transmission electron microscopy studies of moraxella (branhamella) catarrhalis
TRANSCRIPT

IMMUNOLOGY AND MEDICAL MICROBIOLOGY
FEMS Immunology and Medical Microbiology 23 (1999) 57-66
Transmission electron microscopy studies of Moraxella (Branhamella) ca tarrhalis
Margaret Fitzgerald ‘, Riona Mulcahy b, Susan Murphy b, Conor Keane ‘, Davis Coakley b, Thomas Scott a,*
a Department of Biological Sciences, Dublin Institute of Technology, Kevin Street, Dublin 8. Ireland ’ Mercer’s Institute for Reseurch on Ageing. St James’s Hospitul. Dublin 8, Ireland
’ Department of Clinicnl Microbiology, St James’s Hospital, Dub&t 8. Ireland
Received 20 June 1998; received in revised form 23 October 1998; accepted 29 October 1998
Abstract
A trypsin-sensitive 200-kDa protein has been reported to be exclusively associated with haemagglutinating isolates of Moraxella (Branhamella) catarrhalis. Transmission electron microscopy studies revealed that haemagglutination by M. catarrhalis to both human and rabbit erythrocytes was mediated by a trypsin-sensitive outer fibrillar coat. This fibrillar layer was absent on non-haemagglutinating isolates examined. Immuno-electron microscopy, using a polyclonal antiserum containing antibodies to the 200-kDa protein as a probe, showed that the 200-kDa protein is present on the outer fibrillar layer of the bacterium. These findings suggest that the haemagglutinin of M. catarrhalis is a 200-kDa protein present on the outer fibrillar coat. 0 1999 IFederation of European Microbiological Societies. Published by Elsevier Science B.V. All rights reserved.
Kqvwords: Mornxella (Brmhamella) catarrhalis: Electron microscopy: Transmission electron microscopy; Immuno-electron microscopy; Adhesive interaction; Hemagglutination
1. Introduction
In recent years Moraxella (Branhamella) catarrha-
lis has been increasingly recognised as an important human pathogen, bei.ng previously dismissed as a commensal of the oro-nasopharyngeal tract [l]. In its pathogenic role, A4. catarrhalis mainly affects the respiratory tract, particularly the bronchopulmo- nary tree, middle ear and sinuses. After Haemophilus
influenzae and Streptococcus pneumoniae, M. catar-
rhalis is now reported as the third most common causative agent in lower respiratory tract infections
in the elderly [2-51 and in both acute otitis media [6-Q and sinusitis [9-121 in children. Sporadic cases of bacteraemia [ 131. meningitis [ 14,151, endocarditis [ 161, pericarditis [ 171, life-threatening tracheitis [ 181 and acute epiglottitis [19] due to the organism have also been reported.
The emergence of M. catarrhalis as an important human pathogen has prompted researchers to inves- tigate the virulence mechanisms utilised by the or- g&ism and therefore elucidate the pathogenesis of
M. catarrhalis infection. Bacterial adherence to mu- * Corresponding author. Tel.: +353 (1) 4024747;
Fax: +353 (1) 4024995; E-mail: [email protected]
0928-8244/99/$19.00 0 1999 Federation of European Microbiological Societies. Published by Elsevier Science B.V. All rights reserved
PII: SO928-8244(98)00121-7

58 M. Fitzggerald et al. I FEMS Inmunology und Medicul Microhiolug~ 33 (1999) 57-66
cosal surfaces is an important first step in the patho- genesis of many infectious diseases [20]. It has been found that M. caturrhulis adheres to human oro- pharyngeal cells and to erythrocytes. Studies have established that the haemagglutinin is a trypsin-sen- sitive protein [21,22] and is associated with tracheal cell adherence [21]. Two phenotypes of the haemag- glutinin have been identified, phenotype I isolates
agglutinate human erythrocytes only, while pheno- type II isolates agglutinate both human and rabbit
erythrocytes [22]. A 200-kDa protein has been found to be associated with haemagglutinating isolates of M. cutarrhulis, with evidence suggesting that this protein is a haemagglutinin of the organism [23].
2. Materials and methods
2.1. Bucteriul isolutes und growth conditions
A number of investigations have used electron mi- croscopy studies, in order to examine both the in vivo and in vitro expression of surface structures by M. cuturrhulis. Fimbriae have been demonstrated on iso- lates of A4. cuturrhulis. However, the role these fila-
mentous appendages play in the adherence of the organism to erythrocytes and human oropharyngeal
cells remains unresolved [24&26]. In a study of 24 fresh clinical isolates of M. caturrhulis by electron microscopy, it was observed that in strains with a higher haemagglutination titre the number of fim- briae was significantly (P < 0.04) more than in strains with a lower haemagglutination titre [26]. However, the findings of other investigations challenge the sig- nificance of the above observations, as no difference in haemagglutination capacity between fimbriate and non-fimbriate isolates of M. cutarrhulis has also been
reported [24]. Thus, it would appear that both fimb- rial and non-fimbrial adhesins are present on A4. cu-
tarrhalis isolates. To date, there are no studies on the
possible characteristics of the non-fimbrial adhesins of M. caturrhtrlis. In this present study transmission electron microscopy (TEM) studies were employed to elucidate further the nature of the interactions in- volved in haemagglutination by M. cutarrhulis.
Haemagglutinating (B4, K29, K38, K48, S407 and S580) and non-haemagglutinating (21 and K16) iso- lates of M. cuturrhalis were examined. Two of three haemagglutination phenotypes of isolate K48 (W and Y) were used in this study, these phenotypes
were described in a previous study [23]. All the M. cuturrhalis isolates were obtained from a collection acquired during a study of the epidemiological as- pects of M. caturrhulis infection and colonisation [27]. Isolates were identified according to the criteria previously outlined [28] and were stored at -70°C using the Protect Bead System (Technical Service Consultants). Prior to all tests isolates were grown on Columbia blood agar at 37°C for 18 h.
2.2. TEM studies of bucteriu
Bacterial suspensions of three haemagglutinating isolates (K29, K48 and S407) and one non-haemag- glutinating isolate (21) were examined by TEM. The suspensions consisting of 1.5 X log cfu ml-’ (equiva- lent to a MacFarland No. 5 turbidity standard) were prepared in l-ml amounts of sterile distilled water and were washed twice. The bacterial pellets were fixed as previously described [29].
Isolates (21, K29, K48 and S407) of M. caturrhalis were treated with 2 mg trypsin ml-’ (Sigma, Cata-
logue No. T 8003) and isolate K29 was also treated with 2 mg pronase ml-’ (Sigma, Catalogue No. P 6911), at 37°C for 2 h, prior to TEM. Following
washing, the pellets were prepared and fixed for TEM as previously described [29]. The sections were cut on a LKB Bromma Ultratome III and ex- amined in an Hitachi H-7100 electron microscope.
1.1. Aims of stud2 2.3. Trunsniission electron microscopq~ qf
huernuggh~tir~ation
The aims of this study were: (i) to determine the The nature of the interactions occurring when M. nature of the interactions between M. caturrhulis and cutarrhalis adheres to both human and rabbit eryth- both human and rabbit erythrocytes by TEM, and rocytes was examined by TEM. Phenotype 1 isolates (ii) to use immuno-electron microscopy to identify S407, B4, K29, and phenotype 11 isolates K38 and the structures on M. cuturrhulis which are associated S580 which were haemagglutinating with human with the 200-kDa protein. erythrocytes were examined by TEM. Additionally,
.M Fitzgerald et al. I FEMS 1mmunolog.v and Medical Microbiology 23 (1999) 57-66 59
phenotype II isolates (K38 and SSSO) which were
haemagglutinating with rabbit erythrocytes were also examined by TEM. A bacterial suspension (500 pl of 5 x 1O’O cfu ml-‘) was added to 500 pl of a 3% (v/v) suspension of human group 0 eryth- rocytes. The mixture was allowed stand at room tem- perature for 5 min. The agglutinated cells were pel-
leted at 200 xg for 2 min. The deposit was fixed as previously described [29].
2.4. Immune-electron microscopy
Bacterial cells of Al. catarrhalis (isolates 21, B4, K16, K48 [phenotypes W and Y], S109, S407 and SSSO) from overnight cultures on Columbia blood agar plates were suspended in Dulbecco A PBS (PBS-A) at a concentration of 1.5 X log cfu ml-’ (equivalent to a Macl’arland No, 5 turbidity stand- ard). Formvar-coated 200-mesh grids were floated
film side down on these bacterial suspensions for 60 s and blotted dry. The preparations were blocked for 15 min in 2% (w/v) skim milk (Difco) prepared in
PBS-A, followed by five 1-min washes in PBS-A. The preparations were incubated with anti-S407 antise- ru m. diluted 1 in 500 in PBS-A for 30 min. This
B
antiserum had been raised to a protein extract of
M. cutarrhalis and had been adsorbed with non-hae- magglutinating isolates of M. catarrhalis [23]. Fol- lowing incubation of the grid preparations with this antiserum, unbound antibodies were removed with five 1-min washes in PBS-A. This was followed by incubation for 30 min with gold (10 nm)-conju- gated goat anti-rabbit IgG (Sigma), diluted 1 in 20 in PBS-A. Unbound gold conjugate was removed by four 1-min washes in PBS-A, followed by two
1-min washes in distilled water. The grids were blot- ted dry. Some grids were stained for 30 s using 1% sodium silicotungstate pH 7.1 (prepared in distilled
water) and blotted dry. Preparations were viewed using an Hitachi H-7000 transmission electron mi- croscope.
3. Results
3.1. TEA4 observations of bacteria
TEM studies were employed to investigate the sur-
face structures on the haemagglutinating and non- haemagglutinating isolates of M. catarrhalis. On ex-
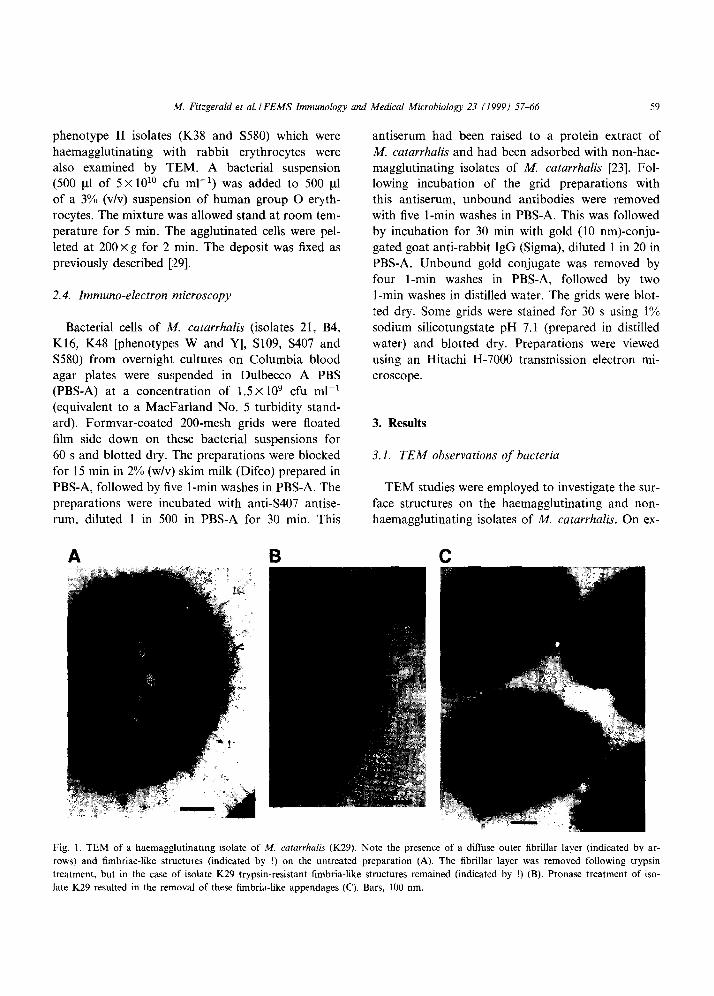
Fig. 1. TEM of a haemagglutinating isolate of M. catarrhalis (K29). Note the presence of a diffuse outer fibrillar layer (indicated by ar-
rows) and fimbriae-like structures (indicated by !) on the untreated preparation (A). The fibrillar layer was removed following trypsin
treatment, but in the case of isolate K29 trypsin-resistant fimbria-like structures remained (indicated by !) (B). Pronase treatment of iso-
late K29 resulted in the removal of these fimbria-like appendages (C). Bars, 100 nm.

60 M. Fitzgerald et al. IFEMS Immunology and Medical Microbiology 23 (1999) 5746
amination of the haemagglutinating isolates (pheno- type I: K29, S407; phenotype II: K48) by TEM and irrespective of haemagglutination phenotype, a dif- fuse outer fibrillar layer was observed on these prep- arations. In addition to this fibrillar layer isolate K29 also exhibited long strands protruding from the sur- face of the cells, which resembled fimbriae (Fig. 1A). When the three haemagglutinating isolates were
treated with pronase and trypsin, the outer fibrillar layer was removed. However, in the case of isolate K29, the fimbrial-like structures remained following trypsin treatment (Fig. 1B). Pronase treatment of K29 resulted in the removal both of these fimbria- like structures and the outer fibrillar layer (Fig. 1C). Haemagglutinating activity of isolate K29 was also lost following pronase treatment.
The outer fibrillar layer observed on the haemag- glutinating isolates of A4. catarrhalis was absent when the non-haemagglutinating isolate (21) was ex- amined by TEM (Fig. 2). However, spicule-like
structures were observed on the surface of this iso- late. Evidence suggests that these structures may play a role in the adherence of the organism to HEp-2 cells (M. Fitzgerald, unpublished observations).
3.2. TEM of haemagglutination
Fig. 2. TEM of a non-haemagglutinating isolate of M. catarrhulis
(21). Note the absence of the diffuse outer fibrillar type coat. but
the presence of spicule-like structures on the cell surface (indi-
cated by arrows). Bar. 100 nm.
TEM studies were also employed to investigate the nature of the interactions occurring when M. catar-
rhalis adheres to both human and rabbit erythro-
cytes. Adherence of isolate S580 to both human and
rabbit erythrocytes appeared to involve the diffuse outer fibrillar layer, with relatively close cell-surface interactions occurring. Fimbrial involvement was not apparent (Fig. 3A,B).
compared it was observed that the outer fibrillar coat on isolate K29 was removed following trypsin treat- ment, while the fimbria-like structure were retained.
These observations are in agreement with the obser- vations made on examining Fig. lA,B.
3.3. Immuno-electron microscopy
Adherence of isolate K29 to human erythrocytes The antiserum to the protein extract of M. catar-
also involved the outer fibrillar layer. An extremely rhalis (isolate S407) from which antibodies other
dense layer was observed in the haemagglutinating than those to the 200-kDa protein had been ad-
bacteria of this isolate and this layer appeared to be sorbed [23] was used in immuno-electron microscopy
involved in the attachment of the organism to hu- to pinpoint where on the bacterial cell this protein
man erythrocytes (Fig. 4A). Haemagglutination by lies. Immuno-electron microscopy demonstrated that
K29 was not abolished following trypsin treatment gold particles attached to the bacterial cell surface
[22] and when this interaction was examined by (isolate S407) and negative staining of this prepara-
TEM, the trypsin-resistant fimbria-like structures tion definitively illustrated that this attachment was
commented on above, appeared to mediate attach- to the outer fibrillar layer of M. catarrhalis (Fig. 5). ment of the trypsin-treated isolate K29 to human Very weak reactions were observed when isolate erythrocytes (Fig. 4B). When Figs. 4A and 4B were S407 was treated with antiserum from which anti-

M. Fitzgerald et al. I FEMS Immunology and Medical Microbiology 23 (1999) 5746 61
Fig. 3. TEM of A4 catarrhalis isolate S580 adhering to human erythrocytes (A) and rabbit erythrocytes (B). Note the involvement of the
bacterium’s diffuse outer fibrillar coat in the attachment of the organism to both erythrocyte types, indicated by arrows. Bars. 100 nm.
bodies specific for the 200-kDa protein had been adsorbed. Additionally, when the adsorbed antise- rum which contained antibodies to the 200-kDa pro- tein was examined with non-haemagglutinating iso- lates (21, K16) by immuno-electron microscopy, very weak or negligible i:nteractions were observed (Fig. 6) emphasising yet iagain the specificity of the 200- kDa protein for haemagglutinating isolates of M.
catarrhalis.
4. Discussion
To support furthler the association between the presence of the 200,-kDa protein and positive hae- magglutination, two haemagglutination phenotypes of isolate K48 (phenotypes W and Y) were probed with the adsorbed antiserum, which contained anti- bodies to the 200-ItDa protein and examined by immuno-electron m:icroscopy. Many gold particles were observed attaching to the strongly haemagglu- tinating phenotype W isolate, but the number of gold particles attaching to the weakly haemagglu-
tinating phenotype Y isolate was reduced (Fig. 7A,B).
In the present study, the TEM examination of M. catarrhalis adhering to both human and rabbit eryth- rocytes revealed that attachment was mediated by a diffuse outer fibrillar coat. In the 1974 edition of
Bergey’s Manual of Determinative Bacteriology, this outer fibrillar layer-like structure was described
as a capsule [30]. The outer fibrillar layer was evident on all the haemagglutinating isolates examined but was absent in the non-haemagglutinating isolates. Trypsin treatment of bacteria prior to TEM exami- nation resulted in the removal of the outer fibrillar layer on haemagglutinating isolates of M. catarrha-
lis, indicating that this structure is protein in nature. Trypsin treatment of haemagglutinating isolates of A4. catarrhalis had previously been shown to render the bacterial cells non-haemagglutinating [21,22]. Thus, the removal of the outer fibrillar layer follow- ing trypsin treatment provides further evidence of its role in haemagglutination by M. catarrhalis.
The authors wish. to stress that the figures pre- Fimbrial involvement was not evident in either the
sented in this article are typical representatives of M. catarrhalis-human erythrocyte interactions (ex- all the fields of view examined in the TEM studies ception isolate K29) or in the M. catarrhalis-rabbit
undertaken. erythrocyte interactions. Currently, there is much un-

62 M. Fikgernld et al. I FEMS Immunology and Medical Microbiology 23 (1999) 5746

M. Fitzgerald et al. I FEMS Immunology and Medical Microbiology 23 (19991 57-66 63
Fig. 4. TEM of M. catarrbalis isolate K29 adhering to human erythrocytes. A distinctive dense outer fibrillar layer present on the un-
treated bacteria appeared 1.0 be involved in adherence, indicated by arrows (A). This layer was dramatically reduced following trypsin
treatment, but instead the trypsin-resistant fimbria-like structures appeared to mediate attachment to human erythrocytes, indicated by ar-
rows (B). Bars, 100 nm. t
certainty regarding thle role fimbriae play in haemag- glutination by M. catarrhalis. In one study, it was observed that strains with a high haemagglutination titre were associated with more fimbriae than strains with a low titre [26]. In that investigation, it was noted that continuous subculture of fresh clinical isolates reduced the level of fimbriation. However, the author failed to comment on the haemagglutina-
tion status of these isolates following continuous subculture. In the present study fresh clinical isolates
were not used and, yet, no reduction in haemagglu- tinating activity following repeated subculture was
observed. Additionally, in a separate study, it was observed that all fimbriated strains did not induce haemagglutination [2.4]. Thus, it is possible that both fimbrial and non-fimbrial haemagglutinins
Fig. 5. Immune-electron microscopy of M. catarrhalis (S407)
shows gold particles attaching to the outer fibrillar layer. Isolate
S407 had been incubated in antiserum (l/500), from which anti-
bodies other than those recognising epitopes on the 200-kDa pro-
tein had been adsorbed. The preparation was then probed with
gold-labelled goat anti-rabbit-gold IgG conjugate (10 nm), fol-
lowed by negative staining with 1% sodium silicotungstate. Bar.
100 nm.
may be found on M. catarrhalis. In the present in- vestigation, TEM observations of isolate K29 lend
support to this hypothesis. It had been previously established that trypsin pre-treatment of isolate K29 was ineffective in abolishing haemagglutination,
whereas pronase treatment of the isolate did abolish haemagglutination [22]. TEM studies of K29 re- vealed that haemagglutination with human erythro- cytes was mediated by both a diffuse outer fibrillar layer and trypsin-resistant fimbria-like structures, thereby supporting the hypothesis that both fimbrial
and non-fimbrial haemagglutinins play a role in hae- magglutination by M. catarrhalis.
A previous study found that a 200-kDa trypsin- sensitive protein was associated exclusively with hae- magglutinating isolates of M. catarrhalis [23]. Using the polyclonal antiserum from which antibodies oth- er than those recognising epitopes on the 200-kDa protein had been adsorbed, it was established by
Fig. 6. Immune-electron microscopy of M. catarrhalis (21) shows
very few gold particles attaching to the outer layer of the bacte-
rium. The non-haemagglutinating isolate (21) had been incubated
in adsorbed antiserum which contained antibodies to the 200-
kDa protein (l/500). followed by probing with gold-labelled goat
anti-rabbit IgG conjugate (10 nm). Bar. 100 nm.

64
A
M. Fitzgerald et al. I FEMS Immunology and Medical Microbiology 23 (1999) 5746
Fig. 7. Immune-electron microscopy of haemagglutinating phenotypes W and Y of isolate K48. Both phenotypes had been incubated in
the adsorbed antiserum, from which antibodies other than those recognising epitopes on the 200-kDa protein (l/500) had been removed.
The preparation was then probed with gold&belled goat anti-rabbit IgG conjugate (10 nm). Note the presence of many gold particles at-
taching to the outer fibrillar layer extending outwards from the cell surface of the strongly haemagglutinating phenotype W (A). The level
of gold particle attachment was reduced in the weakly haemagglutinating phenotype Y (B). Bars, 100 nm.
immunoblot analysis that the 200-kDa protein was surface expressed 1231. In this present study immuno- electron microscopy utilising this antiserum estab-
lished that the 200-kDa protein lies in the outer fi- brillar layer of M. catarrhalis.
The former observation once more highlights the strong association between the 200-kDa protein and the presence of haemagglutinins on M. catarrhafis.
Earlier in this study it had been shown that the outer fibrillar layer present on haemagglutinating isolates of A4. catarrhalis was involved in the M. catarrhalis-
erythrocyte interactions. The presence of the 200- kDa protein in this outer layer has now been dem-
onstrated. Furthermore, it was found following immuno-elec-
tron microscopy that numerous gold particles were seen to be attached to the outer fibrillar layer of the strongly haemagglutinating phenotype W of isolate K48, thereby showing the position and presence of the 200-kDa protein on this isolate. The number of gold particles seen attaching to the weakly haemag- glutinating phenotype Y was reduced. This indicated that expression of the 200-kDa protein in phenotype Y was reduced in comparison to phenotype W. This reduced expression of the 200-kDa protein in pheno-
type Y corresponded with a reduced haemagglutina- tion titre. Consequently, these observations lend yet further support for the hypothesis that the 200-kDa
protein is a haemagglutinin or part of the haemag- glutinin protein.
In conclusion, attachment of AL catarrhalis to both human and rabbit erythrocytes was mediated by a trypsin-sensitive outer fibrillar layer. Immuno- electron microscopy studies revealed that the 200- kDa trypsin-sensitive haemagglutination-associated protein lies in the outer fibrillar layer of A4. catarrha-
lis. Additionally, trypsin-resistant fimbria-like struc- tures present on isolate K29 also appeared to mediate attachment of this bacterium to human erythrocytes.
Thus, haemagglutination by A4. catarrhalis appears to be mediated mainly by fibrillar structures, however fimbriae also appear to have a role. The elucidation of the fibrillar versus the fimbrial role in haemagglu- tination by M. catarrhalis requires further work. In this study different isolates were examined to charac- terise the haemagglutinin of M. catarrhalis. However, it would be appropriate to use isogenic variants of the same strain to further clarify the relative roles of the fibrillar and fimbrial structures in haemagglutina- tion by this microorganism.

M Fitzgerald et al. I FEMS Immunology and Medical Microbiology 23 (1999) 5746 65
4. I. Conclusions
TEM studies revealled that haemagglutination of
human and rabbit erythrocytes by M. catarrhalis iso- lates was mediated by a trypsin-sensitive outer fibril-
lar coat. Immuno-electron microscopy studies indi- cated that the 2.00-kDa haemagglutination- associated-protein was present on this outer fibrillar coat. However, in one isolate of M. catarrhalis it was established that in addition to fibrillar mediated hae- magglutination, fimbrial mediated haemagglutina-
tion was also observed. This finding suggests that both fibrillar and fimbrial structures are involved in haemagglutination by M. catarrhalis.
Acknowledgments
We wish to thank MS Ann Mynes, Electron Mi- croscopy Unit, St James’s Hospital, Dublin and Mr David John, The Electron Microscopy Unit, Trinity College, Dublin for their help with the electron mi- croscopy work. This work was supported by a Health Research Board of Ireland grant to M.F.
References
[l] Catlin, B.W. (1990) Branhamella catarrhalis; an organism
gaining respect as a pathogen. Clin. Microbial. Rev. 3, 293-
320.
[2] Ninane. G., Joly, J. and Kraytman, M. (1978) Bronchopulmo-
nary infection due to Branhamella catarrhalis: 11 cases as-
sessed by transtracheal puncture. Br. Med. J. 1, 276-278.
[3] Mannion, P.T. (1987) Sputum microbiology in a district gen-
eral hospital. The role of Branhamella catarrhalis. Br. J. Dis.
Chest 81. 391-396.
[4] Davies, B.I. and Maesen, F.P.V. (1988) The epidemiology of
respiratory tract pathogens in Southern Netherlands. Eur.
Respir. J. 1, 415420.
[5] Carr. B., Walsh, J.B., Coakley, D.. Mulvihill, E. and Keane,
C. (1991) Prospective hospital study of community acquired
lower respiratory tract infection in the elderly. Respir. Med.
85, 1855187.
[6] van Hare, G.F.. Shurin, P.A.. Marchant, C.D., Cartelli. N.A.,
Johnson, C.E.. Futton, D.. Carlin, S. and Kim, C.H. (1987)
Acute otitis media caused by Branhamella catarrhalis: biology
and therapy. Rev. Infect. Dis. 9, 1627.
[7] Ruuskanen, O., Arola, M.. Putto-Laurila, A.. Mertsola. J.,
Meurman, O., Viljanen. M.K. and Halonen, P. (1989) Acute
otitis media and respiratory virus infections. Pediatr. Infect.
Dis. J. 8, 94-99.
[8] Pichichero, M.E. and Pichichero, CL. (1995) Persistent acute
otitis media: causative pathogens. Pediatr. Infect, Dis. J. 14.
178-183.
[9] Wald, E.R., Milmoe, G.J., Bowen. A.D., Ledesma-Medina, J.,
Salamon, N. and Bluestone, C.D. (1981) Acute maxillary si-
nusitis in children. New Engl. J. Med. 304, 749-754.
[lo] Tinkelman. D.G. and Silk. H.J. (1989) Clinical and bacterio-
logic features of chronic sinusitis in children. Am. J. Dis.
Child. 143, 938941.
[ll] Wald. E.R., Byers, C.. Guerra. N., Casselbrant, M. and Beste,
D. (1989) Subacute sinusitis in children. J. Pediatr. 115, 28-32.
[12] van Cauwenberge, P.B., Vander Mijnsbrugge, A.-M. and In-
gels, K.J.A.O. (1993) The microbiology of acute and chronic
sinusitis and otitis media: a review. Eur. Arch. Otorhinolar-
yngol. 250 (Suppl. l), S3-S6.
[13] Ioannidis, J.P.A., Worthington, M., Griffiths, J.K. and Snyd-
man, D.R. (1995) Spectrum and significance of bacteremia
due to Moraxella catarrhalis. Clin. Infect. Dis. 21, 39&397.
[14] O’Neill, J.H. and Mathieson, P.W. (1987) Meningitis due to
Branhamella catarrhalis. Amt. NZ J. Med. 17, 241-242.
[15] Daoud, A., Abuekteish, F. and Masaadeh, H. (1996) Neona-
tal meningitis due to Moraxella catarrhalis and review of the
literature. Ann. Trop. Paediatr. 16. 1999201.
[16] Turner. H.R.. Taylor, M.R. and Lockwood, W.R. (1985)
Branhamella catarrhalis endocarditis in a patient receiving he-
modialysis. South. Med. J. 78, 1021-1022.
[17] Kostiala. A.A.I. and Honkanen, T. (1989) Branhamella catar-
rhalis as a cause of acute purulent pericarditis. J. Infect. 19,
291-292.
[18] Bodkin, S. and Warde, D. (1993) Moraxella catarrhalis an
unusual pathogen in bacterial tracheitis. Ir. Med. J. 86. 208-
209.
[19] Vernham. G.A. and Crowther, J.A. (1993) Acute myeloid leu-
kaemia presenting with acute Branhamella catarrhalis epiglot-
titis. J. Infect. 26, 93-95.
[20] Beachey, E.H. (1981) Bacterial adherence: adhesin-receptor
interactions mediating the attachment of bacteria to mucosal
surfaces. J. Infect. Dis. 143, 3255345.
[21] Kellens. J., Persoons, M.. Vaneechoutte, M.. van Tiel, F. and
Stobberingh, E. (1995) Evidence for lectin-mediated adherence
of Moraxella catarrhalis. Infection 23, 3741.
[22] Fitzgerald, M., Murphy. S.. Mulcahy, R.. Keane, C., Coakley,
D. and Scott, T. (1996) Haemagglutination properties of Mor-
axella (Branhamella) catarrhalis. Br. J. Biomed. Sci. 53. 257-
262.
[23] Fitzgerald, M.. Mulcahy. R.. Murphy, S.. Keane. C., Coakley.
D. and Scott, T. (1997) A 200 kDa protein is associated with
haemagglutinating isolates of Moraxella (Branhamella) catar-
rhalis. FEMS Immunol. Med. Microbial. 18, 209-216.
[24] Rikitomi, N., Andersson, B., Matsumoto. K., Lindstedt. R.
and Svanborg. C. (1991) Mechanism of adherence of Morax-
eliu (Branhamella) catarrhalis. Stand. J. Infect. Dis. 23. 559-
567.
[25] Ahmed, K. (1992) Fimbriae of Branhamellu catarrhalis as pos-
sible mediators of adherence to pharyngeal epithelial cells.
APMIS 100, 1066-1072.
[26] Ahmed, K., Rikitomi, N. and Matsumoto, K. (1992) Fimbria-

66 M. Fitzgerald et al. I FEMS Immunology and Medical Microbiology 23 (I 999) 5746
tion, haemagglutination and adherence properties of fresh
clinical isolates of Branhantelia catarrhalis. Microbial. Immu-
nol. 36, 1009-1017.
[27] Murphy, S. (1996) Epidemiological, Microbiological and Im-
munological Aspects of M. catarrhalis Infection and Coloni-
sation. Thesis submitted to the University of Dublin for the
award of Doctor of Medicine.
[28] Doern. G.V. and Morse, S.A. (1980) Bran/msella jllieisseria)
cararrhalis: criteria for laboratory identification. J. Clin. Mi-
crobiol. 11. 193-195.
[29] Scott, T.G., Curran, B. and Smyth, C.J. (1989) Electron mi-
croscopy of adhesive interactions between Gardner& vagina-
lis and vaginal epithelial cells, McCoy cells and human red
blood cells. J. Gen. Microbial. 135, 475480.
[30] Reyn, A. (1974) Family I. Neimeriaceae Prevot 1933. In: Ber- gey’s Manual of Determinative Bacteriology. 8th edn. (Bu-
chanan. R.E. and Gibbons, N.E.. Eds.), pp. 427-443. Wil- liams and Wilkins, Baltimore, MD.