
180
200
Naïve 1MED 2MED 3MED
UVB Dose
-20
0
20
40
60
Mechanic
al P
ain
Th
reshold
% D
ecre
ase In
*
**
**
*
Fig. 2. Acute cutaneous UVB exposure produces dose-dependent changes in sensitivity to thermal and mechanical stimulation. Acute cutaneous UVB over-exposure of thevolar surface of the upper limb produces dose-dependent changes in the sensitivity to thermal and mechanical stimulation. (A and C) Show the time course of the changes inthermal and mechanical pain thresholds, respectively, for volunteers irradiated with doses of naïve (sham irradiated), 1, 2 and 3 MED. The peak of sensory changes correlatedwith the peak of erythema at 24 h post-irradiation. (B) Shows the percentage decrease in thermal pain threshold at 24 h post-irradiation. Similarly (D) shows the percentagedecrease in mechanical pain threshold at the same time point. Significant changes in thermal and mechanical sensitivities are seen with doses that produce erythema (1, 2and 3 MED; *p < 0.05, one-way ANOVA followed by a Tukey’s post hoc test; n = 12 for all groups).
528 T. Bishop et al. / European Journal of Pain 13 (2009) 524–532
groups; Fig. 3). The mean reduction in thermal pain thresholdsfollowing thermal burn, topical capsaicin and 3 MED UVBinflammation were 1.5 ± 0.5 �C (3.1 ± 1.1%), 7.1 ± 1.0 �C (15.2 ±2.1%) and 6.5 ± 0.6 �C (13.9 ± 1.3%), respectively. The mean reduc-tion in mechanical pain thresholds following thermal burn, topicalcapsaicin and UVB inflammation were 21.5 ± 8.1 g (13.1 ± 5.0%),63.8 ± 8.8 g (39.0 ± 5.4%) and 68.0 ± 6.4 g (41.6 ± 3.9%), respec-tively.
Moreover, brush-evoked allodynia and pin-prick hyperalgesiawas present in the primary lesion site of all models.
Both the capsaicin and thermal burn models were associatedwith marked ongoing pain during their creation. Spontaneousongoing pain lasted for tens of minutes after the removal of thetopical capsaicin application. In the case of the thermal injurymodel ongoing pain ended when the thermal stimuli ceased. Incontrast there were no reports of spontaneous ongoing pain asso-ciated with UVB exposure or at any time after the lesionsinduction.
3.4. UVB inflammation produces reductions in mechanical painthresholds that are restricted to the primary lesion (Fig. 5)
The lesions produced by UVB are remarkable in that they areprecisely demarcated and restricted to the skin that is exposed tothe irradiation. This is unlike most inflammatory lesions, includingthe capsaicin and thermal burn models, where prominent areas of
flare in adjacent un-injured skin are common. This comparison isapparent in panels b (UVB annulus) and c (annulus application oftopical capsaicin) of Fig. 5. The mechanical allodynia produced fol-lowing UVB inflammation of an annular region of volar forearmskin was highly localised within the boundaries of the primary le-sion (p < 0.05, ANOVA followed by Tukey’s post hoc test; n = 12 forall groups; Fig. 5E). This is in contrast to the pattern of mechanicalallodynia produced following the annular application of topicalcapsaicin where mechanical pain thresholds are reduced in adja-cent un-stimulated secondary areas of skin (p < 0.05, ANOVA fol-lowed by Tukey’s post hoc test; n = 12 for all groups; Fig. 5F).
As expected, the mechanical pain thresholds are reduced by�40% within the UVB irradiated area (reduced by 40.1 ± 6.0% at15 mm from the centre of the annulus and 37.3 ± 6.5% at the edgeof the annulus 23.5 mm from the centre) but unchanged in theuninjured centre of the annulus and at no point outside the annu-lus including those points 26.5 mm from the centre of the annulus(which was only 1.5 mm from the edge of the inflamed skin).
The mechanical pain thresholds recorded from the within theprimary lesion site following topical capsaicin were also signifi-cantly reduced (p < 0.05, one-way ANOVA followed by a Tukey’spost hoc test; n = 12 for all groups); by 34.4 ± 7.6% at 15 mm fromthe centre of the annulus and 27.2 ± 8.3% at the edge of the annulus23.5 mm from the centre. In contrast to the lack of changes seen inthe secondary areas associated with UVB inflammation, mechani-cal pain thresholds were significantly reduced by 24.6 ± 8.9% in
Flu
x (
AU
)
Naï
ve
Therm
al
CAPS
UVB
0
200
400
600
800H
eat P
ain
Thre
shold
(°C
)
Naï
ve
Therm
al
CAPS
UVB
36
38
40
42
44
46
48
Mechanic
al P
ain
Thre
shold
(g)
Naï
ve
Therm
al
CAPS
UVB
60
80
100
120
140
160
180
200
**
*
* *
*
* *#
#
A
B
C
Fig. 3. Changes in cutaneous blood flow, sensitivity to thermal and mechanicalstimulation associated with primary lesion: comparison of UVB-induced inflam-mation and established inflammatory models. The biological activity of all threemodels (thermal, capsaicin and UVB inflammation, 3 MED) was confirmed by theirability to induce significant increases in cutaneous blood flow compared to naïveskin (*p < 0.05, one-way ANOVA followed by a Tukey’s post hoc test; n = 12 for allgroups) and shown in panel (A). (B) Shows the reductions in thermal painthresholds caused by the three models. Significant drops are seen followingcapsaicin and UVB inflammation but not thermal inflammation (*p < 0.05, one-wayANOVA followed by a Tukey’s post hoc test; n = 12 for all groups). Significantreductions in mechanical pain thresholds are seen with all three models comparedto naïve skin (*p < 0.05, one-way ANOVA followed by a Tukey’s post hoc test; n = 12for all groups) and are shown in panel (C). The mechanical pain thresholdsassociated with capsaicin and UVB inflammation are significantly lower than themechanical pain thresholds associated with thermal inflammation (#p < 0.05, one-way ANOVA followed by a Tukey’s post hoc test; n = 12 for all groups).
Therm
al
CAPS
UVB
0
5
10
15
20
25
30
Are
a (
cm
)
2
Therm
al
CAPS
UVB
0
5
10
15
20
25
30
Are
a (
cm
)
2
Therm
al
CAPS
UVB
0
10
20
30
40
50
60
70
Are
a (
cm
)
2
*
*
*
*
*
*
A
B
C
Fig. 4. Emergence of secondary flare, brush-evoked allodynia and pin-prickhyperalgesia: comparison of UVB-induced inflammation (3 MED) and establishedinflammatory models UVB does not produce secondary flare, brush-evokedallodynia or pin-prick hyperalgesia. Panel (A) shows that significant areas ofsecondary flare are associated with thermal and capsaicin inflammation (*p < 0.05,one-way ANOVA followed by a Tukey’s post hoc test; n = 12 for all groups) but notUVB inflammation. Significant brush-evoked allodynia is produced by both thermaland capsaicin inflammation (*p < 0.05, one-way ANOVA followed by a Tukey’s posthoc test; n = 12 for all groups) but not UVB inflammation and shown in panel (B).Panel (C) shows that significant pin-prick hyperalgesia in response to a 10 g vonFrey hair associated with thermal and capsaicin inflammation (*p < 0.05, one-wayANOVA followed by a Tukey’s post hoc test; n = 12 for all groups) but not UVBinflammation.
T. Bishop et al. / European Journal of Pain 13 (2009) 524–532 529
the uninjured centre of the annulus following topical capsaicin(p < 0.05, one-way ANOVA followed by a Tukey’s post hoc test;n = 12 for all groups). Moreover, the pain thresholds are reducedby 21.9 ± 9.5% 1.5 mm from the outer edge of the annulus(26.5 mm from the centre) and by 22.1 ± 7.4% 10 mm from the out-er edge (35 mm from the centre). These secondary changes follow-
ing capsaicin were all statistically significant (p < 0.05, one-wayANOVA followed by a Tukey’s post hoc test; n = 12 for all groups).
4. Discussion
We confirm here that marked primary hypersensitivity consis-tently develops in UVB inflamed skin. In contrast to topical

0.0
15.0
23.5
26.5
35.0
55.0
65.0
Mechanic
al P
ain
Thre
shold
(g)
60
80
100
120
140
160
180
200
0.0
15.0
23.5
26.5
35.0
55.0
65.0
Distance From Centre of Annulus (mm)
Mechanic
al P
ain
Thre
shold
(g)
60
80
100
120
140
160
180
200
0.0
15.0
23.5
26.5
35.0
55.0
65.0
Distance From Centre of Annulus (mm)
Mechanic
al P
ain
Thre
shold
(g)
60
80
100
120
140
160
180
200
0.0
15.0
23.5
26.5
35.0
55.0
65.0
Distance From Centre of Annulus (mm)
* *
**
** *
Fig. 5. Spatial distribution of changes in mechanical pain threshold following establishment of annular UV-B and capsaicin inflammatory lesions. Changes in mechanical painthresholds produced by UVB inflammation are restricted to the primary lesion. Panel (A) shows a diagrammatic representation of the lesions and the points at whichmechanical pain thresholds were taken. Examples of UVB and capsaicin inflammation are shown in (B) and (C), respectively. Panels (D), (E) and (F) show the spatialdistribution of changes in mechanical pain threshold in naïve, UVB inflamed and capsaicin treated skin, respectively. The shaded area in these panels represents the primarylesion. Significant reductions in mechanical pain thresholds compared to naïve skin are restricted to the primary lesion in following UVB inflammation but extend intoadjacent un-injured secondary areas following capsaicin treatment (*p < 0.05, one-way ANOVA followed by a Tukey’s post hoc test; n = 12 for all groups).
530 T. Bishop et al. / European Journal of Pain 13 (2009) 524–532
capsaicin and thermal burn, UVB inflammation did not produceerythematous or sensory changes in secondary skin areas.
In agreement with previous work, we found that doses >1 MEDled to erythema and roughly parallel reductions in thermal andmechanical pain thresholds, developing over several hours, peak-ing at ca. 24 h, before resolving over the following days (Harrisonet al., 2004; Benrath et al., 2001; Bickel et al., 1998; Hoffmannand Schmelz, 1999; Gustorff et al., 2004a). The magnitude andduration of the changes were UVB dose-dependent. Most previousreports have similarly demonstrated monophasic changes in bloodflow and pain thresholds (Harrison et al., 2004; Bickel et al., 1998;Hoffmann and Schmelz, 1999; Gustorff et al., 2004a) though onereport of biphasic changes in blood flow exists, with peaks at 18and 36 h possibly reflecting differences in sampling intervals used(Benrath et al., 2001).
We compared sensory changes in UVB with two recognisedmodels of inflammatory pain, topical capsaicin application and athermal burn injury. The traditional models both cause significantpain during their induction, persisting for tens of minutes follow-ing capsaicin application. In contrast, UVB is not associated withspontaneous pain during induction or at any subsequent time.
All three models produced similar changes at the treatment sitesimilar to previous reports (Pedersen and Kehlet, 1998; Dirks et al.,2003; Norbury et al., 2007 [thermal burn]; Kilo et al., 1994;Mohammadian et al., 1998; Liu et al., 1998 [topical capsaicin];Harrison et al., 2004; Bickel et al., 1998; Hoffmann and Schmelz,1999; Gustorff et al., 2004a [UVR irradiation]). The relatively mod-est consequences of the thermal burn probably result from usingmodest temperature (45 �C). Higher temperatures induce greatersensory change but more adverse events including skin blistering.
The emergence of sensory changes in secondary areas of skinhas been an important and interesting component of existinginflammatory pain models. We sought to characterise the emer-gence of flare, brush-evoked allodynia and pin-prick hyperalgesia
in the three models. In line with previous work, UVB inflammationdid not produce flare (Harrison et al., 2004; Bickel et al., 1998;Hoffmann and Schmelz, 1999; Gustorff et al., 2004a), whereas top-ical capsaicin and thermal injury both did (Pedersen and Kehlet,1998; Dirks et al., 2003; Norbury et al., 2007 [thermal burn]; Kiloet al., 1994; Mohammadian et al., 1998; Liu et al., 1998 [topicalcapsaicin]).
Some literature reports secondary pin-prick hyperalgesia fol-lowing UVB inflammation, using a 150 g von Frey filaments (Sychaet al., 2003, 2005; Gustorff et al., 2004a,b). We were unable to re-peat these findings because most of volunteers found 150 g fila-ments uncomfortable or painful in normal skin. Our data showsthat the average pain threshold, measured in naïve skin with theelectronic von Frey, was 157.4 ± 9.9 g (partial distribution study).Though the previously published work may be semantically cor-rect in using near threshold stimuli it is not common practice.We chose a 10 g von Frey filament to examine secondary hypersen-sitivity as commonly used in clinical and experimental studies(Koltzenburg et al., 1992; Booth and Young, 2000; Bove, 2006). Sig-nificant and robust pin-prick hyperalgesia was produced by topicalcapsaicin and thermal injury but not UVB inflammation.
Brush-evoked allodynia was found in secondary areas after top-ical capsaicin and thermal injury, but not UVB irradiation, suggest-ing the presence of central sensitisation in traditional models butits absence with UVB (Pedersen and Kehlet, 1998; Dirks et al.,2003; Norbury et al., 2007 [thermal burn]; Kilo et al., 1994;Mohammadian et al., 1998; Liu et al., 1998 [topical capsaicin]).
No direct evidence for a given mechanism of hyperalgesia is re-vealed by the present studies. However, there are several plausiblemechanisms potentially contributing to the primary mechanicalhypersensitivity seen here after UVB inflammation. Koppert et al.(2004) examined the role of sensitised mechano-insensitive‘‘silent” nociceptors in the hyperalgesia and mechanically inducedaxon reflex following UVB irradiation. Under normal conditions,

T. Bishop et al. / European Journal of Pain 13 (2009) 524–532 531
neurogenic flare cannot be elicited by phasic mechanical stimula-tion (Schmelz et al., 2000a; Koppert et al., 2004). However, thisfunction of mechano-insensitive nociceptors (Schmelz et al.,2000a) under conditions of UVB inflammation can be seen in sec-ondary skin following impact stimulation of the primary lesion sitesuggesting ‘‘silent” nociceptors are sensitised during UVB inflam-mation (Koppert et al., 2004). On the other hand, the absence ofspontaneous pain and secondary sensory changes around UVB in-flamed skin in the present study might suggests that an involve-ment of mechano-insensitive nociceptors is unlikely. This followsfrom the body of evidence that suggests spontaneous activity inmechano-insensitive C nociceptors is critical in the initiation andmaintenance of central sensitisation and subsequent secondaryallodynia and hyperalgesia (LaMotte et al., 1991; Schmelz et al.,2000b). One explanation would be that UVB leads to peripheralsensitisation but not spontaneous activity in mechano-insensitiveor other nociceptors. In UVB irradiated skin, mechanical stimula-tion leads to axon flares with short latencies (ca. 10 s), suggestingthat mechanically evoked mast cell degranulation is not directlyinvolved (Rukwied and Heyer, 1999; Koppert et al., 2004). Interest-ingly, these investigators noted that repeated mechanical stimula-tion produced increasing pain. The stimulation intervals (20 min)preclude ‘‘wind-up” as a mechanistic explanation. The authors sug-gest mechanical stimulation exaggerates local inflammatory medi-ator release, potentially from mast cells, which thus augmentsperipheral nociceptor function.
Despite evidence for sensitised mechano-insensitive nocicep-tors (Koppert et al. 2004) in the primary hyperalgesia producedby UVB inflammation, these authors could not rule out a contribu-tion of sensitised mechano-sensitive nociceptors which have beenimplicated in studies of other experimental inflammations(Andrew and Greenspan, 1999; Xu et al., 2000).
Another potential mechanism contributing to the erythema andhyperalgesia may be the upregulation of neuropeptides in nocicep-tors. In both rat and human skin, UVB inflammation increases neu-ropeptides (Eschenfelder et al., 1995; Benrath et al., 1995, 2001).However, Koppert et al. (2004) argued that the contribution of neu-ropeptide up-regulation to axon flare in UVB inflammation whichthey investigated at 24 h post-irradiation is negligible as the re-ported increases in neuropeptides were seen at 30–60 h post-irra-diation (Benrath et al., 2001).
There is evidence that bradykinin, a potent algogen producedfollowing tissue injury (for review see McMahon et al., 2006),may mediate UVB hyperalgesia and pain (Eisenbarth et al., 2004).Bradykinin induced pain and erythema was facilitated in UVB in-flamed skin potentially through the up-regulation and de novoexpression of receptors (Perkins and Kelly, 1993; Eisenbarthet al., 2004). Increases in expression occur in other inflammatorymodels and are mediated by the action of cytokines (IL-1; Phagooet al., 1999, 2001) many of which are produced during UVB inflam-mation (Chung et al., 1996; Clydesdale et al., 2001).
The induction of B1 receptor expression during inflammationpromotes sensitisation of nociceptors to mechanical stimuli (Foxet al., 2003, 2005), whereas murine knockouts of B1 receptorsshow reduced inflammatory hyperalgesia (Ferreira et al., 2001). Itis known that BK receptor function is sensitised by inflammatorymediators particularly prostaglandins (Tonussi and Ferreira,1997; Poole et al., 1999), which are produced in UVB inflamed skin(Soter, 1990; Clydesdale et al., 2001), leading to the sensitisation ofcutaneous nociceptors (Liang et al., 2001; Petho et al., 2001).
Central sensitisation cannot be discounted completely as a con-tributory mechanism in UVB-induced hyperalgesia. It may be pos-sible that the large UVB burns used by groups reporting secondaryhyperalgesia induce some spontaneous activity of nociceptors suf-ficient enough to produce secondary hyperalgesia but insufficientto produce a flare response. Moreover, the fact that we find pri-
mary brush-evoked allodynia may suggest that there is a degreeof central sensitisation. The modest doses and area of UVB usedhere, however, do not appear to induce significant centralsensitisation.
Importantly, the type of primary hyperalgesia we report herefollowing UVB-inflammation may exist in other models (thermalburn or capsaicin models) though the clear delineation of primaryand secondary sensory changes, and thus attendant mechanisms, islikely to be masked by the more pronounced contribution of cen-tral sensitisation in traditional models. One might expect that thedegree of central sensitisation induced would depend on theamount of nociceptor activation in any model which in turn wouldbe reflected measures of spontaneous pain. Notably UVB, at least asused here, did not produce spontaneous pain. Strong stimulation ofthe site of UV irradiated skin can produce pain and this of coursecould trigger central sensitisation, akin to the well described rekin-dling of secondary changes with capsaicin or thermal burns (Dirkset al., 2003).
This study contains the first comparison of UVB inflammationwith established inflammatory pain models. We demonstrate that,unlike established models, UVB inflammation predominantly pro-duces changes in sensory processing within the primary lesionwith minimal attendant secondary sensory changes (which are asignificant feature of other inflammatory models of pain). This sug-gests peripheral sensitisation is the predominant mechanistic scaf-fold to the UVB human pain model. In the future UVB inflammationmay provide a model to examine peripheral mechanisms andtreatments of pain.
Acknowledgements
We would like to thank the Guy’s and St. Thomas’ Charity (for-merly Special Trustees of Guy’s and St. Thomas’) for financial sup-port of this work.
References
Ali Z, Meyer RA, Campbell JN. Secondary hyperalgesia to mechanical but not heatstimuli following a capsaicin injection in hairy skin. Pain 1996;68:401–11.
Andrew D, Greenspan JD. Mechanical and heat sensitization of cutaneousnociceptors after peripheral inflammation in the rat. J Neurophysiol1999;82:2649–56.
Benrath J, Eschenfelder C, Zimmerman M, Gillardon F. Calcitonin gene-relatedpeptide, substance P and nitric oxide are involved in cutaneous inflammationfollowing ultraviolet irradiation. Eur J Pharmacol 1995;293:87–96.
Benrath J, Gillardon F, Zimmermann M. Differential time courses of skin blood flowand hyperalgesia in the human sunburn reaction following ultravioletirradiation of the skin. Eur J Pain 2001;5:155–67.
Bickel A, Dorfs S, Schmelz M, Forster C, Uhl W, Handwerker HO. Effects ofantihyperalgesic drugs on experimentally induced hyperalgesia in man. Pain1998;76:317–25.
Booth J, Young MJ. Differences in the performance of commercially available 10-gmonofilaments. Diabetes Care 2000;23:984–8.
Bove G. Mechanical sensory threshold testing using nylon monofilaments: the painfield’s ‘‘tin standard”. Pain 2006;124:13–7.
Charlton E. Ethical guidelines for pain research in humans. Committee on EthicalIssues of the International Association for the Study of Pain. Pain1995;63:277–8.
Chung JH, Youn SH, Koh WS, Eun HC, Cho KH, Park KC, et al. Ultraviolet Birradiation-enhanced interleukin (IL)-6 production and mRNA expression aremediated by IL-1 alpha in cultured human keratinocytes. J Invest Dermatol1996;106:715–20.
Clydesdale GJ, Dandie GW, Muller HK. Ultraviolet light induced injury:immunological and inflammatory effects. Immunol Cell Biol 2001;79:547–68.
Dirks J, Petersen KL, Dahl JB. The heat/capsaicin sensitization model: amethodologic study. J Pain 2003;4(3):122–8.
Eisenbarth H, Rukwied R, Petersen M, Schmelz M. Sensitization to bradykinin B1and B2 receptor activation in UV-B irradiated human skin. Pain2004;110:197–204.
Eschenfelder CC, Benrath J, Zimmermann M, Gillardon F. Involvement of substanceP in ultraviolet irradiation-induced inflammation in rat skin. Eur J Neurosci1995;7:1520–6.
Ferreira J, Campos MM, Pesquero JB, Araujo RC, Bader M, Calixto JB. Evidence for theparticipation of kinins in Freund’s adjuvant-induced inflammatory and

532 T. Bishop et al. / European Journal of Pain 13 (2009) 524–532
nociceptive responses in kinin B1 and B2 receptor knockout mice.Neuropharmacology 2001;41:1006–12.
Fox A, Wotherspoon G, McNair K, Hudson L, Patel S, Gentry C, et al. Regulation andfunction of spinal and peripheral neuronal B1 bradykinin receptors ininflammatory mechanical hyperalgesia. Pain 2003;104:683–91.
Fox A, Kaur S, Li B, Panesar M, Saha U, Davis C, et al. Antihyperalgesic activity of anovel nonpeptide bradykinin B1 receptor antagonist in transgenic miceexpressing the human B1 receptor. Br J Pharmacol 2005;144:889–99.
Fuchs PN, Campbell JN, Meyer RA. Secondary hyperalgesia persists in capsaicindesensitized skin. Pain 2000;84:141–9.
Gustorff B, Hoechtl K, Sycha T, Felouzis E, Lehr S, Kress HG. The effects ofremifentanil and gabapentin on hyperalgesia in a new extended inflammatoryskin pain model in healthy volunteers. Anesth Analg 2004a;98:401–7.
Gustorff B, Anzenhofer S, Sycha T, Lehr S, Kress HG. The sunburn pain model: thestability of primary and secondary hyperalgesia over 10 h in a crossover setting.Anesth Analg 2004b;98:173–7.
Hardy JD, Wolff HG, Goodell H. Experimental evidence on the nature of cutaneoushyperalgesia. J Clin Invest 1950;29:115–40.
Harrison GI, Young AR, McMahon SB. Ultraviolet radiation-induced inflammation asa model for cutaneous hyperalgesia. J Invest Dermatol 2004;122:183–9.
Hoffmann RT, Schmelz M. Time course of UVA- and UVB-induced inflammation andhyperalgesia in human skin. Eur J Pain 1999;3:131–9.
Kilo S, Schmelz M, Koltzenburg M, Handwerker HO. Different patterns ofhyperalgesia induced by experimental inflammation in human skin. Brain1994;117(Pt. 2):385–96.
Klede M, Handwerker HO, Schmelz M. Central origin of secondary mechanicalhyperalgesia. J Neurophysiol 2003;90:353–9.
Koltzenburg M, Lundberg LE, Torebjork HE. Dynamic and static components ofmechanical hyperalgesia in human hairy skin. Pain 1992;51:207–19.
Koltzenburg M, Torebjork HE, Wahren LK. Nociceptor modulated centralsensitization causes mechanical hyperalgesia in acute chemogenic andchronic neuropathic pain. Brain 1994;117(Pt. 3):579–91.
Koltzenburg M. Neural mechanisms of cutaneous nociceptive pain. Clin J Pain2000;16:S131–8.
Koppert W, Brueckl V, Weidner C, Schmelz M. Mechanically induced axon reflex andhyperalgesia in human UV-B burn are reduced by systemic lidocaine. Eur J Pain2004;8:237–44.
LaMotte RH, Shain CN, Simone DA, Tsai EF. Neurogenic hyperalgesia: psychophysicalstudies of underlying mechanisms. J Neurophysiol 1991;66:190–211.
Lewis T. The blood vessels of the human skin and their responses. London: Shaw &Sons; 1927.
Lewis T. Experiments relating to cutaneous hyperalgesia and its spread throughsomatic nerves. Clin Sci (Lond) 1935;2:373.
Liang YF, Haake B, Reeh PW. Sustained sensitization and recruitment of ratcutaneous nociceptors by bradykinin and a novel theory of its excitatory action.J Physiol 2001;532:229–39.
Liu M, Max MB, Robinovitz E, Gracely RH, Bennett GJ. The human capsaicin model ofallodynia and hyperalgesia: sources of variability and methods for reduction. JPain Symptom Manage 1998;16:10–20.
Magerl W, Fuchs PN, Meyer RA, Treede RD. Roles of capsaicin-insensitive nociceptorsin cutaneous pain and secondary hyperalgesia. Brain 2001;124:1754–64.
McMahon SB, Bennett DL, Bevan S. Inflammatory mediators and modulators of pain.In: McMahon SB, Koltzenburg M, editors. Wall and Melzack’s textbook ofpain. Churchill/Livingstone: Elsevier; 2006. p. 49–72.
Meyer RA, Ringkamp M, Campbell JN, Raja SN. Peripheral mechanisms of cutaneousnociception. In: McMahon SB, Koltzenburg M, editors. Wall and Melzack’stextbook of pain. Churchill/Livingstone: Elsevier; 2006. p. 3–34.
Mohammadian P, Andersen OK, Arendt-Nielsen L. Correlation between localvascular and sensory changes following tissue inflammation induced byrepetitive application of topical capsaicin. Brain Res 1998;792:1–9.
Norbury TA, MacGregor AJ, Urwin J, Spector TD, McMahon SB. Heritability ofresponses to painful stimuli in women: a classical twin study. Brain2007;130:3041–9.
Pedersen JL, Kehlet H. Hyperalgesia in a human model of acute inflammatory pain: amethodological study. Pain 1998;74:139–51.
Perkins MN, Kelly D. Induction of bradykinin B1 receptors in vivo in a model ofultra-violet irradiation-induced thermal hyperalgesia in the rat. Br J Pharmacol1993;110:1441–4.
Petho G, Derow A, Reeh PW. Bradykinin-induced nociceptor sensitization to heat ismediated by cyclooxygenase products in isolated rat skin. Eur J Neurosci2001;14:210–8.
Phagoo SB, Poole S, Leeb-Lundberg LM. Autoregulation of bradykinin receptors:agonists in the presence of interleukin-1beta shift the repertoire of receptorsubtypes from B2 to B1 in human lung fibroblasts. Mol Pharmacol1999;56:325–33.
Phagoo SB, Reddi K, Anderson KD, Leeb-Lundberg LM, Warburton D. Bradykinin B1receptor up-regulation by interleukin-1beta and B1 agonist occurs throughindependent and synergistic intracellular signaling mechanisms in human lungfibroblasts. J Pharmacol Exp Ther 2001;298:77–85.
Poole S, Lorenzetti BB, Cunha JM, Cunha FQ, Ferreira SH. Bradykinin B1 and B2receptors, tumour necrosis factor alpha and inflammatory hyperalgesia. Br JPharmacol 1999;126:649–56.
Raja SN, Campbell JN, Meyer RA. Evidence for different mechanisms of primary andsecondary hyperalgesia following heat injury to the glabrous skin. Brain1984;107(Pt. 4):1179–88.
Rukwied R, Heyer G. Administration of acetylcholine and vasoactive intestinalpolypeptide to atopic eczema patients. Exp Dermatol 1999;8:39–45.
Schmelz M, Michael K, Weidner C, Schmidt R, Torebjork HE, Handwerker HO. Whichnerve fibers mediate the axon reflex flare in human skin? Neuroreport2000a;11:645–8.
Schmelz M, Schmid R, Handwerker HO, Torebjork HE. Encoding of burning painfrom capsaicin-treated human skin in two categories of unmyelinated nervefibres. Brain 2000b;123(Pt. 3):560–71.
Soter NA. Acute effects of ultraviolet radiation on the skin. Semin Dermatol1990;9:11–5.
Staahl C, Drewes AM. Experimental human pain models: a review of standardisedmethods for preclinical testing of analgesics. Basic Clin Pharmacol Toxicol2004;95:97–111.
Sycha T, Gustorff B, Lehr S, Tanew A, Eichler HG, Schmetterer L. A simple pain modelfor the evaluation of analgesic effects of NSAIDs in healthy subjects. Br J ClinPharmacol 2003;56:165–72.
Sycha T, Anzenhofer S, Lehr S, Schmetterer L, Chizh B, Eichler HG, et al. Rofecoxibattenuates both primary and secondary inflammatory hyperalgesia: arandomized, double blinded, placebo controlled crossover trial in the UV-Bpain model. Pain 2005;113:316–22.
Tonussi CR, Ferreira SH. Bradykinin-induced knee joint incapacitation involvesbradykinin B2 receptor mediated hyperalgesia and bradykinin B1 receptor-mediated nociception. Eur J Pharmacol 1997;326:61–5.
Torebjork HE, Lundberg LE, LaMotte RH. Central changes in processing ofmechanoreceptive input in capsaicin-induced secondary hyperalgesia inhumans. J Physiol 1992;448:765–80.
Woolf CJ, Salter MW. Plasticity and pain: role of the dorsal horn. In: McMahon SB,Koltzenburg M, editors. Wall and Melzack’s textbook of pain. Churchill/Livingstone: Elsevier; 2006. p. 91–106.
Xu GY, Huang LY, Zhao ZQ. Activation of silent mechanoreceptive cat C and Adeltasensory neurons and their substance P expression following peripheralinflammation. J Physiol 2000;528(Pt. 2):339–48.
Ziegler EA, Magerl W, Meyer RA, Treede RD. Secondary hyperalgesia to punctatemechanical stimuli. Central sensitization to A-fibre nociceptor input. Brain1999;122(Pt. 12):2245–57.