transmembrane signaling in eukaryotes fre- quently ... filecopyright © 1998 by the genetics society...
TRANSCRIPT

Copyright © 1998 by the Genetics Society of America
Genetics
148:
33–47 (January, 1998)
An Essential Function of a Phosphoinositide-Specific Phospholipase CIs Relieved by Inhibition of a Cyclin-Dependent Protein Kinase
in the Yeast
Saccharomyces cerevisiae
Jeffrey S. Flick* and Jeremy Thorner
†
*
Department of Biochemistry, Vanderbilt University School of Medicine, Nashville, Tennessee 37232-0146 and
†
Department of Molecularand Cell Biology, Division of Biochemistry and Molecular Biology, University of California, Berkeley, California 94720-3202
Manuscript received July 24, 1997Accepted for publication September 19, 1997
ABSTRACTThe
PLC1
gene product of
Saccharomyces cerevisiae
is a homolog of the
d
isoform of mammalian phospho-inositide-specific phospholipase C (PI-PLC). We found that two genes (
SPL1
and
SPL2
), when overex-pressed, can bypass the temperature-sensitive growth defect of a
plc1
D
cell.
SPL1
is identical to the
PHO81
gene, which encodes an inhibitor of a cyclin (Pho80p)-dependent protein kinase (Pho85p) complex(Cdk). In addition to overproduction of Pho81p, two other conditions that inactivate this Cdk, a cyclin(
pho80
D
) mutation and growth on low-phosphate medium, also permitted growth of
plc1
D
cells at the re-strictive temperature. Suppression of the temperature sensitivity of
plc1
D
cells by
pho80
D
does not dependupon the Pho4p transcriptional regulator, the only known substrate of the Pho80p/Pho85p Cdk. The sec-ond suppressor,
SPL2
, encodes a small (17-kD) protein that bears similarity to the ankyrin repeat regionspresent in Pho81p and in other known Cdk inhibitors. Both
pho81
D
and
spl2
D
show a synthetic phenotypein combination with
plc1
D
. Unlike single mutants,
plc1
D
pho81
D
and
plc1
D
spl2
D
double mutants were un-able to grow on synthetic complete medium, but were able to grow on rich medium.
Cellular responses after PIP
2
hydrolysis and produc-tion of IP
3
and DAG depend on the cell type and in-clude proliferation, differentiation, and secretion (
Ber-
ridge
1993;
Valius
and
Kazlauskas
1993;
Weiss
and
Littman
1994). In mammalian cells, IP
3
binds to intra-cellular receptors, stimulating the release of sequesteredCa
2
1
, thereby activating Ca
2
1
- and calmodulin-regu-lated protein kinases and phosphoprotein phosphatases(
Clapham
1995). DAG remains in the membrane,where it can activate members of the protein kinase C(PKC) family (
Nishizuka
1992). Both second messen-ger pathways can elicit changes in gene expression(
Clipstone
and
Crabtree
1992;
Franz
et al.
1994;
Hill
and
Treisman
1995;
O’Keefe
et al.
1992).In animal cells, the role of any given PI-PLC isoform
is difficult to assess because of the multiplicity of PI-PLC isotypes present. Moreover, the precise function ofPIP
2
turnover in any given cellular response after re-ceptor activation is complicated because of the simulta-neous recruitment of additional distinct signaling sys-tems, for example, the Ras- and PI-3 kinase–dependentpathways (
Valius
and
Kazlauskas
1993). Genetic ap-proaches have revealed cell type–specific requirementsfor particular PI-PLC isozymes. A Drosophila mutant(
norpA
) deficient in a PI-PLC
b
4 isoform lacks light-stimulated membrane potential in its photoreceptorcells and is blind (
Bloomquist
et al.
1988), defining arole for PIP
2
mobilization by this PI-PLC
b
isoform ininsect phototransduction. Disruption of the mouse PI-
Corresponding author:
Jeffrey S. Flick, Department of Biochemistry,Vanderbilt University School of Medicine, Nashville, TN 37232-0146.E-mail: [email protected]
T
RANSMEMBRANE signaling in eukaryotes fre-quently involves activation of a phosphoinositide-
specific phospholipase C (PI-PLC), which hydrolyzesphosphatidylinositol 4,5-
bis
phosphate (PIP
2
) and gener-ates two second messengers, inositol 1,4,5-
tris
phosphate(IP
3
) and 1,2-diacylglycerol (DAG;
Lee
and
Rhee
1995).Three classes of PI-PLC (
b
,
g
, and
d
) have been charac-terized at the molecular level; each contains a con-served catalytic domain comprised of two segments,designated X and Y, as well as nonconserved segmentsthat confer distinct modes of regulation. Members ofthe PI-PLC
b
family can be stimulated by binding of theG
a
(
Lee
et al.
1992) and G
bg
(
Touhara
et al.
1994;
Wu
et al.
1993) subunits released upon activation of G pro-tein–coupled receptors. Stimulation of the PI-PLC
g
family depends on SH2 domains, which mediate inter-action with and phosphorylation by receptor-tyrosinekinases (
Koch
et al.
1991). In contrast, regulation ofthe PI-PLC
d
family is less well characterized, althoughit has been reported that a mammalian PI-PLC
d
can bestimulated
in vitro
by a GTPase-activating protein (GAP)for the small G protein Rho (
Homma
and
Emori
1995)and by interaction with a novel G
a
h
GTPase (
Feng
et al.
1996).

34 J. S. Flick and J. Thorner
PLC
g
1 gene results in embryonic lethality, indicatingan essential requirement for this isoform in multicellu-lar development (
Ji
et al.
1997). In contrast, loss of a PI-PLC
d
homolog in
Dictyostelium discoideum
did not resultin any apparent phenotype (
Drayer
et al.
1994); conse-quently, the role of phosphoinositide turnover by PI-PLC
d
in this organism remains unknown.As a means to elucidate cellular functions depen-
dent on the activity of a PI-PLC
d
, we have also under-taken a genetic analysis using the budding yeast
Saccha-romyces cerevisiae.
We (
Flick
and
Thorner
1993) andothers (
Payne
and
Fitzgerald-Hayes
1993;
Yoko-o
etal.
1993) isolated a yeast gene,
PLC1
, that encodes a ho-molog of mammalian PI-PLC
d
isoforms. Moreover, we(
Flick
and
Thorner
1993) demonstrated that Plc1p isa PI-PLC, that its activity
in vitro
is Ca
2
1
-dependent andthat, as observed for the mammalian enzymes, its sub-strate selectivity is influenced by the concentration ofCa
2
1
. Yeast cells lacking Plc1p are viable, but growslowly at 30
8
(and below), and display several otherphenotypes, including temperature-sensitive lethality(at 34
8
and above;
Flick
and
Thorner
1993;
Payne
and Fitzgerald-Hayes 1993), sensitivity to hyperos-motic conditions (Flick and Thorner 1993), missegre-gation of chromosomes during mitosis (Payne andFitzgerald-Hayes 1993), and poor utilization of car-bon sources other than glucose (Flick and Thorner
1993; Payne and Fitzgerald-Hayes 1993). In at leastone genetic background, a plc1 null allele is lethal(Yoko-o et al. 1993).
As one approach for identifying the roles that Plc1pplays in cellular physiology, we isolated and character-ized two genes, which, when present in high-copy num-ber, bypass the requirement for Plc1p activity for
growth at 358. One of the genes, PHO81, is a criticalregulatory factor of the PHO regulon (Schneider et al.1994). The second gene, SPL2, encodes a novel proteinthat is regulated in part by the PHO regulon. In S. cerevi-siae, response to phosphate starvation involves thePHO81-dependent inhibition (Schneider et al. 1994)of a cyclin-dependent protein kinase (Cdk) comprisedof the PHO80 and PHO85 gene products (Kaffman etal. 1994; Santos et al. 1995). Pho81p action reducesthe inhibitory phosphorylation of the Pho4p transcrip-tion factor, thereby allowing high-level expression ofgenes encoding secreted acid phosphatase (PHO5),other phosphatases, phosphate transporters, and otherproducts required for efficient phosphate assimilation(Johnston and Carlson 1992).
The findings presented here suggest, first, that Plc1p(or, more likely, the products of the reaction it cata-lyzes) acts to antagonize, directly or indirectly, eitherthe Pho80p/Pho85p Cdk or one of the downstreamsubstrates of this protein kinase. Second, our results in-dicate that the target of Plc1p and Pho80p/Pho85p ac-tion is a novel factor, not the Pho4p transactivator. Fi-nally, the genetic interactions we have uncoveredamong PLC1, SPL2, PHO81, and PHO80/PHO85 showthat the products of these genes participate in overlap-ping regulatory pathways necessary for adaptation tochanging nutrient and temperature conditions.
MATERIALS AND METHODS
Strains, growth conditions, and general methods: The yeaststrains used in this study are listed in Table 1. Strain construc-tion followed standard methods (Rose et al. 1990). Yeast wasgrown on agar plates containing either YPGlc medium (1%
TABLE 1
Yeast strains used in this study
Straina Genotypeb Source
YPH499 MATa Sikorski and Hieter 1989YPH500 MATa Sikorski and Hieter 1989YSS5 MATa pho80D::HIS3 Salama et al. 1994YJF32 MATa plc1D::HIS3 Flick and Thorner 1993YJF132 MATa plc1D::LEU2 Flick and Thorner 1993YJF133 MATa plc1D::LEU2 Flick and Thorner 1993YJF251 MATa plcD::LEU2 pho80D::HIS3 This workYJF252 MATa plc1D::LEU2 pho80D::HIS3 This workYJF277 MATa spl2D::HIS3 This workYJF306 MATa plc1D::LEU2 spl2D::HIS3 This workYJF386 MATa plc1D::LEU2 pho80D::HIS3 spl2D::HIS3 This workYJF552 MATa pho81D::TRP1 This workYJF555 MATa plc1D::HIS3 pho81D::TRP1 This workYJF567 MATa plc1D::LEU2 pho80D::HIS3 pho4D::TRP1 This workBJ3501 MATa pep4::HIS3 prb1-D1 can1 gal2 ADE2 LYS2 TRP1 LEU2 Jones 1991
a All strains are derived from YPH499 or YPH500, except for the protease-deficient strain BJ3501.b Except for strain BJ3501, all strains listed also carry the following mutations: ade2-101oc his3-D200 leu2-D1 lys2-801am trp1-D1 and
ura3-52.

Plc1p Function and the Pho80p/Pho85p Cdk in Yeast 35
yeast extract, 2% peptone, 2% glucose) or a synthetic com-plete medium (SCGlc) that contained (per liter): 1.7 g YeastNitrogen Base without amino acids or ammonium sulfate(Difco, Detroit, MI); 5 g (NH4)2SO4, 20 g glucose, 20 mg eachuracil and l-Arg, 30 mg each l-Tyr and l-Ile, 40 mg adeninesulfate, 50 mg l-Phe, 60 mg l-His, 100 mg each L-Glu andl-Asp, 150 mg each l-Met and l-Val, 200 mg of l-Thr, 260 mgl-Leu, and 400 mg l-Ser. Either uracil or leucine was omittedfrom SCGlc plates to provide selection for the maintenance ofplasmids. Phosphate-depleted YPGlc and SCGlc-Ura were pre-pared as described elsewhere (Bisson and Thorner 1982).Plasmids were introduced into yeast using the lithium acetatetransformation procedure in the presence of carrier DNA(Gietz and Schiestl 1991). Conventional methods were usedfor the construction and manipulation of all plasmids (Ausubel
et al. 1994); Escherichia coli DH5aF9 (GIBCO BRL, Bethesda,MD) was used for the propagation of all plasmids.
Selection of high-copy suppressors: Strain YJF32 (plc1D1::HIS3)was transformed with an S. cerevisiae genomic DNA library(Nasmyth and Tatchell 1980), carried in a 2-mm DNA vec-tor (YEp13), and plated on SCGlc-Leu plates. After incuba-tion at 238 for 12 hr, the plates were transferred to 358 for 5days. From z20,000 transformants, two colonies were ob-tained. Plasmids were rescued from these two yeast clones inbacteria (Robzyk and Kassir 1992) and were retested fortheir suppressor activity by transformation of YJF32 and exam-ination of the resulting transformants for their ability to growas isolated single colonies at 358. One plasmid, pJF115, wasable to suppress the temperature-sensitive phenotype of theplc1D cells and did not contain the PLC1 gene itself, as deter-mined by restriction endonuclease cleavage site analysis. Theinsert DNA in pJF115 was designated SPL1. The selection fordosage suppressors was repeated with a different recipientstrain, YJF132 (plc1D2::LEU2), and by using another S. cerevi-siae genomic library (Carlson and Botstein 1982) carriedon a different 2-mm vector (YEp24). From z18,000 transfor-mants, three colonies grew at 358. After recovery in bacteriaand retesting, all three plasmids were found to reproduciblyrescue the temperature-sensitive growth of YJF132 at 358. Asjudged by restriction enzyme analysis, one plasmid carried thePLC1 gene. The other two plasmids, pJF116 and pJF117, con-tained overlapping fragments of genomic DNA that were notthe same as either PLC1 or SPL1. The insert DNA in pJF116and pJF117 was designated SPL2. Both SPL1- and SPL2-con-taining plasmids were able to suppress the temperature-sensi-tive phenotype of yeast containing either the plc1D1::HIS3 orplc1D2::LEU2 null allele.
Plasmid constructions: The SPL1-containing insert in pJF115was z10 kb. Initial attempts to retrieve conveniently a func-tional subclone were unsuccessful. Therefore, restriction en-zyme digestion and deletion analysis (Sambrook et al. 1989)were used to localize the complementing region within the in-sert. The DNA sequence adjacent to a BamHI restriction sitethat fell within the region responsible for suppressor activitywas determined by standard methods (Biggin et al. 1983).The nucleotide sequence obtained was identical to that re-ported for the PHO81 gene (EMBL/GenBank accession num-ber D13228; Ogawa et al. 1993). On this basis, a 4.7-kb DNAfragment containing the entire PHO81 gene and its promoter(from the HindIII site at position 21011 to the AvaII site atposition 13726, relative to the first base of the ATG initiatorcodon of the PHO81 open reading frame) was inserted intothe HindIII and SmaI sites of the vector YEp351 (Hill et al.1986). The resulting plasmid was found to contain full SPL1suppressor activity and was designated pJF243.
A 1.8-kb BamHI-SalI fragment containing SPL2 suppressoractivity was subcloned from the insert in plasmid pJF117 andligated into the vector YEp351, which had been cleaved with
BamHI and SalI to yield plasmid pJF181. The complete nucle-otide sequence on both strands of this 1.8-kb genomic seg-ment was determined using a Sequenase kit (United StatesBiochemical, Cleveland, OH) and conditions recommendedby the supplier from a series of nested deletions that weregenerated by a procedure described previously (Henikoff
1984). DNA sequences were compiled and analyzed usingLasergene software (DNASTAR Inc., Madison, WI). Databaseswere searched using the BLAST algorithm (Altschul et al.1990).
To express the SPL2 coding sequence from the ADH1 pro-moter, a 0.5-kb NcoI-NdeI fragment from pJF181 (containingnucleotides 21 to 1499 relative to the first base of the ATG ofthe SPL2 open reading frame) was converted to blunt ends bytreatment with the Klenow fragment of Escherichia coli DNApolymerase I and all four deoxyribonucleotide triphosphates,and it was then inserted into the SmaI site of vector pAD4M(Martin et al. 1990). To create an in-frame fusion containinga 17-residue c-myc epitope (-IEEQKLISEEDLLRKRD-cooh;
Evan et al. 1985) attached to the C-terminal residue (codon148) of Spl2, a 1.2-kb EcoRV-BspHI fragment was excised fromplasmid pJF181, converted to blunt ends, and ligated into aderivative of pBluescript containing the Myc tag coding se-quence (pBS-MycTag; constructed by D. Ma) that had beendigested with EcoRI and treated with mung bean nuclease. A1.3-kb BamHI-SalI fragment (nucleotides 2725 to 1445 ofthe SPL2 coding region) containing the resulting SPL2-myc fu-sion was subcloned into BamHI- and SalI-digested YEp352(Hill et al. 1986), yielding plasmid pJF264. To create plas-mid pJF184, a 3.4-kb BamHI fragment containing the entirePLC1 gene (Flick and Thorner 1993) was ligated intoBamHI-digested YEp351.
Physical mapping: The SPL1 and SPL2 loci were assignedto their respective chromosomes by hybridization of appropri-ate 32P-labeled internal probes corresponding to each gene(3-kb BamHI-BamHI fragment of pJF115 and 3.5-kb BamHI-BamHI fragment of pJF116, respectively) to whole yeast chro-mosomes that had been separated by pulsed-field gel electro-phoresis (Chu et al. 1986) and transferred to a nylon membrane(a gift from G. Anderson). The positions of SPL1 and SPL2were located more precisely by hybridization of the same probesto nitrocellulose filters (gift from L. Riles and M. Olson) con-taining an ordered set of yeast genomic DNA segments in-serted into a bacteriophage l vector (Riles et al. 1993).
Construction of null mutations: The one-step gene replace-ment method (Rothstein 1983) was used to introduce allmutant alleles into their corresponding chromosomal loci. Toinactivate the PHO81 gene, plasmid pPHO81D::TRP1B (giftfrom E. O’Neil and E. O’Shea) was digested with NcoI andApaI, and the resulting fragment (in which codons 35–421 ofthe PHO81 coding sequence have been removed and substi-tuted with the TRP1 gene) was used to transform YPH499 totryptophan prototrophy. To disrupt the SPL2 gene, the 149-bp NcoI-BstEII fragment in plasmid pJF181 (codons 1–49 ofthe SPL2 open reading frame) was excised and replaced witha 2.7-kb fragment containing the HIS3 gene derived fromplasmid pJJ217 (Jones and Prakash 1990) to create plasmidpJF230. Plasmid pJF230 was digested with EcoRV and HpaI,and the released fragment was used to transform a diploidstrain, YPH501 (Sikorski and Hieter 1989), to histidine pro-totrophy; sporulation and tetrad dissection of a resulting dip-loid transformant yielded haploid segregants containing thespl2D::HIS3 allele. To eliminate the PHO4 gene, plasmidpPHO4dv (gift from E. O’Shea) was digested with SacI andXhoI, and the resulting fragment (in which codons 1–309 ofthe PHO4 coding sequence have been removed and substi-tuted with the TRP1 gene) was used to transform YPH499 totryptophan prototrophy. Proper integration of all gene trans-
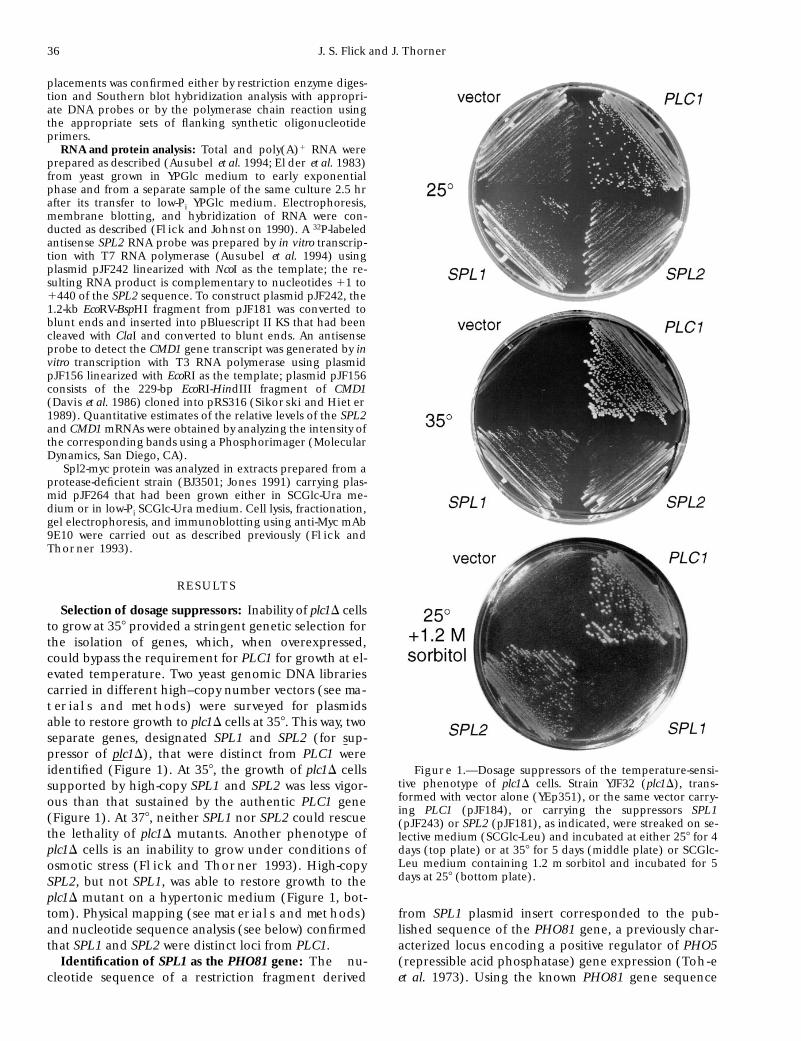
36 J. S. Flick and J. Thorner
placements was confirmed either by restriction enzyme diges-tion and Southern blot hybridization analysis with appropri-ate DNA probes or by the polymerase chain reaction usingthe appropriate sets of flanking synthetic oligonucleotideprimers.
RNA and protein analysis: Total and poly(A)1 RNA wereprepared as described (Ausubel et al. 1994; Elder et al. 1983)from yeast grown in YPGlc medium to early exponentialphase and from a separate sample of the same culture 2.5 hrafter its transfer to low-Pi YPGlc medium. Electrophoresis,membrane blotting, and hybridization of RNA were con-ducted as described (Flick and Johnston 1990). A 32P-labeledantisense SPL2 RNA probe was prepared by in vitro transcrip-tion with T7 RNA polymerase (Ausubel et al. 1994) usingplasmid pJF242 linearized with NcoI as the template; the re-sulting RNA product is complementary to nucleotides 11 to1440 of the SPL2 sequence. To construct plasmid pJF242, the1.2-kb EcoRV-BspHI fragment from pJF181 was converted toblunt ends and inserted into pBluescript II KS that had beencleaved with ClaI and converted to blunt ends. An antisenseprobe to detect the CMD1 gene transcript was generated by invitro transcription with T3 RNA polymerase using plasmidpJF156 linearized with EcoRI as the template; plasmid pJF156consists of the 229-bp EcoRI-HindIII fragment of CMD1(Davis et al. 1986) cloned into pRS316 (Sikorski and Hieter
1989). Quantitative estimates of the relative levels of the SPL2and CMD1 mRNAs were obtained by analyzing the intensity ofthe corresponding bands using a Phosphorimager (MolecularDynamics, San Diego, CA).
Spl2-myc protein was analyzed in extracts prepared from aprotease-deficient strain (BJ3501; Jones 1991) carrying plas-mid pJF264 that had been grown either in SCGlc-Ura me-dium or in low-Pi SCGlc-Ura medium. Cell lysis, fractionation,gel electrophoresis, and immunoblotting using anti-Myc mAb9E10 were carried out as described previously (Flick andThorner 1993).
RESULTS
Selection of dosage suppressors: Inability of plc1D cellsto grow at 358 provided a stringent genetic selection forthe isolation of genes, which, when overexpressed,could bypass the requirement for PLC1 for growth at el-evated temperature. Two yeast genomic DNA librariescarried in different high–copy number vectors (see ma-
terials and methods) were surveyed for plasmidsable to restore growth to plc1D cells at 358. This way, twoseparate genes, designated SPL1 and SPL2 (for sup-pressor of plc1D), that were distinct from PLC1 wereidentified (Figure 1). At 358, the growth of plc1D cellssupported by high-copy SPL1 and SPL2 was less vigor-ous than that sustained by the authentic PLC1 gene(Figure 1). At 378, neither SPL1 nor SPL2 could rescuethe lethality of plc1D mutants. Another phenotype ofplc1D cells is an inability to grow under conditions ofosmotic stress (Flick and Thorner 1993). High-copySPL2, but not SPL1, was able to restore growth to theplc1D mutant on a hypertonic medium (Figure 1, bot-tom). Physical mapping (see materials and methods)and nucleotide sequence analysis (see below) confirmedthat SPL1 and SPL2 were distinct loci from PLC1.
Identification of SPL1 as the PHO81 gene: The nu-cleotide sequence of a restriction fragment derived
from SPL1 plasmid insert corresponded to the pub-lished sequence of the PHO81 gene, a previously char-acterized locus encoding a positive regulator of PHO5(repressible acid phosphatase) gene expression (Toh-e
et al. 1973). Using the known PHO81 gene sequence
Figure 1.—Dosage suppressors of the temperature-sensi-tive phenotype of plc1D cells. Strain YJF32 (plc1D), trans-formed with vector alone (YEp351), or the same vector carry-ing PLC1 (pJF184), or carrying the suppressors SPL1(pJF243) or SPL2 (pJF181), as indicated, were streaked on se-lective medium (SCGlc-Leu) and incubated at either 258 for 4days (top plate) or at 358 for 5 days (middle plate) or SCGlc-Leu medium containing 1.2 m sorbitol and incubated for 5days at 258 (bottom plate).

Plc1p Function and the Pho80p/Pho85p Cdk in Yeast 37
(Ogawa et al. 1993), the portion of the SPL1-contain-ing plasmid insert that contained only the PHO81 cod-ing region and its promoter was subcloned (Figure 2).This DNA fragment carried on a high-copy vector(pJF243; Figure 2) permitted the growth of plc1D cellsat 358 just as well as the original plasmid isolate. To con-firm unequivocally that the SPL1 suppressor activitywas provided solely by the PHO81 gene product, threemutant alleles were constructed. Two truncations ofthe PHO81 open reading frame (plasmids pJF202 andpJF237), even one that removed ,20% of the predictedprotein (codons 957–1179), eliminated SPL1 suppres-sor activity (Figure 2). Likewise, an allele in which weintroduced a frame shift mutation at codon 557 of thePHO81 open reading frame (plasmid pJF251) also to-tally destroyed its SPL1 suppressor activity (Figure 2).
PHO81 is a phosphate-regulated inhibitor of a Cdkcomprised of a cyclin protein encoded by the PHO80gene and a protein kinase catalytic subunit encodedby the PHO85 gene (Schneider et al. 1994). Duringgrowth in normal media (i.e., high phosphate), thePHO4 -encoded transcriptional activator is hyperphos-phorylated by the Pho80p/Pho85p complex, whichleads to exclusion of Pho4p from the nucleus (O’Neil
et al. 1996). Starvation for phosphate stimulates Pho81protein to inhibit Pho80p/Pho85p Cdk activity, thus al-lowing underphosphorylated Pho4p to accumulate inthe nucleus and activate transcription of genes in thePHO regulon, including PHO5 (Schneider et al. 1994).It has been shown previously that increased dosage ofthe PHO81 gene leads to partially constitutive expres-sion of PHO5, even in media containing abundant phos-phate (Yoshida et al. 1989).
SPL1 activity of PHO81 correlates with its ability to in-hibit the Pho80/Pho85 Cdk: Our results suggested thathigh-copy PHO81 might suppress temperature-sensitivegrowth of plc1D cells by inhibiting Pho80p/Pho85p, orvia subsequent induction of a gene of the PHO regulon(or both). Multicopy PHO81 was indeed capable ofcausing derepression of the genes of the PHO regulonin plc1D mutants because, unlike plc1D cells alone,
plc1D mutants carrying SPL1 plasmids secreted elevatedlevels of acid phosphatase, even when grown in highphosphate medium, as assessed by a colorimeteric col-ony overlay assay (data not shown). Domains of Pho81prequired for effective inhibition of Pho80p/Pho85p invivo, as judged by the degree of Pho5p derepression,have been mapped previously (Ogawa et al. 1993). Thecentral region of Pho81p contains six tandem ankyrinrepeats, flanked on their amino-terminal side by a neg-atively acting domain and on their C-terminal side by apositively acting domain. Tandem ankyrin repeats arealso found in p16INK4 and in other low molecular weightinhibitors of mammalian Cdk4 and Cdk6 (Serrano etal. 1993). In addition to its ankyrin repeats, a C-termi-nal segment of Pho81p (residues 772–1179) is requiredfor derepression of PHO5 expression (Ogawa et al. 1995).A plasmid, pJF237, which expressed a form of Pho81pthat retained its ankyrin repeats, but truncated theC-terminal domain (removing amino acids 957–1179),failed to suppress the temperature sensitivity of plc1Dcells (Figure 2) or to derepress acid phosphatase activ-ity (data not shown). Thus, a region of Pho81p previ-ously defined as critical for its ability to inhibit thePho80p/Pho85p enzyme and to cause derepression ofPHO5 was also required for its SPL1 activity.
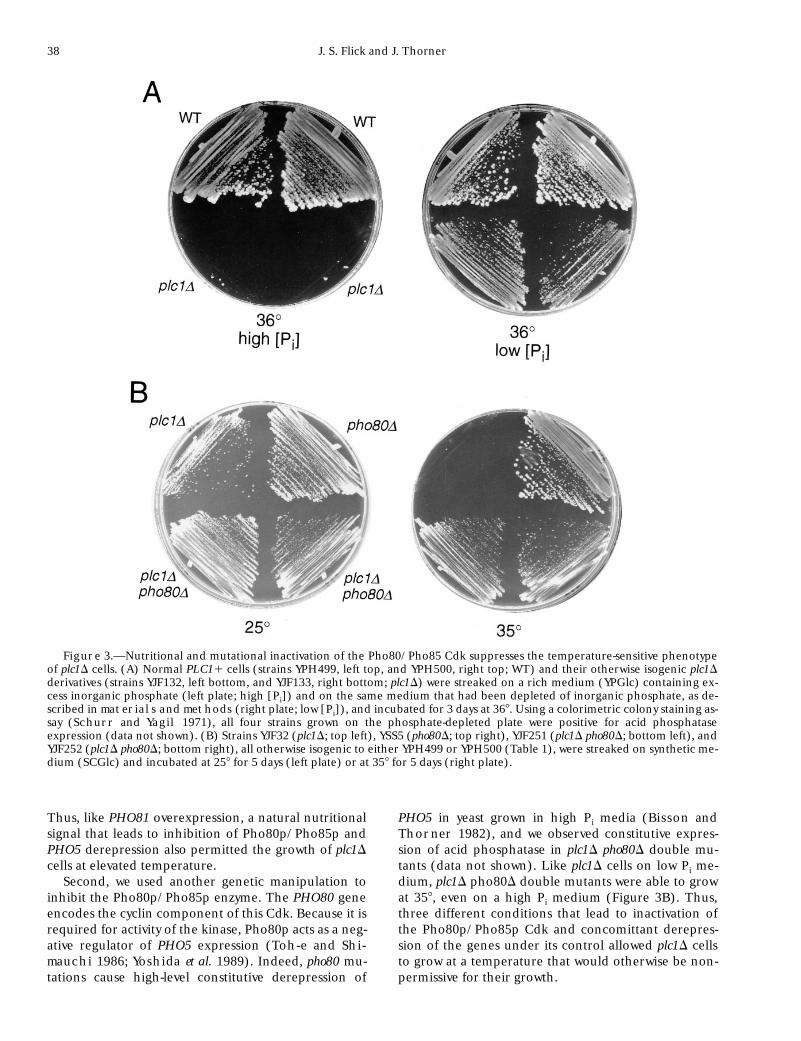
Other conditions that inhibit Pho80p/Pho85p and dere-press the PHO regulon suppress the temperature-sensitivegrowth of plc1D mutants: To determine if PHO81 overex-pression rescued the temperature-sensitive lethality ofplc1D cells solely because of its inhibition of Pho80p/Pho85p and the consequent derepression of genes un-der its control, we examined other conditions thatcause inhibition of this Cdk. First, we simply altered thePi concentration in the growth medium. As shown pre-viously (Flick and Thorner 1993), on a medium con-taining high Pi (where Pho80p/Pho85p activity is highand PHO5 expression is prevented), plc1D cells failed togrow at 358 (Figure 3A, left side). In contrast, on a me-dium containing low Pi (where Pho80p/Pho85p activityis low and PHO5 is expressed), plc1D cells were able topropagate at high temperature (Figure 3A, right side).
Figure 2.—Demonstration that PHO81 encodes SPL1 function. Top line shows a restriction endonuclease cleavage site map ofthe PHO81-containing region [adapted from (Ogawa et al. 1993)] of the DNA insert in the original SPL1 isolate (pJF115). ThePHO81 open reading frame (shaded bar) contains six ankyrin repeats (solid arrowheads). The indicated restriction fragmentswere subcloned into vector YEp351 and retested for their ability (1) or inability (2) to transform strain YJF32 (plc1D) to leucineprototrophy at 358. The insert in plasmid pJF251 contains a frameshift mutation (asterisk) introduced by cleavage with BamHI, fill-ing in with the Klenow fragment of E. coli DNA polymerase I, and religation. Restriction sites: A, AvaII; B, BamHI; E, EcoRI; H, HindIII.
38 J. S. Flick and J. Thorner
Thus, like PHO81 overexpression, a natural nutritionalsignal that leads to inhibition of Pho80p/Pho85p andPHO5 derepression also permitted the growth of plc1Dcells at elevated temperature.
Second, we used another genetic manipulation toinhibit the Pho80p/Pho85p enzyme. The PHO80 geneencodes the cyclin component of this Cdk. Because it isrequired for activity of the kinase, Pho80p acts as a neg-ative regulator of PHO5 expression (Toh-e and Shi-
mauchi 1986; Yoshida et al. 1989). Indeed, pho80 mu-tations cause high-level constitutive derepression of
PHO5 in yeast grown in high Pi media (Bisson andThorner 1982), and we observed constitutive expres-sion of acid phosphatase in plc1D pho80D double mu-tants (data not shown). Like plc1D cells on low Pi me-dium, plc1D pho80D double mutants were able to growat 358, even on a high Pi medium (Figure 3B). Thus,three different conditions that lead to inactivation ofthe Pho80p/Pho85p Cdk and concomittant derepres-sion of the genes under its control allowed plc1D cellsto grow at a temperature that would otherwise be non-permissive for their growth.
Figure 3.—Nutritional and mutational inactivation of the Pho80/Pho85 Cdk suppresses the temperature-sensitive phenotypeof plc1D cells. (A) Normal PLC11 cells (strains YPH499, left top, and YPH500, right top; WT) and their otherwise isogenic plc1Dderivatives (strains YJF132, left bottom, and YJF133, right bottom; plc1D) were streaked on a rich medium (YPGlc) containing ex-cess inorganic phosphate (left plate; high [Pi]) and on the same medium that had been depleted of inorganic phosphate, as de-scribed in materials and methods (right plate; low [Pi]), and incubated for 3 days at 368. Using a colorimetric colony staining as-say (Schurr and Yagil 1971), all four strains grown on the phosphate-depleted plate were positive for acid phosphataseexpression (data not shown). (B) Strains YJF32 (plc1D; top left), YSS5 (pho80D; top right), YJF251 (plc1D pho80D; bottom left), andYJF252 (plc1D pho80D; bottom right), all otherwise isogenic to either YPH499 or YPH500 (Table 1), were streaked on synthetic me-dium (SCGlc) and incubated at 258 for 5 days (left plate) or at 358 for 5 days (right plate).

Plc1p Function and the Pho80p/Pho85p Cdk in Yeast 39
Disruption of the genes for several other Pho85-asso-ciated cyclins does not suppress plc1D: Pho85p has re-cently been shown to associate with nine other putativecyclin proteins, in addition to Pho80p, including:Pcl1p, Pcl2p, Pcl5p, Pcl6p, Pcl7p, Pcl8p, Pcl9p, Pcl10p,and Clg1p (Measday et al. 1997). To determine if lossof other Pho85 Cdk complexes can bypass the tempera-ture sensitivity caused by the plc1D mutation, we gener-ated double mutants of plc1D with pcl1D, pcl2D, pcl5D,and clg1D. Unlike plc1D pho80D cells, none of thesedouble mutants was able to grow at a restrictive temper-ature, indicating that suppression was not a general re-sult of perturbed Pho85p activity or altered cyclin ra-tios. A plc1D pho85D double mutant also failed to growat restrictive temperature (data not shown). This find-ing might mean that a Pho85p-independent activity ofoverproduced Pho81p (or of loss of Pho80p) is respon-sible for suppression of the plc1D mutation. It is morelikely, however, that alteration or reduction of Pho85pactivity is able to suppress the plc1D temperature-sensi-tive defect, whereas total elimination of all Pho85p-dependent Cdk activity has such a deleterious effect oncells as to preclude growth of a plc1D pho85D doublemutant under any condition. In support of this view,pho85D mutants, which lack 10 different Cdk complexes,grow quite poorly even at 308 (Gilliquet and Berben
1993), show defects in glycogen metabolism (Timblin
et al. 1996), and display abnormal cellular morphology(Measday et al. 1997).
Molecular characterization of the SPL2 gene: The SPL2suppressor locus was delimited to a 1.8-kb BamHI-SalIfragment by subcloning restriction fragments from oneof the original plasmid isolates and retesting for sup-pressor activity (Figure 4A). Within this DNA fragment,nucleotide sequence analysis (EMBL/GenBank acces-sion number P38839) delineated a 148-codon openreading frame (YHR136c) that originated from theright arm of chromosome 8 (Johnston et al. 1994).Two different approaches were taken to confirm thatthe 148-codon open reading frame encoded the SPL2suppressor activity. First, a frameshift mutation intro-duced at codon 49 abolished the suppressor activity ofthe subcloned BamHI-SalI fragment (plasmid pJF217;Figure 4A). Second, expression of just the 148-codonsequence from a different promoter (ADH 1) on a highcopy plasmid vector was able to suppress the tempera-ture-sensitive growth defect of plc1D cells (plasmidpJF235; Figure 4A). Furthermore, our subsequent char-acterization (see below) confirmed that SPL2 encodesthe predicted protein.
The deduced SPL2 gene product (Figure 4B) has acalculated molecular mass of 17 kD and, allowing forconservative amino acid replacements, is similar to thesequences of mammalian low molecular weight Cdk in-hibitors and also to the ankyrin repeat region inPho81p (Figure 4C). Similarity between Spl2p and theother proteins is as follows: p16 (29%), p18 (33%), p19
(30%), and Pho81 (27%). By comparison, Far1p, ademonstrated inhibitor of the Cdc28/Cln2 Cdk (Peter
and Herskowitz 1994), does not bear detectable ho-mology to any mammalian Cdk inhibitor yet identified.We noted that the N-terminal 20 residues of Spl2p arecomprised almost exclusively of hydrophobic and un-charged amino acids, and that Gly2 is a potential targetfor N-myristoylation, although Spl2 lacks other consen-sus residues found in efficiently myristoylated proteins(Towler et al. 1988). The remainder of the protein canbe divided into two regions: residues 32–97 constitute ahighly basic segment (net charge 19), and residues 98–148 comprise a highly acidic segment (net charge 211;Figure 4B). Potential phosphorylation sites for severalclasses of protein kinase are also present in Spl2, in-cluding cAMP-dependent protein kinase (Ser59 andSer86), proline-directed protein kinases (Thr5, Ser30and Thr81), and casein kinase II (Ser142 and Ser143).The functional significance of these sites has not beenexplored.
SPL2 gene expression and Spl2 protein level are reg-ulated by phosphate: The SPL2 promoter region con-tains three matches to the consensus (59-CACGTG-39)for binding of the Pho4p transactivator located at posi-tions 2147, 290, and 231 (where 11 is the ATG; Fig-ure 4B). When Pi is limiting, Pho4p is in the nucleus,binds to such sites in the upstream regions of PHO5and the other genes of the PHO regulon, and activatestranscription (Fisher et al. 1991). To determine if itsexpression is regulated by phosphate, a 32P-labeledSPL2 probe was hybridized to size-fractionated RNAprepared from yeast grown in either a high or a low Pimedium. The internal control for loading was a probefor CMD1 mRNA, that encodes calmodulin. The SPL2transcript was polyadenylated and long enough (z0.45kb) to encode the SPL2 open reading frame (Figure5A). In cells grown in high Pi medium, SPL2 mRNA wasexpressed at z30% of the level of the CMD1 mRNA. Af-ter shift to low Pi medium for 2.5 hr, the steady-statelevel of SPL2 mRNA increased 2.7-fold (when normal-ized to CMD1 RNA), indicating that SPL2 expression isinduced and suggesting that its potential Pho4-bindingsites have a functional role. In further support that in-duction occurs at the transcriptional level, expressionof an SPL2-lacZ fusion (containing SPL2 sequencesfrom 2724 to 13) in wild-type cells increased 4.4-foldafter transfer to a low Pi medium, whereas in pho80Dcells, expression was constitutively high and did not in-crease significantly upon transfer to low Pi medium, asexpected if SPL2 is regulated by Pho4p (data notshown).
For detection, sequences encoding a c-Myc epitopewere fused in-frame to the C terminus of the SPL2 cod-ing region. The epitope used is recognized by the mono-clonal antibody 9E10. The epitope-tagged Spl2 (Spl2myc)expressed from the SPL2 promoter suppressed the tem-perature sensitivity of plc1D cells as efficiently as untagged

40 J. S. Flick and J. Thorner
Spl2 (data not shown), demonstrating that Spl2myc wasfunctional. Extracts of yeast expressing Spl2myc fromthe SPL2 promoter on a multicopy plasmid were ana-lyzed by SDS-PAGE and immunoblotting (Figure 5B).Spl2myc migrated with a mobility corresponding to anapparent molecular mass of z23 kD, which was some-what larger than its calculated molecular mass (19.2
kD). However, anomalous migration has been observedfor other polypeptides that are also very highly charged(Benton et al. 1994; Swartzman et al. 1996). As judgedby densitometry, Spl2myc level increased about three-fold in cells grown in a low Pi medium compared to itslevel in an equivalent amount of extract from cellsgrown on the same medium containing high Pi (Figure
Figure 4.—Restriction map, de-duced primary structure, and se-quence comparison of the SPL2 gene.(A) Physical map of the genomic DNA(thin line) in the region containingthe SPL2 gene (shaded bar), which isadjacent to the 59 end of the YCK1gene (hatched box) and transcribedin the same direction (single-headedarrows). The indicated DNA fragmentswere subcloned into vector YEp351and retested for their ability (1) or in-ability (2) to transform YJF32 (plc1D)to leucine prototrophy at 358. Theinsert in plasmid pJF217 contains aframeshift mutation (asterisk) intro-duced by cleavage with BstEII, fillingin with the Klenow fragment of E. coliDNA polymerase I, and religation. Inplasmid pJF235, the SPL2 open read-ing frame was expressed from theADH1 promoter, as described in ma-
terials and methods, and was suffi-cient for complementation (#). Re-striction sites: B, BamHI; Bs, BspHI; Bt,BstEII; N, NcoI; Nd, NdeI; R, EcoRV;and, S, SalI. (B) Nucleotide sequence(where 11 indicates the first base ofthe ATG initiator codon; EMBL/Gen-Bank accession number P38839) andpredicted amino acid sequence (where1 indicates the initiator methionine res-idue) of the SPL2 gene. Three consen-sus Pho4-binding sites in the promoterregion (underlined), basic residues (K,R; boxed), and acidic residues (D, E;circled) are indicated. (C) Comparisonof the primary sequence of Spl2 tothose of known Cdk inhibitors, p16(Serrano et al. 1993), p18 (Guan et al.1994), p19 (Chan et al. 1995), andPho81 (Ogawa et al. 1993). Identitiesand conventional conservative substitu-tions (Tanaka et al. 1990) (A5G5P5S;L5I5V5M; R5K5H; D5E; N5Q;F5Y5W; S5T; N5D; Q5E) betweenSpl2p and any of the other proteinslisted are given as white-on-black let-ters.
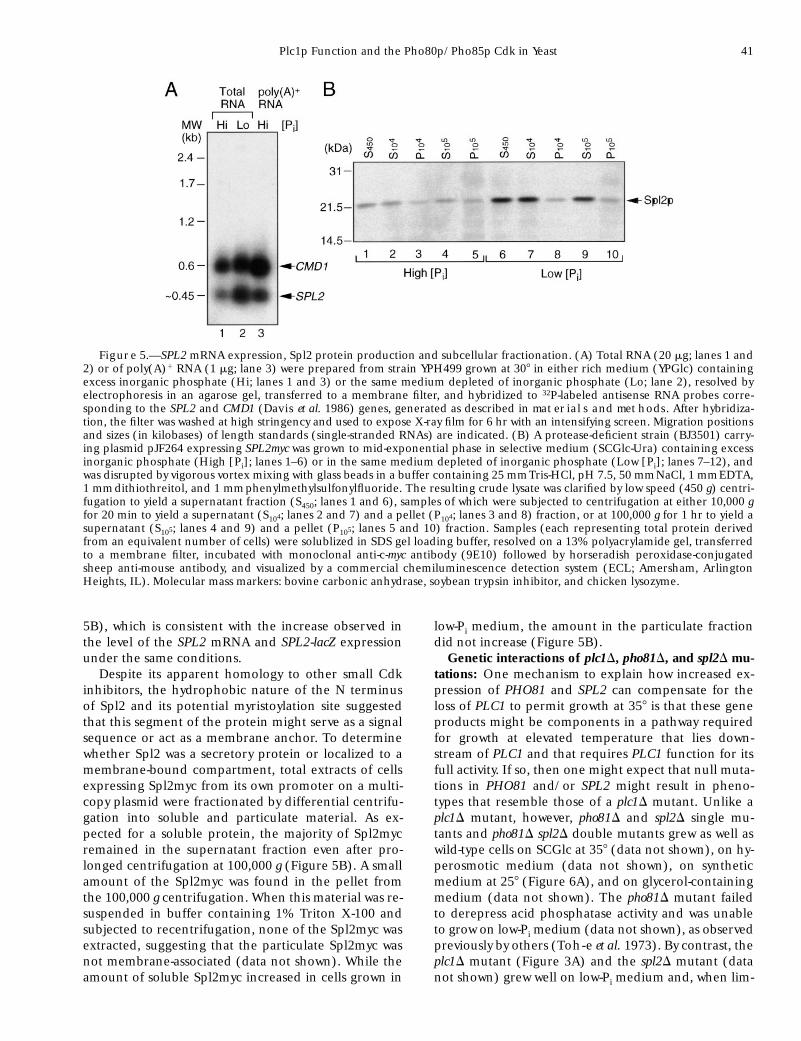
Plc1p Function and the Pho80p/Pho85p Cdk in Yeast 41
5B), which is consistent with the increase observed inthe level of the SPL2 mRNA and SPL2-lacZ expressionunder the same conditions.
Despite its apparent homology to other small Cdkinhibitors, the hydrophobic nature of the N terminusof Spl2 and its potential myristoylation site suggestedthat this segment of the protein might serve as a signalsequence or act as a membrane anchor. To determinewhether Spl2 was a secretory protein or localized to amembrane-bound compartment, total extracts of cellsexpressing Spl2myc from its own promoter on a multi-copy plasmid were fractionated by differential centrifu-gation into soluble and particulate material. As ex-pected for a soluble protein, the majority of Spl2mycremained in the supernatant fraction even after pro-longed centrifugation at 100,000 g (Figure 5B). A smallamount of the Spl2myc was found in the pellet fromthe 100,000 g centrifugation. When this material was re-suspended in buffer containing 1% Triton X-100 andsubjected to recentrifugation, none of the Spl2myc wasextracted, suggesting that the particulate Spl2myc wasnot membrane-associated (data not shown). While theamount of soluble Spl2myc increased in cells grown in
low-Pi medium, the amount in the particulate fractiondid not increase (Figure 5B).
Genetic interactions of plc1D, pho81D, and spl2D mu-tations: One mechanism to explain how increased ex-pression of PHO81 and SPL2 can compensate for theloss of PLC1 to permit growth at 358 is that these geneproducts might be components in a pathway requiredfor growth at elevated temperature that lies down-stream of PLC1 and that requires PLC1 function for itsfull activity. If so, then one might expect that null muta-tions in PHO81 and/or SPL2 might result in pheno-types that resemble those of a plc1D mutant. Unlike aplc1D mutant, however, pho81D and spl2D single mu-tants and pho81D spl2D double mutants grew as well aswild-type cells on SCGlc at 358 (data not shown), on hy-perosmotic medium (data not shown), on syntheticmedium at 258 (Figure 6A), and on glycerol-containingmedium (data not shown). The pho81D mutant failedto derepress acid phosphatase activity and was unableto grow on low-Pi medium (data not shown), as observedpreviously by others (Toh-e et al. 1973). By contrast, theplc1D mutant (Figure 3A) and the spl2D mutant (datanot shown) grew well on low-Pi medium and, when lim-
Figure 5.—SPL2 mRNA expression, Spl2 protein production and subcellular fractionation. (A) Total RNA (20 mg; lanes 1 and2) or of poly(A)1 RNA (1 mg; lane 3) were prepared from strain YPH499 grown at 308 in either rich medium (YPGlc) containingexcess inorganic phosphate (Hi; lanes 1 and 3) or the same medium depleted of inorganic phosphate (Lo; lane 2), resolved byelectrophoresis in an agarose gel, transferred to a membrane filter, and hybridized to 32P-labeled antisense RNA probes corre-sponding to the SPL2 and CMD1 (Davis et al. 1986) genes, generated as described in materials and methods. After hybridiza-tion, the filter was washed at high stringency and used to expose X-ray film for 6 hr with an intensifying screen. Migration positionsand sizes (in kilobases) of length standards (single-stranded RNAs) are indicated. (B) A protease-deficient strain (BJ3501) carry-ing plasmid pJF264 expressing SPL2myc was grown to mid-exponential phase in selective medium (SCGlc-Ura) containing excessinorganic phosphate (High [Pi]; lanes 1–6) or in the same medium depleted of inorganic phosphate (Low [Pi]; lanes 7–12), andwas disrupted by vigorous vortex mixing with glass beads in a buffer containing 25 mm Tris-HCl, pH 7.5, 50 mm NaCl, 1 mm EDTA,1 mm dithiothreitol, and 1 mm phenylmethylsulfonylfluoride. The resulting crude lysate was clarified by low speed (450 g) centri-fugation to yield a supernatant fraction (S450; lanes 1 and 6), samples of which were subjected to centrifugation at either 10,000 gfor 20 min to yield a supernatant (S104; lanes 2 and 7) and a pellet (P104; lanes 3 and 8) fraction, or at 100,000 g for 1 hr to yield asupernatant (S105; lanes 4 and 9) and a pellet (P105; lanes 5 and 10) fraction. Samples (each representing total protein derivedfrom an equivalent number of cells) were solublized in SDS gel loading buffer, resolved on a 13% polyacrylamide gel, transferredto a membrane filter, incubated with monoclonal anti-c-myc antibody (9E10) followed by horseradish peroxidase-conjugatedsheep anti-mouse antibody, and visualized by a commercial chemiluminescence detection system (ECL; Amersham, ArlingtonHeights, IL). Molecular mass markers: bovine carbonic anhydrase, soybean trypsin inhibitor, and chicken lysozyme.

42 J. S. Flick and J. Thorner
ited for phosphate, displayed levels of secreted acidphosphatase comparable to those seen in derepressedwild-type cells (data not shown). The lack of similarityin phenotypes between a plc1D mutant and the pho81Dand spl2D single and double mutants suggests that nei-ther PHO81 nor SPL2 function in a strictly linear path-way downstream of PLC1.
An alternative hypothesis to explain suppression ofthe temperature sensitivity of plc1D cells by high-levelexpression of PHO81 or SPL2 is that PHO81 and SPL2act in an independent pathway(s) that is necessary forgrowth at elevated temperature and that overlaps (or ispartially redundant in function) with the PLC1-depen-dent pathway. If so, double mutants in which both path-ways are disrupted should display more severe defectsthan single mutants in which only one pathway is non-functional. To test this possibility, plc1D pho81D andplc1D spl2D double mutants were constructed and ex-amined, along with the plc1D pho80D double mutant(Figure 3B). On rich medium (YPGlc) at 258, the plc1D
pho81D and plc1D spl2D double mutants grew (Figure6B) at a rate similar to that displayed by the plc1D singlemutant (Figure 6A). Unlike the plc1D single mutant(Figure 6A), however, neither the plc1D pho81D doublemutant nor the plc1D spl2D double mutant was able toform visible colonies on the synthetic medium (SCGlc),even after incubation for 5 days at the same tempera-ture (Figure 6B). In contrast, as might have been antic-ipated from the ability of a pho80D mutation to sup-press the temperature sensitivity of a plc1D mutation(Figure 3B), the plc1D pho80D double mutant grew onthe synthetic medium at a rate similar to that of theplc1D single mutant. Surprisingly, the plc1D pho80Ddouble mutant was unable to form single colonies onrich medium after 5 days at 258. Thus, with respect togrowth on synthetic medium, the plc1D pho81D andplc1D spl2D double mutants did indeed display a muchmore severe phenotype than a plc1D single mutant.
If the loss of SPL2, like the loss of PHO81, leads to in-creased activity of the PHO80/PHO85-encoded Cdk,
Figure 6.—Nutritional effects and genetic interactions among plc1D, spl2D, pho81D, and pho80D mutations (A) Strains YPH499(WT; upper left), YJF32 (plc1D; upper right), YJF277(spl2D; lower left), and YJF552 (pho81D; lower right), constructed as describedin materials and methods, were streaked either on a rich medium (YPGlc; left plate) or on a synthetic complete medium(SCGlc; right plate) and incubated at 258 for 3 and 5 days, respectively. (B) Strains YJF306 (plc1D spl2D; upper left), YJF251(plc1Dpho80D; upper right), YJF386 (plc1D pho80D spl2D; lower left), and YJF555 (plc1D pho81D; lower right), constructed as described inmaterials and methods, were streaked as indicated in A and incubated at 258 for 4 and 5 days, respectively.

Plc1p Function and the Pho80p/Pho85p Cdk in Yeast 43
then the inability of the plc1D pho81D and plc1D spl2Ddouble mutants to grow on SCGlc might be caused byhyperelevation of the Pho80/Pho85 Cdk. To test thispossibility, a plc1D spl2D pho80D triple mutant was con-structed. Indeed, this strain was able to grow on the syn-thetic medium (Figure 6B) just as well as a plc1D singlemutant (Figure 6A). Conversely, this triple mutant wasalso able to grow well on rich medium (Figure 6B), sug-gesting that the inability of the plc1D pho80D cells togrow on YPGlc might be caused, directly or indirectly,by the derepression of SPL2. Taken all together, theseresults suggest that, in addition to their role in support-ing growth at elevated temperature, the functions ofPLC1, PHO81, SPL2, and the PHO80/PHO85-encodedCdk all converge on a pathway required for proper re-sponse to elevated temperature and changing nutrientlevels.
Epistasis relationships in plc1D suppression: Raisingthe dosage of either PHO81 or SPL2 suppressed thetemperature-sensitive growth of a plc1D mutant (Figure1). Because SPL2 is a phosphate-regulated gene (Fig-ures 4 and 5), and because overexpression of PHO81derepresses expression of other genes in the PHO regu-lon (Yoshida et al. 1989), it was possibile that high-copyPHO81 suppressed the plc1D mutation solely because of
stimulation of SPL2 expression. By this model, suppres-sion of a plc1D mutation by a pho80D mutation (Figure3) should also require SPL2 activity. To test this hypoth-esis, the phenotype of the plc1D pho80D spl2D triple mu-tant was compared to that of an otherwise isogenicplc1D pho80D double mutant. As observed before, theplc1D pho80D double mutant was able to grow at 348, atemperature that is nonpermissive for the plc1D singlemutant (Figure 7). Likewise, the plc1D pho80D spl2D tri-ple mutant was also able to grow at 348, demonstratingthat SPL2 is not essential for the ability of a pho80D mu-tation to bypass the plc1D mutation. This result furthersuggests that when the Pho80p/Pho85p Cdk is inacti-vated, the activity of a factor(s) other than SPL2 is stim-ulated that allows the Plc1-deficient cells to grow at anelevated temperature. On the other hand, growth ofthe plc1D pho80D spl2D triple mutant at 348 and 358 wasdiscernibly weaker than that of the plc1D pho80D dou-ble mutant, indicating that elevated SPL2 expression,while not obligatory, contributes to suppression.
When the Pho80p/Pho85p Cdk is inactivated, thereis a dramatic increase in the transcription of PHO4 -dependent genes. Because SPL2 was not essential forsuppression of the temperature-sensitive growth of aplc1D mutant by a pho80D mutation, it seemed reason-
Figure 7.—PHO4 or SPL2 are not essential for suppression of the temperature-sensitive phenotype plc1D cells by a pho80D mu-tation. Strains YJF32 (plc1D; upper left), YJF251 (plc1D pho80D; upper right), YJF567 (plc1D pho80D pho4D; lower left), and YJF386(plc1D pho80D spl2D; lower right), constructed as described in materials and methods, were streaked on synthetic medium(SCGlc) and incubated for 5 days at the indicated temperatures.

44 J. S. Flick and J. Thorner
able to assume that elevated expression of anotherPHO4 -dependent gene(s) was responsible for the sup-pression since, to date, Pho4p is the only known in vivosubstrate of the Pho80p/Pho85p Cdk. To determine ifany genes under Pho4p control were required for sup-pression, a plc1D pho80D pho4D triple mutant was con-structed. As expected, this strain failed to grow on alow-Pi medium and did not express detectable acidphosphatase activity (data not shown). In marked con-trast, the plc1D pho80D pho4D triple mutant grew at 348and at a rate comparable to that of the plc1D pho80Dspl2D triple mutant (Figure 7). Thus, PHO4-dependenttranscription was not required for suppression of theplc1D mutation when the Pho80p/Pho85p Cdk was in-activated by a pho80D mutation. Furthermore, the factthat plc1D pho80D pho4D and plc1D pho80D spl2D strainsgrew similarly and somewhat more weakly than theplc1D pho80D double mutant (Figure 7) provides an ad-ditional demonstration that SPL2 is under PHO4 con-trol, and that SPL2 expression contributes to the abilityof cells to grow at elevated temperatures. Nonetheless,the fact that PHO4 is not required for suppression sug-gests that loss of the Pho80p/Pho85p Cdk suppresses aplc1D mutation because the Pho80p/Pho85p normallyantagonizes the function of an as yet unidentified fac-tor that is required for the ability of cells to grow at ele-vated temperatures.
DISCUSSION
On the basis of the pleiotropic phenotypes displayedby yeast cells lacking a PLC1 gene, we proposed thatPlc1p deficiency causes defects in nutritional andstress-related responses (Flick and Thorner 1993). Asshown here, two genes, PHO81 and SPL2, when overex-pressed, can bypass the need for PLC1 to permit growth.348. Because elevated dosage of either gene sup-pressed a plc1 null allele, Pho81p and Spl2p might rep-resent components in a pathway directly downstreamof Plc1p. However, the genetic interactions of theplc1D, pho81D, and spl2D mutants (Figure 6) suggestthat Plc1p, Pho81p, and Spl2p act independently, butin a partially redundant or overlapping manner. Thus,the functions of Plc1p, Pho81p, and Spl2p appear toconverge on a common target that is required forgrowth at elevated temperature and proper response tonutrient levels.
When Pi is limiting, Pho81p inhibits the Pho80p/Pho85p Cdk (Schneider et al. 1994). Although the in-hibitory action of Pho81p apparently is enhanced bylow-Pi conditions, overexpression of PHO81 causes par-tially constitutive derepression of PHO5, even whencells are grown in high-Pi medium (Yoshida et al.1989). Thus, Pho81p must be capable, even in its basalstate, of inhibiting Pho80p/Pho85p. In fact, it has beenshown by others that overexpression of Pho81p does re-duce Pho80p/Pho85p activity in vitro, that growth in
low-Pi medium causes a PHO81-dependent reduction inPho80p/Pho85p activity in vitro, and that a pho80D mu-tation totally inactivates a PHO85-dependent kinaseactivity (Kaffman et al. 1994; Schneider et al. 1994).Hence, the observed suppression of the temperature-sensitive phenotype of plc1D cells by high-copy PHO81(Figure 1), by growth on low-Pi medium (Figure 3A),and by a pho80D mutation (Figure 3B) is most likelycaused by inhibition of Pho80p/Pho85p activity. There-fore, in a formal genetic sense, Plc1p activity may eithernegatively regulate the Pho80p/Pho85p Cdk or pro-duce a signal that opposes the effect of Pho80p/Pho85p on a downstream target. The interaction of PI-PLC d activity with a phosphate-regulated pathway ap-pears to be conserved. In Schizosaccharomyces pombe, amutation in the plc11 gene, encoding a PI-PLC d ho-molog, is partially suppressed by growth on low-Pi me-dia (Fankhauser et al. 1995).
Pho4p is the only known in vivo target of thePho80p/Pho85p Cdk, and phosphorylation of Pho4pby Pho80p/Pho85p prevents it from stimulating tran-scription by excluding Pho4p from the nucleus (O’Neil
et al. 1996). Hence, inactivation of Pho80p/Pho85pcould perhaps allow growth of plc1D cells at elevatedtemperature because it permits Pho4p to stimulate theexpression of a gene(s) that are required for growth athigh temperature. However, we found that a plc1D pho80Dpho4D triple mutant grew almost as well as the plc1Dpho80D double mutant at a temperature (348) that isnonpermissive for a plc1D single mutant (Figure 7). Theseresults suggest that Pho80p/Pho85p-dependent inhibi-tion (presumably by direct phosphorylation) of anotherfactor, different from Pho4p, is responsible for block-ing growth of plc1D cells at elevated temperature. Infurther support of this conclusion, high-copy PHO4causes robust constitutive expression of PHO5 in plc1Dcells, as in wild-type cells (Yoshida et al. 1989), yet it isonly a very weak suppressor of the temperature sensitiv-ity and does not suppress at all in a plc1D pho81D doublemutant (J. Flick, unpublished results). These observa-tions indicate that the weak suppression by multicopyPHO4 is caused by elevated expression of PHO81 andpossibly SPL2, which are both PHO4 -dependent genes(Yoshida et al. 1989; Creasy et al. 1993; Figure 5), andthe resulting inhibition of the Pho80p/Pho85p Cdk(rather than caused by induction of other PHO genes).
The second plc1D suppressor was the SPL2 gene(Figure 4). SPL2 encodes a 17-kD soluble protein. Al-though both SPL2 mRNA (Figure 5A) and Spl2 protein(Figure 5B) appear to be expressed at a significantbasal level, growth of cells in low-Pi medium caused areadily detectable induction (approximately threefold).However, suppression of the temperative-sensitive de-fect of plc1D cells by high-copy SPL2 did not requirestimulation by low-Pi conditions because suppressionwas observed on high-Pi medium (Figure 1). An spl2Dmutant had no readily discernible growth phenotype

Plc1p Function and the Pho80p/Pho85p Cdk in Yeast 45
(Figure 6), and computer searches determined that nohomolog or related gene exists in the S. cerevisiae ge-nome. Given that both PHO81 and SPL2 were isolatedas dosage suppressors, that both are Pho4p-dependentgenes, that the phenotypes of plc1D pho81D and plc1Dspl2D mutants are very similar, and that SPL2 is not re-quired for suppression by a pho80D mutation, Spl2pmay represent a novel inhibitor of the Pho80p/Pho85pCdk. Indeed, based on its molecular weight and se-quence, Spl2 (p17) resembles several known low molec-ular weight ankyrin-repeat–containing inhibitors (p16,p18, and p19) of mammalian Cdks (Sherr and Rob-
erts 1995), as well as a portion of the ankyrin-repeatregion of Pho81p (Figure 4C). However, neither anspl2D mutation nor high-copy SPL2 (unlike high-copyPHO81) perturb the regulation of PHO5 expression inplc1D yeast (J. Flick, unpublished results). Thus, Spl2pmay block the ability of Pho80p/Pho85p to phosphory-late the novel factor, but not its ability to phosphory-late Pho4p (whereas Pho81p may block the ability ofPho80p/Pho85p to phosphorylate both this factor andPho4). A somewhat analogous situation has been ob-served in animal cells where Cdk inhibition by Cip1/Waf1 inhibits the function of PCNA in DNA replica-tion, but not in DNA repair (Li et al. 1994).
The interactions among PLC1, SPL2, and elementsof the PHO regulon suggest that a factor critical forgrowth .348 is modulated by these gene products. Thissame factor may also be involved in nutrient sensingand/or utilization because either a pho81D or an spl2Dmutation prevents the growth of plc1D cells on syn-thetic medium, but not on rich medium. In support ofa connection between nutrient uptake and/or utiliza-tion and the functions of PLC1, SPL2, and the PHO reg-ulon was our observation that a plc1D pho80D doublemutant was sensitive to rich media, but able to grow onsynthetic medium (Figure 6B). This sensitivity could re-sult from hyperstimulation of amino acid uptake as aconsequence of the loss of both the Pho80p/Pho85pCdk and the loss of PLC1 function. The ability pho80mutations to promote uptake of nutrients other thaninorganic phosphate is highlighted by the fact that thetup7 mutation, isolated on the basis of enhanced dTMPuptake, is allelic to pho80 (Bisson and Thorner 1982).Another connection between nitrogen metabolism andPIP2 turnover is suggested by a report that IP3 and DAGlevels in yeast are increased when a nitrogen source isresupplied to starved cells (Schomerus and Kuntzel
1992). We propose the following working model (Figure 8)
to explain the connection among PLC1, SPL2, and theregulators of PHO gene expression that have been un-covered by the genetic interactions presented here. Inthe absence of PLC1 function, a pathway that promotesgrowth at elevated temperature and a pathway thatcontrols nutrient uptake and/or utilization cannotfunction properly because the activity of the Pho80p/
Pho85p Cdk is too high. Plc1p activity either acts to op-pose the inhibitory action of Pho80p/Pho85p on adownstream target, “X,” or stimulates the function of Xby a convergent pathway. Hence, reduction of Pho80p/Pho85p activity by overproduction of either Pho81p (aknown inhibitor of Pho80/Pho85) or Spl2p (a candi-date Cdk inhibitor), by growth on low-Pi medium or byintroduction of a pho80D mutation, all permit plc1Dcells to grow at restrictive temperature. Alternatively,Spl2 may act to stimulate the activity of X rather thaninhibiting Pho80p/Pho85p activity. Loss of both Plc1pand Pho81p, or both Plc1p and Spl2p, presumablycauses a more severe inactivation of X, explaining theinability of plc1D pho81D and plc1D spl2D double mu-tants to grow on standard synthetic medium, as well asthe relief of this phenotype by a pho80D mutation or byaugmentation with a rich growth medium. Indeed, lossof Pho81p alone has been shown to cause an elevationin Pho80p/Pho85p activity in cell extracts (Schneider
et al. 1994). The novel factor X, and not Pho4p, is re-quired for the induction of these growth pathways be-cause, for example, a pho80D mutation permits growthof plc1D cells at restrictive temperature in both aPHO41 and a pho4D background.
That Pho85 appears to interact with as many as nineother cyclins in addition to Pho80p suggests that differ-ent forms of Pho85p may receive differential inputsfrom cellular metabolism, which allows the cell to as-sess globally its nutritional and physiological state sothe rate of growth and cell division can be adjusted ac-cordingly. PIP2 has roles in membrane structure, secre-tion (through modulation of the activity of Arf pro-teins; Brown et al. 1993; Waksman et al. 1996), andactin cytoskeleton dynamics (through binding to the
Figure 8.—Model for genetic interactions and regulatorycircuitry involving PLC1, SPL2, and the PHO regulon. Nega-tive (inhibitory) interactions are indicated by a bar; positive(stimulatory) interactions are indicated by an arrow. See textfor further explanation.

46 J. S. Flick and J. Thorner
yeast homologs of profilin, cofilin, and gelsolin; Pol-
lard et al. 1994). Therefore, cells may monitor the sup-ply of PIP2 via its Plc1p-mediated turnover and inte-grate this information with the activity of the Pho80p/Pho85p Cdk. In this way, proper coordinate regulationof the rate of cell growth (which requires amino acidsfor protein synthesis and lipids for membrane expan-sion) and commitment to the cell cycle may be achieved.
We thank Erin O’Shea, Lawrence Bergman, Vivian Measday,
Brenda Andrews and Kim Arndt for the generous gifts of plasmidsand strains, J. Michael Bishop for the gift of hybridoma cell linec-myc 9E10, Linda Riles, Maynard Olson, and Mark Johnston forinformation and advice during the physical mapping of SPL1/PHO81and SPL2, Mark Lindsay for excellent technical assistance, Kathi
Glauner for assistance in the analysis of SPL2 gene expression,Randy Schekman for the use of his Phosphorimager, and membersof the Thorner lab for helpful discussions, especially Chau V. Huynh
and Elisabeth A. Schnieders.
This work was supported by a postdoctoral fellowship (DRG-1163)from the Cancer Research Fund of the Damon Runyon-WalterWinchell Foundation, funds from the Department of Biochemistry,Vanderbilt University, and a National Scientist Development Grant(AHA-9630327N) from the American Heart Association (to J.S.F.), byfunds provided by the Berkeley campus Lucille P. Markey Program inBiomolecular Structure and Design, by facilities supplied by the Can-cer Research Laboratory of the University of California, Berkeley, andby National Institutes of Health research grant GM21841 (to J.T.).
LITERATURE CITED
Altschul, S., W. Gish, W. Miller, E. Myers and D. Lipman,
1990 Basic local alignment search tool. J. Mol. Biol. 215: 403–410.
Ausubel, F., R. Brent, R. Kingston, J. Moore, J. Seidman et al.,1994 Current Protocols in Molecular Biology, Greene PublishingAssociates/Wiley-Interscience, New York.
Benton, B. M., J.-H. Zang and J. Thorner, 1994 A novel FK506-and rapamycin-binding protein (FPR3 gene product) in theyeast Saccharomyces cerevisiae is a proline rotamase localized to thenucleolus. J. Cell Biol. 127: 623–639.
Berridge, M. J., 1993 Inositol trisphosphate and calcium signal-ling. Nature 361: 315–325.
Biggin, M. D., T. J. Gibson and G. F. Hong, 1983 Buffer gradientgels and 35S label as an aid to rapid DNA sequence determina-tion. Proc. Natl. Acad. Sci. USA 80: 3963–3965.
Bisson, L., and J. Thorner, 1982 Mutations in the PHO80 geneconfer permeability to 59-mononucleotides in Saccharomyces cere-visiae. Genetics 102: 341–359.
Bloomquist, B. T., R. D. Shortridge, S. Schneuwly, M. Perdew, C.
Montrell et al., 1988 Isolation of a putative phospholipase Cgene of Drosophilia, norpA, and its role in phototransduction.Cell 54: 723–733.
Brown, H., S. Gutowski, C. Moomaw, C. Slaughter and P. Stern-
weis, 1993 ADP-ribosylation factor, a small GTP-dependentregulatory protein, stimulates phospholipase D activity. Cell 75:1137–1144.
Carlson, M., and D. Botstein, 1982 Two differentially regulatedmRNAs with different 59 ends encode secreted with intracellularforms of yeast invertase. Cell 28: 145–154.
Chan, F., J. Zhang, L. Cheng, D. Shapiro and A. Winoto, 1995Identification of human and mouse p19, a novel CDK 4 andCDK6 inhibitor with homology to p16INK4. Mol. Cell. Biol. 15:2682–2698.
Chu, G., D. Vollrath and R. W. Davis, 1986 Separation of largeDNA molecules by contour-clamped homogeneous electricfields. Science 234: 1582–1585.
Clapham, D. E., 1995 Calcium signaling. Cell 80: 259–268.Clipstone, N. A., and G. R. Crabtree, 1992 Identification of cal-
cineurin as a key signaling enzyme in T-lymphocyte activation.Nature 357: 695–697.
Creasy, C. L., S. L. Madden and L. W. Bergman, 1993 Molecularanalysis of the PHO81 gene of Saccharomyces cerevisiae. Nucleic Ac-ids Res. 21: 1975–1982.
Davis, T. N., M. S. Urdea, F. R. Masiarz and J. Thorner, 1986Isolation of the yeast calmodulin gene: calmodulin is an essentialprotein. Cell 47: 423–431.
Drayer, A. L., J. Van der Kaay, G. W. Mayr and P. Van Haastert,
1994 Role of phospholipase C in Dictyostelium: formation ofinositol 1,4,5-trisphosphate and normal development in cellslacking phospholipase C activity. EMBO J. 13: 1601–1609.
Elder, R. T., E. Y. Loh and R. W. Davis, 1983 RNA from the yeasttransposable element Ty1 has both ends in the direct repeats, astructure similar to retrovirus RNA. Proc. Natl. Acad. Sci. USA80: 2432–2436.
Evan, G. I., G. K. Lewis, G. Ramsay and J. M. Bishop, 1985 Isolationof monoclonal antibodies specific for human c-myc proto-onco-gene product. Mol. Cell. Biol. 5: 3610–3616.
Fankhauser, H., M. Schweingruber, E. Edenharter and M.
Schweingruber, 1995 Growth of a mutant defective in a puta-tive phosphoinositide-specific phospholipase C of Schizosaccharo-myces pombe is restored by low concentrations of phosphate andinositol. Curr. Genet. 28: 199–203.
Feng, J.-F., S. G. Rhee and M.-J. Im, 1996 Evidence that phospholi-pase d1 is the effector in the Gh(transglutaminase II)-mediatedsignalling. J. Biol. Chem. 271: 16451–16454.
Fisher, F., P.-S. Jayaraman and C. Goding, 1991 C-Myc and theyeast transcription factor PHO4 share a common CACGTG-bind-ing motif. Oncogene 6: 1099–1104.
Flick, J. S., and M. Johnston, 1990 Two systems of glucose repres-sion of the GAL1 promoter in Saccharomyces cerevisiae. Mol. Cell.Biol. 10: 4757–4769.
Flick, J., and J. Thorner, 1993 Genetic and biochemical character-ization of a phosphatidyl-inositol-specific phospholipase C in Sac-charomyces cerevisiae. Mol. Cell. Biol. 13: 5861–5876.
Franz, B., E. Norby, G. Bren, N. Steffan, C. Paya et al., 1994Calcineurin acts in synergy with PMA to inactivate IkB/MAD3,an inhibitor of NF-kB. EMBO J. 13: 861–870.
Gietz, R. D., and R. H. Schiestl, 1991 Applications of high effi-ciency lithium acetate transformations of intact yeast cells usingsingle-stranded nucleic acids as carrier. Yeast 7: 253–263.
Gilliquet, V., and G. Berben, 1993 Positive and negative regula-tors of the Saccharomyces cerevisiae “PHO system” participate inseveral cell functions. FEBS Lett. 108: 333–340.
Guan, K., C. Jenkins, Y. Li, M. Nichols, X. Wu et al., 1994 Growthsuppression by p18, a p16INK4/MTS1- and p14INK4B/MTS2-related CDK6 inhibitor, correlates with wild-type pRb function.Genes Dev. 8: 2939–2952.
Henikoff, S., 1984 Unidirectional digestion with exonuclease IIIcreates targeted breakpoints for DNA sequencing. Gene 28:351–359.
Hill, C., and R. Treisman, 1995 Transcriptional regulation by ex-tracellular signals: mechanisms and specificity. Cell 80: 199–211.
Hill, J. E., A. M. Myers, T. J. Koerner and A. Tzagoloff,
1986 Yeast/E. coli shuttle vectors with multiple unique restric-tion sites. Yeast 2: 163–167.
Homma, Y., and Y. Emori, 1995 A dual functional signal mediatorshowing RhoGAP and phospholipase C-d stimulating activities.EMBO J. 14: 286–291.
Ji, Q.-S., G. Winnier, K. Niswender, D. Horstman, R. Wisdom et al.,1997 Essential role of the tyrosine kinase substrate phospholi-pase C-g1 in mammalian development. Proc. Natl. Acad. Sci.USA 94: 2999–3003.
Johnston, M., and M. Carlson, 1992 Regulation of carbon andphosphate utilization, pp. 193–282 in The Molecular and CellularBiology of the Yeast Saccharomyces, edited by E. Jones, J. Pringle
and J. Broach. Cold Spring Harbor Laboratory Press, ColdSpring Harbor, NY.
Jones, E. W., 1991 Tackling the protease problem in Saccharomycescerevisiae. Methods Enzymol. 194: 428–453.
Jones, J. S., and L. Prakash, 1990 Yeast Saccharomyces cerevisiae se-lectable markers in pUC18 polylinkers. Yeast 6: 363–366.
Kaffman, A., I. Herskowitz, R. Tjian and E. O’Shea, 1994 Phos-phorylation of the transcription factor PHO4 by a cyclin-CDKcomplex, PHO80-PHO85. Science 263: 1153–1156.

Plc1p Function and the Pho80p/Pho85p Cdk in Yeast 47
Koch, C. A., D. Anderson, M. F. Moran, C. Ellis and T. Pawson,
1991 SH2 and SH3 domains: elements that control interac-tions of cytoplasmic signalling proteins. Science 252: 668–674.
Lee, C. H., D. Park, D. Wu, S. G. Rhee and M. I. Simon, 1992 Mem-bers of the Gq a subunit gene family activate phospholipase C bisozymes. J. Biol. Chem. 267: 16044–16047.
Lee, S. B., and S. G. Rhee, 1995 Significance of PIP2 hydrolysis andregulation of phospholipase C isozymes. Curr. Opin. Cell Biol. 7:183–189.
Li, R., S. Waga, G. Hannon, D. Beach and B. Stillman, 1994 Dif-ferential effects by the p21 CDK inhibitor on PCNA-dependentDNA replication and repair. Nature 371: 534–537.
Martin, G., D. Viskochil, G. Bollag, P. McCabe, W. Crosier et al.,1990 The GAP-related domain of the neurofibromatosis type 1gene product interacts with ras p21. Cell 63: 843–849.
Measday, V., L. Moore, R. Retnakaran, J. Lee, M. Donoviel et al.,1997 A family of cyclin-like proteins that interact with thePho85 cyclin-dependent kinase. Mol. Cell. Biol. 17: 1212–1223.
Nasmyth, K., and K. Tatchell, 1980 The structure of transposableyeast mating type loci. Cell 19: 753–764.
Nishizuka, Y., 1992 Intracellular signaling by hydrolysis of phos-pholipids and activation of protein kinase C. Science 258: 607–614.
O’Keefe, S., J. Tamura, R. Kincaid, M. Tocci and E. O’Neil,
1992 FK506- and CsA-sensitive activation of the interleukin-2promoter by calcineurin. Nature 357: 692–694.
O’Neil, E., A. Kaffman, E. Jolly and E. O’Shea, 1996 Regulationof PHO4 nuclear localization by the PHO80-PHO85 cyclin-CDKcomplex. Science 271: 209–212.
Ogawa, N., K. Noguchi, Y. Yamashita, T. Yasuhara, N. Hayashi etal., 1993 Promoter analysis of the PHO81 gene encoding a 134kDa protein bearing ankyrin repeats in the phosphatase regulonof Saccharomyces cerevisiae. Mol. Gen. Genet. 238: 444–454.
Ogawa, N., K.-I. Noguchi, H. Sawai, Y. Yamashita, C. Yompakdee
et al., 1995 Functional domains of Pho81p, an inhibitor ofPho85p protein kinase, in the transduction pathway of Pi signalsin Saccharomyces cerevisiae. Mol. Cell. Biol. 15: 977–1004.
Payne, W., and M. Fitzgerald-Hayes, 1993 A mutation in PLC1, acandidate phosphoinositide-specific phospholipase C gene fromSaccharomyces cerevisiae, causes aberrant mitotic chromosome seg-regation. Mol. Cell. Biol. 13: 4351–4364.
Peter, M., and I. Herskowitz, 1994 Direct inhibition of the yeastcyclin-dependent kinase Cdc28-Cln by Far1. Science 265: 1228–1231.
Pollard, T., S. Almo, S. Quirk, V. Vinson and E. Lattman,
1994 Structure of actin-binding proteins: insights about func-tion at atomic resolution. Annu. Rev. Cell. Biol. 10: 207–249.
Riles, L., J. E. Dutchik, A. Baktha, B. K. McCauley, E. C. Thayer etal., 1993 Physical maps of the six smallest chromosomes of Sac-charomyces cerevisiae at a resolution of 2.6 kilobase pairs. Genetics134: 81–150.
Robzyk, K., and Y. Kassir, 1992 A simple and highly efficient pro-cedure for rescuing autonomous plasmids from yeast. NucleicAcids Res. 20: 3790.
Rose, M. D., F. Winston and P. Hieter, 1990 Methods in Yeast Genet-ics: A Laboratory Course Manual, Cold Spring Harbor LaboratoryPress, Cold Spring Harbor, NY.
Rothstein, R. J., 1983 One-step gene disruption in yeast. MethodsEnzymol. 101: 202–211.
Salama, S. R., K. B. Hendricks and J. Thorner, 1994 G1 cyclindegradation: the PEST motif of yeast Cln2 is necessary, but notsufficient, for rapid protein turnover. Mol. Cell. Biol. 14: 7953–7966.
Sambrook, J., E. F. Fritsch and T. Maniatis, 1989 Molecular Clon-
ing: A Laboratory Manual, Cold Spring Harbor Laboratory Press,Cold Spring Harbor, NY.
Santos, R. C., N. C. Waters, C. L. Creasy and L. W. Bergman,
1995 Structure-function relationships of the yeast cyclin-depen-dent protein kinase, PHO85. Mol. Cell. Biol. 15: 5482–5491.
Schneider, K. R., R. L. Smith and E. K. O’Shea, 1994 Phosphate-regulated inactivation of the kinase PHO80-PHO85 by the CDKinhibitor PHO81. Science 266: 122–125.
Schomerus, C., and H. Kuntzel, 1992 CDC25-dependent induc-tion of inositol 1,4,5-trisphosphate and diacylglycerol in Saccharo-myces cerevisiae by nitrogen. FEBS Letts. 307: 249–252.
Schurr, A., and E. Yagil, 1971 Regulation and characterization ofacid and alkaline phosphatase in yeast. J. Gen. Microbiol. 65:291–303.
Serrano, M., G. J. Hannon and D. Beach, 1993 A new regulatorymotif in cell-cycle control causing specific inhibition of cyclinD/CDK4. Nature 366: 704–707.
Sherr, C. J., and J. M. Roberts, 1995 Inhibitors of mammalian G1cyclin- dependent kinases. Genes Dev. 9: 1149–1163.
Sikorski, R. S., and P. Hieter, 1989 A system of shuttle vectors andyeast host strains designed for efficient manipulation of DNA inSaccharomyces cerevisiae. Genetics 122: 19–27.
Swartzman, E. E., M. N. Viswanathan and J. Thorner, 1996 ThePAL1 gene product is a peroxisomal ABC transporter in the yeastSaccharomyces cerevisiae. J. Cell Biol. 132: 549–563.
Tanaka, K., M. Nakafuku, T. Satoh, J. Marshall, J. Gibbs et al.,1990 S. cerevisiae genes IRA1 and IRA2 encode proteins thatmay be functionally equivalent to mammalian ras GTPase activat-ing proteins. Cell 60: 803–807.
Timblin, B. K., K. Tatchell and L. W. Bergman, 1996 Deletion ofthe gene encoding the cyclin-dependent kinase Pho85 alters gly-cogen metabolism in Saccharomyces cerevisae. Genetics 143: 57–66.
Toh-e, A., and T. Shimauchi, 1986 Cloning and sequencing of thePHO80 gene and CEN15 of Saccharomyces cerevisaie. Yeast 2: 129–139.
Toh-e, A., Y. Ueda, S. Kakimoto and Y. Oshima, 1973 Isolation andcharacterization of acid phosphatase mutants in Saccharomycescerevisiae. J. Bacteriol. 122: 727–738.
Touhara, K., J. Inglese, J. Pitcher, G. Shaw and R. Lefkowitz,
1994 Binding of G protein bg-subunits to pleckstrin homologydomains. J. Biol. Chem. 269: 10217–10220.
Towler, D., J. Gordon, S. Adams and L. Glaser, 1988 The biologyand enzymology of eukaryotic protein acylation. Annu. Rev. Bio-chem. 57: 69–99.
Valius, M., and A. Kazlauskas, 1993 Phospholipase C-g1 and phos-phatidylinositol 3 kinase are the downstream mediators of thePDGF receptor’s mitogenic signal. Cell 73: 321–334.
Waksman, M., Y. Eli, M. Liscovitch and J. Gerst, 1996 Identifica-tion and characterization of a gene encoding phospholipase Dactivity in yeast. J. Biol. Chem. 271: 2361–2364.
Weiss, A., and D. R. Littman, 1994 Signal transduction by lympho-cyte antigen receptors. Cell 76: 263–274.
Wu, D., A. Katz and M. Simon, 1993 Activation of phospholipase Cb2 by the a and bg subunits of trimeric GTP-binding protein.Proc. Natl. Acad. Sci. USA 90: 5297–5301.
Yoko-o, T., Y. Matsui, H. Yagisawa, H. Nojima, I. Uno et al.,1993 The putative phosphoinositide-specific phospholipase Cgene, PLC1, of the yeast Saccharomyces cerevisiae is important forcell growth. Proc. Natl. Acad. Sci. USA 90: 1804–1808.
Yoshida, K., N. Ogawa and Y. Oshima, 1989 Function of the PHOregulatory genes for repressible acid phosphatase synthesis inSaccharomyces cerevisiae. Mol. Gen. Genet. 217: 40–46.
Communicating editor: M. Johnston