three-dimensional analysis of mitosis and cytokinesis in the … · 2006-11-23 · intermediate...
TRANSCRIPT

4889Research Article
IntroductionGiardia intestinalis, a widespread zoonotic intestinal parasite,has two life-cycle stages: a binucleate, double diploid flagellateor trophozoite form that attaches to the intestinal microvilli,and an infectious cyst form that persists in the environment(Adam, 2001; Gillin et al., 1996). Over several million casesof malabsorptive diarrhea by giardiasis are estimated to occurannually worldwide, and waterborne outbreaks of giardiasisare frequent in areas where unfiltered waters are routinelycontaminated. (Savioli et al., 2006). Although Giardia is wellknown in terms of disease (Adam, 2001; Savioli et al., 2006)and has received much recent attention due to evolutionarycontroversies (Baldauf, 2003; Baldauf et al., 2000; Best et al.,2004; Ciccarelli et al., 2006; Dacks et al., 2002; Graczyk, 2005;Knight, 2004; Sogin et al., 1989), there remains littleknowledge of the assembly and division of the complexgiardial cytoskeletal systems required for the life cycle of theparasite, including the mechanism of mitosis.
Like all diplomonads, Giardia has two diploid nuclei(2n=10), which are both transcriptionally active and identicalin DNA content (Adam et al., 1988; Kabnick and Peattie, 1990;Wiesehahn et al., 1984; Yu et al., 2002). In addition, Giardiais bilaterally symmetrical and possesses a complex anddistinctive microtubule cytoskeleton that establishes anterior-posterior, left-right and dorsal-ventral polarity and,importantly, plays a major role in its virulence (Elmendorf etal., 2003). The microtubule cytoskeleton of the trophozoite, or
intestinal form, is characterized by four main elements: eightflagellar axonemes and basal bodies, the ventral disc, the funisand the median body (Fig. 1A,B). Giardia is unique amongdiplomonads in that it possesses the ventral disc, a novelorganelle composed of an overlapping spiral lamella ofmicrotubules that mediates attachment to the intestinalmicrovilli (or a laboratory substrate), most likely by a suction-based mechanism (Hansen et al., 2006).
To complete cell division, the complex giardial cell mustduplicate and partition both diploid nuclei as well as themultiple cytoskeletal structures. Thus, a detailed analysis ofmitosis in Giardia at both the ultrastructural and molecularlevel is needed to resolve two intriguing questions. First, howdoes Giardia coordinate the division of two equivalent nucleiand a complex microtubule cytoskeleton? Second, howconserved is the mechanism of mitosis in a highly divergentand putatively early branching eukaryote? Understandingchromosome segregation in Giardia, however, has beenhampered principally by the lack of cytological descriptions ofintermediate stages of mitosis, including the inability toidentify the mitotic spindle (Solari et al., 2003). Several priorstudies have sought to identify the stages of mitosis usingprimarily light microscopy and chromatin staining, yet havenot described a mitotic spindle (Cèrva and Nohynkova, 1992;Filice, 1952). Furthermore, recent debate concerning themechanism of giardial cell division has lead to proposals ofunconventional mechanisms of chromosome segregation,
In the binucleate parasite Giardia intestinalis, two diploidnuclei and essential cytoskeletal structures including eightflagella are duplicated and partitioned into two daughtercells during cell division. The mechanisms of mitosisand cytokinesis in the binucleate parasite Giardia arepoorly resolved, yet have important implications for themaintenance of genetic heterozygosity. To articulate themechanism of mitosis and the plane of cell division, we usedthree-dimensional deconvolution microscopy of each stageof mitosis to monitor the spatial relationships of conservedcytological markers to the mitotic spindles, the centromeresand the spindle poles. Using both light- and transmissionelectron microscopy, we determined that Giardia has asemi-open mitosis with two extranuclear spindles thataccess chromatin through polar openings in the nuclearmembranes. In prophase, the nuclei migrate to the cellmidline, followed by lateral chromosome segregation in
anaphase. Taxol treatment results in lagging chromosomesand half-spindles. Our analysis supports a nuclearmigration model of mitosis with lateral chromosomesegregation in the left-right axis and cytokinesis along thelongitudinal plane (perpendicular to the spindles), ensuringthat each daughter inherits one copy of each parentalnucleus with mirror image symmetry. Fluorescence in situhybridization (FISH) to an episomal plasmid confirms thatthe nuclei remain separate and are inherited with mirrorimage symmetry.
Supplementary material available online athttp://jcs.biologists.org/cgi/content/full/119/23/4889/DC1
Key words: Giardia intestinalis, Mitosis, Cytokinesis, Spindle, Basalbody, Centromere
Summary
Three-dimensional analysis of mitosis and cytokinesisin the binucleate parasite Giardia intestinalisMeredith S. Sagolla, Scott C. Dawson*, Joel J. Mancuso and W. Zacheus Cande‡
345 LSA, Department of Molecular and Cell Biology, University of California Berkeley, Berkeley, CA 94720, USA*Present address: Section of Microbiology, University of California Davis, Davis, CA 95616, USA‡Author for correspondence (e-mail: [email protected])
Accepted 22 September 2006Journal of Cell Science 119, 4889-4900 Published by The Company of Biologists 2006doi:10.1242/jcs.03276
Jour
nal o
f Cel
l Sci
ence

4890
including the role of non-spindle microtubular organelles incell division (Benchimol, 2004b; Solari et al., 2003) or ofthe presence of multiple or incongruent planes of division(Benchimol, 2004a; Ghosh et al., 2001; Yu et al., 2002).
Owing to the cytoskeletal complexity of the binucleateGiardia cell, understanding the mechanism of mitosis and celldivision requires the use of three-dimensional (3D) imagingand monitoring of crucial aspects of the spindle such asthe spatial orientation of spindle poles and kinetochoreattachments of chromosomes (at the centromere) to the spindle.Centrin, a ubiquitous protein found in association with basalbodies and centrioles, is located with the centrioles at thespindle poles during mitosis in metazoans (Levy et al., 1996;Salisbury, 1995). To test hypotheses of both the mode andmechanism of mitosis, we imaged mitotic cells in 3D withconserved molecular markers of the mitotic spindle (anti-tubulin), the centromere (cenH3::GFP), and the basal bodiesand the spindle poles (anti-centrin), allowing us to monitor thebehavior of hallmark features of the spindle during mitosis.
This study is the first to observe dual mitotic spindles, toconfirm the role of centrin directly at the spindle poles and toinfer spindle attachment to the chromosomes at the kinetochore(using the centromere histone variant cenH3). By our detailedanalysis of mitotic stages using 3D deconvolution lightmicroscopy and transmission electron microscopy (TEM), wefind that mitosis is semi-open, with two extranuclear centralspindles and microtubules that enter the nuclei through polaropenings in the nuclear envelopes. Chromosomes aresegregated along the left-right (L-R) axis, and cytokinesisoccurs along the longitudinal axis – perpendicular to thespindle – suggesting that daughter cells inherit one copy ofeach parent nucleus.
By monitoring these cytological markers at all stages ofmitosis and cytokinesis, we present a model of cell division inGiardia that: (1) establishes both the spatial orientation and thetimeline of chromosome segregation (mitosis) and cytokinesis;(2) illustrates the timing of the duplication of cytoskeletalstructures including the median body, the ventral disc and theeight flagella; and (3) accounts for the fate of each nucleus
Journal of Cell Science 119 (23)
during cell division. This model of cell division can explainprior studies on nuclear partitioning during cytokinesis, albeitwith an alternative mode of cytokinesis than that originallyproposed (Ghosh et al., 2001).
ResultsTraditionally, identifying individual mitotic stages in Giardiahas been difficult due to the inability to generate synchronizedcultures. Therefore, we first developed a method for enrichingcultures of Giardia for mitotic cells (supplementary materialTable 1) followed by indirect immunofluorescence of keycytological markers to visualize changes in the microtubulecytoskeleton throughout mitosis. To describe mitotic cells weimmunostained mitotic spindles (anti-tubulin) and the spindlepoles (anti-centrin), visualized green fluorescent protein(GFP)-tagged centromeres (cenH3::GFP) and tracked hallmarkfeatures of the spindle during mitosis using 3D deconvolutionmicroscopy (Materials and Methods). Between 50-100 cellswere analyzed for each mitotic stage; representative images arepresented in Fig. 2.
These analyses allow the establishment of a timeline of thecrucial events of mitosis and the determination of the plane ofnuclear division and cytokinesis. Although we emphasize thedynamics of chromosome segregation during mitosis at highlevels of resolution, we also present a timeline of theduplication and partitioning of major cytoskeletal elementsincluding the eight flagella, the median body and the ventraldisc, albeit in less detail.
InterphaseMicrotubule cytoskeleton: Giardia trophozoites have anelaborate microtubule cytoskeleton, consisting of four majorstructural microtubule arrays: a ventral disc, flagellaraxonemes with basal bodies, the funis and the median body (anenigmatic microtubular array of unknown function) (see Fig.1A,B) (see also Elmendorf et al., 2003). In the imagespresented here, the ventral disc is weakly stained relative to theother microtubule organelles.
Chromosomes: in interphase, chromatin is decondensed, as
Fig. 1. Giardia has two equivalent nuclei and a complexmicrotubule cytoskeleton. (A) The microtubule cytoskeleton ofthe trophozoite is characterized by eight flagellar axonemesand basal bodies (afl, anterior; c, caudal; pfl, posterior; vfl,ventral), the ventral adhesive disc (vd), the funis (fn) and themedian body (mb) (diagramed in A). All basal bodies arebetween the nuclei. (B) Anti-tubulin labeling reveals themicrotubule arrays, including the eight flagella, and medianbody. The ventral disk is more weakly stained than the otherstructures. Although images of Giardia are frequentlypresented as two-dimensional projections in dorsal perspectiveas in A, describing cell division in Giardia requires analysis ofcell morphology in three dimensions. (C) The cell is shown in3D from a side angle. The spatial positions of the four nucleifollowing nuclear division are shown arrayed along the left-right (L-R) and dorsal-ventral (D-V) axes. Three possibleplanes of cytokinesis are represented in C; longitudinal[anterior-posterior (A-P)], transverse (L-R) and frontal (D-V).The dorsal-posterior nuclei are marked with asterisks in C,D.
Jour
nal o
f Cel
l Sci
ence

4891Mitosis in Giardia
indicated by the uniform fluorescence of 4,6-diamino-2-phenylindole (DAPI)-stained nuclei (Fig. 2A,D). Althougheukaryotic centromeric sequences vary greatly at thenucleotide level, centromeric nucleosomes contain thespecialized histone H3 variant cenH3 (Malik and Henikoff,2003). To visualize the centromeres of Giardia chromosomes,we identified the cenH3 homolog in Giardia and constructeda GFP fusion protein (S.C.D. and W.Z.C., unpublisheddata). In interphase, cenH3::GFP foci were seen throughoutthe DAPI-stained nuclei, indicating that the cenH3:GFPmarks discrete centromeric loci on the chromosomes (Fig.2A).
Basal bodies: to determine the role of basal bodies inorganizing the giardial mitotic spindles, we monitored the
movement of flagellar basal bodies during interphase andmitosis using anti-centrin immunostaining. The basal bodies ofall eight flagella are located between the two adjacent nucleiin the anterior region of the cell. In interphase trophozoites, weobserved that centrin localized to two clusters between the twonuclei, colocalizing with the flagellar basal bodies (Fig. 2D) ashas been shown previously (Belhadri, 1995; Correa et al.,2004; Meng et al., 1996).
ProphaseMitotic prophase is characterized by extensive chromatincondensation and the nucleation of spindle microtubules. Asin most eukaryotes, we found that individual giardialchromosomes could be resolved following chromatin
condensation in mitotic prophase (Fig. 2B). In prophase,each chromosome contained a single cenH3::GFP focus,indicating that each of the ten chromosomes in eachnucleus has a single centromeric locus. Spindlemicrotubule nucleation was also initiated duringprophase: microtubules first appeared between the twonuclei near the flagellar basal bodies and extendedaround each nucleus individually (Fig. 2B).
An unanticipated repositioning of the nuclei occurredthroughout prophase. The two nuclei migrated to the cellmidline one nucleus on top of the other along the dorsal-ventral axis, with the ventral nucleus slightly anterior tothe dorsal nucleus (supplementary material Movie 1).Three-dimensional reconstructions of these cells showedthe two nuclei remained separate from one another(supplementary material Movie 1). At the completion ofnuclear migration individual chromosomes could bedistinguished. During nuclear migration the spindle
Fig. 2. During mitosis two autonomous spindles areresponsible for chromosome segregation after nuclei migrateto the cell center. The centromere-specific histone cenH3marks centromeres (cenH3:GFP in green, A-C,G-I), andcentrin antibodies mark spindle poles (green, D-F,J-L). TAT1(anti-tubulin) labels the microtubule cytoskeleton includingthe mitotic spindles (all cells, in red) and DAPI labelschromatin (all cells, in blue). Behavior of left (red) and right(blue) nuclei is diagramed above each stage. (A) Interphase;centromeres are discreet foci in each nucleus. (D) Interphase;centrin stains two foci in association with the flagellar basalbodies between the two nuclei. (B) Prophase; spindlemicrotubules appear between the two nuclei and extendaround each nucleus. Centromeres are one spot on eachcondensed chromosome. (E) Prophase; four centrin foci areassociated with the forming spindles. (C) Metaphase;following nuclear migration the nuclei become stacked alongthe dorsal-ventral axis. The microtubules organized into twobipolar spindles. (F) Metaphase; centrin foci are at the fourspindle poles. (G) Anaphase; chromatin is segregated tospindle poles along the left-right axis of the cell withcentromeres clustered at the spindle poles. (J) Anaphase;centrin remains at the spindle poles. (H) Telophase; a linearmicrotubule structure replaces the bipolar spindles andcentromeres remain clustered. (K) Telophase; centrin foci arebetween the two nuclei of each daughter. (I) Cytokinesis;chromatin decondenses and the cleavage furrow forms at theanterior end, creating a heart-shaped cell. (L) Cytokinesis;centrin stains two foci between the nuclei. Cytokinesis dividesthe cell into left and right halves. Bar, 2 �m.
Jour
nal o
f Cel
l Sci
ence

4892
microtubules continued to elongate, encompassing eachnucleus by the end of prophase.
Duplication of the centrin foci also occurred duringprophase. At the onset of prophase and spindle assembly thenumber of centrin foci increased from two to four as the resultof either duplication or separation of the basal bodies (Fig. 2E).During nuclear migration the centrin foci were present at thesites of microtubule nucleation. These foci moved around theperiphery of the nucleus as the spindle microtubules continuedto elongate and the nuclei migrated to the center. When the twonuclei were stacked, two centrin foci, each with its owncomplement of microtubules, were positioned on oppositesides of each nucleus.
MetaphaseUpon completion of nuclear migration, the microtubulessurrounding each nucleus formed two independent bipolarspindles, stacked in the dorsal-ventral plane (Fig. 2C,F). Theopposing poles of each spindle were oriented along the left-right axis of the cell. The chromatin was clustered tightly inthe center of each spindle axis so that individual chromosomeswere not visible (Fig. 2C). We were unable to observe acanonical metaphase alignment of centromeres along ametaphase plate. At this time, centrin localized to each of thefour spindle poles (Fig. 2F).
Anaphase A and BChromosome segregation in both nuclei occurred in twostages: chromatid segregation to the spindle poles in anaphaseA, followed by spindle elongation along the left-right axis ofthe cell in anaphase B (Fig. 2G). In Giardia, anaphase Ainitiates first in one nucleus (data not shown), howeveranaphase B occurs simultaneously between the two nuclei. Asa result of both nuclear migration and lateral chromosomesegregation, the sister chromatids from each nucleus weresegregated to opposite sides (L-R) of the cell. At thecompletion of anaphase, nuclei of different parental originreside on opposite sides of the cell with the daughters of thedorsal nucleus remaining dorsal and slightly posterior withrespect to the daughter nuclei of the ventral nucleus.
During anaphase A, the cenH3::GFP localized to the leadingedge (near the spindle pole) of the segregating DNA, abehavior characteristic of kinetochore attachment tomicrotubules. The cenH3::GFP foci remained tightly clusteredtogether at the spindle poles during anaphase B (Fig. 2G). Thisdistribution pattern implicates centromeres as the site of
microtubule attachment during chromosome segregation inGiardia as in other eukaryotes. Centrin foci remained at thespindle poles throughout anaphase A and anaphase B (Fig. 2J).
Telophase and cytokinesisIn telophase, a microtubule bundle with unfocused endsextended between the nuclei on each side of the cell replacingthe bipolar spindle arrays (Fig. 2H). We interpret this structureas the remaining spindle microtubules following the loss offocused spindle poles. We are not able to rule out formation ofthis structure from de novo microtubule polymerization;however, a de novo origin is unlikely given that intermediatestructures were not observed. The single cenH3::GFP fociobserved in the daughter nuclei indicated that the centromeresremained clustered in telophase nuclei.
During telophase, the two centrin foci on each side of thenuclei moved from their anaphase position near the cellperiphery to their position between each pair of nuclei as seenin interphase (Fig. 2K). By the onset of cytokinesis, the DNAwas decondensed, cenH3::GFP foci were seen throughout thenucleus, the nuclei were adjacent to one another and allcytoskeletal structures were regenerated (described below).
To complete cytokinesis, a furrow formed at the anterior endof the cell creating a heart-shaped four nuclear stage cell (Fig.2I,L). The furrow progressed from anterior to posterior, in thelongitudinal plane, separating the left and right sides of theheart into the two daughter cells. As a result, each daughter cellinherits two nuclei each derived from one left and one rightnucleus of the mother cell.
Patterns of nuclear inheritance using FISH of anepisomal plasmid in mitotic nucleiTo determine the pattern of nuclear partitioning andinheritance, we used fluorescence in situ hybridization (FISH)to visualize an episomal plasmid in the cenH3::GFP strain,which is maintained by one nucleus in each Giardiatrophozoite (Ghosh et al., 2001; Singer et al., 1998; Yu et al.,2002). In each cell, we observed the episomal plasmid FISHsignal in only one nucleus (either the left or the right) aspreviously reported (Ghosh et al., 2001). Since a nucleuscontains more than one episome, by using deconvolutionmicroscopy we are able to detect multiple discrete FISHsignals in the labeled nucleus. In prophase nuclei, during thelate stages of nuclear migration, the FISH signal was observedonly in one nucleus (see dorsal nucleus in Fig. 3B,B’). Givenour previous observation that daughter nuclei from each
Journal of Cell Science 119 (23)
Fig. 3. FISH to an episomalplasmid maintained in onenucleus shows nuclei are inheritedwith mirror image symmetry andthat the nuclei never mix.(A) Interphase; the FISH probehybridizes to the episomecontained in the right nucleus.(B) In the metaphase nuclei,stacked on top of each other in thecell center, FISH signal isdetected in the dorsal nucleus.(B’) Projection of the first eight z-sections, dorsal nucleus, of the cell seen in B contains the FISH signal. (B’’) Projection of the last eight z-sections, ventral nucleus, of the cell shown in B shows the FISH signal is excluded. (C) During cytokinesis, one nucleus in each daughter islabeled with mirror image symmetry between the two cells. Red, DAPI; green, FISH. Bars, 2 �m.
Jour
nal o
f Cel
l Sci
ence

4893Mitosis in Giardia
nucleus are segregated laterally, we would expect only onenucleus in each daughter cell to contain the episome. Aspredicted, in heart-shaped cells entering cytokinesis, only onenucleus in each future daughter cell showed the episomalplasmid FISH signal (Fig. 3C); the two cells showed mirrorimage symmetry of the labeled nuclei as has been previouslyreported (Ghosh et al., 2001).
Giardia has a semi-open mitosisWe used TEM to visualize spindle ultrastructure, to assess theintegrity of the nuclear envelope during mitosis and todetermine the identity of the spindle pole. TEM images of bothearly and late mitotic nuclei showed that the spindles areextranuclear and that the nuclear envelope remains intactthroughout mitosis (Fig. 4A). Microtubules originating from
each spindle pole form a sheath around the nucleusoutside of the nuclear envelope. The nuclear envelopeelongates with the spindle in anaphase B, extendingbetween the spindle poles of fully elongated spindles.The persistence of the nuclear envelope throughoutmitosis precludes mixing of the chromatin betweennuclei. Furthermore, we did not observe fusion of the twonuclei at any mitotic stage.
Although the nuclear envelope is present throughout,this type of mitosis can be classified as semi-open. Theright spindle pole (outlined by the box in Fig. 4A) isshown at greater magnification in Fig. 4B. Cytoplasmicmicrotubules from spindle poles passed through largeopenings in the nuclear membrane (Fig. 4B). In the late-stage (anaphase B) nuclei, the internal (presumablykinetochore) microtubules extended a few microns intothe nucleus near the chromatin, seen as an electron-denseregion near the pole (Fig. 4B).
We observed both basal bodies as well as centrinlocalization at each of the four spindle poles. Eachspindle pole was associated with at least one axoneme(Fig. 4A,C). We imaged at least two basal bodiespositioned at right angles to one other at each spindlepole: one basal body is in cross-section, whereas thesecond basal body with associated axoneme – at a moreperipheral location relative to the spindle pole – is seenin longitudinal section (Fig. 4A,C). The basal bodycross-section revealed a ring of doublet microtubules thatlack a central microtubule pair (Fig. 4C), resemblingthe transition zone of a basal body as described inChlamydomonas (O’Toole et al., 2003). Spindlemicrotubules radiated from one of the basal bodies (seenin cross-section) in proximity to spindle poles (Fig. 4C).In telophase the flagellar basal bodies were positionedbetween the two daughter nuclei on each side of the cell(Fig. 4D).
Microtubule stabilization disrupts mitotic spindlesand causes chromosome missegregationPrior studies have suggested that chromosomesegregation in Giardia occurs through a spindle-independent mechanism (Solari et al., 2003). Todetermine conclusively whether dynamic microtubulesare required for chromosome segregation, we treatedmitotic cells with taxol [which binds to the �-tubulinsubunit of the tubulin heterodimer in polymerizedmicrotubules (Parness and Horwitz, 1981; Rao et al.,1995)] and evaluated segregation defects. Althoughgiardial tubulin subunits are highly divergent, the �-tubulin subunit has retained the amino acid sequenceassociated with taxol binding (see supplementarymaterial Fig. S1). Nearly 100% of mitotic cells treatedwith either 10 �M or 20 �M taxol (n>100) showeddramatic defects in chromosome segregation and
Fig. 4. As shown by TEM, the mitotic spindle is semi-open and basalbodies with axonemes are present at the four spindle poles. (A) Cross-section of a mitotic nucleus; microtubules form a sheath surrounding thenucleus. The nuclear envelope is intact except for large openings at thepoles. Bar, 200 nm. (B) High-magnification view of area outlined by boxin A. Cytoplasmic microtubules enter the nucleus through a large openingin the nuclear membrane. Bar, 50 nm. (B’) Diagram of nuclear membrane(blue) and microtubules (red) seen in B. (C) Spindle microtubules radiatefrom the basal body seen in cross-section, which organizes the spindlemicrotubules. Bar, 300 nm. (D) Basal bodies are located between the twodaughter telophase nuclei. One basal body with associated axoneme isseen in longitudinal section, the others are in cross-section. Bar, 400 nm.bb, basal body; fl, flagellum; n, nucleus.
Jour
nal o
f Cel
l Sci
ence
4894
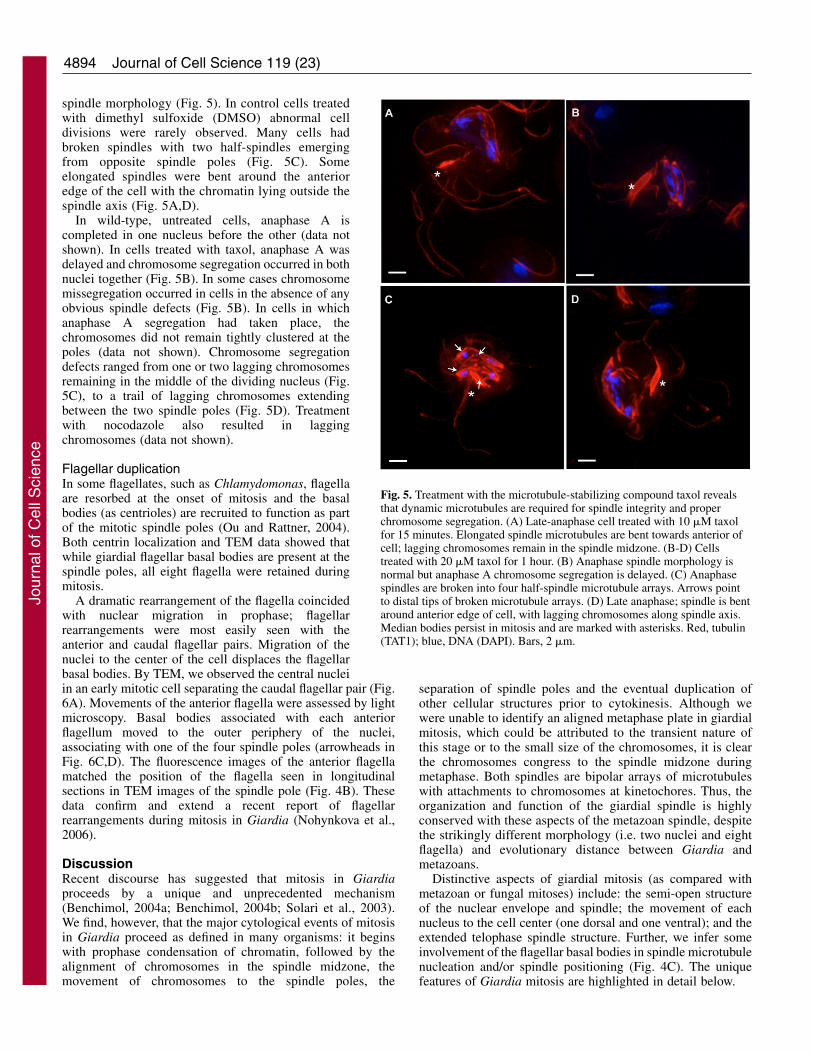
spindle morphology (Fig. 5). In control cells treatedwith dimethyl sulfoxide (DMSO) abnormal celldivisions were rarely observed. Many cells hadbroken spindles with two half-spindles emergingfrom opposite spindle poles (Fig. 5C). Someelongated spindles were bent around the anterioredge of the cell with the chromatin lying outside thespindle axis (Fig. 5A,D).
In wild-type, untreated cells, anaphase A iscompleted in one nucleus before the other (data notshown). In cells treated with taxol, anaphase A wasdelayed and chromosome segregation occurred in bothnuclei together (Fig. 5B). In some cases chromosomemissegregation occurred in cells in the absence of anyobvious spindle defects (Fig. 5B). In cells in whichanaphase A segregation had taken place, thechromosomes did not remain tightly clustered at thepoles (data not shown). Chromosome segregationdefects ranged from one or two lagging chromosomesremaining in the middle of the dividing nucleus (Fig.5C), to a trail of lagging chromosomes extendingbetween the two spindle poles (Fig. 5D). Treatmentwith nocodazole also resulted in laggingchromosomes (data not shown).
Flagellar duplicationIn some flagellates, such as Chlamydomonas, flagellaare resorbed at the onset of mitosis and the basalbodies (as centrioles) are recruited to function as partof the mitotic spindle poles (Ou and Rattner, 2004).Both centrin localization and TEM data showed thatwhile giardial flagellar basal bodies are present at thespindle poles, all eight flagella were retained duringmitosis.
A dramatic rearrangement of the flagella coincidedwith nuclear migration in prophase; flagellarrearrangements were most easily seen with theanterior and caudal flagellar pairs. Migration of thenuclei to the center of the cell displaces the flagellarbasal bodies. By TEM, we observed the central nucleiin an early mitotic cell separating the caudal flagellar pair (Fig.6A). Movements of the anterior flagella were assessed by lightmicroscopy. Basal bodies associated with each anteriorflagellum moved to the outer periphery of the nuclei,associating with one of the four spindle poles (arrowheads inFig. 6C,D). The fluorescence images of the anterior flagellamatched the position of the flagella seen in longitudinalsections in TEM images of the spindle pole (Fig. 4B). Thesedata confirm and extend a recent report of flagellarrearrangements during mitosis in Giardia (Nohynkova et al.,2006).
DiscussionRecent discourse has suggested that mitosis in Giardiaproceeds by a unique and unprecedented mechanism(Benchimol, 2004a; Benchimol, 2004b; Solari et al., 2003).We find, however, that the major cytological events of mitosisin Giardia proceed as defined in many organisms: it beginswith prophase condensation of chromatin, followed by thealignment of chromosomes in the spindle midzone, themovement of chromosomes to the spindle poles, the
separation of spindle poles and the eventual duplication ofother cellular structures prior to cytokinesis. Although wewere unable to identify an aligned metaphase plate in giardialmitosis, which could be attributed to the transient nature ofthis stage or to the small size of the chromosomes, it is clearthe chromosomes congress to the spindle midzone duringmetaphase. Both spindles are bipolar arrays of microtubuleswith attachments to chromosomes at kinetochores. Thus, theorganization and function of the giardial spindle is highlyconserved with these aspects of the metazoan spindle, despitethe strikingly different morphology (i.e. two nuclei and eightflagella) and evolutionary distance between Giardia andmetazoans.
Distinctive aspects of giardial mitosis (as compared withmetazoan or fungal mitoses) include: the semi-open structureof the nuclear envelope and spindle; the movement of eachnucleus to the cell center (one dorsal and one ventral); and theextended telophase spindle structure. Further, we infer someinvolvement of the flagellar basal bodies in spindle microtubulenucleation and/or spindle positioning (Fig. 4C). The uniquefeatures of Giardia mitosis are highlighted in detail below.
Journal of Cell Science 119 (23)
Fig. 5. Treatment with the microtubule-stabilizing compound taxol revealsthat dynamic microtubules are required for spindle integrity and properchromosome segregation. (A) Late-anaphase cell treated with 10 �M taxolfor 15 minutes. Elongated spindle microtubules are bent towards anterior ofcell; lagging chromosomes remain in the spindle midzone. (B-D) Cellstreated with 20 �M taxol for 1 hour. (B) Anaphase spindle morphology isnormal but anaphase A chromosome segregation is delayed. (C) Anaphasespindles are broken into four half-spindle microtubule arrays. Arrows pointto distal tips of broken microtubule arrays. (D) Late anaphase; spindle is bentaround anterior edge of cell, with lagging chromosomes along spindle axis.Median bodies persist in mitosis and are marked with asterisks. Red, tubulin(TAT1); blue, DNA (DAPI). Bars, 2 �m.
Jour
nal o
f Cel
l Sci
ence

4895Mitosis in Giardia
Spindle structureBased on ultrastructural analyses, mitotic spindles in protistshave been classified using several cytological features: thecontinuity of the nuclear envelope during mitosis (open, semi-open and closed); the position of the spindle relative to thenuclear envelope (pleuromitosis versus orthomitosis); and theposition of chromosomes relative to the spindle axis (Raikov,1994). The nuclear envelope sequesters chromatin throughoutmost of the cell cycle. In most plants and animals, a completenuclear envelope breakdown occurs during prophase, allowingmicrotubules direct access to chromatin. In protists, bothkinetoplastids (e.g. trypanosomes) and heteroloboseans (e.g.Naegleria gruberi) retain an intact nuclear envelope, or closedmitosis, with an intranuclear spindle, a trait that is shared withboth fission and budding yeast (O’Toole et al., 2003; Ogbadoyiet al., 2000; Schuster, 1975; Winey et al., 1995). Thetrichomonads also have a closed mitosis with an extranuclearspindle that attaches to kinetochores embedded in the nuclearenvelope (Brugerolle, 1975b; Gomez-Conde et al., 2000).
During giardial mitosis, the nuclear envelope also acts as abarrier between cytoplasmic microtubule arrays and chromatin.By TEM, we provide the first evidence that Giardia has a semi-open mitosis in which microtubules from the two extranuclearspindles penetrate the nucleus through polar openings in thenuclear envelope (Fig. 4B). This is similar to a description ofsemi-open mitosis in a related diplomonad, Hexamita inflata(Brugerolle, 1975a), and is consistent with the diversity of
mitotic spindle structure in protists. Owing to theubiquity of extranuclear spindles in protists, theancestral state of eukaryotic mitosis may be similar toGiardia and trichomonads, an extranuclear spindle thatinteracts with chromatin either through a semi-opennucleus or across an intact nuclear envelope. In thisscenario, both closed mitosis with intranuclear spindles(yeasts) and open mitosis (plants and metazoans) arederived mitotic forms.
Evidence for kinetochore microtubules inchromosome segregationThe mitotic spindle links the microtubule cytoskeletonto chromosomes through kinetochore microtubules tofacilitate chromosome segregation. Two experimentalobservations strongly support the role of kinetochoremicrotubules in chromosome segregation in Giardia: thecenH3::GFP localization pattern in mitosis and thepresence of intranuclear microtubules seen by TEM.
A histone H3 variant cenH3 is found exclusively atcentromeres, and has been proposed as a universalmarker of the eukaryotic centromere (Sullivan et al.,2001). Using a GFP-tagged cenH3 strain, we haveshown that cenH3::GFP localizes to a discrete focus oneach prophase chromosome, allowing us to visualizethe giardial centromere and demonstrate that giardialchromosomes are monocentric (S.C.D. and W.Z.C.,unpublished data). During prophase in other eukaryotes,centromeres containing cenH3 are required to build themitotic kinetochore by recruiting motors, checkpointproteins and additional structural elements (Van Hooseret al., 2001). In giardial mitosis, the clustering of cenH3-marked centromeres at the leading edge of segregatingDNA (Fig. 2G) implies that chromosome segregation ismediated by microtubule attachments at the kinetochore.
The number of microtubules attached to the kinetochoreranges from a single kinetochore microtubule per chromosomein budding yeast to more than 20 per chromosome in somemetazoans (McDonald et al., 1992; Winey et al., 1995).Although we have not directly observed kinetochore-microtubule associations, given the number of microtubulesseen in a single TEM section and the number of kinetochoresavailable for attachment in the nucleus, i.e. ten, we predictmore than one microtubule is attached per kinetochore inGiardia.
Microtubule-stabilizing compounds generate laggingchromosomes on the anaphase spindle in Giardia. Byinhibiting microtubule depolymerization using taxol, we wereable to generate many of the same anaphase B spindle defectsas previously observed in metazoans (Amin-Hanjani andWadsworth, 1991; De Brabander et al., 1986; Jordan et al.,1993). Primarily, we found that taxol treatment caused the lossof interzonal microtubules, resulting in the formation of twohalf-spindles. This observation provides further evidencefor the role of kinetochore microtubules in chromosomesegregation and demonstrates the bipolar organization ofspindle microtubules.
A role for the flagellar basal bodies in spindleorganization and positioningThe morphology of the microtubule organizing centers
Fig. 6. During mitosis the flagella and basal bodies are displaced by themigrating nuclei and become associated with the spindle poles. (A) TEMof a metaphase mitotic cell shows that the caudal flagella (cfl) aredisplaced following nuclear migration. Bar, 1 �m. (B) Interphase; arrowmarks position of basal bodies of anterior flagella. (C) Early prophase;arrow marks position of basal bodies, located at the periphery of themigrating nuclei. (D) Metaphase; the anterior flagella is wrapped aroundthe dorsal nucleus, and the basal body is associated with the right spindlepole. Median body marked by asterisk. Bar, 2 �m.
Jour
nal o
f Cel
l Sci
ence

4896
(MTOCs) of mitotic spindles is diverse among eukaryotes. Thecentrosomes of metazoan cells consist of a pair of centriolessurrounded by pericentriolar material containing factors, suchas gamma tubulin, required for microtubule nucleation (Ou andRattner, 2004). Yeasts lack centrioles but are able to nucleateand organize the mitotic spindle using spindle pole bodies(SPBs) (Jaspersen and Winey, 2004). Flagellated protists suchas Chlamydomonas recruit flagellar basal bodies, structurallyrelated to centrioles, to function as part of the MTOC at thespindle poles following resorption of the flagella. These basalbodies of Chlamydomonas organize the spindle microtubulesin a bipolar array and also appear to play a role in spindlepositioning (Coss, 1974; Ehler et al., 1995). The association offlagellar basal bodies with the spindle poles may serve toensure equal partitioning of basal bodies in each division cycle,thereby preserving the number of flagella in each daughter cell(Dutcher, 2003; Marshall and Rosenbaum, 2000; Heath, 1980;Lechtreck and Grunow, 1999; Wright et al., 1989).
In Giardia, eight basal bodies are inherited by each daughtercell during a mitotic division that includes two spindles and fourspindle poles. How are the 16 giardial basal bodies accountedfor during division and what roles do they play in spindleformation, positioning and cell-polarity determination? Wehypothesize that the polarity of each daughter cell is establishedthrough the association of the eight basal bodies with the fourspindle poles during mitosis. This proposal is based on ouranalysis of spindle structure using TEM, which demonstratesthe association of multiple basal bodies with spindle poles (Fig.4). We presume that one basal body at each pole acts as thecentral structural component of the MTOC (as indicated by thespindle microtubules radiating from the basal body seen incross-section, see Fig. 4C). We predict that careful analysis of�-tubulin in the intermediate stages of mitosis as outlined in ourstudy would reveal �-tubulin as part of the spindle-polecomplex. In a recent study, �-tubulin localization was detectedat basal bodies both before and after nuclear division, althoughspindles were not directly observed (Nohynkova et al., 2000).We found that a second basal body is found at the periphery ofthe spindle-pole region, peripheral to the focused spindlemicrotubules (Fig. 4C). This basal body, with its associatedflagellum, may play an indirect role in spindle nucleation andorganization; its association with a spindle pole may ensure itsproper segregation to the daughter cells. Our data support therecent description of movements of flagellar complexes duringcell division (Nohynkova et al., 2006).
Outside of spindle nucleation, the association of flagellarbasal bodies with spindle poles could establish and maintain cellpolarity in each generation. We propose that specificcombinations of two axonemes and associated basal bodies ateach pole confer a unique positional identity to each of the fourspindle poles. This is supported by both centrin immunostaining(Fig. 2K) and the TEM of telophase cells (Fig. 4D), whichreveal flagellar basal bodies positioned between the twodaughter nuclei on each side of the cell. The specific associationof the eight basal bodies (with different developmental origins)with the four individual spindle poles can provide spatial cuesfor the proper establishment of cell polarity after mitosis.
Nuclear repositioning, cytokinesis and the orientation ofdaughter cellsThe bilaterally symmetrical Giardia trophozoite is complex,
and one would expect mitosis and cytokinesis to be tightlycontrolled and spatially defined processes. Based on ouranalysis of mitotic stages, we present a new model of giardialmitosis, the nuclear migration model, named for therepositioning of the two nuclei during prophase (Fig. 2C). Inthis model, the left and right nuclei are inherited with mirrorimage symmetry in the two daughter cells (Fig. 7). Althoughthis pattern of nuclear inheritance has been predicted byprevious studies (Benchimol, 2004a; Benchimol, 2004b; Cèrvaand Nohynkova, 1992; Filice, 1952; Ghosh et al., 2001; Yu etal., 2002), the nuclear migration model differs in how themirror image symmetry of daughter nuclei is achieved. Toclarify the possible planes of cell division the axes of mitosisand cytokinesis are defined in Fig. 1C.
Three primary observations provide empirical support forour proposed model of mitosis: nuclear migration duringprophase, cytokinesis perpendicular to the spindle axes andmirror image symmetry of nuclei using FISH. To illustrate this,we find that following prophase nuclear migration, nucleardivision occurs along the left-right axis. The two nuclei remainseparate during this time and thus do not exchange geneticinformation. Second, cytokinesis occurs in the longitudinalaxis perpendicular to the axis of nuclear division.
Although it has been proposed that Giardia divides alongmultiple division planes depending on whether the cells areattached or unattached to a substrate (Benchimol, 2004a),this is not consistent with our observations. We examinedpopulations of cells that were both attached and unattached atthe time of fixation, and determined that chromosomesegregation only occurs along the left-right axis of the cell withcytokinesis perpendicular to that plane. In the course of ourexperiments hundreds of mitotic cells were imaged and wenever detected cytokinesis occurring in the dorsal-ventralplane.
Despite differences in terms of the timing and mechanismof cell division among diverse eukaryotes, the cell divisionplane is always perpendicular to the axis of chromosomesegregation as defined by the mitotic spindle (Balasubramanianet al., 2004). Our model for cytokinesis, occurring in thelongitudinal plane perpendicular to the spindle axis, isconsistent with patterns of cytokinesis in other eukaryotes.
Finally, we show that nuclei are inherited with mirror imagesymmetry. In cytokinesis, we observed mirror image symmetryof nuclei containing the episomal FISH signal, as has beenpreviously reported (Ghosh et al., 2001). Given that cytokinesisoccurs in the longitudinal plane, the only outcome for nuclearidentity is mirror image symmetry between the daughter cells.
Prior attempts to define the pattern of nuclear inheritancehave also used FISH of an episomal plasmid to track the fateof either nucleus during cell division, but did not look atintermediate stages of mitosis, particularly during the prophasenuclear migration or the later four nuclear pre-cytokinesis stage(Ghosh et al., 2001; Yu et al., 2002). In addition, these studiespresumed that cytokinesis occurs in the frontal plane or dorsal-ventral axis. Ghosh et al. reported that 50% of the FISH signalwas present in either the left or right nucleus and in one nucleusper daughter in heart-shaped cells undergoing cytokinesis(Ghosh et al., 2001). Based on these observations, Ghosh andcolleagues concluded that frontal cell division occurred,resulting in daughter cells in a ventral-ventral orientation.Alternatively, Yu et al. observed that the same nucleus always
Journal of Cell Science 119 (23)
Jour
nal o
f Cel
l Sci
ence

4897Mitosis in Giardia
contained the episomal plasmid, concluding that the cells werein a dorsal-ventral orientation (Yu et al., 2002). The pattern ofFISH we observe supports the observations of Ghosh et al.,wherein the two nuclei are inherited with mirror imagesymmetry. The fact that no cells contained two labeled nucleiin this or prior studies implies that: (1) there is no nuclearfusion during giardial mitosis; and (2) daughters inherit onecopy of each parental nucleus in each generation.
Implications and possible mechanisms of the nuclearmigration model for the maintenance of heterozygosityin the two nucleiBeyond establishing the plane of cytokinesis, the mode ofGiardia cell division has important implications that bear onthe maintenance of a unique genetic identity for each nucleus.Based on our observations, the nuclei remain physically andgenetically distinct from one another with daughter cellsinheriting one copy of both nuclei. Thus, if neither nuclearfusion events nor meiosis occur, asexually dividingGiardia should theoretically accumulate substantial allelicheterozygosity between each nucleus in a short time, which
has not been observed (Baruch et al., 1996; Kabnick andPeattie, 1990; Lu et al., 1998; Yu et al., 2002). The recentidentification of putative meiotic genes in Giardia (Ramesh etal., 2005) has raised the possibility that rare nuclear fusionsor meiotic events provide a mechanism for chromosomalrecombination and reduction of such heterozygosity. Yetcanonical meiosis, including the presence of characteristiccytological evidence such as a synaptonemal complex, has notbeen directly observed in Giardia trophozoites. One possibilityis that two genetically disparate nuclei could be maintained inGiardia, if during rare points in the life cycle (i.e. encystation)the two nuclei could fuse and homogenize genetic material.Alternatively, the inheritance of genetic material from bothnuclei may somehow be important for cell survival.
What is the mechanism of nuclear migration? Centrin, acalcium binding protein associated with basal bodies andcentrosomes of eukaryotic cells (Salisbury, 1995), mayrepresent a link between flagellar segregation and nuclearmigration. In algal cells, centrin is a major component of thenuclear basal body connector, a fibrous network that physicallylinks the basal bodies to the nucleus (Brugerolle and Mignot,
A-PD-V D-D–V-V
A
P
L R
D
V
*
A-P D-V
† ‡
Fig. 7. The nuclear migration model, to the left and shadedin green, is proposed from this analysis (Figs 2-4). Eachmodel presents three events that determine the pattern ofnuclear inheritance in each division cycle: the plane ofnuclear division, the plane of cytokinesis and theorientation of the daughter cells relative to one another.The schematic depicts the relative positions of the left(red) and right (blue) nuclei before and after mitosis andcytokinesis as seen from the dorsal side of the cell (seealso Fig. 1). Following nuclear migration to the cellmidline, the left-right (L-R) axis of nuclear division yieldstwo daughter nuclei, one from each parental nucleuslocated on opposite (L-R) sides of the cell. Thelongitudinal plane of cytokinesis results in inheritance ofnuclei with mirror image symmetry in the daughter cells.Dorsal-ventral (D-V) cytokinesis leads to inheritance oftwo nuclei of the same parental origin and is not supportedby our data or by previous FISH studies (Ghosh et al.,2001; Yu et al., 2002). Models outlined in previous studiesare presented on the right. Nuclear division was assumedto be in the D-V plane, by an unspecified mechanism,resulting in nuclei of the same parental origin residing onone side (L-R) of the cell (Filice, 1952; Ghosh et al., 2001;Yu et al., 2002; Benchimol, 2004a). Division along thelongitudinal axis results in each daughter cell inheritingnuclei of the same parental origin. Cytokinesis in thefrontal plane (Ghosh et al., 2001; Yu et al., 2002;Benchimol, 2004a) (Fig. 1C) results in either identical ormirror image symmetry of the two nuclei in daughter cellsdepending on the orientation of the cells to one another. A-P, anterior-posterior; D-D, dorsal-dorsal; D-D–V-V, dorsal-dorsal–ventral-ventral. †(Yu et al., 2002; Benchimol,2004a). ‡(Ghosh et al., 2001; Benchimol, 2004a).
Jour
nal o
f Cel
l Sci
ence

4898
2003; Marshall and Rosenbaum, 2000; Salisbury et al., 1988;Wright et al., 1989; Wright et al., 1985). In interphase, weobserved centrin immunolocalization solely at basal bodies(Fig. 2) in contrast to prior studies (Belhadri, 1995; Meng etal., 1996). During mitosis, we determined that centrin localizedonly to basal bodies associated with the four spindle poles (Fig.2). Although centrin is not associated with all eight basalbodies, centrin fibers may connect spindle pole-associatedbasal bodies with other basal bodies in preparation forsegregation during mitosis. Thus, basal body migration andnuclear migration during mitosis may be coordinated events,facilitated by the centrin-dependent attachment of basal bodiesto the nuclear envelope.
ConclusionsDiplomonads have been proposed to represent the earliestdiverging lineage of extant eukaryotes, based on single rRNAand single and/or concatenated protein phylogenies using thebest available phylogenetic methods when an archaealoutgroup is included (Baldauf, 2003; Baldauf et al., 2000;Ciccarelli et al., 2006; Van de Peer et al., 2000). In anyevolutionary analysis, diplomonads remain monophyletic andlack any statistically supported affiliation with late-emerginggroups of eukaryotes or with the majority of so-called excavatetaxa (with the possible exception of retortamonads ortrichomonads) (Simpson, 2003; Simpson et al., 2006; Simpsonet al., 2002).
Regardless of phylogenetic position, giardial mitosis sharescommon aspects of mitosis with other eukaryotes. In this way,insights into mitotic function in Giardia are informative forunderstanding general questions of spindle organization indiverse eukaryotes as compared with commonly studiedexperimental models. In terms of the evolution of the mitoticspindle, the observations of spindle organization indiplomonads such as Giardia with the comparison with otherprotists leads to two main conclusions. First, mitosis in bothGiardia and the trichomonads occurs with extranuclearspindles that interact with chromatin across the nuclearmembrane. Therefore, this may represent a more ancestral stateof mitotic organization than either the open mitosis of plantsand metazoans or closed mitosis of fungi and trypanosomes.Second, monocentric chromosome structure is probablyancestral to holocentric chromosome structure.
At a minimum, understanding both the molecularconservation of structure and function of the dual spindles insuch a highly divergent eukaryote provides a uniqueperspective on the mechanism of mitosis and its evolution.Intriguing prospects remain to be determined; in particular,how conserved is mitosis at the molecular level in Giardia?Also, how do we account for the observed homogeneitybetween the nuclei in populations of Giardia given ourproposed nuclear migration model? In the long-term, a detailedunderstanding of both the mode and mechanism of giardial celldivision will be crucial toward developing anti-giardialcompounds that target the cell division machinery.
Materials and MethodsStrains and culture conditionsGiardia intestinalis trophozoites, strain WBC6, were grown in modified TYI-S-33medium at 37°C (Keister, 1983). Cultures were incubated in 15 ml or 6 ml plasticscrew-cap tubes (Fisher Scientific). Cell cultures were enriched for mitotic cells bygrowing 1-2 days past confluency. Fresh, warmed medium was added to the tube
and cultures were incubated at 37°C for 3-7 hours. Cells were collected at one-hourintervals to collect the maximum number of mitotic cells. We generally observed10-20% mitotic cells in each culture (supplementary material Movie 1).
GFP-tagging of essential markers in mitosisThe giardial cenH3 gene (GenBank acc. no. EAA41448), plus approximately 80 bpof upstream promoter sequence was PCR amplified from genomic DNA andsubcloned into Asc1-Age1 sites of the pMCS-GFP vector. Transformation of Giardiatrophozoites was also as previously described (Singer et al., 1998); however,electroporation conditions were modified using the GenePulserXL (Bio-Rad) at375 mV, 1000 �F and 700 ohms. Following electroporation, the cells were addedto 12 ml fresh medium and allowed to recover at 37°C overnight and selected with10 �g/ml puromycin (Calbiochem) for 4-7 days. Strains were maintained at a finalconcentration of 50 �g/ml puromycin.
Immunolocalization of the microtubule cytoskeleton with GFPfusion proteinsCells were fixed in the culture tubes with 1% paraformaldehyde, centrifuged,washed in PEM buffer (100 mM PIPES, 1 mM EGTA, 0.1 mM MgSO4) andattached to poly-L-lysine-coated coverslips. Cells attached to coverslips includecells that were both attached and unattached at the time of fixation. Cells werepermeabilized in 0.1% Triton X-100 for 10 minutes. Coverslips were washed PEMbuffer and blocked for 30 minutes in PEMBALG [PEM+ 1% bovine serum albumin(BSA), 0.1% sodium azide, 100 mM lysine and 0.5% cold-water fish skin gelatin(Sigma, St Louis, MO)]. We visualized microtubules by incubating coverslips withthe monoclonal �-tubulin antibody TAT1 (Woods et al., 1989) diluted 1:75 inPEMBALG at room temperature overnight. The TAT1 antibody was directly labeled(1:1) with a Zenon fragment conjugated with an Alexa Fluor 555 (Molecular Probes,Eugene, OR) at room temperature for 5 minutes before dilution in PEMBALG.Coverslips were washed in PEMBALG, then PEM before mounting with ProLongAntiFade with DAPI (Molecular Probes).
Direct fixation of the cenH3::GFP fusion protein was performed to visualize GFPprotein localization in conjunction with immunostaining of the cytoskeleton.Trophozoites were cultured as described above, and medium was exchanged with1� HEPES buffered saline (HeBs) for 30 minutes at 37°C, prior to fixation with1% paraformaldehyde.
Centrin antibody 20H5 (a gift of J. L. Salisbury) was directly labeled with theZenon fragment conjugated with the Alexa Fluor 555 (above). Coverslips withmitotic cells were incubated in a 1:50 dilution in PEMBALG overnight at roomtemperature. In double-labeling experiments with �-tubulin, the centrin antibodywas directly labeled with an Alexa Fluor 488-conjugated Zenon fragment. Centrinimmunostaining was performed first, followed by a brief fixation, followed bytubulin staining with TAT1 (Woods et al., 1989).
FISH of the episomal plasmid cenH3::GFP in mitotic cellsFISH probes to the episomally maintained cenH3::GFP plasmid were made byincorporation of Cy3-labeled dUTPs by nick translation (Roche) of an AscI-AgeIdigest of the cenH3::GFP plasmid (removing the cenH3 sequence). Labeled probewas precipitated in LiCl, resuspended in 100% formamide and used atapproximately 10 ng/�l final concentration in 37°C hybridizations of mitotictrophozoites as previously described (Yu et al., 2001).
Fluorescence deconvolution microscopyImages were collected using SoftWorX image acquisition software (AppliedPrecision, Issaquah, WA) on an Olympus IX70 wide-field inverted fluorescencemicroscope with an Olympus UPlanApo 100�, NA 1.35, oil-immersion objectiveand Photometrics CCD CH350 camera cooled to –35°C (Roper Scientific, Tuscon,AZ). Serial sections were acquired at 0.2 �m intervals, and data stacks weredeconvolved using the SoftWorX deconvolution software. For presentationpurposes, two-dimensional projections were created from the 3D data sets using theDeltaVision image analysis software (Applied Precision).
Transmission electron microscopyGiardia trophozoites were grown on Aclar® tabs and were fixed in 2.0%glutaraldehyde in 0.1 M cacodylate buffer, pH 7.2, overnight at room temperature,and post-fixed with 1% OsO4 + 0.8% potassium ferricyanide + 5 mM CaCl2 in 0.1M cacodylate buffer, pH 7.2, for 45 minutes. Aclar tabs with cells were dehydratedwith ethanol and infiltrated with Epon resin. Mitotic trophozoites were embeddedas previously described (Muller-Reichert et al., 2003). Briefly, trophozoites attachedto Aclar tabs were flat embedded on top of coated glass-slides and cured at 60°Cfor two days. Aclar tabs were removed and cells of interest were identified usingphase microscopy and then marked, using a diamond scribe. Regions containingmitotic cells were excised and remounted for sectioning. Ultra-thin sections (50-65nm) were collected and stained with uranyl acetate and lead citrate and imaged ina JEOL 1200 transmission electron microscope.
Taxol-treatment of trophozoitesTaxol (pacitaxel; Sigma) was stored as a 5 mg/ml stock in DMSO at –20°C. For
Journal of Cell Science 119 (23)
Jour
nal o
f Cel
l Sci
ence

4899Mitosis in Giardia
experiments determining the contribution of the microtubule cytoskeleton tochromosome segregation, taxol was diluted directly into culture tubes containingmedia to a working concentration of either 10 �M or 20 �M. An equal concentrationof DMSO was added to control cells.
We would like to acknowledge Heidi Elmendorf and Steve Singer(Georgetown University, Washington, DC), C.C. Wang and colleagues(UCSF) and contributors to the Giardia Genome Project, Mitch Sogin,Hillary Morrison and Andrew MacArthur as well as Keith Gull(Oxford University, UK) and Jeff Salisbury (Mayo Clinic) forplasmids, reagents and methodologies. We also thank members of theCande laboratory for helpful discussions. This work was supported byan NIH grant A1054693 to W.Z.C.
ReferencesAdam, R. D. (2001). Biology of Giardia lamblia. Clin. Microbiol. Rev. 14, 447-475.Adam, R. D., Nash, T. E. and Wellems, T. E. (1988). The Giardia lamblia trophozoite
contains sets of closely related chromosomes. Nucleic Acids Res. 16, 4555-4567.Amin-Hanjani, S. and Wadsworth, P. (1991). Inhibition of spindle elongation by taxol.
Cell Motil. Cytoskeleton 20, 136-144.Balasubramanian, M. K., Bi, E. and Glotzer, M. (2004). Comparative analysis of
cytokinesis in budding yeast, fission yeast and animal cells. Curr. Biol. 14, R806-R818.Baldauf, S. L. (2003). The deep roots of eukaryotes. Science 300, 1703-1706.Baldauf, S. L., Roger, A. J., Wenk-Siefert, I. and Doolittle, W. F. (2000). A kingdom-
level phylogeny of eukaryotes based on combined protein data. Science 290, 972-977.Baruch, A. C., Isaac-Renton, J. and Adam, R. D. (1996). The molecular epidemiology
of Giardia lamblia: a sequence-based approach. J. Infect. Dis. 174, 233-236.Belhadri, A. (1995). Presence of centrin in the human parasite Giardia: a further
indication of its ubiquity in eukaryotes. Biochem. Biophys. Res. Commun. 214, 597-601.
Benchimol, M. (2004a). Mitosis in Giardia lamblia: multiple modes of cytokinesis.Protist 155, 33-44.
Benchimol, M. (2004b). Participation of the adhesive disc during karyokinesis in Giardialamblia. Biol. Cell 96, 291-301.
Best, A. A., Morrison, H. G., McArthur, A. G., Sogin, M. L. and Olsen, G. J. (2004).Evolution of eukaryotic transcription: insights from the genome of Giardia lamblia.Genome Res. 14, 1537-1547.
Brugerolle, G. (1975a). Contribution à l’ étude cytologique et phylétique des diplozoaires(Zoomastigophorea, Diplozoa, Dangeard 1910). V. Nouvelle interpretation del’organisation cellulaire de Giardie. Protistologica 11, 99-109.
Brugerolle, G. (1975b). Etude de la cryptopleuromitose et de la morphogenese de divisionchez Trichomonas vaginalis et chez plusieurs genres de Trichomonadines primitives.Protistologica 11, 457-468.
Brugerolle, G. and Mignot, J. P. (2003). The rhizoplast of chrysomonads, a basal body-nucleus connector that polarises the dividing spindle. Protoplasma 222, 13-21.
Cèrva, L. and Nohynkova, E. (1992). A light microscopic study of the course of cellulardivision of Giardia intestinalis trophozoites grown in vitro. Folia Parasitol. 39, 97-104.
Ciccarelli, F. D., Doerks, T., von Mering, C., Creevey, C. J., Snel, B. and Bork, P.(2006). Toward automatic reconstruction of a highly resolved tree of life. Science 311,1283-1287.
Correa, G., Morgado-Diaz, J. A. and Benchimol, M. (2004). Centrin in Giardia lamblia– ultrastructural localization. FEMS Microbiol. Lett. 233, 91-96.
Coss, R. A. (1974). Mitosis in Chlamydomonas reinhardtii basal bodies and the mitoticapparatus. J. Cell Biol. 63, 325-329.
Dacks, J. B., Marinets, A., Ford Doolittle, W., Cavalier-Smith, T. and Logsdon, J.M., Jr (2002). Analyses of RNA Polymerase II genes from free-living protists:phylogeny, long branch attraction, and the eukaryotic big bang. Mol. Biol. Evol. 19,830-840.
De Brabander, M., Geuens, G., Nuydens, R., Willebrords, R., Aerts, F. and De Mey,J. (1986). Microtubule dynamics during the cell cycle: the effects of taxol andnocodazole on the microtubule system of Pt K2 cells at different stages of the mitoticcycle. Int. Rev. Cytol. 101, 215-274.
Dutcher, S. K. (2003). Elucidation of basal body and centriole functions inChlamydomonas reinhardtii. Traffic 4, 443-451.
Ehler, L. L., Holmes, J. A. and Dutcher, S. K. (1995). Loss of spatial control of themitotic spindle apparatus in a Chlamydomonas reinhardtii mutant strain lacking basalbodies. Genetics 141, 945-960.
Elmendorf, H. G., Dawson, S. C. and McCaffery, J. M. (2003). The cytoskeleton ofGiardia lamblia. Int. J. Parasitol. 33, 3-28.
Filice, F. P. (1952). Studies on the cytology and life history of a Giardia from thelaboratory rat. Univ. Calif. Publ. Zool. 57, 53-146.
Ghosh, S., Frisardi, M., Rogers, R. and Samuelson, J. (2001). How Giardia swim anddivide. Infect. Immun. 69, 7866-7872.
Gillin, F. D., Reiner, D. S. and McCaffery, J. M. (1996). Cell biology of the primitiveeukaryote Giardia lamblia. Annu. Rev. Microbiol. 50, 679-705.
Gomez-Conde, E., Mena-Lopez, R., Hernandez-Jauregui, P., Gonzalez-Camacho, M.and Arroyo, R. (2000). Trichomonas vaginalis: chromatin and mitotic spindle duringmitosis. Exp. Parasitol. 96, 130-138.
Graczyk, T. K. (2005). Is Giardia a living fossil? Trends Parasitol. 21, 104-107.Hansen, W. R., Tulyathan, O., Dawson, S. C., Cande, W. Z. and Fletcher, D. A. (2006).
Giardia lamblia attachment force is insensitive to surface treatments. Eukaryotic Cell5, 781-783.
Heath, I. B. (1980). Variant mitoses in lower eukaryotes: indicators of the evolution ofmitosis? Int. Rev. Cytol. 64, 1-80.
Jaspersen, S. L. and Winey, M. (2004). The budding yeast spindle pole body: structure,duplication, and function. Annu. Rev. Cell Dev. Biol. 20, 1-28.
Jordan, M. A., Toso, R. J., Thrower, D. and Wilson, L. (1993). Mechanism of mitoticblock and inhibition of cell proliferation by taxol at low concentrations. Proc. Natl.Acad. Sci. USA 90, 9552-9556.
Kabnick, K. S. and Peattie, D. A. (1990). In situ analyses reveal that the two nuclei ofGiardia lamblia are equivalent. J. Cell Sci. 95, 353-360.
Keister, D. B. (1983). Axenic culture of Giardia lamblia in TYI-S-33 mediumsupplemented with bile. Trans. R. Soc. Trop. Med Hyg. 77, 487-488.
Knight, J. (2004). Giardia: not so special, after all? Nature 429, 236-237.Lechtreck, K. F. and Grunow, A. (1999). Evidence for a direct role of nascent basal
bodies during spindle pole initiation in the green alga Spermatozopsis similis. Protist150, 163-181.
Levy, Y. Y., Lai, E. Y., Remillard, S. P., Heintzelman, M. B. and Fulton, C. (1996).Centrin is a conserved protein that forms diverse associations with centrioles andMTOCs in Naegleria and other organisms. Cell Motil. Cytoskeleton 33, 298-323.
Lu, S. Q., Baruch, A. C. and Adam, R. D. (1998). Molecular comparison of Giardialamblia isolates. Int. J. Parasitol. 28, 1341-1345.
Malik, H. S. and Henikoff, S. (2003). Phylogenomics of the nucleosome. Nat. Struct.Biol. 10, 882-891.
Marshall, W. F. and Rosenbaum, J. L. (2000). How centrioles work: lessons from greenyeast. Curr. Opin. Cell Biol. 12, 119-125.
McDonald, K. L., O’Toole, E. T., Mastronarde, D. N. and McIntosh, J. R. (1992).Kinetochore microtubules in PTK cells. J. Cell Biol. 118, 369-383.
Meng, T. C., Aley, S. B., Svard, S. G., Smith, M. W., Huang, B., Kim, J. and Gillin,F. D. (1996). Immunolocalization and sequence of caltractin/centrin from the earlybranching eukaryote Giardia lamblia. Mol. Biochem. Parasitol. 79, 103-108.
Muller-Reichert, T., Hohenberg, H., O’Toole, E. T. and McDonald, K. (2003).Cryoimmobilization and three-dimensional visualization of C. elegans ultrastructure.J. Microsc. 212, 71-80.
Nohynkova, E., Draber, P., Reischig, J. and Kulda, J. (2000). Localization of gamma-tubulin in interphase and mitotic cells of a unicellular eukaryote, Giardia intestinalis.Eur. J. Cell Biol. 79, 438-445.
Nohynkova, E., Tumova, P. and Kulda, J. (2006). Cell division of giardia intestinalis:flagellar developmental cycle involves transformation and exchange of flagella betweenmastigonts of a diplomonad cell. Eukaryotic Cell 5, 753-761.
O’Toole, E. T., Giddings, T. H., McIntosh, J. R. and Dutcher, S. K. (2003). Three-dimensional organization of basal bodies from wild-type and delta-tubulin deletionstrains of Chlamydomonas reinhardtii. Mol. Biol. Cell 14, 2999-3012.
Ogbadoyi, E., Ersfeld, K., Robinson, D., Sherwin, T. and Gull, K. (2000). Architectureof the Trypanosoma brucei nucleus during interphase and mitosis. Chromosoma 108,501-513.
Ou, Y. and Rattner, J. B. (2004). The centrosome in higher organisms: structure,composition, and duplication. Int. Rev. Cytol. 238, 119-182.
Parness, J. and Horwitz, S. B. (1981). Taxol binds to polymerized tubulin in vitro. J.Cell Biol. 91, 479-487.
Raikov, I. B. (1994). The diversity of forms of mitosis in protozoa: a comparative review.Eur. J. Protistol. 30, 252-269.
Ramesh, M. A., Malik, S. B. and Logsdon, J. M., Jr (2005). A phylogenomic inventoryof meiotic genes; evidence for sex in Giardia and an early eukaryotic origin of meiosis.Curr. Biol. 15, 185-191.
Rao, S., Orr, G. A., Chaudhary, A. G., Kingston, D. G. and Horwitz, S. B. (1995).Characterization of the taxol binding site on the microtubule. 2-(m-Azidobenzoyl)taxolphotolabels a peptide (amino acids 217-231) of beta-tubulin. J. Biol. Chem. 270, 20235-20238.
Salisbury, J. L. (1995). Centrin, centrosomes, and mitotic spindle poles. Curr. Opin. CellBiol. 7, 39-45.
Salisbury, J. L., Baron, A. T. and Sanders, M. A. (1988). The centrin-basedcytoskeleton of Chlamydomonas reinhardtii: distribution in interphase and mitoticcells. J. Cell Biol. 107, 635-641.
Savioli, L., Smith, H. and Thompson, A. (2006). Giardia and cryptosporidium join the‘neglected diseases initiative’. Trends Parasitol. 22, 203-208.
Schuster, F. L. (1975). Ultrastructure of mitosis in the amoeboflagellate Naegleriagruberi. Tissue Cell 7, 1-11.
Simpson, A. G. (2003). Cytoskeletal organization, phylogenetic affinities and systematicsin the contentious taxon Excavata (Eukaryota). Int. J. Syst. Evol. Microbiol. 53, 1759-1777.
Simpson, A. G., Roger, A. J., Silberman, J. D., Leipe, D. D., Edgcomb, V. P., Jermiin,L. S., Patterson, D. J. and Sogin, M. L. (2002). Evolutionary history of “early-diverging” eukaryotes: the excavate taxon Carpediemonas is a close relative of Giardia.Mol. Biol. Evol. 19, 1782-1791.
Simpson, A. G., Inagaki, Y. and Roger, A. J. (2006). Comprehensive multigenephylogenies of excavate protists reveal the evolutionary positions of “primitive”eukaryotes. Mol. Biol. Evol. 23, 615-625.
Singer, S. M., Yee, J. and Nash, T. E. (1998). Episomal and integrated maintenance offoreign DNA in Giardia lamblia. Mol. Biochem. Parasitol. 92, 59-69.
Sogin, M. L., Gunderson, J. H., Elwood, H. J., Alonso, R. A. and Peattie, D. A. (1989).Phylogenetic meaning of the kingdom concept: an unusual ribosomal RNA fromGiardia lamblia. Science 243, 75-77.
Jour
nal o
f Cel
l Sci
ence

4900
Solari, A. J., Rahn, M. I., Saura, A. and Lujan, H. D. (2003). A unique mechanism ofnuclear division in Giardia lamblia involves components of the ventral disk and thenuclear envelope. Biocell 27, 329-346.
Sullivan, B. A., Blower, M. D. and Karpen, G. H. (2001). Determining centromereidentity: cyclical stories and forking paths. Nat. Rev. Genet. 2, 584-596.
Van de Peer, Y., Baldauf, S. L., Doolittle, W. F. and Meyer, A. (2000). An updated andcomprehensive rRNA phylogeny of (crown) eukaryotes based on rate-calibratedevolutionary distances. J. Mol. Evol. 51, 565-576.
Van Hooser, A. A., Ouspenski, I. I., Gregson, H. C., Starr, D. A., Yen, T. J., Goldberg,M. L., Yokomori, K., Earnshaw, W. C., Sullivan, K. F. and Brinkley, B. R. (2001).Specification of kinetochore-forming chromatin by the histone H3 variant CENP-A. J.Cell Sci. 114, 3529-3542.
Wiesehahn, G. P., Jarroll, E. L., Lindmark, D. G., Meyer, E. A. and Hallick, L. M.(1984). Giardia lamblia: autoradiographic analysis of nuclear replication. Exp.Parasitol. 58, 94-100.
Winey, M., Mamay, C. L., O’Toole, E. T., Mastronarde, D. N., Giddings, T. H.,
McDonald, K. L. and McIntosh, J. R. (1995). Three-dimensional ultrastructuralanalysis of the Saccharomyces cerevisiae mitotic spindle. J. Cell Biol. 129, 1601-1615.
Woods, A., Sherwin, T., Sasse, R., MacRae, T. H., Baines, A. J. and Gull, K. (1989).Definition of individual components within the cytoskeleton of Trypanosoma bruceiby a library of monoclonal antibodies. J. Cell Sci. 93, 491-500.
Wright, R. L., Salisbury, J. and Jarvik, J. W. (1985). A nucleus-basal body connectorin Chlamydomonas reinhardtii that may function in basal body localization orsegregation. J. Cell Biol. 101, 1903-1912.
Wright, R. L., Adler, S. A., Spanier, J. G. and Jarvik, J. W. (1989). Nucleus-basalbody connector in Chlamydomonas: evidence for a role in basal body segregation andagainst essential roles in mitosis or in determining cell polarity. Cell Motil.Cytoskeleton 14, 516-526.
Yu, L. Z., Birky, C. W., Jr and Adam, R. D. (2002). The two nuclei of giardia eachhave complete copies of the genome and are partitioned equationally at cytokinesis.Eukaryotic Cell 1, 191-199.
Journal of Cell Science 119 (23)
Jour
nal o
f Cel
l Sci
ence