transient stimulation deoxyribonucleic acid- ribonucleic …jvi.asm.org/content/6/1/58.full.pdf ·...
TRANSCRIPT

JOURNAL OF VIROLOGY, JUIY 1970, p. 58-68 Vol. 6, No. 1Copyright © 1970 American Society for Microbiology Printed in U.S.A.
Transient Stimulation of Deoxyribonucleic Acid-Dependent Ribonucleic Acid Polymerase andHistone Acetylation in Human EmbryonicKidney Cultures Infected with Adenovirus
2 or 12: Apparent Induction of HostRibonucleic Acid Synthesis
NADA LEDINKOPutnam Memorial Hospital Institute for Medical Research, Benniiigtonl, Vermont 05201
Received for publication 2 March 1970
The synthesis of cell-specific ribonucleic acid (RNA) appeared to be stimulatedin human embryonic kidney (HEK) cultures infected with adenovirus 2 or 12.Deoxyribonucleic acid (DNA)-RNA hybridization experiments revealed that by44 to 70 hr after infection with either virus, the relative amount of pulse-labeledRNA capable of hybridizing with HEK cell DNA increased considerably; such RNAwas detected in both nuclear and cytoplasmic fractions. The main increase inapparent host RNA synthesis was preceded by (i) a relatively early transientstimulation of the DNA-dependent RNA polymerase activity in isolated nuclei, and(ii) a small but consistently observed increase in the rate of acetylation of lysine-richand arginine-rich histone fractions. The Mn2+-(NH4)2SO4 and Mg2+-activated RNApolymerase reactions measured in nuclei isolated from cells infected with adenovirus2 or 12 were stimulated at about the same time; a rapid loss of polymerase activityfollowed. The augmentation of the two RNA polymerase reactions found in adeno-virus 12-infected cells was independent of protein synthesis. After the initial increase,the acetylation rate of histones of cells infected with adenovirus 2 or 12 declined,until late in infection it was approximately 40 to 70% of the control cell rate.
Adenovirus infection of human embryonickidney (HEK) cultures appears to induce aninitial small burst, followed by an inhibition ofhost deoxyribonucleic acid (DNA) synthesis rela-tively early after infection (5, 17, 32, 36). Thisspecific inhibition does not involve degradationof cell DNA into acid-soluble or acid-insolublematerial (17). The inhibition of the replication ofthe host genome has been noted as well in otheradenovirus-infected human cells systems (9, 10,21, 25). The effect of adenovirus infection on thetranscription of the host chromosome has alsobeen investigated. In studies with KB cells, it wasfound that the overall rate of ribonucleic acid(RNA) synthesis increases initially to a smallextent relatively early after infection (25) and isthen inhibited (9, 25). Both viral- and cellular-specific RNA species, however, are synthesizedin appreciable amounts even late in infection (2,9, 33).
Infection of HEK cells with adenovirus 2 or12 results in a significant stimulation of the rateof total RNA synthesis throughout most of theperiod of virus maturation and an increase inthe net RNA content of infected cells (17). Someof the reasons underlying the enhancement ofRNA metabolism were studied. In this communi-cation, evidence is presented which suggests thatinduction of cell-specific RNA synthesis occurs inHEK cells infected with adenovirus 2 or 12; thisinduction is apparently preceded by transientincreases of the Mg2+-activated and Mn2+-(NH4)SO4-activated DNA-dependent RNA po-lymerase reactions and in the capacity for histoneacetylation.
MATERIALS AND METHODSCells and virus. The cultivation of stocks of non-
oncogenic type 2 and oncogenic type 12 humanadenoviruses and the plaque method of virus assay
58
on May 7, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

INDUCTION OF HOST RNA SYNTHESIS
have been described elsewhere (15). The multiplicityof infection given refers to input multiplicity (plaque-forming units adsorbed per cell).
Infection of cells. Confluent or nearly confluentmonolayer HEK cell cultures, containing about 2 X106 cells in a 60-mm petri dish, were washed twicewith Hanks balanced salt solution, and 0.2 ml of virussuspension was added onto the cell layer. The givenmultiplicity was high enough to infect all or practicallyall of the cells, as determined by the infective centerassay (15). After adsorption for 1 hr at 37 C, the cellswere washed twice, covered with 3 ml of Eagle'sminimal essential medium containing 4%c fetal calfserum and 1 mM arginine, and incubated at 37 C in ahumidified 5%10 C02-air mixture. Control cultures werepropagated under the same conditions, but no viruswas added. Ten or more cultures were collected forvarious determinations at stated times.
DNA-dependent RNA polymerase activity. Nucleiand cytoplasm were prepared by using proceduresdescribed previously (16). Isolated nuclei were assayedfor the Mg2+-activated and Mn2+-(NH4)2SO4-acti-vated RNA polymerase reactions (14, 34). The enzymeactivity was assayed by measuring the conversion of3H-cytosine triphosphate (CTP) into an acid-insolubleproduct. This conversion was directly proportionalto the amount of enzyme added to the reactionmixture. The incubation mixture for the Mn2+-activated RNA polymerase reaction, carried out inthe presence of (NH4)2SO4, contained, in 0.5 ml: 50,umoles of tris(hydroxymethyl)aminomethane (Tris)-hydrochloride buffer (pH 7.9); 1.5 /Mmoles of MnC12;10 Amoles of NaF; 2.5 ,umoles of 2-mercaptoethanol;0.05 ml of a solution of (NH4)2SO4, saturated at roomtemperature and adjusted to pH 7.5 with NH40H;60 ,ug each of guanosine triphosphate (GTP), uridinetriphosphate (UTP), and adenosine triphosphate(ATP); 0.05 ,uc of 3H-CTP (tetralithium salt; 1.26c/mmole); and 0.1 to 0.2 ml of a nuclear suspensioncontaining 100 to 200 ,g of protein.The incubation mixture for the Mg2+-activated
reaction, carried out in the absence of (NH4)2SO4,contained, in 0.5 ml: 50 ,umoles of Tris-hydrochloridebuffer (pH 7.9); 2.5 ,umoles of MgCl2; 30,moles ofKCI; 3 Mmoles of NaF; 10 ,moles of cysteine; 2.5Amoles of 2-mercaptoethanol; 60 ug each of GTP,UTP, and ATP; 0.05 MAc of 3H-CTP (1.26 c/mmole);and 0.1 to 0.2 ml of a nuclear suspension containing100 to 200,ug of protein.The assay mixtures were incubated for 30 min at
37 C, at which time the reaction was stopped by theaddition of 5 ml of 0.5 N HC104 at 0 C. The acid-insoluble radioactivity was measured as describedelsewhere (18). Cytoplasmic extracts, or mixtures ofnuclear and cytoplasmic fractions, were assayed in thepresence of an ATP-generating system containing10,moles of phosphoenolpyruvate and 0.1 mg ofpyruvate kinase (11).
Protein determination. Protein was estimated by themethod of Lowry et al. (20) by using crystallinebovine serum albumin as the standard.
Thymidine kinase assay. The assay for thymidinekinase has been described in detail (15).
Extraction of HEK-cell DNA. HEK-cell DNA was
extracted from frozen pellets of noninfected cells byusing procedures outlined previously, with the excep-tion that three phenol extractions were made beforethe ribonuclease treatment (17).
Extraction of labeled nuclear or cytoplasmic RNA.At different times, HEK cell cultures were labeledwith uridine-5-3H (10 ,c/ml, 21.3 c/mmole) for 2 hrat 37 C. The cultures were then washed three timeswith cold Hanks balanced salt solution, and nucleiand cytoplasm were isolated by using the procedureof Penman (24). For hybridization experiments,nuclear RNA was extracted by using the methodof Penman (24), and the cytoplasmic RNA wasisolated by the method of Peacock and Dingman (23).The rate of labeling of the nuclear or cytoplasmic
RNA was determined from the incorporation of 3H-uridine into RNA as described previously (14). Thecultures were labeled and then fractionated intonuclear and cytoplasmic fractions as described above.DNA-RNA hybridization. The procedure was
based on the method of Gillespie and Spiegelman(8). Labeled RNA in 0.2 X SSC (0.15 M NaCl plus0.015 M sodium citrate) was heated before use for 15min at 100 C and then cooled rapidly in ice. HEK-cell DNA immobilized on membrane filters wasannealed with RNA in a final volume of 1.5 ml of 2X SSC at 66 C for 20 hr. The radioactivity remainingafter ribonuclease treatment was measured. Theamount of radioactivity bound to blank filters wassubtracted from that bound to DNA filters.
Acetate labeling of histones. Portions of cell cultureswere exposed to sodium acetate-2-'4C (3 MAc/ml, 34to 39 mc/mmole) for 1 hr at 37 C. The cells werefractionated into nuclear and cytoplasmic fractionsby using techniques described previously (16), exceptthat MgCl2 was omitted in the isolation procedure.This was done to avoid a possible loss of histone, sinceit has been shown that the use of MgCl2 during isola-tion of rat liver nuclei resulted in some loss of histone(6). Histone isolation was performed as outlinedbelow.
Evidence that the labeled acetate was incorporatedas acetyl groups attached to histones was indicated bythe following results. The labeled acetate was notremoved when the total histones, isolated as describedbelow, were treated with chloroform-methanol (2:1)to remove any lipids. Furthermore, over 80%7o of the"4C-acetate remained after treatment of histones with16% trichloroacetic acid at 90 C for 15 min to removeany polysaccharides and nucleic acids. In addition,since control and infected cultures contained approxi-mately the same amount of total histone (unpublisheddata), infection apparently did not induce net histonesynthesis. This suggests that any incorporation oflabeled acetate as amino acids, e.g., glutamate oraspartate, will be similar in both control and infectedcultures. Finally, the presence of radioactive acetylgroups in histones was verified by acid hydrolysis ofthe histones and steam distillation of labeled aceticacid by using methods described previously (1). Mostof the radioactivity was recovered in this manner.
Preparation of total histone. The isolated nuclei,prepared in the absence of MgCl2 as outlined above,were washed with 88% ethanol-0.01 N HCI to remove
VOL. 6, 1970 59
on May 7, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

LEDINKO
a tryptophan-containing protein fraction (27). Aftercentrifugation at 1,000 X g for 10 min at 0 C, thenuclei were resuspended in 0.5 ml of 0.25 N HCl for30 min at 0 C (29). After the nuclei were collected bycentrifugation, a second extraction was carried out,and the extracts were combined and tested for proteincontent.
Preparation of histone fractions. Using nucleiisolated in the absence of MgCl2 as outlined above,fractionation of histones into Fl, F2a, F2b, and F3fractions was performed by using method 2 of Johns(13), as modified by Sekeris et al. (31). To avoidpossible losses entailed by the isolation procedure,the following carrier histones, 10 mg each of calfthymus types II, III, and IV, were added at thebeginning of the extraction procedure to the nucleisuspended in 12 ml of absolute ethanol-1.25 N HCI(4:1). The radioactivity of aliquot portions of eachfraction was tested as described elsewhere (15).
RESULTSGrowth of adenoviruses 2 and 12 in HEK cells.
The growth characteristics of adenovirus 2 and 12in HEK cell cultures have been described (15).New mature adenovirus 2 appeared at approxi-mately 18 hr postinfection, and new adenovirus12 appeared at about 22 hr. Synthesis of adeno-virus 2 was complete at 40 to 50 hr, and that ofadenovirus 12 was complete at 50 to 60 hr.
Requirements for DNA-dependent RNA polym-erase activity. Isolated mammalian cell nucleihave been reported to synthesize different RNAspecies depending on whether the assay systemcontains Mg2+ at low ionic strength or Mn2+ plusa high concentration of (NH4)2S04 to raise theionic strength (26, 35). In the presence of Mg2+at low ionic strength, the nuclei synthesized pre-dominantly a guanine-cytosine-rich (ribosomal-like) RNA (22, 26, 35), whereas in the presence ofMn2+ at high ionic strength a more DNA-likeRNA was made (26, 35). These findings appearto reflect the presence of distinct nucleolar andnucleoplasmic DNA-dependent RNA polymer-ases (30).The Mg2+-activated and Mn2+-(NH4)2SO4-
activated RNA polymerase reactions were as-sayed in isolated noninfected HEK cell nuclei(Table 1). In both assay systems, all four ribo-nucleoside triphosphates were needed for optimalactivity, and most of the enzyme activity was lostwhen deoxyribonuclease, ribonuclease, or acti-nomycin D was added to the nuclei. A smallamount of activity did not appear to be depend-ent on the presence of all four ribonucleosidetriphosphates, suggesting some homopolymerformation. The ATP-generating system (phos-phoenolpyruvate-pyruvate kinase) did not appearto be essential. Omission of the (NH4)2S04 fromthe Mn2+-(NH4)2SO4 assay system resulted in a70% loss of enzyme activity. Addition of
(NH4)2S04 to the Mg2+-containing assay mix-ture, on the other hand, had no appreciableeffect.
Stimulation of RNA polymerase activity wasconsiderably more pronounced using the Mn2+-(NH4)2S04 assay, as compared to the Mg2+-con-taining system; the factor of increase was approx-imately 7. Similar findings were reported withisolated rat liver nuclei (35). This stimulation ofRNA synthesis has been related primarily to theionic strength of the assay medium which waspresumably responsible for a dissociation of pro-teins from DNA (12).
Changes in DNA-dependent RNA polymerasesduring infection. Changes in the Mn2+-(NH4)2S04-activated RNA polymerase activity were studied atvarious times after infection of HEK cells withadenovirus 2 or 12. Infection with either virusinduced a transient activation of the Mn2+-(NH4)2S04-activated RNA polymerase (Fig. 1).An approximately twofold increase in enzymeactivity was observed at 14 hr after infectionwith adenovirus 2 and at 21 hr after adenovirus12 infection. Thereafter, a relatively rapid lossof the RNA polymerase activity occurred. By 70hr postinfection, the activity in infected cellsdecreased to about 40 to 60% below control cellactivity. In contrast, the activity of thymidinekinase began to increase at approximately 20 hrafter infection of HEK cells by adenovirus 2 or12 and reached a maximum at about 50 hr post-infection, as was reported previously (15).
0 20 40 60 80 20 40 60 80
10
2
0
FIG. 1. DNA-dependent RNA polymerase andthymidine kinase activities in HEK cells infected withadenovirus 2 or 12. The multiplicity of adenovirus 2was 22 to 30 and that of adenovirus 12 was 14 to 21.Solid line: the Mn2+-(NH4)2SO4-activated RNA poly-merase activity relative to that of noninfected cells.The activity of control cell nuclei at 14 hr postinfectionwas 81 to 90 pmoles of 3H-CTP incorporated intoacid insoluble material per mg protein in 30 min at37 C. Thereafter, the activity either remained the sameor decreased by 50% of this value by 70 hr. Brokenline: thymidine kinase activity expressed as picomolesof thymidine phosphorylated per microgram proteinin 30 min at 37 C.
60 J. VIROL.
on May 7, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

INDUCTION OF HOST RNA SYNTHESIS
Preliminary results indicated that no differ-ences were detected between the MnI+-(NH4)2S04-activated polymerases of adenovirus12-infected (21-hr nuclei) and control noninfectednuclei with respect to their requirements forribonucleoside triphosphates and (NH4)2SO4(Table 1). In addition, an approximately equalsensitivity to the addition of deoxyribonuclease,ribonuclease, or actinomycin D was found.The activities of the Mn2-(NH4)2SO4- and
Mg'+-activated RNA polymerases were thencompared at different times after adenovirus 2or 12 infection (Fig. 2). A transient stimulationof both RNA polymerase activities was observedat about the same time. The Mg2+-activatedRNA polymerase subsequently decreased toreach the control cell level by approximately 50hr after infection with either adenovirus andapparently remained at approximately this levelup to 70 hr. The significance of the disparity ofthe inhibition of the two RNA polymerase reac-tions is not known, but it may be suggested that
TABLE 1. Factors affecting DNA-dependent RNApolymerase activity in isolated noninfected
HEK cell nuclei
Assay system for R-NApolymerase activitya
Reaction mixtureMn2+- g2
(NH4) 2SO4 Mg'+Complete........ 78 11Omit UTP................... 24 3.8Omit GTP................... 18 4.2Omit ATP................... 17 2.4Omit UTP, GTP, ATP....... 25 4.1
Add deoxyribonuclease (25jig) at time zero ..... ...... 31 3.1
Add ribonuclease (50 ,ug) at30 min'.................... 21 2.6
Add actinomycin D (5,g) attime zero.................. 16 3.2
Omit (NH4)2SO4............. 24Add (NH4)2SO4 10Add phosphoenolpyruvate (5
,umoles) plus pyruvate ki-nase (10,ug) ................ 62 9.1
a Picomoles of 3H-CTP incorporated into acid-insoluble material per milligram protein in 30 minat 37 C. The rate of incorporation of 3H-CTP waslinear with time for approximately 10 min forboth assay systems. Approximately 90% of thereaction was complete in 30 min with the Mn2+-(NH4)2SO4 system, and approximately 80% wascomplete in 30 min with the Mg2+ assay system.
I Additional incubation was carried out at 37 Cfor 30 min.
0o 20 40 60 80 20 40 60 80
HOURS
FIG. 2. Mn12+-(NH4) 2SO4- and Mg2+-activated RNApolymerase activities of HEK cells inifected withadenovirus 2 or 12 relative to that of nonzinfected cells.See legend of Fig. I for coniditions of inzfectioni and theMn2+-(NH4)2SO4 activity of conitrol cells throughoutthe experiment. The Mg2+-activated RNA polymeraseactivity of conztrol cell nuclei at 14 hr postinfection was12 to 14 pmoles of 3H-CTP incorporated into acid-insoluble material per mg protein in 30 min at 37 C.The activity did not decrease appreciably during theexperimental period.
the activities of the two polymerases are regu-lated by different control mechanisms.The addition of 100 to 200 ,ug of highly po-
lymerized salmon sperm DNA to the two assaysystems did not increase the activity of the RNApolymerase in the control nuclei and nuclei iso-lated from cells infected with adenovirus 2 or 12for 70 hr.
Effect of puromycin on the adenovirus-inducedstimulation of DNA-dependent RNA polymeraseactivity. Puromycin, a potent inhibitor of proteinsynthesis (15), was used to determine whetherthe increase in RNA polymerase activity foundin HEK cells infected with adenovirus 12 de-pended on new protein synthesis. Puromycinadded to a final concentration of 4 X 10- M inthe culture medium abolished the induction ofthymidine kinase activity, as was reported previ-ously (15), but had no apparent effect on themagnitude of increase of the Mn*-(NH4)2SO4- orMg'+-activated RNA polymerase reactions(Table 2). The stimulation of the two RNApolymerase activities was not, therefore, depend-ent on protein synthesis.
Tests for inhibitor of Mn2+-(NH4)SO4-acti-vated RNA polymerase. To learn whether thecytoplasm of infected cells contained an inhibitorof the Mn2-(NH4)2SO4-activated RNA polym-erase, nuclei isolated from noninfected cells weremixed with the cytoplasm isolated from cells in-fected with adenovirus 12 for 70 hr. (The infectednuclei from these cells had only 40% of theenzyme activity found in nuclei isolated from
VOL. 6, 1970 61
on May 7, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

LEDINKO
TABLE 2. Effect of puromycini on the adenovirus12-iniduced stimuilationi of DNA-dependent
RNA polymerase activity"
RNA polymerase activity, Ratio ofactivity of
Time Time of -infected toafter puro- Noninfected Adenovirus 12- noninfectedvirus mycin cells infected cells cellsaddi - addi- _
tion tion Mn2/ Mn2V Mn2±/(NH4) 2 Mg' (NIH4) 2 (NH4)2 MIg+SO4 SO4 SO4
hlr kr
14 -c 91 14 84 12 0.92 10.8614 91 14 84 12 0.92 0.86
21 -c 83 14 159 25 1.92 1.7914 58 12 110 21 1.89 1.75
a The multiplicity of adenovirus 12 was 25.Puromycin hydrochloride (4 X 10-5 M) was addedto infected or control cultures at 14 hr postinfec-tion. The amount of puromycin added inhibited80 to 90%7 of protein synthesis, as determined bymeasuring the uptake of 3H-L-leucine into acid-insoluble material (15).
b RNA polymerase activity measured by theMn'+-(NH4)2SO4 or Mg2+ assay systems wasexpressed as picomoles of 3H-CTP incorporatedinto acid-insoluble material per milligram of pro-tein in 30 min at 37 C.
c No puromycin added.
control cells, whereas the infected cytoplasmcontained approximately 3%6 of the activity foundin the nuclei.) The activity of the mixture wasapproximately the value expected from the sumof the activities of the individual fractions in themixture. No inhibitor was thus detected in thecytoplasmic fractions prepared from infectedcells.
Attempts to demonstrate a dissociable inhibi-tor in the nuclei of infected cells were made bymixing control nuclei with nuclei isolated fromcells infected with adenovirus 12 for 70 hr. Adissociable inhibitor, however, was not detectedsince the activity of the Mn2+-(NH4)2SO4-acti-vated RNA polymerase of the mixture was similarto the value expected from the sum of the activi-ties of both components.
Induction of host RNA synthesis in HEK cellsinfected with adenovirus 2 or 12. The possibilitywas considered that the observed stimulation ofRNA polymerase activity was related to anadenovirus-mediated effect on the host DNAtemplate, either to increase the fraction of DNAactive in RNA synthesis or to increase the tem-plate efficiency of certain loci. The followingfindings suggest that cell-specific RNA wasinduced after adenovirus infection.A stimulation of total RNA synthesis in HEK
cultures infected with adenovirus 2 or 12 hasbeen reported (17). Cultures infected for 38 hrwith either virus incorporated three to four timesas much 3H-uridine into RNA as did noninfectedcultures. Moreover, the net RNA synthesized byinfected cultures exceeded that of control cells(17). In the present experiments, a study wasmade of the rate of incorporation of 3H-uridineinto the nuclear and cytoplasmic RNA fractionsprepared from noninfected cells and from cellsinfected by adenovirus 2 or 12 for various times(Table 3). Stimulation by infection with eithervirus of the incorporation of the label into RNAisolated from both nuclear and cytoplasmic frac-tions was observed relatively early after infection(20 to 22 hr) as well as late (68 to 70 hr); theincrease varied by a factor of approximately 2to 4.The amount of 3H-uridine incorporated into
the acid-soluble pool of HEK cells infected withadenovirus 2 or 12 for 38 to 63 hr was two tothree times that of noninfected cells (17). Itremains to be determined whether infectionaffects the equilibration of 3H-uridine with thetotal cellular UTP pools. The estimation of therate of RNA synthesis in the present experimentsis based on the assumption that the specificactivity of the RNA precursor in the acid-soluble pool was the same in the infected andcontrol cells.The possibility existed that the RNA synthesis
in the infected cells continued at the preinfection
TABLE 3. Inzcorporationz of 'H-uridine for 2 hr inttoniuclear and cytoplasmic RNA fractionts of
adenzovirus 12-inzfected and noninfrctedHEK cell cultuiresa
Fraction of counts/min ofCounts/min of 3H-uridine incorporated per3H-uridine infected cell relative to control
incorporated p>erTime 103 noninfectedpost- cells Adenovirus Adenovirus
infection 2-infected 12-infected
Nucleus Cyto- Nucleus Cyto Nucleus Cyto-plasm plasm plasm
hr
1-3 16.60 5.83 1.02 0.88 0.93 1.1420-22 13.16 4.94 1.36 1.46 1.57 1.9144-46 11.64 2.99 2.17 2.20 2.46 3.9468-70 8.47 2.19 1.94 3.47 2.25 2.73
a HEK cells were infected with adenovirus 2at a multiplicity of 32 or with adenovirus 12 at amultiplicity of 21. At indicated times, replicateinfected and control cultures were exposed for 2hr to 3H-uridine (10lc/ml, 21.3 c/mmole). Therate of RNA synthesis was determined from theincorporation of 3H-uridine into RNA.
62 J. VIROL.
on May 7, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

INDUCTION OF HOST RNA SYNTHESIS
TABLE 4. Inzductioni of cell RNA synthesis in HEK cell cultures intfectedwith adenzoviruts 2 or 12a
Cell fraction
NucleusControlAdenovirus 2-infectedAdenovirus 12-infected
CytoplasmControlAdenovirus 2-infectedAdenovirus 12-infected
NucleusControlAdenovirus 2-infectedAdenovirus 12-infected
CytoplasmControlAdenovirus 2-infectedAdenovirus 12-infected
NucleusControlAdenovirus 2-infectedAdenovirus 12-infected
|CytoplasmControlAdenovirus 2-infectedAdenovirus 12-infected
NucleusControlAdenovirus 2-infectedAdenovirus 12-infected
CytoplasmControlAdenovirus 2-infectedAdenovirus 12-infected
Counts per minper,ug of extracted
RNA
39.0 X 10340.3 X 10338.7 X 103
2.2 X 1032.1 X 1032.4 X 103
35.2 X 10339.1 X 10340.4 X 103
1.7 X 1032.0 X 1032.7 X 103
18.4 X 10344.4 X 10329.1 X 103
0.75 X 1031.9 X 103
2.6 X 103
15.7 X 10326.5 X 10325.5 X 103
0.85 X 1031.8 X 1031.4 X 103
Hybrid
Counts/min
1,9401,979 (1.02)b1,843 (0.95)
2,4402,269 (0.93)2,538 (1.04)
1,3152,262 (1.72)2,801 (2.13)
1,2251,948 (1.59)1,936 (1.58)
8801,549 (1.76)2,086 (2.37)
4601,412 (3.07)4,490 (9.76)
260317 (1.22)333 (1.28)
70378 (5.4)
1,267 (18.1)
Per cent ofinput
0.300.280.32
0.720.780.82
0.260.320.40
0.700.820.58
0.280.220.28
0.580.720.99
0.250.210.23
0.770.690.70
a HEK cells were infected with adenovirus 2 at a multiplicity of 32 or with adenovirus 12 at a multi-plicity of 21. Samples of 5 X 107 infected as well as control noninfected cells were labeled for 2 hr with750 ,c of 3H-uridine (21.3 c/mmole). The RNA was extracted from nuclear and cytoplasmic fractionsand dissolved in 0.1 X SSC. Approximately 15 to 25 ,ug of nuclear RNA or 100 to 180 ,g of cytoplasmicRNA was hybridized to 18,ug of HEK cell DNA. This variation in concentration range for each RNAdid not appear to affect appreciably the amount of hybrid formed. Saturating RNA concentrations werenot determined. Incubation of HEK cell 3H-RNA with salmon sperm DNA (50,Mg) did not result inhybridization.
b Fraction of 3H-RNA hybridized relative to control.
rate, but, since adenovirus-infected cells do notdivide (15), the increase of RNA on a per cellbasis gave the appearance that there was a
stimulation of the rate of RNA synthesis. Thispossibility is not supported by the following con-
siderations. The specific activity (counts per minper microgram) of the nuclear and cytoplasmicRNA extracted from cells infected for 46 or 70hr was substantially higher than that of the
control cells (Table 4). The number of non-infected cells, moreover, increased only by a
factor of 1.14 in 70 hr. During this time, the rateof RNA synthesis of control cultures progres-sively declined (Table 3) as did that of DNAsynthe3is (15). These observations indicate thatthe dense HEK cultures employed were approach-ing or were in the stationary phase of growth.To determine whether host RNA synthesis was
Timepostinfection
hIr
1-3
20-22
44-46
68-70
VOL. 6, 1970 63
on May 7, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

LEDINKO
induced, DNA-RNA hybridization experimentswere carried out. Control and adenovirus-infected cultures were given 3H-uridine for 2-hrperiods at various times. The cultures were thenfractionated into nuclear and cytoplasmic frac-tions. The RNA was extracted from each frac-tion and hybridized with cell DNA (Table 4).Labeled RNA that hybridized with cell DNAwas detected in both nuclear and cytoplasmicfractions. Approximately 1%CO or less of theradioactive RNA formed hybrids with HEK cellDNA. The per cent of total counts hybridizedwas 0.21 to 0.40 for the nuclear RNA speciesand 0.58 to 0.99 for the cytoplasmic RNAspecies. The reason for this low hybridization isnot known; one possibility is that only a smallproportion of the DNA sequences was repre-sented in the RNA. At 20 to 22 hr and at 44 to46 hr, the amount of newly synthesized hostnuclear RNA of cells infected with adenovirus 2or 12 was about twice that of the control cells.Between 44 and 70 hr after infection by adeno-virus 2, the amount of radioactivity present incytoplasmic RNA that hybridized with hostDNA, relative to control RNA, increased by afactor of 3 to 5; after adenovirus 12 infection,there was an increase by a factor of 10 to 18. Atthese times, the rates of total RNA as well as oftotal DNA synthesis of infected cultures substan-tially exceeded those of control cultures (15, 17);a significant increase in thymidine kinase andDNA polymerase activities in infected cells wasalso found (15, 16).
Histone acetylation in HEK cells infected withadenovirus 2 or 12. The possibility was consideredthat the number of host DNA sites available fortranscription increased after adenovirus infec-tion. In attempting to understand the control oftranscription in eukaryotic cells, it has beenspeculated that both histone and nonhistone pro-teins play a role (7). According to one attractivehypothesis, at least some of the histones maydetermine the state of differentiation, and non-histone proteins may determine the rate of RNAsynthesis in open parts of the genome; the re-moval of histone inhibition may be achieved bychemical modification of side chains, such asacetylation (7). Acetate incorporation into his-tones (acetylation) has been shown to be anormal function of histone metabolism (1, 26,27). Moreover, the activation of RNA synthesisin various systems, such as in lymphocytestreated with phytohemagglutinin, has been asso-ciated with histone acetylation (27, 28).To learn whether acetylation of histones could
be important in the process of host RNA synthe-sis in HEK cultures infected with adenovirus 2 or
12, control and infected cultures were exposedfor 1 hr to acetate-2-14C. The four main histonefractions, designated Fl, F2a, F2b, and F3 (13),were extracted from the isolated nuclei, and acomparison was made of the labeled acetateuptake into these fractions. The Fl and F2b frac-tions have been characterized as relatively lysine-rich, and the F2a and F3 fractions have beencharacterized as relatively arginine-rich (4).The rate of acetylation of the lysine-rich his-
tones of noninfected control cells declined grad-ually during a period of 68 hr, and, at this time,the incorporation of acetate was approximately50% of the initial rate (Fig. 3). The acetylationrate of the arginine-rich histones, on the otherhand, decreased more rapidly to approximately40% of the initial rate by 42 hr (Fig. 3). At 68hr, the rate of RNA synthesis of control cellswas approximately 50% or less of the initial rate(Fig. 3). The decrease in the rate of histone
I-
I-
l,c
Il I C II
0 20 '0 60 80 20 40 60 80HOLRS
FIG. 3. (A) Rate of acetylation of lysine-rich F1 andF2b and arginine-rich F2a and F3 histone fractions ofnoninfected and adenovirus 2-infected HEK cells. Seefootnote a of Table 5 for infection conditions. Atvarious times, samples of approximately 4 X 107replicate infected and control cells were exposed for Ihr to sodium acetate-2-'4C (3 ,c/ml, 34 to 39 mclmmole). The histone fractions were extracted from theisolated nuclei. Counts/min of labeled acetate are
given in terms of total histone. (B) Rate of overall RNAsynthesis of control and adenovirus 2-infected HEKcells. Portions of cell cultures were exposed to uridine-5-3H at indicated times (2 ,uc/ml, 3.6 clmmole) forI hr at 37 C, and the rate of RNA synthesis was
determined (14).
64 J. VIROL.
I
C->vQ
iz
.F.jAZEm
9
on May 7, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from
INDUCTION OF HOST RNA SYNTHESIS
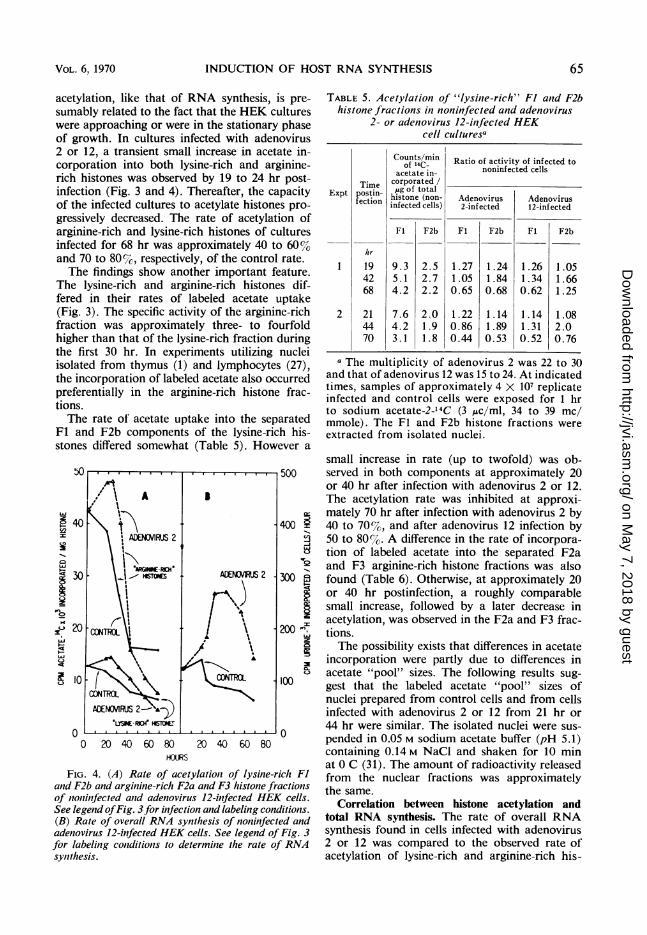
acetylation, like that of RNA synthesis, is pre-sumably related to the fact that the HEK cultureswere approaching or were in the stationary phaseof growth. In cultures infected with adenovirus2 or 12, a transient small increase in acetate in-corporation into both lysine-rich and arginine-rich histones was observed by 19 to 24 hr post-infection (Fig. 3 and 4). Thereafter, the capacityof the infected cultures to acetylate histones pro-gressively decreased. The rate of acetylation ofarginine-rich and lysine-rich histones of culturesinfected for 68 hr was approximately 40 to 60%oand 70 to 80%7, respectively, of the control rate.The findings show another important feature.
The lysine-rich and arginine-rich histones dif-fered in their rates of labeled acetate uptake(Fig. 3). The specific activity of the arginine-richfraction was approximately three- to fourfoldhigher than that of the lysine-rich fraction duringthe first 30 hr. In experiments utilizing nucleiisolated from thymus (1) and lymphocytes (27),the incorporation of labeled acetate also occurredpreferentially in the arginine-rich histone frac-tions.The rate of acetate uptake into the separated
Fl and F2b components of the lysine-rich his-stones differed somewhat (Table 5). However a
50 I I I I. -. 500
A
,640~ 400~lAD9O0/US 2
30 rosADOE?O RI2 3W
o0 2\XI
10 CON.OL100TR
v ~~~~~~~~~~v0 20 40 60 80 20 40 60 80
HOURS
FIG. 4. (A) Rate of acetylation of lysine-rich F1and F2b and arginine-rich F2a and F3 histone fractionsof noninfected aind adenovirus 12-infected HEK cells.See legend ofFig. 3for infection and labeling conditions.(B) Rate of overall RNA synthesis of noninfected andadenovirus 12-infected HEK cells. See legend of Fig. 3for labeling conditions to determine the rate of RNAsynthesis.
TABLE 5. Acetylation of "lysine-rich" F1 and F2bhistone fractions in noninfected and adenovirus
2- or adenovirus 12-infected HEKcell culturesa
TimeExpt postin-
fection
1
2
hr
194268
214470
Counts/minof 14C-
acetate in-corporated /
sg of totalhistone (non-infected cells)
IFl F2b
9.35.14.2
7.64.23.1
2.52.72.2
2.01.91.8
Ratio of activity of infected tononinfected cells
Adenovirus2-infected
Fl F2b
1.271.050.65
1.220.860.44
1.241.840.68
1.141.890.53
Adenovirus12-infected
Fl F2b
1.261.340.62
1.141.310.52
1.051.661.25
1.082.00.76
aThe multiplicity of adenovirus 2 was 22 to 30and that of adenovirus 12 was 15 to 24. At indicatedtimes, samples of approximately 4 X 107 replicateinfected and control cells were exposed for 1 hrto sodium acetate-2-1"C (3 4c/ml, 34 to 39 mc/mmole). The Fl and F2b histone fractions were
extracted from isolated nuclei.
small increase in rate (up to twofold) was ob-served in both components at approximately 20or 40 hr after infection with adenovirus 2 or 12.The acetylation rate was inhibited at approxi-mately 70 hr after infection with adenovirus 2 by40 to 70%,, and after adenovirus 12 infection by50 to 80%7. A difference in the rate of incorpora-tion of labeled acetate into the separated F2aand F3 arginine-rich histone fractions was alsofound (Table 6). Otherwise, at approximately 20or 40 hr postinfection, a roughly comparablesmall increase, followed by a later decrease inacetylation, was observed in the F2a and F3 frac-tions.The possibility exists that differences in acetate
incorporation were partly due to differences inacetate "pool" sizes. The following results sug-gest that the labeled acetate "pool" sizes ofnuclei prepared from control cells and from cellsinfected with adenovirus 2 or 12 from 21 hr or44 hr were similar. The isolated nuclei were sus-pended in 0.05 M sodium acetate buffer (pH 5.1)containing 0.14 M NaCl and shaken for 10 minat 0 C (31). The amount of radioactivity releasedfrom the nuclear fractions was approximatelythe same.
Correlation between histone acetylation andtotal RNA synthesis. The rate of overall RNAsynthesis found in cells infected with adenovirus2 or 12 was compared to the observed rate ofacetylation of lysine-rich and arginine-rich his-
VOL. 6, 1970 65
on May 7, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

66 LEDINKO
TABLE 6. Acetylation of "arginine-rich" F2a andF3 histone fractions in noninfected andadenovirus 2- or adenovirus 12-infected
HEK cell culturesa
TimeExpt postin-
fection
1
2
hr
194268
214470
Counts/min of14C-acetate
incorporated/Ag of total
histone (non-infected cells)
Ratio of activity of infected tononinfected cells
Adenovirus2-infected
F2a F3 F2a F3
30.710.011.6
43.421.816.8
7.96.24.4
4.13.92.7
1.250.790.45
1.260.780.37
1.201.270.53
1.151.250.39
Adenovirus12-infected
F2a F3
1.241.180.61
1.240.960.68
1.061.320.72
1.051.370.36
a See footnote a of Table 5. The F2a and F3histone fractions were extracted from isolatednuclei.
tones (Fig. 3 and 4). At 30 to 50 hr after infec-tion with either virus, the rate of total RNA syn-thesis per cell reached a maximal level of approx-imately three times that of noninfected cells.Even at 68 hr postinfection, infected culturesincorporated approximately twice as much 3H-uridine into RNA as did control cultures. Theoverall increase in the acetylation of histones,therefore, appeared to precede the main increasein total RNA synthesis.
Tests for greater fragility of nuclei from in-fected cells. The possibility was considered thatthe greater decrease in the acetylation rate of thelysine-rich and arginine-rich histones found ininfected than in noninfected cells was related toa loss of histones from the nuclei of infectedcells. The following experiment was performed inan attempt to ascertain whether nuclei from in-fected cells were more fragile than those fromnoninfected cells. Control cultures and cultures in-fected with adenovirus 2 or 12 for 69 hr werewashed and exposed for 1 hr to a mixture of14C-lysine and 3H-arginine. The cultures were thenfractioned into nuclear and cytoplasmic frac-tions; from the nuclei, the Fl, F2a, F2b, and F3histones were isolated. From the cytoplasm, thetotal histones were extracted with acid in thepresence of carrier histones and precipitated withacetone; then the above four histone fractionswere isolated from the precipitate.The total amount of labeled lysine incorporated
into the nuclear Fl and F2b, or of labeled argi-nine into the F2a and F3 fractions, was comparedwith the amount incorporated into these frac-tions isolated from the cytoplasm of infected and
J. VIROL.
TABLE 7. Test for loss of labeled Fl, F2a, F2b, andF3 histone fractions from the nucleus to thecytoplasm of noninfected and adenovirus
2- or adenovirus 12-infected HEKcell culturesa
Per cent cytoplasmic-labeled histonefraction found relative to nuclear fraction
Histonefraction
Noninfected Adenovirus Adenovirus2-infected 12-infected
Fl .........|. 1.8 1.6 7.7F2a 1.3 1.5 1.8F2b 1.5 6.4 4.5F3........ 0.9 1.5 2.1
a The multiplicity of adenovirus 2 was 22 andthat of adenovirus 12 was 15. At 69 hr after infec-tion, samples of approximately 2 X 107 controlas well as infected cells were washed and incu-bated at 37 C in Hanks balanced salt solutioncontaining 2 mm glutamine, 1% dialyzed fetalcalf serum, and both '4C-lysine (2.5 MAc/ml, 312mc/mmole) and 3H-arginine (5 ,uc/ml, 18 c/mmole).One hour later, the cultures were fractionatedinto nuclear and cytoplasmic fractions. From thenuclei, the Fl, F2a, F2b, and F3 histones wereisolated. From the cytoplasmic fraction, totalhistones were extracted with acid in the presenceof carrier histones, and the acid-soluble histoneswere then precipitated with 10 volumes of acetone.The precipitate was subsequently fractionatedinto the above four histone fractions. The totalamount of l4C-lysine incorporated into the Fland F2b fractions and of 3H-arginine incorporatedinto the F2a and F3 fractions was then deter-mined.
control cells (Table 7). In noninfected cells, onlyapproximately 1 to 2% of the radioactivity pres-ent in each nuclear fraction was found in thecorresponding fraction in the cytoplasm. A some-what higher proportion of labeled histones wasfound in the cytoplasm of infected than of non-infected cells. Approximately 2 to 6% and 2 to8% of the label present in the different nuclearhistones was recovered in the cytoplasm of cul-tures infected with adenovirus 2 and adenovirus12, respectively. The slightly greater loss of labelednuclear histones obtained from infected than fromnoninfected cells, however, was not sufficient toaccount for the degree of inhibition of histoneacetylation by infection (Tables 5 and 6).
DISCUSSION
The mechanisms involved in the control of genefunction in eukaryotic cells are unknown, but thefinding that acetylation is apparently a normalfunction of histone metabolism (1, 27, 28) has ledto speculation on the possible role of acetylated
on May 7, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

INDUCTION OF HOST RNA SYNTHESIS
histones in nuclear function. Arginine-rich his-tones have been shown to inhibit DNA-dependentRNA polymerase activity in vitro; acetylation ofthe histones removes the inhibitory activity (1).Furthermore, during periods of gene activation invarious systems, such as in regenerating liver orlymphocytes treated with phytohemagglutinin, theaccelerated acetylation of histones takes placebefore the increase in RNA synthesis (27, 28).These findings are consistent with the possibilitythat alterations of acetyl side chains of histonesplay a role in the modification of chromatinstructure and in the control of gene expression (1,7). The binding of histone to DNA, for example,may be altered by histone acetylation, providinga means of switching the RNA synthesis on or off.Even before the main increase in synthesis ofhost-specific RNA was found in adenovirus-in-fected HEK cells, an increase in the rate of histoneacetylation was detected, suggesting that a switch-ing on of RNA synthesis followed a prior altera-tion in the deoxyribonucleoprotein. It must stillbe determined, however, whether the increasedhistone acetylation was indeed responsible foralterations in gene activity.The heightened nuclear activity of adenovirus-
infected HEK cells, as evidenced by an increasein the rate of host RNA synthesis, may also be inpart due to an increase in the DNA-dependentRNA polymerase activity. The temporary increasein enzymatic activity, which was unaffected by thepresence of puromycin, could reflect the bindingof the enzyme to defined sites on the host DNAtemplate. The subsequent inhibition of RNApolymerase activity, also found in monkey cellsabortively infected with adenovirus 2 (14) and inKB cells infected with adenovirus 5 (2), could notbe explained by such relatively obvious possi-bilities as the unavailability or degradation ofprimer DNA (17) or the degradation of the RNAproduct, since control and infected cell nuclearfractions contained a similar small amount ofribonuclease activity (18). The decrease in polym-erase activity was followed relatively shortlythereafter by a decrease in histone acetylation. Ithas already been suggested that the acetylation ofhistones might be involved in the possible regula-tion of the inhibitory action of histones on theDNA primer activity for RNA synthesis. Theapproximate coincidence of the decrease in RNApolymerase activity and histone acetylation givessome support for this possibility.Recent studies on the structure of the bacterial
DNA-dependent RNA polymerase have revealedthat the enzyme contains a subunit which acts inconjunction with the other components to initiatetranscription at selected sites on the DNA (3).The decrease in polymerase activity observed in
adenovirus-infected cells could also result fromthe replacement of a factor(s) associated with theenzyme to bring about some modification oftranscription specificity. The role of the host RNApolymerase in the replication of adenovirus re-mains to be elucidated.
ACKNOWLEDGMENTS
This investigation was supported by Public Health Servicegrant CA10409-03 from the National Cancer Institute. N. L. isa Research Career Development Awardee from the NationalCancer Institute (5K03 CA 05278-04 VR).
The continued interest and support of Helene Toolan aregratefully acknowledged.
LITERATURE CITED
1. Allfrey, V. G., R. Faulkner, and A. E. Mirsky. 1964. Acetyla-tion and methylation of histones and their possible rolein the regulation of RNA synthesis. Proc. Nat. Acad. Sci.U.S.A. 51:786-794.
2. Bello, L. J., and H. S. Ginsberg. 1969. Relationship betweendeoxyribonucleic acid-like ribonucleic acid synthesisand inhibition of host protein synthesis in type 5 adeno-virus-infected KB cells. J. Virol. 3:106-113.
3. Burgess, R. R., A. A. Travers, J. J. Dunn, and E. K. F.Bautz. 1969. Factor stimulating transcription by RNApolymerase. Nature (London) 221:43-46.
4. Dick, C., and E. W. Johns. 1969. The biosynthesis of thefive main histone fractions of rat thymus. Biochim. Bio-phys. Acta 174:380-386.
5. Doerfier, W. 1969. Nonproductive infection of baby ham-ster kidney cells (BHK21) with adenovirus type 12. Virology38:587-606.
6. Dounce, A. L., and R. Ickowicz. 1969. The effect of themethod of isolation of cell nuclei on the ratios of nuciearproteins to DNA. Arch. Biochem. Biophys. 131:359-368.
7. Georgiev, G. P. 1969. Histones and the control of geneaction. Annu. Rev. Genetics 3:155-180.
8. Gillespie, D., and S. Spiegelman. 1965. A quantitative assayfor DNA-RNA hybrids with DNA immobilized on amembrane. J. Mol. Biol. 12:829-842.
9. Ginsberg, H. S., L. J. Bello, and A. J. Levine. 1967. Controlof biosynthesis of host macromolecules in cells infectedwith adenoviruses, p. 547-572, In J. S. Colter and W.Paranchych (ed.), Symposium on the molecular biologyof viruses. Academic Press Inc., New York.
10. Hodge, L. D., and M. D. Scharff. 1969. Effect of adenoviruson host cell DNA synthesis in synchronized cells. Virology37:554-564.
11. Holland, J. J., and J. A. Peterson. 1964. Nucleic acid andprotein synthesis during poliovirus infection of humancells. J. Mol. Biol. 8:556-573.
12. Hurlbert, R. B., E. G. Miller, and C. L. Vaughan. 1969.Control of RNA polymerase reactions in isolated nucleiand nucleoli. Advan. Enzyme Regul. 7:219-233.
13. Johns, E. W. 1964. Studies on histones. 7. Preparative methodsfor histone fractions from calf thymus. Biochem. J.92:55-59.
14. Ledinko, N. 1966. Changes in metabolic and enzymatic ac-tivities of monkey kidney cells after infection with adeno-virus. 2. Virology 28:679-692.
15. Ledinko, N. 1967. Stimulation of DNA synthesis and thy-midine kinase activity in human embryonic kidney cellsinfected by adenovirus 2 or 12. Cancer Res. 27:1459-1469.
16. Ledinko, N. 1968. Enhanced deoxyribonucleic acid polym-erase activity in human embryonic kidney cultures in-fected with adenovirus 2 or 12. J. Virol. 2:89-98.
17. Ledinko, N., and C. K. Y. Fong. 1969. Kinetics of nucleicacid synthesis in human embryonic kidney cultures in-
VOL. 6, 1970 67
on May 7, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

68 LEDINKO
fected with adenovirus 2 or 12: inhibition of cellulardeoxyribonucleic acid synthesis. J. Virol. 4:123-132.
18. Ledinko, N., and H. W. Toolan. 1968. Cortisone inhibitionof nuclear RNA polymerase of the human neoplasm,H. Emb. Rh. 4I, transplanted in rats. Endocrinology83:371-375.
19. Levine, A. J., and H. S. Ginsberg. 1968. Role of adenovirusstructural proteins in the cessation of host-cell biosyntheticfunctions. J. Virol. 2:430-439.
20. Lowry, 0. H., N. J. Rosebrough, A. L. Farr, and R. J. Ran-dall. 1951. Protein measurement with the Folin phenolreagent. J. Biol. Chem. 193:265-275.
21. Mantyjarvi, R., and W. C. Russell. 1969. Studies on DNAsynthesis in cells infected with adenovirus. J. Gen. Virol.5:339-350.
22. Maul, G. G., and T. H. Hamilton. 1967. The intranuclearlocalization of two DNA-dependent RNA polymeraseactivities. Proc. Nat. Acad. Sci. U.S.A. 57:1371-1378.
23. Peacock, A. C., and C. W. Dingman. 1967. Resolution ofmultiple ribonucleic acid species by polyacrylamide gelelectrophoresis. Biochemistry 6:1818-1827.
24. Penman, S. 1966. RNA metabolism in the HeLa cell nucleus.J. Mol. Biol. 17:117-130.
25. Pinia, M., and M. Green. 1969. Biochemical studies onadenovirus multiplication. XIV. Macromolecule and en-zyme synthesis in cells replicating oncogenic and non-oncogenic human adenovirus. Virology 38:573-586.
26. Pogo, A. O., V. C. Littau, V. G. Allfrey, and A. E. Mirsky1967. Modification of ribonucleic acid synthesis in nucleiisolated from normal and regenerating liver: some effectsof salt and specific divalent cations. Proc. Nat. Acad.Sci. U.S.A. 57:743-750.
27. Pogo, B. G. T., V. G. Allfrey, and A. E. Mirsky. 1966. RNAsynthesis and histone acetylation during the course of gene
J. VIROL.
activation in lymphocytes. Proc. Nat. Acad. Sci. U.S.A.55:805-812.
28. Pogo, B. G. T., A. 0. Pogo, V. G. Allfrey, and A. E. Mirsky.1968. Changing patterns of histone acetylation and RNAsynthesis in regeneration of the liver. Proc. Nat. Acad.Sci. U.S.A. 59:1337-1344.
29. Robbins, E., and T. W. Borun. 1967. The cytoplasmic syn-thesis of histones in HeLa cells and its temporal relation-ship to DNA replication. Proc. Nat. Acad. Sci. U.S.A.57:409-416.
30. Roeder, R. G., and W. J. Rutter. 1969. Multiple forms ofDNA-dependent RNA polymerase in eukaryotic organ-isms. Nature (London) 224:234-237.
31. Sekeris, C. E., K. E. Sekeri, and D. Gallwitz. 1967. Themethylation of the histones of rat liver nuclei in vitro.Hoppe-Seyler's Z. Physiol. Chem. 348:1660-1666.
32. Takahashi, M., T. Ogino, K. Baba, and M. Onaka. 1969.Synthesis of deoxyribonucleic acid in human and hamsterkidney cells infected with human adenovirus types 5 and12. Virology 37:513-520.
33. Thomas, D. C., and M. Green. 1966. Biochemical studieson adenovirus multiplication. XI. Evidence of a cyto-plasmic site for the synthesis of viral-coded proteins.Proc. Nat. Acad. Sci. U.S.A. 56:243-246.
34. Widnell, C. C., and J. R. Tata. 1964. A procedure for theisolation of enzymically active rat-liver nuclei. Biochem.J. 92:313-317.
35. Widnell, C. C., and J. R. Tata. 1966. Studies on the stimula-tion by ammonium sulfate of the DNA-dependent RNApolymerase of isolated rat liver nuclei. Biochim. Biophys.Acta 123:478-492.
36. Yamashita, T., anid H. Shimojo. 1969. Induction of cellularDNA synthesis by adenovirus 12 in human embryo kidneycells. Virology 38:351-355.
on May 7, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from