trigeminal central sensitization and its modulation in ... · trigeminal central sensitization and...
TRANSCRIPT

Trigeminal Central Sensitization and Its Modulation in Acute and Chronic Orofacial Pain Models
by
Pavel Cherkas
A thesis submitted in conformity with the requirements for the degree of Master of Science
Faculty of Dentistry University of Toronto
© Copyright by Pavel Cherkas 2014

ii
Trigeminal Central Sensitization and Its Modulation in Acute and Chronic Orofacial Pain Models
Pavel Cherkas
Master of Science
Faculty of Dentistry
University of Toronto
2014
Abstract This study aimed to examine whether trigeminal nerve injury induces chronic nociceptive
behaviour and central sensitization (CS) in functionally identified medullary dorsal horn (MDH)
nociceptive neurons in mice, and whether CS in acute and chronic orofacial pain models and
nociceptive behaviour in the chronic model are affected by systemic administration of
pregabalin. Infraorbital nerve injury induced chronic facial mechanical allodynia as well as
MDH CS; acute noxious tooth pulp stimulation also induced MDS CS. Systemic administration
of pregabalin attenuated the nerve injury-induced allodynia as well as the MDH CS in both the
chronic and acute pain models. These findings reveal that MDH CS occurs in mouse models of
acute and chronic orofacial pain and that pregabalin may prove useful clinically in acute and
chronic orofacial pain states.

iii
Acknowledgements
I would like to express my inmost gratitude to a number of individuals whose support has been
crucial for the accomplishment of this work.
I would especially like to thank Dr. Barry J. Sessle, my supervisor and mentor, who taught me
how to conduct scientific research and give it a concise written form.
I would also like to thank Dr. Shimon Friedman for his unstinting mentoring and help over my
academic and clinical years at the University of Toronto.
I would like to thank Dr. Limor Avivi-Arber for her kind assistance in preparing this work.
Furthermore, I would like to express my appreciation to my colleague and friend at the
University of Toronto, Dr. Vidya Varathan.
I wish to extend my thanks to my family for their continued understanding and support
throughout these years.
Finally, my words of gratitude go to Ilona, my patient and loving wife who walked this journey
with me.
This study was supported in part by grants from the American Association of Endodontists
Foundation and the Canadian Academy of Endodontics Endowment Fund.

iv
Table of Contents
Abstract ii
Acknowledgements iii
Table of Contents iv
Abbreviations vi
Chapter 1: General Introduction 1
1.1 Introduction 1
1.2 Pain 1
1.2.1 Multidisciplinarity and socioeconomical impact of pain 2
1.2.2 Acute vs. chronic pain 2
1.2.3 Classification of pain: nociceptive, inflammatory and neuropathic 3
1.2.4 Classification and prevalence of orofacial pain 3
1.2.5 Trigeminal neuropathic pain (atypical odontalgia and atypical facial pain) 4
1.3 Pain management 5
1.3.1 Non-pharmacological management of orofacial pain 5
1.3.2 Pharmacological management of orofacial pain 6
1.3.3 Anticonvulsant drug pregabalin 7
1.4 Mechanisms and models of orofacial pain 8
1.4.1 Peripheral processes of the trigeminal system 8
1.4.2 Central processes in the trigeminal system 10
1.4.3 Central sensitization 12

v
1.4.4 Models of orofacial pain 13
1.4.5 Modulation of orofacial pain mechanisms 16
1.5 Statement of the problem and rationale 16
Chapter 2: Project Specific Aims and Hypotheses 17
Chapter 3: Articles 18
3.1 Article 1 18
Pregabalin Blocks Central Sensitization in Medullary Dorsal Horn in a Rodent
Model of Acute Tooth Pulp Inflammatory Pain
3.2 Article 2 36
Prolonged Nociceptive Behavior and Central Sensitization are Attenuated by
Pregabalin in a Mouse Trigeminal Neuropathic Pain Model
Chapter 4: Discussion 50
4.1 Central sensitization and nociceptive behavior 50
4.2 Nociceptive behaviour: contralateral effects 51
4.3 Effects of pregabalin 51
4.4 Clinical implications 53
4.5 Study strengths and limitations 54
4.6 Future research directions 56
Chapter 5: Conclusions 58
References 59

vi
Abbreviations
AIDS Acquired immunodeficiency syndrome
ANOVA Analysis of variance
ATP Adenosine triphosphate
c-Fos Cellular oncogene Fos
CGRP Calcitonin gene-related peptide
CNS Central nervous system
CNX Cervical nerve transection
CO2 Carbon dioxide
COX Cyclooxygenase
CS Central sensitization
EMG Electromyography
GABA Gamma-Aminobutyric acid
GFAP Glial fibrillary acidic protein
HIV Human immunodeficiency virus
i.p. Intraperitoneal
IAN Inferior alveolar nerve
IANR Inferior alveolar nerve regeneration
IANX Inferior alveolar nerve injury
IASP International Association for the Study of Pain
IL-I Interleukin-1beta
ION-CCI Infraorbital nerve ligation
IONX Infraorbital nerve transection
MDH Medullary dorsal horn
MO Mustard oil
MWT Mechanical nociceptive withdrawal thresholds
NMDA N-methyl-D-aspartate
NS Nociceptive-specific
P2X Purinoceptor subtype 2X
p38 MAPK P38 mitogen-activated protein kinases
RCT Root canal therapy

vii
RF Mechanoreceptive field
RFm Reticular formation
RM Repeated measures
SEM Standard error of mean
SGC Satellite glial cells
TMD Temporomandibular disorder
WDR Wide dynamic range

1
Chapter 1 General Introduction
1.1 Introduction
According to the IASP, pain is an unpleasant multidimensional sensory and emotional experience
associated with actual or potential tissue damage, or described in terms of such damage (1). Pain
is the most frequent chief complaint for which patients come to a dental office. While some of
the pain complaints can be easily managed by providing an adequate dental treatment usually
followed by over-the-counter medications, others require comprehensive evaluation and
diagnosis with long-lasting therapeutic and psychological approaches. Unrelieved pain can cause
changes in the patient’s life, additional autonomic symptoms and changes in the peripheral and
central nervous systems. Pain has also a motivational component that may affect the patient’s
behaviour, reflexes and sleep. Understanding the physiology of pain will provide better
therapeutic methods for pain management, improving the patient’s experience and preventing
negative outcomes associated with the pain experience.
1.2 Pain
Being a multidimensional experience, pain is associated with actual or potential tissue damage.
Pain has a sensory component that translates and transmits nociceptive signals via the peripheral
and central nervous systems up to a final destination in the cortical regions such as the
somatosensory cortex where the processing related to “pain sensation” (which is always a
psychological state) takes place. The affective or emotional component of pain comes into play
in the unpleasantness of pain. The motivational component of pain provides a conscious
manifestation of a pre-conscious perception of threat to body tissues that motivates a subject to
avoid or remove the threat (2). The cognitive component of pain is related to the patient’s
personal experiences and beliefs. If the patient believes in a negative outcome of the treatment,
he or she may catastrophize the actual experience and perceive pain to be of a higher intensity
(e.g., painful experience from a dental office visit may intensify pain experience several years
later).

2
1.2.1 Multidisciplinarity and socioeconomical impact of pain
The prevalence of chronic pain is considerably high, estimated to be within the range of 12-30 %
in Europe, and about 20% of the adult population in Canada, according to recent surveys (3-5).
The estimated cost in lost productive time from chronic pain conditions is over US$ 60 billion
annually (6). The burdens of related drug abuse, depression, and complications of opioid
treatment are more difficult to quantify (6). A recent Canadian survey has demonstrated that
more than 50% of patients with chronic pain have reported reduced quality of life, job loss or
reduced job responsibilities and about 30% indicated increased rates of depression (7). In
Canada, the personal financial costs for patients with pain is estimated around $1,500 per month
(3). Chronic pain conditions carry a huge economic cost of several billion dollars per year in the
United States and Canada (3, 8-10).
Also noteworthy is that more than 40 million people undergo surgical procedures every year in
the United States. A high proportion of these patients undergo major surgical procedures that
result in nerve damage with the potential for the subsequent development of chronic pain
conditions (6). About 1.7 million people in the United States are survivors of limb loss, and each
year over 130 000 new amputees are added to that number (6). There are estimates that around
50–80% of these patients experience significant, long-term phantom or residual limb pain (11).
Up to 10% of these patients develop severe, life-changing, chronic pain. Analogous findings of
chronic pain after trigeminal nerve damage are reviewed below.
1.2.2 Acute vs. chronic pain
Pain is commonly divided into two categories: acute and chronic pain. Acute pain usually carries
a “protective” role to warn the body of potential or real tissue damage. The majority of acute
pain conditions can be successfully treated and will heal uneventfully. However, there are reports
indicating that about 20% of acute pain conditions can transition into a chronic state (12-18).
Furthermore, chronic pain can be associated with a variety of chronic diseases and disorders such
as arthritis, diabetes, cancer, HIV/AIDS or be an “independent entity” such as migraine,
temporomandibular disorders (TMD), trigeminal neuralgia and fibromyalgia (12). It is generally
accepted that chronic pain does not confer a positive biological role in most cases.

3
1.2.3 Classification of pain: nociceptive, inflammatory and
neuropathic
Nociception is a process of detection and signalling the presence of a noxious stimulus.
Nociceptive pain is an alarm mechanism that identifies and signals the presence of a damaging or
potentially damaging stimulus. This mechanism takes place when noxious stimuli activate
primary afferent neurons (nociceptors) that innervate peripheral tissues such as skin, bone,
muscle, connective tissues, vessels and viscera. There are two types of primary afferent neurons
that normally transmit nociceptive signals: unmyelinated (C fibres) and myelinated (A fibres).
The axons of these neurons carry sensory information from the peripheral tissues to the dorsal
horn of the spinal cord and trigeminal brainstem sensory nuclear complex. Inflammatory pain is
produced following persistent tissue injury, as occurs after trauma, surgery or during acute and
chronic inflammatory diseases (19). In these conditions, damaged cells and inflammatory cells
recruited to the site of damage release substances that activate, and/or sensitize, peripheral
nociceptors (12, 20, 21).
Neuropathic pain is a condition caused as a direct consequence of a lesion or a disease of the
parts of the nervous system that normally signal pain (22-24). This pain can be associated with
such conditions as traumatic nerve injury, diabetic peripheral neuropathy, AIDS, post-herpetic
neuralgia, or pain originating in the central nervous system, as in spinal cord injury and multiple
sclerosis and stroke (19).
1.2.4 Classification and prevalence of orofacial pain
There is a consensus on three major steps that are used to classify orofacial pain (25, 26). During
the first step, cluster analysis identifies different entities among all patients. The second step is
based on diagnostic criteria for each of the previously identified groups. During the third step, a
group of experts decide on unaddressed cases after the first two steps (25, 26).
The prevalence of toothache pain in the general population has been reported to be around 12%
(27). Acute pain occurring within one week following root canal treatment (RCT) has been
reported in 1.6% to 6.6% of all RCT-treated patients (16). Occurrence of persistent pain for up to
6 months after RCT has been reported in 3% to 12% of patients (13-16, 28). Extrapolation of this

4
data to an estimate for the United States (and Canadian) populations indicates that in the United
States approximately 870,000 (96,000 in Canada) of the new cases of persistent pain occur
following relatively common dental treatment each year, with 550,000 (61,000 in Canada) cases
of such pain not having an identifiable local reason explaining why it is present (15). The
prevalence of TMD pain ranges from 9% to 15% in the adult population and this pain appears to
be two times as common in women than in men (29, 30). Similarly, migraine headache is more
prevalent in women and affects about 20% of women and 7% of men (31, 32). The prevalence of
non-migranious headaches is relatively high and can be experienced by 60% to 80% of adults
(31). The incidence of trigeminal neuralgia, a condition characterized by a sudden, brief
paroxysmal stabbing pain, is about 27 patients per 100,000 persons annually (33, 34). Other pain
conditions in the orofacial region (atypical odontalgia and atypical facial pain) are discussed in
the next section.
1.2.5 Trigeminal neuropathic pain (atypical odontalgia and
atypical facial pain)
Root canal therapy frequently involves the extirpation of pulp tissue and injury of the nerves
supplying the pulp. Two persistent pain conditions in which pulp nerve injury has been
implicated are atypical odontalgia and atypical facial pain. Atypical odontalgia is defined as pain
in or around a tooth, which is not related to any dental cause and is often mistaken as toothache
and treated with multiple dental treatments (35, 36). Atypical odontalgia has also been defined by
the International Headache Society as a subgroup of persistent idiopathic facial pain, also
including atypical facial pain. Atypical odontalgia and atypical facial pain share the definition of
‘persistent facial pain that does not have the characteristics of the cranial neuralgias and is not
attributed to another disorder’ (37). These pains may in fact constitute a sub-set of trigeminal
neuropathic pain resulting from injury of sensory fibres supplying the extirpated pulp and have
been well characterized by Baad-Hansen (35) and List et al. (38). Physiological testing (38) has
shown that patients with atypical odontalgia have peripheral and central sensitization changes.
Characteristically, atypical odontalgia pain persists during most of the day, it is non-paroxysmal
(18, 35-38) and it can affect both sexes and all adult ages, although there is a preponderance of
women in their mid-40s who are affected. It has been suggested that genetic predisposition and
environmental influences can contribute to the severity of the pain (18, 35-39). The diagnosis of

5
atypical odontalgia and atypical facial pain is often difficult, and is based on the exclusion of
conditions with known pathophysiology in the teeth or adjacent structures (35, 37). Baad-Hansen
et al. have suggested that the management of atypical odontalgia and atypical facial pain is
primarily based on expert opinion and case reports and that these conditions are often difficult to
manage effectively. Therefore, it is very important to study the mechanisms involved in the
development and maintenance of atypical odontalgia and other orofacial neuropathic conditions
and how they might be effectively managed.
1.3 Pain management
Based on the survey of the American Dental Association, the most common orofacial pain
treatments provided by dentists are provision of occlusal appliances, occlusal adjustment
(equilibration), thermal packs, medications and diet counseling (40). Historically, many of the
treatment modalities have been based on personal opinions. The current clinician’s approach
promotes an evidence-based concept of treatment with a focus on translation of the new
scientific evidence into clinical settings. It is based on the integration of personal clinical
experience with the best available evidence emerging from current research. The area of
orofacial pain management includes non-pharmacological and pharmacological approaches.
1.3.1 Non-pharmacological management of orofacial pain
In the case of dental pain, management of pain associated with dentin exposure may include
occlusion of dentinal tubules to prevent intra-tubular fluid flow or application of potassium ions
that reduce the excitability of pulpal primary afferents (41). Treatment of reversible pulpitis due
to caries or defective restoration includes removal of caries and provision of a new restoration
with integral marginal seal (42). The definitive management of pain associated with irreversible
pulpitis is RCT. Another approach is the extraction of the affected tooth (42). Pain associated
with periodontal diseases is treated by irrigation, root planning and pocket elimination. In some
periodontal situations, tooth extraction may be required (43). Pain due to the apical or
periodontal abscesses is quite often treated as an emergency by surgical incision and drainage
(44).

6
Non-pharmacological treatment options for patients with persistent pain in the orofacial region
may include occlusal therapies such as the adjustment of teeth, provision of bite appliances,
orthodontic and fixed prosthodontic treatments with or without surgical corrections. For long-
term management, the provision of an inter-occlusal appliance can benefit the patient. Other
treatment modalities for patients with persistent orofacial pain may include physiotherapy in the
orofacial area (45-47).
There is some progress in management of chronic orofacial pain states such as headache TMD
and trigeminal neuralgia by using the traditional acupuncture (48). Cognitive behavioural therapy
is another approach that has been shown to improve outcomes for patients with TMD (49).
1.3.2 Pharmacological management of orofacial pain
Pharmacological management of orofacial inflammatory pain aims to block or reduce the
nociceptive input from the peripheral site (e.g. tooth pulp, bone, soft tissue), block nociceptive
impulse propagation along the peripheral nerve and reduce neuroplastic changes in the central
nervous system. Administration of short and long-lasting local anaesthetics following tissue
manipulations during dental procedures prevents generation and propagation of these nociceptive
impulses along the primary afferents. Drugs that block peripheral sensitization induced by
inflammatory process are used to minimize the nociceptive input from the periphery.
Medications that block the synthesis of prostaglandins or cyclooxygenase (COX) enzymes peri-
operatively are another approach to minimize the peripheral nociceptive input. To attenuate
changes that can occur following peripheral acute inflammatory processes, medications that act
in the central nervous system such as acetaminophen and opioids also may be used. However,
opioids have many adverse effects such as nausea, vomiting and drowsiness (50). Non-opioid
analgesics include the following medications: salicylates (aspirin, diflunisal), acetaminophen,
non-steroid anti-inflammatory drugs such as ibuprophen and naproxen and COX-2 inhibitors
such as celecoxib (51). A combination of analgesics is another pharmacological approach to deal
with inflammatory pain that does not respond to a single agent alone (52). The rationale behind
this approach is to affect several mechanisms involved in the development and maintenance of
inflammatory pain.

7
Treatment of neuropathic pain is complex and challenging, and cannot be restricted to the
implicated peripheral tissue. Pharmacological treatment for neuropathic pain includes
medications such as opioids, antidepressants, anticonvulsants and topical medications.
Opioids such as morphine, oxycodone, fentanyl, hydrocodone and codeine act principally via the
-receptor. However, there is a risk of drug abuse from prescribing opioids, and some degree of
tolerance to opioids has been demonstrated; therefore, more attention is required from a clinician
prescribing this type of medication (53).
The tricyclic antidepressants such as amitriptyline, nortriptyline, imipramine, desipramine and
doxepin are often effective in the management of diabetic peripheral neuralgia, post-herpetic
neuralgia and post-mastectomy chronic pain (54). The mechanism of action of these drugs is
related to the blockage of reuptake of noradrenaline and serotonin in the brain that are released in
response to pain. Thus, tricyclic antidepressants provide a prolonged inhibitory action via
mechanisms involving noradrenaline and serotonin.
Topical drugs such as lidocaine and capsaicin act peripherally to induce analgesia and have been
demonstrated to be effective in relieving pain in post-herpetic neuralgia (55).
Anticonvulsant drugs such as carbamazepine and gabapentin have been shown to be effective in
treating patients with diabetic neuropathy, malignancy-related pain, post-herpetic neuralgia and
any neuropathic pain states including trigeminal neuralgia (39, 55-58).
1.3.3 Anticonvulsant drug pregabalin
Recent studies have demonstrated that pregabalin, an anticonvulsant drug (a potent α2δ-calcium
channel blocker) that influences glutamatergic neurotransmission, is effective in treating
neuropathic pain conditions (55, 56, 58-63). However, pregabalin still has to undergo detailed
investigation in patients with trigeminal neuropathic as well as in acute inflammatory pain
models. Systemic effects of pregabalin can significantly inhibit ectopic discharges from injured
afferent neurons (64). Our group has demonstrated that pregabalin can decrease sensorimotor
responses and glutamate release in an acute orofacial inflammatory pain model in rats (65). The
effects of pregabalin on neuronal activity in an acute orofacial inflammatory pain model have not
been studied yet. There is evidence indicating that intrathecal application of pregabalin reduces

8
the enhanced noxious stimulus-induced spinal release of glutamate seen in neuropathic rats.
There are also reports indicating that pregabalin at varying doses reduces nociceptive responses
in the spinal dorsal horn neurons in rat pain models (66-69). We have previously demonstrated a
dose-dependent effect of pregabalin in reversing the facial mechanical allodynia and medullary
dorsal horn (MDH) central sensitization present at postoperative day 7 following partial infra-
orbital nerve injury (70).
Pregabalin does not appear to have been tested in acute inflammatory and long-lasting trigeminal
neuropathic pain models; therefore, there is no scientific underpinning supporting or contesting
its clinical use for trigeminal neuropathic pain conditions. More studies are required to document
and understand the underlying mechanisms of pregabalin actions in acute inflammatory and
long-lasting neuropathic pain models.
1.4. Mechanisms and models of orofacial pain
Orofacial pain is not only a “simple” transmission of the nociceptive input up to the cortical
areas along the peripheral and central parts of the nervous system, but is strongly associated with
cellular changes that take place along that pathway (71-74). Furthermore, not only nociceptive
neurons, but also different types of neuronal and glial cells are involved in acute and chronic pain
states (71, 72, 74). Peripheral and central sensitizations are contributing factors to the
hyperalgesia, allodynia, spontaneous and referred pain and pain spread that may be manifested in
pain conditions (71-73).
1.4.1 Peripheral processes in the trigeminal system
The orofacial region including the teeth, skin, temporomandibular joint and orofacial
musculature are innervated mainly by branches of the trigeminal nerve. Among the primary
afferent nerve fibres that terminate in sense organs (receptors), there is a population of slowly
conducting small-diameter primary afferents with free nerve endings which are activated by
noxious stimuli. Activation of these nociceptors generates action potentials in these afferents that
conduct these signals into the central nervous system and that can result in pain.
Damage to peripheral tissues induces the release of chemical mediators such as serotonin,
histamine, and tumor necrosis factor-alpha from mast cells, macrophages and immune cells (12,

9
20, 21). Nociceptive afferents themselves also are capable of releasing neurotransmitters such as
substance P and calcitonin gene-related peptide (CGRP). Additionally, sympathetic efferents
innervating peripheral blood vessels and skin can release noradrenaline and activate nociceptive
afferent. These chemicals can activate certain receptors on the nociceptive afferents and result in
the release of second messengers inside the cell. Another way by which nociceptive afferents can
be activated is by application of inflammatory irritants such as mustard oil (MO) and capsaicin
that activate TRPA1 and TRPV1 receptors respectively. Subsequently, the activated nociceptive
afferents may become hyperexcitable to the noxious and non-noxious stimuli. Since damage-
induced chemical mediators may spread through tissues, the changes in nociceptive afferent
sensitivity can also occur in neighbouring primary nociceptive afferents. The increased
excitability of the nociceptive endings can lead to spontaneous activity, lowered activation
thresholds and increased responsiveness to noxious stimuli. This process of increased sensitivity
of primary afferents due to changes that take place after tissue damage is called peripheral
sensitization. Peripheral sensitization may contribute to such clinical conditions as allodynia,
hyperalgesia and spontaneous pain.
Many primary afferents innervate the tooth pulp. What sets the tooth pulp apart from other
peripheral tissues is its very low compliance due to dentinal boundaries. This factor may
significantly contribute to increased sensitivity in states such as reversible and irreversible
pulpitis.
The majority (exceptions are jaw muscle spindle afferents and some mechanosensitive afferents
supplying periodontal tissues) of somatosensory primary afferents that innervate orofacial tissues
have their cell bodies in the trigeminal ganglion. The central projections of these primary afferent
cell bodies enter the brainstem and may ascend or descend in the trigeminal spinal tract from
which they give off collaterals that terminate in one or more subdivisions of the trigeminal
brainstem sensory nuclear complex (12, 71-73).
There is growing evidence that neurons and satellite glial cells (SGCs) in the trigeminal ganglion
undergo changes following acute and chronic peripheral injury; analogous changes occur in the
dorsal root ganglion in the spinal somatosensory system (75-79). Prolonged discharges of
primary afferents can increase the expression of a variety of sodium channels in the trigeminal
ganglion neurons that can lead to an increase in the excitability of trigeminal nerve afferents
(80). The tetrodotoxin-resistant sodium channels and potassium channels are thought to be

10
involved in an enhancement of trigeminal neuronal activity following trigeminal nerve injury
(81). Peripheral injury induces gene expression, neuropeptide generation and increased neuronal
excitability in the trigeminal ganglion (12, 75, 78). Neurons and SGC may release neuropeptides
such as CGRP and substance P that affect their activity (82). Novel findings from several groups
indicate that gap junctions between SGCs in the trigeminal ganglion may be important in
spreading excitatory signals between glial cells and neurons (78, 79). Alterations of
neuropeptides, receptors, cytokines, and growth factors in trigeminal neurons are thought to be
possible mechanisms that cause an increase in the excitability of trigeminal neurons following
trigeminal nerve injury (83).
As a result of all these processes described above, the increased nociceptive activity can generate
an increased afferent barrage into the central nervous system. There, additional functional
changes can occur in central nociceptive processing and contribute to the pain experience.
1.4.2 Central processes in the trigeminal system
The trigeminal primary afferents activate second-order neurons within the trigeminal brainstem
sensory nuclear complex which can be subdivided into the principal or main sensory nucleus and
the spinal tract nucleus which comprises three subnuclei (oralis, interpolaris, caudalis; Fig. 1,
(12, 71-73, 84)). Subnucleus caudalis extends into the cervical spinal cord where it merges with
the spinal dorsal horn (12, 71-73). Due to the high functional and anatomical similarity between
spinal dorsal horn and trigeminal subnucleus caudalis, latter has been designated as the
‘medullary dorsal horn’ or MDH. Many neurons in the four components of the trigeminal
brainstem complex contribute to ascending nociceptive or non-nociceptive pathways involved in
the somatosensory function or modulation (12, 71-73). The trigeminal brainstem complex has a
somatotopic or topographic organization (12, 71-73, 85).
Many of the small-diameter primary afferents terminate in the MDH (12, 71, 73, 85). Noxious
stimulation of peripheral orofacial tissues results in the release from the nociceptive central
endings of substance P, CGRP, glutamate and somatostatin which act on receptors of second-
order sensory neurons to produce a long-latency, sustained excitation of these neurons (12, 71,
85). Numerous findings indicate that MDH serves as the principal brainstem relay site of
trigeminal nociceptive information to higher brain centres involved in the discrimination of pain
and also to local brainstem neurons involved in nociceptive reflexes (72, 73).

11
There are anatomical and electrophysiological similarities between MDH and the spinal dorsal
horn (12, 71, 73, 85). For example, MDH has a laminated structure similar to the spinal dorsal
horn, and like the spinal dorsal horn, nociceptive neurons occur in the MDH and can be
categorized into two main groups on the basis of their cutaneous (or mucosal) receptive field
properties: nociceptive-specific (NS) neurons, which receive small-diameter afferent inputs from
A-delta and/or C fibres and which respond only to noxious stimuli (e.g. pinch and heat) applied
to a localized craniofacial receptive field; and wide dynamic range (WDR) or convergent
neurons, which may receive large-diameter and small-diameter A-fibre inputs as well as C-fibre
inputs and which are excited by non-noxious (e.g. tactile) stimuli as well as by noxious stimuli.
The NS and WDR neurons are concentrated in the superficial (I/II) and deep (V/VI) laminae of
MDH.
Many trigeminal brainstem neurons project to the thalamus either directly, or indirectly via
polysynaptic pathways that may involve the reticular formation (12, 71, 85). These projections
carry signals that reach the higher brain centres involved in the somatosensory perception (e.g.
touch and pain) and other functions (e.g. emotion and motivation). The projections from the
trigeminal brainstem complex to the thalamus can result in the activation of neurons in thalamus
which directly transmits signals to neurons in the overlying somatosensory cerebral cortex.

12
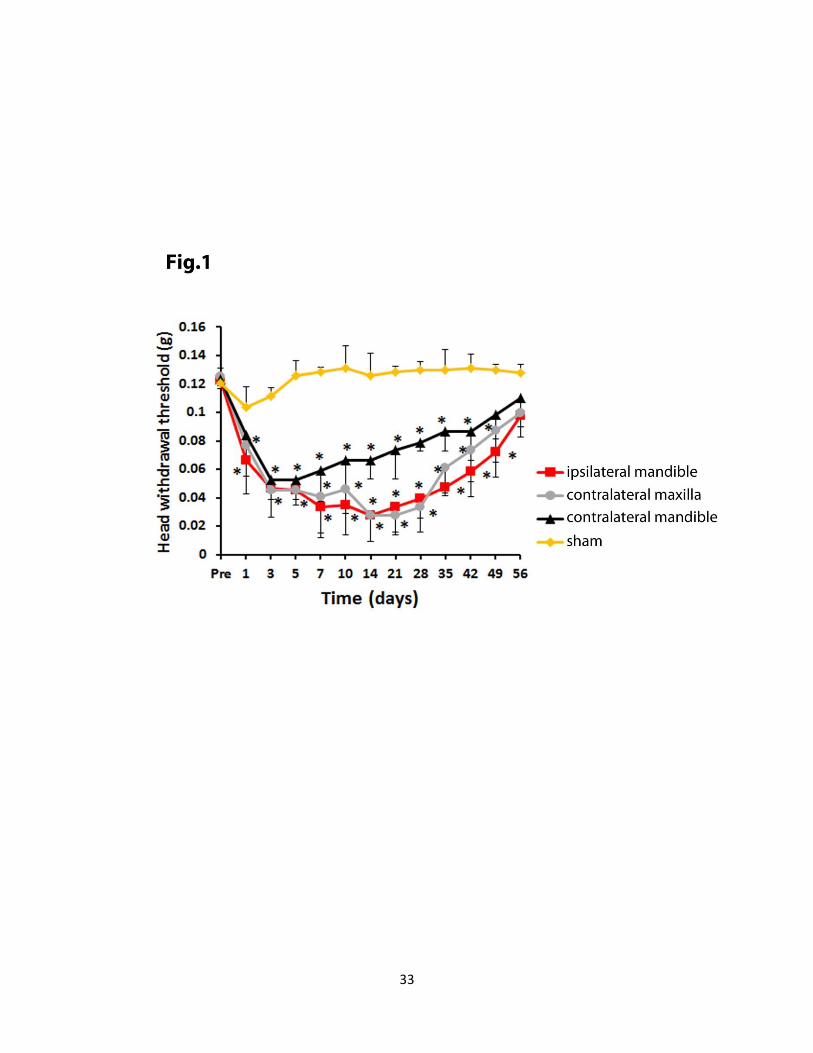
Figure 1. Major trigeminal somatosensory pathways from the orofacial region. Trigeminal
primary afferents project via the trigeminal ganglion to second-order neurons in the trigeminal
brainstem sensory nuclear complex. These neurons may project to neurons in higher levels of the
brain (for example, in the thalamus) or to neurons in brainstem regions such as the cranial nerve
motor nuclei or the reticular formation (RF). Not shown are the projections of some cervical
nerve and cranial nerve VII, IX, X, and XII afferents to the trigeminal complex and the
projection of many V, VII, IX, and X afferents to the solitary tract nucleus (from Sessle (12)).
1.4.3 Central sensitization
There are two major means by which neuroplasticity can be induced in central somatosensory
pathways; first, an increased nociceptive afferent input (e.g. by direct stimulation of peripheral
nerves by an injury or by inflammation) and second, a decreased afferent input (e.g. through
nerve damage resulting in deafferentation). As a result, an increased neuronal excitability may
occur, accompanied by pain behaviour. Pain associated with changes in the central nervous
system has been viewed as a reflection of a centrally based “functional plasticity” or “central
sensitization” (12, 71, 73, 74, 85, 86).

13
Central sensitization of nociceptive neurons can be produced by nerve inflammation or damage,
such as that associated with pulpectomy or transection of dental nerve fibres, and is reflected as
an increase in nociceptive neuronal mechanoreceptive field (RF) size, a decrease in mechanical
activation threshold and an increase in spontaneous activity and in responses to noxious RF
stimuli. Central sensitization thus reflects a hyperexcitability of nociceptive processes in the
central nervous system and has been implicated as an important mechanism in acute as well as
chronic pain conditions following injury or inflammation of peripheral tissues (12, 71, 73, 74,
85, 86, 88).
Injury of primary afferents that results in central sensitization can involve several processes. For
example, the generation of abnormal impulses by affected primary afferents, formation of
neuroma and consequently generation of abnormal peripheral discharges, sprouting of the
afferents into neighbouring tissues, abnormal expression of different receptors by primary
afferents, development of physical contacts among sympathetic efferents and nociceptive
afferents, sprouting of nociceptive afferents in the central nervous system can be the underlying
mechanisms of induction and maintenance of central sensitization.
Central sensitization has been well documented in nociceptive neurons in MDH, but can also
occur in other nociceptive neurons along the trigeminal nociceptive pathway (e.g. subnucleus
oralis, ventrobasal thalamus, etc., (89, 90)). In addition, several studies have demonstrated the
involvement not only of neurons but also of non-neural cells (e.g., glia, and cells of immune
system) in the development and maintenance of orofacial neuropathic pain states (12, 74, 79).
Futhermore, several chemical mediators such as glutamate and endogenous ATP and their
receptors have been shown to be essentially involved in the initiation of central sensitization in
the MDH in rodent models of acute and chronic pulpitis pain (84, 91-94) and chronic trigeminal
neuropathic pain (70, 95).
1.4.4 Models of orofacial pain
The nociceptive neurons in MDH and other higher centres can be activated not only by noxious
mechanical and/or thermal stimuli applied to the orofacial region, but also by application of
algesic chemicals and inflammatory irritants to the orofacial tissues such as the tooth pulp of rat
(84, 93, 96, 97). In the trigeminal system, application of algesic chemicals and inflammatory

14
irritants into orofacial tissues can markedly increase the RF and responses of NS and WDR
neurons in the MDH and reduce mechanical activation threshold (MAT, (12, 71)). Injection of
inflammatory irritants into cutaneous or deep orofacial tissues can induce acute nociceptive
behaviour in humans and animals (12). Depending on the inflammatory irritant, the nociceptive
behaviour associated with inflammation may last for hours or even months. Central sensitization
in the inflammatory pain models is usually reversible, but can be associated with pain behaviour
that lasts for hours or even longer.
Several studies have demonstrated that application of MO to the tooth pulp of rat induces
electromyography (EMG) activity in jaw muscles, expression of c-Fos (a marker of neuronal
activity) and increased neuronal excitability in the MDH (12, 65, 98). In this model increased
neuronal excitability of the MDH nociceptive neurons involves activation of NMDA and P2X
receptors, and several mediators such as serotonin, NMDA, and IL-I (12, 92, 93, 97)There is
also evidence that brainstem astrocytes and microglia are involved in the developing and
maintenance of central sensitization in acute and chronic pulpitis models in rats (79, 94). Pre-
emptive administration of microglial inhibitors can prevent the development of central
sensitization in rats (79, 99).
Several models of trigeminal neuropathic pain have also been developed in rats and mice: the
inferior alveolar nerve (IAN) injury (IANX) model, the infraorbital nerve ligation (ION-CCI)
model, the infraorbital nerve transection (IONX) model (70, 100), the inferior alveolar nerve
regeneration (IANR) model, and the cervical nerve transection (CNX) model (83, 101, 102).
There is a modulation of spike discharges in the primary afferent neurons following such
peripheral nerve injury in rats (83, 103). It has been shown in rats that primary afferent neurons
are sensitized and the activation threshold in these neurons became lower following the long-
lasting abnormal spike generation in injured primary afferents for more than several weeks (104).
This prolonged discharge can increase expression of a variety of sodium channels in the
trigeminal ganglion neurons that can lead to an increase in the excitability of trigeminal nerve
afferents (80). The tetrodotoxin-resistant sodium channels and potassium channels are thought to
be involved in an enhancement of trigeminal ganglion neuronal activity following trigeminal
nerve injury in rats (81).

15
The alteration of chemical mediators including neuropeptides, receptors, cytokines, and growth
factors in trigeminal ganglion neurons is thought to be a possible mechanism that causes an
increase in the excitability of trigeminal ganglion neurons and increased input to the MDH
nociceptive neurons following trigeminal nerve injury in rats (83). Glial fibrillary acidic protein
(GFAP) expression increases in the trigeminal ganglion following nerve injury, and becomes
detectable by immunocytochemistry following nerve injury in rodents (105). The increase in
GFAP in SGCs in the trigeminal ganglion after nerve injury may be triggered by increased
glutamate released in the sensory ganglion, resulting from increased neuronal firing in rats and
mice (75, 106). It has previously been demonstrated that the number of gap junctions between
SGCs in the trigeminal ganglion of rodents increases following trigeminal nerve injury,
suggesting that the changes in SGC gap junctions can be a factor in generating or maintaining
neuropathic pain (75, 78).
After the long-lasting hyperactivity of the primary afferent neurons, a barrage of action potentials
is conveyed to the central nervous system (CNS), resulting in the production of central
sensitization of the brain stem nociceptive neurons (70, 83, 94, 107). There are several reports
indicating that hyperactive astroglial cells in the MDH of rats are significantly involved in the
central sensitization of trigeminal nociceptive neurons (108, 109). Hyperactive astroglial cells
were found in the rat MDH following IAN transection and this was also associated with an
increased activity of nociceptive neurons. The proposed mechanism of astroglial involvement in
the development and maintenance of trigeminal central sensitization is via excessive release of
glutamine which is taken up in the primary afferent terminals via glutamate transporters,
resulting in an increase in the glutamate release at the synaptic cleft.
Another important population of non-neuronal cells that has been shown to be involved in
neuropathic pain mechanisms in rats is microglial cells. These cells are activated 1–3 days after
IAN transection, whereas the activation of astroglial cells takes 7–14 days after that (94, 109). It
has recently been reported that astroglial and microglial cells have specific interactions and
communicate with each other. The astroglial and microglial cell interactions may be involved in
the hyper-activation of the MDH nociceptive neurons following trigeminal nerve injury in rats
(108).

16
1.4.5 Modulation of orofacial pain mechanisms
The central sensitization that takes place in the nociceptive neurons in the MDH following
peripheral injury is a reflection of neuroplasticity. There are also other modulatory mechanisms
of somatosensory transmission and these are not limited to the MDH area but can also occur in
thalamic and cortical areas (12, 71, 73, 74, 89, 90). Most of the modifications of the ascending
somatosensory transmission, however, take place at the level of trigeminal brainstem complex.
Each subdivision of the trigeminal brainstem complex receives numerous peripheral inputs and
interconnections from other parts of the brain. These interactions that are derived from the
periphery are termed afferent inhibition, while those from the other brain regions are called
descending modulation. There are interconnections among neurons of MDH, ascending
modulatory influences of MDH on more rostral regions, and descending influences from regions
such as the periaqueductal gray and cerebral cortex to the trigeminal brainstem complex (71).
Since many of the sensory brainstem neurons are involved in somatosensory and autonomic
reflexes, modification of sensory transmission can affect motor and autonomic functions. For
example, our recent study has demonstrated that application of the inflammatory irritant MO to
the rat’s tooth pulp induces spontaneous EMG activity in the jaw-opening and jaw-closing
muscles (65).
1.5 Statement of the problem and rationale
There have been no reports in the literature documenting whether injury of the mouse ION can
induce long-lasting facial nociceptive behaviour and central sensitization in functionally
identified trigeminal nociceptive neurons in the MDH. There have also been no studies
documenting the effect of the anticonvulsant drug pregabalin on long-lasting nociceptive
behaviour and central sensitization following ION injury, and whether pregabalin is effective in
attenuating central sensitization in functionally identified MDH nociceptive neurons in the
trigeminal nociceptive system in an acute rodent inflammatory orofacial pain model. Thus there
is a need to collect pre-clinical data on the possible therapeutic effectiveness of pregabalin in
treating orofacial acute and chronic orofacial pain conditions.

17
Chapter 2 Project Specific Aims and Hypotheses
The specific aims of this study were to test (i) if trigeminal nerve injury in mice produces
prolonged nociceptive behaviour and induces central sensitization in functionally identified
nociceptive neurons in the MDH and (ii) if systemic administration of pregabalin can reverse
these nociceptive changes and (iii) if MDH mustard oil-induced central sensitization in rats and
mice can be attenuated by systemic administration of pregabalin.
Our two working hypotheses were:
1) injury of the ION in mice induces nociceptive behaviour and central sensitization in
functionally identified nociceptive neurons in the MDH
2) Pregabalin affects central sensitization in nociceptive neurons in the MDH in acute and
chronic pain models in mice and rats and reverses nociceptive behaviour in a chronic pain
model in mice

18
Chapter 3 Articles
3.1 Article 1
(Submitted for publication)
Prolonged Nociceptive Behavior and Central Sensitization are Attenuated by Pregabalin in
a Mouse Trigeminal Neuropathic Pain Model
Pavel S. Cherkas, DMD, PhD1,2
, Vidya Varathan, BDS, PhD2, Shimon Friedman, DMD
1, Limor
Avivi-Arber, DMD, PhD2,3
, Barry J. Sessle, MDS, PhD2,4
1 Discipline of Endodontics, Faculty of Dentistry, University of Toronto, Toronto, Ontario,
Canada
2 Discipline of Oral Physiology, Faculty of Dentistry, University of Toronto, Toronto, Ontario,
Canada
3 Discipline of Prosthodontics, Faculty of Dentistry, University of Toronto, Toronto, Ontario,
Canada
4 Department of Physiology, Faculty of Medicine, University of Toronto, Toronto, Ontario,
Canada
Correspondence to:
Pavel S. Cherkas, DMD, PhD,
Faculty of Dentistry, Departments of Oral Physiology and Endodontics, University of Toronto,
124 Edward Street, Toronto, Ontario, M5G 1G6, Canada
Tel: +1 416 979 4910; fax: +1 416 979 4936; e-mail: [email protected]
Funding disclosure: This research was supported in part by a Research Grant from the
American Association of Endodontists Foundation, Canadian Academy of Endodontics
Endowment Fund, NIH grant DE-04786 and Pfizer Canada.
Dr. Sessle was awarded a 2009-2011 research grant from Pfizer Canada.

19
Abstract
Introduction: Chronic post-endodontic pain may be generated by trigeminal nerve injury. Since
central sensitization may underlie the development of chronic pain, this study aimed to examine
in mice if (i) trigeminal nerve damage produces prolonged nociceptive behavior and induces
central sensitization in functionally identified nociceptive neurons in the medullary dorsal horn
(MDH) and (ii) if systemic administration of pregabalin can modulate these nociceptive
changes.
Methods: Facial mechanical nociceptive withdrawal thresholds (MWT) were tested in adult A/J
male mice with von Frey filaments applied to facial skin, pre-operatively and post-operatively up
to day 56 following infraorbital nerve transection (IONX) or sham surgery. On post-operative
days 7, 21 and 49, MWT were assessed before and after administration of pregabalin (75mg/kg
i.p., n=11/group) or isotonic saline (vehicle control, n=11/group). MDH nociceptive neurons
were also recorded at similar post-operative days (n=7/group), and their mechanical activation
threshold (a decrease of which reflects central sensitization) was assessed before and after
pregabalin administration.
Results: The MWT values were significantly reduced in the IONX group (p<0.05; ANOVA,
followed by post-hoc tests) compared to sham, up to post-operative day 49. At post-operative
days 7, 21 and 49, pregabalin (but not saline) reversed the IONX-induced reduction of MWT. A
decreased mechanical activation threshold in MDH nociceptive neurons was also recorded, and
this effect too was reversed by pregabalin (but not saline).
Conclusions: This study demonstrated that pregabalin attenuated IONX-induced mechanical
allodynia and the associated MDH central sensitization in mice and indicated that pregabalin
might be useful in management of orofacial neuropathic pain conditions.

20
Introduction
Persistent pain conditions carry a huge socioeconomic burden, at a financial cost of several
billion dollars per year in both the United States and Canada (1, 2). A specific persistent pain
condition, chronic post-endodontic pain may be induced by trigeminal nerve injury, associated
with extirpation of pulp tissue and injury of the nerves supplying the pulp during root canal
treatment (RCTx). It has been estimated that more than 22 million endodontic procedures (15.1
million RCTxs) are performed annually in the United States alone (3). Acute pain occurs within
one week of RCTx in 3% to 11.6% of all treated patients (4-7); it may become chronic in 3 to 6%
of patients (6-12), if persistent post-endodontic pain develops in 5% of treatments, then more
than a million patients will experience a chronic neuropathic pain for a period of 6 months or
longer. Post-endodontic pain is typically treated with various short-term analgesics taken orally
when the pain becomes severe. This regimen is highly effective in the majority of patients (13)
but not in all those experiencing post-endodontic pain, especially chronic post-endodontic pain
which is often difficult to manage effectively.
Chronic pain involves a process of central sensitization which refers to an increase in the
excitability of nociceptive neurons within the central nervous system (CNS) and is reflected as a
decrease in mechanical activation threshold and an increase in neuronal mechanoreceptive field
(RF) size and response to noxious RF stimuli. Central sensitization has been implicated as an
important process in acute as well as chronic pain conditions following injury or inflammation of
peripheral tissues, including trigeminal nerve injury (14, 15). We have previously demonstrated
that glutamate and endogenous ATP and their receptors are essentially involved in the initiation
of central sensitization in the trigeminal subnucleus caudalis (also termed the medullary dorsal
horn, MDH) in rodent models of acute pulpitis pain (16-19) and chronic trigeminal neuropathic
pain (20, 21).
There has been limited study of recently developed analgesic drugs in these models. Recent
studies have shown that pregabalin, an anticonvulsant drug (a potent α2δ-calcium channel
blocker) that influences glutamatergic neurotransmission (22, 23), is effective in treating many
neuropathic and inflammatory chronic pain conditions in both patients and animals (24-26). We
have demonstrated that nociceptive sensorimotor behavioral responses and the medullary release
of glutamate in the acute inflammatory tooth pulp pain model can be attenuated in a dose-

21
dependent manner by systemic administration of pregabalin (27). To our knowledge, however,
pregabalin has not been tested in a mouse trigeminal neuropathic pain model in order to provide
pre-clinical data bearing on its potential use for trigeminal neuropathic pain conditions in
humans. Therefore, the aims of this study were to test in mice if (i) trigeminal nerve damage
produces prolonged nociceptive behavior and induces central sensitization in functionally
identified nociceptive neurons in the medullary dorsal horn (MDH) and (ii) if systemic
administration of pregabalin can modulate these nociceptive changes.

22
Materials and Methods
Animals
One hundred and twenty-six male A/J mice (Jackson laboratories, USA) aged 8-12 weeks and
weighing 18-24 g, were used in this study. All experimental procedures were approved by the
University of Toronto Animal Care committee in accordance with the Ontario Animal Research
Act (Canada). All efforts were used to limit the number of animals required for the study.
Surgical procedure
Under isoflurane anesthesia (4% induction, 2–2.5% maintenance), an incision was made in the
right maxillary gingivo-buccal groove. The infra-orbital nerve (ION), which supplies the
maxillary whisker pad, mucosa and anterior teeth, was exposed and dissected free from the
surrounding tissue. The ION was lifted from the maxillary bone and totally sectioned (IONX)
with iridectomy scissors. In sham mice, the ION was exposed but not sectioned. Post-
operatively, mice were monitored daily.
Behavioral testing
A total of 66 mice were used in the behavioral experiments [sham and IONX; IONX-pregabalin
at post-operative days 7, 21 or 49 and IONX-saline (11 per group)]. Mice were initially trained
for at least 3 consecutive days to stay in a plastic cage and protrude their snout through a hole in
the cage wall, during mechanical stimulation of the maxillary or mandibular facial area with von
Frey filaments, as previously described (28). Escape responses to the mechanical stimulation
were demonstrated as a sudden backward withdrawal movement of the head from the cage
opening. Baseline values of the animal’s mechanical withdrawal threshold (MWT) were obtained
by slowly applying graded von Frey filaments to the contralateral maxillary or mandibular and
ipsilateral mandibular areas of the face. The MWT at the maxillary or mandibular area was
defined as the lowest filament intensity that evoked three or more escapes out of five stimulation
trials with intervals of more than 10 seconds. Behavioral scoring was carried out at days 1-3
before IONX (baseline values), at day 0 (1 h before IONX) and up to post-operative day 56. On
post-operative days 7, 21 and 49, MWT was assessed before and after intraperitoneal (i.p.)
administration of pregabalin (Pfizer, Canada; 75 mg/kg i.p., n=11) or isotonic saline (vehicle

23
control, n=11). The pregabalin dosage chosen has been shown to be effective in other pain
models (20, 27). Experiments were carried out in ‘blind’ design in all groups.
Electrophysiological recording and stimulation procedures
A total of 60 mice were used in the electrophysiological study [sham at post-operative days 7, 21
or 49 (3 per group); IONX at post-operative days 7, 21 or 49 (3 per group); IONX-pregabalin
and IONX-saline at post-operative days 7, 21 or 49 (7 per group)]. Two to three neurons in each
animal were studied in the IONX and sham groups, but only one neuron was studied in each
animal in those IONX groups receiving pregabalin (or saline) due to the long-lasting effect of
pregabalin. Evoked single neuron activity was recorded in histologically verified sites in the deep
laminae of the right MDH (lateral: 0.8-1.0 mm; posterior: 0.8-1.0 mm), as previously described
(17, 18, 29). Briefly, neuronal responses to stimulation of the orofacial region were amplified
and displayed on an oscilloscope and computer and the data were later analyzed off-line.
Mechanical (brush, pressure and pinch) and noxious thermal stimuli were applied to classify
neurons recorded as nociceptive-specific (NS) if they responded exclusively to the noxious
stimulation, as previously described (17, 18). Each NS neuron's cutaneous orofacial receptive
field (RF) was determined with nonserrated forceps, and its activation threshold to mechanical
stimulation of its RF was assessed by force-monitoring forceps. Central sensitization was
reflected as a decrease in activation threshold, as previously described (17, 18). Pregabalin (75
mg/kg) or saline was administered (i.p.) after the baseline properties of each neuron were
obtained in each animal group at 7, 21 or 49 days following IONX. The neuronal properties were
re-assessed at 60 minutes after pregabalin or saline administration, as previously reported (20).
Recording sites were marked by electrolytic lesions (anodal current of 8 µA for 13 s) and
verified histologically (Fig. 3B).
Sample size calculation
Based on the results of a pilot study, we calculated the sample size required to compare
populations means with 95% power, of 11 mice for behavioral testing and 6 neurons for
electrophysiological experiments per group.

24
Analysis
Data were tested for normality (Kolmogorov–Smirnov test) and equal variance. Data are
reported as mean ± SEM. For the behavioral tests, differences in MWT between the baseline
(average of the 3 preoperative day values) and each postoperative day were analyzed with1-way
analysis of variance (ANOVA) followed by the Bonferroni post-hoc test. The effects of
pregabalin compared with saline (as vehicle control) were tested by 1-way ANOVA followed by
the Bonferroni post-hoc test. Differences between the groups (sham, IONX, IONX after
pregabalin and IONX after saline) were analyzed with 2-way ANOVA followed by Dunnett's
test. For neuronal recordings, differences within the same post-operative day group (7, 21 or 49
days) in neuronal mechanical activation threshold between the groups (sham, IONX, IONX after
pregabalin and IONX after saline) were analyzed with 1-way ANOVA followed by the
Bonferroni post-hoc test. The level of significance was set at a P < 0.05.
Results
Behavioral testing
Nociceptive MWT to mechanical stimulation of the contralateral maxillary and ipsilateral and
contralateral mandibular areas were tested in the IONX and sham-operated animals (Fig. 1).
When compared with sham animals, the IONX group demonstrated a significant (2-way
ANOVA, p< 0.05) decrease in MWT in the ipsilateral mandibular, contralateral maxillary and
mandibular area. The decrease in MWT, considered to reflect mechanical allodynia, lasted up to
day 42 following IONX for all three facial sites and up to day 49 for the contralateral maxillary
and ipsilateral mandibular areas (2-way ANOVA, p< 0.05).
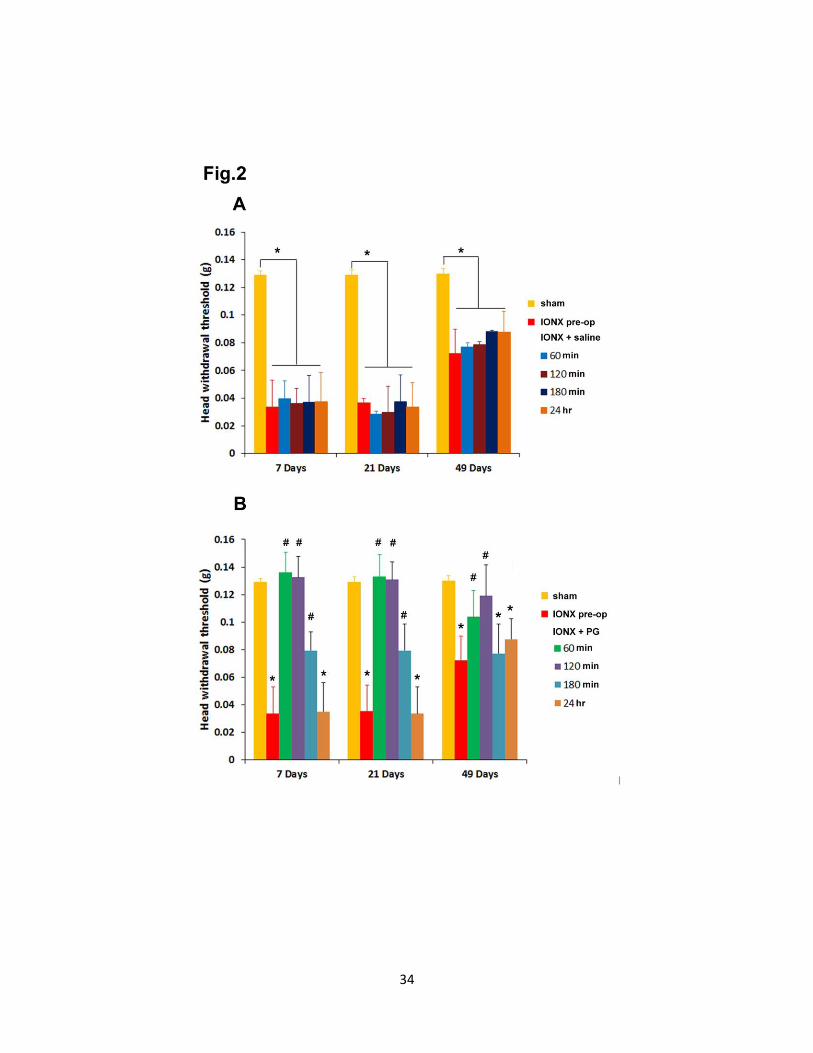
The effects on nociceptive behavior of pregabalin or vehicle (control) were studied in 3 different
groups of IONX animals tested at 7, 21 or 49 days following IONX (Fig. 2). Administration of
pregabalin significantly reversed the IONX-induced decreased MWT values to control levels for
two hours at days 7, 21 and 49 (2-way ANOVA, p<0.05). Administration of vehicle (control) did
not reversed the IONX-induced decreased MWT to control levels (2-way ANOVA, p<0.05).

25
Electrophysiological recording
Eighty-four functionally identified NS neurons were studied in the sham (control) and in the
IONX groups at days 7, 21 and 49. All neurons were located in the deep laminae of the MDH
(Fig 3, B). The mechanical activation threshold of single nociceptive MDH neurons in sham
animals at days 7, 21 and 49 did not change significantly (1-way ANOVA followed by
Bonferroni post-hoc test, Fig.3 A). The IONX produced significant decrease in mechanical
activation threshold at days 7, 21 and 49 (1-way ANOVA followed by Bonferroni post-hoc test),
indicative of central sensitization in the neurons. Administration of pregabalin 60 minutes before
neuronal evaluation significantly reversed the IONX-induced central sensitization at day 7, 21
and 49 (1-way ANOVA followed by Bonferroni post-hoc test, p>0.05). Administration of saline
(vehicle control) did not significantly impact on the IONX-induced decreased mechanical
activation threshold (1-way ANOVA followed by Bonferroni post-hoc test, p>0.05).
Discussion
This study has demonstrated for the first time that injury of the mouse ION produced long-
lasting facial nociceptive behavior, reflected as a decrease in MWT, and also induces central
sensitization in trigeminal nociceptive neurons in the MDH, reflected as a decrease in
mechanical activation threshold. This is the first report to document that administration of
pregabalin (but not saline as vehicle control) reduced the long-lasting nociceptive behavior and
central sensitization at days 7, 21 and 49 after IONX. These findings indicated that pregabalin
might be effective in treating orofacial neuropathic pain conditions, which could be of significant
clinical importance.
Central sensitization of central nociceptive neurons, reflected as a decrease in mechanical
activation threshold and increases in their responses to noxious RF stimuli, has been implicated
as an important process in acute and chronic orofacial pain conditions following injury or
inflammation of peripheral tissues (16, 17, 30, 31). The present findings were consistent with our
recent results (20) indicating that pregabalin is effective in attenuation of facial mechanical
hypersensitivity and central sensitization in a rat model of trigeminal neuropathic pain (20).
Also, in another recent study, we have documented in a model of acute inflammatory dental pain
that pregabalin is effective in a dose-dependent manner in significantly attenuating the

26
nociceptive sensorimotor behavioral responses and medullary release of glutamate evoked by
application of the inflammatory irritant mustard oil to the rat tooth pulp (27). Reports have
demonstrated the effectiveness of pregabalin in the spinal nociceptive system since it can reduce
nociceptive behavioral responses in rat models of neuropathic pain (22, 32). Our previous study
(20) demonstrates in the rat trigeminal neuropathic pain model a dose-dependent effect of
pregabalin in reversing the facial mechanical allodynia and MDH central sensitization at an early
post-operative stage (day 7) but the present study demonstrated that pregabalin might be
effective at longer post-operative times, even when the nociceptive behavior and central
sensitization have become well maintained.
One limitation of this work is the unknown site and mode of action of pregabalin (33) that were
not addressed in this study, although it has been proposed that pregabalin can reduce the influx
of calcium by binding to the α2δ protein of calcium channels. This action reduces the release of
neurotransmitters such as glutamate, norepinephrine and substance P (34-37). Another limitation
of this study was use of general anaesthesia for the MDH study and use of awake animals for the
testing of nociceptive behaviour. General anaesthesia may have an effect on neuronal properties
by blocking normal conductivity and altering membrane permeability. Despite this, the MDH
and behavioural data as well as effects of pregabalin on them were nonetheless complementary.
Systemic administration of pregabalin can significantly inhibit ectopic discharges from injured
peripheral sensory neurons (38), this could partially explain the effect of pregabalin in the
present study with the drug reducing or preventing afferent input to the MDH NS neurons. In a
previous study, we have demonstrated that administration of pregabalin abolishes spontaneous
activity in NS MDH neurons in a trigeminal neuropathic pain model in rats (20) which is
consistent with this possibility. Nevertheless, there is an additional possibility of direct
pregabalin action in the central nervous system since intrathecal application of pregabalin
reduces the enhanced noxious stimulus-induced spinal release of glutamate seen in neuropathic
rats (22). Pregabalin has affinity to receptors in the cortex, olfactory bulb, hypothalamus,
amygdala, hippocampus, cerebellum and dorsal horn of the spinal cord (39).
Detailed clinical studies of the potential use of pregabalin in orofacial pain states are limited
(40). However, several reports have demonstrated the analgesic success of pregabalin in the
treatment of paresthesia following inferior alveolar nerve damage (41), glossopharyngeal (42)
and lacrimal (43) neuralgias as well as post-traumatic neuropathic facial pain (44). The current

27
study provided pre-clinical data suggesting that pregabalin might be effective in the management
of trigeminal neuropathic pain conditions, including chronic post-endodontic pain.
Conclusions
This study demonstrated for the first time that pregabalin attenuated the long-lasting IONX-
induced mechanical allodynia and associated MDH central sensitization induced by trigeminal
nerve injury. It provided pre-clinical evidence to support the use of pregabalin in treating
orofacial neuropathic pain conditions.

28
References.
1. Stewart WF, Ricci JA, Chee E, Morganstein D, Lipton R. Lost productive time and cost
due to common pain conditions in the US workforce. JAMA 2003;290:2443-2454.
2. Sessle BJ. Unrelieved pain: a crisis. Pain Res Manag 2011;16:416-420.
3. Survey of Dental Services Rendered, Chicago, American Dental Association. 2006.
4. Al-Negrish AR, Habahbeh R. Flare up rate related to root canal treatment of
asymptomatic pulpally necrotic central incisor teeth in patients attending a military
hospital. J Dent 2006;34:635-640.
5. Eleazer PD, Eleazer KR. Flare-up rate in pulpally necrotic molars in one-visit versus two-
visit endodontic treatment. J Endod 1998;24:614-616.
6. Nixdorf DR, Moana-Filho EJ, Law AS, McGuire LA, Hodges JS, John MT. Frequency of
persistent tooth pain after root canal therapy: a systematic review and meta-analysis. J
Endod 2010;36:224-230.
7. Nixdorf DR, Moana-Filho EJ, Law AS, McGuire LA, Hodges JS, John MT. Frequency of
nonodontogenic pain after endodontic therapy: a systematic review and meta-analysis. J
Endod 2010;36:1494-1498.
8. Campbell RL, Parks KW, Dodds RN. Chronic facial pain associated with endodontic
therapy. Oral Surg Oral Med Oral Pathol 1990;69:287-290.
9. Keenan AV. Only a small percentage of patients experience persistent pain for more than
6 months after root canal therapy. J Evid Based Dent Pract 2010;10:235-236.
10. Marbach JJ, Hulbrock J, Hohn C, Segal AG. Incidence of phantom tooth pain: an atypical
facial neuralgia. Oral Surg Oral Med Oral Pathol 1982;53:190-193.
11. Ng YL, Glennon JP, Setchell DJ, Gulabivala K. Prevalence of and factors affecting post-
obturation pain in patients undergoing root canal treatment. Int Endod J 2004;37:381-391.
12. Polycarpou N, Ng YL, Canavan D, Moles DR, Gulabivala K. Prevalence of persistent
pain after endodontic treatment and factors affecting its occurrence in cases with
complete radiographic healing. Int Endod J 2005;38:169-178.
13. Su Y, Wang C, Ye L. Healing rate and post-obturation pain of single- versus multiple-
visit endodontic treatment for infected root canals: a systematic review. J Endod;37:125-
132.
14. List T, Leijon G, Svensson P. Somatosensory abnormalities in atypical odontalgia: A
case-control study. Pain 2008;139:333-341.
15. Sessle BJ. Acute and chronic craniofacial pain: brainstem mechanisms of nociceptive
transmission and neuroplasticity, and their clinical correlates. Crit Rev Oral Biol Med
2000;11:57-91.
16. Chiang CY, Li Z, Dostrovsky JO, Hu JW, Sessle BJ. Glutamine uptake contributes to
central sensitization in the medullary dorsal horn. Neuroreport 2008;19:1151-1154.

29
17. Chiang CY, Park SJ, Kwan CL, Hu JW, Sessle BJ. NMDA receptor mechanisms
contribute to neuroplasticity induced in caudalis nociceptive neurons by tooth pulp
stimulation. J Neurophysiol 1998;80:2621-2631.
18. Chiang CY, Zhang S, Xie YF, Hu JW, Dostrovsky JO, Salter MW, et al. Endogenous
ATP involvement in mustard-oil-induced central sensitization in trigeminal subnucleus
caudalis (medullary dorsal horn). J Neurophysiol 2005;94:1751-1760.
19. Hu B, Chiang CY, Hu JW, Dostrovsky JO, Sessle BJ. P2X receptors in trigeminal
subnucleus caudalis modulate central sensitization in trigeminal subnucleus oralis. J
Neurophysiol 2002;88:1614-1624.
20. Cao Y, Wang H, Chiang CY, Dostrovsky JO, Sessle BJ. Pregabalin suppresses
nociceptive behavior and central sensitization in a rat trigeminal neuropathic pain model.
J Pain 2013;14:193-204.
21. Miyamoto M, Tsuboi Y, Takamiya K, Huganir RL, Kondo M, Shinoda M, et al.
Involvement of GluR2 and GluR3 subunit C-termini in the trigeminal spinal subnucleus
caudalis and C1-C2 neurons in trigeminal neuropathic pain. Neurosci Lett 2011;491:8-12.
22. Kumar N, Laferriere A, Yu JS, Leavitt A, Coderre TJ. Evidence that pregabalin reduces
neuropathic pain by inhibiting the spinal release of glutamate. J Neurochem
2010;113:552-561.
23. Quintero JE, Dooley DJ, Pomerleau F, Huettl P, Gerhardt GA. Amperometric
measurement of glutamate release modulation by gabapentin and pregabalin in rat
neocortical slices: role of voltage-sensitive Ca2+ alpha2delta-1 subunit. J Pharmacol Exp
Ther 2011;338:240-245.
24. Christensen D, Gautron M, Guilbaud G, Kayser V. Effect of gabapentin and lamotrigine
on mechanical allodynia-like behaviour in a rat model of trigeminal neuropathic pain.
Pain 2001;93:147-153.
25. Lindsay TJ, Rodgers BC, Savath V, Hettinger K. Treating diabetic peripheral neuropathic
pain. Am Fam Physician 2010;82:151-158.
26. Tzellos TG, Toulis KA, Goulis DG, Papazisis G, Zampeli VA, Vakfari A, et al.
Gabapentin and pregabalin in the treatment of fibromyalgia: a systematic review and a
meta-analysis. J Clin Pharm Ther 2010;35:639-656.
27. Narita N, Kumar N, Cherkas PS, Chiang CY, Dostrovsky JO, Coderre TJ, et al. Systemic
pregabalin attenuates sensorimotor responses and medullary glutamate release in
inflammatory tooth pain model. Neuroscience 2012;218:359-366.
28. Saito K, Hitomi S, Suzuki I, Masuda Y, Kitagawa J, Tsuboi Y, et al. Modulation of
trigeminal spinal subnucleus caudalis neuronal activity following regeneration of
transected inferior alveolar nerve in rats. J Neurophysiol 2008;99:2251-2263.
29. Hu JW. Response properties of nociceptive and non-nociceptive neurons in the rat's
trigeminal subnucleus caudalis (medullary dorsal horn) related to cutaneous and deep
craniofacial afferent stimulation and modulation by diffuse noxious inhibitory controls.
Pain 1990;41:331-345.

30
30. Sessle BJ. Peripheral and central mechanisms of orofacial inflammatory pain. Int Rev
Neurobiol 2011;97:179-206.
31. Sessle BJ. Peripheral and central mechanisms of orofacial pain and their clinical
correlates. Minerva Anestesiol 2005;71:117-136.
32. Hurley RW, Chatterjea D, Rose Feng M, Taylor CP, Hammond DL. Gabapentin and
pregabalin can interact synergistically with naproxen to produce antihyperalgesia.
Anesthesiology 2002;97:1263-1273.
33. Bialer M. Why are antiepileptic drugs used for nonepileptic conditions? Epilepsia
2012;53 Suppl 7:26-33.
34. Cunningham MO, Woodhall GL, Thompson SE, Dooley DJ, Jones RS. Dual effects of
gabapentin and pregabalin on glutamate release at rat entorhinal synapses in vitro. Eur J
Neurosci 2004;20:1566-1576.
35. Dooley DJ, Donovan CM, Pugsley TA. Stimulus-dependent modulation of
[(3)H]norepinephrine release from rat neocortical slices by gabapentin and pregabalin. J
Pharmacol Exp Ther 2000;295:1086-1093.
36. Dooley DJ, Mieske CA, Borosky SA. Inhibition of K(+)-evoked glutamate release from
rat neocortical and hippocampal slices by gabapentin. Neurosci Lett 2000;280:107-110.
37. Fink K, Dooley DJ, Meder WP, Suman-Chauhan N, Duffy S, Clusmann H, et al.
Inhibition of neuronal Ca(2+) influx by gabapentin and pregabalin in the human
neocortex. Neuropharmacology 2002;42:229-236.
38. Chen SR, Xu Z, Pan HL. Stereospecific effect of pregabalin on ectopic afferent
discharges and neuropathic pain induced by sciatic nerve ligation in rats. Anesthesiology
2001;95:1473-1479.
39. Bian F, Li Z, Offord J, Davis MD, McCormick J, Taylor CP, et al. Calcium channel
alpha2-delta type 1 subunit is the major binding protein for pregabalin in neocortex,
hippocampus, amygdala, and spinal cord: an ex vivo autoradiographic study in alpha2-
delta type 1 genetically modified mice. Brain Res 2006;1075:68-80.
40. Zakrzewska JM. Medical management of trigeminal neuropathic pains. Expert Opin
Pharmacother 2010;11:1239-1254.
41. Lopez-Lopez J, Estrugo-Devesa A, Jane-Salas E, Segura-Egea JJ. Inferior alveolar nerve
injury resulting from overextension of an endodontic sealer: non-surgical management
using the GABA analogue pregabalin. Int Endod J 2012;45:98-104.
42. Kitchener JM. Glossopharyngeal neuralgia responding to pregabalin. Headache
2006;46:1307-1308.
43. Pareja JA, Cuadrado ML. Lacrimal neuralgia: So far, a missing cranial neuralgia.
Cephalalgia 2013:doi: 10.1177/0333102413488000.
44. Singh RK, Sinha VP, Pal US, Yadav SC, Singh MK. Pregabalin in post traumatic
neuropathic pain: Case studies. Natl J Maxillofac Surg 2012;3:91-95.

31
Figure legends
Figure 1. Time course of the MWT in bilateral maxillary and mandibular facial areas after
IONX. In the control (sham) group (n=11), the thresholds did not differ from pre-operative
values bilaterally in mandibular area (yellow line). In the IONX groups (n =11), bilateral
mechanical allodynia was established at day 1 after surgery. The MWT to mechanical
stimulation of the ipsilateral mandibular (red line), contralateral maxillary (grey line) and
mandibular (black) area were significantly (n=11, 2-way ANOVA, p<0.05) different from the
control (sham) values and lasted for up to day 42 following IONX. The MWT to mechanical
stimulation of the ipsilateral mandibular (red line) and contralateral maxillary (grey line) were
significantly different from the control (sham) values for up to 49 days post-operatively. *P < .05
for comparison between sham group values and values at the different time points after IONX
(2-way ANOVA). All values are shown as mean ± SEM.
Figure 2. The effects on the nociceptive behavior of pregabalin (PG, 75 mg/kg, i.p., B) or saline
(vehicle control, A) were determined at 7, 21 and 49 days post-operatively. Administration of
pregabalin (75 mg/kg, i.p., B) but not saline (vehicle control, A) significantly reversed the
IONX-induced decreased MWT values to control levels for two hours at days 7, 21 and 49
(n=11, 2-way ANOVA followed by Dunnett's test, p<0.05). *P < .05 for comparison between
sham group values and values at the different time points after IONX (2-way ANOVA followed
by Dunnett's test). #P < .05 for comparison between the pre-treatment (pre-tx) IONX values and
values at the different time points after pregabalin administration (1-way ANOVA followed by
Bonferroni post-hoc test). All values are shown as mean ± SEM.
Figure 3. (A) Changes in mechanical activation threshold (MAT) of MDH nociceptive neurons
at 7, 21 and 49 days following IONX. Each group consists of 7 NS neurons.The IONX
significantly (1-way ANOVA followed by Bonferroni post-hoc test) decreased the MAT
compared to control values at all tested days (red bars). Administration of pregabalin (75 mg/kg,
i.p., PG, green bars) significantly (1-way ANOVA followed by Bonferroni post-hoc test)
reversed the IONX-induced decreased MAT values at days 7, 21 and 49. Saline (vehicle control,
blue bars) did not affect the IONX-induced decreased MAT values at days 7, 21 and 49
following IONX (1-way ANOVA followed by Bonferroni post-hoc test). (B) Histologically
confirmed neuronal recording sites in sham and experimental groups. The sites were plotted onto

32
a section of the caudal medulla (−4.4 mm behind interaural line). Sp 5, trigeminal spinal tract;
MDH, medullary dorsal horn (trigeminal subnucleus caudalis). All values are shown as mean ±
SEM. *P < .05.
33
34

35

36
3.2 Article 2
(Submitted for publication)
Pregabalin Blocks Central Sensitization in Medullary Dorsal Horn in a Rodent Model of
Acute Tooth Pulp Inflammatory Pain.
P.S. Cherkas1, M. Miyamoto
1,3, V. Varathan
1, B.J. Sessle
1,2
1. Faculty of Dentistry, Department of Oral Physiology,
2. Faculty of Medicine, Department of Physiology, University of Toronto, Toronto, Ontario,
Canada.
3. Department of Physiology, Nihon University School of Dentistry, Tokyo, Japan
Correspondence to:
Pavel S. Cherkas, DMD, PhD,
Faculty of Dentistry, Departments of Oral Physiology and Endodontics, University of Toronto,
124 Edward Street, Toronto, Ontario, M5G 1G6, Canada
Tel: +1 416 979 4910; fax: +1 416 979 4936; e-mail: [email protected]
Funding disclosure: This study was supported by NIH grant DE-04786 and Pfizer Canada.

37
Abstract
Acute inflammation of the tooth pulp can induce nociceptive sensorimotor behavioural responses
as well as glutamate release in the rat medullary dorsal horn (MDH) and central sensitization
reflected as increases in receptive field (RF) size and responses to noxious stimuli and a decrease
in mechanical activation threshold in functionally identified MDH nociceptive neurons. The aim
of this study was to test if MDH central sensitization in this acute inflammatory pain model in
rats and mice can be attenuated by systemic administration of pregabalin, a potent α2δ-calcium
channel agent that is used for the management of especially neuropathic pain. Male adult rats and
mice were anaesthetized with α-chloralose (50-75 mg/kg)/urethane (1-1.125 g/kg) i.p.. The right
maxillary first molar pulp and the dorsal surface of the caudal medulla were surgically exposed.
The activity of single neurons was recorded at histologically verified sites in the MDH, and
mechanical (brush, pressure, and pinch) stimuli were applied to functionally identified
nociceptive-specific (NS) neurons. Pulp application of mineral oil (vehicle) produced no
evidence of central sensitization in NS neurons, compared to their baseline properties. However,
application of mustard oil (MO) to the pulp induced MDH central sensitization in rats and mice,
as reflected in significant increases in RF size (40-130%, n= 6/group, repeated measures
ANOVA, p<0.05) and responses to noxious stimuli (80-200%, n=6/group, p<0.05) and
decreased mechanical activation threshold (45-75%, N=6/group, p<0.05). Compared to vehicle
control treatment, administration of pregabalin (100 mg/kg, i.p.) markedly reduced the MO-
induced central sensitization in rats and mice to baseline levels in all measured parameters: RF
size (n=6/group, 2-way ANOVA, p<0.05), responses to noxious stimuli (n=6/group, 2-way
ANOVA, p<0.05) and mechanical activation threshold (n=6/group, 2-way ANOVA, p<0.05).
These results indicate that pregabalin can block central sensitization in the MDH in a rodent
acute inflammatory pain model. Further understanding of the mechanisms involved in
pregabalin-induced reduction in MDH central sensitization in inflammatory, as well as
neuropathic, orofacial pain models may lead to broader clinical applications of pregabalin and
other anticonvulsant drugs in pathological pain states.
Highlights
• Application of mustard oil to the tooth pulp induces trigeminal central sensitization
Pregabalin blocks central sensitization in trigeminal nociceptive neurons

38
Abbreviations: MDH, medullary dorsal horn; NS, nociceptive-specific
Keywords: pregabalin, central sensitization, plasticity, trigeminal, dorsal horn, pain
Introduction
We have previously shown in an acute tooth pulp inflammatory pain model that application of
the inflammatory irritant mustard oil (MO) to the rodent molar tooth pulp induces central
sensitization in trigeminal subnucleus caudalis (also termed the medullary dorsal horn, MDH)
and involves N-methyl-D-aspartate (NMDA) glutamatergic receptor mechanisms [10]. Central
sensitization of central nociceptive neurons is reflected as an increase in their mechanoreceptive
field (RF) size, a decrease in mechanical activation threshold and an increase in responses to
noxious RF stimuli, and has been implicated as an important process in acute and chronic
orofacial pain conditions following injury or inflammation of peripheral tissues [15, 32-34].
Pregabalin is an anticonvulsant drug that is known to decrease glutamate release by binding to
the α2δ subunit of voltage-dependent calcium channels [26, 30]. Recent evidence indicates that
pregabalin is effective in treating many chronic inflammatory and especially neuropathic pain
conditions in patients and reducing nociceptive behaviour in animal pain models [21, 23-26, 28,
31, 35-38]. Recently, we have shown that nociceptive sensorimotor behavioral responses and the
medullary release of glutamate evoked by MO application to the rodent tooth pulp can be
attenuated in a dose-dependent manner by systemic administration of pregabalin [28]. Also, we
have documented that pregabalin attenuates the mechanical allodynia and MDH central
sensitization in a rat trigeminal neuropathic pain model [4]. However, it is unclear whether
pregabalin interferes with central sensitization in functionally identified MDH nociceptive
neurons in conditions of acute inflammatory pain. Therefore, the aim of this study was to test if
MDH central sensitization in this acute inflammatory pain model in rats and mice can be
attenuated by systemic administration of pregabalin. Some of the data has been presented in
abstract form [6-8].
Material and Methods
All procedures and surgeries were approved by the University of Toronto Animal Care
Committee in accordance with the regulations of the Ontario Animal Research Act (Canada).

39
Animal preparation
Adult male Sprague-Dawley rats (280-400g, n=18) were initially anesthetized by intraperitoneal
α-chloralose (50 mg/kg)/urethane (1g/kg) and adult male C57BL6 mice (18-24g, n=18) by α -
chloralose (75 mg/kg)/urethane (1.125 g/kg). The animals were prepared as previously described
in detail [10, 12]. Briefly, the coronal pulp of the right maxillary first molar was exposed and
covered with a saline-soaked cotton pellet, and then the dorsal surface of the caudal medulla was
surgically exposed. Each animal then received a continuous intravenous infusion of a mixture of
70% α -chloralose/urethane solution (0.2 g/ml) and 30% pancuronium solution (1mg/ml in rats
and 0.1mg/ml in mice) at a rate of 0.4 or 0.04 ml/h, respectively. The animal was artificially
ventilated throughout the whole experiment and heart rate, expired CO2 level, and rectal
temperature were continuously monitored and maintained at physiological levels.
Electrophysiological recording and stimulation procedures
Spontaneous and evoked single neuron activity was recorded in histologically verified sites in
the deep laminae of the right MDH (lateral: 1.4-2.0 mm; posterior: 1.5-2.0 mm referred to the
obex in rats and lateral: 0.8-1.0 mm; posterior: 0.8-1.0 mm in mice), as previously described in
detail [10, 12, 20]. Briefly, neuronal responses to stimulation of the orofacial region were
amplified and displayed on an oscilloscope and computer and the data were later analyzed off-
line with Spike 2 software (Cambridge Electronic Desighn, Science Park, Milton Road,
Cambridge, UK). Mechanical (brush, pressure and pinch) and noxious thermal (radiant heat 51–
53°C) stimuli were applied to classify neurons recorded in the deep laminae of MDH as
nociceptive-specific (NS) if they responded exclusively to the noxious stimulation, as previously
described [10, 12]. Each NS neuron's cutaneous orofacial RF was determined with nonserrated
forceps, and its activation threshold to mechanical stimulation of its RF was assessed by force-
monitoring forceps. Its responses were also recorded to graded noxious pressure applied to the
RF by the forceps (50 g, 75 g and 100 g in rats and 10 g, 15 g and 26 g in mice applied in
ascending order, each for 5 s at an interval of >45 s), and the number of spikes evoked by each of
these graded stimuli were summed. The level of any spontaneous activity of a NS neuron was
determined over an initial 1-min recording period. As previously documented [10, 12], central
sensitization was reflected as an increase in spontaneous activity, RF size and responses to

40
noxious stimuli, and a decrease in activation threshold. Recording sites were marked by
electrolytic lesions (anodal current of 8 µA for 13 s) and verified histologically.
Experimental paradigm
Only one neuron was studied in each experiment due to the long-lasting effects of MO and
pregabalin. Two assessments of neuronal properties were carried out before each animal received
a bolus (1 ml in rats, 0.5 ml in mice, i.p.) of isotonic saline or pregabalin (100 mg/kg). The dose
of pregabalin was chosen on the basis of that used in the early studies [4, 37] and on our recent
sensorimotor behavioral study defining the dose-response relationship of pregabalin on the
behaviour [28]. Then, 30 min later, MO (n=6) or its vehicle (mineral oil, n=6) was applied in
each animal (at room temperature) to the exposed pulp which was then sealed with CAVIT
(ESPE, Seefeld/Oberbayren, Germany). Starting at three minutes after the solution was applied
to the pulp, neuronal properties were assessed at 10 min intervals over the next 50 min.
Statistical analyses
Statistical analyses were based on normalized data (in percentages) of orofacial RF size,
responses to graded pressure or pinch stimuli and mechanical activation threshold and
spontaneous activity. Differences between baseline values and values at different time-points
after MO or vehicle (mineral oil) application and after pregabalin or vehicle and MO application
were treated by repeated measures (RM) analysis of variance (ANOVA) or ANOVA on ranks,
followed by Dunnett's test. Differences between the groups of the same species were treated by
2-way ANOVA followed by Dunnett's test. The level of significance was set at a P value of less
than 0.05. All values are presented as mean ± SEM.
Results
A total of 36 functionally identified NS neurons in rats (n=18) and mice (n=18) were studied. In
both rats and mice, one group of neurons (n=6) was tested with MO application to the pulp
preceded by saline (i.p.), another (n=6) with MO to the pulp preceded by pregabalin, and another
(n=6) with mineral oil to the pulp preceded by saline. The recording sites of all NS neurons were
histologically verified and were located in the deep laminae of MDH (see Fig. 1G, H). The
baseline level of spontaneous activity was low (0–2 spikes/min) in all NS neurons in both rats

41
and mice. Application of mineral oil (vehicle) to the tooth pulp in rats and mice produced no
evidence of central sensitization in any of the NS neurons (data not shown), compared to their
baseline properties. Only one neuron in rats and two in mice had baseline activity (0.03-0.05 Hz)
before and after mineral oil application. In contrast to the lack of the effect of mineral oil, MO
application to the pulp produced an immediate response in 50% (rats) and 83% (mice) of the NS
neurons tested. In these neurons, there was a brief burst (latency 10–30 s; duration 3–4 min) of
discharges followed by a long-lasting (20–30 min) firing (3–4 spikes/min) that was significantly
(RM ANOVA, p<0.05) higher than baseline level. In 34% (rats) and 17% (mice) of the neurons,
MO induced no immediate burst of firing and only a mild (1–3 spikes/min) long-lasting firing; in
the remaining neurons it produced no changes in background activity. Administration (i.p.) of
saline control had no effect on baseline activity of the NS neurons prior and after MO application
to the tooth pulp in both rats and mice. Administration (i.p.) of pregabalin had also no effect on
baseline activity of the NS neurons prior to MO application, but significantly (2-way ANOVA,
p<0.05) reduced all MO-induced immediate and long-lasting background activity in both rats
and mice.
Orofacial RF size
Application of MO to the pulp induced significant increases in RF size in the NS neurons in rats
(130%, RM ANOVA, p<0.05) and in mice (40%, RM ANOVA, p<0.05). Application of mineral
oil to the pulp did not induce any significant increases in RF size in the NS neurons in either rats
or mice (RM ANOVA, p<0.05). The effect of MO on orofacial RF size was significantly larger
in rats than in mice (Fig.1 A, B). In addition, a transient novel tactile RF appeared in two of the
six neurons following MO application in both rats and mice (data not shown), consistent with our
earlier findings in rats [4, 10] . Compared to saline (control) treatment, administration of
pregabalin significantly (2-way ANOVA, p<0.05) reversed the MO-induced increased RF size to
baseline levels in both rats and mice (Fig. 1 A, B).
Mechanical activation threshold
Application of MO to the pulp significantly (RM ANOVA, p<0.05) decreased mechanical
activation threshold in rats (45%, Fig. 1C) and in mice (75%, Fig. 1D). Application of mineral
oil to the pulp did not decrease mechanical activation threshold in the NS neurons in either rats

42
or mice (RM ANOVA, p>0.05). The effect of MO on mechanical activation threshold lasted
significantly longer in mice than in rats (Fig.1 C, D; 2-way ANOVA, p<0.05). Administration of
pregabalin (but not saline control) significantly reversed the MO-induced decreased mechanical
activation threshold to baseline levels in both rats and mice (p<0.05, 2-way ANOVA, Figs. 1 C,
D).
Responses to noxious stimuli
Application of MO to the pulp significantly (RM ANOVA, p<0.05) increased responses to the
noxious stimuli in rats (200%, Fig. 1E) and in mice (80%, Fig. 1F). Application of mineral oil to
the pulp did not induced any significant increases in responses to the noxious stimuli in the NS
neurons in either rats or mice (RM ANOVA, p>0.05). The effect of MO on responses to the
noxious stimuli lasted significantly longer in mice than in rats (Fig.1 E, F; 2-way ANOVA,
p<0.05). Compared to saline (control) treatment, administration of pregabalin significantly
reversed the MO-induced increased responses to the noxious stimuli to baseline levels in both
rats and mice (p<0.05, 2-way ANOVA, Fig. 1 E, F).
Discussion
This is the first report to document that pregabalin is effective in attenuating central sensitization
of functionally identified dorsal horn nociceptive neurons in either spinal or trigeminal
nociceptive systems in an acute rodent inflammatory pain model. Consistent with our previous
findings in rats, we found that application of the inflammatory irritant MO to the rat molar tooth
pulp could induce MDH central sensitization of NS neurons that was reflected as increases in RF
size, spontaneous activity and responses to noxious RF stimuli and a decrease in mechanical
activation threshold [9-12, 22], but we have also shown for the first time that MO application to
the molar pulp also induces MDH central sensitization in mice. The effects of pregabalin that
were documented are also consistent with our recent study in this model of acute inflammatory
pain [28] where we found that pregabalin was effective in a dose-dependent manner in
significantly attenuating the nociceptive sensorimotor behavioural responses and medullary
release of glutamate evoked by the application of MO to the rat tooth pulp. In the current study,
we have shown that administration of pregabalin was effective in significantly reducing all the
parameters of central sensitization tested in the MDH nociceptive neurons in this model as well

43
as in the analogous acute tooth pulp inflammatory pain model in mice. Also, in another recent
study [4], we demonstrated that pregabalin effectively attenuated facial mechanical
hypersensitivity and MDH central sensitization in a rat model of trigeminal neuropathic pain.
Although delineation of the mechanisms and sites of action of pregabalin was not an aim of this
study, it is known that pregabalin is a structural analog of gamma-aminobutyric acid (GABA),
but it is inactive at GABA-A or -B receptors, is not converted into a GABA or a GABA
antagonist, and it does not affect GABA uptake [17]. Indeed, the exact mechanism of action of
pregabalin is not completely clear. One explanation is that by tightly binding to the α2δ protein,
pregabalin reduces the influx of calcium, thereby reducing the release of neurotransmitters,
including glutamate, norepinephrine, and substance P [13, 14, 16].
Systemic effects of pregabalin can significantly inhibit ectopic discharges from injured afferent
neurons, which suggests that the analgesic effect of pregabalin on neuropathic pain is potentially
mediated, at least in part, by a peripheral inhibitory action on the generation of ectopic
discharges caused by nerve injury [5]. In our acute inflammatory model, we have shown that
application of MO to the tooth pulp can induce spontaneous activity in some NS MDH neurons,
which could conceivably be due to ectopic afferent discharges, and that administration of
pregabalin completely prevented the MO-induced spontaneous responses. Therefore, it is
possible that the effects of pregabalin observed in the present study were at least partly mediated
by a reduction of ectopic afferent activity and direct reduction or elimination of the nociceptive
afferent input to the MDH, thereby reducing the central sensitization of the MDH neurons. Our
findings that pregabalin reduced MO-induced activity of the MDH nociceptive neurons is
consistent with this possibility, although it is likely pregabalin also acts in the central neural
system to produce this effect. There is previous evidence indicating that intrathecal application of
pregabalin reduces the enhanced noxious stimulus-induced spinal release of glutamate seen in
neuropathic rats [26], supporting the possibility of a central action of pregabalin. In addition,
identification of pregabalin binding sites in mice has revealed that the highest concentration
occurred in the cortex, olfactory bulb, hypothalamus, amygdala, hippocampus, cerebellum and
dorsal horn of the spinal cord [3], further supporting a central action of pregabalin.
There are reports indicating that pregabalin at varying doses reduces nociceptive behavioural
responses in the spinal nociceptive system in rat pain models [2, 18, 21, 26]. We have
previously demonstrated a dose-dependent effect of pregabalin in reversing the facial mechanical

44
allodynia and MDH central sensitization present at postoperative day 7 following partial infra-
orbital nerve injury [4]. A notable finding of the present study was that pregabalin had no effect
on the baseline neuronal properties of the MDH nociceptive neurons. This suggests that
pregabalin can attenuate central sensitization in MDH nociceptive neurons without affecting
normal nociceptive processing, consistent with our previous findings in the trigeminal
neuropathic pain model [4].
Detailed clinical studies of the potential use of pregabalin in orofacial pain states are limited, but
there are reports of clinical analgesic effects of pregabalin for acute pain following third molar
extraction [19] and for paresthesia following inferior alveolar nerve damage [27], and that
pregabalin can be successfully used to manage glossopharyngeal [23], lacrimal [29] and
postherpetic [1] neuralgias as well as post-traumatic facial pain due to a peripheral nerve injury
[35]. Also, there is an expert recommendation for management of orofacial neuropathic pain
conditions by pregabalin [38]. However, no randomized clinical trials have been conducted to
address the effects of pregabalin in orofacial neuropathic or acute inflammatory conditions.
Nevertheless, in animal models, we have shown it is effective in a trigeminal neuropathic pain
model as noted above [4], and that pregabalin can also prevent both the medullary release of
glutamate and prolonged jaw and tongue muscle activity produced by application of MO to the
pulp [28]. Furthermore, our current findings demonstrate that pregabalin effectively attenuates
nociceptive neuronal hyperexcitability reflecting central sensitization in the rodent model of
acute inflammatory tooth pain, and provides pre-clinical data supporting its potential clinical use
in the treatment of orofacial inflammatory pain states as well as neuropathic pain conditions in
humans; further clinically based studies are needed to test its usefulness in such inflammatory
pain states.
Acknowledgements
The authors acknowledge Ms. Susan Carter and Mr. Yathavan Varathan for their technical
assistance. This work was supported by grants from Pfizer Canada and US National Institutes of
Health DE04786. We thank Pfizer Canada for providing the pregabalin used in this study. BJS is
the holder of a Canada Research Chair.

45
References
[1] A. Achar, P.P. Chakraborty, S. Bisai, A. Biswas, T. Guharay, Comparative study of
clinical efficacy of amitriptyline and pregabalin in postherpetic neuralgia, Acta
dermatovenerologica Croatica 20 (2012) 89-94.
[2] K. Bannister, S. Sikandar, C.S. Bauer, A.C. Dolphin, F. Porreca, A.H. Dickenson,
Pregabalin suppresses spinal neuronal hyperexcitability and visceral hypersensitivity in
the absence of peripheral pathophysiology, Anesthesiology 115 (2011) 144-152.
[3] F. Bian, Z. Li, J. Offord, M.D. Davis, J. McCormick, C.P. Taylor, L.C. Walker, Calcium
channel alpha2-delta type 1 subunit is the major binding protein for pregabalin in
neocortex, hippocampus, amygdala, and spinal cord: an ex vivo autoradiographic study in
alpha2-delta type 1 genetically modified mice, Brain research 1075 (2006) 68-80.
[4] Y. Cao, H. Wang, C.Y. Chiang, J.O. Dostrovsky, B.J. Sessle, Pregabalin suppresses
nociceptive behavior and central sensitization in a rat trigeminal neuropathic pain model,
The journal of pain 14 (2013) 193-204.
[5] S.R. Chen, Z. Xu, H.L. Pan, Stereospecific effect of pregabalin on ectopic afferent
discharges and neuropathic pain induced by sciatic nerve ligation in rats, Anesthesiology
95 (2001) 1473-1479.
[6] P.S. Cherkas, N. Kumar, T.J. Coderre, Y. Cao, H. Wang, C. Chiang, J.O. Dostovsky, B.J.
Sessle, Effects of pregabalin on nociceptive behaviour and evoked glutamate release in a
trigeminal neuropathic pain model. Canadian Association for Neuroscience Meeting,
Vancouver, (2012).
[7] P.S. Cherkas, N. Kumar, M. Miyamoto, T.J. Coderre, J.O. Dostrovsky, B.J. Sessle,
Pregabalin blocks central sensitization and glutamate release in rodent medullary dorsal
horn associated with acute tooth pulp. 13th World Congress on Pain, Montreal, (2010).
[8] P.S. Cherkas, V. Varathan, M. Miyamoto, J.O. Dostrovsky, B.J. Sessle, Pregabalin
attenuates nociceptive behavior induced by trigeminal nerve injury in rodents. Canadian
Pain Society Meeting, Niagara Falls, (2011).
[9] C.Y. Chiang, Z. Li, J.O. Dostrovsky, B.J. Sessle, Central sensitization in medullary
dorsal horn involves gap junctions and hemichannels, Neuroreport 21 (2010) 233-237.
[10] C.Y. Chiang, S.J. Park, C.L. Kwan, J.W. Hu, B.J. Sessle, NMDA receptor mechanisms
contribute to neuroplasticity induced in caudalis nociceptive neurons by tooth pulp
stimulation, Journal of neurophysiology 80 (1998) 2621-2631.
[11] C.Y. Chiang, J. Wang, Y.F. Xie, S. Zhang, J.W. Hu, J.O. Dostrovsky, B.J. Sessle,
Astroglial glutamate-glutamine shuttle is involved in central sensitization of nociceptive
neurons in rat medullary dorsal horn, The Journal of neuroscience 27 (2007) 9068-9076.
[12] C.Y. Chiang, S. Zhang, Y.F. Xie, J.W. Hu, J.O. Dostrovsky, M.W. Salter, B.J. Sessle,
Endogenous ATP involvement in mustard-oil-induced central sensitization in trigeminal
subnucleus caudalis (medullary dorsal horn), Journal of neurophysiology 94 (2005) 1751-
1760.
[13] M.O. Cunningham, G.L. Woodhall, S.E. Thompson, D.J. Dooley, R.S. Jones, Dual
effects of gabapentin and pregabalin on glutamate release at rat entorhinal synapses in
vitro, The European journal of neuroscience 20 (2004) 1566-1576.

46
[14] D.J. Dooley, C.M. Donovan, T.A. Pugsley, Stimulus-dependent modulation of
[(3)H]norepinephrine release from rat neocortical slices by gabapentin and pregabalin,
The Journal of pharmacology and experimental therapeutics 295 (2000) 1086-1093.
[15] R. Dubner, Plasticity in central nociceptive pathways. H. Merskey, J.D. Loeser, R.
Dubner (Eds.), The Paths of Pain 1975–2005, IASP Press, Seattle (2005) 101–115
[16] K. Fink, D.J. Dooley, W.P. Meder, N. Suman-Chauhan, S. Duffy, H. Clusmann, M.
Gothert, Inhibition of neuronal Ca(2+) influx by gabapentin and pregabalin in the human
neocortex, Neuropharmacology 42 (2002) 229-236.
[17] J.E. Frampton, R.H. Foster, Pregabalin: in the treatment of generalised anxiety disorder,
CNS drugs 20 (2006) 685-693; discussion 694-685.
[18] J. Hendrich, C.S. Bauer, A.C. Dolphin, Chronic pregabalin inhibits synaptic transmission
between rat dorsal root ganglion and dorsal horn neurons in culture, Channels 6 (2012)
124-132.
[19] C.M. Hill, M. Balkenohl, D.W. Thomas, R. Walker, H. Mathe, G. Murray, Pregabalin in
patients with postoperative dental pain, European journal of pain 5 (2001) 119-124.
[20] J.W. Hu, Response properties of nociceptive and non-nociceptive neurons in the rat's
trigeminal subnucleus caudalis (medullary dorsal horn) related to cutaneous and deep
craniofacial afferent stimulation and modulation by diffuse noxious inhibitory controls,
Pain 41 (1990) 331-345.
[21] R.W. Hurley, D. Chatterjea, M. Rose Feng, C.P. Taylor, D.L. Hammond, Gabapentin and
pregabalin can interact synergistically with naproxen to produce antihyperalgesia,
Anesthesiology 97 (2002) 1263-1273.
[22] K. Itoh, C.Y. Chiang, Z. Li, J.C. Lee, J.O. Dostrovsky, B.J. Sessle, Central sensitization
of nociceptive neurons in rat medullary dorsal horn involves purinergic P2X7 receptors,
Neuroscience 192 (2011) 721-731.
[23] J.M. Kitchener, Glossopharyngeal neuralgia responding to pregabalin, Headache 46
(2006) 1307-1308.
[24] K.P. Kumar, D.K. Kulkarni, I. Gurajala, R. Gopinath, Pregabalin versus tramadol for
postoperative pain management in patients undergoing lumbar laminectomy: a
randomized, double-blinded, placebo-controlled study, Journal of pain research 6 (2013)
471-478.
[25] N. Kumar, P.S. Cherkas, V. Varathan, M. Miyamoto, C.Y. Chiang, J.O. Dostrovsky, B.J.
Sessle, T.J. Coderre, Systemic pregabalin attenuates facial hypersensitivity and noxious
stimulus-evoked release of glutamate in medullary dorsal horn in a rodent model of
trigeminal neuropathic pain, Neurochemistry international 62 (2013) 831-835.
[26] N. Kumar, A. Laferriere, J.S. Yu, A. Leavitt, T.J. Coderre, Evidence that pregabalin
reduces neuropathic pain by inhibiting the spinal release of glutamate, Journal of
neurochemistry 113 (2010) 552-561.
[27] J. Lopez-Lopez, A. Estrugo-Devesa, E. Jane-Salas, J.J. Segura-Egea, Inferior alveolar
nerve injury resulting from overextension of an endodontic sealer: non-surgical
management using the GABA analogue pregabalin, International endodontic journal 45
(2012) 98-104.
[28] N. Narita, N. Kumar, P.S. Cherkas, C.Y. Chiang, J.O. Dostrovsky, T.J. Coderre, B.J.
Sessle, Systemic pregabalin attenuates sensorimotor responses and medullary glutamate
release in inflammatory tooth pain model, Neuroscience 218 (2012) 359-366.

47
[29] J.A. Pareja, M.L. Cuadrado, Lacrimal neuralgia: So far, a missing cranial neuralgia,
Cephalalgia (2013) doi: 10.1177/0333102413488000.
[30] J.E. Quintero, D.J. Dooley, F. Pomerleau, P. Huettl, G.A. Gerhardt, Amperometric
measurement of glutamate release modulation by gabapentin and pregabalin in rat
neocortical slices: role of voltage-sensitive Ca2+ alpha2delta-1 subunit, The Journal of
pharmacology and experimental therapeutics 338 (2011) 240-245.
[31] R. Sabatowski, R. Galvez, D.A. Cherry, F. Jacquot, E. Vincent, P. Maisonobe, M.
Versavel, G. Study, Pregabalin reduces pain and improves sleep and mood disturbances
in patients with post-herpetic neuralgia: results of a randomised, placebo-controlled
clinical trial, Pain 109 (2004) 26-35.
[32] B. Sessle, Orofacial pain. H. Merskey, J.D. Loeser, R. Dubner (Eds.), The Paths of Pain
1975–2005, IASP Press, Seattle (2005) 131–150.
[33] B.J. Sessle, Acute and chronic craniofacial pain: brainstem mechanisms of nociceptive
transmission and neuroplasticity, and their clinical correlates., Crit Rev Oral Biol Med 11
(2000) 57-91.
[34] B.J. Sessle, Peripheral and central mechanisms of orofacial inflammatory pain,
International review of neurobiology 97 (2011) 179-206.
[35] R.K. Singh, V.P. Sinha, U.S. Pal, S.C. Yadav, M.K. Singh, Pregabalin in post traumatic
neuropathic pain: Case studies, National journal of maxillofacial surgery 3 (2012) 91-95.
[36] T.G. Tzellos, K.A. Toulis, D.G. Goulis, G. Papazisis, V.A. Zampeli, A. Vakfari, D.
Kouvelas, Gabapentin and pregabalin in the treatment of fibromyalgia: a systematic
review and a meta-analysis, Journal of clinical pharmacy and therapeutics 35 (2010) 639-
656.
[37] T. Yokoyama, Y. Maeda, K.M. Audette, K.A. Sluka, Pregabalin reduces muscle and
cutaneous hyperalgesia in two models of chronic muscle pain in rats, J Pain 8 (2007) 422-
429.
[38] J.M. Zakrzewska, Medical management of trigeminal neuropathic pains, Expert opinion
on pharmacotherapy 11 (2010) 1239-1254.

48
Legend to Figures
Figure 1. Blockade by pregabalin of the MO-induced MDH central sensitization in rats (A, C, E)
and mice (B, D, F).
(A, B) Changes in NS neuronal RF size. Application of MO to the tooth pulp in both rats (A) and
mice (B) produced significant increases in pinch RF size (p<0.05, RM ANOVA; * p<0.05,
Dunnett's test, n=6). Administration of pregabalin (but not saline) reversed the MO-induced
increases in neuronal RF size (p>0.05, RM ANOVA, n=6). (C, D) Changes in mechanical
activation threshold of NS neurons. Application of MO to the tooth pulp in both rats (C) and
mice (D) produced significant decreases in mechanical activation threshold (p<0.05, RM
ANOVA; * p<0.05, Dunnett's test, n=6). Administration of pregabalin (but not saline)
significantly blocked the MO-induced decrease in threshold and increases in responses (p>0.5,
RM ANOVA, n=6). (E, F) Changes in neuronal responses to noxious stimuli. Application of MO
to the tooth pulp in both rats (E) and mice (F) produced significant increases in neuronal
responses to noxious stimuli (p<0.05, RM ANOVA; * p<0.05, Dunnett's test, n=6).
Administration of pregabalin (but not saline) significantly blocked the MO-induced increases in
responses (p>0.5, RM ANOVA, n=6). Note that the baseline RF size, activation threshold and
responses to noxious stimuli did not change significantly following pregabalin administration in
both rats and mice (p>0.5, RM ANOVA). Post-hoc analysis indicated that there were significant
differences in values at post-MO time-points between the saline (control) and pregabalin
administrations (# p<0.05, 2-way ANOVA followed by Dunnett test, n=6). (G, H) The recording
sites of the NS neurons were histologically verified in the deep laminae of MDH in rats (G) and
mice (H); Sp 5, trigeminal spinal tract.

49

50
Chapter 4 Discussion
This is the first documentation of long-lasting facial nociceptive behaviour associated with
central sensitization in functionally identified trigeminal nociceptive neurons in the MDH
following injury of the mouse ION. The study also demonstrates for the first time that
administration of pregabalin (but not saline as vehicle control) reduces the long-lasting
nociceptive behaviour and central sensitization at days 7, 21 and 49 following ION injury. This
is also the first report to document that the anticonvulsant drug pregabalin is effective in
attenuating central sensitization of functionally identified MDH nociceptive neurons in the
trigeminal nociceptive system in an acute rodent inflammatory pain model. These findings
provide pre-clinical evidence suggesting that pregabalin may be effective in treating orofacial
acute and chronic pain conditions, which is of significant clinical importance as noted below.
4.1 Central sensitization and nociceptive behaviour
The present study is a continuation of an ongoing series of experiments conducted in our
laboratory that has explored the effects of the inflammatory irritant MO on nociceptive processes
in the rodent MDH. Consistent with our previous findings in rats, we found that application of
MO to the rat molar tooth pulp could induce MDH central sensitization of NS neurons that was
reflected as increases in neuronal RF size, spontaneous activity and responses to noxious RF
stimuli and a decrease in mechanical activation threshold (84, 92, 96, 99, 110). In addition, we
have also shown for the first time that MO application to the molar pulp similarly induces MDH
central sensitization in mice. Central sensitization has been implicated as an important process in
acute and chronic orofacial pain conditions following injury or inflammation of peripheral
tissues (12, 71, 73, 74).
The current findings from the neuropathic pain model experiments are also consistent with
previous studies of our laboratory that have documented the occurrence of central sensitization
of NS neurons following partial ION injury in rats (70). In this study we found that damage of
the ION induces long-lasting bilateral facial mechanical allodynia and central sensitization in
functionally identified NS nociceptive neurons in the medullary dorsal horn in mice.

51
4.2 Nociceptive behaviour: contralateral effects
In the present study we demonstrated that injury of the mouse ION produces long-lasting facial
nociceptive behaviour evoked not only by stimulation of the ipsilateral mandibular skin but also
by stimulation of the contralateral maxillary and mandibular skin. Contralateral as well as
ipsilateral nociceptive behaviour experienced as hyperalgesia and allodynia may reflect
neuroplastic changes manifested as central sensitization of nociceptive neurons in MDH and
other central nervous system areas (12, 71, 73, 74). The scope of the current work was not
intended to determine the mechanisms underlying these neuroplastic changes but one study has
suggested that the development of contralateral orofacial hyperalgesia in a chronic orofacial pain
model is mediated through descending facilitatory influences of the rostral ventromedial medulla
on the spinal trigeminal nucleus (111). Another study has demonstrated that trigeminal subnuclei
interpolaris and caudalis play a unique role in the development of contralateral allodynia in the
deep orofacial tissues and that it involves glial cells and inflammatory cytokines (112). These
mechanisms would explain the contralateral behaviour in our present experiments where
unilateral injury to ION produced long-lasting facial nociceptive behaviour also on the
contralateral side, but further studies are required to identify the mechanisms involved in the
development and maintenance of contralateral hyperalgesia and allodynia in the current IONX
model.
4.3 Effects of pregabalin
There are reports indicating that pregabalin at varying doses reduces nociceptive behavioural
responses in the spinal nociceptive system in rat pain models (66-69). The effects of pregabalin
that were documented in the present study are also consistent with findings in our recent study in
an acute inflammatory pain model (65) where we found that pregabalin was effective in a dose-
dependent manner in significantly attenuating the nociceptive sensorimotor behavioural
responses and medullary release of glutamate evoked by the application of MO to the rat tooth
pulp. In the current study, we have shown that administration of pregabalin was effective in
significantly reducing the effect of central sensitization in MDH nociceptive neurons in this
model in rats as well as in the acute tooth pulp inflammatory pain model in mice.

52
A notable finding of the present study was that pregabalin had no effect on the baseline neuronal
properties of the MDH nociceptive neurons. This suggests that pregabalin can attenuate central
sensitization in MDH nociceptive neurons without affecting normal nociceptive processing,
consistent with our previous findings in the trigeminal neuropathic pain model in rats (70). In
that study we also demonstrated a dose-dependent effect of pregabalin in reversing the facial
mechanical allodynia and MDH central sensitization at an early post-operative stage (day 7). The
present data are consistent with this finding but the study in mice has additionally revealed that
pregabalin may be effective at longer post-operative times when the nociceptive behaviour and
central sensitization have become well maintained.
The site and mode of action of pregabalin are still not fully explained (113) and we did not
address them in this study. Several mechanisms of action have been proposed. One is that
pregabalin binds to the α2δ protein of calcium channels and reduces the influx of calcium. This
action reduces the release of neurotransmitters, such as glutamate, norepinephrine, and substance
P (114-117). Systemic administration of pregabalin can significantly inhibit ectopic discharges
from injured peripheral sensory neurons (64) which could at least partially explain the effect of
pregabalin in the present study. Systemic administration of pregabalin could have reduced or
prevented afferent input to the MDH NS neurons, which is consistent with current findings in the
acute inflammation model and our group’s demonstration that administration of pregabalin
abolished spontaneous activity in NS MDH neurons in a trigeminal neuropathic pain model in
rats (70). In addition, there is a possibility of direct action of pregabalin in the central nervous
system. It has been demonstrated that intrathecal application of pregabalin reduces the enhanced
noxious stimulus-induced spinal and MDH release of glutamate seen in neuropathic rats (69,
118). Also, pregabalin has affinity to its receptors in the cortex, olfactory bulb, hypothalamus,
amygdala, hippocampus, cerebellum and dorsal horn of the spinal cord (119). In addition,
pregabalin may also affect glial cell function since it has been reported to suppress proliferation
of microglia and astrocytes observed in a spinal cord injury model in rats (120). All these
findings strongly suggest that pregabalin could be effective in reducing behavioural allodynia
and central sensitization in MDH NS neurons in the current trigeminal neuropathic pain model in
mice via a central mode of action.

53
4.4 Clinical implications
Our findings that acute inflammation of the tooth pulp and chronic injury of the ION induce
nociceptive behaviour and central sensitization in NS neurons in MDH are of significant clinical
importance. Untreated pulp inflammation or TMDs, injury to the trigeminal nerve during RCT
or apical surgery, periodontal surgery, microsurgery, orthodontic tooth movement, tooth
extraction, implant placement or orthognathic surgery may induce long-lasting chronic pain
states which are frequently very difficult to manage (14, 15, 17, 28, 121). The current findings
provide, at least partial, insights into of mechanisms that take place in the trigeminal nociceptive
system following acute or chronic nerve injury.
The current work has demonstrated that injury of the mouse ION produces long-lasting facial
nociceptive behaviour not only ipsilaterally but also contralaterally. These finding are important
for understanding of certain clinical situations that may manifest contralateral pain spread. For
example, persistent orofacial pain following dental treatment or tooth extraction, implant
placement, persistent pain associated with TMD and other chronic clinical conditions in the
orofacial region may induce contralateral pain spread (122-124). These clinical conditions are
extremely difficult to treat. Understanding the mechanisms of development and maintenance of
contralateral pain spread in trigeminal pain states may provide a significant improvement in
clinical management of such pain states.
Not only has this study produces insights into the mechanisms underlying the development and
maintenance of acute and chronic pain states, but it also suggests a possible management
approach to such conditions. There are numerous reports in the literature demonstrating the use
of pregabalin in management of several chronic pain states involving the spinal somatosensory
system. Pregabalin has been found effective in treating neuropathic conditions associated with
cancer (57, 125), management of fibromyalgia (126) and post-herpetic neuralgia (55, 56). Also
pregabalin has been found effective in management of central neuropathic pain (127), following
spinal cord injury (128) and diabetic peripheral neuropathy (62).There are however only a
limited number of clinical studies of the use of pregabalin in orofacial pain states, but there are
some reports of analgesic effects of pregabalin for acute pain following third molar extraction
(129) and for paresthesia following inferior alveolar nerve damage (130). Some studies have
demonstrated successful use of pregabalin in management of glossopharyngeal (58), lacrimal

54
(131) and post-herpetic (56) neuralgias as well as post-traumatic facial pain due to a peripheral
nerve injury (60). Additionally, there is an expert recommendation for management of orofacial
neuropathic pain conditions by pregabalin (63). Up to now, no randomized clinical trials have
been conducted to address the effects of pregabalin in orofacial neuropathic or acute
inflammatory conditions. The current study provides further pre-clinical data suggesting that
pregabalin may be effective in the management of acute inflammatory tooth pain as well as
trigeminal neuropathic pain conditions, including chronic post-endodontic pain. In animal
models, we have shown that pregabalin is effective in a trigeminal neuropathic pain model as
noted above (70), and that pregabalin can also prevent the medullary release and MDH central
sensitization as well as prolonged jaw and tongue muscle activity produced by application of MO
to the pulp (65). These pre-clinical findings collectively also point to the need for further
clinically based studies to test its effectiveness in acute and chronic orofacial pain states.
4.5 Study strengths and limitations
The novel findings of this study that demonstrate a long-lasting facial nociceptive behaviour and
central sensitization in functionally identified trigeminal nociceptive neurons in the MDH
following injury of the mouse ION is a strength of this work. The demonstration of therapeutic
effects of pregabalin in this chronic model of a long-lasting nociceptive behaviour and central
sensitization is another strength of this work.
The confounding factors that could have altered the observed nociceptive behaviour and central
sensitization in the MDH nociceptive neurons in this study (e.g. exposure to oral operatory
procedures, pain induced by soft tissue trauma, general anaesthesia) were controlled by the use
of a Sham group. The Sham group experienced the general anaesthesia and all the operatory
procedures performed on the IONX group except for the ION transection and showed no
significant changes in the animal behaviour or electrophysiological properties of the NS neurons.
In addition, application of mineral oil to the tooth pulp served as a vehicle control in the acute
inflammatory model. Administration of saline (vehicle control) controlled for the systemic
effects of pregabalin. The experimental vehicle control group demonstrated no significant
changes in the nociceptive behaviour or electrophysiological properties of the NS neurons.
Another strength was the correlation between nociceptive behaviour and neuronal changes in

55
MDH, emphasizing a potential underlying mechanism contributing to nociceptive neuropathic
pain behaviour.
Despite these strengths, this study had some limitations. These include the unknown site of
action of the systemically administered pregabalin and the uncertainty whether the effect was due
to its direct peripheral action or an analgesic effect in the central nervous system, and exactly
what central nervous system sites were affected by pregabalin. This limitation was discussed
previously in sub-section 4.3.
Another limitation of this study was use of general anaesthesia for the MDH study and use of
awake animals for the testing of nociceptive behaviour. General anaesthesia may have an effect
on neuronal properties by blocking normal conductivity and altering membrane permeability.
Despite this, the MDH and behavioural data as well as effects of pregabalin on them were
nonetheless complementary.
An additional limitation of this study that restricts the generalizability of the findings was the use
of male animals in all experiments. This was done deliberately since fluctuations in female sex
hormones may cause variability in nociceptive responses and thus introduce confounding effects
(87). Studies in female animals analogous to those conducted in male animals in this project
represent a future research direction that may provide insights into effects of pregabalin in
females and into possible sex difference in these effects.
Another limitation is that this study evaluated the effects of inflammation or trigeminal injury
only on nociceptive neurons in the MDH and only on NS neurons in the deep MDH laminae.
Therefore, it cannot provide a complete picture of the central sensitization that occurs at the
higher brain levels (e. g. thalamus, somatosensory cortex, etc.) and whether wide dynamic range
and NS neurons in superficial laminae of the MDH are affected in these models. Evaluation of
the central sensitization in these brain sites is necessary to provide a more comprehensive picture
of the neuroplastic changes that take place in acute or chronic orofacial pain states and the
potential effects of pregabalin at these sites.

56
4.6 Future research directions
The novel findings in this study provide valuable information on the potential clinical
applications of pregabalin in pathological pain states. However, more studies are needed to
understand the mechanism of action and whether the effects of pregabalin are due to central or
peripheral actions, or both, in acute and chronic orofacial pain states, and as noted above, and
whether there are sex differences in its effects.
Another approach in management of acute and chronic orofacial pain is identification of new
drugs and their potential sites of action. Other future directions could test the effects of agents
that might potentially influence nociceptive behaviour and MDH central sensitization in the
acute and chronic orofacial pain models. For example, acetaminophen is a non-steroidal anti-
inflammatory drug that is widely used over-the-counter as analgesic and antipyretic. It could be
used to see if it reduces acute and chronic nociceptive pain behaviour and MDH central
sensitization. Other candidate agents are antibiotics. There are several review articles indicating
that there is no benefit of using antibiotics in acute inflammatory conditions in endodontics in
cases without systemic complications (132, 133), but the emphasis has been on the antimicrobial
properties of the antibiotics. Non-microbial properties of these substances associated with pain
mechanisms have never yet been studied. One of the antibiotics used in endodontics,
minocycline, inhibits microglial function via p38 MAPK and produces analgesia in neuropathic
pain models (134-137). While minocycline has yet to undergo detailed investigation in animal
models of chronic or acute orofacial pain, studies have recently demonstrated that it attenuates
the MDH central sensitization induced in the acute pulp inflammatory and chronic neuropathic
pain models in rats (99, 138) Therefore, studying the effects of minocycline on sensorimotor
behaviour and trigeminal central sensitization in a dose-dependent manner in an acute
inflammatory pain model may be of interest to support its clinical use for treatment of trigeminal
pain, including endodontic pain.
An additional gap in knowledge of trigeminal pain mechanisms is the role of genetic factors.
Humans display highly variable sensitivity to pain, including variable responses in developing a
chronic pain state to identical injuries or pathologies, including those related to endodontic
therapy or trigeminal nerve injury. Also, humans may vary in their response to drugs used to treat

57
their pain conditions, e.g., due to individual differences in absorption and processing of
chemicals. These individual differences in developing pain conditions as well as in responses to
analgesic drug may depend at least in part on individual’s genotype. The current literature on
mouse pain models indicates the importance of the mouse genotype in mediating nociceptive
sensitivity and establishing a predisposition to neuropathic pain following neural injury (139-
142). In preliminary experiments we have found differences between two genetically different
mouse strains (C57BL/6 and A/J) in orofacial nociceptive behaviour and MDH central
sensitization induced by trigeminal nerve injury (143), which suggests genetic factors may be
involved in the predisposition to developing orofacial nociceptive behaviour and MDH central
sensitization. The findings might possibly also account for differences between patients in the
efficacy of current treatment regimens for trigeminal neuropathic pain conditions. Therefore, pre-
clinical studies to identify genetic factors in establishing and maintenance of acute and chronic
pain and the effects of pain-relieving agents are of great importance.

58
Chapter 5 Conclusions
The current work has demonstrated that application of the inflammatory irritant MO to the molar
tooth pulp induces central sensitization in trigeminal MDH neurons in rats and mice. This work
also shows that injury of the mouse ION produces long-lasting facial nociceptive behaviour and
central sensitization in trigeminal MDH neurons in mice. The results of this study also indicate
that administration of pregabalin can block central sensitization in the MDH in a rodent acute
inflammatory pain model as well as attenuate the long-lasting IONX-induced mechanical
allodynia and associated MDH central sensitization. These findings provide insights into
mechanisms that may contribute to acute and chronic orofacial pain states, and also pre-clinical
data supporting the potential clinical use of pregabalin in the treatment of orofacial inflammatory
pain states as well as neuropathic pain conditions in humans.

59
References
1. IASP. Classification of Chronic Pain, Second Edition, IASP Task Force on Taxonomy,
edited by H. Merskey and N. Bogduk, IASP Press, Seattle. 1994.
2. Auvray M, Myin E, Spence C. The sensory-discriminative and affective-motivational
aspects of pain. Neurosci Biobehav Rev 2010;34:214-223.
3. Sessle BJ. Unrelieved pain: a crisis. Pain Res Manag 2011;16:416-420.
4. Boulanger A, Clark AJ, Squire P, Cui E, Horbay GL. Chronic pain in Canada: have we
improved our management of chronic noncancer pain? Pain Res Manag 2007;12:39-47.
5. Moulin DE, Clark AJ, Speechley M, Morley-Forster PK. Chronic pain in Canada--
prevalence, treatment, impact and the role of opioid analgesia. Pain Res Manag 2002;7:179-184.
6. Van de Ven TJ, John Hsia HL. Causes and prevention of chronic postsurgical pain. Curr
Opin Crit Care 2012;18:366-371.
7. Nanos. www.painexplained.ca. In.; 2007/2008.
8. Stewart WF, Ricci JA, Chee E, Hahn SR, Morganstein D. Cost of lost productive work
time among US workers with depression. JAMA 2003;289:3135-3144.
9. Stewart WF, Ricci JA, Chee E, Morganstein D. Lost productive work time costs from
health conditions in the United States: results from the American Productivity Audit. J Occup
Environ Med 2003;45:1234-1246.
10. Stewart WF, Ricci JA, Chee E, Morganstein D, Lipton R. Lost productive time and cost
due to common pain conditions in the US workforce. JAMA 2003;290:2443-2454.
11. Sherman RA, Sherman CJ, Parker L. Chronic phantom and stump pain among American
veterans: results of a survey. Pain 1984;18:83-95.
12. Sessle BJ. Peripheral and central mechanisms of orofacial inflammatory pain. Int Rev
Neurobiol 2011;97:179-206.
13. Keenan AV. Only a small percentage of patients experience persistent pain for more than
6 months after root canal therapy. J Evid Based Dent Pract 2010;10:235-236.
14. Marbach JJ, Hulbrock J, Hohn C, Segal AG. Incidence of phantom tooth pain: an atypical
facial neuralgia. Oral Surg Oral Med Oral Pathol 1982;53:190-193.
15. Nixdorf DR, Moana-Filho EJ, Law AS, McGuire LA, Hodges JS, John MT. Frequency of
nonodontogenic pain after endodontic therapy: a systematic review and meta-analysis. J Endod
2010;36:1494-1498.
16. Nixdorf DR, Moana-Filho EJ, Law AS, McGuire LA, Hodges JS, John MT. Frequency of
persistent tooth pain after root canal therapy: a systematic review and meta-analysis. J Endod
2010;36:224-230.
17. Polycarpou N, Ng YL, Canavan D, Moles DR, Gulabivala K. Prevalence of persistent
pain after endodontic treatment and factors affecting its occurrence in cases with complete
radiographic healing. Int Endod J 2005;38:169-178.
18. Zakrzewska JM, McMillan R. Trigeminal neuralgia: the diagnosis and management of
this excruciating and poorly understood facial pain. Postgrad Med J 2011;87:410-416.
19. Fornasari D. Pain mechanisms in patients with chronic pain. Clin Drug Investig 2012;32
Suppl 1:45-52.
20. Petho G, Reeh PW. Sensory and signaling mechanisms of bradykinin, eicosanoids,
platelet-activating factor, and nitric oxide in peripheral nociceptors. Physiol Rev 2012;92:1699-
1775.

60
21. Sacerdote P, Levrini L. Peripheral mechanisms of dental pain: the role of substance P.
Mediators Inflamm 2012;2012:951920.
22. Baron R. Peripheral neuropathic pain: from mechanisms to symptoms. Clin J Pain
2000;16:S12-20.
23. Ellis A, Bennett DL. Neuroinflammation and the generation of neuropathic pain. Br J
Anaesth 2013;111:26-37.
24. Stemkowski PL, Smith PA. Sensory neurons, ion channels, inflammation and the onset of
neuropathic pain. Can J Neurol Sci 2012;39:416-435.
25. Woda A, Tubert-Jeannin S, Bouhassira D, Attal N, Fleiter B, Goulet JP, et al. Towards a
new taxonomy of idiopathic orofacial pain. Pain 2005;116:396-406.
26. McNeill C, Dubner R, Woda A. What is Pain and How Do We Classify Orofacial Pain
In: Orofacial Pain From Basic Science to Clinical Management: The Transfer of Knowledge in
Pain Research to Education 2008.
27. Lipton JA, Ship JA, Larach-Robinson D. Estimated prevalence and distribution of
reported orofacial pain in the United States. J Am Dent Assoc 1993;124:115-121.
28. Campbell RL, Parks KW, Dodds RN. Chronic facial pain associated with endodontic
therapy. Oral Surg Oral Med Oral Pathol 1990;69:287-290.
29. LeResche L, Mancl LA, Drangsholt MT, Huang G, Von Korff M. Predictors of onset of
facial pain and temporomandibular disorders in early adolescence. Pain 2007;129:269-278.
30. Nilsson IM, List T, Drangsholt M. Prevalence of temporomandibular pain and subsequent
dental treatment in Swedish adolescents. J Orofac Pain 2005;19:144-150.
31. Wober-Bingol C. Epidemiology of migraine and headache in children and adolescents.
Curr Pain Headache Rep 2013;17:341.
32. Merikangas KR. Contributions of epidemiology to our understanding of migraine.
Headache 2013;53:230-246.
33. Katusic S, Beard CM, Bergstralh E, Kurland LT. Incidence and clinical features of
trigeminal neuralgia, Rochester, Minnesota, 1945-1984. Ann Neurol 1990;27:89-95.
34. Katusic S, Williams DB, Beard CM, Bergstralh EJ, Kurland LT. Epidemiology and
clinical features of idiopathic trigeminal neuralgia and glossopharyngeal neuralgia: similarities
and differences, Rochester, Minnesota, 1945-1984. Neuroepidemiology 1991;10:276-281.
35. Baad-Hansen L. Atypical odontalgia - pathophysiology and clinical management. J Oral
Rehabil 2008;35:1-11.
36. Baad-Hansen L, Pigg M, Ivanovic SE, Faris H, List T, Drangsholt M, et al. Intraoral
somatosensory abnormalities in patients with atypical odontalgia--a controlled multicenter
quantitative sensory testing study. Pain 2013;154:1287-1294.
37. Greene CS, Murray GM. Atypical odontalgia: an oral neuropathic pain phenomenon. J
Am Dent Assoc 2011;142:1031-1032.
38. List T, Leijon G, Svensson P. Somatosensory abnormalities in atypical odontalgia: A
case-control study. Pain 2008;139:333-341.
39. Zakrzewska JM. Differential diagnosis of facial pain and guidelines for management. Br
J Anaesth 2013;111:95-104.
40. Glass EG, Glaros AG, McGlynn FD. Myofascial pain dysfunction: treatments used by
ADA members. Cranio 1993;11:25-29.
41. Gillam D, Chesters R, Attrill D, Brunton P, Slater M, Strand P, et al. Dentine
hypersensitivity--guidelines for the management of a common oral health problem. Dent Update
2013;40:514-516, 518-520, 523-514.

61
42. Zero DT, Zandona AF, Vail MM, Spolnik KJ. Dental caries and pulpal disease. Dent Clin
North Am 2011;55:29-46.
43. Taba M, Jr., Souza SL, Mariguela VC. Periodontal disease: a genetic perspective. Braz
Oral Res 2012;26 Suppl 1:32-38.
44. Siqueira JF, Jr., Rocas IN. Microbiology and treatment of acute apical abscesses. Clin
Microbiol Rev 2013;26:255-273.
45. Feine JS, Lund JP. An assessment of the efficacy of physical therapy and physical
modalities for the control of chronic musculoskeletal pain. Pain 1997;71:5-23.
46. Feine JS, Widmer CG, Lund JP. Physical therapy: a critique. Oral Surg Oral Med Oral
Pathol Oral Radiol Endod 1997;83:123-127.
47. de Toledo EG, Jr., Silva DP, de Toledo JA, Salgado IO. The interrelationship between
dentistry and physiotherapy in the treatment of temporomandibular disorders. J Contemp Dent
Pract 2012;13:579-583.
48. Ahn CB, Lee SJ, Lee JC, Fossion JP, Sant'Ana A. A clinical pilot study comparing
traditional acupuncture to combined acupuncture for treating headache, trigeminal neuralgia and
retro-auricular pain in facial palsy. J Acupunct Meridian Stud 2011;4:29-43.
49. Aggarwal VR, Tickle M, Javidi H, Peters S. Reviewing the evidence: can cognitive
behavioral therapy improve outcomes for patients with chronic orofacial pain? J Orofac Pain
2010;24:163-171.
50. Vella-Brincat J, Macleod AD. Adverse effects of opioids on the central nervous systems
of palliative care patients. J Pain Palliat Care Pharmacother 2007;21:15-25.
51. Botting RM. Inhibitors of cyclooxygenases: mechanisms, selectivity and uses. J Physiol
Pharmacol 2006;57 Suppl 5:113-124.
52. Blondell RD, Azadfard M, Wisniewski AM. Pharmacologic therapy for acute pain. Am
Fam Physician 2013;87:766-772.
53. Williams JT, Ingram SL, Henderson G, Chavkin C, von Zastrow M, Schulz S, et al.
Regulation of mu-opioid receptors: desensitization, phosphorylation, internalization, and
tolerance. Pharmacol Rev 2013;65:223-254.
54. Cohen SP, Abdi S. New developments in the use of tricyclic antidepressants for the
management of pain. Curr Opin Anaesthesiol 2001;14:505-511.
55. Gan EY, Tian EA, Tey HL. Management of herpes zoster and post-herpetic neuralgia.
Am J Clin Dermatol 2013;14:77-85.
56. Achar A, Chakraborty PP, Bisai S, Biswas A, Guharay T. Comparative study of clinical
efficacy of amitriptyline and pregabalin in postherpetic neuralgia. Acta Dermatovenerol Croat
2012;20:89-94.
57. Fallon MT. Neuropathic pain in cancer. Br J Anaesth 2013;111:105-111.
58. Kitchener JM. Glossopharyngeal neuralgia responding to pregabalin. Headache
2006;46:1307-1308.
59. Sabatowski R, Galvez R, Cherry DA, Jacquot F, Vincent E, Maisonobe P, et al.
Pregabalin reduces pain and improves sleep and mood disturbances in patients with post-herpetic
neuralgia: results of a randomised, placebo-controlled clinical trial. Pain 2004;109:26-35.
60. Singh RK, Sinha VP, Pal US, Yadav SC, Singh MK. Pregabalin in post traumatic
neuropathic pain: Case studies. Natl J Maxillofac Surg 2012;3:91-95.
61. Srivastava A, Shah S, Maseeh A, Vyasa B, Balaji M, Bhatter S, et al. A clinical study to
compare the efficacy and safety of pregabalin sustained release formulation with pregabalin

62
immediate release formulation in patients of diabetic peripheral neuropathic pain. Indian J
Endocrinol Metab 2012;16:S477-479.
62. Tesfaye S, Selvarajah D. Advances in the epidemiology, pathogenesis and management
of diabetic peripheral neuropathy. Diabetes Metab Res Rev 2012;28 Suppl 1:8-14.
63. Zakrzewska JM. Medical management of trigeminal neuropathic pains. Expert Opin
Pharmacother 2010;11:1239-1254.
64. Chen SR, Xu Z, Pan HL. Stereospecific effect of pregabalin on ectopic afferent
discharges and neuropathic pain induced by sciatic nerve ligation in rats. Anesthesiology
2001;95:1473-1479.
65. Narita N, Kumar N, Cherkas PS, Chiang CY, Dostrovsky JO, Coderre TJ, et al. Systemic
pregabalin attenuates sensorimotor responses and medullary glutamate release in inflammatory
tooth pain model. Neuroscience 2012;218:359-366.
66. Hurley RW, Chatterjea D, Rose Feng M, Taylor CP, Hammond DL. Gabapentin and
pregabalin can interact synergistically with naproxen to produce antihyperalgesia.
Anesthesiology 2002;97:1263-1273.
67. Hendrich J, Bauer CS, Dolphin AC. Chronic pregabalin inhibits synaptic transmission
between rat dorsal root ganglion and dorsal horn neurons in culture. Channels (Austin)
2012;6:124-132.
68. Bannister K, Sikandar S, Bauer CS, Dolphin AC, Porreca F, Dickenson AH. Pregabalin
suppresses spinal neuronal hyperexcitability and visceral hypersensitivity in the absence of
peripheral pathophysiology. Anesthesiology 2011;115:144-152.
69. Kumar N, Laferriere A, Yu JS, Leavitt A, Coderre TJ. Evidence that pregabalin reduces
neuropathic pain by inhibiting the spinal release of glutamate. J Neurochem 2010;113:552-561.
70. Cao Y, Wang H, Chiang CY, Dostrovsky JO, Sessle BJ. Pregabalin suppresses
nociceptive behavior and central sensitization in a rat trigeminal neuropathic pain model. J Pain
2013;14:193-204.
71. Sessle BJ. Peripheral and central mechanisms of orofacial pain and their clinical
correlates. Minerva Anestesiol 2005;71:117-136.
72. Sessle BJ. Orofacial pain. H. Merskey, J.D. Loeser, R. Dubner (Eds.), The Paths of Pain
1975–2005, IASP Press, Seattle (2005), pp. 131–150. 2005.
73. Dubner R, Ren K. Brainstem mechanisms of persistent pain following injury. J Orofac
Pain 2004;18:299-305.
74. Salter MW. Cellular neuroplasticity mechanisms mediating pain persistence. J Orofac
Pain 2004;18:318-324.
75. Cherkas PS, Huang TY, Pannicke T, Tal M, Reichenbach A, Hanani M. The effects of
axotomy on neurons and satellite glial cells in mouse trigeminal ganglion. Pain 2004;110:290-
298.
76. Kushnir R, Cherkas PS, Hanani M. Peripheral inflammation upregulates P2X receptor
expression in satellite glial cells of mouse trigeminal ganglia: a calcium imaging study.
Neuropharmacology 2011;61:739-746.
77. Warwick RA, Hanani M. The contribution of satellite glial cells to chemotherapy-
induced neuropathic pain. Eur J Pain 2013;17:571-580.
78. Hanani M. Satellite glial cells: more than just 'rings around the neuron'. Neuron Glia Biol
2010;6:1-2.
79. Chiang CY, Dostrovsky JO, Iwata K, Sessle BJ. Role of glia in orofacial pain.
Neuroscientist 2011;17:303-320.

63
80. Eriksson J, Jablonski A, Persson AK, Hao JXK, P.F, Wiesenfeld-Hallin Z, Xu XJ, et al.
Behavioral changes and trigeminal ganglion sodium channel regulation in an orofacial
neuropathic pain model. Pain 2005;119:82-94.
81. Nakagawa K, Takeda M, Tsuboi Y, Kondo M, Kitagawa J, Matsumoto S, et al. Alteration
of primary afferent activity following inferior alveolar nerve transection in rats. Mol Pain
2010;6:9.
82. Takeda M, Takahashi M, Matsumoto S. Contribution of the activation of satellite glia in
sensory ganglia to pathological pain. Neurosci Biobehav Rev 2009;33:784-792.
83. Iwata K, Miyachi S, Imanishi M, Tsuboi Y, Kitagawa J, Teramoto K, et al. Ascending
multisynaptic pathways from the trigeminal ganglion to the anterior cingulate cortex. Exp Neurol
2011;227:69-78.
84. Chiang CY, Park SJ, Kwan CL, Hu JW, Sessle BJ. NMDA receptor mechanisms
contribute to neuroplasticity induced in caudalis nociceptive neurons by tooth pulp stimulation. J
Neurophysiol 1998;80:2621-2631.
85. Sessle BJ. Acute and chronic craniofacial pain: brainstem mechanisms of nociceptive
transmission and neuroplasticity, and their clinical correlates. Crit Rev Oral Biol Med
2000;11:57-91.
86. Latremoliere A, Woolf CJ. Central sensitization: a generator of pain hypersensitivity by
central neural plasticity. J Pain 2009;10:895-926.
87. Greenspan JD, Craft RM, LeResche L, Arendt-Nielsen L, Berkley KJ, Fillingim RB, et
al.Studying sex and gender differences in pain and analgesia: a consensus report. Pain 2007; 132:
S26-45.
88. Sessle BJ. Acute and chronic craniofacial pain: brainstem mechanisms of nociceptive
transmission and neuroplasticity, and their clinical correlates. Crit Rev Oral Biol Med
2000;11:57-91.
89. Zhang S, Chiang CY, Xie YF, Park SJ, Lu Y, Hu JW, et al. Central sensitization in
thalamic nociceptive neurons induced by mustard oil application to rat molar tooth pulp.
Neuroscience 2006;142:833-842.
90. Park SJ, Zhang S, Chiang CY, Hu JW, Dostrovsky JO, Sessle BJ. Central sensitization
induced in thalamic nociceptive neurons by tooth pulp stimulation is dependent on the functional
integrity of trigeminal brainstem subnucleus caudalis but not subnucleus oralis. Brain Res
2006;1112:134-145.
91. Chiang CY, Li Z, Dostrovsky JO, Hu JW, Sessle BJ. Glutamine uptake contributes to
central sensitization in the medullary dorsal horn. Neuroreport 2008;19:1151-1154.
92. Chiang CY, Zhang S, Xie YF, Hu JW, Dostrovsky JO, Salter MW, et al. Endogenous
ATP involvement in mustard-oil-induced central sensitization in trigeminal subnucleus caudalis
(medullary dorsal horn). J Neurophysiol 2005;94:1751-1760.
93. Hu B, Chiang CY, Hu JW, Dostrovsky JO, Sessle BJ. P2X receptors in trigeminal
subnucleus caudalis modulate central sensitization in trigeminal subnucleus oralis. J
Neurophysiol 2002;88:1614-1624.
94. Tsuboi Y, Iwata K, Dostrovsky JO, Chiang CY, Sessle BJ, Hu JW. Modulation of
astroglial glutamine synthetase activity affects nociceptive behaviour and central sensitization of
medullary dorsal horn nociceptive neurons in a rat model of chronic pulpitis. Eur J Neurosci
2011;34:292-302.

64
95. Miyamoto M, Tsuboi Y, Takamiya K, Huganir RL, Kondo M, Shinoda M, et al.
Involvement of GluR2 and GluR3 subunit C-termini in the trigeminal spinal subnucleus caudalis
and C1-C2 neurons in trigeminal neuropathic pain. Neurosci Lett 2011;491:8-12.
96. Chiang CY, Wang J, Xie YF, Zhang S, Hu JW, Dostrovsky JO, et al. Astroglial
glutamate-glutamine shuttle is involved in central sensitization of nociceptive neurons in rat
medullary dorsal horn. J Neurosci 2007;27:9068-9076.
97. Cherkas PS, Dostrovsky JO, Sessle BJ. Activation of peripheral P2X receptors is
sufficient to induce central sensitization in rat medullary dorsal horn nociceptive neurons.
Neurosci Lett 2012;526:160-163.
98. Holland GR. Experimental trigeminal nerve injury. Crit Rev Oral Biol Med 1996;7:237-
258.
99. Itoh K, Chiang CY, Li Z, Lee JC, Dostrovsky JO, Sessle BJ. Central sensitization of
nociceptive neurons in rat medullary dorsal horn involves purinergic P2X7 receptors.
Neuroscience 2011;192:721-731.
100. Cherkas PS, Kumar N, Coderre TJ, Cao Y, Wang H, Chiang C, et al. Effects of
pregabalin on nociceptive behaviour and evoked glutamate release in a trigeminal neuropathic
pain model. In: Canadian Association for Neuroscience Meeting. Vancouver; 2012.
101. Strassman AM, Burstein R. A new animal model of headache: Ongoing pain vs stimulus-
evoked hypersensitivity. Cephalalgia 2013;33:1073-1074.
102. Vos BP, Strassman AM, Maciewicz RJ. Behavioral evidence of trigeminal neuropathic
pain following chronic constriction injury to the rat's infraorbital nerve. J Neurosci
1994;14:2708-2723.
103. Tsuboi Y, Takeda M, Tanimoto T, Ikeda M, Matsumoto S, Kitagawa J, et al. Alteration
of the second branch of the trigeminal nerve activity following inferior alveolar nerve transection
in rats. Pain 2004;111:323-334.
104. Kitagawa J, Takeda M, Suzuki I, Kadoi J, Tsuboi Y, Honda K, et al. Mechanisms
involved in modulation of trigeminal primary afferent activity in rats with peripheral
mononeuropathy. Eur J Neurosci 2006;24:1976-1986.
105. Ohara PT, Vit JP, Bhargava A, Romero M, Sundberg C, Charles AC, et al. Gliopathic
pain: when satellite glial cells go bad. Neuroscientist 2009;15:450-463.
106. Liu CN, Michaelis M, Amir R, Devor M. Spinal nerve injury enhances subthreshold
membrane potential oscillations in DRG neurons: relation to neuropathic pain. J Neurophysiol
2000;84:205-215.
107. Saito K, Hitomi S, Suzuki I, Masuda Y, Kitagawa J, Tsuboi Y, et al. Modulation of
trigeminal spinal subnucleus caudalis neuronal activity following regeneration of transected
inferior alveolar nerve in rats. J Neurophysiol 2008;99:2251-2263.
108. Chiang CY, Sessle BJ, Dostrovsky JO. Role of astrocytes in pain. Neurochem Res
2012;37:2419-2431.
109. Okada-Ogawa A, Suzuki I, Sessle BJ, Chiang CY, Salter MW, Dostrovsky JO, et al.
Astroglia in medullary dorsal horn (trigeminal spinal subnucleus caudalis) are involved in
trigeminal neuropathic pain mechanisms. J Neurosci 2009;29:11161-11171.
110. Chiang CY, Li Z, Dostrovsky JO, Sessle BJ. Central sensitization in medullary dorsal
horn involves gap junctions and hemichannels. Neuroreport 2010;21:233-237.
111. Chai B, Guo W, Wei F, Dubner R, Ren K. Trigeminal-rostral ventromedial medulla
circuitry is involved in orofacial hyperalgesia contralateral to tissue injury. Mol Pain 2012;8:78.

65
112. Shimizu K, Guo W, Wang H, Zou S, LaGraize SC, Iwata K, et al. Differential
involvement of trigeminal transition zone and laminated subnucleus caudalis in orofacial deep
and cutaneous hyperalgesia: the effects of interleukin-10 and glial inhibitors. Mol Pain
2009;5:75.
113. Bialer M. Why are antiepileptic drugs used for nonepileptic conditions? Epilepsia
2012;53 Suppl 7:26-33.
114. Cunningham MO, Woodhall GL, Thompson SE, Dooley DJ, Jones RS. Dual effects of
gabapentin and pregabalin on glutamate release at rat entorhinal synapses in vitro. Eur J
Neurosci 2004;20:1566-1576.
115. Dooley DJ, Donovan CM, Pugsley TA. Stimulus-dependent modulation of
[(3)H]norepinephrine release from rat neocortical slices by gabapentin and pregabalin. J
Pharmacol Exp Ther 2000;295:1086-1093.
116. Dooley DJ, Mieske CA, Borosky SA. Inhibition of K(+)-evoked glutamate release from
rat neocortical and hippocampal slices by gabapentin. Neurosci Lett 2000;280:107-110.
117. Fink K, Dooley DJ, Meder WP, Suman-Chauhan N, Duffy S, Clusmann H, et al.
Inhibition of neuronal Ca(2+) influx by gabapentin and pregabalin in the human neocortex.
Neuropharmacology 2002;42:229-236.
118. Kumar N, Cherkas PS, Varathan V, Miyamoto M, Chiang CY, Dostrovsky JO, et al.
Systemic pregabalin attenuates facial hypersensitivity and noxious stimulus-evoked release of
glutamate in medullary dorsal horn in a rodent model of trigeminal neuropathic pain. Neurochem
Int 2013;62:831-835.
119. Bian F, Li Z, Offord J, Davis MD, McCormick J, Taylor CP, et al. Calcium channel
alpha2-delta type 1 subunit is the major binding protein for pregabalin in neocortex,
hippocampus, amygdala, and spinal cord: an ex vivo autoradiographic study in alpha2-delta type
1 genetically modified mice. Brain Res 2006;1075:68-80.
120. Ha KY, Carragee E, Cheng I, Kwon SE, Kim YH. Pregabalin as a neuroprotector after
spinal cord injury in rats: biochemical analysis and effect on glial cells. J Korean Med Sci
2011;26:404-411.
121. Benoliel R, Kahn J, Eliav E. Peripheral painful traumatic trigeminal neuropathies. Oral
Dis 2011:doi: 10.1111/j.1601-0825.2011.01883.x.
122. Sessle BJ. Neural mechanisms and pathways in craniofacial pain. Can J Neurol Sci
1999;26 Suppl 3:S7-11.
123. Avivi-Arber L, Lee JC, Sessle BJ. Chapter 9--face sensorimotor cortex neuroplasticity
associated with intraoral alterations. Prog Brain Res 2011;188:135-150.
124. Svensson P, Graven-Nielsen T. Craniofacial muscle pain: review of mechanisms and
clinical manifestations. J Orofac Pain 2001;15:117-145.
125. Laird B, Colvin L, Fallon M. Management of cancer pain: basic principles and
neuropathic cancer pain. Eur J Cancer 2008;44:1078-1082.
126. Calandre EP, Rico-Villademoros F. The role of antipsychotics in the management of
fibromyalgia. CNS Drugs 2012;26:135-153.
127. Scherder EJ, Plooij B. Assessment and management of pain, with particular emphasis on
central neuropathic pain, in moderate to severe dementia. Drugs Aging 2012;29:701-706.
128. Rekand T, Hagen EM, Gronning M. Chronic pain following spinal cord injury. Tidsskr
Nor Laegeforen 2012;132:974-979.
129. Hill CM, Balkenohl M, Thomas DW, Walker R, Mathe H, Murray G. Pregabalin in
patients with postoperative dental pain. Eur J Pain 2001;5:119-124.

66
130. Lopez-Lopez J, Estrugo-Devesa A, Jane-Salas E, Segura-Egea JJ. Inferior alveolar nerve
injury resulting from overextension of an endodontic sealer: non-surgical management using the
GABA analogue pregabalin. Int Endod J 2012;45:98-104.
131. Pareja JA, Cuadrado ML. Lacrimal neuralgia: So far, a missing cranial neuralgia.
Cephalalgia 2013:doi: 10.1177/0333102413488000.
132. Matthews DC, Sutherland S, Basrani B. Emergency management of acute apical
abscesses in the permanent dentition: a systematic review of the literature. J Can Dent Assoc
2003;69:660.
133. Mohammadi Z. Systemic, prophylactic and local applications of antimicrobials in
endodontics: an update review. Int Dent J 2009;59:175-186.
134. Cho IH, Chung YM, Park CK, Park SH, Lee H, Kim D, et al. Systemic administration of
minocycline inhibits formalin-induced inflammatory pain in rat. Brain Res 2006;1072:208-214.
135. Cui Y, Liao XX, Liu W, Guo RX, Wu ZZ, Zhao CM, et al. A novel role of minocycline:
attenuating morphine antinociceptive tolerance by inhibition of p38 MAPK in the activated
spinal microglia. Brain Behav Immun 2008;22:114-123.
136. Ma F, Zhang L, Lyons D, Westlund KN. Orofacial neuropathic pain mouse model
induced by Trigeminal Inflammatory Compression (TIC) of the infraorbital nerve. Mol Brain
2012;5:44.
137. Mostafeezur RM, Zakir HM, Yamada Y, Yamamura K, Iwata K, Sessle BJ, et al. The
effect of minocycline on the masticatory movements following the inferior alveolar nerve
transection in freely moving rats. Mol Pain 2012;8:27.
138. Shibuta K, Suzuki I, Shinoda M, Tsuboi Y, Honda K, Shimizu N, et al. Organization of
hyperactive microglial cells in trigeminal spinal subnucleus caudalis and upper cervical spinal
cord associated with orofacial neuropathic pain. Brain Res 2012;1451:74-86.
139. Mogil JS, Wilson SG, Bon K, Lee SE, Chung K, Raber P, et al. Heritability of
nociception II. 'Types' of nociception revealed by genetic correlation analysis. Pain 1999;80:83-
93.
140. Mogil JS, Wilson SG, Bon K, Lee SE, Chung K, Raber P, et al. Heritability of
nociception I: responses of 11 inbred mouse strains on 12 measures of nociception. Pain
1999;80:67-82.
141. LaCroix-Fralish ML, Mo G, Smith SB, Sotocinal SG, Ritchie J, Austin JS, et al. The
beta3 subunit of the Na+,K+-ATPase mediates variable nociceptive sensitivity in the formalin
test. Pain 2009;144:294-302.
142. Mogil JS, Miermeister F, Seifert F, Strasburg K, Zimmermann K, Reinold H, et al.
Variable sensitivity to noxious heat is mediated by differential expression of the CGRP gene.
Proc Natl Acad Sci U S A 2005;102:12938-12943.
143. Cherkas PS, Varathan V, Miyamoto M, Dostrovsky JO, Sessle BJ. Pregabalin attenuates
nociceptive behavior induced by trigeminal nerve injury in rodents. In: Canadian Pain Society
Meeting. Niagara Falls; 2011.