tomato mosaic virus replication protein suppresses...
TRANSCRIPT

JOURNAL OF VIROLOGY, Oct. 2003, p. 11016–11026 Vol. 77, No. 200022-538X/03/$08.00�0 DOI: 10.1128/JVI.77.20.11016–11026.2003Copyright © 2003, American Society for Microbiology. All Rights Reserved.
Tomato Mosaic Virus Replication Protein Suppresses Virus-TargetedPosttranscriptional Gene Silencing
Kenji Kubota,1 Shinya Tsuda,2 Atsushi Tamai,1 and Tetsuo Meshi1,3,4*Department of Botany, Graduate School of Science, Kyoto University, Kyoto 606-8502,1 Department of Plant Pathology, National
Agricultural Research Center, Tsukuba 305-8666,2 Plant Physiology Department, National Institute of Agrobiological Sciences,Tsukuba 305-8602,3 and CREST, Japan Science Technologies, Kawaguchi 332-0012,4 Japan
Received 3 April 2003/Accepted 15 July 2003
Posttranscriptional gene silencing (PTGS), a homology-dependent RNA degradation system, has a role indefending against virus infection in plants, but plant viruses encode a suppressor to combat PTGS. Usingtransgenic tobacco in which the expression of green fluorescent protein (GFP) is posttranscriptionally silenced,we investigated a tomato mosaic virus (ToMV)-encoded PTGS suppressor. Infection with wild-type ToMV (Lstrain) interrupted GFP silencing in tobacco, coincident with visible symptoms, whereas some attenuatedstrains of ToMV (L11 and L11A strains) failed to suppress GFP silencing. Analyses of recombinant virusescontaining the L and L11A strains revealed that a single base change in the replicase gene, which causes anamino acid substitution, is responsible for the symptomless and suppressor-defective phenotypes of theattenuated strains. An agroinfiltration assay indicated that the 130K replication protein acts as a PTGSsuppressor. Small interfering RNAs (siRNAs) of 21 to 25 nucleotides accumulated during ToMV infection,suggesting that the major target of the ToMV-encoded suppressor is downstream from the production ofsiRNAs in the PTGS pathway. Analysis with GFP-tagged recombinant viruses revealed that the suppressorinhibits the establishment of the ToMV-targeted PTGS system in the inoculated leaves but does not detectablysuppress the activity of the preexisting, sequence-specific PTGS machinery there. Taken together, these resultsindicate that it is likely that the ToMV-encoded suppressor, the 130K replication protein, blocks the utilizationof silencing-associated small RNAs, so that a homology-dependent RNA degradation machinery is not newlyformed.
RNA silencing, including posttranscriptional gene silencing(PTGS) in plants, is a homology-dependent RNA degradationsystem occurring in the cytoplasm (41) and characterized bythe presence of 21- to 25-nucleotide (nt) small interferingRNAs (siRNAs) (11). Biochemical and genetic analyses haveshown that the core mechanisms of PTGS are shared amongeukaryotes (30, 42, 46). PTGS is induced by double-strandedRNAs (dsRNAs) or structured single-stranded RNAs(ssRNAs), which are processed into siRNAs by RNase III-likeenzymes, such as Dicer in Drosophila. A sort of siRNA isincorporated into the sequence-specific degradation complex(RNA-induced silencing complex [RISC]) and thereby guidesthe RISC to destroy target RNA that contains a sequencecomplementary to the siRNA in the RISC. In some organisms,including plants, a host RNA-dependent RNA polymerase(RdRp) is also involved in PTGS (5, 24), presumably convert-ing the target ssRNA into dsRNA, which is the substrate usedby Dicer homologs to generate secondary siRNAs. In plants,PTGS generates mobile signals with sequence-specific infor-mation that spread through plasmodesmata and phloem todistant organs, establishing systemic silencing (23).
Plant viruses are strong inducers and targets of PTGS; there-fore, defending plants against viruses is thought to be a role ofPTGS (18, 45). Consequently, many plant viruses have evolved
to encode a suppressor of PTGS to combat this resistancemechanism (41, 44). The suppressor proteins identified so farare structurally and functionally diverse. Potyvirus helper com-ponent proteinase (HC-Pro), one of the first identified sup-pressors (1, 4, 13), probably inhibits the maintenance step ofPTGS at or upstream from the production of siRNA (16, 17)and/or downstream from the production of siRNA (14, 32).Cucumovirus 2b, which was also one of the first suppressorsidentified (4), suppresses systemic silencing, possibly after thegeneration of the silencing signal (8). Potato virus X (PVX)p25 prevents systemic silencing (43), presumably through theinhibition of the class of long (�25-nt) siRNAs (10). Recently,tombusvirus p19 was shown to bind siRNAs in vitro, and so itmay prevent the spread of mobile silencing signals (35). Thecoat protein (CP) of turnip crinkle virus probably functions atan earlier step of PTGS, upstream from siRNA production(32).
Tomato mosaic virus (ToMV) is a positive-sense ssRNAvirus belonging to the Tobamovirus genus. The ToMV genomeencodes at least four proteins: two replicase components(130K and 180K proteins), a movement protein (MP), and CP(22). The 130K and 180K proteins are translated from thegenomic RNA, and MP and CP are translated from the re-spective subgenomic mRNAs. The 180K protein is synthesizedby read-through of the leaky termination codon of the 130Kprotein gene. Like Tobacco mosaic virus (TMV, the type mem-ber of the Tobamovirus genus), ToMV infects tobacco plantssystemically, causing mosaic symptoms, which are character-ized by intermingled light and dark green regions (20).
* Corresponding author. Mailing address: Plant Physiology Depart-ment, National Institute of Agrobiological Sciences, Kannondai 2-1-2,Tsukuba 305-8602, Japan. Phone: 81-29-838-8361. Fax: 81-29-838-8347. E-mail: [email protected].
11016
on June 21, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

Previously, Voinnet et al. (44) showed that TMV infectionreverses green fluorescent protein (GFP) silencing systemicallyin leaves of Nicotiana benthamiana and suggested, based on thesuppression pattern, that TMV may counteract PTGS by pre-venting the spread of systemic signals as well as by a furthermechanism. However, wild-type TMV and ToMV cause severesystemic necrosis in N. benthamiana approximately 1 weekafter infection, a fact which imposes a limitation on the inves-tigation of these tobamovirus suppressor functions in the nor-mal infection process. In this work, we investigated the activityof the ToMV-encoded suppressor by using transgenic tobaccoin which GFP is expressed constitutively or in which the ex-pression of GFP is silenced posttranscriptionally. We foundthat infection with ToMV interrupted GFP silencing, in par-allel with the visible symptoms, through the action of a sup-pressor. An agroinfiltration assay indicated that the 130K rep-lication protein functions as a suppressor of PTGS. We alsofound that the suppressor probably exerts its effects at a spe-cific step after siRNA production to prevent the de novo for-mation of the RISC in ToMV-infected leaves.
MATERIALS AND METHODS
Transgenic plants. N. tabacum cv. Samsun and N. benthamiana were trans-formed with Agrobacterium tumefaciens LBA4404 harboring plasmid pBK-erG3(see below) by the leaf disk method essentially as described previously (7). Thetobacco and N. benthamiana lines constitutively expressing an endoplasmic re-ticulum (ER)-localized, highly fluorescent GFP variant (erG3GFP) (38) wereG3Sm1 and G3Nb3, respectively, which carry the transgene homozygously at asingle locus. For the GFP-silenced tobacco line (G3Sm2), T2 plants and a few T3lines showing a strong phenotype were used; these had various combinations ofmultiple copies of the transgene that differed in transgene number and in thelocus of integration. Because the same results were obtained with different T2and T3 plants, a T3 line (G3Sm2-1) carrying three copies of the transgenehomozygously and showing a strong phenotype was used in the later stages of theexperiments.
Inoculation with viruses and infectious transcripts. The ToMV strains usedwere wild-type strain L (27) and L-derived attenuated strains L11 and L11A (25).Infectious ToMV and PVX transcripts were derived from MluI-cut pTL-seriesplasmids and SpeI-cut pTX-series plasmids (see below), respectively, with T7RNA polymerase (Takara Bio). As the standard method, ToMV and its deriv-atives (including in vitro transcripts) were suspended in 10 mM sodium phos-phate (pH 7.0) and inoculated onto the single fifth true leaf of Samsun plantswhen it was 7.5 to 9.0 cm long. Unless otherwise specified, the inocula containedinfectious viruses at concentrations that caused approximately 100 to 150 localnecrotic lesions on Xanthi nc tobacco leaves. Inoculated plants were grown at 26or 28°C (16 h of light). Recombinant PVX was first amplified in N. benthamiana,and homogenates of the inoculated leaves were used to inoculate tobacco leaves.
Construction of plasmids. pTLW3 carries the full-length ToMV cDNA plus alinker-derived 25-bp sequence (between the MluI and the EcoRI sites) ofpLFW3 (21) downstream of a T7 promoter between the PstI and EcoRI sites ofpUC18. The 2.93-kb BstXI-AccIII fragment (nt 832 to 3759) of pLFA1 (21) wasreplaced with the corresponding fragment of pTLW3 to create pTLJA1. The2.47-kb NheI-KpnI fragment (nt 1926 to 4394) was exchanged between pTLW3and pTLJA1 to yield pTLJ and pTLA. The 1.41-kb KpnI-BstEII fragment (nt4395 to 5799) of pTLW3 was replaced with the corresponding fragment (fromthe KpnI site to the end of the G3GFP open reading frame [ORF]) of piL.G3(38) to yield pTL.G3. The 0.59-kb fragment (from the filled-in BstEII site to theMluI site) of pTL.G3 was replaced with an NcoI (filled-in)-MluI fragment ofpTLW3 to create pTLBN.G3. The KpnI-MluI fragment of pTLBN.G3 containingthe G3GFP ORF was introduced into pTLJ to yield pTLBN.G3(J). pTX.erG3was constructed by replacing the ApaI-XhoI fragment of pP2C2S (3) with thecorresponding fragment of piX.erG3 (37). pTX.GUS was constructed by insert-ing the SmaI-SacI fragment of pBI121 (Clontech), encoding �-glucuronidase(GUS), into the SalI site of pP2C2S after blunting. Note that the erG3GFP genein pTX.erG3 and pBK-erG3 (see below) encodes the signal peptide at the Nterminus and the ER retention signal at the C terminus, whereas the G3GFPgene in pTLBN.G3 and pTLBN.G3(J) encodes cytosolic GFP.
The XbaI-SacI fragment of pBI-erG3, which encodes erG3GFP (38), was
inserted between the corresponding sites of pBI121 (Clontech) to create pBK-erG3. The coding region for HC-Pro of potato virus Y (PVY; Japanese O strain)was amplified by reverse transcription (RT)-PCR with primers 5�-GCTCTAGAAACATGGCCTCAAATGCTGAGAATTTTTGGAAGGGTCTG-3� and 5�-CGGGATCCTTAACCAACTCTATAGTGTTTTATATCAGACTC-3�, cut withXbaI and BamHI (shown in italic type in the primers), and cloned into pMD1 (apBI121-derived vector with a modified cloning site; supplied by B. Baker) tocreate pMD-YHCP. The Met and Ala codons and a termination codon wereincluded in the primers. pBI-L130NRT contains the 5� noncoding region and the130K protein ORF of pTLW3. An additional termination codon was positionedjust after the leaky termination codon of the 130K protein ORF. pBI-L130FS wascreated by filling in the XbaI site located in the 130K protein ORF of pBI-L130NRT (nt 1002 of the ToMV genome). pBI-L130J contains the pTLJ-derived130K protein ORF.
GFP observation (imaging) and GUS staining. GFP signals were observedunder an epifluorescence stereomicroscope (MZ-FLIII; Leica) equipped with afilter set (GFP Plant; Leica) and a charge-coupled device camera (DXM1200;Nikon). Plants were excited with a halogen lamp (Intralux 6000-1; Volpi AG)through a no. 24902 filter (440 to 480 nm) and then photographed with a digitalcamera (Camedia E-20; Olympus) equipped with a cut filter (520 nm, SC52; FujiFilm). This procedure resulted in a low level of red autofluorescence. Alterna-tively, plants were irradiated with the mercury lamp of the epifluorescencestereomicroscope through the GFP filter (450 to 490 nm) and then photographedwith a charge-coupled device camera (DC500; Leica) equipped with a zoom lens(70 to 180 mm, Zoom-Micro Nikkor; Nikon) and a band-pass filter (500 to 550nm; Omega Optical, Inc.) to reduce red autofluorescence. GUS staining wasperformed as described previously (19).
RNA analysis. Leaves, excluding major veins, were homogenized in liquidnitrogen. Total RNAs were extracted with TRIzol reagent (Invitrogen) accordingto the manufacturer’s instructions. For Northern blot analysis of GFP mRNA,total RNAs were glyoxylated, separated on a 1% agarose gel, and then trans-ferred to a Hybond-N membrane (Amersham Biosciences). Prehybridization andhybridization were performed as described previously (11) with a DNA probederived from the erG3GFP ORF (the EcoRI-ClaI fragment).
For small-RNA analysis, high-molecular-weight RNAs were removed fromtotal RNAs (isolated from 0.5 to 2 g of leaf tissues) by centrifugation in thepresence of 5% polyethylene glycol and 0.5 M NaCl (11). Low-molecular-weightRNAs were recovered by ethanol precipitation, suspended in formamide, andseparated on a 15% polyacrylamide–7 M urea gel. The amounts of low-molec-ular-weight RNAs loaded were adjusted on the basis of the ethidium bromide-stained bands, which contained the major species of tRNAs, 5S rRNAs, and 5.8SrRNA, migrating near the �400-bp DNA marker on a 1% agarose gel (17).
After blotting onto Hybond-NX (Amersham Biosciences), hybridization wasperformed at 40°C overnight with riboprobes as described previously (17). Ri-boprobes corresponded to the erG3GFP ORF or the 3�-terminal 0.8-kb region ofthe ToMV genome and were prepared with T7 or T3 RNA polymerase from theappropriate subclones, followed by partial alkaline hydrolysis into 50- to 100-nt(on average) fragments. Size markers were prepared by filling in the SpeI-cut endof pBluescript in the presence of [�-32P]dCTP and cutting with SacII (20-ntoligonucleotide) or EcoRI (25-nt oligonucleotide). Synthetic RNAs with thesame sequences as the above 20- and 25-nt oligonucleotides were also used insome experiments. The oligonucleotide DNA markers migrated �1 base fasterthan the corresponding RNA markers. Radioactive bands were detected with aTyphoon 8600 imager (Amersham Biosciences).
Specific amplification of transgene-derived GFP mRNA and GFP-tagged viralgenomes by RT-PCR was performed as follows. Central portions (2 mm indiameter) of GFP lesions induced by TLBN.G3 or TLBN.G3(J) were punchedout under an epifluorescence stereomicroscope and subjected to total RNAextraction as described above. Disks of the same diameter were punched fromareas outside the GFP lesions on the same inoculated leaves or from mock-inoculated leaves.
cDNAs were synthesized with Ready-to-Go RT-PCR beads (Amersham Bio-sciences) and with oligo(dT)12–16 or a 20-nt oligonucleotide complementary tothe 3� end of ToMV RNA as a primer. PCR primers used to amplify theerG3GFP mRNA sequence were 5�-ATGAAGACTAATCTTTTTCTCTTTCTCATC-3�, corresponding to the N terminus of erG3GFP (part of the signalpeptide), and 5�-TTAAAGCTCATGTTTGTATAGTTCATC-3�, correspondingto the C terminus (including the ER retention signal). Primers used to amplifyactin mRNA were 5�-ATGGCAGACGGTGAGGATATTCA-3� and 5�-GCCTTTGCAATCCACATCTGTTG-3�. Primers used to specifically detect TLBN.G3and TLBN.G3(J) were an oligonucleotide corresponding to nt 5271 to 5300 ofToMV and 5�-TGTGGGAGTTGAAGTTGTATTCCAA-3�, which correspondsto an internal portion of the G3GFP ORF. Amplification was carried out with
VOL. 77, 2003 ToMV REPLICATION PROTEIN SUPPRESSES PTGS 11017
on June 21, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from
ExTaq DNA polymerase (Takara Bio) for 22 cycles (for viral RNA or GFPmRNA) or 35 cycles (for actin mRNA) of 94°C for 30 s, 55°C for 30 s, and 72°Cfor 1 min. Each PCR (50 �l) contained cDNAs derived from total RNAsequivalent to 0.1 GFP lesion.
Protein analysis. Leaf tissues were homogenized in 100 mM Tris-HCl (pH6.8)–4% sodium dodecyl sulfate (SDS)–12% 2-mercaptoethanol. To detect the130K protein, protease inhibitor CompleteMini (Roche-Boehringer) was in-cluded. After debris was removed, supernatants were boiled and mixed with anequal volume of 20% glycerol containing 0.02% bromophenol blue. To detectCP, proteins were separated by SDS– 15% polyacrylamide gel electrophoresis(PAGE) and stained with Coomassie brilliant blue. To immunodetect GFP orthe ToMV 130K protein, proteins were separated by SDS–12.2% PAGE orSDS–7.5% PAGE, respectively, and transferred to a polyvinylidene difluoridemembrane (Millipore). The membrane was treated with anti-GFP antibodies(Clontech) or anti-130K protein antibodies (9) as primary antibodies and horse-radish peroxidase-conjugated goat anti-rabbit immunoglobulin G antibody (Am-ersham Biosciences) as the secondary antibody, and immunoreactive bands werevisualized by enhanced chemiluminescence (Amersham Biosciences).
Agroinfiltration. A. tumefaciens C58C1/pGV2260 harboring the appropriateplasmid was cultured, set to an optical density at 600 nm of 1.0, and used toinfiltrate small leaves of �3-week-old N. benthamiana plants essentially as de-scribed previously (6). The infiltration mixture contained two parts Agrobacte-rium carrying an empty vector, pOCA18 (28), or a plasmid encoding a suppressorand one part Agrobacterium carrying pBK-erG3, which induces local silencing.Infiltrated plants were kept at 22°C for 1 day to ensure transformation and thenkept at the same or different temperatures for testing of the temperature de-pendency of silencing induction (36).
RESULTS
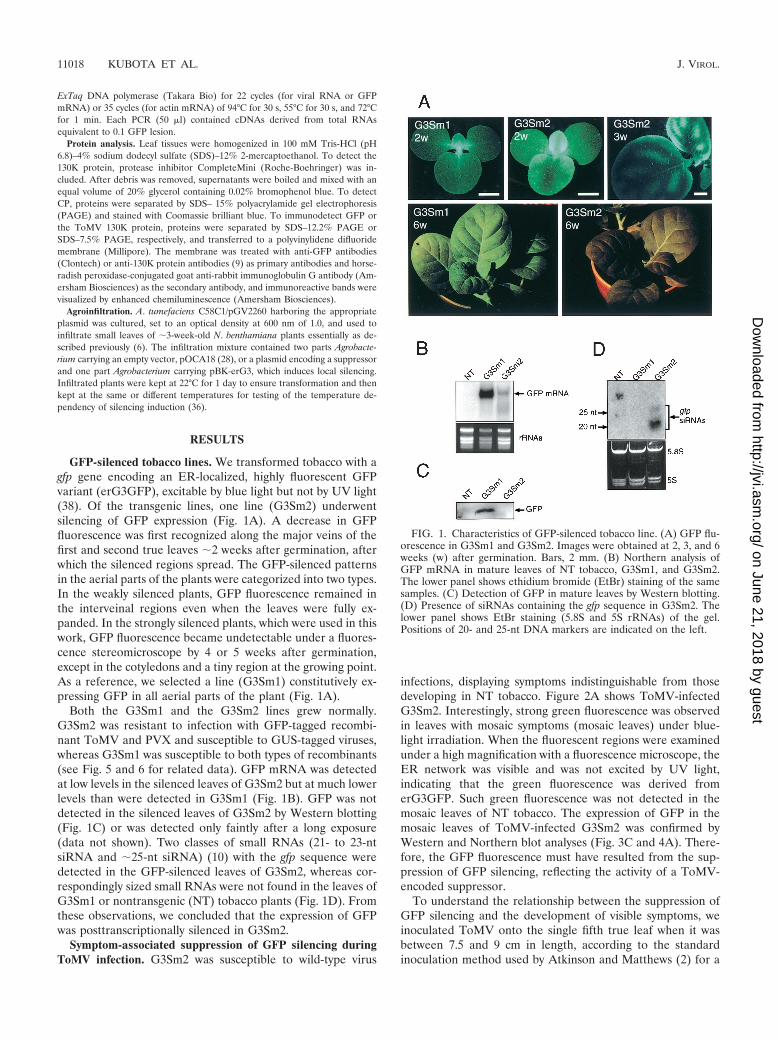
GFP-silenced tobacco lines. We transformed tobacco with agfp gene encoding an ER-localized, highly fluorescent GFPvariant (erG3GFP), excitable by blue light but not by UV light(38). Of the transgenic lines, one line (G3Sm2) underwentsilencing of GFP expression (Fig. 1A). A decrease in GFPfluorescence was first recognized along the major veins of thefirst and second true leaves �2 weeks after germination, afterwhich the silenced regions spread. The GFP-silenced patternsin the aerial parts of the plants were categorized into two types.In the weakly silenced plants, GFP fluorescence remained inthe interveinal regions even when the leaves were fully ex-panded. In the strongly silenced plants, which were used in thiswork, GFP fluorescence became undetectable under a fluores-cence stereomicroscope by 4 or 5 weeks after germination,except in the cotyledons and a tiny region at the growing point.As a reference, we selected a line (G3Sm1) constitutively ex-pressing GFP in all aerial parts of the plant (Fig. 1A).
Both the G3Sm1 and the G3Sm2 lines grew normally.G3Sm2 was resistant to infection with GFP-tagged recombi-nant ToMV and PVX and susceptible to GUS-tagged viruses,whereas G3Sm1 was susceptible to both types of recombinants(see Fig. 5 and 6 for related data). GFP mRNA was detectedat low levels in the silenced leaves of G3Sm2 but at much lowerlevels than were detected in G3Sm1 (Fig. 1B). GFP was notdetected in the silenced leaves of G3Sm2 by Western blotting(Fig. 1C) or was detected only faintly after a long exposure(data not shown). Two classes of small RNAs (21- to 23-ntsiRNA and �25-nt siRNA) (10) with the gfp sequence weredetected in the GFP-silenced leaves of G3Sm2, whereas cor-respondingly sized small RNAs were not found in the leaves ofG3Sm1 or nontransgenic (NT) tobacco plants (Fig. 1D). Fromthese observations, we concluded that the expression of GFPwas posttranscriptionally silenced in G3Sm2.
Symptom-associated suppression of GFP silencing duringToMV infection. G3Sm2 was susceptible to wild-type virus
infections, displaying symptoms indistinguishable from thosedeveloping in NT tobacco. Figure 2A shows ToMV-infectedG3Sm2. Interestingly, strong green fluorescence was observedin leaves with mosaic symptoms (mosaic leaves) under blue-light irradiation. When the fluorescent regions were examinedunder a high magnification with a fluorescence microscope, theER network was visible and was not excited by UV light,indicating that the green fluorescence was derived fromerG3GFP. Such green fluorescence was not detected in themosaic leaves of NT tobacco. The expression of GFP in themosaic leaves of ToMV-infected G3Sm2 was confirmed byWestern and Northern blot analyses (Fig. 3C and 4A). There-fore, the GFP fluorescence must have resulted from the sup-pression of GFP silencing, reflecting the activity of a ToMV-encoded suppressor.
To understand the relationship between the suppression ofGFP silencing and the development of visible symptoms, weinoculated ToMV onto the single fifth true leaf when it wasbetween 7.5 and 9 cm in length, according to the standardinoculation method used by Atkinson and Matthews (2) for a
FIG. 1. Characteristics of GFP-silenced tobacco line. (A) GFP flu-orescence in G3Sm1 and G3Sm2. Images were obtained at 2, 3, and 6weeks (w) after germination. Bars, 2 mm. (B) Northern analysis ofGFP mRNA in mature leaves of NT tobacco, G3Sm1, and G3Sm2.The lower panel shows ethidium bromide (EtBr) staining of the samesamples. (C) Detection of GFP in mature leaves by Western blotting.(D) Presence of siRNAs containing the gfp sequence in G3Sm2. Thelower panel shows EtBr staining (5.8S and 5S rRNAs) of the gel.Positions of 20- and 25-nt DNA markers are indicated on the left.
11018 KUBOTA ET AL. J. VIROL.
on June 21, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

detailed description of the development of tobacco mosaicdisease. The leaves were numbered sequentially from the in-oculated leaf (L0) to the upper leaves.
GFP fluorescence was not restored to leaves L0 to L2. TheGFP signal was first observed on L3 and L4 at 3 or 4 dayspostinoculation (dpi), along with class I to IV veins (Fig. 2C).GFP expression had been silenced in these leaves at the timeof inoculation. In class III and IV veins, GFP fluorescence wasassociated with visible vein clearing. Mosaic symptoms devel-oped in L4 and the upper leaves, where GFP fluorescence wasdetected, coinciding well with yellow tissues (Fig. 2A and B).No green fluorescence was detected in the tiny dark greenislands that were surrounded by yellow tissues. However, largedark green tissues often contained irregularly shaped, lightyellow regions that exhibited weak green fluorescence. OnceGFP silencing was broken, GFP fluorescence remained untilthe leaves were apparently senescent.
A replicase gene mutation results in a symptomless pheno-type and defective suppressor activity. That the suppression ofGFP silencing or the activity of the ToMV-encoded suppressorwas associated with mosaic symptoms suggested the possibilitythat some symptomless mutants may be suppressor defective.On examining some attenuated strains, we found that ToMVL11 and L11A represent this type of virus, spreading systemi-cally without causing symptoms or restoring GFP fluorescencein G3Sm2 (Fig. 3A). The L11A genome differs from the wild-type L genome at 10 positions, including three coding changes(21, 25). L11, the parental attenuated strain of L11A, has thefirst mutation at nt 1117 (25), suggesting that this is the caus-ative mutation. However, because the L11 genome has beensequenced only partially, it was possible that another mutationin the L11 genome was responsible for the defect in suppressorfunction. Therefore, we prepared several recombinant virusesto identify with certainty the mutation that causes defectivesuppressor activity.
TLJA1 (in vitro transcript from pTLJA1), which differs fromTLW3 (the wild-type construct equivalent to L) at four posi-tions, including all three coding changes (Fig. 3A), displayed
the same phenotype as L11A in every characteristic examined;e.g., it caused no systemic symptoms on Samsun tobacco,formed normal local lesions on Xanthi nc tobacco with the Ngene and, importantly, failed to suppress GFP silencing inG3Sm2. TLJ contains one of the four substitutions of TLJA1,and TLA contains the other three (Fig. 3A). TLJ caused nomosaic symptoms and did not restore GFP fluorescence inG3Sm2 (Fig. 3B). On the other hand, TLA caused mild mosaicsymptoms, and GFP fluorescence was detected in systemicallyinfected leaves of TLA-infected G3Sm2, coinciding with thevisible symptoms, although its intensity was weaker than thatseen on L- or TLW3-infected mosaic leaves (Fig. 3B).
To confirm that TLJ and TLJA1 caused systemic infectionbut did not restore GFP expression in G3Sm2, leaf extractswere subjected to protein analysis (Fig. 3C). Yellow tissuescontain a high level of virus, whereas tiny dark green islandsare virus free or contain a low level of virus (2). Consistent withthis description, high levels of CP were detected in yellowtissues of TLW3- or TLA-induced mosaic leaves, where GFPwas also expressed at high levels (Fig. 3C, lanes 6 and 9). Indark green islands, CP was often detected at low levels and,correspondingly, GFP levels were slightly increased (Fig. 3C,lanes 5 and 8). In the nonsymptomatic leaves of TLJ- orTLJA1-infected G3Sm2, no increase in GFP levels was ob-served, even though CP accumulated (Fig. 3C, lanes 7 and 10).It is therefore likely that the mutation at nt 1117 impairs theactivity of a factor that functions in the active suppressionprocess.
Although TLJ and TLJA1 caused no systemic symptoms inleaves and did not restore GFP fluorescence in the correspond-ing leaves of G3Sm2, faint GFP fluorescence was detectedaround regions showing very weak vein clearing in L3 and L4,but only at an early stage of infection (data not shown), as wasthe case for L11 and L11A. Interestingly, this GFP expressiondid not persist and became fully silenced within a few days.Such transient restoration of GFP expression was more pro-nounced when these viruses were inoculated onto small leaves,�3 cm or less in length (Fig. 3D). Infection with either sup-pressor-active or suppressor-defective constructs restored GFPfluorescence on these small L0 leaves as well as on a few upperleaves, accompanied by slight chlorotic or vein-clearing symp-toms. A few days later, these GFP signals in TLJ- or TLJA1-inoculated plants weakened (Fig. 3D) and then disappeared.No GFP fluorescence was detected in newly emerging leaves,as was the case after infection with the standard inoculationmethod. In TLW3-infected plants, GFP fluorescence did notdisappear once it was restored. Therefore, the mutation at nt1117 did not completely inactivate ToMV-encoded suppressoractivity.
Production of ToMV-specific small RNAs in infected leaves.To better understand how the ToMV-encoded suppressorfunctions, we analyzed ToMV- and gfp-derived RNAs in upperleaves of mock-, TLW3-, or TLJ-inoculated plants. TotalRNAs for the TLW3 genome were detected at high levels,severalfold higher than for the TLJ genome (Fig. 4A, upperpanel), as expected from a previous report for L11A (26). GFPmRNA was detected at similar levels in mock-, TLW3-, orTLJ-inoculated G3Sm1 plants (Fig. 4A, lower panel, lanes 4 to6). In G3Sm2 leaves, TLW3 infection increased the expressionof GFP mRNA (Fig. 4A, lane 8), reflecting the suppression of
FIG. 2. Symptom-associated suppression of GFP silencing inG3Sm2 infected with ToMV. (A) GFP fluorescence pattern on mosaicleaves at 11 dpi. (B) Close-up view at 12 dpi of a mosaic leaf, in whichGFP fluorescence is visible in yellow tissues. (C) Restoration of GFPexpression in L3 around veins with vein-clearing symptoms at 6 dpi.Paired views in panels A and B were taken under white-light irradia-tion (left panels) and blue-light irradiation (right panels). Bars, 2 mm.
VOL. 77, 2003 ToMV REPLICATION PROTEIN SUPPRESSES PTGS 11019
on June 21, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

GFP silencing in yellow tissues (Fig. 2 and 3). GFP mRNA wasdetected at low steady-state levels in mock- and TLJ-infectedG3Sm2 leaves, which were nonsymptomatic and displayed noGFP fluorescence (Fig. 4A, lanes 7 and 9).
Low-molecular-weight RNAs were enriched from totalRNAs and then subjected to small-RNA analysis (Fig. 4B).ToMV-derived small RNAs were detected infection specifi-cally in either TLW3- or TLJ-infected leaves. No clear differ-ence was found in the overall distribution patterns of TLW3-and TLJ-derived small RNAs. The minus-strand small RNAsmigrated at 21 to 23 nt and at �25 nt, like gfp siRNAs inG3Sm2. The �25-nt ToMV siRNA band was often faint (inthree independent series of experiments), compared with thecorresponding band of gfp siRNAs. The plus-strand smallRNAs of ToMV migrated at 21 to 24 nt and also containedabundant longer species (�25 nt). gfp siRNAs were detectedonly in G3Sm2 (Fig. 4B, lanes 7 to 9) and were not generatedin G3Sm1 during infection (Fig. 4B, lanes 4 to 6).
These observations indicate that ToMV-specific siRNAs(21- to 25-nt species) were produced after infection. However,because it was unclear when the leaves were infected system-ically and how the viruses were distributed there (in particular,in symptomless leaves of TLJ-infected plants), the timing anddistribution of ToMV siRNAs in the upper leaves were notresolved. Therefore, we examined siRNA production in L0 ofTLW3- and TLJ-inoculated G3Sm2 over time (Fig. 4C and D).TLW3 accumulated vigorously (Fig. 4C, lanes 5 to 8). BecauseTLW3 levels were very high at 6 and 8 dpi, the total RNAsloaded were adjusted on the basis of the amounts of rRNAsstained (Fig. 4C and D). The accumulation of TLJ reached anearly maximum level at approximately 4 dpi (Fig. 4C, lanes 9to 12), consistent with previous reports for L11A (15, 26). GFPmRNA levels did not change significantly upon infection, con-sistent with the observation that no GFP fluorescence wasrestored in L0.
TLW3-derived small RNAs were detected at 2 dpi, and theirlevels increased rapidly as viruses accumulated (Fig. 4D, lanes5 to 8). Therefore, virus-derived small RNAs were generatedin infected tissues from an earlier stage of infection, suggestingthat the siRNA production step was not the major target of theToMV-encoded suppressor. TLJ-derived small RNAs accumu-lated to levels similar to those of TLW3-derived small RNAs(Fig. 4D, lanes 9 to 12). However, the ratio of siRNA togenomic RNA appeared to be higher in TLJ-infected leavesthan in TLW3-infected leaves after infection proceeded (e.g.,�4 dpi). gfp siRNA levels were not detectably influenced byinfection (Fig. 4D, lanes 5 to 12).
ToMV-encoded suppressor inhibits de novo formation of thePTGS system in L0. ToMV small RNAs accumulated in L0,
FIG. 3. Mapping the mutation responsible for the suppressor-de-fective phenotype of ToMV L11A. (A) Schematic representation of thegenomic structures of ToMV and its derivatives. Nucleotides differingfrom wild-type ToMV (ToMV-L and TLW3) are indicated by opentriangles (silent substitutions) and closed triangles (substitutions caus-ing coding changes). The activity that interrupts GFP silencing in thesystemically infected leaves of G3Sm2 is indicated at the right: �,symptom-associated suppression of GFP silencing; �, symptomlessinfection without suppression of GFP silencing. (B) Symptoms andsuppression of GFP silencing in L5 of G3Sm2 infected with TLW3,TLJ, or TLA. Leaves were photographed under white-light irradiation(upper panels) and blue-light irradiation (lower panels). Dark greentissues are marked by asterisks. Bar, 4 mm. (C) No increase in GFPlevels in TLJ- or TLJA1-infected leaves. Proteins extracted from 1 mgof leaf tissue were separated by SDS-PAGE, followed by Coomassiebrilliant blue staining to detect CP (upper panel) or by Western blot-
ting to detect GFP (lower panel). Lanes 1 to 3, healthy leaves (the fifthtrue leaves); lanes 4 to 10, upper leaves (L5) of G3Sm2 inoculated withbuffer (lane 4) or the in vitro transcripts indicated above the panel(lanes 5 to 10). D and Y, tiny dark green islands and yellow tissues,respectively. (D) Transient restoration of GFP silencing in TLJ-in-fected G3Sm2. TLJ was inoculated onto a small leaf (L0), and a typicalGFP patch appearing on L1 was photographed at 5 dpi (left panel). At10 dpi, most GFP patches had disappeared, or the intensity of GFPfluorescence had decreased (middle panel). The right panel shows L1of TLW3-inoculated G3Sm2 at 10 dpi. Bar, 4 mm.
11020 KUBOTA ET AL. J. VIROL.
on June 21, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

whereas GFP silencing was not interrupted in L0. A simpleexplanation is that the ToMV-encoded suppressor inhibits denovo formation of the sequence-specific RNA degradation ma-chinery. However, in G3Sm2 plants showing the weak silencingphenotype, GFP expression was not silenced in the interveinalregions even when the leaves became senescent. Therefore, itwas still possible that the PTGS system was not efficientlyestablished de novo in L0 that we used under standard inocu-lation conditions. Therefore, we examined directly whether thecells in L0 were capable of establishing the ToMV-targeted
PTGS machinery de novo and whether the ToMV-encodedsuppressor inhibited its establishment.
For this purpose, we prepared two constructs carrying theGFP ORF between the duplicated CP subgenomic promoters,TLBN.G3 and TLBN.G3(J) (Fig. 5A). TLBN.G3 carries thewild-type replicase gene, and TLBN.G3(J) has a mutation re-sponsible for the suppressor-defective phenotype. If PTGS isactivated upon ToMV infection in L0 and if the replicase geneis involved in the suppression of PTGS there, GFP signals fromTLBN.G3(J) will weaken even in L0 in due course. Unlike
FIG. 4. RNA analysis of ToMV-infected tobacco. (A) Detection of genomic RNA and GFP mRNA in systemically infected leaves. Plants (NTtobacco, G3Sm1, or G3Sm2) were mock inoculated (lanes m) or inoculated with TLW3 (lanes L) or TLJ (lanes J) and then grown at 28°C. RNAswere extracted from L6 and L7 (excluding class I and II veins) at 12 dpi. The same amounts of total RNAs were loaded in the lanes. GenomicRNA was visualized by ethidium bromide (EtBr) staining (upper panel), and GFP mRNA was visualized by Northern blot analysis (lower panel).(B) Detection of small RNAs derived from the total RNAs shown in panel A. The gel was stained with EtBr prior to blotting (bottom panel). Theamounts of RNAs loaded were adjusted based on EtBr staining of 5.8S and 5S rRNAs. The same membrane was reprobed to detect ToMV- andgfp-specific RNAs, indicated at the right. (�) and (�), small RNAs with plus (sense) and minus (antisense) polarities, respectively. Positions of20- and 25-nt RNA markers are indicated by closed and open arrowheads, respectively. (C) Profiles of accumulation of genomic RNA and GFPmRNA in L0 of G3Sm2. Total RNAs were extracted at the indicated dpi and analyzed as described for panel A. The amounts of RNAs loadedwere adjusted based on rRNA levels; accordingly, the RNA amounts loaded in lanes 7 and 8 were 1.3- and 1.7-fold higher, respectively, than thoseloaded in the other lanes. (D) Detection of small RNAs in L0. Small RNAs in each lane were derived from the total RNAs in the correspondinglane in panel C. The amounts of RNAs loaded were adjusted based on EtBr staining of 5.8S and 5S rRNAs. The gel was processed as describedfor panel B.
VOL. 77, 2003 ToMV REPLICATION PROTEIN SUPPRESSES PTGS 11021
on June 21, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from
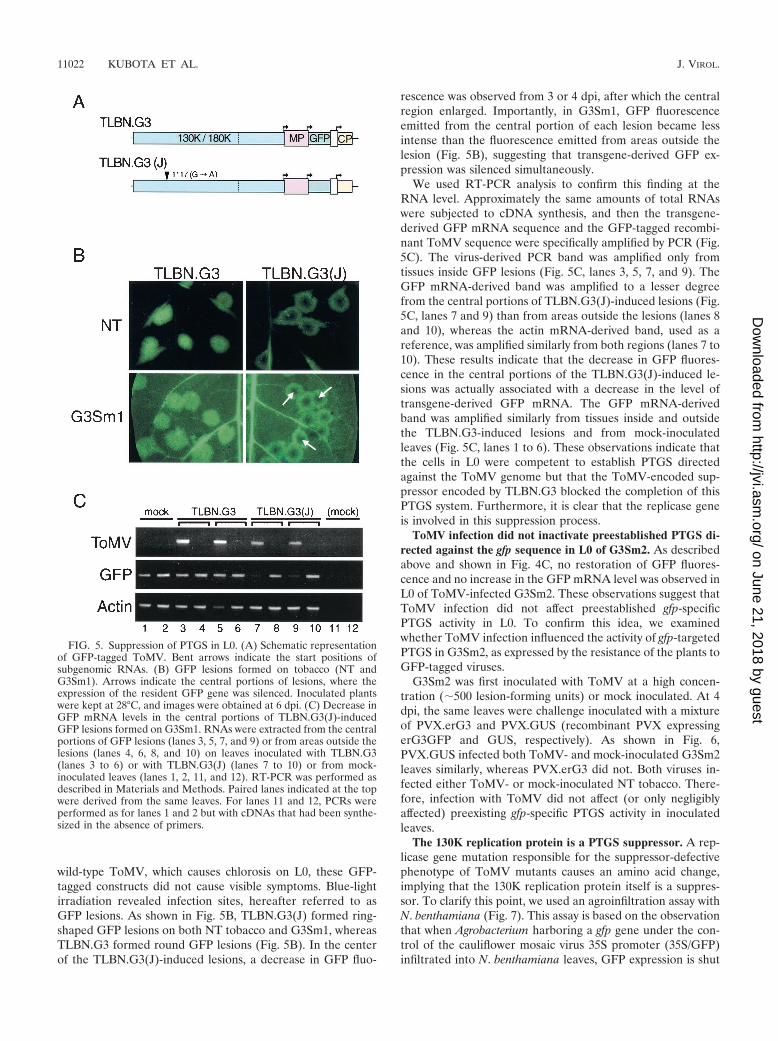
wild-type ToMV, which causes chlorosis on L0, these GFP-tagged constructs did not cause visible symptoms. Blue-lightirradiation revealed infection sites, hereafter referred to asGFP lesions. As shown in Fig. 5B, TLBN.G3(J) formed ring-shaped GFP lesions on both NT tobacco and G3Sm1, whereasTLBN.G3 formed round GFP lesions (Fig. 5B). In the centerof the TLBN.G3(J)-induced lesions, a decrease in GFP fluo-
rescence was observed from 3 or 4 dpi, after which the centralregion enlarged. Importantly, in G3Sm1, GFP fluorescenceemitted from the central portion of each lesion became lessintense than the fluorescence emitted from areas outside thelesion (Fig. 5B), suggesting that transgene-derived GFP ex-pression was silenced simultaneously.
We used RT-PCR analysis to confirm this finding at theRNA level. Approximately the same amounts of total RNAswere subjected to cDNA synthesis, and then the transgene-derived GFP mRNA sequence and the GFP-tagged recombi-nant ToMV sequence were specifically amplified by PCR (Fig.5C). The virus-derived PCR band was amplified only fromtissues inside GFP lesions (Fig. 5C, lanes 3, 5, 7, and 9). TheGFP mRNA-derived band was amplified to a lesser degreefrom the central portions of TLBN.G3(J)-induced lesions (Fig.5C, lanes 7 and 9) than from areas outside the lesions (lanes 8and 10), whereas the actin mRNA-derived band, used as areference, was amplified similarly from both regions (lanes 7 to10). These results indicate that the decrease in GFP fluores-cence in the central portions of the TLBN.G3(J)-induced le-sions was actually associated with a decrease in the level oftransgene-derived GFP mRNA. The GFP mRNA-derivedband was amplified similarly from tissues inside and outsidethe TLBN.G3-induced lesions and from mock-inoculatedleaves (Fig. 5C, lanes 1 to 6). These observations indicate thatthe cells in L0 were competent to establish PTGS directedagainst the ToMV genome but that the ToMV-encoded sup-pressor encoded by TLBN.G3 blocked the completion of thisPTGS system. Furthermore, it is clear that the replicase geneis involved in this suppression process.
ToMV infection did not inactivate preestablished PTGS di-rected against the gfp sequence in L0 of G3Sm2. As describedabove and shown in Fig. 4C, no restoration of GFP fluores-cence and no increase in the GFP mRNA level was observed inL0 of ToMV-infected G3Sm2. These observations suggest thatToMV infection did not affect preestablished gfp-specificPTGS activity in L0. To confirm this idea, we examinedwhether ToMV infection influenced the activity of gfp-targetedPTGS in G3Sm2, as expressed by the resistance of the plants toGFP-tagged viruses.
G3Sm2 was first inoculated with ToMV at a high concen-tration (�500 lesion-forming units) or mock inoculated. At 4dpi, the same leaves were challenge inoculated with a mixtureof PVX.erG3 and PVX.GUS (recombinant PVX expressingerG3GFP and GUS, respectively). As shown in Fig. 6,PVX.GUS infected both ToMV- and mock-inoculated G3Sm2leaves similarly, whereas PVX.erG3 did not. Both viruses in-fected either ToMV- or mock-inoculated NT tobacco. There-fore, infection with ToMV did not affect (or only negligiblyaffected) preexisting gfp-specific PTGS activity in inoculatedleaves.
The 130K replication protein is a PTGS suppressor. A rep-licase gene mutation responsible for the suppressor-defectivephenotype of ToMV mutants causes an amino acid change,implying that the 130K replication protein itself is a suppres-sor. To clarify this point, we used an agroinfiltration assay withN. benthamiana (Fig. 7). This assay is based on the observationthat when Agrobacterium harboring a gfp gene under the con-trol of the cauliflower mosaic virus 35S promoter (35S/GFP)infiltrated into N. benthamiana leaves, GFP expression is shut
FIG. 5. Suppression of PTGS in L0. (A) Schematic representationof GFP-tagged ToMV. Bent arrows indicate the start positions ofsubgenomic RNAs. (B) GFP lesions formed on tobacco (NT andG3Sm1). Arrows indicate the central portions of lesions, where theexpression of the resident GFP gene was silenced. Inoculated plantswere kept at 28°C, and images were obtained at 6 dpi. (C) Decrease inGFP mRNA levels in the central portions of TLBN.G3(J)-inducedGFP lesions formed on G3Sm1. RNAs were extracted from the centralportions of GFP lesions (lanes 3, 5, 7, and 9) or from areas outside thelesions (lanes 4, 6, 8, and 10) on leaves inoculated with TLBN.G3(lanes 3 to 6) or with TLBN.G3(J) (lanes 7 to 10) or from mock-inoculated leaves (lanes 1, 2, 11, and 12). RT-PCR was performed asdescribed in Materials and Methods. Paired lanes indicated at the topwere derived from the same leaves. For lanes 11 and 12, PCRs wereperformed as for lanes 1 and 2 but with cDNAs that had been synthe-sized in the absence of primers.
11022 KUBOTA ET AL. J. VIROL.
on June 21, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

off by local PTGS within a few days after infiltration, whereaslocal PTGS is inhibited when a suppressor-encoding gene iscointroduced (41).
When the 35S/GFP gene was agroinfiltrated into G3Nb3, atransgenic N. benthamiana line constitutively expressing GFP,GFP fluorescence in the infiltrated zone decreased from �2days after infiltration. The concurrent silencing of GFP expres-sion from the resident gene was first observed in marginalregions, where GFP levels were apparently lower than in non-infiltrated regions (Fig. 7A). When the ToMV 130K protein orthe well-known suppressor, HC-Pro of PVY (4), was coex-pressed, GFP silencing was inhibited and, consequently, ahigher level of fluorescence was observed in the infiltratedzone (Fig. 7A). GFP expression was not prolonged when aframeshift mutant of the 130K protein [130K (FS)] was ex-pressed with the 35S/GFP gene (Fig. 7A). The expression ofthe 130K protein in the infiltrated zone was confirmed byWestern blotting (Fig. 7B, lane 4). No enhanced expression ofthe resident GFP gene occurred when the 130K protein wasexpressed alone (data not shown). Therefore, the ToMV 130Kprotein acted as a suppressor of PTGS in the absence of rep-licating virus.
The TLJ-encoded 130K protein, designated 130K(J), alsoexhibited suppressor activity under the assay conditions shownin Fig. 7A, although this activity was weaker than that of thewild-type 130K protein (data not shown). However, it is pos-sible that the effect of the mutation is temperature dependent,because we found that the central holes of TLBN.G3(J)-in-duced GFP lesions on tobacco were larger at 28°C than atlower temperatures. Furthermore, it was recently shown thatPTGS is more strongly induced at higher temperatures in N.benthamiana (36). Therefore, we examined the activity of the130K(J) protein at 28°C.
As shown in Fig. 7C, the 130K(J) protein still suppressed
local GFP silencing; however, the resulting GFP fluorescencedeclined earlier than when the wild-type 130K protein wasexpressed. Western blotting showed that the intensity of greenfluorescence reflected the accumulation of GFP in the infil-trated zone (Fig. 7D), confirming that the suppressor activityof the 130K(J) protein is weaker than that of the wild-type130K protein. Furthermore, the amounts of immunoreactiveGFP (Fig. 7D) were in good agreement with the amounts ofGFP mRNA in the corresponding samples extracted a daybefore (Fig. 7E). Small RNAs with the gfp sequence accumu-lated in the infiltrated zone, irrespective of whether the 130Kprotein was expressed (Fig. 7F), supporting the idea that theToMV-encoded suppressor inhibits the PTGS pathway at astep after the generation of siRNAs in infected cells.
The suppressor activity exhibited by the wild-type 130K pro-tein was weaker than the activity of HC-Pro (Fig. 7C to E).Incomplete inhibition of silencing by the wild-type 130K pro-tein in N. benthamiana was also confirmed by the observationthat TLBN.G3 induced ring-shaped GFP lesions on both NTand GFP-transgenic N. benthamiana leaves, although the cen-tral holes were smaller than those of TLBN.G3(J)-inducedlesions (data not shown).
DISCUSSION
In this work, we investigated ToMV-encoded suppressoractivity by using GFP-constitutive (G3Sm1) and GFP-silenced(G3Sm2) transgenic tobacco lines. Wild-type ToMV (L andTLW3) infection resulted in the suppression of GFP silencingin G3Sm2 together with visible systemic symptoms in theleaves, whereas no suppression of GFP silencing was observedin the nonsymptomatic upper leaves of G3Sm2 infected withthe L-derived attenuated strains, L11 or L11A. Mapping thecausative mutation that results in the suppressor-defectivephenotype of the attenuated strains showed that the replicasegene is involved in the suppression of PTGS (Fig. 3). Anagroinfiltration assay indicated that the 130K replication pro-tein itself is able to suppress local silencing in the absence ofother ToMV-encoded proteins (Fig. 7).
PTGS is known to be associated with the accumulation ofsiRNAs of 21 to 25 nt (10, 11). As in animals, some sorts ofsiRNAs (the class of short siRNAs) are thought to be incor-porated into the RISC to guide the complex to the target inplants (11, 42). In ToMV-infected leaves, this class of siRNAswere found with both polarities, and their levels increased asthe virus accumulated (Fig. 4), suggesting that ToMV-directedPTGS was initiated in the infected cells. These findings alsosuggest that the major target of the ToMV-encoded suppressoris downstream from the production of siRNAs. Analyses withGFP-tagged ToMV constructs demonstrated that the suppres-sor of wild-type ToMV prevents the establishment of theToMV-directed PTGS system (Fig. 5). Furthermore, resis-tance to GFP-tagged PVX in the GFP-silenced leaves ofG3Sm2 was not broken by ToMV infection (Fig. 6), indicatingthat ToMV infection did not detectably inhibit the activity ofthe preexisting sequence-specific PTGS machinery, namely,the RISC. Taken all, it is likely that the ToMV suppressor actsafter the production of siRNAs and inhibits their utilization, sothat the RISC machinery is not newly formed.
The replicase mutation impaired the activity that prevents
FIG. 6. Effect of ToMV infection on the preestablished PTGS sys-tem. NT tobacco or G3Sm2 was mock inoculated or inoculated withToMV. At 4 dpi, when ToMV had spread over the inoculated leaves,a mixture of PVX.GUS and PVX.erG3 was challenge inoculated. Theexpression of GFP and GUS was examined 3 days later. Leaves werephotographed under white-light irradiation (WL, top panels) and un-der blue-light irradiation (GFP, bottom panels). The GUS stainingpatterns of the same regions are shown (middle panels). Bar, 1 cm forall panels.
VOL. 77, 2003 ToMV REPLICATION PROTEIN SUPPRESSES PTGS 11023
on June 21, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

FIG. 7. Suppression of local GFP silencing by ToMV 130K replication protein. GFP-transgenic N. benthamiana (G3Nb3) was agroinfiltratedwith the 35S/GFP gene and a gene for the ToMV 130K replication protein (130K), its frameshift mutant [130K(FS)], the TLJ 130K protein[130K(J)], or PVY HC-Pro. Vector, cointroduction of a vector. (A) GFP fluorescence visualized under blue-light irradiation. The infiltrated plantwas kept at 22°C for 5 days. Arrows indicate the silencing of resident gene-derived GFP expression around the infiltrated zone. (B) Expression ofthe 130K and 130K(J) proteins in infiltrated zones. Proteins were extracted from infiltrated plants kept at 22°C for 2 days. (C) GFP fluorescencevisualized as in panel A. The plant was kept at 22°C for 1 day to establish transformation and at 28°C for 3 days. (D) Detection of GFP in infiltratedzones of plants treated as in panel C. (E) Northern analysis of GFP mRNA in infiltrated zones. The plants were kept at 22°C for 1 day and at 28°Cfor 2 days. The lower panel shows ethidium bromide (EtBr) staining of the same samples. (F) Analysis of siRNAs containing the gfp sequence ininfiltrated zones. Small RNAs in each lane were derived from total RNAs in the corresponding lane of panel E. The gel was stained with EtBrprior to blotting (bottom panel). Positions of 20- and 25-nt RNA markers are indicated by closed and open arrowheads, respectively.
11024 KUBOTA ET AL. J. VIROL.
on June 21, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

de novo formation of the virus-targeting RISC (Fig. 5). How-ever, agroinfiltration experiments indicated that the mutationin the replication protein weakened the suppressor activity butdid not completely abolish it (Fig. 7). Incomplete inhibition ofthe PTGS pathway would explain, at least partly, why TLJ andthe other attenuated strains with the same replicase mutationcan infect systemically but accumulate to lower levels thanwild-type ToMV. The accumulation of L11 and L11A in inoc-ulated leaves ceases a few days after inoculation, when the rateof L11 and L11A RNA synthesis is down-regulated (15, 26).These findings are consistent with our observation thatTLBN.G3(J) formed ring-shaped GFP lesions (Fig. 5), inwhich a decrease in GFP fluorescence was apparent at 3 or 4dpi.
Suppression of PTGS depends on both the activity of thehost PTGS machinery and the activity of a given suppressor. Ithas recently been shown that PTGS is more strongly induced athigher temperatures in N. benthamiana (36) and, therefore,suppression of PTGS by a suppressor is more difficult at highertemperatures. This would explain the observation that the cen-tral holes of TLBN.G3(J)-induced GFP lesions on tobaccowere larger at higher temperatures (Fig. 5B). However, it isalso possible that the mutation itself has a temperature-sensi-tive nature. In relation to this, it should be noted that TLJ andother attenuated strains showed the same symptomless andsuppressor-defective phenotypes on the systemically infectedleaves of tobacco lines at all temperatures examined.
It is generally accepted that siRNAs are generated from theprecursor dsRNA by the action of Dicer or its homologs (39,41, 42). In transgene-mediated PTGS, dsRNA may be formedby the annealing of sense and antisense transcripts or directlyderived from an inverted repeat-containing transcript (40).dsRNA could also be formed from structured ssRNAs throughthe amplification system in which host-encoded RdRp is in-volved (5, 24). The ToMV dsRNA, or the precursor ofsiRNAs, may be generated by virus-encoded RdRp duringreplication or by the host amplification system, which mayrecognize the abundantly synthesized (sub)genomic RNAs inthe infected cells as aberrant ssRNAs. The minus-strandToMV siRNAs, which migrated like the sense and antisensegfp siRNAs detected in G3Sm2 (Fig. 4B and D), probablyreflect the Dicer-like activity in the infected cells. On the otherhand, the plus-strand small RNAs contained longer species(�25 nt) in addition to 21- to 25-nt siRNAs. These �25-ntRNAs must be derived from single-stranded (sub)genomicRNAs, because no corresponding minus-strand RNAs accu-mulated. These plus-strand, �25-nt RNA fragments might beexclusively degradation intermediates. However, it is possiblethat some might be used in the amplification cycle of PTGS(e.g., as primers or templates for dsRNA synthesis).
Irrespective of how ToMV dsRNA and siRNAs are gen-erated in infected cells, the dsRNA must be produced afterreplication proceeds. During infection by ToMV, the 130Kand 180K replication proteins are translated directly fromthe genomic RNA. Of these, the 180K protein contains theRdRp domain and is, therefore, absolutely required for theminus-strand synthesis (12). The accumulation of the 130Kprotein is more than 10-fold greater than that of the 180Kprotein and accordingly, the 130K protein thus producedwould have a good chance of inactivating the host PTGS
pathway, before the replication machinery is set up andsubsequently synthesizes a large amount of progeny RNAs.Therefore, it is a reasonable strategy for the 130K protein toact as the suppressor to prevent the establishment of PTGSthat targets its own genome.
GFP silencing of G3Sm2 was differently suppressed byToMV infection depending on the positions of the leaves.Under standard inoculation conditions, GFP silencing wasnot disrupted in L0 (Fig. 4C), where the preexisting RISCactivity was not, or negligibly, affected by infection (Fig. 6).This observation suggests that the RISC in mature leavesmay have rather a long half-life. In contrast, GFP silencingwas interrupted in the vein-clearing regions and also in thesmall inoculated leaves (Fig. 2C and 3D). In these parts ofthe plant, GFP expression was apparently silenced at thetime of inoculation. This indicates that ToMV can block thesilencing system at work in some tissues (e.g., sink leaves)and suggests that ToMV may suppress PTGS by weakeningpreexisting RISC activity. This suppression might resultfrom an active process in which a virus-encoded or -inducedfactor is involved, or from a passive process that results fromthe disturbance of the intracellular milieu caused by theabrupt invasion and subsequent replication of the virus.Another possibility is that in young tissues, the RISC mighthave a short half-life or that siRNA in the RISC might berapidly exchanged. Whichever the case, the ability to inhibitthe PTGS system, albeit transiently as seen in TLJ-infectedplants (Fig. 3D), would facilitate the establishment of infec-tion, because the invading virus would have time to producesufficient replication protein to suppress the PTGS directedagainst its own genome.
Mosaic symptoms develop only on leaves that were small orinvisible at the time of inoculation (20). Suppression of GFPsilencing in the yellow tissues of the mosaic leaves may beexplained by the inhibition of siRNA utilization; infection at anearlier stage of leaf development would result in a decrease inthe number of active RISC, because it would be diluted duringproliferation. It is also possible that the PTGS system is con-currently inhibited in mosaic leaves by an as-yet-unidentifiedmechanism, as seen in the vein-clearing regions.
ToMV L11A has been used successfully as an attenuatedstrain to protect tomato plants from infection by severe strains(29). Our finding that the replicase mutation in L11A results ina suppressor-defective phenotype suggests that the cross-pro-tection observed in L11A-infected plants is primarily PTGS-mediated; i.e., PTGS directed against the L11A genome pre-vents infection of any virus with sufficient homology to L11A.The attenuated phenotype of L11 and L11A may also be attrib-utable to their defective suppressor activity, because viralsuppressors often determine pathogenicity (31, 33, 34, 44).However, the replicase gene mutation responsible for the sup-pressor-defective phenotype did not completely abolish sup-pressor activity (Fig. 7C). Furthermore, we have found thatsome symptomless strains are suppressor-active insofar as theywere tested with G3Sm2 (data not shown). Therefore, we can-not exclude the possibility that the symptomless and suppres-sor-defective phenotypes of L11 and L11A derive from a singlemutation but are mechanistically distinct.
VOL. 77, 2003 ToMV REPLICATION PROTEIN SUPPRESSES PTGS 11025
on June 21, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

ACKNOWLEDGMENTS
We thank D. Baulcombe for a PVX vector, B. Baker for Agrobac-terium strains and binary vectors, V. Vance for a protocol, and M.Ishikawa for an anti-130K protein antibody.
This work was supported by grants-in-aid from the Ministry of Ed-ucation, Culture, Sports, Science, and Technology of Japan.
REFERENCES
1. Anandalakshmi, R., G. J. Pruss, X. Ge, R. Marathe, A. C. Mallory, T. H.Smith, and V. B. Vance. 1998. A viral suppressor of gene silencing in plants.Proc. Natl. Acad. Sci. USA 95:13079–13084.
2. Atkinson, P. H., and R. E. Matthews. 1970. On the origin of dark green tissuein tobacco leaves infected with tobacco mosaic virus. Virology 40:344–356.
3. Baulcombe, D. C., S. Chapman, and S. Santa Cruz. 1995. Jellyfish greenfluorescent protein as a reporter for virus infections. Plant J. 7:1045–1053.
4. Brigneti, G., O. Voinnet, W. X. Li, L. H. Ji, S. W. Ding, and D. C. Baulcombe.1998. Viral pathogenicity determinants are suppressors of transgene silenc-ing in Nicotiana benthamiana. EMBO J. 17:6739–6746.
5. Dalmay, T., A. Hamilton, S. Rudd, S. Angell, and D. C. Baulcombe. 2000. AnRNA-dependent RNA polymerse gene in Arabidopsis is required for post-transcriptional gene silencing mediated by a transgene but not by a virus.Cell 101:543–553.
6. Erickson, F. L., S. Holzberg, A. Calderon-Urrea, V. Handley, M. Axtell, C.Corr, and B. Baker. 1999. The helicase domain of the TMV replicase pro-teins induces the N-mediated defence response in tobacco. Plant J. 18:67–75.
7. Grayburn, W. S. 1997. Plant transformation techniques and vectors, p. 361–376. In W. V. Dashek (ed.), Methods in plant biochemistry and molecularbiology. CRC Press LLC, Boca Raton, Fla.
8. Guo, H.-S., and S.-W. Ding. 2002. A viral protein inhibits the long rangesignaling activity of the gene silencing signal. EMBO J. 21:398–407.
9. Hagiwara, Y., K. Komoda, T. Yamanaka, A. Tamai, T. Meshi, R. Funada, T.Tsuchiya, S. Naito, and M. Ishikawa. 2003. Subcellular localization of hostand viral proteins associated with tobamovirus RNA replication. EMBO J.22:344–353.
10. Hamilton, A., O. Voinnet, L. Chappell, and D. Baulcombe. 2002. Two classesof short interfering RNA in RNA silencing. EMBO J. 21:4671–4679.
11. Hamilton, A. J., and D. C. Baulcombe. 1999. A species of small antisenseRNA in posttranscriptional gene silencing in plants. Science 286:950–952.
12. Ishikawa, M., T. Meshi, F. Motoyoshi, N. Takamatsu, and Y. Okada. 1986.In vitro mutagenesis of the putative replicase genes of tobacco mosaic virus.Nucleic Acids Res. 14:8291–8305.
13. Kasschau, K. D., and J. C. Carrington. 1998. A counterdefensive strategy ofplant viruses: suppression of posttranscriptional gene silencing. Cell 95:461–470.
14. Kasschau, K. D., Z. Xie, E. Allen, C. Llave, E. J. Chapman, K. A. Krizan, andJ. C. Carrington. 2003. P1/HC-Pro, a viral suppressor of RNA silencing,interferes with Arabidopsis development and miRNA function. Dev. Cell4:205–217.
15. Kiho, Y., and M. Nishiguchi. 1984. Unique nature of an attenuated strain oftobacco mosaic virus: autoregulation. Microbiol. Immunol. 28:589–599.
16. Llave, C., K. D. Kasschau, and J. C. Carrington. 2000. Virus-encoded sup-pressor of posttranscriptional gene silencing targets a maintenance step inthe silencing pathway. Proc. Natl. Acad. Sci. USA 97:13401–13406.
17. Mallory, A. C., L. Ely, T. H. Smith, R. Marathe, R. Anandalakshmi, M.Fagard, H. Vaucheret, G. Pruss, L. Bowman, and V. B. Vance. 2001. HC-Prosuppression of transgene silencing eliminates the small RNAs but not trans-gene methylation or the mobile signal. Plant Cell 13:571–583.
18. Marathe, R., R. Anandalakshmi, T. H. Smith, G. J. Pruss, and V. B. Vance.2000. RNA viruses as inducers, suppressors and targets of post-transcrip-tional gene silencing. Plant Mol. Biol. 43:295–306.
19. Martin, T., R.-V. Wohner, S. Hummel, L. Willmitzer, and W. B. Frommer.1992. The GUS reporter system as a tool to study plant gene expression, p.23–43. In S. R. Gallagher (ed.), GUS protocols: using the GUS gene as areporter of gene expression. Academic Press, Inc., San Diego, Calif.
20. Matthews, R. E. F. 1991. Plant virology, 3rd ed. Academic Press Ltd., Lon-don, England.
21. Meshi, T., M. Ishikawa, F. Motoyoshi, K. Semba, and Y. Okada. 1986. Invitro transcription of infectious RNAs from full-length cDNAs of tobaccomosaic virus. Proc. Natl. Acad. Sci. USA 83:5043–5047.
22. Meshi, T., Y. Watanabe, and Y. Okada. 1992. Molecular pathology of to-bacco mosaic virus revealed by biologically active cDNAs, p. 149–186. In
T. M. A. Wilson and J. W. Davies (ed.), Genetic engineering with plantviruses. CRC Press, Inc., Boca Raton, Fla.
23. Mlotshwa, S., O. Voinnet, M. F. Mette, M. Matzke, H. Vaucheret, S. W. Ding,G. Pruss, and V. B. Vance. 2002. RNA silencing and the mobile silencingsignal. Plant Cell 14:S289–S301.
24. Mourrain, P., C. Beclin, T. Elmayan, F. Feuerbach, C. Godon, J.-B. Morel,S. Nicic, N. Picault, K. Remoue, M. Sanial, T.-A. Vo, and H. Vaucheret. 2000.Arabidopsis SGS2 and SGS3 genes are required for posttranscriptional genesilencing and natural virus resistance. Cell 101:533–542.
25. Nishiguchi, M., S. Kikuchi, Y. Kiho, T. Ohno, T. Meshi, and Y. Okada. 1985.Molecular basis of plant viral virulence; the complete nucleotide sequence ofan attenuated strain of tobacco mosaic virus. Nucleic Acids Res. 13:5585–5590.
26. Nishiguchi, M., F. Motoyoshi, and N. Oshima. 1990. Comparison of virusproduction in infected plants between an attenuated tomato strain (L11A)and wild strain (L) of tobacco mosaic virus. Ann. Phytopathol. Soc. Jpn.56:691–694.
27. Ohno, T., M. Aoyagi, Y. Yamanashi, H. Saito, S. Ikawa, T. Meshi, and Y.Okada. 1984. Nucleotide sequence of the tobacco mosaic virus (tomatostrain) genome and comparison with the common strain genome. J. Bio-chem. 96:1915–1923.
28. Olszewski, N. E., F. B. Matrtin, and F. M. Ausubel. 1988. Specialized binaryvector for plant transformation: expression of the Arabidopsis thalianaAHAS gene in Nicotiana tabacum. Nucleic Acids Res. 16:10765–10782.
29. Oshima, N. 1981. Control of tomato mosaic disease by attenuated virus. Jpn.Agric. Res. Q. 14:222–228.
30. Plasterk, R. H. A. 2002. RNA silencing: the genome’s immune system.Science 296:1263–1265.
31. Pruss, G., X. Ge, X. M. Shi, J. C. Carrington, and V. B. Vance. 1997. Plantviral synergism: the potyviral genome encodes a broad-range pathogenicityenhancer that transactivates replication of heterologous viruses. Plant Cell9:859–868.
32. Qu, F., T. Ren, and T. J. Morris. 2003. The coat protein of turnip crinklevirus suppresses posttranscriptional gene silencing at an early initiation step.J. Virol. 77:511–522.
33. Ratcliff, F., B. D. Harrison, and D. C. Baulcombe. 1977. A similarity betweenviral defense and gene silencing in plants. Science 276:1558–1560.
34. Ratcliff, F. G., S. A. MacFarlane, and D. C. Baulcombe. 1999. Gene silencingwithout DNA: RNA-mediated cross-protection between viruses. Plant Cell11:1207–1215.
35. Silhavy, D., A. Molnar, A. Lucioli, G. Szittya, C. Hornyik, M. Tavazza, andJ. Burgyan. 2002. A viral protein suppresses RNA silencing and binds si-lencing-generated, 21- to 25-nucleotide double-stranded RNAs. EMBO J.21:3070–3080.
36. Szittya, G., D. Silhavy, A. Molnar, Z. Havelda, A. Lovas, L. Lakatos, Z.Banfalvi, and J. Burgyan. 2003. Low temperature inhibits RNA silencing-mediated defence by the control of siRNA generation. EMBO J. 22:633–640.
37. Tamai, A., and T. Meshi. 2001. Cell-to-cell movement of Potato virus X: therole of p12 and p8 encoded by the second and third open reading frames ofthe triple gene block. Mol. Plant-Microbe Interact. 14:1158–1167.
38. Tamai, A., and T. Meshi. 2001. Tobamoviral movement protein transientlyexpressed in a single epidermal cell functions beyond multiple plasmodes-mata and spreads multicellularly in an infection-coupled manner. Mol. Plant-Microbe Interact. 14:126–134.
39. Tang, G., B. J. Reinhart, D. P. Bartel, and P. D. Zamore. 2003. A biochem-ical framework for RNA silencing in plants. Genes Dev. 17:49–63.
40. Vaucheret, H., C. Beclin, and M. Fagard. 2001. Post-transcriptional genesilencing in plants. J. Cell Sci. 114:3083–3091.
41. Voinnet, O. 2001. RNA silencing as a plant immune system against viruses.Trends Genet. 17:449–459.
42. Voinnet, O. 2002. RNA silencing: small RNAs as ubiquitous regulators ofgene expression. Curr. Opin. Plant Biol. 5:444–451.
43. Voinnet, O., C. Lederer, and D. C. Baulcombe. 2000. A viral movementprotein prevents spread of the gene silencing signal in Nicotiana benthami-ana. Cell 103:157–167.
44. Voinnet, O., Y. M. Pinto, and D. C. Baulcombe. 1999. Suppression of genesilencing: a general strategy used by diverse DNA and RNA viruses of plants.Proc. Natl. Acad. Sci. USA 96:14147–14152.
45. Waterhouse, P. M., M.-B. Wang, and T. Lough. 2001. Gene silencing as anadaptive defence against viruses. Nature 411:834–842.
46. Zamore, P. D. 2002. Ancient pathways programmed by small RNAs. Science296:1265–1269.
11026 KUBOTA ET AL. J. VIROL.
on June 21, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from