title: efficient metabolic exchange and electron transfer within a
TRANSCRIPT

1
Title: Efficient metabolic exchange and electron transfer within a syntrophic TCE degrading co-1
culture of Dehalococcoides mccartyi 195 and Syntrophomonas wolfei 2
Running Head: Syntrophic exchanges between D. mccartyi and S. wolfei 3
Authors: Xinwei Mao,1 Benoit Stenuit,
1* Alexandra Polasko,
1 Lisa Alvarez-Cohen
1,2 4
1Department of Civil and Environmental Engineering, College of Engineering, University of California, 5
Berkeley, CA, 94720; 6
2 Earth Sciences Division, Lawrence Berkeley National Laboratory, Berkeley, CA 94720 7
8
Corresponding author: 9
Lisa Alvarez-Cohen 10
Civil and Environmental Engineering 11
726 Davis Hall 12
University of California 13
Berkeley, CA 94720-1710 14
Phone: (510)643-5969 15
17
*Present address: Catholic University of Louvain, Earth & Life Institute, Applied Microbiology and 18
Bioprocess Cluster, Louvain-la-Neuve, Belgium 19
20
Key words: chlorinated solvents, reductive dechlorination, Dehalococcoides, syntrophy, cell aggregates, 21
bioremediation 22
23
24
AEM Accepted Manuscript Posted Online 9 January 2015Appl. Environ. Microbiol. doi:10.1128/AEM.03464-14Copyright © 2015, American Society for Microbiology. All Rights Reserved.
on March 27, 2018 by guest
http://aem.asm
.org/D
ownloaded from

2
Abstract 25
Dehalococcoides mccartyi 195 (strain 195) and Syntrophomonas wolfei (S. wolfei) were grown 26
in a sustainable syntrophic co-culture using butyrate as electron donor and carbon source and 27
TCE as electron acceptor. The maximum dechlorination rate (9.9 ± 0.1 µmol d-1
) and cell yield 28
(1.1 ± 0.3 × 108
cells µmol-1
Cl-) of strain 195 maintained in co-culture were respectively 2.6 29
times and 1.6 times higher than those measured in the pure culture. Strain 195 cells concentration 30
was about 16 times higher than S. wolfei in the co-culture. Aqueous H2 concentrations ranged 31
from 24 to 180 nM during dechlorination and increased to 350 ± 20 nM when TCE was depleted, 32
resulting in cessation of butyrate fermentation by S. wolfei with a theoretical Gibbs free energy 33
of -13.7 ± 0.2 kJ mol-1
. Carbon monoxide in the co-culture was around 0.06 µmol per bottle, 34
which was lower than that observed for strain 195 in isolation. The minimum H2 threshold value 35
for TCE dechlorination by strain 195 in the co-culture was 0.6 ± 0.1 nM. Cell aggregates during 36
syntrophic growth were observed by scanning electron microscopy. The interspecies distances to 37
achieve H2 fluxes required to support the measured dechlorination rates were predicted using the 38
Fick’s law and demonstrated the need for aggregation. Filamentous appendages and EPS-like 39
structures were present in the intercellular spaces. The transcriptome of strain 195 during 40
exponential growth in the co-culture indicated increased ABC-transporter activities compared 41
with the pure culture, while the membrane-bound energy metabolism related genes were 42
expressed at stable levels. 43
44
45
46
on March 27, 2018 by guest
http://aem.asm
.org/D
ownloaded from

3
Introduction 47
Groundwater contamination by trichloroethene (TCE), a potential human carcinogen, 48
poses a serious threat to human health and can lead to the generation of vinyl chloride (VC), 49
which is a known human carcinogen (1). Strains of Dehalococcoides mccartyi are the only 50
known bacteria that can completely degrade TCE to the benign end product ethene. 51
Biostimulation of indigenous Dehalococcoides sp. and bioaugmentation using Dehalococcoides-52
containing cultures are recognized as the most reliable in situ bioremediation technologies 53
resulting in complete dechlorination of TCE to ethene (2). However, the mechanisms that 54
regulate the activity of D. mccartyi within natural ecosystems and shape its functional robustness 55
under disturbed environments are poorly understood due to multi-scale microbial community 56
complexity and heterogeneity of biogeochemical processes involved in the sequential 57
degradation (3-4). D. mccartyi exhibits specific restrictive metabolic requirements for a variety 58
of exogenous compounds, such as hydrogen, acetate, corrinoids, biotin and thiamine, which can 59
be supplied by other microbial genera through a complex metabolic network (1,5-8). Therefore, 60
the growth of D. mccartyi is more robust within functionally-diverse microbial communities that 61
are deterministically assembled than in pure cultures (5,8-9). Previous studies have shown that D. 62
mccartyi uses hydrogen as its sole electron donor for TCE dechlorination and out-competes other 63
terminal electron-accepting processes such as methanogenesis and acetogenesis at low hydrogen 64
partial pressures (10-11). Interspecies hydrogen transfer between D. mccartyi and supportive 65
organisms is a key process of electron flow that drives reductive dechlorination in the 66
environment. Although dechlorinating microbial communities have been extensively studied 67
over the past decades (4,10,12-13), community assembly processes in the highly specialized 68
ecological niche of Dehalococcoides and the network of interactions between D. mccartyi, its 69
on March 27, 2018 by guest
http://aem.asm
.org/D
ownloaded from

4
syntrophic partners and other co-existing community members have yet to be deciphered. The 70
optimization of D. mccartyi-based bioremediation systems to treat TCE-contaminated 71
groundwater would be facilitated by a systems-level understanding of interspecies electron, 72
energy and metabolite transfers that shape the structural and functional robustness of TCE-73
dechlorinating microbial communities. 74
To date, only a few studies of D. mccartyi-containing constructed co-cultures and tri-75
cultures have been published (5,7,14-16). A study with D. mccartyi 195 (strain 195) revealed that 76
its growth in defined medium could be optimized by providing high concentrations of vitamin 77
B12 and that over the short term, the strain could be grown to higher densities in co-cultures and 78
tri-cultures with fermenters Desulfovibrio desulfuricans and/or Acetobacterium woodii that 79
convert lactate to generate the hydrogen and acetate required by D. mccartyi (5). Recent studies 80
demonstrated interspecies corrinoid transfer between Geobacter lovleyi and D. mccartyi strains 81
BAV1 and FL2 (16) and interspecies cobamide transfer from a corrinoid-producing methanogen 82
and acetogen to D. mccartyi (7). Another study showed that strain 195 can grow in a long-term 83
sustainable syntrophic association with Desulfovibrio vulgaris Hildenborough (DvH) as a co-84
culture as well as with hydrogenotrophic methanogen Methanobacterium congolense (MC) as a 85
tri-culture (15). The maximum dechlorination rates and cell yield of strain 195 were enhanced 86
significantly in the defined consortia. Another unexpected syntrophic association has recently 87
been discovered between carbon monoxide (CO)-producing strain 195 and CO-metabolizing 88
anaerobes which enhance the growth and dechlorination activity of strain 195 by preventing the 89
accumulation of toxic CO as an obligate by-product from acetyl-CoA cleavage (17). Although 90
these studies demonstrated the robust growth of D. mccartyi-containing co-cultures on PCE/TCE 91
with higher dechlorination activity and cell yields than isolates, a clear and mechanistic 92
on March 27, 2018 by guest
http://aem.asm
.org/D
ownloaded from

5
understanding of metabolic cross-feeding and electron transfer between D. mccartyi and its 93
syntrophic partner(s) is needed to establish predictive models for dechlorination activity. 94
Butyrate fermentation is an endergonic reaction under standard conditions (Table 1) that 95
can only be carried out for energy generation by syntrophic microorganisms growing with H2 96
consumers (18-20). Syntrophomonas wolfei is a model butyrate fermenter that grows 97
syntrophically with hydrogenotrophic methanogens (19-21). S. wolfei can also grow without a 98
syntrophic partner on crotonate as the sole energy source through its disproportionation to 99
acetate and butyrate (Table 1) (22). In previous studies, Syntrophomonas spp. have been detected 100
in butyrate-fed dechlorinating enrichment cultures in significant relative abundance (13,23). In 101
this study, a TCE and butyrate-fed syntrophic co-culture of strain 195 with S. wolfei was 102
established and maintained to study the physiology and transcriptome of syntrophically growing 103
D. mccartyi. Spatial architecture and the physical proximity of the cells were analyzed in the co-104
culture S. wolfei/strain 195 and another syntrophic co-culture DvH/strain 195 (15). The 105
knowledge gained from this study provides us with a more fundamental understanding of the 106
metabolic exchange and energy transfer among the key players of TCE-dechlorinating 107
communities. 108
Material and methods 109
Bacterial cultures and growth conditions 110
Bacterial co-cultures of strain 195 and S. wolfei (5% vol/vol inoculation of each bacterium) were 111
initially established in 160 mL serum bottles containing 100 mL defined medium (5) with TCE 112
supplied at a liquid concentration of 0.6 mM (corresponding to 78 µmol TCE per bottle), 10 mM 113
crotonic acid, 100 µg L-1
vitamin B12 and N2/CO2 (90:10 v/v) headspace at 34 °C with no 114
on March 27, 2018 by guest
http://aem.asm
.org/D
ownloaded from

6
agitation. Cultures were subsequently transferred (5% vol/vol inoculation) for growth on 4 mM 115
butyric acid with 0.6 mM TCE as the electron acceptor. The established syntrophic co-culture 116
was continuously and stably maintained on butyrate in 100 mL defined medium over 45 sub-117
culturing events before the experiments were performed. The electron donor-limited condition 118
(on a hydrogen production/consumption basis) was achieved by feeding the co-culture 0.25 mM 119
butyrate (25 µmol per bottle that could theoretically generate 50 µmol H2 based on stoichiometry) 120
and 0.6 mM TCE (78 µmol per bottle that requires 234 µmol H2 to reductively dechlorinate TCE 121
to ethene). The strain 195 isolate was grown in defined medium with H2/CO2 (90:10 v/v) 122
headspace, 0.6 mM TCE as electron acceptor and 2 mM acetate as carbon source. Pure S. wolfei 123
was grown on crotonate in 160 mL serum bottles as described previously (Beaty et al. 1987). Co-124
culture DvH and strain 195 were grown in the same medium with the substitutions of 5 mM 125
lactate. Pure culture and co-culture purities were routinely checked by phase-contrast microscopy 126
and DGGE (Denaturing Gradient Gel Electrophoresis). 127
Chemical analysis 128
Chloroethenes and ethene were measured by FID-gas chromatograph using 100 µL headspace 129
samples, and hydrogen and carbon monoxide was measured by RGD-gas chromatography using 130
300 µL headspace sample as described previously (23-24). Organic acids, including butyrate and 131
acetate, were analyzed with a high-performance liquid chromatograph as described previously
132
(23). Mass of each compound was calculated based on gas-liquid equilibrium by using Henry’s 133
law constants at 34 °C according to: mass (μmol/bottle) = Cl×Vl + Cg×Vg, 𝑘Hcc = 𝐶l/𝐶g. Cl: liquid 134
phase concentration (μM); Vl : volume of liquid phase (L); Cg: gas phase concentration (μM); Vg: 135
volume of gas phase (L); 𝑘Hcc : Henry’s law constant (unitless). 136
on March 27, 2018 by guest
http://aem.asm
.org/D
ownloaded from

7
Scanning electron microscopy (SEM) 137
The co-cultures (S.wolfei and strain 195 grown on butyrate, and DvH and strain 195 on lactate) 138
provided with 0.6 mM TCE were collected when cell growth had ceased and treated by the 139
following standard protocols for SEM observation (http://em-140
lab.berkeley.edu/EML/protocols/psem.php). Briefly, cultures were collected by slowly filtering 1 141
mL fresh liquid sample through a 0.2µm GTTP filter (IsoporeTM
membrane, polycarbonate, 142
hydrophilic, 25 mm in diameter from Millipore), followed by chemical fixation, dehydration, 143
critical point drying and conductive coating (25). SEM images were obtained according to 144
standard operation protocols (26). The maximum interspecies distances between strain 195 and 145
S.wolfei that would enable the observed dechlorination rates for butyrate fermentation were 146
estimated using Fick’s diffusion law according to the procedure described by Ishii (27): 147
148
JH2 = H2 flux across the total surface areas (As,tot) of S. wolfei (pmol m-2
cell d-1
); 149
As = 2.1× 10-12
m2 cell
-1, S. wolfei surface area calculated based on the assumption of 0.25 by 150
2.5 µm cells (based on SEM observation). As,tot =As× S.wolfei cell number 151
DH2 = 6.31 10-5
cm2 s
-1, molecular diffusion coefficient in water for hydrogen at 35 °C (28); 152
CH2-sw = maximum H2 concentration enabling exergonic fermentation (μM) (the highest H2 153
level at which S.wolfei can ferment butyrate.); 154
CH2-195 = theoretical minimum H2 concentration useable by strain 195 for energy generation 155
(μM) (the lowest H2 level at which strain 195 can dechlorinate TCE); 156
JH2= 𝐷H2
×𝐶H2−𝑠𝑤 − 𝐶𝐻2−195
d𝑠𝑤−195
on March 27, 2018 by guest
http://aem.asm
.org/D
ownloaded from

8
dsw-195 = allowed interspecies distance for accomplishing syntrophic oxidation at an observed 157
substrate utilization rate (μm). 158
DNA extraction and cell number quantification 159
1.5 mL liquid samples were collected for cell density measurements and cells were harvested by 160
centrifugation (21,000 × g, 10 min at 4°C). Genomic DNA was extracted from cell pellets using 161
Qiagen DNeasy Blood and Tissue Kit according to the manufacturer’s instructions for Gram-162
positive bacteria. qPCR using SYBR Green-based detection reagents was applied to quantify 163
gene copy numbers of each bacterium with S.wolfei 16S rRNA gene primers (forward primer 5’-164
GTATCGACCCCTTCTGTGCC-3’, and reverse primer 5’-CCCCAGGCGGGATACTTATT-3’) 165
(19), and D. mccartyi tceA gene primers (forward primer 5’-166
ATCCAGATTATGACCCTGGTGAA-3’ and reverse primer 5’-167
GCGGCATATATTAGGGCATCTT-3’), as previously described (29). The electron equivalents 168
(µmol) diverted to biomass were calculated based on empirical biomass formula C5H7O2N (113 169
g mol-1
cell, 20 e- eq mol
-1 cells). 170
RNA preparation 171
Cultures were sampled for RNA on day 6 during exponential growth when around 75% of 78 172
µmol TCE was dechlorinated (~20 µmol TCE remained). In order to collect sufficient material 173
for transcriptomic microarray analysis, 18 bottles of pure strain 195 and 18 bottles of the co-174
culture were inoculated and grown from triplicate bottles of the isolate and co-culture, 175
respectively. For each biological triplicate, cells from six bottles were collected by vacuum 176
filtration on day 6 during active dechlorination for the co-culture and day 12 for the strain 195 177
pure culture (200 mL culture per filter, 0.2-µm autoclaved GVWP filter (Durapore membrane, 178
on March 27, 2018 by guest
http://aem.asm
.org/D
ownloaded from

9
Millipore, Billerica, MA). Each filter was placed in a 2 mL orange-cap micro-centrifuge tube, 179
frozen with liquid nitrogen and stored at -80 °C until further processing. 180
RNA was extracted using the phenol-chloroform method described previously (31) with the 181
following minor modifications. The ratio of phenol (pH 4.0)-chloroform-isoamylalcohol used for 182
extraction was 25:24:1 (vol:vol), and the RNA pellet was re-suspended in 100 µL of nuclease-183
free water. RNA samples were purified following manufacturer’s instructions with the AllPrep 184
DNA/RNA Mini Kit (Qiagen). Additional DNA contamination was removed with Turbo DNA 185
free kit (Ambion, Austin, TX) according to the manufacturer’s instructions. The quality of RNA 186
samples was checked by electrophoresis (1.0 µL RNA sample), and the concentrations of RNA 187
samples were quantified using a nano-photometer (IMPLEN, Westlake Village, CA, USA). The 188
ratio of A260/A280 for all samples was between ~1.80-2.0. Purified RNA was stored at -80 °C 189
prior to further use. 190
Transcriptomic microarray analysis 191
A complete description of the Affymetrix GeneChip microarray used in this study has been 192
reported elsewhere (4). Briefly, the chip contains 4,744 probe sets that represent more than 98% 193
of the ORFs from four published Dehalococcoides genomes (strain 195, VS, BAV1, and 194
CBDB1). cDNA was synthesized from 9 µg RNA, then each cDNA sample was fragmented, 195
labeled and hybridized to each array. All procedures were performed with minimal modifications 196
to the protocols in section 3 of the Affymetrix GeneChip Expression Analysis Technical Manual 197
(Affymetrix, Santa Clara, CA http://www.affymetrix.com). Microarray data analysis methods 198
were described previously (15,32). 199
on March 27, 2018 by guest
http://aem.asm
.org/D
ownloaded from

10
Results 200
Syntrophic co-culture degradation characteristics 201
TCE did not inhibit growth of the S. wolfei isolate at concentrations up to 0.6 mM 202
(Supplemental material, Part 1, Fig. S1); therefore, co-cultures were maintained with 0.6 mM 203
TCE and 4 mM butyrate. After subculturing this co-culture 45 times, the maximum 204
dechlorination rate of strain 195 was approximately 2.6 fold (9.9 ± 0.1 µmol d-1
) (Fig. 1a) greater 205
than the strain 195 isolate (3.8 ± 0.1 µmol d-1
) (15). The calculation of strain 195 cell yield was 206
based on the metabolic reductive TCE dechlorination to VC, and the cell yield of strain 195 was 207
1.6 times greater in the co-culture (1.1 ± 0.3 × 108 cells µmol
-1 Cl
-) (Fig. 1b) compared to the 208
pure culture (6.8 ± 0.9 × 107 cells µmol
-1 Cl
-) (15), similar to results from a co-culture containing 209
a sulfate-reducing bacterium (9.0 ± 0.5 × 107
cells µmol-1
Cl-) (15) (Table 2). The initial 210
concentration of butyrate (from 1 mM to 20 mM) did not affect the dechlorination rate (data not 211
shown), showing that high butyrate concentrations are not inhibitory to dechlorination. Cell 212
numbers of strain 195 (1.3 ± 0.2 × 108 cells mL
-1) were consistently about 16 times higher than S. 213
wolfei (7.7 ± 0.1 × 106
cells mL-1
) in the co-cultures growing on butyrate. In contrast, when 214
maintaining the co-culture on crotonate, the cell number ratio was about 1.3:1 (Supplemental 215
material, Part 1, Fig. S2). In the syntrophic co-culture with DvH growing on lactate, the cell 216
number ratio of strain 195 to DvH was reported to be about 5:1 (15). 217
When the ratio of butyrate to TCE in the co-culture was maintained under electron 218
acceptor limitation with 79 ± 6.7 µmol TCE and 440 ± 29 µmol butyrate (measured by HPLC) as 219
the electron donor, H2 levels remained steady at ~24-180 nM during active dechlorination (from 220
day 0 to day 7) and increased to 350 ± 20 nM right after TCE and cis-DCE were consumed on 221
on March 27, 2018 by guest
http://aem.asm
.org/D
ownloaded from

11
day 8 (Fig. 1c and Supplemental material, Part 2, Table S2). This H2 concentration was similar to 222
the level (480 nM) that resulted in cessation of S. wolfei growth on butyrate in isolation (33). At 223
the end of the experiment (day 18), 88.4% of the amended TCE could be accounted for in 224
dechlorination products VC (7.4± 0.3 % molar equivalents) and ethene (81.0± 0.1% molar 225
equivalents). The missing 11.6% of initial TCE was likely due to the numerous sampling events 226
during the experiment, as confirmed by losses in the abiotic controls (8.7% loss of initial TCE 227
added). In addition, 115 ± 3.0 µmol butyrate was consumed while 234 ± 9 µmol acetate and 3.9 228
±1.1 µmol H2 remained in the bottles (in the abiotic control, the amount of butyrate decreased 26 229
µmol with no production of acetate or H2). This indicates that around 88.6% of electrons (0.886, 230
this corresponds to 𝑓𝑒° close to 0.84 from Fig. 1d) (in the form of H2) generated from butyrate 231
fermentation (ca. 230 µmol H2) supported dechlorination. A calculation based on biomass cell 232
numbers (Assumed 0.5 gram dry weight per gram cell, i.e. water accounts for 50% of the cell 233
weight. The cell formula was C5H7O2N, with 20 electron equivalent per mol biomass) indicates 234
that 7.4 % (17 µmol H2) of the electrons from butyrate fermentation (in the form of H2) were 235
diverted to biomass production, giving a total electron recovery of about 98 %. This result 236
demonstrates that butyrate was efficiently used as the electron donor in the co-culture with high 237
electron transfer efficiency between the two bacteria. 238
The calculated Gibbs free energy of reaction (∆𝐺𝑟) (butyrate fermentation and associated 239
hydrogen generation by S.wolfei) was -21.7 ± 0.3 kJ mol-1
on day 6 and decreased to -13.7 ± 0.2 240
kJ mol-1
on day 8 when TCE and cis-DCE was depleted. This number was smaller than the 241
hypothetical minimum energy (-20 kJ mol-1
) required by a bacterium to exploit the free energy 242
change in a reaction and support growth (34) (detailed calculations in Supplemental material, 243
Part 2). After TCE and cis-DCE depletion, we observed a slight decrease in butyrate (36.0 ± 6.6 244
on March 27, 2018 by guest
http://aem.asm
.org/D
ownloaded from

12
µmol) during co-metabolic VC dechlorination (Fig. 1c) with H2 concentrations increasing to a 245
stable level of 1.2 ± 0.3 µM after day 10 while the cell numbers of strain 195 and S. wolfei were 246
observed to decrease (Fig. 1b and 1c). The calculated ∆𝐺𝑟 of butyrate fermentation during the 247
cometabolic process was less negative and reached -5.7 ± 1.4 kJ mol-1
by the end of the 248
experiment (Supplemental material, Part 2, Fig. S3). 249
In order to determine the hydrogen threshold of strain 195 in the co-culture, it was grown 250
under electron donor-limited conditions (TCE fed in excess). H2 concentrations dropped to 251
thresholds of 0.6 ± 0.1 nM at which point TCE was not further degraded by strain 195 and 252
butyrate depletion ceased (Supplemental material, Part 3, Figure S4). In order to validate our 253
method for quantifying the minimum H2-threshold concentration, we measured the hydrogen 254
threshold of the sulfate reducing microorganism Desulfovibrio vulgaris Hildenborough (DvH), 255
and obtained 15.2 ± 1.4 nM, which is in agreement with literature results (14.8 nM) for this 256
bacterium (35). 257
We tested the inhibitory effect of CO on S. wolfei by exposing cells to 0.6 to 8 µmol CO 258
per bottle and observed no significant effect on cell growth (Fig. 2a). CO accumulation for the S. 259
wolfei isolate growing on crotonate was 0.02 µmol per bottle throughout the experiment (Fig. 2b), 260
which was at the same level as the abiotic control (data not shown). In the co-culture growing on 261
butyrate, CO was measured at around 0.06 µmol per bottle. This was lower than that detected for 262
the strain 195 isolate (0.5 ± 0.1 µmol per bottle after one dose of TCE (77 µmol) was depleted) 263
(17). Furthermore, we found that pure S. wolfei could consume CO from 14.5 ± 1.5 µmol to 0.4 264
± 0.1 µmol in 42 days (Fig. 2c) when growing on crotonate. 265
on March 27, 2018 by guest
http://aem.asm
.org/D
ownloaded from

13
Formation of cell aggregates 266
Strain 195 and S. wolfei isolates as well as the co-cultures of S. wolfei and strain 195 267
maintained on butyrate were grown to late exponential to early stationary phase before samples 268
were analyzed by scanning electron microscopy (SEM). Figures 3a and 3b show specific cell 269
morphology of the isolates with strain 195 growing in individual coccus-shapes with diameters 270
of around 0.5-1 µm while most S. wolfei cells grew individually or in pairs as 0.25 - 0.5 by 2.5 - 271
5 µm rods as observed in previous studies (1,21). When butyrate was amended to the co-culture 272
as electron donor with TCE as electron acceptor, the cells form cell aggregates with size ranges 273
from 2 µm to 10 µm (Fig. 3c-e). In relatively small aggregates, cells were found connected with 274
flagellum-like filaments (Fig. 3c,d). In large aggregates, EPS-like structures were observed (Fig. 275
3d,e). Cell aggregates were also observed during exponential growth phase (Supplemental 276
material, Part 4, Figure S5). However, when strain 195 was grown as a co-culture with DvH on 277
lactate, the two strains grew together but did not form obvious cell aggregates (Fig. 3f). 278
In order to determine whether the co-cultures may be capable of sharing electrons via 279
direct interspecies electron transfer (DIET), the crotonate and butyrate co-cultures were sent to 280
Professor Lovley’s laboratory for conductivity testing (36). The co-cultures exhibited three 281
orders of magnitude lower conductivity than Geobacter, indicating that H2 rather than DIET is 282
the transfer mechanism. In addition, since S. wolfei could potentially use both hydrogen and 283
formate as electron carriers for interspecies electron transfer (37), we tested the expression levels 284
of several formate dehydrogenase genes (fdhA: Swol_0786, Swol_0800, Swol_1825) in co-285
cultures grown on crotonate or butyrate and S. wolfei grown in isolation. The relative expression 286
of fdhA subunits to gyrB gene was generally in the range of 0.6-1.2 units for all three conditions 287
on March 27, 2018 by guest
http://aem.asm
.org/D
ownloaded from

14
(38), confirming that hydrogen rather than formate was the exclusive electron shuttle between 288
these two bacteria. 289
The theoretical maximum interspecies distance for an observed rate of molecular transfer 290
between two microorganisms can be calculated using Fick’s diffusion law (20). Here, this 291
distance was calculated for co-culture growth on butyrate (Table 3, with detailed calculations in 292
Supplemental material, Part 4) by defining JH2 as the hydrogen flux between S. wolfei and strain 293
195. JH2 was calculated from the total surface areas of S. wolfei and substrate oxidation rate 294
(hydrogen production rate = 2 × butyrate oxidation rate) measured in the co-culture experiment 295
using equation 2 in Table 1. CH2-sw (0.35 ± 0.02 µM) is the maximum H2 concentration 296
immediately outside of an S. wolfei cell when fermentation by S. wolfei stopped on butyrate and 297
CH2- 195 is the theoretical minimum H2 concentration above which strain 195 can gain energy, 298
estimated in this study as 0.6 ± 0.1 nM. dsyn-195 is then the calculated maximum interspecies 299
distance for accomplishing syntrophic oxidation at the observed substrate oxidation rate. 300
The theoretical mean cell-cell distance of randomly dispersed cells is calculated based on 301
the total cell numbers (quantified using qPCR) suspended in the unstirred liquid culture (Table 3). 302
In this study, cell settling was not observed for the strain 195 isolate or co-cultures growing on 303
butyrate (strain 195 and S. wolfei) or lactate (strain 195 and DvH). Considering that the cell 304
number ratio of strain 195 to S. wolfei was about 20:1 on day 4 (Fig. 1c), strain 195 cells account 305
for the majority of all cells in the bottle, therefore the average cell-cell distance between 306
suspended strain 195 and S. wolfei cells (i.e., >> 27.1 µm, Table 3) would be larger than the 307
calculated theoretical maximum cell-cell distance for achieving butyrate fermentation (Table 3). 308
In order to accomplish syntrophic butyrate oxidation at the rate observed, the average 309
interspecies distance must be much less than the distance between randomly dispersed cells 310
on March 27, 2018 by guest
http://aem.asm
.org/D
ownloaded from

15
(Table 3), necessitating the formation of aggregates. In addition, according to the 311
thermodynamically consistent rate law, the calculated free energy available for S. wolfei growth 312
on day 4 was -27.4 ± 1.1 kJ mol-1
which is small (Supplemental material, Part 2, Fig. S3), 313
indicating that close physical contact between the two species is particularly important for 314
efficient syntrophic butyrate fermentation. In contrast, when strain 195 was grown with DvH as a 315
co-culture, the maximum calculated interspecies distance was large enough (755 μm) to enable 316
interspecies hydrogen transfer between the two bacteria (Supplemental material, Part 4, Table 317
S4), and aggregates were not formed. 318
Strain 195 Transcriptome during syntrophic growth 319
Transcriptomic microarray analysis comparing the co-culture growing on butyrate and 320
strain 195 growing in isolation identified 214 genes that were differentially transcribed (Fig. 4, 321
Table S7). Among these differentially expressed genes, 18 were up-regulated and 196 were 322
down-regulated in the syntrophic co-culture compared to the strain 195 isolate. Among the up-323
regulated genes, most significantly expressed genes (signal level 5000~20,000) belonged to 324
transport and metabolism functions including several types of ABC (ATP binding cassette) 325
transporters. Genes located within an operon of a Fec-type ABC transporter (from DET1173 to 326
DET1176) which are involved in periplasmic iron-binding were up-regulated 2.7 to 4.2 times, 327
respectively; genes (DET1491, DET1493) from a cluster encoding a peptide ABC transporter 328
responsible for ATP binding were up-regulated 2.1 and 2.3 times, respectively; genes 329
(DET0140-0141) encoding a phosphate ABC transporter and ATP-binding protein were up-330
regulated 2.0 to 2.6 times, respectively. There were also a few genes with unknown functions up-331
regulated at high signal levels compared to the isolate. DET1008, with highest homology to a 332
on March 27, 2018 by guest
http://aem.asm
.org/D
ownloaded from

16
gene encoding a cell-division initiation protein (39) was up-regulated 3.2 times and acetyl-CoA 333
synthase (DET1209) was up-regulated 2.4 times at a high signal level (~20,000). 334
Genes associated with membrane-bound oxidoreductase complexes, which are related to 335
energy metabolism, such as RDases, hydrogenases, molybdopterin oxidoreductases, putative 336
formate dehydrogenases, as well as NADH-oxidoreductases, showed no significant differential 337
expression patterns between the co-culture and pure culture throughout the experiment (Table 338
S5). Among the 196 down-regulated genes in the co-culture compared to the isolate, 75 were 339
genes with signal levels lower than 1000 in both treatments (suggesting low expression level), 340
and most of them encode for hypothetical proteins (Table S5). Many of the down-regulated 341
genes were not related to energy metabolism, such as those encoding a phage domain protein 342
(DET0354); mercuric reductase (DET0732); Mg chelatase-like protein (DET0986) and 343
virulence-associated proteinE (DET1098). 344
Discussion 345
In the strain 195 and S. wolfei syntrophic co-culture studied here, strain 195 grew 346
exponentially at the rate of 0.69 d-1
with a doubling time of 1.0 day, calculated from cell numbers 347
(tceA copies) from day 2 to day 6. This doubling time is shorter than previously reported for the 348
isolates of D. mccartyi sp. (31,40-42). The dechlorination rate was 9.9 ± 0.1 µmol d-1
which is 349
similar to the value observed with strain 195 and DvH co-cultures (11.0 ± 0.01 µmol d-1
, (15)) 350
and higher than the dechlorination rates observed with other D. mccartyi-containing co-cultures 351
(4.2-6.8 µmol d-1
) (5,7,14-16). The cell yield of strain 195 in this co-culture (1.1 ± 0.3 × 108 cells 352
µmol-1
Cl- released) was similar to that observed in other D. mccartyi co-culture studies, e.g., 9.0 353
on March 27, 2018 by guest
http://aem.asm
.org/D
ownloaded from

17
± 0.5 ×107 cells µmol
-1 Cl
-1 (15) and 9.0 ± 1.4 ×10
7 cells µmol
-1 Cl
- (7). Given that the minimum 354
free energy change required for ATP synthesis is in the range of 15-25 kJ mol-1
for most 355
syntrophic fermentations (43), only a small amount of energy saved as ATP is expected to be 356
available during S. wolfei syntrophic butyrate fermentation (19). Indeed, S. wolfei in the co-357
culture was growing very near the thermodynamic threshold (delta G from -41.9 to -5.7 kJ mol-1
), 358
and consequently the cell numbers remained at low levels (1:16 ratio with strain 195 cells) when 359
growing on butyrate compared to growth on crotonate. The ratio of calculated free energy 360
available for strain 195 to reduce TCE to VC (-272.0 kJ mol-1
) and S. wolfei (-43.6 kJ mol-1
, 361
Table S2) growing on butyrate was about 6.2:1 at the beginning of the experiment. Considering 362
that the cell size of S. wolfei is about 2.5 times larger than strain 195, the theoretical cell 363
production ratio (based on free energy) of strain 195 to S. wolfei (15.6:1) is similar to our 364
observation (16:1). While growing on crotonate, the ratio of available energy for strain 195 (-365
295.3kJ mol-1
) and S. wolfei (-465.2 kJ mol-1
) was 0.6, and the theoretical cell production ratio of 366
1.2:1 is similar to our observation of 1.3:1 (Supplemental material, Part 1). Roden and Jin (44) 367
found a linear correlation between microbial growth yields (YXS, g cell mol-1
substrate) and 368
estimated catabolic ∆𝐺𝑟 for metabolism of short-chain fatty acids and H2 coupled metabolic 369
pathways. Table 4 compares the growth yields calculated using the Roden and Jin (44) method 370
(YXS,cal) with observed growth yields (YXS,obs) of syntrophs growing in a variety of co-cultures. 371
Our results confirmed that it is possible to estimate syntrophic cell yields by percentage error of 372
less than 100% by applying an empirical equation for the listed syntrophic co-culture studies (44), 373
suggesting that the growth yield of syntrophic bacteria and the ratio maintained in the co-cultures 374
were mainly controlled by thermodynamics. Furthermore, we found H2 concentrations to be the 375
on March 27, 2018 by guest
http://aem.asm
.org/D
ownloaded from

18
key factor affecting the ∆𝐺𝑟 in the co-cultures, which in turn affects predictions of microbial 376
growth yields (calculation in Tables S2 and S6). 377
When TCE was supplied in excess to the co-culture (on a H2 production/consumption 378
basis), the H2 level dropped to the threshold concentration of 0.6 ± 0.1 nM and dechlorination 379
ceased. Although the calculated ∆𝐺𝑟available for dechlorination by strain 195 was still negative 380
(-145.5 kJ mol-1
) at this concentration (Supplemental material, Part 5, Table S6), it did not 381
support growth. Thermodynamic calculations indicate that hydrogen concentrations would have 382
to reach liquid concentrations of 10-27
nM to drive the ∆𝐺𝑟 for strain 195 positive, a value that is 383
unrealistic in the environment (45). Therefore, the minimum H2 threshold for strain 195 cells is 384
likely not based on thermodynamics, but is rather based upon other factors such as enzyme 385
binding affinity and specificity (46). This finding is consistent with previously published H2 386
thresholds for dechlorination by Dehalcoccoides-containing communities (10,42), and falls in the 387
same range as thresholds for sulfate reduction (47). Other dechlorinating isolates have been 388
reported to have minimum H2 thresholds around ~0.04-0.3 nM (11,47,48). In previous studies of 389
S. wolfei growing with H2-oxidizing hydrogenotrophic methanogens, sulfate reducers, and nitrate 390
reducers, the estimated energy available for S. wolfei ranged from 0.9 to 15.0 kJ mol-1
(18) when 391
butyrate fermentation ceased and the ratio of acetate to butyrate was around 900. While in this 392
study we observed estimated energy available for S. wolfei was -13.7 ± 0.2 kJ mol-1
with the 393
acetate to butyrate ratio of 0.4:1. Previous studies have shown calculated ∆𝐺𝑟 value fell in the 394
range (-10 to -15 kJ mol-1
) observed in H2-dependent terminal electron accepting processes under 395
starvation conditions in sulfate reducing bacteria and methanogenic archaea (49,50,63). 396
Furthermore, the value obtained in this study was close to the average threshold value of butyrate 397
on March 27, 2018 by guest
http://aem.asm
.org/D
ownloaded from

19
metabolism (-13.8 ± 1.2 kJ mol-1
) reported in co-culture S. aciditrophicus and Desulfovibrio 398
strain G11 at different acetate to butyrate ratios by Jackson and McInerney (51). 399
It is interesting that the amount of butyrate in the bottles continued to decrease slightly 400
(36 µmol) after TCE was depleted and ethene was being produced from VC (from day 8 to day 401
18). According to thermodynamics, microbial metabolism ceases when the ∆𝐺𝑟 available from a 402
reaction becomes positive. At the beginning of the experiment, the ∆𝐺𝑟 available for S. wolfei 403
butyrate fermentation was negative (-41.9 ± 1.5 KJ mol-1
), and on day 6, this number increased 404
to -21.7 ± 0.3 kJ mol-1
, then on day 8, it increased further to -13.7 ± 0.2 kJ mol-1
(Supplemental 405
material, Fig. S3), which is less than the minimum energy (-20 kJ mol-1
) required by a bacterium 406
to synthesize ATP (34). During this time, the cell numbers decreased indicating cell death and 407
lysis. It has been reported that butyrate fermentation by S. wolfei stops when aqueous H2 408
concentrations reach 480 nM (33). In this work, the H2 concentration was 350 ± 20 nM in the co-409
culture on day 7 when all TCE was reduced to VC. However, the concentration of H2 steadily 410
increased to 1,200 ± 300 nM by the end of the experiment on day 18, indicating that additional 411
butyrate fermentation occurred although cell growth had ceased, suggesting that regulation of the 412
fermentation enzymes may not be stringent. During active dechlorination, H2 concentrations 413
remained at levels below 200 nM (Fig. 1b) indicating that the H2 generation rate was at about the 414
same level as the consumption rate (Fig. 1d) and that the growth rate of the two species was 415
strictly coupled by hydrogen transfer. 416
In strain 195, acetyl-CoA is cleaved in an incomplete Wood-Ljungdahl pathway to 417
provide the methyl group for methionine biosynthesis, whereby carbon monoxide (CO) is 418
produced as a byproduct that accumulates and eventually inhibits D. mccartyi growth and 419
dechlorination (17,52). However, in microbial communities, CO can serve as an energy source 420
on March 27, 2018 by guest
http://aem.asm
.org/D
ownloaded from

20
for many anaerobic microorganisms (53). A previous study showed a high CO concentration 421
(15%) to have an inhibitory effect on S. wolfei (Sieber et al. 2014). Here we found that not only 422
did low levels of CO not exhibit inhibitory effects on S. wolfei cell growth, but S. wolfei could 423
consume CO (µmol per bottle) while growing on crotonate. CO levels in the co-culture growing 424
on butyrate were maintained at low levels during the feeding cycle rather than accumulating as it 425
does with the 195 isolate. Therefore it is possible that CO serves as a supplemental energy source 426
for S. wolfei during syntrophic fermentation with strain 195. This finding is interesting since 427
there were no anaerobic carbon monoxide dehydrogenase genes annotated in the S. wolfei 428
genome (19). However Swol_1136 and Swol_1818, which were originally annotated as iron-429
sulfur cluster binding domain-containing protein and 2Fe-2S binding protein in S. wolfei, were 430
more recently annotated in the KEGG database (54) as carbon-monoxide dehydrogenase small 431
sub-units for CO conversion to CO2, suggesting the genomic potential for CO metabolic ability 432
in S. wolfei. 433
Formation of cell aggregates has been shown to be a distinctive feature of obligate 434
syntrophic communities containing acetogenic bacteria and methanogenic archaea (20). 435
Clustering of cells with decreasing inter-microbial distances leads to increased fluxes and 436
increased specific growth rates (55). Previous studies reported that the syntrophic co-culture of a 437
propionate degrader and a hydrogenotrophic methanogen could form cell aggregates while 438
growing on propionate for optimal hydrogen transfer (27,56). A similar phenomenon was also 439
observed in mixed cultures containing propionate degraders and methanogens enriched from an 440
anaerobic bio-waste digester where flocs formed showing that reducing the interspecies distances 441
by aggregation was advantageous in complex ecosystems (57). In engineered dechlorinating 442
systems, D. mccartyi species have been observed in biofilms within a membrane bioreactor (58) 443
on March 27, 2018 by guest
http://aem.asm
.org/D
ownloaded from

21
and in bioflocs maintained in a continuous flow bio-reactor fed with butyrate (13). Based on 444
morphological observation in this study, strain 195 cells grown in isolation are more likely to 445
grow in planktonic form and when grown with DvH as a syntrophic co-culture on lactate, no 446
significant aggregates were observed. However when grown with S. wolfei on butyrate, the co-447
cultures formed aggregates during syntrophic growth. Thermodynamic calculations show that 448
syntrophic butyrate oxidation is endergonic unless the circumstantial H2 partial pressure is 449
maintained very low (Table 1 and Supplemental material, Table S2). Furthermore fdh 450
dehydrogenase expression in S. wolfei under different growth conditions confirmed that these 451
genes were not up-regulated in in the co-culture grown on butyrate compared to growth on 452
crotonate, or in isolation (38). This is the first study to report D. mccartyi forming cell aggregates 453
with a syntrophic partner. In addition, because the average inter-microbial distances are smaller 454
for syntrophic aggregates, the H2 and metabolite flux between cells would be expected to 455
increase, leading to higher material transfer efficiencies, which could partly explain the observed 456
increased cell yields of strain 195. Another reason for the observed increased cell yields of strain 457
195 is likely due to the continuous removal of CO, which was shown to exert an inhibitory effect 458
on D.mccartyi growth (17). 459
In a previous study of Methanococcus maripaludis growing in syntrophic association 460
with Desulfovibrio vulgaris under hydrogen limitation in chemostats (59), M. maripaludis was 461
growing close to the thermodynamic threshold for growth, and the acetyl-CoA synthase 462
transcripts levels of M. maripaludis in the co-culture decreased compared to the isolate. While in 463
our study, the acetyl-CoA synthase transcripts levels of strain 195 in the co-culture increased 464
compared to the isolate. A recent study (60) showed a division of metabolic labor and mutualistic 465
on March 27, 2018 by guest
http://aem.asm
.org/D
ownloaded from

22
interactions were the reason for increased rates of growth in a vast majority of cross-feeding 466
strains. The benefit of cross-feeding was larger than the cost (60, 64). 467
Acknowledgements 468
We Acknowledge Professor Michael McInerney at University of Oklahoma kindly provided us 469
the pure strain Syntrophomonas wolfei for this study. We also thank Dr. Nikhil S. Malvankar and 470
Professor Derek Lovley at University of Massachusetts carried out the experiments of 471
conductivity test for the co-cultures. This work was funded through research grants from NIEHS 472
(P42-ES04705-14) and NSF (CBET-1336709) 473
474
475
476
Reference 477
478
1. MaymóGatell X, Chien YT, Gossett JM, Zder SH. 1997. Isolation of a bacterium that 479
reductively dechlorinates tetrachloroethene to ethene. Science 276 (5318): 1568-1571. 480
2. McCarty PL. 2010. Groundwater contamination by chlorinated solvents: History, 481
Remediation Technologies and Strategies. p 1-24. In Stroo HF, Ward CH. (ed), In Situ 482
Remediation of Chlorinated Solvent Plumes. Chapter 1. Springer Science Publishing, 483
New York City, NY. 484
3. Lu X, Wilson JT, Kampbell DH. 2006. Relationship between Dehalococcoides DNA in 485
ground water and rates of reductive dechlorination at field scale. Water Research 40 (16): 486
on March 27, 2018 by guest
http://aem.asm
.org/D
ownloaded from

23
3131-3140. 487
4. Lee PKH, Warnecke F, Brodie EL, Macbeth TW, Conrad ME, Andersen GL, 488
Alvarez-Cohen L. 2012. Phylogenetic microarray analysis of a microbial community 489
performing reductive dechlorination at a TCE-contaminated site. Envion Sci Technol 46 490
(2): 1044-1054. 491
5. He J, Holmes VF, Lee PK, Alvarez-Cohen L. 2007. Influence of vitamin B-12 and 492
cocultures on the growth of Dehalococcoides isolates in defined medium, Appl Environ 493
Microbiol 73 (9): 2847-2853. 494
6. Men YJ, Seth EC, Yi S, Allen, RH, TAga ME, Alvarez-Cohen L. 2014. Sustainable 495
growth of Dehalococcoides mccartyi 195 by corrinoid salvaging and remodeling in 496
defined lactate-fermenting consortia. Appl Environ Microbiol 80 (7): 2133-2141. 497
7. Yan J, Im J, Yang Y, Löffler FE. 2013. Guided cobalamin biosynthesis supports 498
Dehalococcoides mccartyi reductive dechlorination activity. Philos Trans R Soc Lond B 499
Biol Sci 368 (1616): 1-23. 500
8. Schipp CJ, Marco-Urrea E, Kublik A, Seifert J, Adrian L. 2013. Organic cofactors in 501
the metabolism of Dehalococcoides mccartyi strains. Philos Trans R Soc Lond B Biol Sci 502
368 (1616): 1-12. 503
9. Ziv-El, M, Delgado AG, Yao Y, Kang D, Nelson KG, Halden RU, Krajmalnik-504
Brown R. 2011. Development and characterization of DehaloR^2, a novel anaerobic 505
microbial consortium performing rapid dechlorination of TCE to ethene. Appl Microbiol 506
Biotechnol 92 (5): 1063-1071. 507
10. Yang YR, McCarty PL. 1998. Competition for hydrogen within a chlorinated solvent 508
dehalogenating anaerobic mixed culture, Environ Sci Technol 32 (2): 3591-3597. 509
on March 27, 2018 by guest
http://aem.asm
.org/D
ownloaded from

24
11. Löffler FE, Tiedje JM, Sanford RA. 1999. Fraction of electrons consumed in electron 510
acceptor reduction and hydrogen thresholds as indicators of halorespiratory physiology. 511
Appl Environ Microbiol 65 (9): 4049-4056. 512
12. Duhamel M, Wehr SD, Yu L, Rizvi H, Seepersad D, Dworatzed S, Cox EE, 513
Edwards EA. 2002. Comparison of anaerobic dechlorinating enrichment cultures 514
maintained on tetrachloroethene, trichloroethene, cis-dichloroethene and vinyl chloride. 515
Water Research 36 (17): 4193-4202. 516
13. Rowe AR, Lazar BJ, Morris RM, Richardson RE. 2008. Characterization of the 517
community structure of a dechlorinating mixed culture and comparisons of gene 518
expression in planktonic and biofloc-Associated “Dehalococcoides” and 519
Methanospirillum Species. Appl Environ Microbiol 74 (21): 6709-6719. 520
14. Cheng D, Chow WL, He J. 2010, A Dehalococcoides-containing co-culture that 521
dechlorinates tetrachloroethene to trans-1,2-dichloroethene. ISME Journal 4 (1): 88-97. 522
15. Men YJ, Feil H, Verberkmoes NC, Shah MB, Johnson DR, Lee PK, West 523
KA, Zinder SH, Andersen GL, Alvarez-Cohen L. 2011. Sustainable syntrophic growth 524
of Dehalococcoides ethenogenes strain 195 with Desulfovibrio vulgaris Hildenborough 525
and Methanobacterium congolense: global transcriptomic and proteomic analyses. ISME 526
J 6 (2): 410-421. 527
16. Yan J, Ritalahti KM, Wagner DD, Löffler FE. 2012. Unexpected specificity of 528
interspecies cobamide transfer from Geobacter spp. to organohalide-respiring 529
Dehalococcoides mccartyi strains. Appl Environ Microbiol 78 (18):6630-6636 . 530
17. Zhuang WQ, Yi S, Bill M, Brisson VL, Feng X, Men Y, Conrad ME, Tang YJ, 531
Alvarez-Cohen L. 2014. Incomplete Wood-Ljungdahl pathway facilitates one carbon 532
on March 27, 2018 by guest
http://aem.asm
.org/D
ownloaded from

25
metabolism in organohalide-respiring Dehalococcoides mccartyi. PNAS 111 (17): 6419-533
6424. 534
18. Jin Q. 2007. Control of hydrogen partial pressures on the rates of syntrophic microbial 535
metabolisms: a kinetic model for butyrate fermentation. Geobiology 5 (1): 35-48. 536
19. Sieber JR, Sims DR, Han C, Kim E, Lykidis A, Lapidus AL, McDonnald E, Rohlin 537
L, Culley DE, Gunsalus R, McInerney MJ. 2010. The genome of Syntrophomonas 538
wolfei: new insights into syntrophic metabolism and biohydrogen production. Environ 539
Microbiol 12 (8): 2289-2301. 540
20. Stams AJM, Worm P, Sousa DZ, Alves MM, Plugge CM. 2012. Syntrophic 541
Degradation of Fatty Acids by Methanogenic Communities, p.127-142. In Hallenbeck 542
P.C. (ed), Microbial Technologies in Advanced Biofuels Production. Wageningen, The 543
Netherlands. 544
21. McInerney MJ, Bryant MP, Hespell RB, Costerton JW. 1981. Syntrophomonas wolfei 545
gen. nov. sp. nov., an anaerobic, syntrophic, fatty acid-oxidizing bacterium. Appl. 546
Environ Microbiol 41(4): 1029-1039. 547
22. Beaty PS, McInerney MJ. 1987. Growth of Syntrophomonas wolfei in pure culture on 548
crotonate. Arch Microbiol 147 (4): 389-393. 549
23. Freeborn RA, West KA, Bhupathiraju VK, Chauhan S, Rahm BG, Richardson RE, 550
Alvarez-Cohen L. 2005. Phylogenetic analysis of TCE-dechlorinating consortia enriched 551
on a variety of electron donors. Environ Sci Technol 39(21): 8358-8368. 552
24. Lee PKH, Johnson DR, Holmes VF, He J, Alvarez-Cohen L. 2006. Reductive 553
dehalogenase gene expression as a biomarker for physiological activity 554
of Dehalococcoides spp. Appl Environ Microbiol 72 (9): 6161-6168. 555
on March 27, 2018 by guest
http://aem.asm
.org/D
ownloaded from

26
25. Gorby YA, Yanina S, McLean JS, Rosso KM, Moyles D, Dohnalkova A, Beveridge 556
TJ, Chang IS, Kim BH, Kim KS, Culley DE, Reed SB, Romine MF, Saffarini DA, 557
Hill EA, Shi L, Elias DA, Kennedy DW, Pinchuk G, Watanabe K, Ishii S, Logan B, 558
Nealson KH, Fredrickson JK. 2006. Electrically conductive bacterial nanowires 559
produced by Shewanella oneidensis strain MR-1 and other microorganisms. PNAS 103 560
(30): 9536-9542. 561
26. Dykstra M J, Reuss LE. 1992. Biological Electron Microscopy- Theory, Techniques, 562
and Troubleshooting. Chapter 9. p 223-245. Scanning Electron Microscopy 2nd
ed, ASM 563
Press, Washington DC. 564
27. Ishii S, Kosaka T, Hori K, Hotta Y, Watanabe K. 2005. Coaggregation Facilitates 565
Interspecies Hydrogen Transfer between Pelotomaculum thermopropionicum and 566
Methanothermobacter thermautotrophicus, Appl Environ Microbiol 71 (12):7828-7845. 567
28. Haynes WM. 2013. CRC Handbook of Physics and Chemistry, 95th
edition ISBN 568
9781482208672 569
29. Johnson DR, Lee PKH, Holmes VF, Alvarez-Cohen L. 2005. An internal reference 570
technique for accurately quantifying specific mRNAs by real-time PCR with application 571
to the tceA reductive Dehalogenase gene. Appl Environ Microbiol 71 (7): 3866-3871. 572
30. Duhamel M, Edwards EA. 2007. Growth and yields of dechlorinators, acetogens, and 573
methanogens during reductive dechlorination of chlorinated ethenes and 574
dihaloelimination of 1,2-dichloroethane. Environ Sci Technol 41(7):2303-2310. 575
31. Johnson DR, Brodie EL, Hubbard, Alan E, Andersen GL, Zinder SH, Alvarez-576
Cohen L. 2008. Temporal Transcriptomic Microarray Analysis of Dehalococcoides 577
ethenogenes Strain 195 during the Transition into Stationary Phase. Appl Environ 578
on March 27, 2018 by guest
http://aem.asm
.org/D
ownloaded from

27
Microbiol 74 (9): 2864-2872. 579
32. West KA, Lee PKH, Johnson DR, Zinder SH, Alvarez-Cohen L. 2013. Global gene 580
expression of Dehalococcoides within a robust dynamic TCE-dechlorinating community 581
under conditions of periodic substrate supply. Biotechnol and Bioeng 110 (5): 1333-1341. 582
33. Wallrabenstein C, Schink B. 1994. Evidence of reversed electron transport in 583
syntrophic butyrate and benzoate oxidation by Syntrophomonas wolfei and Syntrophus 584
buswellii. Arch Microbiol 162 (1):136-142. 585
34. Schink B. 1997. Energetics of syntrophic cooperation in methanogenic degradation. 586
Microbiol. Mol Biol Rev 61 (2): 262-280. 587
35. Cord-Ruwisch R, Seitz HJ, Conrad R. 1988. The capacity of hydrogenotrophic 588
anaerobic bacteria to compete for traces of hydrogen depends on the redox potential of 589
the terminal electron acceptor. Arch Microbiol 149(4): 350-357. 590
36. Shrestha PM, Rotaru AE, Aklujkar M, Liu F, Shrestha M, Summers ZM, 591
Malvankar N, Flores DC, Lovley DR. 2013. Syntrophic growth with direct interspecies 592
electron transfer as the primary mechanism for energy exchange. Environ Microbiol Rep 593
5 (6): 904-910. 594
37. Sieber JR, Le HM, McInerney MJ. 2014. The importance of hydrogen and formate 595
transfer for syntrophic fatty, aromatic and alicyclic metabolism. Environ Microbiol 16 (1): 596
177-188. 597
38. Crable B R. 2013. Ph.D dissertation. Enzyme systems involved in interspecies hydrogen 598
and formate transfer between syntrophic fatty and aromatic acid degraders and 599
methanospirillum hungatei. University of Oklahoma. 600
39. Ueda K, Yamashita A, Ishikawa J, Shimada M, Watsuji TO, Morimura K, Ikeda H, 601
on March 27, 2018 by guest
http://aem.asm
.org/D
ownloaded from

28
Hattori M, Beppu T. 2004. Genome sequence of Symbiobacterium thermophilum, an 602
uncultivable bacterium that depends on microbial commensalism. Nucl Acids Res 32 (16): 603
4937-4944. 604
40. He J, Sung Y, Krajmalnik-Brown R, Rittalahti KM, Löffler FE. 2005. Isolation and 605
characterization of Dehalococcoides sp. strain FL2, a trichloroethene (TCE)-and 1,2-606
dichlroethene-respiring anaerobe. Environ Microbiol 7(9): 1442-1450. 607
41. Sung Y, Ritalahti KM, Apkarian RP, Löffler FE. 2006. Quantitative PCR confirms 608
purity of strain GT, a novel trichloroethene-to-ethene-respiring Dehalococcoides isolate. 609
Appl Environ Microbiol 72 (3): 1980-1987. 610
42. Löffler FE, Yan J, Ritalahti KM, Adrian L, Edwards EA, Konstantinidis KT, 611
Muller JA, Fullerton H, Zinder SH, Spormann AM. 2013. Dehalococcoides mccartyi 612
gen. nov., sp. nov., obligately organohalide-respiring anaerobic bacteria relevant to 613
halogen cycling and bioremediation, belong to a novel bacterial class, Dehalococcoidia 614
classis nov., order Dehalococcoidales ord. nov. and family Dehalococcoidaceae fam. 615
nov., within the phylum Chloroflexi. Int J Syst Evol Microbiol 63(Pt 2):625-635. 616
43. Stams AJM. and Plugge CM. 2009. Electron transfer in syntrophic communities of 617
anaerobic bacteria and archaea. Nat Rev Microbiol 7 (8): 568-577. 618
44. Roden EE, Jin Q. 2011. Thermodynamics of Microbial Growth Coupled to Metabolism 619
of Glucose, Ethanol, Short-Chain Organic Acids, and Hydrogen. Appl Environ Microbiol 620
77 (5): 1907-1909. 621
45. Heimann AC, Jakobsen R. 2006. Experimental evidence for a lack of thermodynamic 622
control on hydrogen concentrations during anaerobic degradation of chlorinated ethenes. 623
Environ Sci Technol 40(11) : 3501-3507. 624
on March 27, 2018 by guest
http://aem.asm
.org/D
ownloaded from

29
46. Dolfing, J. 2003. Thermodynamic considerations for dehalogenation. p. 89-114. In 625
Häggblom, M. M., Bossert, I.D., (ed), Dehalogenation: Microbial Processes and 626
Environmental Applications.Kluwer Academic Publishers: Boston/Dordrecht/London 627
47. Luijten ML, Roelofsen W, Langenhoff AA, Schraa G, Stams AJ. 2004. Hydrogen 628
threshold concentrations in pure cultures of halorespiring bacteria and at a site polluted 629
with chlorinated ethenes. Environ Microbiol 6 (6): 646-650. 630
48. Lu XX, Tao S, Bosma T, Gerritse J. 2001. Characteristic hydrogen concentrations for 631
various redox processes in batch study. J Environ Sci Health A Tox Hazard Subst 632
Environ Eng 36 (9): 1725-1734. 633
49. Hoehler TM, Alperin MJ, Alber DB, Martens CS. 2001. Apparent minimum free 634
energy requirements for methanogenic Archaea and sulfate-reducing bacteria in an 635
anoxic marine sediment. FEMS Microbiol Ecol 38 (1): 33-41. 636
50. Hoehler TM. 2004. Biological energy requirements as quantitative boundary conditions 637
for life in the subsurface. Geobiology 2 (4): 205-215. 638
51. Jackson BE, McInerney M J. 2002. Anaerobic microbial metabolism can proceed close 639
to therymodynamic limits. Nature 415 (6870): 454-456. 640
52. Tang YJ, Yi S, Zhuang W, Zinder SH, Keasling JD, Alvarez-Cohen L. 2009. 641
Investigation of Carbon Metabolism in "Dehalococcoides ethenogenes" Strain 195 by 642
Use of Isotopomer and Transcriptomic Analyses. J Bacteriol 191(16): 5224-5231. 643
53. Oelgeschläger E and Rother M. 2008. Carbon monoxide-dependent energy metabolism 644
in anaerobic bacteria and archaea. Arch Microbiol 190 (3): 257-269. 645
54. Kanehisa M, Goto S, Sato Y, Kawashima M, Furumichi M, Tanabe M. 2014. Data, 646
information, knowledge and principle: back to metabolism in KEGG. Nucleic Acids Res 647
on March 27, 2018 by guest
http://aem.asm
.org/D
ownloaded from

30
42 (1) : 199–205. 648
55. Schink B, Thauer RK. 1988. Energetics of syntrophic methane formation and the 649
influence of aggregation, p. 5-17. In Lettinga G, Zehnder A. J. B., Grotenhuis J. T. C., 650
and Hulshoff L. W. (ed), Granular Anaerobic Sludge: Microbiology and Technology. 651
Pudoc. Wageningen, The Netherlands. 652
56. Ishii S, Kosaka T, Hotta Y, Watanabe K. 2006. Simulating the contribution of 653
coaggregation to interspecies hydrogen fluxes in syntrophic methanogenic consortia. 654
Appl Environ Microbiol 72 (7): 5093-5096. 655
57. Felchner-Zwirello M, Winter J, Gallert C. 2013. Interspecies distances between 656
propionic acid degraders and methanogens in syntrophic consortia for optimal hydrogen 657
transfer. Appl Microbiol Biotechnol 97 (20): 9193-9205. 658
58. Chung J, Krajmalnik-Brown R, Rittmann BE. 2008. Bioreduction of trichloroethene 659
using a hydrogen-based membrane biofilm reactor. Environ Sci Technol 42 (2): 477-483. 660
59. Walker CB, Redding-Johanson AM, Baidoo EE, Rajeev L, He Z, Hendrickson EL, 661
Joachimiak MP, Stolyar S, Arkin AP, Leigh JA, Zhou J, Keasling JD, 662
Mukhopadhyay A, Stah DA. 2012. Functional responses of methanogenic archaea to 663
syntrophic growth. ISME J 6 (11): 2045-2055. 664
60. Pande S, Merker H, Bohl K, Reichelt M, Schuster S, de Figueiredo LF, Kaleta C 665
and Kost C. 2014. Fitness and stability of obligate cross-feeding interactions that emerge 666
upon gene loss in bacteria. The ISME J 8 (5): 953-962. 667
61. Traore, A. S., M. L. Fardeau, C. E. Hatchikian, J. Legall, and J. P. Belaich. 1983. 668
Energetics of growth of a defined mixed culture of Desulfovibrio vulgaris and 669
Methanosarcina barkeri - Interspecies hydrogen transfer in batch and continuous cultures. 670
on March 27, 2018 by guest
http://aem.asm
.org/D
ownloaded from

31
Appl. Environ Microbiol 46 (5):1152-1156. 671
62. Beaty PS, McInerney MJ. 1989. Effects of organic-acid anions on the growth and 672
metabolism of Syntrophomonas-wolfei in pure culture and in defined consortia. Appl 673
Environ Microbiol 55 (4): 977-983. 674
63. Heimann A, Jakobsen R, Blodau C. 2010. Energetic constraints on H2-dependent 675
terminal electron accepting processes in anoxic environments: a review of observations 676
and model approaches. Environ Sci Technol 44 (1): 24-33. 677
64. Mee MT, Collins JJ, Church GM, Wang HH. 2014. Syntrophic exchange in synthetic 678
microbial communities. PNAS 111 (20): 2149-2156. 679
680
on March 27, 2018 by guest
http://aem.asm
.org/D
ownloaded from

0 2 4 6 8 10 12 14 16 180
20
40
60
80
100
Time (day)
Eth
enes
(µ
mol/bottle
)
a
0 2 4 6 8 10 12 14106
107
108
109
1010
1011
Time (day)
cell n
um
ber
/bottle
b
0 100 200 3000
50
100
150
200
250
butyrate consumed in H2 equivalent (mmol)Chlo
rinat
ed e
then
es r
educe
d in H
2 e
quiv
alen
t (m
mol)
d fe=0.84, R2=0.9772
0 2 4 6 8 10 12 14 16 180
100
200
300
400
500
600
0
1
2
3
4
5
Time (day)
Fat
ty a
cids
(µm
ol/bottle
)
c
Liq
uid
H2 co
ncen
tration (µ
M)
on March 27, 2018 by guest
http://aem.asm
.org/D
ownloaded from
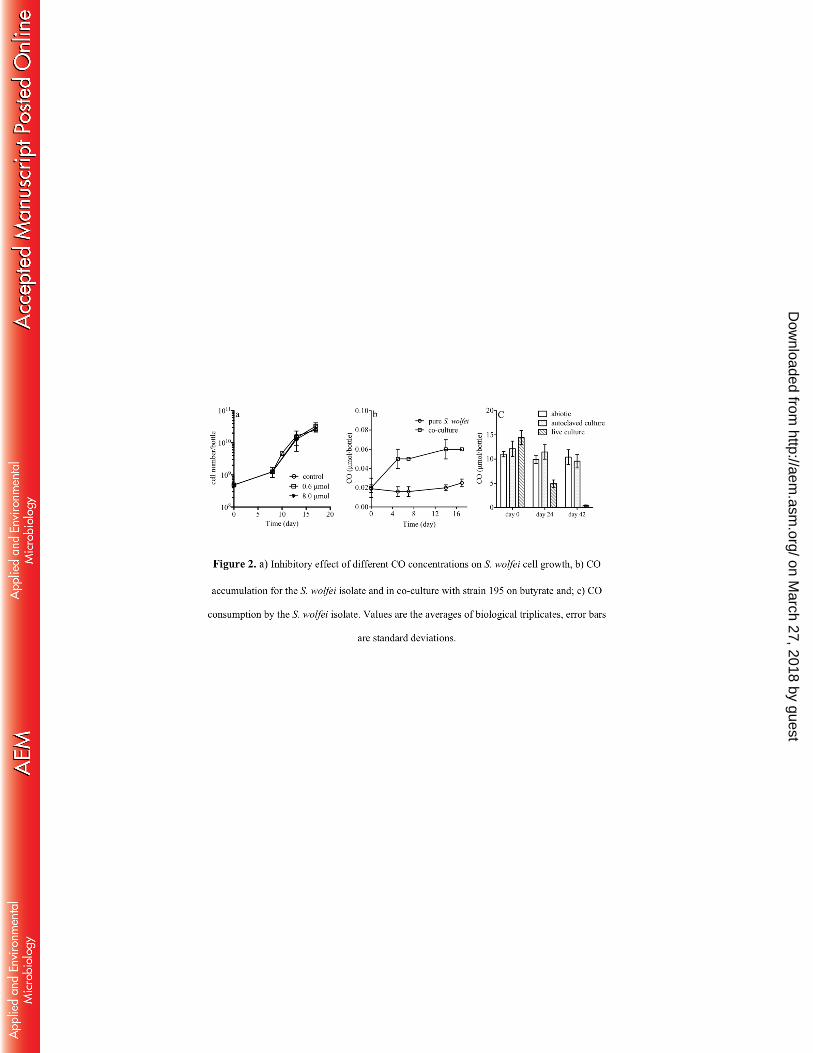

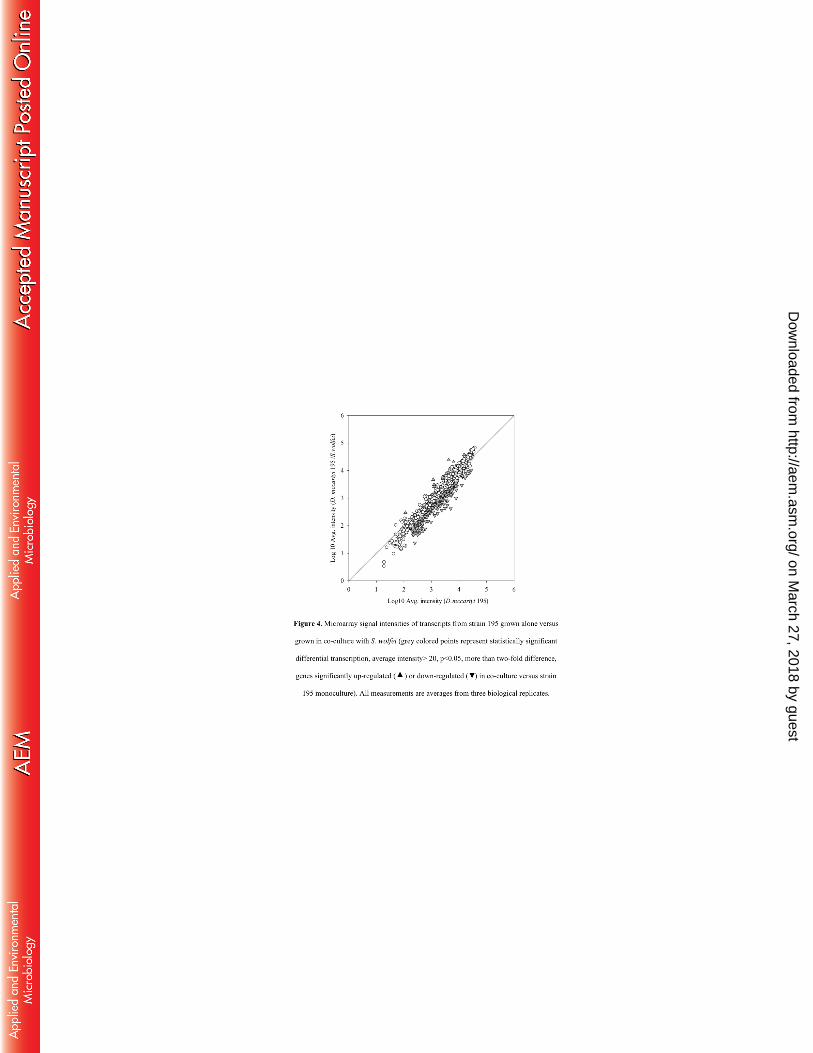
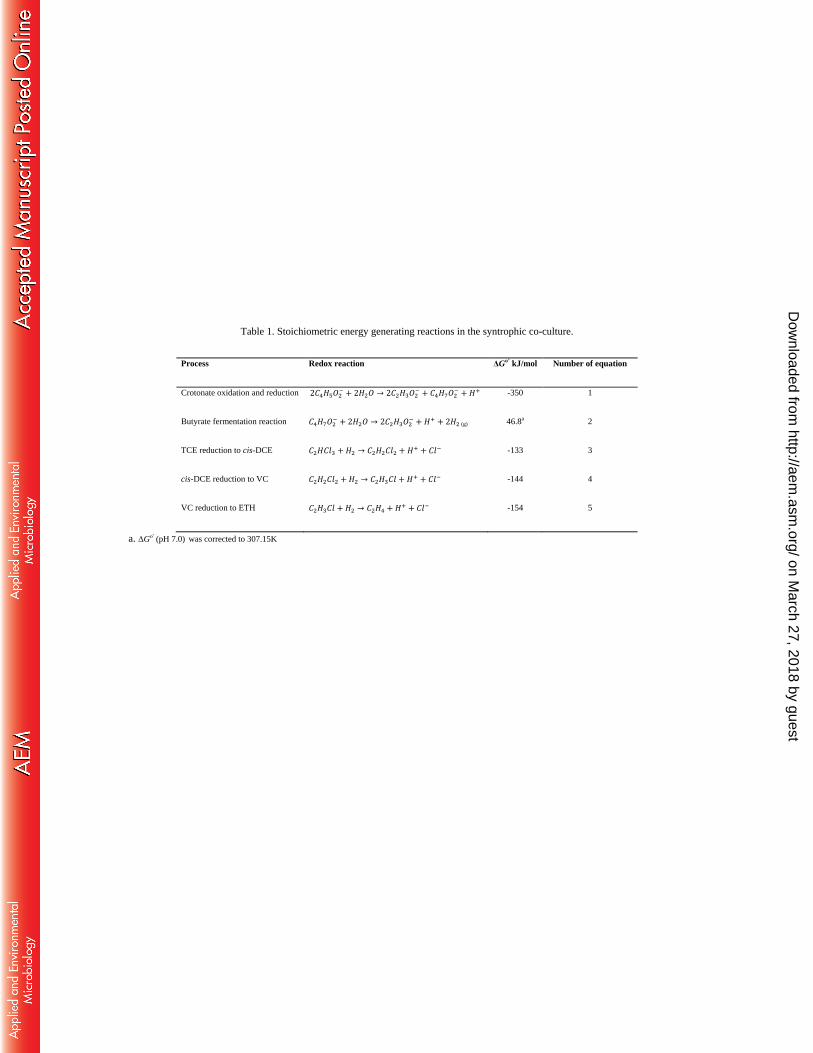
Table 1. Stoichiometric energy generating reactions in the syntrophic co-culture.
Process Redox reaction ΔGo’
kJ/mol Number of equation
Crotonate oxidation and reduction 2𝐶4𝐻5𝑂2− + 2𝐻2𝑂 → 2𝐶2𝐻3𝑂2
− + 𝐶4𝐻7𝑂2− + 𝐻+ -350 1
Butyrate fermentation reaction 𝐶4𝐻7𝑂2− + 2𝐻2𝑂 → 2𝐶2𝐻3𝑂2
− + 𝐻+ + 2𝐻2 (g) 46.8a 2
TCE reduction to cis-DCE 𝐶2𝐻𝐶𝑙3 + 𝐻2 → 𝐶2𝐻2𝐶𝑙2 + 𝐻+ + 𝐶𝑙− -133 3
cis-DCE reduction to VC 𝐶2𝐻2𝐶𝑙2 + 𝐻2 → 𝐶2𝐻3𝐶𝑙 + 𝐻+ + 𝐶𝑙− -144 4
VC reduction to ETH 𝐶2𝐻3𝐶𝑙 + 𝐻2 → 𝐶2𝐻4 + 𝐻+ + 𝐶𝑙− -154 5
a. ΔGo’ (pH 7.0) was corrected to 307.15K
on March 27, 2018 by guest
http://aem.asm
.org/D
ownloaded from


Table 2. Dechlorination rate and cell yield of D. mccartyi strains in various co-culture studies
Co-culturesa Dechlorination
rate
(µmol d-1)
Specific dechlorination
rate of strain 195
(µmol cell-1 d-1)
D. maccartyi cell yield
(cells µmol-1 Cl- released)
Reference
Strain 195 and Desulfovibrio desulfuricans b 0.4 2.3 × 10-10 2.4 × 108 5
Strain 195 and Sedimentibacter sp. c 6.8 N.A. 4.2 × 108 14
Strain 195 and Desulfovibrio vulgaris Hildenborough 11 ± 0.01 N.A. 9.0 ± 0.5 × 107 15
Strain BAV1 and Geobacter lovleyi d
Strain FL2 and Geobacter lovleyi
6.1
4.2
7.9 × 10-10
1.5 × 10-9
6.7 × 107
3.3 × 107
16
Strain BAV1 and Sporomusa sp. strain KB1 e
Strain GT and Sporomusa sp. strain KB1
Strain FL2 and Sporomusa sp. strain KB1
3.3
2.1
2.5
2.7 × 10-10
1.4 × 10-10
7.2 × 10-10
2.1 ± 0.2 × 108
2.1 ± 0.3 × 108
9.0 ± 1.4 × 107
7
Strain 195 and S. wolfei 9.9 ± 0.1 7.6 ×10-10 1.1 ± 0.3 ×108 This study
on March 27, 2018 by guest
http://aem.asm
.org/D
ownloaded from

aDehalococcoides mccartyi strain 195 (strain 195), Dehalococcoides mccartyi strain BAV1 (strain BAV1), Dehalococcoides mccartyi strain GT
(strain GT), Dehalococcoides mccartyi strain FL2 (strain FL2). b Calculated from Fig. 4B and Table 2 therein.
c Calculated from Fig. 1C and Fig. 3 therein.
d Calculated from Fig. 2 and Table 1 therein.
e Calculated from Fig. 6 and Table 1 therein.
on March 27, 2018 by guest
http://aem.asm
.org/D
ownloaded from
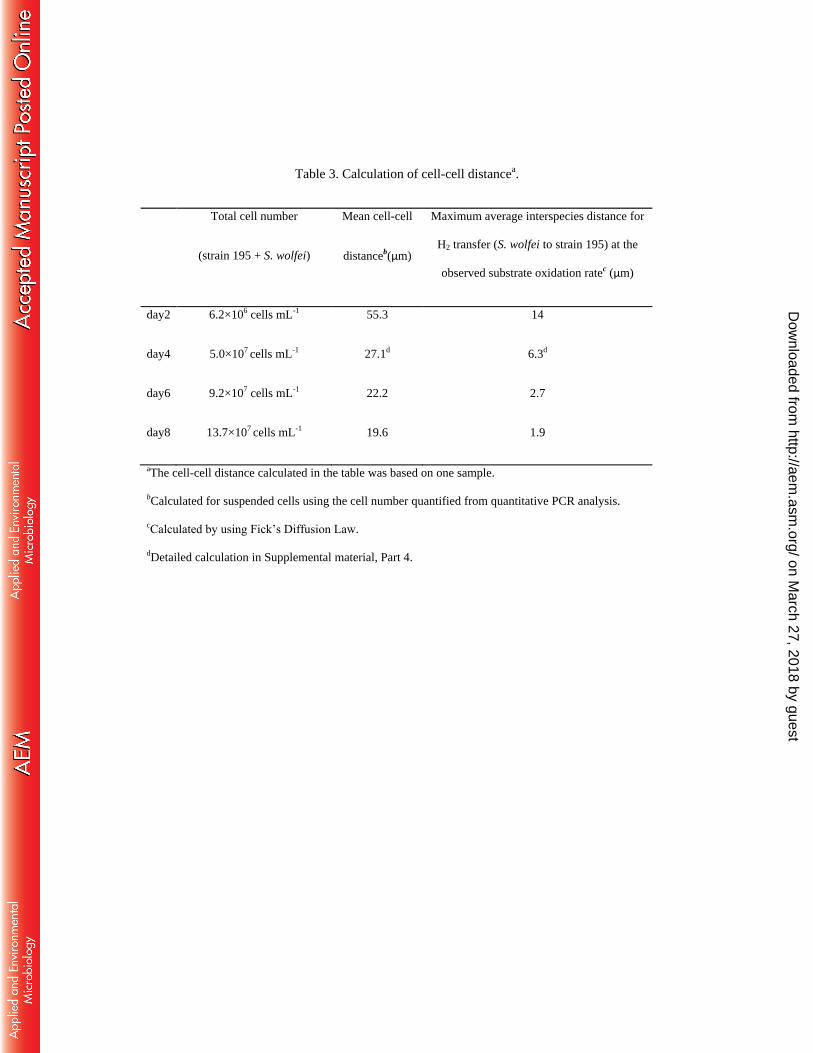
Table 3. Calculation of cell-cell distancea.
Total cell number
(strain 195 + S. wolfei)
Mean cell-cell
distanceb(µm)
Maximum average interspecies distance for
H2 transfer (S. wolfei to strain 195) at the
observed substrate oxidation ratec (µm)
day2 6.2×106 cells mL-1 55.3 14
day4 5.0×107 cells mL-1 27.1d 6.3d
day6 9.2×107 cells mL-1 22.2 2.7
day8 13.7×107 cells mL-1 19.6 1.9
aThe cell-cell distance calculated in the table was based on one sample.
bCalculated for suspended cells using the cell number quantified from quantitative PCR analysis.
cCalculated by using Fick’s Diffusion Law.
dDetailed calculation in Supplemental material, Part 4.
on March 27, 2018 by guest
http://aem.asm
.org/D
ownloaded from
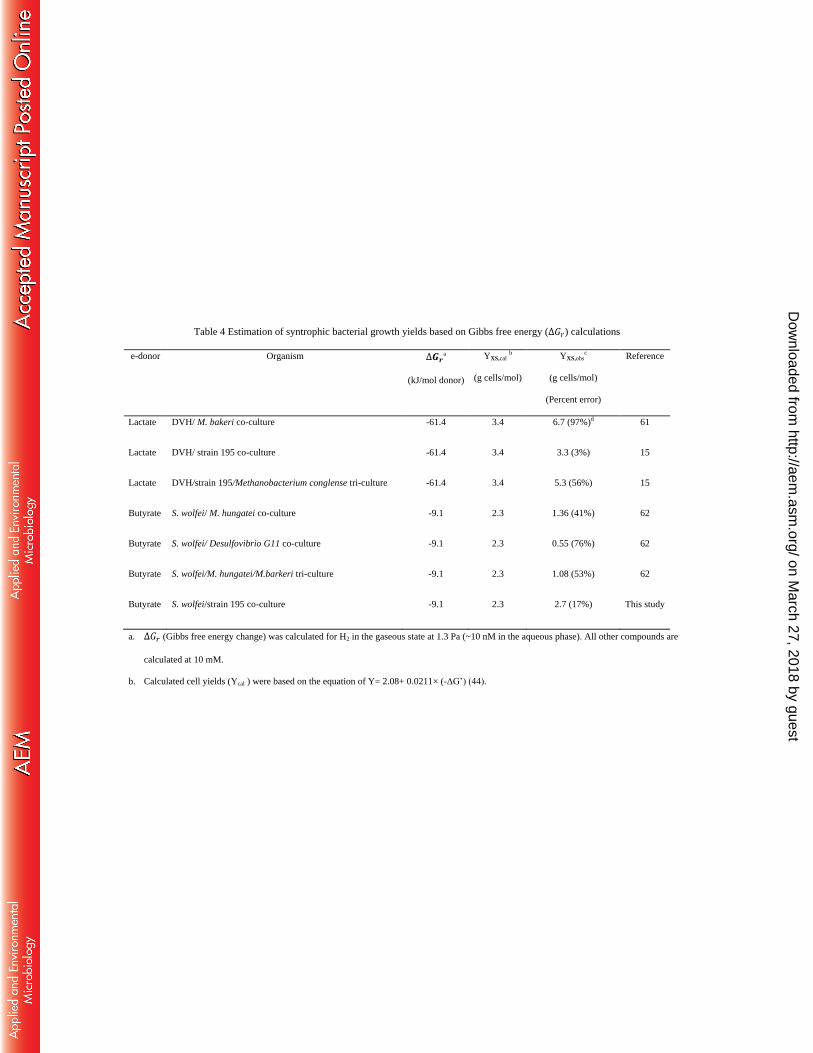
Table 4 Estimation of syntrophic bacterial growth yields based on Gibbs free energy (∆𝐺𝑟) calculations
e-donor Organism ∆𝑮𝒓a
(kJ/mol donor)
YXS,cal b
(g cells/mol)
YXS,obsc
(g cells/mol)
(Percent error)
Reference
Lactate DVH/ M. bakeri co-culture -61.4 3.4 6.7 (97%)d 61
Lactate DVH/ strain 195 co-culture -61.4 3.4 3.3 (3%) 15
Lactate DVH/strain 195/Methanobacterium conglense tri-culture -61.4 3.4 5.3 (56%) 15
Butyrate S. wolfei/ M. hungatei co-culture -9.1 2.3 1.36 (41%) 62
Butyrate S. wolfei/ Desulfovibrio G11 co-culture -9.1 2.3 0.55 (76%) 62
Butyrate S. wolfei/M. hungatei/M.barkeri tri-culture -9.1 2.3 1.08 (53%) 62
Butyrate S. wolfei/strain 195 co-culture -9.1 2.3 2.7 (17%) This study
a. ∆𝐺𝑟 (Gibbs free energy change) was calculated for H2 in the gaseous state at 1.3 Pa (~10 nM in the aqueous phase). All other compounds are
calculated at 10 mM.
b. Calculated cell yields (Ycal ) were based on the equation of Y= 2.08+ 0.0211× (-ΔG’) (44).
on March 27, 2018 by guest
http://aem.asm
.org/D
ownloaded from

c. Observed cell yields (Yobs) were based on directly measured cell masses (61) or masses converted from protein concentrations by assuming 2 g
cells/ g protein (62), or converted from cell numbers based on the assumption of 6×10-13 g per syntrophic cell (15) (44) (62).
d. Absolute error E = 3.3 g cells mol-1; Relative error = 0.97 ; Percent error = 97%
on March 27, 2018 by guest
http://aem.asm
.org/D
ownloaded from