transport characteristics of the loop of henle - springer · chapter 33 transport characteristics...
TRANSCRIPT

CHAPTER 33
Transport Characteristics of the Loop of Henle
Maurice B. Burg and John L. Stephenson
1. Introduction
The loop of Henle was early identified as a structure essential for the concentration of the urine, since it occurs only in birds and mammals, the two classes that produce urine with osmolality higher than the blood. Subsequent study of the loop of Henle has mainly involved its role in urinary concentrating mechanisms. The latter have been analyzed most satisfactorily in terms of mathematical models that integrate the loop of Henle with the vascular system and other nephron segments, such as the distal tubule and collecting ducts. Prerequisite to considering the specific models, however, is a knowledge of the anatomical relations of the loop of Henle and of the pertinent experimental findings. This information, outlined briefly in the following, is contained in several recent reviews(1-4) to which the reader is referred for further details and extensive bibliography.
Maurice B. Burg and John 1. Stephenson • National Heart, Lung, and Blood Institute, National Institutes of Health, Bethesda, Maryland 20014
2. Anatomy
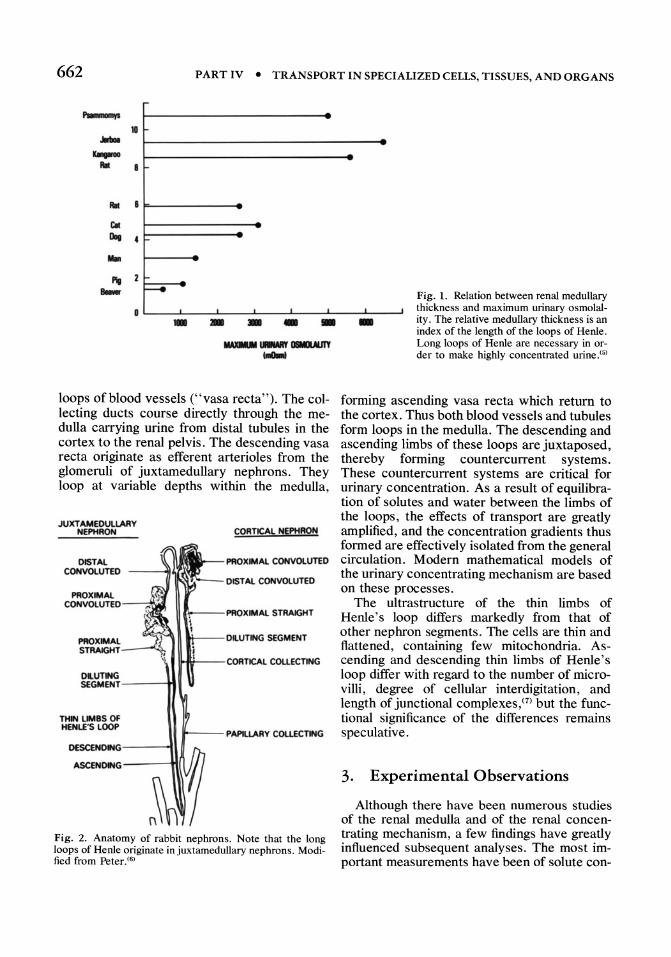
Knowledge of the anatomy of the renal medulla has provided clues important for understanding the urinary concentrating mechanism. The loop of Henle extends into the renal medulla connecting the proximal to the distal tubule. Its length varies from species to species, as well as between different nephrons in the same species. The relative length of the loop of Henle can be gauged from the thickness of the medulla. As shown in Fig. 1,(0) the maximum urinary concentration of a given species is directly related to the relative thickness of its medulla. Evidently, long loops of Henle are required to achieve very high osmolality in the urine. Within a given species there may also be considerable variation in the lengths of Henle's loops between different nephrons. Cortical nephrons have short loops, whereas juxtamedullary nephrons have long loops, as shown for the rabbit in Fig. 2.(6) Although the fraction of the nephrons that have long loops varies from species to species, this does not correlate with maximum urinary concentration.(5)
Within the renal medulla, in addition to the loop of Henle, are located collecting ducts and
661
T. E. Andreoli et al. (eds.), Physiology of Membrane Disorders© Plenum Publishing Corporation 1978
662 PART IV • TRANSPORT IN SPECIALIZED CELLS, TISSUES, AND ORGANS
"-rI 1G .....
KMena All
All • ~
CII -0.. 4
MIn
"" a.. r---e -- - - -loops of blood vessels ("vasa recta"). The collecting ducts course directly through the medulla carrying urine from distal tubules in the cortex to the renal pelvis. The descending vasa recta originate as efferent arterioles from the glomeruli of juxtamedullary nephrons. They loop at variable depths within the medulla,
JUXTAMEDUlLARY NEPHRON
DISTAl.
CORTICAL NEPHRON
CONVOlUTED ------.,'''[, PROXIMAl. CONVOlUTED
DISTAl. CONVOlUTED
PROXIMAl.
PROXIMAL STRAlGHT--~
SEGMENT
1HIt--- CCIRTlCAl COlLECTING
DUITING SEGMENT---H
THIN lIMBS Of HENlE'S lOOP
DESCENDING---11
ASCENDING --- -tt,
~--P~RYCOll£CTlNG
Fig. 2. Anatomy of rabbit nephrons. Note that the long loops of Henle originate injuxtamedullary nephrons. Modified from Peter. (6)
- Fig. 1. Relation between renal medullary thickness and maximum urinary osmolality. The relative medullary thickness is an index of the length of the loops of Henle. Long loops of Henle are necessary in order to make highly concentrated urine.(5)
forming ascending vasa recta which return to the cortex. Thus both blood vessels and tubules form loops in the medulla. The descending and ascending limbs of these loops are juxtaposed, thereby forming countercurrent systems. These countercurrent systems are critical for urinary concentration. As a result of equilibration of solutes and water between the limbs of the loops, the effects of transport are greatly amplified, and the concentration gradients thus formed are effectively isolated from the general circulation. Modern mathematical models of the urinary concentrating mechanism are based on these processes.
The ultrastructure of the thin limbs of Henle's loop differs markedly from that of other nephron segments. The cells are thin and flattened, containing few mitochondria. Ascending and descending thin limbs of Henle's loop differ with regard to the number of microvilli, degree of cellular interdigitation, and length of junctional complexes,m but the functional significance of the differences remains speculative.
3. Experimental Observations
Although there have been numerous studies of the renal medulla and of the renal concentrating mechanism, a few findings have greatly influenced subsequent analyses. The most important measurements have been of solute con-
CHAPTER 33 • TRANSPORT CHARACTERISTICS OF LOOP OF HENLE 663
centrations in the various structures in the medulla and of their permeabilities to solutes and water. The principal experimental observations are as follows.
3.1. The Concentration of Solutes Is Elevated in the Renal Medulla during Antidiuresis
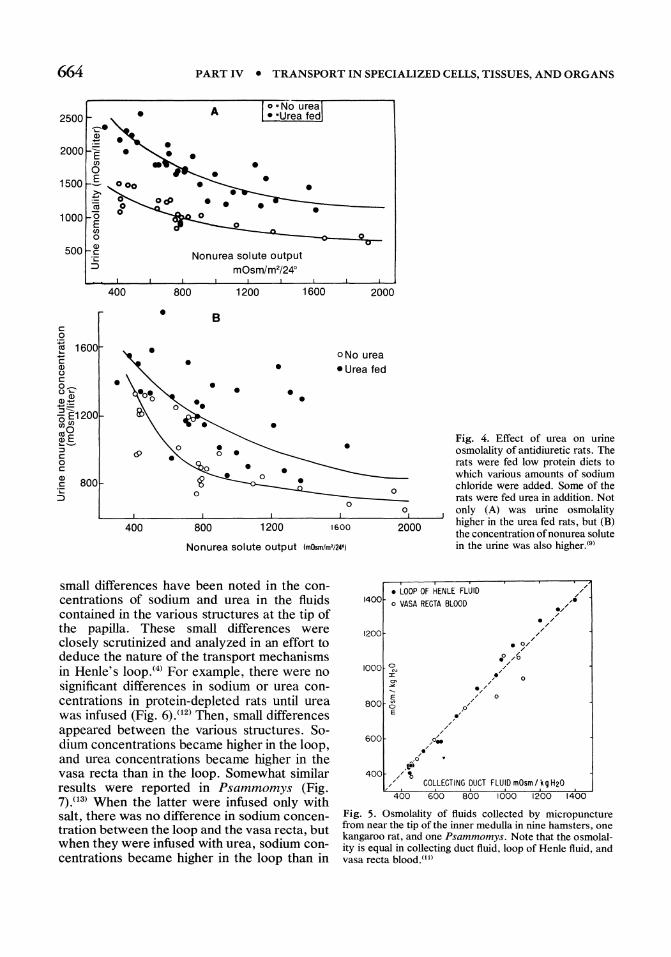
During antidiuresis, the concentration of solutes in the tissue of the renal medulla is much higher than in systemic blood, approximating that in the urine (Fig. 3A).(8) The osmolality increases progressively from the base of the outer medulla to the tip of the inner medulla. Urea and sodium chloride account for most of the osmolality. In contrast, during water diuresis the concentration of the solutes in the medulla is much reduced, approaching that in plasma (Fig. 3B). It is impossible to deduce the mechanism of concentration from such measurements, since the whole tissue which is analyzed obviously includes tubule fluid, extracellular fluid, and intracellular fluid. It is evident, however, that urinary concentration is related to the osmolality of the medullary tissue, and that the osmolality of the medulla is highest at its tip.
3.2. Urea Enhances Maximum Urinary Concentrating Ability
As already noted, there is a high concentration of urea both in urine and in medullary
A
2IDI
1l1li
Inner Outer URINE MEDULlA URINE
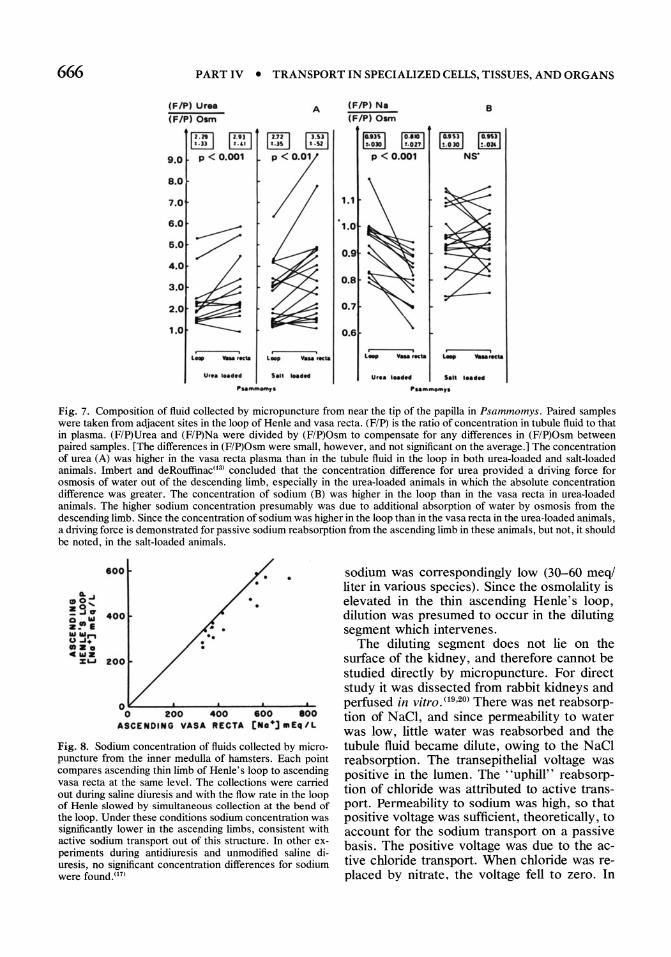
tissue, when urine is maximally concentrated. When rats are depleted of urea by feeding a low protein diet, the maximal concentration of the urine is reduced (Fig. 4A). (9) Subsequent administration of urea corrects the defect. The increase in urine concentration does not simply reflect a higher urea concentration in the urine. The concentration of nonurea solutes is also increased in the urine of rats (Fig. 4B). Therefore, urea has a special role in urine concentration.
3.3. At the Tip of the Papilla during Antidiuresis, Fluids in Loops of Henle, Collecting Ducts, and Blood Are Equiosmolal, but Differ Slightly in Composition
The composition of fluid contained in tubules and blood vessels in the inner medulla was measured by micropuncture. The renal pelvis was opened and fluid was sampled from superficial structures near the tip of the papilla. The method is limited by the inaccessibility of structures more than a millimeter or so from the tip of the papilla in most species. Also, when the renal pelvis is opened, there are changes in the composition of the fluid surrounding the papilla which may impair the concentrating mechanism. (10) The principal finding was that fluids from loops of Henle, collecting ducts, and vasa recta blood had equal osmolalities near the tip of the papilla (Fig. 5).(11)
Despite identical osmolalities, however,
B
MEDULlA
Fig. 3. Composition of the renal medulla. Medullas from rats were sliced and the solute concentrations in the tissue were measured at different depths. Note (A) that in antidiuresis the solute concentrations rose progressively, approaching those in urine at the tip of the inner medulla, and that urea made up a large fraction of the tissue osmolality. Also, note (B) that tissue solute concentrations were much reduced during diuresis. (8)
664 PART IV • TRANSPORT IN SPECIALIZED CELLS, TISSUES, AND ORGANS
2500 • A
2000
1500 • • 1000
Q)
o -ou----"O:..,.o.--
500 c .;: ::J
400 800
• c 0
~ 1600 c • Q) u c • o~ u ..... Q)2 +-':=
-5E1200 ",,,, ",0 e?..s ::J <9 c • 0 c Q)
800 c .;: ::J
400
Nonurea solute output mOsm/m2/24°
1200
B
• • • •
1600
• •
800 1200
2000
oNo urea • Urea fed
•
o 0
o
1600 2000
Nonurea solute output ImOsmim'i24')
Fig. 4. Effect of urea on urine osmolality of antidiuretic rats. The rats were fed low protein diets to which various amounts of sodium chloride were added. Some of the rats were fed urea in addition. Not only (A) was urine osmolality higher in the urea fed rats, but (B) the concentration of nonurea solute in the urine was also higher. (9)
small differences have been noted in the concentrations of sodium and urea in the fluids contained in the various structures at the tip of the papilla. These small differences were closely scrutinized and analyzed in an effort to deduce the nature of the transport mechanisms in Henle's 100p.(4) For example, there were no significant differences in sodium or urea concentrations in protein-depleted rats until urea was infused (Fig. 6),<12) Then, small differences appeared between the various structures. Sodium concentrations became higher in the loop, and urea concentrations became higher in the vasa recta than in the loop. Somewhat similar results were reported in Psammomys (Fig. 7).03) When the latter were infused only with salt, there was no difference in sodium concentration between the loop and the vasa recta, but when they were infused with urea, sodium concentrations became higher in the loop than in
• LOOP OF HENLE FLUID 1400 0 VASA RECTA BLOOD
1200
1000 ~ :x:
E 800 0
E
600
o
400 600 800 1000 1200 1400
Fig. 5. Osmolality of fluids collected by micropuncture from near the tip of the inner medulla in nine hamsters, one kangaroo rat, and one Psammomys. Note that the osmolality is equal in collecting duct fluid, loop of Henle fluid, and vasa recta bloodyn

CHAPTER 33 • TRANSPORT CHARACTERISTICS OF LOOP 9F HENLE 665
the vasa recta. Evidently, when the supply of urea is sufficient, sodium concentration is higher in the loop than in the vasa recta. This observation may have important implications for understanding the transport processes in the loop. The relatively high sodium concentration at the tip of the loop under these conditions was attributed to absorption of additional water (in excess of sodium) from the descending limb.o2,13) Further, it has been proposed that the sodium concentration gradient which is formed provides a driving force for sodium reabsorption from the ascending limb.o4,15) It should be noted, however, that no sodium concentration difference was detected in the absence of urea loading, despite the fact that the urine remained concentrated. Evidently, the sodium concentration difference is dependent on urea, but it is not clear that a sodium concentration difference is required for concentrating the urine.
3.4. Sodium Is Reabsorbed in the Thin Ascending Henle's Loop, but the Mechanism Involved Is Not Clear
In order to sample fluid from the base of the papilla in rats by micropuncture, a partial nephrectomy was performed exposing more of the papilla than is normally accessible. In this preparation, there was net reabsorption of . sodium by the ascending limbY6) Since reabsorption of sodium in this location is theoretically important for urinary concentration, the mechanism of the sodium absorption has been an important subject of investigation. It may be at least in part by active transport since, under special conditions, it occurs against a concentration difference in hamsters (Fig. 8).(17) However, under normal conditions of antidiuresis in rats(l6) and hamsters,(l7) the sodium concentrations is not significantly lower in the thin ascending Henle's loop than in the adjacent vasa recta, and compelling evidence for active sodium transport under these conditions is lacking. Similarly, when thin ascending limbs from rabbits were perfused in vitro with equal concentrations of sodium in the perfusate and bath, the sodium concentration in the collected fluid was unchanged; this experiment provided no evidence for active sodium transport. (18) Since significant concentration differences for so-
• I • i
• II. WI CD II. WI CD
Fig. 6. Concentrations of sodium and urea in fluids collected by micropuncture from near the tip of the inner medulla in rats on low-protein diets before (control) and after urea infusions. Before urea infusion the concentrations in thin descending loop of Henle (DL), vasa recta (VR), and collecting ducts (CD) were approximately equal. After urea infusion, the concentration of urea rose in all structures and became significantly higher in vasa recta blood than in descending loop of Henle. Sodium concentration rose significantly in the descending loop of Henle , but the absolute difference in concentration compared to vasa recta blood was not statistically significant. (12)
dium between the thin ascending limb and the vasa recta have not been consistently detected, it is not clear how much, if any, of the sodium reabsorption in the thin ascending limb is caused ordinarily by active transport.
3.5. Fluid in the Diluting Segment ("Thick Ascending Limb of Henle's Loop") Is Diluted by Active Chloride Reabsorption
The initial segment of the distal tubule, located at the end of Henle's loop, is the diluting segment (thick ascending limb of Henle's loop). Its function was deduced originally from micropuncture studies. Fluid at the beginning of the distal convoluted tubule was dilute even during antidiuresis (Fig. 9),<11) and the concentration of
666 PART IV • TRANSPORT IN SPECIALIZED CELLS, TISSUES, AND ORGANS
IF/P) Ur •• A (F/P) 0""
II][] I ·U ~ t . 4.' [ill] t .JS ~ t .Sf
9.0 P < 0.001
7; B.O
7.0
6.0
~ 6.0
4.0
~ 3.0
2.0
1.0
~--~, ~,----~,
.. ...., ... Iecil .. .., YMe rtC" u., ....... Sal1 ... ilM
IF/P) N.
IF/P) 0""
I"'" I 1°"" I :toO» t-on p < 0.001
8
10.1" I ,0.11'1 ~ .alO ~ . DII
NS'
l""~--_---';_ ... "'" --_--.:....,
Uf •• I .. ,,, .... "" ........ ,. Fig. 7. Composition of fluid collected by micropuncture from near the tip of the papilla in Psammomys. Paired samples were taken from adjacent sites in the loop of Henle and vasa recta. (F/P) is the ratio of concentration in tubule fluid to that in plasma. (F/P)Urea and (F/P)Na were divided by (F/P)Osm to compensate for any differences in (F/P)Osm between paired samples. [The differences in (F/P)Osm were small, however, and not significant on the average.] The concentration of urea (A) was higher in the vasa recta plasma than in the tubule fluid in the loop in both urea-loaded and salt-loaded animals. Imbert and deRouffinaco3l concluded that the concentration difference for urea provided a driving force for osmosis of water out of the descending limb, especially in the urea-loaded animals in which the absolute concentration difference was greater. The concentration of sodium (B) was higher in the loop than in the vasa recta in urea-loaded animals. The higher sodium concentration presumably was due to additional absorption of water by osmosis from the descending limb. Since the concentration of sodium was higher in the loop than in the vasa recta in the urea-loaded animals, a driving force is demonstrated for passive sodium reabsorption from the ascending limb in these animals, but not, it should be noted, in the salt-loaded animals.
800
~~ ..... !9~ 400 a .... a_ I ...... ,.., u ...... .a. c ... a
:z;u 200
o~----~----~----~----~ o 200 400 100 aoo ASCENOING VASA "ECrA [No·] .. Eca/L
Fig. 8. Sodium concentration of fluids collected by micropuncture from the inner medulla of hamsters. Each point compares ascending thin limb of Henle's loop to ascending vasa recta at the same level. The collections were carried out during saline diuresis and with the flow rate in the loop of Henle slowed by simultaneous collection at the bend of the loop. Under these conditions sodium concentration was significantly lower in the ascending limbs, consistent with active sodium transport out of this structure. In other experiments during antidiuresis and unmodified saline diuresis, no significant concentration differences for sodium were foundY7)
sodium was correspondingly low (30-60 meq/ liter in various species). Since the osmolality is elevated in the thin ascending Henle's loop, dilution was presumed to occur in the diluting segment which intervenes .
The diluting segment does not lie on the surface of the kidney, and therefore cannot be studied directly by micropuncture. For direct study it was dissected from rabbit kidneys and perfused in vitro. (19,20) There was net reabsorption of N aCI, and since permeability to water was low, little water was reabsorbed and the tubule fluid became dilute, owing to the N aCl reabsorption. The transepithelial voltage was positive in the lumen. The "uphill" reabsorption of chloride was attributed to active transport. Permeability to sodium was high, so that positive voltage was sufficient, theoretically, to account for the sodium transport on a passive basis. The positive voltage was due to the active chloride transport. When chloride was replaced by nitrate, the voltage fell to zero. In
CHAPTER 33 • TRANSPORT CHARACTERISTICS OF LOOP OF HENLE 667
contrast, when sodium was replaced by choline, the voltage persisted.
During water diuresis, the distal tubules and collecting ducts are relatively impermeable to water, so the hypotonicity of the urine is accounted for largely by excretion of the fluid formed in the diluting segment. Thus, active chloride transport in the diluting segment provides energy to dilute the urine. Paradoxically, it may also provide energy for the concentration of urine, despite the fact that the tubule fluid that is produced is dilute. Diluting segments, as well as distal tubules and cortical collecting tubules, are relatively impermeable to urea. Therefore, reabsorption of NaCI separates the salt that is absorbed from the urea that remains in the tubule lumen. The potential energy created by the separation of salt from urea in the diluting segment may in principle be utilized to transport sodium out of the thin ascending limbs of Henle's loop.
I I
1 0 09 08
07
06
05
0 4
03
•• • ••
• ••
U/p ; ••
OSMOL ALIT Y ~. ... -i
• •
'X. DISTAL CONVOLUTION URINE , I I '---J
o 20 40 60 110 100
90 80
70
60
~o
4 0
30
20
15
10
Fig. 9. Osmolality ratio (tubule fluid/plasma) of fluid collected by micropuncture in seven rats during antidiuresis. Fluid from the early distal tubule was dilute despite the fact that the urine was concentrated. (lI)
0.2 11.4 OJ! OJ 1.1) 12
FRACOON OF Fl1BlED I.HA REMAINING
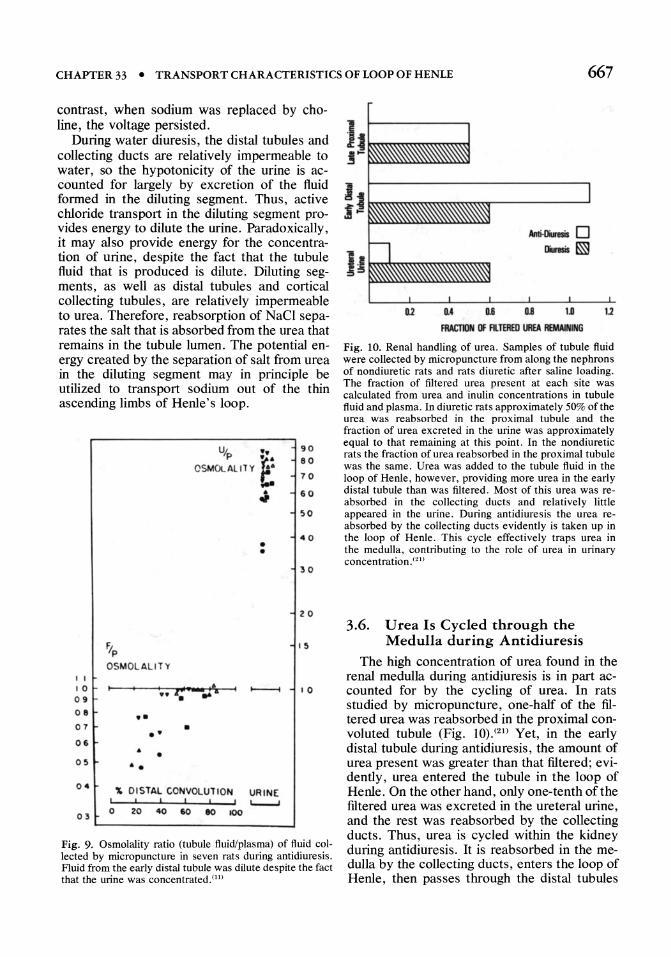
Fig. 10. Renal handling of urea. Samples of tubule fluid were collected by micropuncture from along the nephrons of nondiuretic rats and rats diuretic after saline loading. The fraction of filtered urea present at each site was calculated from urea and inulin concentrations in tubule fluid and plasma. In diuretic rats approximately 50% of the urea was reabsorbed in the proximal tubule and the fraction of urea excreted in the urine was approximately equal to that remaining at this point. In the nondiuretic rats the fraction of urea reabsorbed in the proximal tubule was the same. Urea was added to the tubule fluid in the loop of Henle, however, providing more urea in the early distal tubule than was filtered. Most of this urea was reabsorbed in the collecting ducts and relatively little appeared in the urine. During antidiuresis the urea reabsorbed by the collecting ducts evidently is taken up in the loop of Henle. This cycle effectively traps urea in the medulla, contributing to the role of urea in urinary concentration. (21)
3.6. U rea Is Cycled through the Medulla during Antidiuresis
The high concentration of urea found in the renal medulla during antidiuresis is in part accounted for by the cycling of urea. In rats studied by micropuncture, one-half of the filtered urea was reabsorbed in the proximal convoluted tubule (Fig. 10).(21) Yet, in the early distal tubule during antidiuresis, the amount of urea present was greater than that filtered; evidently, urea entered the tubule in the loop of Henle. On the other hand, only one-tenth of the filtered urea was excreted in the ureteral urine, and the rest was reabsorbed by the collecting ducts. Thus, urea is cycled within the kidney during antidiuresis. It is reabsorbed in the medulla by the collecting ducts, enters the loop of Henle, then passes through the distal tubules

668 PART IV • TRANSPORT IN SPECIALIZED CELLS, TISSUES, AND ORGANS
into the collecting ducts again. As noted earlier, salt is separated from urea in the diluting segment, storing potential energy. As a result of this urea "cycle," urea and salt mix again in the medulla and the stored potential energy is available for concentrating the urine.
3.7. The Descending and Ascending Thin Limbs of Henle's Loop in Rabbits Have Different Permeability Properties
The permeability properties of the thin limbs of Henle's loops have been studied directly by perfusing isolated rabbit tubules in vitro. (18,22)
Thin descending limbs of Henle's loop were found to have a high permeability to water and low permeability to salt and urea (Fig. 11).(22)
Fluid traversing the thin descending limb in vivo passes successively through regions of the medulla in which the osmolality is increasing. In the absence of active transport, osmotic equilibration of the tubule fluid could occur passively either by uptake of solute or abstraction of water. The latter presumably occurs in rabbits since, when fluid in the bath surrounding the tubules in vitro was hyperosmotic, equilibration occurred almost entirely by loss of water from the tubule fluid.(22) In contrast, uptake of solute, sodium chloride and urea, is apparently substantial in rats(l2,23) and Psammomys, (13,24) based on micropuncture studies. In the micropuncture studies, the flux of solute into the lumen of the thin descending limb was inferred in part from comparison of fluid taken from the end of the proximal convoluted tubules to that at the tip of the loop of Henle and
Fig. 11. Permeability of thin descending limb of Henle's loop compared to thin ascending limb. Isolated perfused rabbit tubules were studied in vitro. Note that permeability to water was much higher in the descending limb; permeability to solute was much higher in the ascending limb. Also, permeability to sodium exceeded that to urea in the ascending limb .oS,22)
Water (osmotic)
Sod ium
Urea
beginning of the distal convoluted tubule. Although the results are difficult to explain on any other basis, the conclusion lacks certainty for two reasons: (a) Because oflimitations inherent in the micropuncture technique, the proximal and distal tubule samples are by necessity from cortical nephrons, whereas the loop samples are from juxtamedullary nephrons. Therefore, it is conceivable that the results are due to differences between the populations of nephrons rather than solute entry into the descending limb. (b) Between sites accessible for micropuncture there were other nephron sements in addition to the thin descending limbs (e.g., proximal straight tubules and/or ascending limbs), making it difficult to be sure what portion of the transport occurred in the thin descending limbs. *
Additional evidence for solute entry into the descending limb of Henle's loop was derived from comparison of inulin concentration and osmolality of fluid collected from the tip of the loop. Samples collected from kidneys producing more concentrated urine had a higher osmolality, but little change in inulin concentration. (24) Therefore, the increased osmolality was attributed to entry of additional solute into the descending limb rather than abstraction of additional water.
In summary, despite the reservations just indicated, the evidence for solute entry into the descending thin limbs in rats and Psammomys is formidable. As will be discussed, the differ-
* Along this line, Kawamura and Kokkat25) reported that rabbit proximal straight tubules perfused in vitro secreted urea into the tubule lumen, If a similar process occurs in rat and Psammomys in vivo. it could explain at least part of the excess of urea found in Henle's loop.
DESCENDING LIMB ASCENDING LIMB
Xlcr'
Parm •• bllity (em sec- I)
3
CHAPTER 33 e TRANSPORT CHARACTERISTICS OF LOOP OF HENLE 669
1.1
1.0 o
o 0 0 0 0
-.---------~--------."-.~ .... -.- •• -•••••••• .?.-o--~.-..Q. •. P. •• Q ••
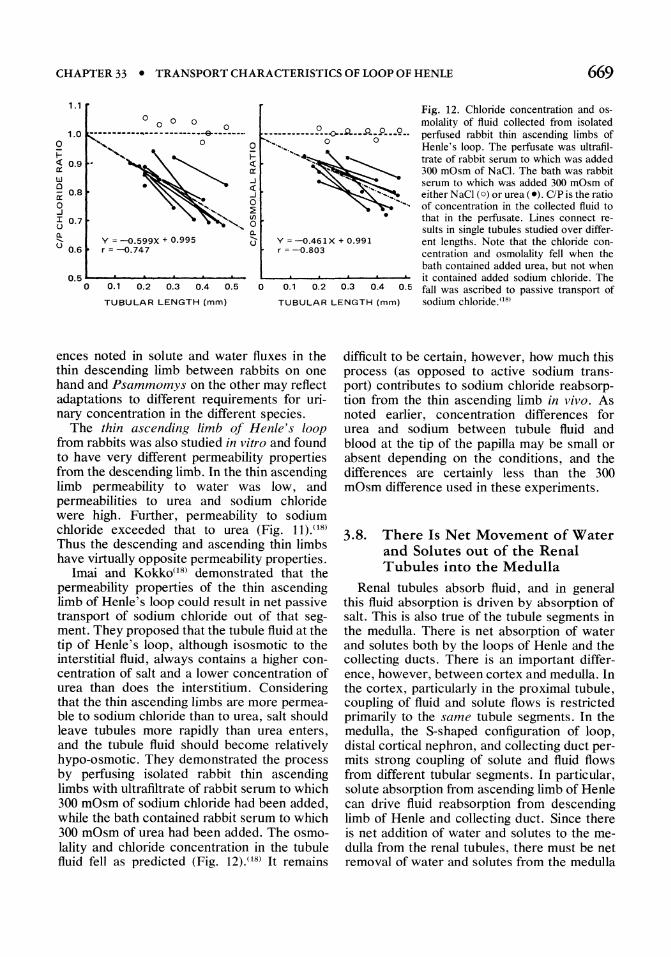
Fig. 12. Chloride concentration and osmolality of fluid collected from isolated perfused rabbit thin ascending limbs of Henle's loop. The perfusate was ultrafiltrate of rabbit serum to which was added 300 mOsm of NaCl. The bath was rabbit serum to which was added 300 mOsm of either NaCI (0) or urea (e). c/Pis the ratio of concentration in the collected fluid to that in the perfusate. Lines connect re-
o 0 ..... 0 0
.... ~.'~~ I-~ 0.9
W o a: 0.8
9 3 0.7
" ~ a: ..J c:( ..J o ~ I/)
o Q.
., ... , •
Q.
l) 0.6 Y = -o.599X + 0.995 r = -0.747
l) Y = -0.461 X + 0.991 r = -0.803
sults in single tubules studied over different lengths. Note that the chloride concentration and osmolality fell when the bath contained added urea, but not when it contained added sodium chloride. The 0.5 ...... -"-----"'-----....... o 0.1 0.2 0.3 0.4 0.5 o 0.1 0.2 0.3 0.4
TUBULAR LENGTH (mm)
0.5 fall was ascribed to passive transport of sodium chloride. (18) TUBULAR LENGTH (mm)
ences noted in solute and water fluxes in the thin descending limb between rabbits on one hand and Psammomys on the other may reflect adaptations to different requirements for uri· nary concentration in the different species.
The thin ascending limb of Henle's loop from rabbits was also studied in vitro and found to have very different permeability properties from the descending limb. In the thin ascending limb permeability to water was low, and permeabilities to urea and sodium chloride were high. Further, permeability to sodium chloride exceeded that to urea (Fig. 11).08)
Thus the descending and ascending thin limbs have virtually opposite permeability properties.
Imai and Kokko(l8) demonstrated that the permeability properties of the thin ascending limb of Henle's loop could result in net passive transport of sodium chloride out of that seg· ment. They proposed that the tubule fluid at the tip of Henle's loop, although isosmotic to the interstitial fluid, always contains a higher con· centration of salt and a lower concentration of urea than does the interstitium. Considering that the thin ascending limbs are more permea· ble to sodium chloride than to urea, salt should leave tubules more rapidly than urea enters, and the tubule fluid should become relatively hypo·osmotic. They demonstrated the process by perfusing isolated rabbit thin ascending limbs with ultrafiltrate of rabbit serum to which 300 mOsm of sodium chloride had been added, while the bath contained rabbit serum to which 300 mOsm of urea had been added. The osmo· lality and chloride concentration in the tubule fluid fell as predicted (Fig. 12).08) It remains
difficult to be certain, however, how much this process (as opposed to active sodium trans· port) contributes to sodium chloride reabsorp· tion from the thin ascending limb in vivo. As noted earlier, concentration differences for urea and sodium between tubule fluid and blood at the tip of the papilla may be small or absent depending on the conditions, and the differences are certainly less than the 300 mOsm difference used in these experiments.
3.8. There Is Net Movement of Water and Solutes out of the Renal Tubules into the Medulla
Renal tubules absorb fluid, and in general this fluid absorption is driven by absorption of salt. This is also true of the tubule segments in the medulla. There is net absorption of water and solutes both by the loops of Henle and the collecting ducts. There is an important differ· ence, however, between cortex and medulla. In the cortex, particularly in the proximal tubule, coupling of fluid and solute flows is restricted primarily to the same tubule segments. In the medulla, the S·shaped configuration of loop, distal cortical nephron, and collecting duct per· mits strong coupling of solute and fluid flows from different tubular segments. In particular, solute absorption from ascending limb of Henle can drive fluid reabsorption from descending limb of Henle and collecting duct. Since there is net addition of water and solutes to the me· dulla from the renal tubules, there must be net removal of water and solutes from the medulla

670 PART IV • TRANSPORT IN SPECIALIZED CELLS, TISSUES, AND ORGANS
4. 1} • AHL ~ J ___ DH~_~ .... _~ _
CD =9
Fig. 13. Kuhn's countercurrent multiplier. The system consists ofloop of Henle and collecting duct. Concentration is built up in the loop by water moving from descending limb (DHL) to ascending limb (ARL) or solute from AHL to DHL. Water and solute move between DHL and collecting duct (CD), so that CD and DHL attain nearly the same osmolality. There is no provision in the model for the transport of water and solute from medullary nephron segments to the interstitium and vasa recta:=:>Water movement;". solute movement. Adapted from Hargitay and Kuhn'2" and Kuhn and Ramel(2"' for Stephenson.'34)
by the blood vessels. In the steady state, this addition and removal must exactly balance. The quantitative aspects of this balance are the essence of the concentrating mechanism.
4. Functional Role of the Loop of Henle in the Concentration of Urine
Theoretical models of the concentrating mechanism have played an essential role in the development of our understanding of the function of the loop of Henle. Thus, the elaborate counterflow system of the loop was regarded as an accident of evolutionary packaging(26) until Kuhn and his co-workers, in a series of brilliant papers, analyzed analogous counterflow systems theoretically and experimentally and pointed out that counterflow permitted a small single effect to be multiplied many-fold to produce a highly concentrated fraction of the outflow. (27-30)
The model in the form that became widely accepted is illustrated in Fig. 13. The basic single membrane process is active salt transport from ascending limb of Henle to descending limb of Henle. This process is multiplied by counterflow to build up a concentration gradient that increases from the cortex to the tip of the papilla. This model predicted the high osmolality of all medullary structures that was
found in both slice studies and by micropuncture, and also predicted that, in the concentrating kidney, either net water addition to the loop of Henle or net solute addition to the collecting duct must occur.
N either of these predictions is supported by observation. In fact, experimental data show that in both the concentrating and diluting kidney, there is net absorption of water and solutes from both loops of Henle and collecting duct. There is no place for this water and solute to go except into the vasa recta (see Fig. 14). It is clear that any adequate model of the concentrating mechanism must as a first requirement satisfy mass conservation and that to do this the vasa recta must be incorporated into the model. Early in the development of the Kuhn models, the idea was promulgated that the vasa recta, by countercurrent exchange, prevents "washout" of the medullary gradient developed by countercurrent multiplication.'31-33) This concept was grafted onto the Kuhn solute cycling multiplier, but with no provision for mass conservation. This hybrid model became the orthodox view of the concentrating mechanism, but this view is valid only for concentration by pure solute cycling.
Here we present an alternative model of the concentrating mechanism, which has been developed in the past few years. (14,34-36) In this model, the basic process is not solute cycling from ascending to descending limb of Henle, but volume absorption from the loop of Henle and collecting duct driven by solute absorption from loop of Henle and collecting duct. In this model, the basic concentrating unit is not the loop of Henle, but an integrated nephrovascular unit consisting of loop of Henle, vasa recta, collecting duct, and distal cortical nephron (Fig. 14).
The quantitative analysis of this nephrovascular system in terms of membrane parameters requires detailed numerical calculations. A partial analysis of this type has been carried OUt.(37,38) Qualitative understanding of the model can be attained without such detailed calculations by making some limiting assumptions about the water and solute permeabilities of the tubules and capillaries. This leads to the central core model of the medulla. It is in terms of this model and basic mass conservation relations that we analyze loop of Henle function.

CHAPTER 33 • TRANSPORT CHARACTERISTICS OF LOOP OF HENLE 671
4.1. Central Core Model of the Nephrovascular Counterflow System
In order to understand how the coupled counterflow system of tubule and vasa recta concentrates by solute-driven volume absorption, we will exploit systematically some simple conservation relations. In the steady state at any given medullary level, the net volume taken up by the vasa recta must equal net volume absorbed by the tubules between that level and the papillary tip. Thus, we have our first conservation relation:
volume absorbed = volume outflow - volume inflow (1)
A similar conservation relation obtains for each individual solute and for the total osmolality, i.e. ,
solute absorbed = solute outflow - solute inflow (2)
Ifwe neglect axial diffusion, we have, for either tubule or capillary,
solute flow = v olume flow x concentration (3)
Let us now center our attention on solute removal by the vasa recta. In general, concentrations in ascending and descending vasa recta will differ, and solute removal is given by the difference in the products of each concentration and the corresponding volume flow. If, however, solute permeabilities of the vasa recta are large, their solute concentration will tend to equilibrate with the surrounding interstitium. Thus, at a given medullary level, concentration of every solute in interstitium and ascending and descending vasa recta will be nearly the same. For Eq. (2) we can then write for each medullary level
solute absorbed = (volume outflow - volume inflow) x concentration (4)
which can be combined with Eq. (1) to give
solute absorbed = volume absorbed x concentration (5)
This model is functionally equivalent to merging ascending and descending vasa recta and interstitium into a single tube, the central
Fig. 14. Diagram to illustrate the main feature of solute and water movement in the S-shaped counterflow system. Salt without water is transported out of AHL by a combination of active and passive means. This salt increases the osmolality of the medullary interstitium. Some of the salt may enter DHL to be recycled but primarily it osmotically extracts water from DHL and CD. Urea, concentrated by water removal along the nephron, diffuses from CD to the interstitium; palt enters the loop of Henle to be recycled, part is returned to the cortex by the vasa recta.
core, which is closed at the papillary tip and open at the corticomedullary junction where it is supposed to discharge into the systemic circulation (Figs. 15 and 16).
How does the system concentrate? If everywhere along the loop of Henle and collecting duct absorption was isotonic or nearly so, as in the proximal tubule, the isotonic absorbate would flow up the core and no concentration would occur. The system can concentrate because of the different permeability properties of ascending and descending limbs of Henle. Here the essential feature is the relative water impermeability of ascending limb. Given this, the system can concentrate by water extraction, solute addition, or any combination of the two.
We will first consider concentration by water

672 PART IV • TRANSPORT IN SPECIALIZED CELLS, TISSUES, AND ORGANS
IIOTONIC
DILUTE
18DTOIIIIC
IIOTOIIIIC PLAaA
OHL
AVR
'~=~~----CORE'-------OVR ~ __ -4 _________________ ~ __ ,_'
tit,
extraction, both because it is simpler to explain and because no correct theoretical analysis is possible with the classical models. Let us assume that both descending limb and collecting duct are solute impermeable but water permeable, and that ascending limb is water impermeable but transports solute from lumen to core by some unspecified mechanism (Fig. 16). These transport characteristics permit the ascending limb (AHL) to supply solute without water to the core. This solute raises the osmolality of the core, and water without solute moves from the descending limb and collecting duct into the core. The net result is to dilute the fluid in the ascending limb and to concentrate the fluid in the descending limb, collecting duct, and core. In addition, the combined absorption of solute and water generates a volume flow up the core counter to the flow in collecting duct and descending limb. Given sufficiently large hydraulic permeabilities of descending limb of Henle (DHL) and collecting duct (CD), the counterflowing solutions in core and these
Fig. 15. (a) Fully connected central core model. With sufficiently large solute and water permeabilities of ascending vasa recta (A VR) and descending vasa recta (DVR), concentrations in A VR, DVR, and interstitium become identical at a given medullary level. Solute removal then is given by the product of the concentration by flow difference in A VR and DVR. Functionally A VR, DVR, and interstitium are merged into a single tube closed at the papillary tip and open at the junction of medulla and cortex; (b) shows various core segments being hypothetically merged to give the final flow tube configuration in (c) . From StephensonY4)
structures will equilibrate osmotically at every medullary level. In accordance with Eq. (4), the product of the core flow by the core concentration at a given medullary level must equal solute absorbed from the ascending limb between that level and the papillary tip. Stated inversely, this means that, if CM(x) is the common osmolality of core, descending limb and collecting duct, we have at that level:
core flow = solute absorbed/CM(x) (6)
But, by Eq. 0), the core flow equals the water reabsorbed from the descending limb and collecting duct; hence,
water absorbed = solute absorbedl C M(X) (7)
or
(8)
where Tv(x) is water abs"orbed and T ~x) is solute absorbed between level x and the tip. Since no solute leaves DHL or CD, this water

CHAPTER 33 • TRANSPORT CHARACTERISTICS OF LOOP OF HENLE 673
absorption concentrates the fluid in DHL and CD according to the relation
CM(L) FDHL(X) + FCD(x) ----CM(x) FDHdL) + FCD (L)
FDHL(X) + FCD(x) (9)
FDHdx) + FCD(x) - Tv(x)
where C M(L) is the common osmolality of structures at the papillary tip, FDHL(X) axial volume flow in DHL at level x. and FCD(x) axial volume flow in CD. Dividing numerator and denominator of the right-hand side of Eq. (9) by the sum of the volume flows yields
CM(L) ( TvCx) )-1 (10) CM(x) = 1 - FDHL(X) + FCD(x)
Thus the concentration ratio, rex) = CuCL)! C uCx), generated by the counterflow system or any segment of it, depends on the fractional volume absorption Tv(x)![FDHL(X) + FCD(x)]. This volume absorption from the descending limb is driven by solute absorption from the ascending limb according to Eq. (8). When this relation is substituted into Eq. (10), we obtain
( T uCx) )-1 r( x) = 1 - ::-=--:--:---=-=--:--:-=-:::--:--:
[FDHL(X) + FCD(x)]CM(x) (11)
The term [FDHdx) + FCD(x)]Cu(x) is the combined solute inflow to the descending limb and collecting duct. Thus, the concentration ratio developed by the system or a segment of it is a function of the fractional solute absorption. This relation carries over in a slightly modified form to solute cycling and mixed modes of operation, but in Eq. (11) we regard TM(x)! CuCx) as solute-driven volume absorption. For a given solute transport out of AHL to extract a maximum volume from DHL and CD, it is essential that the outflowing solutions in ascending vasa recta (A VR) equilibrate not only with inflow in the descending vasa recta (DVR) , but also with inflow in DHL and CD. When complete equilibration of concentrations in A VR and DVR by the highly efficient counterflow system is assumed, the core model emerges and the prime function of the vascular loop is unmasked. It serves as a mixing and expansion chamber'that links solute transport out of the ascending limb of Henle to volume transport out of the descending limb of Henle
AHL ~
~:......-_C_0-1:F-_E __ J DHL
CD
Fig. 16. Fundamentals of the concentrating mechanism. Solute without water from ascending limb increases the osmolality of the vascular interstitial core and extracts water without solute from descending limb and collecting duct. Solute and water mix and expand in the core. The large hydraulic permeabilities of DHL and CD permit the counterflowing solutions in core and nephrons to equilibrate osmotically, thereby maximizing water extraction and concentration at the papillary tip. From Stephenson.(34)
and collecting duct. In order to gain a clearer understanding of the loop of Henle function in this nephrovascular counterflow system, we extend this analysis to include concentration in the descending limb by solute cycling or by a mixture of solute cycling and water extraction.
4.2. Concentration by Solute Cycling and Mixed Mode Operation in Central Core Model
If the ascending limb of Henle is water impermeable, its transmural volume flow vanishes everywhere and ,axial volume flow in the ascending limb remains constant and numerically equal to volume flow entering from the descending limb of Henle at the papillary tip. Adopting the sign convention that flow toward the tip IS positive and away is negative, we have
FAHL(X) = -FDHL(L) (12)
Likewise, since there is no net addition to the algebraic sum of the volume flows in core, DHL, and CD, this sum must remain constant, or
FDHL(X) + FCD(x) + Fcore(x) = const (13)
Equation (13) must hold at the tip where Fcore(L) = 0; therefore,
FDHL(X) + FCD(x) + Fcore(x)
= FDHL(L) + F cD(L) (14)

674 PART IV • TRANSPORT IN SPECIALIZED CELLS, TISSUES, AND ORGANS
Solute transport out of AHL equals total net solute uptake by the core, DHL, and collecting duct. Denoting these transports by Jcore(X) , JDHdx), JCD(x), and JAHdx), we have at every medullary level
Jcore(x) + JDHdx)+ JCD(x) = -JAHdx) (15)
For AHL we have the basic conservation equation for axial solute flow:
d[F AHdx)C AHL(X)] _ -J () dx - AHL X (16)
which, by Eq. (12), becomes
dCAHL(X) FDHdL) dx = J AHL(X) (17)
For the sum of the axial solute flows in DHL, core, and CD, we have the differential equation:
d[Lcore.DHL,CD Fi(X) Ci(x)]
dx
is net solute transport out of AHL between level x and the tip of the papilla.
Equations (20) and (21) can be manipulated algebraically to give numerous relations useful in understanding the concentrating mechanism. The concentration ratio r(x) attained by the segment of medulla between a given level and the tip of the papilla is given by
rex) == C~L) C~x)
-(1 TAHL(X) )-1 [FDHdL) + FCD(L)]C~L) (23)
Equation (23) is identical to Eq. (11), except that in Eq. (23) transport out of AHL is referred to solute flow in DHL and CD at the tip. In this form the equation is valid whether or not solute is cycled between core and DHL or CD. Equation (23) can be cast into the dimensionless form
rex) = [1 - fT(X)(1 - fU)]-l (24)
= -[Jcore(x) + JDHL(X) + JCD(x)] (18) where
If we assume that core, DHL, and CD have the common osmolality CM(x) and substitute from Eq. (14) and (15), we obtain
dC~x) [FDHL(L) + FCD(L)] dx = JAHdx) (19)
Equations (17) and (19) are the linear differential equations with constant coefficients that relate concentration gradients in medullary structures to transport out of the ascending limb of Henle.
Integration of Eq. (17) and (19) gives the equations
FDHdL)[CM(L) - CAHdx)]
= fJAHL(X)dX=TAHL(X) (20)
and
[FDHL(L) + FCD(L)][ CM(L) - CM(x)]
= f J AHdx) dx = T AHL (x) (21)
where
(22)
fAx) == TAHL(X)/[FDHL(L) C~L)] (25)
is fractional solute transport out of the AHL system and
fu= FCD(L)I[FDHL(L) + FCD(L)] (26)
is fractional urine flow. * Micropuncture studies show that fluid enter
ing the distal nephron from AHL is hypotonic. From Eq. (20) and (21) we calculate the relative
*This analysis of the medullary counterflow system sets an upper limit on the concentrating effect of given net solute transport out of AHL. This ratio is not attained because of various dissipative processes: (a) axial diffusion of solute; (b) incomplete osmotic equilibration between ascending and descending flows; (c) water reabsorption from AHL; and (d) hypotonicity of CD inflow. An analysis(36J that takes into account these proces ses leads to the modified normalized equation for the concentration ratio
r = [I - fTO - JU)(1 - fWWl
where fw is the fractional dissipation,
fw == (W + TAHL.VCPM - Fcn(O) ilCcn)ITAHL
where W is the solute that is unavailable for concentration because of axial diffusion down the concentration gradient or hypertonicity of A VR outflow, T AHL, v is net water reabsorption from AHL, C pM is osmolality of DHL inflow, and ilCCD is hypotonicity of CD inflow,

CHAPTER 33 • TRANSPORT CHARACTERISTICS OF LOOP OF HENLE 675
hypotonicity of ascending limb at any level in the medulla to be
C~x) - CAHdx)
= [FDH~(L) - FDHL(L) ~ FCD(L)]TAHd X )
(27)
We now are in a position to understand the general function of the loop of Henle. We remind the reader that in deriving the foregoing equations we have introduced no assumptions about solute exchange between the interstitial vascular core of the concentrating unit and the tubules except that permeabilities of descending limb and collecting duct are such that they equilibrate osmotically with the core. This equilibration can be by a combination of water extraction and solute cycling. Overall, the concentrating unit splits the descending limb and collecting duct inflow into three outflows: ascending limb, final urine, and the absorbate that is returned to the systemic circulation via the core. The total vohime of this absorbate depends on the net solute absorption from loop of Henle and collecting duct, but the development of a concentration gradient is dependent on net solute transport without water from ascending limb of Henle. If solute transport out of ascending limb of Henle, J AHL(X) in the equations, falls to zero, then the concentration gradient goes to zero and at every medullary level and in all structures concentrations will be identical and equal to C,y(O) , which is to say nearly isotonic with plasma.
Overall, the loop of Henle supplies solute in excess of volume to the core. The measure of this excess solute is the dilution of ascending limb outflow; thus,
excess solute = iFAHL(O)I[CM(O) - CAHL(O)] (28)
~her~ iF AHL(O) I is the absolute value of ascendmg 11mb volume flow. Substituting from Eq. (26) and using (27), we find
excess solute = fuT AHL(O) (29)
Thus the excess solute supplied by the loop is the product of the fractional urine flow and total net solute transport out of the ascending limb. Since the absorbate returned to the systemic circulation by the core is isotonic or
nearly so, the sum of ascending limb outflow and final urine must be isotonic. Hence, in the overall operation of the concentrating unit it is the excess solute supplied by the loop that extracts water without solute from the collecting duct system. This water is the negative free water T'iI 2o of Smith.
Permeability studies on isolated tubules imply that rabbits must concentrate almost entirely by water extraction.(18,22) Micropuncture studies on Psammomys suggest that they concentrate descending limb fluid primarily by solute additionY3,24,39) Studies on rats and hamsters(11,23) suggest a mixed mode operation.
The general effect of solute entering into either descending limb of Henle or collecting ~uct is to increase solute flow at the papillary tIp
[FDHdL) + FCD(L)]CM(L)
If T AHL(O) remains fixed, this decreases the ratio
TAHdO)/{[FDHL(L) + FCD(L)]CM(L)}
and as can be seen from Eq. (11), the concentration ratio C,y(L)/CM(O). Conversely, solute extraction from either descending limb of Henle or collecting duct will tend to increase the concentration ratio. Put another way, and referred to the dimensionless equation (24), solute absorption from descending limb tends to increase frCO) and solute reabsorption from collecting duct tends to decrease fu. Both tend to increase the overall concentration ratio rand tota~ water reabsorption from the loop and collectmg duct. In general, solute cycling from core to nephron is a dissipative process and tends to decrease urinary concentration and to decrease water reabsorption and so increase urinary flow. If, however, as solute cycling is increased, ascending limb solute transport T.~HL(O) increases so as to maintain the fractional transport.fT(O) more or less constant, and fractional urine flow .fu remains constant, the concentration will not fall, the urine flow will increase, and the kidneys will be able to excrete a relatively large volume of relatively concentrated urine.
It seems plausible that, depending on adaptive needs, a species may make greater or lesser use of water extraction versus solute cycling to concentrate descending limb flow. Thus, a desert rodent with an extreme water

676 PART IV • TRANSPORT IN SPECIALIZED CELLS, TISSUES, AND ORGANS
conservation problem might be expected to concentrate by water extraction. On the other hand, Psammomys, which has no particular water conservation problem, but because of its diet of halophile plants must excrete a large excess of salt, might be expected to utilize solute cycling to maximize its osmolal clearance. These shifts from water extracting to solute cycling modes of operation must of course be reflected in variations in the relative water and salt permeabilities of descending limb of Henle.
4.3. Augmentation of Concentration by Urea Cycling
As we have noted, in many mammals maximum concentrating ability appears to be related to urea cycling. In the cycle, water and salt absorption along the loop of Henle and distal nephron leaves urea behind and relatively concentrated in the tubular fluid. In the medullary collecting duct, which is relatively permeable to urea, this concentrated urea is absorbed and enters the vascular core, where it contributes to the solute concentration. Part of the urea is removed by the volume flow up the core and part enters the loop to be recycled.
How can this urea cycle augment the concentration ratio? By Eq. (24), it must do so by one of two means: It must either increase the fractional transport JrlO) or decrease the fractional urine flow fu. Theoretical analysis of this process using the central core model showed that if the descending limb of Henle is relatively impermeable to urea, cycling depresses core salt concentration relative to loop salt concentration. This augments salt transport out of the ascending limb of Henle and thus increases fT(0).(35) To understand this process (Fig. 17), we note first that at any level the net urea supplied to the core between that level and the tip must equal the product of net volume flow up the core by core urea concentration; thus, Turea = Feore x [urea core], where Turea is the net urea supplied to the core between the given level and the tip. Similarly, for salt we have the relation
TsaJt = F core x [salt core] (30)
From these two relations we find that any level
[salt core] [salt core] + [urea core]
Tsalt + T urea (31)
But if core and descending limb equilibrate osmotically and the reflection coefficients of both salt and urea are near unity, then
[salt core] + [urea core] = [salt DHL] + [urea DHL] = C M(X) (32)
From Eqs. (31) and (32) we derive
[salt DHL] - [salt core]
= [salt DHL] - C M(X) TsaJt (33) TsaJt + T urea
From Eq. (33), it can be seen that if the salt concentration in the descending limb of Henle and total osmolality are held fixed, the effect of urea transport from collecting duct into the core is to depress core salt concentration relative to descending limb salt concentration. In other words, the effect of urea cycling is to add a positive increment to the difference between core and descending limb salt concentration:
Msalt] = [salt DHL] - [salt core] (34)
When descending limb flow is reversed at the tip to become ascending limb flow, this same positive increment remains to increase ascending limb salt concentration relative to core salt concentration. This increment must increase salt transport out of the ascending limb if it is salt permeable. To see this, let us assume that salt transport out of ascending limb is given by
a J AHL = --,--------::--:-::-::-::-~,..__-
1 + bl[NaCl]AHL
+ hs{[N aCl]AHL - [N aCl]eorc} (35)
where a is the maximum rate of transport, b the Michaelis constant, and hs the salt permeability. If salt is the only solute in the system, it follows from Eq. (27) that core concentration is greater than ascending limb concentration and the passive transport term hs{[NaCl]AHL -[NaCl]eore} will be negative. If urea is now cycled from collecting duct to core, a positive increment will be added to this term and hence to J AHL. If sufficient urea is cycled. the differ-

CHAPTER 33 • TRANSPORT CHARACTERISTICS OF LOOP OF HENLE 677
---_ co
---.. SALT
c::> WATER
~ UREA
--_ FLOW
---_ DHL
Fig. 17. Increase of concentration in central core model by urea cycling. Urea from CD and salt from AHL and CD mix in the core and extract water from CD and DHL. The combined volume expansion of the core mixture and contraction of DHL fluid depresses core salt concentration relative to DHL salt concentration. The concentration difference adds a positive increment to transport of salt out of AHL by passive diffusion and so enhances concentration. From Stephenson and Mejia (unpublished).
ence [NaCl]AHL - [NaCl]core can become positive and there can be net transport out of ascending limb of Henle with no active transport.
For urea cycling to augment the concentration ratio in this way, there are some obvious permeability requirements. The urea-rich solution entering the collecting duct must be generated in the outer medulla and cortex by pumping salt out of the nephron and leaving urea behind. This implies both active salt transport in these segments of the tubule and relative urea impermeability. For the urea to be delivered to the interstitial vascular core in the inner medulla, it must be retained in the collecting duct in the outer medulla, but permeate the collecting duct in the inner medulla. To extract water osmotically from the loop of Henle, urea must not permeate the descending limb too readily. Finally, for the depressed salt concentration in the core to induce significant passive salt transport out of ascending limb of Henle, its salt permeability must be large. Experimental studies on isolated rabbit tubules showed that qualitatively they satisfied the required permeability properties. Reasoning from these permeability data, Kokko and Rector postulated that urea diffusing into the inner medulla from the collecting duct could induce passive salt transport out of the thin ascending limb; and that concentration in the inner medulla
could occur without active transport,(1o) thus reaching the same conclusion that had been predicted from the theoretical analysis of the central core model. (14) This recent independent and essentially simultaneous postulation of the passive mechanism from the central core model and permeability data was preceded by many years by Kuhn and Ryffel's enunciation of the principle of concentration by mixing. (27) They demonstrated the process experimentally in model sucrose and phenol systems, and suggested the kidney might somehow use the mixing process in the concentration of urine. In Kuhn's subsequent work on the medullary counterflow system, the idea was never developed in any consequential way.
Since the postulation of the passive mechanism, the attention of physiologists has centered on whether it can entirely account for concentration in the inner medulla. (4) What has not been emphasized is that even in the presence of active transport of salt out of ascending limb the additional passive salt transport generated by urea cycling is an important feature of the concentrating process, because it is the mechanism whereby urea cycling augments concentrating ability.
If ascending limb is impermeable to salt, model studies show that infusion of urea can lead only to osmotic diuresis and decreased

678 PART IV • TRANSPORT IN SPECIALIZED CELLS, TISSUES, AND ORGANS
concentration. Urea cycling reduces the extent of this diuresis, but does not enhance concentration until the passive mechanism is introduced into the model (Stephenson and Mejia, unpublished) .
Although theoretical analysis,(14,32) computer simulation, (34) and experiments on isolated tubules(18) have all shown that in principle it is possible for concentration in the inner medulla to occur entirely by passive means, theoretical analysis and computer simulation also have shown that there are many limitations on a completely passive mechanism. Among other things, urea leakage into the loop sharply limits the mechanism. Thus it is not very surprising that a species such as Psammomys which appears to concentrate mainly by solute cycling seems to make little use of the mechanism. (When Psammomys were urea loaded, the concentration of the urine decreased(13) rather than increased as in other species.) This therefore appears to be an exception to the generalization that urea augments urinary concentration.
S. Summary
The principal function of the loop of Henle is to supply solute in excess of water for the concentration of fluid in the collecting duct system. The S-shaped configuration of loop, distal nephron, and collecting duct permits this solute to be delivered to the medullary interstitium where it abstracts water from the collecting duct by osmosis. The excess solute and abstracted water mix and expand to near isotonicity in the interstitial vascular core. It is this volume expansion that supplies the immediate work required for concentrating the final urine. The water abstracted by the excess solute in general is only a small fraction of the total volume of nearly isotonic fluid that is reabsorbed from collecting duct and loop together and returned to the systemic circulation by the vascular core. For the efficient operation of the system it is essential that at a given medullary level, interstitium, ascending and descending vasa recta, descending limb of Henle, and collecting duct all equilibrate osmotically with one another. This is accomplished by the highly efficient nephrovascular counterflow system and the large solute and/or hydraulic
permeabilities of the various structures. For the ascending limb to supply solute in excess of water it is essential that it be able to maintain a relatively hypotonic fluid. This requires that the ascending limb both be relatively water impermeable and have net outward total solute transport. In the outer medulla this is accomplished by active salt transport that exceeds backleak of salt. In the inner medulla the role of active transport is moot, but urea cycling in the concentrating kidney depresses the relative salt concentration in the core and either decreases or reverses the backleak. In either case, an increment in solute transport out of ascending limb occurs and augments the concentration gradient generated along the medulla. These qualitative features of the concentrating mechanism and the functional role of the loop of Henle seem established, but a detailed correlation of the transport characteristics of the loop of Henle with adaptive and homeostatic mechanisms has not yet been carried out. Such a correlation requires both more data and more detailed modeling.
References
1. Wirz, H., and R. Dirix. 1973. Urinary concentration and dilution. In: Handbook of Physiology, Section 8: Renal Physiology. J. Orloff and R. Berliner, eds. Am. Physiol. Soc. Washington, D.C. pp. 415-430.
2. Marsh, D. 1971. Osmotic concentration and dilution of the urine. In: The Kidney: Morphology, Biochemistry, Physiology, Vol. III. C. Rouiller and A. Muller, eds. Academic Press, New York. pp. 71-126.
3. Jamison, R. 1974. Countercurrent systems. In: MTP International Reviews of Science, Vol. 6: Kidney and Urinary Tract Physiology. K. Thurau, eds. Butterworth, London. pp. 199-245.
4. Berliner, R. W. 1976. The concentrating mechanism in the renal medulla. Kidney Int. 9:214-222.
5. Schmidt-Nielson, G., and R. O'Dell. 1961. Structure and concentrating mechanism in the mammalian kidney. Am. J. Physiol. 200:1119-1124.
6. Peter, I. 1909. Untersuchungen uber Bau und Entwicklung der Niere. Fischer, Jena.
7. Schwartz, M., and M. Venkatachalam. 1974. Structural differences in thin limbs of Henle: Physiological implications. Kidney Int. 6:103-208.
8. Hai, M., and S. Thomas. 1969. Influence of pre-hydration on the changes in renal tissue composition induced by water diuresis in the rat. J. Physiol. 205:599-618.
9. Crawford, J. A. Doyle, and J. Probst. 1959. Service of urea in water conservation. Am. J. Physiol. 196:545-548.
10. Schutz, W., and J. Schnermann. 1972. Pelvic urine

CHAPTER 33 • TRANSPORT CHARACTERISTICS OF LOOP OF HENLE 679
composition as a determinant of inner medullary solute concentration and urine osmolarity. Pfluegers Arch. 334:154-166.
11. Gottschalk, C., and M. Mylle. 1959. Micropuncture study of the mammalian urinary concentrating mechanism: Evidence for the countercurrent hypothesis. Am. J. Physiol. 196:927-936.
12. Pennell, J., V. Sanjana, N. Frey, and C. Jamison. 1975. The effect of urea infusion on the urinary concentrating mechanism in protein-depleted rats. J. Clin. Invest. 55:399-409.
13. Imbert, M., and C. de Rouffignac. 1976. Role of sodium and urea in the renal concentrating mechanism in Psammomys obesus. Pfluegers Arch. 36:107-114.
14. Stephenson, J. L. 1972. Concentration of urine in a central core model of the renal counterflow system. Kidney Int. 2:85-94.
15. Kokko, J. P., and F. C. Rector. 1972. Countercurrent multiplication system without active transport in inner medulla. Kidney Int. 2:214-223.
16. Jamison, R. 1968. Micropuncture study of segments of thin loop of Henle in the rat. Am. J. Physiol. 215:236-242.
17. Marsh, D., and S. Azen. 1975. Mechanism of NaCI reabsorption by hamster thin ascending limbs of Henle's loop. Am. J. Physiol. 228:71-79.
18. Imai, M., and J. Kokko. 1974. Sodium, urea, and water transport in the thin ascending limb of Henle. Generation of osmotic gradients by passive diffusion of solutes. J. CUn Invest. 53:393-402.
19. Burg, M., and N. Green. 1973. Function of the thick ascending limb of Henle's loop. Am. J. Physiol. 224:659-668.
20. Rocha, A. S., and J. P. Kokko. 1973. Sodium chloride and water transport in the medullary thick ascending limb of Henle. Evidence for active chloride transport. J. Clin Inves. 52:612-623.
21. Lassiter, W., C. Gottschalk, and M. Mylle. 1964. Net trans tubular movement of water and urea in saline diuresis. Am. J. Physiol. 204:669-673.
22. Kokko, J. 1970. Sodium chloride and water transport in the descending limh of Henle. J. Clin. Invest. 49: 1838-1846.
23. Pennel, J., F. Lacy, and R. Jamison. 1974. An in vivo study of the concentrating process in the descending limb of Henle's loop. Kidney Int. 5:337-347.
24. Rouffignac, C. de. 1972. Physiological role of Henle in urinary concentration. Kidney Int. 2:297-303.
25. Kawamura, S., and J. Kokko. 1976. Urea secretion by the straight segment of the proximal tubules. J. CUn. Invest. 58:604-612.
26. Smith, H. W. 1956. Principles of Renal Physiology. Oxford Univ. Press, London and New York.
27. Kuhn, W., and K. Ryffel. 1942. Herstellung Konzentrierter Losungen aus verdunnten durch blosse membranwirkung. Ein modellversuch zur funktion der niere. Hoppe Seylers Z. Physiol. Chem. 276:145-178.
28. Hargitay, B., and W. Kuhn. 1951. Das Multiplikationsprinzip als Grundlage der Hamkonzentrierung in der Niere. Z. Electrochem. Angew. Phys. Chem. 55:539-558.
29. Kuhn, W., and A. Ramel. 1959. Zweierlei Gleichgewichtspotentiale an ionendurchlassigen, ionen-intranichtpermutierenden Membranen (Ruhe- und Aktionspotential). Helv. Chim. Acta 42:293-305.
30. Kuhn, W., and A. Ramel. 1959. Aktiver Salztransport als moglicher (und wahrscheinlicher) Einzeleffekt bei der Hamkonzentrierung in der Niere. Helv. Chim. Acta 42:628-660.
31. Wirz, H. 1953. Der osmotische Druck des Blutes in der Nierenpapille. Helv. Physiol. Acta 11:20-29.
32. Berliner, R. W., N. G. Levinsky, D. G. Davidson, and M. Eden. 1958. Dilution and concentration of the urine and the action of antidiuretic hormone. Am. J. Med. 24:730-744.
33. Ullrich, K. J., K. Kramer, and J. Boylan. 1%2. Present knowledge of the countercurrent system in the mammalian kidney. In: Heart, Kidney and Electrolytes. C. K. Friedberg, ed. Grune & Stratton, New York. pp. 1-37.
34. Stephenson, J. L. 1973. Concentrating engines and the kidney. 1. Central core model of the renal medulla. Biophys. J. 13:512-545.
35. Stephenson, J. L. 1973. Concentrating engines and the kidney. II. Multisolute central core systems. Biophys. J. 13:545-567.
36. Stephenson, J. L. 1976. Concentrating engines and the kidney. III. Canonical mass balance equation for multinephron medels of the renal medulla. Biophys. J. 16:1273-1286.
37. Stephenson, J. L., R. P. Tewarson, and Raymond Mejia. 1974. Quantitative analysis of mass and energy balance in non-ideal models of the renal counterflow system. Proc. Natl. Acad. Sci. U.S.A. 71: 1618-1622.
38. Stephenson, J. L., R. Mejia, and R. P. Tewarson. 1976. Model of solute and water movement in the whole kidney. Proc. Natl. Acad. Sci. U.S.A. 73:252-256.
39. Rouffignac, C. de, and F. Morel. 1969. Micropuncture study of water, electrolytes, and urea movements along the loops of Henle in PSllmmomys. J. CUn. Invest. 48:474-486.