timp-1 deficiency leads to lethal partial hepatic ischemia and reperfusion injury
TRANSCRIPT

TIMP-1 Deficiency Leads to Lethal Partial HepaticIschemia and Reperfusion Injury
Sergio Duarte, Takashi Hamada, Naohisa Kuriyama, Ronald W. Busuttil, and Ana J. Coito
Hepatic ischemia and reperfusion injury (IRI) remains an important challenge in clinicalorthotopic liver transplantation (OLT). Tissue inhibitor of metalloproteinase-1 (TIMP-1) is themajor endogenous regulator of matrix metalloproteinase-9 (MMP-9). In this study we investi-gated the functional significance of TIMP-1 expression in a well-established mouse model ofpartial liver IRI. Compared to wildtype mice, TIMP-12/2 mice showed further impaired liverfunction and histological preservation after IRI. Notably, TIMP-1 deficiency led to lethal liverIRI, as over 60% of the TIMP-12/2 mice died postreperfusion, whereas all TIMP-11/1 micerecovered and survived surgery. Lack of TIMP-1 expression was accompanied by markedlyhigh levels of MMP-9 activity, which facilitates leukocyte transmigration across vascularbarriers in hepatic IRI. Indeed, TIMP-12/2 livers were characterized by massive leukocyte infil-tration and by up-regulation of proinflammatory mediators, including tumor necrosis factoralpha, interferon-gamma, and inducible nitric oxide synthase post-IRI. The inability of TIMP-12/2 mice to express TIMP-1 increased the levels of active caspase-3 and depressed the expres-sion of Bcl-2 and the phosphorylation of Akt, emphasizing an important role for TIMP-1expression on hepatocyte survival. Using independent parameters of regeneration, 5-bromo-deoxyuridine incorporation, proliferating cell nuclear antigen expression, and histone H3 phos-phorylation, we provide evidence that hepatocyte progression into S phase and mitosis wasimpaired in TIMP-1-deficient livers after IRI. Inhibition of the cell cycle progression by TIMP-1 deficiency was linked to depressed levels of cyclins-D1 and -E and to a disrupted c-Met signal-ing pathway, as evidenced by reduced phosphorylated c-Met expression and elevated c-Metectodomain shedding postliver IRI. Conclusion: These results support a critical protectivefunction for TIMP-1 expression on promoting survival and proliferation of liver cells and onregulating leukocyte recruitment and activation in liver IRI. (HEPATOLOGY 2012;00:000-000)
Hepatic ischemia/reperfusion injury (IRI)occurs during trauma, shock, orthotopic livertransplantation (OLT), and other surgical
procedures where the blood supply to the liver is tem-porarily interrupted.1 Hepatic IR-related damage is theresult of various factors that include leukocyte migra-tion, release of cytokines, and free radicals.1,2
Leukocytes migration across endothelial and extrac-ellular matrix (ECM) barriers is dependent on cellularadhesion-release and focal matrix degradation mecha-nisms.3 Although adhesion molecules are importantfor the successful leukocyte transmigration by provid-ing leukocyte attachment to the endothelium, there isa growing body of evidence suggesting that matrixmetalloproteinases (MMP) are critical for facilitatingleukocyte movement across vascular barriers.3 In thisregard, our previous studies showed an important rolefor leukocyte-expressed MMP-9, or gelatinase B, as a
Abbreviations: ALT, alanine transaminase; AST, aspartate transaminase;BrdU, 5-bromodeoxyuridine; ECM, extracellular matrix; IFN-c, interferon-gamma; iNOS, inducible nitric oxide synthase; IRI, ischemia and reperfusioninjury; MMP-9, matrix metalloproteinase-9; OLT, orthotopic livertransplantation; PCNA, proliferating cell nuclear antigen; TIMP-1, tissueinhibitor of metalloproteinase-1; TNF-a, tumor necrosis factor alpha.From the Dumont-UCLA Transplant Center, Division of Liver and Pancreas
Transplantation, Department of Surgery, David Geffen School of Medicine atUCLA, Los Angeles, CA.Received September 2, 2011; accepted March 4, 2012.Supported by grants from the National Institutes of Health (NIH), National
Institute of Allergy and Infectious Diseases (NIAID) R01AI057832, UCLA AcademicSenate, and the Pfleger Foundation (to A.J.C.). S.D. was supported in part by adoctoral fellowship from the Fundacco para a Ciencia e Tecnologia (FCT), Portugal.Address reprint requests to: Dr. Ana J. Coito, Dumont-UCLA Transplant
Center, 77-120 CHS, Box: 957054, Los Angeles, CA 90095-7054.E-mail: [email protected]; .CopyrightVC 2012 by the American Association for the Study of Liver Diseases.View this article online at wileyonlinelibrary.com.DOI 10.1002/hep.25710Potential conflict of interest: Nothing to report.Additional Supporting Information may be found in the online version of
this article.
1

key mediator of leukocyte transmigration leading toliver injury.4
Tissue inhibitors of metalloproteinases (TIMPs) area family of naturally occurring inhibitors of MMPs.Alterations in the MMP-TIMP balance have beenlinked to pathological conditions that require disrup-tion of the basement membrane, such as tumor inva-sion, angiogenesis, and wound healing.5 There are atleast four identified members (TIMP 1-4) in theTIMP family, varying in tissue-specific expression andin their ability to inhibit various MMPs.6 Among thedifferent TIMPs, TIMP-1 is of particular interest;TIMP-1 is a 28.5-kDa soluble glycoprotein known toinhibit MMP-9 with high affinity, without interactingwith MMP-2, or gelatinase A (the other member ofthe gelatinase family), as it lacks the required C-termi-nal MMP-2-interacting residues.7,8
In addition to its ability to inhibit MMP activity,TIMP-1 possesses other biological activities, such ascell growth regulation, that are just beginning to berecognized and characterized.9 The specific effects ofTIMPs likely depend on the cell context and on thepathological condition. Although TIMP-1 has beendetected in the plasma of patients after liver transplan-tation,10 and in rat liver grafts after IRI,11 its role inliver IRI, or in OLT, remains to be established. There-fore, in the present study we used mice lacking TIMP-1 to examine the significance of TIMP-1 expression inhepatic IRI.
Materials and Methods
Mice and Model of Hepatic IRI. Male TIMP-1�/�
knockout (KO) mice in the C57BL/6 background(B6.129S4-Timp1tm1Pd/J) and respective TIMP-1þ/þ
wildtype (WT) C57BL-6 controls were obtained fromthe Jackson Laboratory. Hepatic IRI was performed asdescribed.4 Briefly, arterial and portal venous bloodsupplies were interrupted to the cephalad lobes of theliver for 90 minutes using an atraumatic clip and micewere sacrificed after reperfusion. The animal studieswere performed according to approved guidelines bythe American Association of Laboratory Animal Care.Assessment of Liver Damage. Serum alanine trans-
aminase (ALT) and serum aspartate transaminase(AST) levels were measured with an autoanalyzer byANTECH Diagnostics (Los Angeles, CA), asdescribed.4 Liver specimens were fixed with a 10%buffered formalin solution, embedded in paraffin, andprocessed for hematoxylin and eosin (H&E) staining;to determine the percentage of necrotic area, 10 ran-
dom sections per slide were evaluated in duplicateusing National Institutes of Health (NIH) Image-J.Immunohistochemistry. Immunostaining was per-
formed in cryostat sections as described.4,11 Mac-1(M1/70) and Ly-6G (1A8), from BD Biosciences,TIMP-1 (Ab86482; Abcam), MMP-9 (AF909; R&DSystems), and cleaved-caspase-3 (ASP175; Cell Signal-ing) antibodies were used at optimal dilutions. Sec-tions were blindly evaluated by counting 10 high-pow-ered fields (HPFs)/section in triplicate. Dual/triplestaining was detected by immunofluorescence withAlexa Fluor 594-red antigoat immunoglobulin G(IgG) (HþL) (Molecular Probes), and Texas Red anti-rat IgG (HþL) antibodies (Vector Laboratories). AlexaFluor 488 phalloidin (Molecular Probes) and Vecta-shield mounting media with DAPI (Vector Laborato-ries) were used for F-actin and nuclear staining, respec-tively. Slides were analyzed using a Leica ConfocalMicroscope (UCLA Brain Research Institute).Parameters of Regeneration. Mice were injected in-
traperitoneally with 50 mg/kg of 5-bromodeoxyuridine(BrdU) (Sigma) 2 hours prior to liver harvest asdescribed.12 BrdU incorporation, proliferating cell nu-clear antigen (PCNA), and phosphorylated histone H3were detected by immunohistochemistry in paraffinsections using anti-BrdU (Bu20a; Neomarkers), anti-PCNA (PC-10; Neomarkers), and anti-pH3 (Ser10;Cell Signaling) antibodies. Proliferation indexes weredetermined in triplicate and quantified under light mi-croscopy by counting 10, randomly chosen, HPFs/sec-tion. Data are expressed as the percentage of BrdU,PCNA, or pH3 stained hepatocytes per total numberof hepatocytes.Myeloperoxidase (MPO) Assay. MPO activity was
evaluated in frozen tissue homogenized in an iced so-lution of 0.5% hexadecyltrimethyl-ammonium and 50mmol/L of potassium phosphate buffer solution.4 Aftercentrifugation the supernatants were mixed in a solu-tion of hydrogen peroxide-sodium acetate and tetra-methyl benzidine (Sigma). The quantity of enzymedegrading 1 lmol/L of peroxide/min at 25�C/g of tis-sue was defined as 1U of MPO activity.Western Blot and Zymography Analysis. Western
blots and Zymography were performed asdescribed.4,11 Proteins (40 lg/sample) in sodium do-decyl sulfate (SDS)-loading buffer were electropho-resed through 12% SDS-polyacrylamide gel electro-phoresis (PAGE) and transferred to PVDFmembranes. Membranes were incubated with specificantibodies against cleaved caspase-3 (ASP175), phos-pho-AKT (D9E; C31E5E), AKT (C67E7), phospho-c-Met (D26 and 130H2), c-Met (25H2) (Cell
2 DUARTE ET AL. HEPATOLOGY, Month 2012

Signaling), Bcl-2 (Abcam), and cyclin D1 (BD Bio-sciences). After development, membranes were strippedand reblotted with antiactin antibody (Santa Cruz).Gelatinolytic activity was detected in liver extracts
(100 lg) by 10% SDS-PAGE contained 1 mg/mL ofgelatin (Invitrogen) under nonreducing conditions. Af-ter incubation in development buffer (50 mmol/L Tris-HCl, 5 mmol/L CaCl2, and 0.02% NaN3, pH 7.5),gels were stained with Coomassie brilliant blue R-250(Bio-Rad) and destained with methanol/acetic acid/water (20:10:70). Prestained molecular weight markers(Bio-Rad) and MMP-9 (BIOMOL International)served as standards. Relative quantities of protein weredetermined using a densitometer (NIH Image Jsoftware).RNA Extraction and Reverse Transcriptase Poly-
merase Chain Reaction (PCR). RNA was extractedfrom livers with Trizol (Life Technologies) asdescribed.4 Reverse transcription was performed using5 lg of total RNA in a first-strand complementaryDNA (cDNA) synthesis reaction with SuperScript IIIRNaseH Reverse Transcriptase (Life Technologies) asrecommended by the manufacturer. The cDNA prod-uct was amplified by PCR using primers specific foreach target cDNA.Data Analysis. Data in the text and figures are
expressed as mean 6 standard deviation. Statisticalcomparisons between groups of normally distributeddata were performed with Student’s t test using statisti-cal package SPSS (Chicago, IL). Kaplan-Meier analysiswas used to determine statistical significance of the dif-ferences in mouse survival. P < 0.05 was consideredstatistically significant.
Results
Time Course of TIMP-1 Expression in WildtypeLivers Post-IRI. TIMP-1 messenger RNA (mRNA)was almost undetectable in naive livers and it was sig-nificantly up-regulated in TIMP-1þ/þ livers from 3hours to 7 days postreperfusion (Fig. 1A). TIMP-1protein expression was mildly detected in TIMP-1þ/þ
naive livers and it was markedly increased in livers af-ter 6 hours of reperfusion, particularly at 24 hours and48 hours post-IRI (Fig. 1B). Immunofluorescenceanalysis showed TIMP-1 staining in the surviving pa-renchyma predominantly around the portal triads ofwildtype livers (Fig. 1C); TIMP-1þ staining wasmostly detected in cells along hepatic sinusoids, likelyhepatic stellate cells (HSCs), and in scattered hepato-cytes. In vitro studies have linked TIMP-1 productionto HSC and to hepatocytes.13 Conversely, TIMP-1
staining was absent in TIMP-1�/� livers after IRI (Fig.1C).TIMP-1-Deficient Mice Had Reduced Survival
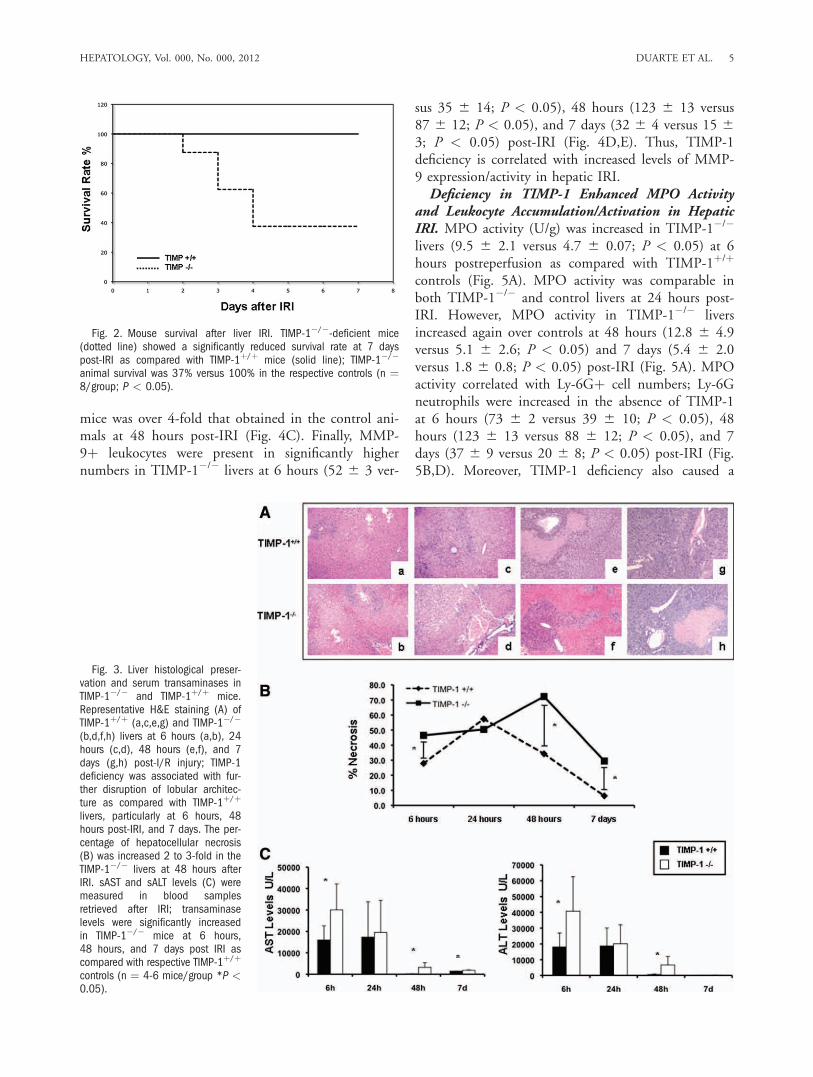
Rate After Hepatic IRI. To test the significance ofTIMP-1 expression in liver IRI, our experimentsincluded TIMP-1-deficient and respective wildtype(TIMP-1þ/þ) control mice. The model of partial liverIRI is nonlethal14; regardless of the significant liverdamage detected during the first few days of hepaticIRI, virtually every animal survives after reperfusion.Notably, TIMP-1 deficiency resulted in an unantici-pated reduced survival rate post-IRI (37% versus100%; P < 0.05). Only three out of the eight TIMP-1�/� mice survived after reperfusion, whereas all eightTIMP-1þ/þ WT animals recovered from injury andsurvived up to 7 days post-IRI (Fig. 2). TIMP-1�/�
mice failed to recover from the injury and succumbedbetween the second and fourth day post-IRI. There-fore, these results indicate an important role forTIMP-1 expression in hepatic IRI.Liver Damage Was Increased in TIMP-1-Deficient
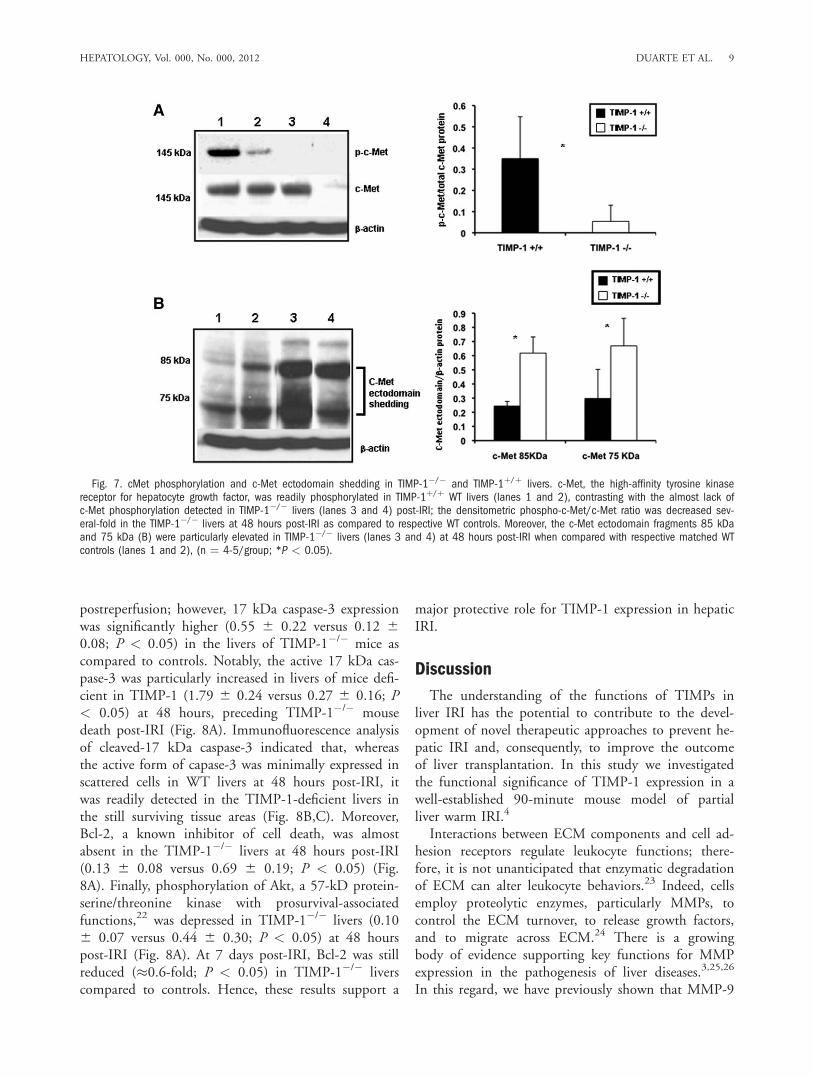
Mice After IRI. There were no detectable differencesin liver histology and transaminase levels between na-ive TIMP-1�/� and naive WT mice. WT livers werecharacterized by significant sinusoidal congestion andextensive necrosis after reperfusion; however, TIMP-1deficiency was associated with further lobular architec-ture disruption at 6 hours, 48 hours, and 7 days post-IRI (Fig. 3A). Indeed, TIMP-1�/� mice demonstrated2 to 3-fold higher levels of hepatocellular necrosis (P< 0.05) when compared with TIMP-1þ/þ mice at 48hours post-IRI (Fig. 3B). TIMP-1�/� mice that sur-vived surgery showed improved liver histology at 7days post-IRI; however, levels of liver necrosis werestill higher in these mice when compared to respectiveWT controls (Fig. 3A,B). The serum transaminase lev-els (U/L) were significantly increased in TIMP-1 miceat 6 hours (sAST: 30,040 6 12,104 versus 16,033 66,598, P < 0.05; sALT: 40,660 6 21,970 versus18,148 6 8,727, P < 0.05), 48 hours (sAST: 3,2906 2,170 versus 197.75 6 82.44, P < 0.05; sALT:6,720 6 5,298 versus 571.25 6 348.9, P < 0.05),and 7 days (sAST: 1,909 6 155 versus 1,472 6 62, P< 0.05; sALT: 254 6 88 versus 119 6 42, P < 0.05)post-IRI (Fig. 3C). Altogether, these data emphasizethe concept that TIMP-1 has a protective function inhepatic IRI.MMP-9 Expression and Activity Was Up-regu-
lated in TIMP-1-Deficient Livers After IRI. TIMP-1�/� mice showed significantly up-regulated MMP-9/b-actin mRNA expression at 6 hours (0.44 6 0.17versus 0.20 6 0.11; P < 0.05), 48 hours (0.53 6
HEPATOLOGY, Vol. 000, No. 000, 2012 DUARTE ET AL. 3

0.15 versus 0.29 6 0.07; P < 0.05), and 7 days (0.486 0.13 versus 0.19 6 0.14; P < 0.05) after IRI (Fig.4A). Moreover, zymography analysis showed thatMMP-9 activity was almost undetected in naive liversand highly expressed in TIMP-1�/� and WT livers
post-IRI; however, MMP-9 activity was markedly up-regulated in the livers of TIMP-1�/� mice after 6hours (P < 0.05) and 48 hours (P < 0.05) of reperfu-sion as compared to controls (Fig. 4B). Indeed, theMMP-9 activity increase observed in the TIMP-1�/�
Fig. 1. Time course of TIMP-1 expression in TIMP-1þ/þ livers post-IRI. TIMP-1 mRNA expression (A) was almost absent in naive WT livers(lane 1) and it was significantly up-regulated at 3 hours (lanes 2 and 3), 6 hours (lanes 4 and 5), 24 hours (lanes 6 and 7), 48 hours (lanes8 and 9), and 7 days (lanes 10 and 11) after liver IRI. TIMP-1 protein (B) was mildly expressed in naive WT livers (lanes 1 and 2) and in liversat 3 hours (lanes 3 and 4) and 7 days (lanes 10 and 11) post-IRI; however, it was markedly increased in livers at 6 hours (lanes 5 and 6), 24hours (lanes 7 and 8), and 48 hours (lanes 9 and 10) after IRI. (C) Representative immunofluorescence staining in TIMP-1þ/þ (a,c,e,g) andTIMP-1�/� (b,d,f,h) livers at 6 hours post-IRI; TIMP-1 in red (a,b; Alexa Fluor 594), F-actin in green (c,d; Alexa Fluor 488 phalloidin), nuclearstain in blue (e,f; Dapi), and staining overlay (g,h); TIMP-1-positive staining was mostly detected in the surviving parenchyma surrounding thevasculature of WT livers post-IRI, whereas TIMP-1 staining was undetectable in the TIMP-1�/� livers (arrows denote TIMP-1 staining; n ¼ 4/group; *P < 0.05 relative to naive livers).
4 DUARTE ET AL. HEPATOLOGY, Month 2012
mice was over 4-fold that obtained in the control ani-mals at 48 hours post-IRI (Fig. 4C). Finally, MMP-9þ leukocytes were present in significantly highernumbers in TIMP-1�/� livers at 6 hours (52 6 3 ver-
sus 35 6 14; P < 0.05), 48 hours (123 6 13 versus87 6 12; P < 0.05), and 7 days (32 6 4 versus 15 63; P < 0.05) post-IRI (Fig. 4D,E). Thus, TIMP-1deficiency is correlated with increased levels of MMP-9 expression/activity in hepatic IRI.Deficiency in TIMP-1 Enhanced MPO Activity
and Leukocyte Accumulation/Activation in HepaticIRI. MPO activity (U/g) was increased in TIMP-1�/�
livers (9.5 6 2.1 versus 4.7 6 0.07; P < 0.05) at 6hours postreperfusion as compared with TIMP-1þ/þ
controls (Fig. 5A). MPO activity was comparable inboth TIMP-1�/� and control livers at 24 hours post-IRI. However, MPO activity in TIMP-1�/� liversincreased again over controls at 48 hours (12.8 6 4.9versus 5.1 6 2.6; P < 0.05) and 7 days (5.4 6 2.0versus 1.8 6 0.8; P < 0.05) post-IRI (Fig. 5A). MPOactivity correlated with Ly-6Gþ cell numbers; Ly-6Gneutrophils were increased in the absence of TIMP-1at 6 hours (73 6 2 versus 39 6 10; P < 0.05), 48hours (123 6 13 versus 88 6 12; P < 0.05), and 7days (37 6 9 versus 20 6 8; P < 0.05) post-IRI (Fig.5B,D). Moreover, TIMP-1 deficiency also caused a
Fig. 2. Mouse survival after liver IRI. TIMP-1�/�-deficient mice(dotted line) showed a significantly reduced survival rate at 7 dayspost-IRI as compared with TIMP-1þ/þ mice (solid line); TIMP-1�/�
animal survival was 37% versus 100% in the respective controls (n ¼8/group; P < 0.05).
Fig. 3. Liver histological preser-vation and serum transaminases inTIMP-1�/� and TIMP-1þ/þ mice.Representative H&E staining (A) ofTIMP-1þ/þ (a,c,e,g) and TIMP-1�/�
(b,d,f,h) livers at 6 hours (a,b), 24hours (c,d), 48 hours (e,f), and 7days (g,h) post-I/R injury; TIMP-1deficiency was associated with fur-ther disruption of lobular architec-ture as compared with TIMP-1þ/þ
livers, particularly at 6 hours, 48hours post-IRI, and 7 days. The per-centage of hepatocellular necrosis(B) was increased 2 to 3-fold in theTIMP-1�/� livers at 48 hours afterIRI. sAST and sALT levels (C) weremeasured in blood samplesretrieved after IRI; transaminaselevels were significantly increasedin TIMP-1�/� mice at 6 hours,48 hours, and 7 days post IRI ascompared with respective TIMP-1þ/þ
controls (n ¼ 4-6 mice/group *P <0.05).
HEPATOLOGY, Vol. 000, No. 000, 2012 DUARTE ET AL. 5

substantial increase of infiltrating Mac-1 macrophagesat 6 hours (67 6 3 versus 37 6 10; P < 0.05), 24hours (73 6 2 versus 41 6 8; P < 0.05), 48 hours(154 6 34 versus 101 6 15; P < 0.05), and 7 days(64 6 19 versus 30 6 5; P < 0.05) post-IRI (Fig.5C,D). The extent of leukocyte infiltration correlatedwith proinflammatory cytokine expression; tumor ne-crosis factor alpha (TNF-a) (0.66 6 0.15 versus 0.376 0.28; P < 0.05), interleukin (IL)-1b (1.08 6 0.29versus 0.75 6 0.24 P < 0.05), and interferon-gamma(IFN-c) (1.08 6 0.29 versus 0.75 6 0.24; P < 0.05)were significantly up-regulated in TIMP-1�/� livers at
6 hours post-IRI (Fig 5E). TIMP-1�/� livers at 48hours (IL-1b: 0.21 6 0.04 versus 0.10 6 0.02; P <0.05) and 7 days (IL-1b: 0.20 6 0.04 versus 0.14 60.03 and TNF-a: 0.32 6 0.07 versus 0.21 6 0.04; P< 0.05) post-IRI were also characterized by signifi-cantly increased proinflammatory cytokine expression.Further, inducible nitric oxide synthase (iNOS) expres-sion, which associates with liver injury,15 showed an�2.5-fold increase (P < 0.05) in 6-hour TIMP-1�/�
livers. In contrast, IL-10, well known for its protectiverole in hepatic IRI,16 was down-regulated in TIMP-1�/� livers at 48 hours (0.26 6 0.13 versus 0.65 6
Fig. 4. MMP-9 expression and activity in TIMP-1�/� and TIMP-1þ/þ mice. MMP-9 mRNA expression (A), as detected by RT-PCR analysis,was significantly up-regulated in TIMP-1�/� mice at 6 hours, 48 hours, and 7 days after IR injury as compared to the respective WT controls.MMP-9 activity (B,C), analyzed by zymography in TIMP-1þ/þ (lanes 1, 3, 4, 7, 8, 11, 12, 15, and 16) and TIMP-1�/� (lanes 2, 5, 6, 9, 10,13, 14, 17, and 18) livers; MMP-9 activity was almost absent in naive livers of TIMP-1þ/þ (lane 1) and TIMP-1�/� (lane 2) mice and highlydetectable in TIMP-1þ/þ and TIMP-1�/� livers at 6 hours (lanes 3-6), 24 hours (lanes 7-10), 48 hours (lanes 11-14), and 7 days (lanes 15-18) post-IRI; however, compared to controls, MMP-9 activity was markedly up-regulated in TIMP-1�/� livers at 6 hours, 48 hours, and 7 days af-ter IRI. MMP-9þ cells (D,E) in WT controls (a,c,e) and TIMP-1�/� livers (b,d,f) at 6 hours (a,b), 24 hours (c,d), and 48 hours (e,f) post-IRI;MMP-9þ cells were detected in significantly higher numbers in TIMP-1�/� livers, particularly at 6 hours, 48 hours, and 7 days postreperfusion(n ¼ 4-5/group; *P < 0.05).
6 DUARTE ET AL. HEPATOLOGY, Month 2012

0.14; P < 0.05) and 7 days (0.43 6 0.21 versus 0.826 0.14; P < 0.05) post-IRI.TIMP-1 Deficiency Did Not Alter the Expression
of Major Chemokines in Hepatic IRI. To determinewhether TIMP-1 deficiency affects chemokine expres-sion, we assessed major cell activating chemokineslinked to liver IRI (Fig. 5F). CXCL-1 (1.16 6 0.19versus 1.02 6 0.03) and CXCL-2 (0.24 6 0.18 versus0.24 6 0.06) were comparably expressed in bothTIMP-1�/� and wildtype livers at 6 hours post-IRI.Moreover, TIMP-1�/� and WT livers also expressedsimilar levels of MCP-1 (0.86 6 0.11 versus 0.66 60.20) and SDF-1 (0.45 6 0.13 versus 0.45 6 0.02) 6hours postreperfusion. The expression levels of thesechemokines were also comparable in TIMP-1�/� andWT livers at 24 hours, 48 hours, and 7 days post-IRI(data not shown).Deficiency in TIMP-1 Impaired Liver Regenera-
tion After IRI. To determine whether TIMP-1 defi-ciency interferes with cell proliferation, the percentageof cells in S phase, the BrdU and PCNA labelingindexes, and the percentage of phosphorylated histoneH3 (P-H3)-positive cells, the mitotic index (MI), were
evaluated after liver IRI. BrdU (0.53 6 0.11 versus1.70 6 0.13; P < 0.05), PCNA (0.51 6 0.46 versus5.02 6 2.98; P < 0.05), and MI (0.50 6 0.46versus 2.96 6 1.67) indexes were modestly detected at24 hours post-IRI, with decreased proliferation indexesin the TIMP-1�/� livers when compared to controls.Although BrdU (0.92 6 0.11 versus 6.46 6 0.24;P < 0.05), PCNA (2.65 6 0.33 versus 26.96 6 2.74;P < 0.05), and MI (1.87 6 1.71 versus 10.74 61.82; P < 0.05) indexes were still almost negligible inTIMP-1�/� livers at 48 hours post-IRI, they were sig-nificantly increased in TIMP-1þ/þ controls (Fig. 6A-C). Several TIMP-1�/� animals died between the sec-ond and fourth day post-IRI; nonetheless, TIMP-1�/�
mice that survived surgery exhibited some evidence ofdelayed liver regeneration, as the MI (7.16 6 2.47versus 3.39 6 1.17) was enhanced in these animals at7 days post-IRI. Moreover, cyclin D1, a regulator ofthe G1-to-S phase transition,17 and cyclin E, also nec-essary for entry into S phase,18 were down-regulated atmRNA levels in TIMP-1�/� livers (cyclin D1: 0.21 60.04 versus 0.53 6 0.11; P < 0.05; cyclin E: 0.44 60.32 versus 1.18 6 0.42; P < 0.05) at 48 hours post-
Fig. 5. Intrahepatic MPO enzyme activityand leukocyte infiltration/activation in TIMP-1�/� and TIMP-1þ/þ mice. MPO enzymaticactivity (A), an index of neutrophil infiltration,was markedly up-regulated in TIMP-1�/� liv-ers when compared to WT controls at 6 hours,48 hours, and 7 days after IRI. Ly-6Gþ neu-trophil infiltration (B) was significantlyincreased in livers of TIMP-1�/� mice at 6hours, 48 hours, and 7 days post-IRI. Mac-1þ macrophages (C) were detected in highernumbers in livers of TIMP-1�/� mice at 6hours, 24 hours, 48 hours, and 7 days post-IRI. (D) Immunostaining of Ly-6G neutrophils(left) and Mac-1 macrophages (right) in TIMP-1þ/þ livers (a) and TIMP-1�/� (b) at 6 hoursafter IRI. Proinflammatory mediators (E) inTIMP-1þ/þ and TIMP-1�/� livers; TNF-a, IL-1b, IFN-c, and iNOS mRNA levels were signifi-cantly up-regulated in TIMP-1�/� deficient liv-ers at 6 hours post-IRI as compared torespective controls. Chemokine gene evalua-tion (F) showed comparable expressions ofCXCL-1, CXCL-2, MCP-1, and SDF-1 in TIMP-1þ/þ and TIMP-1�/� livers after reperfusion(n ¼ 4-5/group; *P < 0.05).
HEPATOLOGY, Vol. 000, No. 000, 2012 DUARTE ET AL. 7

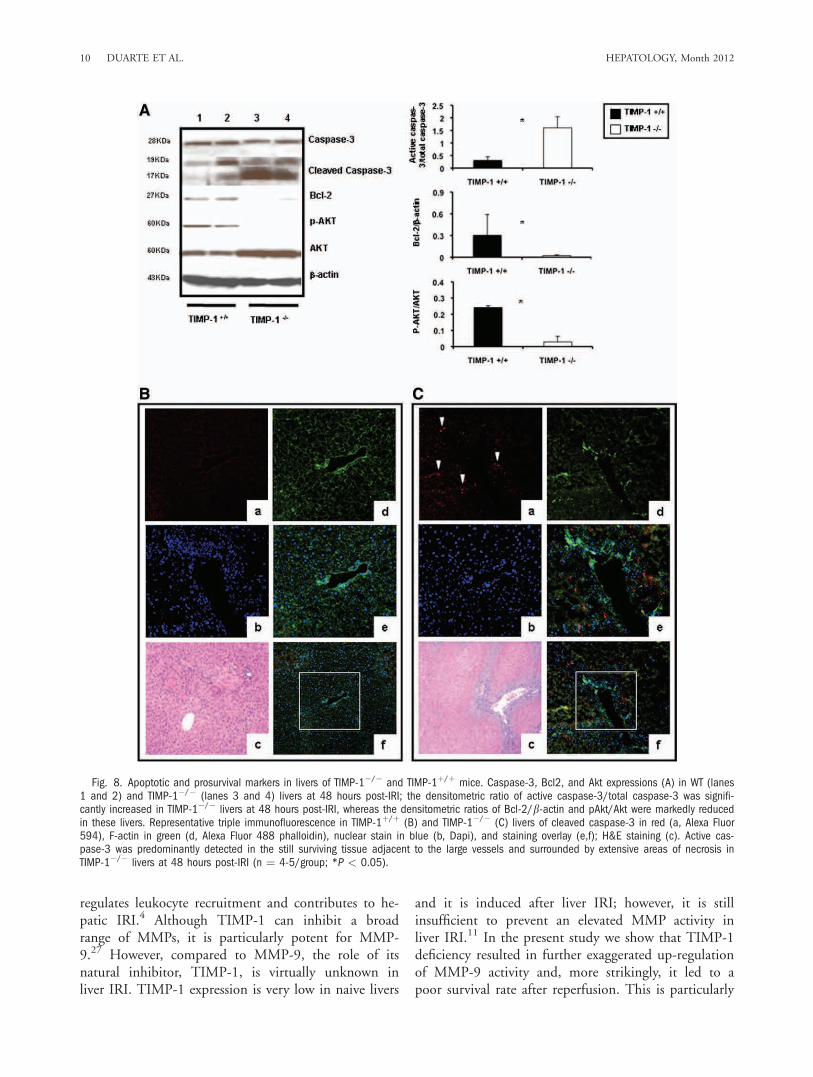
reperfusion (Fig 6D). Cyclin D1 was almost absent inTIMP-1�/� livers at the protein level (0.20 6 0.26versus 1.19 6 0.25; P < 0.05), contrasting with analmost 6-fold increased expression detected in WT liv-ers at 48 hours post-IRI (Fig. 6E). c-Met-HGF inter-actions result in c-Met phosphorylation, which is thecentral stimulus for the G1-S progression of hepato-cytes.19 The inability of TIMP-1�/� mice to expressTIMP-1 led to markedly decreased HGF/c-Met signal-ing, as evidenced by the markedly reduced levels ofphosphorylated c-Met (0.05 6 0.07 versus 0.35 60.20; P < 0.05) in their livers at 48 hours post-IRI(Fig. 7A). Further, c-Met ectodomain shedding, a pro-
cess by which proteins are proteolytically released fromthe cell surface, negatively regulates c-Met signaling.20
In our settings, the absence of TIMP-1 resulted in sig-nificantly enhanced c-Met ectodomain shedding inliver IRI (Fig. 7B). Therefore, these results evidencethat loss of TIMP-1 interferes with liver regenerationafter IRI.Lack of TIMP-1 Exacerbates Caspase-3 Activation
in Liver IRI. Caspase-3 is expressed in tissues as aninactive 32-kDa precursor, which is cleaved to generatea 17-kDa mature active form during apoptosis.21 Theactive caspase-3 was absent in naive livers andincreased in TIMP-1�/� and WT livers at 6 hours
Fig. 6. Expression of hepaticregenerative markers in TIMP-1�/�
and TIMP-1þ/þ mice. HepatocyteBrdU incorporation (A), PCNA label-ing (B), and phosphorylated his-tone H3-positive cells (C) in TIMP-1þ/þ (a,c) and TIMP-1�/� (b,d)livers at 48 hours post-IRI; PCNAstaining (c,d) is shown at highermagnification to better illustratepositive (c) and virtually negative(d) PCNA hepatocyte-labeling in thesurviving parenchyma of TIMP-1þ/þ
and TIMP-1�/� livers, respectively.TIMP-1�/� livers showed markedlydiminished BrdU, PCNA, and mitoticlabeling indexes as compared tocontrols. The densitometric ratios ofcyclin D1/b-actin and cyclin E/b-actin mRNA (D) were significantlydepressed in TIMP-1�/� livers at48 hours post-IRI. Cyclin D1 at theprotein level (E) was also pro-foundly depressed in TIMP-1�/�
livers at 48 hours post-IRI (n ¼ 4-5/group; *P < 0.05).
8 DUARTE ET AL. HEPATOLOGY, Month 2012
postreperfusion; however, 17 kDa caspase-3 expressionwas significantly higher (0.55 6 0.22 versus 0.12 60.08; P < 0.05) in the livers of TIMP-1�/� mice ascompared to controls. Notably, the active 17 kDa cas-pase-3 was particularly increased in livers of mice defi-cient in TIMP-1 (1.79 6 0.24 versus 0.27 6 0.16; P< 0.05) at 48 hours, preceding TIMP-1�/� mousedeath post-IRI (Fig. 8A). Immunofluorescence analysisof cleaved-17 kDa caspase-3 indicated that, whereasthe active form of capase-3 was minimally expressed inscattered cells in WT livers at 48 hours post-IRI, itwas readily detected in the TIMP-1-deficient livers inthe still surviving tissue areas (Fig. 8B,C). Moreover,Bcl-2, a known inhibitor of cell death, was almostabsent in the TIMP-1�/� livers at 48 hours post-IRI(0.13 6 0.08 versus 0.69 6 0.19; P < 0.05) (Fig.8A). Finally, phosphorylation of Akt, a 57-kD protein-serine/threonine kinase with prosurvival-associatedfunctions,22 was depressed in TIMP-1�/� livers (0.106 0.07 versus 0.44 6 0.30; P < 0.05) at 48 hourspost-IRI (Fig. 8A). At 7 days post-IRI, Bcl-2 was stillreduced (�0.6-fold; P < 0.05) in TIMP-1�/� liverscompared to controls. Hence, these results support a
major protective role for TIMP-1 expression in hepaticIRI.
Discussion
The understanding of the functions of TIMPs inliver IRI has the potential to contribute to the devel-opment of novel therapeutic approaches to prevent he-patic IRI and, consequently, to improve the outcomeof liver transplantation. In this study we investigatedthe functional significance of TIMP-1 expression in awell-established 90-minute mouse model of partialliver warm IRI.4
Interactions between ECM components and cell ad-hesion receptors regulate leukocyte functions; there-fore, it is not unanticipated that enzymatic degradationof ECM can alter leukocyte behaviors.23 Indeed, cellsemploy proteolytic enzymes, particularly MMPs, tocontrol the ECM turnover, to release growth factors,and to migrate across ECM.24 There is a growingbody of evidence supporting key functions for MMPexpression in the pathogenesis of liver diseases.3,25,26
In this regard, we have previously shown that MMP-9
Fig. 7. cMet phosphorylation and c-Met ectodomain shedding in TIMP-1�/� and TIMP-1þ/þ livers. c-Met, the high-affinity tyrosine kinasereceptor for hepatocyte growth factor, was readily phosphorylated in TIMP-1þ/þ WT livers (lanes 1 and 2), contrasting with the almost lack ofc-Met phosphorylation detected in TIMP-1�/� livers (lanes 3 and 4) post-IRI; the densitometric phospho-c-Met/c-Met ratio was decreased sev-eral-fold in the TIMP-1�/� livers at 48 hours post-IRI as compared to respective WT controls. Moreover, the c-Met ectodomain fragments 85 kDaand 75 kDa (B) were particularly elevated in TIMP-1�/� livers (lanes 3 and 4) at 48 hours post-IRI when compared with respective matched WTcontrols (lanes 1 and 2), (n ¼ 4-5/group; *P < 0.05).
HEPATOLOGY, Vol. 000, No. 000, 2012 DUARTE ET AL. 9
regulates leukocyte recruitment and contributes to he-patic IRI.4 Although TIMP-1 can inhibit a broadrange of MMPs, it is particularly potent for MMP-9.27 However, compared to MMP-9, the role of itsnatural inhibitor, TIMP-1, is virtually unknown inliver IRI. TIMP-1 expression is very low in naive livers
and it is induced after liver IRI; however, it is stillinsufficient to prevent an elevated MMP activity inliver IRI.11 In the present study we show that TIMP-1deficiency resulted in further exaggerated up-regulationof MMP-9 activity and, more strikingly, it led to apoor survival rate after reperfusion. This is particularly
Fig. 8. Apoptotic and prosurvival markers in livers of TIMP-1�/� and TIMP-1þ/þ mice. Caspase-3, Bcl2, and Akt expressions (A) in WT (lanes1 and 2) and TIMP-1�/� (lanes 3 and 4) livers at 48 hours post-IRI; the densitometric ratio of active caspase-3/total caspase-3 was signifi-cantly increased in TIMP-1�/� livers at 48 hours post-IRI, whereas the densitometric ratios of Bcl-2/b-actin and pAkt/Akt were markedly reducedin these livers. Representative triple immunofluorescence in TIMP-1þ/þ (B) and TIMP-1�/� (C) livers of cleaved caspase-3 in red (a, Alexa Fluor594), F-actin in green (d, Alexa Fluor 488 phalloidin), nuclear stain in blue (b, Dapi), and staining overlay (e,f); H&E staining (c). Active cas-pase-3 was predominantly detected in the still surviving tissue adjacent to the large vessels and surrounded by extensive areas of necrosis inTIMP-1�/� livers at 48 hours post-IRI (n ¼ 4-5/group; *P < 0.05).
10 DUARTE ET AL. HEPATOLOGY, Month 2012

interesting in that the model of partial liver IRI isnonlethal.14 Indeed, all TIMP-1þ/þ mice survived he-patic IRI despite the significant damage detected inthe livers after reperfusion; in contrast, only three outof eight TIMP-1�/� mice survived more than 4 daysafter liver IRI. In general, TIMP-1�/� mice showedadditional impairment of liver function and moresevere lesions, which likely led to their death betweenthe second and fourth day postreperfusion.Although infiltrating leukocytes are recognized as
mediators of hepatic IRI,3,28 the mechanisms involvedin their recruitment to sites of inflammatory stimulationin liver are still far from being understood. TIMP-1�/�
livers showed massive leukocyte accumulation post-IRI.This latter feature, together with the findings thatMMP-9 enzymatic activity was significantly increased inthe TIMP-1�/� livers, strongly support an importantregulatory role for TIMP-1 on leukocyte recruitment inhepatic IRI. Local concentrations of TIMP-1 are impor-tant for regulating MMP-9 activity in vivo,29 andTIMP-1 has also been implicated in leukocyte infiltra-tion into the damaged brain.30 In addition to amplifiedleukocyte migration, TIMP-1-deficient mice showed sig-nificantly increased levels of proinflammatory mediatorsafter liver injury. IFN-c and iNOS, which have beenlinked to tissue injury, including hepatic injury,15,31
were markedly up-regulated in the TIMP-1�/� liverspost-IRI. Moreover, TNF-a, whose expression is oftenassociated with neutrophil infiltration and liver dam-age,32 was also significantly increased in the TIMP-1livers after reperfusion.Impaired liver regeneration/repair is one of the most
frequent features in acute liver failure. Adult hepato-cytes, which make up to 80% of hepatic cells, arelong-lived and normally do not undergo cell division;however, they maintain the ability to proliferate inresponse to injury.33 Using three independent parame-ters of regeneration (BrdU, PCNA, and MIs), we pro-vide evidence that hepatocyte progression into S phaseand mitosis was disrupted in TIMP-1-deficient miceduring the first 48 hours post-IRI. Cyclins D1 and E,which are necessary for entry into S phase,17,18 wereprofoundly depressed in the TIMP-1-deficient liverspost-IRI. It is known that inhibition of cyclin D1leads to growth arrest and to impaired hepatic regener-ation.34 It is perhaps important to stress that the roleof TIMP-1 in liver regeneration may depend on thetype of injury, as TIMP-1 can negatively affect regen-eration after substantial hepatic resection.35
Our results agree with previous findings indicatingthat TIMP-1 has a growth-promoting activity in abroad variety of cells,9,36,37 including in hepatocytes,38
and that TIMP-1 can stimulate the HGF/cMet path-way by inhibiting MMP-mediated c-Met shedding.39
Activation of the HGF/cMet signaling pathwayrequires phosphorylation of c-Met, which is neededfor efficient liver regeneration.40 In our settings, theinability of TIMP-1�/� mice to express TIMP-1 ledto virtually undetectable phosphorylated c-Met levelsafter liver reperfusion. Further, TIMP-1 deficiencyresulted in increased proteolytic cMet ectodomainshedding, which may account in part for the reducedlevels of phosphorylated c-Met postliver IRI; soluble c-Met shed ectodomains act as decoy receptors by inter-fering with HGF binding to c-Met.20 Therefore, ourwork strongly supports the view that TIMP-1�/� livershave an impaired capability to regenerate after IRI.In addition to impaired liver regeneration, cell death
by necrosis, apoptosis, or necroapoptosis is a promi-nent feature of liver IRI.14,41 The expression of TIMP-1 was detected in the surviving parenchyma of WTmice after the ischemic insult, suggesting a potentialrole for TIMP-1 in conferring resistance to cell death.Indeed, TIMP-1�/�-deficient livers exhibited increasedliver necrosis, particularly at 48 hours post-IRI. More-over, caspase-3 activation, the executor of apoptosis,42
was significantly increased in TIMP-1�/� livers ascompared with control littermates after IRI, and it wasaccompanied by a decrease in Bcl-2 expression.Although morphologic alterations of apoptosis aremostly mediated by caspases,42 Bcl-2 is an integralmembrane antiapoptotic protein expressed even inhealthy cells.43 In this regard, it has been reported thatTIMP-1 can inhibit apoptosis in a wide variety of celltypes, including stellate cells,44 B cells,45 epithelialcells,46 and mesangial cells47 through MMP-dependentand -independent mechanisms. Moreover, it has alsobeen shown that exogenous TIMP-1 confers resistanceagainst apoptosis in isolated endothelial cells by way ofactivation of the PI-3/Akt signaling pathway.48 Akt is a57-kD protein-serine/threonine kinase with prosurvivalfunctions.22 In our settings, lack of TIMP-1 expressionresulted in almost completely depleted Akt phospho-rylation, without changing total Akt protein levels,suggesting that TIMP-1 activates the Akt signalingpathway in hepatic IRI. TIMP-1 inhibition of celldeath can also be mediated by way of its regulatoryrole on MMP enzymatic activity. The ECM proteolysismediated by MMPs can lead to detachment of livercells, resulting in apoptosis by a phenomenon called‘‘anoikis.’’49 Indeed, we have previously shown thatMMP-9, in addition to facilitating leukocyte infiltra-tion in livers after IRI, induces hepatocyte apoptosisafter IRI.15
HEPATOLOGY, Vol. 000, No. 000, 2012 DUARTE ET AL. 11

In summary, these studies demonstrate an importantprotective role for TIMP-1 expression in liver IRI.Overall, we show that TIMP-1 has relevant functionsin promoting cell survival and proliferation of livercells and on regulating leukocyte recruitment and acti-vation in liver IRI. The inability of TIMP-1�/� miceto express TIMP-1 resulted in enhanced liver damageand in lethal hepatic IRI. Moreover, our data providethe rationale for studies, currently under developmentin our laboratory, aimed at efficiently overexpressingTIMP-1 in vivo as a potential therapeutic approach toimprove hepatic IRI.
References1. Selzner N, Rudiger H, Graf R, Clavien PA. Protective strategies against
ischemic injury of the liver. Gastroenterology 2003;125:917-936.
2. Busuttil RW, Tanaka K. The utility of marginal donors in liver trans-plantation. Liver Transpl 2003;9:651-663.
3. Coito AJ. Leukocyte transmigration across endothelial and extracellularmatrix protein barriers in liver ischemia/reperfusion injury. Curr OpinOrgan Transplant 2011;16:34-40.
4. Hamada T, Fondevila C, Busuttil RW, Coito AJ. Metalloproteinase-9deficiency protects against hepatic ischemia/reperfusion injury.HEPATOLOGY 2008;47:186-198.
5. Chirco R, Liu XW, Jung KK, Kim HR. Novel functions of TIMPs incell signaling. Cancer Metastasis Rev 2006;25:99-113.
6. Egeblad M, Werb Z. New functions for the matrix metalloproteinasesin cancer progression. Nat Rev Cancer 2002;2:161-174.
7. Bjorklund M, Koivunen E. Gelatinase-mediated migration and invasionof cancer cells. Biochim Biophys Acta 2005;1755:37-69.
8. Morgunova E, Tuuttila A, Bergmann U, Tryggvason K. Structuralinsight into the complex formation of latent matrix metalloproteinase 2with tissue inhibitor of metalloproteinase 2. Proc Natl Acad Sci U S A2002;99:7414-7419.
9. Stetler-Stevenson WG. Tissue inhibitors of metalloproteinases in cell signal-ing: metalloproteinase-independent biological activities. Sci Signal 2008;1:re6.
10. Kuyvenhoven JP, Molenaar IQ, Verspaget HW, Veldman MG, PalaretiG, Legnani C, et al. Plasma MMP-2 and MMP-9 and their inhibitorsTIMP-1 and TIMP-2 during human orthotopic liver transplantation.The effect of aprotinin and the relation to ischemia/reperfusion injury.Thromb Haemost 2004;91:506-513.
11. Moore C, Shen XD, Gao F, Busuttil RW, Coito AJ. Fibronectin-alpha4beta1 integrin interactions regulate metalloproteinase-9 expres-sion in steatotic liver ischemia and reperfusion injury. Am J Pathol2007;170:567-577.
12. Lesurtel M, Graf R, Aleil B, Walther DJ, Tian Y, Jochum W, et al. Pla-telet-derived serotonin mediates liver regeneration. Science 2006;312:104-107.
13. Torres L, Garcia-Trevijano ER, Rodriguez JA, Carretero MV, BustosM, Fernandez E, et al. Induction of TIMP-1 expression in rat hepaticstellate cells and hepatocytes: a new role for homocysteine in liverfibrosis. Biochim Biophys Acta 1999;1455:12-22.
14. Yadav SS, Sindram D, Perry DK, Clavien PA. Ischemic preconditioningprotects the mouse liver by inhibition of apoptosis through a caspase-dependent pathway. HEPATOLOGY 1999;30:1223-1231.
15. Hamada T, Duarte S, Tsuchihashi S, Busuttil RW, Coito AJ. Induciblenitric oxide synthase deficiency impairs matrix metalloproteinase-9 ac-tivity and disrupts leukocyte migration in hepatic ischemia/reperfusioninjury. Am J Pathol 2009;174:2265-2277.
16. Yoshidome H, Kato A, Edwards MJ, Lentsch AB. Interleukin-10 sup-presses hepatic ischemia/reperfusion injury in mice: implications of acentral role for nuclear factor kappaB. HEPATOLOGY 1999;30:203-208.
17. Sherr CJ. Cancer cell cycles. Science 1996;274:1672-7.
18. Ohtsubo M, Theodoras AM, Schumacher J, Roberts JM, Pagano M.Human cyclin E, a nuclear protein essential for the G1-to-S phase tran-sition. Mol Cell Biol 1995;15:2612-2624.
19. Michalopoulos GK, DeFrances MC. Liver regeneration. Science 1997;276:60-66.
20. Petrelli A, Circosta P, Granziero L, Mazzone M, Pisacane A, FenoglioS, et al. Ab-induced ectodomain shedding mediates hepatocyte growthfactor receptor down-regulation and hampers biological activity. ProcNatl Acad Sci U S A 2006;103:5090-5095.
21. Han Z, Hendrickson EA, Bremner TA, Wyche JH. A sequential two-step mechanism for the production of the mature p17:p12 form of cas-pase-3 in vitro. J Biol Chem 1997;272:13432-13436.
22. Mullonkal CJ, Toledo-Pereyra LH. Akt in ischemia and reperfusion.J Invest Surg 2007;20:195-203.
23. Sternlicht MD, Werb Z. How matrix metalloproteinases regulate cellbehavior. Annu Rev Cell Dev Biol 2001;17:463-516.
24. Woessner JF Jr. That impish TIMP: the tissue inhibitor of metallopro-teinases-3. J Clin Invest 2001;108:799-800.
25. Dechene A, Sowa JP, Gieseler RK, Jochum C, Bechmann LP, El FoulyA, et al. Acute liver failure is associated with elevated liver stiffness andhepatic stellate cell activation. HEPATOLOGY 2010;52:1008-1016.
26. Bissell DM. Inflammation and hepatic fibrosis. Semin Liver Dis 2010;30:211-214.
27. Gardner J, Ghorpade A. Tissue inhibitor of metalloproteinase (TIMP)-1: the TIMPed balance of matrix metalloproteinases in the central nerv-ous system. J Neurosci Res 2003;74:801-806.
28. Jaeschke H, Farhood A, Smith CW. Neutrophils contribute to ische-mia/reperfusion injury in rat liver in vivo. FASEB J 1990;4:3355-3359.
29. Gough PJ, Gomez IG, Wille PT, Raines EW. Macrophage expressionof active MMP-9 induces acute plaque disruption in apoE-deficientmice. J Clin Invest 2006;116:59-69.
30. Crocker SJ, Whitmire JK, Frausto RF, Chertboonmuang P, SolowayPD, Whitton JL, et al. Persistent macrophage/microglial activation andmyelin disruption after experimental autoimmune encephalomyelitis intissue inhibitor of metalloproteinase-1-deficient mice. Am J Pathol2006;169:2104-2116.
31. Lappas CM, Rieger JM, Linden J. A2A adenosine receptor inductioninhibits IFN-gamma production in murine CD4þ T cells. J Immunol2005;174:1073-1080.
32. Yang SQ, Lin HZ, Lane MD, Clemens M, Diehl AM. Obesity increasessensitivity to endotoxin liver injury: implications for the pathogenesis ofsteatohepatitis. Proc Natl Acad Sci U S A 1997;94:2557-2562.
33. Taub R. Liver regeneration: from myth to mechanism. Nat Rev MolCell Biol 2004;5:836-847.
34. Natarajan A, Wagner B, Sibilia M. The EGF receptor is required forefficient liver regeneration. Proc Natl Acad Sci U S A 2007;104:17081-17086.
35. Mohammed FF, Pennington CJ, Kassiri Z, Rubin JS, Soloway PD,Ruther U, et al. Metalloproteinase inhibitor TIMP-1 affects hepatocytecell cycle via HGF activation in murine liver regeneration. HEPATOLOGY
2005;41:857-867.
36. Hayakawa T, Yamashita K, Tanzawa K, Uchijima E, Iwata K. Growth-promoting activity of tissue inhibitor of metalloproteinases-1 (TIMP-1)for a wide range of cells. A possible new growth factor in serum. FEBSLett 1992;298:29-32.
37. Fata JE, Leco KJ, Moorehead RA, Martin DC, Khokha R. Timp-1 isimportant for epithelial proliferation and branching morphogenesisduring mouse mammary development. Dev Biol 1999;211:238-254.
38. Kopitz C, Gerg M, Bandapalli OR, Ister D, Pennington CJ, Hauser S,et al. Tissue inhibitor of metalloproteinases-1 promotes liver metastasisby induction of hepatocyte growth factor signaling. Cancer Res 2007;67:8615-8623.
39. Schelter F, Kobuch J, Moss ML, Becherer JD, Comoglio PM, Boccac-cio C, et al. A disintegrin and metalloproteinase-10 (ADAM-10) medi-ates DN30 antibody-induced shedding of the met surface receptor.J Biol Chem 2010;285:26335-26340.
12 DUARTE ET AL. HEPATOLOGY, Month 2012

40. Huh CG, Factor VM, Sanchez A, Uchida K, Conner EA, ThorgeirssonSS. Hepatocyte growth factor/c-met signaling pathway is required forefficient liver regeneration and repair. Proc Natl Acad Sci U S A 2004;101:4477-4482.
41. Malhi H, Gores GJ, Lemasters JJ. Apoptosis and necrosis in the liver: atale of two deaths? HEPATOLOGY 2006;43:S31-S44.
42. Cohen GM. Caspases: the executioners of apoptosis. Biochem J 1997;326(Pt 1):1-16.
43. Cory S, Adams JM. The Bcl2 family: regulators of the cellular life-or-death switch. Nat Rev Cancer 2002;2:647-656.
44. Murphy FR, Issa R, Zhou X, Ratnarajah S, Nagase H, Arthur MJ,et al. Inhibition of apoptosis of activated hepatic stellate cells by tissueinhibitor of metalloproteinase-1 is mediated via effects on matrix metal-loproteinase inhibition: implications for reversibility of liver fibrosis.J Biol Chem 2002;277:11069-11076.
45. Guedez L, Stetler-Stevenson WG, Wolff L, Wang J, Fukushima P, MansoorA, et al. In vitro suppression of programmed cell death of B cells by tissueinhibitor of metalloproteinases-1. J Clin Invest 1998;102:2002-2010.
46. Liu XW, Taube ME, Jung KK, Dong Z, Lee YJ, Roshy S, et al. Tissueinhibitor of metalloproteinase-1 protects human breast epithelial cellsfrom extrinsic cell death: a potential oncogenic activity of tissue inhibi-tor of metalloproteinase-1. Cancer Res 2005;65:898-906.
47. Lin H, Chen X, Wang J, Yu Z. Inhibition of apoptosis in rat mesangialcells by tissue inhibitor of metalloproteinase-1. Kidney Int 2002;62:60-69.
48. Boulday G, Fitau J, Coupel S, Soulillou JP, Charreau B. Exogenous tis-sue inhibitor of metalloproteinase-1 promotes endothelial cell survivalthrough activation of the phosphatidylinositol 3-kinase/Akt pathway.Ann N Y Acad Sci 2004;1030:28-36.
49. Frisch SM, Screaton RA. Anoikis mechanisms. Curr Opin Cell Biol2001;13:555-562.
HEPATOLOGY, Vol. 000, No. 000, 2012 DUARTE ET AL. 13