ultrastructure and cell wall composition in cell division cycle mutants ofschizosaccharomyces pombe...
TRANSCRIPT

Antonie van Leeuwenhoek 59: 155-165, 1991. �9 1991 Kluwer Academic Publishers. Printed in the Netherlands.
Ultrastructure and cell wall composition in cell division cycle mutants of Schizosaccharomyces pombe deficient in septum formation
Pedro Mateos & Angel Domfnguez* Departamento de Microbiologia y Genrtica. Universidad de Salamanca, E-37071 Salamanca, Spain (* requests for offprints)
Received 26 June 1990; accepted in revised form 3 December 1990
Key words: cell division cycle, Schizosaccharomyces pombe, cell wall composition, septum formation
Abstract
A number of temperature-sensitive cdc- mutants of Schizosaccharomycespombe that are affected in septum formation were analyzed with respect to their ultrastructure and the composition of their cell wall polymers. One mutant strain, cdc 16-116, has a cell wall composition similar to the wild type (strain 972 h-). However two other mutants, cdc 4 and cdc 7, show a higher galactomannan content and a lower et-glucan content. In all the mutants tested, total glucose incorporation, protein, RNA and DNA synthesis increased similarly to wild type over 31/2 h. After 2-3 h of incubation at the non permissive temperature- 35 ~ C- , cell numbers remained constant although, increases in optical densities at 600 nm were observed. According to scanning electron microscopy, the mutants had aberrant shapes after 5h of incubation at 35 ~ C, Transmission electron microscopy showed that cdc 3 is unable to complete septum formation, cdc 4 showed the most varied morphological shapes and aberrant depositions of cell wall material, cdc 8 exhibited a deranged plasma membrane and cell wall regions near of cell poles; an abnormal septum and several nuclei, cdc 7 showed elongated cells with several nuclei and with an apparently normal cell wall completely lacking in septum and septal material, cdc 16 showed more than one septum per cell.
Introduction
Temperature-sensitive mutations blocking specific steps in the cell division cycle of Saccharomyces cerevisiae (Hartwell et al. 1970) and Schizosaccha- romyces pombe (Nurse et al. 1976) have been iso- lated. The use of these mutants has greatly in- creased our knowledge of the cell division cycle in eukaryotic cells (for recent reviews see Pardee 1989; Murray & Kirschner 1989; Hartwell & Wein- ert 1989). However, most of the recent work has been directed towards genes that play a key role in the control of the eukaryotic cell cycle (Lee & Nurse 1988; Wittenberg & Reed 1988). Other as- pects, such as cytokinesis and cell separation and
the genes that control these processes are less well known (Pringle & Hartwell 1981; Wheals 1987). With respect to yeast cell wall composition, most work has been carried out on S. cerevisiae (Cabib et al. 1982; Slater 1981; Hernandez et al. 1989) al- though some excellent papers on the structure of glucans and cell wall growth ofS. pombe have been published (Manners & Meyer 1977; Streiblovh 1981).
It has been proposed that CDC 24 controls the location of chitin on S. cerevisiae (Sloat & Pringle 1978), although other authors suggest that the acti- vation of chitin synthetase zymogen may be a non specific response to cell-cycle arrest and in fact several cdc mutants seem to produce this kind of

156
effect (Roberts et al. 1983). A filamentous ring which appears during bud enlargement at the neck between mother and daughter cells seems crucial for cytokinesis in S. cerevisiae because the cdc mutants 3, 11 and 12, defective in cytokinesis, lack this ring (Byers & Goetsch 1976). A similar sugges- tion has been advanced for several cdc mutants of S. pombe defective in septum formation (Streiblov~ 1981). The cdc mutants of S. cerevisiae defective in cytokinesis have shown no obvious defects in cell wall structure (Dominguez et al. 1982), and neither have defects in cell wall polymers been described for the cdc mutants of S. pombe with abnormal septa (Nurse et al. 1976; Streiblov~ 1981). We re- port here that the cell wall in S. pombe cdc 4 and cdc 7 shows a relatively higher content in galacto- mannan and a lower content in ct-glucan, indicating that the latter polymer may be the most important component in septum formation. By transmission electron microscopy cdc 8 exhibits defects in the cell wall and/or plasma membrane near to the poles, which appear swollen.
Materials and methods
Organisms and growth conditions Schizosaccharomyces pombe 972 h-, cdc 3-6, cdc 4-8, cdc 7-24, cdc 8-27 and cdc 16-116 were kindly supplied by Dr. P. Nurse Department of Biochem- istry, University of Oxford, U.K. Growth media and conditions for temperature shifts were those used by Nurse et al. (1976). Cell numbers were determined as described in Mitchinson (1970). Dry weights were determined as described previously (Rodriguez & Dominguez 1984).
Analytical procedures DNA was determined by the diphenylamine reac- tion (Bostock 1970). Total glucose incorporation, protein and RNA were determined as described previously (Dominguez et al. 1978).
Labelling and analysis of cell wall components The amounts of galactomannan, a-glucan and 13- glucan synthesized were determined by the method
of Manners & Meyer (1977) after addition of (U- 14C) glucose to the growth medium.
Electron microscopy Electron microscopy was performed exactly in the same way described by Rodriguez & Dominguez (1984).
Chemicals (W-14C) uracil (sp act 62 mCi/mmol), L-(U-14C) leucine (sp act 348 mCi/mmol) and D-(U-14C)glu - cose (sp act 224 mCi/mmol) were obtained from the Radiochemical Centre Amersham, Bucks, UK.
Results
Macromolecular synthesis in cdc mutants
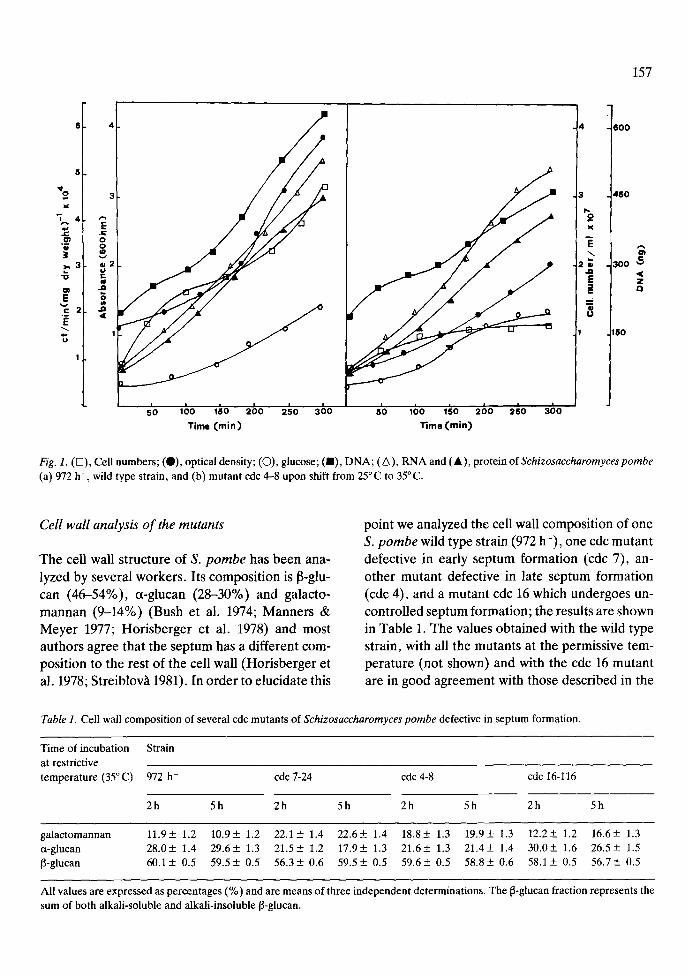
Exponentially growing cultures in liquid minimal medium at 25 ~ C of the wild type strain 972 h- and of the cdc 3, 4, 7, 8, 16 mutants were shifted to 35 ~ C for 5 h. At the times indicated samples were taken and cell numbers, optical density, total glucose incorporation, protein, RNA and DNA synthesis were measured. The results of the wild type strain, 972 h-, and one mutant, cdc 4 are shown in Fig. 1. In all the mutants tested an increase in glucose incorporation, DNA, RNA and protein synthesis was observed. The results in mutants cdc 3, cdc 7 (not shown, Nurse et al. 1976) and cdc 4 (Fig. lb) were similar to those obtained with the wild type up to 31/2 h. A slow decrease (between 15-30%) in glucose incorporation, RNA and protein synthesis was found at 5 h of incubation in the cdc 4 mutant (Fig. lb). Mutant cdc 16 halted DNA synthesis after 31/2h of incubation at 35~ (not shown), in agreement with the results described by Minet et al. (1979). Cell numbers remained constant after one hour of incubation at the restrictive temper- ature in cdc 3, and after three hours in the case of cdc 4 (Fig. lb). Thus, macromolecular synthesis is probably due to cell elongation, as could be dem- onstrated by electron microscopy (see later) and by the increase in optical density at 600 nm (Fig. lb).
157
5
1<
"7 4
t.-
> , 3 " 0
E iP
6 .
50 100 150 200 250 300 50 100 150 200 250 300
Time ( ra in ) Time ( m i n )
|
I
600
450
300 " "
Z a
150
Fig. 1. (D), Cell numbers; (0 ) , optical density; (O), glucose; (11), DNA; (A), RNA and ( � 9 protein of Schizosaccharomyces pombe (a) 972 h- , wild type strain, and (b) mutant cdc 4-8 upon shift from 25~ to 35 ~ C.
Cell wall analysis of the mutants
The cell wall structure of S. pombe has been ana- lyzed by several workers. Its composition is 13-glu- can (46-54%), ~t-glucan (28-30%) and galacto- mannan (9-14%) (Bush et al. 1974; Manners & Meyer 1977; Horisberger et al. 1978) and most authors agree that the septum has a different com- position to the rest of the cell wall (Horisberger et al. 1978; Streiblovh 1981). In order to elucidate this
point we analyzed the cell wall composition of one S. pombe wild type strain (972 h-), one cdc mutant defective in early septum formation (cdc 7), an- other mutant defective in late septum formation (cdc 4), and a mutant cdc 16 which undergoes un- controlled septum formation; the results are shown in Table 1. The values obtained with the wild type strain, with all the mutants at the permissive tem- perature (not shown) and with the cdc 16 mutant are in good agreement with those described in the
Table 1. Cell wall composition of several cdc mutants of Schizosaccharornyces pombe defective in septum formation.
Time of incubation Strain at restrictive temperature (35~ 972 h- cdc 7-24 cdc 4-8 cdc 16-116
2h 5h 2h 5h 2h 5h 2h 5h
galactomannan 11.9+ 1.2 10.9+ 1.2 22.1+ 1.4 22.6_+ 1.4 18.8+ 1.3 19.9+ 1.3 12.2+ 1.2 16.6_+ 1.3 ct-glucan 28.0_+ 1.4 29.6_+ 1.3 21.5 + 1.2 17.9 4- 1.3 21.6_+ 1.3 21.4 + 1.4 30.0 4- 1.6 26.5-+ 1.5 13-glucan 60.1___ 0.5 59.5+ 0.5 56.3-+ 0.6 59.5_+ 0.5 59.6+ 0.5 58.8_+ 0.6 58.1+ 0.5 56.7+ 0.5
All values are expressed as percentages (%) and are means of three independent determinations. The 13-glucan fraction represents the sum of both alkali-soluble and alkali-insoluble 13-glucan.
158
literature (Bush et al. 1974; Manners & Meyer 1977), although in our hands the amount of 6- glucan was 2--4% higher. By contrast, the other two mutants, cdc 7 and cdc 4, although with a normal content in f~-glucan, showed significant var- iations in the amounts of galactomannan (which increased by almost 100%) and ct-glucan (which decreased by 30%). Both effects, that are pro- duced in similar amounts of cell wall material, sug- gest that the septum has a higher a-glucan content than the rest of the cell wall.
Cell morphology
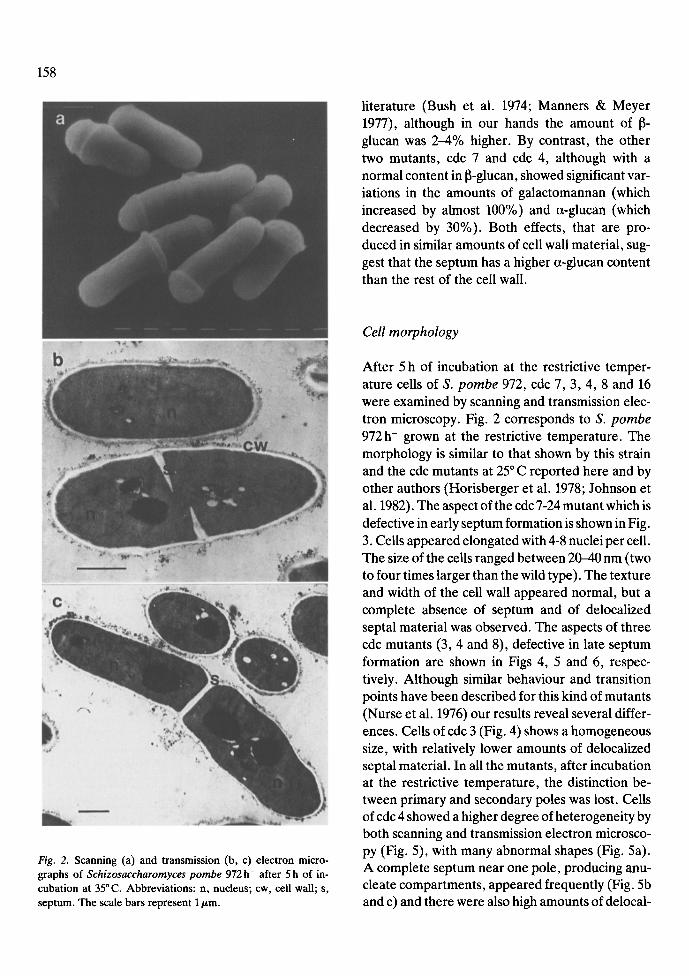
Fig. 2. Scanning (a) and transmission (b, c) electron micro- graphs of Schizosaccharomyces pombe 972 h- after 5 h of in- cubation at 35~ Abbreviations: n, nucleus; cw, cell wall; s, septum. The scale bars represent 1/~m.
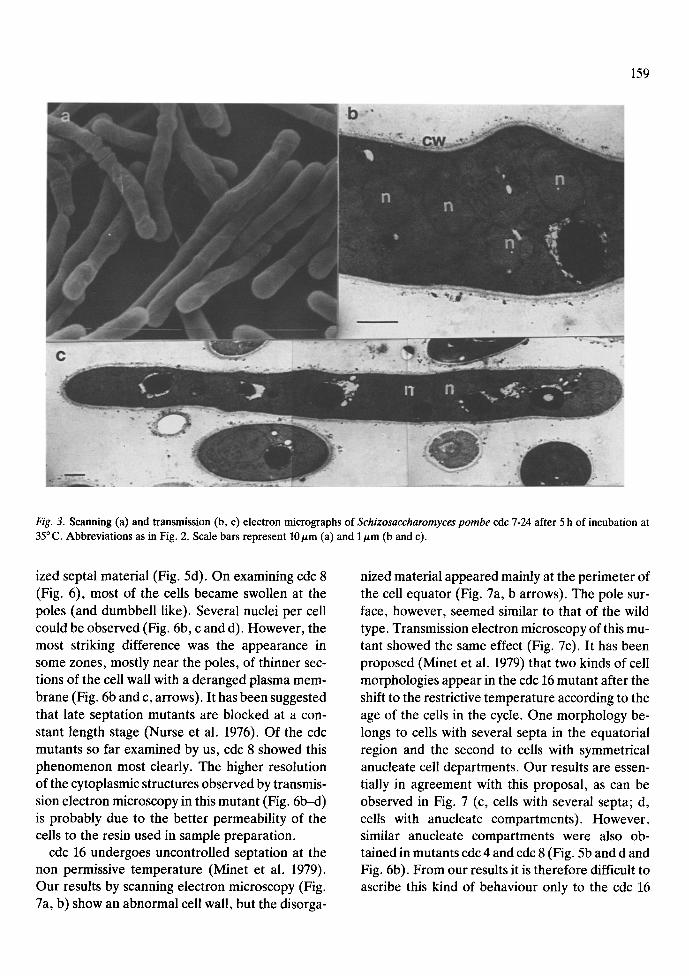
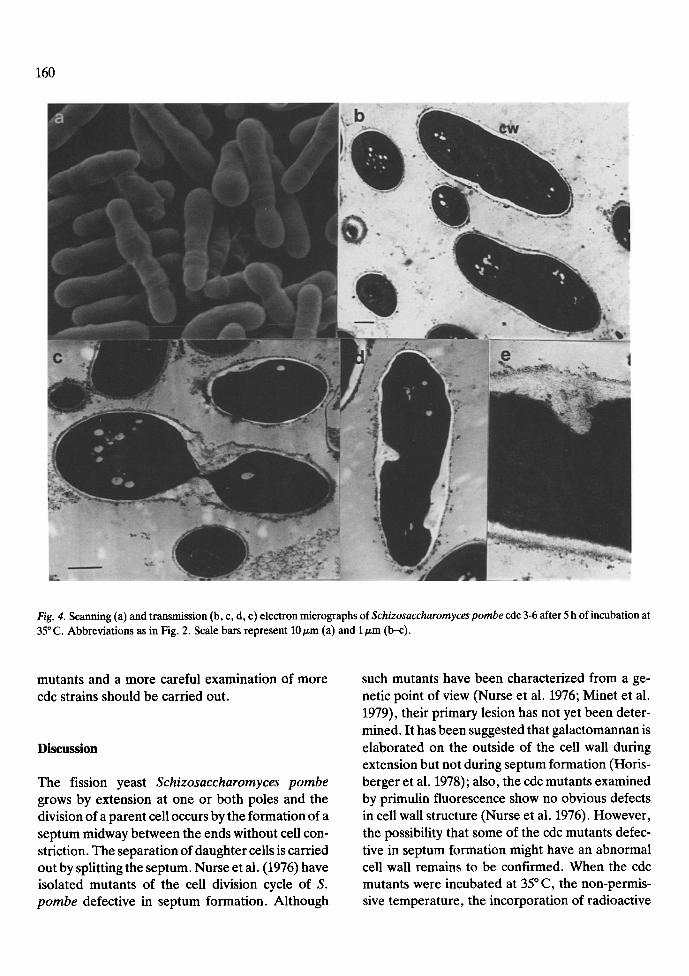
After 5 h of incubation at the restrictive temper- ature cells of S. pombe 972, cdc 7, 3, 4, 8 and 16 were examined by scanning and transmission elec- tron microscopy. Fig. 2 corresponds to S. pombe 972 h- grown at the restrictive temperature. The morphology is similar to that shown by this strain and the cdc mutants at 25 ~ C reported here and by other authors (Horisberger et al. 1978; Johnson et al. 1982). The aspect of the cdc 7-24 mutant which is defective in early septum formation is shown in Fig. 3. Cells appeared elongated with 4-8 nuclei per cell. The size of the cells ranged between 20-40 nm (two to four times larger than the wild type). The texture and width of the cell wall appeared normal, but a complete absence of septum and of delocalized septal material was observed. The aspects of three cdc mutants (3, 4 and 8), defective in late septum formation are shown in Figs 4, 5 and 6, respec- tively. Although similar behaviour and transition points have been described for this kind of mutants (Nurse et al. 1976) our results reveal several differ- ences. Cells of cdc 3 (Fig. 4) shows a homogeneous size, with relatively lower amounts of delocalized septal material. In all the mutants, after incubation at the restrictive temperature, the distinction be- tween primary and secondary poles was lost. Cells of cdc 4 showed a higher degree of heterogeneity by both scanning and transmission electron microsco- py (Fig. 5), with many abnormal shapes (Fig. 5a). A complete septum near one pole, producing anu- cleate compartments, appeared frequently (Fig. 5b and c) and there were also high amounts of delocal-
159
Fig. 3. Scanning (a) and transmission (b, c) electron micrographs of Schizosaccharomyces pombe cdc 7-24 after 5 h of incubation at 35 ~ C. Abbreviations as in Fig. 2. Scale bars represent 10/~m (a) and 1/zm (b and c).
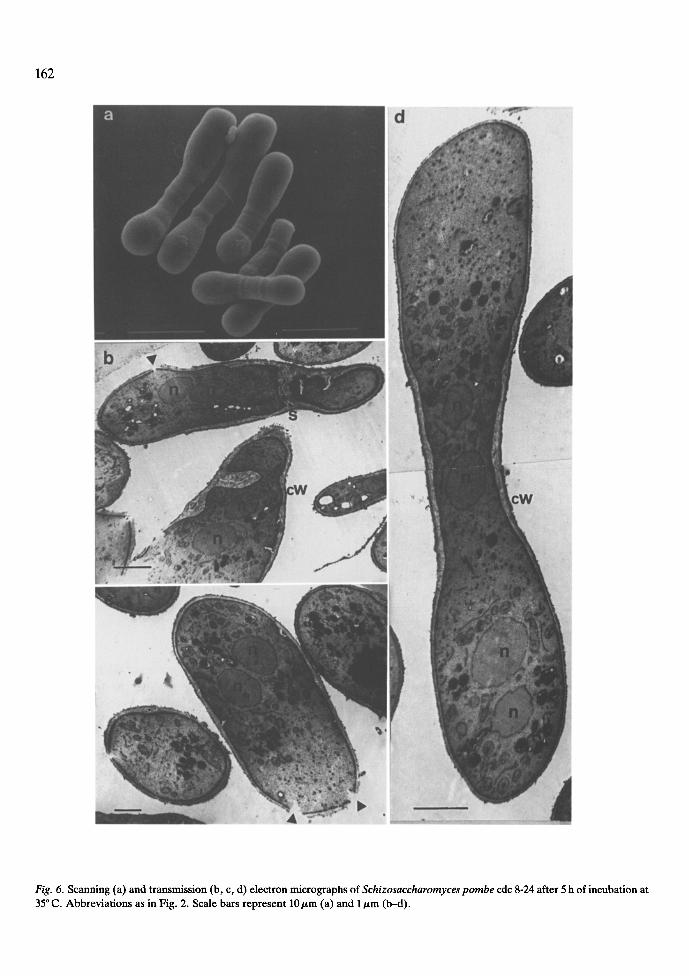
ized septal material (Fig. 5d). On examining cdc 8 (Fig. 6), most of the cells became swollen at the poles (and dumbbell like). Several nuclei per cell could be observed (Fig. 6b, c and d). However, the most striking difference was the appearance in some zones, mostly near the poles, of thinner sec- tions of the cell wall with a deranged plasma mem- brane (Fig. 6b and c, arrows). It has been suggested that late septation mutants are blocked at a con- stant length stage (Nurse et al. 1976). Of the cdc mutants so far examined by us, cdc 8 showed this phenomenon most clearly. The higher resolution of the cytoplasmic structures observed by transmis- sion electron microscopy in this mutant (Fig. 6b--d) is probably due to the better permeability of the cells to the resin used in sample preparation.
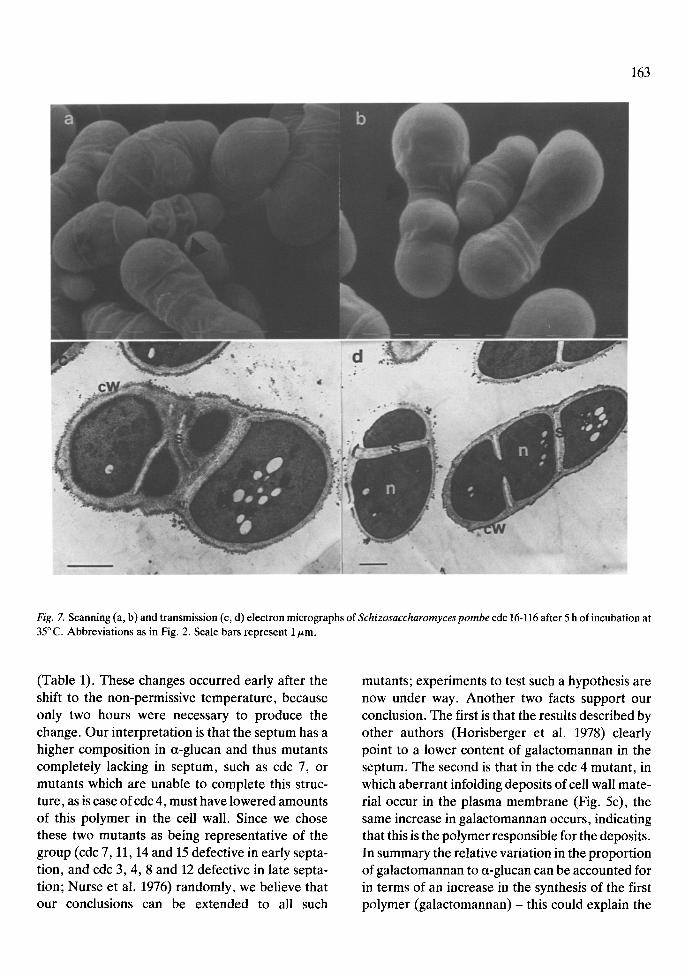
cdc 16 undergoes uncontrolled septation at the non permissive temperature (Minet et al. 1979). Our results by scanning electron microscopy (Fig. 7a, b) show an abnormal cell wall, but the disorga-
nized material appeared mainly at the perimeter of the cell equator (Fig. 7a, b arrows). The pole sur- face, however, seemed similar to that of the wild type. Transmission electron microscopy of this mu- tant showed the same effect (Fig. 7c). It has been proposed (Minet et al. 1979) that two kinds of cell morphologies appear in the cdc 16 mutant after the shift to the restrictive temperature according to the age of the cells in the cycle. One morphology be- longs to cells with several septa in the equatorial region and the second to cells with symmetrical anucleate cell departments. Our results are essen- tially in agreement with this proposal, as can be observed in Fig. 7 (c, cells with several septa; d, cells with anucleate compartments). However, similar anucleate compartments were also ob- tained in mutants cdc 4 and cdc 8 (Fig. 5b and d and Fig. 6b). From our results it is therefore difficult to ascribe this kind of behaviour only to the cdc 16
160
Fig. 4. Scanning (a) and transmission (b, c, d, e) electron micrographs of Schizosaccharomyces pombe cdc 3-6 after 5 h of incubation at 35 ~ C. Abbreviations as in Fig. 2. Scale bars represent 10/zm (a) and 1 #m (b--c).
mutants and a more careful examination of more cdc strains should be carried out.
Discussion
The fission yeast Schizosaccharomyces pombe grows by extension at one or both poles and the division of a parent cell occurs by the formation of a septum midway between the ends without cell con- striction. The separation of daughter cells is carried out by splitting the septum. Nurse et al. (1976) have isolated mutants of the cell division cycle of S. pombe defective in septum formation. Although
such mutants have been characterized from a ge- netic point of view (Nurse et al. 1976; Minet et al. 1979), their primary lesion has not yet been deter- mined. It has been suggested that galactomannan is elaborated on the outside of the cell wall during extension but not during septum formation (Horis- berger et al. 1978); also, the cdc mutants examined by primulin fluorescence show no obvious defects in cell wall structure (Nurse et al. 1976). However, the possibility that some of the cdc mutants defec- tive in septum formation might have an abnormal cell wall remains to be confirmed. When the cdc mutants were incubated at 35 ~ C, the non-permis- sive temperature, the incorporation of radioactive

161
Fig. 5. Scanning (a) and transmission (b, c, d) electron micrographs of Schizosaccharomyces pombe cdc 4-8 after 5 h of incubation at 35~ Abbreviations as in Fig. 2. Scale bars represent 10/,~m (a) and 1/zm (b--d).
precursors into macromolecules was similar to what was observed in the wild type over at least 31/2 h (Fig. 3, Nurse et al. 1976; Minet et al. 1979). However our cell wall fractionation revealed signif- icant differences in cell wall composition between the wild type strain and the cdc mutants defective in septum formation (Table 1). The values obtained with the 972h- strain and with the multiseptate cdc mutant 16 are in good agreement with those de- scribed in the literature (Bush et al. 1974; Manners & Meyer 1977), although in our experiments we observed a small increase (between 2--4%) in the amount of [3-glucan with respect to the values de- scribed by other authors. Whether this is real or
just an artefact of our sample preparation remains to be elucidated. However, since the discrepancy is very low and the results could be reproduced in all samples, we believe that the following conclusions are in no way compromised.
A different picture emerged when we analyzed the cell wall composition of the mutants defective in septum formation. Regardless of whether the mutants were defective in early septation (cdc 7) or late septation (cdc 4) significant and compensatory changes in the cell wall polymers occurred. While the amount of fl-glucan remained constant a de- crease in the amount of a-glucan and an increase in the amount of galactomannan were observed
162
Fig. 6. Scanning (a) and transmission (b, c, d) electron micrographs of Schizosaccharomyces pombe cdc 8-24 after 5 h of incubation at 35 ~ C. Abbreviations as in Fig. 2. Scale bars represent 10 btm (a) and 1 ~m (b-d).
163
Fig. 7. Scanning (a, b) and transmission (c, d) electron micrographs of Schizosaccharomyces pombe cdc 16-116 after 5 h of incubation at 35 ~ C. Abbreviations as in Fig. 2. Scale bars represent 1/zm.
(Table 1). These changes occurred early after the shift to the non-permissive temperature, because only two hours were necessary to produce the change. Our interpretation is that the septum has a higher composition in ~t-glucan and thus mutants completely lacking in septum, such as cdc 7, or mutants which are unable to complete this struc- ture, as is case ofcdc 4, must have lowered amounts of this polymer in the cell wall. Since we chose these two mutants as being representative of the group (cdc 7, 11, 14 and 15 defective in early septa- tion, and cdc 3, 4, 8 and 12 defective in late septa- tion; Nurse et al. 1976) randomly, we believe that our conclusions can be extended to all such
mutants; experiments to test such a hypothesis are now under way. Another two facts support our conclusion. The first is that the results described by other authors (Horisberger et al. 1978) clearly point to a lower content of galactomannan in the septum. The second is that in the cdc 4 mutant, in which aberrant infolding deposits of cell wall mate- rial occur in the plasma membrane (Fig. 5c), the same increase in galactomannan occurs, indicating that this is the polymer responsible for the deposits. In summary the relative variation in the proportion of galactomannan to ~-glucan can be accounted for in terms of an increase in the synthesis of the first polymer (galactomannan) - this could explain the

164
deposition of wall material in the cdc 4 mutant - or because the synthesis of a-glucan is reduced. This latter assumption justifies why in the case of cdc 7 there is no increase in the content of cell wall mate- rial (since there is no septum, the proportion of a-glucan would decrease). This could also explain the result obtained with the cdc 4 mutant. Results supporting such a notion have been obtained with mutants in which a decrease can be observed in the galactomannan content. These mutants are oval in shape and in them the proportion of septum mate- rial compared with the material in the rest of the wall is greater (Ribas, 1989). However it is not clear why the multiseptate mutant cdc 16 does not have a higher alpha-glucan content compared to the wild type strain. Our results at the moment can only be explained by assuming that the total pro- portion of the cell wall polymers of multiseptate cells remains constant and experiments to elucidate this point are currently being carried out.
The cytological images of the terminal pheno- types of the mutants are complex (Figs 3, 4, 5, 6 and 7). Cultures of cdc 7 (Fig. 4) show a uniform aspect with an apparently normal cell wall and without any kind of abnormal deposits. Irregular forms or delocalized wall material are common features in cdc 3, 4 and 8. However, while cdc 3 samples are almost completely lacking in septum, some asym- metric septa producing anucleate compartments were observed in cdc 4 (Fig. 5b and d) and cdc 8 (Fig. 6b). Although swollen poles are formed in all the mutants, this effect is more clearly appreciated in cdc 8 (Fig. 6a). Also, in this mutant deranged plasma membrane and cell walls were observed (Fig. 6b and c). This fact could be explained by a fight against the constant volume stage (Mitchison 1970) or, more likely, by an unbalanced synthesis of the cell wall polymers, probably at the sites at which new septum is to be developed. Although defects in the assembly and/or function of the ring of cytoplasmic micro filaments have been implicat- ed in this mutant (Streiblov~ 1981), in our transmis- sion electron micrographs of intact untreated cells these structures are almost impossible to observe, due to the extremely hard cell wall. The behaviour of the multiseptate mutant cdc 16 (Fig. 7) agrees
with previously described results (Minet et al. 1979, Streiblov~ 1981).
The findings of the present study seem to be consistent with the suggestion that the plasma membrane would be stimulated to induce septation in the absence of nuclear division by mutants cdc 4, 8 and 16 (Streiblov~ 1981). We also suggest that there are two kind of mutants defective in late septation; those which are able to complete septum formation (although asymmetrically), such as cdc 4 and cdc 8, and others which, with larger amounts of wall material, are unable to close the septum, like cdc 3.
Acknowledgements
We wish to thank N. Skinner for revising the En- glish version of the manuscript.
References
Bostock CJ (1970) DNA synthesis in the fission yeast Schizo- saccharomyces pombe. Exp. Cell. Res. 60:16-26
Bush DA, Horisberger M, Herman I & Wussch P (1974) The wall structure of Schizosaccharomycespombe. J. Gen. Micro- biol. 81:199-206
Byers B & Goetsch L (1976) Loss of the filamentous ring in cyt0kinesis-defective mutants of budding yeast. Abstracts of the First International Congress on Cell Biology. J. Cell. Biol. 70(2): 35a
Cabib E, Roberts R & Bowers B (1982) Synthesis of the yeast cell wand and its regulation. Ann. Rev. Biochem. 51:763-793
Dominguez A, Varona RM, Viilanueva JR & Sentandreu R (1982) Mutants of Saccharomyces cerevisiae cell division cycle defective in cytokinesis. Biosynthesis of '.the cell wall and morphology. Antonie van Leeuwenhoek 48:145-157
Dominguez A, Villanueva JR & Sentandreu R (1978) Inositol deficiency in Saccharomyces cerevisiae NCYC 86. Antonie van Leeuwenhoek 44:25-34
Hartwell LH, Culotti J & Reid B (1970) Genetic control of the cell-division cycle in yeast, I. Detection of mutants. Proc. Natl. Acad. Sci. USA 66:352-359
Hartwell LH & Weinert TA (1989) Checkpoints: controls that ensure the order of cell cycle events. Science 246:629-634
Hernandez LM, BaUou L, Alvarado E, Tsai P-K & Ballou CE (1989) Structure of the phosphorylated N-linked oligosaccha- rides from the mnn9 and mnnl0 mutants of Saccharomyces cerevisiae. J. Biol. Chem. 264:13648-13659
Horisberger M, Vonlanthen M & Rosset J (1978) Localization

of ct-galactomannan and of wheat germ agglutinin receptors in Schizosaccharomyces pombe. Arch. Microbiol. 119: 107- 111
Johnson BF, Calleja GB, Yoo BY, Zuker M & McDonald IJ (1982) Cell division: Key to cellular morphogenesis in the fission yeast, Schizosaccharornyces. Int. Rev. Cytol 75: 167- 207
Lee MG & Nurse P (1988) Cell cycle control genes in fission yeast and mammalian cells. Trends Genet. 4:287-290
Manners DJ & Meyer MT (1977) The molecular structures of some glucans from the cell walls of Schizosaccharomyces pombe. Carbohydr. Res. 57:189-203
Minet M, Nurse P, Thuriaux P & Mitchison JM (1979) Un- controlled septation in cell division cycle mutant of the fission yeast Schizosaccharomycespombe, J. Bacteriol. 137:440--446
Mitchinson JM (1970) Physiological and cytological methods for Schizosaccharomyces pombe. In: Prescott EM (Ed) Methods in Cell Physiol, Vol IV (pp 131-165). Academic Press New York
Murray AW &Kirschner MW (1989) Dominoes and clocks: the union of two views of the cell cycle. Science 246:614-621
Nurse P, Thuriaux P & Nasmyth K (1976) Genetic control of the cell-division cycle in the fission yeast Schizosaccharomyces pombe. Mol. Gen. Genet. 146:167-178
Pardee AB (1989) GI Events and regulation of cell prolifer- ation. Science 246:603-608
Pringle J & Hartwell LH (1981) The Saccharomyces cerevisiae
165
cell cycle. In: Strathern J, Jones E, Broach J (Eds) The Molecular Biology of the Yeast Saccharomyces (pp 97-142). Cold Spring Harbor Laboratory, New York
Ribas JC (1989) Estudios sobre la biosfntesis de113(1,3)glucano de la pared celular de Schizosaccharomyces pombe. Ph.D. thesis, University of Salamanca
Roberts RL, Bowers B, Slater ML & Cabib E (1983) Chitin synthesis and localization in cell division cycle mutants of Saccharomyces cerevisiae. Mol. Cell. Biol. 3:922-930
Rodriguez C & Dominguez A (1984) The growth characteristics of Saccharomycopsis lipolytica: morphology and induction of mycelium formation. Can. J. Microbiol. 30:605-612
Slater ML (1981) Molecular biology of budding. In: Arnold WN (Ed) Yeast Cell Envelopes: Biochemistry, Biophysics and Ultrastructure, Vol II (pp 65-77). CRC Press, Boca Raton, Florida
Sloat BP & Pringle JR (1978) A mutant of yeast defective in cellular morphogenesis. Science 200:1171-1173
Streiblovh E (1981) Fission. In: Arnold WN (Ed) Yeast Cell Envelopes: Biochemistry, Biophysics and Ultrastructure, Vol II (pp 79-92). CRC Press, Boca Raton, Florida
Wheals AE (1987) Biology of the cell cycle in yeasts. In: Rose AH, Harrison JS (Eds) The Yeasts, Vol II (pp 283-390). Academic Press, London
Wittenberg C & Reed SI (1988) Control of the yeast cell cycle is associated with assembly disassembly of the cdc 28 protein kinase complex. Cell 54:1061-1072