transcriptional repression by the proline rich homeodomain ... · the proline-rich homeodomain...
TRANSCRIPT

M4:04488 revised version
The Proline-Rich Homeodomain (PRH) protein recruits
members of the Groucho/TLE protein family to co-
repress transcription in haematopoietic cells
Tracey E. Swingler, Kirstin L. Bess, Jing Yao1, Stefano Stifani1 and
Padma-Sheela Jayaraman*
Department of Biochemistry, University of Bristol, University Walk,
Bristol BS81TD, U.K. 1Montreal Neurological Institute, McGill University, 3801 rue University,
Montreal, Quebec H3A2B4, Canada.
Running title: Transcriptional repression by PRH
*corresponding author
Phone: (+44-117-9289708)
Fax: (+ 44-117-9288274)
E-mail: [email protected]
Abbreviations used: PRH, Proline-Rich Homeodomain; TLE, transducin-like Enhancer of
split; GAL4AD, activation domain of GAL4; GAL4DBD, DNA binding domain of GAL4:
GST, glutathione S-transferase; TK, thymidine kinase; UAS, Upstream Activating
Sequence; 5-FOA, 5-Fluoroorotic Acid; Ura, Uracil; His, Histidine; X-gal, 5-bromo-4-
chloro-3-indolyl-β-D-galactopyranoside; MCS, multiple cloning site.
by guest on February 3, 2019http://w
ww
.jbc.org/D
ownloaded from

SUMMARY
The Proline-Rich Homeodomain protein (PRH/Hex) is important in the control
of cell proliferation and differentiation and in the regulation of multiple processes in
embryonic development. We have shown previously that PRH contains two domains
that can independently bring about transcriptional repression. The PRH homeodomain
represses transcription by binding to TATA box sequences, whilst the proline-rich N-
terminal domain of PRH can repress transcription when attached to a heterologous
DNA-binding domain. The Groucho/Transducin-like Enhancer of split (TLE) family
of proteins are transcriptional co-repressors that interact with a number of DNA-bound
transcription factors and play multiple roles in development. Here we demonstrate that
the proline-rich N-terminal domain of PRH binds to TLE1 in vitro and in yeast two-
hybrid assays. We show that PRH and TLE proteins are co-expressed in
haematopoietic cells and interact in co-immunoprecipitation assays. We demonstrate
that TLE1 increases repression by PRH in transient transfection assays and that
titration of endogenous TLE proteins by co-expression of Grg5, a natural trans-
dominant negative protein, alleviates transcriptional repression by PRH. Finally we
show that a mutation in the PRH N-terminal domain that blocks the PRH-TLE1
interaction in vitro eliminates co-repression. We discuss these results in terms of the
roles of PRH and TLE in cell differentiation and development.
Key words: Transcriptional repression, PRH, HEX, TLE, co-repressor,
haematopoiesis, trans-dominant negative,
2
by guest on February 3, 2019http://w
ww
.jbc.org/D
ownloaded from

INTRODUCTION
The Proline-Rich Homeodomain (PRH) protein also known as Hex
(Haematopoietically expressed), is an orphan homeodomain protein that functions as
an important regulator of haematopoiesis (1). PRH was first identified in human and
avian haematopoietic cells (1). PRH is strongly expressed in pluripotent
haematopoietic progenitors, in erythromyeloid and B-cell progenitors but not in T-
cell lineages (2-5) and is down-regulated in most haematopoietic lineages during
differentiation (3;6). PRH interacts with the growth control protein PML (7) and has
been shown to regulate cell growth or differentiation in a number of different tissues
(8-10). Up-regulation of PRH expression is linked with several lymphoid leukaemias
(11;12). In mice transplanted with bone marrow transduced with a PRH expressing
retrovirus PRH can act as an oncogene and cause T-cell derived lymphomas (11;12).
However PRH also acts as a tumour suppressor in some haematopoietic lineages.
PRH can inhibit oncogenic transformation by the translation initiation factor eIF4E
by disruption of the mRNA transport activity of eIF4E through a direct interaction
with eIF4E in the nucleus (13). Disruption of the nuclear localisation of PRH is
associated with the loss of this key regulatory function in a subset of myeloid
leukaemias (14). Thus the effects of PRH on growth and differentiation are both dose-
dependent and context-dependent. PRH also has an important role in the regulation of
early embryonic patterning (4;15;16). Indeed it plays a central role in the formation of
the vertebrate head and the formation of many endoderm-derived organs such as liver
and thyroid (5).
PRH functions as a transcriptional repressor in haematopoietic, liver, thyroid
and embryonic stem cells (15;17-19). However, it has also been reported to activate
transcription of its own gene in thyroid cells (20). PRH represses transcription in
3
by guest on February 3, 2019http://w
ww
.jbc.org/D
ownloaded from

haematopoietic cells by binding to TATA box sequences and TBP using the PRH
homeodomain. In addition however, the proline-rich PRH N-terminal repression
domain can repress transcription independently of the DNA binding activity of the
homeodomain by as yet unidentified mechanisms (21). Whilst the PRH N-terminal
repression domain is known to bind to PML, eIF4E and the proteosome subunit C8.
These interactions have not been shown to influence transcriptional repression in vivo
(7;13;22).
The TLE proteins are members of a family of co-repressor proteins that
includes the murine Grg proteins and the Drosophila Groucho protein. Groucho/TLE
family proteins are involved in many developmental decisions including: neuronal
and epithelial differentiation, segmentation and sex determination, differentiation of
haematopoietic, osteoblast and pituitary cells (23-27). Members of the Groucho /TLE
family do not have DNA binding activity but are instead recruited to DNA by
interactions with DNA binding proteins. In general, these proteins use a C-terminal
region known as the WD repeat domain to interact with DNA-binding proteins
(28;29) and an N-terminal glutamine-rich Q domain for tetramerisation (30-32).
Some members of the TLE family of proteins, such as Grg5, lack the WD repeat
regions but retain the oligomerisation Q domain. Over-expression of Grg5 can relieve
TLE-mediated co-repression, presumably because this protein retains the ability to
tetramerise with full-length TLE proteins (32-34). Once recruited to a promoter, the
Groucho/TLE proteins can bring about transcriptional repression by recruiting histone
deacetylases (35-37) or by directly interacting with histones (38), or with the basal
transcriptional machinery (39;40).
Here we show that the PRH N-terminal repression domain can interact with
TLE1 in vitro and in vivo using a short sequence of amino acids that corresponds to an
4
by guest on February 3, 2019http://w
ww
.jbc.org/D
ownloaded from

Engrailed homology (Eh-1) motif. Moreover we demonstrate that the interaction
between TLE1 and PRH is required for enhanced repression of transcription by PRH.
Furthermore, we show that titration of endogenous TLE proteins by Grg5 decreases
repression by PRH. Thus TLE proteins co-repress transcription with PRH.
5
by guest on February 3, 2019http://w
ww
.jbc.org/D
ownloaded from

EXPERIMENTAL PROCEDURES
Bacterial expression plasmids
A GST-tagged avian PRH N-terminus (GST-PRHN1-141) expression vector has
been described previously (21). PCR fragments encoding amino acids 1-125 or 61-
141 of avian PRH and flanked by 5’ SalI and 3’ SpeI restriction sites were cloned into
pGEX20T (Pharmacia) downstream of the GST moiety to create pGEX-PRHN61-141
and pGEX-PRHN1-125. The GST-tagged human PRH N-terminus expression vector
pGEX-MycPRHN1-132 was created as follows. Briefly, a DNA sequence encoding the
Myc-tagged human PRH N-terminus (amino acids 1-132) was cloned as a BamHI-
StuI fragment from pMUG1-MycPRH (see below) into pGEX20T (Pharmacia). The
PRH fragment was inserted between the unique BamHI site and the XbaI site in the
vector by first modifying the XbaI restriction site with Klenow enzyme to blunt the
XbaI restriction site. The pGEX-MycPRH F32E N1-132 plasmid that expresses the PRH
N-terminus carrying the F32E mutation was created exactly as described above except
that the PRH N-terminal fragment was obtained from pMUG1-MycPRH F32E (see
below). The DNA sequence of these plasmids and the plasmids described below were
verified by DNA sequencing.
Mammalian expression and reporter plasmids
The pTK-PRH and pTK-GAL reporter plasmids have been described
previously (21). pSV-β-Galactosidase Control Vector (pSV-lacZ) was obtained from
Promega. pBSK II-HPRH is a vector carrying the full-length human PRH cDNA and
was a gift from Dr. G. Manfioletti (University of Trieste). The mammalian expression
plasmid pFlag-Grg5 (pCMV-Tag1-AES) expresses the Grg5 protein tagged with the
pFlag epitope and has been described previously (41) and was a gift from Dr. T.
6
by guest on February 3, 2019http://w
ww
.jbc.org/D
ownloaded from

Okamoto (Nagoya University). The mammalian expression plasmid pCMV2-
FlagTLE1 contains TLE1 coding sequence in frame with the Flag epitope. The
FlagTLE1 construct was generated by first digesting a pBluescript-TLE1 plasmid
with BanII, followed by removal of protruding ends with T4 DNA polymerase and
recovery of a 1.6 kb fragment encoding the N-terminal region of TLE1. This fragment
was subcloned into pCMV2-Flag digested with EcoRV. A second pBluescript-TLE1
restriction fragment obtained after digestion with SmaI and encoding the remaining
half of TLE1 was then ligated in frame into the first ligation product to generate
pCMV2-FlagTLE1 expressing full-length TLE1.
The mammalian expression plasmid pMUG1-MycPRH expresses full-length
human PRH and was created as follows: pUHD15-1 (42) was modified by replacing
the sequence between the unique BamHI and EcoRI sites with a linker which destroys
these two restriction sites and contains a multiple cloning sequence (MCS). The
sequence of the linker is as follows: 5’ A ATT GGA TCC ATG GGA ATT CGA
GGT CGA CAG TGA 3’.The linker contains a translational start signal (bold) and
BamHI , NcoI, EcoRI, and SalI restriction sites. The resulting pMUG1 plasmid,
contains the CMV promoter with a MCS downstream. A BamHI-SmaI double-
stranded oligonucleotide encoding a Myc tag (Myc 9E10 epitope)
(5’GATCCATGGAACAAAAACTCATCTCAGAAGAGGATCTG 3’) and a SmaI-
EcoRI fragment from pBSK-HPRH carrying the human PRH coding sequence from
amino acid 7 was inserted between the BamHI and EcoRI sites in pMUG1. This
results in an expression construct where the PRH coding sequence was placed in
frame with the Myc tag and the ATG in the MCS.
7
by guest on February 3, 2019http://w
ww
.jbc.org/D
ownloaded from

Yeast plasmids
pACT2 contains the GAL4 AD upstream of a MCS (Clontech). To create
pACT2-HPRH a SmaI –EcoRI fragment from pBSK-HPRH carrying the PRH coding
sequence from amino acid 7, was ligated into pACT2 between the BamHI and EcoRI
restriction sites. The BamHI site in pACT2 was filled in using Klenow enzyme to
allow blunt end ligation with the PRH SmaI site. pGBT9-TLE1 contains the TLE1
cDNA in frame with the GAL4 DBD and has been described previously (31;31;43).
pAS2-1 contains the GAL4 DBD upstream of a MCS (Clontech). To create pAS2-1-
PRHN1-132 an EcoRI-StuI fragment from pBSK-HPRH encoding the N-terminal 132
amino acids of PRH was ligated between the EcoRI and SmaI sites of pAS2-1. An
oligonucleotide was inserted between the EcoRI site in pAS2-1 and the internal SmaI
site in HPRH to achieve the correct reading frame (5’
CATGCAGTACCCGCACCCC 3’). To create the deletion mutant pAS2-1-PRHN1-98,
the pAS2-1-PRHN1-132 construct was digested with BamHI and partially digested with
ApaI. An ApaI-BamHI oligonucleotide (5’ CGCCGCGCCCACG 3’) was then ligated
between the ApaI site located at amino acid 98 within the PRH amino acid sequence
and the unique BamHI site in the vector pAS2-1. pGAD424-TLE1 contains the full-
length TLE1 coding sequence and expresses a GAL4-TLE1 fusion protein.
pGAD424-TLE1Q-SP and pGAD424-TLE1WD contain the Q (amino acids 1-135),
Q-SP (amino acids 1-435) and WD (amino acids 444-770) domains of TLE1,
respectively. These plasmids have all been described previously (31;43).
Yeast two-hybrid assay
The two-hybrid assay (44) was carried out in yeast strain MaV203 (Clontech).
The two-hybrid assay and β-galactosidase assays in liquid culture were performed as
8
by guest on February 3, 2019http://w
ww
.jbc.org/D
ownloaded from

described by Fields and Song (44) and the Clontech manual. Colony lift assays were
performed by transferring yeast colonies onto 3MM filter paper. The yeast were then
lysed by rapidly freeze thawing in liquid nitrogen and then placed on filter paper
saturated with Z-buffer (60mM Na2HPO4, 40mM NaH2PO4, 10mM KCl, 1mM
MgCl2, 50mM β-mercaptoethanol) containing X-gal indicator (0.32mg/ml). The blue
colour for each transformant was compared after incubation of the filter at 37°C for
3hrs.
Mutagenesis
The QuikChange kit (Stratagene) was used for the mutagenesis of pMUG1-
MycPRH to PRH F32E and was used according to the manufacturer’s instructions.
The resulting mutant was fully sequenced to confirm the sequence change.
Expression and purification of tagged-PRH proteins
The human and avian GST-PRH fusion proteins were expressed in BL21
pLysS cells (Novagen). Fusion protein expression was induced with 1mM IPTG.
Cells were harvested and lysed by incubation with 100µl of lysozyme (1mg/ml) for
20 mins followed by sonication in PBS/1% Triton-X-100. GST-PRHN fusion proteins
were purified over glutathione-Sepharose 4B beads (Sigma) according to the
manufacturer’s instructions and snap frozen in liquid nitrogen. Aliquots of these
proteins were eluted with 10mM glutathione and assayed for purity by SDS-PAGE
followed by staining with Coomassie blue. Proteins were quantified using the Bio-
RAD phosphoric acid protein assay.
9
by guest on February 3, 2019http://w
ww
.jbc.org/D
ownloaded from

In vitro binding assays
The plasmid used for the in vitro transcription and translation of TLE1,
pcDNA3-TLE1, has been described previously (27;43). Transcription and translation
was carried out using a TNT kit (Promega) according to the manufacturer’s protocol.
Approximately 20 µg of each of the GST fusion proteins, or an equimolar amount of
GST protein as judged by Coomassie staining, were bound to 50µl of glutathione-
Sepharose 4B beads (Sigma) according to the manufacturer’s instructions. To assay
for specific interactions, 10µl of [35S] methionine-labelled in vitro translated protein
was added and incubated in binding buffer (20mM Hepes pH7.8, 200mM KCl, 5mM
MgCl2, 0.5mM DTT, 0.5% NP40, 50ng/µl BSA), with gentle agitation, for 60 mins at
4°C. The beads were then washed six times with 1ml of binding buffer. Bound
proteins were eluted by boiling the beads in protein sample buffer containing 1% SDS
and analysed by SDS-PAGE and fluorography.
Pull-downs and western blotting
Whole cell extract from 2 x 108 K562 cells was made as follows: the cell
pellet was washed in PBS twice and then resuspended in 1ml high salt lysis buffer
(500mM NaCl, 50mM Tris pH 7.5, 0.1% SDS, 0.1% NP-40). The cell suspension was
drawn up and down six times with a 3 X Monojet needle (1.1x50mm, 19Gx2”) and
then incubated on ice for 5 mins. The extract was centrifuged at maximum speed for 5
mins at 4°C in an Eppendorf microcentrifuge. Whole cell extract was added to
approximately 10µg of GST-HPRHN protein or 10µg of GST protein bound to
glutathione resin. After two hours at 4°C with tumbling, the resin was collected by
centrifugation in an Eppendorf microcentrifuge, washed three times in 1ml of wash
buffer (150mM NaCl, 50mM Tris pH 7.5, 0.1% SDS, 0.1% NP-40) and then
10
by guest on February 3, 2019http://w
ww
.jbc.org/D
ownloaded from

resuspended in 50µl 2X SDS loading buffer. All operations were carried out at 4°C
and in the presence of protease inhibitors. After SDS PAGE the proteins were
immunoblotted onto Immobilon-P membrane (Sigma). TLE proteins were detected
using a rat monoclonal pan TLE antibody raised against the conserved carboxyl-
terminal WD40 domain of Groucho/TLEs (25;45) and an ECL kit (Amersham). HC8
was detected with a mouse monoclonal antibody (Affiniti).
Immunofluoresence
K562 cells were grown to a density of 1x 106 cells/ml. 10mls of cells were
collected by centrifugation and resuspended in 1ml of PBS. Glass slides were coated
with poly-L-lysine (Sigma) for 10 mins at 20°C and then washed with water. The
washed K562 cells were incubated on the coated slide for 10 mins at 20°C and then
washed in PBS. The cells were fixed in paraformaldehyde for 30 mins, and then
rinsed twice in PBS and permeabilised by incubation in 0.1% Triton-X-100 in PBS
for 10 mins. The cells were then rinsed with PBS and incubated with PBSA (3% BSA
in PBS) for 20 mins to block non-specific antibody binding. After rinsing in PBS,
antibody staining was performed with a 1:10 dilution of a rat monoclonal pan TLE
antibody and a 1:10 dilution of a mouse polyclonal anti-PRH antibody for 1 hour at
20°C. The cells were rinsed in PBS twice, in PBSA twice, and then incubated with
secondary antibodies for 1 hour at 20°C. PRH was detected with a TRITC donkey
anti-mouse secondary antibody (Stratec) that had been preadsorbed for
immunoreactivity against rat antibodies. MycPRH was detected with a monoclonal
antibody (Santa Cruz). TLE was detected with a 1:100 dilution of a biotinylated rabbit
anti-rat secondary antibody (Vector Laboratories Inc.) and a 1:100 dilution of a
Fluorescein-labelled goat anti-Biotin tertiary antibody (Vector Laboratories Inc.).
11
by guest on February 3, 2019http://w
ww
.jbc.org/D
ownloaded from

Immunostained cells were viewed on a Leica DM IRBE confocal microscope and
imaging performed using Leica Confocal Software Version 2.00.
Cell culture and transient transfection assays
K562 cells were grown in glutamine added DMEM media (Sigma)
supplemented with 10% foetal calf serum (FCS) to a density of approximately 1x106
cells/ml. The cells were then collected by centrifugation and then resuspended in
media plus 10% FCS to a density of 5x107 cells/ml. 5µg of the luciferase reporter
plasmid with 5µg of the β-galactosidase reporter plasmid and the amount of
expression plasmids indicated in the Results, were electroporated into 1x107 cells
using a BIORAD Genepulser (200V, 975µF). The cells were rested for 10 mins and
then incubated overnight in 10ml of supplemented media. After 24 hours the cells
were harvested and luciferase activity assayed using the Promega Luciferase Assay
System according to the manufacturer’s instructions. β-galactosidase assays were
performed as an internal control for transfection efficiency. After subtraction of the
background the luciferase counts were normalised against the β-galactosidase value.
Co-immunoprecipitation assays
K562 cells (2 x 107) were co-transfected with 2µg pMUG1-MycPRH and
either 2µg pCMV2-FlagTLE1 or 2µg pCMV-Flag-Grg5 as described above. Five
transfections were pooled (1 x 108 cells) to make the nuclear extract for each co-
immunoprecipitation. Nuclear extracts were made by the method of Dignam et al.
(46) with modifications described by Dorn et al. (47). Nuclear extracts were incubated
with 4µl of a monoclonal anti-Myc9E10 antibody (Santa Cruz) for 30 mins at 4°C.
100µl of a 50% slurry of Protein G beads (Sigma) was then incubated with the nuclear
12
by guest on February 3, 2019http://w
ww
.jbc.org/D
ownloaded from

extracts for a further 90 mins at 4°C. After this time, the resin was collected by
centrifugation in an Eppendorf microcentrifuge (13,000 rpm for 1 min), washed three
times in 1ml of wash buffer B (150mM NaCl, 50mM Tris pH 7.5, 0.2% NP-40) and
then resuspended in 50 µl 2X SDS loading buffer. All operations were carried out at
4°C and in the presence of protease inhibitors. After SDS PAGE the proteins were
immunoblotted onto Immobilon-P membrane and TLE and Grg5 proteins were
detected using anti-Flag antibodies (Sigma). Approximately 10% of the nuclear
extract (200µg protein) was used for Western blotting experiments as described
above.
13
by guest on February 3, 2019http://w
ww
.jbc.org/D
ownloaded from

RESULTS
PRH and TLE1 interact in yeast
PRH contains two motifs that could be involved in an interaction with TLE
proteins. Amino acids 30-39 (TPFYIEDILG) contains a sequence that matches the
Engrailed homology (Eh-1) motif (FxIxxIL) identified originally in the Drosophila
homeodomain proteins Engrailed and Goosecoid and shown to mediate the interaction
of both these proteins with Groucho (48;49). The second is a putative ‘Runt/Hairy’
motif (LLWSPF amino acids 124-129 in PRH) which loosely resembles the
tryptophan containing motifs WRPY and WRPW that are used to recruit the Groucho
protein to the Runt and Hairy transcription factors respectively (50;51). To investigate
whether PRH might interact with TLE proteins we made use of the yeast two-hybrid
assay (Fig. 1). The human PRH cDNA was placed in frame with GAL4 activation
domain in the vector pACT2 to create pACT2-PRH (see EXPERIMENTAL
PROCEDURES). pGBT9-TLE1 contains the full-length TLE1 cDNA (amino acids 1-
770) in frame with the GAL4 DNA binding domain (31;43;43). pACT2-PRH and
pGBT9-TLE1 were co-transformed into yeast strain MaV203. This strain contains
integrated copies of the LacZ and His reporter genes under the control of GAL4-
dependent promoters. A functional interaction between the two hybrids in this strain
would be expected to produce β-galactosidase activity and histidine prototrophs.
Although expression of pGBT9-TLE1 with a vector containing the GAL4AD or
expression of pACT2-PRH with a vector containing the GAL4DBD did not result in
the production of β-galactosidase activity (Fig. 1B, columns 1 and 2), significant β-
galactosidase activity was detected when pGBT9-TLE1 and pACT2-PRH were co-
expressed (Fig. 1B, column 3). These data suggest that a functional transcription
factor is produced only when both proteins are present and therefore that PRH
14
by guest on February 3, 2019http://w
ww
.jbc.org/D
ownloaded from

interacts with TLE1 in this assay. However, the co-expression of PRH and TLE1
results in very poor growth of the transformed yeast strain when histidine is added to
the growth media and no growth in the absence of histidine (not shown). This
suggests that the interaction of the full-length PRH and TLE proteins is toxic for this
yeast strain.
To confirm that the PRH-TLE1 interaction occurs and to establish whether the
N-terminal domain of PRH is responsible for the interaction we repeated the yeast
two-hybrid assay with a truncated PRH construct (Fig. 1C). In this case, full-length
TLE1 (1-770) was fused to the GAL4 activation domain in pGAD424 to create
pGAD424-TLE1 (31;43). A fragment of the human PRH cDNA encoding the first 98
amino acids of PRH was cloned into the GAL4 DNA binding domain vector pAS2-1
to create pAS2-1-PRHN1-98 (see EXPERIMENTAL PROCEDURES). This fragment
of PRH lacks the Runt/Hairy-like motif (LLWSPF) described earlier but contains the
putative Eh-1 motif. These constructs were then transformed into yeast. To inhibit
leaky expression of the histidine gene transformants were assayed for growth on
dropout media plates containing 50mM 3-amino-triazol (50mM 3-AT) and
subsequently assayed for lacZ expression. Yeast co-transformed with PRHN1-98 and
TLE1 grew well on -HLT (+50mM 3AT) dropout medium whereas yeast transformed
with only one of the partners and the corresponding empty vector either did not grow
or grew very poorly on this medium (Fig. 1C). Moreover only yeast co-transformed
with PRHN1-98 and TLE1 resulted in colonies that produced β-galactosidase activity in
X-gal colony lift assays (Fig. 1C, compare sectors 7 and 2 on X-gal). Thus it appears
that the N-terminal 98 amino acids of PRH are sufficient for the interaction of PRH
with TLE1 and that the ‘Runt/Hairy’-like sequence found in PRH is not required for
the interaction.
15
by guest on February 3, 2019http://w
ww
.jbc.org/D
ownloaded from

The TLE1 protein is 770 amino acids long and has several functional domains
(Fig. 1A) (45). The N-terminal region of the protein (amino acids 1-135) is a
glutamine-rich tetramerisation domain known as the Q domain. The middle of the
protein (amino acids 135- 435) contains a glycine/proline rich G/P domain, a nuclear
localisation and CKII phosphorylation region (CcN) and a Ser/Thr/Pro-rich SP
domain. The C-terminus of the protein contains a WD-repeat domain (amino acids
444-770) that can mediate protein-protein interactions with transcription factors
(28;29;31;52). To determine which regions of TLE1 are involved in the interaction
between PRH and TLE1 a series of TLE1 deletion mutants were fused to the GAL4
activation domain in pGAD and co-transformed into yeast with pAS2-1-PRHN1-98.
The TLE deletion mutants pGAD-TLEQ (1-135), pGAD-TLEQ/SP (1-435) and
pGAD-TLEWD (444-770) have been described previously (31). Interestingly, when
co-transformed with pAS2-1-PRHN1-98 all of the TLE deletion mutants allowed
growth of the corresponding yeast transformants on –HLT (+50mM 3AT) media and
also resulted in β-galactosidase activity in X-gal filter lift assays (Fig. 1C, sectors 7-
10) and in liquid β-gal assays (not shown). These data suggest that PRHN 1-98 is able to
interact with both the Q domain at the N-terminus of TLE1 and also with the WD-
repeat domain at the C-terminus of TLE1.
To confirm the growth phenotypes of the yeast co-transformed with these
constructs we made use of the Ura- (+5-Fluoroorotic acid) counter-selection assay
(53). Yeast strain MaV203 is Ura- and contains an integrated copy of the Ura3 gene
under the control of a Gal4-dependent UAS. Only yeast carrying interacting proteins
become Ura+ and can convert 5-Fluoroorotic acid (5-FOA) to the highly toxic
compound 5-fluorouracil. As can be seen from the data in Fig. 1C, yeast containing
any of the TLE1 proteins and PRHN 1-98 die on media containing 5-FOA (Fig. 1C,
16
by guest on February 3, 2019http://w
ww
.jbc.org/D
ownloaded from

sectors 7-10, on FOA). In contrast, yeast transformed with the two empty vectors
pAS2-1 and pGAD424 (sector 1), or each of the TLE1 proteins with pAS2-1 (sectors
2-5), or PRHN 1-98 with pGAD424 (sector 6) show very limited growth on -HLT
(+3AT) media, no β-galactosidase activity, and growth on media containing 5-FOA.
Taken together these data confirm that PRH and TLE1 can interact in yeast.
Furthermore, these data suggest that both the Q domain and the WD domains of TLE1
are involved in the interaction with the first 98 amino acids of PRH,
Immunostaining of PRH and TLE proteins in K562 cells
To investigate further the biological significance of the interaction between
PRH and TLE proteins we next examined the distribution of these proteins in K562
haematopoietic cells. PRH is known to be strongly expressed in this cell line and
other PRH interacting proteins have been identified from yeast two-hybrid screens
with cDNAs obtained from this cell line (22;54). The K562 cell line was originally
obtained from a patient with Chronic Myeloid Leukaemia (CML) in blast crisis and in
culture these cells spontaneously give rise to multiple cell types, larger myeloblasts
and smaller myeloid cells at various stages of differentiation (55). To determine the
intracellular localisation of the endogenous PRH and TLE proteins in the K562 cell
line we used confocal laser microscopy and immunofluoresence. Figure 2A shows
endogenous TLE staining of K562 cells with a fluorescein labelled pan-TLE antibody
(green signal) and Figure 2B shows DNA staining with DAPI (blue signal). TLE is
present in both the nucleus and the cytoplasm of K562 blasts and smaller myeloid
cells. The staining pattern of the cells is not however uniform. For example, cells 1
and 2 both show a predominant cytoplasmic staining for TLE and cells 3 and 4 show
predominant nuclear staining for TLE (compare DAPI and fluorescein staining). The
17
by guest on February 3, 2019http://w
ww
.jbc.org/D
ownloaded from

differences in sub-cellular localisation of TLE proteins in these cells may reflect the
differentiation status of these cells. Certainly TLE proteins are known to become
more strongly associated with the nuclear compartment during the neural
differentiation of P19 embryonic carcinoma cells (56). Fig. 2C shows endogenous
PRH staining of the same cells with a TRITC labelled anti-PRH mouse polyclonal
antibody (red signal). This antibody stains both the nucleus and the cytoplasm of
K562 cells. In all cases when cells were stained without the primary antibody there
was little, if any, detectable immunofluoresence (data not shown). As expected, co-
immunofluoresence experiments using TRITC labelled anti-PRH mouse polyclonal
antibodies and the same fluorescein labelled pan-TLE rat monoclonal antibodies show
that a significant proportion, but not all K562 cells, contain both PRH and TLE
proteins in the nucleus (Fig. 2D). Figure 2 also shows a high magnification image of a
K562 cell, where endogenous PRH is present in the nucleus and cytoplasm (Fig. 2F,
TRITC staining) and endogenous TLE proteins are strongly nuclear (Fig. 2G, FITC
staining). Co-immunofluoresence of this cell shows that both endogenous PRH and
TLE proteins are present in the nucleus (Fig. 2H).
Since both TLE and PRH proteins can be found in both cytoplasmic and
nuclear compartments, we wanted to determine whether over-expressed TLE1 and
PRH proteins are predominantly cytoplasmic or nuclear. Transfection of K562 cells
with either pMUG1-MycPRH or pFLAGTLE1 followed by immunostaining with a
Myc monoclonal antibody or the rat panTLE antibody respectively, results in around
10% of cells expressing the transfected proteins. However, in both cases
immunofluoresence experiments show that the transfected proteins are located in the
nucleus (data not shown). Similarly, when K562 cells are co-transfected with both
PRH and TLE1 expression plasmids, both MycPRH and FlagTLE1 are strongly
18
by guest on February 3, 2019http://w
ww
.jbc.org/D
ownloaded from

localised to the nucleus. Figure 2I shows a high magnification transmitted light image
of a transfected K562 cell. Figures, 2J, and 2K show PRH (TRITC), TLE (FITC), and
DAPI staining of the same cell. Figure 2L shows co-localisation of TLE1 with PRH in
the nucleus of a transfected K562 cell.
PRH and TLE proteins interact in vitro and in K562 haematopoietic cells
To provide biochemical evidence for the interaction between PRH and TLE1
we carried out in vitro binding studies and pull-down experiments. The equivalent N-
terminal regions of human PRH (amino acid 1-132) and avian PRH (amino acids 1-
141) were expressed in bacteria as GST fusion proteins and partially purified. In vitro
transcription and translation was used to produce labelled TLE1. Glutathione beads
carrying GST or the GST-PRHN proteins were incubated with labelled TLE1 and then
washed extensively. Bound protein was eluted by boiling in SDS-PAGE loading
buffer and analysed by SDS-PAGE. Figure 3A shows that labelled TLE1 binds to the
human GST-PRHN1-132 (lane 3) and avian GST-PRHN 1-141 (lane 4) fusion proteins but
binds only very weakly, if at all, to GST alone (lane 2). Thus both the human and the
avian PRH N-terminal domains bind to TLE1 in vitro. To extend our in vitro binding
studies we determined whether the purified human PRH N-terminus is able to interact
with endogenous TLE proteins in K562 whole cell extracts. Human GST-PRHN 1-132
or GST alone were immobilised on glutathione-Sepharose beads and incubated with
K562 whole cell extracts. After extensive washing, bound proteins were eluted by
boiling in SDS-PAGE loading buffer and separated by SDS-PAGE. The bound
proteins were then probed with the pan TLE rat monoclonal antibody in a Western
blot. The GST-PRHN1-132 protein is clearly able to retain the endogenous TLE proteins
present in the whole cell extracts (Fig. 3B, lane 3). However, an equivalent amount of
19
by guest on February 3, 2019http://w
ww
.jbc.org/D
ownloaded from

GST (as judged by Coomassie staining) does not retain TLE (Fig. 3B, lane 2). We
conclude that the human PRH N-terminal protein purified from bacterial cells is able
to interact with the endogenous TLE proteins present in K562 cell extracts.
To confirm these results co-immunoprecipitation studies were carried out
using full-length PRH tagged with the Myc9E10 epitope (pMUG1-MycPRH) and
TLE1 tagged with a Flag epitope (pCMV2-FlagTLE1) (Fig. 3C). K562 cells were
transiently co-transfected with MycPRH and FlagTLE1 plasmids and nuclear extracts
were made from the transfected cells. Proteins in these nuclear extracts were
separated on SDS-PAGE gels and probed with an antibody raised against the Flag
epitope. Flag-tagged TLE1 proteins with a molecular weight of approximately 90kD
were detected with this antibody in Western blotting experiments (Fig. 3C, lane 1).
Shorter exposures show the presence of a doublet of TLE1 proteins (data not shown).
The same nuclear extract was incubated with either Protein G-Sepharose beads alone,
or with an anti-Myc mouse monoclonal antibody bound to Protein G-Sepharose
beads. After extensive washing the bound proteins were separated on SDS-PAGE gels
and probed with the anti-Flag antibody. In the presence of the Myc antibody, a
doublet of the same molecular weight as that of the TLE1 proteins is co-
immunoprecipitated (Fig. 3C, lane 3). This doublet is not present when Protein G-
Sepharose beads alone are incubated with nuclear extracts made from the transfected
cells (Fig. 3C, lane 2). Thus, FlagTLE1 can interact with MycPRH in haematopoietic
cells.
Some apparent protein-protein interactions occur indirectly and are observed
because two proteins both independently bind to contaminating DNA present in the
nuclear extract. For example, Oct-2 and the non-specific DNA binding subunits of the
Ku protein co-immunoprecipitate only in the presence of DNA, whereas the
20
by guest on February 3, 2019http://w
ww
.jbc.org/D
ownloaded from

interaction between Rb and E1A is not dependent on the presence of DNA (57).
Ethidium bromide (EtBr) intercalates into DNA and can abolish protein-DNA
interactions without disrupting protein-protein interactions. To determine whether the
PRH-TLE1 interaction requires the presence of DNA we carried out the co-
immunoprecipitation experiment with MycPRH and FlagTLE1 described above in the
presence of different concentrations of EtBr. The direct protein-protein interactions
between Rb and E1A are insensitive to EtBr concentrations of 0.2mg/ml (57). Figure
3C, shows that TLE co-immunoprecipitates with MycPRH even in the presence of
0.4mg/ml of EtBr (Fig. 3C, compare lanes 3 and 4). Since Ethidium bromide does not
appear to significantly affect the co-immunoprecipitation of TLE1 and PRH it is
likely that this is a protein-protein interaction that does not require promoter DNA.
To establish whether transfected MycPRH interacts with endogenous TLE
proteins, nuclear extracts were made from K562 cells alone or K562 cells transfected
with MycPRH. Proteins in these nuclear extracts were separated on SDS-PAGE gels
and probed with the rat pan-TLE antibody raised against the C-terminus of TLE
proteins. In the presence of transfected MycPRH, nuclear TLE proteins with a
molecular weight of approximately 90kD were detected in Western blotting
experiments (Fig. 3D, lane 1). Nuclear extracts expressing transfected MycPRH were
incubated with an anti-Myc mouse monoclonal antibody bound to Protein G-
Sepharose beads (Fig. 3D, lane 4) or incubated with Protein G-Sepharose beads alone
(Fig. 3D, lane 3). As a further control untransfected K562 nuclear extracts were
incubated with an anti-Myc mouse monoclonal antibody bound to Protein G-
Sepharose beads (Fig. 3D, lane 2). After washing the bound proteins were separated
on SDS-PAGE gels and probed with the pan-TLE antibody. Small amounts of TLE
proteins non-specifically bound to Protein G-Sepharose beads are detected by the pan-
21
by guest on February 3, 2019http://w
ww
.jbc.org/D
ownloaded from

TLE antibody even in the absence of the Myc antibody (Fig. 3C, lane 3) or in the
absence of transfected MycPRH (Fig. 3C, lane 2). However, in the presence of
transfected MycPRH, a doublet of the same molecular weight as that of the
endogenous TLE proteins is strongly co-immunoprecipitated by the Myc antibody
(Fig. 3D, lane 4). We conclude that TLE proteins can associate with PRH both in vitro
and in haematopoietic cells.
TLE proteins can co-repress transcription with PRH
To determine the biological significance of the interaction between PRH and
TLE1 we investigated whether TLE1 could function as a co-repressor of transcription
with PRH. We have shown previously that the reporter plasmid pTK-PRH is
repressed by chicken PRH in avian haematopoietic BM2 cells (21) and by human
PRH in K562 cells (22). The pTK-PRH reporter and the control reporter plasmid
pSV-β-gal were transiently transfected into K562 cells with 100ng of pMUG1-
MycPRH. As can be seen in Figure 4A, this amount of pMUG1-MycPRH decreases
pTK-PRH promoter activity to approximately 60% of its unrepressed activity
(compare columns 1 and 2). Co-transfection with 200ng of the TLE1 expression
plasmid pCDNA3-TLE1 results in increased repression by PRH (Fig. 4A, compare
columns 2 and 3). This effect although small, is highly reproducible. Transfection of
pCDNA3-TLE1 and the pTK-PRH reporter alone does not decrease reporter activity,
and if anything there is an increase in promoter activity (Fig. 4A, compare columns 1
and 4). This data suggests that TLE1 can co-repress transcription, albeit weakly, with
PRH in transient transfection assays.
There are a number of possible reasons why we might not observe very strong
co-repression between PRH and TLE1 in transient transfection assays. K562 cells
22
by guest on February 3, 2019http://w
ww
.jbc.org/D
ownloaded from

strongly express several TLE family members (TS and PSJ unpublished observations)
and one possibility is that transfection of the TLE1 plasmid into K562 cells does not
strongly increase the amount of TLE1 proteins in the cells available to interact with
PRH. A complementary approach to examine co-repression between PRH and TLE
proteins in these cells is to remove endogenous TLE proteins and measure any effects
on PRH activity. Several studies have shown that Grg5, a naturally occurring protein
that is related to the first 200 amino acids of Groucho/TLE, can tetramerise with TLE
proteins and function as a dominant negative regulator (32;32-34;34). We therefore
examined whether we could relieve repression by PRH by using Grg5 to titrate out
endogenous TLE proteins. Transfection of 100ng of pMUG1-MycPRH decreases
pTK-PRH promoter activity to approximately 60% of its unrepressed activity
(Fig. 4B, compare columns 1 and 2). Co-transfection of Grg5 with PRH relieves
repression by PRH in a dose-dependent fashion. In the presence of 5µg pFlag-Grg5
reporter activity returns to approximately 75% of its unrepressed level (Fig. 4B, lanes
3-5). The presence of pFlag-Grg5 did not similarly increase reporter activity in the
absence of PRH (Fig. 4B, lanes 6-8). In fact, 5µg pFlag-Grg5 decreases reporter
activity in the absence of PRH to approximately 65-70% of its normal level.
However, Flag-Grg5 does not appear to significantly affect the activity of the viral
CMV promoter that is used to express both MycPRH and Flag-Grg5 (data not shown).
Thus, repression of the TK-PRH promoter by PRH can be relieved by co-expression
of Grg5. To examine whether Grg5 can itself interact with PRH in K562 cells, we
carried out co-immunoprecipitation studies using pMycPRH and pFlag-Grg5. We
were unable to demonstrate co-immunoprecipitation of Grg5 and PRH, although in a
parallel experiment TLE1 and PRH were co-immunoprecipitated under wash
conditions of the same stringency (data not shown). Thus, whilst we cannot rule out
23
by guest on February 3, 2019http://w
ww
.jbc.org/D
ownloaded from

the possibility that Grg5 might bind directly to PRH and inhibit PRH-dependent
repression, our experiments indicate that most likely, the titration of TLE proteins by
Grg5 results in decreased repression by PRH. We conclude that endogenous TLE
proteins do co-repress with PRH and that over-expressed TLE1 can co-repress with
PRH.
Mapping the TLE interaction motif in PRH
The human and avian PRH proteins are strongly conserved (1) and both
proteins contain an Eh-1 motif within the first 60 amino acids of the PRH N terminus
(Fig. 5A). To determine whether this motif is required for the interaction of TLE
proteins with PRH we made use of three avian GST-PRHN fusion proteins in in vitro
binding assays. The N-terminus of avian PRH (amino acids 1-141) or N-terminal
fragments of avian PRH (amino acids 1-125 and 61-141) were expressed in bacteria
as GST fusion proteins and purified. In vitro transcription and translation was used to
produce labelled TLE1 and binding assays were carried out as described above.
Figure 5B shows that labelled TLE1 binds to the GST-PRHN 1-141 (lane 3) and GST-
PRHN 1-125 (lane 5) fusion proteins but does not bind to GST-PRHN 61-141 (lane 4) or to
GST alone (lane 2). These data confirm the interaction data in yeast which showed
that the ‘Runt/Hairy’ motif is not required for the PRH-TLE interaction. Furthermore,
these data strongly suggest that the Eh-1 domain located within the first 60 amino
acids of PRH is required for the interaction. To confirm these in vitro binding studies
we determined whether the purified GST-PRHN deletion proteins are able to bind to
endogenous TLE proteins in pull-down experiments. Bound proteins were probed
with pan TLE rat monoclonal antibody or with a mouse monoclonal antibody raised
against the C8 subunit of the proteosome. Figure 5C shows that endogenous TLE
24
by guest on February 3, 2019http://w
ww
.jbc.org/D
ownloaded from

proteins bind to GST-PRHN 1-141 (lane 3) and GST-PRHN 1-125 (lane 5) but do not bind
to GST-PRHN 61-141 (lane 4) or GST alone (lane 2). In contrast with this, Figure 5C
shows that another PRH interacting protein, proteosome subunit C8 binds to GST-
PRHN 1-141 (lane 3), GST-PRHN 1-125 (lane 5) and GST-PRHN 61-141 (lane 4) but not to
GST alone (lane 2). Thus whilst all the fusion proteins appear to be functional for
protein-protein interactions with C8, the GST-PRHN fusion protein lacking the first 60
amino acids of PRH is unable to interact with endogenous TLE proteins.
To establish whether the Eh-1 motif in PRH is responsible for the interaction
of PRH with TLE, we mutated phenylalanine 32 to glutamic acid (F32E) in the Myc-
tagged Human PRH mammalian expression plasmid pMUG1-MycPRH to create
pMUG1-MycPRHF32E. This amino acid is the most conserved within the Eh-1 motif
and changing this amino acid to glutamic acid has previously been shown to abolish
the interaction of the homeodomain proteins Goosecoid and Engrailed with Groucho
(49;58). The wild type human PRH N terminus (amino acids 1-132) and the mutated
PRH N terminus were transferred to a GST expression vector and GST-PRHN1-132 and
GST-PRHN1-132 F32E fusion proteins were expressed in bacteria and purified. In vitro
binding assays were carried out with these fusion proteins and in vitro transcribed and
translated TLE1 as described above. Figure 5D shows that labelled TLE1 binds to the
human GST-PRHN1-132 (lane 3) but binds only very weakly, if at all, to GST-PRHN 1-
132 F32E (lane 4) or to GST alone (lane 2). Pull-down experiments were also carried
out as described above with these fusion proteins and K562 nuclear extracts. Figure
5E shows that endogenous TLE proteins bind to the human GST-PRHN1-132 (lane 2)
but binds only very weakly, if at all, to GST-PRHN 1-132 F32E (lane 3) or to GST alone
(lane 1). In contrast, Figure 5E shows that proteosome subunit C8 binds to GST-
PRHN1-132 (lane 2) and GST-PRHN 1-132 F32E (lane 3) but binds only very weakly, if at
25
by guest on February 3, 2019http://w
ww
.jbc.org/D
ownloaded from

all, to GST alone (lane 1). Thus we conclude that the Eh-1 domain in PRH mediates
the interaction between PRH and TLE proteins present in K562 cells.
To confirm that the Eh-1 domain mediates the interaction between PRH and
TLE1 that occurs during co-repression of the TK promoter, we carried out transient
co-transfection assays. The pTK-PRH reporter and the control reporter plasmid pSV-
β-gal were transiently transfected into K562 cells with 100ng of pMUG1-
MycHPRHF32E. Figure 5F shows that pMUG1-MycHPRHF32E decreases pTK-PRH
promoter activity to approximately 60% of its unrepressed activity (compare columns
1 and 2). However, co-transfection of 200ng of the TLE1 expression plasmid
pCDNA3- TLE1 with pMUG1- MycHPRHF32E does not result in increased repression
by PRHF32E (Fig. 5F, compare columns 2 and 3). This data suggests that the Eh-1
domain mediates the interaction between PRH and TLE1 in vivo.
26
by guest on February 3, 2019http://w
ww
.jbc.org/D
ownloaded from

DISCUSSION
We have shown previously that the PRH N-terminal region contains a
transcriptional repression domain with potential Groucho/TLE interaction motifs (21).
Here we have demonstrated that PRH interacts with TLE proteins in haematopoietic
cells, in yeast and in vitro and that an Eh-1 motif in the PRH N-terminal region
mediates the interaction of these proteins. It has been documented previously that
Grg5 binds to TLE proteins and functions as a trans-dominant negative (32-34). We
have shown that titration of endogenous TLE proteins by Grg5 results in decreased
transcriptional repression by PRH suggesting that PRH and TLE associate in cells.
Furthermore over-expression of TLE1 increases repression by PRH but does not
increase repression by a PRH protein containing a mutation in the Eh-1 motif that
eliminates binding to TLE1 in vitro. Taken together, these observations strongly
suggest that PRH and TLE proteins interact in K562 haematopoietic cells and that
TLE proteins function as co-repressors for PRH. It is of interest to note that the PRH
F32E mutation does not significantly decrease the ability of PRH to repress
transcription in transient transfection assays. This situation is similar to that reported
by Tolkunova et al., for Engrailed (En) (48). En has multiple repression domains and
repression mechanisms. En interacts with the Groucho co-repressor and the
interaction is essential for full repression in Drosophila embryos but the Eh-1 F to E
mutation in En resulted in less than a 10% reduction of En repression activity in
standard transient transfection assays (48). Like En, PRH has several repression
mechanisms and therefore the full effect of this mutation might only be clearly seen in
genomic DNA in the natural in vivo context.
Groucho/TLE-dependent repressors have been found to function as long-range
repressors, that is, they block promoter function in a distance and orientation-
27
by guest on February 3, 2019http://w
ww
.jbc.org/D
ownloaded from

independent manner. The ability to recruit TLE proteins to a promoter represents an
important mechanism for PRH-dependent transcriptional repression. There are at least
two domains in Groucho/TLE proteins that are known to interact with DNA bound
transcription factors: the N-terminal Q domain and the C-terminal WD-repeat domain.
The Q domain (amino acids 1-135) is a tetramerisation domain that is essential for
transcriptional repression (23;30) and sufficient for the interaction of TLE with the
transcription factors TCF and Blimp-1 (PRD1-BF1) (32). In contrast with the
interaction of TLE proteins with TCF and Blimp-1, the WD-repeat domain in
Groucho is essential and sufficient for direct interactions with the Engrailed and Hairy
transcription factors (59). However, the Q and WD-repeat domains in TLE1 are both
involved in interactions of TLE proteins with the Runt-related transcription factor
RUNX2/Cbfa1 and the winged-helix protein BF-1 (60;61). Similarly, we have shown
that the PRH N-terminal domain interacts with both the Q domain and the WD-repeat
domain in TLE1. It has been suggested that the employment of several TLE protein-
protein interactions domains allows greater specificity of interaction (62).
Alternatively, the use of more than one TLE protein-protein interaction domain might
confer greater stability to a TLE-PRH complex. Certainly the stable interaction of
another homeodomain protein, Pax5, with TLE proteins requires two separate
domains in TLE1 and two separate domains in Pax5 (63).
Although the TLE proteins were not originally identified in the haematopoietic
compartment, it has become apparent that TLE proteins interact with a number of
transcription activator proteins that are found in haematopoietic cells, including the
human Runt domain protein, AML1 (also known as RUNX-1/CEBP-2/
PEBP2)(27;64), T-cell factor (TCF) also known as Lymphoid Enhancer Factor (LEF-
1) (64), and the B-cell specific factors PRDI-BF1 and Pax5 (32;63). In each case the
28
by guest on February 3, 2019http://w
ww
.jbc.org/D
ownloaded from

interaction of the transcription activator with TLE proteins results in transcriptional
repression. Thus, TLE proteins are likely to be co-repressors for PRH in a number of
haematopoietic lineages. In addition, both PRH and TLE are expressed in a variety of
tissues in the developing embryo. PRH is expressed in the pre-gastrulation embryo
and is one of the earliest markers for dorsoventral patterning (4). Later, in the early
embryo, PRH is expressed in the anterior endoderm (head organiser region) adjacent
to tissues expressing the Goosecoid homeodomain protein (15), that co-represses
transcription with TLE proteins (49). Interestingly, PRH is essential for forebrain
development (16) and TLE proteins have been implicated in neuronal differentiation
(65) and dorsoventral patterning of the neural tube (66). Expression of PRH has also
been detected in endothelial precursor cells (4;67) and in osteoblasts (3). TLE
expression occurs post-gastrulation in mesoderm derivatives that go on to produce the
endothelial cells of the lining of the heart, muscle, and bone (25). In Drosophila
Groucho is part of the Notch signalling cascade. It is believed that the role of this
signalling pathway is to bring about a halt in embryonic neurogenesis so that cells that
were committed to become neuroblasts are made competent to enter the epidermal
lineage instead (68). Studies with TLE proteins have shown that they play a similar
developmental role (25). TLE expression is elevated in undifferentiated or
transformed epithelial cells and down-regulated as epithelial cells differentiate. This
suggests that TLE proteins are involved in the maintenance of the undifferentiated
state (69). Significantly, Notch signalling also plays a fundamental role in
haematopoietic development; in general Notch signalling promotes self-renewal and
inhibits differentiation (70). PRH is found in haematopoietic and endothelial
progenitors (3;4), and a variety of tissues derived from endoderm including thyroid
and liver (5). However PRH is absent in terminally differentiated haematopoietic cells
29
by guest on February 3, 2019http://w
ww
.jbc.org/D
ownloaded from

and/or endothelial cells (3;4). Furthermore, our studies on myeloid stem cells have
shown that PRH expression is down-regulated during differentiation of early myeloid
progenitors towards myeloblasts or erythrocytes (6). Thus, PRH expression is
associated with the undifferentiated state in haematopoietic and endothelial cells.
Here we have shown that undifferentiated haematopoietic blasts contain nuclear PRH
and TLE proteins. It is tempting to speculate that the interplay of TLE proteins and
PRH in these cells contributes to the control of cell differentiation. Further work will
be required to reveal in what other cell types TLE proteins act as co-repressor proteins
for PRH and the importance of the TLE1-PRH interaction in differentiation and
development.
30
by guest on February 3, 2019http://w
ww
.jbc.org/D
ownloaded from

AKNOWLEDGEMENTS
With many thanks to Dr. Kevin Gaston for comments on the manuscript and for
many useful discussions. Thanks also to Dr. Mark Jepson for help with the confocal
laser microscope studies at the Bristol University MRC Cell Imaging Facility. K. L.
Bess and T. E. Swingler were supported by B.B.S.R.C. studentships. Dr. S. Stifani is
a Scholar of the Fonds de la Recherche en Sante du Quebec. Dr. P. -S. Jayaraman is
grateful to the MRC for a Career Development Award.
31
by guest on February 3, 2019http://w
ww
.jbc.org/D
ownloaded from

FIGURE LEGENDS
Figure 1 TLE1 binds to PRH in yeast cells.
(A) A schematic representation of the PRH and TLE1 proteins (not to scale). (B) Top-
A schematic of the interacting fusion proteins GAL4DBD-TLE1 and GAL4AD-PRH
at the GAL4 UAS. Below- A bar chart of the β-galactosidase activity obtained after
transformation with (1) pACT2-PRH and pAS2-1, (2) pACT2 and pGBT9-TLE1, and
(3) pACT2-PRH and pGBT9-TLE1. The experiment was performed several times and
the results of one experiment performed in triplicate are shown. The data is presented
as the mean and standard error. (C) Top- A schematic of the fusion proteins
GAL4DBD-PRHN and GAL4AD-TLE1 at the GAL4 UAS. Below- Growth
phenotypes on -LT media, -HLT (+ 50mM 3AT) media, -LT (+ 0.2% 5FOA) media,
and β-galactosidase activity of yeast transformants in a colony-lift assay (X-gal). The
sectors contain: (1) pAS2-1 and pGAD424, (2) pAS2-1 and pGADTLE1, (3) pAS2-1
and pGADTLE1 WD, (4) pAS2-1 and pGADTLE1 Q, (5) pAS2-1 and pGADTLE1
Q-SP (6) pAS2-1-PRHN1-98 and pGAD424, (7) pAS2-1-PRHN1-98 and pGADTLE1, (8)
pAS2-1-PRHN1-98 and pGADTLE1 WD, (9) pAS2-1-PRHN1-98 and pGADTLE1 Q,
(10) pAS2-1-PRHN1-98 and pGADTLE1 Q-SP.
Figure 2 PRH and TLE expression in K562 cells.
(A) Endogenous TLE staining of K562 cells using a FITC labelled pan-TLE antibody
(low magnification x40). (B) DNA staining of the same cells with DAPI. (C)
Endogenous PRH staining of the same cells using a TRITC labelled anti-PRH mouse
polyclonal antibody. (D) Co-immunofluoresence of endogenous PRH and TLE
proteins using TRITC labelled anti-PRH mouse polyclonal antibodies and FITC
labelled pan-TLE rat monoclonal antibodies. (E) High magnification (x100)
32
by guest on February 3, 2019http://w
ww
.jbc.org/D
ownloaded from

brightfield image of a single K562 cell. (F) Endogenous PRH in the nucleus and
cytoplasm of the same cell (FITC staining). (G) Endogenous TLE in the nucleus of
the same cell (TRITC staining). (H) Co-immunofluoresence of endogenous PRH and
TLE proteins in the same cell. (I) High magnification (x100) brightfield view of
transfected K562 cells. (J) Transfected MycPRH (TRITC staining). (K) Transfected
TLE1 (FITC staining). (L) Co-localisation of TLE1 with MycPRH in the transfected
cell.
Figure 3 TLE proteins binds to PRH in vitro and in vivo.
(A) In vitro transcribed and translated TLE1 (1) was incubated with glutathione beads
coated with GST (2), GST-Human PRHN (3), or GST-Avian PRHN (4). Bound
proteins were eluted using glutathione, separated by SDS-PAGE and visualised using
fluorography and a PhosphorImager. (B) Lane 1 shows a Western analysis of
endogenous TLE proteins in K562 cell nuclear extract. Lanes 2 and 3 show a Western
blot of endogenous TLE binding to glutathione beads coated with GST alone or GST-
Human PRHN, respectively. The sizes of marker proteins are indicated. (C) Lane 1
shows a Western of Flag-tagged TLE1 in K562 cell nuclear extract made after
transfection with pCMV2-FLAGTLE1 and pMug1-MycPRH. Lanes 2 and 3 show
TLE1 in the same extract after immunoprecipitation with Protein G beads (2) or
Protein G beads and Myc9E10 antibody (3). The blot was probed with an anti-Flag
antibody. Lane 4 is exactly as lane 3 except that all incubations and washes were
performed in the presence of 0.4 mg/ml Ethidium bromide. (D) Lane 1 shows a
Western of endogenous TLE proteins in K562 cell nuclear extract made after
transfection with pMug1-MycPRH. Lane 2 shows endogenous TLE proteins in
nuclear extract from untransfected cells after immunoprecipitation with Protein G
33
by guest on February 3, 2019http://w
ww
.jbc.org/D
ownloaded from

beads and Myc9E10 antibody. Lanes 3 and 4 show endogenous TLE proteins in the
nuclear extract from transfected cells after immunoprecipitation with Protein G beads
or Protein G beads and Myc9E10 antibody.
Figure 4 PRH and TLE proteins co-repress transcription.
(A) K562 cells were transiently transfected with 5µg of a luciferase reporter plasmid
(pTK-PRH) containing 5 PRH binding sites upstream of the minimal TK promoter (1-
4). pMug1-MycPRH (100ng) was co-transfected into the same cells either alone or
together with pCDNA3-TLE1 (200ng), (2) and (3), respectively. Luciferase activity
was normalised for transfection efficiency using a co-transfected plasmid expressing
β-galactosidase. The data is presented as promoter activity relative to the reporter
alone and the values represent the mean and standard error of at least three
experiments. (B) K562 cells were transiently transfected with 5µg pTK-PRH (1-8).
100ng of a PRH expression vector (pMug1-MycPRH) was co-transfected into the
same cells (2-5) with 0.5µg (3), 1µg (4) or 5µg (5) of the Grg5 expression vector
pFlag-Grg5. As a control the same amounts of pFlag-Grg5 were transfected into K562
cells in the absence of PRH (6-8). Luciferase activity is normalised and presented
exactly as in (A).
Figure 5 The PRH Eh-1 motif mediates interaction and co-repression with TLE
proteins
(A) A schematic representation of the PRH proteins showing the putative Eh-1 motif
located between amino acids 30 and 39. (B) In vitro transcribed and translated TLE1
(1) was incubated with glutathione beads coated with GST (2), GST-PRHN 1-141 (3),
GST-PRHN 61-141 (4), or GST-PRHN 1-125 (5). Bound proteins were eluted using
34
by guest on February 3, 2019http://w
ww
.jbc.org/D
ownloaded from

glutathione, separated by SDS-PAGE and visualised using fluorography and a
PhosphorImager. (C) The top panel shows a pull-down experiment using GST-PRH
proteins and the endogenous TLE proteins from K562 cell nuclear extract after
Western blotting with the pan-TLE antibody. The bottom panel shows the same
membrane after stripping and reprobing with an HC8 antibody. (D) In vitro
transcribed and translated TLE1 (1) was incubated with glutathione beads coated with
GST (2), GST-PRHN 1-141 (3), or GST-PRHN 1-141 F32E (4). Bound proteins were
eluted and visualised as above. (E) The top panel shows a pull-down experiment using
GST-PRHN 1-141 and GST-PRHN 1-141 F32E and the endogenous TLE proteins from
K562 cell nuclear extract after Western blotting with a TLE antibody. The bottom
panel shows that same membrane after stripping and reprobing with an HC8 antibody.
(F) K562 cells were transiently transfected with 5µg of pTK-PRH (1-4). pMug1-
MycPRH F32E (100ng) was co-transfected into the same cells either alone or together
with pCDNA3-TLE1 (200ng), (2) and (3), respectively. Luciferase activity is
normalised and presented exactly as in Figure 4.
35
by guest on February 3, 2019http://w
ww
.jbc.org/D
ownloaded from

REFERENCESReferences
1. Crompton, M. R., Bartlett, T. J., MacGregor, A. D., Manfioletti, G., Buratti, E., Giancotti, V., and Goodwin, G. H. (1992) Nucleic.Acids.Res. 20, 5661-5667
2. Bedford, F. K., Ashworth, A., Enver, T., and Wiedemann, L. M. (1993) Nucleic.Acids.Res. 21, 1245-1249
3. Manfioletti, G., Gattei, V., Buratti, E., Rustighi, A., De Iuliis, A., Aldinucci, D., Goodwin, G. H., and Pinto, A. (1995) Blood 85, 1237-1245
4. Thomas, P. Q., Brown, A., and Beddington, R. S. (1998) Development 125, 85-94
5. Bogue, C. W., Ganea, G. R., Sturm, E., Ianucci, R., and Jacobs, H. C. (2000) Dev.Dyn. 219, 84-89
6. Jayaraman, P., Frampton, J., and Goodwin, G. (2000) Leuk.Res. 24, 1023-1031
7. Topcu, Z., Mack, D. L., Hromas, R. A., and Borden, K. L. (1999) Oncogene 18, 7091-7100
8. Guo, Y., Chan, R., Ramsey, H., Li, W., Xie, X., Shelley, W. C., Martinez-Barbera, J. P., Bort, B., Zaret, K., Yoder, M., and Hromas, R. (2003) Blood
9. Nakagawa, T., Abe, M., Yamazaki, T., Miyashita, H., Niwa, H., Kokubun, S., and Sato, Y. (2003) Arterioscler.Thromb.Vasc.Biol. 23, 231-237
10. Obinata, A., Akimoto, Y., Omoto, Y., and Hirano, H. (2002) Dev.Growth Differ. 44, 281-292
11. George, A., Morse, H. C., III, and Justice, M. J. (2003) Oncogene 22, 6764-6773
12. Hansen, G. M. and Justice, M. J. (1999) Oncogene 18, 6531-6539
13. Topisirovic, I., Culjkovic, B., Cohen, N., Perez, J. M., Skrabanek, L., and Borden, K. L. (2003) EMBO J. 22, 689-703
14. Topisirovic, I., Guzman, M. L., McConnell, M. J., Licht, J. D., Culjkovic, B., Neering, S. J., Jordan, C. T., and Borden, K. L. (2003) Mol.Cell Biol. 23, 8992-9002
15. Brickman, J. M., Jones, C. M., Clements, M., Smith, J. C., and Beddington, R. S. (2000) Development 127, 2303-2315
16. Martinez Barbera, J. P., Clements, M., Thomas, P., Rodriguez, T., Meloy, D., Kioussis, D., and Beddington, R. S. (2000) Development 127, 2433-2445
36
by guest on February 3, 2019http://w
ww
.jbc.org/D
ownloaded from

17. Tanaka, T., Inazu, T., Yamada, K., Myint, Z., Keng, V. W., Inoue, Y., Taniguchi, N., and Noguchi, T. (1999) Biochem.J. 339, 111-117
18. Pellizzari, L., D'Elia, A., Rustighi, A., Manfioletti, G., Tell, G., and Damante, G. (2000) Nucleic Acids Res. 28, 2503-2511
19. Guiral, M., Bess, K., Goodwin, G., and Jayaraman, P. S. (2001) J.Biol.Chem. 276, 2961-2970
20. Puppin, C., D'Elia, A. V., Pellizzari, L., Russo, D., Arturi, F., Presta, I., Filetti, S., Bogue, C. W., Denson, L. A., and Damante, G. (2003) Nucleic Acids Res. 31, 1845-1852
21. Guiral, M., Bess, K., Goodwin, G., and Jayaraman, P. S. (2001) J.Biol.Chem. 276, 2961-2970
22. Bess, K. L., Swingler, T. E., Rivett, J., Gaston, K., and Jayaraman, P. S. (2003) Biochem.J. 374, 667-675
23. Courey, A. J. and Jia, S. (2001) Genes Dev. 15, 2786-2796
24. Dasen, J. S., Barbera, J. P., Herman, T. S., Connell, S. O., Olson, L., Ju, B., Tollkuhn, J., Baek, S. H., Rose, D. W., and Rosenfeld, M. G. (2001) Genes Dev. 15, 3193-3207
25. Dehni, G., Liu, Y., Husain, J., and Stifani, S. (1995) Mech.Dev. 53, 369-381
26. Eastman, Q. and Grosschedl, R. (1999) Curr.Opin.Cell Biol. 11, 233-240
27. Levanon, D., Goldstein, R. E., Bernstein, Y., Tang, H., Goldenberg, D., Stifani, S., Paroush, Z., and Groner, Y. (1998) Proc.Natl.Acad.Sci.U.S.A 95, 11590-11595
28. Chen, G. and Courey, A. J. (2000) Gene 249, 1-16
29. Pickles, L. M., Roe, S. M., Hemingway, E. J., Stifani, S., and Pearl, L. H. (2002) Structure.(Camb.) 10, 751-761
30. Chen, G., Nguyen, P. H., and Courey, A. J. (1998) Mol.Cell Biol. 18, 7259-7268
31. Grbavec, D., Lo, R., Liu, Y., and Stifani, S. (1998) Eur.J.Biochem. 258, 339-349
32. Ren, B., Chee, K. J., Kim, T. H., and Maniatis, T. (1999) Genes Dev. 13, 125-137
33. Roose, J., Molenaar, M., Peterson, J., Hurenkamp, J., Brantjes, H., Moerer, P., van de, W. M., Destree, O., and Clevers, H. (1998) Nature 395, 608-612
34. Wang, J. C., Waltner-Law, M., Yamada, K., Osawa, H., Stifani, S., and Granner, D. K. (2000) J.Biol.Chem. 275, 18418-18423
37
by guest on February 3, 2019http://w
ww
.jbc.org/D
ownloaded from

35. Chen, G., Fernandez, J., Mische, S., and Courey, A. J. (1999) Genes Dev. 13, 2218-2230
36. Boddy, M. N., Freemont, P. S., and Borden, K. L. (1994) Trends Biochem.Sci. 19, 198-199
37. Choi, C. Y., Kim, Y. H., Kwon, H. J., and Kim, Y. (1999) J.Biol.Chem. 274, 33194-33197
38. Palaparti, A., Baratz, A., and Stifani, S. (1997) J.Biol.Chem. 272, 26604-26610
39. Yu, X., Li, P., Roeder, R. G., and Wang, Z. (2001) Mol.Cell Biol. 21, 4614-4625
40. Zhang, H. and Emmons, S. W. (2002) Genetics 160, 799-803
41. Tetsuka, T., Uranishi, H., Imai, H., Ono, T., Sonta, S., Takahashi, N., Asamitsu, K., and Okamoto, T. (2000) J.Biol.Chem. 275, 4383-4390
42. Gossen, M. and Bujard, H. (1992) Proc.Natl.Acad.Sci.U.S.A 89, 5547-5551
43. Grbavec, D., Lo, R., Liu, Y., Greenfield, A., and Stifani, S. (1999) Biochem.J. 337, 13-17
44. Fields, S. and Song, O. (1989) Nature 340, 245-246
45. Stifani, S., Blaumueller, C. M., Redhead, N. J., Hill, R. E., and Artavanis-Tsakonas, S. (1992) Nat.Genet. 2, 343
46. Dignam, J. D., Lebovitz, R. M., and Roeder, R. G. (1983) Nucleic.Acids.Res. 11, 1475-1489
47. Dorn, A., Benoist, C., and Mathis, D. (1989) Mol.Cell Biol. 9, 312-320
48. Tolkunova, E. N., Fujioka, M., Kobayashi, M., Deka, D., and Jaynes, J. B. (1998) Mol.Cell Biol. 18, 2804-2814
49. Jimenez, G., Verrijzer, C. P., and Ish-Horowicz, D. (1999) Mol.Cell Biol. 19, 2080-2087
50. Aronson, B. D., Fisher, A. L., Blechman, K., Caudy, M., and Gergen, J. P. (1997) Mol.Cell Biol. 17, 5581-5587
51. Paroush, Z., Finley, R. L., Jr., Kidd, T., Wainwright, S. M., Ingham, P. W., Brent, R., and Ish-Horowicz, D. (1994) Cell 79, 805-815
52. Gehring, W. J., Affolter, M., and Burglin, T. (1994) Annu.Rev.Biochem. 63:487-526, 487-526
53. Vidal, M., Braun, P., Chen, E., Boeke, J. D., and Harlow, E. (1996) Proc.Natl.Acad.Sci.U.S.A 93, 10321-10326
38
by guest on February 3, 2019http://w
ww
.jbc.org/D
ownloaded from

54. Jayaraman, P. S., Frampton, J., and Goodwin, G. (2000) Leuk.Res. 24, 1023-1031
55. Koeffler, H. P. and Golde, D. W. (1980) Blood 56, 344-350
56. Husain, J., Lo, R., Grbavec, D., and Stifani, S. (1996) Biochem.J. 317 ( Pt 2), 523-531
57. Lai, J. S. and Herr, W. (1992) Proc.Natl.Acad.Sci.U.S.A 89, 6958-6962
58. Smith, S. T. and Jaynes, J. B. (1996) Development 122, 3141-3150
59. Jimenez, G., Paroush, Z., and Ish-Horowicz, D. (1997) Genes & Development 11, 3072-3082
60. McLarren, K. W., Lo, R., Grbavec, D., Thirunavukkarasu, K., Karsenty, G., and Stifani, S. (2000) J.Biol.Chem. 275, 530-538
61. Yao, J., Lai, E., and Stifani, S. (2001) Mol.Cell Biol. 21, 1962-1972
62. Du, Z., Cong, H., and Yao, Z. (2001) Biochem.Biophys.Res.Commun. 282, 701-706
63. Eberhard, D., Jimenez, G., Heavey, B., and Busslinger, M. (2000) EMBO J. 19, 2292-2303
64. Imai, Y., Kurokawa, M., Tanaka, K., Friedman, A. D., Ogawa, S., Mitani, K., Yazaki, Y., and Hirai, H. (1998) Biochem.Biophys.Res.Commun. 252, 582-589
65. Yao, J., Liu, Y., Lo, R., Tretjakoff, I., Peterson, A., and Stifani, S. (2000) Mech.Dev. 93, 105-115
66. Muhr, J., Andersson, E., Persson, M., Jessell, T. M., and Ericson, J. (2001) Cell 104, 861-873
67. Newman, C. S., Chia, F., and Krieg, P. A. (1997) Mech.Dev. 66, 83-93
68. Artavanis-Tsakonas, S., Rand, M. D., and Lake, R. J. (1999) Science 284, 770-776
69. Liu, Y., Dehni, G., Purcell, K. J., Sokolow, J., Carcangiu, M. L., Artavanis-Tsakonas, S., and Stifani, S. (1996) Genomics 31, 58-64
70. Ohishi, K., Katayama, N., Shiku, H., Varnum-Finney, B., and Bernstein, I. D. (2003) Semin.Cell Dev.Biol. 14, 143-150
39
by guest on February 3, 2019http://w
ww
.jbc.org/D
ownloaded from

Figure 1
770
A
B
C
PRH N +TLE1 Q-SP
10 12
3
456
7
8
9
PRH N +TLE1 Q
PRH N +TLE1 WD
PRH N +TLE1
PRH N +pGAD424
V +pGAD424
V +TLE1
V + TLE1 WD
V + TLE1 Q
V +TLE1 Q-SP
1-LT
X-Gal-LT +0.2% 5-FOA
-HLT +50mM 3-AT
TLE1-AD
DBD-PRH N
REPORTERGAL UAS
lacZ / His
DBD-TLE1
PRH-AD
0
0.2
0.4
0.6
0.8
1.0
β -ga
lact
osid
ase
activ
ity( a
rbita
ry u
nits
)
1 2 3
REPORTERGAL UAS
lacZ
PRHRepression
1 137 197 270HD
1 137 197 270HD
TLE1
1 383128SP
- + + + - +
WD-repeatQ
DBD-TLE1PRH-AD
by guest on February 3, 2019http://w
ww
.jbc.org/D
ownloaded from

Figure 2
2
3
1
42
3
1
4
2
3
1
4
2
3
1
4
2
3
1
4
A
C D
B
F HGE
J LKI
by guest on February 3, 2019http://w
ww
.jbc.org/D
ownloaded from
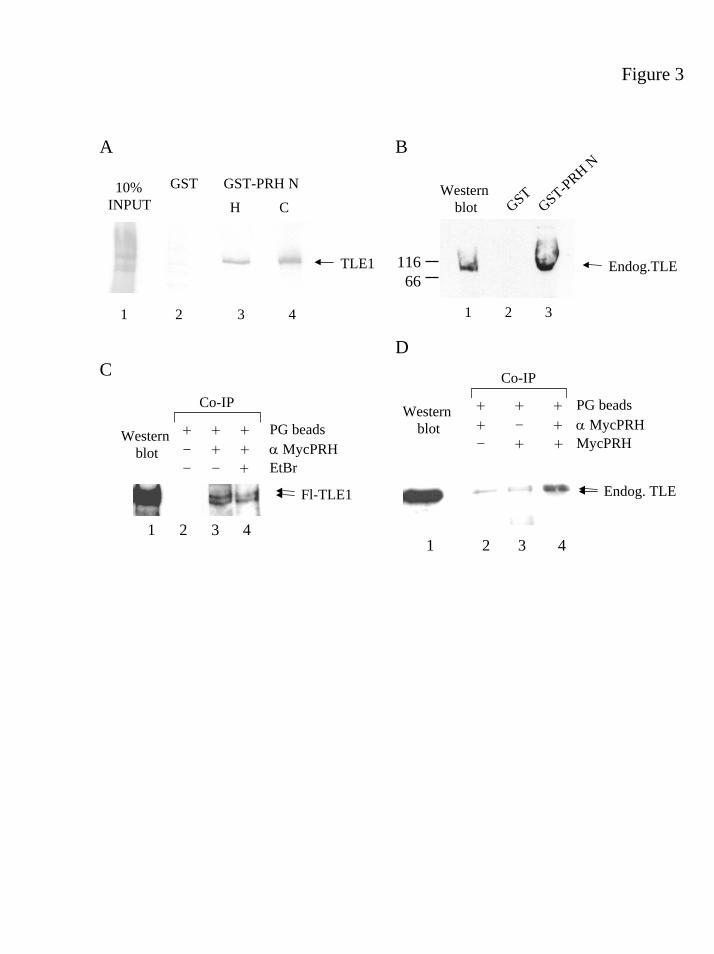
Figure 3
10% INPUT
TLE1
GSTH C
GST-PRH N
1 2 3 4
A
C
B
D
Endog. TLE
+ + + PG beads+ - + α MycPRH- + + MycPRH
Westernblot
Co-IP
1 2 3 4
11666
1 2 3
Endog.TLE
Westernblot GST
GST-PRH N
Fl-TLE1
1 2 3 4
+ + + PG beads- + + α MycPRH- - + EtBr
Westernblot
Co-IP
by guest on February 3, 2019http://w
ww
.jbc.org/D
ownloaded from
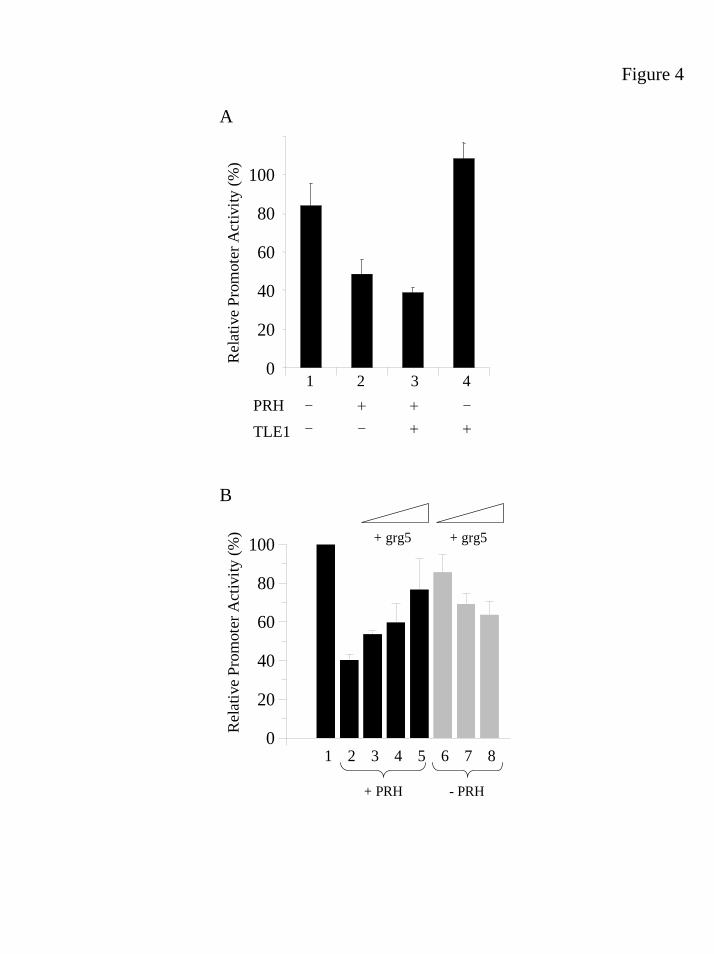
Figure 4
0
20
40
60
80
100
+ PRH
+ grg5 + grg5
Rel
ativ
e Pr
omot
er A
ctiv
ity (%
)
1 2 3 4 5 6 7 8
A
- PRH
A
B
0
20
40
60
80
100
Rel
ativ
e Pr
omot
er A
ctiv
ity (%
)
1 3 42--
PRH
TLE1
++
+-
-+
by guest on February 3, 2019http://w
ww
.jbc.org/D
ownloaded from

Figure 5
A
C
E
TLE130%
INPUT GSTGST-PRH N
GST-PRH N
F32E
1 2 3 4
TLE1
B
D
F
TLE130%
INPUT GST
1 2 3 4 5
GST-PRH 1-141
GST-PRH 61-14
1
GST-PRH 1-125
TLE1
GSTK562 extract
1 2 3 4 5
Endog. TLE
GST-PRH 1-141
GST-PRH 61-14
1
GST-PRH 1-125
1 2 3 4 5
HC8*
GSTGST-PRH N
GST-PRH N
F32E
1 2 3
Endog.TLE
HC8*
1 2 3
TPFYIEDILG putative Eh-1 motif30 39
Repression
1 137 196 270
HD
1 137 270
HD
. .
- + + -- - + +
0
20
40
60
80
100
Relat
ive Pr
omote
r Acti
vity (
%)
PRH F32E TLE1
1 3 42
by guest on February 3, 2019http://w
ww
.jbc.org/D
ownloaded from

JayaramanTracey E. Swingler, Kirstin L. Bess, Jing Yao, Stefano Stifani and Padma-Sheela
groucho/TLE protein family to co-repress transcription in haematopoietic cellsThe proline-rich homeodomain (PRH) protein recruits members of the
published online June 7, 2004J. Biol. Chem.
10.1074/jbc.M404488200Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
by guest on February 3, 2019http://w
ww
.jbc.org/D
ownloaded from